
Variabilidad radial de algunos caracteres anatómicos de...
12
Variabilidad radial de algunos caracteres anatómicos de Aspidosperma quebracho blanco Este estudio determina el patrón de variación de los caracteres anatómicos estructurales del leño en Aspidosperma quebracho blanco, una de las especies de mayor importancia comercial del Parque Chaqueño Seco, Argentina. Se determinó la incidencia de la fuente de variación sitio, árbol, y distancias radiales sobre la variabilidad de los caracteres estructurales densidad básica, número y diámetro de poros, largo de vasos y fibras. El diseño consistió en un muestreo multietápico con submuestras de orden jerárquico. Para el análisis estadístico se utilizó un modelo anidado con efecto aleatorio en sitio y árboles y fijo en distancias radiales a la médula. La estimación de los componentes de varianza fue realizada por el método de Máxima Verosimilitud Restringida. Los resultados mostraron que la distancia a la médula es la fuente de variación más importante en los caracteres estudiados. El sitio no tiene influencia excepto en la longitud de las fibras. La longitud de fibras aumenta rápida- mente en dirección a la corteza, evidenciando la presencia de leño juvenil. La densidad básica no mostró ser un índice eficiente para valorar la calidad de la madera. 3 Palabras claves: xilema, variación, radial, quebracho blanco. RESUMEN Key words: Wood, variation, radial, quebracho blanco. Radial wood variation patterns of Aspidosperma quebracho blanco, one of the most important species of the Dry Chaco Regi on, and their influence on wood quality are reported in this study. Pattern variations of wood density, frequency and diameter of pore vessels and fiber length were examined using sites, trees and radial distances as sources of variation. Univariate analysis was performed using a nested design, with random effects on sites and trees and fixed distances to the pith. Variance Components were estimated by Restricted Maximum Likelihood. The results indicated that distance to the pith was the most important source of variation for the traits studied. With the exception of fiber length, the wood anatomical features measured were not affected by site. Fiber length increased rapidly from pith to bark and defined the juvenile wood zone. Wood density was not found to be a good indicator of wood quality due to the great variability between trees. SUMMARY ARTICULOS Radial wood variation in some anatomical features in Aspidosperma quebracho blanco JUANA GRACIELA MOGLIA, CARLOS RAUL LOPEZ Instituto de Silvicultura y Manejo de Bosques, Facultad de Ciencias Forestales, Universidad Nacional de Santiago del Estero, Avda. Belgrano 1912 (S), Cp. 4200 Santiago del Estero, Argentina. Email: [email protected] BOSQUE 22(2): 3-14, 2001
Transcript of Variabilidad radial de algunos caracteres anatómicos de...

Variabilidad radial de algunos caracteres anatómicos de Aspidosperma quebracho blanco
Este estudio determina el patrón de variación de los caracteres anatómicos estructurales del leño en Aspidosperma quebracho blanco, una de las especies de mayor importancia comercial del Parque Chaqueño Seco, Argentina. Se determinó la incidencia de la fuente de variación sitio, árbol, y distancias radiales sobre la variabilidad de los caracteres estructurales densidad básica, número y diámetro de poros, largo de vasos y fibras. El diseño consistió en un muestreo multietápico con submuestras de orden jerárquico. Para el análisis estadístico se utilizó un modelo anidado con efecto aleatorio en sitio y árboles y fijo en distancias radiales a la médula. La estimación de los componentes de varianza fue realizada por el método de Máxima Verosimilitud Restringida. Los resultados mostraron que la distancia a la médula es la fuente de variación más importante en los caracteres estudiados. El sitio no tiene influencia excepto en la longitud de las fibras. La longitud de fibras aumenta rápidamente en dirección a la corteza, evidenciando la presencia de leño juvenil. La densidad básica no mostró ser un índice eficiente para valorar la calidad de la madera.
3
Palabras claves: xilema, variación, radial, quebracho blanco.
RESUMEN
Key words: Wood, variation, radial, quebracho blanco.
Radial wood variation patterns of Aspidosperma quebracho blanco, one of the most important species of the Dry Chaco Region, and their influence on wood quality are reported in this study.
Pattern variations of wood density, frequency and diameter of pore vessels and fiber length were examined using sites, trees and radial distances as sources of variation. Univariate analysis was performed using a nested design, with random effects on sites and trees and fixed distances to the pith. Variance Components were estimated by Restricted Maximum Likelihood. The results indicated that distance to the pith was the most important source of variation for the traits studied. With the exception of fiber length, the wood anatomical features measured were not affected by site. Fiber length increased rapidly from pith to bark and defined the juvenile wood zone. Wood density was not found to be a good indicator of wood quality due to the great variability between trees.
SUMMARY
A R T I C U L O S
Radial wood variation in some anatomical features in Aspidosperma quebracho blanco
JUANA GRACIELA MOGLIA, CARLOS RAUL LOPEZ
Instituto de Silvicultura y Manejo de Bosques, Facultad de Ciencias Forestales, Universidad Nacional de Santiago del Estero,
Avda. Belgrano 1912 (S), Cp. 4200 Santiago del Estero, Argentina. Email :[email protected]
BOSQUE 22(2): 3-14, 2001

En zonas áridas y semiáridas, donde la hume
dad es la principal causante del crecimiento cícli
co anual, la actividad cambial está influida más
por el régimen de las lluvias que por la temperatu
ra (Fhan 1964).
En Argentina Aspidosperma quebracho blanco
(Schlecht) Apocinaceae es una de las especies de
mayor importancia comercial en el Parque Cha-
queño Seco. Es la especie más difundida de la
Región del Chaco, y ocupa el 3 8 % de un total de
8 millones de has de bosque nativo, extendiéndose
también a la zona de transición con el Monte y a
la Mesopotamia (Dimitri et al., 1997). Se encuen
tra entre las especies nativas que aportan el mayor
volumen de material aserrado.
Aunque el quebracho blanco ha sido util izado
predominantemente para la elaboración de los pro
ductos tradicionales (carbón, leña y durmientes) ,
el color y veteado de su madera jun to al acabado
ópt imo en las piezas trabajadas la colocan entre
las más interesantes para uso en carpintería, parqués
y tornería.
O B J E T I V O S . De te rminar el patrón de var iación de
los caracteres estructurales del leño en sent ido
radial .
• Determinar la incidencia de los árboles y el si
tio sobre la variabilidad de los caracteres es
tructurales del leño.
• Estudiar la adecuación de la densidad básica
como probable indicador de la calidad de la
madera.
M A T E R I A L E S Y M E T O D O S
Se utilizaron 10 ejemplares adultos de Aspi
dosperma quebracho blanco, con diámetros supe
riores a los 30 cm, extraídos de dos sitios de la
Región Chaqueña Seca, Santiago del Estero, Ar-
g e n t i n a .
C A R A C T E R I Z A C I Ó N D E L Á R E A D E E S T U D I O El área
de estudio se inscribe en la Región Chaqueña .
Según Hueck (1978), el Chaco es una gigantesca
región de 800.000 k m 2 cubierta por el bosque seco
ininterrumpido más grande del territorio sudame
ricano. El cl ima pertenece al t ipo subtropical de
estepas y montes esteparios de t ipo continental con
veranos muy cal ientes e inviernos fríos y con
4
La industria de la madera requiere materia pri
ma uniforme para la elaboración de productos de
calidad. Su aptitud c o m o materia prima está aso
ciada con el grado de variabilidad de los caracte
res anatómicos estructurales. En consecuencia, es
necesario conocer los patrones de variación anató
micos estructurales y su relación con las fuentes
de variación ambientales .
La identificación de la variabilidad y su ade
cuación a los estándares de calidad de los usuarios
es de fundamental importancia para poder optimizar
el uso de la madera de acuerdo a las necesidades
industriales. Este logro mejora la eficiencia de la
materia pr ima y el control de calidad del producto
(Zobel y Van Buijtenen, 1989).
En general, las propiedades y características de
uso de una madera pueden definirse de manera
satisfactoria mediante pruebas directas, pero debi
do a que son costosas y en muchos casos destruc
tivas, solo pueden ser aplicadas en escala reduci
da. Sin embargo, hay ciertos rasgos de la madera
que son buenos indicadores de sus propiedades y
usos, y que permiten ser utilizados como índices
de calidad (Burley y Wood , 1979).
La densidad básica identifica la calidad de una
madera y permite est imar su dureza, porosidad,
compor tamiento de los esfuerzos mecánicos, ex
tensión de los cambios dimensionales , calidad del
acabado y rendimiento. Por esta razón se la em
plea como criterio de evaluación, permit iendo pre
decir las posibil idades de uso y la calidad de los
productos a obtener (Burley y Wood , 1979).
La densidad básica está influida por el espesor
y estructura de los anillos, t amaño de las células,
espesor de la pared y proporción de los diferentes
tejidos que conforman la estructura de la madera
(Zobel y Talbert , 1984). Deb ido a esto, son tam
bién rasgos claves cuya medición y evaluación de
su variabilidad pueden servir como índices de la
calidad de la madera (Burley y Wood , 1979).
I N T R O D U C C I O N
La estructura de la madera varía entre espe
cies, dentro de cada especie, entre árboles, y aun
dentro de cada ejemplar, además de las fuentes
geográficas (Zobel y Van Buijtenen, 1989). Por lo
que la estructura de la madera es notablemente
heterogénea como resultado de la influencia de
factores intrínsecos (genéticos) y extrínsecos (am
bientales). La naturaleza e intensidad de la influen
cia de estos factores sobre los elementos anatómi
cos pueden diferir según la especie o el género.
JUANA GRACIELA MOGLIA, CARLOS RAUL LOPEZ
períodos de lluvia y de sequía bien delimitados
(Boletta et al, 1989).
La vegetación dominante es bosque subtropical
de xerófilos. El quebrachal prevalece como comu
nidad cl imáxica, con Schinopsis quebracho colo
rado (quebracho colorado santiagueño) y Aspi
dosperma quebracho blanco (quebracho blanco).
En la actualidad estos bosques se encuentran apro
vechados en diferentes intensidades, existen áreas
a) Campo experimental La María (Sitio 1). Este
sitio se encuentra ubicado a 27 km de la ciudad de
Sant iago del Estero. Las precipitaciones se con
centran en diciembre, enero y febrero y el balance
hídrico es negat ivo en todos los meses del año
(Juárez, 1998). Las precipitaciones t ienen una dis
tr ibución más uniforme que en el Sitio 2, con
máx imas en dic iembre y en marzo . Los suelos se
clasifican como Regosols ( F A O - U N E S C O , 1988)
ci tado por Lorenz (1995). La altitud promedio es
de 190 m, con una leve pendiente hacia el Oeste.
La textura del suelo corresponde a las clases fran
co l imoso y franco arenoso (Lorenz, 1995).
b) El Desierto (Sitio 2). Se encuentra en el De
par tamento Copo a 400 km al NE de la c iudad de
Santiago del Estero. Las precipitaciones se encuen
tran concentradas en el pr imer tr imestre y el ba
lance hídrico es negat ivo en todos los meses del
año (Juárez, 1998). La prec ip i tac ión t iene dos
máximas correspondientes a d ic iembre (más acen
tuado) y marzo .
Los suelos se ubican en la Paleo-Planicie Aluvial
del Río Salado, y se clasifican como Moll isoles .
El estrato arbustivo contiene mayor número de
especies que en el La María .
Las características cl imáticas del Sitio 1 y 2 se
detallan en el cuadro 1:
5
muy afectados por el fuego y ya no existen rodales
vírgenes (Hueck, 1978).
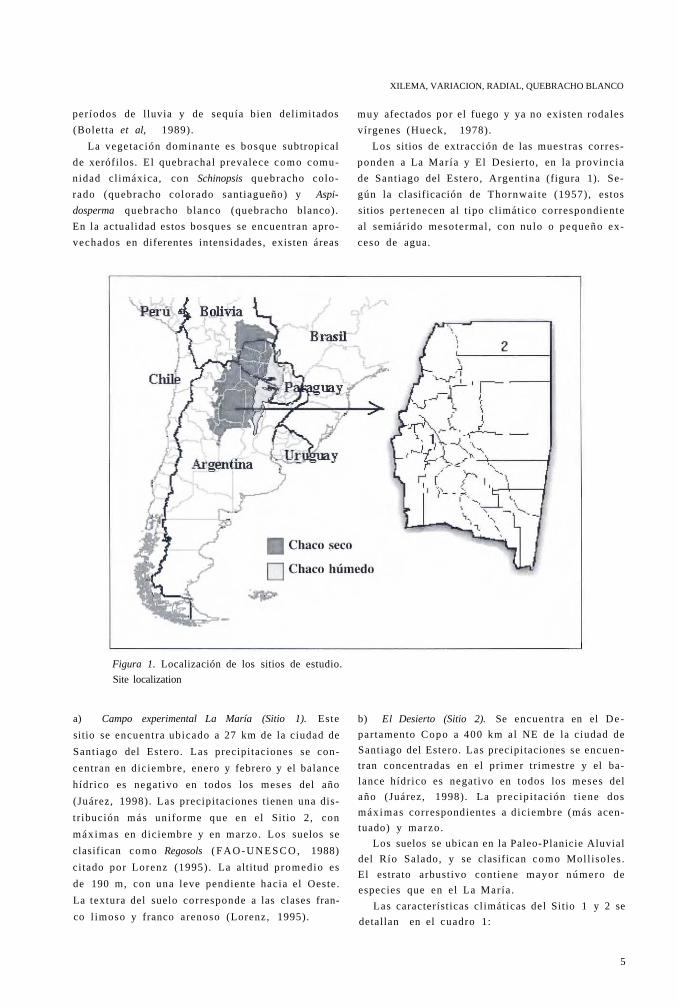
Los sitios de extracción de las muestras corres
ponden a La Mar ía y El Desierto, en la provincia
de Santiago del Estero, Argent ina (figura 1). Se
gún la clasificación de Thornwai te (1957), estos
sitios pertenecen al t ipo cl imático correspondiente
al semiárido mesotermal , con nulo o pequeño ex
ceso de agua.
Figura 1. Localización de los sitios de estudio.
Site localization
XILEMA, VARIACION, RADIAL, QUEBRACHO BLANCO

CUADRO 1
Datos climáticos La María, a 28° 03' LS y 64° 15' LO, 169 m s.n.m. (Sitio 1) y de Los Pirpintos, a 20° 75' LS y 62° 04' LO (Sitio 2).
C l i m a t i c da ta of La M a r í a , a 28° 0 3 ' LS y 64° 15' L O , 169 m s .n .m. (Si te 1) a n d
L o s P i rp in tos a 2 0 ° 7 5 ' LS y 6 2 ° 0 4 ' LO (Si te 2 ) .
Temperatura media del mes
más cálido (Enero)
Temperatura media del mes
más frío (Julio)
Precipitación anual
Evapotrans-piración
anual
Déficit anual
aproximado
Sitio 1 26,1° C 10,6° C 579 ram 1011 432 mm
Sitio 2 28,0° C 15,6° C 614 mm 1174 560 mm
Fuente: E . E . A . - I N T A . La M a r í a S a n t i a g o del Es t e ro y D i r ecc ión de R e c u r s o s H í d r i c o s de S a n t i a g o del E s t e r o .
Para la obtención de las muestras se escogieron
10 individuos adultos y sanos, sin daños visibles y
de fuste recto, según las recomendaciones de Burley
y Wood (1979). Los ejemplares seleccionados co
rresponden a los árboles dominantes debido a su
sensibilidad a los factores del ambiente.
En cada individuo se midió altura total y altura
de fuste, considerada hasta la pr imera ramifica
ción, y el diámetro a 1,30 m.
Se extrajeron dos probetas de 2 cm de arista,
sobre el radio Norte de la sección transversal, a
1,30 m de altura. Una de ellas se destinó para la
determinación de la densidad básica y la otra para
los preparados microscópicos . Se tomaron cinco
p robe ta s a par t i r de 2 cm d e s d e la m é d u l a
Butterfield et al. (1993), y dejando un cm entre
cada una de las muestras .
La densidad básica, se determinó según la Nor
ma I R A M 9544 (1977), a 12% de humedad.
O B T E N C I O N D E P R E P A R A D O S M I C R O S C O P I C O S . Las
probetas fueron hervidas cuatro horas por día,
durante c inco días, para su ablandamiento. Las
secciones obtenidas se t iñeron con coloración
triple de acridina-crisoidina y azul de astra. Luego
de deshidratarlas, en una secuencia creciente de
alcoholes, se montaron en entellán.
Los macerados se prepararon colocando astillas
de madera en una parte de ácido acético glacial y
dos partes de peróxido de hidrógeno, según la téc
nica de Berlyn y Mishke (1976). Posteriormente
fueron t e ñ i d o s c o n sa f ran ina y m o n t a d o s en
entellán. Las descripciones microscópicas se rea
lizaron de acuerdo a las recomendaciones del Co
mité de I A W A (International Association of W o o d
Anatomists , 1989).
Las variables utilizadas fueron:
1. Densidad básica.
2. Frecuencia de poros (número de poros por
m m 2 ) .
3. Diámetro tangencial de poros .
4. Longitud de vasos.
5. Longitud de fibras.
A N A L I S I S E S T A D I S T I C O D E L O S D A T O S
Análisis descriptivo. Fueron est imadas las medi
das de dispersión, tendencia central y el análisis
exploratorio de datos para comprobar los supues
tos de análisis paramétr icos y aumentar la eficien
cia de la prueba de " F " .
Análisis de la varianza. Se realizó mediante un
modelo mixto, con efecto aleatorio de sitios y ár
boles, y fijo de distancias a la médula .
El muestreo fue mult ietápico con submuestras de
orden jerárquico, Van Lar (1991) . El proceso de
muestreo comprende las variables clasificatorias
"sit ios", "árboles" y "distancias radiales" anidadas
en forma sucesiva.
El modelo lineal aditivo correspondiente es:
JUANA GRACIELA MOGLIA, CARLOS RAUL LOPEZ

si = efecto del i és imo sitio.
A i j = efecto del j és imo árbol dentro del i és imo sitio.
Dijk = efecto del k és ima distancia, dentro del j
és imo árbol dentro del i és imo sitio.
La participación porcentual de las variables cla-sificatorias sobre la variabilidad de los caracteres estudiados fue calculada en base a los componentes de varianza est imados por el método de Máxi ma Verosimil i tud Restr ingida (SAS. V.G.12) .
R E S U L T A D O S Y D I S C U S I O N
Análisis descriptivo. En el cuadro 2 se muestran las medidas de posición y dispersión de las variables analizadas por t ratamiento y sitio.
CUADRO 2
Promedios y coeficientes de variación (C.V.) por distancias a la médula de las variables analizadas.
M e a n s a n d var ia t ion coeff ic ient ( C . V . ) o f the ana l i zed va r iab les by d i s t ance to the p i th
Variables DI D2 D3 D4 D5
Densidad básica Media 0,809 0,813 0,813 0,792 0,784 Densidad básica
C.V. 5,40 6,65 5,58 6,03 5,86
Número de poros Media 15,04 9,67 8,27 7,80 7,41 Número de poros
C.V. 25,44 23,63 16,88 5,98 16,68
Diámetro de poros Media 78,74 82,39 87,18 87,89 99,39 Diámetro de poros
C.V. 30,48 16,46 15,19 12,28 11,96
Largo de vasos Media 286,28 298,48 319,32 341,51 348,23 Largo de vasos
C.V. 5,77 9,55 10,60 8,61 10,29
Largo de fibras Media 1.221,36 1.376,38 1.485,36 1.447,82 1.534,49 Largo de fibras
C.V. 5,90 9,87 8,25 5,41 5,23
7
Las variables Número de poros y Diámetro de
poros fueron procesadas uti l izando su transforma
ción logarítmica.
Los Coeficientes de Variación (CV) resultaron
bajos especialmente en el largo de fibras y la
densidad básica y son similares a los encontrados
por Helinska Rackzkowska y Fabisiak (1991) y
Manwil ler (1974) en Quercus.
Análisis de la Varianza (ANOVA). En el cuadro 3
se mues t ran los r e su l t ados de los anál i s i s de
varianza de los caracteres estudiados. Sólo existen
diferencias significativas entre sitios para la longi
tud de fibras, lo que denota la poca influencia de
esta fuente de variación en las respuestas de las
variables analizadas.
XILEMA, VARIACION, RADIAL, QUEBRACHO BLANCO

CUADRO 3
Resultados del ANOVA aplicados a las características estudiadas.
A N O V A results of studied characteristics.
Variables Fuente de variación
Variables Sitio Arboles
Densidad n.s. **
Número de poros n.s. n.s.
Diámetro de poros n.s. *
Longitud de vasos n.s. *
Longitud de fibras * n.s.
n.s. = no significativo * = significativo al 95% de probabilidades ** = significativo al 99% de probabilidad.
La densidad básica no muestra diferencias sig
nificativas entre sitios. Esto no concuerda con los
estudios de Zhanner (1968), citado por Zobel y
Van Buijtenen (1989), que establecen que el sitio
tiene influencia sobre la densidad en las especies
de porosidad difusa de alta densidad como que
bracho blanco. Según este autor, en los sitios con
mayor humedad se forma mayor cantidad de va
sos sobre una matriz con fibras de paredes más
delgadas, lo que produce densidad básica más baja.
La longitud de fibras fue la única variable que
mostró diferencias significativas entre sitios, sien
do un 8% más largas en el sitio que tiene menor
disponibilidad de agua de acuerdo a su balance
hídrico. Esta aparente contradicción puede deber
se a la forma de crecimiento en longitud de las
fibras. La menor velocidad de crecimiento permite
la e longación de las células antes de volver a
multiplicarse. Si la velocidad de crecimiento es
rápida, la célula madre producida por la célula
cambial inicial recibe el est ímulo para dividirse
antes de agrandarse (Larson, 1994). En este sen
tido, He l in ska -Rackzkowska y Fabis iak (1991)
encontraron en Quercus que los árboles dominan
tes, cuyo crecimiento es mayor que los suprimi
dos, tienen fibras significativamente más cortas.
Sin embargo Baas (1986) encontró en Quercus
que el crecimiento muy lento produce reducción
de los e lementos .
Si bien no hay diferencias significativas en los
elementos de conducción, el diámetro de poros es
un 12%, menor en el Sitio 2, que tiene menor
disponibilidad de agua. Esto concuerda con Tyree
et al. (1984) y Tyree y Sperry (1989), que sostie
nen que a nivel intraespecífico existe correlación
negativa entre el diámetro de poros y la pérdida de
conductividad hidráulica por efecto de la fuerte
tensión hídrica que provoca la aridez.
La disminución del diámetro de poros con el
défici t h íd r ico ha s ido desc r i t a por Ca r lqu i s t
(1977), Barajas-Morales (1985), Woodcock e Ignas
(1994). La respuesta insignificante de las varia
bles anatómicas del quebracho blanco con el sitio
puede deberse a la presencia de un sistema subsi
diario de conducción (traqueidas vasicéntricas), que
hacen innecesario una modificación acentuada del
sistema vascular.
Las variables densidad, diámetro de poros y lon
gitud de vasos, muestran diferencias significativas
entre árboles dentro de cada sitio. Respecto a la
densidad básica, estos resultados concuerdan con
Panshin y De Zeew (1980), Zobel y Van Buijtenen
(1989), Zhang y Zong (1990), quienes encontraron
que la mayor proporción de variación de la densi
dad básica se debe a la diferencia entre árboles.
Lei et al. (1996) no encontraron diferencias sig
nificativas entre árboles en las variables longitud
de fibras y diámetro de vasos. La mayor causa de
variación para estas variables se encuentra dentro
del árbol en función de la edad.
Componentes de varianza. Las varianzas de las
variables clasificatorias es t imadas para cada ca
rácter se muestran en la Cuadro 4. La representa
ción porcentual de la variación asociada a cada
fuente permite catalogar la incidencia de las mis
mas a la variabilidad del carácter en estudio.
Varianza explicada por cada fuente de variación expresado en porcentaje.
Explained variance, in percentage, for each source of variation.
Variables Sitio Arboles Distancia
Densidad 0,0 82,1 17,9
Número de poros 2,6 1,4 96,0
Diámetro de poros 7,6 26,0 66,4
Longitud de vasos 0,0 19,4 80,6
Longitud de fibras 20,7 3,0 76,3
8
CUADRO 4
JUANA GRACIELA MOGLIA, CARLOS RAUL LOPEZ

Densidad básica. Este carácter no se ve afectado por el sitio y su variabil idad está más expresada por las diferencias entre árboles que por las distancias a la médula (figura 2). Aeckermann (1995) confirma que la causa de la variabilidad de la densidad básica en Quercus, a edades fijas en 7 sitios se debe a la diferencia entre árboles.
Ferreira y Kageyama (1994) encontraron en Eucalyptus grandis que el efecto de la edad (asimilable a distancias) es más importante que el efecto de sitios en la variación de la densidad básica.
Figura 2 Porcentaje de variación explicada por los efec
tos del modelo
Variance percentage explained by the model
La tendencia decreciente en sentido radial de densidad básica de quebracho blanco se muestra en la figura 3. Esta tendencia es similar a la encontrada por Coronel (1984) en esta misma especie. Este decrecimiento radial puede deberse a que el duramen contiene extractivos y material incrustante que aumenta su densidad. De acuerdo a Besold et al. (1988), el contenido de extractivos del quebracho blanco es de 7 ,7%.
Corone l (1984) encont ró diferencias significa
t ivas para la dens idad bás ica en quebracho b lan
co entre la zona interna (0,905 k g / m 3 ) , y (0 ,867
k g / d m 3 ) en la zona externa, basándose en las dife
rencias entre albura y duramen. Los valores calcu
lados por este autor son mayores que los encontra
dos en el presente trabajo.
Los patrones de variación de la densidad bási
ca, en sentido radial, han sido ampl iamente estu
diados en coniferas. En latifoliadas hay mayor va
riabilidad en los caracteres estructurales del leño,
por lo que pueden encontrarse diversos patrones
de variación de densidad. Según Kol lman (1959),
el valor máx imo de densidad básica en latifoliadas
se encuentra en el centro del t ronco. Este autor
señala que en maderas de porosidad difusa, espe
cialmente de densidad básica baja como las del
género Populus, existe una tendencia creciente de
densidad en los pr imeros años y luego d isminuye.
E s t e m i s m o c o m p o r t a m i e n t o s e e n c o n t r ó e n
Quercus y en haya (Kollman, 1959).
La tendencia decreciente observada en Aspidos
perma quebracho blanco también fue encontrada
en Populus por Yanchuck et al. (1983), y en Acer
sacharium por Saydak (1967), ci tados por Peszlen
(1994), y en Hyeronima y Vochysia por Butterfield
et al. (1993).
Número de poros. En este carácter hay gran inci
dencia de las diferencias entre distancias dentro
del individuo sobre su variabilidad. La incidencia
de las diferencias entre árboles y sitio es insignifi
cante, según se observa en el figura 4.
9
Figura 3. Variación radial de la densidad básica.
Radial variation of basic density
Figura 4. Porcentaje de variación explicada por los efec
tos del modelo.
Variance percentage explained by the model
XILEMA, VARIACION, RADIAL, QUEBRACHO BLANCO
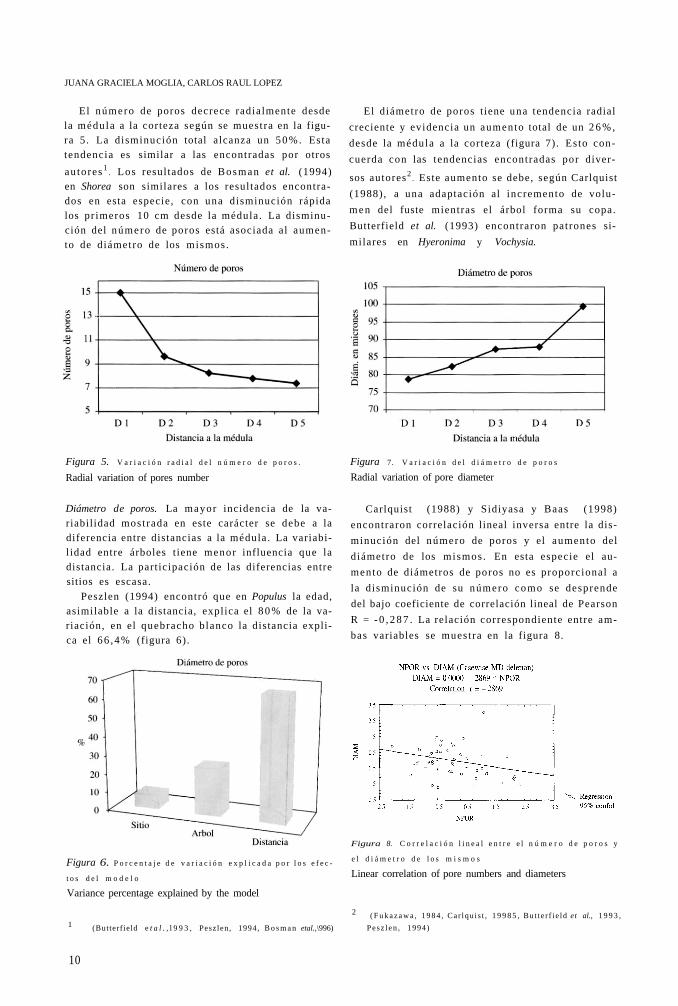
El número de poros decrece radialmente desde la médula a la corteza según se muestra en la figura 5. La disminución total alcanza un 5 0 % . Esta tendencia es similar a las encontradas por otros
au tores 1. Los resultados de Bosman et al. (1994)
en Shorea son similares a los resultados encontrados en esta especie, con una disminución rápida los pr imeros 10 cm desde la médula . La disminución del número de poros está asociada al aumento de diámetro de los mismos .
El diámetro de poros t iene una tendencia radial
creciente y evidencia un aumento total de un 2 6 % ,
desde la médula a la corteza (figura 7). Esto con
cuerda con las tendencias encontradas por diver
sos autores2. Este aumento se debe, según Carlquist
(1988), a una adaptación al incremento de volu
m e n del fuste mientras el árbol forma su copa.
Butterfield et al. (1993) encontraron patrones si
milares en Hyeronima y Vochysia.
Diámetro de poros. La mayor incidencia de la variabilidad most rada en este carácter se debe a la diferencia entre distancias a la médula . La variabil idad entre árboles t iene menor influencia que la distancia. La part icipación de las diferencias entre sitios es escasa.
Peszlen (1994) encontró que en Populus la edad, asimilable a la distancia, explica el 8 0 % de la variación, en el quebracho blanco la distancia explica el 66 ,4% (figura 6).
1 (Butterf ield e t a l . , l 9 9 3 , Peszlen, 1994, B o s m a n etal.,\996)
Carlquist (1988) y Sidiyasa y Baas (1998)
encontraron correlación lineal inversa entre la dis
minución del número de poros y el aumento del
diámetro de los mismos . En esta especie el au
mento de diámetros de poros no es proporcional a
la disminución de su número como se desprende
del bajo coeficiente de correlación lineal de Pearson
R = - 0 , 2 8 7 . La relación correspondiente entre am
bas variables se muest ra en la figura 8.
2 ( F u k a z a w a , 1 9 8 4 , Ca r lqu i s t , 1 9 9 8 5 , Bu t t e r f i e ld et al., 1 9 9 3 ,
P e s z l e n , 1994)
10
Figura 8. C o r r e l a c i ó n l i n e a l e n t r e e l n ú m e r o d e p o r o s y
e l d i á m e t r o d e l o s m i s m o s
Linear correlation of pore numbers and diameters Figura 6. P o r c e n t a j e d e v a r i a c i ó n e x p l i c a d a p o r l o s e f e c
t o s d e l m o d e l o
Variance percentage explained by the model
Figura 7. V a r i a c i ó n d e l d i á m e t r o d e p o r o s
Radial variation of pore diameter
Figura 5. V a r i a c i ó n r a d i a l d e l n ú m e r o d e p o r o s .
Radial variation of pores number
JUANA GRACIELA MOGLIA, CARLOS RAUL LOPEZ
Administrador
Línea
Administrador
Línea

Longitud de vasos. Existe gran part icipación de
las distancias a la médula en la expresión de la
variabil idad en este carácter. Hay escasa variación
entre árboles y ninguna entre sitios, como se ob
serva en la figura 9.
La longitud de vasos tiene un patrón de varia
ción creciente en sentido radial (figura 10) con
ascenso total de 17%. Este aumento no es propor
cional al evidenciado por el diámetro de los mis
mos y concuerda con lo observado por Fukazawa
(1984) y Carlquist (1988).
La longitud de los vasos es similar a la longitud
de las células cambiales fusiformes de las que
derivan, sobre todo en especies de porosidad difu
sa como el quebracho blanco, similar a lo obser-
vado por Chalk y Chat taway (1934) ci tado por
Larson (1994). A medida que el árbol crece sus
células t ambién lo hacen para conduci r m a y o r
vo lumen de agua (Larson, 1994). El pat rón de
variación radial de los e lementos vasculares es
menos acentuado que el de las fibras, y concuerda
con lo observado por León y Espinoza de Pernía
(1998) en Cordia thaisiana.
Longitud de fibras. En este carácter, la diferencia
entre distancias dentro del individuo es la princi
pal causa de variación, aunque también t ienen in
cidencia las diferencias entre sitios (figura 11).
Peszlen (1994) encontró en Populus que la edad
explica el 7 6 % de la variación para esta variable.
En la figura 12 se observa la tendencia crecien
te en la longitud de fibras.
11
Figura 9. Porcentaje de variación explicada por los efec
tos del modelo.
Variance percentage explained by the model
Figura 11. Porcentaje de variación explicada por los
efectos del modelo.
Variance percentage explained by the model
Figura 1 2 . Variación radial de la longitud de fibras.
Radial variation of fibre length.
Figura 10. Variación radial de la longitud de vasos.
Radial variation of vessels length.
XILEMA, VARIACION, RADIAL, QUEBRACHO BLANCO

I sma i l et al. ( 1 9 9 5 ) en Neolanckia cadamo Antrocephalus
chinensis; en Cupresus sempervirens P a r e s k e v o u p o u l o u
( 1 9 9 1 ) . En Populus ( J a m e et al, 1 9 4 3 ; S c a r a m u z z i , 1 9 5 5 ;
I n o k u m a et al., 1 9 5 6 ; B o c e Ka i se r , 1 9 6 1 ; M a r t o n et al.,
1 9 6 8 ; H o l t y M u r p h e , 1 9 7 8 ; M u r p h e e t al, 1979 ; Y a n c h u k
et al., 1984) , t o d o s c i t ados po r P e s z l e n ( 1 9 9 4 ) .
Teniendo en consideración la diferencia de as
censo rápido, luego más lento en la tendencia de
ascenso en la longitud de fibras, es posible distin
guir la presencia de madera juveni l en el quebra
cho blanco. La fase de ascenso rápido se eviden
cia hasta la distancia D 3 , que corresponde a un
rango de 9,5 a 10,5 cm de distancia de la médula ,
que corresponden a edades que varían entre 31 a
40 años, según el árbol considerado. Aunque esta
edad es avanzada si se compara con aquellas en
contradas en Pinus y otras especies (10 a 14 años),
es similar a la encontrada en Quercus (Helinska -
Raczkowska y Fabisiak, 1991), donde la madera
juvenil se prolonga hasta los 30 años, y que coin
cide con la edad de madurez sexual de esta espe
c ie . T a m b i é n o t ros a u t o r e s 4 d e t e r m i n a r o n en
Quercus que la zona juveni l comprende 30 a 40
años, basándose también en la variación en senti
do radial de la longitud de fibras, resultado similar
al encontrado en quebracho blanco. Lei et al. (996)
encontraron que el pasaje de madera juveni l a
adulta (determinado por el cambio de pendiente
de la curva) variaba entre 10 y 26 años en Quercus
garrayana, dependiendo del e lemento anatómico
considerado.
En latifoliadas, la presencia de madera juveni l
es difícil de determinar y depende de la especie.
Las especies más longevas forman madera juveni l
durante un t iempo más prolongado, y sus cambios
no son tan severos como en aquellas especies de
rápido crecimiento (Zobel y Van Buijtenen, 1989).
Así, Butterfield et al. (1993) no encontraron defi
nición de clara de madera juveni l en Hyeronima
alchornoides y Vochysia guatemaltensis. En ála
mos se encontró un rápido ascenso de la longitud
de fibras en los primeros 10 a 20 años y luego una
disminución que evidenciaba la presencia de ma
dera juven i l 5 .
En los gráficos de tendencias de las figuras se
observa que las tendencias de las variables, n ú m e
ro y diámetro de poros, longitud de vasos, tam
bién cambian su pendiente en la distancia D3 (9 a
11 cm). Hasta las distancias (16 a 18 cm de radio),
que corresponden a edades alrededor de los 72
años y según se observa en los gráficos, no pudo
observarse la estabilización de los e lementos , cita-
4 H a m i l t o n , 1 9 6 1 ; F a r m e r , 1 9 6 9 ; T a y l o r , 1 9 7 9 ; P e t r i c y
Scukanec , 1980; F u r u k a w a et al, 1983 , c i tados por He l in ska -
R a c z k o w s k a y F a b i s i a k ( 1 9 9 1 ) . 5 S c a r a m u z z i , 1 9 5 5 ; B o y c e y K a y s e r , 1 9 6 1 , c i t a d o s p o r
P e s z l e n (1994 ) .
12
El incremento total en la longitud de fibras en
quebracho blanco es de 20 ,4%. Hel inska-Rackz-
kowska y Fabisiak (1991) encontraron resultados
similares en Quercus donde la madera madura
presenta longitudes de fibras y vasos, entre 10 a
2 0 % mayores que las de la madera juvenil . En
Triplochiton scleroxlon, Teng A m o o k o et al, cita
dos por Zobel y Van Buijtenen (1989), encontra
ron aumento de 3 0 % para longitud de fibras mien
tras que Butterfield et al. (1993) observaron au
mentos de 2 7 7 % en Hyeronima alchornoides.
Entre los promedios D1 y D2 y D2 y D3 la
longitud de las fibras incrementa 7% y 5,6% res
pect ivamente. En géneros como Quercus, Fagus,
Fraxinus se observaron patrones ascendentes con
tinuos hasta los 100 años (Zobel et al, 1989).
El aumento evidenciado por la longitud de fi
bras es más acentuado que el de longitud de va
sos. Lei et al. (1996) y Bosman et al. (1994) en
contraron resu l tados s imi lares . Es to se debe a
que mientras los vasos siguen la tendencia de cre
cimiento de las células iniciales fusiformes, las
fibras se elongan poster iormente debido a su cre
cimiento intrusivo (Larson, 1994).
La tendencia creciente de la longitud de fibras
con respecto a las distancias a la médula como la
de quebracho blanco fue observada por diferentes
autores 3 , y sigue un patrón similar al de la longi
tud de vasos. Según Zobel y Van Buijtenen (1989),
las fibras t ienen el pat rón de variación más con
sistente entre los e lementos ana tómicos en la ma
dera.
De las variables consideradas en este análisis,
la mayoría tienen c o m o fuente de variación princi
pal un porcentaje superior al 7 0 % , a la distancia a
la médula . Esto reafirma lo encontrado por diver
sos autores como Panshin y De Zeew (1980); Zobel
y Van Buijtenen (1989) que sostienen que la ma
yor fuente de variación se encuentra en el árbol
mismo.
La fuente de variación árbol resultó ser impor
tante en la densidad básica, lo que implica que
este parámetro no es adecuado y no puede ser
utilizado como un índice de calidad, debido a la
alta variabilidad entre árboles y su escasa variabi
lidad en sentido radial .
3
JUANA GRACIELA MOGLIA, CARLOS RAUL LOPEZ
Administrador
Línea
Administrador
Línea

da por varios autores como característica de la
madera madura .
Giménez (1998) encontró en Schinopsis que
bracho colorado que los caracteres estructurales
no se estabil izaban aun a los 70 años de edad.
Coincidentemente, los componentes de varianza
más importantes de la densidad básica y del por
centaje de fibras (Moglia 2000) son las diferencias
entre árboles.
La causa de la escasa variabilidad de la densi
dad básica, en esta especie, puede atribuirse justa
mente a que sigue la misma tendencia del porcen
taje de fibras que no muestra tendencia definida
en función de la distancia radial (Moglia, 2000) .
C O N C L U S I O N E S Y R E C O M E N D A C I O N E S
Los resultados obtenidos justifican las siguien
tes conclusiones:
• La distancia a la médula es la fuente de varia
ción más importante en la mayoría de los carac
teres anatómicos estudiados, lo que ratifica que
la variabilidad dentro del árbol es más importan
te que la variabilidad entre árboles o sitios.
• La densidad básica no mostró ser un índice efi
ciente para valorar la calidad de la madera de
bido a la alta variabilidad entre árboles.
• Los caracteres anatómicos considerados no es
tán afectados por el sitio, excepto la longitud de
las fibras que es influida por su crecimiento
intrusivo, produciendo fibras más largas en el
sitio con menor disponibil idad de agua.
• La longitud de fibras incrementa rápidamente
en dirección a la corteza hasta una edad aproxi
mada de 35 años, debido a la presencia de leño
juveni l . En edades superiores, el incremento es
menor . Hasta las edades estudiadas los e lemen
tos no están estabil izados.
• La longitud de vasos aumenta en forma más
acentuada hasta una edad aproximada de 35
años. Estas variables tienen edades de madura
ción similares a la longitud de fibras, corrobo
rando la presencia de leño juveni l .
• La dis tancia a la médu la resul tó ser eficiente,
debido a su corre lación con la edad cambial ,
para evaluar la var iabi l idad en la madera en
sent ido radial cuando los anil los son poco vi
sibles .
A E C K E R M A N N , F . 1995 . " T h e r e l a t i onsh ip b e t w e e n forest
site and in t ra r ing w o o d dens i ty factors for Quercus robur,
n . s .w . f rance" . In specia l I S S U E : si te c lass i f ica t ion a n d
e v a l u a t i o n . I U F R O s u b g r o u p S 1 . 0 2 . 0 6 . M e e t i n g h e l d
19 -23 . Oct . 1993 . Annales des Sciences Forestiers 52 (6) :
6 3 5 - 6 5 2 .
B A A S , P . 1986 . E c o l o g i c a l pa t t e rns i n x y l e m a n a t o m y . In T.J .
G ivn i sh (ed.) . On the economy of plant form and function:
3 2 7 - 3 5 2 . C a m b r i d g e , Un ive r s i t y P r e s s . C a m b r i d g e , N e w
Y o r k .
B A R A J A S - M O R A L E S , J . 1985 W o o d s t ruc tura l d i f fe rences
b e t w e e n t rees of t w o t rop ica l fores ts in M e x i c o . IAWA
Bulletin. N e w Ser i e s N° 8 3 5 5 - 3 6 4 .
B E N D T S E N , B . A. , J . R . S E N F T . 1989 . " M e c h a n i c a l a n d
ana tomica l proper t ies in individual g rowth r ings of p lanta t ion
- g r o w n eas te rn c o t t o n w o o d a n d lob lo l ly p i n e " , Wood and
Fiber Sci. 18: 2 3 - 3 8 .
B E R L Y N , B . , J . M I S H K E . 1976 . Botanical Microtechnique
and Cythochemestry. I o w a S ta te U n i v e r s i t y . A m e s I o w a .
B E S O L D , G., M . E . C A R R A N Z A , G . G I A N N U Z Z I . 1 9 8 8
Aná l i s i s q u í m i c o de la m a d e r a y b i o m a s a de e s p e c i e s del
N o r o e s t e a rgen t i no y su p o s i b l e a p r o v e c h a m i e n t o i n d u s
tr ial . Ins t i tu to d e T e c n o l o g í a d e l a M a d e r a . U N S E . I T M -
Ser ie d e P u b l i c a c i o n e s 8 8 0 1 .
B O L E T T A P . E. , L . R . A C U Ñ A , M . L . J U A R E Z . 1989 . A n á
lisis de las carac te r í s t icas c l imá t i cas de la p rov inc i a de S tgo .
del E s t e r o y c o m p o r t a m i e n t o d u r a n t e la s equ í a de la c a m
p a ñ a ag r í co la 1988 /89 . 2 3 p .
B O S M A N , M . T . M . , I . D E K O R T , M . K . G E N D E R E N , P .
B A A S . 1 9 9 4 . " R a d i a l v a r i a t i o n i n w o o d p r o p e r t i e s o f
na tura l ly a n d p lan ta t ion g r o w n L igh t R e d M e r a n t i S h o r e a
( D i p t e r o c a r p a c e a e ) " , IAWA Journal 15 (2 ) : 1 1 1 - 1 2 0 .
B U R L E Y , J., P . J . W O O D . 1979 . M a n u a l S o b r e I n v e s t i g a c i o
nes de E s p e c i e s y P r o c e d e n c i a s c o n R e f e r e n c i a E s p e c i a l a
los T r ó p i c o s . T r o p i c a l Fo re s t ry P a p e r s 10 E 1 0 a D e p a r t m e n t
o f Fo res t ry C o m m o n w e a l t h F o r e s t r y Ins t i tu te . U n i v e r s i t y
of Oxford .
B U T T E R F I E L D , R . P . , R . P . C R O O K , R . A D A M S , R .
M O R R I S . 1 9 9 3 . Rad ia l va r i a t ion i n w o o d spec i f ic g rav i ty ,
f ibre l eng ths a n d vesse l a r ea for t w o Cen t r a l A m e r i c a n
h a r d w o o d s : Hyeronima alchornoides a n d Vochysia
guatemaltensis na tura l a n d p l a n t a t i o n - g r o w n t rees . IAWA
Journal. V o l . 14 (2) 1 5 3 - 1 6 2 .
C A R L Q U I S T , S . 1977 . " E c o l o g i c a l fac tors i n w o o d e v o l u t i o n .
A florist ic a p p r o a c h " , Am. J. Bot. 6 4 : 8 8 7 - 8 9 6 .
C A R L Q U I S T , S. 1988 . Comparative wood anatomy. Springer
Series of wood Science. S p r i n g e r V e r l a g .
C O R O N E L , E . 1984 . L a d e t e r m i n a c i ó n d e a l g u n a s p r o p i e d a
des t ecno lóg icas de Aspidosperma q u e b r a c h o b l a n c o Sche lk t
y su va lo rac ión a t ravés de c o m p a r a c i o n e s e x p e r i m e n t a l e s
13
A G R A D E C I M I E N T O S
A la M s . Cs. Estela Pan, por la colaboración en
la determinación de la densidad básica. A la Ing.
María Luisa Juárez por la realización de los balan
ces hídricos de los sitios. A Fabián Zubrinic , por
la edición y la realización de las figuras. A Andrés
González, por la participación en el apeo y prepa
ración de muestras, y a Mirta Spossetti , por la
realización de los preparados microscópicos .
B I B L I O G R A F I A
XILEMA, VARIACION, RADIAL, QUEBRACHO BLANCO

de las N o r m a s I R A M y D I N pa ra l a c las i f icac ión de l a
m a d e r a a s e r r a d a ba jo las p r i n c i p a l e s c o n s i d e r a c i o n e s de
a p r o v e c h a m i e n t o y a p l i c a c i o n e s a rgen t inas . Tes i s doc to ra l ,
F a c . C i a s . F t a l e s A lbe r t L u d w i g d e F r e i b u r g . A l e m a n i a .
D I M I T R I , M. J., F . R O S A R I O , J . L E O N A R D I S , J . S . B I L O N I .
1997. El nuevo libro del árbol. T o m o I. Edi tor ia l . El A teneo .
F E R R E I R A , M . , P . Y . K A G E Y A M A . 1994 . M e l h o r a m e n t o
G e n é t i c o d a D e n s i d a d e d a M a d e i r a d e E u c a l i p t o . A n a i s d o
3 a C o n g r e s s o F lo res t a l B ra s i l e i ro . V o l . I I 148-152 .
F A H N , A . 1 9 6 4 . " S o m e a n a t o m i c a l a d a p t a t i o n s o f d e s e r t
p l an t s " , P h y m o r p h o l o g y . V o l 14: 9 3 - 1 0 1 .
F U K A Z A W A , K . 1984. Juven i l e w o o d o f h a r d w o o d s j u d g e d by
densi ty var ia t ion. I A W A Bul le t in N e w Ser ies Nº 5 . 6 5 - 7 3 .
G I M E N E Z , A . M . 1 9 9 8 . In f luenc ia d e l a e d a d sobre los ca rac
te res a n a t ó m i c o s y el c r e c i m i e n t o de Schinopsis q u e b r a c h o
c o l o r a d o (Sch lech t . ) Ba rk l . E t M e y e r , A n a c a r d i a c e a e , Tes i s
p r e s e n t a d a c o m o pa r t e de los r equ i s i to s p a r a op ta r por e l
g r a d o d e D o c t o r . U n i v e r s i d a d N a c i o n a l d e T u c u m á n .
H E L I N S K A - R A C Z K O W S K A , L. , E . F A B I S I A K . 1 9 9 1 . Ra
dial var ia t ion and g r o w t h ra te in t he l eng th of the axial
e l e m e n t s of the sess i le o a k w o o d . IAWA Bulletin N e w Se
r ies V o l . 12 (3) : 2 5 7 - 2 6 2 .
H U E C K , K., 1978 . L o s B o s q u e s d e S u d a m é r i c a . S o c i e d a d A le
m a n a d e C o o p e r a c i ó n T é c n i c a G . T . Z . T rad , d e B r u n R .
E s c h b o r n , A l e m a n i a .
I A W A C O M M I T T E E . 1 9 8 9 . " I A W A Li s t o f M i c r o s c o p i c
fea tures for h a r d w o o d iden t i f i ca t ion" . IAWA Bulletin n . s .
10 (3 ) : 2 1 9 - 3 3 2 . In : Wood Structure in Biological and
technological research (eds . ) P. B a a s , A.J . B o l t o n y D . M .
Cat l ing . 1 5 7 - 1 8 1 . L e i d e n Botan ica l Ser ies Nº3. Le iden U n i v .
P ress , T h e H a g u e .
I R A M I N S T I T U T O A R G E N T I N O D E R A C I O N A L I Z A C I O N
D E M A T E R I A L E S . 1977 . M a d e r a de f in ic iones N o r m a N º
9 5 0 2 . C D U : 6 7 4 : 1 6 8 . 1 . N o r m a N º 9 5 4 4 . D e t e r m i n a c i ó n d e
la d e n s i d a d bás i ca .
I S M A I L , J., M . H . J U S O H , M . H . S A H A R I . 1995. A n a t o m i c a l
va r ia t ion in p l a n t e d K e l a m p a y a n Neolamarckia cadamo,
( R u b i a c e a e ) . IAWA Journal. V o l 16 (3) : 2 7 7 - 2 8 8 .
J U A R E Z , M . L . 1998 I n f o r m e sobre b a l a n c e s h íd r icos de las
l oca l i dades La M a r í a y L o s P i rp in tos . ( Inéd i to ) .
K O L L M A N , F . 1959 . T e c n o l o g í a de l a m a d e r a y sus apl ica
c i o n e s . M i n i s t e r i o de Agr i cu l t u r a . Ins t i tu to Fores ta l de In
v e s t i g a c i o n e s y E x p e r i e n c i a s y Se rv i c io s de la M a d e r a .
M a d r i d , E s p a ñ a .
L A R S O N , P. 1994 . The vascular Cambium. Development and
Structure. Springer Series in wood Science. 7 2 0 p.
L E I , H. , M . R . M I L O T A , B . G A R T N E R . 1996 . " B e t w e e n and
wi th in - t r ee va r ia t ion in the a n a t o m y a n d specif ic grav i ty of
w o o d in O r e g o n w h i t e o a k (Quercus garrayana)", IAWA
Journal V o l . 17 (4) : 4 4 5 - 4 6 1 .
L E O N , H . , E S P I N O Z A D E P E R N I A . 1998 . " V a r i a b i l i d a d d e
la m a d e r a de Cordia thaisiana B o r r a g i n á c e a ) en sen t ido
t r ansve r sa l " . Revista Forestal Venezolana 42 (1) : 15 -23 .
L O R E N Z , G . 1995 . " C a r a c t e r i z a c i ó n e c o l ó g i c a d e u n sue lo
Eu t r i c R e g o s o l ba jo b o s q u e e n e l C h a c o S e m i á r i d o A r g e n
t ina" . Revista Quebracho 3: 1 3 - 2 3 .
M A N W I L L E R , F . G . 1974 . " F i b r e l eng th i n s t e m and b r a n c h e s
of smal l h a r d w o o d s on s o u t h e r n p ine s i t e s" . Wood Sci. 7:
130-132 .
M O G L I A , J . G . 2 0 0 0 . V a r i a b i l i d a d radial de los ca r ac t e r e s
a n a t ó m i c o s de Aspidosperma q u e b r a c h o b l a n c o . T e s i s p a r a
l a ob t enc ión del g r a d o de D o c t o r U n i v e r s i d a d N a c i o n a l de
T u c u m á n . 9 5 p .
P A N S H I N , A. J . , C. DE Z E E W . 1980 . Textbook of wood
technology. 4 t h Ed. M c . G r a w - H i l l B o o k C o . N e w Y o r k .
7 0 5 p .
P A R A S K E V O P O U L O U , A . H . 1 9 9 1 . " V a r i a t i o n o f w o o d
s t ruc tu re a n d p r o p e r t i e s of Cupresus sempervirens va r .
hor izonta l is in natural popu la t ions in G r e e c e " . IAWA Bulletin
N e w Ser ies Vo l . 12 (2) : 195 -206 .
P E S Z L E N , I . 1994 . " In f luence o f a g e on se l ec ted a n a t o m i c a l
p rope r t i e s of Populus c l o n e s " . IAWA Journal 15: 3 1 1 - 3 2 1 .
S I D I Y A S A , K.; P . B A A S . 1998 . E c o l o g i c a l a n d s y s t e m a t i c
W o o d a n a t o m y of Alstonia ( A p o c y n a c e a e ) . IAWA Journal.
Vol . 19 (2) : 2 0 7 - 2 2 9 .
T H O R N W A I T E , C . W . 1 9 5 7 . I n s t r u c t i o n s a n d t a b l e s for
c o m p u t i n g poten t ia l evapo t r ansp i r a t i on a n d the w a t e r ba
l a n c e . D r e x e l I n s t i t u t e o f T e c h n o l o g y , P u b l i c a t i o n s i n
C l ima to logy , V o l u m e n X , N u m b e r 3 . P p . 1 8 5 - 3 1 1 . C e n t e r o n
N e w Je r sey , U S A .
T Y R E E , M . T. ; S . D . D A V I S , H . C O C H A R D . 1994. " B i o p h y
sical pe r spec t i ve s of x y l e m evo lu t ion : is t he re a t radeof f of
hydraul ic efficiency for vulnerabi l i ty to dys func t ion?" , IAWA
Journal 15: 3 3 5 - 3 6 0 .
T Y R E E , M . T ; J . S . S P E R R Y . 1989 . " V u l n e r a b i l i t y o f x y l e m
to cav i ta t ion a n d e m b o l i s m " . Ann. Rev. Plant Phys. Mol.
Bio. 4 0 : 19 -38 .
V A N L A A R , A. 1 9 9 1 . Forest Biometry. P u b l i c a t i o n s p o n s o r e d
b y Sapp i F o r e s t s . 5 9 0 p .
W O O D C O C K , D . W . , C . M . I G N A S . 1994 . P r e v a l e n c e o f
w o o d charac te r s i n Eas te rn N o r t h A m e r i c a : W h a t cha rac t e r s
a re m o s t p r o m i s i n g for i n t e r p r e t i n g c l i m a t e s a n d fossi l
w o o d ? , Amer. J. Bot.
Y A N C H U K , A . D. , B . P . D A N C I K , M . M . M I C K O . 1 9 8 3 .
" In t rac lona l var ia t ion in w o o d dens i ty o f t r e m b l i n g A s p e n
in A l b e r t a " . Wood and Fiber Sci. 15 (3) : 3 8 7 - 3 9 4 .
Z H A N G , S . Y. ; Y . Z O N G . 1990 . "Effect o f g r o w t h ra te i n
specif ic g rav i ty of E a s t - L i a n i n g o a k Quercus liaotungensis)
w o o d " , Can. J. For. Res. 2 1 : 2 5 5 - 2 6 0 .
Z O B E L , B. J., J. T A L B E R T . 1984. Applied forest tree improve
ment. John W i l e y y S o n s . N e w Y o r k .
Z O B E L , B . , J . V A N B U I J T E N E N . 1989. Wood variation: Its
causes and control. S p r i n g e r - V e r l a g . Be r l ín .
R e c i b i d o : 1 2 . 0 1 . 2 0 0 0
14
JUANA GRACIELA MOGLIA, CARLOS RAUL LOPEZ


















