Análisis histológico sobre el desarrollo y maduración del...
34
Análisis histológico sobre el desarrollo y maduración del ovario en Baeacris punctulatus (Thunberg) (Orthoptera: Acrididae: Melanoplinae) R E S U M E N — Baeacris punctulatus es una de las especie de Melanoplinae considerada de importancia económica para la Argentina. Los ovarios de insectos están sujetos a cam- bios en su anatomía, morfología y ultraestructura durante toda la vida del animal. Los estudios histológicos sobre desarrollo y maduración de las gónadas en Acrididae de Suda- mérica son escasos. El objetivo de este trabajo es proveer información sobre el proceso de morfogénesis del ovario y la ovogénesis durante el desarrollo embrionario, postembrio- nario y el imago. Las hembras de Baeacris punctulatus obtenidas en cautiverio se fijaron en Bouin y se conservaron en alcohol n-butílico. Los ejemplares completos se incluyeron en paraplast. Se realizaron cortes seriados de 6 μm de espesor. Los cortes se colorearon con Hematoxilina-Eosina, Tricrómico de Mallory y Hematoxilina de Verhoeff. Los resultados demostraron que en esta especie los ovarios se diferencian en el Estadio Embrionario X, identificándose en cada ovariola el germario con ovogonias en mitosis. La meiosis ovogonial se inicia antes de la eclosión de las ninfas. Los ovocitos primarios pasan por tres periodos de crecimiento: el primer periodo abarca desde el Estadio Embrionario XI al Ninfal II, se desarrolla en el germario e incluye desde preleptonema al paquinema. El segundo periodo abarca desde el Estadio Ninfal II a los 5 días de imago, se desarrolla en el vitelario e incluye el subestadio de postpaquinema y estadios de previtelogénesis del diplonema. El tercer periodo de crecimiento abarca desde los 5 a los 15 días de imago, incluye los estadios de vitelogénesis hasta la síntesis de cubiertas externas del ovocito, la envoltura vitelina y el corion. Entre los 15 y 25 días del imago se producen las primeras ovula- ciones y en este periodo la hembra alcanza la madurez sexual. Estos resultados aportan datos sobre la biología reproductiva de Baeacris punctulatus , los cuales pueden contribuir a un mejor manejo de esta especie. Palabras claves: Baeacris punctulatus, Orthoptera, Acrididae, Melanoplinae, Ovogénesis, Histología. A B S T R A C T — “Histological analysis of the development and maturation of ovaries in Baeacris punctulatus (Thunberg) (Orthoptera, Acrididae, Melanoplinae)”. Baeacris punctulatus is a Melanoplinae species of economical importance in Argentina. During the life of an in- sect, ovaries undergo changes characterized by morphological and phisiological cell alte- rations. Histological studies of the development and maturation of gonads in South American Acrididae are rare. The objective of this study is to provide information on the morphogenesis of the ovary and oogenesis during the embrionic, postembrionic and the imago stage of development. Captive females Baeacris punctulatus were obtained. They were fixed in Bouin and preserved in n-butilic alcohol. Specimens were placed in “paraplast” and 6 μm-thick segments were stained with Hematoxylin-Eosin, Trichrome of Mallory and Hematoxylin of Verhoeff. Results show that in this species, ovaries differentiate in the embryonic stage X, observing in the ovarioles the germarium with oogonia in mitosis. The oogonial meiosis begins before the hatching of the nymphs. The primary oocytes go through three stages of development. The first stage takes place between the embryonic stage XI and nymphal stage II. It develops in the germarium and it includes from pre- leptonema to paquinema. The second period goes from nymphal stage II to five days of the adult stage and it occurs in the vitelarium that includes the substage of postpaquinema and stages of previtellogenesis of the diplonema. The third period occurs between the fifth and fifteen days of the imago and it includes stages of vitellogenesis up to the synthesis Michel, Adriana A. Instituto de Morfología Animal, Fundación Miguel Lillo, Miguel Lillo 251, 4000 San Miguel de Tucumán, Argentina. e-mail: [email protected] Recibido: 20/10/08 – Aceptado: 28/05/09 30 Acta zoológica lilloana 52 (1–2): 30–63, 2008
Transcript of Análisis histológico sobre el desarrollo y maduración del...

A. A. Michel: Análisis histológico del ovario en Baeacris punctulatus30
Análisis histológico sobre el desarrollo y maduracióndel ovario en Baeacris punctulatus (Thunberg)(Orthoptera: Acrididae: Melanoplinae)
R E
S
U
M
E
N — Baeacris punctulatus es una de las especie de Melanoplinae considerada
de importancia económica para la Argentina. Los ovarios de insectos están sujetos a cam-bios en su anatomía, morfología y ultraestructura durante toda la vida del animal. Losestudios histológicos sobre desarrollo y maduración de las gónadas en Acrididae de Suda-mérica son escasos. El objetivo de este trabajo es proveer información sobre el procesode morfogénesis del ovario y la ovogénesis durante el desarrollo embrionario, postembrio-nario y el imago. Las hembras de Baeacris punctulatus obtenidas en cautiverio se fijaronen Bouin y se conservaron en alcohol n-butílico. Los ejemplares completos se incluyeronen paraplast. Se realizaron cortes seriados de 6 µm de espesor. Los cortes se colorearoncon Hematoxilina-Eosina, Tricrómico de Mallory y Hematoxilina de Verhoeff. Los resultadosdemostraron que en esta especie los ovarios se diferencian en el Estadio Embrionario X,identificándose en cada ovariola el germario con ovogonias en mitosis. La meiosis ovogonialse inicia antes de la eclosión de las ninfas. Los ovocitos primarios pasan por tres periodosde crecimiento: el primer periodo abarca desde el Estadio Embrionario XI al Ninfal II, sedesarrolla en el germario e incluye desde preleptonema al paquinema. El segundo periodoabarca desde el Estadio Ninfal II a los 5 días de imago, se desarrolla en el vitelario eincluye el subestadio de postpaquinema y estadios de previtelogénesis del diplonema. Eltercer periodo de crecimiento abarca desde los 5 a los 15 días de imago, incluye losestadios de vitelogénesis hasta la síntesis de cubiertas externas del ovocito, la envolturavitelina y el corion. Entre los 15 y 25 días del imago se producen las primeras ovula-ciones y en este periodo la hembra alcanza la madurez sexual. Estos resultados aportandatos sobre la biología reproductiva de Baeacris punctulatus, los cuales pueden contribuira un mejor manejo de esta especie.Palabras claves: Baeacris punctulatus, Orthoptera, Acrididae, Melanoplinae, Ovogénesis,Histología.
A B
S
T
R
A
C
T — “Histological analysis of the development and maturation of ovaries in
Baeacris punctulatus (Thunberg) (Orthoptera, Acrididae, Melanoplinae)”. Baeacris punctulatusis a Melanoplinae species of economical importance in Argentina. During the life of an in-sect, ovaries undergo changes characterized by morphological and phisiological cell alte-rat ions. Histological studies of the development and maturat ion of gonads in SouthAmerican Acrididae are rare. The objective of this study is to provide information on themorphogenesis of the ovary and oogenesis during the embrionic, postembrionic and theimago stage of development. Captive females Baeacris punctulatus were obtained. Theywere fixed in Bouin and preserved in n-butilic alcohol. Specimens were placed in “paraplast”and 6 µm-thick segments were stained with Hematoxylin-Eosin, Trichrome of Mallory andHematoxylin of Verhoeff. Results show that in this species, ovaries differentiate in theembryonic stage X, observing in the ovarioles the germarium with oogonia in mitosis. Theoogonial meiosis begins before the hatching of the nymphs. The primary oocytes gothrough three stages of development. The first stage takes place between the embryonicstage XI and nymphal stage II. It develops in the germarium and it includes from pre-leptonema to paquinema. The second period goes from nymphal stage II to five days ofthe adult stage and it occurs in the vitelarium that includes the substage of postpaquinemaand stages of previtellogenesis of the diplonema. The third period occurs between the fifthand fifteen days of the imago and it includes stages of vitellogenesis up to the synthesis
Michel, Adriana A.Instituto de Morfología Animal, Fundación Miguel Lillo, Miguel Lillo 251, 4000 San Miguelde Tucumán, Argentina. e-mail: [email protected]
Recibido: 20/10/08 – Aceptado: 28/05/09
30Acta zoológica li l loana 52 (1–2): 30–63, 2008

Acta zoológica li l loana 52 (1–2): 30–63, 2008 31
of external covers, vitelline cover and corium. The first ovulations occur between the fif-teenth and twenty-fifth day. It is during this period that the female reaches sexual maturity.These results provide information on the reproductive biology of B. Punctulatus, which maycontribute to better species management.Key Words: Baeacris punctulatus, Orthoptera, Acrididae, Melanoplinae, Oogenesis, His-tology.
INTRODUCCIÓN
La especie Baeacris punctulatus (Thun-berg) forma parte del grupo “punctulatus” delgénero Baeacris. En este grupo se incluyen unconjunto de especies crípticas con una ampliadistribución en Sudamérica, de aspecto exte-rior uniforme pero fácilmente separables porsu distribución geográfica y los caracteresderivados de la genitalia del macho. Estasespecies son en general de tamaño pequeño,de forma frágil y elegante sobre todo el ma-cho, ya que las hembras son más robustas ygrandes. En su coloración predominan loscolores castaño y pardo, siendo las tibias pos-teriores de color rojo vivo (Carbonell y Ron-deros, 1973). Dentro del grupo “punctulatus”y aún del género, Baeacris punctulatus es laespecie de mayor distribución geográfica, yaque ocupa todo el territorio de América delSur al este de los Andes, desde Colombia hastaArgentina (Bentos Pereira, 1989; Ronderos yCigliano, 1991).
Baeacris punctulatus es una de las espe-cies de Melanoplinae considerada de impor-tancia económica para Argentina (Lieber-mann y Schiuma, 1946). Así mismo, se hacomprobado que en esta especie de desarro-llo continuo, con diapausa embriónica facul-tativa (Michel y Terán, 2006), la hembrapresenta una gran susceptibilidad a agentesde control biológico, siendo ésta sustratoadecuado para la esporulación de microspo-ridios en el cuerpo graso. En insectos, el vo-lumen del cuerpo graso en la hembra adultaestá directamente relacionado con el desa-rrollo y maduración del ovario (Villeco,1998; Lange, 2002, 2003; Albert et al.,2004).
En condiciones de cría óptimas para sudesarrollo, en B. puntulatus el desarrolloembrionario tiene una duración media de 20días, que incluyen 11 estadios determinados
por cambios morfogenéticos producidos des-de la puesta hasta la eclosión de las ninfas.La duración media del desarrollo postem-brionario en el macho es de 28 días, mien-tras que para la hembra la duración mediaes de 30 días. En ambos sexos el desarrollopostembrionario incluye 5 estadios ninfalesdesde la eclosión de las ninfas hasta lamuda imaginal, determinados por el análisisdel desarrollo de pterotecas y genitalias ex-ternas. Los adultos matenidos en cámara decría tuvieron una longevidad máxima de 70días. La primera cópula se produce entre los2 y 12 días después de la muda imaginal yla primera puesta 10 días después de la có-pula (Michel y Terán, 2006).
Aunque se realizaron estudios histológi-cos de gónadas en especies sudamericanasde Acridoidea, como los de Cruz-Landim(1994) en Conocephalus saltador (Saussure)(Orthoptera, Conocephalidae), Ferreira etal. (2006) en Cromacris speciosa (Thunberg)(Orthoptera, Romaleidae) y Santos et al.(2006) en Tropidacris collares (Stoll) (Or-thoptera, Romaleidae), los estudios histoló-gicos sobre desarrollo y maduración de lasgónadas en especies de Acrididae de Suda-mérica son escasos. Entre los trabajos querealizan este tipo de estudios, se puede men-cionar el realizado por Michel (2007) sobredesarrollo del testículo y espermatogénesisen Baeacris punctulatus.
En Baeacris punctulatus, la gónada de lahembra está constituida por un par de ova-rios ubicados por encima del tubo digestivo.Cada ovario esta constituido por 10 ovario-las panoísticas, es decir carecen de célulasnutricias especiales (trofocitos) y el epiteliofolicular es el único tejido trófico presente.Cada ovariola, limitada externamente poruna capa de células epiteliales planas, estáformada por tres regiones: el filamento ter-minal que se une a la pared del cuerpo a tra-

A. A. Michel: Análisis histológico del ovario en Baeacris punctulatus32
vés del ligamento suspensorio, el tubo dehuevos en el cual se desarrolla la línea ger-minal y un corto pedicelo que desemboca enel oviducto lateral. El tubo de huevo está for-mado por el germario en el extremo distal,con ovogonias y ovocitos en las primeras eta-pas de la profase I de la meiosis, dispuestosen forma irregular y rodeados por tejido pre-folicular y el vitelario en la región media yproximal, con ovocitos en las diferentes eta-pas de previtelogénesis y vitelogénesis, dis-puestos linealmente y rodeados por una capacontinua de células foliculares (Michel y Te-rán, 2005).
Los ovarios de insectos están sujetos acambios en su anatomía, morfología y ul-traestructura durante toda la vida del animal(Büning, 1994). Considerando que Baeacrispunctulatus es una de las especies de Mela-noplinae de importancia económica paraArgentina y que los estudios sobre morfogé-nesis de la gónada y gametogénesis en Acri-didae de Sudamérica son escasos, el objetivode este trabajo es proveer información sobreel proceso de morfogénesis del ovario y laovogénesis durante el desarrollo embriona-rio, postembrionario y en el imago.
Para cumplir con los objetivos planteadosse realizó un estudio exhaustivo en cortes se-riados de ejemplares fijados enteros, en dife-rentes tiempos desde el momento de la puestahasta los 30 días de imago, lo que permitióobtener una imagen tridimensional de loscambios morfológicos producidos en los ova-rios durante su desarrollo. Por un lógico pro-blema de espacio, solamente se han incluídolas figuras más representativas de los procesodescripto. Los aspectos cualitativos que éstasrepresentan constituyen una síntesis de lagran cantidad de información obtenida.
MATERIAL Y MÉTODOS
La cría de la especie se llevó a cabo apartir de desoves obtenidos de adultos silves-tres capturados con redes entomológicas, enáreas destinadas al pastoreo y banquinas delKm. 28 y 29 de Ruta provincial N° 340 (Tu-cumán) y mantenidos en el laboratorio atemperatura ambiente (20 ± 2° C) en jaulas
provistas de desovadores. La incubación delos desoves y el mantenimiento de las crías serealizó en una cámara con un ambiente re-gulado a 30 ± 1° C, entre 50 y 70% de hu-medad relativa y un fotoperíodo de 14 horasluz. La alimentación de los adultos silvestresy de ninfas y adultos obtenidos en cautiveriose realizó con hojas frescas de Taraxacumofficinale. Los ejemplares de referencia seencuentran depositados en la Colección Ento-mológica del Instituto-Fundación Miguel Li-llo (IFML), Tucumán, Argentina.
Para el estudio histológico, durante eldesarrollo embrionario se tomaron muestrasde 3 huevos cada una, cada 3 horas duranteel primer día y luego cada 24 horas, extra-yéndose hasta 3 muestras por vez. Mientrasque para el estudio durante el desarrollopostembrionario, se tomaron muestras de 2ejemplares hembras cada 48 horas desde elmomento de la eclosión de las ninfas hastala muda imaginal. Para el estudio en elimago, se tomaron muestras de 2 hembrascada una a las 24, 48, 72 y 120 horas de lamuda imaginal y luego cada 5 días hasta los30 días del imago. Además se tomaronmuestras de hembras vírgenes, durante lacópula, y a las 12, 24 y 48 horas después dela primera cópula y en el momento de lapuesta. La obtención de ejemplares de Baea-cris punctulatus en las diferentes etapas deldesarrollo se realizó considerando los esta-dios de desarrollo establecidos por Michel yTeisaire (1996) y Michel y Terán (2006).
Los ejemplares completos se fijaron enBouin (solución acuosa saturada de ácidopícrico, formol puro y ácido acético 70: 25:5) durante 24 horas. Se deshidrataron enuna batería ascendente de etanol y se con-servaron en alcohol n-butílico. Las hembrasse incluyeron en paraplast y se realizaroncortes seriados de 6 µm de espesor con orien-tación sagital y frontal. Se coloreó con he-matoxilina de Erlich-eosina (H-E) (Geurr,1956 citado por Humason, 1979), hematoxi-lina de Verhoeff (H.V.) (según Mallory, 1944citado por Humason, 1979) y tricrómico deMallory (Azan) Heidenhain (M.A.) (Modifi-cado por Konef, 1938, citado por Humason,1979). Las observaciones se realizaron con

Acta zoológica li l loana 52 (1–2): 30–63, 2008 33
un microscopio Leitz-Dialux 20 2B y las mi-crofotografías se tomaron con una cámaraMiranda acoplada con adaptador al tubomonocular de fotografías.
RESULTADOS
1. MORFOGÉNESIS DEL OVARIO Y OVOGÉNESIS
EN EL DESARROLLO EMBRIONARIO
Estadio Embrionario X (17 a 18 días despuésde la puesta).— En Baeacris punctulatus, losovarios se identifican por su organizaciónestructural recién a partir del Estadio X deldesarrollo embrionario, en posición dorsalal tubo digestivo, desde la mitad del 2° seg-mento abdominal al 4° inclusive, siendo ro-deados por abundante tejido graso. En cadaovario, se pueden distinguir una región dor-sal y anterior en la cual se observan 10 gru-pos celulares dispuestos linealmente en sen-tido longitudinal. En corte transversal, cadauno de estos grupos está formado por 6 ovo-gonias dispuestas en forma irregular y ro-deadas por 2 o 3 células mesodérmicas alar-gadas que forman una capa continua alrede-dor de las mismas. Las ovogonias son célu-las pequeñas con un diámetro de 7,5 µm,con grandes núcleos de forma esférica uovoidea y escaso citoplasma levemente ba-sófilo. La mayor parte de las ovogonias seencuentran en interfase, ya que en sus nú-cleos la heterocromatina está condensada engránulos gruesos y separados y la eucromati-na se presenta en forma de finas hebras quecubren completamente el nucleoplasma;siendo escaso el número de ovogonias quehan comenzado a dividirse mitóticamente(Fig. 1). Cada uno de estos grupos celularesconstituye el rudimento del germario de lasovariolas en desarrollo.
En la región ventral y hacia la línea me-dia del cuerpo, en cada ovario se observa uncordón continuo de células grandes, redon-deadas, levemente basófilas, dispuestas enforma lineal y rodeadas por numerosas célu-las mesodérmicas. Estas células tienen undiámetro que oscila entre 10,5 y 12 µm, pre-sentan un núcleo grande y esférico, rodeadopor escaso citoplasma granular levementebasófilo y la cromatina condensada en grá-
nulos gruesos y separados. Por el tamaño ylas características del núcleo, estas célulascorresponden a preovocitos, ya que se en-cuentran en una etapa previa a la iniciaciónde la meiosis, es decir, en un período de re-plicación del DNA o fase S de la interfasepremeiótica. Este cordón de células ventra-les de cada ovario constituye el rudimentodel vitelario, a partir del cual se formará elvitelario al completarse el desarrollo de laovariola (Tabla 1, al final del trabajo). Lascélulas mesodérmicas que rodean a la re-gión ventral de cada ovario se prolongan endirección cefálica en un delgado cordón queconstituye el esbozo de la glándula accesoriade la hembra y en dirección caudal en uncordón que forma el esbozo de la porcióngonádica del oviducto lateral de cada ladodel cuerpo.
Estadio Embrionario XI (20 a 21 días des-pués de la puesta).— En este estadio, cadaovario está constituido por 10 ovariolascompletamente diferenciadas y rodeadas poruna delgada capa de células mesodérmicasaplanadas que constituyen la membrana li-mitante externa, la cual se prolonga distal-mente en un delgado y corto cordón celularque corresponde al filamento terminal de lamismas. Las ovariolas son ligeramente alar-gadas, con un diámetro menor en el extremodistal de las mismas, ubicado en la regióndorsal de cada ovario y rodeado por abun-dante tejido graso, mientras que el extremoproximal de la ovariola de mayor diámetrose ubica en la región ventral del ovario yestá limitado solamente por una delgadacapa de células mesodérmicas.
En estas hembras, las ovogonias ubicadasen el extremo distal de cada ovariola se en-cuentran en activo proceso de división celu-lar, observándose numerosos núcleos en pro-fase y metafase mitótica. Al mismo tiempoen la región ventral de la ovariola se ha re-ducido el número de preovocitos y en cam-bio se observan numerosas células esféricasligeramente mayores, con grandes núcleosrodeados por escaso citoplasma levementebasófilo y cromosomas en diferentes gradosde condensación en el interior de los mis-

A. A. Michel: Análisis histológico del ovario en Baeacris punctulatus34
mos, lo que indicaría el inicio del estadio deprofase I de la meiosis (Fig. 2). Dichas célu-las corresponden a ovocitos primarios en lossiguientes subestadio: a) en preleptonemacon un diámetro aproximado de 16,5 µm, enla etapa inicial de la condensación cromosó-mica con cromosomas largos y delgados queforman un ovillo compacto intensamentebasófilo en el interior del nucleoplasma, b)células en leptonema con un diámetro deaproximadamente 18 µm, con el núcleo me-nos basófilo y cromosomas más condensadosque se presentan como finas hebras con pe-
queños engrosamientos (cromómeros) dis-puestos en forma de cuentas de collar, y c)en menor número se observan ovocitos pri-marios en cigonema, de un tamaño ligera-mente mayor aproximadamente 19,5 µm yen sus núcleos se visualizan cromosomascomo cintas largas y delgadas levementebasófilos, indicando el apareamiento progre-sivo de cromosomas homólogos parcialmen-te condensados. La ligera variación en tama-ños indicaría que estos ovocitos se encuen-tran en el “primer periodo de crecimiento”(Tabla 1).
Figura 1: Corte sagital de la hembra de Baeacris punctulatus en el Estadio Embrionario X,se observa la región dorsal del ovario (O) con esbozos de ovariolas (OV) separados porabundante tejido graso (TC). Cada ovariola está constituida por grupos de ovogonias (o) dis-puestas en forma irregular, rodeados por una delgada capa de células mesodérmicas. H-E.Barra = 18,2 µm.Figura 2: Corte sagital de la región ventral del ovario (O) en el Estadio Embrionario XI, encontinuidad con el esbozo de la porción gonádica del oviducto lateral (PGL). Se observa elextremo proximal de las ovariolas separados por una delgada capa de células mesodérmicas(cm) y en su interior ovocitos primarios (op) en preleptonema (pl) y leptonema (l). H-E.Barra = 1,2 µm.Figura 3: Corte sagital del ovario en el Estadio Ninfal I, se observa la región proximal de lasovariolas (OV) rodeada por células mesodérmicas y cuerpo graso (CGR). En el interior de laovariola se identifican grupos de ovocitos primarios (op) en paquinema (pa) rodeados porcélulas prefoliculares (cp). H-E. Barra = 1,2 µm.Figura 4: Región distal de las ovariolas (OV) de un ejemplar en el Estadio Ninfal II, limitadapor la vaina externa (VE) de células mesodérmicas y tejido graso (TC). En el interior de laovariola, se observan ovogonias (o) en interfase (i) y metafase mitótica (m). H-E. Barra =1,2 µm.

Acta zoológica li l loana 52 (1–2): 30–63, 2008 35
2. MORFOGÉNESIS DEL OVARIO Y OVOGÉNESIS
EN EL DESARROLLO POSTEMBRIONARIO
Estadio Ninfa I (1 a 6 días después de laeclosión de las ninfas).— Al comienzo deldesarrollo postembrionario, los ovarios sonligeramente alargados, se ubican en posi-ción dorsal al tubo digestivo, y abarcan des-de el 2° al 4° segmento abdominal inclusive.En cada ovario se distinguen claramente 10ovariolas completamente diferenciadas, alar-gadas y en forma de huso, dispuestas en for-ma paralela y separadas por una delgadacapa de tejido graso. En estas hembras, cadaovariola está formada por un delgado fila-mento terminal, que se prolonga hasta elextremo posterior y dorsal del primer seg-mento abdominal; y el germario está limita-do externamente por una delgada vaina ex-terna de aproximadamente 8 células epitelia-les en corte transversal.
En la región distal del germario, se ob-servan por corte transversal aproximadamen-te 14 ovogonias dispuestas en forma irregu-lar, la mayoría de las cuales se encuentranen interfase mitótica y un número reducidode ellas, en las restantes etapas de la mito-sis. En la región media y proximal se obser-van 2 preovocitos y 6 u 8 ovocitos primariosen preleptonema, leptonema y cigonema dis-puestos en forma irregular. A continuaciónde estas células, en la región basal del ger-mario, se observan 2 ovocitos primarios enpaquinema inicial de aproximadamente 21µm de diámetro. Estas células son esféricascon un núcleo grande claro y escaso citoplas-ma levemente basófilo alrededor del mismo.En el interior del núcleo se observan cromo-somas largos y anchos como cintas, leve-mente basófilos y con escasos centrómerosvisibles entre ellos (Fig. 3).
Al finalizar el primer estadio ninfal, seobserva un ligero aumento de longitud delas ovariolas, aumentando el número de ovo-gonias a 20 por corte transversal en el extre-mo distal de las mismas, mientras que en elextremo proximal, se observan 2 ovocitosprimarios en paquinema avanzado. En elnúcleo de estos ovocitos se observan cromo-somas más cortos y de una basofilia más in-tensa que en la etapa inicial de este subesta-
dio. En el extremo de cada cromosoma seobservan gránulos fuertemente basófilos quecorresponden a los centrómeros de cromáti-das hermanas. En estas hembras, el diámetrode los ovocitos primarios en los diferentessubestadios de la profase I de la meiosis va-ría entre 14 y 21 µm, desde el extremo distalal proximal del germario, y forman nidos de2 a 4 células rodeados por 3 a 4 células pre-foliculares de forma alargada con grandesnúcleos ovoideos fuertemente basófilos (Ta-bla 1, Fig. 3).
Estadio Ninfal II (7 a 12 días después de laeclosión de las ninfas).— Al comienzo delestadio Ninfal II, en el germario de cada ova-riola se observan las mismas cantidades ytipos celulares del estadio anterior, pero enestos ejemplares las ovogonias próximas alfilamento terminal se disponen en formairregular, mientras que las de la región basalde la ovariola forman grupos de 2 a 6 célu-las. Cada grupo está limitado externamentepor células prefoliculares dispuestas en for-ma desordenada. En ambas zonas, la mayo-ría de las ovogonias se encuentra en interfa-se, profase o prometafase mitótica y soloalgunas en metafase (Fig. 4). Entre los ovo-citos primarios que ocupan el resto del ger-mario, se observan “núcleos picnóticos”, queprobablemente correspondan a restos de ovo-gonias en degeneración que no se diferen-ciaron en ovocitos (Fig. 5).
Al finalizar este estadio, las ovariolas pre-sentan una mayor longitud, observándose enel extremo distal del germario aproximada-mente 30 ovogonias por corte transversal, delas cuales las que se disponen en forma irregu-lar están en interfase, mientras que las restan-tes que forman nidos se encuentran principal-mente en metafase, anafase y telofase mitóti-ca. En el extremo proximal de cada ovariola ya continuación del germario, comienza a for-marse el vitelario, ya que por primera vez seobservan ovocitos primarios en el subestadiode postpaquinema dispuestos en forma linealy rodeados cada uno de ellos por 5 o 6 célulasprefoliculares (Tabla 1). Estos ovocitos (5 o 6por cada ovariola) son células ligeramentemás grandes que las del subestadio anterior,

A. A. Michel: Análisis histológico del ovario en Baeacris punctulatus36
ya que presentan una variación del diámetrode 21 a 24 µm que se manifiesta tanto en lacantidad de citoplasma basófilo como en eltamaño de la vesícula germinal. En estos ovo-citos, los cromosomas no son visibles y la cro-matina es difusa y forma grandes acúmulosen el nucleoplasma. Estas células, por su ima-gen nuclear, se encuentran en una etapa inter-media entre el subestadio de paquinema y elde diplonema, y correspondería al inicio del“segundo período de crecimiento” del ovocito.
Estadio Ninfal III (13 a 16 días después dela eclosión de las ninfas).— En este estadioen cada ovario, se observa un aumento delongitud de las ovariolas, las cuales estánseparadas entre sí por una delgada vainaexterna y todas ellas rodeadas por abundan-te tejido graso. En cada ovariola se distingueen la región distal, el filamento terminal yel germario, y en la región media y proxi-mal, el vitelario. En el germario se observanovogonias con escasa actividad mitótica, yaque la mayoría se encuentra en el periodo deinterfase, y ovocitos primarios en el “primerperiodo de crecimiento” (preleptonema, lep-tonema, cigonema, paquinema inicial yavanzado), los cuales al igual que en el es-tadio ninfal II, forman grupos de 2 a 4 célu-las rodeados por 3 a 4 células prefolicularescada uno. En el vitelario se observan ovoci-tos primarios en el “segundo periodo de cre-cimiento”, los cuales se disponen en formalineal y aumentan progresivamente de tama-ño hacia el extremo proximal de la ovariola,observándose primero 2 o 3 ovocitos esféri-cos en postpaquinema y rodeados cada unoo de a pares por 2 o 3 células prefolicularesy luego 4 a 8 ovocitos primarios al comien-zo del subestadio de diplonema de la meio-sis, los cuales serán considerados en este tra-bajo como ovocitos en el estadio previtelogé-nico I (Tabla 1, Fig. 6). En esta etapa de laovogénesis (diplonema), se produce la pri-mera detención de la meiosis y los ovocitosprimarios pasarán por diferentes estadios deprevitelogénesis y vitelogénesis en los cualesse realizará la incorporación de diferentesmoléculas y nutrientes que le permitirán al-canzar su tamaño definitivo.
Cada uno de los ovocitos en el estadioprevitelogénico I está rodeado por una capacontinua de 4 a 8 células foliculares aplana-das con grandes núcleos ovoideos fuertemen-te basófilos y escaso citoplasma alrededorde los mismos, que constituyen el esbozo deun folículo ovárico. Estos ovocitos previtelo-génicos son de forma cúbica a esférica y pre-sentan un aumento progresivo de tamañoque va desde 30 a 72 µm, lo que se mani-fiesta tanto en la cantidad de citoplasmacomo en el diámetro de la vesícula germi-nal. Dicha vesícula es grande (21 a 36 µm) yse ubica en el centro del ovocito, con unaenvoltura nuclear claramente visible. La cro-matina es difusa y difícil de observar al mi-croscopio óptico, ya que se presenta en for-ma de finas hebras y pequeñas condensacio-nes basófilas (cromómeros) que se distribu-yen uniformemente en el jugo nuclear y au-mentan en número a medida que aumenta eltamaño del ovocito. En la región central ypróxima a la envoltura nuclear se observande 4 a 8 “cuerpos proteinosos” que se identi-fican como acúmulos de diferentes tamañosconstituidos por fina granulación basófila.Entre estos acúmulos se observan 2 o 3 ma-cronucleolos formados por granulacionesfuertemente acidófilas y numerosos micronu-cleolos fuertemente acidófilos alrededor deestos últimos y próximos a la envoltura nu-clear. El citoplasma de los ovocitos en el es-tadio previtelogénico I es basófilo finamentegranular, observándose una disminución pro-gresiva de la basofilia a medida que aumen-ta el tamaño de los mismos. En algunos ca-sos, se observan pequeños gránulos fuerte-mente acidófilos próximos a la envolturanuclear, que probablemente estén relaciona-dos con la presencia de los cuerpos proteino-sos ubicados dentro de la vesícula germinal.Tanto el aspecto de los cromosomas, como lapresencia de los cuerpos proteinosos y el cre-cimiento de la masa celular, indican que es-tos ovocitos se encuentran en una etapa ini-cial del subestadio de diplonema de la meio-sis (Tabla 2, al final del trabajo, y Fig. 6).
Estadio Ninfal IV (17 a 22 días después dela eclosión de las ninfas).— En este estadio,

Acta zoológica li l loana 52 (1–2): 30–63, 2008 37
los ovarios están rodeados por abundantecuerpo graso y han aumentado su volumenabarcando desde el 1° al 4° segmento abdo-minal. Cada ovariola está rodeada por unadelgada capa de células epiteliales planas,con núcleos voluminosos que sobresalen delescaso citoplasma levemente acidófilo, queconstituyen la vaina externa que separa tan-to al germario como al vitelario del tejidograso circundante. La vaina externa se pro-longa en el extremo distal de cada ovariolaen un delgado filamento terminal, y todoslos filamentos de un mismo ovario se unen
distal y dorsalmente en la cavidad abdomi-nal para formar el ligamento suspensorio quese une al diafragma dorsal. El ligamentosuspensorio está formado por células alarga-das cuyos núcleos ovoideos con la cromatinacondensada en gránulos gruesos y separa-dos, están rodeados por abundante citoplas-ma levemente acidófilo.
En el extremo distal del germario, a conti-nuación del filamento terminal se observanaproximadamente 35 ovogonias, la mayoríade las cuales forman grupos de 4 o 5 célulascada uno y se encuentran en interfase mitóti-
Figura 5: Región proximal de las ovariolas (OV) de un ejemplar en el Estadio Ninfal II. Entrelos ovocitos primarios (op) en diferentes estadios de la primera división meiótica, se observannúcleos picnóticos (np). H-E. Barra = 1,2 µm.Figura 6: Corte sagital del ovario en el Estadio Ninfal III, se observan las ovariolas (OV) limi-tadas por la vaina externa (VE). En cada ovariola, se identifica en la región distal se identificael germario (G) y en la región media y proximal el vitelario (VT). En el vitelario, se observanovocitos en postpaquinema (pp) y en el estadio previtelogénico I (prI). Se destacan los cuer-pos proteinosos (cu) en la vesícula germinal de estos ovocitos. H-E. Barra = 18,2 µm.Figura 7: Región proximal de las ovariolas en el Estadio Ninfal IV rodeada por abundantecuerpo graso (CGR). Se observan folículos ováricos constituidos por una capa continua decélulas foliculares planas (cf), con ovocitos en el estadio previtelogénico II (prII). Se destacala vesícula germinal (g) en posición central con numerosos cuerpos proteinosos (cu). H-E.Barra = 18,2 µm.Figura 8: Región proximal de la ovariola en el Estadio Ninfal V. Se observa un folículo basalrodeado por epitelio folicular simple (EP) y en su interior un ovocito primario en el estadioprevitelogénico III (prIII). En la vesícula germinal se destacan los nucleolos (n) entre los cuer-pos proteinosos (cu). H-E. Barra = 18,2 µm.

A. A. Michel: Análisis histológico del ovario en Baeacris punctulatus38
ca, observándose solo algunas células en losestadios de profase y metafase, mientras queen el extremo proximal se observan ovocitosprimarios (en grupos de 2 o 3 células rodea-dos por células prefoliculares) en el primerperíodo de crecimiento, de los cuales en cadacorte pueden observarse aproximadamente 1en preleptonema, 3 en leptonema y 5 en pa-quinema inicial y avanzado (Tabla 1).
En el vitelario además de los ovocitos pri-marios en postpaquinema (3 o 4) y en el esta-dio previtelogénico I (aproximadamente 10células dispuestas linealmente), se observan 4o 5 ovocitos en una etapa más avanzada dediplonema que será considerada como estadioprevitelogénico II. Estos ovocitos son ligera-mente ovoideos y aumentan de tamaño haciael extremo proximal de la ovariola, con undiámetro mayor que oscila entre 72 y 120 µm.Estos ovocitos previtelogénicos no ocupancompletamente la luz del folículo ovárico, elcual está constituido aproximadamente por10 a 26 células foliculares alargadas con nú-cleos ovoides y abundante citoplasma basófiloentre ellos (Tabla 1, Fig. 7).
En los ovocitos en estadio previtelogénicoII, la vesícula germinal es grande y esférica(42 a 54 µm), se ubica en el centro del mis-mo y está rodeada por una envoltura nu-clear fuertemente basófila, observándose enella una mayor cantidad de cromatina des-condensada y de cromómeros que se distri-buyen en forma homogénea sin llegar a cu-brir completamente el jugo nuclear. En elinterior de la vesícula germinal de estos ovo-citos se observan aproximadamente 11 cuer-pos proteinosos de menor tamaño que en elestadio anterior y 2 o 3 macronucleolosfuertemente acidófilos y numerosos micronu-cleolos ubicados entre los cuerpos basófiloso bien superpuestos con ellos. El citoplasmagranular de estos ovocitos presenta una ba-sófilia menor que en el estadio I de previtelo-génesis, observándose pequeños gránulosfuertemente acidófilos próximos a la envol-tura nuclear y algunos de menor tamaño enel resto del citoplasma (Tabla 2, Fig. 7).
Estadio Ninfal V (23 a 30 días después de laeclosión de las ninfas).— En este estadio, los
ovarios están rodeados por gran cantidad decuerpo graso, tubos de Malpighi y traqueo-las, y entre las ovariolas se observa abun-dante tejido graso. En cada ovariola puedeobservarse claramente el límite entre germa-rio y vitelario, marcado por una delgadacapa de células prefoliculares, y dentro delvitelario, cada folículo ovárico está separadopor 2 a 3 capas de células interfolicularesentre los ovocitos en el estadio previtelogéni-co I y de 6 a 8 capas entre los ovocitos enlos estadios previtelogénicos II y III.
En estas hembras, el germario ocupa solouna pequeña zona del extremo distal de cadaovariola, observándose una importante re-ducción en el número de ovogonias (12 a 14por corte longitudinal) y de actividad mitóti-ca de las mismas, ya que la mayoría se en-cuentran en interfase y solo algunas en pro-fase mitótica. A continuación de las ovogo-nias, en el extremo proximal del germario,se observan 6 a 8 ovocitos primarios en pre-leptonema y leptonema y 3 o 4 en paquine-ma. El vitelario en cambio abarca la mayorparte de la ovariola, observándose entre 15 y16 ovocitos primarios en diferentes etapasdel segundo periodo de crecimiento. En elextremo distal del vitelario, se observa lamisma cantidad de ovocitos primarios en lossubestadios de postpaquinema y diplonema(estadios previtelogénicos I y II) que en elcuarto estadio ninfal. Sin embargo, los ovo-citos en el estadio previtelogénico II han au-mentado de tamaño, llegando a tener losmás grandes un diámetro mayor de 210 µm.Estos ovocitos ocupan completamente la luzdel folículo ovárico, el cual llega a tenerhasta 45 células foliculares en los más gran-des. En el extremo proximal del vitelario (acontinuación de los ovocitos anteriores), seobservan 3 o 4 ovocitos alargados (con undiámetro mayor que oscila entre 222 y 288µm) en una etapa posterior del subestadio dediplonema, la cual será considerada comoestadio previtelogénico III (Tabla 1). Estosovocitos ocupan completamente la luz delfolículo ovárico, el cual está constituido porun epitelio cúbico simple de 45 a 65 célulasen activa división. Las células foliculares deestos folículos, con limites celulares clara-

Acta zoológica li l loana 52 (1–2): 30–63, 2008 39
mente visibles, presentan grandes núcleosesféricos con la cromatina condensada engránulos gruesos y finos y escaso citoplasmagranular basófilo alrededor de los mismos.El epitelio folicular descansa sobre una lá-mina basal fuertemente acidófila, que estáen íntimo contacto con la vaina externa decada ovariola y correspondería a la túnicapropia.
En los ovocitos en el estadio previtelogéni-co III, la vesícula germinal, delimitada poruna envoltura nuclear fuertemente basófila,alcanza un diámetro de 60 µm y está ligera-mente desplazada al extremo posterior delovocito. En el interior de la vesícula germinal,la cromatina está completamente desconden-sada, ya que se presenta en forma de finas yabundantes hebras con numerosas condensa-ciones basófilas (cromómeros), que cubrencompletamente el jugo nuclear dando la ima-gen de cromosomas plumulados. Entre la cro-matina, se observan 6 a 7 cuerpos proteinosospequeños y compactos, y 3 a 4 nucleolosgranulares fuertemente acidófilos, ambas es-tructuras ubicadas preferentemente en el cen-tro de la vesícula germinal (Fig. 8).
El ooplasma de los ovocitos en el estadioprevitelogénico III está constituido por finasy abundantes granulaciones basófilas que leconfieren una basofilia menor a la observadaen los ovocitos de estadios anteriores, dondela granulación es más gruesa y menos abun-dante. En el ooplasma perinuclear, al igualque en los otros estadios de previtelogénesis,se observa pequeños gránulos fuertementeacidófilos próximos a la envoltura nuclear.En el ooplasma perinuclear y rodeando a lavesícula germinal de algunos de estos ovoci-tos se puede observar además una o más ve-sículas grandes o “núcleos accesorios”, conuna envoltura idéntica a la envoltura nu-clear del ovocito y en cuyo interior se obser-van pequeños acúmulos de material basófiloy acidófilo (Tabla 2, Fig. 9).
3. MORFOGÉNESIS DEL OVARIO Y OVOGÉNESIS
EN EL IMAGO
Imago 1 (24 y 72 horas después de la mudaimaginal).— En hembras vírgenes, en lasprimeras horas de imago que aún no fueron
copuladas, se observa abundante tejido grasorodeando externamente a cada ovario y, entrelas ovariolas, alrededor del extremo proxi-mal. En el germario de cada ovariola, elnúmero de ovogonias y de ovocitos prima-rios en el primer período de crecimiento esconsiderablemente menor al observado enlos primeros estadios ninfales, ya que lamayoría de las células se encuentran en el“segundo periodo de crecimiento”, por lo queen estas hembras es difícil observar las pri-meras etapas de la ovogénesis. A continua-ción del filamento terminal, en el germario seobservan aproximadamente 5 ovogonias eninterfase o bien en los primeros estadios dela mitosis, 2 o 3 ovocitos primarios en lepto-nema y 4 o 5 en paquinema, ambos tiposcelulares rodeados por células del tejido pre-folicular. Siguiendo al germario de cadaovariola se observa un acúmulo de aproxi-madamente 8 células prefoliculares que loseparan del vitelario.
En estas hembras, en la región distal ymedia del vitelario se observan aproximada-mente 3 o 4 ovocitos primarios en postpa-quinema y 14 ovocitos en el segundo perio-do de crecimiento en los estadios previtelo-genicos I, II y III, ya mencionados en ejem-plares del Estadio Ninfal V, mientras que enel extremo proximal se observan 4 ovocitosen el estadio previtelogénico IV, los cuales aligual que los restantes ovocitos del vitelario,se disponen linealmente y aumentan progre-sivamente de tamaño hacia el extremo de laovariola (Tabla 1). Entre el extremo proxi-mal de cada ovariola y el pedicelo se obser-va un tapón epitelial de mayor espesor queen ejemplares del Estadio Ninfal V, el cualestá constituido por 6 capas de 2 células fo-liculares delgadas, con grandes núcleos enposición externa y escaso citoplasma granu-lar levemente basófilo entre ellos. El epiteliofolicular que rodea a cada ovocito previtelo-génico del vitelario se apoya sobre una del-gada lámina basal fuertemente acidófila otúnica propia, la cual se prolonga a travésdel tejido interfolicular y separa a ambostejidos de la vaina epitelial externa que ro-dea a cada ovariola (Fig. 10).
Los ovocitos en el estadio previtelogénico
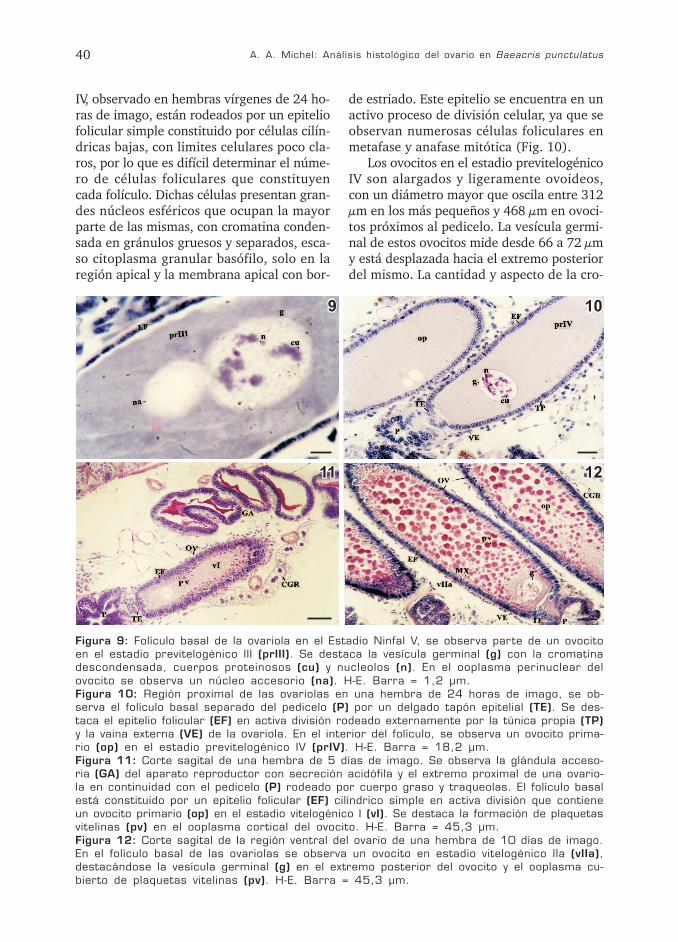
A. A. Michel: Análisis histológico del ovario en Baeacris punctulatus40
IV, observado en hembras vírgenes de 24 ho-ras de imago, están rodeados por un epiteliofolicular simple constituido por células cilín-dricas bajas, con limites celulares poco cla-ros, por lo que es difícil determinar el núme-ro de células foliculares que constituyencada folículo. Dichas células presentan gran-des núcleos esféricos que ocupan la mayorparte de las mismas, con cromatina conden-sada en gránulos gruesos y separados, esca-so citoplasma granular basófilo, solo en laregión apical y la membrana apical con bor-
de estriado. Este epitelio se encuentra en unactivo proceso de división celular, ya que seobservan numerosas células foliculares enmetafase y anafase mitótica (Fig. 10).
Los ovocitos en el estadio previtelogénicoIV son alargados y ligeramente ovoideos,con un diámetro mayor que oscila entre 312µm en los más pequeños y 468 µm en ovoci-tos próximos al pedicelo. La vesícula germi-nal de estos ovocitos mide desde 66 a 72 µmy está desplazada hacia el extremo posteriordel mismo. La cantidad y aspecto de la cro-
Figura 9: Folículo basal de la ovariola en el Estadio Ninfal V, se observa parte de un ovocitoen el estadio previtelogénico III (prIII). Se destaca la vesícula germinal (g) con la cromatinadescondensada, cuerpos proteinosos (cu) y nucleolos (n). En el ooplasma perinuclear delovocito se observa un núcleo accesorio (na). H-E. Barra = 1,2 µm.Figura 10: Región proximal de las ovariolas en una hembra de 24 horas de imago, se ob-serva el folículo basal separado del pedicelo (P) por un delgado tapón epitelial (TE). Se des-taca el epitelio folicular (EF) en activa división rodeado externamente por la túnica propia (TP)y la vaina externa (VE) de la ovariola. En el interior del folículo, se observa un ovocito prima-rio (op) en el estadio previtelogénico IV (prIV). H-E. Barra = 18,2 µm.Figura 11: Corte sagital de una hembra de 5 días de imago. Se observa la glándula acceso-ria (GA) del aparato reproductor con secreción acidófila y el extremo proximal de una ovario-la en continuidad con el pedicelo (P) rodeado por cuerpo graso y traqueolas. El folículo basalestá constituido por un epitelio folicular (EF) cilíndrico simple en activa división que contieneun ovocito primario (op) en el estadio vitelogénico I (vI). Se destaca la formación de plaquetasvitelinas (pv) en el ooplasma cortical del ovocito. H-E. Barra = 45,3 µm.Figura 12: Corte sagital de la región ventral del ovario de una hembra de 10 días de imago.En el folículo basal de las ovariolas se observa un ovocito en estadio vitelogénico IIa (vIIa),destacándose la vesícula germinal (g) en el extremo posterior del ovocito y el ooplasma cu-bierto de plaquetas vitelinas (pv). H-E. Barra = 45,3 µm.

Acta zoológica li l loana 52 (1–2): 30–63, 2008 41
matina en el nucleoplasma es similar a laobservada en los estadios anteriores de pre-vitelogénesis, pero en estos ovocitos, loscuerpos proteinosos son menos compactos yparecen grandes acúmulos basófilos dispues-tos preferentemente en la región central dela misma. Además se observan 2 macronu-cleolos compactos y fuertemente acidófilossuperpuestos con los cuerpos proteinosos ynumerosos micronucleolos junto a la carainterna de la envoltura nuclear. El ooplasmade estos ovocitos está constituido por finasgranulaciones acidófilas con escasas plaque-tas vitelinas, pequeñas y fuertemente acidófi-las en la periferia del ovocito. La membranaplasmática es delgada y fuertemente acidófi-la (Tabla 2, Fig. 10). Tanto el aumento dediámetro de la vesícula germinal y el despla-zamiento de ésta hacia el extremo posteriordel ovocito como la ligera acidofilia delcitoplasma, indicarían la finalización delsegundo periodo de crecimiento (que incluyeovocitos en postpaquinema y en los diferen-tes estadios de previtelogénesis) que tienelugar dentro del subestadio de diplonema dela meiosis.
En el ooplasma de algunos ovocitos pre-vitelogénicos IV, se puede observar un núme-ro variable de “núcleos accesorios” que sepresentan como vesículas de diferentes diá-metros, íntimamente unidas entre sí y enproximidad a la vesícula germinal, rodeadaspor una membrana semejante a la envolturanuclear pero de menor espesor, observándo-se con gran aumento, pequeños canales queatraviesan las membranas de vesículas veci-nas y escaso material granular basófilo en elinterior de las mismas. Las diferencias mor-fológicas existentes entre estos núcleos acce-sorios y los descriptos durante el desarrollopostembrionario probablemente se deban aque se encuentran en una etapa diferente desu formación.
Imago 2 (5 días después de la muda imagi-nal).— En estas hembras se observan impor-tantes cambios en sus gónadas, observándo-se en el extremo proximal del vitelario decada ovariola ovocitos en diplonema en unaetapa inicial del proceso de vitelogénesis,
que serán considerados en este trabajo comoovocitos en el estadio vitelogénico I, indiciodel comienzo del “tercer periodo de creci-miento” del ovocito (Tabla 1).
Los ovocitos primarios en el estadio vite-logénico I están rodeados por un epitelio fo-licular simple con limites celulares poco vi-sibles, de un espesor considerablemente ma-yor al observado en ovocitos al final de laprevitelogénesis y, al igual que en ovocitosprevitelogénicos IV, se encuentra en un activoproceso de división celular, ya que en él seobservan numerosas células en metafase yanafase mitótica. Dicho epitelio está consti-tuido por células cilíndricas altas con gran-des núcleos ovoideos en posición basal, conla cromatina condensada en gránulos grue-sos y separados y envoltura nuclear poco vi-sible. El citoplasma es basófilo finamentegranular y está localizado principalmente enla región apical de la célula y la membranaapical presenta un fino borde estriado. Elepitelio folicular de estos ovocitos se apoyasobre una lámina basal fuertemente acidófi-la, de mayor espesor que en los restantes fo-lículos del vitelario (Fig. 11).
En ovocitos del estadio vitelogénico I seobserva un importante crecimiento que semanifiesta principalmente en la cantidad deooplasma de los mismos, ya que la vesículagerminal mantiene un diámetro similar alobservado en hembras de 24 horas de imago,mientras que el diámetro mayor de estos ovo-citos oscila entre 540 y 594 µm. En estos ovo-citos, la vesícula germinal ha completado sudesplazamiento hacia el extremo posteriordel mismo (próximo al pedicelo de la ovario-la), lo que indicaría el inicio del proceso devitelogénesis. La cromatina se encuentra enforma de finas hebras y abundantes gránulosbasófilos (cromómeros) que cubren la mayorparte del jugo nuclear y la envoltura nucleares claramente visible. En la vesícula germinalde estos ovocitos no se observan cuerpos pro-teinosos y los nucleolos se presentan comopequeñas granulaciones fuertemente acidófi-las o micronucleolos. El citoplasma está cons-tituido en su mayor parte por finas granula-ciones acidófilas, observándose solo en la re-gión cortical pequeñas plaquetas vitelinas

A. A. Michel: Análisis histológico del ovario en Baeacris punctulatus42
fuertemente acidófilas. En este estadio de vite-logénesis, el ovocito comienza a sintetizarparte de su cubierta externa, observándoseentre el epitelio folicular y el oolema, unadelgada matriz extracelular constituida porfinas granulaciones fuertemente acidófila quecorresponde a la envoltura vitelina en forma-ción (Tabla 2, Fig. 11).
En estas hembras, alrededor del extremoproximal de las ovariolas, se observa ungran desarrollo del cuerpo graso, cuyos adi-pocitos de forma cilíndrica, se unen forman-do largos cordones irregulares. Dichas célu-las presentan núcleos esféricos y fuertementebasófilos en posición central, con la croma-tina condensada en gránulos gruesos quecubren completamente el nucleoplasma. Al-rededor del núcleo se observa abundantecitoplasma de aspecto ligeramente esponjoso(Fig. 11).
Imago 3 (10 días después de la muda imagi-nal).— En estas hembras, en el extremoproximal del vitelario de cada ovariola, seobservan ovocitos primarios en una etapamás avanzada del proceso de vitelogénesisque será considerado como estadio vitelogé-nico II (Tabla 1). Estos ovocitos son ovoideosy presentan un rango de crecimiento mayoral observado en los ovocitos en el estadio vi-telogénico I, ya que su diámetro mayor osci-la entre 700 y 1500 µm. La vesícula germi-nal permanece en el extremo posterior delmismo, con un diámetro aproximado de 66µm. En este estadio de vitelogénesis, se dife-rencian dos tipos de ovocitos: 1) los de me-nor tamaño, que serán considerados comoovocitos en estadio vitelogénico IIa, en cuyavesícula germinal se observa parte de la cro-matina en forma de finas hebras y el restocondensada en gránulos fuertemente basófi-los que se concentra en pequeños acúmulosen la región central de la misma, donde seobserva además 6 micronucleolos. La envol-tura nuclear de estos ovocitos es completa-mente lisa (Fig. 12); y 2) los de mayor tama-ño, considerados como ovocitos en estadiovitelogénico IIb, en cuya vesícula germinalse observa mayor cantidad de cromatinacondensada y la envoltura nuclear es ligera-
mente ondulada, fragmentada en algunospuntos y con finas granulaciones basófilasjunto a la cara interna y externa de la mis-ma. Las características de la vesícula germi-nal de estos ovocitos indicarían que en lascélulas en el estadio vitelogénico IIb se hainiciado el proceso de condensación cromó-somica y fragmentación de la envoltura nu-clear. Por lo tanto, estos ovocitos se encuen-tran en la etapa previa a la ruptura de lavesícula germinal, indicando la finalizacióndel subestadio de diplonema. En el citoplas-ma de ambos tipos de ovocitos se observanfinas granulaciones acidófilas y abundantesplaquetas vitelinas esféricas de diferentesdiámetros, variando en cantidad y diámetrosegún el tamaño del ovocito. En el ooplasmade los ovocitos en el estadio vitelogenicos IIbse observa la mayor cantidad de plaquetasvitelinas (Tabla 2, Fig. 13).
Los ovocitos en el estadio vitelogénico IIestán rodeados por un epitelio folicular cons-tituido por células cilíndricas altas con for-ma de botella o clava, con grandes núcleosfuertemente basófilos ubicados en la base dela célula y la cromatina condensada en grá-nulos gruesos que cubren completamente elnucleoplasma. Alrededor del núcleo, se ob-serva escaso citoplasma basófilo finamentegranular, mientras que en la región apicalde la célula se observa abundante citoplas-ma homogéneo fuertemente basófilo. En losovocitos en el estadio vitelogénico IIb, seobserva un halo blanco en la región su-pranuclear de las células foliculares, quecorresponde a la imagen negativa del Golgi,indicando una intensa actividad secretora.En esta zona de la ovariola (al igual que enel resto del vitelario), entre la membranaapical del epitelio folicular y la membranaplasmática del ovocito en el estadio vitelogé-nico II, se observa una delgada matriz extra-celular constituida por finas granulacionesacidófilas (Tabla 2, Figs. 12 y 13).
Imago 4 (12 días después de la muda imagi-nal).— En estas hembras, el folículo basalde cada ovariola contiene un ovocito prima-rio en la etapa final del proceso de vitelogé-nesis incluido en el subestadio de diplonema

Acta zoológica li l loana 52 (1–2): 30–63, 2008 43
de la meiosis, que será considerado como es-tadio vitelogénico III. Como en estos ovocitosaún no se realizó el proceso de coriogénesisque en estos insectos se produce en el inte-rior de cada ovariola antes de la ovulación,fue necesario hacer una subdivisión dentrode este estadio de vitelogénesis y por lo tan-to los ovocitos del folículo basal, que care-cen de corion, serán considerados como ovo-citos en el estadio vitelogénico IIIa. Estosovocitos presentan un diámetro mayor queoscila entre 2,64 a 3,36 µm y por lo tanto seencuentran en la etapa final del tercer perio-do de crecimiento (Fig. 14).
Los ovocitos en el estadio vitelogénicoIIIa, presentan un ooplasma completamentecargados de plaquetas vitelinas de formahexagonal que se unen formando grandesmasas de vitelo en el interior del mismo,observándose restos de citoplasma granularacidófilo solo en la región cortical, donde seobserva una estrecha zona de gránulos dediferentes diámetros fuertemente acidófilosjunto a la cara interna del oolema. En estosovocitos la vesícula germinal no es visible(Tabla 2, Fig. 14). En el estadio vitelogénicoIIIa se completa la síntesis de la envolturavitelina, la cual se identifica como una del-gada matriz extracelular de aspecto homo-géneo y levemente acidófila, ubicada entreel oolema y la membrana plasmática delepitelio folicular (Fig. 15). Cada ovocito enel estadio vitelogénico IIIa está rodeado porepitelio folicular simple, constituido por cé-lulas cúbicas en la mayor parte del folículoque sintetizarán el corion, y células cilíndri-cas en el extremo posterior del ovocito rela-cionadas con la formación del micrópilo yearópilo del huevo. El núcleo de las célulasfoliculares cúbicas es esférico en posiciónbasal, con la cromatina condensada en grá-nulos finos distribuidos en forma homogéneacubriendo completamente el nucleoplasma y1 o 2 nucleolos acidófilos claramente visi-bles. El citoplasma, fuertemente basófilo yfinamente granular, está concentrado princi-palmente en la región apical de la célula.Las células cilíndricas presentan un núcleo decaracterísticas similares a las anteriores,pero el citoplasma es menos basófilo y de
aspecto vacuolar en la región basal y apicalde las mismas (Tabla 2, Fig. 15).
En las ovariolas de hembras de 12 díasde imago, en íntimo contacto con la mem-brana basal del epitelio folicular de los ovo-citos del vitelario se observa una delgadalámina basal levemente acidófila, que co-rresponde a la túnica propia. En cortestransversales de estas hembras, coloreadoscon la técnica de Verhoeff, se determinó quela lámina basal que rodea a los ovocitos delvitelario, está constituida por una gruesacapa interna de fibras de colágeno en inti-mo contacto con la membrana basal del epi-telio folicular y una delgada capa externade fibras elásticas en contacto con la vainaexterna de la ovariola (Fig. 16).
En estas hembras, casi todos los ovocitosdel extremo proximal de las ovariolas seencuentran en la etapa final de la vitelogéne-sis, observándose en cada ovario la madura-ción simultánea de aproximadamente 18ovocitos, lo cual coincide con el límite supe-rior del rango alcanzado en el número dehuevos por desoves de esta especie. Sin em-bargo, en un número reducido de ovariolas,a continuación de los ovocitos en el estadiovitelogénico I, se observa un “cuerpo de re-absorción”, producto de la degeneración deun ovocito vitelogénico II que no ha comple-tado su desarrollo (Fig. 14). En estas hem-bras se observa un ligero adelgazamientodel tapón epitelial que impide la comunica-ción entre el lumen de cada ovariola y la luzdel extremo distal del oviducto lateral res-pectivo, quedando constituido solamente por3 o 4 capas de células con núcleos pequeñosy abundante citoplasma levemente basófiloentre ellos (Fig. 15).
Los “cuerpos de reabsorción” observadosen estas hembras se encuentran en las prime-ras etapas de su formación, ya que son alar-gados y miden aproximadamente 1,5 mm delongitud. Cada uno de estos cuerpos está se-parado del folículo anterior, por tejido inter-folicular, en el cual la mayor parte de lascélulas se disponen en forma irregular, y delpedicelo por un tapón epitelial de célulascompletamente desordenadas, entre las quese observan acúmulos de pigmentos color

A. A. Michel: Análisis histológico del ovario en Baeacris punctulatus44
ámbar producidos por la reabsorción del vi-telo del ovocito degenerado. Cada cuerpo dereabsorción está limitado externamente poruna gruesa túnica propia lisa y fuertementeacidófila, que se pliega profundamente en elextremo anterior (tejido interfolicular) yposterior (tapón epitelial) del mismo. En elinterior de los cuerpos de reabsorción se ob-servan numerosas células foliculares esféri-cas y de disposición irregular, con escaso
citoplasma granular levemente acidófilo ygrandes núcleos esféricos con la cromatinacondensada en gránulos finos que cubrencompletamente el nucleoplasma o bien pe-queños y picnóticos. Entre las células folicu-lares se pueden observar grandes vacuolas yrestos de vitelo fuertemente acidófilo.
Imago 5 (14 días de imago durante la cópulay a las 12, 24 y 48 horas después de la cópu-
Figura 13: Corte sagital de la región ventral del ovario de una hembra de 10 días de imago.Se observa parte de un folículo basal de una ovariola con un ovocito en estadio vitelogénicoIIb (vIIb), se destaca la vesícula germinal (g) en el extremo posterior del ovocito con la en-voltura nuclear (en) ondulada y fragmentada, el ooplasma perinuclear cubierto de plaquetasvitelinas (pv), y externamente al oolema una gruesa matriz extracelular (MX). Se destaca lascaracterísticas citológicas del epitelio folicular (EF) con intensa actividad secretora. H-E.Barra = 1,2 µm.Figura 14: Corte sagital del ovario de una hembra de 12 días de imago. En el folículo basalde las ovariolas, se observan ovocitos en estadio vitelogénico IIIa (vIIIa) con el ooplasmacargado de vitelo (v) y rodeados por un epitelio folicular (EF) cúbico simple con células cilín-dricas en el extremo posterior del ovocito. Se destaca una ovariola con un cuerpo de reab-sorción (CR) en su extremo proximal. H-E. Barra = 288 µm.Figura 15: Región proximal de la ovariola de una hembra de 12 días d imago. Se observa elfolículo basal con un ovocitos en estadio vitelogénico IIIa (vIIIa) separado del oviducto lateral(OL) por un grueso tapón epitelial (TE). Se destaca la formación de gránulos acidófilos (GC)junto a la cara interna del oolema y externamente a esta la envoltura vitelina (MV). Se obser-va la modificación del epitelio folicular (EF) en la región micropilar. H-E. Barra = 45,3 µm.Figura 16: Corte transversal de un ovocito en el estadio vitelogénico IIa (vIIa) de una hembrade 12 días de Imago. Se detaca el ooplasma cargado de vitelo (v). Externamente al epiteliofolicular (EF), se observa la túnica propia (TP) constituida por fibras elásticas. H.V. Barra =6,6 µm.

Acta zoológica li l loana 52 (1–2): 30–63, 2008 45
la).— En hembras fijadas durante la primeracópula, los ovarios están completamente de-sarrollados y en cada ovariola se puede dis-tinguir claramente el germario con escasasovogonias y ovocitos primarios en el primerperiodo de crecimiento y el vitelario conovocitos primarios en el segundo y tercerperiodo de crecimiento, observándose desdeel filamento terminal al pedicelo, las dife-rentes etapas de la ovogénesis descriptas enhembras adultas de diferentes edades (Tabla1). Sin embargo, en el folículo basal decada ovariola de estas hembras, se observaun ovocito primario en la etapa final del es-tadio III de vitelogénesis, considerado comoestadio vitelogénico IIIb. Estos ovocitos man-tienen las dimensiones y características cito-lógicas de los observados en el estadio vite-logénico IIIa, pero están cubiertos externa-mente además de la envoltura vitelina por elcorion. El corion que rodea a estos ovocitoses delgado, fuertemente acidófilo y estáconstituido por una capa interna o endoco-rion, ubicada junto a la envoltura vitelina, yuna capa externa o exocorion, delgada, ho-mogénea, fuertemente acidófila y con depre-siones hexagonales en su superficie externa,en íntimo contacto con las células folicula-res que rodean al ovocito. En estos ovocitosen los cuales se ha completado el proceso decoriogénesis, el epitelio folicular es delgadoy está formado por células cúbicas bajas deescasa actividad secretora. En ellas se obser-van núcleos esféricos ligeramente comprimi-dos sobre la membrana basal, la cromatinaestá condensada en gránulos gruesos y sepa-rados, el citoplasma, ubicado principalmen-te en la región apical, es basófilo finamentegranular. La membrana apical de las célulasfoliculares está en contacto directo con elexocorion (Tabla 2, Fig. 17).
En hembras fijadas durante la cópula, elfolículo basal de cada ovariola está rodeadoexternamente por la túnica propia y la vainaexterna completamente adelgazadas, y acontinuación de este folículo se observa undelgado tapón epitelial constituido por 2 o 3capas de células, que impiden el descenso delos ovocitos maduros a través del pedicelohacia la luz del oviducto lateral.
En los ovarios de hembras aisladas delmacho y fijadas a las 12, 24 y 48 horas des-pués de la primera cópula, se observaron enel interior de los 2 últimos folículos del vite-lario, ovocitos en los estadios vitelogénicosIIIa y IIIb y a continuación de éstos un grue-so tapón epitelial que impide el descenso delos ovocitos maduros.
Imago 6 (15, 20 y 25 días después de lamuda imaginal).— Las observaciones reali-zadas en gónadas y demás órganos reproduc-tivos de estas hembras (ya copuladas), mos-traron un desarrollo completo de la ovogé-nesis dentro del ovario, observándose en elinterior del folículo basal de cada ovariola,ovocitos en el estadio vitelogénico IIIa o IIIb(con o sin corion) y en los ovarios de algu-nas de estas hembras indicios de la primeraovulación. En estas hembras, cada ovariolacontiene la serie completa de las diferentesetapas de la ovogénesis, desde ovogonias enel extremo distal del germario hasta ovocitosprimarios en el estadio vitelogénico II en elextremo proximal del vitelario, ya que losovocitos en el estadio vitelogénico III fueronovulados (Tabla 1). A continuación del últi-mo folículo de cada ovariola se observa unaestrecha zona de tejido interfolicular (concélulas aplanadas dispuestas en capas para-lelas) que lo separa de un corto conducto deparedes gruesas y luz estrecha ubicado en labase de la ovariola. Cada uno de estos con-ductos corresponde a un “cuerpo lúteo”, for-mado por la reducción progresiva del folícu-lo basal de la ovariola, luego de la ovula-ción de un ovocito maduro en el estadio vite-logénico IIIb.
La longitud de cada cuerpo lúteo varíasegún el grado de contracción del folículo,pero en general, están constituidos por 3 omás capas de células foliculares en degene-ración que rodean a una pequeña cavidadcentral. Las células foliculares son pequeñasy ovoideas y, en ellas, se observan grandesnúcleos fuertemente basófilos con la croma-tina condensada en gránulos gruesos quecubren completamente el nucleoplasma yescaso citoplasma levemente acidófilo alre-dedor de los mismos. Cada cuerpo lúteo está

A. A. Michel: Análisis histológico del ovario en Baeacris punctulatus46
limitado externamente por una delgada túni-ca propia, que forma pequeños pliegues enel extremo superior e inferior del folículo,producidos por su proceso de contracción.
En ovarios de hembras fijadas entre los15 y 25 días de imago, en el extremo proxi-mal de algunas ovariolas, a continuacióndel tapón epitelial que separa el folículo
basal del pedicelo, se observaron restos defolículos con grandes vacuolas cargadas depigmentos amarillos. En estas hembras seobservaron ovocitos en el estadio vitelogéni-co IIIb (con el extremo micropilar dirigidohacia el extremo posterior del abdomen) enla región distal y media de los oviductos la-terales, con un corion de aspecto diferente al
Figura 17: Corte sagital del ovario de una hembra de 14 días de imago fijada durante la cópu-la. Se observa parte del folículo basal de dos ovariolas con ovocitos en estadio vitelogénico IIIb(vIIIb), cargados de vitelo (v). Entre el epitelio folicular (EF) y la envoltura vitelina (MV) de cadaovocito, se observa un corion formado por una gruesa capa interna o endocorion (COI) y unadelgada capa externa o exocorion (COE). Externamente al epitelio folicular de cada ovariola seobserva la túnica propia (TP) y la vaina externa (VE). H-E. Barra = 1,2 µm.Figura 18: Corte sagital de una hembra de 16 días de imago. Se observa el extremo proxi-mal de una ovariola con ovocitos en el estadio vitelogénico II (vII), un pequeño cuerpo lúteo(CL) y a continuación del pedicelo (P), la región distal del oviducto lateral (OL) en cuya luz seobserva un ovocitos en estadio vitelogénico IIIb (vIIIb). En estos ovocitos, en los que el vitelose ha desprendido por el procesamiento histológico, se observa un aumento de espesor ycambio de coloración del exocorion (COE). H-E. Barra = 288 µm.Figura 19: Corte sagital de una hembra de 30 días de imago. Se observa el extremo proxi-mal de las ovariolas con ovocitos primarios (op) en estadio vitelogénicoIIIa (vIIIa) y a continua-ción de estos un cuerpo lúteo (CL), producto de la ovulación de un ovocito maduro. H-E.Barra = 288 µm.Figura 20: Extremo proximal de la ovariola de una hembra de 30 días de imago, donde seobserva parte de un folículo basal con un ovocito en estadio vitelogénicoIIIa (vIIIa), separadodel cuerpo lúteo (CL) por un delgado tapón epitelial (TE). Se destaca las modificaciónes delepitelio folicular que rodea al ovocito con el del cuerpo lúteo, con células foliculares (cf) dis-puestas en forma irregular y numerosos núcleos picnótico (np). Externamente al epitelio fo-licular se observa la túnica propia (TP) plegada a nivel del tapón epitelial y la vaina externa(VE) lisa. Se destaca el ooplasma del ovocito cargado de grandes plaquetas vitelinas (v) ypequeños gránulos acidófilos (GC) junto a la cara interna del oolema. H-E. Barra = 18,2 µm.

Acta zoológica li l loana 52 (1–2): 30–63, 2008 47
observado en ovocitos maduros dentro de laovariola. El corion de estos ovocitos, presen-ta una coloración ligeramente pardusca y unimportante aumento del espesor del exo yendocorion, lo cual se acentúa más en elextremo posterior del ovocito o región mi-cropilar (Fig. 18).
Aunque en estas hembras, todas las ova-riolas presentan un cuerpo lúteo a continua-ción del folículo basal, el número de ovoci-tos dentro del oviducto lateral respectivo fuemenor al número de ovocitos ovulados. Porotra parte, en estas hembras no se observa-ron ovocitos maduros ni en la luz del oviduc-to medio ni en la de la cámara genital.
Imago 7 (30 días de imago y al finalizar lapuesta).— Las observaciones realizadas enestas hembras, muestran en el folículo basalde cada ovariola, un ovocito en el estadiovitelogénico IIIa (sin corion), separado delpedicelo por un cuerpo lúteo reducido. Cadacuerpo lúteo contiene abundantes células fo-liculares, cuyos núcleos presentan un altogrado de condensación de la cromatina obien son picnóticos y están rodeados por uncitoplasma vacuolar levemente acidófilo. Lapresencia de un ovocito completamente de-sarrollado y un cuerpo lúteo reducido (pro-ducto de la primera ovulación) en el extremoproximal de cada una de las ovariolas deestas hembras, indicaría que en estos ovariosse estaría por producir la segunda ovulación(Figs. 19 y 20). En estos ejemplares no seobservaron ovocitos en el estadio vitelogéni-co III ni en oviductos ni en cámara genital.Sin embargo, se observó una amplia comu-nicación entre la luz de los oviductos latera-les y el oviducto medio y un aumento de diá-metro de la dilatación dorsal de la cámaragenital, lo que indicaría el paso a través deestos conductos de ovocitos recientementeovulados.
DISCUSIÓN Y CONCLUSIONES
En este trabajo, se realiza una descripciónhistológica sobre el desarrollo del ovario deBaeacris punctulatus, en la cual se pone demanifiesto las etapas más importantes en su
proceso de diferenciación durante el desarro-llo embrionario, postembrionario y el imago.Al mismo tiempo, se establecen parámetrosmorfofisiológicos para la identificación delas células sexuales durante las diferentes eta-pas del proceso de ovogénesis. Las caracterís-ticas observadas en el desarrollo y madura-ción del ovario de Baeacris punctulatus secomparan con los aspectos mencionados so-bre este proceso para otras especies de Acridi-dae, como Anacridium aegyptium, Melano-plus differentialis, Locusta migratoria migra-torioides, Poecilocerus pictus, Schistocercagregaria y para otros insectos.
Mediante las observaciones realizadas, sedeterminó que en Baeacris punctulatus losovarios se identifican por su organizaciónestructural a partir del Estadio X del desarro-llo embrionario. En este estadio, cada ovarioestá constituido por el número total de esbo-zos que forman las ovariolas de la hembraadulta.
En el Estadio Embrionario XI, antes de laeclosión de las ninfas, en cada esbozo deovariola se identifica el rudimento del ger-mario con ovogonias en mitosis, preovocitosy ovocitos primarios en preleptonema, lepto-nema y cigonema de la profase I de la meio-sis. En Baeacris punctulatus como en Anacri-dium aegyptium (Colombo, 1953) los pre-ovocitos, por sus características citológicas,corresponden a ovogonias en interfase pre-meiótica. Sin embargo, la determinación dela iniciación de la meiosis ovogonial antesde la eclosión de las ninfas en Baeacris punc-tulatus no fue mencionada hasta el momentoen otras especies de Acrididae, ya que lostrabajos realizados por Nelsen (1934) enMelanoplus differentialis, Roonwal (1937) yGallois (1979a, 1979b) en Locusta migrato-ria migratorioides, Colombo (1953, 1955) yColombo y Bassato (1957) en Anacridiumaegyptium, mencionan que tanto en el ma-cho como en la hembra de estas especies, lameiosis comienza entre el primer y segundoestadio ninfal.
Los ovocitos primarios en insectos aligual que en vertebrados, durante la ovogé-nesis experimentarán una serie de transfor-maciones morfológicas e histoquímicas, tan-

A. A. Michel: Análisis histológico del ovario en Baeacris punctulatus48
to a nivel citoplasmático como nuclear, queles permite crecer y madurar. Los cambiosnucleares llevarán al final de la meiosis a laformación de un núcleo haploide y los cam-bios citoplasmáticos permiten la acumula-ción de los materiales necesarios para diri-gir el desarrollo embrionario temprano(Karp, 1987; Villecco, 1998; Alberts et al.,2004). En Baeacris punctulatus, al igual queen Anacridium aegyptium (Colombo, 1953),una vez iniciada la meiosis, los ovocitos pri-marios pasarán por tres periodos de creci-miento: el primero incluye desde el comien-zo de la profase I con el subestadio de pre-leptonema hasta el final del subestadio depaquinema, el segundo abarca el subestadiode postpaquinema y el período de previtelo-génesis del subestadio de diplonema, y eltercero abarca el periodo de vitelogénesisdel subestadio de diplonema de la profase dela primera división de maduración. Al finaldel tercer período de crecimiento, el ovocitoha completado su desarrollo en el ovario yestá en condiciones de ser ovulado.
Durante el primer estadio del desarrollopostembrionario de la hembra de Baeacrispunctulatus, las ovariolas son ligeramentealargadas y están constituidas por el fila-mento terminal y la región del germario conovogonias y preovocitos rodeados por célu-las mesodérmicas indiferenciadas y gruposde ovocitos primarios en preleptonema, lep-tonema, cigonema y paquinema rodeadospor células prefoliculares. En esta especie, aligual que en Anacridium aegyptium (Colom-bo, 1953), el paquinema por su larga dura-ción se dividió en dos etapas, las cuales seidentifican con facilidad por el comporta-miento de los cromososmas a lo largo delmismo. Mientras el leptonema y cigonemaabarcan periodos cortos que duran horas, elpaquinema tiene una duración considerable-mente mayor, abarcando varios días dentrodel desarrollo postembrionario. La variaciónde tamaño y la disposición con respecto alas células prefoliculares observadas en losovocitos primarios durante el primer estadioninfal de la hembra de B. punctulatus, indi-carían que estas células se encuentran en elprimer periodo de crecimiento.
Durante el segundo estadio ninfal, las ova-riolas crecen en longitud debido a la activamultiplicación de las células mesodérmicas ylas células de la línea germinal que en ellas sedesarrollan. Las ovogonias y los preovocitosubicados lejos del filamento terminal adquie-ren una cierta ordenación con respecto a lascélulas prefoliculares. Sin embargo, entre lasovogonias ubicadas en diferentes zonas delgermario, no existen diferencias citológicassignificativas, como sucede con las esperma-togonias en el macho de esta especie (Michel,2007), que justifiquen la clasificación de lasovogonias en primarias, secundarias y tercia-rias como lo establece Colombo (1953) paraAnacridium aegyptium.
En Baeacris punctulatus, entre los ovoci-tos en el primer periodo de crecimiento, seobservan numerosos núcleos picnóticos, quepueden haberse originado de ovogonias o decélulas prefoliculares que no continuaron sudesarrollo. Este material cromatínico extra-celular también fue observado por Colombo(1953) en ovariolas de igual estadio ninfalde A. aegyptium, y este autor afirma que di-cho material se origina de ovogonias en de-generación y cumple una importante funciónen la iniciación de la meiosis tanto en laovogénesis como en la espermatogénesis deesta especie. En estudios previos realizadossobre espermatogénesis en B. punctulatus(Michel, 2007), no se observaron núcleospicnóticos en los folículos testiculares, por loque en esta especie este material cromatínicoextracelular solamente podría intervenir enla iniciación de la meiosis ovogonial.
Al finalizar el segundo estadio ninfal, enel extremo proximal de cada ovoriola, seobservaron ovocitos primarios en postpaqui-nema dispuestos linealmente y rodeados porcélulas prefoliculares, por lo que en estaetapa del desarrollo postembrionario de B.punctulatus comienza a diferenciarse el vite-lario y se inicia el segundo periodo de creci-miento del ovocito. Las características cito-lógicas de los ovocitos en postpaquinema enesta especie es similar a la observada paraotros Acrididae como lo mencionan Colombo(1953), Bonhag (1958) y Chapman (1969).Coincidimos con estos autores en que estas

Acta zoológica li l loana 52 (1–2): 30–63, 2008 49
células por su forma, tamaño y relación conlas células prefoliculares, son fácilmenteidentificables en el extremo distal del vitela-rio de ovariolas panoísticas en desarrollo.
Durante el tercer estadio ninfal, se produ-cen importantes cambios en los ovarios deBaeacris punctulatus, ya que en la región delvitelario por primera vez se observan ovocitosprimarios en el estadio previtelogenico I delsubestadio de diplonema. El subestadio diplo-nema en insectos como en los restantes gruposde invertebrados y vertebrados es una faseextremadamente larga y de intensa actividadmetabólica, que se manifiesta tanto por elcomportamiento de los cromosomas comopor el aumento de la masa celular. Duranteeste subestadio, el crecimiento del ovocito de-penderá en gran parte de la actividad sinteti-zadora de las células foliculares que formanuna capa epitelial continua alrededor delmismo y se mantienen unidas por contactosen hendiduras que permiten el intercambio depequeñas moléculas originadas a partir deltejido circundante y que serán los precursoresde las proteínas vitelinas (Bonhag, 1958;Chapman, 1969; Karp, 1987; Saxena et al.,1988; Büning, 1994; Albert et al., 2004). EnB. punctulatus, el diplonema incluye el perio-do de previtelogenésis con cuatro estadios,que abarcan desde el comienzo del diplonemahasta la aparición de las primeras vesículasvitelinas y un periodo de vitelogénesis con tresestadios, en los cuales el ovocito acumulagran cantidad de proteínas vitelinas en sucitoplasma, dando lugar a un crecimientopronunciado en un corto periodo de tiempo,que finaliza con la formación de las cubiertasexternas del ovocito.
Los ovocitos en el estadio previtelogénicoI están rodeados por un epitelio folicularplano y se identifican fácilmente por su ta-maño, forma, basofília citoplasmática y lascaracterísticas de la vesícula germinal en lacual comienza el proceso de desespiraliza-ción de la cromatina, se forman los cuerposproteinosos y se visualizan claramente losnucleolos. En Baeacris punctulatus, los cuer-pos proteinosos estuvieron presentes en todoslos estadios de previtelogénesis, aunque sumorfología y ubicación en la vesícula ger-
minal fue cambiando en las diferentes eta-pas del proceso. Estas estructuras basófilastambién fueron mencionadas para ovocitosprevitelogénicos de Anacridium aegyptiumcomo aparato siderofilo o endonúcleos (Co-lombo, 1953), de Gryllus como cuerpos pro-teinosos o endocuerpos y de Xenopus comoorganelas esféricas (Büning, 1994). Aunqueestas estructuras varían en forma y cantidadsegún el grupo, coincidimos con ambos au-tores en que los cuerpos proteinosos estánestrechamente relacionados con las hebrasde DNA y junto con los nucleolos cumplenuna importante función en la síntesis de RNAdurante la profase meiótica del ovocito.
Durante el cuarto estadio ninfal de Baea-cris punctulatus, los ovarios alcanzan el ta-maño del adulto y están rodeados por abun-dante cuerpo graso, especialmente en la re-gión proximal de las ovariolas. Coincidimoscon Villeco (1998) y Albert et al. (2004) enque este aumento de volumen del cuerpo gra-so está directamente relacionado con el pro-ceso de previtelogénesis que en ellas se desa-rrolla. En el vitelario, los ovocitos primariosde mayor tamaño se encuentran en el segun-do estadio de previtelogénesis y se identifi-can por una marcada disminución de la ba-sofília citoplasmática, un aumento de espe-sor en la envoltura nuclear, aumento en lacantidad de cromatina descondensada y enel número de cuerpos proteinosos en la vesí-cula germinal.
Durante el quinto estadio ninfal de lahembra de Baeacris punctulatus, los ovariosestán rodeados por abundante cuerpo graso,gran cantidad de tubos de Malphigi y gran-des tráqueas. En las ovariolas se reduce lalongitud del germario y aumenta la del vite-lario como consecuencia de una variación enel número de células que cada uno de elloscontiene. En este estadio se diferencia el teji-do prefolicular que separa ambas regionesde la ovariola y el tejido interfolicular quesepara los folículos ováricos dentro del vite-lario, aunque en esta especie ambos tejidospresentan características citológicas simila-res. Coincidimos con Bonhag (1958) en lanecesidad de diferenciarlos según la posiciónque éstos ocupan en la ovariola.

A. A. Michel: Análisis histológico del ovario en Baeacris punctulatus50
En el quinto estadio ninfal, los ovocitosdel extremo proximal de la ovariola alcan-zan el estadio previtelogénico III y están ro-deados por un epitelio folicular cúbico sim-ple en activa división, que descansa sobreuna delgada lámina basal en íntimo contac-to con la vaina externa de la ovariola. Lavesícula germinal está ligeramente desplaza-da hacia el extremo posterior de la célula,la cromatina adquiere la característica for-ma de cromosomas plumulados, los cuerposproteinosos son pequeños y compactos y losnucleolos adquieren un aspecto granular. Elooplasma de estos ovocitos es de menor ba-sofília que en el estadio anterior de previte-logénesis y en su región perinuclear se for-man núcleos accesorios. Según Büning(1994), los cromosomas plumulados hansido identificados en algunos ovarios panoís-ticos de insectos usando métodos clásicos.Sin embargo, las hélices son muy pequeñas ydelicadas y solamente en algunas especiesde Orthoptera, como Locusta migratoria yDecticus albifrons, la estructura de las héli-ces de ovocitos en los últimos estadios deprevitelogénesis pueden ser comparables conla de los anfibios, mientras que en otros ova-rios panoíticos como Acheta domesticus, laimagen no es clara. Las imágenes obtenidasen las vesículas germinales de ovocitos alfinal del periodo de previtelogénesis deBaeacris punctulatus son idénticas a las ob-servadas en anfibios, por lo que coincidimoscon Büning (1994) en que el concepto gene-ral de cromosomas plumulados utilizadospara anfibios, puede ser también utilizadopara ovocitos de insectos con ovariolas pa-noísticas.
Bonhag (1958) menciona la formaciónde núcleos accesorios en el citoplasma corti-cal de ovocitos vitelogénicos de insectos conovariolas meroísticas como Hemiptera, Le-pidoptera, Coleoptera, Diptera y especial-mente en Hymenoptera, en los cuales ellosactuarían como precursores de los cuerposde vitelo. Hopkins (1964) en cambio analizala estructura y función de los núcleos acceso-rios durante los estadios de vitelogénesis deBombus terrestris y sostiene que ellos se ori-ginan a partir de la vesícula germinal, con-
tienen RNA y juegan un papel importante enla síntesis del vitelo y formación de la en-voltura vitelina. Mientras que Büning(1994) menciona la formación de núcleosaccesorios durante los estadios de previtelo-génesis y vitelogénesis en el citoplasma deovocitos de Hymenoptera, Mallophaga, Dip-tera (Chironomus, Pryne) y Mecoptera (Pano-rpa) y sostiene que aunque el proceso de for-mación y estabilización de los mismos es si-milar en los diferentes grupos mencionados,la estructura y la distribución en el ovocitoes diferente en cada uno de ellos y probable-mente estén involucrados en diferentes pro-cesos celulares. Según este autor, los núcleosaccesorios se originan por compartimentali-zación de la vesícula germinal durante losestadios de previtelogénesis y vitelogénesis yjuntos con los cuerpos proteinosos cumplenuna importante función como almacena-miento y transporte de ribonucleoproteínas yproteínas nucleares indispensables para lasprimeras etapas de embriogénesis. Los nú-cleos accesorios identificados en ovocitosprevitelogénicos de Baeacris punctulatus pre-sentan una estructura y distribución similara la observada en Eomenacanthus strami-neus (Mallophaga). Cabe mencionar que elhallazgo en B. punctulatus constituye la pri-mera cita para Orthoptera. Considerandoque los estudios realizados hasta el momen-to en los diferentes grupos de insectos nohan permitido determinar claramente la fun-ción y el destino de los núcleos accesorios,será necesario realizar posteriores estudioscon técnicas de inmunocitoquímica e histo-químicas, con el fin de contribuir al conoci-miento del papel que juegan estas estructu-ras durante el desarrollo del ovocito y delembrión.
En las primeras 24 horas, después de lamuda imaginal de la hembra Baeacris punctu-latus, se reduce el número de ovogonias y deovocitos primarios en el primer período decrecimiento, por lo que es difícil identificarlas primeras etapas de la ovogénesis. Los ovo-citos de mayor tamaño se encuentran en elestadio previtelogénico IV y están rodeadospor un epitelio folicular cilíndrico bajo enactiva multiplicación. Estos ovocitos son

Acta zoológica li l loana 52 (1–2): 30–63, 2008 51
alargados y ligeramente ovoideos, la vesículagerminal desplazada hacia el extremo poste-rior de la célula, presenta un gran desarrollode los cromosomas plumulados, grandescuerpos proteinosos y macronucleolos. En elooplasma cortical de estos ovocitos comien-zan a formarse las primeras plaquetas viteli-nas, mientras que en la región perinuclearproliferan los núcleos accesorios en íntimocontacto con la vesícula germinal. En las ova-riolas de estas hembras, aumenta el espesorde la lámina basal del epitelio folicular querodea a cada ovocito del vitelario y por lotanto comienza a formarse la túnica propia.Bonhag (1958), Chapman (1969) y Sahai yBanerjee (1987) sostienen que en insectos engeneral y en particular en Poecilocerus pictus,la túnica propia se extiende a lo largo de todala ovariola (tanto en la región del germariocomo del vitelario) y por lo tanto tiene undoble origen, a partir de las células epitelia-les que forman la vaina externa y de las célu-las foliculares. En B. punctulatus, la túnicapropia está presente solamente en la regióndel vitelario, por lo que sería sintetizada solopor las células foliculares y como lo mencio-nan estos autores, en esta región de la ovario-la, la vaina externa y la túnica propia cum-plen una importante función en el transportede nutrientes desde el cuerpo graso a travésdel epitelio folicular al ovocito en desarrollopara la síntesis de vitelo. Asimismo, la pre-sencia de fibras colágenas en la cara internade la túnica propia observada en la regiónproximal del vitelario, corroboraría que estalámina basal cumple una función de sostén,mientras que la presencia de fibras elásticasen la capa externa indicaría que la túnicapropia también cumple una importante fun-ción en el transporte del ovocito a través delvitelario.
A los 5 días de vida adulta se produce unimportante aumento en la cantidad de cuer-po graso que rodea a cada ovario y cambiala morfología de las células adiposas querodean la región proximal de las ovariolas,lo cual como mencionan Nieto Nafria yMier Durante (1985) está directamente rela-cionado con la iniciación del proceso de vi-telogénesis en los ovocitos ubicados en el
folículo basal de esta región, ya que en estaespecie como en los restantes insectos, elcuerpo graso actúa como un órgano de reser-va de glucógeno, proteínas y lípidos para serutilizados en periodos de gran actividadmetabólica.
En Baeacris punctulatus, al igual que enAnacridium aegyptium (Colombo, 1953), aliniciarse el proceso de vitelogénesis se iniciael tercer período de crecimiento del ovocito,el cual culminará cuando la célula comple-tamente cargada de vitelo haya alcanzado sutamaño definitivo y esté en condiciones deser ovulada. En hembras de 5 días, los ovoci-tos ubicados en el folículo basal de cadaovariola se encuentran en el estadio vitelogé-nico I y están rodeados por un epitelio foli-cular simple constituido por células cilíndri-cas altas en activa división y una gruesa lá-mina basal que le confiere mayor espesor ala túnica propia en esta región del vitelario.Los ovocitos en el estadio vitelogénico I seidentifican fácilmente por su tamaño y for-ma y por su citoplasma granular acidófilocon numerosas plaquetas vitelinas en la re-gión cortical. La vesícula germinal en estosovocitos está ubicada en el extremo micropi-lar o posterior de la célula, con la cromatinacompletamente descondensada, con numero-sos micronucleolos y una envoltura nuclearclaramente visible. Estas observaciones coin-ciden con las de Büning (1994), quien sostie-ne que el desplazamiento de la vesícula ger-minal hacia su posición definitiva en el hue-vo, es el principal indicio de la iniciación delproceso de vitelogénesis. Del mismo modo,la presencia de una delgada matriz extrace-lular acidófila entre el oolema y la membra-na plasmática del epitelio folicular, indica-ría que en el primer estadio de la vitelogéne-sis de B. punctulatus comienza a sintetizarsela envoltura vitelina, la cual como en losrestantes insectos, formará parte de la cu-bierta externa del ovocito en el momento deser ovulado (Albert et al., 2004).
A los 10 días de imago de la hembra deBaeacris punctulatus, el ovocito primario delfolículo basal de cada ovariola ha alcanza-do el segundo estadio de vitelogénesis. Esteestadio incluye dos etapas sucesivas, las cua-

A. A. Michel: Análisis histológico del ovario en Baeacris punctulatus52
les se diferencian por el tamaño del ovocito,las características de la vesícula germinal(tanto de la cromatina como de la envolturanuclear), la cantidad de plaquetas vitelinasacumuladas en el citoplasma y en el estadofuncional de las células foliculares que lorodean. Lusis (1963) analiza el proceso devitelogénesis en Schistocerca gregaria y lodivide en cuatro estadios, de los cuales elsegundo correspondería solo al estadio vite-logenico IIa de B. punctulatus, ya que esteautor no observa ovocitos en la etapa previaa la ruptura de la vesícula germinal, la cualcoincidimos con Büning (1994) y Albert etal. (2004), indica la finalización del procesode vitelogénesis y por lo tanto del estadio dediplonema de la profase I de la meiosis.
La segunda etapa del estadio vitelogénicoII de Baeacris punctulatus fue también obser-vada por Sahai y Banerjee (1987) en Poecilo-cerus pictus. En el segundo estadio de vitelo-génesis de B. punctulatus, las células folicu-lares adquieren forma de clava, con un nú-cleo grande en posición basal e intensa baso-filia citoplasmática en la región apical, des-tacándose en el epitelio folicular de los ovo-citos en el estadio vitelogenico IIb, el grandesarrollo del aparato de Golgi en la regiónsupranuclear de sus células, lo que indicaríasu intensa actividad secretora (Bonhag,1958; Albert et al., 2004). Las característi-cas citológicas del epitelio folicular en elsegundo estadio de vitelogénesis de Baeacrispunctulatus, coinciden con las observadas enetapas previas a la síntesis del corion de Poe-cilocerus pictus. Sin embargo, en B. punctu-latus, no se han observado células folicula-res binucleadas como las mencionadas porChapman (1969) en otros insectos, y Sahai yBanerjee (1987) y Saxena et al. (1988) en P.pictus. Las observaciones realizadas en losrestantes órganos reproductores de estashembras, en las cuales aún no se ha comple-tado el desarrollo del ovocito en el ovario,mostraron que sus espermatecas estaban car-gadas de abundante fluido seminal con es-permatozoides, por lo que coincidimos conGillot (1980) en que las hembras adultas deAcrididae pueden ser copuladas antes de al-canzar su madurez sexual.
Entre los 12 y 14 días de vida adulta dela hembra de Baeacris punctulatus, los ovo-citos primarios que llegan al folículo basalde cada ovariola se encuentran en el últimoestadio de vitelogénesis. Durante este esta-dio, los ovocitos pasarán por dos etapas su-cesivas: la primera, considerada como esta-dio vitelogenico IIIa, en la cual se completala síntesis de vitelo y de la envoltura viteli-na, y el ovocito alcanza su tamaño definitivofinalizando así el tercer período de creci-miento, y la segunda etapa, consideradacomo estadio vitelogénico IIIb, en la cual seproduce la síntesis del corion a partir de lascélulas foliculares. Al finalizar el periodo devitelogénesis, en B. punctulatus como en losrestantes insectos, los ovocitos primarios es-tarán en condiciones de iniciar el proceso demaduración ovocitaria, es decir, la reanuda-ción de la meiosis por un aumento local dela ecdisona en el ovocito, lo que le permitirácompletar su desarrollo dentro del ovario ytransformarse en un ovocito maduro en con-diciones de ser ovulado (Büning, 1994; Vi-llecco, 1998; Albert et al., 2004). Debido ala gran cantidad de vitelo acumulada en elooplasma de los ovocitos en el estadio vitelo-genico III, no se pudo determinar la caracte-rísticas del material cromosómico de estascélulas. Sin embargo, las características dela vesícula germinal en ovocitos en el esta-dio vitelogénico IIb en los cuales se observauna envoltura nuclear fragmentada en algu-nos puntos, indicaría que en los ovocitos enel estadio vitelogénico IIIb, que correspondeal final del período de vitelogénesis de B.punctulatus, ya se ha producido la rupturade la vesícula germinal e iniciado el subes-tadio de diacinesis o etapa inicial de la ma-duración ovocitaria.
Los ovocitos en el estadio vitelogénicoIIIa se identifican fácilmente porque la vesí-cula germinal no es visible y el ooplasmaestá completamente cargado de vitelo, ob-servándose en la región cortical una delgadazona de gránulos fuertemente acidófilos.Externamente al ovocito y en íntimo contactocon el oolema, se observa la envoltura viteli-na. Bonhag (1958), Lusis (1963), Sahai(1984), y Saxana et al. (1988) analizan el

Acta zoológica li l loana 52 (1–2): 30–63, 2008 53
proceso de vitelogénesis en ovocitos termi-nales de diferentes especies de Acrididae y serefieren a la síntesis de vitelo y al proceso decoriogénesis, pero no mencionan la presen-cia de gránulos acidófilos como los observa-dos en el ooplasma cortical de ovocitos en elestadio vitelogénico IIIa de Baeacris punctu-latus, los cuales podrían corresponder a losgránulos corticales presentes en muchos óvu-los incluidos los de mamíferos. Según Albertet al. (2004), los gránulos corticales, aligual que la envoltura vitelina, son sinteti-zados por el propio ovocito y cumplen unaimportante función durante la fecundación,ya que al producirse el impacto espermático,éstos liberan su contenido por exocitosis yproducen una alteración de la cubierta exter-na del ovocito evitando la polispermia. Laenvoltura vitelina, en cambio, tiene comofunción proteger al ovocito de lesiones me-cánicas y actuar como barrera especificapermitiendo solo el paso de espermatozoidesde la misma especie.
En el estadio vitelogénico IIIa de Baeacrispunctulatus, se producen importantes cam-bios a nivel del epitelio folicular, los cuales,coincidiendo con Chapman (1969) y Albertet al. (2004), están directamente relaciona-dos con el proceso de coriogénesis. En estosovocitos la mayor parte del epitelio folicularestá constituido por células cúbicas con unaintensa basofília citoplasmática, mientrasque las células ubicadas en el extremo poste-rior del ovocito, que están relacionadas conla formación del micrópilo y earópilo delhuevo, son cilíndricas con un citoplasma le-vemente basófilo y de aspecto vacuolar. Enel estadio vitelogénico IIIb, el ovocito estárodeado por un corion con dos capas fácil-mente diferenciables por su morfología y es-pesor, y como consecuencia del proceso decoriogénesis el epitelio folicular es de menorespesor y escasa actividad secretora.
El epitelio folicular en ambas etapas deltercer estadio de vitelogénesis, presenta unalámina basal de gran espesor constituida poruna gruesa capa interna de fibras de coláge-no y una delgada externa de fibras elásticas.El aumento de espesor de la lámina basaldel epitelio folicular durante el desarrollo
del ovocito en el vitelario indicaría que enBaeacris punctulatus la túnica propia, ade-más de ser una estructura mecánica quemantiene la forma de la ovariola, cumpleuna importante función en el transporte denutrientes y el acceso de hormonas durantela previtelogénesis. Por otra parte, el mayorespesor de la túnica propia en el extremoproximal del vitelario indicaría que tambiéncontribuye al desplazamiento del ovocitodentro de la ovariola y a su eliminación du-rante la ovulación, lo cual coincidiría conlos conceptos de túnica propia establecidospor Bonhag (1958), Chapman (1969) y Bü-ning (1994) para otras especies de Acrididaecon ovariolas panoísticas.
Entre los 15 y 30 días de vida adulta, to-das las hembras de Baeacris punctulatus con-tienen ovariolas cargadas de ovocitos madu-ros, observándose en algunas de estas hem-bras indicios de la primera ovulación, esdecir, el proceso que involucra la liberacióndel ovocito completamente desarrollado delfolículo basal de la ovariola, la ruptura deltapón epitelial, el ingreso al pedicelo y elpaso hacia la luz del extremo distal del ovi-ducto lateral (Chapman, 1969). En los ova-rios de estas hembras, se observó un escasonúmero de ovariolas que contenían en subase grandes vacuolas cargadas de pigmen-tos amarillos, producto de la acumulaciónde lípidos por la reabsorción de ovocitosanormales. Estos restos de folículos corres-ponden a los cuerpos de reabsorción mencio-nados para otras especies de Acrididae comoAnacridium aegyptium (Colombo, 1957),Locusta migratoria migratorioides y Schisto-cerca gregaria (Singh, 1958; Lusis, 1963;Uvarov, 1966; Chapman, 1969).
En algunas hembras de 30 días Imago deBaeacris punctulatus, el extremo proximalde la ovariola mostraba un ovocito primarioen el estadio vitelogénico III y un cuerpolúteo reducido producto de la primera ovula-ción, lo que indicaría que en estas hembras,se estaba por producir una segunda ovula-ción. La estructura histológica del cuerpolúteo observada en las hembras de esta espe-cie coincide con la descripta por Uvarov(1966), Chapman (1969), Sahai y Banerjee

A. A. Michel: Análisis histológico del ovario en Baeacris punctulatus54
(1987) y Büning (1994), para diferentes gru-pos de insectos.
Mediante las observaciones realizadas enhembras de 15 a 30 días de vida adulta deBaeacris punctulatus, se determinó que enesta especie como en el resto de los Orthop-tera (Uvarov, 1966; Chapman, 1969), todaslas ovariolas ovulan simultáneamente; sien-do Anacridium aegyptium (Colombo, 1955)la única especie de Acrididae en la cual sedeterminó que las ovariolas de un mismoovario ovulan en tiempos diferentes. La ma-duración y ovulación simultánea de los ovo-citos en todas las ovariolas de estas hembras,sumado a la escasa cantidad de ovocitosanormales que en ellas se producen cuandolas hembras son mantenidas en condicionesóptimas para el desarrollo, son indicios dela gran capacidad reproductiva de la espe-cie Baeacris punctulatus.
En esta especie como en Melanoplus di-fferentialis (Nelsen, 1934), después de laovulación, los ovocitos son almacenados enla región distal y media de los oviductos la-terales con el área micropilar dirigida haciael extremo posterior del abdomen, observán-dose en Baeacris punctulatus una modifica-ción del espesor y la coloración de las capasexternas e internas del corion de estos ovoci-tos. Según observaciones realizadas en estu-dios previos por Michel y Terán (2005), estasmodificaciones del corion son producidaspor la secreción proteica que sintetizan lascélulas epiteliales de la región distal y me-dia de estos conductos. En hembras de 15 a30 días de imago, la cantidad de ovocitosalmacenados en los oviductos laterales fuemenor al número de cuerpos lúteos observa-dos en el ovario y esto sumado al hecho deno haber encontrado ovocitos durante su tra-yecto por el oviducto medio y la cámara ge-nital, indicarían que en Baeacris punctulatusel descenso de los ovocitos maduros a travésde los oviductos, la fecundación y posterioreliminación de la cámara genital se produceen sucesivas etapas hasta completar el deso-ve durante la ovoposición.
Aunque en Baeacris punctulatus no sepudo observar la etapa de maduración ovo-citaria en la cual se produce la ovulación,
las características de la vesícula germinal deovocitos del segundo estadio de vitelogénesisnos permite inferir que en esta especie, comoen los restantes insectos (Bonhag, 1958;Chapman, 1969; Büning, 1994; Albert et al.,2004), los ovocitos son ovulados en la meta-fase de la primera división de maduración,estadio en el cual se produciría la segundadetención de la meiosis hasta su fecundaciónen la cámara genital de la hembra.
En Baeacris punctulatus, la posición delos ovocitos en el oviducto lateral después dela ovulación, indicaría que al ingresar éstosa la cámara genital de la hembra se ubica-rían dentro de la dilatación dorsal, regióndescripta en estudios previos por Michel yTerán (2005), con el micrópilo en contactodirecto con la abertura del conducto esper-matecal. En esta especie, como en otros in-sectos (Gillot, 1980; Conn, 1993; Albert etal., 2004), esta orientación del ovocito faci-litará la penetración del esperma durante lafecundación. Una vez producido el impactoespermático, se reanudará la meiosis y desdesu eliminación de la cámara genital hastacompletar la ovoposición, el ovocito setransformará en un óvulo maduro tras locual podría producirse la anfimixis. Segúnestudios previos (Michel y Teisaire, 1996), enB. punctulatus, la unión de los pronucleos seproduce después de la puesta, ya que en estaespecie el primordio embriónico es visiblerecién 48 horas después de la puesta.
Los resultados obtenidos en este trabajoindicarían que la hembra de Baeacris punc-tulatus, bajo condiciones controladas de la-boratorio, alcanza la madurez sexual entrelos 15 y 25 días del imago, período en elcual los ovocitos completan su desarrollo enel ovario y se produce la primera ovulación.Estos resultados se corresponden con las ob-servaciones de cría y con los estudios demorfogénesis, en los cuales se establece queentre los 15 y 25 días después de la mudaimaginal, se completa el desarrollo de losórganos reproductores de la hembra, hayuna intensa actividad secretora en las glán-dulas accesorias y en los oviductos laterales,y se producen las primeras ovoposiciones(Michel, 2001; Michel y Terán, 2006).

Acta zoológica li l loana 52 (1–2): 30–63, 2008 55
Según Turk y Barrera (1979), en Dichro-plus punctulatus (=Baeacris punctulatus) lamadurez sexual en ambos sexos se alcanzacon la primera cópula a los 10 días de vidaadulta. Considerando los resultados obteni-dos en este trabajo y los conceptos de madu-rez sexual establecidos por Uvarov (1966) yGillot (1980), tanto para Orthoptera comopara otros insectos, en Baeacris punctulatus,la primera cópula (que en condiciones ópti-mas para el desarrollo se produce entre las48 horas y 12 días de la muda imaginal), esun indicio de la madurez sexual solo en elmacho, ya que entre los 5 y 10 días despuésde la muda imaginal del macho, se completael proceso de morfogénesis de los órganosreproductores y la espermatogénesis en lostestículos produciéndose abundantes esper-matozoides que se liberan a los conductosdeferentes y se almacenan en las vesículasseminales. Además en este período, las glán-dulas accesorias del macho están completa-mente diferenciadas y producen abundantesecreción, que permitirá la formación delespermatóforo durante la cópula (Michel,2001; 2007).
La información aportada en este trabajocontribuye a un mejor conocimiento del de-sarrollo y la reproducción de Baeacris punc-tulatus, lo cual será de utilidad para la apli-cación de métodos de control de los Melano-plinae de importancia económica para laArgentina.
AGRADECIMIENTOS
Este trabajo se basa en parte de los resul-tados obtenidos en mi Tesis Doctoral, por loque quiero agradecer a mi director de tesis,el Dr. Héctor R. Terán. Agradezco también alConsejo de Investigación de la UniversidadNacional de Tucumán, por el apoyo efectua-do durante la realización de la misma.
LITERATURA CITADA
Albert, B., Bray, D., Lewis, J., Raff, M.,Roberts, K y Watson, J. D. 2004. Bio-logía Molecular de la Célula. Omega,S.A. Barcelona, 1387 pp.
Bentos Pereira, A. 1989. Distribución geo-gráfica de las especies del género Di-chroplus Stal (Orthoptera, Acrididae,Melanoplinae). Revista Brasilera deEntomología, 33(1): 31-47.
Bonhag, P. F. 1958. Ovarian structure andvitellogenesis in insects. Annual Reviewof Entomology, 3: 137-160.
Büning, J. 1994. The insect ovary: Ultras-tucture, previtellogenic growth and evolu-tion. Chapman y Hall, London, 400 pp.
Carbonell, C. S. y Ronderos, R. A. 1973.Las especies del grupo punctulatus delgénero Dichropus Stal (Orthoptera, Acri-didae). Revista del Museo de La Plata,11: 359-398.
Chapman, R. F. 1969. The Insects, Structu-re and Function. Elsevier, New York,819 pp.
Colombo, G. 1953. L’oogenesi negli Ortotte-ri: I. Ricerche istologiche e citometrichein Anacridium aegyptium L. dalla schiusaall’imagine. Acta Zoologica, Stockholm,34: 191-232.
Colombo, G. 1955. L’oogenesis negli Ortot-teri. II. Ricerche sull accrescimento de-gli oociti di Anacridium aegyptium L. Ar-chivio Zoologico Italiano, Napoli, 40:235-263.
Colombo, G. y Bassato, M. 1957. La diffe-renziazione delle gonadi negli embrionidella cavallette. Ricerche istologiche suembrioni di Anacridium aegyptium L. (Or-thoptera, Acridoidea). Bollettino di Zoolo-gia, 24: 275-285.
Conn, D. B. 1993. Atlas of InvertebrateReproduction and Development. Wiley-Liss, New York, 252 pp.
Cruz-Landim, C. 1994. Ultrastructural stu-dies on the spermatogenesis of Conoce-phalus saltator (Saussure). (Orthoptera,Conocephalidae): citoplasm eliminationfrom spermatid. Revista Do Centro DeCiencias Biomedicas Da UniversidadeFederal de Uberlandia, 10(1): 21-28.
Ferreira, A. V. S., Wanderley-Teixeira, V.,Santos, F. A. B., Veiga, A. F. S. L. yTeixeira, A. A. C. 2006. Histologia doaparelho reprodutor masculino e morfo-metria da população celular dos folículostesticulares de Chromacris speciosa(Thunberg, 1824) (Orthoptera: Romalei-dae) submetido a três fotoperíodos. Ar-quivos Instituto de Biología, Sao Paulo,73(3): 349-355.
Gallois, D. 1979a. Données chronologiquessur la morphogénese de l’appareil génitalfemelle de Locusta migratoria migrato-rioides (R. et F.) (Orthoptera, Acrididae).Acrida, 8(3): 117-131.

A. A. Michel: Análisis histológico del ovario en Baeacris punctulatus56
Gallois, D. 1979b. Etude chronologique del’organogénese de l’appareil génital mâlechez Locusta migratoria migratorioides(R. et F.) (Orthoptera, Acrididae). BulletinBiologique de la France et de la Belgi-que, 113: 31-51.
Gillot. C. 1980. Entomology. Plenum Press,New York (USA), 729 pp.
Hopkins, C. R. 1964. The histochemistry andfine structure of the accessory nuclei inBombus terrestris. Quarterly Journal Mi-croscopical Science, 105: 475-480.
Humason, G. L. 1979. Animal Tissue Tech-niques. W. H. Freeman and Company-San Francisco, 661 pp.
Karp, G. 1987. Biología Celular. McGraw-Hill, 950 pp.
Lange, C. E. 2002. Foro. El desarrollo deNosema locustae (Protozoa: Microspori-dia) para el control biológico de tucuras(Orthoptera: Acridoidea) y las conse-cuencias de su utilización en la Argenti-na. Revista Sociedad Entomológica Ar-gentina, 61(1-2): 1-9.
Lange, C. E. 2003. Niveles de esporulaciónexperimentales y naturales de Nosemalocustae (Microsporidia) en especies detucuras y langostas (Orthoptera: Acri-doidea) de la Argentina. Revista Socie-dad Entomológica Argentina, 62(1-2):15-22.
Liebermann, J. y Schiuma, R. 1946. Las«tucuras» más perjudiciales de nuestraagricultura y ganadería. Publicación Ins-tituto Sanidad Vegetal, año II, ser. B,7:1-62.
Lusis, O. 1963. The histology and histoche-mistry of development and resortion inthe terminal oocytes of the desert lo-cust, Schistocerca gregaria. QuarterlyJournal of Microscopical Science,104:57-68.
Michel, A. A. 2001. Morfogénesis del siste-ma reproductor y gametogénesis enBaeacris punctulatus (Thunberg) (Orthop-tera, Acrididae). Tesis Doctoral. Facultadde Ciencias Naturales, Universidad Na-cional de Tucumán, 310 pp.
Michel, A. A. 2007. Aspectos histológicossobre el desarrollo del testículo y la es-permatogénesis en Baeacris punctulatus(Orthoptera, Acrididae, Melanoplinae).Acta Zoológica Lilloana, 51(1): 67-85.
Michel, A. A. y Teisaire, E. S. 1996. Crono-logía del desarrollo embrionario normalde Baeacris punctulatus (Thunberg) (Or-thoptera, Acrididae). Revista Chilena deEntomología, 23: 29-41.
Michel, A. A. y Terán, H. R. 2005. Análisismorfológico del sistema reproductor fe-
menino de Baeacris punctulatus (Thun-berg) (Orthoptera: Acrididae: Melanopli-nae). Revista Sociedad Entomológica Ar-gentina, 64(3): 107-117.
Michel, A. A. y Terán, H. R. 2006. Cría encautiverio de Baeacris punctulatus (Thun-berg) (Orthoptera, Acrididae, Melanopli-nae). Acta Zoológica Lilloana, 50(1-2):131-134.
Nelsen, O. E. 1934. The development of theovary in the grass-hopper, Melanoplusdifferentialis (Acrididae, Orthoptera). Jo-urnal of Morphology, 55: 515-43.
Nieto Nafria, J. M. y Mier Durante, M. P.1985. Tratado de Entomología. OmegaS.A., 599 pp.
Ronderos, R. y Cigliano, M. M. 1991. TheAndean Dichoplini: cladistic analysis withdescription of Keyacris n. gen. and Pon-deracris n. gen. (Orthoptera, Acrididae,Melanoplinae). Transactions of the Ame-rican Entomological Society, 117(3-4):167-191.
Roonwal, M. L. 1937. Studies on the embr-yology of the African migratory locust,Locusta migratoria migratorioides Reicheand Frm. (Orthoptera, Acrididae). II Or-ganogeny. Philosophical transactions ofthe Royal Society of London, Serie B,227: 175-244.
Sahai, Y. N. 1984. Histochemical studies ofoocyte development in Poecilocerus pic-tus Fabr. Folia Morphologica, 32(2):160-170.
Sahai, Y. N. y Banerjee, S. 1987. Histologi-cal and Cytological studies of develop-ment of the oocyte of Poecilocerus pic-tus Fabr. (Acrididae,Orthoptera). FoliaMorphologica, 35(2): 182-193.
Santos, F. D., Veiga, A. F. S. L., Alves, L.C., Wanderley-Teixeira, V. y Teixeira A.A. C. 2006. Aspectos histológicos doaparelho reproductor femenino e morfo-metria dos ovários de Tropidacris colla-ris (Stoll, 1813) (Orthoptera: Romalei-dae) submetido a trés fotoperíodos. Ar-quivos do Instituto Biologico, Sao Paulo,73(3): 343-348.
Saxena, R. C., Jain, S., Saxena, A. y Sahai,Y. N. 1988. Towards the contributionof follicular epitelium in the growing oocy-te of Poecilocerus pictus (Orthoptera).A cytochemical study. Beitrage ZurEntomologie, 38(1): 229-231.
Singh, T. 1958. Ovulation and corpus luteumformation in Locusta migratoria migrato-rioides Reiche and Fairmaire and Schis-tocerca gregaria (Forskal). Transactionsof the Royal Entomological Society ofLondon, 110: 1-20.

Acta zoológica li l loana 52 (1–2): 30–63, 2008 57
Turk, S. y Barrera, M. 1979. Acrididos delNOA III. Estudio bio-ecológico sobre sieteespecies del género Dichroplus Stal. (Or-thoptera, Acrididae). Acta Zoológica Li-lloana, 35: 785-805.
Uvarov, B. 1966. Grassohoppers and Lo-custs. A Handbook of General Acridolo-
gy. Cambridge University Press, vol 1,481 pp.
Villeco, E. I. 1998. Ovogénesis en anfibios:Interacciones celulares. Tesis Doctoral,Facultad de Bioquímica, Química y Far-macia, Universidad Nacional de Tucu-mán, 125 pp.
Ver tablas 1 y 2 en páginas ss.

A. A. Michel: Análisis histológico del ovario en Baeacris punctulatus58
Estadiosdel desarrollo
Regiones de laovariola
Tiposcelulares
Estadios ysubestadios dela ovogénesis
Nºde células
Germario
Germario
Germario
Germario
...........................
Vitelario
Germario
...........................
Vitelario
Ovogonias...........................
Preovocitos
Ovogonias
...........................Preovocitos
...........................Ovocitos primarios“primer periodo de
crecimiento”
Ovogonias...........................
Preovocitos...........................
Ovocitos primarios“primer periodo creci-
miento”
Ovogonias
...........................Ovocitos primarios“primer periodo de
crecimiento”
...........................Ovocitos primarios
“segundo periodo decrecimiento”
Ovogonias...........................Ovocitos primarios“primer periodo de
crecimiento”
...........................Ovocitos primarios
“segundo periodo decrecimiento”
Interfase...........................
Interfase premeiótica
InterfaseProfase
Metafase...........................
Interfase premeiótica...........................
PreleptonemaLeptonemaCigonema
Interfase...........................
Interfase premeiotica...........................
PreleptonemaLeptonemaCigonema
Paquinema inicialPaquinema vanzadado
InterfaseProfase
PrometafaseMetafase
...........................Preleptonema
LeptonemaCigonema
Paquinema inicialPaquinema avanzado...........................
Postpaquinema
Interfase...........................
PreleptonemaLeptonemaCigonema
Paquinema inicialPaquinema avanzado...........................
PostpaquinemaDiplonema:
previtelogénico I
6...........................
4
10
...........................4
...........................
4 a 6
14 a 20...........................
2...........................
6 a 8
22
30
...........................
8 a 10
...........................5 a 6
30...........................
8 a 10
...........................2 a 3
4 a 8
Embrionario X
Embrionario XI
Ninfal I
Ninfal II
Ninfal III
Tabla 1 (continúa en págs ss). Regiones de la ovariola y estadios de la ovogénesis durante eldesarrollo embrionario, postembrionario y el imago de Baeacris punctulatus.

Acta zoológica li l loana 52 (1–2): 30–63, 2008 59
Estadiosdel desarrollo
Regiones de laovariola
Tiposcelulares
Estadios ysubestadios dela ovogénesis
Nºde células
Ninfal IV
Ninfal V
Imago de 24 a 72horas
Imago 5 días
Germario
...........................
Vitelario
Germario
...........................
Vitelario
Germario
..........................
Vitelario
Germario
...........................
Vitelario
Ovogonias
...........................
Ovocitos primarios“primer periodo de
crecimiento”...........................
Ovocitos primarios“segundo periodo de
crecimiento”
Ovogonias
...........................Ovocitos primarios“primer periodo de
crecimiento”...........................
Ovocito primario“segundo periodo de
crecimiento”
Ovogonias...........................Ovocitos primarios“primer periodo de
crecimiento”...........................
Ovocitos primarios“segundo periodo de
crecimiento”
Ovogonias...........................Ovocitos primarios“primer periodo de
crecimiento”...........................
Ovocitos primarios“segundo periodo de
crecimiento”
...........................Ovocitos primarios“tercer periodo de
crecimiento”
InterfaseProfase
Metafase...........................
PreleptonemaLeptonema
Paquinema inicialPaquinema avanzado...........................
PostpaquinemaDiplonema:
previtelogénico Iprevitelogénico II
InterfaseProfase
...........................PreleptonemaLeptonemaPaquinema
...........................Postpaquinema
Diplonema:previtelogenico Iprevitelogénico IIprevitelogénico III
Interfase...........................
LeptonemaPaquinema
...........................Postpaquinema
Diplonema:previtelogénico Iprevitelogénico IIprevitelogénico IIIprevitelogenico IV
Interfase...........................
LeptonemaPaquinema
...........................Postpaquinema
Diplonema:previtelogénico Iprevitelogénico IIprevitelogénico IIIprevitelogenico IV
...........................Diplonema:
vitelogénico I
35
...........................1323
...........................3 a 4
8 a 104 a 5
...........................
3 a 4...........................
3 a 4
84
3 a 4
5...........................
2 a 34 a 5
...........................3 a 4
14
4
5...........................
2 a 34 a 5
...........................3 a 4
14
4...........................
1
12 a 14
6 a 8

A. A. Michel: Análisis histológico del ovario en Baeacris punctulatus60
Imago 10 días
Imago 12 días
Imago 14 días
Germario
...........................
Vitelario
Germario
...........................
Vitelario
Germario
...........................
Vitelario
Estadiosdel desarrollo
Regiones de laovariola
Tiposcelulares
Estadios ysubestadios dela ovogénesis
Nºde células
Ovogonias...........................Ovocitos primarios“primer periodo de
crecimiento”...........................
Ovocitos primarios“segundo periodo de
crecimiento”
...........................
Ovocitos primarios“tercer periodo de
crecimiento”
Ovogonias...........................Ovocitos primarios“primer periodo de
crecimiento”...........................
Ovocitos primarios“segundo periodo de
crecimiento”
...........................
Ovocitos primarios“tercer periodo de
crecimiento”
Ovogonias...........................Ovocitos primarios“primer periodo de
crecimiento”...........................
Ovocitos primarios“segundo periodo de
crecimiento”
Ovocitos primarios“tercer periodo de
crecimiento”
Interfase...........................
LeptonemaPaquinema
...........................Postpaquinema
Diplonema:previtelogénico Iprevitelogénico IIprevitelogénico IIIprevitelogenico IV
...........................Diplonema:
vitelogénico Ivitelogénico IIavitelogénico IIb
Interfase...........................
LeptonemaPaquinema
...........................Postpaquinema
Diplonema:previtelogénico Iprevitelogénico IIprevitelogénico IIIprevitelogenico IV
...........................Diplonema:
vitelogénico Ivitelogénico IIavitelogénico IIbvitelogénico IIIa
Interfase...........................
LeptonemaPaquinema
...........................Postpaquinema
Diplonema:previtelogénico Iprevitelogénico IIprevitelogénico IIIprevitelogenico IV
...........................Diplonema:
vitelogénico Ivitelogénico IIavitelogénico IIbvitelogénico IIIb
5...........................
2 a 34 a 5
...........................3 a 4
14
4...........................
111
5...........................
2 a 34 a 5
...........................3 a 4
14
4...........................
1111
5...........................
2 a 34 a 5
...........................3 a 4
14
4...........................
1111

Acta zoológica li l loana 52 (1–2): 30–63, 2008 61
Estadiosdel desarrollo
Regiones de laovariola
Tiposcelulares
Estadios ysubestadios dela ovogénesis
Nºde células
Interfase...........................
LeptonemaPaquinema
...........................Postpaquinema
Diplonema:previtelogénico Iprevitelogénico IIprevitelogénico IIIprevitelogenico IV
...........................Diplonema:
vitelogénico Ivitelogénico IIavitelogénico IIb
“cuerpo luteo 1”
Interfase...........................
LeptonemaPaquinema
...........................Postpaquinema
Diplonema:previtelogénico Iprevitelogénico IIprevitelogénico IIIprevitelogenico IV
...........................Diplonema:
vitelogénico Ivitelogénico IIavitelogénico IIbvitelogénico IIIa
“cuerpo luteo 1”
5...........................
2 a 34 a 5
...........................3 a 4
14
4...........................
1111
5...........................
2 a 34 a 5
...........................3 a 4
14
4...........................
11111
Imago 15 a 25 días
Imago 30 días
Germario
...........................
Vitelario
Germario
...........................
Vitelario
Ovogonias.....................................
Ovocitos primarios“primer periodo de
crecimiento”.....................................
Ovocitos primarios“segundo periodo de
crecimiento”
Ovocitos primarios“tercer periodo de
crecimiento”
Ovogonias.....................................
Ovocitos primarios“primer periodo de
crecimiento”.....................................
Ovocitos primarios“segundo periodo de
crecimiento”
.....................................
Ovocitos primarios“tercer periodo de
crecimiento”
Tabla 1 (continuación y final). Regiones de la ovariola y estadios de la ovogénesis durante eldesarrollo embrionario, postembrionario y el imago de Baeacris punctulatus.

A. A. Michel: Análisis histológico del ovario en Baeacris punctulatus62
Estadiosdel ovocito
Cantidad, formay medida
Vesículagerminal
Ooplasma Folículoovárico
Granular basófilointenso a moderado,con pequeños grá-
nulos acidófilospróximos a la envol-
tura nuclear.
Granular levementebasófilo, con esca-
sos gránulos fuerte-mente acidófilos
próximos a la envol-tura nuclear.
Con finas y abun-dantes granulacio-
nes levemente basó-filas, en la regiónperinuclear peque-
ños gránulos fuerte-mente acidófilos y
1 o 2 “núcleosaccesorios”.
Con finas granula-ciones acidófilas yescasas plaquetasvitelinas en la re-gión periférica y
“núcleos accesorios”en número variablepróximos a la vesí-
cula germinal.
Con finas granula-ciones acidófilas ynumerosas plaque-
tas vitelinas solo enla periferia del
mismo. Rodeando alovocito una delgada
envoltura vitelinacon finas granula-ciones fuertemente
acidófilas.
Epitelio simpleconstituído por 4 a8 células aplana-das, con núcleos
fuertemente basófi-los y citoplasma
levemente basófilo.
Epitelio simple con10 a 26 células decaracterísticas idén-ticas al estadio I.
Epitelio simple con45 a 65 célulascúbicas, núcleos
esféricos con cro-matina condensada
en gránulos gruesosy finos, escaso
citoplasma basófilofinamente granular.
Epitelio simple, concélulas cilíndricas
bajas, núcleos esfé-ricos con cromatinaen gránulos gruesosy separados, cito-plasma granular
basófilo.
Epitelio simple concélulas cilíndricas
altas, núcleos ovoi-des en posición
basal, cromatina engránulos gruesos yseparados, citoplas-ma con finas granu-laciones basófilas,lámina basal del
epitelio fuertementeacidófila.
Previtelogénico I
Previtelogénico II
Previtelogénico III
Previtelogénico IV
Vitelogenico I
4 a 8 ovocitoscúbicos a esféricoscon un Ø mayor de
30 a 72 µm.
5 ovocitos ligera-mente ovoides conun Ø mayor de 72
a 210 µm
3 a 4 ovocitosalargados con un Ø
mayor de 222 a288 µm.
4 ovocitos alarga-dos y ligeramenteovoides con un Ømayor de 312 a
468 µm.
un ovocito alargadocon un Ø mayor de
540 a 594 µm
Ø 21 a 36 µm, decontornos lisos, en
posición central,cromatina en finashebras y abundan-tes cromómeros, 4a 8 “cuerpos protei-nosos”, 2 a 3 ma-cronucleolos y nu-merosos micronu-
cleolos.
Ø 42 a 54 µm, enposición central,envoltura nuclear
fuertemente basófi-la, mayor cantidadde cromatina en
hebras y cromóme-ros, 11 “cuerpos
proteinosos” y 2 a3 micronucleolos.
Ø 60 µm,ligeramente despla-zada hacia el extre-
mo posterior delovocito, cromatina
completamente des-espiralizada, 6 a 7“cuerpos proteino-sos” y 3 a 4 nu-cleolos granulares
fuertemente acidófi-los.
Ø 66 a 72 µm,de contornos lisos
y ubicada en elextremo posterior
del ovocito, cromati-na igual al estadioanterior, los “cuer-pos proteinosos” se
presentan comograndes acúmulos
basófilos en elcentro de la misma2 macronucleolos,
compactos y centra-les, micronucleolosnumerosos junto ala cara interna de
la envoltura.
Ø igual al anterior,de contorno liso y
ubicada en el extre-mo posterior, cro-
matina en finashebras y cromóme-
ros que cubrencompletamente eljugo nuclear y nu-merosos micronu-
cleolos fuertementeacidófilos.
Tabla 2. Características del ovocito primario en los estadios de previtelogénesis y vitelogéne-sis de Baeacris punctulatus

Acta zoológica li l loana 52 (1–2): 30–63, 2008 63
Estadiosdel ovocito
Cantidad, formay medida
Vesículagerminal
Ooplasma Folículoovárico
Vitelogénico IIa
Vitelogénico IIb
Vitelogénico IIIa
Vitelogénico IIIb
1 ovocito alargado,con un Ø mayor enel límite inferior delrango 697,82 µm a
1,5 mm
1 ovocito alargado,con un Ø mayor enel límite superiordel rango 697,82
µm a 1,5 mm.
1 ovocito alargadocon un Ø mayor de
2,640 a 3,360mm.
Igual al anterior.
Ø igual al anterior,contorno liso, croma-tina en finas hebrasy el resto en gránu-
los fuertementebasófilos que formanacúmulos en el cen-tro, 6 micronucleo-los centrales, envol-tura nuclear fuerte-
mente basófila.
Mayor cantidad decromatina condensa-da, envoltura nuclearondulada, fragmenta-da en algunos puntosy con finos gránulosen la cara interna y
externa de la misma.
No es visible.
Igual al anterior.
Con finas granula-ciones acidófilas yabundantes plaque-
tas vitelinas dediferentes diámetrosy fuertemente acidó-
filas.
Con plaquetas viteli-nas de mayor diá-metro y en mayorproporción que en
el estadio IIa.
Completamentecargado de plaque-tas vitelinas, que-dando una delgada
franja de citoplasmacon finas granula-
ciones acidófilas enla periferia del
ovocito, en la cualse ubican gránuloscorticales de dife-
rente diámetro.Envoltura vitelina
homogénea levemen-te acidófila.
Igual al anterior,pero recubiertos
externamente por undelgado corion fuer-temente acidófilo.
Epitelio simple,células cilíndricasaltas en forma de
botella, con grandesnúcleos ovoidesbasales y fuerte-mente basófilos,
cromatina en gránu-los gruesos, cito-plasma alrededor
del núcleo con finasgranulaciones basó-filas y en la regiónapical fuertementebasófilo. Lamina
basal con una capainterna de fibrascolágenas y unaexterna de fibras
elásticas.
Igual al anterior,pero con imagennegativa del golgien la región su-
pranuclear. Láminabasal igual al ante-
rior.
Epitelio simple concélulas cúbicas connúcleos esféricos
basales, cromatinaen gránulos finos y
1 o 2 nucleolosacidófilos, citoplas-ma granular fuerte-
mente basófilo;células cilíndricas
en la región micro-pilar, con núcleosesféricos basalescon cromatina en
gránulos finos, 1 o2 nucleolos y cito-
plasma vacuolarbasófilo.
Epitelio simple concélulas cúbicas
bajas, con núcleosesféricos comprimi-dos sobre la mem-brana basal, croma-tina condensada engránulos gruesos yseparados, citoplas-ma apical con finasgranulaciones basó-
filas.
Tabla 2 (continuación y final). Características del ovocito primario en los estadios de previte-logénesis y vitelogénesis de Baeacris punctulatus