5. Citoesqueleto Enero 2013
81
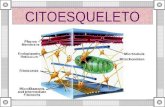
Citoesqueleto • La capacidad de las células eucarióticas de adoptar una gran variedad de formas y llevar a cabo movimientos direccionales y coordinados depende de una red muy compleja de filamentos proteicos que se extienden a través del citoplasma. Este recibe el nombre de citoesqueleto, aunque a diferencia del esqueleto óseo, es una estructura sumamente dinámica que se reorganiza continuamente mientras la célula cambia de forma y se divide y responde a su entorno.
-
Upload
lalo-esparza-huerta -
Category
Documents
-
view
57 -
download
0
Transcript of 5. Citoesqueleto Enero 2013

Citoesqueleto • La capacidad de las células eucarióticas de
adoptar una gran variedad de formas y llevar a cabo movimientos direccionales y coordinados depende de una red muy compleja de filamentos proteicos que se extienden a través del citoplasma. Este recibe el nombre de citoesqueleto, aunque a diferencia del esqueleto óseo, es una estructura sumamente dinámica que se reorganiza continuamente mientras la célula cambia de forma y se divide y responde a su entorno.

• (A) filamentos intermedios de queratina teñidos de verde mediante inmuno fluorescencia.
• (B) dibujo de una microfotografía donde se observan los haces de filamentos intermedios

Célula en cultivo fijada y marcada con el azul de Coomasie, colorante específico de proteínas. Podemos observar la gran variedad de estructuras filamentosas que se extienden a través de la célula.

Citoesqueleto • El citoesqueleto es el responsable directo de
movimientos tales como el deslizamiento de las células sobre el sustrato, la contracción muscular, y todos los cambios de forma que ocurren durante el desarrollo embrionario de los vertebrados; también proporciona la maquinaria para los movimientos intracelulares, tales como el transporte de los orgánulos desde un lugar a otro en el citoplasma y la segregación de los cromosomas durante la mitosis.

• Las diversas actividades del citoesqueleto dependen de tres tipos de filamentos proteicos –los filamentos de actina, los microtúbulos y los filamentos intermedios. Cada tipo de filamento esta formado por una subunidad proteica distinta: actina para los filamentos de actina, tubulina para los microtúbulos, y una familia de proteínas fibrosas relacionadas, tales como vimentina o láminas, para los filamentos intermedios.
Citoesqueleto

• En cada uno de los tres principales tipos de proteínas de citoesqueleto, estas se ensamblan formando un filamento lineal, que puede ir de un lado de la célula hasta el lado opuesto. Estos filamentos conectan complejos proteicos y orgánulos de regiones distintas de la célula. Además forma el soporte mecánico. Y forma un armazón interno que mantiene todo el volumen citoplasmático.
Citoesqueleto

• Los filamentos de actina (también conocidos como micro filamentos) son polímeros helicoidales, enroscados de dos en dos, de la proteína actina. Aparecen como estructuras flexibles, con un diámetro de 5 a 9 nm, que están organizadas en una gran variedad de haces, de redes bidimensionales y de geles tridimensionales. Aunque los filamentos de actina están dispersos por el citoplasma de la célula, están altamente concentrados en el córtex, justo por debajo de la membrana plasmática.

• Los microtúbulos son cilindros huecos formados por una proteína tubulina. Su diámetro externo es de 25 nm y son mucho mas rígidos que los filamentos de actina. Los microtúbulos son largos y rectos y típicamente disponen de un extremo unidos a un centro organizador de microtúbulos (MTOC) llamado centrosoma.

• Los filamentos intermedios son estructuras parecidas a cuerdas, de un diámetro de aproximadamente de 10 nm; están formados por las proteínas de los filamentos intermedios. Uno de los tipos de filamentos intermedios forma una red llamada lámina nuclear que se localiza debajo de la membrana nuclear interna. Otros filamentos se extienden a lo largo del citoplasma proporcionando resistencia mecánica y sosteniendo la tensión mecánica de los tejidos epiteliales mediante la unión de los citoplasmas de las células vecinas a través de uniones celulares.

Filamentos intermedios• Los filamentos intermedios tienen un alto grado
de fuerza tensional y su función principal consiste
en conferir resistencia a las células contra el estrés
mecánico asociado con el estiramiento. Se les
denomina intermedio porque su diámetro esta
entre los filamentos delgados de actina y los
filamentos más gruesos de miosina.

• Forman una red que rodea al núcleo y se extiende hacia la
periferia para abarcar el total del citoplasma. En la
periferia se encuentran anclados a la membrana en el sitio
de uniones intercelulares como los desmosomas . También
se localizan dentro del núcleo; una red filamentos
intermedios denominada lámina nuclear, sustenta y
refuerza la envoltura nuclear en todos las eucariotas.
Filamentos intermedios

• Esta microfotografía revela la estructura reticular formada por los filamentos intermedios de queratina (verde), los cuales rodean al núcleo y se extienden a través del citoplasma celular. Otra proteína fue teñida de azul para mostrar la localización de los limites celulares.

• Los filamentos intermedios se asemejan a cuerdas formadas por hebras largas y retorcidas de proteínas. El monómero proteico de un filamento intermedio (A) está compuesto por un dominio bastoniforme central y una región globular en cada extremo. Los monómeros se asocian de a pares para formar dímeros (B), y luego los dímeros se alinean de dos en dos para formar un tetrámero escalonado (C). Los tetrámeros pueden unirse por sus extremos (D) y ensamblarse para dar lugar a una estructura helicoidal que tiene 8 cadenas de tetrámeros que constituyen el filamento intermedio cordoniforme (E). Microfotografía electrónica del filamento final.

• Los filamentos intermedios presentes en el citoplasma pueden clasificarse en 4 categorías: 1) filamentos de queratina en las células epiteliales; 2) vimentina y filamentos relacionados con ésta en las células del tejido conectivo, las células musculares y las células del sostén del sistema nervioso; 3) neuro- filamentos en las células nerviosas, y 4) láminas nucleares, los cuales otorgan resistencia a la membrana nuclear de las células animales.
Filamentos intermedios

Filamentos intermedios


• Los filamentos intermedios que constituyen la lámina nuclear rígida están constituidas por una clase de proteínas llamadas laminas. A diferencia de los filamentos intermedios encontrados en citoplasma sumamente estables, los de la lámina nuclear se desensamblan y se reconstituyen durante cada división nuclear, cuando la membrana nuclear se rompe durante la mitosis y se regenera en cada una de las células hijas.
Filamentos intermedios


Filamentos intermedios

Filamentos intermedios

• La enfermedad genética humana epidermolisis bullosa simple, las mutaciones en los genes de las queratinas que se expresan normalmente en la capa basal de la epidermis desorganizan la red de filamentos de cito queratina en estas células, transformándolas en células extremada- mente sensibles a lesiones mecánicas: una ligera presión puede provocar al rotura de las células basales mutantes, y la piel se llena de ampollas en la zona afectada.
Filamentos intermedios

• La proteína defectuosa se ensambla con las moléculas de queratina y desorganiza la red de filamentos de queratina en las capas celulares basales. Micrografías ópticas de piel normal (A) y mutante (B) muestran que las ampollas se producen a partir de la ruptura de las células en la capa basal de la epidermis mutante.
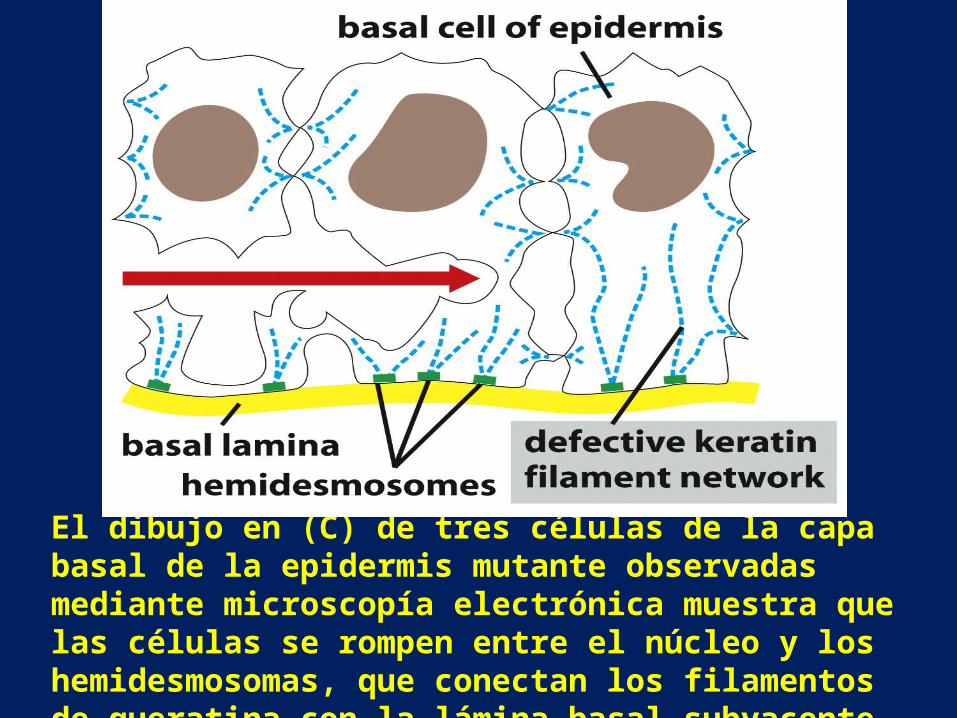
El dibujo en (C) de tres células de la capa basal de la epidermis mutante observadas mediante microscopía electrónica muestra que las células se rompen entre el núcleo y los hemidesmosomas, que conectan los filamentos de queratina con la lámina basal subyacente.

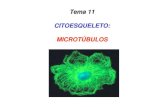
• Los microtúbulos desempeñan un papel
crucial en la organización de todas las células
eucariontes. Se trata de tubos proteicos
huecos, largos relativamente rígidos, que
tienen la capacidad de desensamblarse con
rapidez en un sitio y ensamblarse de nuevo.
Microtúbulos

Microtúbulos
• En una célula animal típica, los microtúbulos se originan en una estructura pequeña localizada cerca del centro de la célula , denominada centrosoma.
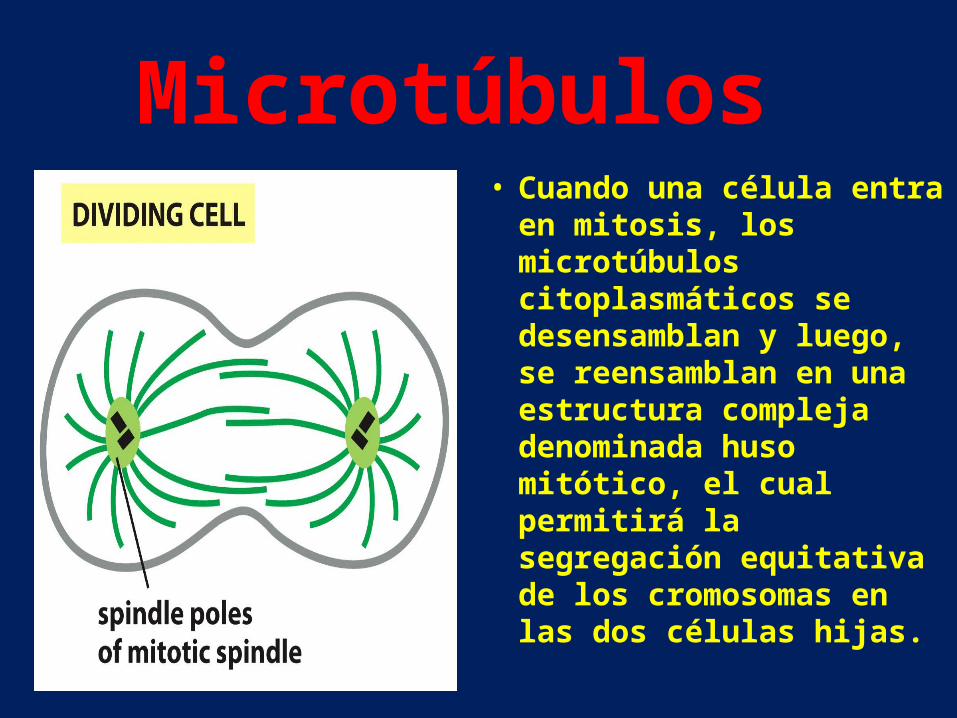
• Cuando una célula entra en mitosis, los microtúbulos citoplasmáticos se desensamblan y luego, se reensamblan en una estructura compleja denominada huso mitótico, el cual permitirá la segregación equitativa de los cromosomas en las dos células hijas.
Microtúbulos
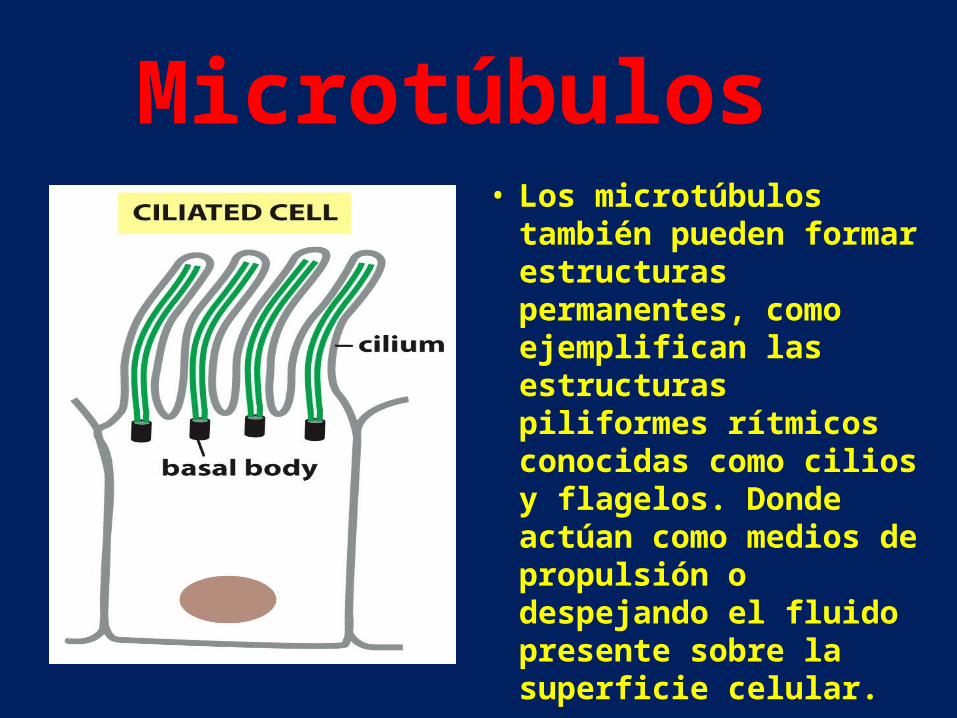
• Los microtúbulos también pueden formar estructuras permanentes, como ejemplifican las estructuras piliformes rítmicos conocidas como cilios y flagelos. Donde actúan como medios de propulsión o despejando el fluido presente sobre la superficie celular.
Microtúbulos

• Los microtúbulos están formados por subunidades de
(moléculas de tubulina), cada una de las cuales es un dímero
compuesto por dos proteínas globulares muy semejantes
denominadas α-tubulina y β-tubulina, que están firmemente
unidas entre sí con enlaces covalentes. Los dímeros de
tubulina se apilan unidas por enlaces no covalentes para
formar la pared del microtúbulo cilíndrico hueco.
Microtúbulos

Microtúbulos • (A) el esquema muestra una
molécula de tubulina (un dímero αβ) y un protofilamento, y su localización en la pared el microtúbulo. (B y C). Esquema de un microtúbulo que revela la agrupación de las moléculas de tubulina en la pared de microtúbulos. (D) corte transversal de un microtúbulo con su anillo de 13 subunidades, c/u de las cuales se corresponde un dímero de tubulina distinto. (E) vista longitudinal de un microtúbulo con un microscopio electrónico.

Microtúbulos • Los microtúbulos son estructuras polares: un
extremo (extremo más) es capaz de crecer a gran velocidad, mientras que el otro extremo (extremo menos) tiene tendencia a perder subunidades si no está estabilizado. En la mayoría de las células, el extremo menos de los microtúbulos está estabilizado mediante la unión a una estructura que recibe el nombre de centrosoma, y los extremos con crecimiento rápido están entonces libres para añadir moléculas de tubulina. El centrosoma suele estar localizado cerca del núcleo, en la zona central de la célula.

Microtúbulos

• Los centrosomas contienen cientos de
elementos anulares que se originan a partir
de otro tipo de tubulina, la ϒ-tubulina y cada
anillo de ϒ-tubulina es el punto de partida o
sitio de nucleación, del crecimiento del
microtúbulo.
Microtúbulos



• Cada filamento del microtúbulo crece o se retrae independientemente de sus vecinos. La disposición de un centrosoma se modifica continuamente a medida que crecen nuevos microtúbulos (flechas rojas) y se retaren los microtúbulos preexistentes (flechas azules).
Microtúbulos

• Los dímeros de tubulina portadores de GTP se unen más firmemente entre sí que los dímeros de tubulina de GDP. Por lo tanto, los microtúbulos que tienen dímeros de tubulina recién incorporados a su extremo con GTP unido tienden a seguir creciendo.
Microtúbulos

• Si el crecimiento del microtúbulo es lento, las subunidades presentes en este casquete de GTP hidrolizarán el GTP a GDP antes de que las nuevas subunidades de tubulina cargadas con GTP hayan tenido tiempo de unirse. Por lo tanto, se pierde el casquete de GTP; las subunidades que contiene GDP se liberan del extremo libre, de manera que el microtúbulo comienza a retraerse en forma continua.
Microtúbulos

• Un microtúbulo recién formado persistirá sólo si sus
dos extremos están protegidos contra la
despolimerización. En las células los extremos
menos de los microtúbulos suelen estar protegidos
por los centros organizadores. Los extremos más
están libres pero pueden ser estabilizados por otras
proteínas.
Microtúbulos

• (A) célula no polarizada con nuevos microtúbulos que crecen y se retraen en muchas direcciones desde un centrosoma. Algunos de estos microtúbulos se encuentran con proteínas (B) de encapuchamiento y estabilizarlos. Esta estabilización selectiva determinará una reorientación de la disposición de los microtúbulos (C) y convertirá a la célula en una forma muy polarizada.

• Mucha de las disposiciones de los microtúbulos
celulares son lábiles y esta labilidad es imprescindible
para que puedan desarrollar su función. El huso
mitótico es la diana de una gran variedad de drogas
antimitóticas específicas que actúan interfiriendo en el
recambio de tubulina de los microtúbulos y el acervo
de tubulina libre.
Microtúbulos

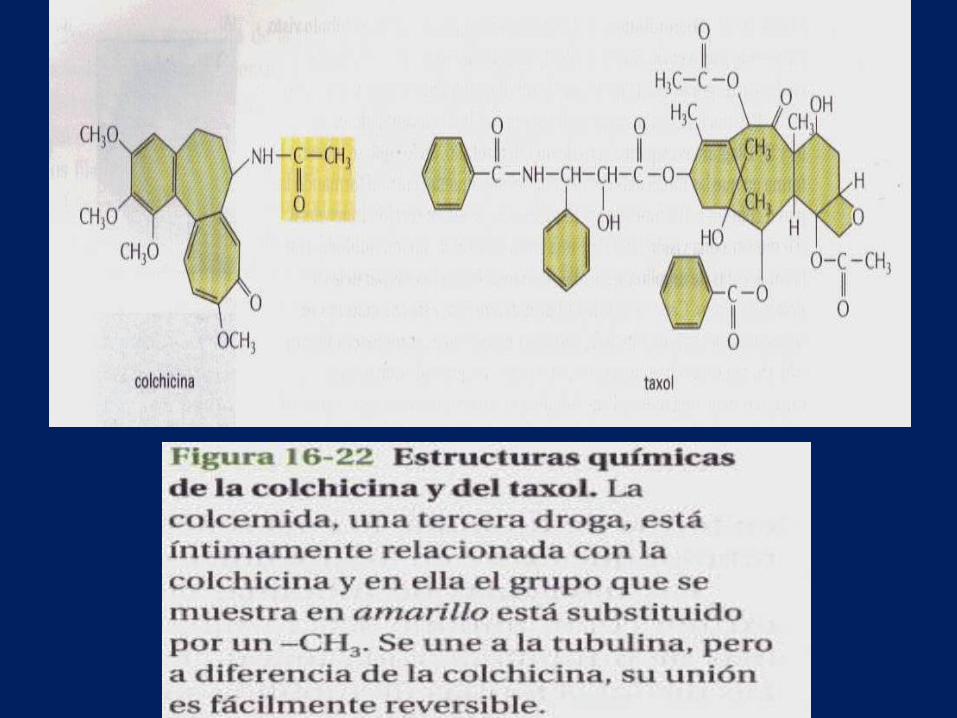
• Una de estas drogas es la colchicina, alcaloide que se
extrae del azafrán silvestre y que ha sido utilizada
como planta medicinal para el tratamiento de la
gota. Cada molécula de colchicina se une a una
molécula de tubulina e impide su polimerización,
pero no puede unirse a la tubulina una vez la
tubulina ha polimerizado formando un microtúbulo.
Microtúbulos

• La exposición de una célula en división la colchicina o
a la colcemida, droga relacionada con ella, produce
una desaparición rápida del huso mitótico, e indica
que el equilibrio químico se mantiene mediante el
recambio continuo de subunidades entre los
microtúbulos del huso mitótico y el acervo de
tubulina libre.
Microtúbulos

• El taxol que se extrae de la corteza del tejo, tiene un efecto compuesto. Se une fuertemente a los microtúbulos y los estabiliza; cuando se añade a células, provoca que muchas de las moléculas de tubulina libre se ensamblen formando microtúbulos. La estabilización de los microtúbulos con taxol detiene la mitosis de las células en división, lo cual indica que durante la mitosis los microtúbulos deben ser capaces no solo de polimerizar sino también de despolimerizar.
Microtúbulos

Fármacos que afectan los microtúbulos y los filamentos
Fármacos específicos de microtúbulos Acción
Taxol Se une y estabiliza los microtúbulos
Colchicina, colcemida Se une a las subunidades e impide la polimerización
Vinblastin, vincristin Se une a las subunidades e impide la polimerización
Fármacos específicos de actina Acción
Faloidina Se une y estabiliza los filamentos
Citocalasina Recubre los extremos más de los filamentos
Latrunculina Se una a las subunidades e impide la polimerización

• El citoplasma esta en continuo movimiento. Las
mitocondrias y otros orgánulos membranosos
cambian sus posiciones mediante movimientos
saltatorios periódicos. Estos movimientos son
provocados por proteínas motoras, las cuales se
unen a los microtúbulos o a los filamentos de
actina para moverse a lo largo de estos
filamentos.
Microtúbulos

• La primera proteína motora descubierta fue
la miosina, una proteína que se desliza a lo
largo de los filamentos de actina que es
abundante en el músculo esquelético donde
constituye la parte más importante del
aparato contráctil.
Microtúbulos

• Los movimientos son generados por
proteínas motoras , que utilizan la energía
derivada de ciclos repetidos de hidrólisis de
ATP y viajan sostenidamente a lo largo del
filamento de la actina o del microtúbulo en
una sola dirección.
Microtúbulos
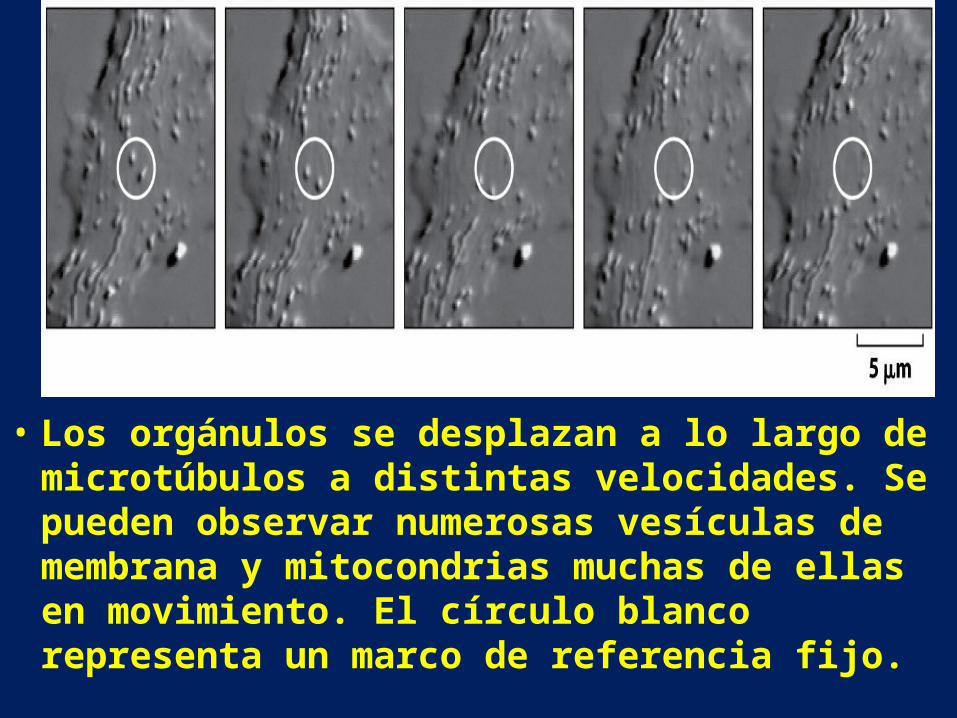
• Los orgánulos se desplazan a lo largo de microtúbulos a distintas velocidades. Se pueden observar numerosas vesículas de membrana y mitocondrias muchas de ellas en movimiento. El círculo blanco representa un marco de referencia fijo.

Microtúbulos • Las proteínas motoras que se mueven a lo
largo de los microtúbulos son distintas a las miosinas y pertenecen a dos familias: las quinesinas, que generalmente se mueven hacia el extremo más de los microtúbulos (a partir del centrosoma), y las dineínas, que se desplazan hacia el extremo menos (hacia el centrosoma). Al igual que las miosinas, cada tipo de proteína motora dependiente de microtúbulos transporta una carga determinada con la cual se desplaza.
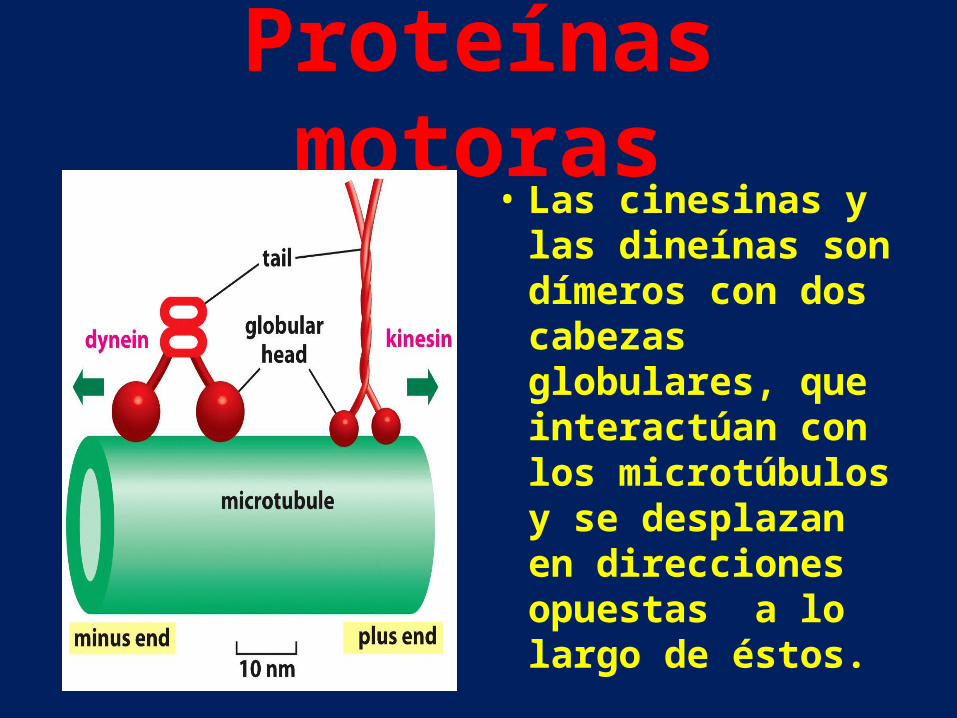
Proteínas motoras• Las cinesinas y las
dineínas son dímeros con dos cabezas globulares, que interactúan con los microtúbulos y se desplazan en direcciones opuestas a lo largo de éstos.

• Las cabezas globulares de cinesina y dineína son enzimas que hidrolizan ATP. Esta reacción aporta la energía para un ciclo de cambios conformacionales de la cabeza que le permite desplazarse a lo largo del microtúbulo mediante un ciclo de unión, separación y nueva unión.
Proteínas motoras
d
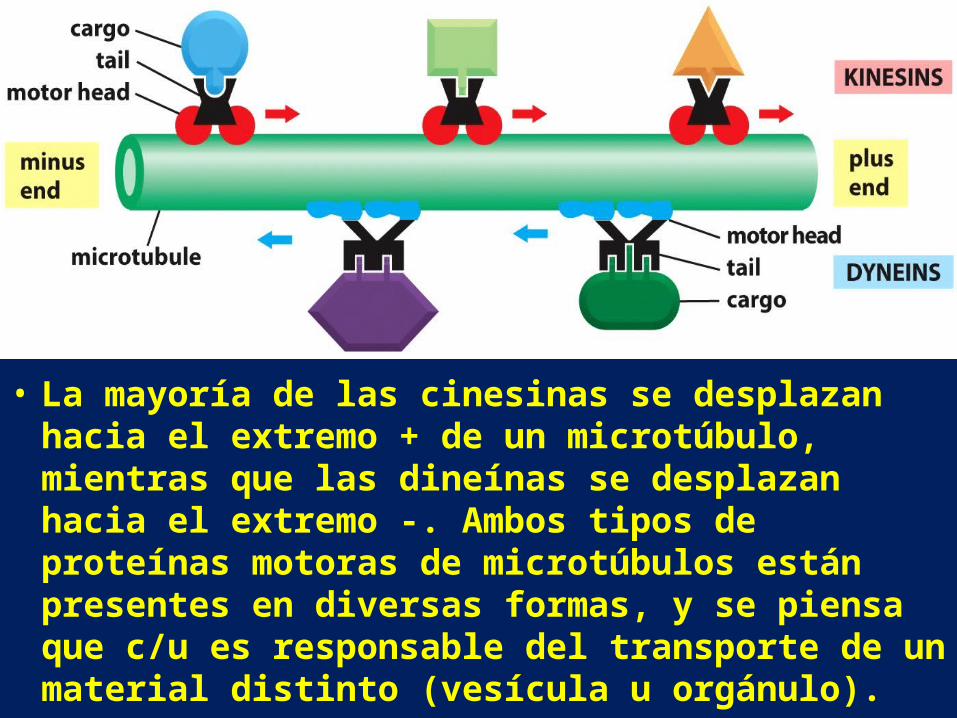
• La mayoría de las cinesinas se desplazan hacia el extremo + de un microtúbulo, mientras que las dineínas se desplazan hacia el extremo -. Ambos tipos de proteínas motoras de microtúbulos están presentes en diversas formas, y se piensa que c/u es responsable del transporte de un material distinto (vesícula u orgánulo).

• Las proteínas motoras microtúbulos-dependientes juegan un papel muy importante en el posicionamiento de los orgánulos membranoso dentro de la célula eucariota. Los túbulos membranoso del retículo endoplásmico, están alineados con los microtúbulos y se extienden casi hasta la periferia celular; el complejo de Golgi, en cambio, está localizado cerca del centrosoma
Microtúbulos


Flagelos y cilios • Los cilios y los flagelos están formados por
microtúbulos estables movidos por la dineína.
Los cilios son estructuras piliformes de 25 µm
de diámetro, cubiertas de membrana
plasmática, que parten de la superficie de
varios tipos de células eucariotas.

• Cada cilio contiene una porción central
formada por un haz de microtúbulos estables
que crecen a partir de un cuerpo basal
localizado en el citoplasma; el cuerpo basal
actúa como centro organizador del cilio.
Cilios

• Cilios semejantes a pelos revisten la superficie de numerosas células eucariotas.

• Los cilios desplazan agua sobre la superficie
de una célula o propulsan células aisladas a
través de un medio líquido. Por ejemplo:
algunos protozoos usan los cilios para recoger
partículas alimenticias, y otros los emplean
como medio de locomoción.
Cilios

• En las células epiteliales que tapizan las vías
respiratorias (más de mil millones por cm2) los
cilios barren capas de mucus que contienen
partículas de polvo atrapadas y células
muertas hacia la faringe donde son deglutidas
y finalmente eliminadas del organismo.
Cilios

Cilios

Flagelos • Los flagelos que impulsan a los espermatozoides
y a muchos protozoos presentan una estructura
interna similar a la de los cilios, pero son más
largos. Los flagelos desplazan a la célula,
propagando ondas regulares a lo largo de toda
su extensión que impulsan a las células a través
de un medio líquido.

Flagelos • Los flagelos propulsan a
la célula mediante movimientos ondulatorios repetitivos. Movimiento ondulatorio de un solo flagelo de un espermatozoide de un tunicado.

Flagelos • Los microtúbulos de cilios y flagelos revelan
nueve dobletes de microtúbulos dispuestos
en forma anular alrededor de una par de
microtúbulos simple, con una disposición de
9 + 2 que es característica de casi todos los
cilios y flagelos eucariotas.

• Microfotografía electrónica del corte transversal de un flagelo de Chlamydomonas que ilustra la disposición 9 + 2 característica de los microtúbulos.

• Los microtúbulos se asocian a diferentes proteínas que se proyectan en posiciones regulares a lo largo de su haz. Alguna actúan como uniones cruzadas que mantienen unido el haz de microtúbulos; otras generan al fuerza que causa la incurvación del cilio.

Flagelos y cilios • La mas importante de las proteínas accesorias es la
dineína que provoca el movimiento de incurvación
de la parte central. La dineína ciliar esta unida por su
cola a un microtúbulo, mientras que su cabeza
interactúa con un microtúbulo adyacente generando
una fuerza deslizante entre ambos filamentos.

• Dados los múltiples enlaces que mantienen
juntos a los dobletes de microtúbulos
adyacentes, lo que sería un simple
movimiento de deslizamiento paralelo entre
microtúbulos libres, se convierte en un
movimiento ondulatorio en el cilio.
Flagelos y cilios

• Los dobletes de microtúbulos se deslizan entre sí con un movimiento telescopado por la acción repetitiva de las dineínas asociadas.
Flagelos y cilios

• En un flagelo intacto, los dobletes están conectados entre sí por uniones proteicas flexibles, de modo que la acción de las dineínas produce incurvación en lugar de deslizamiento.
Flagelos y cilios

• Los filamentos de actina se encuentran en todas las
células eucariotas y son esenciales para muchos de sus
movimientos sobre todo los relacionados con la
superficie celular. Por ejemplo en ausencia de filamentos
de actina una célula animal no podría desplazarse a lo
largo de una superficie, englobar una partícula de gran
tamaño mediante fagocitosis ni dividirse en dos células.
Micro filamentos

• Los filamentos de actina se asocian con proteínas
fijadoras que posibilitan que los filamentos cumplan
diversas funciones celulares. Los filamentos de
actina pueden dar lugar a la formación de
estructuras rígidas y relativamente permanentes
como las micro vellosidades del ribete en cepillo
que tapizan el intestino.
Micro filamentos
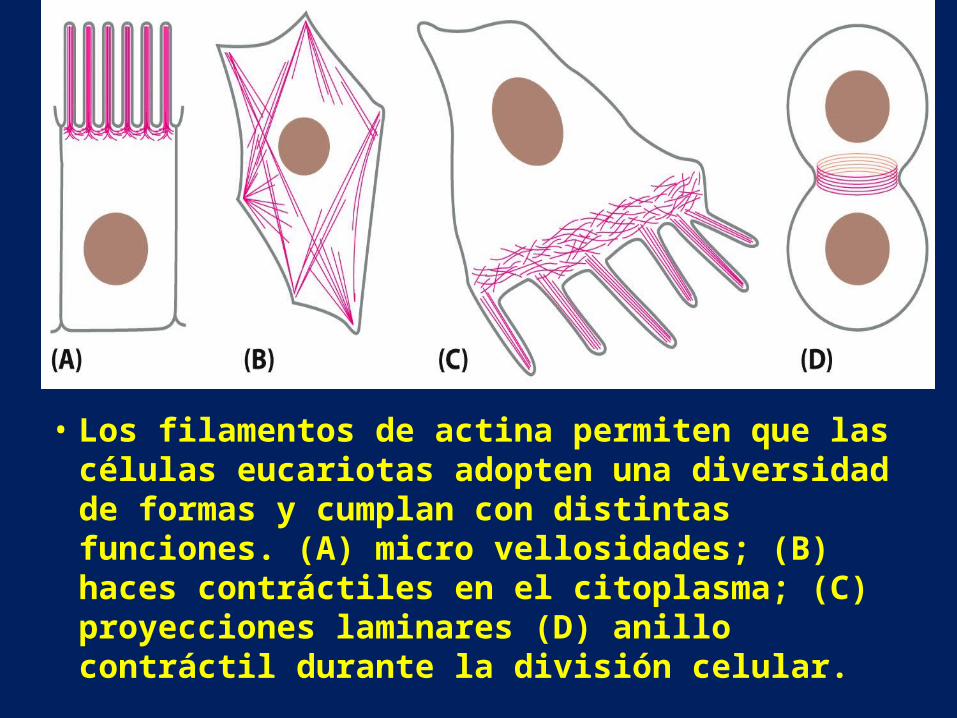
• Los filamentos de actina permiten que las células eucariotas adopten una diversidad de formas y cumplan con distintas funciones. (A) micro vellosidades; (B) haces contráctiles en el citoplasma; (C) proyecciones laminares (D) anillo contráctil durante la división celular.

• Los filamentos de actina se visualizan como hebras de
7 nm de diámetro. Cada filamento es una cadena
retorcida de moléculas globulares de actina idénticas,
que apuntan en la misma dirección a lo largo de la
cadena del eje. Un filamento de actina tiene una
polaridad estructural, con un extremo + y un extremo
-.
Micro filamentos

Micro filamentos

• Como en los microtúbulos, muchas de la funciones
desempañadas por los filamentos de actina, como su
participación en la motilidad celular, requieren la
capacidad de ensamblarse y desensamblarse. Las
citocalasinas impiden la polimerización de la actina,
mientras como la faloidina, estabilizan los filamentos de
actina y los protegen de la despolarización.
Micro filamentos

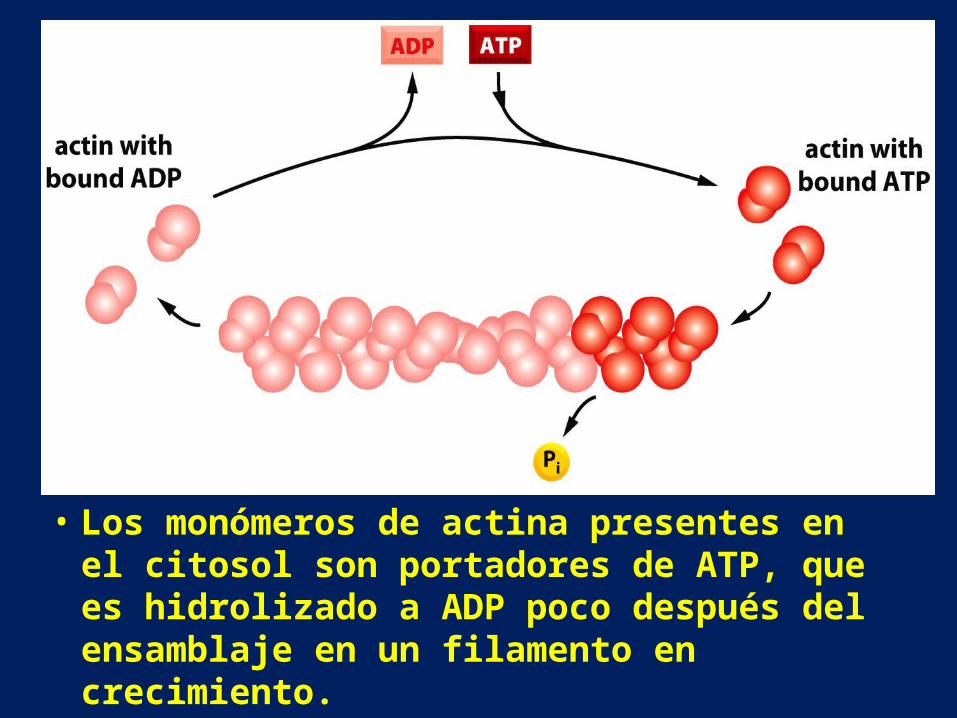
• Los monómeros de actina presentes en el citosol son portadores de ATP, que es hidrolizado a ADP poco después del ensamblaje en un filamento en crecimiento.
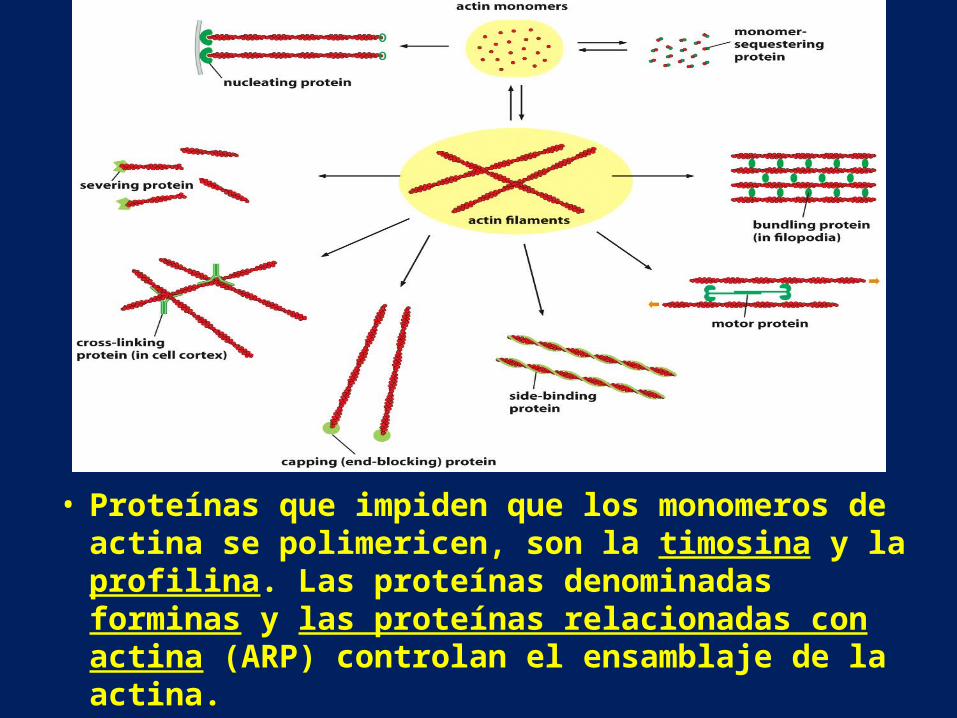
• Proteínas que impiden que los monomeros de actina se polimericen, son la timosina y la profilina. Las proteínas denominadas forminas y las proteínas relacionadas con actina (ARP) controlan el ensamblaje de la actina.

Micro filamentos • Los filamentos de actina actúan formando redes o
haces. Los filamentos de actina que descansan
debajo de la membrana plasmática, por ejemplo
están asociados a una red e proteínas que se unen
a la actina formando el córtex celular.

Micro filamentos • Mecanismos de
deslizamiento.1. La célula emite protrusiones
en su frente o borde activo.2. Estas protrusiones se
adhieren a la superficie sobre la cual la célula se desliza.
3. El resto de la célula se arrastra hacia adelante por tracción sobre los puntos de apoyo.