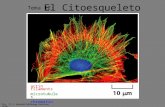
14. Citoplasma-Citoesqueleto
24
UNIDAD N° 2: UNIDAD N° 2: Temas 4 y 5: CITOPLASMA Y Temas 4 y 5: CITOPLASMA Y CITOESQUELETO CITOESQUELETO Blgo. Ms. Pablo Chuna Mogollón Profesor Auxilar T.C. Area Biología Departamento Académico de Ciencias - UPAO
-
Upload
aurelia-gonzales-calle -
Category
Documents
-
view
110 -
download
5
Transcript of 14. Citoplasma-Citoesqueleto

UNIDAD N° 2: UNIDAD N° 2:
Temas 4 y 5: CITOPLASMA Y Temas 4 y 5: CITOPLASMA Y CITOESQUELETO CITOESQUELETO
Blgo. Ms. Pablo Chuna Mogollón
Profesor Auxilar T.C.
Area Biología
Departamento Académico de Ciencias - UPAO

El citoplasma puede ser dividido en dos espacios: el citosol y el encerrado en el interior de los organelos.
El citosol o matriz citoplasmática es considerado el verdadero medio intracelular, que se extiende desde la envoltura nuclear hasta la membrana plasmática y que llena el espacio no ocupado por el sistema de endomembranas, las mitocondrias y los peroxisomas.
Em promedio el citosol reprenta el 50% del volumen del citoplasma, cifra que aumenta en las células embrio-narias y en las menos diferenciadas.
El pH del citosol es de 7.2.
FUNCIONES CELULARES QUE OCURREN EN EL CITOSOL
• Reacciones metabólicas Ej. Reacciones enzimáticas de glucólisis, glucogenólisis• Reacciones de Biosíntesis Ej. Biosíntesis de aminoácidos, nucleótidos, ácidos grasos,
glucogenogénesis,etc.• Tráfico intracelular Ej. RER- Golgi - Secreción• Respuestas intracelulares a moléculas de señalización celular • Múltiples reacciones en las que intervienen el ATP, GTP, RNAt, RNAm y el RNAr.• Localización de elementos del citoesqueleto, las chaperonas, proteosomas, inclusiones, etc.
CITOSOL

Para que el plegamiento de una proteína sea correcto se necesita la intervención de unas moléculas llamadas chaperonas, que se designan así porque acompañan a las proteínas y previenen sus plegamientos prematuros y cuidan que sean correctos. Chaperonas
Proteína plegada
PolipéptidoCompletoliberado
CHAPERONAS
PROTEOSOMAS
En el citosol existen estructuras que destruyen a las
proteínas. Así cuando una proteína debe
desaparecer, porque se ha plegado mal, se ha
dañado o su función ha concluido, es degradada por
un complejo enzimático llamado proteosoma.

CITOESQUELETO
El citoesqueleto comprende un armazón proteico filamentoso desplegado por todo el citosol.
Está integrado por tres clases de filamentos: microtúbulos, microfila-mentos y filamentos intermedios, y un conjunto de proteínas acceso-rias, clasificadas como regulado-ras, ligadoras y motoras.
Las proteínas reguladoras controlan el nacimiento, el alargamiento y la desaparición de los tres filamentos.
Las proteínas ligadoras conectan a los filamentos entre sí o con otros componentes de las células.
Las proteínas motoras sirven para trasladar macromoléculas y organelos de un punto a otro del citoplasma, o hace que dos fila-mentos contiguos se deslicen.

Los microtúbulos son filamentos que se hallan en casi todas las células eucariotas y poseen un diámetro de 25 nm.
Se caracterizan por su aspecto tubular, rectilíneos y uniformes.
En los cortes transversales presentan una configuración anular con una pared de 6 nm de espesor y una luz central uniformemente clara.
Son polímeros compuestos por unidades proteicas llamadas tubulinas. Cada tubulina es un heterodímero de 110 a 120 Kda, cuyas dos subunidades son: α-tubulina y β-tubulina.
MICROTUBULOS

De acuerdo con su localización, se clasifican en:
1.Citoplasmáticos, presentes en la célula en interfase.
2.Mitóticos, correspondientes a las fibras del huso mitótico.
3. Ciliares, localizados en el eje de los cilios.
4.Centriolares, pertenecientes a los cuerpos basales y los centriolos.
Los microtúbulos nacen en una estructura contigua al núcleo denominada centrosoma.
(se le llama también centro organizador de los microtúbulos o MTOC)
El centrosoma esta compuesto por un par de centriolos o diplosoma (gr. Diploos= doble y soma= cuerpo) y una sustancia amorfa
que lo circunda la matriz centrosomica.
La matriz contiene un complejo de proteinas
reguladoras denominadas γ-tubulinas

Los heterodímeros pueden agregarse (polimerizarse) o retirarse (despolimerizarse) por ambos extremos. Durante la polarización el microtúbulo se larga, y durante la despolimerización se acorta.
Uno de los extremos del microtúbulo se llama más [+], el otro, menos [-]. Estas designaciones se deben a que por el extremo [+] el microtúbulo se alarga y se acorta más rápidamente que por extremo [-].
En el extremo [-] de los microtúbulos se encuentra el complejo γ tubulina que tiene forma anular, y se comporta como un molde a partir del cual se nuclean las primeras 13 tubulinas.
El microtúbulo comienza a crecer por su extremo [+] al agregarse nuevas tubulinas provenientes del depósito de tubulinas del citosol.

Proteínas asociadas a los microtúbulos (MAPs).
Las MAPs parecen colaborar en el ensamblaje de los dímeros para formar microtúbulos, y también participan en la estabilización del microtúbulo y en la relación entre microtúbulos adyacentes.
Dos familias de proteínas motoras, las quinesinas y las dineínas, transportan las vesículas limitadas por membrana, las proteínas y los organelos a lo largo de los microtúbulos.
Casi todas las quinesinas desplazan cargas hacia el extremo [+] de los microtúbulos, mientras que las dineínas transportan cargas hacia el extremo [-] (t. retrógrado).

En el cuerpo neuronal y en el axon se ha identificado una MAP reguladora llamada
tau (τ) que inhibe la despolimerización de las tubulinas en los extremos de los microtúbulos. Ejerce también una función ligadora, ya que establece puentes entre los microtúbulos contiguos y les confiere estabilidad.
Las tau contienen un número determinado de fosfatos, cuyo aumenta altera su funciona-miento normal. Esto ocurre en la enfermedad de Alzheimer, caracterizada por un deterioro neuronal progresivo a consecuencia de la inestabilidad de los microtúbulos.
ALZHEIMER

La colchicina, un medicamento utilizado par el tratamienro de la gota, actúa uniéndose a las tubulina e impide su polimerización, lo que lleva –al no formarse el capuchón- a la desaparición de los microtúbulos
Los alcaloide de la Vinca (vincristina y vinblastina) tienen acción similar a la de la colchicina, aunque lo hacen casi selectivamente sobre las fibras del huso, de ahí que se les utilice para bloquear las divisiones en las células neoplásicas en el tratamiento del cáncer.
El taxol es otra droga usada para tratar el cáncer, pues impide la despolimerización de las fibras del huso e induce su crecimiento descontrolado, incompatible con la división celular.
AGENTES QUE AFECTAN A LOS MICROTUBULOS
Colchicina

• Forma celular. La posición del RE, aparato de Golgi y las mitocondrias se ha relacionado con los microtúbulos. Asimismo es evidente su papel en la vaina caudal de espermátides, en los axones y dendritas, y en la banda marginal de eritrocitos nucleados.
• Transporte celular. Los microtúbulos actúan como un soporte o carril por la superficie del cual proteínas motoras transportan diversas estructuras. Ej. el transporte axónico rápido anterógrado transporta materiales desde el soma neural hacia el terminal axónico. El transporte axónico rápido retrógrado mueve vesículas recubiertas que se reciclan, etc.
• Exocitosis (secreción) y Endocitosis. Los microtúbulos intervienen en el transporte de vesículas de endocitosis por el interior de la célula y en la secreción más tardía, de los gránulos desde el aparato de Golgi hasta la membrana plasmática.
• Movimiento de los cromosomas.
FUNCIONES DE LOS MICROTUBULOS

CILIOS
Los cilios son apéndices delgados de 0,25 m de diámetro y varios micrones de largo, que surgen de la superficie de diversos tipos celulares.
Cada uno está compuesto por un eje citosólico, la matriz ciliar, envuelto por unas prolongación de la membrana plasmática. En medio de dicha matriz, se encuentra un armazón filamentoso regular llamado axonema.
Cada cilio nace en un cuerpo basal o cinetosoma (gr. Kineetos= movible, soma= cuerpo), que es una estructura idéntica a un centriolo del diplosoma.

El movimiento ciliar es producido por el axonema, que en un corte transversal los microtúbulos del axonema muestran una configuración especial, conocida como “9 + 2”.
El axonema contiene proteínas ligadoras y proteínas motoras.
Las proteínas ligadoras unen a los dobletes entre sí y los sostienen en sus posiciones en el interior del cilio, como las nexinas y las proteínas radiales.
Las proteínas motoras representadas por la dineinas, que forman puentes inestables entre los dobletes contiguos.
El síndrome de kartagener se debe a una o más mutaciones de los genes que codifican a la dineína ciliar o a otras proteínas accesorias del axonema. Por consecuencia los cilios y los flagelos son inmóviles, lo que provoca cuadros de bronquitis crónica y esterilidad en la mujer y en el varón.

Los centriolos y los cuerpos basales poseen una estructura idéntica. Constituyen cilindros huecos abiertos en sus extremos. La pared del centriolo o del cuerpo basal está formada por 9 unidades microtubulares cada una compuesta por tres microtúbulos fusionados entre sí.
Los centriolos del diplosoma se deiferencian de los cuerpos basales porque:
a. Los centriolos se localizan cerca del núcleo y los segundos cerca de la superficie celular.
b. Los centriolos poseen matriz centrosómica y los cuerpos basales no la poseen.
c. Los centriolos se presentan de a dos, ambos perpendiculares entre sí, y los cuerpos basales están formados por una sola unidad.
CENTRIOLOS

MICROFILAMENTOS O FILAMENTOS DE ACTINA
Los microfilamentos poseen un diámetro de 7-8nm y son más flexibles que los microtúbulos. Suelen asociarse en haces de modo que raramente se les ve aisladas.
Sobre la base de su distribución en la célula, se clasifican en:
a.Corticales, los que se ubican por debajo de la membrana plasmática.
b.Transcelulares, dado que atraviesan el citoplasma en todas las direcciones.
Los microfilamentos son polímeros constituidos por el agregado de monómeros. Cada monómero se halla asociados a un ADP o a un ATP, su estructura terciaria es globular, de ahí que reciba el nombre de proteína G.
A igual que los microtúbulos, los microfilamentos poseen un extremo [+] y un extremo [-], por el primero se alargan y se acortan más rápidamente que por el segundo

Cada microfilamento comienza a formarse a partir de un núcleo de tres monómeros de actina G que se combina entre sí.
El alargamiento del núcleo originario se produce como consecuencia del agregado sucesivo de nuevos monómeros en los extremos [+] y [-] del filamento.
La polimerización requiere que la actina G contenga un ATP.
La droga citocalasina B provoca la despolimerización de los filamentos de actina debido a que se une a dos sus extremos y bloquea su crecimiento, con la consiguiente desparición de los capuchones de actinas con ATP.
Las faloidinas, por su parte, se combinan lateralmente con los microfilamentos de actina, estabilizándolos
Citocalasina B

• Contracción de células musculares y no musculares• Microfilamentos de las microvellosidades intestinales.• Movimiento ameboide.• Movimiento de fibroblastos• Anillo ecuatorial contráctil de la citocinesis.• Fusión de organelos membranosos.
FUNCION DE LOS MICROFILAMENTOS

Los filamentos intermedios son estructuras fuertes, insolubles, resistentes a los cambios de temperatura y dispuestas en densas redes tridimensionales en el citoplasma que se relacionan y forman parte de las uniones intercelulares.
Los filamentos intermedios forman una red continua tendida entre la membrana plasmática y la envoltura nuclear, alrededor de la cual componen una malla filamentosa compacta.
La composición química de los filamentos intermedios es diversa.
FILAMENTOS INTERMEDIOS
Tipo P. M. (Kd) Diámet. Tipos celulares
Queratina 40 -70 8 nm Células epiteliales
Desmina 53 10 nm Células musculares
Vimentina 57 10 nm Células de origen mesenquimatoso y algunas cél. epiteliales
Neurofilamentos 62 - 110 10 nm Neuronas
Gliofilamentos 50 8 nm Astrocitos y células de schwann
Otros filamentos
Nestina
Láminofilamentos
200
62 - 72
10 nm
redes
Neuronas embrionarias
Lámina nuclear (todas las células)

Distribución de filamentos Intermedios en el cuerpo humano

Los filamentos intermedios son polímeros lineales cuyos monómeros son proteínas que presentan una estructura en hélice α fibrosa.
Las proteínas fibrosas están integradas por una sucesión de secuencias idénticas de 7 aa cada una, lo que les permite combinarse entre sí lado con lado y formar dímeros lineales.
Los dímeros vuelven a combinarse entre sí para generar tetrámeros.
Los tetrámeros se conectan por sus extremos y dan lugar a estructuras cilíndricas alargadas llamadas protofila-mentos, los cuales se adosan por sus lados y componen una estructura fibrilar de 10 nm de diámetro

Epidermolisis ampollar simple
Epidermolisis ampollar simple (EBS)
Hiperqueratosis epidermolitica (HE)

Acumulacion y ensamble anormal deneurofilamentos
Perdida progresiva de neuronas motoras
Atrofia muscular, parálisis, y muerte
El citoplasma de las neuronas contiene haces flojos de filamentos intermedios cuyo eje longitudinal se orienta en paralelo con el del axon. Estos neurofilamentos estan formados por tres proteinas distintas: NF-L, NF-H y NF-M
Esclerosis amiotrófica lateral
En noventa por ciento de los niños con progeria tienen una mutación en el gen que codifica la proteína lamina A de la lámina nuclear:
Síndrome de Hutchinson Gilford

CARACTERÍSTICASMICROTUBULOS MICROFILAMENTOS FILAMENTOS
INTERMEDIOS
Distribución Todas las eucariotas Todas las eucariotas Células animales
Dimensiones 20-25 nm de diámetro 6-8 nm de diámetro 10 nm de diámetro
Estructura Tubo rígido y hueco Filamento helicoidal Fibras gruesas similares a cuerdas
Características Algunos son lábiles y transitoriosOtros son permanentes
Delgados, filamentos flexibles
Fuertes estructuras estables
Localización en célula En cilios, flagelos, huso mitótico
Eelementos contráctiles de músculos, microvilli, anillo contráctil en células en división
En hemidesmosomas, desmosomas, lámina nuclear
Subunidades incorporadas en un polímero
Heterodímero GTP tubu-lina-αβ, Tubulina γ para nucleación
Monómeros ATP-actina Varias proteínas fibrosas
Sitio preferencial de la incorporación
Extremo (+) (tubulina-β) Extremo (+) Interno
Polaridad Sí Sí No
Equilibrio dinámico Sí Sí No

CARACTERÍSTICASMICROTUBULOS MICROFILAMENTOS FILAMENTOS
INTERMEDIOS
Proceso de ensamble Cada dímero de tubulina
une GTP antes de ser
incorporado en microtú-
bulo, la polimerización
requiere Mg2+,
Monómeros de actina-G
son agregados para el
crecimiento del filamen-
to, requiere de presencia
de Mg2+, K+.
Los monómeros forman
dímeros helicoidales, los
que a su vez se asocian
para formar tetrámeros,
que se unen extremo
con extremo para formar
protofilamentos.
Proteínas motoras Cinesinas, dineínas Miosinas Ninguna
Principal grupo de
proteínas relacionadas
Proteínas asociadas a
los microtúbulos (MAP)
Proteínas de unión con
actina
Plaquinas
Fármacos que actúan
sobre polimerización-
despolimerización
Colchicina
Nocodazol
Taxol
Citocalasina
Faloidinas
Funciones principales Transporte intracelular,
soporte
Motilidad, contractilidad Soporte estructural
Enfermedades en que
se ven alteradas
Síndrome de Kartagener
(falta brazos de dineína)
Síndrome de Young
(malformación de
enlaces radiales)
Epidermolisis ampollar
(mutación en gen de
queratina)
Esclerosis amiotrófica
lateral (ensamble anor-
mal de neurofilamentos)