
1. INTRODUCCIÓN El tomate (Lycopersicon esculentum Mill.), es ...
47
1. INTRODUCCIÓN El tomate (Lycopersicon esculentum Mill.), es una de las hortalizas de mayor importancia mundial. Nuestro país posee 20.000 hectáreas de cultivo, de ellas el 40% está destinado al consumo fresco y un 60% al procesamiento de otros productos (URRUTIA, 2000). Existen diferentes sistemas de producción de tomate: hidropónico, botado, bajo plástico, por nombrar algunos. La presente investigación se centra en la producción bajo plástico, donde la mayor superficie de producción se encuentra en la zona central con 1.073 hectáreas (INE, 1997). La producción bajo plástico presenta una limitación lumínica debido principalmente a las épocas de cultivo, la transparencia de la cubierta (polietileno), y la alta densidad de plantación utilizada. El cultivo de variedades indeterminadas de tomate requiere manejar la planta con un eje sin ramificaciones. Para ello se realiza poda de brotes axilares. En algunas circunstancias también se requiere de poda de hojas para regular el crecimiento vegetativo y reproductivo. La defoliación se realiza en presencia de hojas envejecidas que constituyen un gasto para la planta, así como las que presenten una lesión y pueden ser fuente de entrada para patógenos y en último caso las que generan un exceso de cobertura. Este manejo permite mejorar las condiciones de iluminación, aumentando la fotosíntesis al interior de la canopia y la temperatura ambiente. Los manejos de deshoje tienen un importante efecto sobre la intercepción de luz al interior de la canopia. La hipótesis de esta investigación plantea que el incremento de la iluminación y de la eficiencia de las hojas remanentes a causa del deshoje permitirían un aumento de la producción y calidad de los frutos.
Transcript of 1. INTRODUCCIÓN El tomate (Lycopersicon esculentum Mill.), es ...

1. INTRODUCCIÓN
El tomate (Lycopersicon esculentum Mill.), es una de las hortalizas de mayor
importancia mundial. Nuestro país posee 20.000 hectáreas de cultivo, de ellas el 40%
está destinado al consumo fresco y un 60% al procesamiento de otros productos
(URRUTIA, 2000).
Existen diferentes sistemas de producción de tomate: hidropónico, botado, bajo
plástico, por nombrar algunos. La presente investigación se centra en la producción
bajo plástico, donde la mayor superficie de producción se encuentra en la zona central
con 1.073 hectáreas (INE, 1997).
La producción bajo plástico presenta una limitación lumínica debido principalmente a
las épocas de cultivo, la transparencia de la cubierta (polietileno), y la alta densidad
de plantación utilizada. El cultivo de variedades indeterminadas de tomate requiere
manejar la planta con un eje sin ramificaciones. Para ello se realiza poda de brotes
axilares. En algunas circunstancias también se requiere de poda de hojas para regular
el crecimiento vegetativo y reproductivo. La defoliación se realiza en presencia de
hojas envejecidas que constituyen un gasto para la planta, así como las que presenten
una lesión y pueden ser fuente de entrada para patógenos y en último caso las que
generan un exceso de cobertura. Este manejo permite mejorar las condiciones de
iluminación, aumentando la fotosíntesis al interior de la canopia y la temperatura
ambiente.
Los manejos de deshoje tienen un importante efecto sobre la intercepción de luz al
interior de la canopia. La hipótesis de esta investigación plantea que el incremento de
la iluminación y de la eficiencia de las hojas remanentes a causa del deshoje
permitirían un aumento de la producción y calidad de los frutos.

2
El presente ensayo tiene por objetivo evaluar el efecto de la defoliación sobre el
rendimiento y calidad del fruto, así como la acumulación de biomasa, y parámetros
físicos como la temperatura y radiación.

3
2. REVISIÓN BIBLIOGRÁFICA
2.1. Descripción de la especie:
El tomate (Lycopersicon esculentum Mill.) es una planta dicotiledónea perteneciente
a la familia Solanácea. Su origen ha sido propuesto para la región andina de Perú,
Bolivia, Ecuador y el norte de Chile. Esta especie presenta un comportamiento
perenne en clima tropical y anual en clima templado.
2.1.1. Descripción morfológica de la planta
La planta presenta una raíz pivotante y es capaz de alcanzar cerca de tres metros de
profundidad y una extensión lateral de 1,5 metros. Cuando el sistema radicular sufre
algún daño tiene la capacidad de emitir raíces adventicias en la porción basal del tallo
(CORFO y UNIVERSIDAD CATÓLICA DE CHILE, 1986).
El tallo es anguloso, alcanza 4 cm de diámetro en su base y está recubierto por
tricomas, en su mayoría de origen glandular y que le otorgan el olor característico. En
las primeras etapas es erguido, luego debido al peso, toma un hábito rastrero
(MAROTO, 1994).
Las hojas están dispuestas alternadamente sobre el tallo, son compuestas e
imparipinnadas, generalmente tienen de siete a nueve foliolos lobulados o dentados y
también están cubiertas de tricomas (MAROTO, 1994).
La flor del tomate es perfecta. Los pétalos y los sépalos se encuentran dispuestos en
forma helicoidal en un número de cinco o más. En cada inflorescencia se agrupan tres
a diez flores formando el racimo floral (RODRÍGUEZ, TABARES y MEDINA,
1984).

4
El fruto es una baya, que dependiendo del cultivar, presenta distintos colores como
rojo, rosado, violáceo o amarillo. De igual manera su forma varía desde achatada a
pera. La superficie puede ser lisa o presentar surcos más o menos profundos (CORFO
y UNIVERSIDAD CATÓLICA DE CHILE, 1986).
2.1.2. Organización del tallo durante el crecimiento vegetativo y reproductivo
Al inicio del crecimiento el tallo principal genera de seis a 12 hojas en la porción
proximal antes que la yema principal se transforme en una inflorescencia. Las hojas
crecen lateralmente con una filotaxia 2/5.
El crecimiento reproductivo siguiente se produce a partir de la yema axilar de la
última hoja, la cual produce un tallo secundario que crece como una prolongación del
primario, desplazando lateralmente la inflorescencia. Los sucesivos segmentos del
tallo se desarrollan de igual forma, generando una inflorescencia cada tres hojas
(CHAMARRO, 1995).
El desarrollo del tallo es variable y está dado de acuerdo al cultivar. Según esto
existen dos hábitos de crecimiento: Primero el de tipo determinado, en el cual el
crecimiento del eje principal está detenido por la formación de una inflorescencia
terminal, después de haber formado una cierta cantidad de inflorescencias (dos o
tres), cada una o dos hojas, y desarrolla fuertes brotes axilares desde la base del tallo
y, segundo el de tipo indeterminado, en el que el alargamiento del eje principal es
continuo y produce inflorescencias laterales frecuentemente cada tres hojas, durante
gran parte de la vida de la planta (KINET y PEET, 1997).

5
2.1.3. Morfología de la hoja
Las hojas del tomate son pinnadas compuestas de siete a 11 foliolos laterales y un
gran foliolo terminal. Los foliolos laterales son usualmente peciolados, lobulados
irregularmente y con bordes dentados. Las hojas de tomate son de tipo dorsiventral.
El tejido parenquimático está recubierto por una epidermis superior y otra inferior, las
cuales no contienen cloroplastos. Inmediatamente debajo y perpendicularmente a la
epidermis superior se encuentra el parénquima en empalizada, que posee en el
citoplasma numerosos cloroplastos. El mésofilo esponjoso se encuentra situado entre
el mésofilo en empalizada y la epidermis inferior, y contiene un número de
cloroplastos cercano al del parénquima en empalizada. Los nervios primarios y
secundarios tienen una estructura similar al tallo y poseen un floema externo y un
floema interno (CHAMARRO, 1995).
Las hojas se originan a partir del primordio foliar, que aparece inicialmente como una
pequeña protuberancia sobre la cúpula del ápice, creciendo cerca de 200 µm antes de
los primeros signos de formación de los foliolos laterales (HO y HEWITT, 1986). En
cambio, el foliolo terminal se forma de un meristema marginal a lo largo de los
flancos del primordio, en el extremo distal. Los demás foliolos se desarrollan de
forma similar, a partir de grupos de células que forman pequeñas protuberancias
sobre los flancos del primordio (COLEMAN y GREYSON, 1976 y PICKEN et al.,
1986, citados por CHAMARRO, 1995).
La iniciación de las hojas se produce a intervalos de dos a tres días, en función de las
condiciones ambientales (CHAMARRO, 1995).

6
2.1.4. Desarrollo del fruto
El crecimiento acumulativo del fruto se puede representar en forma de una curva
sigmoídea dividida en tres períodos. El período inicial dura cerca de dos semanas,
durante el cual su crecimiento es lento. Es seguido por tres a cinco semanas de
crecimiento rápido hasta el estado verde maduro. Finalmente, un período de
crecimiento lento por dos semanas más, en el cual ocurren intensos cambios
metabólicos (CHAMARRO, 1995).
La división celular está limitada a la fase inicial de crecimiento lento, durante la cual
empieza la elongación celular. La tasa de crecimiento relativo del fruto alcanza su
máximo hacia el final de la primera semana, y luego declina durante el período de
rápido crecimiento, el que se origina únicamente de la elongación celular (HO y
HEWITT, 1986).
Durante el desarrollo de frutos de tomate la economía de hidratos de carbono está
determinada por todos los aspectos que involucra la relación fuente-depósito en la
planta. Esto implica que la producción de fotoasimilados se realiza en los órganos
fuente, siendo posteriormente particionados en las hojas, transportados y exportados
en forma de disacáridos a depósitos alternativos, y finalmente importados hacia los
frutos depósitos y metabolizados en su interior. Aunque el nivel de hidratos de
carbono en los frutos es producto de la interacción de todos estos procesos, el destino
de los fotoasimilados que se importan a los frutos es controlado por el metabolismo
propio de ellos (SCHAFFER et al., 1999).
El tamaño final del fruto está estrechamente relacionado con el número y peso de las
semillas, además del número de lóculos (HO y HEWITT, 1986). Según OWEN y

7
AUNG (1990), existe una relación lineal entre diámetro final del fruto y el diámetro
del ovario en antesis.
2.1.4.1. Maduración del fruto
Durante el período final de crecimiento lento del fruto, el color, sabor, aroma, textura
y composición cambian marcadamente (GRIERSON y KADER, 1986). Durante esta
etapa, la respiración aumenta hasta alcanzar un nivel máximo llamado pico
climatérico. Simultáneamente se produce un incremento en la producción de etileno,
que tiene una profunda influencia en el desarrollo del proceso de maduración
(CHAMARRO, 1995). La textura del fruto se modifica a través de la degradación de
las paredes celulares por acción de varias enzimas, de las cuales, la más importante es
la poligalactouronasa, resultando una textura blanda y jugosa. El cambio de color del
fruto, empieza dos a tres días después del estado verde maduro, y se desarrolla
progresivamente de amarillo a naranja y rojo, producto de la transformación de
cloroplastos a cromoplastos con la acumulación de pigmentos (GRIERSON y
KADER, 1986)
La calidad del fruto está principalmente representada por sus características de sabor
y color. El sabor del tomate está determinado principalmente por los niveles de
azúcares y ácidos. Los azúcares, glucosa y fructosa constituyen el 65% de los sólidos
solubles totales del fruto. Por su parte, el ácido málico y cítrico representan el 13% de
la materia seca del fruto. Al inicio del crecimiento del fruto, predomina el ácido
málico, mientras que el ácido cítrico sólo representa el 25% de los ácidos orgánicos.
El pH del jugo maduro oscila entre 4 y 4.8 (CHAMARRO, 1995).

8
En los frutos de tomate, el color se forma básicamente por dos pigmentos
carotenoides, siendo el licopeno el de mayor importancia frente al β-caroteno
(CHAMARRO, 1995).
2.1.5. Aspectos del desarrollo del fruto que inciden en la producción
El éxito comercial de un cultivo de tomate no sólo depende de la cantidad de fruta
cosechada, sino también de la calidad de los frutos. El rendimiento de una planta de
tomate depende tanto del número como del peso de los frutos (HO y HEWITT, 1986)
y de la relación que exista entre ellos (CASTRO, 1991).
El mejoramiento en calidad se logra mediante el cruzamiento de distintos cultivares, y
la cantidad de la fruta por el uso de tecnologías, siendo un buen ejemplo, los
invernaderos de condiciones ambientales controladas (HO y HEWITT, 1986).
El rendimiento final también está determinado por la relación entre el número y
calibre de la fruta. Así por ejemplo, con raleo de frutos ─ consistente en eliminar el
20% de ellos ─ el calibre aumenta en un 6%, pero el rendimiento total disminuye en
un 25% (CASTRO, 1991).
2.2. Factores medioambientales que afectan el desarrollo del cultivo:
2.2.1. Efecto de la luminosidad en el crecimiento y desarrollo
La integral de radiación diaria es más importante que la calidad de la luz y el
fotoperíodo para el crecimiento de la planta de tomate (PICKEN, STEWART y
KLAPWIJK, 1986). La floración del tomate no es muy sensible al largo del día, y
usualmente es descrita como una planta de día corto cuantitativa (HO y HEWITT,

9
1986), sin embargo, según KINET y PEET (1997), el tomate es considerado como
una planta de día neutro.
KINET (1977) estudió el efecto de diferentes condiciones de fotoperíodo e intensidad
lumínica sobre el desarrollo de la inflorescencia en tomate. En su experimento utilizó
intensidades de luz de 9.000 y 18.000 ergs cm-2 s-1. El estudio demostró que al
aumentar la integral de luz se acelera la iniciación floral, y se promueve el desarrollo
de la inflorescencia. También aumenta la tasa de producción de hojas y el crecimiento
del tallo. A igual intensidad lumínica, la iniciación floral fue más temprana y mejoró
el desarrollo de la inflorescencia en fotoperíodo corto. No hubo efecto del fotoperíodo
sobre la producción de hojas. Sin embargo, el crecimiento del tallo aumentó entre un
30 a 50% en día largo respecto al día corto.
La aparición de la primera inflorescencia se ve retardada en condiciones de baja
luminosidad. Al momento de la aparición macroscópica de la primera inflorescencia,
una baja irradiación (por ejemplo, 9 W/m2 por 8 horas diarias) reduce
progresivamente el grado de desarrollo de la primera flor, logrando el aborto total en
un período de 10 días. En forma similar, cuando las plantas crecen en ambientes con
integrales de luz sub-óptima se reduce el número de flores por racimo, y perjudica el
desarrollo de las mismas (HO, 1996).
2.2.2. Efectos de la luz en la productividad
La alta densidad de plantación utilizada en el tomate de invernadero tiene una fuerte
influencia en la producción, debido a la competencia de las plantas por la luz. Con
respecto a los diferentes materiales de cubierta usados en invernadero, se ha
demostrado que una pérdida de 1% de luz equivale a 1% de disminución en la
producción (HO y HEWITT, 1986). Se ha demostrado que tanto el tamaño del fruto

10
como el contenido de sólidos solubles están directamente relacionados con la
radiación solar recibida, ya que ésta influye sobre la producción de fotoasimilados,
los que después serán dirigidos a los diferentes depósitos de la planta (HO y
HEWITT, 1986).
En los estudios desarrollados por COCKSHULL, GRAVES y CAVE (1992) sobre la
influencia del sombreamiento artificial en el rendimiento del cultivo de tomate en
invernaderos, se encontró que en las primeras 14 semanas de cosecha hay una
relación directa entre la radiación solar recibida y el rendimiento. En dicho
experimento se estableció que el tomate produce 2.01 kg de peso fresco de fruta por
cada 100 MJ de radiación solar incidente en el cultivo. En cambio, a partir de la
semana número 15, el sombreamiento no parece afectar las formas básicas de
respuesta del cultivo a la radiación solar o del particionamiento de asimilados entre
los órganos reproductivos y vegetativos. El número de frutos por racimo se
correlacionó linealmente con la radiación solar recibida, especialmente cerca del
momento de la primera antesis de cada racimo, cuando la radiación fue menor a 1.5
MJ m-2 d-1.
2.2.3. Efectos de la temperatura sobre el crecimiento y desarrollo
Las temperaturas óptimas para el cultivo del tomate son de: 24-25°C/15-18°C
(día/noche). Bajo los 12°C se paraliza el desarrollo vegetativo, y con temperaturas
inferiores a los 7°C se necesita una ayuda artificial de calefacción (RODRÍGUEZ,
TABARES y MEDINA, 1984). La temperatura promedio diaria determina la
producción de tomate, la tasa de expansión del área foliar y el área foliar específica
(DE KONING, 1988).

11
La temperatura también controla los procesos de particionamiento de asimilados. Por
ejemplo, cuando la temperatura del aire es de 30°C/24°C (día/noche), una mayor
cantidad de asimilados se translocan hacia los racimos a expensas del desarrollo
radicular. Por otro lado, en condiciones de 17°C/12°C existe una mayor translocación
hacia las raíces a expensas de los racimos (SHISHIDO y HORI, 1979, citados por
HO, 1996).
2.2.4. Temperatura en el desarrollo reproductivo
La temperatura tiene una importancia fundamental en la velocidad de desarrollo de
las flores después de su iniciación. Por ejemplo, las flores de la primera inflorescencia
se desarrollan más rápidamente a una temperatura media de 20°C que a 16°C, y
además se promueve el desarrollo de la segunda inflorescencia (CALVERT, 1964).
La tasa de crecimiento del fruto está directamente relacionada con la temperatura,
cuyo rango óptimo nocturno es de 18 a 20°C (HO y HEWITT, 1986), pero esta
relación es influenciada por los cambios en los niveles hídricos y en menor grado por
la tasa de fotosíntesis neta. Con incrementos de temperatura se puede esperar una
mayor actividad metabólica, y de esta manera una mayor entrada de carbono y agua,
dando como resultado un aumento en la expansión celular (PEARCE, GRANCE y
HARWICK, 1993). Por otro lado, el rendimiento de frutos se reduce por temperaturas
del aire bajo los 18°C (HO y HEWITT, 1986).
Cuando el fruto se desarrolla entre 10 y 30ºC, la tasa de crecimiento se correlaciona
positivamente con la temperatura; bajo esa condiciones el diámetro aumenta en 2-5
µm h-1°C-1 (PEARCE, GRANCE y HARWICK, 1993). Similares resultados
obtuvieron ADAMS, COCKSHULL y CAVE (2001) al estudiar el tiempo entre

12
antesis y cosecha, con temperaturas de 14-16ºC. En dicho estudio la temperatura y el
tiempo de desarrollo presentó un coeficiente de correlación de 0.98.
2.3. Materia seca:
HALBROOKS y WILCOX (1980) señalan que la acumulación de materia seca en la
planta es relativamente lenta hasta el día 56 después de la plantación. Posteriormente
entre el día 56 a 70, la acumulación de peso seco total es rápida debido al desarrollo
vegetativo. El desarrollo del fruto ocurre entre los días 70 y 105, en donde el aumento
del peso total de la planta se debe a la acumulación de peso seco de los frutos. Este
último es más o menos constante desde el día 56 a 100 y como promedio acumulan
5,55 g·día-1.
2.3.1. Efecto de la defoliación en la ganancia de materia seca
La defoliación parcial de la planta modifica el patrón normal de distribución de la
materia seca. MARTÍNEZ et al. (1999) investigaron el aumento de peso en las hojas
remanentes en tomate, por efecto de la defoliación. El tratamiento consistió en la
poda de una y dos hojas por simpodio. Los resultados demostraron que la eliminación
de más de una hoja entre racimos produce un incremento en el peso seco de las hojas
que permanecen en la planta, con peso promedio de 30 g·hoja-1, en comparación con
15 g.hoja-1 en las plantas con poda de una hoja.
La defoliación también altera la proporción de la materia seca en estructuras
reproductivas versus vegetativas. ANDRIOLO et al. (2001) evaluaron los efectos de
la defoliación en la acumulación de materia seca y su distribución hacia el fruto. Los
tratamientos realizados consistieron en dejar una, dos y tres hojas por simpodio,
encontrando que la materia seca total fue superior en las plantas con tres hojas por

13
simpodio, pero en la materia seca del fruto no se detectó diferencias significativas
entre los tratamientos. Ellos concluyeron que la materia seca extra acumulada en las
plantas no defoliadas no se distribuyó a los frutos, sino que permaneció en las hojas
principales.
2.4. Relación fuente-depósito:
El patrón de crecimiento de una planta de tomate sugiere que hay una priorización en
la distribución de asimilados entre los órganos. Los depósitos tienen diferentes
habilidades para atraer asimilados (fuerza de depósito). Así, la prioridad de un órgano
para recibir asimilados es el resultado de la competencia entre los órganos depósitos
(HO, 1984, citado por HO, 1996). En la planta de tomate la distribución de los
asimilados desde la hoja hacia órganos en crecimiento se modifica a medida que la
planta se desarrolla. En estado de fructificación, los frutos se transforman en los
principales depósitos de asimilados, por encima de las hojas jóvenes, flores y raíces
(HO y HEWITT, 1986).
Cada una de las estructuras de la planta se nutre por hojas y la asignación de las hojas
a los órgano de la planta se van modificando a lo largo del crecimiento. Así por
ejemplo, la primera inflorescencia es abastecida principalmente por las hojas uno,
tres, seis y ocho; el ápice es abastecido por las primeras cuatro hojas y las raíces por
las hojas cinco a nueve. Las hojas uno, tres, seis y ocho entregan asimilados a
distintos órganos, y todas están conectadas al mismo conjunto de haces vasculares.
Probablemente, cada racimo recibe asimilados principalmente de las hojas en las dos
líneas adyacentes que están justo por debajo y por encima del racimo. Ello se debe a
la falta de una conexión vascular directa entre los racimos y las hojas opuestas. Una
cantidad relativamente menor de asimilados de estas hojas sería entregada a los
racimos. Como los frutos atraen asimilados localmente, el suministro de ellos hacia el

14
ápice y las raíces puede ser atribuido a unas pocas hojas superiores y basales,
respectivamente. Este patrón de distribución de asimilados durante la fructificación
consiste en una mayor acumulación de materia seca en los frutos (HO, 1996).
Bajo condiciones normales, la hoja es en sí misma uno de los más importantes
depósitos de asimilados. El porcentaje de exportación de los fotoasimilados de la hoja
es cercano al 15%, durante el crecimiento de dicho órgano. En una hoja madura sólo
el 50% de los fotoasimilados son transportados fuera de la hoja. Los fotoasimilados
de las hojas van preferentemente a la fruta del simpodio al cual pertenecen. En el caso
que las fuentes excedan los depósitos dentro de una unidad, los excesos de asimilados
pueden translocarse fuera de ésta (TANAKA y FUJITA, 1974).
Además de las limitaciones anatómicas descritas, la relación fuente-depósito se puede
modificar por factores exógenos. Dentro de éstos se puede mencionar la luz,
concentración de anhídrido carbónico, temperatura, nutrición, condiciones hídricas y
poda (HO y HEWITT, 1986).
2.4.1. Relación fuente-depósito en plantas defoliadas
Como se describió en la sección 2.4., el transporte de asimilados se concentra entre
las hojas de un determinado simposio y su fruta. La exportación fuera del simpodio se
presenta cuando hay una producción que excede la demanda local. La traslocación
inter-unidades puede llegar a ser mayor cuando las hojas o el racimo de una unidad
son removidos. Durante la etapa de maduración los frutos representan el mayor
depósito de asimilados, y su tasa de asimilación neta se incrementa al remover el 30%
del área foliar y decrece cuando la remoción es del 50% o 100% de las hojas
(TANAKA y FUJITA, 1974).

15
Según SLACK (1986), la capacidad que tiene una hoja de importar o exportar
asimilados depende también del momento en que se realice la defoliación. Así, al
realizar el manejo cuando la carga frutal en la planta es cercana al nivel máximo,
podría provocar disturbios del balance entre las fuentes y depósitos, resultando en una
pérdida de rendimiento. Al contrario, cuando los frutos están iniciando su
maduración, será posible mantener un balance en la planta durante el período de
maduración.
SHISHIDO, KUMAKURA y HORI (1993) estudiaron los cambios en las relaciones
fuente-depósito por efecto de la defoliación en tomate. Por medio del uso de
marcadores de 14C, encontraron que cuando existe una relación fuente-depósito entre
hojas la eliminación de la hoja depósito provoca que la fuente remanente realice un
ajuste compensatorio y desarrolle una nueva relación fuente-depósito. Ello provoca
un aumento en el porcentaje de distribución de asimilados, a expensas de la raíz, que
sostendría el crecimiento de la planta en el futuro. Estos autores señalan que cuando
se remueve un depósito que tiene una relación fuerte a una hoja-fuente, y al crearse la
nueva relación, la hoja-fuente mantiene el porcentaje de la exportación de 14C, y
aumenta la distribución de asimilados a las hojas en crecimiento y el ápice. Al
contrario, cuando un depósito tiene una relación débil y se encuentra alejado de la
hoja fuente, no se genera un cambio importante.
Por causa de la remoción de hojas, el potencial metabólico se expresa al máximo, se
incrementa la tasa de asimilación neta (TANAKA, FUJITA y KIKUCHI 1974) y la
actividad fotosintética de las restantes hojas (TANAKA, FUJITA y SHIOYA 1974).
Las plantas de tomate tienen una relación fuente-depósito potencial lo
suficientemente alta como para asegurar el desarrollo pleno de todos los puntos de
crecimiento (TANAKA, FUJITA y KIKUCHI 1974).

16
2.5. Manipulaciones hortícolas para regular la cantidad de hojas.
AUNG y KELLY (1966) señalan que el propósito de la poda es regular la forma y
tamaño de la planta así como la floración y fructificación. El principio implicado en
esta práctica es la relación entre el crecimiento vegetativo y reproductivo. Una de las
posibles razones que justifican el uso de la poda de hojas es el gran número de hojas
axilares presentes en la planta durante la fase de crecimiento vegetativo, que a
menudo tarda el inicio de la fase reproductiva.
Los investigadores LEOPOLD y LAM (1960) y DECOTEAU (1990) encontraron
una relación entre la precocidad y la poda de hojas, determinando un efecto
promovedor de la floración de la primera inflorescencia al remover las hojas más
jóvenes, mientras que la poda de las hojas maduras presenta un efecto inhibitorio
sobre la misma. LEOPOLD y LAM (1960) realizaron un estudio sobre influencia del
deshoje sobre la precocidad en variedades tardías y tempranas, utilizando diferentes
niveles de deshoje y en dos estados fenológicos de la hoja (joven y adulta),
encontraron que este manejo provoca una mayor precocidad en las variedades
tempranas y una respuesta opuesta en las variedades tardías.
Según AUNG y KELLY (1966) los resultados obtenidos por efecto de la defoliación
pueden ser influenciados por condiciones medioambientales. En condiciones de alta
intensidad lumínica y temperatura, como en verano, se puede acelerar la tasa de
crecimiento de las plantas y así promover la edad fisiológica de las hojas,
disminuyendo el efecto que genera la defoliación sobre la planta. La remoción de
hojas maduras en la planta de tomate resulta en una mayor tasa de asimilación neta
(TAN) en las hojas remanentes con respecto a las hojas jóvenes de la misma planta o
en las hojas de plantas no defoliadas. La alta TAN podría ser explicada sobre la base

17
de una rápida tasa en el incremento de área foliar de las hojas remanentes por efecto
del deshoje.
DECOTEAU (1990) estudió el efecto de la decapitación, remoción de hojas del tallo
principal y de los brotes laterales, realizando una defoliación de las hojas principales
y eliminando por completo las hojas inmaduras (longitud menor a 2.5 cm) de los
brotes axilares. El área foliar para el tratamiento de remoción de hojas axilares fue de
3440 cm², para la remoción de hojas principales fue de 3020 cm² y finalmente el
control resultó en 4700 cm². El autor concluyó que la poda de hojas principales
reduce el área foliar de la planta en un 36%, y no estimula el desarrollo adicional de
área foliar con respecto del control. Con la remoción de hojas de los brotes axilares se
redujo el área foliar total de la planta en un 27%, pero aumentó en el tamaño de las
hojas principales en un 33%, en comparación con el control.
2.5.1. Efecto de la defoliación en el rendimiento
El rendimiento depende de la fase del crecimiento en la que se realiza la poda de
hojas. El rendimiento no se reduce cuando el deshoje se realiza en la fase vegetativa,
mientras que si se efectúa durante la fase reproductiva se espera una reducción del
mismo en forma proporcional al grado de defoliación (TANAKA y FUJITA, 1974).
MARTÍNEZ et al. (1998) estudiaron el efecto de la remoción de una hoja entre
simpodios en tres momentos fenológicos del cultivo: en floración, en caída de pétalos
y en fruto con diámetro de 10 mm. Las plantas defoliadas presentan un rendimiento
promedio de 5 kg/planta, mientras que el control sólo obtuvo 4 kg/planta.
Por otro lado, SLACK (1983) evaluó el efecto de la remoción de hojas entre racimos
en el rendimiento. Los resultados obtenidos no mostraron una pérdida en la mayoría

18
de sus tratamientos, pero la poda de dos hojas entre racimos sucesivos causó una
pérdida de rendimiento cercana al 30% debido principalmente a la producción de
fruto de calibre pequeño.
ELIZONDO (1999) sometió a cuatro tipos de deshoje a plantas de tomate
indeterminado. Los manejos de poda que realizó son: eliminación de las cinco hojas
basales a 20 días post plantación, poda de la hoja que cubre el racimo al momento de
la aplicación de auxinas sintéticas, poda de dos hojas sobre cada racimo una semana
después de la aplicación de auxinas sintéticas y poda de la hoja que cubre a cada
racimo una semana después de la aplicación de auxinas sintéticas. En su investigación
pudo concluir que el deshoje de cinco hojas basales produce una pérdida de 15% del
rendimiento. El resto de los tratamientos no presentan diferencias significativas en el
rendimiento.
Los investigadores WOLK, KRETCHMAN y ORTEGA (1983) estudiaron la
respuesta del tomate a la defoliación. Los tratamientos que realizaron consistieron en
la remoción de las hojas en tres momentos fenológicos (primera inflorescencia,
floración completa y fruto con un diámetro de 2.5 cm), frente a tres niveles de
deshoje (25%, 50% y 80%). La defoliación del 25% o 50% anterior a la floración o
en antesis parece no tener influencia en el rendimiento. Sin embargo con
defoliaciones de 25% o 50% cuando la fruta se encuentra con un diámetro de 2.5 cm,
genera una reducción del 10% del rendimiento.
STACEY (1983) investigó el efecto de la defoliación en el rendimiento de tomate.
Sus resultados indican que la planta puede tolerar una considerable defoliación por
plagas o enfermedades, particularmente en las hojas basales del tallo, sin presentar
una pérdida importante en rendimiento. Así, una remoción de un cuarto del área foliar
de la hoja tiene un efecto menor en el número de frutos cosechados que la

19
eliminación de la mitad o de las tres cuartas partes de la lámina, resultando en una
reducción de la cantidad de fruta en 17% y 34%, respectivamente.
2.6. Manejos comerciales de poda:
Los manejos que se realizan en la planta de tomate corresponden a la eliminación de
brotes axilares, hojas y ápices (ROJAS, 1992).
En la poda de hojas existen tres criterios a considerar. En primer lugar se eliminan las
hojas denominadas parásitas que se ubican en la porción basal del tallo, cuando los
frutos han madurado. Esta condición se debe al envejecimiento natural o por la menor
llegada de luz, a causa del sombreamiento causado por el aumento en altura de la
planta. Estas hojas no son capaces de realizar la fotosíntesis eficientemente, logrando
un gasto de energía mayor que la producida. Segundo, se eliminan las hojas dañadas
por plagas o enfermedades, o por algún factor mecánico, químico o físico. Finalmente
en algunos casos es factible eliminar la hoja que cubre el racimo; esta labor se realiza
por razones de crecimiento vigoroso con internudos cortos en el cual la planta genera
hojas muy grandes que cubren completamente al racimo (ROJAS, 1992).

20
3. MATERIAL Y MÉTODO
3.1. Ubicación del ensayo:
El ensayo se realizó en el Fundo La Palma perteneciente a Inversiones Quintil S.A,
ubicado en la comuna de Quillota, provincia de Quillota, V región (Longitud 71°
Oeste, Latitud 32°50´ Sur) y a 130 metros sobre el nivel del mar.
3.2. Características de clima y suelo:
La zona de Quillota pertenece al clima Mediterráneo Marino, agroclima Quillota. Se
encuentra en el sector poniente del valle del Aconcagua. El régimen térmico de esta
zona se caracteriza por una temperatura media anual de 15.3°C, con una máxima
media del mes más cálido (enero) de 27°C y una mínima media del mes más frío
(julio) de 5.5°C. El período libre de heladas aprovechable es de nueve meses, de
septiembre a mayo. La suma anual de temperaturas, base 5°C, es de 3.700 grados/días
y 10°C, 1900 grados/días. Las horas fríos de marzo a noviembre llegan a 500. La
temperatura media mensual se mantiene sobre los 10°C (NOVOA et al., 1989).
Según MARTÏNEZ (1981), los suelos del sector de La Palma pertenecen a la serie La
Palma que se caracteriza por ser suelos sedimentarios, profundos y de origen aluvial,
de texturas arcillosas, de color pardo y buen drenaje.
3.3. Material vegetal:
Se utilizó un cultivo comercial de tomate en invernadero frío, denominado tomate de
otoño, durante el período del 15 de enero al 15 de julio de 2002. Se usó la variedad de
tomate FA-593 caracterizada por ser del tipo larga vida poseedor del gen rin y de
crecimiento indeterminado.

21
La siembra se realizó en contenedores el día 10 de diciembre de 2001. Posteriormente
el trasplante se realizó el 15 de enero de 2002. Las plantas se dispusieron en mesas de
plantación de 80 cm de ancho, con distancia de plantación de 20 cm sobre la hilera
(Anexo 1). La densidad de plantación fue de 28.000 plantas por hectárea.
La conducción se realizó a eje simple, dispuestas en “V” y el sistema de entutorado se
realizó por medio de una cinta gareta. Las plantas cuelgan de una estructura de
soporte compuesta por alambres a 2 m de altura a lo largo de la mesa de plantación.
Se utilizaron invernaderos fríos de madera, tipo dos aguas, de 4 metros de alto, 34
metros de largo y 6 metros de ancho agrupados en módulos de 30 capillas. El sistema
de ventilación del invernadero cuenta con lucarna cenital de proyección vertical y
ventanas ubicadas en la parte superior de los dos frentes del invernadero. La cubierta
utilizada fue de polietileno térmico (0.2 mm de espesor) de dos temporadas.
Los manejos culturales se realizaron de acuerdo a las técnicas actuales para la
producción forzada de tomates bajo plástico en la zona de Quillota (ALLENDES,
MENDOZA y ROJAS, 1992). A continuación se detallan algunos de ellos
considerados de mayor relevancia para este caso. Para permitir la cuaja óptima de los
frutos, se utilizó el producto de nombre comercial Procarpyl (i.a. Ácido-4-cloro
feoxiacético; (2-naftiloxy) acetamida). La dosis del producto comercial de 4 ml·l-1 fue
asperjada sobre el racimo floral cuando presentó un 50% de floración. Este manejo se
complementó con la vibración manual de las plantas en forma periódica. La poda de
los brotes axilares se realizó cuando alcanzaron un largo aproximado de 3 cm.
Se definió una carga frutal de cinco frutos por racimos, y las plantas se despuntaron
sobre el sexto racimo floral.

22
La fertirrigación se realizó de acuerdo a la solución nutritiva de Steiner modificado
(DUIMOVIC, 1993).
3.4. Tratamientos:
El manejo de poda de hojas consistió en retirar una de ellas por simpodio. El
momento de realizar el manejo estuvo dado por la condición de desarrollo del racimo
que corona a cada simpodio. Éste debe encontrarse con frutos de 10 mm de diámetro
ecuatorial (Martínez et al., 1998). Así por ejemplo, se retiró una hoja entre el primer y
segundo racimo, cuando los frutos del segundo racimo tenían un diámetro ecuatorial
de 10 mm, y así sucesivamente.
Los tratamientos consistieron en someter a las plantas a similares tipos de deshoje, en
una misma etapa fenológica, pero con una diferente distribución en la planta, como se
detalla a continuación:
T0: Testigo (sin deshoje)
T1: Poda de los simpodios 1- 2- 3
T2: Poda de los simpodios 4-5-6
T3: Poda de los simpodios 1-2-3-4-5-6
3.5. Mediciones:
Las mediciones de rendimiento, tamaño y calidad interna del fruto sólo se realizaron
sobre los racimos 1-2-3-4, ya que condiciones climáticas y sanitarias impidieron la
cosecha de los racimos 5 y 6.En las mediciones de calidad del fruto se realizaron
combinando los racimos 1-2 y 3-4, debido a que la cosecha de los racimos presentó
variabilidad en el tiempo.

23
3.5.1. Rendimiento, tamaño, crecimiento y calidad de frutos
La cosecha de los frutos se realizó dos veces por semana, desde los 45 hasta los 68
días después del transplante. El índice de cosecha correspondió a tomate rosado, es
decir, con un 30 a 50% de la superficie con coloración rosada o roja. Al momento de
la recolección se pesó cada uno de los frutos para obtener su calibre y el rendimiento
final. Los frutos se clasificaron según su peso en las siguientes categorías
(ELIZONDO, 1999):
Extra: Fruto con peso mayor a 250 g
Primera: Fruto con peso entre 150 y 250 g
Segunda: Fruto con peso entre 150 y 100 g
Tercera: Fruto con peso entre 100 y 80 g
Se midió el diámetro ecuatorial de los frutos con un pie de metro y con una
frecuencia de siete días, desde los 45 hasta los 136 días después del transplante. La
medición se realizó en dos plantas de la mesa central de cada unidad experimental,
realizando un seguimiento de dos frutos de cada racimo.
Color: se realizaron cuatro lecturas de color, en puntos equidistantes a lo largo del
diámetro ecuatorial, sobre la epidermis de cada fruto. La determinación se hizo por
medio de un colorímetro por reflexión (Minolta, modelo CR-200), que proporciona
valores de L, a y b basados en la tabla de colores de Hunter. Estos parámetros
indican: L, luminosidad, a el rango de color verde a rojo y b rango que va desde azul
a amarillo. Con los valores promedio obtenidos se calculó los parámetros Croma
(saturación) y Ángulo de matiz (Tono) por medio de las siguientes fórmulas
(McGUIRE, 1992):

24
pH: Los frutos de cada muestra se trituraron de manera de obtener una pulpa, sobre la
cual se realizó la medición con un pH-metro marca Schott modelo Handylab 1.
Sólidos Solubles: Se midió en el suero obtenido al decantar la pulpa del fruto por
medio de un refractómetro de mano de 10° Brix marca Atago modelo ATC-1.
Biomasa aérea: Se determinó la materia seca a tres plantas por unidad de muestreo.
Cada planta fue sometida a un secado en horno a 60 °C por 72 horas. Además, se
midió el peso fresco de 15 plantas.
3.5.2. Parámetros ambientales
Radiación fotosintéticamente activa: Se realizó con un censor marca Li-Cor, cada
siete días, entre las 10:00 AM y 11:00 AM, obteniendo una lectura en cada una de las
unidades de muestreo. La medición se efectuó a una altura de 32 cm sobre el suelo,
que corresponde a la altura promedio del primer racimo.
Temperatura: Se utilizaron termógrafos marca Ibutton modelo Thermochron. Se
dispusieron dos termógrafos en una planta de cada tratamiento, los que se ubicaron en
la parte basal (entre racimos 1-2) y distal (racimos 5-6). Los sensores se fijaron al
tallo mediante una envoltura de papel metálico y cinta adhesiva.
22)( bacCroma +=
( )baarctghtizÁngulodema /)( =°

25
3.6. Unidad experimental:
La unidad experimental está representa una parcela de forma rectangular de 48 m2,
está compuesta por tres mesas de cultivo y posee un número de 120 plantas.
La unidad La unidad de muestreo corresponde a una sección de tres metros lineales
sobre la mesa central de cada unidad experimental y está compuesta por 15 plantas
(Anexo 2).
3.7. Diseño experimental:
Se ocupó un diseño completamente al azar, con cuatro tratamientos y cinco
repeticiones. La distribución espacial de las parcelas se muestra en el Anexo 3. En los
cuadros de la sección 4 (Presentación y Discusión de Resultados) se especifica en
número de sub-muestras utilizadas para cada parámetro evaluado.
3.8. Análisis de datos:
Los datos se evaluaron mediante el análisis de Varianza (ANDEVA). Cuando el valor
P asociado al estadístico de prueba fue menor o igual a 0.05 se usó el test de
separación de medias de Tukey, con α = 0.05.

26
4. PRESENTACIÓN Y DISCUSIÓN DE RESULTADOS
4.1. Rendimiento:
En el presente ensayo no se detectaron diferencias significativas entre los
tratamientos con respecto al rendimiento (Cuadro 1).
El rendimiento final de una planta de tomate depende del número y del peso de los
frutos (HO y HEWITT, 1986) y de la relación entre ellos (CASTRO, 1991). Al igual
que en el presente ensayo, ELIZONDO (1999) y SLACK (1983), en sus ensayos de
defoliación no obtuvieron resultados significativos. El primero obtuvo un rendimiento
promedio de 100 ton/ha en cada tratamiento y el segundo logró un valor promedio de
diez kg·planta-1.
CUADRO 1. Efecto del deshoje en el rendimiento medido en ton/ha de fruta cosechada de los primeros cuatro racimos de tomate de otoño, var. FA-593. Quillota, 2002.
Tratamiento Rendimiento ton/ha *
Control
T1: Poda de los simpodios 1- 2- 3
T2: Poda de los simpodios 4-5-6
T3: Poda de los simpodios 1-2-3-4-5-6
71NS
75
79
81 S: no significativo (P> 0.05) * Promedio de la muestra de 15 plantas de la mesa central de cada unidad experimental.
STACEY (1983) y WOLK, KRETCHMAN y ORTEGA (1983) en sus ensayos de
defoliación encontraron diferentes respuestas del rendimiento, obteniendo un
aumento, mantención y disminución del mismo. En general, no sólo el nivel de
defoliación podría afectar el rendimiento, sino que también la etapa fenológica en la
cual este manejo se efectúa.

27
MARTÍNEZ et al. (1998) realizaron una poda de hojas equivalente a la realizada en
el presente ensayo. A diferencia del presente ensayo, ellos determinaron que la poda
de la tercera hoja entre cada racimo produce un aumento en el rendimiento.
La mantención del rendimiento equivalente al control en las plantas defoliadas puede
explicarse por una mayor eficiencia fotosintética de las hojas remanentes. TANAKA
y FUJITA (1974) plantean que la defoliación parcial (30% de las hojas) de tomate
aumenta la tasa fotosintética de las hojas restantes en las plantas, compensando
parcialmente la asimilación de las hojas removidas. Lo anterior es corroborado por
AUNG y KELLY (1966), que indican que la remoción de hojas maduras en el tomate
produce una mayor tasa de asimilación neta, probablemente producto del incremento
de área foliar de las hojas remanentes.
La teoría del aumento en la eficiencia fotosintética de las hojas remanentes planteada
por TANAKA y FUJITA (1974),) quedó demostrada en el estudio de SHISHIDO,
KUMAKURA y HORI (1993), que plantean que cuando se realiza una remoción de
hojas y se elimina el depósito de la relación fuente-depósito, la fuente remanente
realiza un ajuste compensatorio y desarrolla una nueva relación. Ello provoca un
aumento en el porcentaje de distribución de asimilados a las hojas jóvenes, a
expensas de la raíz que sostendría el crecimiento de la planta en el futuro.
4.2. Distribución de calibres:
En cada categoría de tamaño no se detectó diferencias significativas entre las medias
de los tratamientos (P>0.05) (Figura 1). El promedio de frutos grandes (primera)
correspondió a 55%, mientras que los mas pequeños (tercera) equivalen al 5%.
ELIZONDO (1999) tampoco encontró diferencias en el calibre de la fruta entre los
tratamientos de deshoje.

28
0
10
20
30
40
50
60
Extra Primera Segunda Tercera Cuarta
Categorías de tamaño
% d
e fr
uta
en c
ada
cate
goría
T0
T1
T2
T3
T0: testigo (sin deshoje)
T1: Poda de los simpodios 1- 2- 3
T2: Poda de los simpodios 4-5-6
T3: Poda de los simpodios 1-2-3-4-5-6
FIGURA 1. Efecto del deshoje sobre el calibre. Los frutos cosechados de clasificaron en cinco categorías de tamaño de tomate de otoño, var. FA-593. Quillota,
2002.
Tratamientos

29
4.3. Descarga de la fruta:
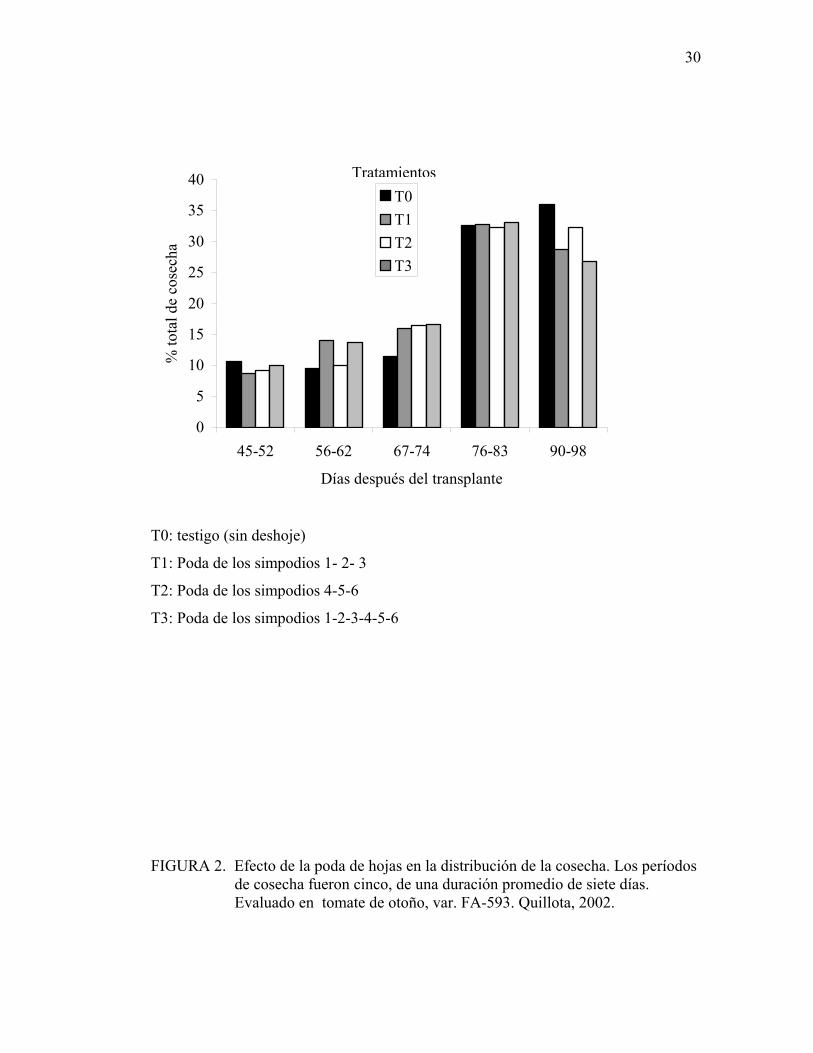
La cosecha se extendió por un período de cinco semanas. En las tres primeras
semanas, en el mes de abril, se cosechó un tercio del rendimiento total, y en las dos
últimas semanas (mes de mayo) se recolectó el resto de los frutos. Los resultados del
análisis estadístico indican que no existen diferencias significativas entre los
tratamientos al comparar la descarga de fruta en cada período de cosecha (Figura 2).
ELIZONDO (1999) tampoco encontró diferencias en la descarga de la fruta entre los
tratamientos de deshoje, evaluando los seis primeros racimos del cultivo.
Para el cultivo de tomate de otoño existe un factor de tipo económico que tiene
importancia, asociado a la estacionalidad de precio y oferta. De acuerdo a los datos
compilados por ODEPA (2002) para el período 1999-2001, a medida que la cosecha
se realiza en los meses de mayo y junio, la oferta de tomate se reduce de 15 millones
de kilos a sólo 5 millones, y los precios experimentan un aumento de $120 (mayo) a
$220 (junio) (Anexo 4). La estrategia comercial debería enfocarse a desplazar la
cosecha hacia el final del período de cultivo.
4.4. Crecimiento del fruto:
Los resultados demostraron que no existen diferencias significativas en la tasa de
crecimiento del fruto, así como en el número de días desde la cuaja a cosecha
(Cuadro 2). Este período de tiempo varió entre 51 y 53.4 días y la tasa de crecimiento
promedio fue de 0.6 mm mm-1·día-1. Los valores de días de cuaja a cosecha se ubican
dentro del rango mencionado por CHAMARRO (1995), quien precisa que el tiempo
necesario para que el ovario fecundado se desarrolle a un fruto maduro es de 7 a 9
semanas, en función del cultivar y las condiciones ambientales.
30
0
5
10
15
20
25
30
35
40
45-52 56-62 67-74 76-83 90-98
Días después del transplante
% to
tal d
e co
sech
aT0T1T2T3
T0: testigo (sin deshoje)
T1: Poda de los simpodios 1- 2- 3
T2: Poda de los simpodios 4-5-6
T3: Poda de los simpodios 1-2-3-4-5-6
FIGURA 2. Efecto de la poda de hojas en la distribución de la cosecha. Los períodos de cosecha fueron cinco, de una duración promedio de siete días. Evaluado en tomate de otoño, var. FA-593. Quillota, 2002.
Tratamientos

31
CUADRO 2. Efectos de la defoliación sobre el tiempo de desarrollo del fruto evaluado en los parámetros de tasa de crecimiento y días de cuaja a cosecha del fruto, en tomate otoño var. FA-593. Quillota, 2002.
Tratamiento
Días de cuaja a
cosecha*
Tasa crecimiento
(mm·mm-1·día-1)*
Control
T1: Poda de los simpodios 1- 2- 3
T2: Poda de los simpodios 4-5-6
T3: Poda de los simpodios 1-2-3-4-5-6
53.2NS
53.4
50.5
51
0.6 NS
0.7
0.6
0.6 NS: no significativo (P> 0.05) Promedio de una submuestra de 2 frutos de cada racimo de dos platas de la mesa central de cada
unidad experimental.
4.5. Sólidos Solubles:
Los resultados de los niveles de sólidos en los racimos frutales se presentan en el
Cuadro 3. Para esta variable no hubo diferencias significativas. En promedio el valor
de sólidos solubles fue de 3.87 °Brix.
CUADRO 3. Efecto del deshoje sobre en nivel de sólidos solubles (grados °Brix) de cada tratamiento medido en tomate de otoño, var. FA-593. Quillota,
2002. Sólidos solubles (°Brix)*
Tratamiento Racimo 1- 2 Racimo 3 - 4
Control
T1: Poda de los simpodios 1- 2- 3
T2: Poda de los simpodios 4-5-6
T3: Poda de los simpodios 1-2-3-4-5-6
3.76 NS
3.98
3.32
3.92
3.96 NS
4.08
3.98
3.94 NS: no significativo (P> 0.05) * Promedio de una muestra de 8 frutos de la mesa central de cada unidad experimental.

32
WOLK, KRETCHMAN y ORTEGA (1983) estudiaron la respuesta del tomate a la
defoliación. Ellos señalan que en la medida que la defoliación fue más intensa (en tres
niveles: 25%, 50% y 80%) el contenido de sólidos solubles disminuye
progresivamente al aumentar el deshoje. Sin embargo, en este ensayo no se
observaron diferencias significativas entre el tratamiento control y el nivel más alto
de deshoje. TANAKA, FUJITA y SHIOYA (1974), sugieren que la planta puede
ajustar la tasa de fotosíntesis en respuesta al deshoje, debido a que la actividad de las
hojas restantes se incrementa y compensa total o parcialmente la disminución del área
foliar.
4.6. Valor de pH:
Los valores de pH no presentan diferencias significativas para los diferentes
tratamientos que fueron sometidas las plantas (Cuadro 4).
CUADRO 4. Efecto del deshoje sobre el valor de pH de cada tratamiento medido en tomate de otoño, var. FA-593. Quillota, 2002.
pH
Tratamiento Racimo 1-2 Racimo 3-4
Control
T1: Poda de los simpodios 1- 2- 3
T2: Poda de los simpodios 4-5-6
T3: Poda de los simpodios 1-2-3-4-5-6
4.34NS
4.31
4.33
4.28
4.37 NS
4.31
4.30
4.31 NS: no significativo (P> 0.05) * Promedio de una muestra de 8 frutos de la mesa central de cada unidad experimental.
El valor promedio de pH fue de 4.31. Este resultado concuerda con lo descrito por
WOLK, KRETCHMAN y ORTEGA (1983), quienes señalan que el deshoje no afecta

33
el pH. Los valores obtenidos en este ensayo están dentro del rango de 4 a 4,8 indicado
por CHAMARRO (1995) como el normal para tomates de consumo fresco.
4.7. Color:
El parámetro de color externo del fruto no presenta diferencias significativas para
cada uno de sus componentes (luminosidad, croma y tono), como se indica en el
Cuadro 5. Los experimentos de deshoje realizados por otros autores no incluyen una
evaluación del color (ELIZONDO, 1999; STACEY, 1983 y MARTÍNEZ, 1998).
CUADRO 5: Efecto del deshoje sobre el color interno del fruto, medido en los parámetros de luminosidad, croma y tono en tomate de otoño, var. FA- 593. Quillota, 2002.
Luminosidad* Croma* Tono*
Racimo Racimo Racimo
Tratamiento 1-2 3-4 1-2 3-4 1-2 3-4
Control
T1: Poda de los simpodios 1- 2- 3
T2: Poda de los simpodios 4-5-6
T3: Poda de los simpodios 1-2-3-4-5-6
54,5NS
53,0
52,8
52,9
52,9 NS
52,9
54,3
53,9
23,7 NS
29,7
21,7
30,1
25,4 NS
28,9
25,1
30,1
67,3 NS
62,2
75,9
57,4
68,8NS
61,0
78,4
62,1
NS: no significativo (P> 0.05) Promedio de una muestra de 8 frutos de la mesa central de cada unidad experimental.
4.8. Biomasa aérea:
En el peso fresco y seco de la parte aérea de la planta no se encontró diferencias
significativas. El peso seco promedio fue de entre 175 g·planta-1; el peso fresco
promedio fue de 1.4 kg·planta-1 (Cuadro 6).

34
CUADRO 6. Efecto del deshoje sobre la biomasa aérea, medida en el peso fresco y materia seca en tomate de otoño, var. FA-593. Quillota, 2002.
Tratamiento Materia seca (g)* Peso fresco (kg)**
Control
T1: Poda de los simpodios 1- 2- 3
T2: Poda de los simpodios 4-5-6
T3: Poda de los simpodios 1-2-3-4-5-6
168,6NS
182,2
165,1
184,0
1,4NS
1,4
1,4
1,3
NS: no significativo (P> 0.05) *Promedio de una submuestra de 3 plantas de la unidad de muestreo de cada unidad experimental. ** Promedio de la muestra de 15 plantas de la mesa central de cada unidad experimental.
En el presente ensayo la medición de materia seca se realizó sobre las estructuras
aéreas de la planta (tallo y hojas), ya que es complejo extraer raíces del suelo;
además, proporcionalmente las raíces representan un 5% del peso seco de la planta.
En forma similar, el peso seco de las hojas en 2.4 veces el peso del tallo (VÁSQUEZ,
1994).
La distribución de la materia seca en la planta podría ser explicado por MARTÍNEZ
et al. (1999). Estos investigadores utilizaron una metodología de evaluación donde
sólo midieron la ganancia de peso seco de las hojas. Según sus resultados no se
produjo una diferencia de peso seco, por que los manejos de poda no superaron a una
hoja por simpodio.
Los datos de esta investigación se contradicen por lo planteado por los investigadores
ANDRIOLO et al. (2001), quienes utilizando una metodogía similar al presente
ensayo en los manejos de poda y medición de materia seca (tallo y hojas),
determinaron que existen diferencias de peso seco entre los tratamientos y el control
por efecto del deshoje. Ellos concluyeron que la materia seca extra acumulada en las

35
plantas no defoliadas no se distribuyó a los frutos, sino que permaneció en las hojas
principales.
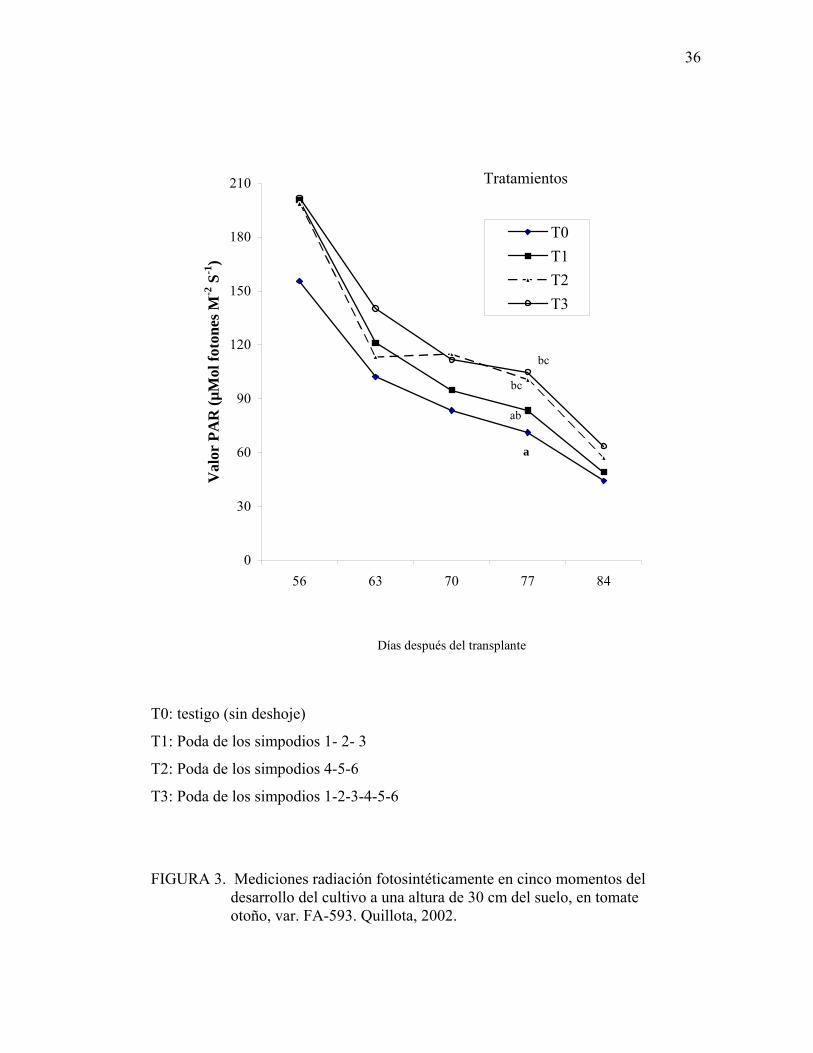
4.9. Intercepción de radiación fotosintéticamente activa:
Se realizaron 6 mediciones de radiación fotosintéticamente activa durante todo el
período del ensayo. Sólo en la medición realizada a los 77 días post-transplante se
detectaron diferencias significativas (Figura 3). El mayor nivel de radiación PAR se
logró con los tratamientos 2 (poda de los simpodios cuatro al seis) y 3 (poda de los
simpodios uno al seis), con 30% más de radiación que el testigo.
La importancia del aumento de la iluminación al interior de la canopia queda de
manifiesto por la investigadores COCKSHULL, GRAVES y CAVE (1992) indican
que existe una relación entre la radiación solar incidente y la ganancia de peso fresco
de la fruta. De acuerdo a ello, la mayor radiación al interior de la canopia observada
en este ensayo podría haber afectado positivamente el rendimiento, situación que no
ocurrió.
4.10. Temperatura:
Los valores de temperatura se presentan ordenados en rangos en las Figuras 4 y 5. En
general, todos los tratamientos tanto en la porción distal como basal del tallo, siguen
el mismo patrón térmico durante el cultivo. Se determinó que la temperatura
disminuye levemente en la parte basal de la planta comparada con los datos de
temperatura de la parte superior. La mayor parte del tiempo la temperatura fluctuó
entre 15 a 20°C tanto en la porción basal y distal de la planta. La temperatura
promedio diaria durante el período del cultivo fue de 18°C. Dicha temperatura
representa una limitación moderada para lograr la máxima expresión del potencial
36
0
30
60
90
120
150
180
210
56 63 70 77 84
Días después del transplante
Val
or P
AR
(µM
ol fo
tone
s M-2
S-1
)T0T1T2T3
a
ab
bc
bc
T0: testigo (sin deshoje)
T1: Poda de los simpodios 1- 2- 3
T2: Poda de los simpodios 4-5-6
T3: Poda de los simpodios 1-2-3-4-5-6
FIGURA 3. Mediciones radiación fotosintéticamente en cinco momentos del desarrollo del cultivo a una altura de 30 cm del suelo, en tomate otoño, var. FA-593. Quillota, 2002.
Tratamientos

37
0100200300400500600700800900
1000
0 a 5 5 a 10 10 a 15 15 a 20 20 a 25 25 a 30 30 a 35
Rango de temperatura (°C)
N° d
e ho
ras
T0T1T2T3
T0: testigo (sin deshoje)
T1: Poda de los simpodios 1- 2- 3
T2: Poda de los simpodios 4-5-6
T3: Poda de los simpodios 1-2-3-4-5-6
FIGURA 4. Registro de temperaturas durante el período de cultivo. Los sensores se ubicaron de la parte basal de la planta. Los datos se ordenaron en rangos de temperatura. Evaluado en tomate de otoño, var. FA-593. Quillota, 2002.
Tratamientos

38
0100200300400500600700800900
1000
0 a 5 5 a 10 10 a 15 15 a 20 20 a 25 25 a 30 30 a 35
Rango de temperatura (°C)
N° d
e ho
ras
T0T1T2T3
T0: testigo (sin deshoje)
T1: Poda de los simpodios 1- 2- 3
T2: Poda de los simpodios 4-5-6
T3: Poda de los simpodios 1-2-3-4-5-6
FIGURA 5. Registro de temperaturas durante el período de cultivo. Los sensores se ubicaron de la parte distal de la planta. Los datos se ordenaron en rangos de temperatura. Evaluado en tomate de otoño, var. FA-593. Quillota, 2002.
Tratamientos

39
productivo de la especie, ya que la temperatura óptima es de 24-25ºC durante el día
(RODRÍGUEZ, TABARES y MEDINA, 1984).
4.11. Consideraciones finales:
Los manejos de poda generan mejores condiciones de iluminación al interior de la
canopia. La planta de tomate muestra una plasticidad y capacidad de adaptación
frente a los manejos de deshoje, manteniendo el rendimiento aún cuando se remueve
el 30% del follaje.
El deshoje podría reducir la incidencia de enfermedades, ya que crea un ambiente
mejor ventilado y con menor déficit de presión de vapor. Dicha condición podría
restringir el desarrollo de hongos patógenos como Botrytis cinerea y Cladosporium
fulvum a causa de la mejora de las condiciones ambientales (BLANCARD, 1990).
Finalmente, otro aspecto positivo de la poda podría ser una disminución en el
consumo de agua e insumos como fertilizantes y pesticidas.

40
5. CONCLUSIONES
El manejo de eliminación de una hoja por simpodio no afecta negativamente el
rendimiento, la calidad de la fruta, ni la acumulación de biomasa aérea.
El rendimiento no se vio afectado negativamente dado que la planta fue capaz de
compensar la pérdida de sus hojas.
En cuanto a la calidad del fruto dado por el nivel de sólidos solubles y pH no se
observaron diferencias significativas.
Por último, la biomasa aérea no sufrió variaciones debido a que los manejos de poda
no superaron a una hoja por simpodio.
En la medición de radiación realizada al final del ciclo productivo, se determinó que
la poda de una hoja en los simpodios uno al seis y la poda de una hoja en los
simpodios cuatro al seis genera un aumento del 30% en la radiación
fotosintéticamente activa al interior de la canopia, en comparación con el testigo.

41
6. RESUMEN
El cultivo de tomate bajo invernadero presenta limitaciones del ingreso de la luz principalmente por la alta densidad de plantación y por el sombreamiento producido por las cubiertas. El objetivo de la presente investigación fue evaluar el efecto de la poda de hojas (deshoje) sobre el ingreso de luz, productividad y calidad de la fruta. El ensayo se realizó en el Fundo La Palma perteneciente a Inversiones Quintil S.A, ubicado en la comuna de Quillota, provincia de Quillota V región, durante el período del 15 de enero al 15 julio de 2002. Se utilizó un cultivo comercial de tomate de la variedad FA-593 de crecimiento indeterminado, bajo condiciones de invernadero frío. El manejo de poda consistió en retirar una hoja de cada simpodio, con frutos de 10 mm de diámetro ecuatorial en el racimo que corona a cada simpodio. Los tratamientos consistieron en: testigo (sin deshoje), poda de los simpodios 1- 2- 3, poda de los simpodios 4-5-6, poda de los simpodios 1-2-3-4-5-6. Se evaluó la precocidad, rendimiento, calidad de la fruta, biomasa aérea, radiación fotosintéticamente activa y temperatura. El diseño experimental fue completamente al azar, con cinco repeticiones. Cada repetición consistió en un conjunto de 120 plantas (48 m2). Los resultados de los tratamientos de deshoje indican que no hubo efecto sobre el rendimiento, precocidad, distribución de calibres, ni en la calidad de la fruta (color, pH, sólidos solubles). La medición de radiación fotosintéticamente activa realizada al final del ciclo productivo fue un 30% mayor con la poda de los simpodios 1-2-3-4-5-6 y la poda de los simpodios 4-5-6 comparado con el control.

42
7. LITERATURA CITADA
ADAMS, S. R.; COCKSHULL, K. E. and CAVE, C. J. 2001. Effect of temperature on the growth and development of tomato fruits. Annals of Botany 88: 869-877 ANDRIOLO, L.; FALCAO, L.; DUARTE, T. and SKREBSKY, E. 2001.
Defoliation of greenhouse tomato plants and its effects on dry matter accumulation and distribution to fruits. Acta Horticulturae 559: 123-126
ALLENDES, H. 1992. Control de plagas y enfermedades en tomates. In: Curso producción de tomate forzado y al aire libre. Universidad Católica de Valparaíso, Facultad de Agronomía. Talca, 9 de julio de 1992. pp.A1-A17
_____________. 1992. Mano de obra. In: Curso producción de tomate forzado y al
aire libre. Universidad Católica de Valparaíso, Facultad de Agronomía. Talca, 9 de Julio de 1992. pp.A18-A23
AUNG, L. and KELLY, W. 1966. Influence of defoliation on vegetative, floral and
fruit development in tomatoes (Lycopersicon esculentum Mill). Proceedings of the American Society for Horticultural Science 89: 563-570.
BLANCARD, D. 1990. Enfermedades del tomate; observar identificar luchar. Madrid, Mundi-Prensa. 212p.
CALVERT, A. 1964. The effects of air temperature on growth of young tomato plants in natural light conditions. Proceedings of the American Society for Horticultural Science 34: 154-162.
CASTRO, M. 1991. Efecto de la densidad de plantas, el raleo de frutos y poda de
racimos sobre la producción del tomate (Lycopersicon esculentum Mill.). Taller de Licenciatura Agr. Quillota, Universidad Católica de Valparaíso, Facultad de Agronomía. 76p.
CORPORACIÓN DE FOMENTO DE LA PRODUCCIÓN-UNIVERSIDAD
CATÓLICA DE CHILE. 1986. Monografías hortícolas. Tomate, arveja, brócoli, zanahoria. Santiago, PUC-CORFO. 99p.
CHAMARRO, J. 1995. Anatomía y fisiología de la planta. In: Nuez, F. ed. El
cultivo del tomate. Madrid, Mundi-Prensa. pp.44-91

43
COCKSHULL, K.; GRAVES, C. and CAVE, C. 1992. The influence of shading on yield of glasshouse tomatoes. Journal of Horticultural Science 67: 11-24.
DECOTEAU, D. 1990. Tomato leaf development distribution as influenced by leaf
removal and decapitation. HortScience 52: 681-684. DE KONING, A. N. 1988. The effect of different day/night temperature regimes on
growth, development and yield of glasshouse tomatoes. Journal of Horticultural Science 63: 465-471.
DUIMOVIC, A. 1993. Fertirrigación en cultivos forzados con los ojos abiertos.
Método “El Maestro”. Producción de Hortalizas protegidas bajo plástico. Santiago, INIA. Noviembre 1993. pp.6.1-6.15 (Serie La Platina n°50).
ELIZONDO, R. 1999. Evaluación técnica de diferentes tipos de deshoje y
determinación de su influencia en la rentabilidad del tomate cultivado para primor, variedad FA-144, en invernadero frío. Taller de Licenciatura Agr. Quillota, Universidad Católica de Valparaíso, Facultad de Agronomía. 56p.
GRIENSON, D. and KADER, A. 1986. Fruit ripening and quality. In: Atherton, J. and Rudich, J. eds. The Tomato Crop: a scientific basis for improvement. London, Chapman & Hall. pp.241-280
HALBROOKS, M.C. and WILCOX, G.E. 1980. Tomato plant development and
elemental accumulation. Journal of the American Society for Horticultural Science 105(6): 826-828.
HO, L. 1996. Tomato In: Zamski, E.; Schaffer, A. eds. Photoassimilate distribution
in plant and crops. New York, Marcel Dekker, Inc. pp. 709-728 _____. and HEWITT, D. 1986. Fruit development. In: Atherton, J. and Rudich, J.
eds. The Tomato Crop. London, Chapman & Hall. pp.201-241 INSTITUTO NACIONAL DE ESTADÍSTICAS. 1997. VI Censo Nacional
Agropecuario. Santiago, INE. 300p. KINET, J. and PEET, M. 1997. Tomato in: H.C. Wien ed. The Physiology of
Vegetable Crops. New York, CABI. pp.207-258 ________. 1977. Effects of light conditions on the development of the inflorescence
in tomato. Scientia Horticulturae 6: 15-26.

44
LEOPOLD, A.C. and LAM, S.L. 1960. A leaf factor influencing tomato earliness. Proceedings of the American Society for Horticultural Science 76: 543-547.
MAROTO, J. 1994. Horticultura Herbácea Especial. 4ª. ed. Madrid, Ediciones
Mundi-Prensa. 611 p. MARTÍNEZ, A. 1981. Proyecto de implantación de sistemas de riegos tecnificados
en la estación experimental “La Palma”, Quillota. Tesis Ing. Agr. Quillota, Universidad Católica de Valparaíso, Facultad de Agronomía. 106p.
MARTÍNEZ, S.; GRIMALDI, M.; GARBI, M. ARTUR, M. y BELTRANO, J. 1999.
Las hojas remanentes en tomate (Lycopersicon esculentum Mill) varian su peso seco por efecto de la defoliación. Agrícola Vergel. 18(214): 676-678.
____________.; ____________.; ________. y ________. 1998. Efecto de la
defoliación en tres momentos fonológicos sobre el rendimiento en tomate (Lycopersicon esculentum Mill.) en invernadero. Agricultura Técnica (Chile) 61: 522-526.
McGUIRE, R. 1992. Reporting of objetive color measurements. HortScience 27:
1254-1255. MENDOZA, H. 1992. Nutrición y fertilización mineral en cultivo de tomate bajo
invernadero y al aire libre. In: Curso producción de tomate forzado y al aire libre. Universidad Católica de Valparaíso, Facultad de Agronomía. Talca, 9 de Julio de 1992. pp.C1-C10
_____________. 1992. Uso de hormonas y bioestimulantes en tomate bajo
invernadero. In: Curso producción de tomate forzado y al aire libre. Universidad Católica de Valparaíso, Facultad de Agronomía. Talca, 9 de Julio de 1992. pp.C11-C34
NOVOA, R.; VILLASECA, S.; Del CANTO, P.; ROUANET, J.; SIERRA, C.; Del
POZO, A. 1989. Mapa agroclimático de Chile. Santiago, INIA. 221p. OFICINA DE ESTUDIOS Y POLÍTICAS AGRARIAS. 2002. Estadísticas de
agricultura Chilena series de precios agropecuarios. (on line). www.odepa.gob.cl
OWEN, H. and AUNG, L. 1990. Genotypic and chemical influences on fruit growth
of tomato. HortScience 25: 1255-1257.

45
PEARCE, B.D.; GRANCE, R.I. and HARWICK, K. 1993. The growth of young tomato fruit. I. Effects of temperature and irradiance on fruit grown in controlled environments. Journal of Horticultural Science 68: 1-11.
PICKEN, A.; STEWART, A. AND KLAPWIJK, D. 1986. Germination and
vegetative develoment. In: Atherton, J. and Rudich, J. (eds). The Tomato Crop. London, Chapman & Hall. pp.111-166
RODRÍGUEZ, R.; TABARES, J.M. y MEDINA, J.A. 1984. El cultivo moderno del
tomate. Madrid, Mundi-Prensa. 552p. ROJAS, B. 1992. Densidades y manejo de plantas. In: Curso producción de tomate
forzado y al aire libre. Universidad Católica de Valparaíso, Facultad de Agronomía. Talca, 9 de Julio de 1992. pp.B1-B8
_________. 1992. Riego por goteo y fertirrigación en tomate. In: Curso producción
de tomate forzado y al aire libre. Universidad Católica de Valparaíso, Facultad de Agronomía. Talca, 9 de Julio de 1992. pp.B9-B18
SCHAFFER, A.; PETREIKOV, M.; MIRON, D.; FOGELMAN M.; SPIEGELMAN
M.; BNEI-MOSHE, Z. and SHEN, S. 1999. Modification of carbohydrate content in developing tomato fruit. HortScience 34: 1024-1026.
SHISHIDO, Y.; KUMAKURA, H. and HORI, Y. 1993. Changes in source-sink
interaction by defoliation and darking of sourse and sink leaf in tomato (Lycopersicon esculentum Mill.). Journal of the Japanene Society for Horticultural Science 62: 95-102.
SLACK, G. 1986. The effects of leaf removal on the tomato and yield of glasshouse
tomatoes. Journal of Horticultural Science 61: 353-360. STACEY, D. 1983. The effects of artificial deloliation on yield of tomato plants and
its relevance to pest damage. Journal of Horticultural Science 58: 117-120. TANAKA, A. and FUJITA, K. 1974. Nutriophysiological studies on the tomato
plant. IV. Source-Sink relationship and structure of the source-sink unit. Soil Science and Plant Nutrition 20: 305-315.
___________.; ____________. and KIKUCHI, K. 1974. Nutriophysiological studies
on the tomato plant III. Photosynthetic rate of individual leaves in relation to dry matter production of plant nutrition . Soil Science and Plant Nutrition 20: 173-183.

46
___________.; ____________. and SHIOYA, M. 1974. Nutriophysiological studies
on the tomato plant. II. Translocation of photosynthates. Soil Science Plant Nutrition 20: 163-171.
URRUTIA, G. 2000. Tomate para consumo fresco: mercado y perspectivas.
Agroeconómico 54: 50-54. VÁSQUEZ, C. 1994. Evaluación de sistema MAFF-modificado de fertilización
versus sistema de fertilización tradicional en dos sistemas de plantación, y confección de curvas de extracción del cultivo de tomate en ambos sistemas. Taller de Licenciatura Agr. Quillota, Universidad Católica de Valparaíso, Facultad de Agronomía. 106p.
WOLK, J.O.; KRETCHMAN, D.W. and ORTEGA, D.G. 1983. Response of tomato
to defoliation. Journal of the American Society for Horticultural Science 108: 536-540.

47
ANEXOS


















