
Caracterización de Siete Especies Marino-Costeras de Importancia Estratégica en Centroamérica

CARACTERIZACIÓN DE ESPECIES DE MALASSEZIA COMO MICROBIOTA
COMENSAL EN LA PIEL DE EQUINOS
DIANA MILENA RODRÍGUEZ SANDOVAL
PONTIFICIA UNIVERSIDAD JAVERIANA
FACULTAD DE CIENCIAS
CARRERA DE BACTERIOLOGÍA
BOGOTÁ
2014

CARACTERIZACIÓN DE ESPECIES DE MALASSEZIA COMO MICROBIOTA
COMENSAL EN LA PIEL DE EQUINOS
DIANA MILENA RODRÍGUEZ SANDOVAL
TRABAJO DE GRADO
Presentado como requisito
Para optar al título de
BACTERIÓLOGA
DIRECTOR:
Dra. RUBIELA CASTAÑEDA SALAZAR M.V., MSc.
CO-DIRECTOR:
Dra. MELVA YOMARY LINARES LINARES BACT., MSc.
PONTIFICIA UNIVERSIDAD JAVERIANA
FACULTAD DE CIENCIAS
CARRERA DE BACTERIOLOGÍA
BOGOTÁ
2014

NOTA DE ADVERTENCIA
Resolución No 13 de Julio de 1946.
“La universidad no se hace responsable por los conceptos emitidos por sus alumnos en
sus trabajos de tesis. Solo velará porque no se publique nada contrario al dogma y a la
moral católica y porque las tesis no contengan ataques personales contra persona alguna,
antes bien se vea en ellas el anhelo de buscar la verdad y la justicia.”

DEDICATORIA
La dedico profundamente a Dios y a la Virgen por darme fortaleza, e iluminar cada día de mi vida
A mi Madre por su dedicación, amor, comprensión y esfuerzo por brindarme un buen futuro
A mi Primo Hermano Carlos Alberto por su dedicación, apoyo, y compañía,
A mis Abuelitos Bárbara, Rita y Leonardo por su amor, ejemplo y apoyo incondicional
A mi Novio Juan Pablo Ramírez por su colaboración, amor y paciencia
A mi Padre, Tíos y Primos por ser parte fundamental en mi crecimiento

AGRADECIMIENTOS
Agradezco a mis profesores que a lo largo de mi carrera han dejado su huella plasmada y me han inculcado grandes valores, convirtiéndome en una persona perseverante y triunfadora
A la Universidad Javeriana y al grupo de investigación UNIDIA los cuales me permitieron la realización de este trabajo
A la Dra. Rubiela Castañeda Salazar por sus conocimientos, por creer en mí, por su apoyo, su paciencia,
dedicación y cariño
A la Dra. Melva Yomary Linares Linares por su apoyo, su valiosa orientación, por sus conocimientos, su paciencia y cariño
A mis amigos Sabine Damme y Juan Camilo Galvis por su compañía, consejos, amistad y por todos los
momentos que compartimos para lograr juntos nuestro objetivo

TABLA DE CONTENIDO
PAGINA
RESUMEN 1
1. INTRODUCCIÓN, PLANTEAMIENTO DEL PROBLEMA, JUSTIFICACIÓN 2
2. MARCO TEORICO 3
2.1 Características del genero Malassezia 3
2.2 Identificación Macroscópica y Microscópica del genero Malassezia 4
2.3. Identificación de especies del genero Malassezia 4
2.3.1 Pruebas bioquímicas 5
2.3.2 Otras pruebas complementarias 7
2.4 Malassezia en Equinos 8
3. OBJETIVOS 10
3.1 Objetivo General 10
3.2 Objetivos Específicos 10
4. METODOLOGÍA 11
4.1 Población de estudio 11
4.2 Muestreo, transporte y procesamiento de las muestras 11
4.3
4.4
Control de calidad y Mantenimiento de las cepas
Análisis de la información
13
13
5. RESULTADOS Y DISCUSIÓN 13
5.1 Examen directo 13
5.2 Descripción macroscópica 14
5.3 Descripción microscópica 15
5.4 Identificación de especies del genero Malassezia mediante pruebas
bioquímicas
16
6. CONCLUSIONES 23
7. RECOMENDACIONES 23
8. REFERENCIAS 24

ÍNDICE DE TABLAS
PÁGINA
Tabla 1. Identificación de las especies del genero Malassezia 8
Tabla 2. Descripción macroscópica de las colonias identificadas como
Malassezia
14
Tabla 3. Mediciones microscópicas de las especies identificadas como
Malassezia a partir de los aislamientos obtenidos.
16
Tabla 4. Identificación de los aislamientos obtenidos de acuerdo con las
pruebas bioquímicas.
19
Tabla 5. Especies aisladas según variables como raza, género y edad. 22

ÍNDICE DE FIGURAS
PÁGINA
.
Figura 1. Clasificación de las especies del Género Malassezia según el requerimiento de lípidos para su crecimiento.
4
Figura 2. Identificación bioquímica de especies de Malassezia
6
Figura 3. Distribución de las especies de Malassezia en diferentes
localizaciones de la piel en equinos.
9
Figura 4.
Medidas microscopicas identificadas como Malassezia Tinción de Gram 100x
15
Figura 5. Resultados pruebas bioquímicas aislamientos identificados como Malassezia
17

1
CARACTERIZACIÓN DE ESPECIES DE MALASSEZIA COMO MICROBIOTA
COMENSAL EN LA PIEL DE EQUINOS
RESUMEN
Objetivo. Caracterizar las especies del género Malassezia en la piel y canal auditivo
externo de equinos remitidos a la Clínica de Grandes Animales de la Facultad de
Medicina Veterinaria y de Zootecnia de la Universidad Nacional de Colombia. Materiales
y métodos. Se obtuvieron 82 muestras de hisopados de canal auditivo externo (izquierdo
y derecho), así como de diferentes regiones de la piel que incluyeron: prepucio, glándula
mamaria e ingle, a partir de 22 equinos remitidos a la Clínica de Grandes Animales de la
Facultad de Medicina Veterinaria y de Zootecnia de la Universidad Nacional de Colombia,
durante el periodo comprendido entre Diciembre de 2013 y Abril de 2014. Las muestras
fueron procesadas mediante examen directo, y cultivo en agar Dixon modificado. A partir
de los crecimientos en los que se observaron colonias morfológicamente compatibles con
Malassezia spp. se evaluaron características macroscópicas y microscópicas como
tamaño, textura, color, forma, borde y superficie de la colonia, se realizaron pruebas
bioquímicas como catalasa, ureasa, asimilación de Tweenes, producción de β-
glucosidasa, cremophor-EL y pruebas complementarias como crecimiento en agar
Sabouraud, producción de fosfolipasa en agar Sabouraud con yema de huevo al 10% y
crecimiento a 37°C y a 40°C. Cepas de referencia. Se utilizaron las cepas de referencia
M. furfur CBS 7019, M. pachydermatis CBS 1879,M. sympodialis CBS 7222, M. slooffiae
CBS 7956 pertenecientes a la colección de microorganismos del Grupo de Micología y
Fitopatología de la Universidad de Los Andes. Resultados. De las 82 muestras
procesadas, el examen directo fue positivo en 17,07% (n=14), observando estructuras
levaduriformes de formas y tamaños diferentes. Al evaluar las características del cultivo y
realizar las pruebas bioquímicas y complementarias, 8,53% (n=7) fueron identificadas
como Malassezia, de las cuales 6,09% (n=5) se pudieron caracterizar a nivel de especie
como M, sympodialis, M. pachydermatis, M, slooffiae, M. obtusa, M globosa y 2,44%(n=2)
como Malassezia spp. Adicionalmente, se identificaron otras levaduras como Candida
spp. en 6,10% (n=5) y Cryptococcus spp en 2,44% (n=2) de las muestras. Conclusión.
La prevalencia de Malassezia establecida para la población evaluada fue del 18,2%
(n=4), logrando caracterizar hasta especie M. sympodialis, M. pachydermatis, M. slooffiae
M. obtusa, M. globosa como microbiota normal de la piel en los equinos evaluados.

2
1. INTRODUCCIÓN, PLANTEAMIENTO DEL PROBLEMA Y JUSTIFICACIÓN
El género Malassezia comprende un grupo de levaduras unicelulares que forman parte de
la microbiota fúngica normal de la piel de humanos y animales de sangre caliente
(Kaneko, 2007). Fisiológicamente tienen afinidad por los lípidos, los cuales utilizan como
única fuente de carbono, a excepción de Malassezia pachydermatis, la cual es no
lipodependiente (Hernández, 2005).
Se han descrito casos en los que esta microbiota comensal puede ocasionar patologías
en humanos como: pitiriasis versicolor (PV), dermatitis seborreica (DS) y dermatitis
atópica (DA), comportándose como patógeno oportunista si se presenta alteración en la
piel u otros factores que puedan favorecer el paso de flora comensal a patógena como
humedad, desórdenes en la queratinización e inmunosupresión, entre otros (Hernández,
2005).
En los animales, Malassezia ha adquirido importancia por su asociación a procesos
patológicos como patógeno oportunista emergente (Hernández,2005), siendo M.
pachydermatis el agente etiológico aislado con mayor frecuencia en diferentes patologías
como otitis externa en los caninos y dermatitis en los felinos (Garau,2005 ;Castellá et al.
2005) recientemente, dos nuevas especies han sido descritas como flora comensal en la
piel de animales sanos, M. caprae (caprinos) y M. equina (equinos).(Cabañes et al. 2007).
En Colombia no existen reportes sobre la presencia de Malassezia spp como flora
comensal en la piel de los equinos, motivo por el cual en este estudio se propone realizar
una caracterización preliminar de las especies de Malassezia presentes en la piel de los
equinos remitidos a la Clínica de Grandes animales de la Facultad de Medicina
Veterinaria y de Zootecnia de la Universidad Nacional de Colombia durante el período
comprendido entre Diciembre de 2013 a Abril de 2014.

3
2. MARCO TEÓRICO
2.1 Características del género Malassezia
Las levaduras del género Malassezia se clasifican dentro del phylum Basidiomicota,
subphylum Ustilaginomycotina, clase Exobasidiomycetes, orden Malasseziales, (Erchiga
et al. 2008), familia Malasseziaceae (Gaitanis et al. 2012). Estas levaduras hacen parte de
la microbiota comensal cutánea de humanos y animales (Hernández, 2005) y se
reproducen por gemación unipolar dejando una prominente y característica cicatriz en la
célula madre (Giusiano, 2006).
Fisiológicamente, Malassezia se caracteriza por ser lipidodependiente, debido a que tiene
un defecto en la capacidad de sintetizar ácidos grasos saturados C12–C16, lo que se
manifiesta en el requerimiento de una fuente exógena de estos ácidos para su desarrollo,
a excepción de M. pachydermatis que es la única especie del género que no requiere de
sustancias lipídicas para su crecimiento (Giusiano, 2006). Por lo anterior, los medios
comúnmente utilizados son Agar Dixon y Agar Leeming y Notman, medios selectivos que
son efectivos para la recuperación y aislamiento de las levaduras del genero Malassezia
(Hernández 2005; Kaneko et al. 2007).
Hasta el momento se han descrito 14 especies, de las cuales 13 son lipodependientes y
una no lipodependiente (Figura 1). En animales se han reportado diferentes especies
lipodependientes, como M. sloofiae, M. globosa, M. sympodialis y la especie no
lipodependiente M. pachydermatis en el conducto auditivo externo de caninos y felinos;
(Salah et al 2010); M. nana en bovinos con o sin otitis externa (Hirai et al. 2004) y M.
sympodialis, M. slooffiae, M. furfur y M. pachydermatis en el conducto auditivo externo de
porcinos (Nardoni et al. 2010). Adicionalmente, en la piel de equinos, en la región
intermamaria en las yeguas, así como en la fosa prepucial en machos, se ha reportado la
presencia de Malassezia equina (White et al. 2006). Otros reportes de literatura describen
la presencia de diferentes especies de Malassezia asociadas a varios sitios anatómicos:
M. pachydermatis, en ingle, dorso, ano; M. furfur en axila ingle dorso ano, M. restricta en
axila, ingle, M. sympodialis en axila, M. slooffiae en ingle, ano, M .obtusa en ano y canal
auditivo externo, M. globosa en axila, ingle y ano. (White 2006; Crespo et al. 2002).

4
Figura 1. Clasificación de las especies del género Malassezia según el requerimiento de
lípidos para su crecimiento.
2.2 Identificación Macroscópica y Microscópica del genero Malassezia
La morfología macroscópica es variable, observándose colonias pequeñas, de aspecto
cremoso a amarillo, algunas veces lisas o ligeramente rugosas umbilicadas; brillantes u
opacas, planas, con borde liso, textura cremosa, esféricas, ovales o cilíndricas y de
tamaños diferentes de acuerdo con la especie evaluada (Hernández , 2005).
Al microscopio se aprecian Blastoconidias que pueden ser ligeramente alargadas, u
ovales de 2-4 μm de largo por 1-2 μm de ancho aproximadamente, con gemas pequeñas.
(Bonifaz, 2012)
2.3 Identificación de especies del género Malassezia
Para la identificación de aislamientos del género Malassezia spp, se realizan diferentes
pruebas de laboratorio que permiten llegar a identificar la especie entre ellas se citan las
siguientes:
Especies de Malassezia
Lipodepedientes
M.furfur (Baillon,1889)
M.globosa,M.obtusa,M.restricta,M.sloofiae
(Guého,Midgley & Guillot ,1.996)
M.sympodialis ( Simmons & Guého, 1990)
M.dermatis, M.japónica,M.yamatoensis
( Sugita et al.2003)
M.nana ( Hirai A, et al.2004)
M.caprae,M.equina(Cabañes FJ, et al.2007)
M.cuniculi ( Cabañes et al.2011)
No Lipodependiente
M.pachydermatis
(Dodge, 1935)

5
2.3.1 Pruebas bioquímicas
- Catalasa: Esta prueba permite evaluar si el microorganismo posee o no la enzima
catalasa que descompone el peróxido de hidrogeno en oxígeno y agua. La prueba se
considera positiva, si se observa formación de burbujas (Khosravi et al. 2008), siendo M.
restricta la única especie de este género que es catalasa negativa (Guého et al. 1996,
Hernández, 2005; Kaneko et al. 2007; Cabañes 2011), aunque la reacción de M.
pachydermatis para esta prueba es variable (Khosravi et al. 2008)
- Ureasa: Por medio de esta prueba se evalúa la capacidad del microorganismo para
hidrolizar la úrea formando dos moléculas de amoniaco por medio de la reacción
enzimática de la ureasa. La prueba se considera positiva si el medio vira a color fucsia
debido al indicador rojo de fenol (Pérez et al. 2002).
- β- glucosidasa: Prueba que se utiliza para evidenciar si el microorganismo produce o
no la enzima β- glucosidasa, que hidroliza la beta esculina en glucosa y esculetina, por
reacción con una sal de hierro formando un complejo de color castaño oscuro o negro
(Mayzer et al. 1997). Para esta prueba se considera una reacción positiva el viraje a color
negro en el medio de cultivo (Khosravi et al. 2008).
- Asimilación del Cremophor-EL: Esta prueba determina la capacidad del
microorganismo para asimilar el Cremophor-EL (Aceite de castor). Se considera una
reacción positiva si luego de 5 días de incubación a 32°C se observa alrededor del pozo
un halo de asimilación y una reacción negativa si hay ausencia del mismo (Figura 2)
(Kaneko et al. 2007). Siendo M. furfur la única especie reportada como positiva (Mayzer,
1997; Hernández, 2005; Giusianio, 2006), aunque últimamente para M.pachydermatis se
ha reportado una reacción variable para asimilar el Cremophor (Ashbee, 2007).
- Asimilación del Tween: Esta prueba evalúa la capacidad del microorganismo para
asimilar suplementos nutricionales lipídicos como Tween 20, 40, 60, 80 (Khosravi, 2008).
La lectura se realiza evidenciando la asimilación de cada tween por parte del
microorganismo después de 7 días de incubación a 32°C (Figura 2) (Hernández, 2005;
Kaneko et al. 2007), cada especie presenta distinta afinidad por los Tweenes: M. furfur y
M. dermatis asimilan todos los Tweens, M. sympodialis y M. pachydermatis no asimilan el
Tween 20, M. obtusa, M. globosa y M. restricta no asimilan ningún Tween (Guého et al.
1996; Giusianio, 2006; Cabañes et al. 2007; Crespo et al. 2008; Nardoni et al. 2010; Salah
et al. 2010).

6
Esculina (+) M. obtusa
M.sympodialis
Esculina (-) M.furfur M. slooffiae
M. restricta, M. globosa
MALASSEZIA
Cultivos en Agar Sabouraud Glucosa
Tween test
M.sympodialis,M.caprae M.furfur M.slooffiae, M.nana M.obtusa, M.restricta,
M.globosa,
M.japonica,M.cuniculi
M. yamatoensis, M.dermatis,
M.equina
M.pachydermatis
80
0
20
0
60
0
40
0
Crecimiento
No crecimiento Cremophor- EL
Interpretación:
Figura2. Identificación bioquímica de especies de Malassezia Tomada y adaptada : (Hernández, 2005)
Positivo M.pachydermatis Reacción catalasa (-)
M.restricta

7
2.3.2 Otras pruebas complementarias
- Crecimiento en Agar Sabouraud: Por medio de esta prueba se evalúa la capacidad del
microorganismo para crecer en agar Sabouraud sin suplementos lipídicos (Guého et al.
1996), siendo M. pachydermatis la única especie de este género que crece en agar
Sabouraud (Guého et al. 1996; Sugita et al. 2003; Hirai et al. 2004; Hernádez ,2005;
Kaneko, 2007; Cabañes &Castellá, 2011).
- Producción de fosfolipasa en agar Sabouraud con yema de huevo: Permite
determinar si el microorganismo produce la enzima fosfolipasa (Pini et al. 2011). La
lectura de la prueba se realiza mediante la medición del halo formado alrededor de la
colonia (Ortiz, 2012). Los resultados de la actividad enzimática se expresan como una
razón (Pz) que se determina midiendo el diámetro de la colonia y el diámetro de la colonia
más el halo de hidrólisis que se generó alrededor de ella (Price et al. 1982; Pini et al.
2011). Se han establecido rangos que permiten clasificar la actividad fosfolipasa así:
PZ<0.64: muy alta, PZ ≥ 0.64 y <1: alta y PZ=1: nula (Cafarchia & Otranto, 2004). La
actividad fosfolipasa juega un papel importante como factor de virulencia, una alta
actividad fosfolipasa puede estar involucrada en el mecanismo por el cual estas levaduras
pueden generar procesos inflamatorios (Ortiz, 2013).
- Crecimiento en agar Dixon a temperaturas de 37 y 40 oC: Otra forma para poder
identificar las especies del genero Malassezia es por la capacidad del microorganismo
para crecer a diferentes temperaturas (Guého et al. 1996; Hernández, 2005) a 37°c se ha
reportado crecimiento débil para las especies M. globosa, M. obtusa, M.restricta, M.
equina y M.nana; buen crecimiento para: M.pachydermatis, M.furfur M. sympodialis, M.
slooffiae, M. dermatis, M, japónica, M. yamatoensis y M. cuniculi. Mientras que a 40°C se
ha reportado crecimiento para todas las especies excepto M. globosa, M. obtusa, M.
restricta, M. japonica, M. equina y M. caprae. Para las dos temperaturas se ha reportado
crecimiento para las especies M. furfur, M. pachydermatis, M. sympodialis, M. slooffiae, M.
dermatis, M. yamatoensis y M. cuniculi (Guého et al. 1996; Sugita et al. 2003; Hirai et al.
2004; Hernández, 2005; Kaneko, 2007; Cabañes, 2011).
En la tabla 1 se resumen los resultados de las pruebas bioquímicas y complementarias
para la identificación de las especies de Malassezia.

8
Tabla 1. Identificación de las especies del genero Malassezia
(+) Crecimiento, (-) no crecimiento, +/- Crece débilmente
Tomada y adaptada de (Guého et al. 1996; Sugita et al.2003; Hirai et al.2004;Hernández, 2005; Cabañes, 2011)
2.4 Malassezia en Equinos
Las infecciones de tipo micótico son de rara ocurrencia en equinos, a excepción de las
ocasionadas por Dermatofitos (Pérez et al. 2000); sin embargo actualmente existen
reportes sobre la participación de levaduras del género Malassezia que han ido
adquiriendo importancia debido a que aunque se pueden encontrar como microbiota
comensal en diferentes áreas del cuerpo, pueden causar o estar asociadas a
determinados desórdenes dermatológicos (Cafarchia, 2013), siendo las levaduras
lipodependientes las más comúnmente reportadas como flora normal (Crespo et al. 2002).
Aunque existen pocos estudios sobre la presencia de Malassezia spp como microbiota
comensal en diferentes regiones anatómicas de la piel de los equinos, algunos reportes
evidencian la presencia de especies de Malassezia en diferentes localizaciones de la
siguiente forma: M. pachydermatis, en ingle, dorso, ano, canal auditivo externo; M.furfur
Especie
Catalasa
Urea
β- glucosidasa
Sabouraud
40ºC
37ºC
Tween Cremophor 20 40 60 80
M. slooffiae +++ + - - + + + + + - -
M. sympodialis +++ + + - + + - + + + -
M. obtusa +++ + + - - + - - - - -
M. furfur +++ + - - + + + + + + +
M. pachydermatis +++ + - + + + - + + + +/-
M.globosa +++ + - - - - - - - - -
M.restricta - + - - - - - - - - -
M.equina +++ + - - - - - + + + -
M.dermatis +++ + - - + + + + + + -
M.nana +++ + - - +/- +/- +o- + + +/- -
M.japonica +++ + v - - + - +/- + - -
M.yamatoensis +++ + v - + + + + + + -
M.caprae +++ + - - +/- - - +/- +/- +/- -
M.cuniculi +++ + + - + + - - - - -

9
en axila, ingle, dorso, ano y canal auditivo externo; M. restricta en axila, ingle y canal
auditivo externo, M.sympodialis en axila; M. slooffiae en ingle, ano, canal auditivo externo;
M .obtusa en ano y canal auditivo externo y M. globosa en axila, ingle y ano (White 2006;
Crespo et al. 2002). Adicionalmente, Cabañes & Boekhout en el año 2007, reportaron el
aislamiento de una nueva especie de Malassezia a partir de muestras del ano de un
equino sano, la cual fue clasificada como M. equina, que se caracteriza
macroscópicamente por presentar colonias rugosas color crema de 2-3 mm, con células
elipsoidales u ovales, de 3-4.5 x 2.2-3.5 μm , catalasa positiva, β- glucosidasa negativa,
asimila los Tweenes 40, 60 y 80, no asimila el cremophor EL, presenta un crecimiento
débil a 37°C; se considera que guarda una relación con M. sympodialis ya que se ha
demostrado un grado de similitud en sus nucleótidos al realizarles análisis moleculares en
la secuencia D1/D2 en los dominios del gen 26S rRNA en la región ITS 1 (Cabañes et al.
2007). La distribución de las diferentes especies según el sito anatómico se detalla en la
figura 3.
Tomada de: http://www.howrse.co.uk/
Adaptada de: (Crespo et al. 2002)
Figura 3. Distribución de las especies de Malassezia en diferentes localizaciones de la
piel en equinos.
Ano M. pachydermatis, M.slooffiae, M.globosa. M obtusa, M.furfur, M.equina
Dorso M. pachydermatis, M.furfur
Axila M.furfur, M.sympodialis
M.globosa, M.restricta
Ingle
M. pachydermatis
M.furfur
M.slooffiae, M.globosa,
M.restricta
Canal auditivo externo
M. pachydermatis, M.furfur M.slooffiae, M.obtusa,
M.restricta

10
En cuanto a la participación de Malassezia en procesos patológicos en equinos, existen
dos reportes de caso. En el primero de ellos, un macho presentaba lesiones dérmicas
en la cara, el cual tenía una condición pre existente de Alopecia areata, sin embargo, al
realizar examen directo y crecimiento en agar sabouraud se identificaron levaduras del
genero Malassezia spp, las cuales fueron consideradas como un patógeno oportunista,
por lo cual se enfatizó sobre la necesidad de estudiar el papel de Malassezia spp como
microbiota normal en la piel de los equinos o si está relacionada como patógeno en un
proceso de enfermedad como el reportado en este caso. (Paterson ,2002)
En el otro caso reportado, se examinaron yeguas que presentaban prurito intenso en la
glándula mamaria a las cuales les fue realizado un examen citológico que reveló
numerosas levaduras las cuales no pudieron ser identificadas hasta especie, lo cual
sugeriría su posible papel como patógeno oportunista en estos animales; sin embargo se
examinaron también las yeguas sanas, a las cuales se les identificó un gran número de
levaduras en la glándula mamaria las cuales sí pudieron ser identificadas hasta especie
encontrándose M. slooffiae y M equina (White, 2005). Posteriormente, en otro estudio
realizado en 11 equinos (5 hembras y 6 machos castrados) se aisló a partir de la piel, en
particular en la región intermamaria de las yeguas y la fosa prepucial de los machos, 7
levaduras pertenecientes al género Malassezia, dentro de las cuales predominó la
especie M. equina con lo cual corroboraron que las levaduras de este género hacían parte
de la microbiota normal de los equinos evaluados (White et al. 2006).
3. OBJETIVOS
3.1 OBJETIVO GENERAL: Caracterizar las especies del género Malassezia en el canal
auditivo externo y la piel de equinos remitidos a la Clínica de Grandes Animales de la
Facultad de Medicina Veterinaria y de Zootecnia de la Universidad Nacional de Colombia.
3.2 OBJETIVOS ESPECÍFICOS:
Identificar aislamientos de Malassezia spp en muestras de canal auditivo externo y
piel de equinos sanos.
Determinar la prevalencia de Malassezia spp. en piel de equinos remitidos a la
Clínica de Grandes animales de la Facultad de Medicina Veterinaria y de
Zootecnia de la Universidad Nacional de Colombia durante el periodo comprendido
entre Diciembre de 2013 a Abril de 2014.
Relacionar la presencia de Malassezia spp.frente a variables de edad, sexo y raza.

11
4. METODOLOGÍA
4.1 Población de estudio: Para este estudio se evaluaron 22 Equinos remitidos a la
Clínica de Grandes Animales (CGA) de la Facultad de Medicina Veterinaria y de
Zootecnia de la Universidad Nacional de Colombia, durante los meses de Diciembre de
2013 a Abril de 2014.
4.2 Muestreo, Transporte y Procesamiento de las muestras: Se realizaron hisopados
de la piel del prepucio, glándula mamaria, ingle e hisopados de canal auditivo externo,
obteniéndose 82 muestras en total, las cuales fueron transportadas al laboratorio de la
PUJ, en tubo estéril, a temperatura ambiente y en el menor tiempo posible. A cada
muestra se le realizó examen directo mediante la elaboración de un frotis en lámina
portaobjetos, se fijó y se coloreó con tinción de Gram para ser observado al microscopio e
identificar la presencia de estructuras levaduriformes o blastoconidias redondas u ovales
de tamaño variado, estructuras filamentosas y/o bacterias. Posterior a esto cada muestra
fue sembrada en Agar Dixon modificado (Extracto de malta 36 g, Peptona 6 g, Ox bile 20
g, Tween 40 10 ml, Glicerol 2 ml, Ácido oleico 2 ml, Agar 12 g, Agua desionizada 1.000
ml) (López ,2007) y se incubó a 320C durante 5 días.
De las siembras en las que se observaron colonias compatibles con Malassezia se hizo
un nuevo aislamiento para obtener colonias puras y de esta forma poder realizar la
descripción macroscópica del crecimiento observado, evaluando las diferentes
características de los aislamientos obtenidos como tamaño, textura, color; forma; así
como el borde y la superficie de la colonia. Para la descripción microscópica se utilizó el
microscopio Leica DM 100 LED versión 2.1.0 el cual permitió medir el ancho y el largo de
las células en micrómetros, por medio de un extendido teñido con coloración de Gram; por
último, para la identificación de especies del género Malassezia se realizaron las
siguientes pruebas bioquímicas:
- Catalasa: La prueba se realizó extendiendo una colonia sobre la superficie de una
lámina portaobjetos a la cual se adicionó una gota de peróxido de hidrógeno al 30%. La
prueba se consideró positiva, si se observaba formación de burbujas, y positivo débil, si
la formación de burbujas se observaba entre 5 y 10 segundos (Khosravi et al.2008)

12
- Ureasa: La prueba se realizó inoculando la levadura en la superficie del medio de cultivo
(agar úrea), posteriormente se incubó a 32oC, durante 7 días. La prueba se consideró
positiva si el medio viraba a color fucsia debido al indicador rojo de fenol (Pérez et al.
2002)
-β- glucosidasa: La prueba se realizó inoculando el microorganismo por punción en el
medio, se incubo a 32oC durante 7 días. Para esta prueba se consideró una reacción
positiva el viraje a color negro en el medio de cultivo. (Khosravi et al.2008)
-Asimilación del Cremophor-EL: La prueba se realizó a partir de una suspensión
concentrada del microorganismo, de la cual se adicionaron 400µl a 10 ml de agar
mycosel esta mezcla se depositó en una caja de Petri pequeña hasta que se solidificó,
posteriormente se realizó un pozo en el centro, donde se agregaron 50µl de Cremophor.
Para esta prueba se consideró una reacción positiva si luego de 7 días de incubación a
32°C se observaba alrededor del pozo un halo de asimilación (Kaneko, 2007).
-Asimilación del Tween: A partir de una suspensión del microorganismo en una
concentración de 0,5 en la escala de MacFarland, se adicionaron 2 ml a un tubo con 16ml
de agar sabouraud adicionado con 0,05% de cloranfenicol y 0.05% cycloheximida
(Mayzer, 1997; Khosravi et al. 2008). Esta preparación se depositó en una caja de Petri
grande hasta su solidificación. Posteriormente, se hicieron 4 pozos de (2mm) a los que se
adicionaron 7µl de cada Tween. La lectura se realizó evidenciando el crecimiento o
asimilación del microorganismo frente a cada Tween después de 7 días de incubación a
32°C (Kaneko, 2007; Hernández, 2005).
Pruebas complementarias
-Crecimiento en agar sabouraud: Se realizó siembra superficial de cada uno de los
aislamientos en tubo inclinado con agar sabouraud, se incubaron a 32°C por 7 días, para
observar si hubo crecimiento (Kaneko, 2007; Hernández, 2005)
-Crecimiento en agar Dixon a temperaturas de 37 y 40oC: Se sembró el
microorganismo en agar Dixon y se incubó a 37°C y a 40°C. Se observó si hubo
crecimiento del microorganismo después de 7 días (Guého et al. 1996; Kaneko, 2007).

13
-Producción de fosfolipasa en agar Sabouraud con yema de huevo: La prueba se
realizó en agar Sabouraud adicionado con 10% de yema de huevo, para esto se
realizaron cuatro inoculaciones del microorganismo en puntos equidistantes, se incubó a
32oC por 7 días, después de los cuales se hizo la medición de los halos y se calcularon
los índices Pz (Countinho, 2005)
- Características microscópicas: Se realizó la medición del largo y ancho de las células
en micrómetros en un extendido de los aislamientos obtenidos teñido con coloración de
Gram, para lo cual se utilizó el microscopio Leica DM 100 LED versión 2.1.0
4.3 Control de calidad y Mantenimiento de las cepas: Como control de calidad se
utilizaron las cepas de referencia M. furfur CBS 7019, M. pachydermatis CBS 1879, M.
sympodialis CBS 7222, M. slooffiae CBS 7956 pertenecientes a la colección de
microorganismos del Grupo de Micología y Fitopatología de la Universidad de Los Andes,
las cuales se conservaron en el medio Skim milk a -20°C, al igual que los aislamientos
obtenidos.
4.4 Análisis de la información: Los datos obtenidos fueron registrados utilizando tablas
de Excel y se determinó la prevalencia general de Malassezia en la población evaluada
utilizando la formula:
5. RESULTADOS Y DISCUSIÓN
De las 82 muestras procesadas, 12,2% (n=10) correspondieron a hisopados de prepucio,
14,6% (n=12) de glándula mamaria, 19,5% (n=16) de ingle y 53,6% (n=44) de canal
auditivo externo izquierdo y derecho.
5.1 Examen directo: Al examen directo con tinción de Gram se observaron levaduras de
formas y tamaños diferentes en el 17,07% (n=14) del total de las muestras de las cuales
el 8,53% (n=7) fueron identificadas como Malassezia, el 6,09%(n=5) como Cándidas y el
2,43%(n=2) como Cryptococcus spp, a las cuales se les realizó la descripción
macroscópica, microscópica y las pruebas bioquímicas para su identificación.
Adicionalmente, en el 40,24% (n=33) de las muestras se observaron en el directo
bacterias y en el 42,68% (n=35) se observaron hifas.
Numero de eventos P= Número de individuos totales
14
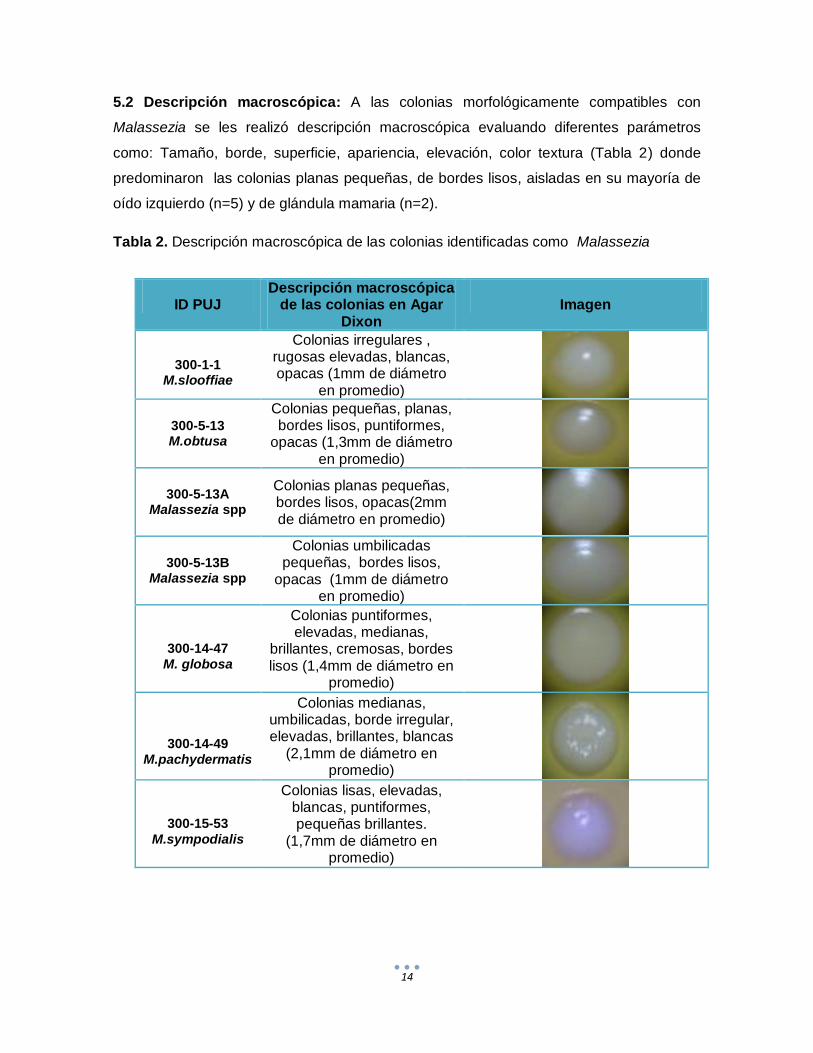
5.2 Descripción macroscópica: A las colonias morfológicamente compatibles con
Malassezia se les realizó descripción macroscópica evaluando diferentes parámetros
como: Tamaño, borde, superficie, apariencia, elevación, color textura (Tabla 2) donde
predominaron las colonias planas pequeñas, de bordes lisos, aisladas en su mayoría de
oído izquierdo (n=5) y de glándula mamaria (n=2).
Tabla 2. Descripción macroscópica de las colonias identificadas como Malassezia
ID PUJ
Descripción macroscópica de las colonias en Agar
Dixon
Imagen
300-1-1
M.slooffiae
Colonias irregulares , rugosas elevadas, blancas, opacas (1mm de diámetro
en promedio)
300-5-13 M.obtusa
Colonias pequeñas, planas, bordes lisos, puntiformes,
opacas (1,3mm de diámetro en promedio)
300-5-13A Malassezia spp
Colonias planas pequeñas, bordes lisos, opacas(2mm de diámetro en promedio)
300-5-13B Malassezia spp
Colonias umbilicadas pequeñas, bordes lisos,
opacas (1mm de diámetro en promedio)
300-14-47
M. globosa
Colonias puntiformes, elevadas, medianas,
brillantes, cremosas, bordes lisos (1,4mm de diámetro en
promedio)
300-14-49
M.pachydermatis
Colonias medianas, umbilicadas, borde irregular, elevadas, brillantes, blancas
(2,1mm de diámetro en promedio)
300-15-53
M.sympodialis
Colonias lisas, elevadas, blancas, puntiformes, pequeñas brillantes.
(1,7mm de diámetro en promedio)

15
Al comparar los resultados obtenidos en la descripción macroscópica de los aislamientos
identificados como Malassezia spp. con lo reportado por diferentes autores los cuales
mencionan unas características propias para cada especie de Malassezia, se encontró
que hubo coincidencia en cuanto a la descripción de las colonias y los parámetros
evaluados como: forma, bordes, elevación y textura, sin embargo se observaron
diferencias en cuanto al tamaño de las colonias, esto debido probablemente a
imprecisiones en la medición ocasionadas por la distribución de las colonias (Gueho et
al.1996; Sugita et al. 2003; Hirai et al. 2004; Hernández, 2005; Cabañes et al. 2007;
Arenas, 2008; Crespo et al. 2008).
6.3 Descripción microscópica: Las medidas microscópicas para los aislamientos
identificados como Malassezia (Figura 4) mostraron que M. pachydermatis presenta el
tamaño más grande 2,4 μm de ancho x 3,2 μm de largo, y M. obtusa el tamaño más
pequeño 1,5 μm de ancho x 2,2 μm de largo (Tabla 3).
Figura 4. Medidas microscopicas identificadas como Malassezia Tinción de Gram 100X
Al comparar estos resultados con los reportes de literatura se evidenció que los valores
obtenidos coinciden parcialmente con los rangos descritos para las diferentes especies
identificadas, las principales diferencias se observaron en el largo de las células, lo que
posiblemente se debe a imprecisiones en la medición ocasionadas por la distribución de
las mismas en el frotis; esto evidencia que este único parámetro no es confiable para
establecer una clasificación adecuada de estas levaduras (Guého et al, 1996; Sugita et
al. 2003; Hirai et al. 2004;Hernández , 2005; Cabañes et al, 2007; Arenas, 2008; Crespo
et al, 2008)

16
Tabla 3. Mediciones microscópicas de las especies identificadas como Malassezia a partir
de los aislamientos obtenidos
ID PUJ Promedio Especie
identificada
Promedio reportado en la literatura
Ancho μm Largo μm Ancho μm Largo μm
300-1-1 2,3 2 M.slooffiae 2,5 8
300-5-13 1,5 2,2 M.obtusa 1,5-2 4-6
300-5-13A 2 3,2 Malassezia spp 1,2 10
300-5-13B 2 3,01 Malassezia spp 1,2 10
300-14-47 2 2,7 M.globosa 2,5 8
300-14-49 2,4 3,2 M.pachydermatis 2-2,5 4-5
300-15-53 2,3 3,1 M.sympodialis 1,5-2,5 2.5-6
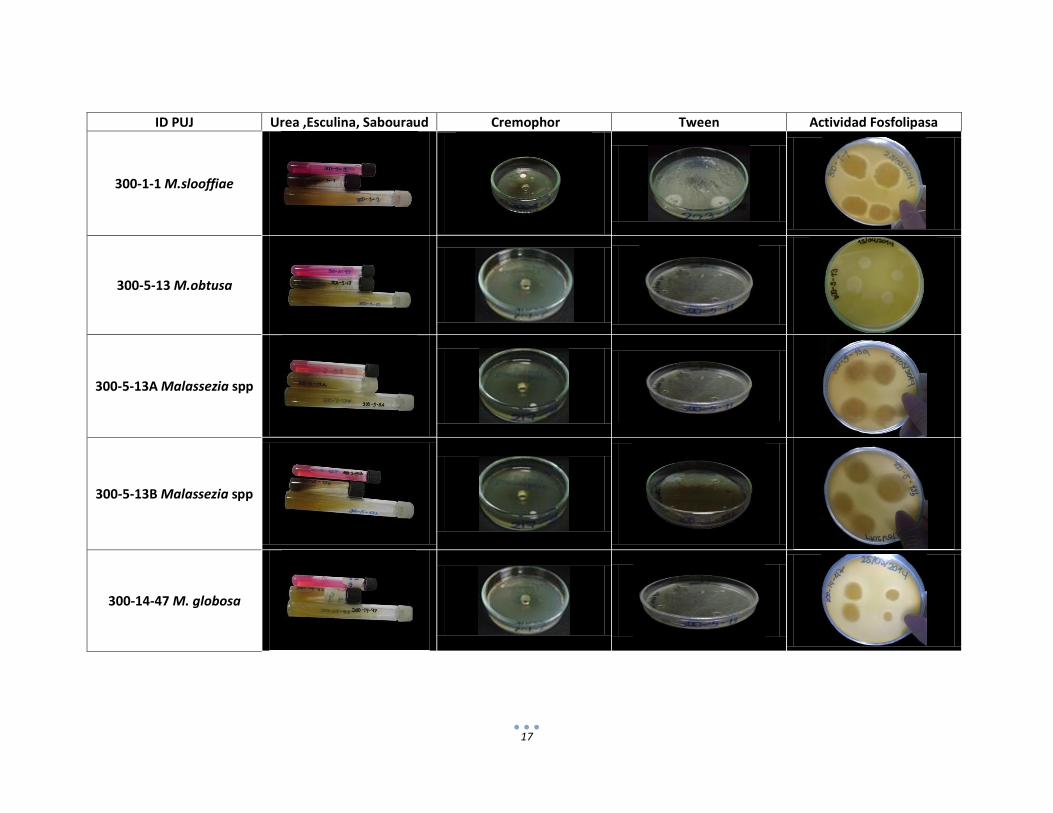
5.4 Identificación de especies del género Malassezia mediante pruebas bioquímicas
Por medio de las pruebas bioquímicas se realizó la identificación hasta especie evaluando
en cada una de éstas los diferentes requerimientos para su crecimiento y desarrollo. Las
principales características que permitieron la identificación de uno de los aislamientos
como M. pachydermatis incluyeron su capacidad para crecer a una temperatura de 32°C
en agar sabourad y la asimiliación de todos los Tweenes, al igual que su crecimiento en
Cremophor (Tabla 4). Aunque M. globosa y M. obtusa comparten ciertas características
como su incapacidad tanto para crecer a 40°C como para la asimilación de los Tweenes,
la prueba de β- glucosidasa permitió su diferenciación pues M. globosa no presenta la
enzima lo que la diferencia de M. obtusa que si la presenta.( Hernández, 2005; Giusiano,
2006) Por su parte, M. slooffiae solamente asimila los Tweenes 20, 40 y 60 y no presenta
la enzima β- glucosidasa, a diferencia de M. sympodialis que si presenta la enzima β-
glucosidasa y adicionalmente asimila los Tweenes 40, 60 y 80; estos hallazgos coinciden
con los diferentes reportes de literatura descritos para la identificación de las diferentes
especies de Malassezia de acuerdo con los resultados de las pruebas bioquímicas
reportadas por varios autores (Guého et al. 1996; Sugita et al. 2003; Harai et al. 2004;
Giusiano, 2006; Kaneko et al. 200; Salah et al. 2010).
17
ID PUJ Urea ,Esculina, Sabouraud Cremophor Tween Actividad Fosfolipasa
300-1-1 M.slooffiae
300-5-13 M.obtusa
300-5-13A Malassezia spp
300-5-13B Malassezia spp
300-14-47 M. globosa

18
ID PUJ Urea, Esculina ,Sabouraud Cremophor Tween Actividad Fosfolipasa
300-14-49 M.pachydermatis
300-15-53 M.sympodialis
Figura 5. Resultados pruebas Bioquímicas aislamientos identificados como Malassezia

19
Tabla 4. Identificación de los aislamientos obtenidos de acuerdo con las pruebas bioquímicas
ID PUJ Sitio Toma de la
muestra
Catalasa
Urea
β-
glucosidasa
Agar
Sabouraud
37ºC
Tween
Cremophor
Actividad
Fosfolipasa
Especie de Malassezia
40ºC 20 40 60 80
300-1-1 Canal auditivo
izquierdo
+++ +++ - - + +++ - + + - - + Pz=0,66
M. slooffiae
300-5-13 Canal auditivo
izquierdo
+++ +++ + - - + - - - - - - M.obtusa
300-5-13A Canal auditivo
izquierdo
+++ +++ - - + + - - - - - - Malassezia spp
300-5-13B Canal auditivo
izquierdo
+++ +++ + débil - + + + - - - - - Malassezia spp
300-14-47 Canal auditivo
izquierdo
+++ +++ - - - + - - - - - + Pz=0,40
M.globosa
300-14-49 Glándula mamaria
+++ ++ + +++ + + + + + + + + Pz=0,75
M. pachydermatis
300-15-53 Glándula mamaria
+++ +++ + - + + - + + - - - M. sympodialis

20
De acuerdo con los resultados obtenidos de las pruebas bioquímicas se pudo determinar
que de los aislamientos obtenidos el 2,43% (n=2) correspondió a Malassezia spp, las
cuales no pudieron identificarse hasta especie debido a que los perfiles bioquímicos no
coincidieron con lo reportado por literatura para las diferentes especies. Los demás
aislamientos si pudieron ser identificados hasta especie: M. pachydermatis, M. globosa,
M. obtusa, M slooffiae y M. sympodialis con un porcentaje del 1,22% (n=1) para cada una
de ellas. Estos datos concuerdan con reportes de literatura en los que se menciona que
estas especies son las más comúnmente aisladas en la piel de equinos sanos. (White,
2006; Crespo et al. 2002).
Las especies identificadas en el presente estudio fueron M. slooffiae, M. obtusa, M.
globosa, M. pachydermatis, M. sympodialis y Malassezia spp, aisladas de Oído Izquierdo
y Glándula mamaria de equinos sanos. De acuerdo con los resultados obtenidos en este
estudio y con lo reportado por White 2005 y Crespo et al. 2010 se aislaron en los equinos
las mismas especies exceptuando M. furfur y M. restricta que no fueron identificadas,
encontrando una concordancia en cuanto al sitio anatómico como lo es el canal auditivo
externo tanto para M. obtusa, como para M. slooffiae. Mientras que las especies M.
pachydermatis y M. sympodialis fueron aisladas de glándula mamaria, lo cual no coincide
con lo reportado por White, 2005 y White et al. 2006, quienes aislaron M. slooffiae y
M.equina en el primer estudio y M.equina en el segundo a partir de la glándula mamaria
de yeguas sanas.
En el presente estudio se evidenciò que 42,8% (n=3) de las especies presentó actividad
fosfolipasa positiva entre ellas: M.slooffiae con un Pz=0,66, M.globosa con un Pz=0,40 y
M.pachydermatis con un Pz=0,70, lo cual reveló que M. pachydermatis fue la única
especie que presentó actividad fosfolipasa intensa, mientras que un 57,1% (n=4) de las
especies no tuvieron actividad fosfolipasa. Estos resultados coinciden con lo reportado en
la literatura donde se evidencia que M. pachydermatis es una de las especies que
presenta alta actividad fosfolipasa tanto en animales sanos como en animales con lesión
(Cafarchia & Otranto, 2004; Pini, 2011; Ortiz, 2013), por otro lado M. sympodialis muestra
una buena actividad fosfolipasa cuando es aislado de piel sana lo cual también se pudo
corroborar a través de este estudio. M. obtusa M. slooffiae, M. globosa, M. restricta no
presentan actividad fosfolipasa en animales sanos según lo reportado por Pini, 2013, lo
cual difiere con los resultados obtenidos ya que para este estudio M. slooffiae y M.
globosa si presentaron actividad fosfolipasa positiva (rango Pz= <1.0 y ≥0,64)

21
La prevalencia general para la población evaluada fue de 18,2%, identificando las
especies M. slooffiae, M. obtusa, M.globosa, M. pachydermatis, M. sympodialis
coincidiendo con lo reportado por diferentes autores Crespo et al.2002; Giusiano,2006;
White,2006 quienes reportan la presencia de estas levaduras como las especies más
aisladas en la microbiota normal de la piel de equinos.
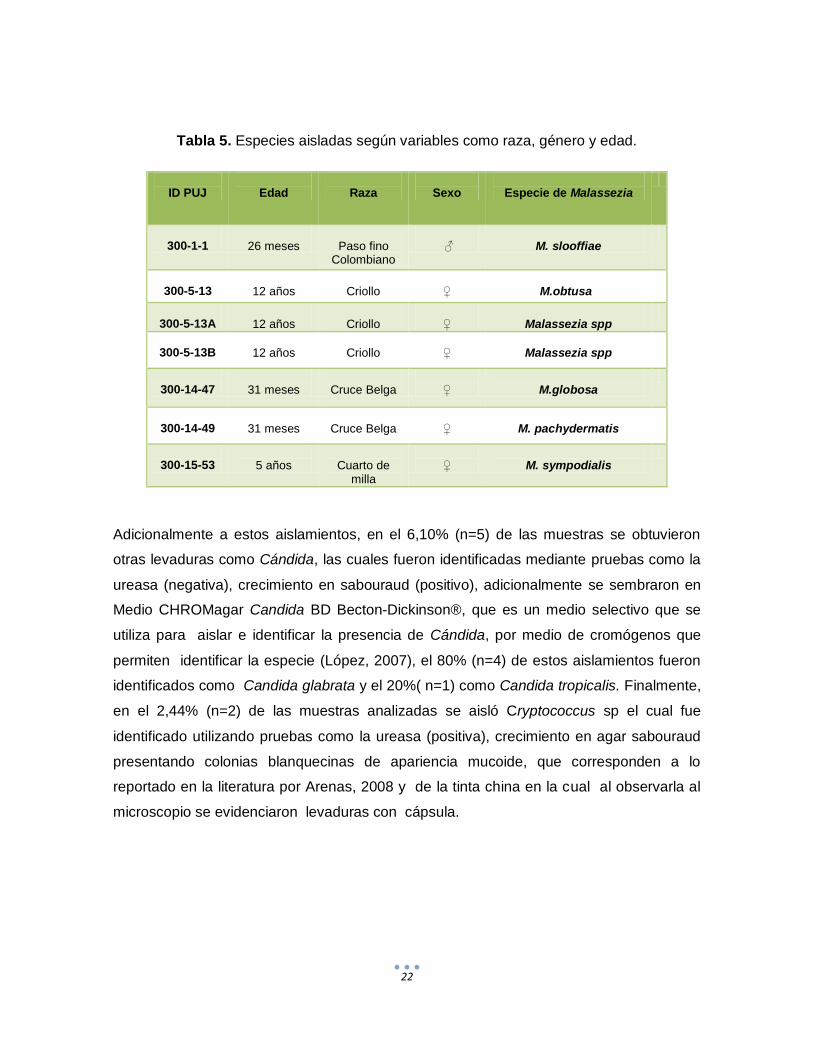
De acuerdo con lo observado en este estudio (Tabla 5) no fue posible establecer una
relación entre los aislamientos y las variables edad, sexo y raza debido al bajo número de
aislamientos obtenidos ya que el 71,42% (n=5) de ellos se obtuvieron de dos equinos 3 de
un equino de raza criolla y 2 de un animal de raza cruce belga, 1 aislamiento se obtuvo de
un equino de raza paso fino y 1 de un equino de raza cuarto de milla, lo que sesgaría el
análisis no solamente en cuanto a la variable raza sino también a las variables sexo y
edad.
Para la variable raza el 14,2% (n=1) de los animales correspondieron a Paso Fino
Colombiano, el 42,8% (n=3) fueron Criollos, el 28,5% (n=2) cruce Belga y el 14,2% (n=1)
Cuarto de Milla, en los animales evaluados en este estudio el mayor número de
aislamientos se hizo en un animal de la raza Criolla; sin embargo debido al escaso
número de aislamientos no es posible inferir que esta raza tenga una mayor presentación
de Malassezia como microbiota normal en la piel. En cuanto a la variable sexo en este
estudio se observó una mayor tendencia de presentación de Malassezia en las hembras
42,8% (n=3) y solo 1 macho (14.2%), sin embargo con estos resultados no es posible
inferir que en las hembras haya una mayor predisposición a la presentación de
Malassezia dados los escasos aislamientos obtenidos; para la variable edad estas
oscilaron entre 26 meses y 12 años, lo que indica una amplia variabilidad en las edades
de presentación, al igual que para las otras dos variables no es posible hacer inferencia
alguna dados los escasos aislamientos del microorganismo.
22
Tabla 5. Especies aisladas según variables como raza, género y edad.
ID PUJ
Edad
Raza
Sexo
Especie de Malassezia
300-1-1
26 meses
Paso fino
Colombiano
♂
M. slooffiae
300-5-13
12 años
Criollo
♀
M.obtusa
300-5-13A
12 años
Criollo
♀
Malassezia spp
300-5-13B
12 años
Criollo
♀
Malassezia spp
300-14-47
31 meses
Cruce Belga
♀
M.globosa
300-14-49
31 meses
Cruce Belga
♀
M. pachydermatis
300-15-53
5 años
Cuarto de
milla
♀
M. sympodialis
Adicionalmente a estos aislamientos, en el 6,10% (n=5) de las muestras se obtuvieron
otras levaduras como Cándida, las cuales fueron identificadas mediante pruebas como la
ureasa (negativa), crecimiento en sabouraud (positivo), adicionalmente se sembraron en
Medio CHROMagar Candida BD Becton-Dickinson®, que es un medio selectivo que se
utiliza para aislar e identificar la presencia de Cándida, por medio de cromógenos que
permiten identificar la especie (López, 2007), el 80% (n=4) de estos aislamientos fueron
identificados como Candida glabrata y el 20%( n=1) como Candida tropicalis. Finalmente,
en el 2,44% (n=2) de las muestras analizadas se aisló Cryptococcus sp el cual fue
identificado utilizando pruebas como la ureasa (positiva), crecimiento en agar sabouraud
presentando colonias blanquecinas de apariencia mucoide, que corresponden a lo
reportado en la literatura por Arenas, 2008 y de la tinta china en la cual al observarla al
microscopio se evidenciaron levaduras con cápsula.

23
6. CONCLUSIONES
La prevalencia de Malassezia para los animales evaluados en este estudio fue de
18,2%.
Se determinó la presencia de M. sympodialis, M. pachydermatis, M. slooffiae, M.
obtusa y M. globosa como microbiota normal en la piel de los equinos evaluados.
Los sitios anatómicos con mayor presencia de Malassezia en los animales
evaluados fueron glándula mamaria y canal auditivo.
Se evidenció una tendencia de mayor presencia de Malassezia en las hembras
No se estableció una relación entre la presencia de Malassezia spp y las
variables edad, sexo y raza.
7. RECOMENDACIONES
Para futuros estudios se recomienda realizar otras pruebas adicionales como
técnicas moleculares que permitan la identificación de los aislamientos que no
pudieron ser identificados hasta especie.
Por otro lado continuar con el estudio muestreando más equinos y en más zonas
de la piel como se ha reportado en la literatura como ano, dorso con el fin de
seguir caracterizando especies de Malassezia como microbiota normal en nuestro
país.

24
8. REFERENCIAS
Arenas R (2008) Micología médica ilustrada. Mc Graw Hill, México DF, México
Ashbee H. (2007) Update on the genus Malassezia. Medical Mycology 45:287-303
Boekhout T, Guého E, Mayser P, Velegraki A. (2010) Malassezia and the skin. Springer-Verlag, Heidelberg, Germany
Bonifaz A. (2012) Micología médica básica. Mc Graw Hill, México DF, México
Cabañes J, Theelen B, Castellá G, Boekhout T. (2007) Two new lipi-dependent Malassezia species from domestic animals. Vet Mycology 7:1064
Cabañes F, Vega S, Castellá G. (2011) Malassezia cuniculi sp. nov., a novel yeast species isolated from rabbit skin. Medical Mycology, 49:40–48
Cafarchia C & Otranto D. (2004) Association between phospholipase production by Malassezia pachydermatis and skin lesions. Journal of Clinical Microbiology 42(10): 4868–4869
Cafarchia C, Figueredo L, Otranto D. (2013) Fungal diseases of horses. Vet
Mycology 6078
Castellá G, Hernández J, Cabañes F. (2005) Genetic typing of Malassezia pachydermatis from different domestic animals. Vet Microbiology 108 : 291–296
Crespo M, Abarca M, Cabañes F. (2002) Occurrence of Malassezia spp. in horses and domestic ruminants. mycoses 45, 333–337
Crespo V, Gómez E, Crespo M. (2008) La pitiriasis versicolor y las levaduras del género Malassezia. Actas Dermosifiliográficas 99:764-771
Gaitanis G, Velegraki A, Frangoulis E, Mitroussia A, Tsigonia A. (2002) Identification of Malassezia species from patient skin scales by PCR-RFLP. Clinical Microbiology and Infection 8:162-173
Garau M, Palacio A, García J. (2005) Prevalence of Malassezia spp. in healthy
pigs. Mycoses, 48, 17–20
Giusiano G. (2006) Malassezia: Estado del conocimiento y perspectivas en su estudio. Revista Argentina de Microbiología 38:41-48
Guého E, Midgley G, Guillot J. (1996) The genus Malassezia with description of
four new species. Anton.Leeuw.Int J.G69:337-355
Hernández J. (2005) Caracterización molecular de especies del género Malassezia. Tesis de Doctorado. Facultad de Veterinaria, Universidad Autónoma
de Barcelona, España

25
Hirai A, Kano R, Makimura K, Duarte E, Hamdam J et al. (2004) Malassezia nana, a novel lipid-dependent yeast species isolated from animals. International Journal of Systematic and Evolutionary Microbiology 54:623-627
Kaneko T, Makimura k, Abe M, Shiota R, Nakamura Y, Kano R,Hasegawa A, Sugita T, Shibuya S, Watanabe S, Yamaguchi H, Abe S, Okamura N. (2007) Revised Culture-Based System for Identification of Malassezia Species. Journal of clinical microbiology, p. 3737–3742
Khosravi A, Eidi S, Ziglari T, Bayat M. (2008) Isolation and differentiation of Malassezia species isolated from healthy and affected small animals, ear and skin. World Journal of Zoology 3(2):77-80
López A, Sánchez A, Gil J. (2007) Fundamentos básicos para el diagnóstico micológico. Revista Iberoamericana de Micología. http://www.guia.reviberoammicol.com/ Obtenido 10 enero 2014
Mayser P, Haze P, Papavassilis C, Pickel M, Gruender K et al. (1997) Differentiation of Malassezia species: selectivity of Cremophor EL, castor oil and ricinoleic acid for M. furfur. British Journal of Dermatology 137:208-213
Nardoni S, Merildi V, Frangioni S, Ariti G, Verin R, Vannucci P, Mancianti F. (2010)
Isolation and characterization of Malassezia spp. in healty swine of different breeds. Veterinary microbiology. 141.,155-158
Nell A, James S, Bond C, Hunt B, Herrtage M. (2002) Identification and distribution of a novel Malassezia species yeast on normal equine skin. Vet. Rec.150:395–398.
Ortiz G, Carmen M, Alfonso J. Carrillo M, y Paya M. (2012) Producción de fosfolipasa y proteinasa en cepas de Malassezia pachydermatis aisladas de perros con otitis y sin otitis. Rev Iberoam Micol. 30(4):235–238
Paterson S. (2002) Identification of Malassezia from a horse’s skin case report. Equine vet. Educ. 14 (3) 121-125
Perez C, Goitia K, Mata S, Hartung C, Colella M , Reyes H, Hernandez C, Villaroel
M, Ontiveros J, Magaldy S, Suarez R. (2002) Use of the stuart urea broth for the urease test, like proof in the identification of yeast. Revista de la sociedad venezolana de microbiología. 22: 2
Pini G, Faggi E. (2011) Extracellular phospholipase activity of Malassezia strains isolated from individuals with and without dermatological disease. Rev Iberoam Micol 28(4):179-182
Price M, Wilkinson I, Gentry L. (1982) Plate method for detectionof phospholipase activity in Candida albicans. Sabouraudia 20: 7-14

26
Salah B I, Makni F, Cheikhrouhou F, Neji S, Sellami H, Ayadi A. (2010) Les levures du genere Malassezia : pathologie, milieux d’isolement et d’identification. Malassezia species: Pathology, isolation and identification media. Journal de mycologie Médicale. 20: 53-60
Sugita T, Takashima M, Kodama M, Tsuboi R, Nishikawa A. (2003) Description of a new yeast species Malassezia japonica and its detection in patients with atopic dermatitis and healthy subjects. Journal of Clinical Microbiology 41:4695-4699
White SD. (2005) Equine Bacterial and Fungal Diseases: A Diagnostic and Therapeutic Update. Clin Tech Equine Pract 4:302-310
White, S.D, Vandenabeele, S.I., Drazenovich, N.L, Foley, J.E. (2006) Malassezia species isolated from the intermammary and preputial fossa areas of horses. J. Vet. Intern. Med. 20, 395–3


















