5.5.5. Características generales del anabolismo celular ... · 4. La fotosíntesis fue causante...
14
5.5.5. Características generales del anabolismo celular: divergencia metabólica y necesidades energéticas. 5.5.5.1. Concepto e importancia biológica de la fotosíntesis. Fotosíntesis y evolución 5.5.5.2. Etapas de la fotosíntesis y su localización. 5.5.5.3. Quimiosíntesis. 20. Diferenciar las fases de la fotosíntesis y localizarlas intracelularmente. 21. Identificar los substratos y los productos que intervienen en las fases de la fotosíntesis y establecer el balance energético de ésta. 22. Reconocer la importancia de la fotosíntesis en la evolución. 23. Reconocer que parte de la materia obtenida en los procesos biosintéticos derivados de la fotosíntesis se utiliza en las vías catabólicas. 2.5.5.5.1. Concepto e importancia biológica de la fotosíntesis La fotosíntesis es un proceso por el cual las plantas convierten la luz solar en hidratos de carbono, azúcares y almidón. Este proceso es la base sobre la que se sustenta la vida en el Planeta, ya que los organismos animales dependen de los organismos fotosintéticos. A comienzos del siglo XIX ya se sabía que las plantas usaban CO2, agua y obtenían glúcidos y oxígeno: Luz SOLAR 6 CO 2 + 6 H 2 O C 6 H 12 O 6 + 6O 2 CLOROFILA ,aunque esta reacción es correcta, no se dice nada acerca de los detalles del proceso: ¿Qué papel desempeña la luz solar? ¿Cómo se unen los carbonos? ¿De dónde procede el O2, del CO2 o del H2O? ¿En qué lugar o lugares de las células vegetales ocurren estos procesos? Una de las primeras pruebas que se realizaron para comprender estos fenómenos, fue en la que se usó oxígeno marcado radioactivamente. Usando agua con O 18 se cultivaron plantas; se pudo comprobar que el oxígeno producido era O 18 . Se hizo la misma experiencia usando CO2 marcado radioactivamente y con agua normal; el resultado fue que el oxígeno producido no estaba marcado radioactivamente. Por lo tanto el oxígeno que producía la planta se obtenía del agua. La ecuación revisada en la actualidad es la siguiente: Luz SOLAR 6 CO 2 + 12 H 2 O C 6 H 12 O 6 + 6O 2 + 6 H 2 O CLOROFILA De esta manera se puede entender de dónde sale todo el oxígeno en la fotosíntesis; el agua de la izquierda actúa como reactivo y el agua de la derecha es metabólica.
Transcript of 5.5.5. Características generales del anabolismo celular ... · 4. La fotosíntesis fue causante...

5.5.5. Características generales del anabolismo celular: divergencia metabólica y necesidades energéticas. 5.5.5.1. Concepto e importancia biológica de la fotosíntesis. Fotosíntesis y evolución 5.5.5.2. Etapas de la fotosíntesis y su localización. 5.5.5.3. Quimiosíntesis. 20. Diferenciar las fases de la fotosíntesis y localizarlas intracelularmente. 21. Identificar los substratos y los productos que intervienen en las fases de la fotosíntesis y establecer el balance energético de ésta. 22. Reconocer la importancia de la fotosíntesis en la evolución. 23. Reconocer que parte de la materia obtenida en los procesos biosintéticos derivados de la fotosíntesis se utiliza en las vías catabólicas.
2.5.5.5.1. Concepto e importancia biológica de la fotosíntesis La fotosíntesis es un proceso por el cual las plantas convierten la luz solar en hidratos de carbono, azúcares y almidón. Este proceso es la base sobre la que se sustenta la vida en el Planeta, ya que los organismos animales dependen de los organismos fotosintéticos. A comienzos del siglo XIX ya se sabía que las plantas usaban CO2, agua y obtenían glúcidos y oxígeno:
Luz SOLAR
6 CO2 + 6 H2O C6H12O6 + 6O2
CLOROFILA
,aunque esta reacción es correcta, no se dice nada acerca de los detalles del proceso:
¿Qué papel desempeña la luz solar?
¿Cómo se unen los carbonos?
¿De dónde procede el O2, del CO2 o del H2O?
¿En qué lugar o lugares de las células vegetales ocurren estos procesos? Una de las primeras pruebas que se realizaron para comprender estos fenómenos, fue en la que se usó oxígeno marcado radioactivamente. Usando agua con O18 se cultivaron plantas; se pudo comprobar que el oxígeno producido era O18. Se hizo la misma experiencia usando CO2 marcado radioactivamente y con agua normal; el resultado fue que el oxígeno producido no estaba marcado radioactivamente. Por lo tanto el oxígeno que producía la planta se obtenía del agua. La ecuación revisada en la actualidad es la siguiente:
Luz SOLAR
6 CO2 + 12 H2O C6H12O6 + 6O2 + 6 H2O
CLOROFILA
De esta manera se puede entender de dónde sale todo el oxígeno en la fotosíntesis; el agua de la izquierda actúa como reactivo y el agua de la derecha es metabólica.

La fotosíntesis es seguramente el proceso bioquímico más importante de la Biosfera por varios motivos:
1. La síntesis de materia orgánica a partir de la inorgánica se realiza fundamentalmente mediante la fotosíntesis; luego irá pasando de unos seres vivos a otros mediante las cadenas tróficas, para ser transformada en materia propia por los diferentes seres vivos.
2. Produce la transformación de la energía luminosa en energía química, necesaria y utilizada por los seres vivos
3. En la fotosíntesis se libera oxígeno, que será utilizado en la respiración aerobia como oxidante.
4. La fotosíntesis fue causante del cambio producido en la atmósfera primitiva, que era anaerobia y reductora.
5. De la fotosíntesis depende también la energía almacenada en combustibles fósiles como carbón, petróleo y gas natural.
6. El equilibrio necesario entre seres autótrofos y heterótrofos no sería posible sin la fotosíntesis.
Se puede concluir que la diversidad de la vida existente en la Tierra depende principalmente de la fotosíntesis.
Antes de comenzar dos palabras acerca de los pigmentos fotosintéticos y los fotosistemas:
La energía luminosa no la pueden usar los seres animales; por lo tanto, son los pigmentos de los seres vegetales los que hacen dicha función. El proceso de conversión de dicha energía se realiza en los cloroplastos, concretamente en los pigmentos fotosintéticos.
PIGMENTOS FOTOSINTÉTICOS:
CLOROFILAS (a o b), XANTOFILAS Y CAROTENOS.
En las moléculas de clorofila se distinguen un anillo porfirínico que contienen magnesio (capta la luz) y una larga cadena isoprenoide denominada fitol (con la que se unen a la membrana de los tilacoides). Gracias a estas cadenas fitólicas, las moléculas de clorofila están ordenadas y asociadas a las membranas del tilacoide. Los dobles enlaces del anillo

de porfirina se alteran cuando recibe la luz solar y los electrones saltan a orbitales más alejados del núcleo atómico. Podríamos imaginar que el “spin” electrónico es mayor cuando recibe la energía solar y esto le permite “saltar” a otros orbitales. En ese estado, los electrones pueden ceder energía cuando regresan a su orbital original. Esta energía cedida es puramente química y no luminosa, aunque si aislamos los cloroplastos en un tubo de ensayo, podría observarse algún fenómeno de fluorescencia cuando los electrones vuelven a sus orbitales. Si las moléculas de clorofila están correctamente ordenadas por sus “colas” fitólicas, la energía no se emite en forma de luz sino que se concentrará en los centros primarios de reacción (CPR)
La clorofila a es el pigmento implicado en la conversión de la energía luminosa en energía química. La clorofila a y b absorben las longitudes de onda correspondientes a la luz ultravioleta, azul, naranja, y roja; los carotenoides absorben las longitudes de onda violeta, azul y verde. Las xantofilas evitan la fotooxidación de la clorofila.
Cuando la luz solar llega a las hojas de los vegetales, la luz se descompone en diversas longitudes de onda que los distintos pigmentos absorben. El efecto conjunto de la excitación de diversos tipos de pigmentos hace más eficaz la captación de la energía solar.

Como se observa en este espectro de absorción, existen longitudes de onda muy eficaces para obtener una respuesta fisiológica.
FOTOSISTEMAS: Son unidades estructurales y funcionales de la membrana de los tilacoides; en ellas se produce la captación de la energía luminosa. Están formados por un complejo clorofílico denominado “antena” (unido a proteínas de membrana) y un centro reactivo o centro primario de reacción (CPR), situado en una proteína transmembrana. La misión de este CPR es transferir electrones hacia la cadena trasportadora de la membrana tilacoidal. Cuando la energía solar excita a las moléculas de la antena, entran en resonancia con otras próximas, de tal manera que toda la energía se transfiere al CPR. Desde aquí, un electrón de alta energía salta a la cadena de transportadores. El hueco dejado se rellena con otro electrón de baja energía cedido por un donador de electrones, (agua u otras moléculas). Después de las experiencias de Emerson & Arnold (1932), (estudiando el rendimiento fotosintético de plantas sometidas a un solo tipo de longitud de onda), se pudo comprobar que existen en las plantas dos tipos de fotosistemas.
EL FOTOSISTEMA I y EL FOTOSISTEMA II, (PSI y PS II) PSI: Se observan en las membranas de los tilacoides que no están apilados, no tienen contacto con el estroma. En su centro de reacción presenta dos moléculas de clorofila que absorben a 700nm (P700), (absorbe en el rojo largo, RL)

PSII: Están en las granas de los tiliacoides. Presenta dos moléculas de clorofila que absorben a 680nm (P680), (absorbe en el rojo corto, RC). También posee el denominado complejo reductor de O2 que es una manganoproteína capaz de romper las moléculas de agua, liberando oxígeno y usando los electrones del enlace para paliar el déficit electrónico cuando se produce el electrón de alta energía en el CPR.
FASE LUMÍNICA DE LA FOTOSÍNTESIS Ocurre en la membrana de los tilacoides, donde están las moléculas de clorofila agrupadas formando los fotosistemas. Transporte de electrones: Los dos fotosistemas son iluminados a la vez, esto quiere decir que dos electrones de alta energía salen de los respectivos CPR y son aceptados por otras moléculas. Debido al distinto potencial de reducción los PSI y PSII, están ordenados espacialmente; esquemáticamente se ha hecho popular el denominado esquema en Z, donde se puede

apreciar la relación existente entre ambos fotosistemas y comprender el fenómeno de las fotofosforilaciones. El flujo de electrones va desde el PSII activado hasta el PSI sin activar, y desde éste hasta la NADP+
FOTOFOSFORILACIÓN NO CÍCLICA o acíclica: Como puede comprobarse en el esquema anterior, el flujo electrónico va desde el PSII hasta la nicotidamina pasando por una serie de transportadores. Se obtiene O2, ATP (durante el transporte por la plastoquinona y plastocianina) y nicotidamina reducida (NADPH) cuando le llegan electrones procedentes de la ferredoxina. Como en el transporte en las mitocondrias, las moléculas encargadas son complejos proteicos ordenados por su potencial de reducción. Para mantener este circuito se necesita continuamente un dador de electrones en el PSII (el agua) y un aceptor final de los mismos (la nicotidamina). Ambos fotosistemas funcionan sincrónicamente. FOTOFOSFORILACIÓN CÍCLICA: El proceso se inicia con la absorción de luz por parte del PSI, éste cede los electrones a la ferredoxina, después los electrones irán al citocromo b6f y de éste a la plastocianina. Desde aquí los electrones son cedidos de nuevo al PSI, ocupando el hueco que dejó la clorofila cuando fue iluminada con la longitud de onda adecuada. Obviamente no se obtiene oxígeno, sí hay formación de ATP y no se obtiene nicotidamina reducida.


FASE OSCURA DE LA FOTOSÍNTESIS En esta fase se usará el ATP y la NADPH obtenida en la fase anterior para reducir el carbono y sintetizar glúcidos. Las reacciones necesarias se hacen independientemente de la luz solar, por lo tanto se hace tanto de día como de noche. La reducción del carbono se hace en el estroma del cloroplasto y gracias a una serie de reacciones cíclicas que en conjunto se denomina ciclo de Calvin-Benson
El CO2 proveniente de la atmósfera llega al estroma del cloroplasto a través de los estomas del envés de las hojas, después difunde al tejido parenquimático y posteriormente llega a las células. Desde aquí difunde hacia el estroma del cloroplasto.

1. La molécula inicial del ciclo es la ribulosa 1-5 bifosfato (RuBP), ésta se une al CO2, formando un compuesto de seis átomos de carbono que se escinde rápidamente en dos moléculas de ácido 3-fosfoglicérico (3PG) de tres átomos de carbono (plantas C3). Para que todo eso pueda ocurrir se necesita el concurso de la ribulosa bifosfato carboxilasa oxigenasa, también conocida como enzima rubisco.
2. Reducción del PGA a gliceraldehído 3-fosfato (G3P), usando NADPH y ATP de la etapa anterior. Por cada tres moléculas de CO2 fijado se producen seis de G3P. Sólo una de ellas se usará en la síntesis de glúcidos en el citosol.
3. Las cinco moléculas restantes de G3P se convertirán en tres de RuBP, que servirán de aceptoras de nuevas moléculas de CO2. Esto requiere la hidrólisis de tres moléculas de ATP.
Balance energético Esta obtención de glúcidos es muy costosa, energéticamente hablando. La fijación de tres CO2 con producción de de una de G3P, requiere el gasto de 9 ATP y seis de NADPH, Por lo tanto son necesarias dos vueltas de ciclo para producir un mol de glucosa. La ecuación global sería la siguiente: 6CO2 + 12 NADPH + 12 H+ +18 ATP hexosa + 12 NADP+ + 18 ADP + 18 Pi
La fotorrespiración y el ciclo de Calvin-Benson en plantas C4
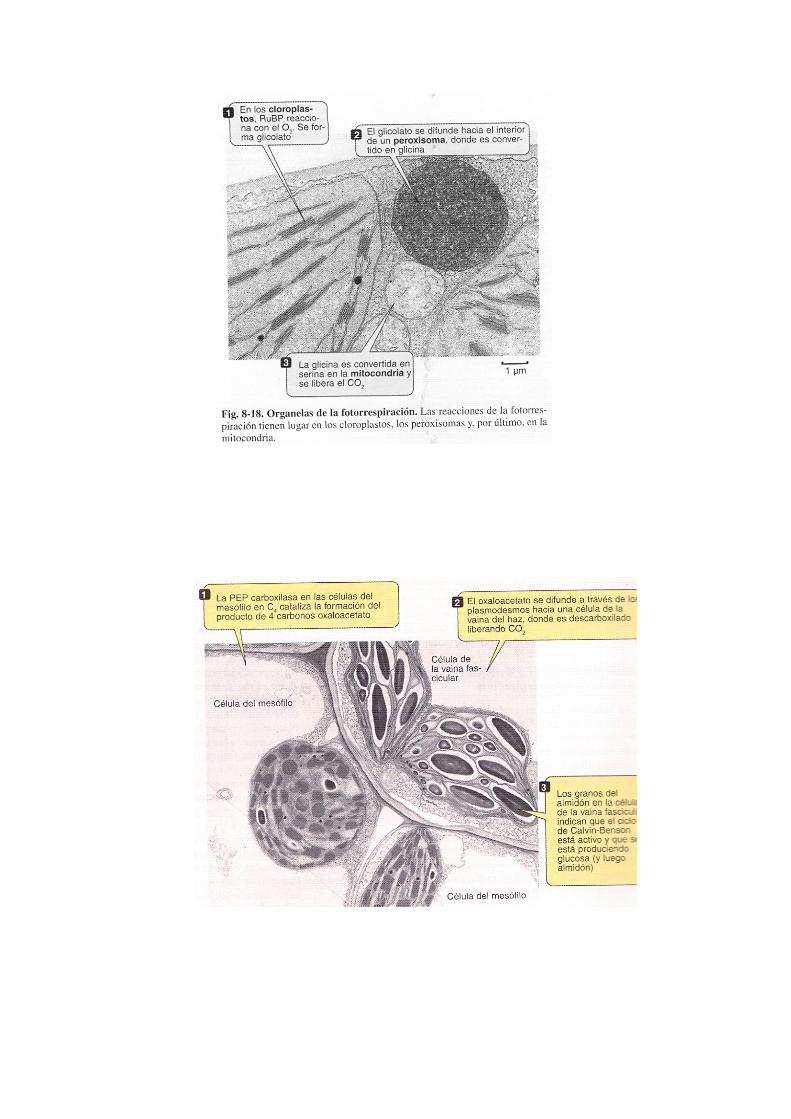
La enzima RUBISCO, es capaz de trabajar como oxigenasa en vez de carboxilasa, de esta forma cataliza la unión del O2 a la RuBP. De esta forma comienza la fotorrespiración, que es un proceso dependiente de la luz, consume O2 y rinde CO2. Este proceso implica a diversos orgánulos: cloroplasto, peroxisoma y mitocondria.
Cuando el O2 se une a la RuBP se forma ácido glicólico que difunde hacia el peroxisoma; aquí se forma H2O2 usando oxígeno atmosférico y el propio ácido glicólico, obteniendo glicina. Posteriormente la glicina se incorpora al ciclo de Krebs en la mitocondria, rindiendo CO2 Este proceso no tiene ninguna función metabólica conocida, aunque resulta evidente que provoca una pérdida de la eficacia fotosintética puesto que al aumentar la concentración de oxígeno, la enzima rubisco se comporta como oxigenasa haciendo esta ruta del ácido glioxílico. Parece ser que la fotorrespiración podría proteger la planta de la fotooxidación de las membranas de los tilacoides; de tal manera que, cuando hay una baja concentración de CO2 o aumento de la concentración de O2, no se realiza el ciclo de
Calvin-Benson.

Plantas C4 (solución evolutiva a la fotorrespiración) Algunas plantas son capaces de concentrar CO2 en sus células fotosintéticas, son las plantas C4. Este tipo de vegetales cuando captan el CO2 forman compuestos de cuatro átomos de carbono (C4) y además poseen una estructura anatómica denominada Kranz (las células del mesófilo rodean a las células del tejido perivascular) .
Las células del mesófilo están adaptadas a concentrar CO2 hacia las células del tejido perivascular que es donde se realiza el ciclo de Calvin-Benson. Las plantas C4 crecen rápidamente y son típicas de ambientes cálidos y húmedos. (Maíz, Trigo, Tabaco)
RUTA DE HATCH-SLACK-KORCHAK (HSK) En este ciclo se observa como el CO2 es fijado de manera provisional en las células del mesófilo, posteriormente va al tejido perivascular que lo usará para el ciclo de Calvin-Benson. El CO2 atmosférico lo capta ácido fosfoenolpirúvico (PEP) gracias al enzima fosfoenolpiruvato carboxilasa, formando el oxalacelato. Mediante el enzima malato deshidrogenasa y gastando NADPH2, se forma malato que llega al tejido perivascular. Posteriormente, este ácido málico se descarboxila y el CO2 obtenido se incorpora al ciclo de Calvin-Benson. Así los azúcares formados van a parar directamente al floema por el que circula la savia elaborada. El ácido málico descarboxilado se transforma en ácido pirúvico y consumiendo ATP de la primera etapa de la fotosíntesis llega a formar ácido fosfoenol-pirúvico, completando el ciclo.
FLOEMA
Malato

CICLO C.A.M (Metabolismo ácido de las crasuláceas): Estas plantas tienen dos carboxilaciones separadas temporalmente: Fijación nocturna de CO2. Esta primera fase se da en la noche (ciclo C4), cuando tienen los estomas abiertos. A través de ellos la planta capta CO2 atmosférico y la fosfoenolpiruvato carboxilasa lo incorpora por carboxilación al fosfoenolpiruvato (PEP) que se transforma en oxalacetato (OAA) con el desprendimiento de un grupo fosfato; el oxaloacetato formado de la prefijación de CO2 es reducido en el citosol a malato mediante la NAD-malato deshidrogenasa, el malato es bombeado con gasto de energía a las vacuolas, donde se va acumulando como ácido málico y es almacenado, provocando que el contenido vacuolar sea muy ácido (cerca de pH 3) durante la noche.
Fijación diurna de CO2: Con la salida del sol, los estomas se cierran previniendo la pérdida de agua e impidiendo la adquisición de CO2. El ácido málico sale de la vacuola y se descarboxila liberando el CO2 y ácido pirúvico el cual es devuelto al ciclo tras ser fosforilazdo con ATP, produciendo nuevamente fosfoenolpiruvato. Ya que los estomas están cerrados, el CO2 liberado internamente no puede escapar de la hoja y en lugar de esto es reducido a carbohidrato por la operación del ciclo de Calvin.
Este mecanismo de concentración de dióxido de carbono permite disminuir la probabilidad de que entre un O2 en el sitio activo de la RuBisCO por lo que la eficiencia fotosintética es mayor. Las plantas CAM suelen ser crasas (no todas) o plantas que viven en un régimen hídrico escaso (orquídeas epifitas)

FACTORES QUE INFLUYEN EN LA FOTOSÍNTESIS
El rendimiento fotosintético se puede medir en función de la concentración de CO2
asimilado por unidad de tiempo, o bien, por la cantidad de O2 desprendido por unidad de tiempo.
1) Concentración de CO2 ambiental: Al aumentar la [CO2], aumenta el rendimiento fotosintético hasta alcanzar un valor máximo.
2) Concentración de O2 ambiental: El aumento de la concentración de oxígeno disminuye el rendimiento fotosintético ya que la enzima Rubisco actuaría como oxigenasa en vez de carboxilasa.
3) Humedad: Cuando la humedad disminuye, las células guarda de los estomas se
cierran y entonces no puede entrar el CO2, con lo cual el rendimiento fotosintético disminuye.
4) Temperatura: Como cualquier actividad enzimática, el aumento de la temperatura, aumenta la probabilidad de choque entre las moléculas. Cuando se desnaturalizan las proteínas se pierde dicha actividad.

5) Intensidad luminosa: Depende de los pigmentos que estén presentes en la planta. Existen adaptaciones a intensidades luminosas bajas y altas (plantas de sol y de sombra)
6) Tipo de luz: El rendimiento óptimo se realiza con luz roja o azul. Si la luz aumenta por encima de 680nm no interviene el PSII y por lo tanto disminuye el rendimiento fotosintético; no se realiza el ciclo de Calvin.
QUIMIOSÍNTESIS Es una actividad propia de las bacterias cuya nutrición no depende de la luz solar, sino de la energía química que se desprende de una oxidación del propio organismo sobre sustratos inorgánicos sencillos (amoniaco, nitritos, sulfuros, ión ferroso, hidrógeno, etc.) Como fuente de carbono pueden usar CO2 del medio.
Bacterias quimiosintéticas de nitrógeno: Se encuentran en el suelo y en el agua. Las Nitrosomonas y Nitrocysstis oxidan el amoniaco a nitrito y las bacterias del género Nitrobacter completan la oxidación a nitrato. Ambas bacterias se complementan, por esta razón se encuentran en el mismo ambiente.
Bacterias quimiosintéticas del azufre. Tiobacterias y bacterias sulfurosas, (quimiolitotrofas no fotosintéticas), oxidan el azufre, el ácido sulfídrico o el tiosultato. Como resultado de esta actividad se sintetiza ácido sulfúrico. Para equilibrar el pH de los suelos excesivamente calcáreos se echa azufre en ellos, las bacterias hacen el resto ya que al acidificar los suelos se convierte el carbonato de calcio en yeso que es soluble en agua.
Bacterias quimiosintéticas del hierro. Oxidan el hierro ferroso a férrico (Thiobacillus ferrooxidans) típico de las aguas de lavados de minerales de hierro.
Bacterias quimiosintéticas del hidrógeno. Usan el H2 como fuente de energía. Dentro de este grupo se encuentran especies que pueden utilizar compuestos orgánicos como fuente de carbono, además de CO2, Hydrogenomonas (autótrofos facultativos)