UNIVERSIDAD NACIONAL AGRARIA LA MOLINA · PDF file1 universidad nacional agraria la molina ....
149
1 UNIVERSIDAD NACIONAL AGRARIA LA MOLINA FACULTAD DE CIENCIAS “ABUNDANCIA RELATIVA Y DIVERSIDAD DE LA AVIFAUNA COMO INDICADORES DEL ESTADO DEL ECOSISTEMA: CAMPUS DE LA UNIVERSIDAD NACIONAL AGRARIA LA MOLINA” Presentado por: LUCÍA NATALIA CASTILLO PALACIOS TESIS PARA OPTAR EL TÍTULO DE BIÓLOGO Lima – Perú 2013
-
Upload
hoangkhanh -
Category
Documents
-
view
222 -
download
0
Transcript of UNIVERSIDAD NACIONAL AGRARIA LA MOLINA · PDF file1 universidad nacional agraria la molina ....

1
UNIVERSIDAD NACIONAL AGRARIA
LA MOLINA
FACULTAD DE CIENCIAS
“ABUNDANCIA RELATIVA Y DIVERSIDAD DE LA AVIFAUNA
COMO INDICADORES DEL ESTADO DEL ECOSISTEMA:
CAMPUS DE LA UNIVERSIDAD NACIONAL AGRARIA LA
MOLINA”
Presentado por:
LUCÍA NATALIA CASTILLO PALACIOS
TESIS PARA OPTAR EL TÍTULO DE
BIÓLOGO
Lima – Perú
2013

2
I. INTRODUCCION
El campus de la Universidad Nacional Agraria La Molina, ubicado en el
distrito de La Molina, Provincia de Lima, Departamento de Lima, (12º05’ S, 76º57’
O, 251 m), con un área aproximada de 230 Ha; es un área variada, conformada por
áreas agrícolas (diversos cultivos permanentes y temporales), áreas naturales
(bosques y matorrales) y zonas urbanas (edificios, pistas, restaurantes, jardines, etc.)
acompañado por un constante tránsito de personas (estudiantes, docentes y
trabajadores) que interactúan de manera directa e indirecta con la avifauna del lugar
(Takano & Castro, 2007; Takano, 2010; Corrales, 2012). Toda ésta variedad presente
en el campus hace importarte pensarlo como un ecosistema complejo, el cual
también se ve afectado por procesos antrópicos; es primordial conocer el estado de
este ecosistema, así como también definir un indicador biológico para su diagnóstico.
Una característica resaltante del campus de la UNALM es su variada avifauna, la
cual ha sido objeto de estudio en años previos. En 1992, Quinteros evaluó los
patrones de uso temporal y espacial de los campos de maíz por las aves granívoras,
enfocándose en la preocupación de estas especies como especies plagas. Takano y
Castro (2007) presentaron una lista taxonómica actualizada para esa fecha, así como
un análisis sobre la abundancia y diversidad de ciertas especies, nuevamente en
campos de cultivo y otras zonas del campus. Mientras que Takano (2010) trabajó
nuevamente en campos de cultivo de maíz estimando el daño que las aves causaban
sobre dichos campos, así como la diversidad de las aves. Por otro lado, Corrales
(2012) buscó conocer la percepción de la comunidad humana de la UNALM
(docentes, alumnos y trabajadores) sobre la fauna vertebrada urbana que habita el
campus, incluyendo las aves. Finalmente Castillo et al. (en prensa) realizó una
evaluación inicial del cambio de la comunidad de aves presente en el campus de la
Universidad Nacional Agraria La Molina a partir de estudios realizados en los años

3
1992, 2001, 2005, 2006 y 2007, y de evaluaciones realizadas entre los años 2009-
2011.
Es importante recordar que el distrito de La Molina ha cambiado con el paso de los
años, la creciente urbanización en dicho distrito se ha incrementado en los últimos
años por la demanda de viviendas, dejando pocos lugares de descanso, alimentación
y reproducción para la avifauna local (Corrales, 2012). Es así que, el campus de la
UNALM, con toda su oferta alimenticia anual y espacios amplios de reposo y
reproducción, podría estar supliendo y cubriendo de cierta manera éstos espacios
depredados por el crecimiento urbano.
Por otro lado, se encuentra la comunidad de aves perenne (Quinteros, 1992; Takano
y Castro, 2007) de la cual es necesario conocer su dinámica, en términos de
abundancia y diversidad, y como ésta se relaciona con las formaciones vegetales y
estructuras urbanas de dicho ecosistema; especialmente por las interacciones ave-
humano que se puedan dar como por ejemplo especies potencialmente plagas; es
importante estudiar el impacto del cambio espacial y temporal de la diversidad y
abundancia en el ecosistema y su dinámica.
La dinámica de esta comunidad de aves es también desconocida, es decir la
fluctuación de las variables comunitarias que las describen, tanto en el tiempo como
en el espacio, y como se relacionan con el paisaje de la UNALM, la distribución de
las variables podría ser usada como indicador del estado actual del ecosistema
(Torres, 2006). Es más, por la constante variación de la estructura del paisaje de la
UNALM, es necesario conocer las zonas del campus y épocas del año, con mayor y
menor diversidad así como abundancia, para que en el futuro sea posible un
planeamiento organizado y coherente del espacio, considerando el campus como un
ecosistema complejo ya que es el campus de la UNALM también susceptible a la
urbanización o crecimiento de las zonas urbanas como laboratorios o edificios que se
han construido en los últimos años en el campus.

4
Por otro lado, si bien se conoce que las actividades forestales y agrícolas tienen un
efecto empobrecedor sobre los ambientes naturales, las consecuencias globales de
los cambios en la diversidad de especies son poco conocidos. En general, el papel de
la diversidad de especies en el funcionamiento del ecosistema es poco conocido y se
cuenta con pocos estudios sobre las implicaciones funcionales de la reducción de
diversidad. Aquí radica el interés conservacionista de muchos, las alteraciones que
pueden existir por la desaparición o introducción de especies. En esto el ser humano
es el principal actor, no solo con la modificación de la morfología del paisaje, sino
también con la introducción de nuevas especies a las que se les conoce como
exóticas, estas especies pueden incrementar en un primer momento la diversidad,
pero a largo plazo la tendencia global indica que podrían bien desplazar a las
especies nativas, modificando la estructura e interacciones en el ecosistema. (Vilá,
1998)
La principal interrogante es, si es posible tener aproximaciones sobre los procesos de
los ecosistemas a partir de las características de las especies o si la variación en sus
abundancias implica alteraciones en los procesos de los ecosistemas, es así que
integrando los conocimientos de ecología de poblaciones con los de ecología de
ecosistemas sería posible develar el papel de las especies en los ecosistemas (Vilá,
1998; Lawton, 1994). Todo esto para formar bases ecológicas en la conservación de
los ecosistemas naturales, siendo la prioridad el mantenimiento de los procesos
ecológicos en su estado más genuino.
Se sabe que la diversidad frecuentemente se usa como un indicador del buen
funcionamiento del ecosistema (Magurran, 1989), por lo que el estudio de la
diversidad de una comunidad así como su estructura y composición, ayuda a
vislumbrar el funcionamiento íntegro del ecosistema, descrito a partir de su
componente avifauna.

5
El presente trabajo tiene como objetivo principal analizar la abundancia relativa y la
diversidad de la Avifauna del campus de la Universidad Nacional Agraria La Molina
como indicadores del estado de dicho ecosistema. Los objetivos específicos son: a)
determinar la abundancia relativa y la diversidad de las aves del campus de la
Universidad Nacional Agraria La Molina, b) elaborar una lista taxonómica
actualizada de las especies de aves presentes en el campus de la UNALM, c)
determinar los grupos funcionales existentes y las especies clave de cada grupo
funcional descrito, d) describir la relación de las aves con el ecosistema campus de
la UNALM a partir de su distribución espacial y temporal.
Entonces, es importante que este ecosistema Campus de la Universidad Nacional
Agraria La Molina, con habitas y ambientes variados, sea estudiado desde el punto
de vista de un sistema complejo, con una avifauna residente de la cual es primordial
conocer su dinámica espacial y temporal, así como su asociación al hábitat.
Finalmente, esta investigación intenta contribuir al uso de las aves como indicadores
biológicos del estado general del ecosistema campus de la UNALM, usando como
herramienta su diversidad y abundancia relativa, así como, brindar información
importante para futuras investigaciones en temas relacionados a la avifauna y su
dinámica además de ampliar el campo de investigación en temas de ecología urbana.

6
II. REVISIÓN DE LITERATURA
2.1 PARÁMETROS DE LAS COMUNIDADES BIÓTICAS
Una comunidad biótica se define como cualquier conjunto de poblaciones que
coexisten en un espacio determinado (Krebs, 1985). Según Magurran (1989), se
caracterizan por la interacción entre dichos organismos y el espacio que los define
como una comunidad. Una comunidad estará entonces representada por aquel
ensamblaje de especies que convive en un tiempo y espacio determinado.
Según Smith (2001) existen dos visiones contrapuestas sobre comunidad, la idea
expuesta por Clements, concepto holista que ve a la comunidad como un todo,
donde existe una asociación de especies y cada especie es parte de una unidad
integrada o “superorganismo” como lo llama Krebs (1985), el cual es natural y no es
producto de una clasificación humana. Por el contrario, Gleason expone más bien
un concepto individualista de comunidad, donde éste es el producto de la
coincidencia en los requerimientos y tolerancias ambientales de las especies, y
cualquier clasificación de ésta es artificial y no una descripción natural de la
estructura de la naturaleza.
Las características de una comunidad se ven expresadas en las características de
cada especie que la compone, entre estas se pueden nombrar: diversidad de especies,
estructura y formas de crecimiento, abundancia relativa y la estructura trófica
(Krebs, 1985). Estas características no son estáticas o inamovibles, al contrario
varían tanto en el tiempo como en el espacio generando dinámica en los ecosistemas.
Se hace necesario la medida de estas características para un mejor entendimiento de
las comunidades, así como la conservación de sus procesos y hábitats (Moreno,
2001).

7
Dentro de estas características que definen a la comunidad, es posible que la más
usada y nombrada sea la Diversidad, concepto antiguo ampliamente discutido,
referido principalmente a la frecuencia y heterogeneidad de las especies. Es así que
se define a la diversidad a partir de dos componentes, la Riqueza de Especies,
concepto más simple y antiguo de diversidad, referido al número de especies
diferentes en la comunidad y el componente estructural, que expresa la desigualdad
de las abundancias de cada especie, denominado Equitatividad. Estos componentes
de la diversidad reaccionarán de modo distinto dependiendo de los componentes
geográficos que conformen el ecosistema (Krebs, 1989; Margalef, 1991).
El concepto más simple y básico de la Diversidad es el de Riqueza de Especies o
variedad, el cual se refiere simplemente al número de especies o componentes
diferentes en una comunidad, sin importar el aporte que cada especie tenga en la
comunidad, es decir no considera si son especies comunes o raras en el ambiente
(Moreno, 2001; Odum & Barrett, 2006; Schmitz, 2007). Sin embargo, se puede
presentar el problema de que no siempre se pueda enumerar a todas las especies
presentes en una comunidad (Krebs, 1989) además de estar relacionado al tamaño de
la muestra (Sánchez, 1992).
Por otro lado, se tiene el componente estructural o de distribución de las unidades, la
equitatividad. En la estructura de la comunidad habrán especies dominantes con
mayores abundancias y especie con abundancias menores. Hipotéticamente, se
asume la existencia de una comunidad ideal donde todas las especies son igualmente
comunes (Krebs, 1989; Odum & Barrett, 2006).
La existencia de estos dos componentes de la Diversidad hace que existan a su vez
muchas maneras de cuantificarlos mediantes índices (Torres, 2006). Probablemente,
el más usado sea el Índice de Shannon- Weaver (H’), medida que expresa la
uniformidad de los valores de importancia a través de todas las especies de las

8
muestras, es decir es una medida del orden, todo esto dentro de la teoría de la
información, suponiendo que la diversidad es susceptible a medirla igual que la
información. También se le conoce como el grado de incertidumbre en predecir a
qué especie pertenecerá cierto individuo extraído al azar de un grupo
suficientemente grande (Moreno, 2001). Se considera que la población a medir es
“indefinidamente grande” (infinita y que los individuos se muestrean al azar)
(Pielou, 1975 citado por Magurran, 1989). Una gran desventaja de este índice es su
incapacidad para distinguir las diferencias estructurales.
Otro índice, en este caso utilizado para la estimación de la Riqueza Específica es el
Índice de Margalef, el cual es una función lineal entre el número de especies y el
número total de individuos presente (Magurran, 1989).
Por otro lado, la equidad es conocida como la medida de uniformidad que mide la
abundancia relativa de los individuos entre las especies (Smith, 2001), es una
relación entre la diversidad observada y la diversidad máxima.
La diversidad, dependiendo del tamaño de la muestra con que se trabaje, puede ser
de tres tipos: Alfa, Beta y Gamma.
La Diversidad Alfa, hace referencia a la diversidad dentro de las comunidades en un
hábitat, generada a partir de procesos evolutivos; también denominada puntual o
intra - hábitat. Su medida se puede realizar con el índice de Shannon-Weaver u otros
existentes. Por otro lado, la Diversidad Beta es una medida de la diferencia existente
entre distintos hábitats (diversidad inter – hábitats), es decir se refiere al grado de
reemplazamiento de especies o cambio biótico a través de gradientes ambientales.
Está basada en proporciones y usualmente es empleada para la comparación de la
diversidad entre comunidades diferentes. Mientras más alto sea el valor de la
diversidad beta mayor desorganización existirá en el ecosistema. Es posible
cuantificarla mediante fórmulas matemáticas, sin embargo se hace más sencillo el

9
uso de medidas de similitud como los Índices de Similitud/Disimilitud, los cuales
son los más usados, expresan el grado de semejanza de dos muestras en base a las
especies presentes en cada una de ellas; no toman en cuenta las abundancias
(Magurran, 1989; Moreno, 2001; Rodríguez, 2004).
Finalmente, la Diversidad Gamma, que se define como la diversidad a niveles
mayores como por ejemplo el paisaje. Esta resulta de la diversidad alfa de las
comunidades individuales (alfa promedio) y del grado de diferencia entre estas
comunidades (diversidad beta) (Moreno, 2001).
Por otro lado, se conoce como Abundancia Relativa a la cantidad de individuos de
una especie dentro de una comunidad, la cual varía según el tiempo y el espacio, en
relación a una medida de tiempo o de espacio, por lo que se puede determinar la
contribución de dicha especie al conjunto de la comunidad (Smith, 2001). También
la podemos definir como las proporciones relativas de diferentes especies en la
comunidad, ya que ninguna comunidad biótica está formada por grupos de especies
con abundancias iguales; siendo un alcance de muchos ecólogos la idea que existen
pocas especies dominantes y muchas especies raras (Magurran, 1989; Schmitz,
2007).
Un forma de expresar mejor el termino de abundancia Relativa es mediante el Índice
de Abundancia Relativa al Espacio (IARE), el cual es un índice operacional basado
en la idea de que la Abundancia Relativa es una función de la Densidad Real, siendo
la primera una buena estimadora de la segunda, ya que permite conocer cambios en
la población sin que sea necesario conocer el tamaño real de la población, siendo
importante mantener los métodos de medida constantes para reducir el error
(Quinteros, Sánchez & Tovar, 2002).

10
2.2 LAS AVES COMO INDICADORES BIOLÓGICOS
Un indicador biológico es un organismo estrechamente asociado con ciertas
condiciones cuya presencia es indicador de la existencia de dichas condiciones, es
por esto que son utilizados para la descripción de ambientes y para la detección de
cambios en estos (Morrison, 1986 citado por Koskimies, 1989).
Para el caso de estudios ambientales se suele utilizar a las aves, según sea el
objetivo. Normalmente se considera a las aves especialistas (especies acuáticas,
endémicas y migratorias) como buenos indicadores de los cambios ambientales.
(González, 2000a; Ramírez, 2000).
Las aves han sido usadas como indicadores a lo largo del tiempo, ya que se les ha
identificado como buenos detectores de los cambios ambientales en los diferentes
tipos de hábitats que existen, gracias a varios factores como el vasto conocimiento
de su ecología y biología, así como las relaciones que guardan con las formaciones
vegetales, sus papeles protagónicos en las redes tróficas para diferentes ecosistemas
y la facilidad de su estudio y registro, no siendo necesario el despliegue de muchos
recursos humanos y económicos (González, 2000a; Ramírez, 2000; Padoa-Schioppa
et al., 2005; Torres, Quinteros & Takano, 2006).
Sin embargo, su uso es aun discutido ya que es cuestionable si una sola especie
puede describir el comportamiento o tendencia de todo un conjunto, es más bien un
componente de la comunidad como la abundancia lo que podría dar una visión más
amplia de los cambios además de ser aplicada a otros espacios (Canterbury et al,
2000), siendo mucho más confiable incluso utilizar la abundancia de un gremio que
de una especie como indicador (Verner, 1984 citado por Canterbury et al, 2000).
Dichos gremios o grupos funcionales se pueden ver afectados por los cambios
poblacionales de una o dos especies que lo conforman, lo que los hace útil como

11
indicador, volviendo a la idea que el análisis de una sola especie no es suficiente
(Canterbury et al, 2000).
Así también, la alta o baja diversidad de un ecosistema se expresa en la alta o baja
diversidad de un solo grupo taxonómico de tal ecosistema (Margalef, 1998 citado
por Véliz, 2002). Por último, la diversidad y sus diversos índices son buenos
indicadores del buen funcionamiento del ecosistema, incluso como indicador de la
calidad ecológica (Magurran, 1989; Torres, 2006).
2.3 MÉTODO DE CONTEO POR TRANSECTOS EN FRANJA PARA EL
CENSADO DE AVES
Este método es un caso particular del Método de Transecto en Línea; consiste
en establecer transectos dentro del área de estudio, donde el observador se mueve a
lo largo de una línea o transecto con una velocidad aproximadamente constante,
registrando las especies que aparecen durante este tiempo (Ralph et al, 1996). Las
especies que se consideran en el conteo son aquellas registradas dentro de una
distancia constante a la cual se le denomina franja, el ancho de dicha franja debe ser
determinada antes del conteo. Normalmente, se establecen los transectos de acuerdo
a la distribución de las aves en el espacio y a la conveniencia del investigador.
El presente método presenta dos supuestos; se cuantifican solamente los individuos
que están dentro del ancho del transecto previamente definido y debe existir la
seguridad de contar a todos los individuos que están dentro de este ancho
(Mandujano, 1994).
Una ventaja del método es que se puede cubrir más terrero de manera más rápida
registrando un mayor número de aves y teniendo menor probabilidad de contar dos
veces el ave, lo cual aumentaría el error (Bibby, 1998). Por otro lado, es posible que

12
se presenten errores en el momento de estimar la distancia a la que se encuentra el
ave, para esto los observadores deberán ser previamente capacitados.
Según Bibby (1998), también es necesario evitar el conteo doble, ya que su efecto
puede traer consecuencias negativas, especialmente si el doble conteo se dio a lo
largo de un mismo transecto; para evitar esta situación es necesario tener en cuenta
las aves que ya se contaron, observando sus movimientos. Es por esto que no se
cuantifica ninguna ave que provenga de la espalda del observador, ya que al no
conocer su origen, se toma el riesgo que sea un mismo individuo que dio la vuelta.
Finalmente, es recomendable empezar el conteo 15 minutos después de la salida del
sol, y continuarlo entre 3 a 4 horas después del amanecer. Para la mayoría de
latitudes templadas, es recomendable realizar el censo entre 5 y 9 am (Ralph et al,
1996), recordando que no se debe realizar la evaluación en períodos de lluvia y/o
neblina, o frío intenso, ya que la actividad de las aves se ve reducida grandemente.
2.4 GRUPOS FUNCIONALES Y ESPECIE CLAVE
Ante los procesos de pérdida de biodiversidad que actualmente se presentan en
el mundo, se requiere transcender más allá que solamente un enfoque en especies
particulares, sino poseer un entendimiento de las relaciones entre la biodiversidad y
la función del ecosistema. Es así que la atención se centra o debe centrarse en los
grupos funcionales más que en especies únicas (Walker, 1992).
Los grupos funcionales son grupos de especies que tienen el mismo rol dentro de
una comunidad, manteniendo y regulando los procesos del ecosistema. Las
características con las que se clasifican a los grupos funcionales varían dependiendo
del tipo de ecosistema y objetivo del estudio, dicha característica es común entre los
organismos que conforman el grupo funcional, como morfología, fisiología o

13
comportamiento, y no está relacionada necesariamente a la relación evolutiva o
taxonómica entre especies (Vilá, 1998; Steneck, 2001).
Por otro lado, se denominan especies claves a aquellas especies que no
necesariamente son las dominantes en el ecosistema (aunque suelen ser las más
abundantes dentro de su grupo funcional), sino también aquellas que están altamente
interconectadas, con un impacto importante y cuantificable en los procesos del
mismo, así como en la diversidad de la comunidad. Dicho estatus, es modificable
según las circunstancias ecológicas (Vilá, 1998; Davic, 2003).
El término fue introducido por primera vez por Paine en 1969, a partir de estudios
realizados con comunidades en costas rocosas de California, donde al retirar una
especie de estrella de mar (Pisaster ochraceus) el cual era el principal depredador, el
número de especies presentes en dicha sección disminuyó de 15 a 8 especies, esto
fue evidencia suficiente para plantear la idea que la desaparición de una especie
clave en el ecosistema llevaría a una pérdida de diversidad global mucho mayor que
la desaparición de una especie conocida como paraguas o carismática (Payton et al,
2002; Davic, 2003).
El desarrollo de la hipótesis de especies claves, se da a partir del concepto de que la
distribución y densidad de las especies está desproporcionada, además de ser
afectada por las actividades de algunas especies en niveles tróficos altos. Una idea
de esta hipótesis es que cada comunidad o ecosistema tiene no una, sino varias
especies claves, éstas suelen ser aquellas que dominan estructura y dinámica en la
comunidad; por lo tanto la distribución de abundancias de la comunidad tendrá una
distribución sesgada y no normal; es difícil pensar que sea posible que todas las
especies tienen la misma importancia relativa en la comunidad, que existan especies
con una gran influencia en otras especies sin tener fuertes interacciones con otras
especies (Mills et al, 1993; Menge, 2001; Payton et al, 2002).

14
En términos de conservación de la diversidad biológica, es importante el concepto
de grupos funcionales, ya que al estar conformados por varias especies, en la
eventualidad de decline o desaparición debido a efectos intra específicos
(enfermedades, respuestas climáticas, etc) de una especie estos actúan como
amortiguadores. Por definición, las especies que conforman un grupo funcional son
ecológicamente equivalentes unas con otras, esta equivalencia permitirá una
compensación funcional por otros miembros otra especie que no son afectados; este
fenómeno conocido como redundancia ecológica provee al ecosistema de la
resiliencia necesaria para enfrentar perturbaciones, permitiéndole así preservarse en
el tiempo (Walker, 1995; Steneck, 2001). Es más, Sánchez (1992) menciona que es
hasta cierto punto impráctico el enfoque en una sola taxa biológica, es más útil
analizar dicha taxa con objetivos mayores de conservación de toda la comunidad
biótica; es más se menciona un mecanismo de recambio y compensación de
especies, que si bien es cierto desestabiliza la taxa en estudio pero hace estable las
características funcionales que son importantes para dicha comunidad (Estabilidad
Funcional).
Por otro lado, desde el punto de vista de las especies claves, Payton et al (2002)
expone que un sistema dependiente de las especies claves sería más vulnerable
puesto que la desaparición de dichas especies traería consigo cambios estructurales y
funcionales en la comunidad; por otro lado también afirma que es posible si existen
varias especies que comparten un mismo gremio trófico, podrían tener una función
compensatoria dado el caso que una especie desapareciera, e incluso otra llegaría a
ocupar el papel de especie clave que antes no tenía.
Según Mills, Soulé y Doaks (1993) es posible separar a las especies en hasta cinco
categorías las cuales no son excluyentes una de las otras. Estas son:

15
2.4.1. ESPECIE CLAVE DEPREDADORA
Puede considerarse especie clave a un depredador, si dicha especie depreda a
un consumidor primario y dicho consumidor es capaz de excluir otras especies de
esta comunidad (competidor); también se considera clave si el depredador controla
la población de otra especie clave que sea un consumidor primario, cuyo alimento
sean plantas hospederas importantes. Por último, es considerada especie clave a
aquel predador que sea controlador de alguna especie plaga.
2.4.2. ESPECIE CLAVE PRESA
Se considera especie clave a una presa cuando su desaparición deja
vulnerable a otras presas, también si su desaparición amenaza a la población de
depredadores de dicha especie. La desaparición de una especie clave presa produce
un incremento en la diversidad de la comunidad (Holt, 1977 citado por Mills et al,
1993).
2.4.3. ESPECIE CLAVE MUTUALISTAS
Son aquellas que mantienen relaciones mutualistas con otras especies y cuya
supervivencia depende de la supervivencia de la otra especie como por ejemplo
especies polinizadoras (Gilbert, 1980 citado por Mills et al, 1993).
2.4.4. ESPECIE CLAVE HOSPEDERAS
Son aquellas que albergan de cierta manera a las especies mutualistas y
dependen ecológicamente de ellas, en ese sentido serían las plantas polinizadas
(Gilbert, 1980 citado por Mills et al, 1993).

16
2.4.5. ESPECIE CLAVE MODIFICADORA
Son aquellas cuyas actividades modifican o afectan el hábitat sin ni siquiera
haber interferido con la cadena alimenticia. Si ésta modificación en el hábitat afecta
la supervivencia de otras especies, entonces es una especie clave.
Finalmente, es así como el trabajo y estudio a nivel de grupos funcionales, y
consecuentemente especies claves, provee información que trasciende barreras
taxonómicas y evolutivas, la cual podría proveer de un tipo de herramienta para la
predicción del cambio en un ecosistema (Steneck, 2001).
2.5 LAS AVES Y LA URBANIZACIÓN
La urbanización se define, según Bowman et al. (2001) como la modificación
por parte del ser humano de paisajes naturales, con la posibilidad de provocar un
profundo efecto en procesos ecológicos a diferentes escalas, desde grandes paisajes
hasta simples individuos. Esta urbanización se incrementa de manera global, se sabe
que actualmente las ciudades representan el 3% del uso de tierras en el mundo y para
el año 2025 se estima que la población mundial alcanzará los cuatro billones de
habitantes (Marzluff et al, 2001 citado por Leveau & Leveau, 2004), entonces esta
urbanización representa una serie de potenciales implicancias para la diversidad
biológica (Marzluff, 1998; Faeth et a.l, 2011).
Las comunidades de aves parecen distribuirse sin problema a lo largo de hábitats
urbanos, además de ser sensibles a los cambios producidos en dichos hábitats (Blair,
1996; McGregor-Fors, 2010).
Es más, éste proceso de urbanización afecta y transforma la estructura física del
hábitat además de alterar sus procesos ecológicos y evolutivos, lo que produce una

17
consecuente alteración en las comunidades biológicas de éstas áreas (Ramírez-
Albores, 2008). Las aves no son inmunes a estos cambios, todo lo contrario, la
urbanización afecta a las aves de manera directa cambiando sus procesos ecológicos
y hábitat, así como sus fuentes de alimento; e indirectamente afectando a sus
predadores, patógenos y competidores (González, 2004).
La mayoría de estudios indican que ante el proceso de urbanización se produce una
reducción general de la diversidad, abundancia y riqueza de especies de fauna
terrestre, en especial las aves, taxa que ha sido más estudiada junto con los
artrópodos, para la cual se ha registrado un incremento en la abundancia pero una
reducción en la riqueza y diversidad; este incremento en la abundancia normalmente
se debe al ingreso de especies exóticas al ecosistema, que pueden llegar a ser hasta el
80% del total de especies (Blair, 1999, Chace & Walsh, 2004; Blair & Launer, 1997
citado por Faggi & Perepelizin, 2006; Faeth et al.,2011; Nolazco, 2012). Por otro
lado, mucho autores han observado que en las zonas urbanas la riqueza es menor que
en las periferias sub urbanas (Faggi & Perepelizin, 2006).
Si bien es cierto los procesos de fragmentación y urbanización tiene un efecto
negativo en la diversidad y riqueza de especies (Almazán–Nuñez & Hinterholzer-
Rodríguez 2010), existen algunos casos donde la riqueza de aves aumenta para
zonas con niveles medios de urbanización, recordando siempre que los efectos de la
urbanización serán diferentes dependiendo del grupo taxonómico que se estudie, así
como las variables geográficas, espaciales y económicas del lugar (Faeth et al.,
2011).
Sin embargo, también puede ocurrir el hecho que unas pocas especies se beneficien
de esta urbanización y se adapten a ella, tolerando los cambios que trae consigo,
como especies sedentarias, con una amplia variedad de requerimientos alimenticios
(principalmente granívoras y omnívoras), altamente sociables y con preferencia de
anidamiento en estructuras artificiales (cavidades). (Chace & Walsh, 2004; Faggi &

18
Perepelizin, 2006; Croci et al, 2008; McGregor-Fors et al., 2009; Lancaster y Rees
1979 citado por Almazán–Nuñez & Hinterholzer-Rodríguez, 2010). Estas pocas
especies, según estudios a escala continental, estarían de cierta manera repitiéndose
en diferentes ciudades en distintos lugares del continente, a éste fenómeno se le
denomina homogenización biótica (Leveau & Leveau, 2006).
Por otro lado, son solo las ciudades temperadas las que tienen la mayor cantidad de
estudios en ecología urbana y no las de latitudes tropicales (Faeth et al., 2011). El
Perú no es la excepción a este vacío de información (Santana, 2011), siendo las aves
nuevamente las más estudiadas con la mayor cantidad de publicaciones en diferentes
parques de Lima, así como en el campus de la UNALM (Quinteros, 1992; González,
1998; González, 2002; González, 2004; Torres, Quinteros & Takano, 2006; Takano
& Castro, 2007; Takano, 2010; Corrales, 2012); Piura es otra ciudad que tiene
estudios en el campus universitario de la Universidad Nacional de Piura (Chávez et
al, 1999; Chávez, 2004) y finalmente un estudio en Lambayeque (Chanamé et al,
2010) quedando aún muchos vacíos de información sobre este tema.

19
III. MATERIALES Y MÉTODOS
3.1. UBICACIÓN DEL ÁREA DE ESTUDIO
El área de estudio es el correspondiente al campus de la Universidad Nacional
Agraria La Molina, ubicada en el distrito de La Molina, Provincia de Lima,
Departamento de Lima (12º05’ S, 76º57’ O, 251 m). El campus ocupa un área
aproximada de 230 hectáreas. Presenta una temperatura anual promedio de 20 °C,
humedad relativa promedio de 84% y una precipitación anual de 11.9 mm (Takano &
Castro, 2007; Takano, 2010).
Es en este paisaje es que ubica el campus de la UNALM (Figura 1), conformado por
áreas agrícolas (diversos cultivos permanentes y temporales), áreas naturales
(matorrales y pequeños bosques) y áreas urbanas (salones, laboratorios, edificios,
pistas, restaurantes, etc) donde se puede encontrar un constante tránsito de personas
que interactúan de manera directa e indirecta con la fauna del lugar.

20
Figura 1: Imagen satelital del campus de la Universidad Nacional Agraria La
Molina (Lima-Perú) con los 40 transectos evaluados. Fuente: Laboratorio de
Planeamiento Ambiental y Ordenamiento Territorial
3.2. MATERIALES
• De Campo
Se utilizaron los siguientes materiales en las evaluaciones de campo:
- Binoculares Bushnell 10x50
- Fichas de datos.
- Mapa simple del campus de la UNALM
- Reloj
- Lápiz y borrador

21
- Guías de identificación de aves: Birds of Peru (Schulenberg et al., 2007) y Las
Aves del Departamento de Lima (Koepcke, 1964).
- Cámara Fotográfica
- GPS
- Cinta Métrica
- Cintas de marcaje
• Programas y otros
- ArcGIS 10
- Golden Software SURFER 10
- Paquete Estadístico PAST
- Microsoft Office: Excel, Word.
- Materiales de escritorio
3.3. METODOLOGÍA
3.3.1. DETERMINACIÓN DE LA ABUNDANCIA RELATIVA Y LA
DIVERSIDAD DE LAS AVES DEL CAMPUS DE LA UNIVERSIDAD
NACIONAL AGRARIA LA MOLINA.
La metodología utilizada para el evaluación de las aves fue el de conteo total,
ya que el área de la UNALM es un área relativamente pequeña, lo que hizo posible
cubrirla en su totalidad.
Para facilidad de los muestreos se realizó una división imaginaria del campus en 4
zonas, por lo que cada equipo de observación, conformado por un máximo de tres
personas (observador, anotador y apoyo) recorrían los diferentes transectos de cada
zona, en el mismo orden secuencial durante los 12 meses que duró la evaluación.
22
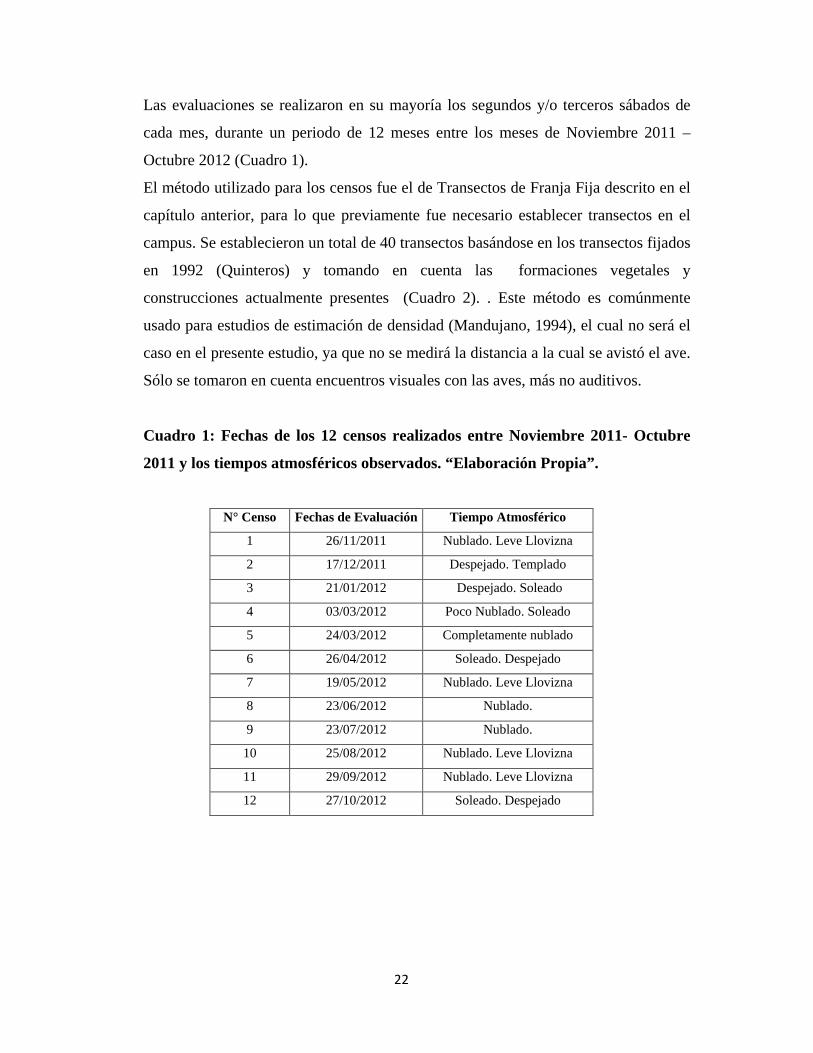
Las evaluaciones se realizaron en su mayoría los segundos y/o terceros sábados de
cada mes, durante un periodo de 12 meses entre los meses de Noviembre 2011 –
Octubre 2012 (Cuadro 1).
El método utilizado para los censos fue el de Transectos de Franja Fija descrito en el
capítulo anterior, para lo que previamente fue necesario establecer transectos en el
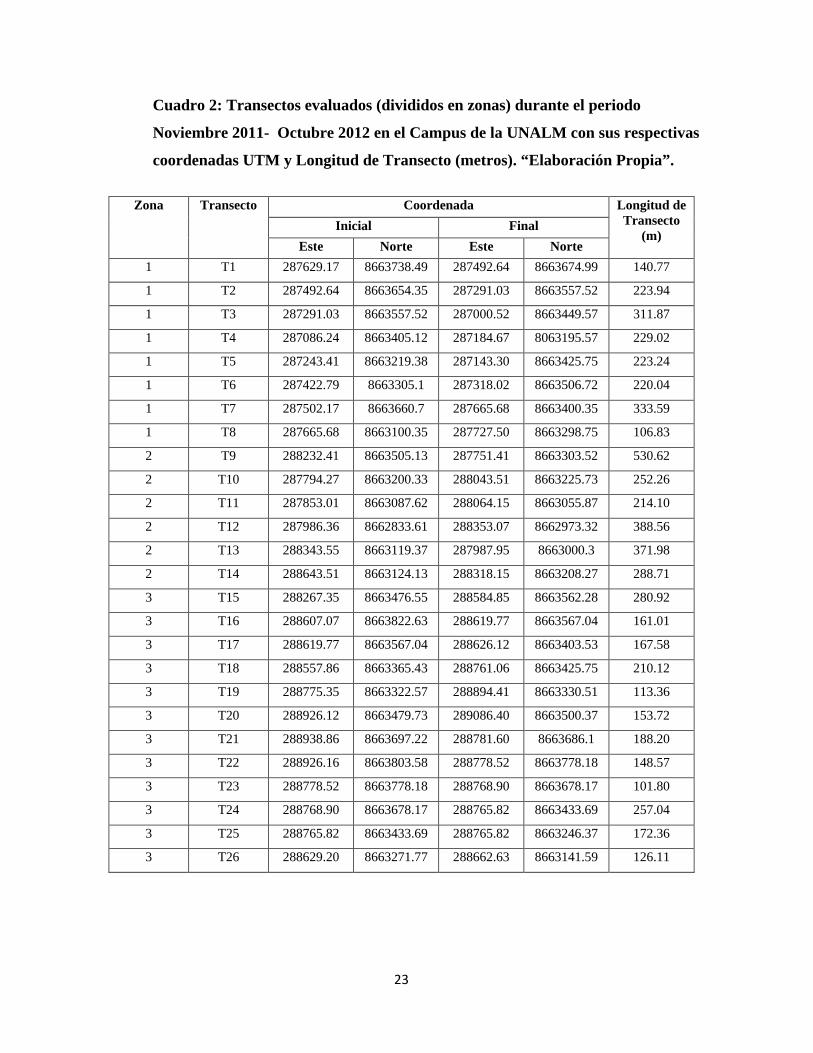
campus. Se establecieron un total de 40 transectos basándose en los transectos fijados
en 1992 (Quinteros) y tomando en cuenta las formaciones vegetales y
construcciones actualmente presentes (Cuadro 2). . Este método es comúnmente
usado para estudios de estimación de densidad (Mandujano, 1994), el cual no será el
caso en el presente estudio, ya que no se medirá la distancia a la cual se avistó el ave.
Sólo se tomaron en cuenta encuentros visuales con las aves, más no auditivos.
Cuadro 1: Fechas de los 12 censos realizados entre Noviembre 2011- Octubre
2011 y los tiempos atmosféricos observados. “Elaboración Propia”.
N° Censo Fechas de Evaluación Tiempo Atmosférico
1 26/11/2011 Nublado. Leve Llovizna
2 17/12/2011 Despejado. Templado
3 21/01/2012 Despejado. Soleado
4 03/03/2012 Poco Nublado. Soleado
5 24/03/2012 Completamente nublado
6 26/04/2012 Soleado. Despejado
7 19/05/2012 Nublado. Leve Llovizna
8 23/06/2012 Nublado.
9 23/07/2012 Nublado.
10 25/08/2012 Nublado. Leve Llovizna
11 29/09/2012 Nublado. Leve Llovizna
12 27/10/2012 Soleado. Despejado
23
Cuadro 2: Transectos evaluados (divididos en zonas) durante el periodo
Noviembre 2011- Octubre 2012 en el Campus de la UNALM con sus respectivas
coordenadas UTM y Longitud de Transecto (metros). “Elaboración Propia”.
Zona Transecto Coordenada Longitud de
Transecto (m)
Inicial Final Este Norte Este Norte
1 T1 287629.17 8663738.49 287492.64 8663674.99 140.77
1 T2 287492.64 8663654.35 287291.03 8663557.52 223.94
1 T3 287291.03 8663557.52 287000.52 8663449.57 311.87
1 T4 287086.24 8663405.12 287184.67 8063195.57 229.02
1 T5 287243.41 8663219.38 287143.30 8663425.75 223.24
1 T6 287422.79 8663305.1 287318.02 8663506.72 220.04
1 T7 287502.17 8663660.7 287665.68 8663400.35 333.59
1 T8 287665.68 8663100.35 287727.50 8663298.75 106.83
2 T9 288232.41 8663505.13 287751.41 8663303.52 530.62
2 T10 287794.27 8663200.33 288043.51 8663225.73 252.26
2 T11 287853.01 8663087.62 288064.15 8663055.87 214.10
2 T12 287986.36 8662833.61 288353.07 8662973.32 388.56
2 T13 288343.55 8663119.37 287987.95 8663000.3 371.98
2 T14 288643.51 8663124.13 288318.15 8663208.27 288.71
3 T15 288267.35 8663476.55 288584.85 8663562.28 280.92
3 T16 288607.07 8663822.63 288619.77 8663567.04 161.01
3 T17 288619.77 8663567.04 288626.12 8663403.53 167.58
3 T18 288557.86 8663365.43 288761.06 8663425.75 210.12
3 T19 288775.35 8663322.57 288894.41 8663330.51 113.36
3 T20 288926.12 8663479.73 289086.40 8663500.37 153.72
3 T21 288938.86 8663697.22 288781.60 8663686.1 188.20
3 T22 288926.16 8663803.58 288778.52 8663778.18 148.57
3 T23 288778.52 8663778.18 288768.90 8663678.17 101.80
3 T24 288768.90 8663678.17 288765.82 8663433.69 257.04
3 T25 288765.82 8663433.69 288765.82 8663246.37 172.36
3 T26 288629.20 8663271.77 288662.63 8663141.59 126.11
24
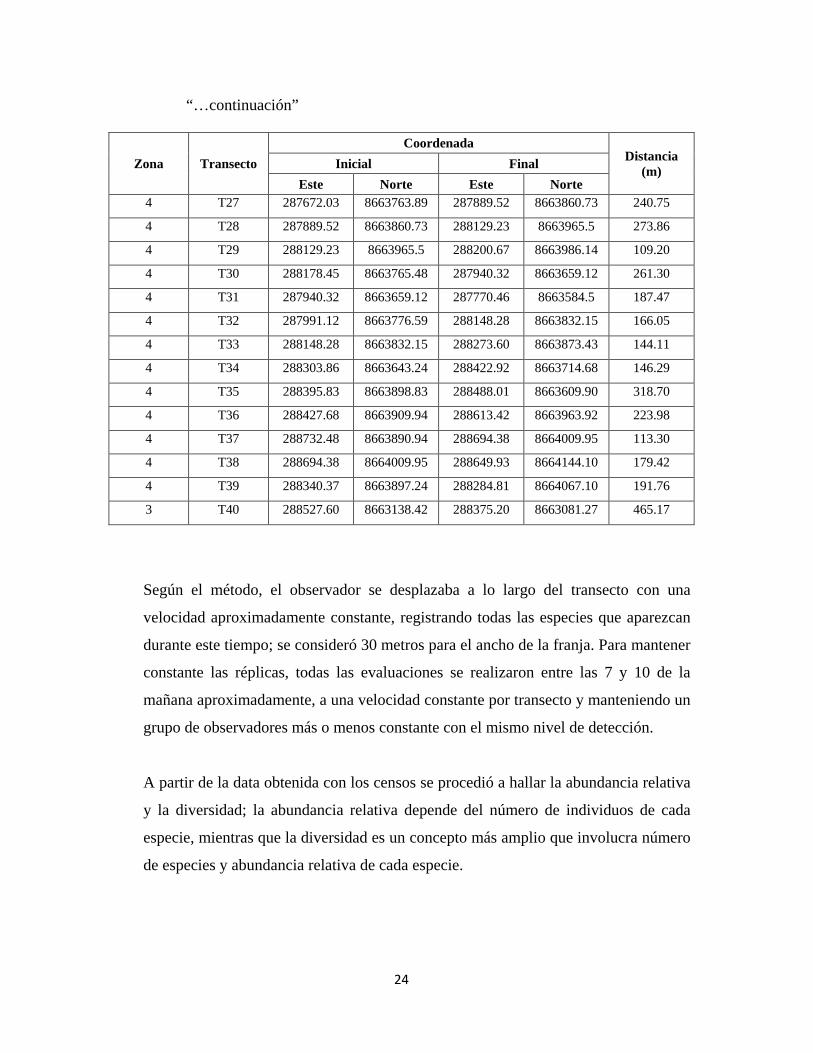
“…continuación”
Zona Transecto Coordenada
Distancia (m) Inicial Final
Este Norte Este Norte 4 T27 287672.03 8663763.89 287889.52 8663860.73 240.75
4 T28 287889.52 8663860.73 288129.23 8663965.5 273.86
4 T29 288129.23 8663965.5 288200.67 8663986.14 109.20
4 T30 288178.45 8663765.48 287940.32 8663659.12 261.30
4 T31 287940.32 8663659.12 287770.46 8663584.5 187.47
4 T32 287991.12 8663776.59 288148.28 8663832.15 166.05
4 T33 288148.28 8663832.15 288273.60 8663873.43 144.11
4 T34 288303.86 8663643.24 288422.92 8663714.68 146.29
4 T35 288395.83 8663898.83 288488.01 8663609.90 318.70
4 T36 288427.68 8663909.94 288613.42 8663963.92 223.98
4 T37 288732.48 8663890.94 288694.38 8664009.95 113.30
4 T38 288694.38 8664009.95 288649.93 8664144.10 179.42
4 T39 288340.37 8663897.24 288284.81 8664067.10 191.76
3 T40 288527.60 8663138.42 288375.20 8663081.27 465.17
Según el método, el observador se desplazaba a lo largo del transecto con una
velocidad aproximadamente constante, registrando todas las especies que aparezcan
durante este tiempo; se consideró 30 metros para el ancho de la franja. Para mantener
constante las réplicas, todas las evaluaciones se realizaron entre las 7 y 10 de la
mañana aproximadamente, a una velocidad constante por transecto y manteniendo un
grupo de observadores más o menos constante con el mismo nivel de detección.
A partir de la data obtenida con los censos se procedió a hallar la abundancia relativa
y la diversidad; la abundancia relativa depende del número de individuos de cada
especie, mientras que la diversidad es un concepto más amplio que involucra número
de especies y abundancia relativa de cada especie.

25
Para el caso de la Abundancia Relativa se dividió el número de individuos
registrados en el transecto entre la distancia de dicho transecto, teniendo así la
expresión “Número de aves registradas por unidad de longitud recorrida” (Ind/m)
(Sánchez et a.l, 1988 citado por Quinteros, 1992) para estandarizar los resultados de
este dato se le multiplicó por 100, teniendo así el Índice de Abundancia Relativa al
Espacio (IARE) expresado en “Número de Individuos registrados en 100 metros”.
Por otro lado, la Diversidad utilizada en primera instancia es la diversidad alfa, la
cual se refiere a la diversidad dentro de las comunidades en un hábitat, generada a
partir de procesos evolutivos (Magurran, 1989; Moreno, 2001). Para su cálculo se
utilizó el Índice de Diversidad de Shannon – Weaver (H), índice muy común y
utilizado en el cálculo de la diversidad, medida que expresa la uniformidad de los
valores de importancia a través de todas las especies de las muestras (Moreno,
2001). Se expresa matemáticamente como:
Donde pi es la proporción de individuos de la especie i-ésima.
Los valores de H´ oscilan entre 0 si solo hay una especie en la comunidad y log S los
cuales no sobrepasa el valor de 6 o 7 bits.
Una gran desventaja de este índice es su incapacidad para distinguir las diferencias
estructurales, por lo que ha sido muy criticado (Magurran, 1989).
Se describe a la Riqueza Específica (S), la cual también es ampliamente usada, como el
número de especies diferentes de aves y se le estimó utilizando el Índice de Margalef que
convierte el número de especies por muestra a una proporción a la que las especies se
añaden por expansión de la muestra (Moreno, 2001) Matemáticamente se expresa de la
siguiente manera:
H’= - Σpi (log2pi)

26
Donde S es el número de especies y N el número total de individuos.
Si Dmg presenta valores menores a 2 se considera baja riqueza y para valores mayores a 5
es un indicativo de una alta riqueza.
Por último, se hallaron valores para Equitatividad (J) o grado de uniformidad, la cual es
posible hallarla a partir de la diversidad máxima, situación hipotética en que todas las
especies de la comunidad presenten la misma abundancia (Krebs, 1989; Rodríguez,
2004), su expresión matemática es:
Siendo Hmax la diversidad máxima.
Donde S es la Riqueza de Especies.
El valor de J varía entre 0, si no existe equidad, y 1 si todas las especies presentan la
misma abundancia.
3.3.2. ELABORACIÓN UNA LISTA TAXONÓMICA ACTUALIZADA DE LAS
ESPECIES DE AVES PRESENTES EN EL CAMPUS DE LA UNALM.
Con la información colectada de los censos mensuales así como observaciones
fuera de éstas evaluaciones, se pudo elaborar una lista taxonómica con las especies de
aves presentes en el campus de la UNALM, basándose en la clasificación de Plengue
(2012). Para esto se usó como base las listas anteriores de 1992 (Quinteros), 2007 (Castro
& Takano) y del 2011 (Castillo et al., en prensa).
Dmg= (S-1) / ln N
Hmax= log2 S
J=H’/Hmax

27
3.3.3. DETERMINACIÓN LOS GRUPOS FUNCIONALES EXISTENTES Y LAS
ESPECIES CLAVE DE CADA GRUPO FUNCIONAL DESCRITO.
Para la determinación de los grupos funcionales se utilizó el criterio de gremios
alimenticios, es decir clasificar a las especies de acuerdo a su tipo de dieta. Basándose en
la literatura revisada y las observaciones directas del comportamiento alimenticio de las
aves del campus durante los censos e independientemente en cafetines, kioskos, etc.
(Giraudo, 2006; Martínez et al, 2010).
Se elaboraron histogramas con las abundancias relativas de las especies de cada grupo
funcional, determinando las especies más abundantes; a partir de las abundancias
relativas de cada especie representadas en los histogramas se determinó cuáles son las
especies clave sugeridas de cada grupo funcional, analizándolas con más detalle en
términos de su distribución espacial mediante mapas desarrollados con el programa
SURFER. Adicionalmente, se realizaron pruebas de regresión múltiple para cada especie
clave, teniendo como variables independientes a las abundancias de los diferentes
elementos del hábitat y como variable dependiente al Índice de Abundancia Relativa al
Espacio promedio de la especie en los 40 transectos; ésto se realizó con el propósito de
evaluar la relación de la distribución de las especies claves con el hábitat.
3.3.4. DESCRIPCIÓN DE LA RELACIÓN DE LAS AVES CON EL
ECOSISTEMA CAMPUS DE LA UNALM A PARTIR DE SU
DISTRIBUCIÓN ESPACIAL Y TEMPORAL DE LA ABUNDANCIA Y LA
DIVERSIDAD
Primero, fue importante contar con una descripción del hábitat; para esto se
describió las formaciones vegetales circundantes a los transectos en cada evaluación
(izquierda y derecha), así como estructuras artificiales y otras características importantes
del hábitat como presencia o ausencia de basura, muros, cuerpos de agua, entre otras.

28
Se tuvo especial atención a los cultivos como maíz, algodón y hortalizas, el estado de
este (arado o reposo) así como la fenología de los mismos, ya sea estado vegetativo o
reproductivo, entendiendo estado vegetativo como el momento en que la planta carece de
flor o fruto y siendo el estado reproductivo la presencia de estos elementos. Con estos
datos se armó una matriz que permitió un análisis más detallado utilizando índices de
diversidad y análisis multivariados como el Análisis Clúster y el Análisis de
Componentes Principales.
El Análisis Clúster, nombre que se le da a un conjunto de técnicas cuyo principal fin es el
de agrupar objetos de acuerdo a sus características, de tal manera que los conglomerados
o grupos que forma tienen alto grado de homogeneidad interna y un alto grado de
heterogeneidad externa, haciendo que gráficamente los objetos muy parecidos sean
próximos y los diferentes alejados. El análisis clúster funciona basándose en la medida de
similitud media dentro de los conglomerados, la técnica más utilizada para este fin son
las distancias euclidianas entre cada par de observaciones, a menor la distancia más
fuerte el grado de similaridad, se suele usar para datos cuantitativos y no de
presencia/ausencia (Pedroza & Dicovskyi, 2007; Hanneman, sf). Por otro lado el Análisis
de Componentes Principales explica la estructura de las varianzas y covarianzas de un
conjunto de variables mediante combinaciones lineales entre estas a las cuales se les
denomina componentes principales, este método busca simplificar los datos para tener
una visión más clara para el análisis e interpretación (Hernández Rodríguez, 1998).
Como ya se mencionó anteriormente, las variables comunitarias, obtenidas a partir de las
evaluaciones mensuales, tuvieron datos para los 12 meses del año y para todo el campus,
lo que permitió realizar un análisis temporal y espacial de abundancia relativa y de la
diversidad la comunidad de aves.
Para el análisis temporal de las variables se realizaron gráficos en Excel, para observar la
variación de dichas variables mensualmente, se calculó el Coeficiente de Variabilidad,
empleado para la comparación de dos o más distribuciones con el propósito de comparar

29
cual presenta mayor o menor variabilidad relativa (Martínez, 2006); también se calculó
los Intervalos de Confianza para la media con la siguiente formula (Berenson et al.,
2006):
Donde
I.C: Intervalo de Confianza
X: Valor Promedio
𝒕 ∝/𝟐: Valor Tabular de la distribución t (α=0.05; g.l = n-1)
𝑺: Desviación Estándar
𝑵: Tamaño de la Muestra
Y para el Coeficiente de Variabilidad se utilizó la siguiente fórmula (Martínez, 2006):
Donde
C.V es el Coeficiente de Variación
X: Valor Promedio
𝑺: Desviación Estándar
En el caso del análisis espacial se trabajó con mapas de contorno con el programa
SURFER 10, para cada mes tanto para la Abundancia Relativa como para la Diversidad.
Adicionalmente, se realizó un analisis multivariado utilizando el Análisis Clúster y
Análisis de Componetes Principales; tanto para los meses evaluados como para los
transectos utilizando la abundancia relativa y la diversidad con el programa PAST.
Tambíen se realizo un análisis de regresión simple entre la diversidad de los elemenos
del hábitata y la diversidad de las aves en los 40 transectos evaluados.
𝑰.𝑪 = 𝑿 ± 𝒕 ∝/𝟐(𝑺/√𝑵)
C.V =S/X*100

30
Finalmente, basándose en la Ley de Taylor, la cual es la relación entre la varianza
muestral y la media muestral de las densidades poblaciones, es un método útil para la
deteccion de patrones de variabilidad espacio-temporal, se realizó un análisis similar pero
utilizando los datos de la comunidad y no evaluando cada especie como la ley lo
demanda. La fórmula que expresa ésta relación es la siguiente, expuesta por Taylor en
1961 ( Giraldo et al., 2002; Castañeda, 2007):
Donde S2: Varianza muestral
m: Media muestral
Posteriormente, Mc Ardle et al. (1990) reemplazaron la varianza por el coeficiente de
variación ya que no se ve afectado por ceros y se permite realizar comparaciones entre
especies, obteniendo la siguiente formula (Giraldo et al., 2002):
Donde
CV es el Coeficiente de Variación.
Es el valor de “b” el que describe el comportamiento de la variabilidad espacial (bs) o
temporal (bt) de las abundancias medias. Se denomina hot-spots si bs>2 y bt>2, refugios
si bs<2 y bt<2; y por último hot-spots fijos y móviles si es que bs>2 y bt<2 (Giraldo et
al., 2002).
S2 = a mb
CV = a0.5 m(0.5b-1)
log10CV= 0.5 log10 a + (0.5b-1) log10m

31
IV. RESULTADOS Y DISCUSIÓN
4.1 COMPOSICIÓN DE LA COMUNIDAD DE AVES EN EL CAMPUS DE LA
UNALM
Del total de evaluaciones realizadas durante la investigación se determinó un total
de 44 especies para el Campus de la UNALM, agrupadas en 22 Familias y 13 Órdenes y
una especie Incertae sedis (inc. sed) es decir no clasificada la cual es Saltator
striatipectus (Saltador Rayado) (Cuadro 3). La Familia con el mayor número de especies
fue Thraupidae con ocho especies, seguida por Tyrannidae con cinco especies,
finalmente Icteridae y Columbidae con 4 especies cada una (Figura 2).
Por otro lado, del total de especies encontradas 38 son residentes permanentes para esta
región; Columba livia (Paloma Doméstica) no es nativa para Perú y 5 no son nativas para
la región de Lima: Aratinga erythrogenys (Cotorra de Cabeza Roja), Forpus coelestis
(Periquito Esmeralda) y Sicalis flaveola (Chiringüe Azafranado) los cuales provienen de
la costa norte del país; mientras que el Coereba flaveola (Mielero Común) es originario
de la costa norte y selva central. Finalmente Thraupis episcopus (Tangara Azuleja) es
una especie proveniente de la costa y selva norte.
Como se aprecia en el Cuadro 4, de las 44 especies observadas durante las evaluaciones,
ocho se encuentran en el Apéndice II de la Categorización del Convención sobre el
Comercio Internacional de Especies Amenazadas de Flora y Fauna Silvestre (CITES), lo
cual significa que éstas especies sin encontrarse en peligro de extinción actual, podrían
llegar a estarlo si no se controla su comercio de manera estricta.

32
Cuadro 3: Especies de aves observadas durante las evaluaciones mensuales
realizadas entre Noviembre 2011 – Octubre 2012 y observaciones personales en el
Campus de la Universidad Nacional Agraria La Molina (UNALM). “Elaboración
Propia”.
Orden Familia Nombre Científico* Nombre común*
Suliformes Phalacrocoracidae Phalacrocorax brasilianus
Cormorán Neotropical
Pelecaniformes Ardeidae Bubulcus ibis Garcita Bueyera
Cathartiformes Cathartidae Coragyps atratus
Gallinazo de Cabeza Negra
Accipitriformes Accipitridae Parabuteo unicinctus
Gavilán Mixto
Falconiformes Falconidae Falco
sparverius Cernícalo Americano
Falco femoralis Halcón Aplomado
Gruiformes Rallidae Pardirallus sanguinolentus
Rascón Plomizo
Charadriiformes Burhinidae Burhinus
superciliaris Alcavarán Huerequeque
Charadriidae Charadrius vociferus
Chorlo Gritón
Columbiformes Columbidae
Zenaida auriculata
Tortolita Orejuda
Zenaida meloda Tortolita Melódica
Columba livia Paloma Doméstica
Columbina cruziana
Tortolita Peruana
Psittaciformes Psittacidae Aratinga
erythrogenys Cotorra de Cabeza Roja
Forpus coelestis Periquito Esmeralda
Cuculiformes Cuculidae Crotophaga sulcirostris
Garrapatero de Pico Estriado
Strigiformes Strigidae Athene cunicularia
Lechuza Terrestre
Apodiformes Trochilidae
Amazilia amazilia
Colibrí de Vientre Rufo
Myrtis fanny Estrellita de Collar Púrpura
Thaumastura cora
Colibrí de Cora
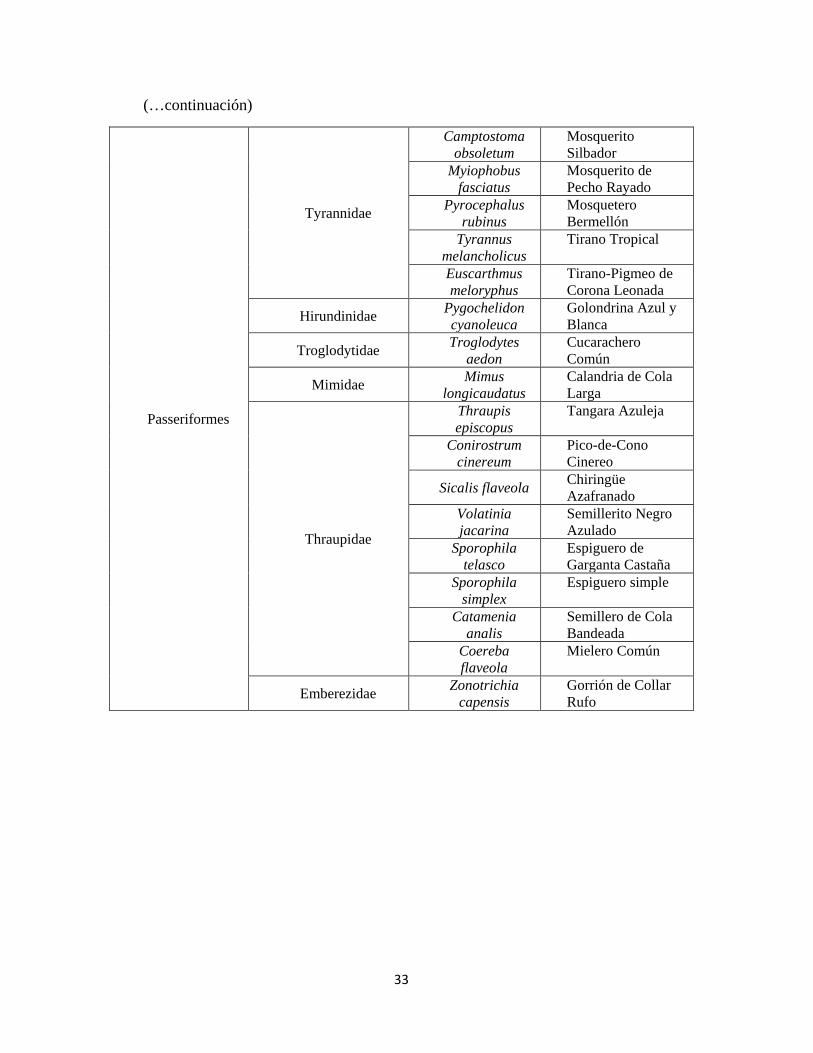
33
(…continuación)
Passeriformes
Tyrannidae
Camptostoma obsoletum
Mosquerito Silbador
Myiophobus fasciatus
Mosquerito de Pecho Rayado
Pyrocephalus rubinus
Mosquetero Bermellón
Tyrannus melancholicus
Tirano Tropical
Euscarthmus meloryphus
Tirano-Pigmeo de Corona Leonada
Hirundinidae Pygochelidon cyanoleuca
Golondrina Azul y Blanca
Troglodytidae Troglodytes aedon
Cucarachero Común
Mimidae Mimus longicaudatus
Calandria de Cola Larga
Thraupidae
Thraupis episcopus
Tangara Azuleja
Conirostrum cinereum
Pico-de-Cono Cinereo
Sicalis flaveola Chiringüe Azafranado
Volatinia jacarina
Semillerito Negro Azulado
Sporophila telasco
Espiguero de Garganta Castaña
Sporophila simplex
Espiguero simple
Catamenia analis
Semillero de Cola Bandeada
Coereba flaveola
Mielero Común
Emberezidae Zonotrichia capensis
Gorrión de Collar Rufo

34
(…continuación)
*Nombre Comunes y clasificación según Plengue (2012)
Cuadro 4: Especies de aves categorizadas en el Apéndices II- CITES. “Elaboración Propia”.
Nombre Científico Nombre común
Amazilia amazilia Colibrí de Vientre Rufo
Aratinga erythrogenys Cotorra de Cabeza Roja
Athene cunicularia Lechuza Terrestre
Falco femoralis Halcón Aplomado
Falco sparverius Cernícalo Americano
Forpus coelestis Periquito Esmeralda
Myrtis fanny Estrellita de Collar Púrpura
Parabuteo unicinctus Gavilán Mixto
Thaumastura cora Colibrí de Cora
Passeriformes
Icteridae
Molothrus bonariensis
Tordo Brilloso
Dives warszewiczi
Tordo de Matorral
Icterus graceannae
Bolsero de Filos Blancos
Sturnella bellicosa
Pecho Colorado Peruano
Fringillidae Sporagra magellanica
Jilguero Encapuchado
Passeridae Passer domesticus
Gorrión Casero
Incertae sedis Saltator Striatipectus
Saltador rayado
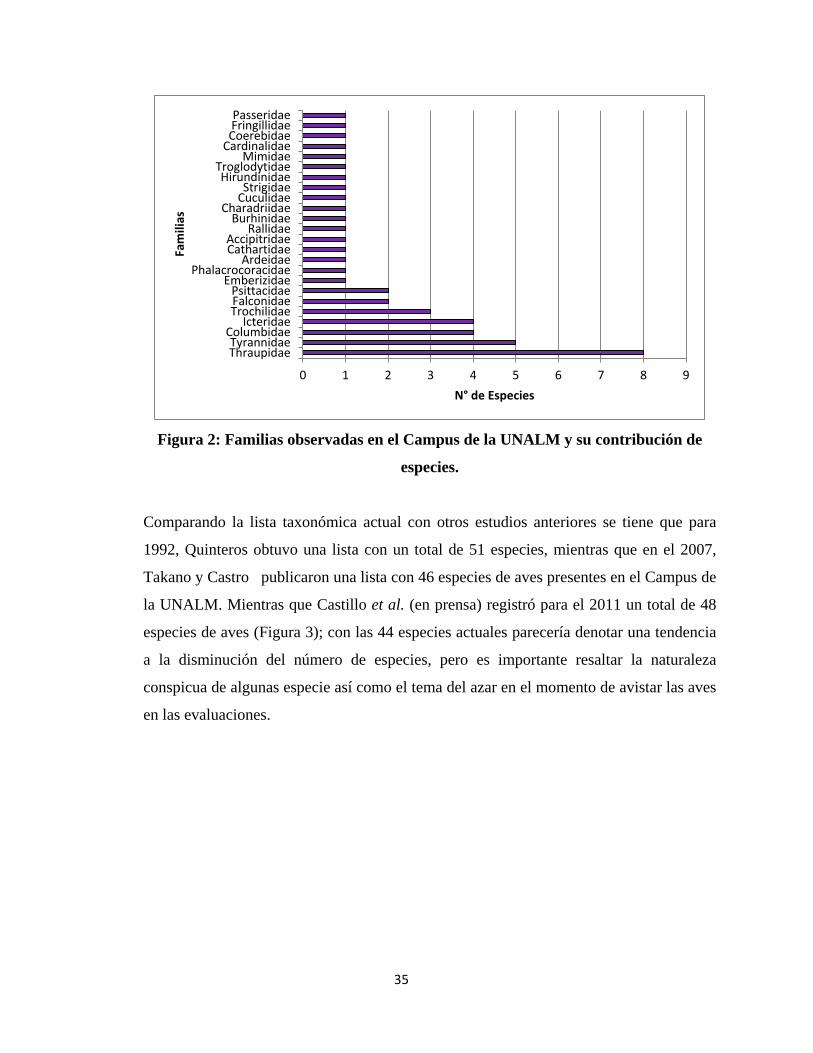
35
Figura 2: Familias observadas en el Campus de la UNALM y su contribución de
especies.
Comparando la lista taxonómica actual con otros estudios anteriores se tiene que para
1992, Quinteros obtuvo una lista con un total de 51 especies, mientras que en el 2007,
Takano y Castro publicaron una lista con 46 especies de aves presentes en el Campus de
la UNALM. Mientras que Castillo et al. (en prensa) registró para el 2011 un total de 48
especies de aves (Figura 3); con las 44 especies actuales parecería denotar una tendencia
a la disminución del número de especies, pero es importante resaltar la naturaleza
conspicua de algunas especie así como el tema del azar en el momento de avistar las aves
en las evaluaciones.
0 1 2 3 4 5 6 7 8 9
ThraupidaeTyrannidae
ColumbidaeIcteridae
TrochilidaeFalconidaePsittacidae
EmberizidaePhalacrocoracidae
ArdeidaeCathartidaeAccipitridae
RallidaeBurhinidae
CharadriidaeCuculidaeStrigidae
HirundinidaeTroglodytidae
MimidaeCardinalidaeCoerebidaeFringillidaePasseridae
N° de Especies
Fam
ilias

36
Figura 3. Número de especies presentes en el Campus de la UNALM de acuerdo
diferentes estudios realizados (Quinteros, 1992; Castro & Takano, 2007; Castillo et
al, en prensa) y el estudio actual (Noviembre 2011-Octubre 2012)
Desde 1992, hay diez especies no registradas para el periodo de evaluaciones actual
(Noviembre 2011 – Octubre 2012); desde el 2007 hay un total de ocho especies de aves
que ya no se han vuelto a avistar en el Campus. Finalmente, desde el 2010 (Castillo et
al., en prensa) hay 4 especies no registradas en comparación del presente estudio (Anexo
5). Sin embargo, resulta pertinente mencionar que especies como Psilopsiagon aurifrons
(Perico Coordillerano), Rhodopis vesper (Colibrí de Oasis), Sporophila luctuosa
(Espiguero Negro y Blanco) y Anairetes reguloides (Torito de Cresta Pintada) han sido
avistadas en evaluaciones posteriores a las comprendidas en este estudio, lo que dejaría la
posibilidad abierta de ampliar la lista.
51 46 48
44
303540455055
Quinteros(1992)
Castro &Takano (2007)
Castillo (2009-2011)
Tesis (2011-2012)
Núm
ero
de E
spec
ies
(S)

37
Figura 4: Dendrograma de presencia y ausencia de especies de aves del campus de
la UNALM de lista anteriores (Quinteros, 1992; Takano &Castro, 2007; Castillo et
al, en prensa) y el estudio actual (Noviembre 2011-Octubre 2012). Índice de Jaccard
“Elaboración Propia”.
Con el fin de poder analizar más detalladamente la diferencia de la composición de las
especies de las diferentes listas taxonómicas elaboradas en estudios anteriores
(Quinteros, 19992; Takano & Castro, 2007; Castillo et al., en prensa) se realizó un
Análisis Clúster, teniendo como resultado el dendrograma de la Figura 4, para distancias
mayores a 0.60 se forman dos grupos, uno conformado por la listas del 2007 (Takano
&Castro), la lista comprendida entre el 2009 -2011 (Castillo et al., en prensa) y el estudio
actual realizado entre Noviembre 2011 - Octubre 2012, y por otro lado se tiene la lista de
1992 (Quinteros); se puede observar claramente como ésta última se aleja del resto, lo
que muestra que la composición de especies de 1992 es muy diferente a la composición
de especies de aves de la última década.
Finalmente, en las evaluaciones realizadas entre Noviembre del 2011 y Octubre del 2012
se registró un total de 44 especies de aves para el Campus de la UNALM, pertenecientes

38
a 22 Familias y 13 Órdenes, además de una especie aun no clasificada. La Familia con el
mayor número de especies fue Thraupidae con 8 especies. Por otro lado se aprecia una
variación en el número de especies presenten en comparación a listas presentadas en años
anteriores (Quinteros, 1992; Takano y Castro, 2007; Castillo et al., en prensa)
presentándose una leve reducción en el número de especies encontradas a la fecha.
Finalmente, en cuanto a la composición de especies de dichas listas se puede resaltar que
la lista de 1992 (Quinteros) presenta más diferencias en relación a las listas del 2007
(Takano y Castro), del 2011 (Castillo et al., en prensa) y el estudio actual, posiblemente
porque el estudio de 1992 (Quinteros) evalúo campos de cultivo a diferencia de las otras
listas, donde se evaluó el campus en su totalidad.
4.2 DESCRIPCIÓN DEL HÁBITAT CIRCUNDANTE Y ZONIFICACIÓN DEL
CAMPUS
Como se describió en la metodología, durante los censos se registró las
formaciones vegetales circundantes, así como la presencia de construcciones (facultades,
aulas, laboratorios, etc) y entre otras características resaltantes del paisaje. Se
determinaron 25 diferentes tipos elementos del hábitat los cuales se mencionan en el
Cuadro 5.

39
Cuadro 5. Elementos del Hábitat descritos para el campus de la Universidad
Nacional Agraria La Molina. “Elaboración Propia”
Elementos Abreviatura
Grupo de Arboles A Arbustos Arb
Cuerpo de agua Agua Presencia de Basura o Desmonte Bas
Borde vivo B.V Bosque Bosq
Campo de Cultivo en arado CC1 Campo de Cultivo en barbecho CC2
Campo de Cultivo en Estado Vegetativo de Algodón CC3-A Campo de Cultivo en Estado Reproductivo de Algodón CC4-A
Campo de Cultivo en Estado Vegetativo de Frutales CC3-F Campo de Cultivo en Estado Reproductivo de Frutales CC4-F Campo de Cultivo en Estado Vegetativo de Hortalizas CC3-H
Campo de Cultivo en Estado Reproductivo de Hortalizas CC4-H Campo de Cultivo en estado Vegetativo de cultivo cualquiera CC3-O
Campo de Cultivo en Estado Reproductivo de cultivo cualquiera CC4-O Campo de Cultivo en Estado Vegetativo de Maíz CC3-M
Campo de Cultivo en Estado Reproductivo de Maíz CC4-M Construcciones Cons
Establo Est Jardines Ja
Herbáceas/maleza Herb/Mal Matorral Mat
Muro Mu Tierra Desnuda Ti.Des
Como se observa en la Figura 5 en cuanto a formaciones vegetales y estructuras
dominantes, Borde Vivo es la más dominante, seguido de Jardines y Árboles, también es
importante mencionar las Construcciones compuesta por salones, facultades y
laboratorios. Los campus de cultivo, para su mejor análisis fueron divididos de acuerdo a
la fenología en que se encontraban y tipo de cultivo (Maíz, Algodón, Frutales, Hortalizas
u otros), es por eso que pierden dominancia en el gráfico, pero si se tomaran en cuenta
campos de cultivo como formación única destinada a la plantación de algún cultivo,
sería las formaciones vegetales las dominantes.
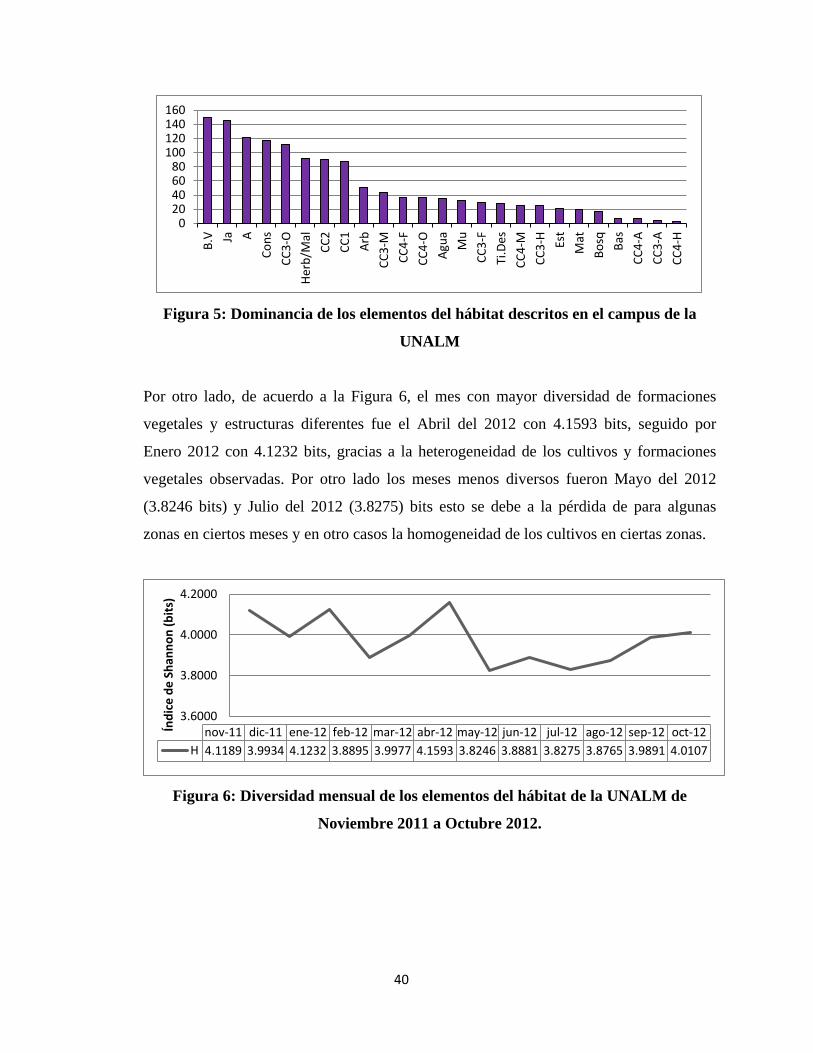
40
Figura 5: Dominancia de los elementos del hábitat descritos en el campus de la
UNALM
Por otro lado, de acuerdo a la Figura 6, el mes con mayor diversidad de formaciones
vegetales y estructuras diferentes fue el Abril del 2012 con 4.1593 bits, seguido por
Enero 2012 con 4.1232 bits, gracias a la heterogeneidad de los cultivos y formaciones
vegetales observadas. Por otro lado los meses menos diversos fueron Mayo del 2012
(3.8246 bits) y Julio del 2012 (3.8275) bits esto se debe a la pérdida de para algunas
zonas en ciertos meses y en otro casos la homogeneidad de los cultivos en ciertas zonas.
Figura 6: Diversidad mensual de los elementos del hábitat de la UNALM de
Noviembre 2011 a Octubre 2012.
020406080
100120140160
B.V Ja A
Cons
CC3-
OHe
rb/M
alCC
2CC
1Ar
bCC
3-M
CC4-
FCC
4-O
Agua M
uCC
3-F
Ti.D
esCC
4-M
CC3-
H Est
Mat
Bosq Ba
sCC
4-A
CC3-
ACC
4-H
3.6000
3.8000
4.0000
4.2000
Índi
ce d
e Sh
anno
n (b
its)
nov-11 dic-11 ene-12 feb-12 mar-12 abr-12 may-12 jun-12 jul-12 ago-12 sep-12 oct-12H 4.1189 3.9934 4.1232 3.8895 3.9977 4.1593 3.8246 3.8881 3.8275 3.8765 3.9891 4.0107

41
La diversidad de las formaciones vegetales y estructuras no varió grandemente a los
largo de los transectos evaluados como se observa en la Figura 7, presentando valores
con niveles de diversidad que varían en su mayoría entre 3.083 bits/ind y 3.4697 bits/ind,
es decir se mantiene un nivel de diversidad más o menos constante; sin embargo lo que sí
varía es la composición de las formaciones, siendo el Transecto N° 9 el más diverso con
3.4697 bits pues presenta gran cantidad de diferentes formaciones que suelen variar a lo
largo del año y el menos diverso el Transecto N°40 con 2.4483 bits el cual presenta
cultivo de hortalizas casi todo el año.
Figura 7: Diversidad de los Elementos del Hábitat en los 40 transectos evaluados de
Noviembre 2011 a Octubre 2012.
Como se explicó anteriormente, con fines prácticos se dividió el campus de la
universidad en 4 zonas como se muestra en la Figura 8, cada zona presenta características
propias, así como formaciones vegetales y estructuras únicas.
2.20002.40002.60002.80003.00003.20003.40003.6000
T1 T2 T3 T4 T5 T6 T7 T8 T9 T10
T11
T12
T13
T14
T15
T16
T17
T18
T19
T20
T21
T22
T23
T24
T25
T26
T27
T28
T29
T30
T31
T32
T33
T34
T35
T36
T37
T38
T39
T40
Índi
ce d
e Sh
anno
n (b
its)
Transectos

42
Figura 8. Imagen satelital del campus de la Universidad Nacional Agraria La
Molina (Lima-Perú) mostrando las 4 zonas evaluadas. Fuente: Laboratorio de
Planeamiento Ambiental y Ordenamiento Territorial
• Zona1: Comprendida desde el Transecto N° 1 hasta el Transecto N° 8, es una
zona totalmente agrícola con campos de cultivo, especialmente de maíz (Zea
mays) y otros cultivos rotativos. Se registra la presencia de muros por estar en el
límite del campus, así como numerosos cercos vivos de plátano.
• Zona 2: Incluye desde el Transecto N° 9 hasta el Transecto N° 14. También
incluye campos de cultivo como maíz (Zea mays), Yuca (Manihot sp.), y
pequeñas parcelas de cultivos variados. Además se encuentra el cuerpo de agua
más grande presente en el campus así como un pequeño cerro parcialmente
cubierto de vegetación y rodeado de desmonte. Finalmente, presenta parcelas de
matorrales y bosques; cabe resaltar que en el ingreso al transecto N° 13 existe un
botadero de desmonte y basura.

43
• Zona 3: Comprende desde el Transecto N° 15 hasta el Transecto N° 26 y
también el Transecto N° 40. Es una zona también predominantemente agrícola,
con presencia de campos de cultivo de hortalizas, frutales como lúcuma (Pouteria
obovata), mandarinas (Citrus nobilis), así como cultivos variados. Hay presencia
de bordes de vegetación, árboles y maleza. Por último, es importante resaltar que
el Transecto N° 15 tiene coincidencia con una parte de los establos.
• Zona 4: Esta zona incluye la parte urbana de la universidad dominada
principalmente por las áreas académicas y administrativas, con gran presencia de
jardines, árboles y arbustos. Incluye cuerpos de agua pequeños, así como el Jardín
Botánico “Octavio Velarde Núñez”; también abarca parte del establo (Transecto
N° 34) y áreas del denominado Campo Ferial. Comprende desde el Transecto N°
27 hasta el Transecto N° 39.
En conclusión, se encontraron 27 diferentes elementos del hábitat entre artificiales y
naturales para el campus de la UNALM, siendo Borde Vivo, Jardín, Árboles y
Construcciones los más dominantes; sin embargo, el campus de UNALM se presenta
como un ecosistema predominantemente agrícola aún. En cuanto al análisis temporal el
mes más diverso fue Abril 2012 (4.1593 bits) y Mayo 2012 el menos diverso (3.8246
bits). Finalmente, la distribución espacial no varía demasiado, teniendo el Transecto N° 9
como el más diverso y el Transecto N° 40 como el menos diverso.
4.3 ANÁLISIS TEMPORAL Y ESPACIAL DE LA ABUNDANCIA RELATIVA
La información recogida de los censos mensuales sirvió para el cálculo del Índice
de Abundancia Relativa al Espacio (IARE) cuyas unidades fueron “Individuos por cada
100 metros recorridos” (Ind/100m). En promedio, según se aprecia en la Figura 9, la
especie más abundante para todo el periodo de evaluación es Zenaida meloda (Tortolita
Melódica) con 102.16 Ind/100m, seguido por Columbina cruziana (Tortolita Peruana)

44
con 87.55 Ind/100m, Pygochelidon cyanoleuca (Golondrina Azul y Blanca) con 66.29
Ind/100m y Dives warszewiczi (Tordo de Matorral) con 36.95 Ind/100m. Por otro lado,
las especies poco abundantes son muchas, siendo las menos abundantes Thaumastura
cora (Colibrí de Cora) con 0.0289 Ind/100m, Icterus graceannae (Bolsero de Filos
Blancos) con 0.06 Ind/100m, Phalacrocorax brasilianus (Cormorán Neotropica) con
0.08 Ind/100m y Pardirallus sanguinolentus (Rascón Plomizo) con 0.08 Ind/100m por
mencionar algunas.

45
Figura 9: Dominancia de especies de aves según su Índice de Abundancia Relativa
al Espacio (IARE) (Promedio Anual de Noviembre 2011- Octubre 2012)
0.03 0.06 0.08 0.08 0.09 0.09 0.21 0.26 0.29 0.30 0.45 0.51 0.78 0.96 0.96 1.39 1.56 1.72 1.91 2.30 3.50 3.83 4.27 5.26 5.95 6.33 7.52 9.32 9.59
12.91 13.04 13.54 13.57 13.88
18.20 19.90
22.27 30.14
36.95 66.29
87.55 102.16
0.00 20.00 40.00 60.00 80.00 100.00 120.00
Thaumastura coraIcterus graceannae
Phalacrocorax brasilianusPardirallus sanguinolentus
Myrtis fannyFalco femoralis
Columba liviaSaltator striatipectus
Conirostrum cinereumCharadrius vociferus
Bubulcus ibisCatamenia analis
Myiophobus fasciatusPasser domesticusAthene cunicularia
Falco sparveriusBurhinus superciliarisParabuteo unicinctus
Tyrannus melanhcolicusSturnella bellicosa
Zonotrichia capensisCamptostoma obsoletum
Coereba flaveolaAmazilia amazilia
Thraupis episcopusSicalis flaveola
Molothrus bonariensisTroglodytes aedon
Aratinga erytrogenysVolatinia jacarina
Pyrocephalus rubinusCarduelis magellanica
Crotophaga sulcirostrisForpus coelestis
Mimus longicaudatusCoragyps atratus
Zenaida auriculataSporophila sp
Dives warszewicziPygochelidon cyanoleuca
Columbina cruzianaZenaida meloda
IARE Promedio (Ind/100m)
Espe
cies
46
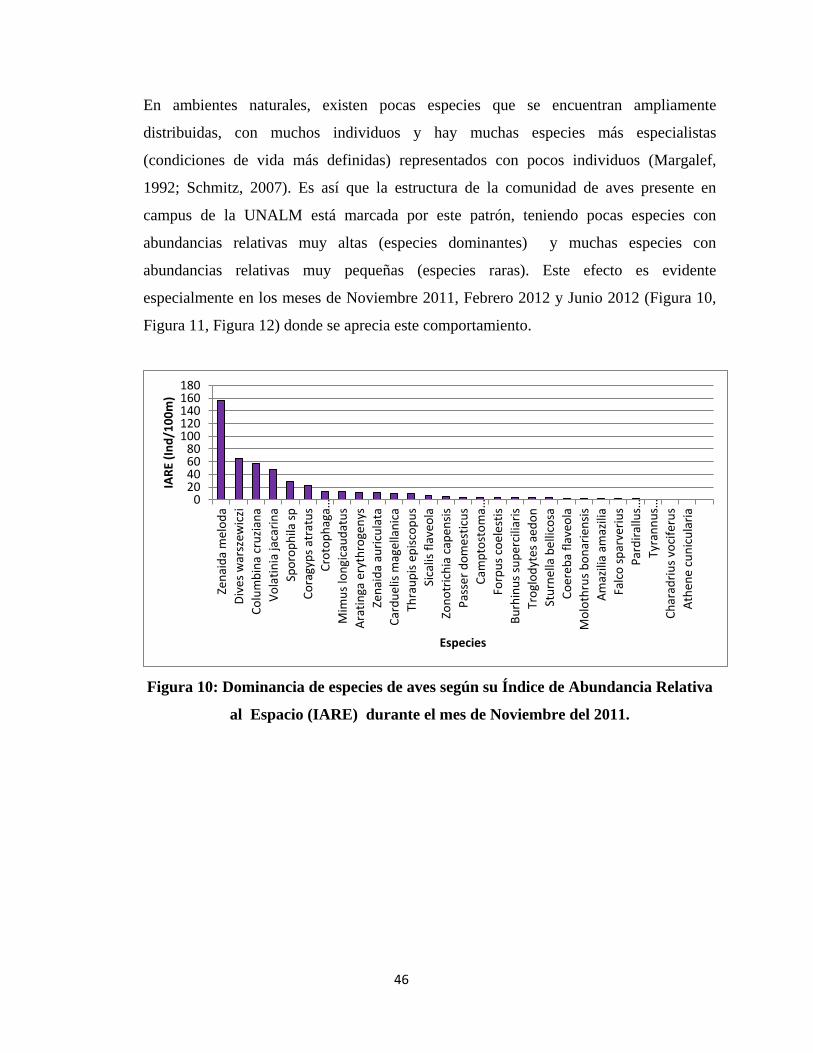
En ambientes naturales, existen pocas especies que se encuentran ampliamente
distribuidas, con muchos individuos y hay muchas especies más especialistas
(condiciones de vida más definidas) representados con pocos individuos (Margalef,
1992; Schmitz, 2007). Es así que la estructura de la comunidad de aves presente en
campus de la UNALM está marcada por este patrón, teniendo pocas especies con
abundancias relativas muy altas (especies dominantes) y muchas especies con
abundancias relativas muy pequeñas (especies raras). Este efecto es evidente
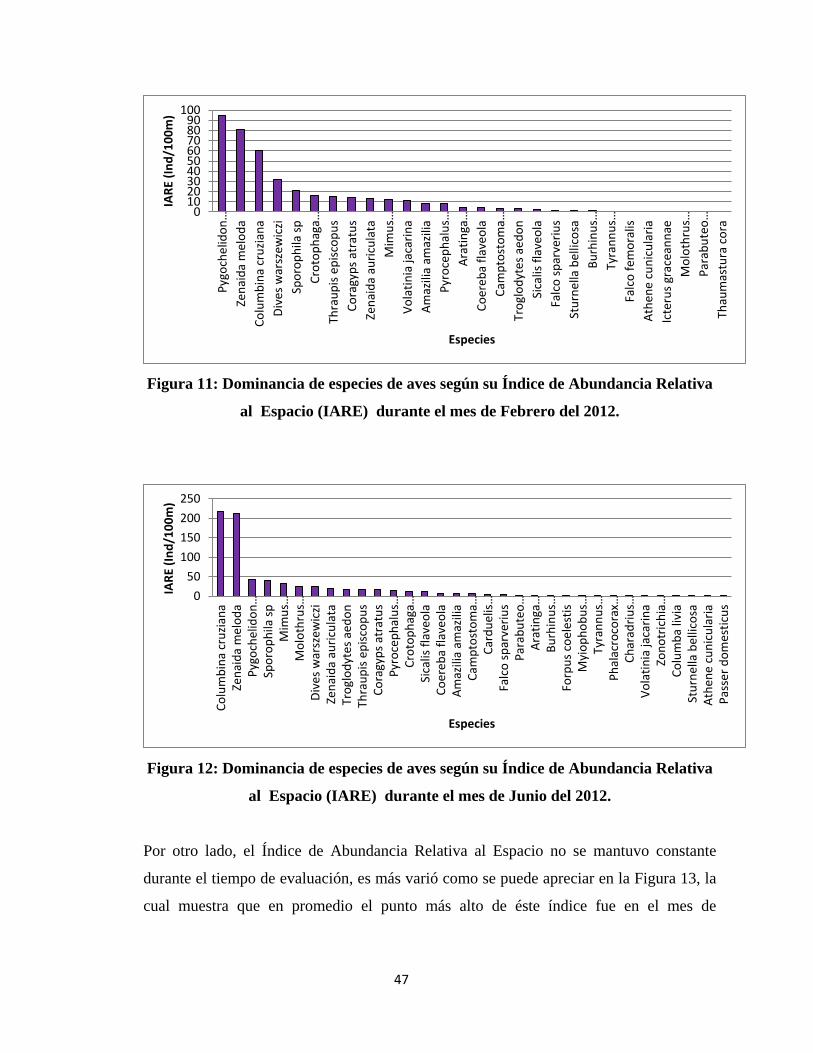
especialmente en los meses de Noviembre 2011, Febrero 2012 y Junio 2012 (Figura 10,
Figura 11, Figura 12) donde se aprecia este comportamiento.
Figura 10: Dominancia de especies de aves según su Índice de Abundancia Relativa
al Espacio (IARE) durante el mes de Noviembre del 2011.
020406080
100120140160180
Zena
ida
mel
oda
Dive
s war
szew
iczi
Colu
mbi
na c
ruzia
naVo
latin
ia ja
carin
aSp
orop
hila
spCo
ragy
ps a
trat
usCr
otop
haga
…M
imus
long
icau
datu
sAr
atin
ga e
ryth
roge
nys
Zena
ida
auric
ulat
aCa
rdue
lis m
agel
lani
caTh
raup
is ep
iscop
usSi
calis
flav
eola
Zono
tric
hia
cape
nsis
Pass
er d
omes
ticus
Cam
ptos
tom
a…Fo
rpus
coe
lest
isBu
rhin
us su
perc
iliar
isTr
oglo
dyte
s aed
onSt
urne
lla b
ellic
osa
Coer
eba
flave
ola
Mol
othr
us b
onar
iens
isAm
azili
a am
azili
aFa
lco
spar
veriu
sPa
rdira
llus…
Tyra
nnus
…Ch
arad
rius v
ocife
rus
Athe
ne c
unic
ular
ia
IARE
(Ind
/100
m)
Especies
47
Figura 11: Dominancia de especies de aves según su Índice de Abundancia Relativa
al Espacio (IARE) durante el mes de Febrero del 2012.
Figura 12: Dominancia de especies de aves según su Índice de Abundancia Relativa
al Espacio (IARE) durante el mes de Junio del 2012.
Por otro lado, el Índice de Abundancia Relativa al Espacio no se mantuvo constante
durante el tiempo de evaluación, es más varió como se puede apreciar en la Figura 13, la
cual muestra que en promedio el punto más alto de éste índice fue en el mes de
0102030405060708090
100
Pygo
chel
idon
…Ze
naid
a m
elod
aCo
lum
bina
cru
ziana
Dive
s war
szew
iczi
Spor
ophi
la sp
Crot
opha
ga…
Thra
upis
episc
opus
Cora
gyps
atr
atus
Zena
ida
auric
ulat
aM
imus
…Vo
latin
ia ja
carin
aAm
azili
a am
azili
aPy
roce
phal
us…
Arat
inga
…Co
ereb
a fla
veol
aCa
mpt
osto
ma…
Trog
lody
tes a
edon
Sica
lis fl
aveo
laFa
lco
spar
veriu
sSt
urne
lla b
ellic
osa
Burh
inus
…Ty
rann
us…
Falc
o fe
mor
alis
Athe
ne c
unic
ular
iaIc
teru
s gra
cean
nae
Mol
othr
us…
Para
bute
o…Th
aum
astu
ra c
ora
IARE
(Ind
/100
m)
Especies
050
100150200250
Colu
mbi
na c
ruzia
naZe
naid
a m
elod
aPy
goch
elid
on…
Spor
ophi
la sp
Mim
us…
Mol
othr
us…
Dive
s war
szew
iczi
Zena
ida
auric
ulat
aTr
oglo
dyte
s aed
onTh
raup
is ep
iscop
usCo
ragy
ps a
trat
usPy
roce
phal
us…
Crot
opha
ga…
Sica
lis fl
aveo
laCo
ereb
a fla
veol
aAm
azili
a am
azili
aCa
mpt
osto
ma…
Card
uelis
…Fa
lco
spar
veriu
sPa
rabu
teo…
Arat
inga
…Bu
rhin
us…
Forp
us c
oele
stis
Myi
opho
bus…
Tyra
nnus
…Ph
alac
roco
rax…
Char
adriu
s…Vo
latin
ia ja
carin
aZo
notr
ichi
a…Co
lum
ba li
via
Stur
nella
bel
licos
aAt
hene
cun
icul
aria
Pass
er d
omes
ticus
IARE
(Ind
/100
m)
Especies

48
Septiembre del 2012 (19.68 Ind/100m) seguido de cerca por el mes de Junio (18.63
Ind/100m); mientras que su punto más bajo lo alcanza en el mes de Diciembre del 2011
con 7.26 Ind/100m en promedio y Enero 2012 con 9.38 Ind/100m.
También es notable el cambio brusco que hay entre los meses de Noviembre 2011 (12.56
Ind/100m) y Diciembre del 2011 donde el IARE disminuye, siendo incluso este el punto
más bajo como se menciona antes, este efecto se invierte en el mes de Enero donde el
índice vuelve a aumentar.
Figura 13: Variación Temporal de la Abundancia (IARE Promedio Anual ,
Ind/100m) de Noviembre del 2011 a Octubre del 2012 para las especies presentes en
el Campus de la UNALM.
Los valores de Intervalo de Confianza (Anexo 7) son relativamente pequeños, lo que
indica que los promedios del IARE son hasta cierto punto precisos para los meses
evaluados, especialmente para Diciembre, Enero y Octubre, sin embargo para los meses
de Junio y Septiembre los intervalos se agrandan, teniendo en cuenta que estos fueron los
meses más abundantes este tamaño de los intervalos se explica, ya que el conteo de gran
NOV DIC ENE FEB MAR ABR MAY JUN JUL AGO SEP OCTIARE Promedio 12.56 7.26 9.38 10.39 15.27 14.78 12.93 18.63 13.02 12.39 19.68 10.01
0.0
5.0
10.0
15.0
20.0
25.0
30.0
35.0
IARE
Pro
med
io (I
nd/1
00m
)
Meses

49
cantidad de individuos disminuye la precisión del conteo incrementando el error, y
consecuente aumentando el tamaño del Intervalo de Confianza.
Utilizando el programa SURFER 10, se elaboraron mapas de contorno, con las
coordenadas de los puntos medios de los transectos y el Índice de Abundancia Relativa al
Espacio Promedio, con el fin de apreciar la distribución de la misma en el campus. Para
esto, también se ubicaron ocho puntos referenciales dentro del campus (P1: Puerta N°1,
PP: Puerta Principal, Fac: Facultad de Ciencias, JB: Jardín Botánico, C: Cerro, Po: Pozo
de agua, PF: Puerta Ferrero, OVH: Observatorio Meteorológico) y se marcaron algunos
límites del campus (números subrayados).
Como se aprecian en la Figura 17, 18, 20 y desde la Figura 22 hasta la Figura 32 la
distribución espacial de la abundancia relativa en la universidad tampoco es equitativa,
no se encuentran valores de abundancia iguales para todo el campus, existe lo que se
denominan núcleos, lugares donde se concentra la abundancia de especies en el campus.
Primero, para una visión general de la distribución de la abundancia se elaboró un mapa
con el IARE promedio para todo el periodo evaluado. Como se observa en la Figura 14
hay un núcleo importante de abundancia el cual según la Figura 15 correspondería a la
zona de los establos y cultivos cercanos (Zona 3 y Zona 4), siendo así los transecto más
abundantes el Transecto N° 34 (40.67 Ind/100m) y el Transecto N° 15 (33.07
Ind/100m), los cuales están ubicados en zona de establos y granja. También hay una
abundancia moderada en la zona de cultivos correspondiente a la Zona 1 y parte de la
Zona 2, así como en la zona de frutales (Zona 3).

50
Figura 14: Distribución de la Abundancia Relativa (IARE Promedio) de la avifauna
registrada en el campus de la UNALM (Noviembre 2011 – Octubre 2012)
Figura 15: Variación de la Abundancia Relativa (IARE Promedio) en los 40
transectos evaluados en el campus de la UNALM en el periodo evaluado
(Noviembre 2011 - Octubre 2012)
Analizando más de cerca cómo se distribuye espacialmente la Abundancia Relativa para
los meses más y menos abundantes se puede observar en la Figura 18 correspondiente al
mes de Diciembre 2011 y la Figura 19 correspondiente al mes de Enero 2012, meses
0
10
20
30
40
50
T1 T2 T3 T4 T5 T6 T7 T8 T9 T10
T11
T12
T13
T14
T15
T16
T17
T18
T19
T20
T21
T22
T23
T24
T25
T26
T27
T28
T29
T30
T31
T32
T33
T34
T35
T36
T37
T38
T39
T40
IARE
Pro
m (I
Nd/
100m
)
Transectos

51
menos abundantes, se puede apreciar que si bien es cierto presentan valores bajos de
abundancia relativa, estos están moderadamente distribuidos.
Enero muestra un núcleo en lo que corresponde a los campos de cultivo de la Zona 1, en
los Transectos N° 2 y N° 6; mientras que Diciembre presenta un núcleo en el Transecto
N° 20 (Zona 3) el cual corresponde a campos de cultivo y otro en la Zona 2 (Transecto
N° 9) (Figura 20). Esto es congruente con lo mostrado en la Figura 16, donde Diciembre
y Enero son los meses que muestran un Coeficiente de Variación (CV) menor en todo el
año de evaluación (80.799 % y 72.104 % respectivamente) respaldando la aseveración
que la abundancia no se encuentra agregada en un solo punto sino mejor distribuida en el
campus.
Para Septiembre 2012, mes más abundante, con valores altos de abundancia distribuidos
en casi todos los transectos (Figura 30) se tienen dos núcleos de abundancia altos en lo
que correspondería al Transecto N° 34 y Transecto N° 15 (granjas y establos) y en el área
de cultivos de la Zona 3 (Transecto N° 24 y Transecto N° 25) y otro núcleo entre la
Puerta Principal y la Facultad de Ciencias (Transecto N° 27) (Figura 31). El Coeficiente
de Variación para este mes es de 94.727 %, el cual es relativamente bajo, esto
respaldaría la idea que la distribución de la abundancia no está altamente agregada.
Sin embargo, Junio 2012 cuenta una historia totalmente diferente, siendo uno de los
meses con mayor abundancia de individuos en todo el año de evaluación, sin embargo es
evidente que toda esta abundancia está concentrada en un solo núcleo (Figura 26),
concentrada en el Transecto N° 34 con 216.01 Ind/100m (Figura 27) el cual se encuentra
ubicado en una zona con campos de cultivo y establos en la Zona 3. La especie
dominante en este transectos fue para este mes Columbina cruziana (Tortolita Peruana),
la cual es posible encontrarla en grandes cantidades alimentándose alrededor del ganado
cuando este a su vez está comiendo. Como se puede observar en la Figura 16, Junio es el
mes con mayor Coeficiente de Variación (182.27%), lo que indicaría un alto grado de
agregación de la abundancia relativa en el campus y una alta variabilidad en la muestra.
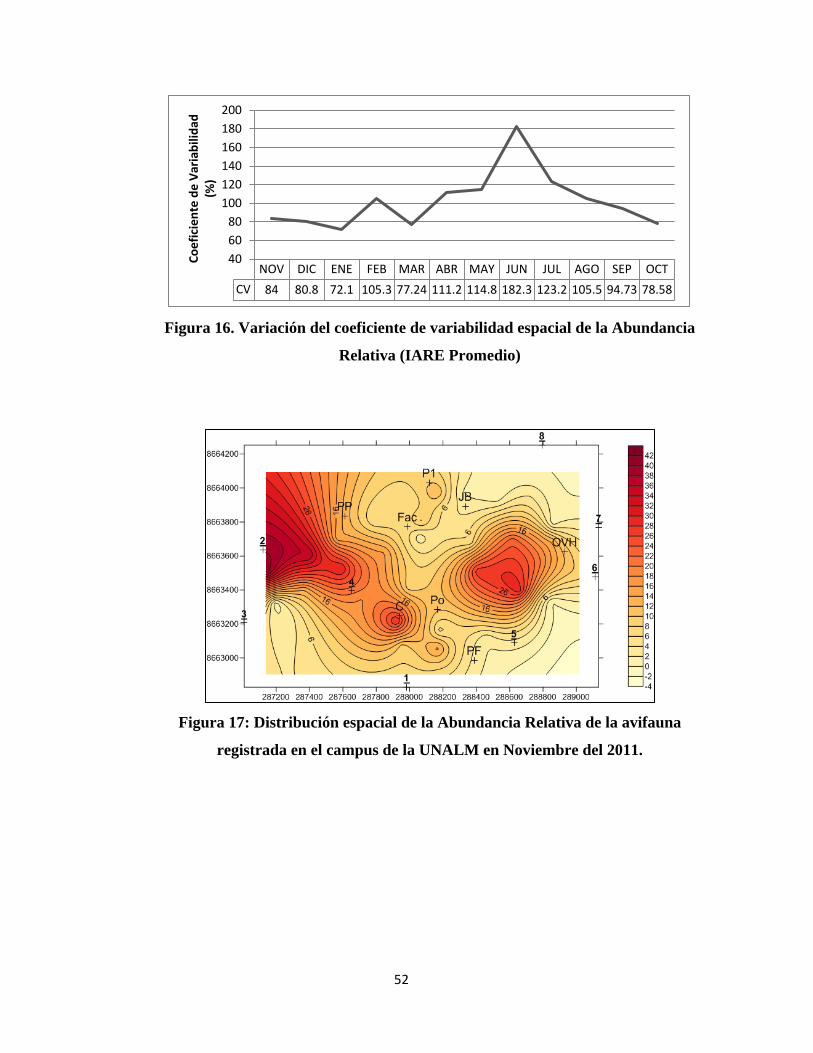
52
Figura 16. Variación del coeficiente de variabilidad espacial de la Abundancia
Relativa (IARE Promedio)
Figura 17: Distribución espacial de la Abundancia Relativa de la avifauna
registrada en el campus de la UNALM en Noviembre del 2011.
NOV DIC ENE FEB MAR ABR MAY JUN JUL AGO SEP OCTCV 84 80.8 72.1 105.3 77.24 111.2 114.8 182.3 123.2 105.5 94.73 78.58
406080
100120140160180200
Coef
icie
nte
de V
aria
bilid
ad
(%)

53
Figura 18: Distribución espacial de la Abundancia Relativa de la avifauna
registrada en el campus de la UNALM en Diciembre del 2011.
Figura 19: Variación de la Abundancia Relativa en los 40 transectos evaluados en el
campus de la UNALM en el mes de Diciembre del 2012.
0.00
5.00
10.00
15.00
20.00
25.00
30.00
T1 T2 T3 T4 T5 T6 T7 T8 T9 T10
T11
T12
T13
T14
T15
T16
T17
T18
T19
T20
T21
T22
T23
T24
T25
T26
T27
T28
T29
T30
T31
T32
T33
T34
T35
T36
T37
T38
T40
IARE
(In
d/10
0m)
Transectos

54
Figura 20: Distribución de la Abundancia Relativa de la avifauna registrada en el
campus de la UNALM en Enero del 2012.
Figura 21: Variación de la Abundancia Relativa en los 40 transectos evaluados en el
campus de la UNALM en el mes de Enero del 2012.
0.005.00
10.0015.0020.0025.0030.0035.00
T1 T2 T3 T4 T5 T6 T7 T8 T9 T10
T11
T12
T13
T14
T15
T16
T17
T18
T19
T20
T21
T22
T23
T24
T25
T26
T27
T28
T29
T30
T31
T32
T33
T34
T35
T36
T37
T38
T39
T40
IARE
(Ind
/100
m)
Transectos
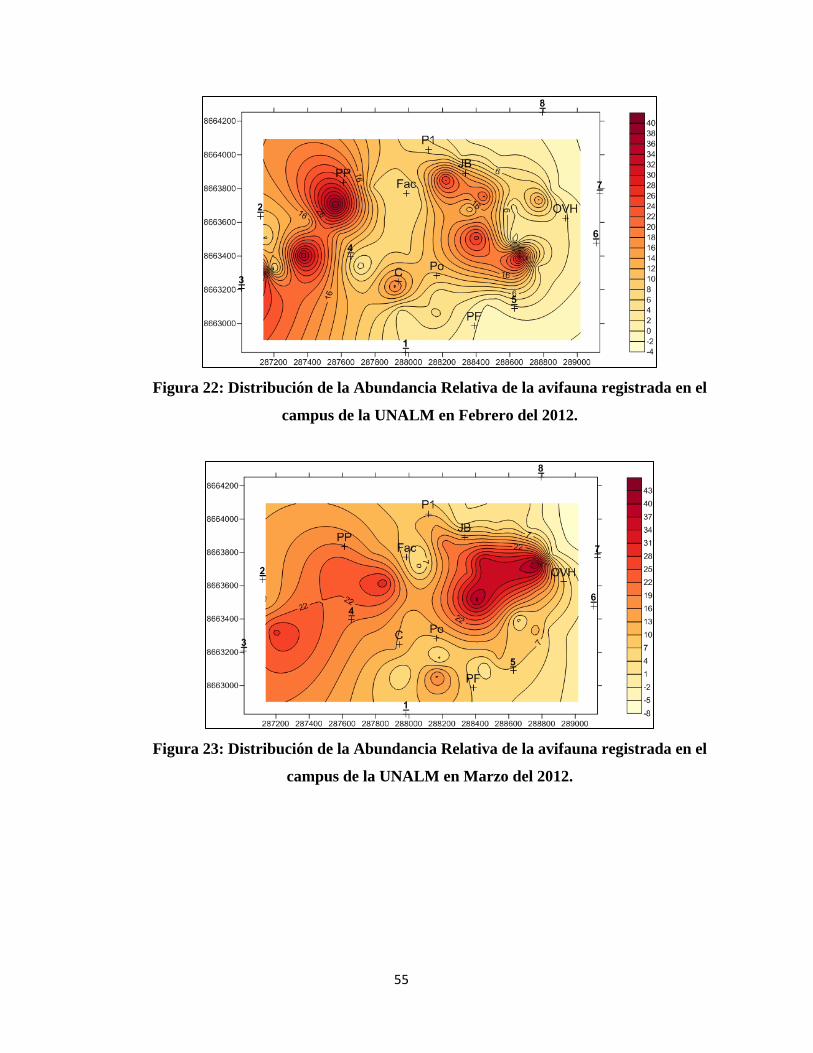
55
Figura 22: Distribución de la Abundancia Relativa de la avifauna registrada en el
campus de la UNALM en Febrero del 2012.
Figura 23: Distribución de la Abundancia Relativa de la avifauna registrada en el
campus de la UNALM en Marzo del 2012.

56
Figura 24: Distribución de la Abundancia Relativa de la avifauna registrada en el
campus de la UNALM en Abril del 2012.
Figura 25: Distribución de la Abundancia Relativa de la avifauna registrada en el
campus de la UNALM en Mayo del 2012.

57
Figura 26: Distribución de la Abundancia Relativa de la avifauna registrada en el
campus de la UNALM en Junio del 2012.
Figura27: Variación de la Abundancia Relativa en los 40 transectos evaluados en el
campus de la UNALM en el mes de Junio del 2012.
0.0050.00
100.00150.00200.00250.00
T1 T2 T3 T4 T5 T6 T7 T8 T9 T10
T11
T12
T13
T14
T15
T16
T17
T18
T19
T20
T21
T22
T23
T24
T25
T26
T27
T28
T29
T30
T31
T32
T33
T34
T35
T36
T37
T38
T39
T40
IARE
(In
d/10
0m)
Título del eje

58
Figura 28: Distribución de la Abundancia Relativa de la avifauna registrada en el
campus de la UNALM en Julio del 2012.
Figura 29: Distribución de la Abundancia Relativa de la avifauna registrada en el
campus de la UNALM en Agosto del 2012.

59
Figura 30: Distribución de la Abundancia Relativa de la avifauna registrada en el
campus de la UNALM en Septiembre del 2012.
Figura 31: Variación de la Abundancia Relativa en los 40 transectos evaluados en el
campus de la UNALM en el mes de Septiembre del 2012.
0
20
40
60
80
100
T1 T2 T3 T4 T5 T6 T7 T8 T9 T10
T11
T12
T13
T14
T15
T16
T17
T18
T19
T20
T21
T22
T23
T24
T25
T26
T27
T28
T29
T30
T31
T32
T33
T34
T35
T36
T37
T38
T39
T40
IARE
(In
d/10
0m)
Transectos

60
Figura 32: Distribución de la Abundancia Relativa de la avifauna registrada en el
campus de la UNALM en Octubre del 2012.
Con el fin de analizar de cierta manera los patrones de variabilidad espacio-temporal se
realizó un análisis basándose en la Ley de Taylor.
Como se puede observar en la Figura 33, el B espacial (Bs) es mayor a dos (Bs= 2.9974)
(Anexo 8) lo que sugeriría un comportamiento tipo Hot Spot, mientras que el B temporal
(Bt) es igual a dos (Bt=2.0054) lo cual significaría que no existe relación entre las medias
y las varianzas (Figura 34); sin embargo sus respectivos coeficientes de determinación
son demasiado bajos para tener interpretación alguna.
Figura 33: Diagrama de dispersión del Bs para el Índice de Abundancia Relativa al
Espacio.
y = 0.4987x + 1.4426 R² = 0.2976
1.81.9
22.12.22.3
0.8 0.9 1 1.1 1.2 1.3 1.4
Log
Med
ia
Log CV

61
Figura 34: Diagrama de dispersión del Bt para el Índice de Abundancia Relativa al
Espacio.
La distribución de las especies en el espacio es rara vez, equitativa, las especies tienden a
agregarse debido a sus requerimientos y ecología (Smith, 2001). Es así que, como lo
muestra la Figura 35, es la Zenaida meloda (Tortolita Melódica) la especie mejor
distribuida en todo el campus con presencia en casi 30 transectos en promedio durante el
tiempo de evaluación, de los 40 transectos evaluados, la Zenaida meloda se adapta a
diferentes ambientes que el campus de la UNALM le ofrece tal vez por su naturaleza
oportunista (Nolazco, 2012); esta especie se mantiene como la especie mejor distribuida
en el campus desde los estudios realizados por Takano & Castro (2007), así como para
las evaluaciones realizadas por el Laboratorio de Ecología de Procesos en el 2011 (
Castillo et al., en prensa). Otras especies con un rango moderado de distribución son
Dives warszewiczi (Tordo de Matorral) con casi 18 transectos usados, Pygochelidon
cyanoleuca (Golondrina Azul y Blanca) también con casi 18 transectos y Pyrocephalus
rubinus (Mosquetero Bermellón) con 17 transectos aproximadamente.
Por otro lado, entre las especies con una distribución muy restringida con valores de uso
de transectos por debajo de la unidad son varias, se puede destacar Icterus graceannae
(Bolsero de Filos Blancos) especie invasora proveniente de la costa norte del Perú
(Schulenberg et al., 2007) que en estudios anteriores también fue registrada como una
especie de distribución restringida (Takano & Castro, 2007). Algunas especies asociadas
y = 0.0028x + 1.857 R² = 3E-05 1
1.5
2
2.5
0.2 0.7 1.2 1.7Lo
g M
edia
Log CV

62
a cuerpos de agua también presentan un rango de distribución reducido como Charadrius
vociferus (Chorlo Gritón), Pardirallus sanguinolentus (Rascón Plomizo) y
Phalacrocorax brasilianus (Cormorán Neotropical) (Schulenberg et al., 2007; Takano y
Castro, 2007; Castillo et al., en prensa).
Burhinus superciliaris (Alcavarán Huerequeque) y Athene cunicularia (Lechuza
Terrestre) son especies asociados a hábitats más específicos (Schulenberg et al., 2007).
Además, ambas especies tienen comportamientos nocturnos, presentando poca actividad
durante el día, este hecho permitió observarlas en lugares específicos durante los censos
que se realizaban a tempranas horas de la mañana. También se puede mencionar a
Sturnella bellicosa (Pecho Colorado Peruano) con dos transectos en promedio usados,
especie mayormente asociada a campos de cultivo. (Takano, 2010).
Finalmente, es importante recalcar que todas estas especies de distribución agregada
mantienen este comportamiento a comparación de estudios anteriores (Takano & Castro,
2007; Castillo et al., en prensa)
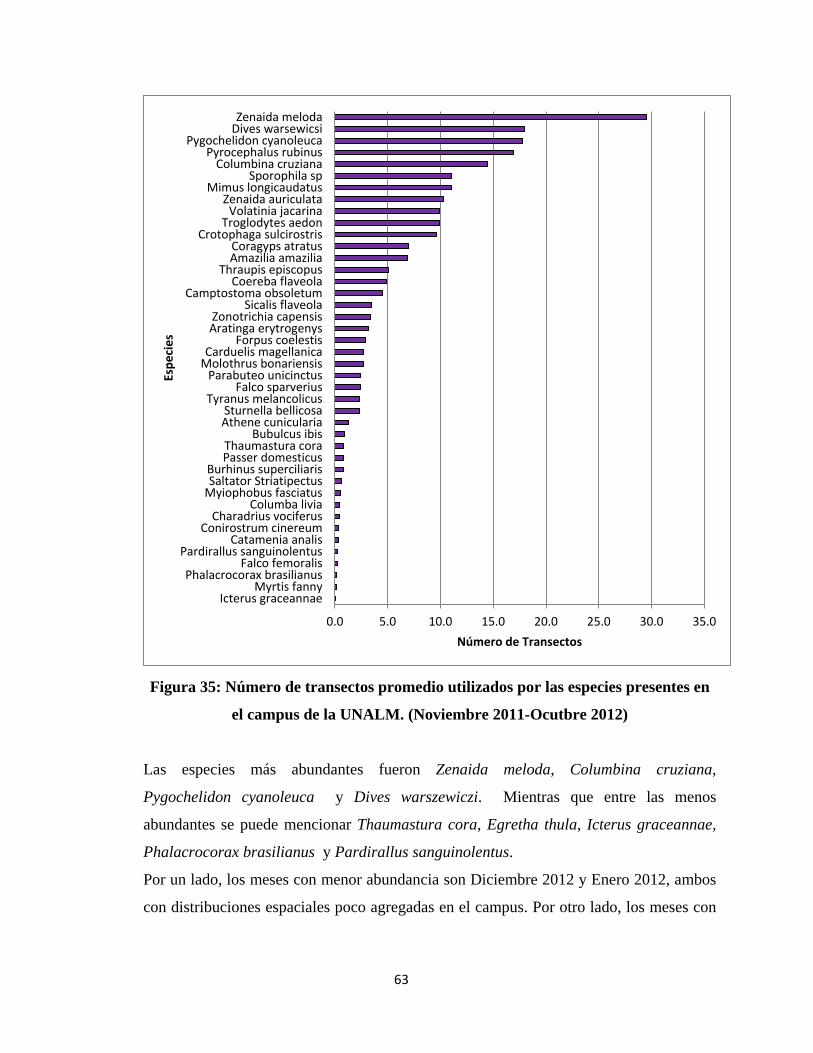
63
Figura 35: Número de transectos promedio utilizados por las especies presentes en
el campus de la UNALM. (Noviembre 2011-Ocutbre 2012)
Las especies más abundantes fueron Zenaida meloda, Columbina cruziana,
Pygochelidon cyanoleuca y Dives warszewiczi. Mientras que entre las menos
abundantes se puede mencionar Thaumastura cora, Egretha thula, Icterus graceannae,
Phalacrocorax brasilianus y Pardirallus sanguinolentus.
Por un lado, los meses con menor abundancia son Diciembre 2012 y Enero 2012, ambos
con distribuciones espaciales poco agregadas en el campus. Por otro lado, los meses con
0.0 5.0 10.0 15.0 20.0 25.0 30.0 35.0
Icterus graceannaeMyrtis fanny
Phalacrocorax brasilianusFalco femoralis
Pardirallus sanguinolentusCatamenia analis
Conirostrum cinereumCharadrius vociferus
Columba liviaMyiophobus fasciatusSaltator StriatipectusBurhinus superciliaris
Passer domesticusThaumastura cora
Bubulcus ibisAthene cuniculariaSturnella bellicosa
Tyranus melancolicusFalco sparverius
Parabuteo unicinctusMolothrus bonariensisCarduelis magellanica
Forpus coelestisAratinga erytrogenysZonotrichia capensis
Sicalis flaveolaCamptostoma obsoletum
Coereba flaveolaThraupis episcopus
Amazilia amaziliaCoragyps atratus
Crotophaga sulcirostrisTroglodytes aedon
Volatinia jacarinaZenaida auriculata
Mimus longicaudatusSporophila sp
Columbina cruzianaPyrocephalus rubinus
Pygochelidon cyanoleucaDives warsewicsiZenaida meloda
Número de Transectos
Espe
cies

64
mayor abundancia son Septiembre 2012 y Junio 2012, Septiembre presenta una
distribución espacial poco agregada, sin embargo Junio presenta una agregación espacial
muy alta con un coeficiente de variabilidad muy alto, todo concentrado en la zona de
establos.
En cuanto a la distribución espacial de la abundancia relativa no es equitativa en el
campus, teniendo un núcleo importante de abundancia en los establos y cultivos cercanos
(Zona 3 y Zona 4), siendo así los transectos más abundantes el Transecto N° 34 y N°
15. También se presenta una abundancia moderada en la zona de cultivos (Zona 1 y
Zona 2); así como en la zona de frutales (Zona 3).
Las especies mejor distribuidas en el campus son Zenaida meloda, Dives warszewiczi
Pygochelidon cyanoleuca , Pyrocephalus tubinus y Columbina cruziana; mientras que
aquellas que presentan una distribución más restringida son Icterus graceannae,
Phalacrocorax brasilianus, Burhinus superciliaris, Athene cunicularia, Charadriius
vociferus, entre otras.
4.4 ANÁLISIS ESPACIAL Y TEMPORAL DE LA DIVERSIDAD
Con los datos obtenidos en las evaluaciones también se calculó un Índice de
Diversidad utilizando el método de Shannon – Weaver con el paquete estadístico PAST,
así como sus componentes: Riqueza Especifica (R.E), estimada con el Índice de Margalef
y la Equitativibilidad (J).
Para el período evaluado correspondiente a Noviembre 2011-Octubre 2012, se puede
apreciar en la Figura 36 que el mes más diverso fue Octubre del 2012 según el índice
usado (2.0489 bits/ind) seguido muy de cerca por Junio del 2012 con 1.9376 bits/ind. Por
otra parte, los meses menos diversos fueron Julio (1.4250 bits/ind) y Mayo (1.5124
bits/ind) del 2012; aunque son los valores más bajos del tiempo evaluado.

65
Por otra parte el Índice de Margalef para Riqueza Específica indica una alta riqueza
especifica en todos los meses, ya que todos superan el valor de dos (considerado de baja
riqueza); siendo Enero del 2012 (4.444) el mes con mayor número de especies y Agosto
del 2012 con una Riqueza Específica de; los meses con menor riqueza específica, los
cuales fueron Marzo (3.747) y Febrero (3.956).
Finalmente, la equidad se mantiene casi constante para todos los meses con un valor
promedio de 0.75.
Figura 36: Variación de la Diversidad (Índice de Shannon – Weaver .H), Riqueza
Específica (Índice de Margalef – R.E) y Equidad (J) en los 40 transectos evaluados
en el campus de la UNALM durante el periodo evaluado (Noviembre 2011 –
Octubre 2012)
Teniendo la diversidad dos componentes que la explican, la riqueza específica y la
equidad, se observa en la Figura 37 y Figura 38 la diversidad está siendo mejor
explicada por la riqueza con un coeficiente de correlación muy alto (R =0.9191) más que
por la equidad que presenta un coeficiente de correlación bajo (R=0.2562), se puede
NOV DIC ENE FEB MAR ABR MAY JUN JUL AGOS SEPT OCTH 1.8681 1.7420 1.9327 1.7710 1.7429 1.7068 1.5124 1.9376 1.4250 1.6878 1.9335 2.0489J 0.721 0.8095 0.8048 0.7383 0.7857 0.7519 0.7495 0.6453 0.6565 0.7715 0.7681 0.8053R.E 4.11 4.137 4.444 3.956 3.747 4.155 3.824 4.392 4.186 4.476 4.058 4.696
00.511.522.533.544.55
0.00
0.50
1.00
1.50
2.00
2.50

66
decir que la comunidad de aves no es una comunidad climática, más bien podría ser una
comunidad intermedia en proceso de sucesión (Smith, 20001)
Figura 37: Relación entre la Equidad (J) y la Diversidad (H) de la aves de la
UNALM para el mes de Octubre del 2012
Figura 38: Relación entre la Riqueza Específica (R.E) y la Diversidad (H) de la aves
de la UNALM para el mes de Octubre del 2012
Además del análisis temporal de la diversidad también fue necesario analizar cómo se
distribuye espacialmente en el campus de la UNALM, se realizó siguiendo el mismo
procedimiento que se usó para el análisis de la abundancia relativa.
Observando la Figura 39, que muestra la distribución de la diversidad promedio para
todo el tiempo evaluado, es notable un núcleo importante de diversidad que comprenden
los ambientes urbanos de la Zona 4 y parte de la Zona 2 (cuerpo de agua, cerro y campos
de cultivo de algodón y maíz).
y = 1.9817x + 0.5083 R² = 0.2562
0
1
2
3
4
0 0.2 0.4 0.6 0.8 1 1.2
Indi
ce d
e Sh
anno
n-W
eave
r (H)
Equitatibilidad (J)
y = 1.0329x + 0.2918 R² = 0.9191
0
1
2
3
4
0 1 2 3 4Indi
ce d
e Sh
anno
n-W
eave
r
Riqueza Específica (R.E)

67
En la Figura 40, se aprecia que el Transecto N° 9 es el más diverso en especies,
Transecto que corresponde a la Zona 2 y que cuenta con varios tipos de formaciones
vegetales, además de estructuras artificiales (laboratorios), vivero, bordes vivos, canal de
regadío, árboles y campos de cultivo. Otros transectos diversos son los comprendidos
también en la Zona 2, el Transecto N° 13 el cual comprende un cerco de cañaveral y
pequeñas parcelas de cultivos experimentales, además de basura al inicio del transecto lo
cual atrae a especies generalistas y carroñeras. Hay otro núcleo pequeño en la zona de
frutales (Zona 3) cerca al Observatorio.
Dentro de las zonas menos diversas podemos apreciar la zona de campos de cultivos que
corresponde a hortalizas en la Zona 3 (Transecto N° 19) y el campo ferial donde hay
tierra descubierta y jardín en la mayor parte incluyendo el Jardín Botánico en la Zona 4
(Transectos N° 36, N° 37, N° 38 y N° 39).
Figura 39: Distribución de la Diversidad promedio de la avifauna registrada en el
campus de la UNALM durante el periodo de evaluación (Noviembre 2011 –
Octubre 2012)

68
Figura 40: Variación de la Diversidad (Índice de Diversidad de Shannon – Weaver)
en los 40 transectos evaluados en el campus de la UNALM (Noviembre 2011-
Octubre 2012)
Analizando más de cerca los meses más y menos diversos, se observa en la Figura 56
correspondiente al mes de Octubre 2012, mes más diverso para el periodo evaluado, que
existen hasta tres núcleos de diversidad, el primero correspondiente a la Zona 4 y parte
de la Zona 2, y otro correspondiente a la zona de Frutales (Transecto N° 21) en la Zona 3
(Figura 57). A esto podemos agregar lo que muestra la Figura 41, Octubre presenta un
bajo Coeficiente de Variabilidad lo que indica que la diversidad no está agregada, siendo
esto coherente con lo que muestra el mapa.
En el caso de Enero 2012, otro mes altamente diverso, los núcleos de diversidad no se
mantienen estáticos, en la Figura 44 se aprecia un núcleo considerable en la Zona 2,
correspondiente a los Transectos N° 9, N° 10 y N° 11; a diferencia de Octubre 2012 se
aprecian varios transectos donde la diversidad es cero, es decir solo se registró solo una
especie. También se aprecia un núcleo en la Zona 1 (Figura 45) correspondiente a los
Transectos N° 2 y N° 3, ambos agrícolas. Su Coeficiente de Variabilidad es bajo (43.27
%) lo que también explicaría un distribución más homogénea de la diversidad, sin
embargo al ser este valor mayor que el de Octubre y como se aprecia en la Figura 41, se
observa una distribución más agregada que el mes antes descrito.
0
0.5
1
1.5
2
2.5
3
T1 T2 T3 T4 T5 T6 T7 T8 T9 T10
T11
T12
T13
T14
T15
T16
T17
T18
T19
T20
T21
T22
T23
T24
T25
T26
T27
T28
T29
T30
T31
T32
T33
T34
T35
T36
T37
T38
T39
T40
Indi
ce d
e Sh
anno
n Pr
omed
io
(bits
)
Transectos
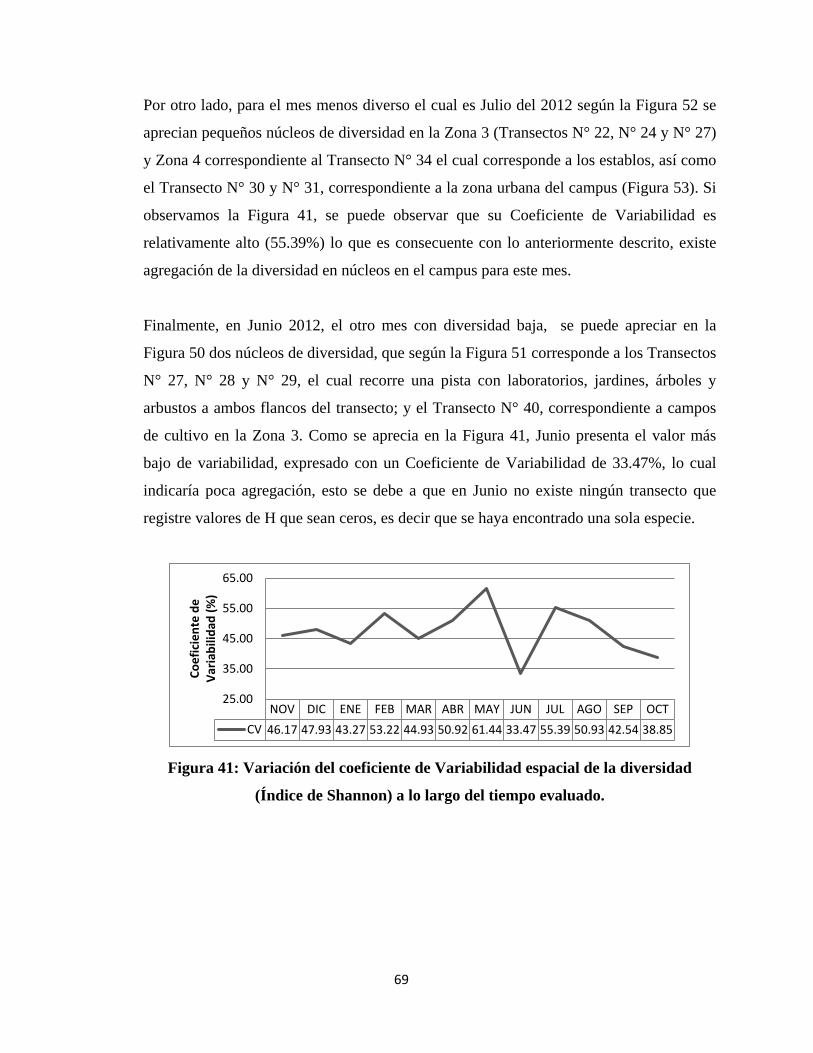
69
Por otro lado, para el mes menos diverso el cual es Julio del 2012 según la Figura 52 se
aprecian pequeños núcleos de diversidad en la Zona 3 (Transectos N° 22, N° 24 y N° 27)
y Zona 4 correspondiente al Transecto N° 34 el cual corresponde a los establos, así como
el Transecto N° 30 y N° 31, correspondiente a la zona urbana del campus (Figura 53). Si
observamos la Figura 41, se puede observar que su Coeficiente de Variabilidad es
relativamente alto (55.39%) lo que es consecuente con lo anteriormente descrito, existe
agregación de la diversidad en núcleos en el campus para este mes.
Finalmente, en Junio 2012, el otro mes con diversidad baja, se puede apreciar en la
Figura 50 dos núcleos de diversidad, que según la Figura 51 corresponde a los Transectos
N° 27, N° 28 y N° 29, el cual recorre una pista con laboratorios, jardines, árboles y
arbustos a ambos flancos del transecto; y el Transecto N° 40, correspondiente a campos
de cultivo en la Zona 3. Como se aprecia en la Figura 41, Junio presenta el valor más
bajo de variabilidad, expresado con un Coeficiente de Variabilidad de 33.47%, lo cual
indicaría poca agregación, esto se debe a que en Junio no existe ningún transecto que
registre valores de H que sean ceros, es decir que se haya encontrado una sola especie.
Figura 41: Variación del coeficiente de Variabilidad espacial de la diversidad
(Índice de Shannon) a lo largo del tiempo evaluado.
NOV DIC ENE FEB MAR ABR MAY JUN JUL AGO SEP OCTCV 46.17 47.93 43.27 53.22 44.93 50.92 61.44 33.47 55.39 50.93 42.54 38.85
25.00
35.00
45.00
55.00
65.00
Coef
icie
nte
de
Varia
bilid
ad (%
)

70
Figura 42: Distribución de la Diversidad de la avifauna registrada en el campus de
la UNALM en Noviembre del 2011.
Figura 43: Distribución de la Diversidad de la avifauna registrada en el campus de
la UNALM en Diciembre del 2011.
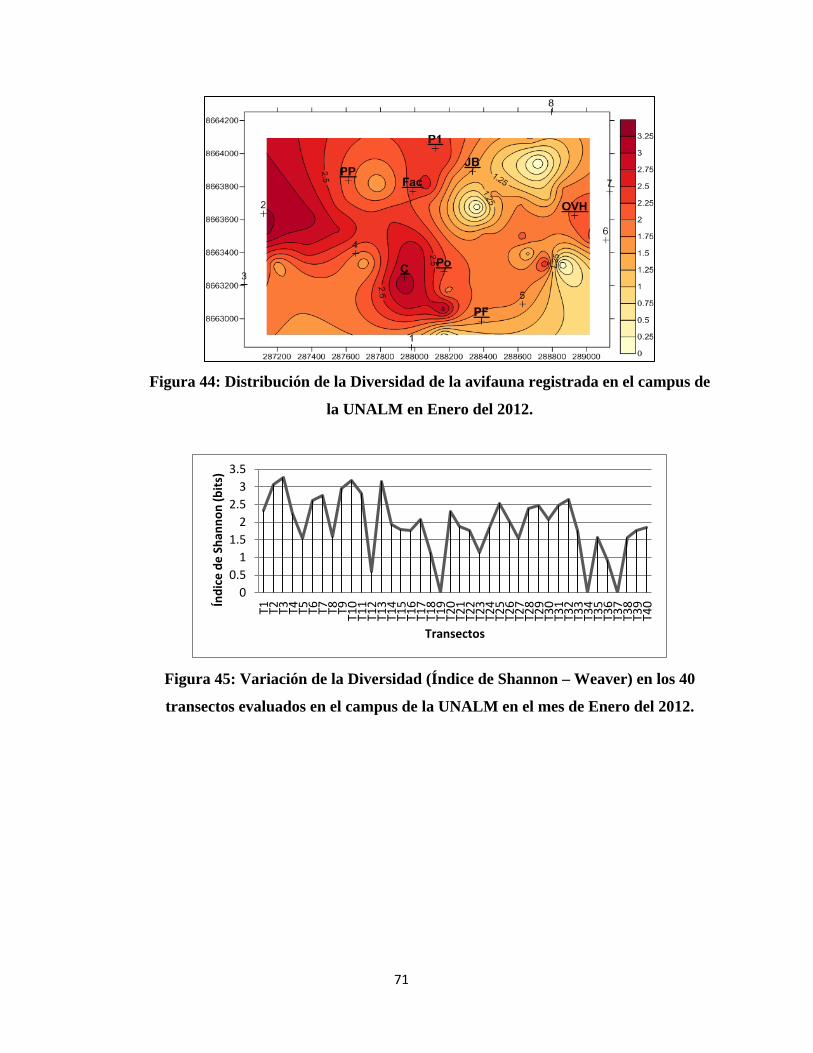
71
Figura 44: Distribución de la Diversidad de la avifauna registrada en el campus de
la UNALM en Enero del 2012.
Figura 45: Variación de la Diversidad (Índice de Shannon – Weaver) en los 40
transectos evaluados en el campus de la UNALM en el mes de Enero del 2012.
00.5
11.5
22.5
33.5
T1 T2 T3 T4 T5 T6 T7 T8 T9 T10
T11
T12
T13
T14
T15
T16
T17
T18
T19
T20
T21
T22
T23
T24
T25
T26
T27
T28
T29
T30
T31
T32
T33
T34
T35
T36
T37
T38
T39
T40Ín
dice
de
Shan
non
(bits
)
Transectos

72
Figura 46: Distribución de la Diversidad de la avifauna registrada en el campus de
la UNALM en Febrero del 2012.
Figura 47: Distribución de la Diversidad de la avifauna registrada en el campus de
la UNALM en Marzo del 2012.
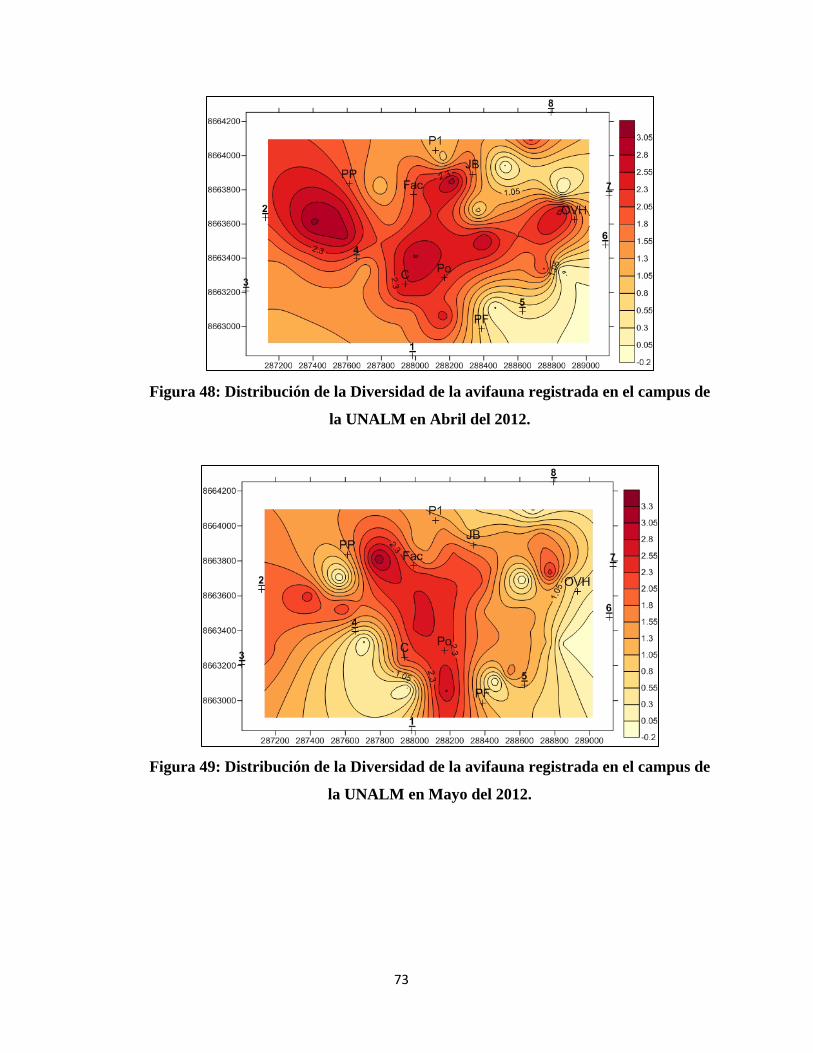
73
Figura 48: Distribución de la Diversidad de la avifauna registrada en el campus de
la UNALM en Abril del 2012.
Figura 49: Distribución de la Diversidad de la avifauna registrada en el campus de
la UNALM en Mayo del 2012.

74
Figura 50: Distribución de la Diversidad de la avifauna registrada en el campus de la
UNALM en Junio del 2012.
Figura 51: Variación de la (Índice de Shannon – Weaver), en los 40 transectos
evaluados en el campus de la UNALM en el mes de Junio del 2012.
00.5
11.5
22.5
33.5
T1 T2 T3 T4 T5 T6 T7 T8 T9 T10
T11
T12
T13
T14
T15
T16
T17
T18
T19
T20
T21
T22
T23
T24
T25
T26
T27
T28
T29
T30
T31
T32
T33
T34
T35
T36
T37
T38
T39
T40Ín
dice
de
Shan
non
(bits
9
Transectos

75
Figura 52: Distribución de la Diversidad de la avifauna registrada en el campus de
la UNALM en Julio del 2012.
Figura 53: Variación de la Diversidad (Índice de Shannon – Weaver) en los 40
transectos evaluados en el campus de la UNALM en el mes de Julio del 2012.
00.5
11.5
22.5
33.5
T1 T2 T3 T4 T5 T6 T7 T8 T9 T10
T11
T12
T13
T14
T15
T16
T17
T18
T19
T20
T21
T22
T23
T24
T25
T26
T27
T28
T29
T30
T31
T32
T33
T34
T35
T36
T37
T38
T39
T40Ín
dice
de
Shan
non
(bits
)
Transectos
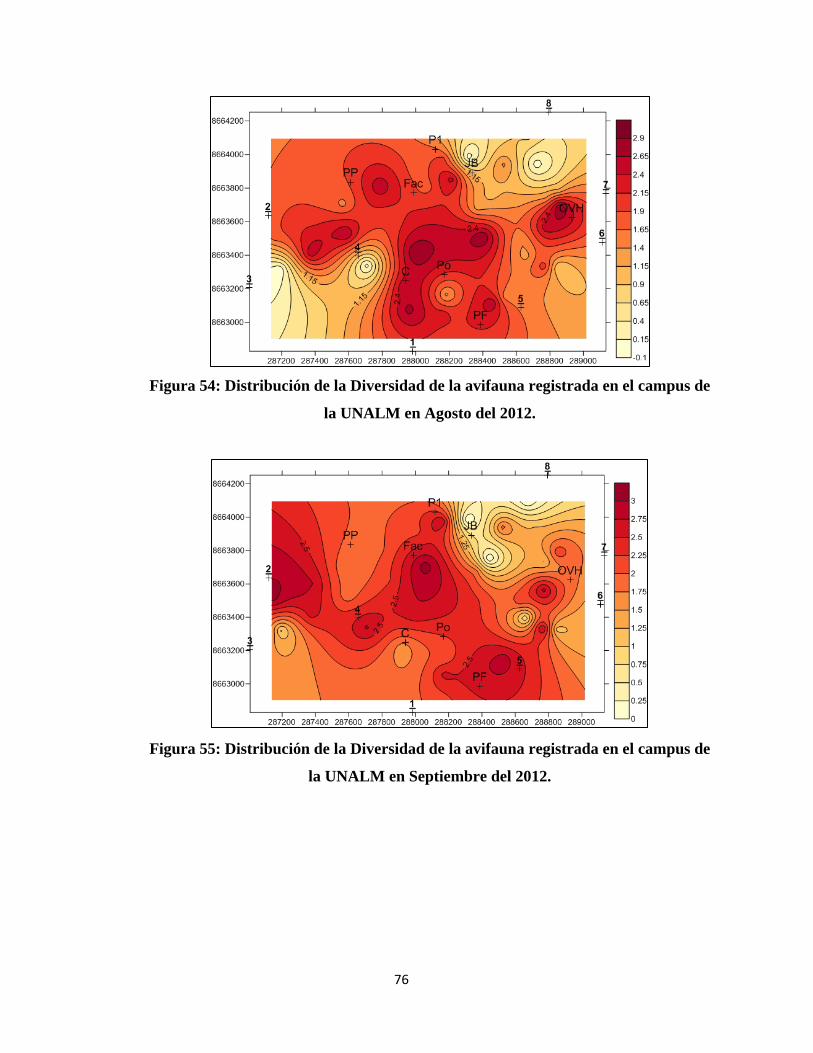
76
Figura 54: Distribución de la Diversidad de la avifauna registrada en el campus de
la UNALM en Agosto del 2012.
Figura 55: Distribución de la Diversidad de la avifauna registrada en el campus de
la UNALM en Septiembre del 2012.

77
Figura 56: Distribución de la Diversidad de la avifauna registrada en el campus de
la UNALM en Octubre del 2012.
Figura 57: Variación de la Diversidad (Índice de Shannon – Weaver), en los 40
transectos evaluados en el campus de la UNALM en el mes de Octubre del 2012.
Para tener una mejor idea del comportamiento espacial y temporal de la comunidad, se
desarrolló un análisis a partir de la Ley de Taylor, de la misma manera que se realizó
para la Abundancia Relativa en la sección anterior.
0
0.5
1
1.5
2
2.5
3
3.5
T1 T2 T3 T4 T5 T6 T7 T8 T9 T10
T11
T12
T13
T14
T15
T16
T17
T18
T19
T21
T22
T23
T24
T25
T26
T27
T28
T29
T30
T31
T32
T33
T34
T35
T36
T38
T39
T40
Índi
ce d
e Sh
anno
n (b
its)
Transectos

78
Como se observa en la Figura 58, el coeficiente de determinación es lo suficientemente
grande para ser aceptable. El B espacial (bs) es menor a dos (Bs= -0.5798) (Anexo 10);
para el caso del comportamiento temporal, con un coeficiente de determinación lo
suficientemente grande para tomarse en cuenta y un B temporal menor a dos (Bt= -
0.7842) (Anexo 10). Teniendo que BS<2 y Bt<2, se entiende que la comunidad se
“refugia”. Es decir, la comunidad en los Transectos o sitios de buena calidad como son
los Transectos N° 9 y N° 13 (Figura 59) es menos variable en el tiempo que en los
Transectos o Sitios de mala calidad (Transecto N° 19 y N° 37); mientras que en los
meses menos favorables como Mayo y Julio la comunidad tiende a refugiarse en algunos
sitios, los más favorables, en este caso los Transectos N° 9 y N° 13, aumentando así la
variabilidad espacial (Gitaldo et al., 2002; Teixeira & Sánchez, 2006).
Figura 58: Diagrama de dispersión del B espacial para el Índice de Diversidad de
Shannon - Weaver.
y = -1.2897x + 1.9895 R² = 0.6914
1.51.61.71.81.9
0.1 0.15 0.2 0.25 0.3 0.35
Log
CV
Log Media
Diagrama de dispersión del B espacial para H

79
Figura 59: Diagrama de dispersión del B temporal para el Índice de Diversidad de
Shannon - Weaver.
Es importante mencionar que existe en la naturaleza una relación entre la complejidad
del hábitat y la diversidad específica, mientras más complejo sea el hábitat o la cobertura
vegetal, mayor será la diversidad y riqueza de la avifauna (González, sf; Nolazco, 2012;
Véliz 2006).
Como se ha podido observar en la mayoría de las figuras anteriores, transectos muy
diversos en su avifauna han sido los transectos más complejos en cuanto a formaciones
vegetales como por ejemplo el Transecto N° 9, el cual es el más diverso en cuanto a
hábitat y avifauna se refiere. Por otro lado, Transectos más homogéneos como el
Transecto N° 40 o con poca cobertura vegetal como el Transecto N° 37, presentan una
menor diversidad de aves. Es por esto, que se realizó un análisis de regresión entre la
diversidad alfa promedio de las aves y la diversidad de los elementos del hábitat, como se
aprecia en la Figura 60 el coeficiente de determinación es demasiado bajo para que exista
una relación consistente; esto se puede deber a la naturaleza de los datos de vegetación,
cuya matriz está construida en base a datos de presencia y ausencia más que datos
cuantitativos, o a la posibilidad que existan otros elementos que definen la distribución
de las especies.
y = -1.3921x + 1.9166 R² = 0.8348
1
1.2
1.4
1.6
1.8
2
2.2
2.4
-0.3 -0.2 -0.1 0 0.1 0.2 0.3 0.4 0.5

80
Figura 60: Relación entre la Diversidad Alfa Promedio de las Aves (bits/ind) y la
Diversidad de los Elementos del Hábitat (bits)
En general, el campus de la UNALM se presenta como un lugar de alta diversidad de
aves, los meses más diversos y con mayor riqueza específica fueron Octubre 2012 y
Enero 2012, los cuales no presentaron una distribución espacial agregada en el campus.
Los meses menos diversos fueron Julio y Junio; para el caso de Julio presenta una alta
agregación espacial de la diversidad, mientras que Junio con un coeficiente de
variabilidad bajo presenta una agregación espacial muy baja. Los meses con menor
riqueza específica fueron Marzo y Febrero. Finalmente, la equidad se mantiene para
todos los meses con un valor promedio de 0.75.
En cuanto a la distribución espacial de la diversidad, ésta no se mantiene constante
durante los diferentes meses de estudio; en promedio existe un núcleo importante en los
ambientes urbanos de la Zona 4 y parte de la Zona 2. Mientras que las zonas menos
diversas están los campos de cultivos que corresponde a la Zona 3 con cultivo de
hortalizas, así como el Campo Ferial y Jardín Botánico en la Zona 4.
El análisis espacio-temporal indica que la comunidad de aves tiende al refugio (Bs<,
Bt<2) durante los meses desfavorables (Mayo y Junio) en los sitios favorables o de
buena calidad (Transectos N° 9 y N° 13).
Finalmente, a pesar de que parecería existir una relación entre la diversidad de los
elementos del hábitat y de la diversidad de las aves, no es posible comprobarla
y = 0.6996x - 0.5416 R² = 0.0544
0.0000
0.5000
1.0000
1.5000
2.0000
2.5000
3.0000
2.2000 2.4000 2.6000 2.8000 3.0000 3.2000 3.4000 3.6000
H (h
abita
t)
H (aves)

81
estadísticamente, posiblemente por la naturaleza de los datos de hábitat o la presencia de
otros factores involucrados.
4.5 ANÁLISIS MULTIVARIADO DE LA ABUNDANCIA RELATIVA,
DIVERSIDAD Y ELEMENTOS DE HABITAT
Con los datos de abundancia relativa, diversidad y la matriz de elementos del
hábitat se realizaron análisis multivariados de Clúster obteniendo dendrogramas con el
fin de explicar las similitudes con los datos de Abundancias Relativas, Diversidad y
Elementos del Hábitat entre meses evaluados y entre los transectos. Para algunos casos se
realizaron también Análisis de Componentes Principales.
4.5.1 ANÁLISIS MULTIVARIADO DE LA ABUNDANCIA RELATIVA
Utilizando el Índice de Abundancia Relativa al Espacio (IARE) se elaboró un
dendrograma (Figura 61) con los meses evaluados para observar las asociaciones
temporales. Se tiene que para distancias de enlace mayor a 1.8 no se forman grupos. Se
pueden observar 3 grupos marcados por debajo de este enlace:
• Grupo 1:
- Noviembre 2011
- Diciembre 2011
- Enero 2012
- Febrero 2012
- Marzo 2012
- Abril 2012
- Mayo 2012

82
• Grupo 2:
- Junio 2012
- Julio 2012
• Grupo 3:
- Agosto 2012
- Septiembre 2012
- Octubre 2012
Figura 61: Dendrograma de datos del Índice de Abundancia Relativa al Espacio de
las aves presentes en el campus de la UNALM (Noviembre 2011-Octubre 2012)
Distancias euclidianas
Por otro lado, se tiene que el análisis One Way NP MANOVA con p-valor representativo
menor a 0.05 (p= 0.0121) existen diferencias significativas entre el Grupo 1 y el Grupo 2,
sin embargo el Grupo 3 no presenta diferencias con ninguno de los otros dos grupos
(Cuadro 6). Junio y Julio son lo meses con mayor coeficiente de variabilidad, es decir los
2.0
1.8
1.6
1.4
1.2
1.0
0.8
0.6
0.4
0.2
0.0
Dista
nce
nov-
11
dic-1
1
ene-
12
feb-
12
mar
-12
abr-1
2
may
-12
jun-1
2
jul-1
2
ago-
12
sep-
12
oct-1
2

83
más agregados espacialmente, esto podría ser una causa para su separación de los otros
dos grupos.
Cuadro 6: One Way NP MANOVA con p=0.0121 para los tres grupos formados a
partir de la Abundancia Relativa. “Elaboración Propia”. Grupo 1 Grupo 2 Grupo 3
Grupo 1 0 0.0295 0.4576
Grupo 2 0.0295 0 0.0978
Grupo 3 0.4576 0.0978 0
Por otro lado, el Análisis de Componentes Principales muestra dos grupos marcados, el
cual corresponde al Grupo 1 y al Grupo 3 que se separan y por otro lado el Grupo 2 que
se desplaza en el componente 1, que estaría explicado en parte por la presencia de
Columbina cruziana y Zenaida meloda (Figura 62). Mientras que el Grupo 3 (meses de
Agosto, Septiembre y Octubre) que se mueve en el componente dos se explica
principalmente por la presencia de Pygochelidon cyanoleuca, ya que en el mes de
Septiembre presenta su mayor abundancia (Anexo 12).

84
Figura 62: Gráfico de Análisis de Componentes Principales para los meses
evaluados usando los Índices de Abundancias Relativas al Espacio.
Para el caso de las agrupaciones espaciales de los transectos, además del Análisis Clúster
también se realizó un Análisis de Compontes Principales. Se puede observar en la Figura
63 que se forma un núcleo importante que agrupa los transectos de la Zona 2 y Zona 3,
principalmente agrícola. Otro núcleo que agrupa en casi su totalidad a los transectos de la
Zona 1, totalmente agrícola, además de dos transectos (Transecto N° 9 y N° 10) de la
Zona 2, los cuales estarán explicados según el Análisis de Componentes Principales, en
su mayoría por las abundancias de Dives warsewicsi, Pygochelidon cyanoleuca y
Zenaida meloda, como lo muestra la Figura 64. También se puede observar como los
Transectos N° 34 y N° 15 se separan del resto, transectos ubicados en los establos que
presentan gran abundancia de la especie Columbina cruziana, lo cual se confirma en la
figura en la que se muestra el Análisis de Componentes Principales, donde también se
separan del resto y están explicados por la abundancia de Columbina cruziana. Hay otro

85
grupo que se forma con los transectos N° 21, N° 26, N° 33 y N° 35 según la figura 63,
que también son explicados en cierta manera por Columbina cruziana como muestra la
Figura 64.
Figura 63: Dendrograma de datos del índice de Abundancia Relativa al Espacio
(IARE) de las aves presentes en los 40 transectos del campus de la UNALM en el
periodo evaluado (Noviembre 2011-Octubre 2012) Distancias euclidianas

86
Figura 64: Gráfico de Análisis de Componentes Principales para los meses evaluados
usando los Índices de Abundancias Relativas al Espacio.
Resumiendo, la abundancia relativa temporalmente se agrupa en tres grupos, Junio y Julio
se diferencian del resto, posiblemente por ser los meses donde la abundancia
espacialmente se agrega más y estar marcado por la presencia de Columbina cruziana, la
cual presenta su mayor abundancia en estos meses.
Finalmente, espacialmente la abundancia relativa se divide en dos grupos importantes,
transectos de la Zona 1, principalmente agrícola explicada por especies como asociadas a
campos de cultivo como Dives warsewicsi, Pygochelidon cyanoleuca y Zenaida meloda;
otro conformado por transectos de la Zona 2 y Zona 3, además de la separación de los
Transectos N° 15 y N° 34, ubicados en los establos y asociados a la alta abundancia de
Columbian cruziana.

87
5.5.2 ANÁLISIS MULTIVARIADO DE LA DIVERSIDAD
Utilizando el Índice de Shannon – Weaver (bits) se elaboró un dendrograma con
los meses evaluados para observar las asociaciones temporales según la diversidad.
Como resultado se obtuvo que para distancias de enlace mayores de 8.0, no se forman
grupos (Figura 65). Se pueden observar 2 grupos marcados por debajo de ésta distancia:
• Grupo 1:
- Noviembre 2011
- Diciembre 2011
- Enero 2012
- Febrero 2012
- Marzo 2012
• Grupo 2:
- Mayo 2012
- Junio 2012
- Julio 2012
- Agosto 2012
- Septiembre 2012
- Octubre 2012
El mes de Abril se separa del resto y no forma parte de ningún grupo, esto se debe a que
dicha evaluación no se pudo completar en su totalidad en una misma fecha, por lo que se
tuvo que realizar en dos fechas diferentes, produciéndose un error que es evidente en el
dendrograma.

88
Figura 65: Dendrograma de datos de la diversidad de las aves presentes en el
campus de la UNALM (Noviembre 2011-Octubre 2012) Distancias Euclidianas
Según la prueba de One Way NP MANOVA, con un p valor no significativo de 0.1094
(p>0.05 no significativo) la prueba no es significativa, no existen diferencias entre ellos.
Por un lado, en el Análisis de Componentes Principales, muestra al Grupo 1 que se
mueve en el componente 3 y está asociado principalmente a especies como Zonotrichia
capensis, Pygochelidon cyanoleuca, Sturnella bellicosa, entre otras. Por otro lado, el
Grupo 2 que se mueve a lo largo del componente 1 se encuentra asociado a la presencia
de especies como Columbina cruziana, Volatinia jacarina, Thraupis episcopus, entre
otras (Figura 66).
8.0
7.2
6.4
5.6
4.8
4.0
3.2
2.4
1.6
0.8
0.0
Dista
nce
nov-
11
dic-1
1
ene-
12
feb-1
2
mar-1
2
abr-1
2
may-
12
jun-1
2
jul-1
2
ago-
12
sep-
12
oct-1
2

89
Figura 66: Gráfico de Análisis de Componentes Principales para los meses
evaluados usando la diversidad de especies.
Para el caso de la organización espacial de los transectos de acuerdo a su diversidad, se
puede observar en el dendrograma de la Figura 67 dos grupos importantes. El primero
compuesto por transectos con campos de cultivo de la Zona 1 y Zona 3, el Transecto N° 11
de la Zona 2 con matorrales, así como transectos de la Zona 4 entre los cuales resalta el
Transecto N° 34, perteneciente a los establos y el Transecto N° 39 el Jardín Botánico.
El otro grupo está compuesto por transectos de naturaleza agrícola de la Zona 1, Zona 2 y
Zona 3, de ésta última se resalta la presencia del Transecto N° 15 ubicado en los establos,
también se observan transectos del área urbana del campus (Zona 4).
Finalmente, se observa la separación del Transecto N° 40 por un lado y los Transectos N°
19 y N° 37 por otro.

90
Figura 67: Dendrograma de datos de la diversidad de las aves presentes en los 40
transectos del campus de la UNALM en el periodo evaluado (Noviembre 2011-
Octubre 2012) Distancias euclidianas
En conclusión, desde el punto de vista de la diversidad, temporalmente no existen
agrupaciones, es decir la diversidad no varía durante el año. Espacialmente hablando, los
transectos se agrupan en dos grupos importantes, con transectos de las diversas zonas del
campus, se nota la separación del Transecto N° 40, por un lado y de los Transectos N° 19
y N° 37, estos dos últimos son considerados como sitios malos en el análisis tipo Taylor,
es decir son lugares con alta variabilidad pero baja diversidad los cuales podrían
desaparecer; por otro lado los lugares considerados en el análisis Taylor indica sitios
consistentemente buenos a los Transectos N° 9, N° 13 y N° 31, los cuales se ubica en el
segundo grupo observado en el dendrograma.

91
5.5.3 ANÁLISIS MULTIVARIADO DE LOS ELEMENTOS DEL HABITAT
Se realizó el análisis de la asociación temporal de los meses evaluados según los
elementos del hábitat, al no observarse grupos consistentes (Figura 68), no se profundizo
en el análisis.
Figura 68: Dendrograma de datos de la elementos del hábitat presentes en el
campus de la UNALM (Noviembre 2011-Octubre 2012) Distancias Euclidianas
Si se observa la Figura 69 se observan dos grupos importantes el primero y más grande lo
conforman transectos ubicados en campos de cultivo diferentes zonas, tanto en la Zona 1
como en la Zona 2 y Zona 3. El otro grupo que se forma a continuación comprende
transecto ubicados en zonas más urbanas (Zona 4 y parte de la Zona 2). Este hecho se ve
respaldado por la Figura 70, donde se aprecia el Análisis de Componentes Principales
que muestra los transectos de naturaleza más agrícola ubicados en el lado izquierdo
asociados a elementos del hábitat como Campo de Cultivo en Barbecho, Campos de
Cultivo en Arado, Campos de Cultivo en Estado Vegetativo y Reproductivo de diversos
cultivos; y los transectos de naturaleza más urbana en el lado derecho asociadas a
30
27
24
21
18
15
12
9
6
3
Dis
tanc
e
ene-
12
feb-
12
may
-12
mar
-12
jun-
12
jul-1
2
ago-
12
sep-
12
oct-1
2
nov-
11
abr-
12
dic-
11

92
elementos del hábitat típicos de la Zona 4, como los son Arboles, Jardines,
Construcciones, etc.
Figura 69: Dendrograma de datos de presencia de los elementos del hábitat en los
40 transectos del campus de la UNALM en el periodo evaluado (Noviembre 2011-
Octubre 2012) Distancias Euclidianas
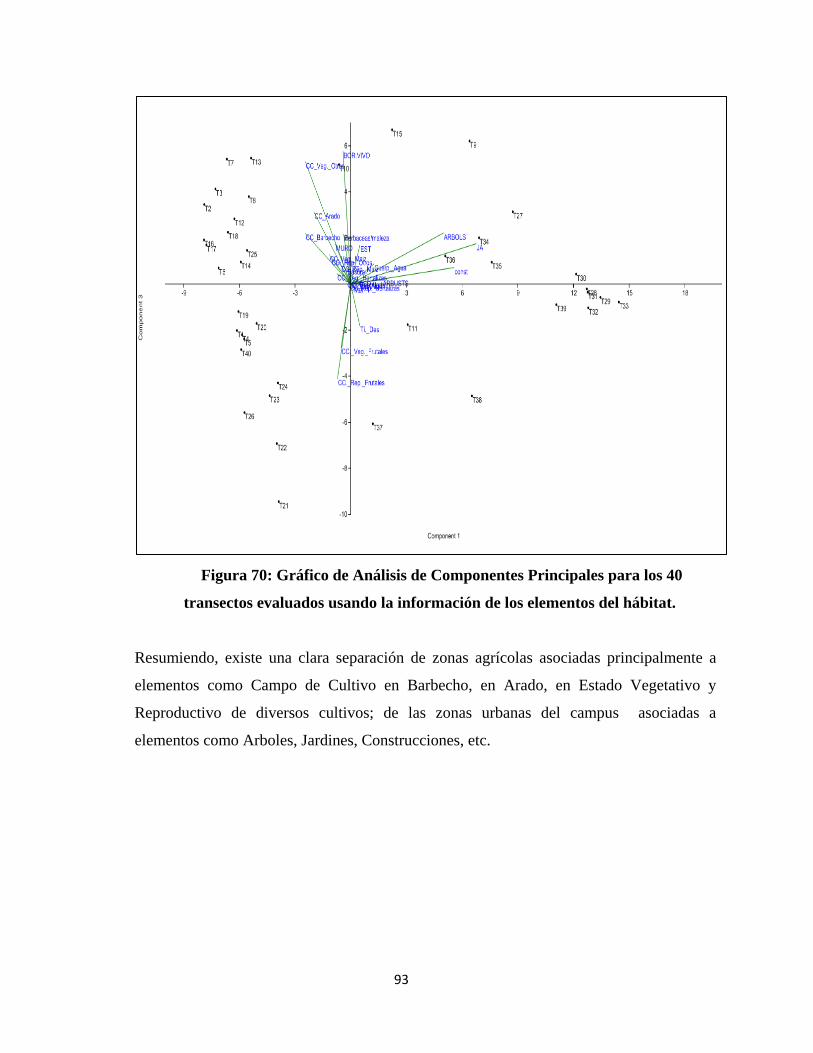
93
Figura 70: Gráfico de Análisis de Componentes Principales para los 40
transectos evaluados usando la información de los elementos del hábitat.
Resumiendo, existe una clara separación de zonas agrícolas asociadas principalmente a
elementos como Campo de Cultivo en Barbecho, en Arado, en Estado Vegetativo y
Reproductivo de diversos cultivos; de las zonas urbanas del campus asociadas a
elementos como Arboles, Jardines, Construcciones, etc.
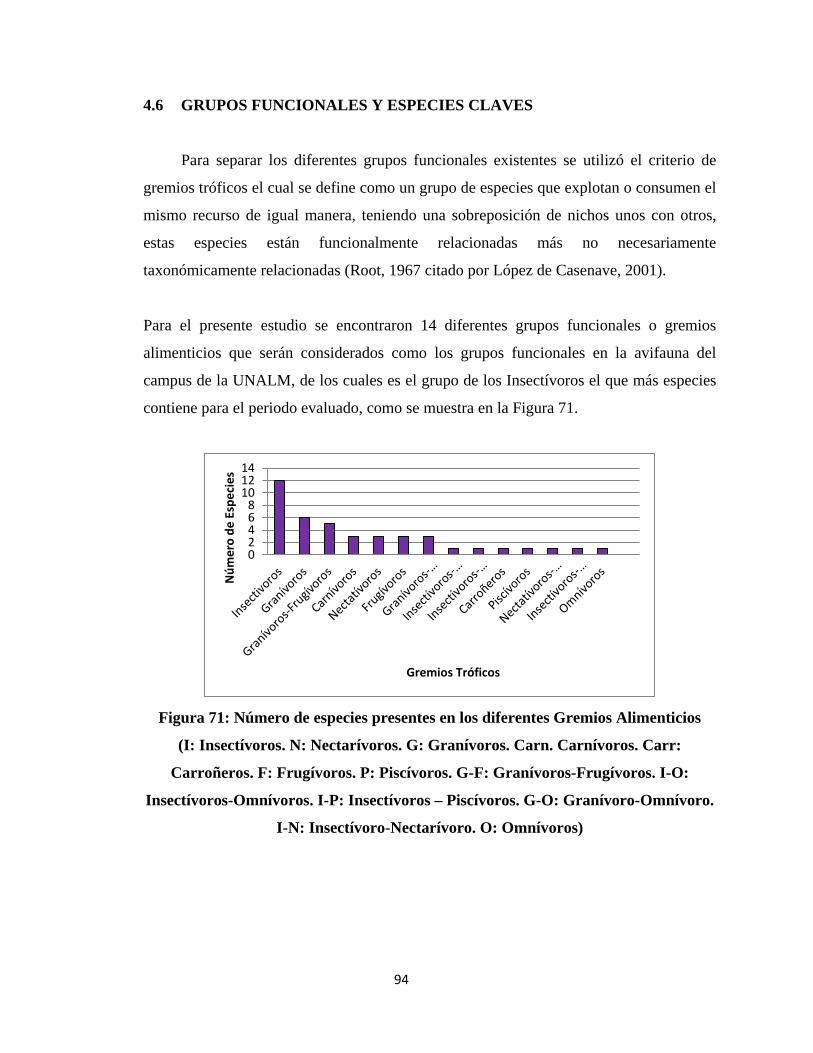
94
4.6 GRUPOS FUNCIONALES Y ESPECIES CLAVES
Para separar los diferentes grupos funcionales existentes se utilizó el criterio de
gremios tróficos el cual se define como un grupo de especies que explotan o consumen el
mismo recurso de igual manera, teniendo una sobreposición de nichos unos con otros,
estas especies están funcionalmente relacionadas más no necesariamente
taxonómicamente relacionadas (Root, 1967 citado por López de Casenave, 2001).
Para el presente estudio se encontraron 14 diferentes grupos funcionales o gremios
alimenticios que serán considerados como los grupos funcionales en la avifauna del
campus de la UNALM, de los cuales es el grupo de los Insectívoros el que más especies
contiene para el periodo evaluado, como se muestra en la Figura 71.
Figura 71: Número de especies presentes en los diferentes Gremios Alimenticios
(I: Insectívoros. N: Nectarívoros. G: Granívoros. Carn. Carnívoros. Carr:
Carroñeros. F: Frugívoros. P: Piscívoros. G-F: Granívoros-Frugívoros. I-O:
Insectívoros-Omnívoros. I-P: Insectívoros – Piscívoros. G-O: Granívoro-Omnívoro.
I-N: Insectívoro-Nectarívoro. O: Omnívoros)
02468
101214
Núm
ero
de E
spec
ies
Gremios Tróficos

95
4.6.1. GRUPO FUNCIONAL INSECTÍVOROS: (I)
Compuesto por aquellas especies que consumen artrópodos terrestres o voladores
(Mols & Visser, 2002 citado por Del Hoyo, 2006). Es el grupo con mayor número de
especies, siendo 12 especies de aves en total. Utilizado el IARE (Ind/100m) se halló la
dominancia de especies dentro de cada grupo funcional; para los Insectívoros
Pygochelidon cyanoleuca (Golondrina Azul y Blanca) es la más abundante (66.29
Ind/100m), como se aprecia en la Figura 72.
Es importante destacar que dentro de este grupo se pueden distinguir dos subgrupos, las
especies insectívoras que hurgan en el suelo para la búsqueda de su alimento como
Molothrus bonariensis (Tordo Brilloso), Burhinus superciliaris (Alcavarán
Huerequeque), Charadrius vociferus (Chorlo Gritón), Troglodytes aedon (Cucarachero
Común) y Pardirallus sanguinolentus (Rascón Plomizo); y las especies que atrapan los
insectos al vuelo como Pygchelidon cyanoleuca (Golondrina Azul y Blanca),
Pyrocephalus rubinus (Mosquetero Bermellón), Myiophobus fasciatus (Mosquerito de
Pecho Rayado), Camptostoma obsoletum (Mosquerito Silbador) y Tyranus
melancholicus (Tirano Tropical) (Santana, 2011; Nolazco, 2012; Ortiz, 2012).
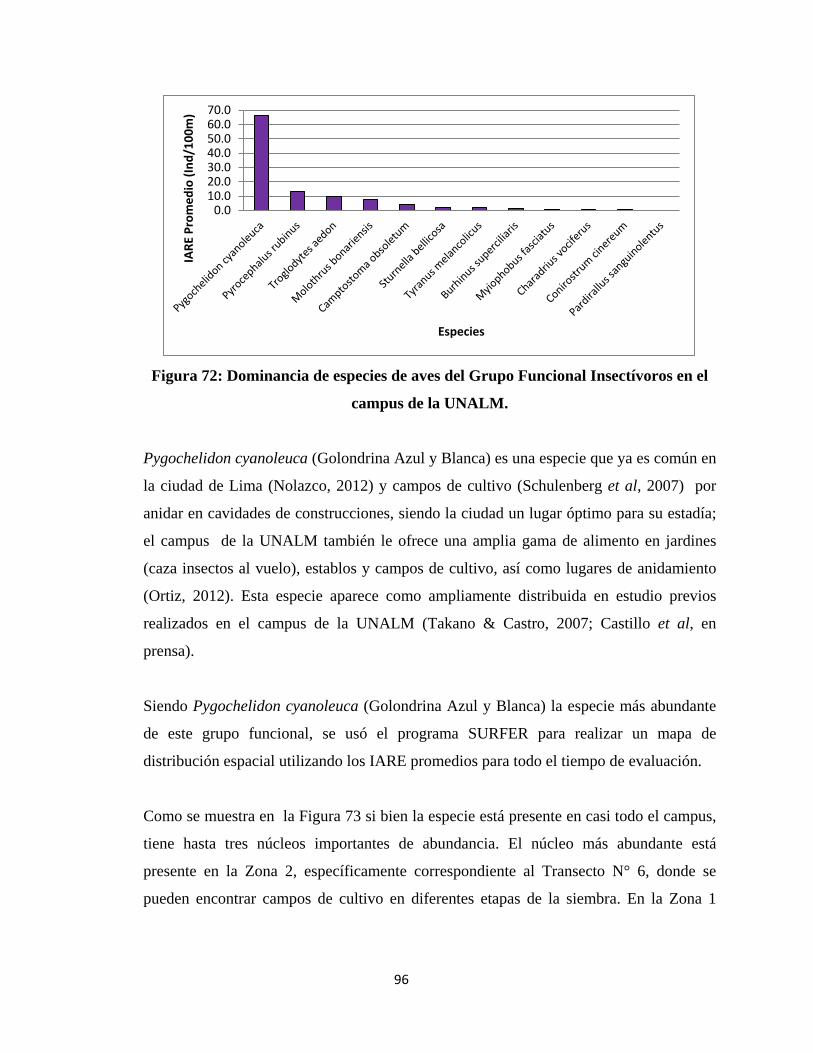
96
Figura 72: Dominancia de especies de aves del Grupo Funcional Insectívoros en el
campus de la UNALM.
Pygochelidon cyanoleuca (Golondrina Azul y Blanca) es una especie que ya es común en
la ciudad de Lima (Nolazco, 2012) y campos de cultivo (Schulenberg et al, 2007) por
anidar en cavidades de construcciones, siendo la ciudad un lugar óptimo para su estadía;
el campus de la UNALM también le ofrece una amplia gama de alimento en jardines
(caza insectos al vuelo), establos y campos de cultivo, así como lugares de anidamiento
(Ortiz, 2012). Esta especie aparece como ampliamente distribuida en estudio previos
realizados en el campus de la UNALM (Takano & Castro, 2007; Castillo et al, en
prensa).
Siendo Pygochelidon cyanoleuca (Golondrina Azul y Blanca) la especie más abundante
de este grupo funcional, se usó el programa SURFER para realizar un mapa de
distribución espacial utilizando los IARE promedios para todo el tiempo de evaluación.
Como se muestra en la Figura 73 si bien la especie está presente en casi todo el campus,
tiene hasta tres núcleos importantes de abundancia. El núcleo más abundante está
presente en la Zona 2, específicamente correspondiente al Transecto N° 6, donde se
pueden encontrar campos de cultivo en diferentes etapas de la siembra. En la Zona 1
0.010.020.030.040.050.060.070.0
IARE
Pro
med
io (I
nd/1
00m
)
Especies

97
también se ubica otro núcleo, cerca de la Puerta Principal (PP) este núcleo corresponde al
Transecto N° 1 el cual también presenta campos de cultivos a ambos lados del transecto.
Por último, se observa un núcleo en la Zona 3, correspondiente a los Transectos N° 16,
N° 17 y N° 18, los cuales corresponden también a campos de cultivo en diferentes fases
de la siembra como barbecho o arado, maíz, hortalizas, entre otros (Figura 74).
El análisis de Regresión Múltiple realizado no mostró significancia, con un p valor no
significativo de 0.06 (p significativo < 0.05), lo que nos dice que el hábitat parecería no
determinar su distribución.
Figura 73: Distribución espacial de la Abundancia Relativa (IARE Promedio) de
Pygochelidon cyanoleuca en el campus de la UNALM en el periodo evaluado
(Noviembre 2011 – Octubre 2012)

98
Figura 74: Variación de la Abundancia Relativa (IARE Promedio) en los 40
transectos evaluados en el campus de la UNALM para Pygochelidon cyanoleuca
durante el periodo evaluado (Noviembre 2011-Octubre 2012)
Se consideró también el análisis espacial de Burhinus superciliaris (Alcavarán
Huerequeque), no por su abundancia que no es dominante si no por ser una especie
reconocida por la población universitaria (Corrales, 2012). Es una especie cuyo hábitat
tradicional son los matorrales desérticos y campos de cultivo recién arados en zonas
bajas de la costa (Schulenberg et al, 2007), donde busca los insectos de los que se
alimenta (Santana, 2011). Se observa en la Figura 75 que presenta un único núcleo
ubicado en la Zona 4, específicamente en los Transectos N° 30 y N° 31 (Figura 76), lo
cual corresponde a transectos que cruzan los salones con jardines y zonas descampadas.
Es en esta zona se han observado nidos de esta especie. Sin embargo, el análisis de
Regresión Múltiple realizado no mostró significancia, ya que se obtuvo un p-valor no
significativo de 0.7 (p significativo < 0.05), lo que nos dice que el hábitat no
determinaría su distribución.
012345678
T1 T2 T3 T4 T5 T6 T7 T8 T9 T10
T11
T12
T13
T14
T15
T16
T17
T18
T19
T20
T21
T22
T23
T24
T25
T26
T27
T28
T29
T30
T31
T32
T33
T34
T35
T36
T37
T38
T39
T40IA
RE P
rom
edio
(Ind
/100
m)
Transectos
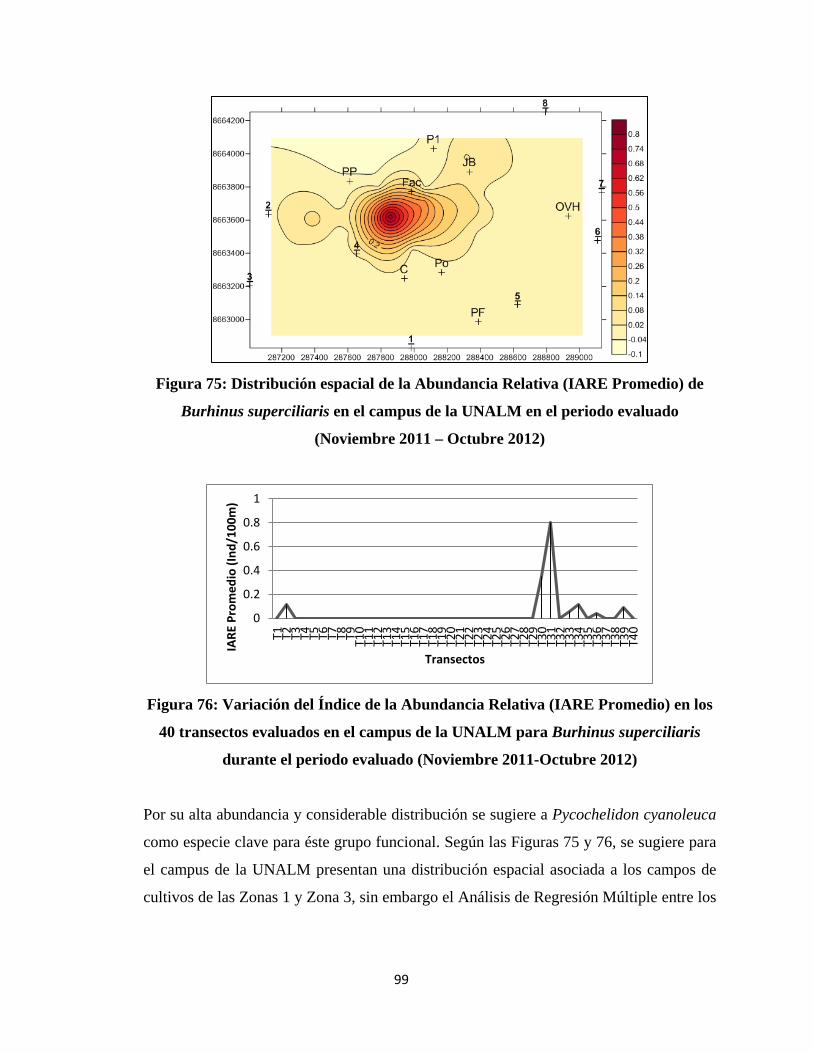
99
Figura 75: Distribución espacial de la Abundancia Relativa (IARE Promedio) de
Burhinus superciliaris en el campus de la UNALM en el periodo evaluado
(Noviembre 2011 – Octubre 2012)
Figura 76: Variación del Índice de la Abundancia Relativa (IARE Promedio) en los
40 transectos evaluados en el campus de la UNALM para Burhinus superciliaris
durante el periodo evaluado (Noviembre 2011-Octubre 2012)
Por su alta abundancia y considerable distribución se sugiere a Pycochelidon cyanoleuca
como especie clave para éste grupo funcional. Según las Figuras 75 y 76, se sugiere para
el campus de la UNALM presentan una distribución espacial asociada a los campos de
cultivos de las Zonas 1 y Zona 3, sin embargo el Análisis de Regresión Múltiple entre los
0
0.2
0.4
0.6
0.8
1
T1 T2 T3 T4 T5 T6 T7 T8 T9 T10
T11
T12
T13
T14
T15
T16
T17
T18
T19
T20
T21
T22
T23
T24
T25
T26
T27
T28
T29
T30
T31
T32
T33
T34
T35
T36
T37
T38
T39
T40
IARE
Pro
med
io (I
nd/1
00m
)
Transectos

100
elementos del hábitat y su abundancia relativa no muestra significancia suficiente para
poder afirmar que el hábitat rige su distribución espacial. Por otro, lado el Burhinus
superciliaris, tampoco parece estar distribuido de acuerdo al hábitat.
4.6.2. GRUPO FUNCIONAL CARNÍVOROS: (CARN)
Son aquellas especies de aves rapaces que se alimentan de otros vertebrados (Del
Hoyo, 2006). Son tres las especies registradas en el campus de la UNALM, de las cuales
la más abundante es Parabuteo unicinctus (Gavilán Mixto) con 1.7230 Ind/100m seguido
de cerca por Falco sparverius (Cernícalo Americano) con 1.3864 Ind/100m; por otro
lado Falco femoralis (Halcón Aplomado) es el menos abundante ya tiene una presencia
ocasional en el campus (Figura 77).
Figura 77. Dominancia de especies de aves del Grupo Funcional Carnívoros en el
campus de la UNALM.
Como se mencionó antes la especie más abundante de este grupo es Parabuteo
unicinctus, especie cuyos hábitats son los bosques secos abiertos, matorrales y campos,
además de ser avistado en áreas urbanas (Schulenberg et al, 2007; Nolazco, 2012). En el
campus de la UNALM está presente en varias zonas, pero presenta un solo núcleo
importante de abundancia (Figura 78), el cual corresponde a la Zona 3, según la Figura
79 este núcleo corresponde a los Transecto N° 19 y Transecto N° 24, también tiene
presencia en la Zona 4 cerca la Puerta N° 1 y en la Zona 2 cerca al cerro y el bosque.
0.0
0.5
1.0
1.5
2.0
Parabuteounicinctus
Falco sparverius Falco femoralis
IARE
Pro
med
io
(Ind/
100m
)
Especies

101
Debido a que los individuos han sido avistados con crías en diferentes épocas del año
sería útil monitorear estas zonas en busca de nidos, además de coincidir estas zonas con
presencia de árboles de alto porte, las cuales son propicias para el anidamiento de esta
especie (Salvador, 2012).
El análisis de Regresión Múltiple nos indica que el hábitat no determinaría su
distribución, puesto que se obtuvo un p valor de 0.18 no significativo (p significativo <
0.05).
Figura 78: Distribución espacial de la Abundancia Relativa (IARE Promedio) de
Parabuteo unicinctus en el campus de la UNALM en el periodo evaluado
(Noviembre 2011 – Octubre 2012)

102
Figura 79: Variación de la Abundancia Relativa (IARE Promedio) en los 40
transectos evaluados en el campus de la UNALM para Parabuteo unicinctus durante
el periodo evaluado (Noviembre 2011-Octubre 2012)
En conclusión, siendo el Parabuteo unicinctus la especie más abundante para este grupo
funcional, se le considera que podría ser la especie clave para este grupo. Según el mapa
de distribución espacial se observa que solo presenta un núcleo importante
correspondiente a la Zona 3 y 2, donde se observan árboles de alto porte los cuales
podrían pensarse los usa para anidar, sin embargo no se cuenta con suficiente respaldo
estadístico para afirmar esto puesto que la prueba de Regresión Múltiple no es lo
suficientemente significativa.
4.6.3. GRUPO FUNCIONAL GRANÍVOROS: (G)
Compuesto por aquellas especies que se alimentan de granos de diferentes
cultivos vegetales, como maíz, cebada, etc y de diferentes tamaños. Para este grupo hay 6
especies, siendo Columbina cruziana (Tortolita Peruana) la más abundante con un Índice
de Abundancia Relativa al Espacio (IARE) de 87.5493 Ind/100m. También en este
grupo, con Índices de Abundancias Relativas al Espacio muchos menores, está Zenaida
auriculata (Tortolita Orejuda) con 22.2741 Ind/100m, Carduelis magellanica (Jilguero
Encapuchado con 13.5357 Ind/100m) y Volatinia jacarina (Semillerito Negro Azulado)
con 12.9137 Ind/100m; también se encuentran en este grupo con abundancias muy
pequeñas Catamenia analis (Semillero de Cola Bandeada) con 0.5083 Ind/100m y
00.10.20.30.40.50.6
T1 T2 T3 T4 T5 T6 T7 T8 T9 T10
T11
T12
T13
T14
T15
T16
T17
T18
T19
T20
T21
T22
T23
T24
T25
T26
T27
T28
T29
T30
T31
T32
T33
T34
T35
T36
T37
T38
T39
T40
IARE
Pro
med
io
(Ind/
100m
)
Transectos

103
Columbia livia (Paloma Doméstica) con 0.2140 Ind/10m, las cuales tiene registros
ocasionales (Figura 80).
Figura 80: Dominancia de especies de aves del Grupo Funcional Granívoros en el
campus de la UNALM.
Para el caso específico de la Columbina cruziana, ésta especie presenta una alta
abundancia, siendo incluso la segunda especie más abundante para todo el campus, sin
embargo tiene una distribución más agregada como ya se había mencionado en secciones
anteriores. Según Schulenberg et al. (2007) se le puede encontrar en valles y todo tipo de
hábitat abiertos como jardines, parques, campos y matorrales. Para el campus de la
UNALM, se le ha registrado comúnmente en tierra desnuda y frecuentemente visita los
establos en búsqueda de alimento, también está presente en campos de cultivo de la Zona
3; es poco vista cerca de la zona urbana del campus (Castro & Takano, 2007; Castillo et
al., en prensa).
Como se puede observar en la Figura 82, existe un solo núcleo impórtate de abundancia
para esta especie, el cual corresponde a la Zona 3 y según la Figura 68 es la ubicada en
los Transectos N° 15 y Transecto N° 34, los cuales corresponden al Establo, como se
había mencionado antes se ha observado a esta especie en grandes cantidades en el
establo alrededor del ganado cuando este se está alimentando, recogiendo granos de la
tierra; si bien no está ausente en el resto de la universidad prefiere campos de cultivo. No
0.0
20.0
40.0
60.0
80.0
100.0
Columbinacruziana
Zenaidaauriculata
Sporagasmagellanica
Volatiniajacarina
Catameniaanalis
ColumbaliviaIA
RE P
rom
edio
(Ind
/100
m)
Especies

104
obstante, según el Análisis de Regresión Múltiple el hábitat no determinaría su
distribución, puesto que se obtuvo un p valor de 0.6 no significativo (p significativo <
0.05).
Figura 81: Distribución espacial de la Abundancia Relativa (IARE Promedio) de
Columbina cruziana en el campus de la UNALM en el periodo evaluado (Noviembre
2011 – Octubre 2012)
Figura 82: Variación del Índice de Abundancia Relativa (IARE Promedio) en los 40
transectos evaluados en el campus de la UNALM para Parabuteo unicinctus durante
el periodo evaluado (Noviembre 2011-Octubre 2012)
0
5
10
15
20
T1 T2 T3 T4 T5 T6 T7 T8 T9 T10
T11
T12
T13
T14
T15
T16
T17
T18
T19
T20
T21
T22
T23
T24
T25
T26
T27
T28
T29
T30
T31
T32
T33
T34
T35
T36
T37
T38
T39
T40
IARE
Pro
med
io
(Ind/
100m
)
Transectos

105
Finalmente, la especie clave de éste grupo de Granívoros podría ser Columbina cruziana
debido a su alta abundancia en comparación con el resto de especies presentes en este
grupo. Ésta especie se encuentra asociada a campos de cultivo y a la zona de establos de
donde se alimenta según observaciones y el mapa de distribución espacial de la figura,
sin embargo, según el Análisis de Regresión Múltiple entre los elementos del hábitat y la
abundancia de la especie en el campus, no habría evidencia estadística para afirmar el
hecho de que la distribución de los elementos del hábitat influencien en la distribución
del ave.
4.6.4. GRUPO FUNCIONAL NECTARÍVOROS: (N)
Conformados por todas aquellas especies que consumen néctar de las flores,
cumpliendo la función de polinizadores en el ecosistema (Del Hoyo, 2006). Son tres las
especies que conforman este grupo funcional todas de la Familia Trochilidae, siendo
Amazilia amazilia (Colibrí de Vientre Rufo) la más abundante con 5.2650 Ind/100m
(Figura 83). Por otro lado, Myrtis fanny (Estrellita de Collar Púrpura) con 0.087
Ind/100m y Thamastura cora (Colibrí de Cora) con 0.029 Ind/100m son poco abundantes
por tener registros raros, no solo para el campus en estudios anteriores si no para la
ciudad de Lima (Takano &Castro, 2007; Nolazco, 2012).
Figura 83: Dominancia de especies de aves del Grupo Funcional Nectarívoros en el
campus de la UNALM.
0.0
1.0
2.0
3.0
4.0
5.0
6.0
Amazilia amazilia Myrtis fanny Thaumastura coraIARE
Pro
med
io (I
nd/1
00m
)
Especies
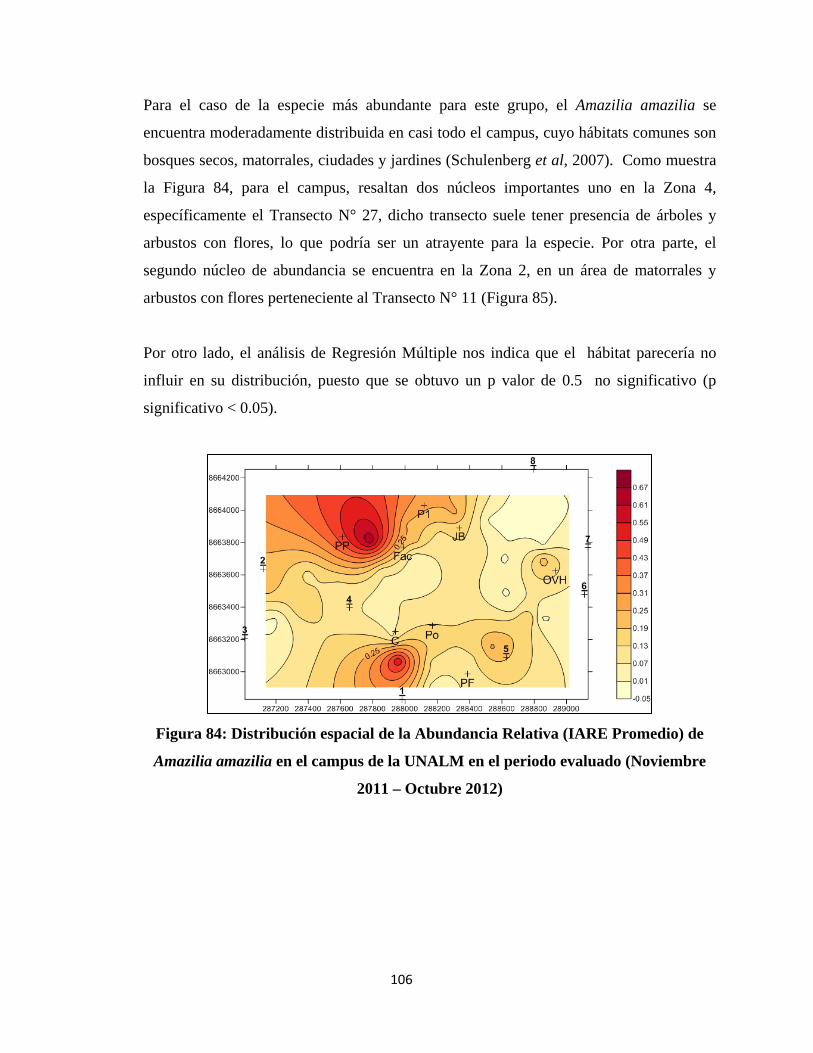
106
Para el caso de la especie más abundante para este grupo, el Amazilia amazilia se
encuentra moderadamente distribuida en casi todo el campus, cuyo hábitats comunes son
bosques secos, matorrales, ciudades y jardines (Schulenberg et al, 2007). Como muestra
la Figura 84, para el campus, resaltan dos núcleos importantes uno en la Zona 4,
específicamente el Transecto N° 27, dicho transecto suele tener presencia de árboles y
arbustos con flores, lo que podría ser un atrayente para la especie. Por otra parte, el
segundo núcleo de abundancia se encuentra en la Zona 2, en un área de matorrales y
arbustos con flores perteneciente al Transecto N° 11 (Figura 85).
Por otro lado, el análisis de Regresión Múltiple nos indica que el hábitat parecería no
influir en su distribución, puesto que se obtuvo un p valor de 0.5 no significativo (p
significativo < 0.05).
Figura 84: Distribución espacial de la Abundancia Relativa (IARE Promedio) de
Amazilia amazilia en el campus de la UNALM en el periodo evaluado (Noviembre
2011 – Octubre 2012)
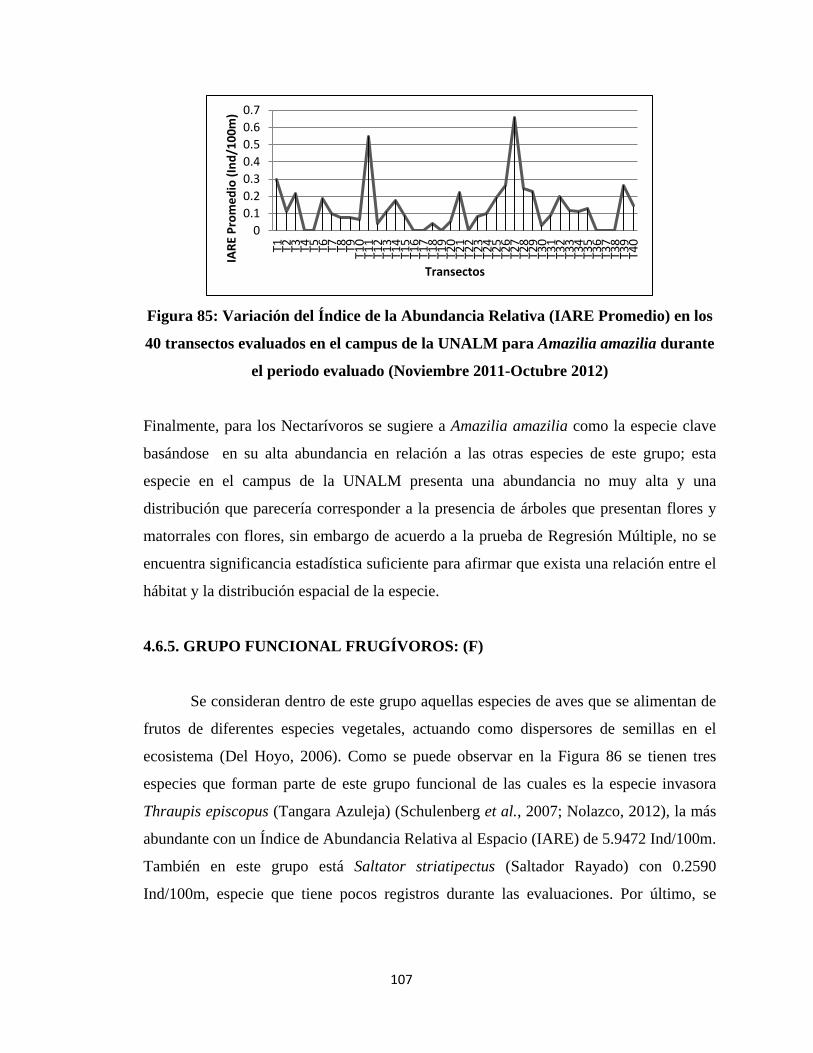
107
Figura 85: Variación del Índice de la Abundancia Relativa (IARE Promedio) en los
40 transectos evaluados en el campus de la UNALM para Amazilia amazilia durante
el periodo evaluado (Noviembre 2011-Octubre 2012)
Finalmente, para los Nectarívoros se sugiere a Amazilia amazilia como la especie clave
basándose en su alta abundancia en relación a las otras especies de este grupo; esta
especie en el campus de la UNALM presenta una abundancia no muy alta y una
distribución que parecería corresponder a la presencia de árboles que presentan flores y
matorrales con flores, sin embargo de acuerdo a la prueba de Regresión Múltiple, no se
encuentra significancia estadística suficiente para afirmar que exista una relación entre el
hábitat y la distribución espacial de la especie.
4.6.5. GRUPO FUNCIONAL FRUGÍVOROS: (F)
Se consideran dentro de este grupo aquellas especies de aves que se alimentan de
frutos de diferentes especies vegetales, actuando como dispersores de semillas en el
ecosistema (Del Hoyo, 2006). Como se puede observar en la Figura 86 se tienen tres
especies que forman parte de este grupo funcional de las cuales es la especie invasora
Thraupis episcopus (Tangara Azuleja) (Schulenberg et al., 2007; Nolazco, 2012), la más
abundante con un Índice de Abundancia Relativa al Espacio (IARE) de 5.9472 Ind/100m.
También en este grupo está Saltator striatipectus (Saltador Rayado) con 0.2590
Ind/100m, especie que tiene pocos registros durante las evaluaciones. Por último, se
00.10.20.30.40.50.60.7
T1 T2 T3 T4 T5 T6 T7 T8 T9 T10
T11
T12
T13
T14
T15
T16
T17
T18
T19
T20
T21
T22
T23
T24
T25
T26
T27
T28
T29
T30
T31
T32
T33
T34
T35
T36
T37
T38
T39
T40
IARE
Pro
med
io (I
nd/1
00m
)
Transectos
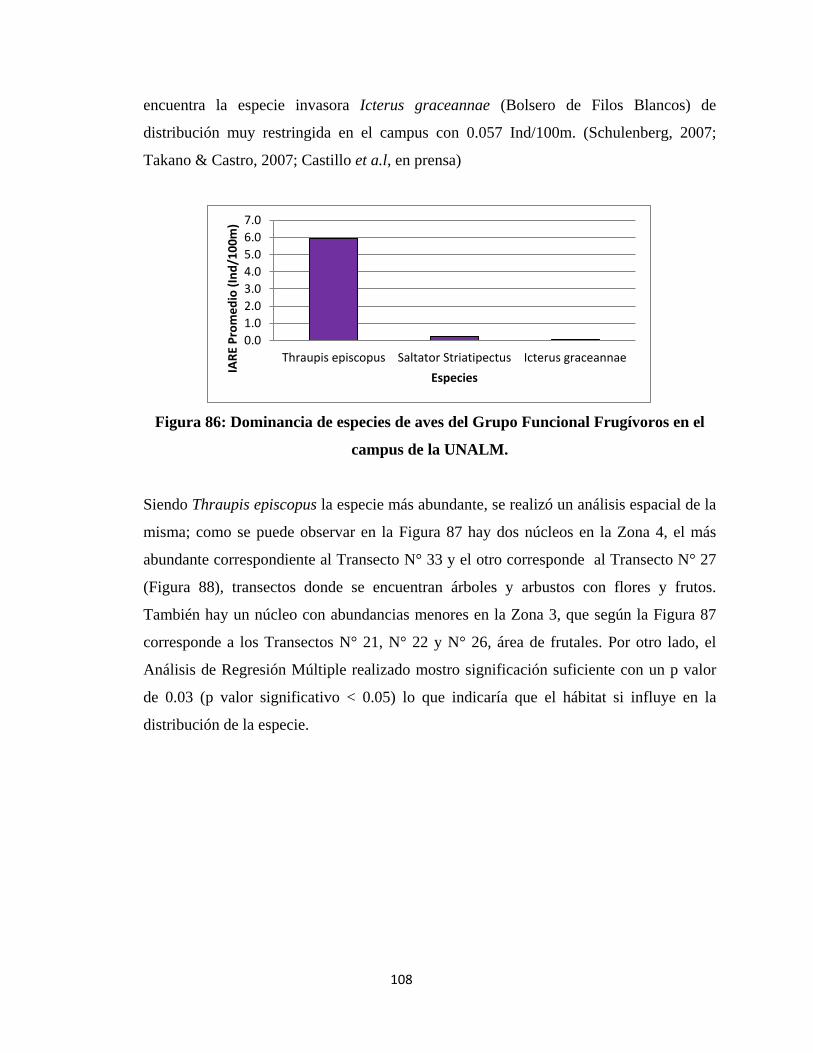
108
encuentra la especie invasora Icterus graceannae (Bolsero de Filos Blancos) de
distribución muy restringida en el campus con 0.057 Ind/100m. (Schulenberg, 2007;
Takano & Castro, 2007; Castillo et a.l, en prensa)
Figura 86: Dominancia de especies de aves del Grupo Funcional Frugívoros en el
campus de la UNALM.
Siendo Thraupis episcopus la especie más abundante, se realizó un análisis espacial de la
misma; como se puede observar en la Figura 87 hay dos núcleos en la Zona 4, el más
abundante correspondiente al Transecto N° 33 y el otro corresponde al Transecto N° 27
(Figura 88), transectos donde se encuentran árboles y arbustos con flores y frutos.
También hay un núcleo con abundancias menores en la Zona 3, que según la Figura 87
corresponde a los Transectos N° 21, N° 22 y N° 26, área de frutales. Por otro lado, el
Análisis de Regresión Múltiple realizado mostro significación suficiente con un p valor
de 0.03 (p valor significativo < 0.05) lo que indicaría que el hábitat si influye en la
distribución de la especie.
0.01.02.03.04.05.06.07.0
Thraupis episcopus Saltator Striatipectus Icterus graceannae
IARE
Pro
med
io (I
nd/1
00m
)
Especies
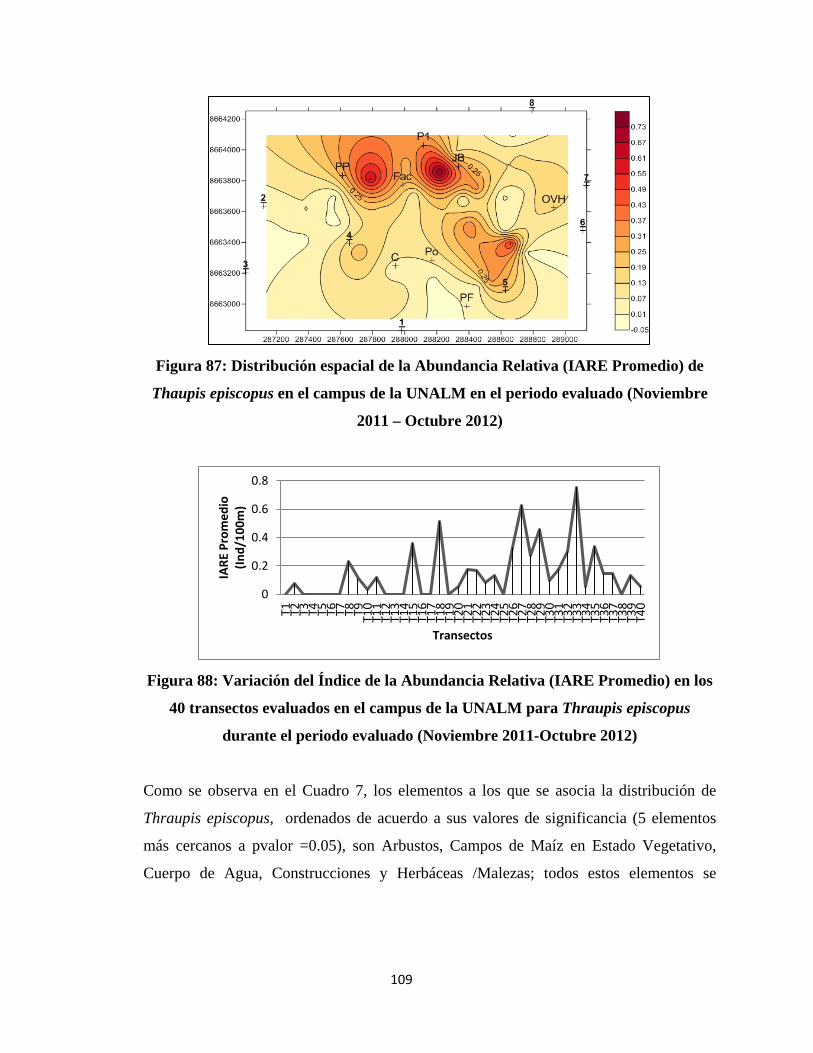
109
Figura 87: Distribución espacial de la Abundancia Relativa (IARE Promedio) de
Thaupis episcopus en el campus de la UNALM en el periodo evaluado (Noviembre
2011 – Octubre 2012)
Figura 88: Variación del Índice de la Abundancia Relativa (IARE Promedio) en los
40 transectos evaluados en el campus de la UNALM para Thraupis episcopus
durante el periodo evaluado (Noviembre 2011-Octubre 2012)
Como se observa en el Cuadro 7, los elementos a los que se asocia la distribución de
Thraupis episcopus, ordenados de acuerdo a sus valores de significancia (5 elementos
más cercanos a pvalor =0.05), son Arbustos, Campos de Maíz en Estado Vegetativo,
Cuerpo de Agua, Construcciones y Herbáceas /Malezas; todos estos elementos se
0
0.2
0.4
0.6
0.8
T1 T2 T3 T4 T5 T6 T7 T8 T9 T10
T11
T12
T13
T14
T15
T16
T17
T18
T19
T20
T21
T22
T23
T24
T25
T26
T27
T28
T29
T30
T31
T32
T33
T34
T35
T36
T37
T38
T39
T40
IARE
Pro
med
io
(Ind/
100m
)
Transectos

110
encuentran presentes en los transectos mostrados en la Figura 88 donde la especie de
distribuye según sus IARE.
Cuadro 7. Elementos del Hábitat significativos para la distribución de Thraupis
episcopus en el campus de la UNALM según el Análisis de Regresión Múltiple
Elementos del Hábitat pvalor
Arbustos 0.060119
CC_Vegetativo_Maiz 0.072834 Cuerpo_Agua 0.080028 Construcción 0.087811 Herbáceas/maleza 0.12905
Finalmente, se propone a Thraupis episcopus como la especie clave para este grupo
funcional debido a su alta abundancia en relación a las otras especies presentes. Ésta
especie se encuentra asociada a formaciones arbóreas, especialmente por la presencia de
flores y frutos, esto tiene un respaldo estadístico en el Análisis de Regresión Múltiple
realizado, donde sí existe una relación entre los elementos del hábitat y la distribución
espacial de la especie.
4.6.6 GRUPO FUNCIONAL CARROÑEROS: (CARR)
Este grupo está formado por aquellos animales que consumen carroña (Houston,
1994 citado por Del Hoyo, 2006). Para el caso del campus de la UNALM se tiene una
sola especie que forma parte de este grupo funcional, el Coragyps atratus (Gallinazo de
Cabeza Negra) con 19 Ind/100m.
Es comúnmente visto en ciudades, pueblos, pastizales y demás zonas abiertas, así como
riberas de ríos (Schulenberg et al., 2007); en la UNALM si se analiza su distribución
espacial como se observa en la Figura 89, existe un solo núcleo de abundancia alta el
cual corresponde al área de establos y alrededores, como el campo deportivo y área del

111
Programa de Carnes como se aprecia en la Figura 90 (Transectos N° 15, N° 16, N°34 y
N°35), zonas de donde posiblemente encuentre alimento. Sin embargo, el Análisis de
Regresión Múltiple realizado nos indica que el hábitat parecería no influir en su
distribución, puesto que se obtuvo un p valor de 0.8 no significativo (p valor
significativo < 0.05).
Figura 89: Distribución espacial de la Abundancia Relativa (IARE Promedio) de
Coragyps atratus en el campus de la UNALM en el periodo evaluado (Noviembre
2011 – Octubre 2012)
Figura 90: Variación del Índice de la Abundancia Relativa (IARE Promedio) en los
40 transectos evaluados en el campus de la UNALM para Coragyps atratus durante
el periodo evaluado (Noviembre 2011-Octubre 2012)
0
2
4
6
T1 T2 T3 T4 T5 T6 T7 T8 T9 T10
T11
T12
T13
T14
T15
T16
T17
T18
T19
T20
T21
T22
T23
T24
T25
T26
T27
T28
T29
T30
T31
T32
T33
T34
T35
T36
T37
T38
T39
T40
IARE
Pro
med
io
(Ind/
100m
)
Transectos

112
Por último, siendo Coragyps atratus el único representante de este grupo, es por defecto
la especie clave sugerida; esta especie en el campus de la UNALM, estaría asociado a
zonas donde es posible encontrar carroña, su principal alimento (establos y mataderos),
sin embargo no existe evidencia estadística suficiente para respaldar ésta aseveración
según los Resultados del Análisis de Regresión.
4.6.7 GRUPO FUNCIONAL PISCÍVOROS: (P)
Incluye todas las especies de aves que se alimentan específicamente de peces e
invertebrados acuáticos (Del Hoyo, 2006); se las separa de los carnívoros por ser
especialistas para ambientes acuáticos. Para el ecosistema estudiado se tiene registrada
una sola especie poco abundante que consume peces (Santana, 2011): Phalacrocorax
brasilianus (Cormorán Neotropical) con 0.076 4 Ind/100m y la cual parece ser visitante
para el campus de manera ocasional.
Según Schulenberg et al. (2007) es una especie asociada a cuerpos de agua como mares,
ríos y humedales. Para el campus de la UNALM, como se observa en la Figura 91, se
presentan dos núcleos de abundancias muy bajas, uno correspondiente al Transecto N°
10 (Figura 92) donde se ubica el Cuerpo de Agua (Zona 2), también se puede observar
escasa presencia en la Zona 4, específicamente en el Transecto N° 39, el cual
corresponde la Jardín Botánico donde también existe un pequeño Cuerpo de Agua.
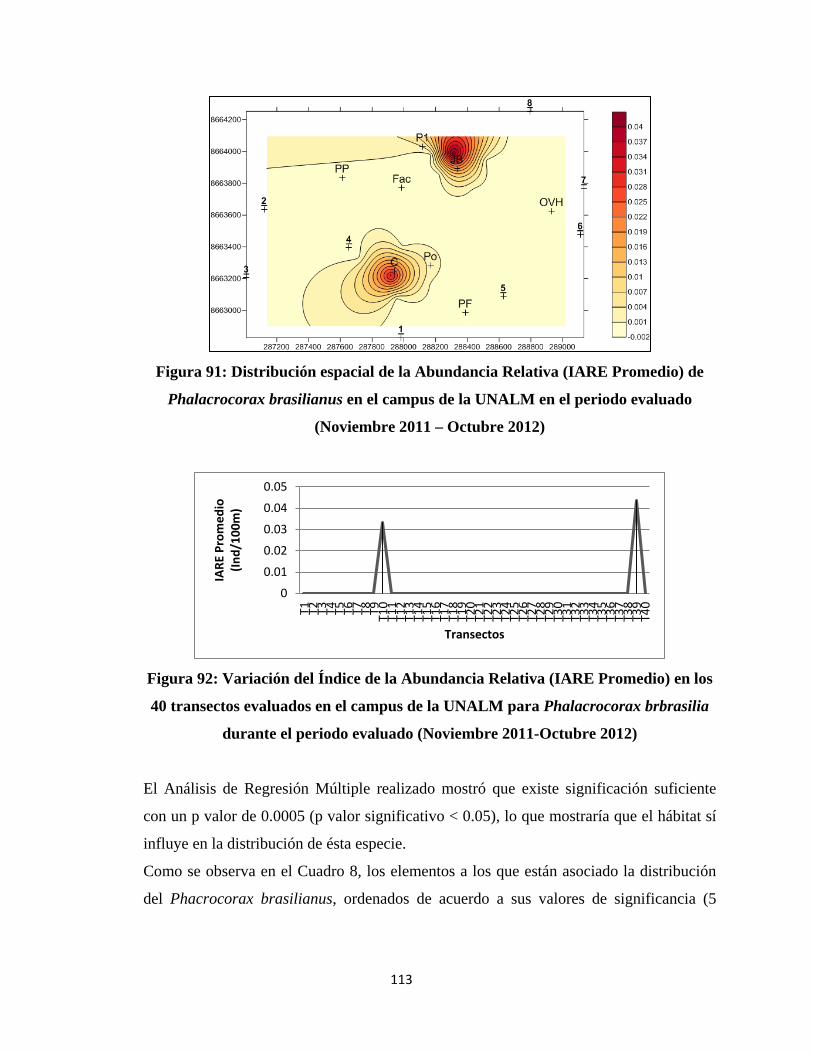
113
Figura 91: Distribución espacial de la Abundancia Relativa (IARE Promedio) de
Phalacrocorax brasilianus en el campus de la UNALM en el periodo evaluado
(Noviembre 2011 – Octubre 2012)
Figura 92: Variación del Índice de la Abundancia Relativa (IARE Promedio) en los
40 transectos evaluados en el campus de la UNALM para Phalacrocorax brbrasilia
durante el periodo evaluado (Noviembre 2011-Octubre 2012)
El Análisis de Regresión Múltiple realizado mostró que existe significación suficiente
con un p valor de 0.0005 (p valor significativo < 0.05), lo que mostraría que el hábitat sí
influye en la distribución de ésta especie.
Como se observa en el Cuadro 8, los elementos a los que están asociado la distribución
del Phacrocorax brasilianus, ordenados de acuerdo a sus valores de significancia (5
0
0.01
0.02
0.03
0.04
0.05
T1 T2 T3 T4 T5 T6 T7 T8 T9 T10
T11
T12
T13
T14
T15
T16
T17
T18
T19
T20
T21
T22
T23
T24
T25
T26
T27
T28
T29
T30
T31
T32
T33
T34
T35
T36
T37
T38
T39
T40
IARE
Pro
med
io
(Ind/
100m
)
Transectos

114
elementos más cercanos a pvalor =0.05), son Arbustos, Cuerpo de Agua, Construcciones,
Matorral y Tierra Descubierta; todos éstos elementos se encuentran presentes en los
transectos mostrados en la figura donde la especie de distribuye según sus IARE,
especialmente Cuerpo de Agua, el cual podría bien ser el elemento que atrae a la especie
al campus.
Cuadro 8. Elementos del Hábitat significativos para la distribución de
Phalacrocorax brasialianus en el campus de la UNALM según el Análisis de
Regresión Múltiple
Elementos del Hábitat p-valor Arbustos 0.0013835
Cuerpo de Agua 0.0014406 Construcción 0.0084055
Matorral 0.036986 Tierra_Descubierta 0.037134
Por ser esta especie un visitante ocasional del campus, según lo observado, no se le
nombra como especie clave, lo que sí es posible afirmar es que existe una relación entre
la distribución espacial de la especie y el hábitat, lo cual está respaldado por la
significancia de la Prueba de Regresión Múltiple realizada.
4.6.8 GRUPO FUNCIONAL GRANÍVOROS – FRUGÍVOROS: (G-F)
Se considera en este grupo mixto a las especies de aves que se alimentan de granos y
a la vez de frutos. Se tienen 5 especies de aves que registran éste comportamiento
alimenticios, siendo el Género Sporophila con las especies Sporophila simplex
(Espiguero Simple) y Sporophila telasco (Espiguero de Garganta Castaña) es el más
abundante en el grupo (30.1281 Ind/100m). Cabe resaltar también a Forpus coelestis
(Periquito Esmeralda) con 13.8813 Ind/100m y Aratinga erytrogenys (Cotorra de Cabeza

115
Roja) con 9.5946 Ind/100m, ambos con abundancias importantes considerando que
ambas son especies invasoras que se han logrado adaptarse (Koepcke, 1964 citado por
Castro & Takano, 2007; Schulenberg et al, 2007) y que suelen ser consideradas plagas de
muchos cultivos, como el maíz (Zeas mays). Con menor abundancia se encuentra
Zonotrichia capensis (Gorrión de Collar Rufo) con 3.5018 Ind/100m y Passer dometicus
(Gorrión Casero) con un IARE de 0.955 Ind/100m, como se observa en la Figura 93.
Figura 93: Dominancia de especies de aves del Grupo Funcional Granívoros -
Frugívoros en el campus de la UNALM.
Según la Figura 94, la distribución de los Sporophila sp, se concentra en la Zona 3,
específicamente en los Transectos N° 23, N° 24 y N° 21 (Figura 95), áreas donde hay
presencia de cultivos de frutales como mandarinas y lúcumas además de maleza y
arbustos. También se puede apreciar presencia de esta especie en otras zonas de cultivo
de la Zona 1 y Zona 2 (Figura 94). Otros estudios (González, 2000b; Takano & Castro,
2007) revelan la preferencia de estas especies por los campos de cultivo.
0.05.0
10.015.020.025.030.035.0
Sporophila sp Forpuscoelestis
Aratingaerytrogenys
Zonotrichiacapensis
Passerdomesticus
IARE
Pro
med
io (I
nd/1
00m
)
Especies
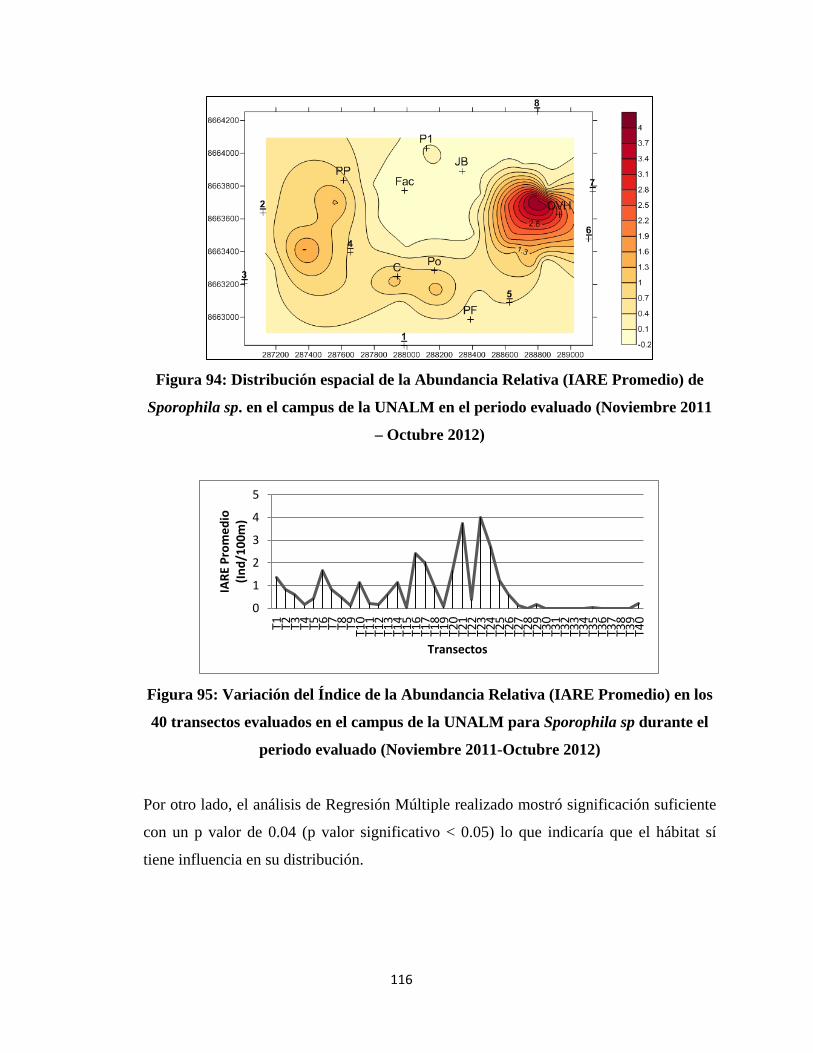
116
Figura 94: Distribución espacial de la Abundancia Relativa (IARE Promedio) de
Sporophila sp. en el campus de la UNALM en el periodo evaluado (Noviembre 2011
– Octubre 2012)
Figura 95: Variación del Índice de la Abundancia Relativa (IARE Promedio) en los
40 transectos evaluados en el campus de la UNALM para Sporophila sp durante el
periodo evaluado (Noviembre 2011-Octubre 2012)
Por otro lado, el análisis de Regresión Múltiple realizado mostró significación suficiente
con un p valor de 0.04 (p valor significativo < 0.05) lo que indicaría que el hábitat sí
tiene influencia en su distribución.
0
1
2
3
4
5
T1 T2 T3 T4 T5 T6 T7 T8 T9 T10
T11
T12
T13
T14
T15
T16
T17
T18
T19
T20
T21
T22
T23
T24
T25
T26
T27
T28
T29
T30
T31
T32
T33
T34
T35
T36
T37
T38
T39
T40
IARE
Pro
med
io
(Ind/
100m
)
Transectos

117
Como se observa en el Cuadro 9, los elementos a los que se asocia su distribución,
ordenados de acuerdo a sus valores de significancia (5 elementos más cercanos al p
valor =0.05), son Campos de Cultivo en Estado Reproductivo de Frutales, Campos de
Cultivo en Estado Vegetativo de Frutales y Borde Vivo; todos estos elementos se
encuentran presentes en los transectos mostrados en la Figura 95, el cual muestra la
distribución del género Sporophila en el campus.
Cuadro 9. Elementos del Hábitat significativos para la distribución de Sporophila
sp. en el campus de la UNALM según el Análisis de Regresión Múltiple
Elementos del Hábitat P valor
CC_Reproductivo_Frutales 0.00016514
CC_Arado 0.052261
CC_Barbecho 0.076414
CC._Vegetativo_Frutales 0.087296
Borde Vivo 0.11981
Adicionalmente, se analizó a Forpus coelestris y Aratinga sp. debido a su asociación a
campos de cultivo y por ser consideradas en algunos casos especies plagas (Takano,
2010).
Por un lado, Forpus coelestis presenta un sólo núcleo importante en la Zona 3 (Figura
96), Transectos N° 23 y N° 24 específicamente según la Figura 97, ésta es un área donde
abundan cultivos de frutales de mandarina y lúcuma, además de arbustos y herbáceas
diversas. Por otra parte, en el resto de la universidad se le puede encontrar, sin embargo
no hay ningún otro punto donde se acumulen como lo hacen en la Zona 3.
El análisis de Regresión Múltiple indica que el hábitat parecería no influir en su
distribución, puesto que se obtuvo un p valor de 0.74 no significativo (p valor
significativo < 0.05).
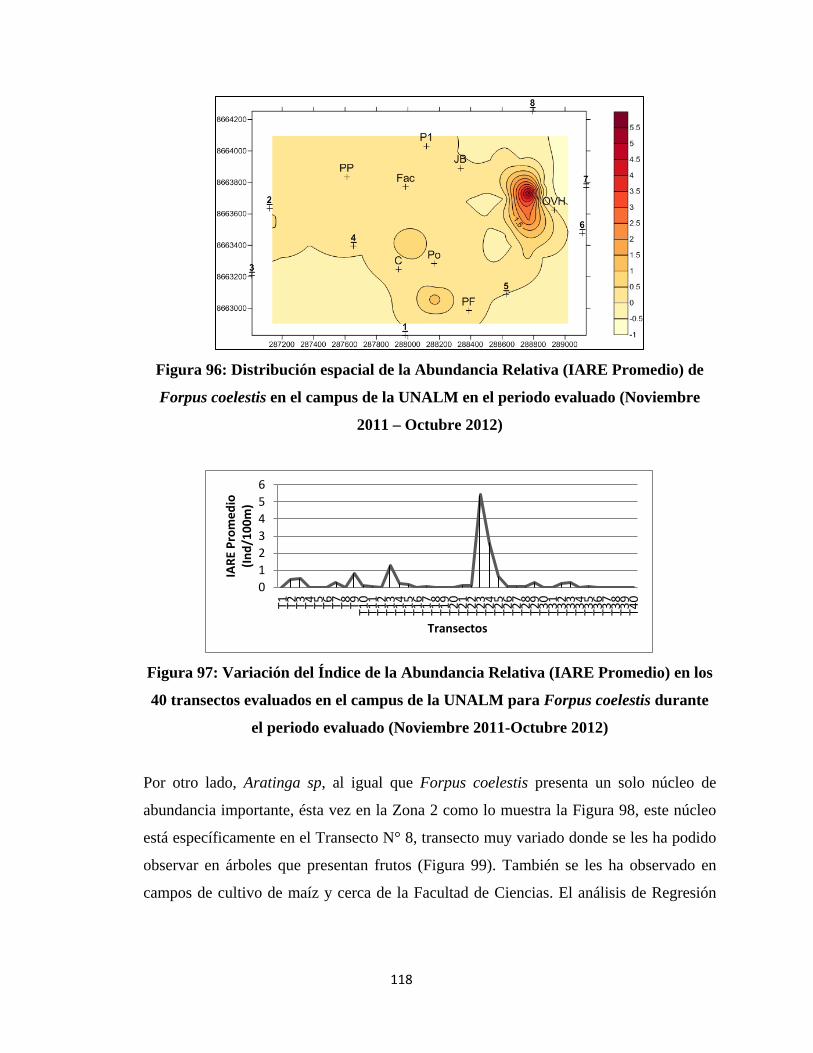
118
Figura 96: Distribución espacial de la Abundancia Relativa (IARE Promedio) de
Forpus coelestis en el campus de la UNALM en el periodo evaluado (Noviembre
2011 – Octubre 2012)
Figura 97: Variación del Índice de la Abundancia Relativa (IARE Promedio) en los
40 transectos evaluados en el campus de la UNALM para Forpus coelestis durante
el periodo evaluado (Noviembre 2011-Octubre 2012)
Por otro lado, Aratinga sp, al igual que Forpus coelestis presenta un solo núcleo de
abundancia importante, ésta vez en la Zona 2 como lo muestra la Figura 98, este núcleo
está específicamente en el Transecto N° 8, transecto muy variado donde se les ha podido
observar en árboles que presentan frutos (Figura 99). También se les ha observado en
campos de cultivo de maíz y cerca de la Facultad de Ciencias. El análisis de Regresión
0123456
T1 T2 T3 T4 T5 T6 T7 T8 T9 T10
T11
T12
T13
T14
T15
T16
T17
T18
T19
T20
T21
T22
T23
T24
T25
T26
T27
T28
T29
T30
T31
T32
T33
T34
T35
T36
T37
T38
T39
T40
IARE
Pro
med
io
(Ind/
100m
)
Transectos
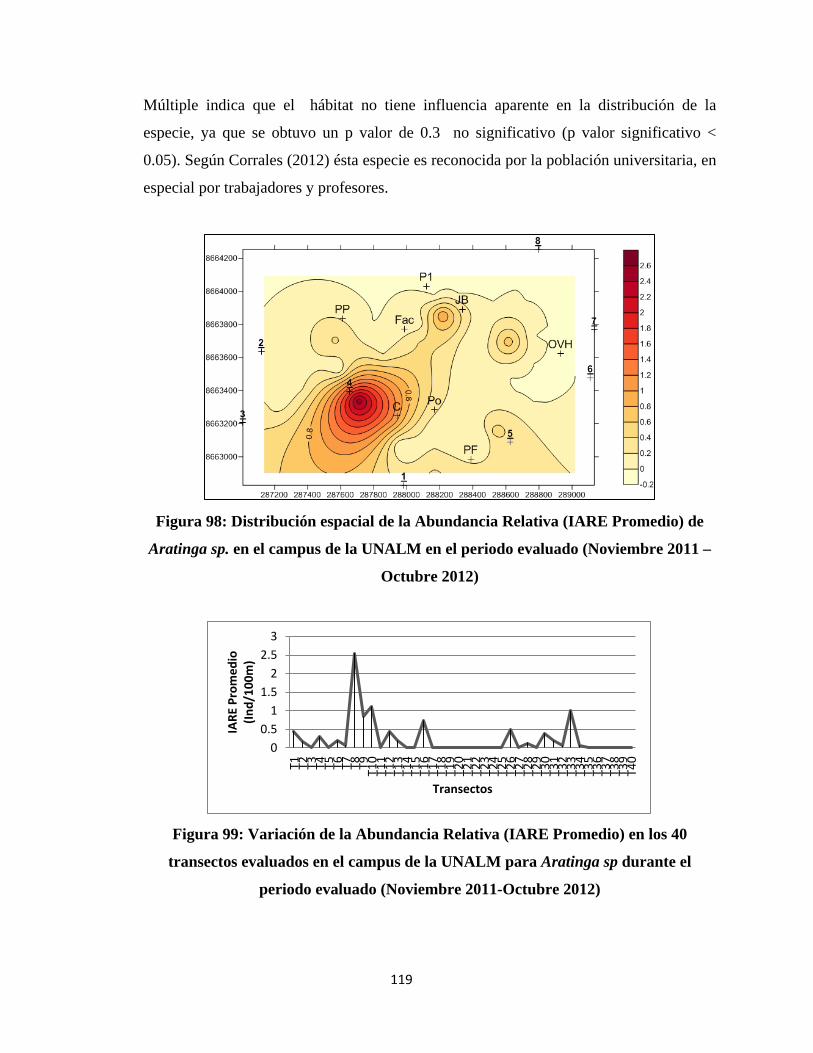
119
Múltiple indica que el hábitat no tiene influencia aparente en la distribución de la
especie, ya que se obtuvo un p valor de 0.3 no significativo (p valor significativo <
0.05). Según Corrales (2012) ésta especie es reconocida por la población universitaria, en
especial por trabajadores y profesores.
Figura 98: Distribución espacial de la Abundancia Relativa (IARE Promedio) de
Aratinga sp. en el campus de la UNALM en el periodo evaluado (Noviembre 2011 –
Octubre 2012)
Figura 99: Variación de la Abundancia Relativa (IARE Promedio) en los 40
transectos evaluados en el campus de la UNALM para Aratinga sp durante el
periodo evaluado (Noviembre 2011-Octubre 2012)
00.5
11.5
22.5
3
T1 T2 T3 T4 T5 T6 T7 T8 T9 T10
T11
T12
T13
T14
T15
T16
T17
T18
T19
T20
T21
T22
T23
T24
T25
T26
T27
T28
T29
T30
T31
T32
T33
T34
T35
T36
T37
T38
T39
T40
IARE
Pro
med
io
(Ind/
100m
)
Transectos

120
Finalmente, debido a su alta abundancia e importante presencia en campos de cultivo se
propone al Género Sporophila como el grupo clave en este gremio. Éste género está
asociado a campos de cultivo así como zonas con presencia de cultivos de frutales, esto
se comprueba con el Análisis de Regresión Múltiple realizado el cual mostró
significancia para la relación entre el hábitat y la distribución espacial de éste género, se
encuentra asociado a campos de cultivo, y en especial a los cultivos de Frutales. Por otro
lado, para Forpus coelestis y Aratinga sp, no existe evidencia estadística suficiente para
demostrar una relación entre el hábitat y la distribución espacial de éstas especies.
4.6.9 GRUPO FUNCIONAL INSECTÍVOROS – OMNÍVOROS: (I-O)
En este grupo se encuentran aquellas especies que principalmente, por su ecología y
según la literatura, son Insectívoros pero que se les ha observado teniendo
comportamientos alimenticios anormales en el campus. Son dos los integrantes: Dives
warsewicsi (Tordo de Matorral) con 36.9534 Ind/100m, su ecología lo sitúa
principalmente en matorrales secos, campos y jardines, pero se ha adaptado muy bien a
ambientes urbanos de Lima donde es una especie muy abundante (Schulenberg et al,
2007; Nolazco, 2012); ha sido observado comiendo desde trozos de papas fritas, pollo
cocido, hot dog, migajas de pan y galletas hasta golosinas artificiales, incluso ha sido
avistado introduciéndose a bolsas llenas de basura y consumiendo menudencia cruda de
pollo. Crotophaga sulcirostris con 13.5705 Ind/100m, el cual según la literatura es un
Insectívoro (Santana, 2011; Nolazco, 2012) pero ha sido avistado consumiendo migajas
de galletas y pan. Se le asocia con campos, pastizales y bordes de bosques, además de ser
comúnmente asociado a campos de cultivo (González, 2000b; Schulenberg et al, 2007).

121
Figura 100: Dominancia de especies de aves del Grupo Funcional Granívoros -
Frugívoros en el campus de la UNALM.
Como se puede apreciar en la Figura 101, presenta un núcleo importante en la Zona 1, de
acuerdo a la Figura 102 este núcleo corresponde a los Transectos N° 1, N° 2, N° 6, N° 7
y N° 8, todos correspondientes a campos de cultivo en su mayoría de maíz. También hay
una núcleo que pretende formarse en la Zona 2, específicamente el área correspondiente
al Transecto N° 12, el cual también corresponde a campos de cultivo de maíz (Figura
101). También se observa presencia en la Zona 4 del campus correspondiente a zonas
urbanas. La prueba de Regresión Múltiple muestra un p valor no significativo de 0.21 (p
valor significativo < 0.05), lo cual indicaría que no existe una relación entre la
distribución del hábitat y la distribución de la especie.
0.0
10.0
20.0
30.0
40.0
Dives warsewicsi Crotophaga sulcirostrisIARE
Pro
med
io (I
nd/1
00m
)
Especies

122
Figura 101: Distribución espacial de la Abundancia Relativa (IARE Promedio) de
Dives warsewicsi en el campus de la UNALM en el periodo evaluado (Noviembre
2011 – Octubre 2012)
Figura 102: Variación de la Abundancia Relativa (IARE Promedio) en los 40
transectos evaluados en el campus de la UNALM para Dives warsewicsi durante el
periodo evaluado (Noviembre 2011-Octubre 2012)
0
1
2
3
4
T1 T2 T3 T4 T5 T6 T7 T8 T9 T10
T11
T12
T13
T14
T15
T16
T17
T18
T19
T20
T21
T22
T23
T24
T25
T26
T27
T28
T29
T30
T31
T32
T33
T34
T35
T36
T37
T38
T39
T40IARE
Pro
med
io (I
nd/1
00m
)
Transectos

123
Para el caso de Crotophaga sulcirostris, en el campus de la UNALM su distribución es
como se muestra en la Figura 103; se pueden observar hasta tres núcleos, el primero en la
Zona 1, correspondiente a los Transectos N° 3 y N° 6 como muestra la Figura 104, en los
cuales existen campos de cultivo principalmente de maíz. También se puede apreciar
otro núcleo en la Zona 2, el cual corresponde al Transecto N° 12, donde también
podemos encontrar campos de cultivo de maíz y el Transecto N° 13 con otros cultivos
variados, así como borde vivo de cañas y basura en el ingreso del transecto. Finalmente,
hay un último núcleo notable en la Zona 3, la cual corresponde a los Transectos N° 15 y
N° 34 los cuales perteneces a los establos. Sin embargo, con un p valor no significativo
de 0.091 (p valor significativo < 0.05), el análisis de Regresión Múltiple indica que el
hábitat no tiene influencia aparente en la distribución de la especie.
Figura 103: Distribución espacial de la Abundancia Relativa (IARE Promedio) de
Crotophaga sulcirostris en el campus de la UNALM en el periodo evaluado
(Noviembre 2011 – Octubre 2012)

124
Figura 104: Variación de la Abundancia Relativa (IARE Promedio) en los 40
transectos evaluados en el campus de la UNALM para Crotophaga sulcirostris
durante el periodo evaluado (Noviembre 2011-Octubre 2012)
La especie clave sugerida es Dives warsewicsi por ser la más abundante de las dos. A
pesar de que el mapa de distribución espacial elaborado en SURFER muestra una
relación de la distribución de la especie con campos de cultivo como lo señala la
literatura revisada, no es posible respaldarlo estadísticamente puesto que el Análisis de
Regresión Múltiple no es significativo.
4.6.10 GRUPO FUNCIONAL GRANÍVOROS – OMNÍVOROS: (G-O)
Aquí se encuentran las especies que son tradicionalmente Granívoras pero que
también por efectos de la urbanización se han adaptado y presentan nuevas dietas. Se
tienen tres especies para este grupo funcional (Figura 102). La más abundante es Zenaida
meloda (Tortolita Melódica) con 102.1554 Ind/100m; a este especie se le ha visto
comiendo también trozos de pollo cocido, papas fritas, golosinas, galletas, pan y muchas
veces ha sido avistada rodeando bolsas y cilindros de basura. Finalmente, se encuentra el
Sicalis flaveola (Chiringüe Azafranado) con 6.3281 Ind/100m el cual también ha sido
observado consumiendo migajas de golosinas como galletas, papas fritas marca Lays,
entre otros.
0
0.5
1
1.5
T1 T2 T3 T4 T5 T6 T7 T8 T9 T10
T11
T12
T13
T14
T15
T16
T17
T18
T19
T20
T21
T22
T23
T24
T25
T26
T27
T28
T29
T30
T31
T32
T33
T34
T35
T36
T37
T38
T39
T40
IARE
Pro
med
io
(Ind/
100m
)
Transectos

125
Figura 105: Dominancia de especies de aves del Grupo Funcional Granívoros -
Omnívoros en el campus de la UNALM.
Zenaida meloda es la especie más abundante de este grupo funcional, sus hábitats suelen
ser campos, jardines y bosques abiertos, además de ser muy abundante en la ciudad de
Lima (Schulenberg et al, 2007; Nolazco, 2012).
Si se observa la Figura 106, el mapa de distribución espacial de la abundancia para la
UNALM, se puede apreciar un núcleo importante en la Zona 3 explicada en parte por el
Transecto N° 34 el cual corresponde al establo (Figura 107). También se observa
presencia importante en la Zona 1 y la Zona 2, mientras que en la Zona 4 urbana es
abundante pero con menores abundancias que en las otras tres zonas. Ésta especie ha sido
reconocida como potencial problema para campos de cultivo por su consumo de granos
(González, 2000b), incluso en los últimos años se está considerando como un problema
dentro de la ciudad de Lima.
0.020.040.060.080.0
100.0120.0
Zenaida meloda Sicalis flaveolaIA
RE P
rom
edio
(In
d/10
0m)
Especies

126
Figura 106: Distribución espacial de la Abundancia Relativa (IARE Promedio) de
Zenaida meloda en el campus de la UNALM en el periodo evaluado (Noviembre
2011 – Octubre 2012)
Figura 107: Variación de la Abundancia Relativa (IARE Promedio) en los 40
transectos evaluados en el campus de la UNALM para Zenaida meloda durante el
periodo evaluado (Noviembre 2011-Octubre 2012)
La prueba de Regresión Múltiple muestra un p valor no significativo de 0.14 (p valor
significativo < 0.05), lo cual indicaría que no existe una relación entre la distribución del
hábitat y la distribución de la especie, lo cual podría respaldar lo anteriormente expuesto
de que la especies es generalista, además de ser la mejor distribuida en el campus.
0
2
4
6
8
10
T1 T2 T3 T4 T5 T6 T7 T8 T9 T10
T11
T12
T13
T14
T15
T16
T17
T18
T19
T20
T21
T22
T23
T24
T25
T26
T27
T28
T29
T30
T31
T32
T33
T34
T35
T36
T37
T38
T39
T40
IARE
Pro
med
io
(Ind/
100m
)
Transectos

127
Finalmente, siendo la especie más abundante para este grupo funcional, se sugiere a
Zenaida meloda como la especie clave de los Granívoros – Omnívoros, además de la
más abundante es la mejor distribuida en el campus, por lo que no se encuentra relación
especifica entre su distribución espacial y el hábitat, ya que no discrimina por su
naturaleza generalista.
4.6.11 GRUPO FUNCIONAL OMNÍVOROS: (O)
Referido a las especies cuya dieta es variada. Tenemos como único integrante al
Minus longicaudatus (18.2039 Ind/100m). Esta especie se caracteriza por consumir de
todo tipo de alimentos como frutos, insectos (Nolazco, 2012) los cuales caza
espantándolos con su cola hasta todo tipo de golosinas, brotes de plantas, y en el campus
de la UNALM se le ha registrado consumiendo papas fritas y migajas de pan; también ha
sido vista rodeando cilindros de basura en el campus.
Según Schulenberg et al (2007) el Mimus longicaudatus (Calandria de Cola Larga) se
distribuye en matorrales desérticos, zonas de bosques abiertos, áreas agrícolas y jardines.
En el campus de la UNALM, como se observa en el mapa de distribución espacial del
IARE (Figura 108) se puede observar que está muy presente en las áreas urbanas del
campus (Zona 4) con dos núcleos importantes en esta zona, uno correspondiente al
Transecto N° 31, el cual se encuentra en la zona urbana de la universidad rodeada de
jardines, arbustos, árboles y salones; otro núcleo también en la zona urbana de la
universidad correspondiente a los Transectos N° 27, N° 28 y N° 29, también rodeado de
jardines, construcciones, árboles, arbustos, entre otros (Figura 109). El análisis de
Regresión Múltiple realizado mostró significación suficiente con un p valor de 0.02 (p
valor significativo < 0.05) lo que indicaría que el hábitat sí presenta influencia en la
distribución del Mimus longicaudatus.

128
Figura 108: Distribución espacial la Abundancia Relativa (IARE Promedio) de
Minus longicaudatus en el campus de la UNALM en el periodo evaluado
(Noviembre 2011 – Octubre 2012)
Figura 109: Variación de la Abundancia Relativa (IARE Promedio) en los 40
transectos evaluados en el campus de la UNALM para Mimus longicaudatus
durante el periodo evaluado (Noviembre 2011-Octubre 2012)
El Cuadro 10 muestra los elementos del hábitat a los que se asocia la distribución del
Mimus longicaudatus de acuerdo a los valores de significancia mostrados (cercanos a p
valor =0.05), éstos son Construcciones, Establo, Herbáceas/Malezas, Cuerpo de Agua y
0
0.5
1
1.5
2
2.5
3
T1 T2 T3 T4 T5 T6 T7 T8 T9 T10
T11
T12
T13
T14
T15
T16
T17
T18
T19
T20
T21
T22
T23
T24
T25
T26
T27
T28
T29
T30
T31
T32
T33
T34
T35
T36
T37
T38
T39
T40IA
RE P
rom
edio
(Ind
/100
m)
Transectos

129
Tierra Descubierta. Todos estos elementos se encuentran presentes en los transectos
mostrados en la Figura 109, el cual muestra la distribución de la especie. Como se
observa en la figura, la especie tiende a distribuirse en zonas urbanas del campus esto se
comprueba con el Análisis de Regresión Múltiple, siendo Construcciones (Salones,
Facultades y Laboratorios) el elementos que mayor está asociado a un alto IARE y por lo
tanto presencia de la especie en el lugar.
Cuadro 10. Elementos del Hábitat significativos para la distribución de Mimus
longicaudatus en el campus de la UNALM según el Análisis de Regresión Múltiple
Elementos del Hábitat P - valor Construcción 0.01264
Establo 0.085429 Herbáceas/maleza 0.41831
Cuerpo Agua 0.44836 Tierra Descubierta 0.45047
Finalmente, ya que Mimus longicaudatus es el único representante de este grupo, es la
especie clave sugerida para los Omnívoros. Según el mapa de distribución a especie se
encuentra a asociada a ambientes urbanos del campus (Zona 4) lo cual es posible
confirmar con el Análisis de Regresión Múltiple significativo, es decir si existe una
dependencia de la distribución espacial de la especie a la distribución de los elementos
del hábitat.
4.6.12 GRUPO FUNCIONAL INSECTÍVOROS – CARNÍVOROS: (I-C)
En este grupo se encuentra las especies que consumen insectos principalmente
pero también complementan su dieta con pequeños vertebrados como reptiles y
mamíferos pequeños de manera oportunista. Solo hay un representante del grupo en el
campus de la UNALM, con un Índice de Abundancia Relativa al Espacio de 0.9583
Ind/100m es la Athene cunicularia (Lechuza Terrestre) con una distribución restringida

130
debido a su ecología, ya que necesita espacios en la tierra para cavar hoyos donde anidar
(Schulenberg et al., 2007; Takano &Castro, 2007; Castillo et al, en prensa). A pesar de
tener una baja abundancia, esta especie es residente del campus por lo que se realizó un
análisis espacial de la distribución de la abundancia, resultando una distribución agregada
ya conocida por trabajos anteriores (Takano, 2007; Castillo et al., en prensa).
Sus hábitats regulares son lugares abiertos como campos y pastizales (Schulenberg et al,
2007). En la Figura 110 se observa un solo núcleo de presencia ubicado en la Zona 4 que
según la Figura 111 correspondería principalmente a los Transectos N° 30 y N° 31 donde
existen jardines y tierra descubierta; en ésta área se han avistados hasta dos nidos de
Athene cunicularia. La prueba de Regresión Múltiple muestra un p valor no significativo
de 0.6 (p valor significativo < 0.05), lo cual indicaría que no existe una relación entre la
distribución del hábitat y la distribución de la especie.
Figura 110: Distribución espacial la Abundancia Relativa (IARE Promedio) de
Athene cunicularia en el campus de la UNALM en el periodo evaluado (Noviembre
2011 – Octubre 2012)
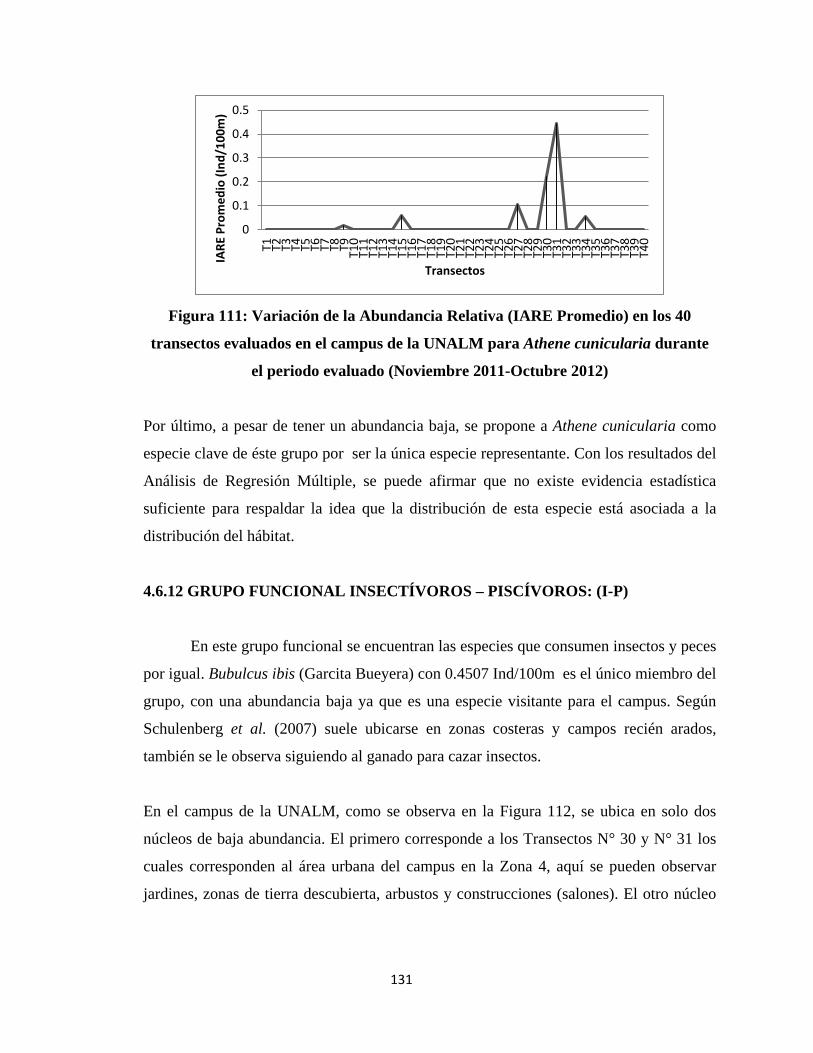
131
Figura 111: Variación de la Abundancia Relativa (IARE Promedio) en los 40
transectos evaluados en el campus de la UNALM para Athene cunicularia durante
el periodo evaluado (Noviembre 2011-Octubre 2012)
Por último, a pesar de tener un abundancia baja, se propone a Athene cunicularia como
especie clave de éste grupo por ser la única especie representante. Con los resultados del
Análisis de Regresión Múltiple, se puede afirmar que no existe evidencia estadística
suficiente para respaldar la idea que la distribución de esta especie está asociada a la
distribución del hábitat.
4.6.12 GRUPO FUNCIONAL INSECTÍVOROS – PISCÍVOROS: (I-P)
En este grupo funcional se encuentran las especies que consumen insectos y peces
por igual. Bubulcus ibis (Garcita Bueyera) con 0.4507 Ind/100m es el único miembro del
grupo, con una abundancia baja ya que es una especie visitante para el campus. Según
Schulenberg et al. (2007) suele ubicarse en zonas costeras y campos recién arados,
también se le observa siguiendo al ganado para cazar insectos.
En el campus de la UNALM, como se observa en la Figura 112, se ubica en solo dos
núcleos de baja abundancia. El primero corresponde a los Transectos N° 30 y N° 31 los
cuales corresponden al área urbana del campus en la Zona 4, aquí se pueden observar
jardines, zonas de tierra descubierta, arbustos y construcciones (salones). El otro núcleo
0
0.1
0.2
0.3
0.4
0.5
T1 T2 T3 T4 T5 T6 T7 T8 T9 T10
T11
T12
T13
T14
T15
T16
T17
T18
T19
T20
T21
T22
T23
T24
T25
T26
T27
T28
T29
T30
T31
T32
T33
T34
T35
T36
T37
T38
T39
T40
IARE
Pro
med
io (I
nd/1
00m
)
Transectos

132
corresponde al Transecto N° 35, donde se encuentra el campo de futbol, cercano a los
establos (Figura 113).
Figura 112: Distribución espacial la Abundancia Relativa (IARE Promedio) de
Bubulcus ibis en el campus de la UNALM en el periodo evaluado (Noviembre 2011
– Octubre 2012)
Figura 113: Variación de la Abundancia Relativa (IARE Promedio) en los 40
transectos evaluados en el campus de la UNALM para Bubulcus ibis durante el
periodo evaluado (Noviembre 2011-Octubre 2012)
0
0.05
0.1
0.15
0.2
0.25
T1 T2 T3 T4 T5 T6 T7 T8 T9 T10
T11
T12
T13
T14
T15
T16
T17
T18
T19
T20
T21
T22
T23
T24
T25
T26
T27
T28
T29
T30
T31
T32
T33
T34
T35
T36
T37
T38
T39
T40
IARE
Pro
med
io (I
nd/1
00m
)
Transectos

133
El Análisis de Regresión Múltiple realizado mostró significación suficiente con un p
valor de 0.048 (p valor significativo < 0.05) lo que indicaría que el hábitat sí se relaciona
con la distribución espacial de Bubulcus ibis.
El Cuadro 11 presenta los elementos del hábitat a los que se asocia la distribución del
Bubulcus ibis ordenados de acuerdo a los valores de significancia mostrados (los 5 más
cercanos a p valor =0.05), éstos son Herbáceas/Malezas, Tierra Descubierta,
Construcciones, Establo y Campos de Cultivos Variados en Estado Vegetativo. Todos
estos elementos se encuentran presentes en los transectos mostrados en la figura.
Cuadro 11. Elementos del Hábitat significativos para la distribución de Bubulcus
ibis en el campus de la UNALM según el Análisis de Regresión Múltiple
Elementos del Hábitat P - valor Herbáceas/maleza 0.0012976 Tierra Descubierta 0.008593 Construcciones 0.013427 Establo 0.013899 CC_Vegetativo_Otros 0.050893
Finalmente, no se elige a Bubulcus ibis como especie clave de este grupo funcional, por
ser una especie avistada muy ocasionalmente, lo que señalaría que es una especie
visitante para el campus. Según el mapa de distribución, se ubica en áreas cercanas a
establos, tierra descubierta y construcciones, esto se comprueba con los resultados de la
Prueba de Regresión Múltiple, lo que indica que sí existe una relación entre la
distribución de la especie y la distribución de los elementos del hábitat para Bubulcus
ibis.

134
4.6.13 GRUPO FUNCIONAL NECTARÍVORO – INSECTÍVORO: (N-I)
Finalmente, este grupo encierra a especies que principalmente consumen néctar y
algunos insectos de manera oportunista. El único miembro de este grupo funcional es la
especie invasora pero abundante en la ciudad, Coereba flaveola (Mielero Común)
(Nolazco, 2012) con un IARE de 4.2699 Ind/100m. Sus hábitats naturales son bosques y
matorrales secos, bordes de bosques y jardines (Schulenberg et al., 2007).
Para el campus de la UNALM, según la Figura 114 correspondiente al análisis espacial
de la abundancia se observan varios núcleos a pesar de que la especie presenta una baja
abundancia promedio. El núcleo mayor corresponde a la Zona 4, específicamente a los
Transectos N° 27, N° 28, N° 29 y N° 30 (Figura 115) transectos con construcciones,
jardines, así como especies arbóreas y arbustivas con flores a ambos lados. También se
observa otro núcleo pequeño en la Zona 2 (Transecto N° 11) correspondiente a una zona
de matorral con arbustos, flores y maleza.
Figura 114: Distribución espacial la Abundancia Relativa (IARE Promedio) de
Coereba flaveola en el campus de la UNALM en el periodo evaluado (Noviembre
2011 – Octubre 2012)
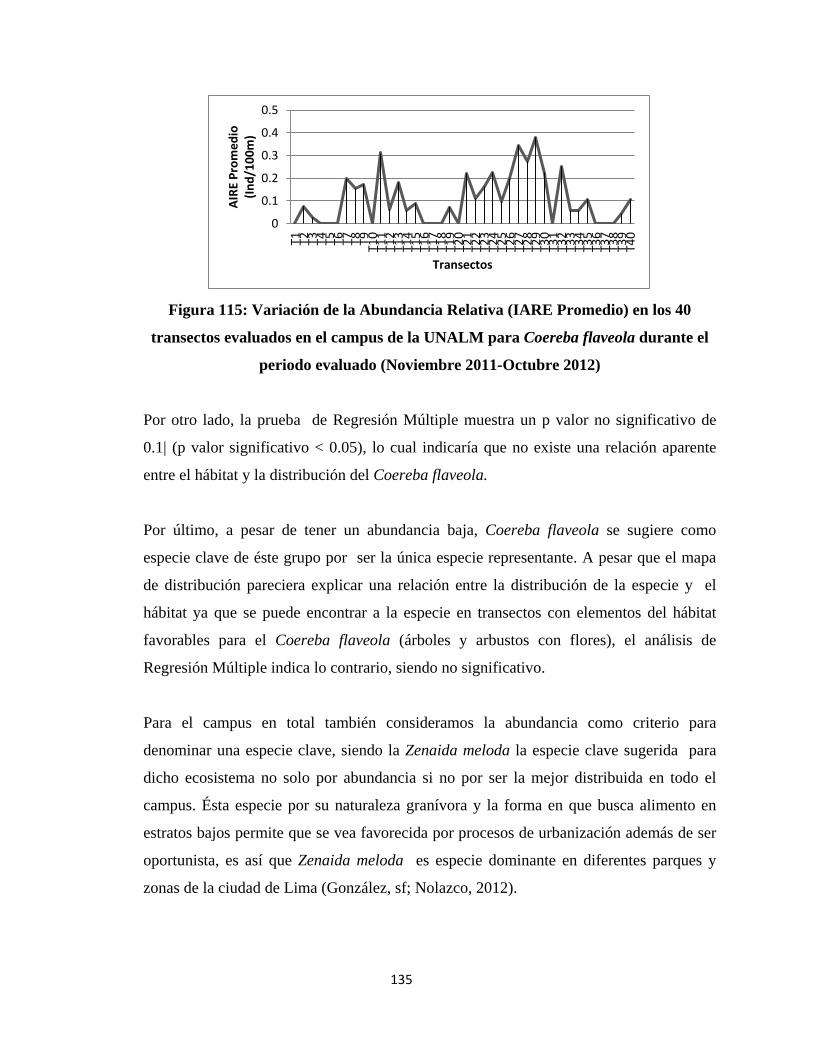
135
Figura 115: Variación de la Abundancia Relativa (IARE Promedio) en los 40
transectos evaluados en el campus de la UNALM para Coereba flaveola durante el
periodo evaluado (Noviembre 2011-Octubre 2012)
Por otro lado, la prueba de Regresión Múltiple muestra un p valor no significativo de
0.1| (p valor significativo < 0.05), lo cual indicaría que no existe una relación aparente
entre el hábitat y la distribución del Coereba flaveola.
Por último, a pesar de tener un abundancia baja, Coereba flaveola se sugiere como
especie clave de éste grupo por ser la única especie representante. A pesar que el mapa
de distribución pareciera explicar una relación entre la distribución de la especie y el
hábitat ya que se puede encontrar a la especie en transectos con elementos del hábitat
favorables para el Coereba flaveola (árboles y arbustos con flores), el análisis de
Regresión Múltiple indica lo contrario, siendo no significativo.
Para el campus en total también consideramos la abundancia como criterio para
denominar una especie clave, siendo la Zenaida meloda la especie clave sugerida para
dicho ecosistema no solo por abundancia si no por ser la mejor distribuida en todo el
campus. Ésta especie por su naturaleza granívora y la forma en que busca alimento en
estratos bajos permite que se vea favorecida por procesos de urbanización además de ser
oportunista, es así que Zenaida meloda es especie dominante en diferentes parques y
zonas de la ciudad de Lima (González, sf; Nolazco, 2012).
0
0.1
0.2
0.3
0.4
0.5
T1 T2 T3 T4 T5 T6 T7 T8 T9 T10
T11
T12
T13
T14
T15
T16
T17
T18
T19
T20
T21
T22
T23
T24
T25
T26
T27
T28
T29
T30
T31
T32
T33
T34
T35
T36
T37
T38
T39
T40
AIRE
Pro
med
io
(Ind/
100m
) Transectos

136
V. CONCLUSIONES
1. En las evaluaciones realizadas entre Noviembre del 2011 y Octubre del 2012 se
registró un total de 44 especies de aves para el Campus de la Universidad Nacional
Agraria La Molina, pertenecientes a 22 Familias y 13 Órdenes, además de una
especie aun no clasificada. Desde el primer estudio realizado en 1992 a la fecha, la
comunidad de aves no ha variado drásticamente en su riqueza pero si en su
composición, lo que llevaría a pensar que ha existido una forma de recambio de
especies. Cabe resaltar que no es posible comparar la diversidad ni la abundancia de
las aves con los estudios realizados anteriormente en el Campus de la UNALM,
debido a la naturaleza y objetivos de dichas investigaciones.
2. Al parecer no existe un efecto estacional marcado en la dinámica de los parámetros
macroscópicos, como los son la abundancia relativa y la diversidad de especies, sin
embargo a nivel de especies si podría haber una influencia del efecto estacional en
sus abundancias.
3. Tanto la diversidad como la abundancia no parecen diferenciarse grandemente entre
mes y mes, sin embargo su distribución en el campus sí cambia. Para el caso de la
abundancia los picos altos en ciertos meses se deben a la abundancia de ciertas
especies en ciertas zonas, como la presencia de Columbina cruziana (Tortolita
peruana) en los establos. Mientras que para la diversidad, las diferencias son menores
posiblemente debido las limitaciones del índice de Shannon-Weaver para apreciar
diferencias estructurales, sin embargo se pueden ubicar zonas de alta diversidad
asociadas posiblemente a hábitats variados y zonas urbanas del campus.
4. El mes de Junio se señala como un mes de altísima abundancia relativa debido a que
la abundancia está dominada por la presencia de Columbina cruziana (Tortolita
peruana) muy concentrada en la zona de establos, pero también de alta diversidad en
relación al resto de meses, lo cual lo hace un mes muy favorable para la avifauna.

137
5. El análisis espacio-temporal indica que la comunidad de aves tiende al refugio
durante los meses desfavorables de Mayo y Julio, este refugio se produce en los sitios
favorables que son los Transectos N° 9 y N° 13, los cuales son los más diversos tanto
en avifauna cómo en elementos del hábitat.
6. En cuanto al hábitat, el campus de la UNALM presenta zonas urbanas y agrícolas,
siendo la agrícola aún dominante. Aquellas zonas muy complejas en hábitat son los
más diversos en aves, posiblemente por ser transiciones de hábitats o eco tonos entre
parches además de ser espacios poco intervenidos por el hombre. Otros transectos en
cambio, muy homogéneos presentan baja diversidad de aves. Esta aparente relación
entre la diversidad de las aves y la complejidad del hábitat no ha podido se
demostrada estadísticamente en el presente estudio. Este hecho podría cambiar a
nivel de especies.
7. Se encontraron 14 grupos funcionales basados la dieta de las especies, dichos grupos
encontrados en el estudio son una expresión de la plasticidad trófica de muchas
especies, ya que si bien es cierto, algunas especies mantienen su dieta regular, otras
son más flexibles y oportunistas con el alimento que encuentran en el campus.
También, se observa la asociación de ciertas especies claves a hábitats específicos
como en el caso de Bubulcus ibis, Mimus longicaudatus Sporophila sp.,
Phalacrocorax brasilianus y Thraupis episcopus; mientras que la mayoría de
especies claves parecerían ser más plásticas en cuanto a su distribución no asociada a
hábitats específicos.
8. Siendo Zenaida meloda la especie más abundante y a la vez la mejor distribuida en el
campus, se considera como especie clave para todo el ecosistema campus de la
UNALM. Por otro lado, ninguna especie clave sugerida se encuentra en ningún
estado de conservación, lo cual replantea las prioridades y criterios en tema de
conservación de especies.

138
9. Es posible afirmar que tanto la Diversidad como la Abundancia Relativa son útiles
para la descripción del estado del ecosistema campus de la UNALM y por lo tanto
indicadores adecuados del estado de dicho ecosistema. Esto se basa en el hecho que
tanto la Diversidad como la Abundancia Relativa varían espacial y temporalmente,
señalando así meses favorables donde las especies tienden a refugiarse, y sitios
desfavorables que podrían llegar a desaparecer; además de tener especies asociadas a
ciertos elementos o características del hábitat. Todo esto en conjunto brinda una
mejor visión de la calidad del ecosistema descrito a partir de su componente avifauna.
Sin embargo, la diversidad presenta más limitantes que la abundancia relativa,
posiblemente por la naturaleza totalitaria del parámetro.

139
VI. RECOMENDACIONES
1. Se recomienda continuar con los censos mensuales en el campus de la UNALM
como una manera de seguir obteniendo data de Abundancia Relativa y Diversidad, lo
cual permitirá realizar estudios comparativos a lo largo de los años.
2. Se debe tener especial atención a las especies clave que el presente estudio sugiere,
así mismo continuar con su monitoreo, realizar estudios de su distribución temporal y
espacial, así como ésta última se relaciona con la distribución del hábitat. Además de
controlar su abundancia y los recursos que consumen. Por otro lado, también sería
conveniente realizar estudios sobre su ecología reproductiva.
3. En base a los resultados del presente estudio, se sugiere iniciar estudios de mayor
precisión sobre la relación aves – elementos del hábitat en el campus de manera más
cuantificable que la realizada en el presente estudio.
4. En ambientes urbanos la dieta de muchas aves se ve alterada debido a la oferta
alimenticia de la ciudad, es importante registrar estos nuevos comportamientos, por lo
que se recomienda realizar seguimientos específicos a dichas especies así como tener
registros visuales de dichos comportamientos.
5. Los establos presentes en el campus de la UNALM se presentan como zonas de alta
abundancia de ciertas especies de aves, lo cual modifica la abundancia de todo el
campus. Por lo que sería conveniente realizar un estudio de la especie dominante de esta
área: Columbina cruziana, para observar horas de mayor y menor actividad, así como los
factores que producen dichos picos de abundancia.

140
6. Continuar con la cuantificación del efecto negativo de las aves a los campos de
cultivos e iniciar con la cuantificación del efecto positivo, ya que algunas especies de
aves pueden ser usadas como controladores biológicos.
7. Se recomienda una mejor reagrupación de los transectos, a fin de tener zonas más
homogéneas en términos de hábitat, ya que han habido cambios en la distribución
espacial del campus desde estudios anteriores.
8. Usar la información sobre diversidad y abundancia relativa en el campus de la
UNALM presentada por ésta investigación como herramienta para una mejor toma de
decisiones sobre el ordenamiento territorial de dicho espacio.
9. Por último, es necesario expandir el área de estudio fuera de los límites de la
UNALM, siendo posible un estudio de efecto borde y de flujo de especies en zonas
aledañas como la laguna de La Molina y otras áreas verdes de considerable importancia
en el distrito.

141
VII. REFERENCIAS BIBLIOGRAFICAS
Almazán–Núñez, RC; Hinterholzer-Rodríguez, A. 2010. Dinámica Temporal de la Avifauna
en un parque urbano de la ciudad de Puebla, México. Huitzil Revista Ornitológica Mexicana
11(1): 26-34.
Bencardino Martínez, C. 1937. Estadística Básica Aplicada. Tercera Edición. ECOE
Ediciones. Bogotá. 406 p.
Berenson, ML; Levine, DM; Krehbiel, TC. 2006. Estadística para la Administración.
Pearson Edition. 619 p.
Bibby, CM. 1998. Expedition field Techinques. Bird surveys. London. Expedition
Advisory Centre. 143p.
Blair, RB. 1996. Land use and avian species diversity along an urban gradient.
Ecological Aplications 6 (2): 506-519.
Bowman, R; Donelly, R; Marzluff, J. 2001. Avian ecology and conservation in an
urbanizing world. Kluwer Academic Publishers. Massachussets.
Canterbury, G; Martin, T; Petit, D; Petit, L; Bradfords, D. 2000. Bird Communities
and Habitat as Ecological Indicators of Forest Condition in Regional Monitoring.
Conservation Biology 14 (2): 544-558.
Castañeda Córdova, LZ. 2007. Caracterización y Evaluación de la Diversidad de la
Artropofauna en Pastizales Bajo el Efecto de una Quema Controlada, en la SAIS Túpac

142
Amaru. Tesis para optar título de Bióloga. Universidad Nacional Agraria La Molina.
Lima- Perú. 127p.
Castillo Palacios, L; Quinteros Carlos, Z; Castañeda Córdova, L. En Prensa. Las Aves
del Campus de la UNIVERSIDAD NACIONAL AGRARIA LA MOLINA. 1992-2011.
Lima-Perú.
Chace, JF; Walsh, JJ. 2004. Urban effects on native avifauna: a review. . Landscape
and Urban Planning. Versión Online. www.elsevier.com/locate/landurbaplan
Chanamé, J, Angulo, E, Carmona, M, Puse, E. 2010. Avifauna del Campus de la
Universidad Nacional Pedro Ruiz Gallo. Lambayeque. Octubre 2007-Abril 2008.
Ciencia, Tecnología y Humanidad 1(1): 47-58.
Congreso Nacional de Biología XV. Simposium Nacional de Educación en Ciencias
Biológicas IX. 2004. Universidad Católica de Santa María de Arequipa). 2004. Arequipa,
Perú. Eds. Chávez, C. Las Aves del Campus de la Universidad Nacional de Piura y sus
alrededores: 6 años mirando al cielo.
III Jornada Nacional de Ornitología. (1999. Calca, Cuzco, Perú) 1999. Chávez, C.;
Balmaceda, J; Viñas, P; Otero, D. (1999). Importancia del campus de la Universidad
Nacional de Piura en la conservación de aves silvestres.
Corrales Orozco, MS. 2012. Percepción de la Comunidad Universitaria sobre los
Vertebrados Urbanos en la Universidad Nacional Agraria La Molina. Tesis Título de
Biólogo. Lima. Perú. Universidad Nacional Agraria La Molina. 105 p.
Croci, S; Butet, A; Clergeau, P. 2008. Does urbanization filter birds on the basics of

143
their biological traits? The Condor 110 (2): 223-240.
Davic, RD. 2003. Liking Keystone Species and Functional Groups: A New
Operational Definition of the Keystone Species Concept. Conservation Ecology 7 (1).
Del Hoyo, J; Elliot, A; Christie, D, Sekerciouglu, C. 2006. Handbook of the birds of
the World. Volume 11: Old World Flycatcher´s to the Old World Warblers. Primera
Edición. Lynx Editions.
Faeth, SH; Bang, C; Saari, S. 2011. Urban biodiversity: patters and mechanisms.
Annals of the New York Academy of Sciences 1223: 69-81.
Faggi, A; Perepelizin, PV. 2006. Riqueza de aves a lo largo de un gradiente de
urbanización en la ciudad de Buenos Aires. Revista del Museo Argentino de Ciencias
Naturales 8(2): 289-297.
Giraldo Mendoza, A; Véliz Rosas, C; Arellano Cruz, G; Sanchez Infantas, E. 2002.
Uso de la Ley de Taylor en el Uso de Patrones Espacio-Temporal en Poblaciones de
Animales: Dos Ejemplos de Aplicación. Ecología Aplicada 1 (1).
Giraudo, L; Kufner, M; Torres, R; Tamburini, D; Briguera, V; Gavier, G. 2006.
Avifauna del bosque chaqueño oriental de la Provincia de Córdova, Argentina. Ecología
Aplicada 5 (1,2):
González, O. 1998. Coereba flaveola. Un ave nueva en el ecosistema de los parques
de la Ciudad de Lima. Ecología 1 (1): 79-83.
González, O. 2000a. Poblaciones de aves como indicadores de los cambios en el
ambiente. Xilema no. 16.

144
González, O. 2000b. Notas sobre distribución y poblaciones de aves en campos de
cultivo del Valle de Majes (Arequipa-Perú). Memorias del IV Congreso
Latinoamericano de Ecología.
González, O. 2002 Distribución y dispersión del mielero (Coereba flaveola, Aves:
Coerebidae) en la ciudad de Lima, Perú. Ecología Aplicada 1(1): 115-116.
González, O. 2004. Ecología de aves urbanas en un parque de la Ciudad de Lima.
Tesis de Maestría. Lima, Perú. Universidad Nacional Mayor de San Marcos. 102 p.
González, O. sf. Variación Espacio-Temporal de la Diversidad de Aves Urbanas en un
Área Verde de la Ciudad de Lima. Revista Científica Dilloniana 4 (1).
Hanneman, RA. Sf. Intoducción a los Métodos del Análisis de Redes Sociales.
Universidad de California Riverside.. Trudicida por Rene Rios. Universidad Católica de
Chile. Capítulo 9. 26 p.
Hernández Rodríguez, O. (1998). Temas de Análisis Estasítico Multivariado. Primera
Edición.Editorial de la Universidad de Costa Rica. 171p.
Koskimies, P. 1989. Birs as a tool in environmental monitoring. Ann. Zool. Fennici,
26, 153-166.
Koepcke, M. 1964. Las Aves del Departamento de Lima. Lima. Perú. Gráfica Morsom
S.A. 127 p.
Krebs, CJ. 1985. Ecología. Segunda Edición. Mexico. Harla, Harper & Row
Latinoamericana. 753 p.

145
Krebs, CJ. 1989. Ecological Methodology. New York. Harper Collins Publishers.
654 p.
Lawton, J. 1994. What do species do in ecosystems? Oikos 71 (3): 367-374
Leveau, CM ; Leveau, LM. 2004. Comunidades de aves e un gradiente urbano de la
ciudad de Mar del Plata, Argentina. Hornero 19 (1) : 13-21.
Leveau, CM ; Leveau, LM. 2006. Ensambles de aves en calles arboladas de tres
ciudades costeras del sudeste de la Provincia de Buenos Aires, Argentina. Hornero 21
(1) : 25-30.
López de Casenave, J. 2001. Estructura Gremial y Organización de un Emsamble de
Aves del Desierto del Monte. Tesis Doctor en Ciencias Biológicas. Buenos Aires.
Argentina.Universidad de Buenos Aires.
X Congreso y XVI Simposio Nacional de Ornitología. (2010) MacGregor-Fors, I.
Ornitología Esquina con Urbanización, entre Ecología y Planeación. Conferencias
Magistrales.
Mandujano, S. 1994. Conceptos Generales del Método de Conteo de Animales en
Transectos. Ciencia 45: 203-211.
Margalef, R. 1991. Ecología. Séptima Edición. Barcelona. Ediciones Omega S.A. 951
p.
Magurran, AE. (1989). Diversidad Ecológica y su Medición. Barcelona. 200 p.
Martínez, O; Oliviera, M; Quiroga, C; Gòmez, I. 2010. Evaluación de la avifauna de
la ciudad de La Paz, Bolivia. Revista Peruana de Biologìa 17 (2): 197-206.

146
Marzluff, JM. 2005. Island Biogeography for an Urbanizing world: How Extinction
and Colonization may Determine Biological Diversity in Human-Dominated Landscapes.
Urban Ecosystems 8: 157-177.
Menge BA; Freidenburg, TL. 2001. Keystones Species, Encyclopedia of Biodiversity
3: 613-631.
Mills, LS; Soulé, ME; Doak, DF. 1993. The Keystone-Species Concept in Ecology
and Conservation. BioScience 43 (4): 219-224Moreno, CE. 2001. Métodos para medir la
biodiversidad. Zaragoza. GORFI S.A. Volumen I.79 p
Odum, EP; Barret, G. 2006. Fundamentos de Ecología. Quinta Edición. Mexico.
Cengage Learning Editores. 598 p.
Ortiz Z, C. 2012. Alimentación Nocturna de la Golondrina Azul y blanca
Pygochelidon cyanoleuca (Hirundinae) en el Perú. The Biologist (Lima) 10 (1), Jan-Jun:
74-75.
Pedroza, H; Dicovskyi, L. (2007) Sistema de Análisis Estadístico con SPSS. IICA.
INTA. Managua. 165 p.
Quinteros Carlos, DZ. (1992) Determinación de los Patrones de uso Temporal y
Espacial de los Cultivos de Maíz por las Aves Granívoras de la UNALM. Tesis título de
Bióloga. Lima. Perú. Universidad Nacional Agraria La Molina. 131 p.
Quinteros, Z; Sánchez, E; Tovar, H. 2002. Relación entre la abundancia relativa y
densidad real entre 3 poblaciones de aves. Ecología Aplicada, 1(1): 95-104.

147
Padoa-Schioppa, E; Beitto, M; Massa, R; Bottoni, L. 2005. Bird communities as
bioindicator: The focal species concept in agricultural landscapes. Ecological Inicators 6:
83-93.
Payton, IJ; Fenner, M; William, GL. c 2002. Keystone species: the concept and its
relevance for conservation and management in New Zealand. Science For Conservation
203.
Plengue, M. (2012) List of the Birds of Perú. Boletin UNOP
PNUMA. Centro de Monitoreo de la Conservación Mundial. 2011. Lista de Especies
CITES. Parte 1. Índice de Especies CITES. (en línea) Ginebra. Suiza. Consultado 28 de
Enero del 2013.
http://www.cites.org/esp/resources/pub/checklist11/Indice_de_especies_CITES.pdf
Ralph,CJ ; Geupel, GR ; Pyle, P ; Martin, TE ; DeSante, DF ; Milá, B. 1996. Manual
de Métodos de Campo para el Monitoreo de aves terrestres. (L. Villaseñor, & I.
Hartasánchez, Trads.) California. Pacific Southwest Research Station.43 p.
Ramírez, A. 2000. Utilidad de las aves como indicadores de la riqueza especìfica
regional de otros taxones. Ardeola. 47 (2) : 221-226.
Ramírez C, D & González M, O. 2001. Análisis de dos Métodos de Muestreo para el
Cálculo de la Diversidad de Especies de Aves Terrestres en la Ciudad de Lima, Perú.
Biota N° 100: 114-123.
Rodriguez, J. c2010. Ecología. Madrid. Ediciones Pirámide. 502 p.

148
Salvador, SA. 2012. Reproducción del Gavilán Mixto (Parabuteo unicinctus) en Villa
María, Córdova, Argentina. Historia Natural. Tercera Serie. 2 (1): 103-115.
Sanchez Infantas, EH. 1992. Diversidad y Estabilidad de Comunidades de Aves en la
Reserva Nacional de Lachay. Tesis Grado Magister Scientiae. Lima. Perú. Universidad
Nacional Agraria La Molina. 111 p.
Santana Vergara, CA. 2011. Patrones de Variación Espacio Temporal Diurno de las
Aves en el Aeropuerto Internacional Jorge Chávez durante los años 2009-2010 y
descripción de su relación con las operaciones de vuelos de aeronaves. Tesis Título de
Biólogo. Lima. Perú. Universidad Nacional Agraria La Molina. 132 p.
Schmitz, OJ. 2007. Ecology and Ecosystem Conservation. Washington. Island
Press.166 p.
Schulenberg, TS; Stotz, DE; Lane, DF; O’Neill, JP; Parker III, TA. 2007. Birds of
Peru. New Jersey. Princenton Field Guides. 656 p.
Smith, R., & Smith, T. 2001. Ecología. Cuarta Edición. Madrid. Pearson Education
S.A. 642 p.
Steneck, RS. 2001. Functional Groups. Encyclopedia of Biodiversity 3: 122-139.
Takano Goshima, F. 2010. Diversidad de aves y estimación del daño que causan sobre la
producción de maíz en el campus de la UNALM. Lima. Perú. Tesis de Licenciatura.
Universidad Nacional Agraria La Molina. 83 p
Takano G, F; Castro I, N. 2007. Avifauna en el Campus de la Universidad Nacional
Agraria La Molina (UNALM), Lima-Perú. Revista de Ecología Aplicada , 6 (1,2)

149
Texeira Roth, V y Sánchez Infantas, E. 2006. Patrones Poblacionales de las
Principales Especies Herbáceas en la Reserva Nacional Lachay. Revista de Ecología
Aplicada. 5 (1,2): 23-27.
Torres Núñez, ME. 2006. Dinámica Temporal de la comunidad de Aves Limicolas
(Haematopodidae, Charadriidae, Scolopacidae, Recurvirostridae y Phalaropodidae) de la
zona Reservada de Pantanos de Villa durante el año 2005. Tesis Título de Biólogo. Lima.
Perú. Universidad Nacional Agraria La Molina. 107 p.
Torres, M., Quinteros, Z. y Takano F. 2006. Variación Temporal de la Abundancia y
diversidad de aves Limícolas en el Refugio de Vida Silvestre Pantanos de Villa. Lima,
Perú. Ecología Aplicada 5 (1,2): 119-125.
Vilà, M. 1998. Efectos de la diversidad de especies en el funcionamiento de los
ecosistemas. Orsis, 13: 105-117.
Véliz Rosas, C. 2002. Resiliencia en Comunidades de aves en la Reserva Nacional de
Lachay luego del evento EL Niño (1997-98)”. Tesis Titulo de Biólogo. Lima. Perú.
Universidad Nacional Agraria La Molina. 128 p
Walker, BH. 1992. Biodiversity and Ecological Redundancy. Conservation Biology 6
(1): 18-23.
Walker, BH. 1995. Conserving Biological Diversity through Ecosystem Resilience.
Conservation Biology 9 (4): 747-752.