
UNIVERSIDAD DE GRANMA FACULTAD DE MEDICINA...
129
UNIVERSIDAD DE GRANMA FACULTAD DE MEDICINA VETERINARIA CENTRO DE ESTUDIO DE PRODUCCIÓN ANIMAL “INVENTARIO DE LA SUCESIÓN VEGETAL SECUNDARIA Y CARACTERIZACIÓN DEL Pennisetum purpureum EN LA AMAZONIA ECUATORIANA” Tesis presentada en opción al grado científico de Doctor en Ciencias Veterinarias Ing. Hernán Alberto Uvidia Cabadiana. MSc. Tutores: Lic. Jorge Ramírez de la Ribera. Dr.C. Ing. Ismael Leonard Acosta. Dr.C. Bayamo MN Año 2015
Transcript of UNIVERSIDAD DE GRANMA FACULTAD DE MEDICINA...

UNIVERSIDAD DE GRANMA
FACULTAD DE MEDICINA VETERINARIA
CENTRO DE ESTUDIO DE PRODUCCIÓN ANIMAL
“INVENTARIO DE LA SUCESIÓN VEGETAL SECUNDARIA Y
CARACTERIZACIÓN DEL Pennisetum purpureum EN LA AMAZONIA
ECUATORIANA”
Tesis presentada en opción al grado científico de Doctor en Ciencias Veterinarias
Ing. Hernán Alberto Uvidia Cabadiana. MSc.
Tutores: Lic. Jorge Ramírez de la Ribera. Dr.C.
Ing. Ismael Leonard Acosta. Dr.C.
Bayamo MN
Año 2015

AGRADECIMIENTOS
Como decía William Osler “En ciencia el reconocimiento se concede al hombre que
convence al mundo, no a aquel a quien se le ocurre la idea”. Cuando en el devenir
del tiempo, surge un evento trascendente en la vida de las personas, y éste lo es
excepcional, es bueno hacer una reflexión, que como decía Menandro “El fruto más
dulce a recoger en esta vida es el del reconocimiento”. La excelencia, implica, por un
lado, la capacidad de liderazgo; el fundamento de principios y valores. Mientras que
por otro, la responsabilidad y el compromiso personal, hacia los demás sean éstos
compañeros, amigos y por supuesto la familia. Sin embargo, la luz de la excelencia,
llega a la plenitud si ésta, está acompañada de humildad. Es decir, eliminar el falso
orgullo por una meta lograda.
A todas las personas e instituciones que de una u otra forma contribuyeron para
alcanzar esta meta tan anhelada, a los profesores de la Facultad de Medicina
Veterinaria, a la Universidad de Granma, a la Secretaria Nacional de Educación
Superior, Ciencia, Tecnología e Innovación (SENESCYT), al Gobierno del Ecuador, a
las autoridades de la Universidad Estatal Amazónica, de manera especial al Dr. Julio
Cesar Vargas Burgos.
Gracias amada esposa Ximena, mis hijos Luis, Jhoeel y mi princesita Johannnita, a
mis padres que cuidan nuestros pasos desde el cielo, a mis hermanos y familiares.
De manera especial al Dr. C. Jorge Ramírez de la Rivera, Dr. C. Ismael Leonard y mi
amigo Dr. C. Danis Verdecia.

DEDICATORIA
A Dios y la memoria de mis padres Luis e Hilda.

Título: Inventario de la sucesión vegetal secundaria y caracterización del
Pennisetum purpureum en la Amazonia Ecuatoriana.
SÍNTESIS
La siguiente investigación tuvo como objetivo identificar y describir las especies que
conforman la sucesión vegetal secundaria de la región Amazónica Ecuatoriana, así
como determinar el rendimiento, indicadores morfológicos y calidad de la especie
Pennisetum purpureum y su relación con la edad, y los factores del clima, al emplear
diferentes métodos de plantación. El experimento se desarrolló en un suelo Inseptisol
subtipo Flavaquentic Eutrudepts. Los resultados ofrecen la identificación de diferentes
géneros en la sucesión secundaria vegetal amazónica, dentro de los que resaltan el
Ipomoea, Desmodium, Vismia, Paspalum, Solanum, entre otros. La caracterización
del Pennisetum purpureum vc Maralfalfa reflejó que para la cobertura aérea (CA),
basal (CB), altura, número de hojas (NH), tallos por plantón (TPP), diámetro de la
hoja (DH) y diámetro del tallo, estuvieron definidos por el número de estaca y la
distancia de siembra en las edades evaluadas. Se determinó la relación funcional de
la edad con indicadores morfológicos y de calidad, para los métodos uno (una estaca
a 0,50m) y dos (una estaca a un metro). La producción total de materia verde y seca
se incrementó con la edad, para (P<0,001), en ambos casos, y se ajustaron
ecuaciones cuadráticas, los valores mas altos se mostraron a los 90 días. Para las
restantes variables morfológicas, tanto para el método uno como el dos, se
mostraron expresiones lineales, cuadráticas y cúbicas, para (P<0,001). La proteína
bruta disminuyó con la edad para (P<0,001), aumentando la fibra y el calcio, el fósforo
presentó para los dos métodos bajos ajustes. Las variables climáticas precipitaciones
y temperatura mostraron altas correlaciones con los indicadores fenológicos y
químicas, fundamentalmente en el método dos. Se establecieron ecuaciones de
regresiones múltiples que permiten calcular la producción de materia verde y seca
total, así como indicadores de calidad, a través de las precipitaciones y la humedad.
La información obtenida permite conocer la sucesión vegetal secundaria de la
Amazonía Ecuatoriana, emplear las ecuaciones matemáticas que expresan la
relación de los indicadores morfológicos y de la calidad con la edad de rebrote y los
factores climáticos, sin el empleo de costosas técnicas de laboratorio.

ÍNDICE
INTRODUCCIÓN.…………………………………………………………………… 1
CAPÍTULO I. REVISIÓN BIBLIOGRÁFICA……………………………………... 5
I.1. La ganadería en el mundo……………………………………………………. 5
I.1.1 Ganadería en América Latina y el Caribe……………………………......... 6
I.2. Características de la Amazonia Ecuatoriana………………………………. 10
I.3. Sucesión vegetal……………………………………………………………… 12
I.3.1. Tipos de sucesión vegetal………………………………………………….. 13
I.3.2. Características de una sucesión………………………..………………… 15
I.4. Caracterización y comportamiento del Pennisetum purpureum en Ecuador 17
I.4.1. Descripción botánica…………………………………………………………… 19
I.4. 2. Métodos de propagación, selección del suelo y preparación……………. 20
I.4.3. Profundidad de plantación………………………………………….………… 21
I.4.4. Densidad y distancia de plantación............................................................ 22
I.4.5 Rendimiento y calidad nutritiva del Pennisetum purpureum vc. Maralfalfa
23
I.5. Factores que afectan el crecimiento y calidad……………………………….. 24
I.5.1. Temperatura…………………………………………………………………….
25
I.5.2 Radiación solar……………………………………………………………….
.………………………………………………………………………….
27
I.5.3 Precipitaciones…………………………………………………………………. 28
I.5.4 Frecuencia y altura de corte o pastoreo………………………………………
30
I.5.5 Carga y tiempo de estancia…………………………………………………… 33
CAPÍTULO II INVENTARIO Y DESCRIPCIÓN DE LA SUCESIÓN VEGETAL
SECUNDARIA EN LA PROVINCIA PASTAZA

II.1. Introducción…………………………………………………………………….. 35
II.2. Materiales y Métodos……………………………………... ………………… 36
II.2.1 Área de la investigación……………………………………………………... 32
II.2.2 Procedimiento………………………………………………………………… 38
II.2.3 Identificación de las especies………………………………………………. 39
II.2.3 Resultados…………………………………………………………..
39
II.2.4. Discusión…………………………………………………….
61
CAPÍTULO III CARACTERIZACIÓN DEL PENNISETUM PURPUREUM VC
MARALFAL EN LA AMAZONÍA ECUATORIANA
III.1 Introducción……………………………………………………………………….
66
III.2. Materiales y Métodos…………………………………………………………….
67
III.2.1. Manejo del experimento………………………………………………………. 67
III.2.2. Preparación del terreno………………………………………………………. 67
III.2.3. Siembra………………………………………………………………………… 68
III.2.4 Toma de muestras……………………………………………………………… 68
III.2.5 Tratamientos y Diseño experimental…………………………………………. 69
III.2.6 Mediciones experimentales…………………………………………………… 69
III.2.7 Análisis estadístico y cálculo…………………………………………………. 69
III.2.8 Procedimiento…………………………………………………………………… 70
III.2.9 Composición química……………..…………………………………………… 70
III.3 Resultados………………………………………………………………………. 71
III.4 Discusión…………………………………………………………………………. 84

CONCLUSIONES………………………………………………………………….. 100
RECOMENDACIONES…………………………………………………………… 101
REFERENCIAS BIBLOGRÁFICAS…………………………………………….. 102

INTRODUCCIÓN
El ecosistema amazónico, en especial su bosque tropical lluvioso, contiene los
hábitats vegetales y animales más ricos y complejos del mundo. En la existencia de
una prolífica flora y fauna junto a extraordinarias variaciones de macro y micro hábitat
radica la característica más importante de esta región.
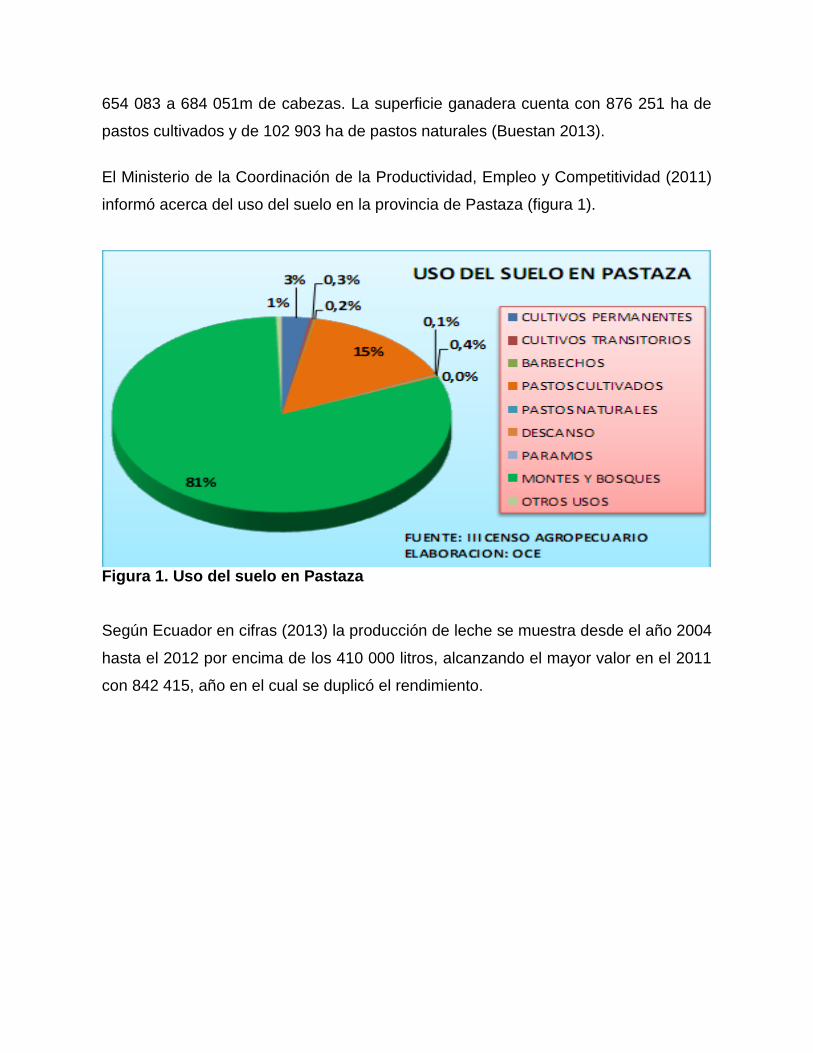
La provincia de Pastaza, ocupa el tercer lugar de la Región Amazónica, con 430 mil
hectáreas de tierra utilizada en usos agropecuarios y afines, lo cual equivale al 28 %
del total de la región y el tres del total del país. Predomina la existencia de montes y
bosques que ocupan el 81 % de la superficie, que corresponden a la selva amazónica
sin producir e inexplorada, seguida de pastos cultivados con el 15 %, y apenas el 3
con cultivos permanentes.
La principal actividad económica de la población Amazónica es la agropecuaria, la
cual, junto con los procesos de migración desde otras áreas del país, han causado
una expansión de la frontera agrícola, acompañada en la mayoría de los casos por
una explotación desmedida y sin control sobre los recursos naturales. Sumado a esto,
la aplicación de políticas orientadas al fomento productivo no acordes a la realidad
socioambiental amazónica, han generado un débil impacto en las condiciones de vida
de la población.
El 28 % de las fincas en la provincia Pastaza pertenecen a pequeños productores. A
pesar de que hay una fuerte concentración de la población en la ciudad de Puyo, sin
embargo, la Población Económicamente Activa (PEA) de Pastaza se encuentra
concentrada también en las zonas rurales, por lo que su principal actividad es la
producción agropecuaria que capta el 37% de la PEA (MCPEC 2011).
La actividad ganadera enfrenta una serie de limitaciones que impiden lograr una alta
producción y productividad. Entre las más importantes destaca la baja capacidad del
ecosistema para resistir las tecnologías actualmente empleadas, donde existen varios
sistemas de producción ganadero con pastoreo a campo abierto, sogueo y
semiestabulado, en terrenos con escasa fertilidad, con pastos poco adaptados a las

condiciones climáticas, estos presentan un deterioro muy visible y progresivo con
baja productividad de biomasa, lo cual trae como consecuencia una disminución
significativa de la producción animal (INIAP 2011).
Por otra parte, la actividad ganadera es vista como una de las principales causantes
del cambio climático (FAO 2013). En el avance de la frontera agrícola de la
Amazonía, la pecuaria se considera como la actividad productiva más predatoria,
porque depende de la sustitución de grandes extensiones de bosques nativos
primarios, por monocultivos de gramíneas forrajeras para la formación de pasturas
(FAO 2009). La intensión de este impuesto lo propició la ley de reforma agraria de
1964 donde se amenazaba a los nuevos habitantes de la Amazonía (colonizadores),
con expropiar fincas que poseyeran el 75% de terreno improductivo lo que obligó a
reemplazar el bosque primario por extensas áreas de pasturas. Estableciendo así
pastos, principalmente Axonophus escoparius, Ramírez et al. (1996) con una baja
carga animal (0,7 a 0,8 UBA.ha-1).
La cantidad y calidad nutricional del forraje disponible, es uno de los factores más
importantes que controlan el consumo animal, y por lo tanto su producción se ve
afectada por una presión de pastoreo constante, de allí que la carga animal/ha/año,
depende fundamentalmente de nuestros conocimientos respeto al manejo técnico del
complejo “Suelo-Planta-Animal”, lo que permite obtener el máximo beneficio técnico,
productivo y económico por unidad de animal, de área y de tiempo, sin afectar
ninguna de estas partes del complejo (Meneses 2011).
Para atenuar esta situación se han realizado grandes esfuerzos en la introducción de
especies y variedades de mayor rendimiento y calidad como el Pennisetum
purpureum vc. Maralfalfa. Sin embardo, su crecimiento, productividad y calidad
están influenciados por las condiciones climáticas de la región.
Es importante destacar que los avances alcanzados en el campo de la nutrición
hacen necesario el conocimiento, cada día más preciso, del valor alimenticio de los
pastos y forrajes, los cuales constituyen en el trópico la mayor fuente de alimentación
de los animales y la más económica por lo que es sumamente importante conocer el

valor alimenticio de los diferentes forrajes verdes o conservados que pueden formar
parte de la ración y que permite exteriorizar el máximo potencial de producción de los
animales (Cáceres y González 2000).
En tal sentido Herrera (2006) argumentó que los pastos resultan ser una fuente
apropiada de alimentos para el vacuno, principalmente en países de clima tropical.
Ello es debido al elevado número de especies que pueden ser utilizadas, posibilidad
de cultivarlos todo el año, la capacidad del rumiante de utilizar alimentos fibrosos, no
compiten como alimento para el ser humano y suele ser una fuente económica.
Problema: La expansión de la frontera agrícola con el desconocimiento de la
sucesión vegetal secundaria, composición florística, y consiguiente utilización para la
alimentación animal en la Amazonía Ecuatoriana, trae consigo el deterioro del
entorno ambiental y la baja productividad de los sistemas. Además, no se cuenta con
la información que permita relacionar los factores climáticos y la edad de rebrote con
el rendimiento e indicadores de la calidad de especies introducidas.
Hipótesis: El detener la expansión de la frontera agrícola y el consiguiente
conocimiento de la sucesión vegetal secundaria, su empleo en la alimentación, así
como contar con la información del rendimiento, composición química y el efecto de
los factores del clima, y la edad, de la especie Pennisetum purpureum vc. Maralfalfa
introducida en la región Amazónica del Ecuador, podría constituir un elemento básico
para un mejor uso y manejo de esta especie en este ecosistema frágil.
Objetivo general
Identificar y describir las especies que conforman la sucesión vegetal secundaria de
una zona en la región Amazónica Ecuatoriana, así como determinar el rendimiento y
calidad de la especie Pennisetum purpureum y su relación con la edad y los factores
del clima expresado a través de ecuaciones matemáticas.

Objetivos específicos
1. Inventariar y describir las especies que conforman la sucesión vegetal
secundaria en áreas frágiles de la Provincia Pastaza.
2. Determinar indicadores del rendimiento, la morfología y calidad del Pennisetum
purpureum vc Maralfalfa al emplear diferentes métodos de plantación.
3. Determinar la relación del rendimiento y la composición química con la edad
del Pennisetum purpureum vc Maralfalfa mediante expresiones matemáticas,
al emplear diferentes métodos de siembra durante el establecimiento.
4. Establecer expresiones matemáticas que permitan relacionar los factores
climáticos (precipitaciones, temperatura afuera y a la sombra, punto de rocío,
humedad y sensación térmica, y el rendimiento e indicadores de la calidad en
el Pennisetum purpureum vc Maralfalfa al emplear diferentes métodos y
densidad de siembra para el establecimiento.
Novedad Científica
Para la Provincia Pastaza del Ecuador se inventariaron y describieron las
especies que conforman la sucesión vegetal secundaria en áreas frágiles, que
pueden ser empleadas en la alimentación animal y en otros fines.
Se caracterizó química y morfológicamente el Pennistum purpureum vc
Maralfalfa al emplear diferentes métodos en su establecimiento.
Se determinaron expresiones matemáticas no existente, que permiten
establecer la relación entre la edad, el rendimiento y la calidad del Pennisetum
purpureum vc Maralfalfa, con los factores del clima, al plantar la especie con
diferentes método y densidad de siembra.
La región cuenta con información no disponible hasta este momento que
permitirá mejorar la preparación de los recursos humanos.

CAPÍTULO I
REVISIÓN BIBLIOGRÁFICA
I.1 La ganadería en el mundo
La alimentación para la población mundial es uno de los desafíos más importantes
para todos los involucrados en el desarrollo de la humanidad, ya que, más de un
billón de personas se sustentan del sector ganadero, y el 70% de los 880 millones de
pobres rurales que viven con menos de un USD por día, dependen al menos
parcialmente de la ganadería para su subsistencia.
Este problema se da por el crecimiento de las poblaciones humanas y, en
consecuencia, a la presión cada vez mayor que ejercen sobre los recursos naturales
FAO (2013a).
Por otra parte, es necesario utilizar de manera más eficiente recursos como el agua,
los combustibles fósiles y los cereales empleados en la cría de animales, reciclar y
disminuir los residuos, establecer un balance positivo en la contribución del ganado al
suministro mundial de alimentos. En estas circunstancias el ganado realiza una
contribución vital al generar un flujo de dinero en efectivo y actuar como amortiguador
económico, siempre y cuando las cadenas de comercialización estén organizadas
para permitir la entrada de productores y comerciantes de pequeña escala, y de
zonas remotas (FAO 2011).
Para las sociedades que se basan en la ganadería como medio de vida más
importante y principal fuente para la seguridad alimentaria, el manejo de los animales
es determinante en la configuración del modo de subsistencia. Estas sociedades
tienen sistemas de producción basados en el pastoreo extensivo, los cuales se
definen como aquellos en los que al menos el 90% del valor total de la producción
proviene de la actividad ganadera, y más del 90 de la materia seca de los piensos
con que se alimenta a los animales provienen de pastizales y forrajes anuales
(Sere y Steinfeld 1996).

El mayor número de personas que dependen del ganado (120 millones) según la
FAO (2011), se encuentra en las sociedades pastoralistas, donde los animales
suministran a sus propietarios leche y ocasionalmente sangre y carne. Se conoce que
diferentes comunidades practican el pastoreo nómada, desplazando a los animales a
través de extensas áreas comunitarias de pastizales. Los ganaderos que crían a los
animales en pastizales en condiciones extensivas son otro ejemplo de sociedades
basadas en la ganadería. Su número es considerablemente inferior al de los
pastores, pero realizan una contribución importante al suministro pecuario total a nivel
nacional y mundial.
I.1.1 Ganadería en América Latina y el Caribe
El proceso de expansión de la ganadería que están viviendo los países de América
Latina, representa tanto una oportunidad como una amenaza para el desarrollo
sustentable de la región. Por un lado, generar riqueza y mitigar la pobreza si se
toman las decisiones políticas adecuadas y se promueven sistemas de producción
ganaderos sustentables y amigables con el ambiente. Por el otro, es una amenaza si
la expansión de la actividad continúa sin considerar los costos ambientales y los
potenciales efectos de marginalización de los pequeños productores (FAO 2013).
Tomando en cuenta la disponibilidad de tierras que caracteriza a la región, el proceso
de expansión ganadera no es un fenómeno aislado del desempeño de los otros
grandes subsectores: la agricultura de granos y la forestaría. Sin lugar a dudas, por
sus dimensiones territoriales y amenazas ambientales la agricultura de granos es el
subsector que más se relaciona con la ganadería en un proceso de crecimiento con
grandes interacciones. Por consiguiente, es necesaria una planificación del
crecimiento, y la intensificación que capitalice las sinergias positivas que la
integración de la agricultura con la pecuaria y eventualmente con la forestación
puedan otorgar a la sostenibilidad y competitividad de los sistemas de producción
(FAO 2014).

Las nuevas tecnologías y prácticas de producción, incluidas razas mejoradas, han
sido adoptadas ampliamente en América Latina y el Caribe durante la última década,
lo cual ha generado un sólido aumento en la producción de carne y leche por cabeza
en todas las especies de ganado (FAO 2013b).
Según la FAO (2014) los inventarios ganaderos están distribuidos de manera
dispareja entre los países de América Latina y el Caribe. Los tres países más grandes
de la región con la mayor cantidad de cabezas reflejan alrededor del 70% del ganado
vacuno y porcino, 64 del ganado lechero, 60 de las aves y casi la mitad de las ovejas.
Brasil continúa siendo líder en la producción de las principales especies de ganado,
con más de la mitad de todos los inventarios de ganado vacuno, lechero y porcino,
alrededor del 40% de los inventarios avícolas y 20 de los inventarios de ovejas. Sin
embargo, el país está a la zaga de muchas otras naciones de América Latina y el
Caribe en lo que a productividad de la carne se refiere.
Por otra parte, la producción de vacuno en México, el segundo mayor productor de
ALC, se mantuvo en alrededor de 1,8 millones de toneladas en el 2011 y el 2012. Es
poco probable que aumente en el 2015, puesto que la industria ganadera se debate
con los efectos de una prolongada sequía y altos precios del forraje
(Hernández et al. 2013).
Destacar, que a medida que ha aumentado la producción ganadera en América
Latina y el Caribe, también se incrementaron los desafíos ambientales, especialmente
en lo que se refiere a las tasas de deforestación. En varios países de la región, se
llevan a cabo transformaciones en los sistemas ganaderos hacia esquemas mixtos
y más intensivos de cultivos/ganadería y producción de lácteos, con inversiones en
infraestructura de transporte y la conversión de tierras de pastoreo en tierras de
labranza (Wassenaar et al. 2007).
América Latina, con sus extensas áreas de pasturas, un régimen climático favorable y
un uso racional de insumos, que incluye granos (cereales, soya) y fertilizantes, cuenta
con todos los ingredientes naturales para ser un importante productor pecuario, para

satisfacer las demandas de alimentos y garantizar la seguridad alimentaria regional y
mundial. Sin embargo, debe existir una política adecuada para el uso de estos y
evitar la degradación de los sistemas pastoriles.
El sector pecuario en América Latina, ha crecido a una tasa anual (3,7%) superior a la
tasa promedio de crecimiento global (2,1). Durante el último tiempo, la demanda total
de carne se incrementó en 2,45, siendo mayor la demanda de ave (4,1), seguida por
la de cerdo (2,67), mientras que la demanda por la carne vacuna se redujo levemente
(0,2). Las exportaciones de carne crecieron a una tasa de 3,2 superiores al
crecimiento de la tasa de producción de 2,75% (FAO 2012).
En las últimas décadas, la ganadería ha tenido un enorme crecimiento,
especialmente en el cono Sur, debido a la expansión de la demanda mundial. Este
acelerado crecimiento ha permitido que se convierta en la región que más exporta
carne bovina y de ave a nivel mundial (Alltech 2015).
Por otra parte, el rápido crecimiento del sector pecuario regional, dos veces superior
al crecimiento promedio mundial, ha ejercido alta presión sobre la base de recursos
naturales, especialmente, la pérdida de cobertura forestal para la producción de
ganado en pastoreo o la producción de granos para los sistemas intensivos de
producción avícola y porcina, situación muy marcada en la Amazonía Ecuatoriana.
Además, es muy probable que de acentuarse los problemas de degradación de
suelos, la producción pecuaria se enfrente a competencia por tierras para la
producción agrícola o agroenergética y corre el riesgo de ser desplazada a zonas
marginales. Los países de la región requieren, por lo tanto, mejorar su capacidad de
análisis y manejo de riesgos en el sector ganadero, y desarrollar estrategias de
producción viables desde el punto de vista técnico, económico, social y ambiental.
Las proyecciones actuales indican que el consumo de carne a nivel mundial se
duplicará en los próximos 20 años. Aunque es una buena noticia para la
seguridad alimentaria de millones de personas, enfrentar esa demanda presionará
el avance de la frontera agrícola-ganadera a zonas de mayor vulnerabilidad
ambiental (FAO 2013a).

Esto puede incrementar los niveles de deforestación en la región, la degradación de
los suelos, la pérdida de biodiversidad y la disminución del recurso hídrico, si no se
toman medidas para evitarlo. Se deben tomar acciones decididas para que el
crecimiento del sector se lleve a cabo de modo sostenible y que contribuya, al mismo
tiempo, a la mitigación del cambio climático, de la pobreza y a la mejora de la salud
humana. Situación está que afecta a la región Amazónica del Ecuador, en especial a
la provincia Pastaza.
Según la FAO (2013b) cerca del 70 % de las áreas de pastoreo de América Latina y
el Caribe se encuentran en proceso de degradación. La deforestación provocada por
el aumento de sistemas extensivos de pastoreo en ciertas zonas es una característica
común en países de América Central y del Sur, sin embargo, hay estrategias
tecnológicas y de manejo para hacer una intensificación sostenible de la producción
pecuaria y evitar la deforestación y ampliación de la frontera ganadera. Es decir, el
sector puede desempeñar un papel clave en la mitigación del cambio climático.
La ganadería puede desarrollar un papel importante tanto en la adaptación al cambio
climático como en mitigar sus efectos en el bienestar de la humanidad. Para
aprovechar el potencial del sector, contribuir a la mitigación y adaptación al cambio
climático mediante una mayor capacidad de vigilar, informar y verificar las emisiones
de la producción pecuaria, será necesario el desarrollo de nuevas tecnologías, donde
los diferentes factores que afectan la calidad y el rendimiento desempeñan un
importante papel. Esto dependerá de las características de cada región.
I.2 Características de la Amazonia Ecuatoriana
En la Amazonía Ecuatoriana el 82% de la superficie con uso agropecuario está
dedicado a pastizales, lo cual demuestra que la ganadería es uno de los rubros de
mayor importancia para la economía campesina (INIAP 1997). Sin embargo, los
niveles de producción, productividad e ingresos en estos sistemas ganaderos son
bajos, por ejemplo; la producción promedio de forraje fue oscilando de 5 a 8

tMS/ha-1/año-1. Esta situación se relaciona a los siguientes factores: suelos pobres en
nutrientes, los pastos utilizados son vulnerables a las plagas y enfermedades, pocos
resistentes a la sombra y baja producción, escasa presencia de árboles y
leguminosas, contaminación de los suelos y agua por actividad petrolera, entre otros
aspectos (INIAP 2011).
Los factores identificados y la necesidad de especies forrajeras de amplia producción
de biomasa, adaptadas a condiciones edafoclimáticas de la Amazonía se consideran
como limitantes para implementar sistemas ganaderos sustentables en la RAE. En la
actualidad la ganadería ocupa 767 306 ha de pastos mejorados y 24 616 ha de
pastos naturales sobre suelos degradados, que causa graves problemas al desarrollo
agropecuario de la zona; por lo que es necesario buscar alternativas sustentables
para recuperar estas áreas degradadas y evitar que se continúen deforestando el
bosques para el establecimiento de nuevas pasturas.
La ganadería bovina en la región Amazónica Ecuatoriana se maneja en base a
técnicas empíricas horizontalmente sin controles ni registros del manejo, pastoreo
controlado con el sogueo. En la zona baja se mantiene la menor cantidad de
animales en sistemas en condiciones idóneas para el bienestar animal, y se
presentan peores condiciones para la implementación de producciones sostenibles,
las principales causas de la baja productividad son: raza o cruces de ganado de bajo
potencial productivo, prácticas incipientes de manejo al ganado, no aparecen
registros de producción y reproducción, mantienen un solo grupo del hato bovino, no
controlan a tiempo las principales enfermedades que afectan a las ganaderías de la
Amazonia, falta de capacitación y transferencia de tecnología, entre otros aspectos
(INIAP 2011).
Estos problemas influyen para que la ganadería de la Amazonía no alcance un
desarrollo armónico y adecuado, sistemas rentables, conveniente niveles de
producción, productividad e ingresos estables. Por otra parte, a partir del año 2004
hasta el 2012, se ha incrementado el ganado en la Amazonía Ecuatoriana de
654 083 a 684 051m de cabezas. La superficie ganadera cuenta con 876 251 ha de
pastos cultivados y de 102 903 ha de pastos naturales (Buestan 2013).
El Ministerio de la Coordinación de la Productividad, Empleo y Competitividad (2011)
informó acerca del uso del suelo en la provincia de Pastaza (figura 1).
Figura 1. Uso del suelo en Pastaza
Según Ecuador en cifras (2013) la producción de leche se muestra desde el año 2004
hasta el 2012 por encima de los 410 000 litros, alcanzando el mayor valor en el 2011
con 842 415, año en el cual se duplicó el rendimiento.

Figura 2. Uso de la frontera agrícola en la provincia de Pastaza
Fuente: (MCPEC 2011)
Por otra parte, en la región Amazónica Ecuatoriana existen varios sistemas de
producción ganadera con pastoreo: a campo abierto, sogueo y semiestabulado,
en terrenos poco fértiles, con pastos no adaptados a las condiciones climáticas, los
mismos que presentan un deterioro muy visible y progresivo con baja productividad
de biomasa, disminuyendo significativamente la producción animal (INIAP 2002).
Es importante destacar que el uso indiscriminado de la tierra reduce los bosques e
incrementan la frontera agrícola (figura 2), lo que trae consigo la aparición de la
vegetación secundaria sucesiva.
I.3 Sucesión vegetal
La mayoría de los tipos de vegetación están sujetos a cambios temporales tanto en
su composición como en la importancia relativa de las formas de vida. Estos cambios
pueden ser clasificados en dos grandes categorías, sucesionales y cíclicos. Durante
los cambios sucesionales existe una modificación progresiva en la estructura y
composición específica de la vegetación, mientras que en los cambios cíclicos tipos
similares de vegetación ocurren en el mismo lugar en intervalos de tiempo. En el caso

de la sucesión, una nueva distinción se pueden establecer considerando si los
cambios ocurren durante la colonización de un nuevo hábitat desprovistos de
vegetación y suelo (sucesión primaria), y aquellos procesos de sucesión que se
producen en la recolonización de un ambiente sometido a disturbios (sucesión
secundaria) (Clark y Clark 1993; Vandermeer et al. 1997; Vandermeer et al. 1998).
El concepto de sucesión vegetal tiene una larga historia, una de las primeras
aproximaciones fue enunciada por Clements (1916) y aplicados a humedales por
Pearsall y Wilson en las décadas del 20 y 30. Odum (1969) extendió la idea de estos
ecólogos, analizando propiedades de los ecosistemas como productividad,
respiración y diversidad. El uso original del término sucesión involucró tres conceptos
fundamentales, la vegetación ocurre en comunidades reconocibles y características,
el cambio en las comunidades en el tiempo ocurre por eventos originados por la
propia biota (sucesión autogénica), y los cambios son lineales y dirigidos hacia un
ecosistema maduro y estable (clímax) (Van der Valk 1985).
En la actualidad este concepto ha continuado evolucionando y
Mitsch y Gosselink (2000); Grime (2001) y Caamal y Armendariz (2002) plantearon
que una sucesión ecológica consiste en el proceso de cambio que sufre un
ecosistema en el tiempo, como consecuencia de los cambios que se producen tanto
en las condiciones del entorno como en las poblaciones que lo integran, dando lugar
a un ecosistema cada vez más organizado y complejo.
I.3.1 Tipos de sucesión vegetal
Existen dos tipos de sucesión vegetal
Sucesión primaria
Sucesión secundaria
Sucesión primaria: Es cuando el proceso de sucesión se inicia de forma natural en
un medio de nueva formación. Este es el caso de la colonización de suelos
volcánicos, lagos naturales de nueva creación, entre otros.

Ejemplo:
Figura 3. Esquema de formación de la sucesión primaria
Fuente: Andrade (2009)
Sucesión secundaria: Es cuando el proceso sucesión se inicia en un medio
previamente ocupado y que ha sido modificado por causas naturales o humanas. Es
el caso de la recuperación natural de un campo de cultivo abandonado, de la
regeneración de un bosque tras un incendio, la recuperación de una laguna
contaminada, entre otros.
Ejemplo:
Figura 4. Esquema de formación de la sucesión secundaria
Fuente: Andrade (2009)
El proceso de sucesión puede durar hasta centenares de años, dependiendo del
ecosistema inicial y de las condiciones en las que se desarrolle (Longman y Jenik
1987; Didham y Lawton 1999).

I.3.2 Características de una sucesión
Cuando se produce una sucesión en condiciones naturales todas siguen unas fases
similares que conducen a cambios parecidos y terminan con comunidades análogas.
Una sucesión presenta varias características:
Aumento del peso y volumen de los organismos del ecosistema o biomasa.
Aumento de la biodiversidad y densidad de organismos.
Aumento de los niveles tróficos del ecosistema y de la complejidad de las
redes tróficas.
Aumenta la resistencia a los cambios que se puedan producir en el
ecosistema.
Fases de una sucesión ecológica:
Fase inicial: Se produce el establecimiento en el suelo de
especies pioneras, organismos de estructura sencilla y
rápida reproducción como líquenes, musgos y algas, que
realizan la fotosíntesis y comienzan a transformar los
componentes del suelo.
Fase media: Se establecen especies vegetales más
complejas, que generan raíces y aportan materia
orgánica al suelo, lo que permite la aparición de gusanos,
artrópodos y otros organismos más complejos.
Fase de maduración: Con el desarrollo del suelo
aparecen arbustos y árboles de mayor porte, que
favorecen la presencia de nuevas especies, tanto
animales como vegetales, completando la red trófica del
ecosistema.

Fase final o clímax: Es el punto de máximo desarrollo
del ecosistema en el que se dan unas condiciones
estables y de equilibrio entre todos sus componentes, con
el máximo de diversidad de especies (Bonet 2004).
Figura 5. Tiempo de desarrollo de las distintas fases en años
Fuente: Andrade (2009)
La desaparición del bosque (figura 5) conduce rápidamente a una asociación abierta
que facilita el paso a la fase de arbustos. La desaparición de los árboles, por ejemplo
de los pinares y encinares, da como resultado la aparición de una serie con
gramíneas, y seguidamente una fase arbustiva en las que dominan los Cistus y
Rosmarinus (Foster 2002).
En la fase arbustiva las condiciones del suelo han tendido a homogeneizarse, con lo
que se facilita el paso a la fase siguiente, constituida por el bosque. La implantación
del bosque se hace según sean las condiciones ambientales de suelo y drenaje
(Guirado y Mendoza 2000).
Por otra parte, en el bosque es preciso distinguir los niveles de población. En la
mayoría de los casos existe un estrato herbáceo generalmente resultado de la acción
del hombre por quema del sotobosque arbustivo, que es el que domina. En los
bosques mediterráneos son frecuentes las especies citadas anteriormente, Cistus y

Rosmarinus entre otras. Por el contrario, en los climas continentales, el estrato bajo lo
constituyen helechos y, en otros casos, erica o brezos. Por este camino hemos
llegado al clímax final, en la que dominan los árboles (Mouquet et al. 2003).
Estas asociaciones de tipo clímax son muy variadas y pueden asimismo tener dentro
de ellas pequeñas microasociaciones o enclaves en puntos en los que, por las
especiales condiciones del suelo o del ambiente, orientación, entre otros aspectos, es
posible la existencia de un microclima, fruto asimismo de una microsucesión. Estos
elementos son importantes no sólo para conocer la exacta tipología de la zona, sino
porque contribuyen a dar variedad y mantener la señal de formas diferenciales entre
clímax (Scheffer y Carpenter 2003).
Una vez alcanzado un tipo de asociación clímax, aunque continúe la competencia, la
comunidad vegetal permanece equilibrada interna y externamente. Sólo un agente de
tipo catastrófico u otro cualquiera de los anteriormente comentados (por ejemplo las
actividades humanas), son capaces de alterar esta situación y dar lugar a la aparición
de una nueva sucesión (Miranda et al. 2004).
Estas especies, que aparecen en esta sucesión que no están identificadas pueden
proporcionar al ganadero fuente de alimentación para sus animales. Sin embargo, el
desconocimiento sobre su empleo continúa incrementando la frontera agrícola,
además de la introducción de nuevas especies, sobre las cuales se desconoce su
rendimiento y calidad en las condiciones de la Provincia Pastaza.
I.4 Caracterización y comportamiento del Pennisetum purpureum en Ecuador
La década de 1950-1960 fue una de las de mayor actividad en el campo de la
sistemática de las Gramíneas, fundamentalmente, porque brindó evidencias acerca
de la filogenia de este grupo de plantas, así como de aspectos de Anatomía,
Entomología, Citología, Histología, Bioquímica y Fisiología (Gold 1968). Según
Gold (1968) y Anon (1997) la taxonomía del Pennisetum purpureum es:

Reino: Plantae
Phylum: Magnoliophyta
Clase: Liliopsida
Orden: Poales
Familia: Poaceae
Subfamilia: Panicoideae
Tribu: Paniceae
Género: Pennisetum
Especie: Pennisetum purpureum
Es un género de plantas anuales o perennes, con hojas comúnmente planas y
panículas densamente espiciformes. Espículas en grupos de 2 ó 3 o solitarias, con un
involucro de cerdas unidas sólo en la misma base y cayendo junto con la espícula;
primera gluma más corta que la espícula, a veces diminuta o ausente; la segunda
gluma igual a la lemna estéril o más corta; lemna fértil lampiña, el margen delgado
encerrando la pálea (Martínez et al. 1996).
Según Herrera (1990) posee hojas anchas y largas que alcanzan de 3,5 y 105cm,
respectivamente y su grueso es de 0,21cm. Su color varía desde el verde claro
(joven) al verde oscuro (maduro). Tiene vellosidades suaves y no muy largas que
pueden alcanzar hasta 122 pelos por campo de observación. El ángulo de inserción
de la hoja con el tallo, medido en la cuarta hoja completamente abierta a partir del
ápice es de 20,5 grados.
En cada planta se pueden encontrar 13 hojas completamente abiertas y a partir de
los 60 días de rebrote comienza la aparición de hojas cloróticas o marchitas (2,4
hojas/planta) en las secciones más próximas al nivel del suelo. Puede alcanzar hasta
4m de altura, pero en condiciones normales de explotación llega a 190cm. Su
crecimiento diario es de 1,07cm. El tallo tiene un diámetro de 1,2cm y su color varía
por las condiciones antes señaladas. Es flexible en estadio joven y rígido cuando

alcanza su madurez. La distancia entre nudos es de 17,2cm y tiene 10 nudos a lo
largo del tallo.
I.4.1 Descripción Botánica
Según Correa et al. (2009), las raíces del cultivar Maralfalfa (Pennisetum purpureum)
son fibrosas y adventicias que surgen de los nudos inferiores de las cañas. Estas
cañas conforman el tallo superficial el cual está compuesto por entrenudos,
delimitados entre sí, por nudos. Los entrenudos en la base del tallo son muy cortos,
mientras que los de la parte superior del tallo son más largos.
Los tallos no poseen vellosidades. Las ramificaciones se producen a partir de los
nudos y surgen siempre a partir de una yema situada entre la vaina y la caña. La
vaina de la hoja surge de un nudo de la caña cubriéndola de manera ceñida. Los
bordes de la vaina están generalmente libres y se traslapan. Es muy común encontrar
bordes pilosos, característica importante en su clasificación (figura 6a). La lígula, que
corresponde al punto de encuentro de la vaina con el limbo, se presenta en corona de
pelos (figura 6b). Mientras que la longitud y el ancho de las hojas pueden variar
ampliamente dentro de una misma planta, la relación entre estas dos medidas parece
ser un parámetro menos variable y muy útil al momento de clasificar las gramíneas
(Häfliger y Scholz 1980). La presencia de pelos en el borde de las hojas, es otro
elemento fundamental en la descripción de esta especie (figura 6c).
Figura 6. Morfología de las hojas del Maralfalfa (Pennisetum purpureum)
Fuente: Andrade (2009)

Los órganos reproductivos consisten en espiguillas típicas del género Pennisetum,
presenta seis brácteas: dos glumas, dos lemas y dos paleas (figura 7). Para su
clasificación a partir de las estructuras podemos señalar las siguientes: las flores
bajas pueden ser estériles y vigorosas o sin estambres, las superiores pueden ser
fértiles, con un tamaño entre la mitad o igual al de las inferiores; las primeras glumas
pueden estar fusionadas con callos, sin rodear la base de la espiga y sin aristas; la
lema de la parte superior es suave, sin arista, de color café a amarillo o púrpura,
glabrosa, con márgenes redondeados o planas, sin aristas; la palea de la flores
superiores están presentes. Poseen tres estambres, y las anteras son oscuras o
grises (figura 7) (Dawson y Hatch 2002).
Figura 7. Inflorescencia del pasto Maralfalfa (Pennisetum purpureum)
Fuente: Andrade (2009)
I.4.2 Métodos de propagación, selección del suelo y preparación
Benítez (1980), informó que Pennisetum purpureum puede propagarse con semilla o
material vegetativo. Según Bernal (1984), esta especie no se propaga por vía sexual;
aunque se ha encontrado semilla viable en un porcentaje de 10.

Se ha empleado el método por cañas, que comprende el tallo entero, despuntado y
desprovisto de hojas; se coloca la caña al fondo del surco y se tapa con tierra. En la
parte aérea de cada nudo emitirá nuevas plantas y en la interna, raíces (Flores 1986).
Ayala (1990) y Martínez (2001) afirman que en un criterio generalizado el género
Pennisetum exige de suelos profundos, bien drenados y de fertilidad media a alta
para lograr la mejor respuesta biológica de la planta. No obstante, este puede crecer
en suelos pedregosos y en una gama amplia de ellos, siempre que se le repongan los
nutrientes, y el drenaje superficial e interno sean buenos. Existe el criterio arraigado
que esta especie requiere la preparación convencional del suelo antes de plantar, que
incluye el surcador y tapado con cultivador que implica realizar seis labores al suelo
durante el proceso de establecimiento.
En el Ecuador el cultivar Maralfalfa se adapta a suelos francos y franco-arcillosos con
un pH de 5,6-7 (INIAP 1999). Se comporta bien en suelos con fertilidad media a
alta, alto contenido de materia orgánica y buen drenaje. Ejerce un efecto
recuperador sobre suelos bien drenados, disminuye la densidad aparente e
incrementa la estabilidad estructural, adermás de aumentar su desarrollo radical y
foliar (Ramírez et al. 2006).
Cuando se comparó el método tradicional de siembra con el de mínima y sin
labranza, Padilla et al. (2004) alcanzaron buenos resultados en el Pennisetum
purpureum vc. CUBA CT-115, este se plantó a vuelta de arado en el mes de junio,
previo al pase de grada media. Sin embargo, no resultó beneficioso plantar sin
preparar el suelo ya que se obtuvo el menor rendimiento y el mayor costo de la
tonelada de MS. Señalando como factor a tener en cuenta la profundidad de plantar.
I.4.3 Profundidad de Plantación
Para condiciones de la Amazonía Ecuatoriana se recomiendan profundidades de 3-
10 cm según Correa et al. (2004) y Ramírez et al. (2006), estos coinciden con los
reportado por Ayala (1990) en condiciones cubanas, donde demostró que los

mayores rendimientos de forraje para el primer corte y total, se obtuvieron cuando la
profundidad de tapado fue de 5 a 10 cm y estas diferencias desaparecieron para el
segundo corte. Los resultados encontrados indicaron que la mejor profundidad de
plantación de la hierba elefante fue de 10 cm, ya que se logró mayor germinación,
altura y población y, como consecuencia, mayor producción de biomasa en la fase de
establecimiento. Aunque es necesario tener en cuenta las condiciones
edafoclimáticas.
Esta profundidad de plantación se puede lograr cuando los tallos se depositan en el
fondo del surco de 20 cm de profundidad y se cubren con una capa de suelo de
10 cm mediante métodos manuales o mecanizados. En suelos pesados, debe
tenerse cuidado de no enterrar demasiado los tallos, esto puede afectarse la
germinación.
I.4.4 Densidad y distancia de plantación
Uno de los factores que más encarece el establecimiento de las especies que se
reproducen por semilla vegetativa, es el gran volumen de semillas que se debe cortar,
recoger y transportar, así como el esparcimiento en el campo. Por lo tanto, cualquier
reducción en las dosis de plantación constituye un ahorro de recursos humanos y
materiales (Herrera 2006).
En estudios realizados en la provincia de Azuay; la producción y el manejo del pasto
Maralfalfa fue superior con 23,66kg/m2 de carga forrajera (materia verde) por cada
metro cuadrado, sembrada a 0,50 x 0,50cm (Cunuhay y Choloquinga 2011). En
Colombia se trabaja con distancias de 0,25cm, con el objetivo de lograr el mayor
número de nudos brotados (Tierra pastos y ganado 2012).
Los trabajos realizados por Padilla et al. (2005) así lo indican, pues el costo de la
semilla y la mano de obra representaron el 48 y 22%, respectivamente de los gastos
totales cuando Pennisetum purpureum se plantó por el método tradicional de

preparación convencional del suelo. Sin embargo, si se emplea labranza mínima
(aradura más grada) y se planta a vuelta de arado, los gastos de semilla se pueden
elevar al 53% y los de mano de obra 22, debido a que se reducen los gastos de
maquinaria y combustible aunque se mantienen los de mano de obra y semilla.
Lo anterior evidencia que esta especie debe ser plantada entre 100 y 120cm entre
surcos, siempre que se garantice la abundante y distribuida germinación a todo lo
largo de los surcos, pues con esto el ahijamiento que produce esta planta asegura las
poblaciones necesarias para obtener altos rendimientos de buena calidad, además de
facilitar las labores de cultivo.
I.4.5 Rendimiento y calidad nutritiva del Pennisetum purpureum vc. Maralfalfa
En Ecuador se registran diferentes investigaciones realizadas en Pennisetum
purpureum vc. Maralfalfa donde reportan determinados indicadores: en la provincia
de Chimborazo, Alzamora (2011) obtuvo una producción promedio de forraje verde a
los 75 días de edad de 64,68 tMV/ha-1, mientras que Andrade (2009) alcanzó una
producción de 127 335 Kg/ha-1.
Mientras que Cárdenas et al. (2012) encontraron producción de materia seca de 8,11;
12,09 y 20,20 t/ha-1, respectivamente para las hojas, tallos y rendimiento total, de ahí
que los valores de producción de MS obtenidos muestran que el pasto Maralfalfa
presentó un incremento lineal, lo que significa que conforme se incrementa la edad de
la planta, existe un incremento en los carbohidratos estructurales (celulosa y
hemicelulosa) y compuestos fenólicos (lignina) influenciando a la producción de MS.
Sin embargo, pueden mediar otras causas como: la disponibilidad de agua, desarrollo
del sistema radicular de la planta y época del año; éstos pueden producir cambios
morfológicos como la disminución de láminas foliares y el aumento de los haces
basculares.

Otros trabajos, reportados por Guamanquishpe (2012) en la provincia de Tungurahua
encontraron 12 890 t/ha-1 de materia verde en el cantón Paute, provincia de Azuay,
Choloquinga y Cunuhay (2011) reportaron 23,66kg/m2 de materia verde. En estas
condiciones la variedad Maralfalfa mostró porcentajes de PB, FB, EE y ELN de 13,5;
30, 40; 2,54 y 40,99%, respectivamente. Mientras Betancur (2004), Carulla et al.
(2004) y Osorio (2004), reportaron valores de 26; 11,4; 68,3; 46,6; 1,9 y 12,02% de
MS, PB, FND, FAD, EE y Ceniza, respectivamente.
Por su parte, Cárdenas et al. (2012), reportaron en las hojas valores de PB, MO,
materia inorgánica, FND,FAD y DMS, en este de cultivar de 12,03; 92,29; 7,69; 63,15;
27,48 y 64,2%, respectivamente. Mientras que en el tallo fueron de 5,90; 93,42; 6,57;
65,66; 34,94 y 53,44%.
Clavero y Razz (2009) al evaluar la calidad nutritiva del pasto Maralfalfa en
condiciones de defoliación encontraron que esta declinó en un período de tres a
nueve semanas de crecimiento. Para cada intervalo los contenidos de nitrógeno total
disminuyeron significativamente, mientras que, la relación nitrógeno soluble y total
disminuyó lentamente de tres a nueve semanas; sin embargo, en todos los
tratamientos hallaron que el 50% del N es soluble, de fácil asimilación. El contenido
de pared celular y lignina incrementaron entre la sexta y la novena. Los mayores
valores de DIVMS (62,45%) fueron obtenidos con tres semanas de crecimiento y
declinó en 10,35 unidades de digestibilidad entre la tercera y novena semana. Las
concentraciones de carbohidratos no estructurales aumentaron ligeramente a medida
que se incrementó el intervalo de corte. Los valores estuvieron en un rango desde
13,5 hasta 20,1% para la tercera y novena semana, respectivamente.
Este estudio indicó que la calidad del pasto Maralfalfa es afectada negativamente a
medida que avanza la madurez de la planta lo cual puede ser debido a incrementos
en la acumulación de material muerto en el perfil de la planta y la lignificación de las
paredes celulares.

Es necesario señalar que tanto los indicadores del rendimiento, como los de calidad
varían en dependencia del suelo, las características climáticas de la región, entre
otros aspectos.
I.5 Factores que afectan el crecimiento y calidad
Los pastos cubren la cuarta parte de la superficie del planeta, ayudan a mitigar el
calentamiento global y evitan la erosión de los suelos, sin embargo los pastizales son
un ecosistema seriamente amenazado por la agricultura, la ganadería y la actividad
forestal. Aun cuando sus servicios ambientales equivalen a unos 900.000 mil millones
de dólares al año, apenas el 0,7% del total mundial tiene algún grado de protección y
en América del Sur sólo alcanza 0,3% (D´Atri 2007).
Con sus 39 millones de kilómetros cuadrados en el mundo (casi una cuarta parte de
la superficie terrestre continental), los pastizales representan uno de los ecosistemas
más extensos del planeta. Distribuidos en casi todos los continentes, se destacan por
su importancia: el sistema de praderas de las Grandes Planicies de América del
Norte, las estepas del este de Europa (pusztas) y de Mongolia, los grassvelds de
Sudáfrica, las planicies de Nueva Zelanda, y las pampas y campos de Argentina,
Uruguay y Brasil (D’Atri 2007).
Los pastos poseen características fisiológicas y morfológicas propias que le brindan
adaptación específica para su crecimiento y calidad. Sin embargo, estos
experimentan modificaciones morfológicas en su rendimiento y calidad cuando
ocurren cambios en las condiciones climáticas, donde la temperatura, la radiación
solar (cantidad y calidad), las precipitaciones y su distribución son los componentes
que más determinan en las condiciones tropicales (Del Pozo 2004).
I.5.1 Temperatura
Los procesos bioquímicos y fisiológicos básicos relacionados con la síntesis,
transporte y degradación de sustancias en las plantas están influenciados por la

temperatura, por el grado de relación que éstas poseen con la cinética de las
reacciones bioquímicas y el mantenimiento de la integridad de las membranas.
No todas las especies de pastos tienen el mismo valor óptimo de temperatura para el
cumplimiento de estas funciones. Así, Baruch y Fisher (1991) informaron que en las
gramíneas tropicales, el óptimo fotosintético se encuentra entre los 35 y 39ºC, y en
las leguminosas entre los 30 y 35ºC, con una alta sensibilidad a las bajas
temperaturas, cuyos efectos negativos en el crecimiento ocurren entre los 0 y 15ºC y
en algunas especies a los 20ºC, si la humedad no es un factor limitante, lo cual está
dado por la baja conversión de azúcares en los tejidos de las plantas, producto de
una disminución en los procesos de biosíntesis y por un déficit energético acarreado
por una reducción en la tasa respiratoria.
Además, se informó por estos autores que cuando las frecuencias de temperaturas
por debajo de los 15ºC se incrementan durante el período de crecimiento, los
asimilatos formados se acumulan gradualmente en los cloroplastos y pueden
afectar la tasa de asimilación y translocación de metabolitos, hasta provocar daños
físicos en el aparato fotosintético que limitará el crecimiento de los pastizales (Baruch
y Fisher 1991).
Uno de los mecanismos estructurales utilizados por los pastos para reducir los
efectos de estrés por altas temperaturas, es el aumento del contenido de la pared
celular, fundamentalmente en lignina, la cual reduce de forma muy marcada la
digestibilidad y la calidad de éstos. Wilson et al. (1991), al estudiar el efecto del
aumento de la temperatura en el contenido de componentes estructurales y la
digestibilidad en las hojas y tallos de C. dactylon, M. maximus vc. Trichoglume, P.
laxacum, L. perenne y M. sativa, encontraron que la reducción en la digestibilidad de
la materia orgánica y la pared celular estaba asociada a un aumento en la
lignificación de sus tejidos. Aunque, Nelson y Moser (1994) argumentaron que
pudieran existir otros mecanismos que influyan en este proceso, pero aún la
información es limitada.

En los trabajos desarrollados con otras especies de pastos, se encontró que el
rendimiento de la D. decumbens en los meses más fríos del año en Cuba (enero -
febrero) osciló entre los 5 y 7 kg MS/ha-1/días en condiciones de secano y sin
fertilización, pero cuando se aplicaron fertilizantes e irrigación, los rendimientos no
sobrepasaron los 24 kg MS/ha-1/día. En el resto de los meses del año, el crecimiento
y producción de los pastos muestran un comportamiento diferente, donde la
temperatura no es un elemento limitante. Esto se corrobora en los trabajos de
Ayala et al. (1989), en el crecimiento del P. purpureum. Este comportamiento podría
ser diferente en la provincia Pastaza, donde las condiciones del clima son muy
similares en todo el año.
I.5.2 Radiación Solar
La radiación solar es un elemento climático que se encuentra estrechamente
relacionado con procesos fisiológicos fundamentales, vinculados con el crecimiento y
los cambios morfológicos que experimentan los pastos y forrajes a través de su
desarrollo. La conversión de energía solar en biomasa en los pastos es variable y
depende de las vías metabólicas a través de las cuales efectúa la fotosíntesis, por lo
que la tasa fotosintética de los pastos es una función de la energía disponible. Las
plantas C3 sometidas a altas intensidades de radiación, pueden afectar la tasa de
asimilación neta y alcanzan el estado de saturación lumínica en sus hojas a valores
de 300-400 J/m2/seg bajo condiciones controladas (Del Pozo 2004).
Las plantas C4, fotosintetizan más por unidad de radiación absorbida y alcanzan la
saturación lumínica a niveles superiores, con eficiencias de hasta 6% (Nelson y
Moser 1994). Sin embargo, en condiciones normales de producción, las hojas
superiores reducen los niveles de radiación al resto del follaje, y sólo una parte de
ellas alcanza su potencial fotosintético, lo que hace que bajo estas condiciones se
logre la saturación lumínica a intensidades superiores.

Por otra parte, la arquitectura del césped también modifica la calidad de la luz en su
interior, provoca variaciones en las respuestas morfológicas a través de su ciclo de
crecimiento, lo cual se manifiesta de forma diferente en dependencia de la especie de
planta, cuyos rasgos fundamentales son: tasa de aparición, elongación y el tamaño
final de los órganos, tasa de senescencia y altura de la planta, entre otras (Sinoquet
y Cruz 1995).
La radiación solar también ejerce su influencia en otros procesos metabólicos de la
planta que determinan en su composición química, ya sea por cambios en la
intensidad como en la calidad de la luz. El aumento en la intensidad de la luz,
favorece los procesos de síntesis y acumulación de carbohidratos solubles en la
planta, mostrando un comportamiento inverso con el resto de los constituyentes
solubles y estructurales, siempre que otros factores no sean limitantes. Buxton y
Fales (1994) señalaron que en las regiones tropicales la reducción de la intensidad de
radiación por el efecto de la sombra cambia la composición química de los forrajes y,
en especial, sus componentes celulares, aunque las respuestas son variables según
la combinación de especies.
Wilson (1993) encontró un aumento en el contenido y rendimiento de nitrógeno en
Megathyrsus y Chloris de 17 y 36%, respectivamente, mientras en Cenchrus,
disminuyó en cuatro cuando los niveles de radiación disminuyeron en 50. En
Cynodon nlemfuensis, la reducción de la radiación mejoró en 3,7 y 2,32 unidades
porcentuales la proteína bruta y el contenido de cenizas, disminuyendo en 4,53 los
componentes estructurales (Del Pozo y Jeréz 1999).
Pezo e Ibrahim (1999) argumentaron que las variaciones producidas en la calidad de
los pastos debido a la radiación, están más relacionadas con los cambios anatómicos
y morfológicos, que por el incremento o disminución de algunos de sus constituyentes
químicos, y que otros factores como las precipitaciones intervienen de forma directa.

I.5.3 Precipitaciones
El volumen de agua caída por las precipitaciones y su distribución a través del año
ejercen efectos notables en el crecimiento y calidad de los pastos, debido a su
estrecha relación con los factores bioquímicos y fisiológicos que regulan estos
procesos biológicos de gran complejidad. El agua es un componente esencial en las
células de las plantas, casi todos los procesos metabólicos dependen de su
presencia; además, se requiere para el mantenimiento de la presión de turgencia, la
difusión de solutos en las células y suministra el hidrógeno y oxígeno que están
involucrados durante el proceso fotosintético (Lösch 1995).
Tanto el exceso como el déficit de precipitaciones pueden provocar estrés en los
cultivos forrajeros. En el caso del primero, generalmente ocurre en los suelos mal
drenados durante la estación lluviosa o en las regiones donde las precipitaciones son
altas durante todo el año, como es el caso de la Amazonía Ecuatoriana. Su efecto
fundamental radica en que causa anoxia en las raíces, afectando su respiración
aeróbica, absorción de minerales y agua. Si este se prolonga en especies no
tolerantes, disminuye la asimilación y translocación del carbono, produciéndose
cambios metabólicos que activan la respiración anaeróbica, lo cual implica una menor
eficiencia energética y bioproductividad en las plantas (Baruch, 1994a).
Por otra parte, modifica la distribución y producción de la biomasa, la tasa de
crecimiento y concentración de minerales en las plantas, aspecto que fue demostrado
por Baruch (1994b) en H. rufa, A. gayanus vc CIAT 621, B. mutica y E. polystachya,
en condiciones de inundación durante 20 a 25 días.
Sin embargo, el estrés por sequía es más común en las regiones tropicales, el cual
afecta el comportamiento fisiológico y morfológico de las plantas. El efecto depende
de su intensidad y el estado de crecimiento y desarrollo de la planta. Entre los efectos
más sensibles se destacan: la reducción de la expansión celular motivada por una
disminución en la presión de turgencia, cierre estomático, la transpiración y, por ende,

la fotosíntesis, aunque en este último con efectos directos en los procesos
enzimáticos y transporte electrónico (Antolín y Sánchez-Díaz 1993), contenido de
clorofila y la estructura de las membranas, las cuales afectan la respiración. El déficit
hídrico modifica la partición de la biomasa entre la parte aérea y radicular en las
plantas (Baruch 1994b), disminuye el área foliar y su duración, al tiempo que
incrementa la senescencia y, como resultado, la abscisión de las hojas.
Por otra parte, bajo estas condiciones de estrés se producen importantes cambios
metabólicos que en muchas especies se consideran como adaptativos, los cuales se
caracterizan por una disminución de la síntesis de proteínas, aumento en la
concentración de aminoácidos libres, especialmente prolina, glicina, betaína,
dipoliaminas y una disminución en la velocidad de síntesis del RNA (Lösch 1995).
Estos cambios metabólicos, generalmente, tienen pocos efectos sobre la calidad de
las plantas y los efectos beneficiosos que se señalan están relacionados con el
proceso de crecimiento. En este sentido, podemos plantear que el aumento en la
calidad de los pastos debido al estrés hídrico está asociado a cambios morfológicos
en las plantas, tales como: reducción en el crecimiento de los tallos y aumento en la
proporción de hojas, elementos característicos en el retraso de la madurez de las
plantas.
El efecto de las precipitaciones en el comportamiento de estos procesos
morfológicos, bioquímicos y fisiológicos relacionados con el crecimiento y la calidad
de los pastos, depende de múltiples factores que están asociados al ambiente, al
suelo y la especie de planta. En este sentido, se ha señalado en la literatura que el
crecimiento de los pastos es una función de la humedad disponible en el suelo y que
éste, a su vez, varía en dependencia de la cantidad y distribución de las
precipitaciones, de la estructura y pendiente de los suelos, de los valores de radiación
y temperatura, así como del área cubierta por la vegetación (Del Pozo 2002).

El comportamiento de los factores del clima en la Provincia Pastaza, se puede decir
que es bastante estable, por lo tanto analizar otros factores sería de gran importancia,
ya que estos influyen en el rendimiento y calidad de los pastos, estos son nombrados
por muchos como factores del manejo.
I.5.4 Frecuencia y altura de corte o pastoreo
El crecimiento y la calidad de los pastos pueden variar considerablemente de acuerdo
con el manejo a que son sometidos, con efectos favorables o no en dependencia de
la especie de planta y las condiciones edafoclimáticas donde se desarrollan. Se
destacan entre ellos la altura de corte o pastoreo, la carga animal y el tiempo de
estancia, entre otros.
La altura y el momento de la cosecha constituyen elementos básicos en el manejo de
los pastos y forrajes, por la influencia que estos ejercen en su comportamiento
morfofisiológico y productivo. Numerosos experimentos se han realizado donde se
estudia la edad y altura de corte o pastoreo con el propósito de profundizar en los
diferentes mecanismos relacionados con la defoliación y el rebrote, así como en sus
respuestas.
Bajo condiciones de corte, Ramos et al. (1987), en un estudio dinámico del
crecimiento de Cynodon nlemfuensis con tres niveles de nitrógeno (0, 200 y 400 kg
N/ha/año) y tres edades de corte, encontraron que, en la medida que aumentó la
edad de corte, la altura y la acumulación de masa seca, aumentaron de forma
significativa, recomendándose cortar entre la quinta y la sexta semana con dosis de
200 a 400 kgN/ha/año, ya que permitió alcanzar los máximos valores de densidad y
rendimiento.
Tergar et al. (1988) evaluaron tres frecuencias de cortes (35, 45 y 55 días) en el
rendimiento de C. dactylon coast cross-1, C. nlemfuensis, D. pentzzi y M. maximus en
un suelo oscuro plástico gleyzado en Puerto Rico; informaron que a los 35 días de
edad sólo C. dactylon coastcross-1 y D. pentzzi expresaron el máximo crecimiento

con valores de hasta 62,0 Kg MS/ha-1/día, mientras que el resto lo hicieron a los 45
días con 63, 64,5 y 47,6 KgMS/ha-1/día para C. nlemfuensis, M. maximus y C.
plectostachyus, respectivamente.
En condiciones de pastoreo la dinámica de crecimiento no sólo depende de las
variaciones del clima y el suministro de nutrientes, sino de la acción de los animales
en el pastoreo, cuyas interacciones son numerosas y complejas, con respuestas
morfológicas y fisiológicas variables, en dependencia del hábito de crecimiento,
mecanismos de propagación y persistencia, y del sistema de manejo empleado en su
explotación.
En sistemas rotacionales, Blanco (1995), en el estudio del crecimiento de
Megathyrsus maximus y Andropogon gayanus, encontró similares respuestas en la
acumulación de materia seca según la edad de rebrote, pero con diferencias en su
cinética de crecimiento, así como en las relaciones con sus componentes
morfológicos. El porcentaje de hojas, el perímetro y diámetro de las macollas fueron
las variables que más se correlacionaron.
Teniendo en cuenta los aspectos antes discutidos, es importante buscar un adecuado
balance entre el rendimiento, la composición química y el contenido de reservas en
las partes bajas y subterráneas de los pastos, que permitan una máxima persistencia
y utilización. La altura de corte o pastoreo es un elemento determinante en la
dinámica de crecimiento de los pastos, por su estrecha relación con la remoción de
los puntos de crecimiento que ocurren durante la cosecha y el balance de
carbohidratos de reservas.
La morfología y el hábito de crecimiento de las especies tienen una gran influencia en
la interrelación entre la defoliación, el IAF residual y la capacidad de intercepción de
la luz, con respuestas diferentes en cada especie de acuerdo con el manejo
impuesto.

En un estudio efectuado por Páez et al. (1995), donde evaluaron diferentes
frecuencias (15, 30, 45 y 60 días) y alturas de cortes (20, 40 y 60 cm) en el
crecimiento y distribución de la biomasa aérea de Megathyrsus maximus, se señaló
que las alturas de 40 y 60 cm proporcionaron una mayor fracción residual de hojas y
por ende un área fotosintéticamente activa y menor movilización de fotoasimilatos
desde las raíces. En especies de hábito de crecimiento rastreras se han reportado
comportamientos similares. No obstante, Mislevy y Pate (1996) y del Pozo (1998)
señalaron que éstas pueden ser cosechadas a bajas alturas y de forma frecuente, sin
que afecte su dinámica de crecimiento, productividad y persistencia, debido a las
características que presenta su sistema asimilativo y la mayor cantidad de puntos de
crecimiento no afectados por el corte o los animales en el pastoreo.
Es importante señalar que los efectos de la altura de corte o pastoreo en el
crecimiento de los pastos son más severos, tanto a corto o a largo plazo, cuando se
realizan muy cerca de la superficie del suelo y de manera frecuente. Aunque es
necesario destacar que en ellos tiene una marcada influencia la carga y tiempo de
estancia.
I.5.5 Carga y tiempo de estancia
En los sistemas de producción basados en pastos, la carga animal es la variable más
importante en el manejo que determina la productividad por animal y por área
(Jones y Sandland 1974). Su efecto fundamental es a través de los cambios que se
producen en la disponibilidad y el consumo de los pastos con influencias marcadas
en la estructura y composición química de la planta, lo cual repercute directamente en
su fisiología.
Fales et al. (1995) encontraron en Dactylis glomerata (L) y Poa pratensis (L)
manejados bajo un sistema rotacional con tres intensidades de pastoreo (35, 49 y 53
vacas días/ha) un efecto positivo en la tasa de crecimiento y acumulación de masa
seca neta y en la calidad, asociado esto último a una reducción en la cantidad de

material senescente y área rechazada por los animales en el pastoreo, cuando se
aumenta la intensidad de este.
En especies tropicales, Turner y Seastedt (1993) hallaron similares respuestas y
destacaron el papel negativo que desempeña una alta frecuencia de pastoreo cuando
los niveles de intensidad son altos, principalmente en la acumulación de las reservas
y en la biomasa aérea de las plantas. Por otra parte, informaron la capacidad que
poseen estas especies para compensar los efectos de la alta intensidad de pastoreo
y señalaron el aumento de la tasa de expansión foliar, índice de área foliar y
asimilación neta como respuesta a la alta intensidad de defoliación; situación similar
encontraron Chacón-Moreno et al. (1995) en Megathyrsus maximus.
Maraschin (1996) señaló que los efectos de la defoliación en la fotosíntesis, fijación
de carbono y translocación y empleo de los carbohidratos de reserva son de carácter
transitorio, cuya duración en Cynodon dactylon puede alcanzar hasta siete días. En
Cynodon nlemfuensis, Del Pozo (1998) encontró que la dinámica de utilización de las
reservas de carbono y su capacidad fotosintética en la planta varió en dependencia
del grado de intensidad de pastoreo con que fue manejado el pastizal.
Por otra parte, debemos señalar que las respuestas productivas en los pastizales por
el aumento de la intensidad de pastoreo no son proporcionales con el nivel de oferta
a los animales, de ahí que dicha técnica deba manejarse de forma tal que no
comprometa el equilibrio suelo-planta-animal. Además, la influencia del clima, así
como la respuesta de especies de reciente introducción a un ecosistemas
determinado, son factores que se debe analizar detenidamente, para dar respuesta
adecuadas con principios de sostenibilidad al manejo de las áreas forrajeras, y evitar
en muchos zonas el incremento de la frontera agrícola. De ahí la importancia de
conocer las especies.

CAPÍTULO II
INVENTARIO Y DESCRIPCIÓN DE LA SUCESIÓN VEGETAL SECUNDARIA EN
LA PROVINCIA PASTAZA
II. 1 Introducción
En pocas décadas, la diversidad biológica ha sido reconocida a nivel nacional e
internacional como un elemento fundamental para el desarrollo de planes de
conservación y el uso sustentable de los recursos naturales. Por lo tanto, su
conocimiento, cuantificación y análisis es fundamental para entender el mundo
natural y los cambios inducidos por la actividad humana (Villareal et al. 2006).
Se estima que alrededor del 25% de la diversidad biológica a nivel mundial se
encuentra en la región andina; los países que comprenden esta región son
considerados como los más diversos y ricos en especies animales y vegetales del
mundo (Mittermeir et al. 1997; Myers et al. 2000). La gran variabilidad ecológica de
los países andinos se debe a la ubicación intertropical, combinada con la presencia
de la Cordillera de los Andes que define un gradiente altitudinal y constituye una
barrera importante que cruza todos los países, y los divide en ámbitos ecológicos de
gran variedad. La variación del clima y la topografía, combinada con factores edáficos
y diferencias en historia geológica, crean una serie de hábitats, cada uno con
elementos florísticos distintos.
Los cambios provocados por la actividad humana en el clima y la cubierta vegetal han
provocado modificaciones importantes en la composición de especies y la diversidad
de los ecosistemas terrestres (IPCC 2002; Tilman y Lehman 2001; Chapin et al.
2000), dando como resultado la formación de nuevos ecosistemas (Hobbs 2006).
Muchas de estas consecuencias pueden ser predecibles. Por suerte, actualmente
disponemos de más y mejor información ambiental que hace unos años, así como de
nuevos métodos estadísticos y de herramientas integradas en los sistemas de

información geográfica (SIG) que permiten realizar predicciones de mayor fiabilidad y
reproducir la distribución de los principales atributos que representan la diversidad
biológica de un territorio. Si el estudio de los cambios en riqueza de biodiversidad es
importante en cualquier lugar del mundo para elaborar estrategias de conservación y
restauración, se torna crítico en las áreas tropicales en las que se encuentran los
puntos calientes de biodiversidad (Myers et al. 2000).
A pesar de las múltiples facetas del concepto, la diversidad biológica puede ser
entendida simplemente como el número de especies presentes en un sitio o región.
Esta aparente simplificación tiene ventajas para la planeación y el desarrollo de
programas de inventarios de biodiversidad, los cuales deben estar enfocados a
responder cuanta diversidad existe, dónde y cómo se distribuye.
Los mecanismos que regulan la biodiversidad a nivel espacial y temporal, pueden
comprenderse a través de estudios comparativos, para lo cual los muestreos dentro
de un inventario deben realizarse con rigor metodológico y deben ser comparables
(Huston 1994). Es importante resaltar que los métodos aplicados para llevar a cabo
inventarios, es decir, las técnicas de muestreo, deben seleccionarse cuidadosamente
y reconocer sus limitaciones para obtener información representativa.
Por ello, el objetivo del presente capítulo es inventariar las especies que conforman
sucesión vegetal secundaria en áreas deterioradas de una zona de la Amazonía
Ecuatoriana; así como describir las especies utilizadas en la alimentación animal y
otros fines.
II.2 Materiales y Métodos
III. 2.1 Área de Investigación
El trabajo se desarrolló en el Centro de Investigación Posgrado y Conservación
Amazónica (CIPCA) de la Universidad Estatal Amazónica, ubicado en el km. 44 vía
Puyo - Tena, Cantón Carlos Julio Arosemena Tola, Provincia de Napo, ubicada en
las siguientes coordenadas: 01 ° 14ʹ 4,105 ʹ ʹ Latitud Sur y 77 ° 53 ʹ 4,27 ʹ ʹ Longitud
Oeste, a una altura de 584 msnm.
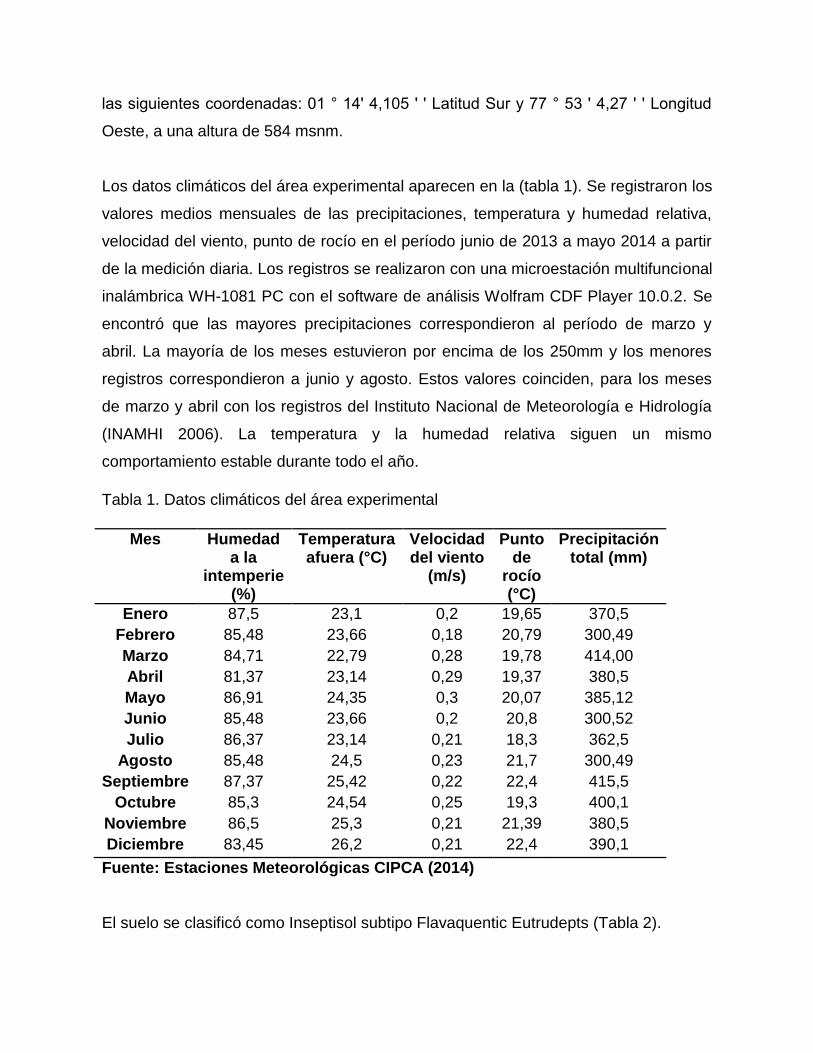
Los datos climáticos del área experimental aparecen en la (tabla 1). Se registraron los
valores medios mensuales de las precipitaciones, temperatura y humedad relativa,
velocidad del viento, punto de rocío en el período junio de 2013 a mayo 2014 a partir
de la medición diaria. Los registros se realizaron con una microestación multifuncional
inalámbrica WH-1081 PC con el software de análisis Wolfram CDF Player 10.0.2. Se
encontró que las mayores precipitaciones correspondieron al período de marzo y
abril. La mayoría de los meses estuvieron por encima de los 250mm y los menores
registros correspondieron a junio y agosto. Estos valores coinciden, para los meses
de marzo y abril con los registros del Instituto Nacional de Meteorología e Hidrología
(INAMHI 2006). La temperatura y la humedad relativa siguen un mismo
comportamiento estable durante todo el año.
Tabla 1. Datos climáticos del área experimental
Mes Humedad a la
intemperie (%)
Temperatura afuera (°C)
Velocidad del viento
(m/s)
Punto de
rocío (°C)
Precipitación total (mm)
Enero 87,5 23,1 0,2 19,65 370,5
Febrero 85,48 23,66 0,18 20,79 300,49
Marzo 84,71 22,79 0,28 19,78 414,00
Abril 81,37 23,14 0,29 19,37 380,5
Mayo 86,91 24,35 0,3 20,07 385,12
Junio 85,48 23,66 0,2 20,8 300,52
Julio 86,37 23,14 0,21 18,3 362,5
Agosto 85,48 24,5 0,23 21,7 300,49
Septiembre 87,37 25,42 0,22 22,4 415,5
Octubre 85,3 24,54 0,25 19,3 400,1
Noviembre 86,5 25,3 0,21 21,39 380,5
Diciembre 83,45 26,2 0,21 22,4 390,1
Fuente: Estaciones Meteorológicas CIPCA (2014)
El suelo se clasificó como Inseptisol subtipo Flavaquentic Eutrudepts (Tabla 2).

Tabla 2. Composición química del Suelo
Fuente: Laboratorio Universidad Técnica de Ambato (2013)
II. 2. 2 Procedimiento
Se levantaron parcelas de 20 x 20 m (400 m2), las cuales fueron georreferenciadas
con la utilización de un GPS RTK precisión de 1 cm, doble frecuencia, 72 canales,
GPS + GLONASS, receptor-100 Hz, teléfono celular integrado, 806 MHz X Scale,
pantalla táctil; cámara integrada, brújula; Windows Mobile 6.1 del sistema operativo.
Las unidades de muestreo se distribuyeron en tres sectores del Centro de
Investigaciones y Posgrado de la Conservación de la Biodiversidad Amazónica a lo
largo de un gradiente altitudinal, comprendido entre los 600 y 900 msnm, que
corresponden al piso altitudinal premontano de la región Andina, estribación oriental
(Aguirre y Yaguana 2012). Se utilizó un muestreo aleatorio simple y se siguió la
metodología de Keels et al. (1997).
ANALISIS Valor
pH extracto suelo:agua 1:2,5 5,50 C.E. extracto suelo:agua 1:2,5(us/cm) 51,6
Textura Franco Arcilloso
Arena, % 44
Limo, % 26
Arcilla, % 30
M.O, % 6,8
N – TOTAL, % 1,3
P, ppm 2,5
K, meq/100 g 0,6
Ca, meq/100 g 7,0
Mg, meq/100 g 6,3
Cu, ppm 6,4
Fe, ppm 94,0
Mn, ppm 3,7
Zn, ppm 1,8
Ca/Mg, meq/100 g 1,1
Mg/K, meq/100 g 10,0
Ca+Mg/K, meq/100 g 21,3

II.2.3 Identificación de las Especies
Para la determinación de las especies resultantes de la sucesión vegetal secundaria
en un área deteriorada, se procedió con la identificación botánica preliminar en el
campo y se confirmó con la literatura apropiada: Manual de Botánica Sistemática,
Etnobotánica y Métodos de Estudio en el Ecuador (Cerón, 1993), Catálogo de Plantas
Vasculares del Ecuador (Jorgensen y León 1999; Neill y Ulloa 2011), Libro Rojo de
las Plantas Endémicas del Ecuador (Valencia et al. 2000) y el sistema de clasificación
propuesta por Angiosperm Phylogeny Group (APG 2003), empleado para la
clasificación de las angiospermas. Además, fueron constatadas en la colección de
muestras del Herbario Ecuatoriano Amazónico (ECUAMZ).
Para las anotaciones, se llevó al campo un cuaderno y lápiz, para registrar la
localidad, altitud, hábitat, fecha entre las más importantes.
II. 3 Resultados
II.3.1 Composición florística
En el entorno que corresponde al bosque húmedo tropical la flora registrada incluyó
un total de 57 familias, 123 géneros y 148 especies, las familias con mayor riqueza de
especies fueron Fabaceae, Moraceae y Myristicaceae con 19, 17 y 10 taxones
respectivamente, que representaron un 79,3 % del total. Por otra parte, 33 familias
para el 56,9 % estuvieron representadas solo por una especie y el resto, 24 familias
con dos o más integrantes, representaron el 40,1 % (Figura 8).
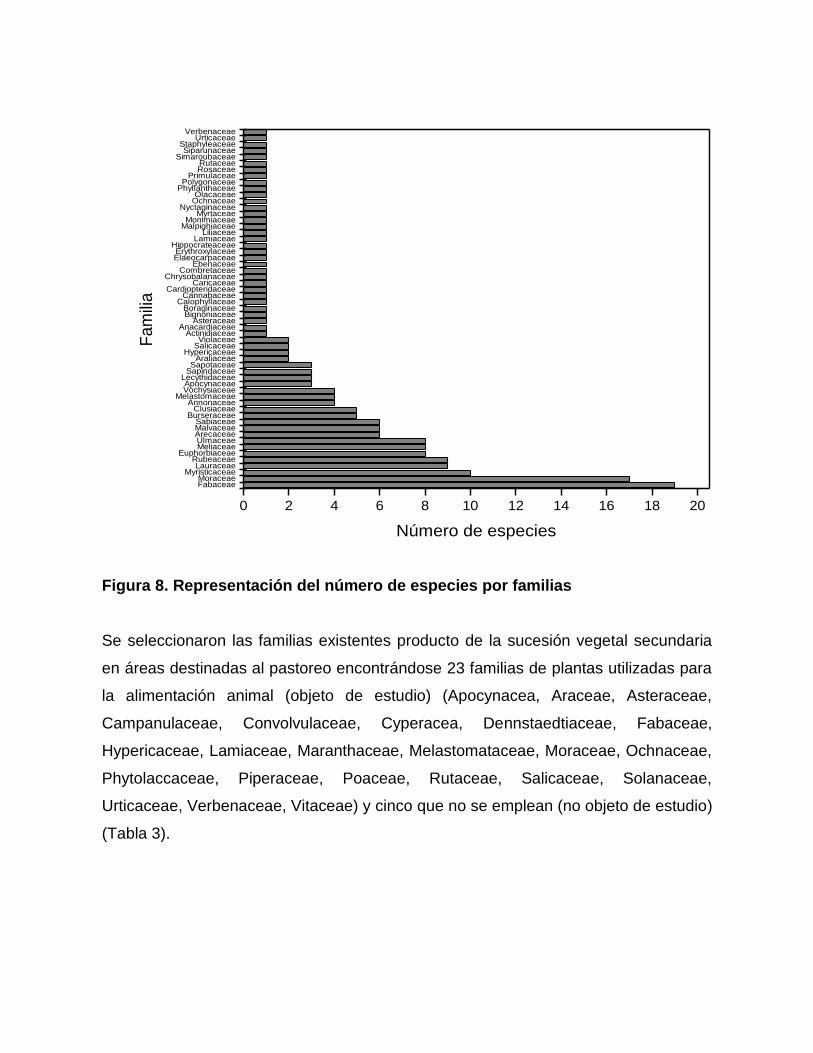
FabaceaeMoraceae
MyristicaceaeLauraceae
RubeaceaeEuphorbiaceae
MeliaceaeUlmaceaeArecaceaeMalvaceaeSabiaceae
BurseraceaeClusiaceae
AnnonaceaeMelastomaceae
VochysiaceaeApocynaceae
LecythidaceaeSapindaceaeSapotaceae
AraliaceaeHypericaceae
SalicaceaeViolaceae
ActinidiaceaeAnacardiaceae
AsteraceaeBignoniaceaeBoraginaceae
CalophyllaceaeCannabaceae
CardiopteridaceaeCaricaceae
ChrysobalanaceaeCombretaceae
EbenaceaeElaeocarpaceaeErythroxylaceae
HippocrateaceaeLamiaceae
LiliaceaeMalpighiaceaeMonimiaceae
MyrtaceaeNyctaginaceae
OchnaceaeOlacaceae
PhyllanthaceaePolygonaceae
PrimulaceaeRosaceaeRutaceae
SimaroubaceaeSiparunaceae
StaphyleaceaeUrticaceae
Verbenaceae
0 2 4 6 8 10 12 14 16 18 20
Número de especies
Fa
mili
a
Figura 8. Representación del número de especies por familias
Se seleccionaron las familias existentes producto de la sucesión vegetal secundaria
en áreas destinadas al pastoreo encontrándose 23 familias de plantas utilizadas para
la alimentación animal (objeto de estudio) (Apocynacea, Araceae, Asteraceae,
Campanulaceae, Convolvulaceae, Cyperacea, Dennstaedtiaceae, Fabaceae,
Hypericaceae, Lamiaceae, Maranthaceae, Melastomataceae, Moraceae, Ochnaceae,
Phytolaccaceae, Piperaceae, Poaceae, Rutaceae, Salicaceae, Solanaceae,
Urticaceae, Verbenaceae, Vitaceae) y cinco que no se emplean (no objeto de estudio)
(Tabla 3).

Tabla 3. Especies identificadas en el área destinada al estudio de la sucesión
vegetal secundaria en suelos deteriorados
II.3.3 Descripción de especies
FAMILIA APOCYNACEAE
Matelea sp
Las Apocynaceae son una familia de las dicotiledóneas que incluye árboles, arbustos,
hierbas, o lianas. Muchas especies son grandes árboles que se encuentran en la
N° FAMILIA NOMBRE CIENTIFICO CATEGORIAS
1 Apocynaceae Matelea sp. Plantas no objeto de estudio
2 Araceae Antthurium sp. Plantas objeto de estudio
3 Asteraceae Veronia sp. Plantas objeto de estudio
4 Campanulaceae Hippobroma longiflora Plantas no objeto de estudio
5 Convolvulaceae Ipomoea batatas Plantas objeto de estudio
6 Convolvulaceae Ipomoea sp. Plantas objeto de estudio
7 Cyperaceae Cyperus ustulatus Plantas objeto de estudio
8 Cyperaceae Scleria sp. Plantas objeto de estudio
9 Dennstaedtiaceae Hypolepis sp. Plantas no objeto de estudio
10 Fabaceae Desmodium heterophyllum Plantas objeto de estudio
11 Hypericaceae Vismia sp. Plantas no objeto de estudio
12 Lamiaceae Hyptis sp. Plantas objeto de estudio
13 Maranthaceae Calathea lutea Plantas objeto de estudio
14 Melastomataceae Miconia sp. Plantas objeto de estudio
15 Moraceae Ficus obtusiuscula Plantas objeto de estudio
16 Ochnaceae Sauvagesia erecta Plantas objeto de estudio
17 Phytolaccaceae Phytolacca americana Plantas objeto de estudio
18 Piperaceae Piper sp. Plantas objeto de estudio
19 Piperaceae Piper aduncum Plantas objeto de estudio
20 Poaceae Paspalum sp. Plantas objeto de estudio
21 Rutaceae Zanthoxylum sp. Plantas objeto de estudio
22 Salicaceae Banara guianensis Plantas no objeto de estudio
23 Solanaceae Solanum sp. Plantas objeto de estudio
24 Solanaceae Witheringia solanácea Plantas objeto de estudio
25 Urticaceae Pouroma cecropiifolia Plantas objeto de estudio
26 Urticaceae Cecropia ficifolia Plantas objeto de estudio
27 Verbenaceae Cithrexylum sp. Plantas objeto de estudio
28 Vitaceae Cissus sp. Plantas objeto de estudio

selva tropical, y la mayoría proceden de los trópicos y los subtrópicos. Algunas son
hierbas perennes de la zona templada (Jardín Botánico de Acapulco 2014).
Matelea (Foto 1) es un género de arbustos perennes con 347 especies
perteneciente a la familia Apocynaceae. Es originario de América (Universidad de
Bayreuth 2013).
Descripción
Son enredaderas sufrútices (en los trópicos), hierbas postradas o erectas (Foto 1)
(en zonas templadas) o, raramente, arbustos, con látex blanco. Brotes lenticelados o
con corteza suberosa; tricomas cortos, erectos o largo, suave o flexuous, incoloro, de
color blanquecino, amarillo, rojizo, de color marrón o de color óxido, tricomas
glandulares presentes o, rara vez, ausente. Hojas largamente pecioladas, hojas
herbáceas o coriáceas (si son glabras), de 1,5 a 30 cm de largo, 1,5 a 21 de ancho,
redondeadas o cuneadas elípticas, ovadas o hastiformes, basalmente cordadas,
lobuladas, ápice agudo, acuminado o atenuado, lisos, o rugosos (Universidad de
Bayreuth 2013).
Las inflorescencias extra-axilares, siempre una por nodo, con 1-10 de flores, simples,
poco pedunculadas, con pedúnculos más largos o casi tan largos como los pedicelos;
indumento de pedúnculos y pedicelos correspondientes con tricomas glandulares
poco frecuentes en los pedúnculos florales; brácteas iconspicuous o ausentes
(Universidad de Bayreuth 2013).
Sinonimia: Matelea abbreviata, Matelea acuminata, Matelea acutissima, Matelea
albiflora, Matelea albomarginata y Matelea angustiloba

FAMILAI ARACEAE
Anthurium sp
El género neotropical Anthurium (Foto 2) consta de 600 a 800 especies, llamadas
comúnmente anturios o calas. Inicialmente se descubrió en Costa Rica. Es originario
de las zonas tropicales y subtropicales de América Central y América del Sur. Es el
género más grande y probablemente más complejo de esta familia. Muchas especies
no se han descrito todavía y se encuentran nuevas cada año. Es el único género
perteneciente a la tribu Anthurieae (USDA 2013).
Descripción
Son plantas perennes, herbáceas o leñosas, erectas, rastreras o trepadoras, de hojas
muy decorativas (Foto 2). Las hojas son de consistencia y grosor notables, ovales, en
forma de corazón o punta de flecha, bastante grande, a veces divididas en lóbulos o
incluso en forma de mano. El espádice, que a menudo se confunde con la flor del
anturio, constituye el elemento de mayor curiosidad: puede ser rojo, purpúreo, verde
manzana, rosa intenso, casi anaranjado, blancas, negras y amarillos que son muy
difíciles de conseguir. Son llamativas y su tamaño varía según su especie
(Croat 1983).
FAMILA ASTERACEAE
Veronia sp
Las asteráceas, también denominadas compuestas (Compositae Giseke, nom. cons.),
reúnen más de 23500 especies repartidas en unos 1600 géneros por lo que son la
familia de Angiospermas con mayor riqueza y diversidad biológica. La familia se
caracteriza por presentar las flores dispuestas en una inflorescencia compuesta

denominada capítulo la cual se halla rodeada de una o más filas de brácteas
(Jeffrey 2007).
Descripción
Pueden ser anuales, bienales o perennes, con una agregación basal de hojas (una
"roseta" basal) o sin ella. Existen especies hidrofíticas (ciertas especies de Bidens o
Cotula), que presentan hojas sumergidas y emergentes (Foto 3). Por otro lado,
existen especies heliofíticas, mesofíticas y xerofíticas (Watson y Dallwitz 2008).
Las Asteráceas han sido por mucho tiempo reconocidas como un grupo natural y a
pesar de que algunos taxónomos han subdividido a la familia en 2 o 3 familias.
Las sinapomorfias de las compuestas incluyen: el cáliz modificado en una estructura
denominada papus, las anteras connadas formando un tubo y los estigmas
modificados para funcionar como cepillos en un mecanismo especializado de
polinización, los ovarios que contienen un solo óvulo de placentación basal y la
producción de lactonas sequiterpénica (Barkley et al. 2008).
Sinonimia
La familia, entonces, ha sido universalmente reconocida y dispuesta en el orden
Asterales de las dicotiledóneas o eudicotiledóneas. Desde el punto de vista de la
taxonomía intrafamiliar, tradicionalmente se han reconocido dos subfamilias:
Asteroideae (o 'Tubuliflorae') y Cichorioideae (o 'Liguliflorae'), desde que en 1894
Hoffmann reconoció a las mismas como dos linajes diferentes. Asteroideae se
caracteriza por la presencia de capítulos radiados y la ausencia de látex. La
subfamilia Cichorioideae, en cambio, se caracteriza por la presencia de látex y por
sus capítulos con flores liguladas (Stevens 2012).

FAMILIA CAMPANULACEA
Hippobroma longiflora
Hippobroma es un género monotípico de plantas perteneciente a la familia
Campanulaceae. Su única especie: Hippobroma longiflora (Foto 4), originaria de
Jamaica y se ha extendido por todo el mundo, naturalizándose en los trópicos
americanos y en Oceanía (Echeverry 2011).
Descripción
Hierbas perennes erectas o decumbentes (Foto 4), con un tamaño de 30 a 90 cm de
alto. Hojas elípticas a oblanceoladas, mayormente 5 a 20 cm de largo y 1 a 5 cm
de ancho, gruesa e irregularmente repando-dentadas con 15 a 20 dientes grandes,
así como también calloso-denticuladas con 4 a 6 veces más dientes pequeños. Flores
solitarias en las axilas de las hojas, con un par de bractéolas basales, filiformes y 2 a
4 mm de largo; hipanto obcónico, 6 a 10 mm de largo, lobos del cáliz más o menos
lineares, márgenes calloso-denticulados (Missouri Botanical Garden 2014).
Sinonimia: Lobelia longiflora, Rapuntium longiflorum, Isotoma longiflora, Laurentia
longiflora, Solenopsis longiflora, Isotoma runcinata y Laurentia longiflora var.
runcinata
FAMILIA CONVOLUVLACEAE
Ipomoea batatas
Ipomoea batatas (Foto 5), llamada comúnmente boniato, batata (del taino), chaco,
papa dulce o camote, es una planta de la familia Convolvulaceae, cultivada en gran
parte del mundo por su raíz tuberosa comestible (Langdon 2001).

Descripción
En el oriente de Venezuela recibe el nombre de chaco, probablemente originario de
lengua guaiquerí. Originaria de los trópicos de Sudamérica y América Central,
cultivada desde hace 8000 años en lo que hoy es Perú, y se han hallado
representaciones de camote en numerosos ceramios precolombinos y restos de las
raíces tuberosas en algunas tumbas. Llegó a Europa en el siglo XVI y se ha difundido
ampliamente en todo el mundo. Según estadísticas, China es el principal productor,
pues cultiva aproximadamente el 80 % del total mundial; le siguen Uganda, Nigeria e
Indonesia. Islas Salomón tiene la mayor producción per cápita del mundo: 160 kg por
persona por año. Es un alimento reconocido como eficaz en la lucha contra la
desnutrición debido a sus características nutritivas, facilidad de cultivo y
productividad (FAO 2009).
Sinonimia: Convolvulus batatas L, Convolvulus tiliaceus, Ipomoea fastigiata
Ipomoea tiliacea y Ipomoea triloba.
Ipomoea sp
Ipomoea (Foto 6) es un género de plantas perteneciente a la familia de las
convolvuláceas, vulgarmente conocidas como campanitas. Comprende 3062
especies oriundas de zonas templadas y cálidas. La mayoría de ellas habita en
África y América tropical. Casi todas las especies del género son enredaderas
herbáceas, aunque existen unas pocas que son arbustivas o arborescentes.
Descripción
Son hierbas perennes (Foto 6), raramente anuales, a veces se encuentran
lignificadas en la base. Pueden ser rizomatosas o tuberosas, pubescentes o glabras.
Los tallos son volubles, raramente decumbentes, de sección poligonal o circular, con
látex hialino, raramente de color blanco (Flora Ibérica 2012).

Tienen una amplia distribución en zonas tropicales y subtropicales de todo el mundo,
abarcando ambos hemisferios. A la llegada de los españoles a América, se conocía el
cultivo de camotes, desde México hasta Perú; restos arqueológicos en este último
país datan de 2.000 años a.c, y aún antes. A la llegada de Magallanes a Oceanía, en
1521, el camote era cultivado en casi todas las islas. Como el término para designarlo
en Oceanía tiene afinidad con el nombre "kumura" que se usa en Perú, y como la
expansión polinésica es mucho más reciente que las culturas americanas en que se
conocía el cultivo, es de asumir que el camote fuera llevado de América a Oceanía.
En Argentina, crecen 52 especies, once de las cuales son endémicas o propias
(De Marzi 2006).
FAMILIA CYPERACEAE
Cyperus ustulatus
Cyperus ustulatus (Foto 7) es el nombre botánico de esta especie perteneciente a la
familia Cyperaceae y es conocida de forma común como: juncia paraguas gigante.
Descripción
Perenne original de Nueva Zelanda puede llegar a alcanzar un metro con veinte
centímetros de altura Cyperus ustulatus (Foto 7) cuenta con poca información fiable
sobre sus flores, pero éstas son presumiblemente de color granate dotadas de
unidades reproductivas hermafroditas (Pérez 2013).
La especie Cyperus ustulatus se desarrollará mejor en suelos con pH ácido, neutro o
alcalino. Su parte subterránea crecerá con vigor en soportes con textura arenosa o
franca, éstos se pueden mantener generalmente húmedos o empapados. Es de suma
importancia regar teniendo en cuenta la información anterior, pero también factores
tales como: exposición al sol, temperatura, textura del suelo, época del año, entre
otros. Todo ello para buscar un equilibrio más o menos constante en la humedad del

soporte. En cuanto a sus necesidades lumínicas, podemos aseverar que es muy
exigente, se debe situarse en un lugar con exposición directa al sol para no repercutir
negativamente en su crecimiento de forma normal (Pérez 2013).
Sinonimia: Cyperus distans, Cyperus esculentus, Cyperus fendlerianus, Cyperus
longus, Cyperus malaccensis, Cyperus rotundus, Cyperus schweinitzii, Cyperus
setigerus, Cyperus squarrosus.
FAMILIA CYPERACEAE
Scleria sp.
Scleria es un género de plantas herbáceas (Foto 8) perteneciente a la familia de las
ciperáceas. Comprende 631 especies descritas y de estas, solo 262 aceptadas.
Descripción
Son plantas anuales o perennes, rizomatosas (Foto 8); con culmos triangular-
triquetros, erectos a inclinados o escandentes, mayormente ásperos; plantas
monoicas. Hojas en 3 series, las inferiores sin lámina, plegadas en las costillas; vaina
cerrada, en el ápice con una lígula y/o contralígula, o éstas no evidentes.
Inflorescencia compuesta, densa o discontinua, de glomérulos o racimos cortos de
espiguillas unisexuales; inflorescencia estaminada con varias flores, estambres 1–3;
inflorescencia pistilada con 1 flósculo terminal directamente abrazado por 2 escamas
grandes y por debajo con unas pocas escamas más pequeñas, estilo variadamente
articulado al ovario, con 3 ramas estigmáticas alargado-lineares. Fruto de contorno
ancho, el cuerpo con superficie dura o variadamente ornamentada, abrazado por un
estípite obpiramidal o prismático, el cual puede o no producir apicalmente un perianto
modificado (el hipoginio) entero o variadamente 3-lobado, fimbriado, ciliado,
crenulado, dentado o tuberculado (Missouri Botanical Garden 2014).
Sinonimia: Scleria abortiva, Scleria acanthocarpa, Scleria achteni, Scleria acriulus y
Scleria adamantina.

FAMILIA DENNSTAEDTIACEAE
Hypolepis sp.
Hypolepis (Foto 9) es un género de helechos perteneciente a la familia
Dennstaedtiaceae. Comprende 141 especies descritas y de estas, sólo 42 aceptadas
(Missouri Botanical Garden 2014).
Descripción
Son una familia de helechos del orden Polypodiales, con rizomas en su mayoría
largamente rastreros (Foto 9). Normalmente sifonostélicos o polistélicos. Pecíolos
usualmente con ápice epipeciolar, usualmente con un haz vascular en forma de
alcantarilla abierto y adaxial. Láminas usualmente grandes, 2-3 pinadas o más
divididas. Indumento de pelos. Venas libres, bifurcadas o pinadas, raramente
anastomosándose y si es así no tienen venillas inclusas. Soros marginales o
submarginales, lineales o discretos. Indusio linear o cupuliforme en el margen de la
lámina, o bien reflejada en los soros. Esporas tetraédricas con marca trilete (Missouri
Botanical Garden 2014).
Sinonimia
Existen 140 especies entre las principales tenemos: H. aculeata, H. alpina, H. alte-
gracillima, H. amaurorachis, H. ambigua, H. anthriscifolia y H. apicilaris

FAMILIA FABACEA
Desmodium heterophyllum
Esta leguminosa es de origen Asiático (Foto 10), fue introducida desde Australia
adaptándose bien desde el nivel del mar hasta los 1200 msnm; con precipitación que
van de 1000 a 4000 mm y temperaturas de 20 a 25°C.
Descripción
Planta de crecimiento matoso (Foto 10), con alturas promedios de 10 a 15 cm, tiene
raíz pivotante, ramificada en la cual se forman los nódulos de bacterias fijadoras de
Nitrógeno en el suelo. Los tallos son abundantes medianamente pubescentes de
color café rojizo o claro, cada uno emite numerosas raicillas que al estar en contacto
con la humedad del suelo, dan origen a nueva plantas, sus hojas son trifoliadas, con
foliolos pequeños y ovalados. Las flores en forma de mariposa son pequeñas de color
lila. Las vainas son pequeñas y aplanadas, al igual que la semilla (González y
Caballero 1989).
Crece bien en suelos de baja fertilidad y muy húmedos como los del Oriente
ecuatoriano, esta especie no tolera sequía, ni inundación prolongada, produce buena
semilla lo cual le permite poblarse rápidamente (González y Caballero 1989). En
Ecuador se ha introducido en la selva baja y alta de la región Amazónica, donde ha
sido seleccionada como promisoria.
FAMILIA HYPERICACEA
Vismia sp.
Vismia (Foto 11) es un género de plantas perteneciente a la familia Hypericaceae.
Como la mayoría de los miembros de Clusiaceae, las especies de este género

contienen xantonas. Comprende 107 especies descritas y de estas, solo 45 aceptada.
Es originario de las zonas tropicales de América y África (Missouri Botanical Garden
2014).
Descripción
Son árboles o arbustos, con tricomas simples y también estrellados o dendroides, y
con hileras glandulares o puntos rojo obscuros a rosados o anaranjados; plantas
hermafroditas (Foto 11). Hojas con puntos glandulares negruzcos. Flores homostilas
o heterostilas; sépalos 5, pubescentes dorsalmente excepto en los angostos
márgenes internos glandulares; pétalos 5, vellosos en el interior, con glándulas
anaranjadas a negras; estambres en 5 fascículos antipétalos cada uno de 10–50
estambres; fascículos estaminodiales 5; ovario 5-locular con placentación axial,
estilos 5, libres. Fruto una baya lisa, glabra, frecuentemente punteado-glandular;
semillas numerosas, cilíndricas (Missouri Botanical Garden 2014).
Sinonimia: Vismia acuminata, Vismia affinis, Vismia amazonica, Vismia angusta,
Vismia angustifolia, Vismia baccifera, Vismia bemerguii, Vismia billbergiana, Vismia
brasiliensis, Vismia buchtienii, Vismia calvescens y Vismia camparaguey.
FAMILIA LAMIACEAE
Hyptis sp
Planta originaria de América tropical, que habita en climas cálidos y semicálidos
desde el nivel del mar hasta los 760 m de altitud. Asociada a vegetación perturbada
derivada de bosques tropicales subperennifolios y perennifolios. Hyptis (Foto 12) es
un género con 694 especies de plantas con flores pertenecientes a la familia
Lamiaceae.

Descripción
Son hierbas o arbustos pequeños, que alcanzan un tamaño de hasta 2.5 m de alto,
aromáticos; tallos jóvenes menudamente puberulentos con tricomas largos en los
nudos (glabros). Hojas lineares, angostamente elípticas u oblongas, 2.2–9.5 cm de
largo y 0.5–2 de ancho, ápice acuminado (redondeado o agudo), base atenuada,
margen irregularmente serrado los 2/3 apicales, envés con tricomas en el nervio
principal, glabrescente; pecíolo 0.1–2 cm de largo. Inflorescencia de verticilastros bien
espaciados y nunca espiciformes, en las axilas de las hojas reducidas, verticilastros
0.2–0.8 cm de largo y 0.5–0.8 mm de ancho (ligeramente más anchos en fruto), 11–
15 flores, verticilastros sésiles, pero cada flor con pedicelo definido, corto y filiforme,
brácteas reemplazadas por prominentes fascículos de tricomas; cáliz 2–2.5 mm de
largo, externamente glabro o menudamente puberulento, internamente glabro,
dientes ampliamente deltoides, erectos, 0.5–0.75 mm de largo; corola blanca
(violeta), tubo 1.5–2 mm de largo, limbo 1–1.5 de largo. Cáliz fructífero 2–3 de largo y
1–2 de ancho, dientes 0.4–1 de largo; nuececillas 1–1.5 de largo, glabras (Foto 12)
(Missouri Botanical Garden 2014).
Sinonímia: Mesosphaerum P. Browne, Brotera Spreng, Condea Adans, Gnoteris Raf
y Hypothronia Schrank.
FAMILIA MELASTOMATACEA
Miconia sp.
Miconia (Foto 13) es un género botánico de plantas perteneciente a la familia
Melastomataceae. Comprende 1350 especies descritas y de estas, solo 709
aceptadas (Missouri Botanical Garden 2014).

Descripción
Son nativos de zonas cálidas y tropicales de América. También habitan en las Islas
Galápagos en Ecuador, que al parecer han sido invadidas par la quina o cascarilla. La
mayoría de las especies son arbustos y pequeños árboles de hasta 15 metros de
altura. Se caracteriza por tener la haz verde oscura y el envés púrpura (Foto 13)
(Missouri Botanical Garden 2014).
Sinonimia: Miconia abbreviata, Miconia aequatorialis, Miconia albican, Miconia
aligera, Miconia alpina, Miconia ascendens, Miconia asplundii, Miconia aspratilis,
Miconia ayacuchensis y Miconia bailloniana.
FAMILIA MORACEAE
Ficus obtusiuscula
El género Ficus (Foto 14) contiene alrededor de 900 taxones específicos e infra-
específicos aceptados de árboles, arbustos y trepadoras de la familia Moraceae, tribu
mono genérica Ficeae, oriundas de la Zona Intertropical, con algunas de ellas
distribuidas por las regiones templadas. La mayoría son perennes, excepto las que
vegetan en latitudes no tropicales y zonas con una larga estación seca (Missouri
Botanical Garden 2014).
Descripción
Una de las características de las especies de este género es la secreción lechosa
llamada látex que segregan al cortar cualquier parte de la planta. Pero el más
característico es el tipo muy particular de inflorescencia que se parece más a un fruto
que a unas flores habituales (Ecología Áreas verdes 2015).

Otra característica importante del género reside en que las yemas terminales de las
hojas están encerradas dentro de un par de estípulas soldadas en un principio y
luego caducas (Foto 14) (Ecología Áreas verdes 2015).
Estos falsos frutos son una estructura especialmente adaptada que se denomina
sicono. Son de forma bulbosa con una pequeña abertura, el ostíolo, en el apex y una
zona hueca en el interior recubierta por pequeñas flores. Estas últimas son
polinizadas por pequeñas avispas que penetran por la abertura para fertilizarlas,
dando frutos diminutos con un núcleo duro (aquenios), vulgarmente llamados pepitas,
rodeados de un mesocarpio carnoso; el conjunto es el higo (Ecología Áreas verdes
2015).
Sinonimia
Unas 830 especies aceptadas de las más de 2.700 descritas y unos 75 taxones infra-
específicos aceptados de los más de 800 descritos: F. abutilifolia, F. aguaraguensis,
F. albert-smithii, F. albipila, F. altissima y F. amazónica.
FAMILIA OCHNACEAE
Sauvagesia erecta
Sauvagesia (Foto 15) es un género de plantas fanerógamas que pertenecen a la
familia Ochnaceae. Comprende 77 especies descritas y de estas, solo 37 aceptadas.
Se distribuye por los trópicos de Sudamérica (Missouri Botanical Garden 2014).
Descripción
Son hierbas anuales o perennes, glabras. Hojas enteras o serrulado-glandulosas,
subsésiles; estípulas persistentes y ciliadas. Flores pequeñas, axilares o raramente
en inflorescencias terminales, blancas a rosadas; sépalos y pétalos 5; estaminodios

en 1–3 verticilos, envolviendo el ovario y los 5 estambres; ovario con 3 placentas
parietales. Fruto una cápsula septicida; semillas numerosas (Foto 15) (Missouri
Botanical Garden 2014).
Sinonimia: Sauvagesia africana, Sauvagesia aliciae, Sauvagesia amoena,
Sauvagesia angustifolia, Sauvagesia brevipetala, Sauvagesia capillaris, Sauvagesia
deflexifolia y Sauvagesia elata.
FAMILIA SALICACEAE
Banara guianensis
Se encuentra en bosques semicaducifolios y bosques de pino, zonas norcentral y
atlántica; a una altitud de 30–1000 msnm; desde Nicaragua hasta Bolivia y Brasil.
Descripción
Son arbustos, árboles (bejucos) (Foto 16), siempre verdes, que alcanza un tamaño de
2–15m de alto, inermes; plantas androdioicas. Hojas alternas, espiraladas, oblongas
o a veces ovadas, con 7–20 cm de largo y 3–7.5 de ancho, el ápice acuminado
(agudo o redondeado), la base truncada (obtusa o redondeada), el margen serrado-
glandular, pinnatinervias; con estípulas diminutas, caducas. La inflorescencia es una
panícula axilar o subterminal, de 9–15 cm de largo, con 20–70 flores, con brácteas
diminutas; sépalos 3, valvados, ovados o elípticos, 2.5–4 mm de largo; pétalos 3,
valvados, oblongos, 2.5–4.5 mm de largo, amarillos; a veces con 1 ó 2 filamentos
estaminales petaloides, estambres hipóginos, numerosos, multiseriados, filamentos
2.5–4 mm de largo, fusionados en la base a un receptáculo densamente velloso,
glándulas del disco ausentes; ovario súpero, estilo simple, 1.5–2.5 mm de largo,
estigma capitado, undulado, ovario rudimentario en las flores estaminadas. Fruto una
baya carnosa, globosa, 7–11 mm de diámetro, con estilo y perianto persistente, verde
a negra, glabra; semillas numerosas (Missouri Botanical Garden 2014).

Sinonimia: Banara fagifolia Vahl, Banara guianensis var. mollis (Poepp.) Eichler,
Banara guianensis var. spruceana Briq, Banara mollis (Poepp.) Tul, Banara
pubescens Spruce ex Benth, Banara pyramidata Rusby, Banara tulasnei J.F. Macbr.,
Kuhlia mollis Poepp y Laetia glandulosa Poepp. ex Tul.
FAMILIA SOLANACEA
Solanum sp.
Las solanáceas (Foto 17) (Solanaceae juss.) son una familia de plantas herbáceas o
leñosas con las hojas alternas, simples y sin estípulas pertenecientes al orden
Solanales, de las dicotiledóneas (Magnoliopsida). Comprende aproximadamente 98
géneros y unas 2700 especies, con una gran diversidad de hábito, morfología y
ecología. La familia es cosmopolita, distribuyéndose por todo el globo con la
excepción de la Antártida. La mayor diversidad de especies se halla en América del
Sur y América Central (Olmstead et al. 1999).
Descripción
Las solanáceas son plantas herbáceas, subarbustos, arbustos, árboles o lianas.
Pueden ser anuales, bienales o perennes, erguidas o decumbentes. Pueden estar
provistas de tubérculos subterráneos. No presentan laticíferos, ni látex, ni jugos
coloreados. Pueden presentar una agregación basal o terminal de hojas o pueden no
tener ninguno de ambos tipos. Las hojas son generalmente alternas o alternadas a
opuestas (o sea, alternas en la base de la planta y opuestas hacia la inflorescencia).
La consistencia de las hojas puede ser herbácea, coriácea, o pueden estar
transformadas en espinas. En general las hojas son pecioladas o subsésiles,
raramente sésiles (Foto 17).

La lámina foliar puede ser simple o compuesta, en este último caso, pueden ser
ternadas o pinnatífidas. La nerviación de las hojas es reticulada y no presentan un
meristema basal. Con respecto a la anatomía foliar, las láminas son en general
dorsiventrales, sin cavidades secretoras. Los estomas se hallan en general
confinados a una de las caras de las hojas, raramente en ambas caras
(Olmstead et al. 1999).
Las flores son en general hermafroditas, si bien hay especies monoicas,
andromonoicas o dioicas (como por ejemplo, algunos Solanum o Symonanthus). La
polinización es entomófila. Las flores pueden ser solitarias o estar agregadas en
inflorescencias cimosas, terminales o axilares. Las flores son de tamaño intermedio,
fragantes (como en Nicotiana), fétidas (Anthocercis) o inodoras. Las flores son
actinomorfas, levemente cigomorfas o marcadamente cigomorfas (como por ejemplo,
las flores con corola bilabiada en Schizanthus). Las irregularidades en la simetría
pueden deberse al androceo, al perianto o a ambos a la vez. Las flores en la gran
mayoría de los casos presentan un perianto diferenciado en cáliz y corola (con 5
sépalos y 5 pétalos, respectivamente), un androceo con 5 estambres y dos carpelos
unidos formando un gineceo con ovario súpero (se dice, entonces, que son
pentámeras y tetracíclicas). Usualmente presentan un disco hipógino (Foto 17)
(Olmstead et al. 1999).
El cáliz es gamosépalo (ya que los sépalos están unidos entre sí formando un tubo),
con los 5 (a veces 4 o 6) segmentos iguales entre sí, es pentalobulado, con los
lóbulos más cortos que el tubo, es persistente y puede muy a menudo ser acrescente.
La corola usualmente presenta 5 pétalos que también se hallan unidos entre sí
formando un tubo. La corola puede ser campanulada, rotada, infundibuliforme o
tubular (Olmstead et al. 1999).

Sinonimia:
De la que existe los siguientes representantes: Solanum (1.330), Lycianthes (200),
Cestrum (150), Nolana (89), Physalis (85), Lycium (85), Nicotiana (76) y Brunfelsia
(45).
FAMILIA SOLANACEA
Witheringia solanácea (Foto 18) crecen de 0 a 1.700 m de altitud. Se encuentran en
regiones tropicales, en países como: Costa Rica, más común en la vertiente Caribe.
También en México, Bolivia, Venezuela y Jamaica (Soto 2015).
Descripción
Hierbas, arbustos o árboles débiles, de 1 a 4m de alto; tallos frecuentemente huecos,
glabros o variadamente pubescentes con tricomas simples, glandulares. Hojas en
pares desiguales, ovadas, de 7 a 18 cm de largo, variadamente puberulentas con
tricomas simples, ápice acuminado, base obtusa; los pecíolos son de un tercio a un
medio de la longitud de la lámina (Soto y Castro 2011).
Inflorescencias de fascículos subsésiles con muchas flores, pedicelos mayormente
delgados, curvadas por abajo de las hojas y erectos en el fruto, hasta 6 mm de largo;
flores, 4-meras; cáliz de 0,5 a 2,0 mm de largo, entero, piloso-glanduloso a glabro;
corola de 5 a 8 mm de largo, es lobada en dos tercios de su longitud, puberulenta
por afuera, glabra por dentro, excepto por un anillo de pubescencia cerca de la
boca, verdosa, café o purpúrea; filamentos insertos en la porción apical del tubo
de la corola, pubescentes, anteras iguales, anchamente ovoides, de 2 a 3 mm de
largo, apicalmente agudas y menudamente apiculadas. Fruto tipo baya globosa
de 4 a 8mm de diámetro; semillas de 1,0 a 1,5mm de diámetro (Foto 18) (Soto y
Castro 2011).

FAMILIA URTICACEA
Cecropia ficifolia
El género Cecropia (Foto 19), nombrado en honor a Cécrope, comprende unas 65
especies de árboles dioicos que se hallan en Centroamérica y Sudamérica.
Anteriormente este género se había clasificado en la familia Moraceae, pero estudios
genéticos lo encontraron más relacionado con la familia Urticaceae, en tanto que
varios expertos prefieren catalogarlo en una familia propia, Cecropiaceae, junto con el
género Coussapoa. Estos árboles suelen denominarse ambaibo, ambay, uarumo,
yagrumo, yarumo o cético (Bernal y Gradstein 2015).
Descripción
Son árboles (Foto 19), frecuentemente con raíces fulcrantes y poco ramificados, tallos
terminales normalmente huecos y septados, habitados por hormigas, con látex
obscuro al secarse. Hojas peltadas, ligera a profundamente palmatilobadas; pecíolos
teretes y acostillados, con pulvínulo grande en la base. Inflorescencias en espigas
densas y carnosas, umbeladas en el ápice de los pedúnculos y envueltas por una
espata decidua; sépalos connados; estambres 2, libres; estigmas fimbriados.
Aquenios 1–3 mm de largo, mayormente 2–3-angulados (Bernal y Gradstein 2015).
Los tallos son huecos y tabicados en los nudos y de ellos se desprenden raíces
zancudas. Contienen un látex tóxico y en sus ambientes nativos forman alianzas con
las hormigas (mirmecófilas). Las hojas son alternas, simples y dispuestas en espiral;
los indígenas suramericanos las utilizan tradicionalmente para incinerarlas y producir
una cal que mezclan con las hojas de coca antes de masticarlas. Son utilizados como
ornamentales en Europa (Bernal y Gradstein 2015).

Sinonimia: Cecropia adenopus, Cecropia angustifolia, Cecropia glaziovii, Cecropia
hololeucan, Cecropia insignis y Cecropia multiflora.
FAMILIA VERBENACEA
Citharexylum sp.
Citharexylum (Foto 20) es un género botánico de plantas con flores con 247 especies
descritas y de estas, solo 113 aceptadas, pertenecientes a la familia de las
verbenáceas. Es nativo de las regiones tropicales y subtropicales de América (Royal
Botanic Garden 2014).
Descripción
Son árboles o arbustos con las hojas opuestas (Foto 20), a veces subopuestas,
ternadas o verticiladas, en ocasiones alternas, simples, margen entero o raramente
dentado, a menudo con glándulas grandes en la base de la lámina. Inflorescencia
racimos o espigas, terminal y axilar, mayormente alargada y con muchas flores
mayormente amarillas o blancas, a veces azules, moradas o lilas, cada flor abrazada
por una bráctea inconspicua; cáliz tubular (no en Nicaragua) o cupuliforme
(cupuliforme agrandado o pateliforme cuando en fruto), truncado, repando o con 5
dientes; corola infundibuliforme o hipocrateriforme, casi regular, en Nicaragua de 5–
8(–10) mm de largo, con 5 lobos casi iguales, 1.5–3 mm de largo; estambres 4,
incluidos, con 1 estaminodio; estilo incluido, estigma cortamente 2-lobado. Fruto
drupáceo, con exocarpo jugoso y endocarpo duro conteniendo 2 pirenos; semillas 2
por cada pireno (Missouri Botanical Garden 2014).
Sinonimia: Citharexylum affine, Citharexylum alainii, Citharexylum albicaule,
Citharexylum altamiranum, Citharexylum andinum, Citharexylum argutedentatum y
Citharexylum berlandieri.

FAMILIA VITACEAE
Cissus sp.
Cissus (Foto 21) es un género de unas 350 especies de lianas leñosas trepadoras en
la familia de la vid (Vitaceae). Tienen una distribución cosmopolita, aunque la mayoría
son de los trópicos de África, el sur de Asia, Australia, Nueva Guinea y las Américas
(Grubenn 2004).
Las especies más comunes, Cissus antarctica y Cissus rhombifolia, son plantas de
jardín. Cissus gongylodes es una planta alimenticia cultivada en la selva desde hace
siglos por los indígenas y denominada por ellos cupá, y en Brasil, cipó-babao. Cissus
quadrangularis es medicinal usada desde la antigüedad en la India. Cissus striata es
una leñosa nativa de Sudamérica (Chile, Brasil y Argentina).
Sinonimia: Cissus alata, Cissus antarctica - uva de canguro, Cissus erosa, Cissus
fuliginea, Cissus gongylodes - cupá, vid de mármol y Cissus hypoglauca.
II.2.4 Discusión
Los bosques amazónicos almacenan múltiples productos naturales entre los que se
destacan gran variedad de alimentos, insumos medicinales y elementos para la
producción artesanal y otros servicios (Vélez 2000; Castro y Galán 2003). Los
efectos producidos por la tala y la quema para suplir la demanda de maderas con
alto valor comercial y las áreas ganaderas constituyen parte de la problemática
ambiental de esta región. Estos procesos extractivos causan pérdida irreversible de
hábitat naturales y recursos genéticos, con efectos secundarios como la
fragmentación, el desplazamiento de la fauna y la alteración de las cadenas tróficas
(IDEAM-FAO 2002). El estudio de las riquezas, diversidad y composición florística de
la sucesión vegetal secundaria que aparecen después de estas prácticas en la

Amazonia Ecuatoriana es de vital importancia, ya que muchas de estas pueden ser
empleadas en la alimentación animal.
La cantidad de familias y de especies inventariadas en este estudio se enmarcan
dentro de la riqueza y diversidad de los bosques húmedos tropicales (Gentry 1995;
Neill 2012; Samaniego 2014). Aunque la riqueza de especies fue menor si se
compara con la registrada para otros bosques Andinos húmedos como los de Murrí
en Colombia, Centinela, Huamaní, Jatun Sacha y Miazí en Ecuador (Phillips y Miller
2002).
Es notorio también que pocos géneros son los que contribuyen sustancialmente a la
riqueza de especies. Esto se corresponde con lo reportado por Gentry (1988) y
Vázquez y Givnish (1998).
Bustamante et al. (2009) al inventariar la riqueza y biodiversidad de la Amazonia
Colombina encontraron 36 especies de árboles y arbustos y 21 herbáceas, donde las
Familias con mayor proporción de especies fueron la Fabaceae y Moraceae. La
riqueza de familias botánicas encontradas en esta investigación coinciden con lo
reportado por Gentry (1982), Sierra (1999), Neill (2000), Lozano (2002),
Yaguano et al. (2012) donde refirieron que estas constituyen elementos distintivos del
bosque húmedo tropical, caracterizado por una flora entre la vegetación de tierras
bajas y de cordillera.
Asimismo, concuerdan con los estudios de Balcazar et al. (2000) y
Cuatrecasas (1958) en la serranía media de Las Quinchas Magdalena, entre 380 y
1 000 m y en gradientes altitudinales de la cordillera Andina.
Mientras que Jardines et al. (2000) encontraron mayor diversidad de especies de
leguminosas naturales en las áreas marginales, especialmente las de crecimiento
voluble, pero dentro del área pastada solo abundaron las plantas de crecimiento
postrado y de porte bajo. Esta característica de algunas leguminosas naturales que

les permite persistir ante el pastoreo frecuente, puede explicar el fracaso de otras
leguminosas seleccionadas y recomendadas para el pastoreo.
Sistachs (1986) y Jardines et al. (2000) plantearon que se conoce que a diferencia de
las leguminosas volubles de clima templado, las tropicales tienen un mayor
rendimiento en el período lluvioso, en el cual pueden alcanzar hasta 75 % del total
anual. También las altas temperaturas de este período coinciden con el óptimo
necesario para el crecimiento de este tipo de planta en el trópico. La humedad del
suelo favorece el crecimiento de las especies y pudiera diferenciar su
comportamiento.
Pinto et al. (2002) al estudiar especies arbóreas y herbáceas forrajeras en el sureste
de México encontraron 66 especies arbóreas y 19 herbáceas relacionadas con el uso
forrajero. Sin embargo, sólo 14 arbóreas, tres enredaderas y cuatro arvenses se
consideraron de mayor importancia forrajera, de acuerdo con su utilización por los
productores. De las enredaderas y arvenses, el puyú (Ipomoea triloba) y la flor
amarilla (Sanvitalia procumbens) se destacaron por su uso forrajero (88,2 y 54,9%,
respectivamente). Una elevada proporción de estas posee un uso múltiple, todas las
especies tuvieron usos o servicios alternos al forrajero. Son usadas como leña (12),
seguidos de las utilizadas como utensilios (10), medicinales (7) y cercas vivas (6).
En esta investigación las especies inventariadas en la sucesión vegetal secundaria
tienen innumerables usos por su riqueza en compuestos secundarios y otros
nutrimentos son empleadas en la medicina, casa y alimentación animal.
La familia Convolvulaceae según Martí et al (2011) la raíz contiene grandes
cantidades de almidón, vitaminas, fibras (celulosa y pectinas) y minerales, y destaca
entre estos el contenido de potasio. En valor energético supera a la patata, se
destaca por la provitamina A (betacaroteno) y las B1, C (ácido ascórbico) y E
(tocoferol). Cuanto más amarillenta es su raíz, más betacaroteno posee, con esta

coloración son muy utilizadas en Asia y África para reducir la deficiencia de vitamina
A en los niños. Su sabor dulce se lo debe a la sacarosa, la glucosa y la fructosa.
Además, su raíz, si bien no posee altos contenidos de proteína, sí es importante en
contenido de lisina. Por esto es que se le utiliza como complemento de diferentes
harinas de cereales. Su contenido de lípidos es bajo. Sus ácidos grasos principales
son el linoleico, el oleico, el esteárico y el palmitoleico. Posee gran cantidad de fibra
digerible, que acelera el tránsito intestinal, previene el cáncer de colon, controla el
nivel de glucosa, reduce el nivel de colesterol y produce sensación de saciedad. Su
piel y su pulpa poseen antioxidantes, por lo que previene enfermedades cardíacas,
diabetes y cáncer (Martí et al. 2011).
En diferentes países se valora su hoja para alimentar tanto a animales como
personas, pues posee importantes niveles de hierro, provitamina A, vitamina B2,
vitamina C y vitamina E, fibra dietaria y polifenoles.
Los géneros Carpodinus, Landolphia, Hancornia, Funtumia y Mascarenhasia fueron
una fuente secundaria de caucho.
Rauvolfia cafra es el "árbol de la Quinina". Rauvolfia serpentina o "raíz de serpiente"
suministra los alcaloides reserpina y rescinamina. Presentan alcaloides carbolínicos y
otros alcaloides indólicos. Se han detectado iridoides en algunos géneros (Plumeria,
Rauwolfia y Allamanda). También presentan proantocianidinas, cianidina y delfinidina.
Pueden presentar kaempferol y/o quercetina. Se ha encontrado ácido ursólico. Los
azúcares son transportados como sacarosa o como oligosacáridos (Watson y
Dallwitz 2014).
En el caso de la Cyperaceae, fue de primordial importancia histórica en el
abastecimiento de papiro, posee tubérculos comestibles por los cuales es cultivada;
algunas otras especies se consumen también, aunque en menor medida. Algunas de
la especies tienen un cierto valor comercial y otras son cultivadas como
ornamentales. Se está incrementando el interés en las especies mayores y de

rápido crecimiento como cultivos de biomasa para la producción de papel y
combustible (Pérez 2013). Una de las representantes de esta familia Scleria sp se
encuentran principalmente en lugares húmedos es utilizada para la alimentación
animal en pastoreo (Missouri Botanical Garden 2014).
Por su parte las Fabaceae ya sean árboles, arbustos o rastreras tiene buena
aceptación por el ganado. Su valor nutritivo en lo que se refiere a proteína cruda está
entre 19,3 a 20,30 y su digestibilidad in vitro de la materia seca, va de 34,1 a 40,4%
(González y Caballero 1989).
El Desmodium heterophyllum por su tamaño bajo, se asocia bien con gramíneas de
igual como Brachiaria humidicola, Brachiaria dictyoneura, Brachiaria decumbens y
Brachiaria ruziziensis, con los que se forma un pastizal de buena producción forrajera
y calidad nutricional, la producción de semilla es abundante lo que hace que forme
una excelente asociación, además de llegar a alterar los componentes de la
composición botánica de la pradera. Es importante para el consumo animal
(González y Caballero 1989).
La importancia de la sucesión secundaria derivada de procesos naturales, se aprecia
por los muchos rasgos que cambian en la comunidad a medida que un ecosistema se
desarrolla y madura: la biomasa, el número de nichos, el grado de crecimiento de los
productores primarios, el grado de diversidad de las especies y el reemplazo de las
mismas, llegando a la etapa de un clímax dinámico (Gliessman 1997).
En las regiones tropicales la sucesión secundaria es un proceso complejo y múltiple,
que puede considerarse más probabilístico que predecible, y que puede desarrollarse
por diferentes rutas desde la fase pionera hasta la madurez. Es común que en los
sistemas agrícolas haya una manipulación continua de la vegetación por medios
culturales como la labranza o el chapeo. Dados los bajos rendimientos de estas
especies es fundamental la introducción de especies forrajeras con el fin de
incrementar la diversidad de ecosistema (Caamal y Armendariz 2002).

Para enfrentar el impacto descrito sobre los ecosistemas, la estrategia debería ser la
creación de un eje de continuidad entre estos y los sistemas de producción
ganaderos; éstos últimos tienen que jugar un papel fundamental en la protección y
restauración de la biodiversidad regional y global de acuerdo con el manejo del
sistema.
Moverse de un agroecosistema uniforme, a uno más diverso y sostenible, no es un
proceso fácil ni inmediato. Por ello, los esfuerzos de reconversión deben realizarse
paso a paso hasta llegar a la meta de la sostenibilidad mediante la diversificación.

CAPÍTULO III
CARACTERIZACIÓN DEL PENNISETUM PURPUREUM VC MARALFALFA EN LA
AMAZONÍA ECUATORIANA
III. 1 Introducción
La pradera es un ente dinámico que requiere de un manejo estratégico para
mantener e incrementar la producción animal. La estacionalidad en la producción de
forraje y la variación en su valor nutritivo son factores que limitan la producción animal
en las zonas tropicales. El manejo eficiente de las especies forrajeras es primordial
para mantener una alta productividad y calidad del forraje, sin propiciar el deterioro de
la pradera (Martínez et al. 2008).
Los avances alcanzados en el campo de la nutrición hacen necesario el
conocimiento, cada día más preciso, del valor alimenticio de los pastos y forrajes, los
cuales constituyen en el trópico la mayor fuente de alimentación de los animales y la
más económica por lo que es sumamente importante conocer el valor alimenticio de
los diferentes forrajes verdes o conservados que pueden formar parte de la ración y
que permite exteriorizar el potencial máximo de producción de los animales (Cáceres
y González 2000).
Las condiciones climáticas de la Amazonía Ecuatoriana se separan de las del resto
del país y se caracterizan fundamentalmente por mayores precipitaciones, cuando en
el resto si son apreciables las estaciones del año.
En este sentido, las expresiones matemáticas pueden ser de gran utilidad para
explicar y predecir el comportamiento de especies de pastos y forrajes ante
determinados factores, como son las lluvias, la humedad, la temperatura, así como la
relación funcional de estos con la edad, al ser una variedad introducida en la
Amazonía Ecuatoriana. Estos estudios son limitados en esta área geográfica.
Por ello, el objetivo del presente capítulo están relacionados con: a) caracterizar y
conocer la relación funcional de la edad con el rendimiento y calidad del Pennisetum
purpureum vc Maralfalfa, así como la influencia de elementos climáticos en la
producción y calidad de esta especie.

III. 2 Materiales y Métodos
III. 2.1 Manejo del Experimento
Se realizó un experimento en las áreas perteneciente al Centro de Investigación,
Postgrado y Conservación de la Biodiversidad de la Amazonia Ecuatoriana (CIPCA),
para evaluar el comportamiento del Pennisetum purpureum vc Maralfalfa. Los datos
climáticos del lugar fueron descritos en el capítulo anterior.
III. 2.2. Preparación del Terreno
Se inició con la selección del terreno; topografía regular, libre de excesiva sombra y
encharcamiento. Fue necesario un muestreo de suelo para el análisis químico del
mismo descrito en el capítulo anterior (tabla 2). Anteriormente se trataba de un
bosque secundario donde todavía se podía visualizar troncos, se procedió a
destroncar (manualmente) y limpiar de malezas existentes, dejando completamente
limpio el terreno.
Se cercó el área total de la parcela con alambre de púa, se realizó la separación de
unidades experimentales con sus respectivos caminos de 1m de ancho. Se realizó el
sorteo de unidades experimentales según el diseño de bloques completos al azar.
La remoción de suelo se efectuó manualmente utilizando azadones en vista de no ser
posible el uso de mecanización, seguido de la aplicación de enmienda previa al
análisis de suelo, posteriormente rastrillado, nivelación y surcada a una profundidad
comprendida entre 3 y 5 cm de acuerdo a las distancias anteriormente ya señaladas
(0.50m y 1m)
Al finalizar la preparación, la parcela tuvo las siguientes características: largo de 22 m
y un ancho de 43 m, el área total es de 946m2. Donde cada unidad experimental
tiene una forma rectangular de 5m de ancho y 6m de largo con un área de 30m2,
separados por caminos de 1m de ancho tanto las unidades experimentales entre sí
como los bloques.

III. 2.3. Siembra
El corte y transporte de material de propagación tuvo lugar un día anterior a la
siembra, desde el cantón Mera-Pastaza, de una pradera establecida hace
aproximadamente 3 años. Las estacas fueron seleccionadas y cortadas de la parte
media de la caña (tallo) de Maralfalfa con 3 nudos, con un largo entre 0.30 a 0.50m
y peso entre 40 y 70 gramos.
La plantación o siembra de las estacas seleccionadas se efectuó depositando
respectivamente en los surcos de las parcelas según el número de estacas,
distancias correspondientes en base al sorteo anteriormente desarrollado. Se realizó
una desinfección del material vegetativo utilizando Vitavax a razón de 10g/l de agua a
través de una bomba de fumigar, para evitar el ataque de posibles plagas o
enfermedades.
Se hizo un conteo de número de estacas en cada unidad experimental con su
correspondiente peso e identificación mediante tarjetas individuales para cada unidad
experimental.
III. 2.4. Toma de muestras
La altura y número de hojas por tallo se midió a partir de la cuarta semana hasta el
final del experimento. Se consideró el número de tallos por macollo, diámetro de
macollo y diámetro de cobertura aérea, largo y ancho de las hojas (parte media) y
diámetro de tallo (cuarto entre nudo) con una frecuencia de 15 días. Las mediciones
se realizaron utilizando un flexómetro.
El rendimiento se midió a las diferentes edades de corte, mediante el corte de 1m2 de
cada unidad experimental, donde se efectuó el pesaje total, la relación hoja-tallo y
material muerto, mediante una balanza digital. Para analizar la composición química
se envió una muestra representativa (1Kg) de pasto verde de planta completa de
cada tratamiento, al laboratorio de Nutrición y Bromatología de la Escuela Superior
Politécnica de Chimborazo (ESPOCH).
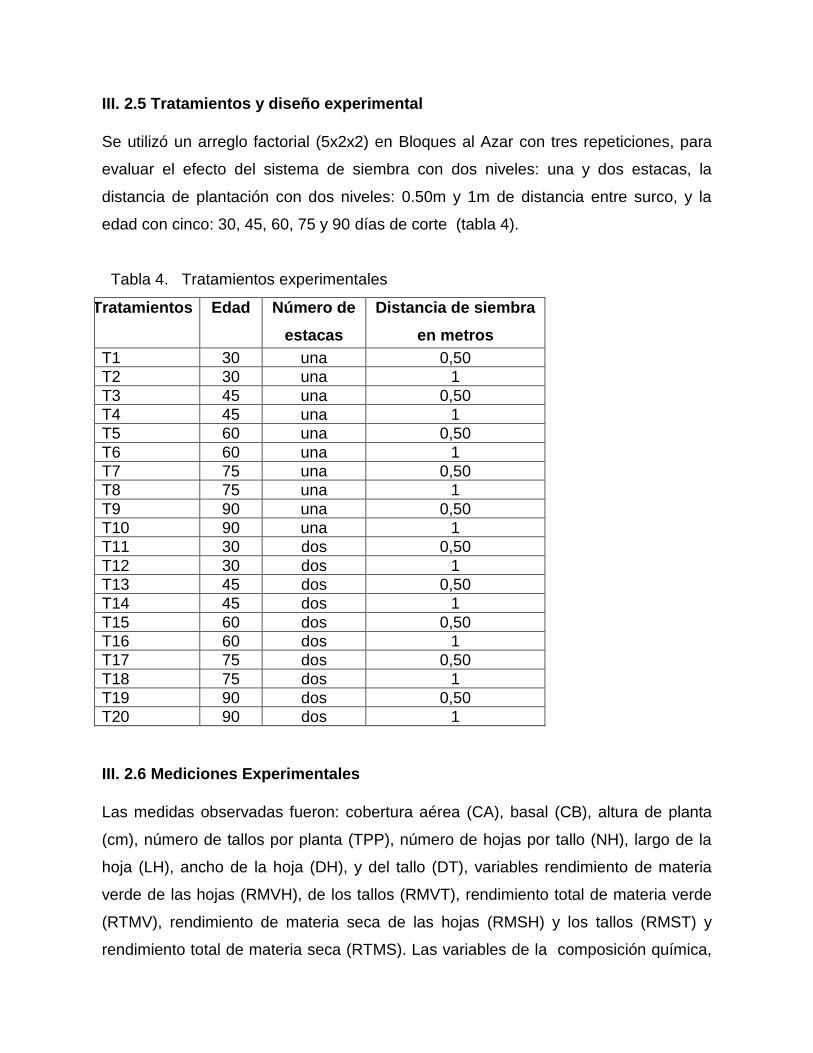
III. 2.5 Tratamientos y diseño experimental
Se utilizó un arreglo factorial (5x2x2) en Bloques al Azar con tres repeticiones, para
evaluar el efecto del sistema de siembra con dos niveles: una y dos estacas, la
distancia de plantación con dos niveles: 0.50m y 1m de distancia entre surco, y la
edad con cinco: 30, 45, 60, 75 y 90 días de corte (tabla 4).
Tabla 4. Tratamientos experimentales
Tratamientos
Edad Número de
estacas
Distancia de siembra
en metros
T1 30 una 0,50
T2 30 una 1
T3 45 una 0,50
T4 45 una 1
T5
60 una 0,50
T6 60 una 1
T7 75 una 0,50
T8 75 una 1
T9 90 una 0,50
T10 90 una 1
T11 30 dos 0,50
T12 30 dos 1
T13 45 dos 0,50
T14 45 dos 1
T15 60 dos 0,50
T16 60 dos 1
T17 75 dos 0,50
T18 75 dos 1
T19 90 dos 0,50
T20 90 dos 1
III. 2.6 Mediciones Experimentales
Las medidas observadas fueron: cobertura aérea (CA), basal (CB), altura de planta
(cm), número de tallos por planta (TPP), número de hojas por tallo (NH), largo de la
hoja (LH), ancho de la hoja (DH), y del tallo (DT), variables rendimiento de materia
verde de las hojas (RMVH), de los tallos (RMVT), rendimiento total de materia verde
(RTMV), rendimiento de materia seca de las hojas (RMSH) y los tallos (RMST) y
rendimiento total de materia seca (RTMS). Las variables de la composición química,

fueron proteína bruta (PB), fibra bruta (FB), grasa bruta (GB), extracto libre de
nitrógeno (ELN), ceniza (Cen), materia seca (MS) y materia orgánica (MO).
III.2.7 Análisis Estadístico y cálculos
Los datos se analizaron según ANOVA, se utilizó el sistema “Statistica versión 8,1”
para Windows. Para comprobar la normalidad de los datos se utilizó la prueba
Kolmogorov Smirnov, la de Bartlet para la homogeneidad de las varianzas, y
Newman–Keuls para determinar diferencias entre las medias. El siguiente modelo
fue utilizado para este análisis: Yijk = µ + ai + sj + spk + (as)ij+(asp) ik + (ssp)jk+(assp)ijk+
eijk, donde Yijk es la variable número de estaca, edad, y distancia de plantación, µ es la
media, ai el número de estacas, sj, la edad, spk distancia de siembra y eijk el error.
Para establecer la relación funcional entre el rendimiento y la edad, se analizaron las
ecuaciones de regresión (lineales, cuadráticas, cúbicas, logarítmicas y Gompertz) y
se utilizó el método descendente. Para la selección de la ecuación de mejor ajuste se
consideró alto valor de R2, alta significación, bajo error estándar de los términos y de
estimación, menor cuadrado medio del error, aporte significativo de los términos y de
la ecuación y bajo coeficiente de indeterminación (1-R2).
Se realizó análisis de correlación lineal entre los indicadores del rendimiento y la
calidad (variables dependientes) y los elementos del clima (variables independientes)
para los tratamientos a 0,50 y un metro sembrados con una estaca acostada. Se
informaron sólo los R2. A partir de estos resultados se establecieron ecuaciones
lineales múltiples entre el RMVH, RMVT, RTMV, RMSH, RMST, RTMS, PB, FB, Ca y
P y los factores del clima. Se informaron sólo los de mejor ajuste para cada
tratamiento.
III. 2.8 Procedimiento
El rendimiento se determinó mediante el corte total de la parcela en cada tratamiento.
Se tuvo en cuenta la composición botánica al expresar el rendimiento de la variedad
en estudio. Posterior al peso en verde de la parcela total, se separaron las hojas y los
tallos verdes, se pesaron de forma individual según Herrera (2006), se secaron en
una estufa de circulación de aire durante 72 horas a 65ºC y permitió determinar

rendimiento de estos. Para esto se emplearon 200g de cada muestra con 4 réplicas
por tratamiento.
III. 2.9 Composición química
Se determinó en los laboratorios de la Universidad Estatal Amazónica y con una
réplica en la Universidad Técnica de Ambato según:
Materia seca (MS): por deshidratación forzada en estufa de circulación a 65 ºC
durante 72 horas (AOAC 1995) y MO a 500 ºC en una mufla. Proteína bruta (PB): por
el método Kjeldhal (AOAC 1995). La FB, GB, ELN, P y Ca de acuerdo con AOAC
(1995).
SECCIÓN A. Caracterización morfológica, del rendimiento e indicadores
químicos y su relación funcional con la edad en el Pennisetum purpureum vc
Maralfalfa.
III.3 Resultados
Los resultados reflejaron que existe interacción entre los diferentes factores
evaluados para (P<0,001), para las variables cobertura área y basal, altura, número
de hojas y tallos por platón (tabla 5).
Tabla 5. Efecto de la edad, distancia de siembra y número de estaca en los
indicadores morfológicos.
Efecto gl CB (%) CA (%) Altura H (cm) NH(U) TPP(U) NE 15 *** *** *** *** ***
Edad 60 *** *** *** *** ***
DP 15 *** *** *** *** ***
NE x Edad 60 *** *** *** *** ***
NE x DP 15 *** *** *** *** ***
Edad x DP 60 *** *** *** *** ***
NE x Edad x DP 60 *** *** *** *** ***
R2 0,99 0,99 0,98 0,99 0,99
X 53,46 133,14 150,42 9,97 6,66
EE± 1,87 6,93 12,40 0,20 0,21 ***P<0,001 gl=grados de libertad, R
2 coeficiente de determinación del modelo

Para las variables largo y ancho de las hojas, diámetro del tallo, rendimiento de
materia verde de hojas y tallos ocurrió algo similar, destacándose los bajos errores
estándares, y altos coeficientes, lo que indica que los factores que influyen en este
comportamiento fueron considerados en el modelo (tabla 6).
Tabla 6 Efecto de la edad, distancia de siembra y número de estaca en indicadores
morfológicos y del rendimiento en tMS/ha-1
Efecto gl LH (cm) AH (cm) DT (cm) RMVH
t/ha
RMVT
t/ha NE 15 *** *** *** *** ***
Edad 60 *** *** *** *** ***
DS 15 *** *** *** *** ***
NE x Edad 60 *** *** *** *** ***
NE x DS 15 *** *** *** *** ***
Edad x DS 60 *** *** *** *** ***
NE x Edad x DS 60 *** *** *** *** ***
R2 0,99 0,98 0,98 0,99 0,99
X 89,11 25,57 12,67 10,78 28,44
EE± 2,04 0,74 0,35 0,58 2,30 ***P<0,001 gl =grados de libertad, coeficiente de determinación del modelo
Por su parte, el rendimiento total de materia verde, de materia seca de hojas y tallos,
así como el total de materia seca reflejó, interacción entre todos los factores para
(P<0,001). Los bajos valores de los errores estándar y altos coeficientes, indicaron
nuevamente que la mayoría de los factores que influyen en este comportamiento
fueron considerados en el modelo (tabla 7).
Tabla 7. Efecto de la edad, distancia de siembra y número de estaca en indicadores
del rendimiento en tMS/ha-1
Efecto gl RTMV RMSH RMST RTMS NE 15 *** *** *** ***
Edad 60 *** *** *** ***
DS 15 *** *** *** ***
NE x Edad 60 *** *** *** ***
NE x DS 15 *** *** *** ***
Edad x DS 60 *** *** *** ***
NE x Edad x DS 60 *** *** *** ***
R2 0,98 0,98 0,99 0,99
X 39,23 2,17 5,70 7,86
EE± 2,60 0,11 0,46 0,52 ***P<0,001 gl =grados de libertad, coeficiente de determinación del modelo

Los diferentes indicadores químicos evaluados reflejaron interacción entre los
factores con diferencias significativas para (P<0,001). Al igual que para las variables
morfológicas y de rendimiento, los errores estándar fueron bajos (tablas 8 y 9).
Tabla 8 Efecto de la edad, distancia de siembra y número de estaca en el porcentaje
de los indicadores químicos.
Efecto gl Cen MO PB FB GB NE 15 *** *** *** *** ***
Edad 60 *** *** *** *** ***
DS 15 *** *** *** *** ***
NE x Edad 60 *** *** *** *** ***
NE x DS 15 *** *** *** *** ***
Edad x DS 60 *** *** *** *** ***
NE x Edad x DS 60 *** *** *** *** ***
R2 0,83 0,83 0,89 0,89 0,90
X 6,81 93,19 9,86 32,42 2,14
EE± 0,19 0,19 0,40 0,63 0,05 ***P<0,001 gl =grados de libertad coeficiente de determinación del modelo
Tabla 9. Efecto de la edad, distancia de siembra y número de estaca en el porcentaje
de los indicadores químicos.
Efecto gl ELN P Ca MS NE 15 *** *** *** ***
Edad 60 *** *** *** ***
DS 15 *** *** *** ***
NE x Edad 60 *** *** *** ***
NE x DS 15 *** *** *** ***
Edad x DS 60 *** *** *** ***
NE x Edad x DS 60 *** *** *** ***
R2 0,99 0,82 0,97 0,95
X 48,77 0,82 0,44 20,05
EE± 0,38 0,04 0,01 0,007 ***P<0,001 gl =grados de libertad, coeficiente de determinación del modelo
Partiendo de la información del análisis de varianza se establecieron las
comparaciones entre la medias al interaccionar los tres factores. Para las variables
cobertura basal y aérea, número de hojas y tallos por plantó, los mayores valores se
presentaron para la edad de 90 días al emplear dos estacas a un metro de distancia,
con diferencias significativas con los restantes tratamientos. En lo referente a la altura
de la planta, ocurrió al utilizar dos estacas a 0,50 m a la edad de 90 días, aunque en
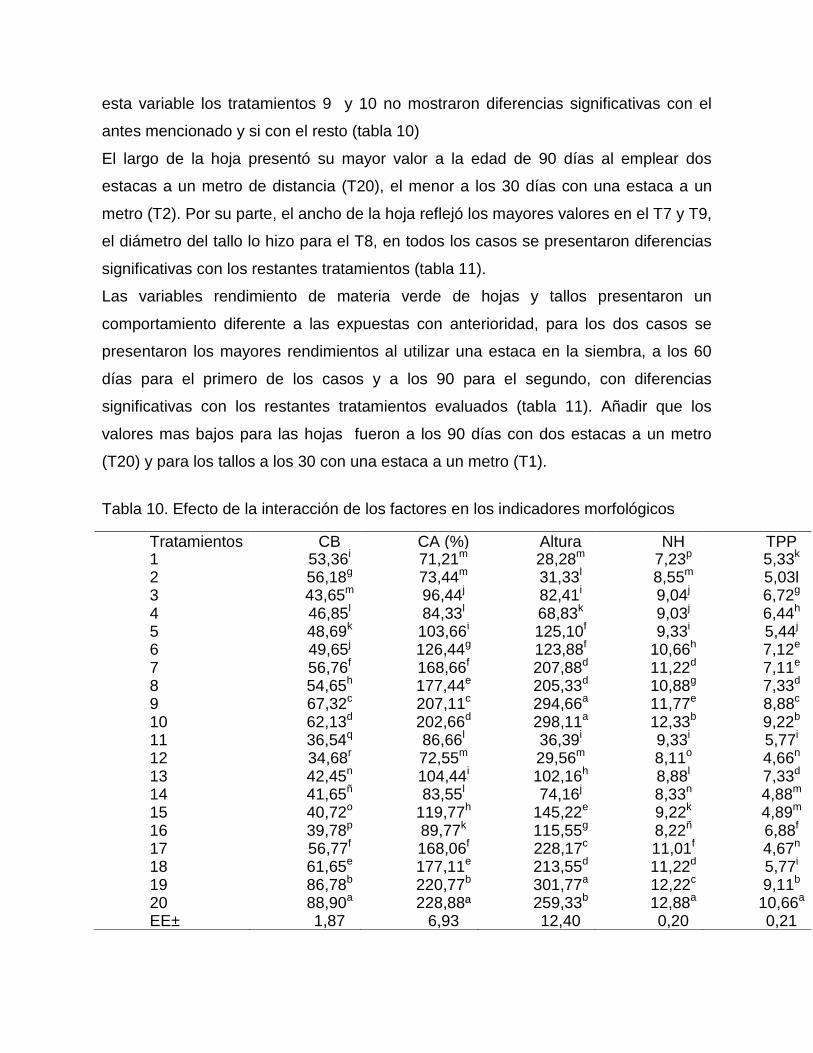
esta variable los tratamientos 9 y 10 no mostraron diferencias significativas con el
antes mencionado y si con el resto (tabla 10)
El largo de la hoja presentó su mayor valor a la edad de 90 días al emplear dos
estacas a un metro de distancia (T20), el menor a los 30 días con una estaca a un
metro (T2). Por su parte, el ancho de la hoja reflejó los mayores valores en el T7 y T9,
el diámetro del tallo lo hizo para el T8, en todos los casos se presentaron diferencias
significativas con los restantes tratamientos (tabla 11).
Las variables rendimiento de materia verde de hojas y tallos presentaron un
comportamiento diferente a las expuestas con anterioridad, para los dos casos se
presentaron los mayores rendimientos al utilizar una estaca en la siembra, a los 60
días para el primero de los casos y a los 90 para el segundo, con diferencias
significativas con los restantes tratamientos evaluados (tabla 11). Añadir que los
valores mas bajos para las hojas fueron a los 90 días con dos estacas a un metro
(T20) y para los tallos a los 30 con una estaca a un metro (T1).
Tabla 10. Efecto de la interacción de los factores en los indicadores morfológicos
Tratamientos CB
(%)
CA (%) Altura
(cm)
NH
(U)
TPP
(U) 1 53,36i 71,21m 28,28m 7,23p 5,33k
2 56,18g 73,44m 31,33l 8,55m 5,03l 3 43,65m 96,44j 82,41i 9,04j 6,72g
4 46,85l 84,33l 68,83k 9,03j 6,44h
5 48,69k 103,66i 125,10f 9,33i 5,44j
6 49,65j 126,44g 123,88f 10,66h 7,12e
7 56,76f 168,66f 207,88d 11,22d 7,11e
8 54,65h 177,44e 205,33d 10,88g 7,33d
9 67,32c 207,11c 294,66a 11,77e 8,88c
10 62,13d 202,66d 298,11a 12,33b 9,22b
11 36,54q 86,66l 36,39i 9,33i 5,77i
12 34,68r 72,55m 29,56m 8,11o 4,66n
13 42,45n 104,44i 102,16h 8,88l 7,33d
14 41,65ñ 83,55l 74,16j 8,33n 4,88m
15 40,72o 119,77h 145,22e 9,22k 4,89m
16 39,78p 89,77k 115,55g 8,22ñ 6,88f
17 56,77f 168,06f 228,17c 11,01f 4,67n
18 61,65e 177,11e 213,55d 11,22d 5,77i
19 86,78b 220,77b 301,77a 12,22c 9,11b
20 88,90a 228,88ª 259,33b 12,88a 10,66a
EE± 1,87 6,93 12,40 0,20 0,21

Letras desiguales en la misma columna difieren para P<0,01
Tabla 11. Efecto de la interacción de los factores en los indicadores morfológicos
y del rendimiento.
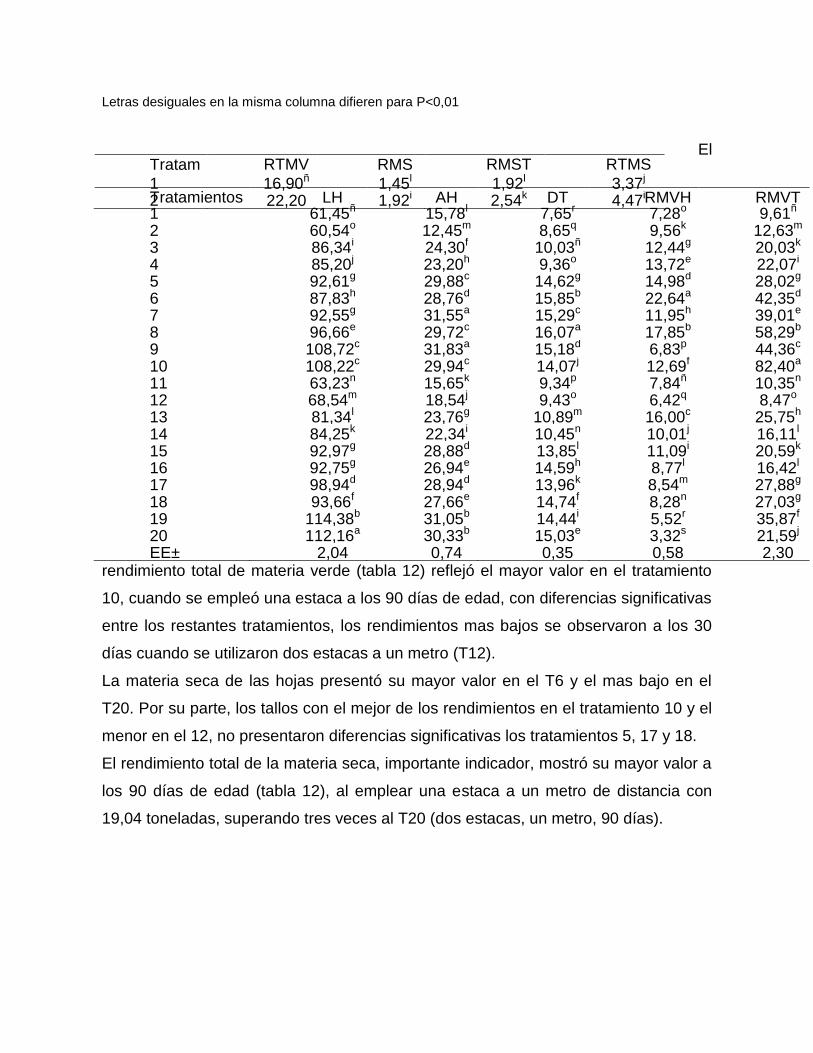
Letras desiguales en la misma columna difieren para P<0,01
El
rendimiento total de materia verde (tabla 12) reflejó el mayor valor en el tratamiento
10, cuando se empleó una estaca a los 90 días de edad, con diferencias significativas
entre los restantes tratamientos, los rendimientos mas bajos se observaron a los 30
días cuando se utilizaron dos estacas a un metro (T12).
La materia seca de las hojas presentó su mayor valor en el T6 y el mas bajo en el
T20. Por su parte, los tallos con el mejor de los rendimientos en el tratamiento 10 y el
menor en el 12, no presentaron diferencias significativas los tratamientos 5, 17 y 18.
El rendimiento total de la materia seca, importante indicador, mostró su mayor valor a
los 90 días de edad (tabla 12), al emplear una estaca a un metro de distancia con
19,04 toneladas, superando tres veces al T20 (dos estacas, un metro, 90 días).
Tratamientos LH
(cm)
AH
(cm)
DT
(cm)
RMVH
t/ha-1
(tt/ha
RMVT
t/ha-1 1 61,45ñ 15,78l 7,65r 7,28o 9,61ñ
2 60,54o 12,45m 8,65q 9,56k 12,63m
3 86,34i 24,30f 10,03ñ 12,44g 20,03k
4 85,20j 23,20h 9,36o 13,72e 22,07i
5 92,61g 29,88c 14,62g 14,98d 28,02g
6 87,83h 28,76d 15,85b 22,64a 42,35d
7 92,55g 31,55a 15,29c 11,95h 39,01e
8 96,66e 29,72c 16,07a 17,85b 58,29b
9 108,72c 31,83a 15,18d 6,83p 44,36c
10 108,22c 29,94c 14,07j 12,69f 82,40a
11 63,23n 15,65k 9,34p 7,84ñ 10,35n
12 68,54m 18,54j 9,43o 6,42q 8,47o
13 81,34l 23,76g 10,89m 16,00c 25,75h
14 84,25k 22,34i 10,45n 10,01j 16,11l
15 92,97g 28,88d 13,85l 11,09i 20,59k
16 92,75g 26,94e 14,59h 8,77l 16,42l
17 98,94d 28,94d 13,96k 8,54m 27,88g
18 93,66f 27,66e 14,74f 8,28n 27,03g
19 114,38b 31,05b 14,44i 5,52r 35,87f
20 112,16a 30,33b 15,03e 3,32s 21,59j
EE± 2,04 0,74 0,35 0,58 2,30
Tratam
ientos
RTMV RMS
H
RMST RTMS
1 16,90ñ 1,45l 1,92l 3,37j
2 22,20m
1,92i 2,54k 4,47i

Tabl
a 12.
Efect
o de
la
inter
acció
n de
los
facto
res
en
los
indic
ador
es
del
rendimiento en t/ha-1/corte.
Letras desiguales en la misma columna difieren para P<0,01
3 32,47i 2,49f 4,01i 6,50g
4 35,79h 2,76e 4,44h 7,19f
5 43,00e 2,99d 5,60g 8,60e
6 65,00c 4,53a 8,48d 13,02c
7 50,96d 2,58f 7,79e 10,18d
8 76,14b 3,59b 11,72b 15,31b
9 51,20d 1,37m 8,90c 10,27d
10 95,13a 2,54f 16,49a 19,04a
11 18,20n 1,57k 2,07l 3,63j
12 14,87o 1,29 1,71m 3,00k
13 41,76f 3,20c 5,16g 8,37e
14 26,13k 2,01h 3,22j 5,23h
15 31,60j 2,21g 4,13i 6,34g
16 25,20l 1,76j 3,29j 5,05h
17 36,42g 1,70j 5.57g 7,28f
18 35,32h 1,67j 5,45g 7,11f
19 41,40f 1,11ñ 7,20f 8,30e
20 24,92l 0,67o 4,33h 5,01h
EE± 2,60 0,11 0,46 0,52

Al evaluar los indicadores químicos (tabla 13), la ceniza mostró los mayores
porcentajes en los tratamientos T6 (60) y T12 (30 días), en ambos la distancia de
siembra fue similar, sin embargo, las estacas variaron, una y dos, respectivamente.
La materia orgánica alcanzó su mayor valor para el T2 (30 días, una estaca, a un
metro). Por su parte, la proteína bruta reflejó los mayores tenores a los 30 días de
edad (tabla 13) al emplear una estaca en la plantación, aunque en las dos distancias
utilizadas (tratamientos 1 y 2), mostrando diferencias con los restantes. Destacar que
para el caso de T5, 6, 8, 12 y 18, no se apreciaron diferencias significativas. En solo
dos tratamientos la proteína estuvo por debajo de 7%.
La fibra bruta reflejó los mayores porcentajes para T8 y T19, con diferencias
significativas entre las restantes. La graso bruta lo hizo para T7 y los tratamientos 1,
3, 6, 9, 10, 11 y 18, nos mostraron diferencias significativas entre ellos (tabla 13).
Tabla 13. Efecto de la interacción de los factores en los indicadores químicos
expresados en %.
Tratamientos Ceniza MO PB FB GB

Letras desiguales en la misma columna difieren para P<0,01
El extracto libre de nitrógeno no estuvo definido por la distancia, número de estacas y
edad (tabla 14), ya que los mayores porcentajes se mostraron para T7 y T12, el mas
bajo fue para el T3. El comportamiento del Calcio estuvo definido por el empleo de
una sola estaca, varió la edad y la distancia, ya que los mayores valores los
alcanzaron los tratamientos 3, 7, 9 y 10. El fósforo, mostró los mejores resultados en
los tratamientos 7, 8, 15, 17 y 18, las diferencias significativas fueron variables, no se
mostró un comportamiento definido, según la edad, el número de estacas o distancia
Para la materia seca ocurrió algo similar, destacar que el menor porcentaje apareció
a los 75 días, cuando se emplearon dos estacas a 0,50m (tabla 14).
Tabla 14. Efecto de la interacción de los factores en los indicadores químicos
expresados en %.
1 7,72c 92,27h 16,03a 24,56j 2,36de
2 6,30f 97,71a 16,55a 24,62j 2,66b
3 8,59b 91,40i 11,38d 37,74b 2,23e
4 7,09d 92,90g 11,56d 31,17g 1,90fg
5 7,24d 92,76g 7,93h 31,70g 2,51c
6 8,63ab 91,37i 7,76h 35,70d 2,30e
7 6,40f 93,61e 7,92h 30,47h 2,96a
8 7,29d 92,70g 7,75h 39,59a 1,55hi
9 6,72e 93,27e 6,03i 37,30b 2,27e
10 4,25h 95,70c 7,58h
36,92bc 2,25e
11 7,17d 92,82g 13,97c 28,34i 2,28e
12 8,84a 91,15i 14,48b 23,50k 1,47i
13 6,49f 93,50e 9,83f 30,57h 1,80g
14 6,82e 93,17f 10,87e 28,55 2,00f
15 7,64c 92,40h 8,45g 32,75f 2,75b
16 6,48f 93,53e 7,92h 34,51e 1,85g
17 2,75i 97,30b 8,46g 36,68c 1,48i
18 7,73c 92,27h 7,93h 31,48g 2,23e
19 7,14d 92,85g 6,55i 39,74a 1,67h
20 5,35g 94,64d 8,68g 32,52f 2,21e
EE± 0,19 0,19 0,40 0,63 0,05
Tratam
ientos
ELN Ca P MS

Letras desiguales en la misma columna difieren para P<0,01
Los resultados anteriores permitieron delimitar que los indicadores tanto morfológicos,
del rendimiento, como químicos fueron de forma generales mayores al emplear una
estaca plantada a un metro. Partiendo de esta información se realizaron modelos de
ajuste con la edad para estas variables, aunque también se hicieron para una estaca
plantada a 0,50m, con el objetivo de relacionar estos indicadores con la edad de la
planta, factor en el cual se apreciaron altas diferencias significativas.
El rendimiento de materia verde para una estaca a 0,50m (método 1) y una estaca a
un metro (método 2) se incrementó con la edad (P<0.01) y se ajustaron ecuaciones
de regresión cuadráticas, los mayores valores se alcanzaron a los 90 días, en ambos
casos. El rendimiento de materia verde en el segundo de los casos superó en el
doble al uno para los 90 días (figura 9).
1 49,32e
f
0,27h 0,36d 19,97f
2 49,87e 0,39g 0,32d 20,11ab
3 40,06m
1,22a 0,53b 20,02e
4 48,27h 0,68f 0,51b 20,09b
5 50,63d 0,81d 0,45c 20,03d
6 45,61j 0,68f 0,36d 20,04d
7 52,26a 1,23a 0,64a 19,98f
8 43,82l 1,08b 0,62a 20,12ab
9 47,66i 1,23a 0,34d 20,06bc
10 48,78g 1,22a 0,46c 20,02e
11 48,23h 0,29h 0,38d 19,99f
12 51,79a
b 0,22i 0,23e 20,18a
13 51,30c 1,08b 0,41c 20,05cd
14 51,75b 0,82d 0,37d 20,01de
15 48,45g
h 0,81d 0,61a 20,09b
16 49,23f 0,94c 0,34d 20,06b
17 50,69d 0,95c 0,63a 17,98g
18 50,63d 0,79e 0,65a 20,16a
19 44,89k 1,09b 0,42c 20,07bc
20 51,29c 0,72e 0,35d 20,12ab
EE± 0,38 0,04 0,02 0,01

y = -0,0105x2 + 1,8465x - 29,16
R2 = 0,99
y = -0,0023x2 + 1,518x - 22,895
R2 = 0,98
0
10
20
30
40
50
60
70
80
90
100
0 10 20 30 40 50 60 70 80 90 100
Edad, días
t/ha/corte
tratamiento 3
tratamiento 1
Figura 9. Efecto de la edad de rebrote en la Rendimiento Total de Materia Verde de
los métodos 1 y 2.
La producción de materia seca se incrementó con la edad (P<0.01) en método (2) y
se ajustó una regresión cuadrática, el mayor valor se reflejó a los 90 días,
sobrepasando las 19 toneladas, duplicando al 1, este último también se ajustó a una
ecuación de regresión cuadrática (figura 10).
y = -0,0005x2 + 0,3073x - 4,646
R2 = 0,98
y = -0,0021x2 + 0,3681x - 5,8053
R2 = 0,99
0
2
4
6
8
10
12
14
16
18
20
0 10 20 30 40 50 60 70 80 90 100
Edad, días
t/ha/cortetratamiento 3
tratamiento 1
Figura 10. Efecto de la edad de rebrote en Rendimiento Total de Materia Seca de los
métodos 1 y 2.
t/ha-1
/corte
t/ha-1
/corte
♦método 1
■método 2
♦método 1
■método 2

Para el resto de las variables en el método 1, el comportamiento y el ajuste varió. Las
variables cobertura basal, largo de la hoja y tallos por plantón, se ajustaron a una
ecuación de regresión cúbica. Las restantes se ajustaron a una ecuación de regresión
cuadrática, excepto el número de hojas y el rendimiento de materia seca de los tallos,
que lo hicieron para una lineal. Los coeficientes en todos los casos sobrepasaron el
0,85, con alta significación para todas las ecuaciones (P<0,001). De forma general los
cuadrados medios del error y los errores estándar fueron bajos, lo que significa la
relación de la edad con las variables evaluadas (tabla 15).
Por otra parte, al establecer la relación funcional entre la edad y los indicadores
morfológicos y del rendimiento en el segundo caso, se pudo apreciar que ajustaron
ecuaciones de regresión cuadrática, excepto para la CB, CA, LH y DT, que fueron
cúbicas Para todos los casos los coeficientes de regresión fueron altos, superior a
0.80, con bajos errores estándar y cuadrados medios del error, lo que demuestra
como la variable independiente explica a la independiente (tabla 16).
Tabla 15. Relación entre la edad de rebrote y los indicadores morfológicos y de
rendimiento en el método 1.
Variabl
es
a b EE± c EE± d EE± R2
1-R2
CMe EE±
CB 132,65 -4,42 0,32 0,07 0,006 0,0001 0,0001 0,99 0,01 0,5 0,71
CA 76,04 -0,91 0,80 0,027 0,007 0,97 0,03 93,8 9,6
Altura -10,24 0,37 0,04 0,03 0,004 0,99 0,01 33,5 5,7
NH 5,21 0,02 0,004 0,95 0,05 0,14 0,36
TPP -4,22 0,59 0,2 -0,01 0,004 0,00006 0,00001 0,85 0,15 0,27 0,52
LH -122,09 10,21 0,29 -0,16 0,005 0,001 0,0001 0,99 0,01 0,41 0,64
AH -9,46 1,03 0,02 -0,006 0,0001 0,99 0,01 0,10 0,32
DT -4,48 0,47 0,06 -0,003 0,001 0,95 0,05 0,53 0,73
RMVH -14,86 0,98 0,03 -0,008 0,0001 0,98 0,02 0,15 0,39
RMVT -14,29 0,86 0,08 -0,002 0,001 0,99 0,01 0,96 0,98
RMSH -3,21 0,2 0,007 -0,002 0,0001 0,98 0,02 0,008 0,28
RMST -1,44 0,11 0,003 0,99 0,01 0,26 0,25
R2
todos a p<0,001
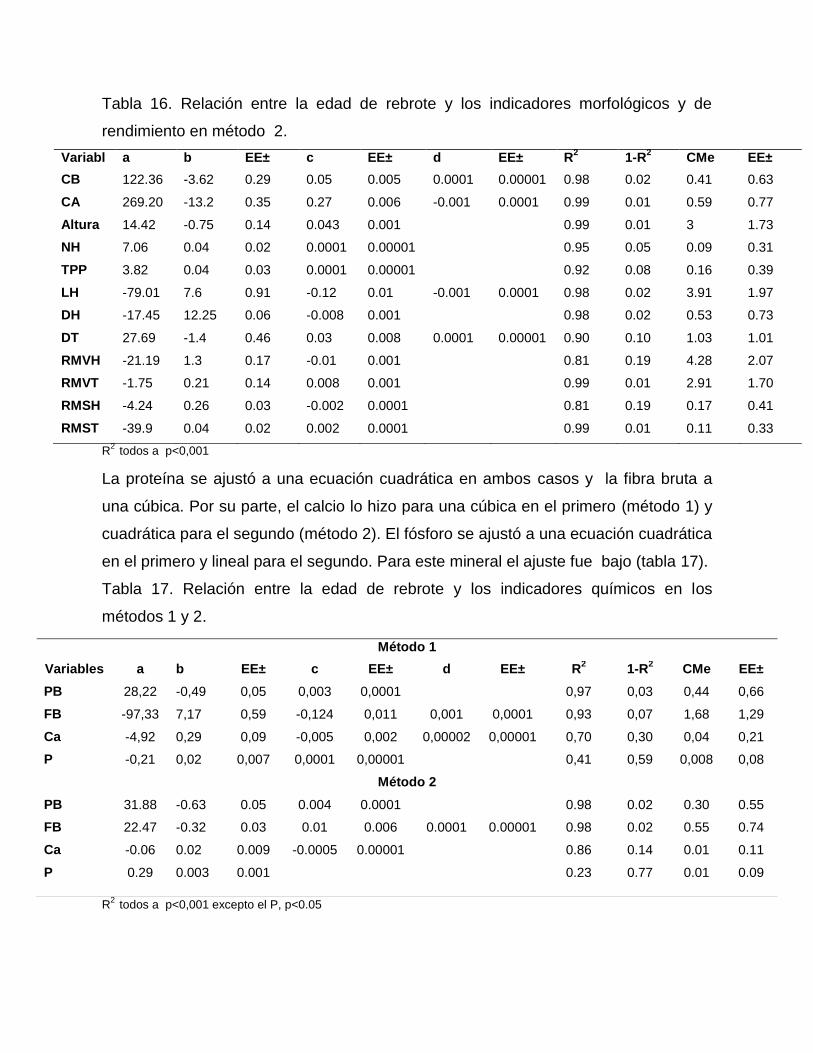
Tabla 16. Relación entre la edad de rebrote y los indicadores morfológicos y de
rendimiento en método 2.
Variabl
es
a b EE± c EE± d EE± R2
1-R2
CMe EE±
CB 122.36 -3.62 0.29 0.05 0.005 0.0001 0.00001 0.98 0.02 0.41 0.63
CA 269.20 -13.2 0.35 0.27 0.006 -0.001 0.0001 0.99 0.01 0.59 0.77
Altura 14.42 -0.75 0.14 0.043 0.001 0.99 0.01 3 1.73
NH 7.06 0.04 0.02 0.0001 0.00001 0.95 0.05 0.09 0.31
TPP 3.82 0.04 0.03 0.0001 0.00001 0.92 0.08 0.16 0.39
LH -79.01 7.6 0.91 -0.12 0.01 -0.001 0.0001 0.98 0.02 3.91 1.97
DH -17.45 12.25 0.06 -0.008 0.001 0.98 0.02 0.53 0.73
DT 27.69 -1.4 0.46 0.03 0.008 0.0001 0.00001 0.90 0.10 1.03 1.01
RMVH -21.19 1.3 0.17 -0.01 0.001 0.81 0.19 4.28 2.07
RMVT -1.75 0.21 0.14 0.008 0.001 0.99 0.01 2.91 1.70
RMSH -4.24 0.26 0.03 -0.002 0.0001 0.81 0.19 0.17 0.41
RMST -39.9 0.04 0.02 0.002 0.0001 0.99 0.01 0.11 0.33
R2
todos a p<0,001
La proteína se ajustó a una ecuación cuadrática en ambos casos y la fibra bruta a
una cúbica. Por su parte, el calcio lo hizo para una cúbica en el primero (método 1) y
cuadrática para el segundo (método 2). El fósforo se ajustó a una ecuación cuadrática
en el primero y lineal para el segundo. Para este mineral el ajuste fue bajo (tabla 17).
Tabla 17. Relación entre la edad de rebrote y los indicadores químicos en los
métodos 1 y 2.
R2
todos a p<0,001 excepto el P, p<0.05
Método 1
Variables a b EE± c EE± d EE± R2
1-R2
CMe EE±
PB 28,22 -0,49 0,05 0,003 0,0001 0,97 0,03 0,44 0,66
FB -97,33 7,17 0,59 -0,124 0,011 0,001 0,0001 0,93 0,07 1,68 1,29
Ca -4,92 0,29 0,09 -0,005 0,002 0,00002 0,00001 0,70 0,30 0,04 0,21
P -0,21 0,02 0,007 0,0001 0,00001 0,41 0,59 0,008 0,08
Método 2
PB 31.88 -0.63 0.05 0.004 0.0001 0.98 0.02 0.30 0.55
FB 22.47 -0.32 0.03 0.01 0.006 0.0001 0.00001 0.98 0.02 0.55 0.74
Ca -0.06 0.02 0.009 -0.0005 0.00001 0.86 0.14 0.01 0.11
P 0.29 0.003 0.001 0.23 0.77 0.01 0.09
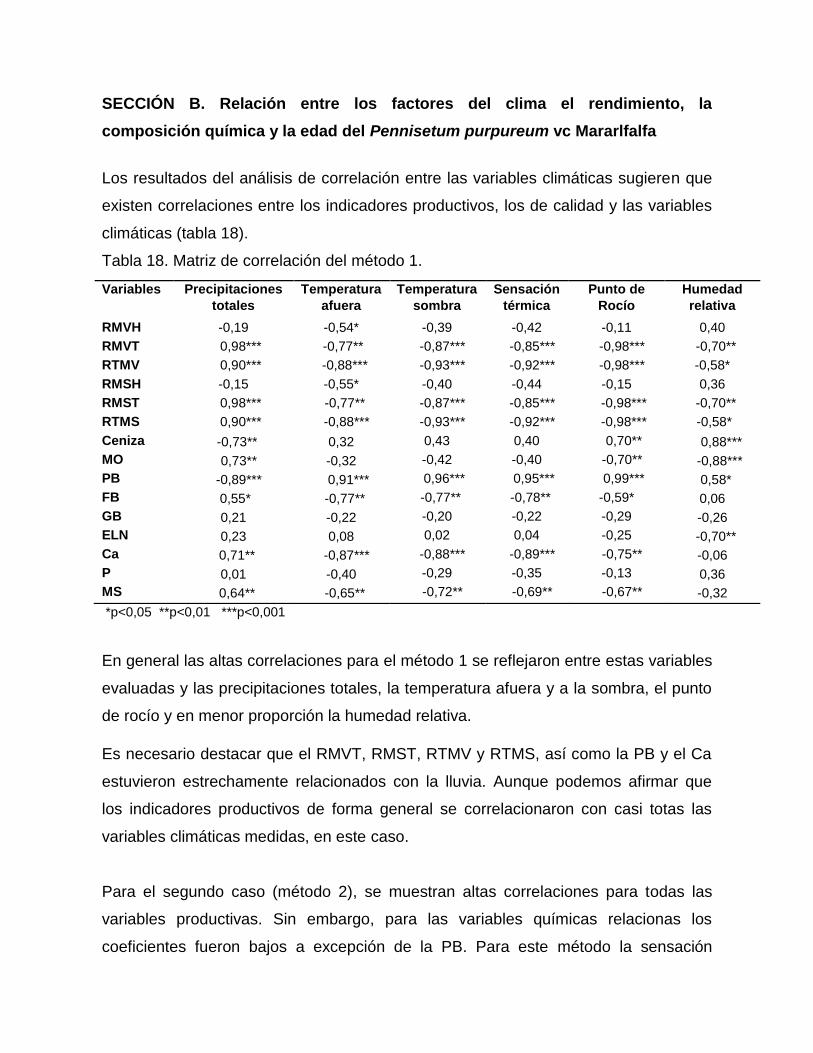
SECCIÓN B. Relación entre los factores del clima el rendimiento, la
composición química y la edad del Pennisetum purpureum vc Mararlfalfa
Los resultados del análisis de correlación entre las variables climáticas sugieren que
existen correlaciones entre los indicadores productivos, los de calidad y las variables
climáticas (tabla 18).
Tabla 18. Matriz de correlación del método 1.
*p<0,05 **p<0,01 ***p<0,001
En general las altas correlaciones para el método 1 se reflejaron entre estas variables
evaluadas y las precipitaciones totales, la temperatura afuera y a la sombra, el punto
de rocío y en menor proporción la humedad relativa.
Es necesario destacar que el RMVT, RMST, RTMV y RTMS, así como la PB y el Ca
estuvieron estrechamente relacionados con la lluvia. Aunque podemos afirmar que
los indicadores productivos de forma general se correlacionaron con casi totas las
variables climáticas medidas, en este caso.
Para el segundo caso (método 2), se muestran altas correlaciones para todas las
variables productivas. Sin embargo, para las variables químicas relacionas los
coeficientes fueron bajos a excepción de la PB. Para este método la sensación
Variables Precipitaciones
totales
Temperatura
afuera
Temperatura
sombra
Sensación
térmica
Punto de
Rocío
Humedad
relativa
RMVH -0,19
-0,54*
-0,39
-0,42
-0,11
0,40
RMVT 0,98***
-0,77**
-0,87***
-0,85***
-0,98***
-0,70**
RTMV 0,90***
-0,88***
-0,93***
-0,92***
-0,98***
-0,58*
RMSH -0,15
-0,55*
-0,40
-0,44
-0,15
0,36
RMST 0,98***
-0,77**
-0,87***
-0,85***
-0,98***
-0,70**
RTMS 0,90***
-0,88***
-0,93***
-0,92***
-0,98***
-0,58*
Ceniza -0,73** 0,32 0,43
0,40
0,70**
0,88***
MO 0,73** -0,32 -0,42
-0,40
-0,70**
-0,88***
PB -0,89*** 0,91*** 0,96***
0,95***
0,99***
0,58*
FB 0,55* -0,77** -0,77**
-0,78**
-0,59*
0,06
GB 0,21 -0,22 -0,20
-0,22
-0,29
-0,26
ELN 0,23 0,08 0,02
0,04
-0,25
-0,70**
Ca 0,71** -0,87*** -0,88***
-0,89***
-0,75**
-0,06
P 0,01 -0,40 -0,29
-0,35
-0,13
0,36
MS 0,64** -0,65** -0,72**
-0,69**
-0,67**
-0,32
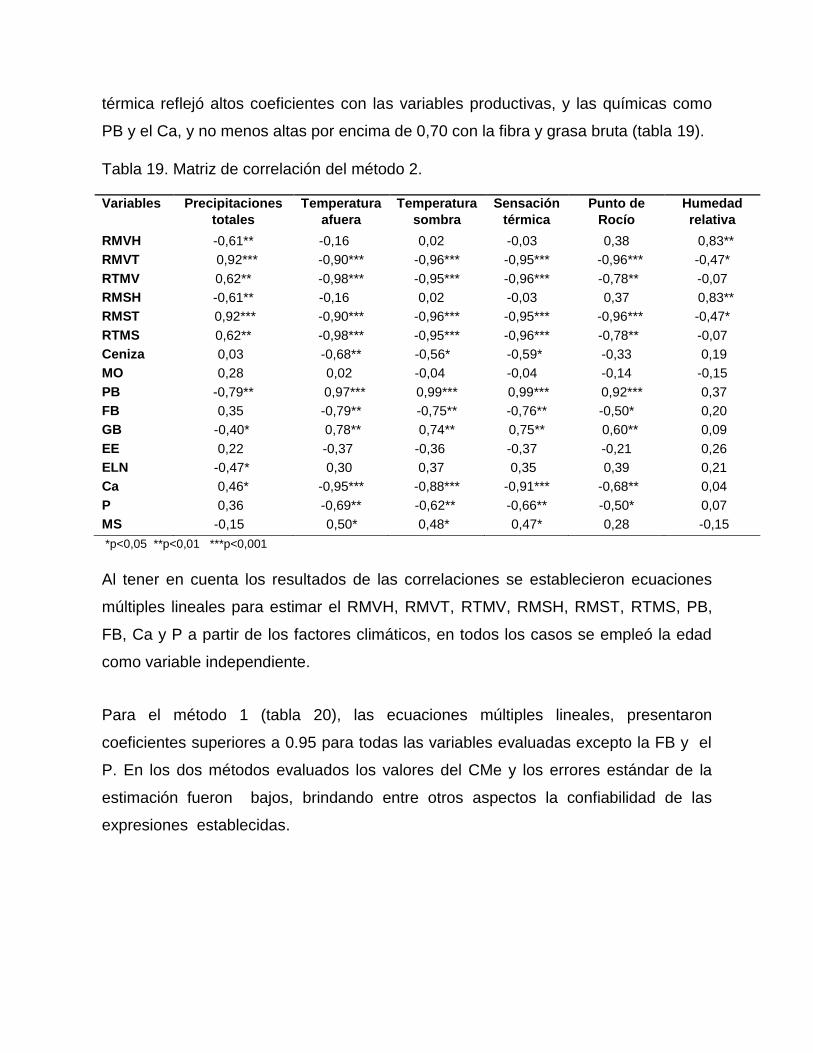
térmica reflejó altos coeficientes con las variables productivas, y las químicas como
PB y el Ca, y no menos altas por encima de 0,70 con la fibra y grasa bruta (tabla 19).
Tabla 19. Matriz de correlación del método 2.
*p<0,05 **p<0,01 ***p<0,001
Al tener en cuenta los resultados de las correlaciones se establecieron ecuaciones
múltiples lineales para estimar el RMVH, RMVT, RTMV, RMSH, RMST, RTMS, PB,
FB, Ca y P a partir de los factores climáticos, en todos los casos se empleó la edad
como variable independiente.
Para el método 1 (tabla 20), las ecuaciones múltiples lineales, presentaron
coeficientes superiores a 0.95 para todas las variables evaluadas excepto la FB y el
P. En los dos métodos evaluados los valores del CMe y los errores estándar de la
estimación fueron bajos, brindando entre otros aspectos la confiabilidad de las
expresiones establecidas.
Variables Precipitaciones
totales
Temperatura
afuera
Temperatura
sombra
Sensación
térmica
Punto de
Rocío
Humedad
relativa
RMVH -0,61** -0,16 0,02 -0,03 0,38 0,83**
RMVT 0,92*** -0,90*** -0,96*** -0,95*** -0,96*** -0,47*
RTMV 0,62** -0,98*** -0,95*** -0,96*** -0,78** -0,07
RMSH -0,61** -0,16 0,02 -0,03 0,37 0,83**
RMST 0,92*** -0,90*** -0,96*** -0,95*** -0,96*** -0,47*
RTMS 0,62** -0,98*** -0,95*** -0,96*** -0,78** -0,07
Ceniza 0,03 -0,68** -0,56* -0,59* -0,33 0,19
MO 0,28 0,02 -0,04 -0,04 -0,14 -0,15
PB -0,79** 0,97*** 0,99*** 0,99*** 0,92*** 0,37
FB 0,35 -0,79** -0,75** -0,76** -0,50* 0,20
GB -0,40* 0,78** 0,74** 0,75** 0,60** 0,09
EE 0,22 -0,37 -0,36 -0,37 -0,21 0,26
ELN -0,47* 0,30 0,37 0,35 0,39 0,21
Ca 0,46* -0,95*** -0,88*** -0,91*** -0,68** 0,04
P 0,36 -0,69** -0,62** -0,66** -0,50* 0,07
MS -0,15 0,50* 0,48* 0,47* 0,28 -0,15

Tabla 20. Modelos lineales simples en Pennisetum purpureum vc Maralfalfa paral
método 1
R2 todos a P<0,001 a=factor independiente, b=edad, c=humedad y d=precipitaciones totales
Para el método 2 (tabla 21), las ecuaciones múltiples lineales, presentaron
coeficientes superiores a 0.95 para todas las variables evaluadas excepto el P. En los
dos casos evaluados los valores del CMe y los errores estándar de la estimación
fueron bajos, brindando entre otros aspectos la confiabilidad de las expresiones
establecidas (tablas 20 y 21).
Tabla 21. Modelos lineales simples en Pennisetum purpureum vc Maralfalfa
tratamiento 3.
R
2 todos a p<0,001
a=factor independiente, b=edad c=humedad para RMVT, RTMV, RMST, RTMS y las variables químicas para RMVH y RMSH, c=precipitaciones totales, d=precipitaciones para el resto, excepto el P que es sensación térmica.
Variables a b EE± c EE± d EE± R2
1-R2
CMe EE±
RMVH -539,9 0,93 0,04 6,4 0,7 -0,05 0,002 0,97 0,03 0,29 0,54
RMVT -283,6 0,8 0,07 3,24 1,24 -0,01 0,004 0,99 0,01 0,85 0,92
RTMV -823,6 1,75 0,04 9,65 0,77 -0,06 0,002 0,99 0,01 0,32 0,57
RMSH -111,4 0,19 0,006 1,32 0,10 -0,01 0,0003 0,98 0,02 0,006 0,08
RMST -56,67 0,16 0,01 0,64 0,23 -0,002 0,0008 0,99 0,01 0,03 0,17
RTMS -164,5 0,34 0,005 1,93 0,08 -0,01 0,0003 0,99 0,01 0,004 0,06
PB 230 -0,46 0,06 -2,46 1,02 0,01 0,003 0,95 0,05 0,58 0,76
FB -1397,6 0,09 0,03 11,81 0,69 0,31 0,001 2,80
Ca -114,32 0,02 0,001 1,36 0,02 0,0001 0,00001 0,99 0,01 0,0004 0,02
P -26,82 0,02 0,006 0,32 0,10 -0,001 0,0003 0,51 0,49 0,006 0,08
Variables a b EE± c EE± d EE± R2
1-R2
CMe EE±
RMVH -4.92 1.44 0.08 -0.07 0.004 0.96 0.04 0.93 0.96
RMVT 1211 1.025 0.03 -14.67 2.62 0.99 0.01 3.89 1.97
RTMV 1022 1.92 0.14 -12.41 0.27 -0.047 0.008 0.99 0.01 2.87 1.69
RMSH -0.98 0.29 0.016 -0.015 0.0009 0.96 0.04 0.03 0.18
RMST 240.5 0.20 0.006 -2.91 0.50 0.99 0.01 0.14 0.38
RTMS 202.24 0.39 0.02 -2.45 0.04 -0.009 0.001 0.99 0.01 0.09 0.30
PB 292.09 -0.62 0.04 -3.17 0.74 0.02 0.002 0.97 0.03 0.30 0.55
FB -393.11 0.92 0.05 4.81 0.93 -0.038 0.003 0.98 0.02 0.48 0.69
Ca -18.16 0.01 0.001 0.21 0.09 0.95 0.05 0.005 0.07
P -73.46 0.01 0.003 0.67 0.13 0.72 0.24 0.76 0.24 0.003 0.05

Discusión
El deterioro de los ecosistemas ganaderos requiere de profundas transformaciones
en su explotación, basadas en principios agroecológicos, donde estos se consideren
como un ecosistema y no como una simple gestión técnico-económica, de tal forma
que sus integrantes (plantas, animales, suelo, clima, hombre y tecnologías) estén
entrelazados sincrónicamente y permitan su utilización efectiva, eficiente y productiva
con resultados económicos, sociales y ambientales positivos. Esta nueva visión
requiere del conocimiento de las leyes que rigen el funcionamiento de cada uno de
los elementos y procesos que forman el sistema, con el propósito de maximizar el
flujo de energía y el reciclaje de materiales (del Pozo 2004).
Basado en este principio se estudió la especie Pennisetum purpureum vc Maralfalfa,
la cual podría ser de utilidad para los sistemas ganaderos de la Amazonía
Ecuatoriana, partiendo de su introducción y que no se dispone de suficiente
información sobre su comportamiento en las condiciones específicas de la Provincia
Pastaza.
Los resultados reflejaron diferencias significativas al interaccionar los factores. Las
variables cobertura aérea (CA), número de hojas (NH), tallos por plantón (TPP),
ancho de la hoja (AH), diámetro del tallo, rendimiento de materia verde y seca de las
hojas, tallos, y del total en cada caso, estuvieron definidos por la distancia y cantidad
de estacas empleadas, donde los mayores valores se presentaron para una estaca
con un metro. Este resultado se puedo deber a la menor competencia por los
nutrientes, principalmente los del suelo y la luz, ya que no constituye las
precipitaciones y humedad del suelo un factor limitante en la Provincia Pastaza. Los
trabajos de Corbea y Martínez (1982), hacen alusión que en las mayores distancias
de siembra, favorecen el crecimiento de la planta y una mejor respuesta.
En condiciones del Ecuador en alturas de 2740 msnm, los trabajos realizados por
Fabian (2009), reflejaron valores muy inferiores a los mostrados en esta investigación
en lo que altura y ancho de la hoja se refiere. Este autor evaluó diferentes clones
obtenidos en el Instituto de Ciencia Animal de Cuba, alcanzando, para el primero de

los casos 72,72 y para el segundo 81,50cm, para los clones 14 y 19,
respectivamente, a los 60 días de edad. Otras investigaciones desarrolladas en Cuba
por Ortiz (2008) en CT 115, dan a conocer valores de 71,2 cm para el largo de la
hoja, inferior a los de este trabajo.
Investigaciones recientes de Herrera et al. (2012) reflejaron alturas inferiores a las
mostradas aquí a las edades de 60 y 90 días, estos autores evaluaron diferentes
clones (115, 14, 18, 19, 20, 21, 24, 30, 36, 39, 41, 42 y 43), y determinaron que la
altura tiende a presentar menor variabilidad entre los clones que entre variedades y
especies, lo que está determinado, entre otros aspectos, por la individualidad
bioquímica y fisiológica de cada planta. Lo cual no sucedió en este trabajo, porque se
evaluó una sola especie. Para el caso del largo de la hojas los clones 115, 21, 34 y
36, mostraron valores similares a los reportados en esta investigación. Destacar que
difieren las variedades y condiciones experimentales, dándole gran plasticidad
ecológica a este género.
Vargas (1988) señaló que los mejores resultados de los indicadores morfológicos se
obtuvieron al evaluar el King grass a distancias de 0,80m sobre 0,60. Por su parte,
Andrade (2009) obtuvo resultados superiores en la altura y rendimiento del
Pennisetum purpureum vc Maralfalfa al emplear distancias de siembra de 0,80m en
el Ecuador, manifestando que en las mayores distancias existen menos
competencias por los nutrientes del suelo y la luz solar, cuando la humedad no es un
factor limitante, lo que explica los resultados expuestos en esta investigación.
Las restantes variables cobertura basal (CB), número y ancho de las hojas y
diámetro del tallo reflejaron su mayores valores al emplear dos estacas a la menor
distancia de siembra, lo que pudo estar favorecido por la competencia de las plantas
por los nutrientes. Sin embargo, esto repercutió en los menores rendimientos de la
especie al sembrarla a esta distancia.
Los trabajos realizados por Navas et al. (2013) al estudiar el Pennisetum purpurem
vc CT 115, no mostraron diferencias al comparar dos distancias de siembra de 70 y

110cm, está última cerca a la empleada en esta investigación (un metro). Estos
autores evaluaron la altura, la cobertura aérea, ancho y largo de la hoja y porcentaje
de hojas y tallos. Manifestando que las no diferencias encontradas se debió a las
pocas repeticiones realizadas en el experimento, cuestión que no sucedió en este
trabajo. Aunque es importante destacar que el género Pennisetum se caracteriza por
la elevada capacidad de convertir la energía luminosa en biomasa y su plasticidad
ecológica. Estos atributos lo convierten en un género con capacidad de producir alta
cantidad de forraje de aceptable calidad (Herrera y Ramos 2006).
En todos los casos que se interaccionó el número de estaca y la distancia de
siembra, se presentaron diferencias significativas. Para los rendimientos de hojas,
tallos y total, ya sea de materia verde y seca, los mejores resultados aparecen para la
última edad evaluada, y muy superior al emplear una estaca a un metro que dos a
esta misma distancia. Resultados superiores a los reportados por los autores (Ortiz
2008, Fabian 2009, Herrera et al. 2012 y Navas et al. 2013).
Murillo et al (2015) en condiciones del Ecuador (provincia de Quevedo), obtuvieron
alturas de 185cm para el cultivar Maralfalfa a los 60 días de edad, con una
disminución del rendimiento de las hojas al incrementar la madurez de la planta.
Sobre este comportamiento Calzada et al (2014) plantearon que de forma general
en las gramíneas forrajeras tropicales cuando avanza la madurez hay una drástica
reducción de la relación hoja-tallo, posiblemente debido al efecto de factores como la
madurez de la planta, la temperatura, la humedad relativa y el manejo del material,
entre otros.
La proporción de las hojas respecto a los tallos es un indicador que permiten
establecer la composición del rendimiento total de la planta, ya que las hojas
muestran: a) alta probabilidad de incrementar el proceso fotosintético, b) mayor
posibilidad de producción de sustancia para el crecimiento, c) mejor acumulación de
reservas para el rebrote, d) mayor cantidad de nutrientes en las hojas que en los
tallos y e) el animal consume mayor cantidad de hojas que tallos (Herrera 2008). El
mayor rendimiento de hojas durante las primeras semanas de rebrote se puede

atribuir a la aparición de nuevos hijos y a la necesidad que tiene la planta de crear las
sustancias necesarias para su desarrollo (Romero et al. 1998). Sin embargo, su
disminución cuando avanza la edad, está asociada según Beliuchenco y Febles
(1980) al incremento del grosor y la longitud del tallo. Esto justifica los resultados de
este experimento, donde la edad tuvo un marcado efecto en casi todas las variables,
máxime cuando se realizaron las interacciones.
Sin embargo, los trabajos realizados por Cárdenas et al. (2012) en Pennisetum
purpureum vc Maralfalfa difieren de lo comentado en el párrafo anterior y de los
resultados de esta investigación, ellos obtuvieron valores de rendimiento de las hojas
cuando las edades de corte fueron superiores a 120 días (3.90t/ha-1), similares al
comportamiento de la especie aquí evaluada pero a los 75 días, para el método 2, no
así para el método 1 que lo mostró a los 45, lo que pudo estar dado por las
diferencias en las condiciones experimentales, entre otros factores, ya que la especie
en estudio fue la misma.
Urbano et al. (2005) obtuvieron un incremento en el rendimiento de 254 kg MS/ha-1
por día de intervalo entre cortes y señalaron que cuando la planta se maneja con
mayores días de recuperación, las reservas para el rebrote aumentaban, lo que
traería consigo mayores rendimientos (Márquez et al. 2007). Las investigaciones de
Calzada et al. (2014) reflejaron rendimientos de 13,42tMS/ha-1/corte a los 90 días de
edad inferiores a los obtenidos en este trabajo, aunque las condiciones
experimentales fueron diferentes.
Otros trabajos, como los de Faría et al. (2007), evaluando la producción forrajera de
cuatro germoplasmas de Pennisetum purpureum, reportan valores similares a los
mostrados en esta investigación. Ellos encontraron rendimientos para el pasto
Maralfalfa (11.19 t/ha-1 tallo) manteniendo una tasa media de crecimiento por encima
de los 113 kg/ha-1/dia-1. Valores obtenidos aquí para la edad de 75 días en el método
2. Destacar que las condiciones experimentales fueron diferentes, lo que muestra la
adaptación de este género a diferentes ecosistemas.

Bayble et al. (2007) en Pennisetum purpureum señalaron incremento del rendimiento
con la edad de la planta y obtuvieron 4,24; 6,67 y 15,20 tMS/ha-1/año a las edades de
60 y 90 días de rebrote, inferiores a los expuestos en este trabajo, donde se
alcanzaron rendimientos de 7,19 y 19,04 para el método 2, a estas mismas edades.
El rendimiento de materia verde y seca de los tallos en los dos métodos, aumentó con
la edad de la planta, lo que pudo estar dado entre otros aspectos por un incremento
en los carbohidratos estructurales (celulosa y hemicelulosa); así como de la lignina,
aunque no fueron medidos en este experimento. Sin embargo, pueden influir otras
causas como: la disponibilidad de agua, desarrollo del sistema radicular de la planta
y época del año; éstos pueden producir cambios morfológicos como la disminución
de láminas foliares y el aumento de los haces basculares (Valenciaga et al. 2009).
Aunque es importante destacar que en la Amazonía Ecuatoriana, las condiciones
climáticas son muy similares en todo el año, lo que trae un crecimiento rápido de la
planta, respuesta a lo sucedido en este trabajo.
En lo referente a las variables de calidad evaluadas, se mostraron altas diferencias
significativas al interaccionar los factores. Cuando se empleó una estaca los
indicadores químicos ceniza, proteína, fibra y grasa bruta, así como P, Ca y materia
seca presentaron los mayores tenores, lo que pudo deberse a los mayores
rendimientos de la materia seca al utilizar este método, ya que estos se presentan en
porcentajes de un kg de dicha materia.
Diferentes estudios reportan el incremento de la parte fibrosa y el calcio, y
disminución de la proteína y el fósforo para diferentes variedades estudiadas en el
trópico (Correa 2006 y Valenciaga 2009). Estos autores basan sus explicaciones en
estudios de la edad para diferentes variedades de Pennisetum, explicando que el
incremento de la madurez de la planta disminuye la calidad, al incrementar la fracción
tallo, la cual posee mayor cantidad de carbohidratos estructurales, cuestión que será
explicada con posterioridad.

La proteína disminuyó con la edad de rebrote en los dos métodos analizados y
puedo estar relacionado con la reducción de la síntesis de compuestos proteicos,
debido al incremento de la síntesis de carbohidratos estructurales (celulosa y
hemicelulosa). Otro factor como la disponibilidad de nitrógeno del suelo, pudo influir
en este comportamiento (tabla 13).
Maya y Durán (2006) reportaron marcada tendencia de la proteína a declinar con la
edad, obteniendo valores de 10,98 y 9,67% a los 28 y 42 días de rebrote,
respectivamente en gramíneas tropicales. Otras investigaciones como las reportadas
por Cárdenas et al. (2012) indicaron descenso de la PB hasta los 60 días según
avanzó la edad de la planta, lo que difieren de este trabajo donde los valores
disminuyeron hasta los 90 días de estudio. Es necesario destacar que para los dos
métodos evaluados hasta la edad de 75 los valores de la PB fueron superiores al 8%.
Por su parte, la FB se incrementó con la edad de la planta sin diferencias
significativas entre 60 y 75 días. Esto se pudo deber a la aparición de nuevos hijos.
Por otra parte, la mayor cantidad de fibra se relaciona con el incremento de la
proporción de los tallos, fracción que posee un alto contenido de lignina, además de
la senescencia de las hojas y acumulación del material muerto (Trejo et al. 2007).
Los minerales estudiados en este trabajo mostraron un comportamiento no definido
con la edad, aunque los porcentajes mas altos para el Calcio se presentaron a los 90
días, sin diferencias significativas con 75. Del Pozo et al (2002) plantearon que el
incremento del calcio con la edad está en dependencia de la acumulación de
carbonatos de Calcio en la pared celular como mecanismo de sostén (fracción tallo).
Esto disminuye el fósforo al reducirse la síntesis de compuestos orgánicos.
La edad o estado de madurez de la planta es, tal vez, el más importante y
determinante en la calidad del forraje. Durante el proceso de crecimiento de la planta,
después del estado foliar inicial existe rápido incremento de materia seca y cambio
continuo en los componentes orgánicos e inorgánicos. En la medida que avanza el
estado de madurez, la formación de los componentes estructurales (celulosa y
hemicelulosa) ocurren con mayor velocidad que el incremento de los carbohidratos

solubles. Además, los componentes nitrogenados progresivamente constituyen
menor proporción de la materia seca (Pirela 2005).
Estos resultados permitieron establecer la relación funcional de la edad con diferentes
indicadores tanto morfológicos, del rendimiento y la calidad, para los métodos 1 y 2.
El rendimiento de materia verde y materia seca se incrementó con la edad y se
ajustaron ecuaciones de regresión cuadrática para los métodos 1 y 2 en las dos
variables, aunque los ajustes fueron mayores para el primero. Del Pozo et al. (2001)
informaron ecuaciones de tercer grado entre el rendimiento y la edad de rebrote del
pasto estrella (Cynodon nlemfuensis), en un suelo ferralítico rojo compacto en la
región occidental de Cuba.
En estos resultados las curvas poseen mayor ajuste que las referidas en este trabajo,
aunque las especies son diferentes y las condiciones experimentales también.
Ramírez (2010) y Ramírez et al (2012) reflejaron ecuaciones de segundo grado para
el Pennisetum purpureum vc CT 169 y el Megathyrsus maximus en los períodos
lluviosos y poco lluvioso, respectivamente, aunque es necesario señalar que las
condiciones experimentales son diferentes, partiendo que las precipitaciones en la
Provincia Pastaza, superan los 4500mm en el año, demostrando la plasticidad
ecológica de este género.
Trabajos mas recientes de Rodríguez et al. (2013) reflejaron ecuaciones de
regresiones lineales, cuadráticas, cúbicas, logísticas y Gompertz, al establecer la
relación funcional de la edad con el rendimiento de materia seca en especies de
Pennisetum en la región occidental de Cuba. Los mayores ajustes y similitud a la
curva sigmoidea de los pastos se pudo deber a los estudios en edades mas
tempranas y tardías a las aquí mostrada, lo que explica el comportamiento de la
especie Maralfalfa en este experimento. Uvidia et al. (2015) reportaron
comportamiento similar del Pennisetum purpureum en el ecuador, al ajustarse las
curvas de producción de materia verde y seca a ecuaciones cuadráticas, lo que
justifica los resultados obtenidos en esta investigación.

Los ajustes para los rendimientos de materia verde y seca de las hojas en los dos
métodos fueron de forma general ecuaciones cuadráticas, lo que coincidió con los
ajustes del rendimiento total. Ramírez et al. (2012) para las hojas y tallos reportaron
expresiones de segundo grado en la especie Brachiaria decumbens con coeficientes
superiores a los mostrados en este trabajo, lo que pudo estar dado por la especie y
las condiciones experimentales, que fueron diferentes, lo que ratifica la aplicabilidad
de los modelos en dependencia de las plantas y condiciones.
Para los dos métodos (1 y 2) de forma general los ajustes para las restantes variables
morfológicas fueron ecuaciones cuadráticas y cúbicas, con altos coeficientes, bajos
errores estándar y cuadrados medios del error, explicando la relación funcional y
veracidad de los modelos en el comportamiento del Pennisetum purpureum vc
Maralfalfa en la Provincia Pastaza.
Los indicadores químicos estudiados se ajustaron a ecuaciones de segundo y tercer
grado a excepción del fósforo en los dos métodos (1 y 2). Los ajustes fueron altos
para la proteína y fibra bruta (tabla 17), lo que pudo estar dado por un
comportamiento mejor definido según avanzó la edad de madurez del material.
Investigaciones desarrolladas en Venezuela por Marcial (2014), reportaron valores
similares de FB para especies de Pennisetum. Sin embargo, el ajuste al relacionar la
edad fueron con ecuaciones lineales, lo que pudo estar dado por la variedad y las
condiciones experimentales diferentes (tabla 17).
En este trabajo se establecieron expresiones matemáticas que permitieron determinar
el contenido de fósforo y calcio de esta variedad al ser sembrada por diferentes
métodos, en dependencia de la edad de rebrote (tabla 18). Los valores que
reportaron autores como Sánchez (2009) dan a conocer porcentajes de P y Ca para
especies de del género y Pennisetum similares a los obtenidos en esta investigación.
Ramírez et al. (2014) obtuvieron ecuaciones cuadráticas al establecer la relación del
Ca y P el fósforo con la edad en el pasto Mulato, aunque destacar que las

condiciones experimentales fueron diferentes, lo que sin duda le confiere importancia
a los modelos de predicción.
Sin dudas, los resultados aquí presentados evidenciaron la influencia que tiene la
edad de rebrote en la producción de esta especie, máxime si se conjuga una
adecuada distancia de siembre y cantidad de estacas. Los mayores valores se
encontraron para el método 2.
Las expresiones matemáticas que se lograron ajustar mostraron cierta uniformidad en
el comportamiento de la variedad, ya que en su generalidad se ajustaron a
expresiones de segundo y tercer grado para los dos métodos, en las variables de
rendimiento y la PB, FB, Ca y P. Sin embargo, al analizar los parámetros de dichas
expresiones se apreciaron valores característicos para cada forma de plantar, lo cual
indicó las características intrínsecas de la especie y su respuesta al medio, según el
método que se emplee para su plantación.
Estos aspectos indicaron que en el momento de trazar estrategias para su manejo
no puede existir uniformidad en su tratamiento y sí tomar como elemento básico los
resultados de la sección A.
Finalmente aunque la sección A no tuvo por objetivo estudiar la influencia de los
factores climáticos, quedó evidenciado su efecto en los indicadores evaluados,
fundamentalmente en los rendimientos de esta especie. Este aspecto se abordó con
mayor profundidad en la sección B.
Los trabajos realizados por Delnum (1966 y 1976) y Dirven (1976) son
investigaciones clásicas donde se determinó el efecto de los factores del clima en el
crecimiento y la composición química de diferentes pastos. Sin embargo, estas
investigaciones se desarrollaron en invernaderos, donde fue posible controlar la
temperatura, la duración, intensidad y calidad de la luz, la humedad relativa y la
cantidad y calidad de agua, entre otros factores.
Es importante destacar que estos estudios fueron de significativa importancia para
establecer elementos básicos fundamentales de la fisiología, crecimiento y desarrollo

de los pastos. Sin embargo, su alcance práctico era limitado debido a las relaciones e
interacciones que se establecen entre esos elementos y el pasto cuando se
desarrolla en las condiciones de campo.
Herrera y Ramos (2006), señalaron que el clima es el conjunto de las condiciones
meteorológicas que suelen darse en una región y es el resultado de la combinación
de varias propiedades físicas de la atmósfera (temperatura, humedad, vientos,
radiaciones, estado eléctrico), que suelen concurrir en la misma y perduran durante
un período de tiempo. Esto reafirmó lo planteado en el párrafo anterior.
Las relaciones establecidas entre los indicadores productivos ( RMVT, RTMV, RMST
y RTMS) y las lluvias caídas indicaron la estrecha dependencia entre estas variables
(tablas 18 y 19).
Investigaciones en Cuba de Herrera y Ramos (2006), Ramirez (2010) y Ramírez et al.
(2012) informaron similar comportamiento en el Napier, King grass y Pennisetum
purpureum vc CT 169, respectivamente, y refirieron en los dos primeros casos, que
en la medida que se incrementó la norma de riego, la relación entre el rendimiento y
esta última disminuyó. Destacar que en la Amazonía Ecuatoriana, y en específico en
la Provincia Pastaza las precipitaciones sobrepasan los 4500mm y pueden alcanzar
los 6000mm en todo el año, con una distribución mensual que sobrepasa los 300mm,
factor que contribuye al buen desarrollo de esta especie, que muestra por su
comportamiento la plasticidad ecológica que posee.
Lösch (1995) planteó que el agua es el componente esencial de las células de las
plantas, que casi todos los procesos metabólicos dependen de su presencia.
Además, se requiere para el mantenimiento de la presión de turgencia, la difusión de
solutos en y entre las células, y suministra el hidrógeno y oxígeno que están
involucrados durante el proceso fotosintético, lo que permite reafirmar las altas
correlaciones con las diferentes variables, tanto de rendimiento como de
composición química medidas.
El efecto de las precipitaciones en el comportamiento de estos procesos
morfológicos, bioquímicos y fisiológicos relacionados con el crecimiento y la calidad

de los pastos, depende de múltiples factores que están estrechamente asociados al
ambiente, al suelo y su humedad y la especie de planta. En este sentido, se señaló
en la literatura que el crecimiento de los pastos es una función de la humedad
disponible en el suelo y que ésta, a su vez, varía en dependencia de la cantidad y
distribución de las precipitaciones, de la estructura y pendiente de los suelos, de
los valores de radiación y temperatura, así como del área cubierta por la vegetación
(Del Pozo 2004).
Sin embargo, llamó la atención que variables como el RMVH y RMSH no presentaron
altas correlaciones con las precipitaciones (tablas 18 y 19) y que en ambos métodos
fueron negativas, esto pudo estar relacionado con un incremento de la fracción tallo al
aumentar la madurez de la planta. Es necesario destacar que para las variables antes
mencionadas las correlaciones con el punto de rocío, aunque fueron negativas
resultaron altas, estableciendo la relación entre este, la temperatura y la humedad, lo
que podría explicar el comportamiento referido en el párrafo anterior. Señalar que la
literatura disponible no mostró resultado alguno donde se correlacionen estas
variables con las características morfológicas de la especie Pennisetum purpureum.
Investigaciones recientes de Uvidia et al (2015) en el Ecuador, reflejaron resultados
similares a los reportados en este trabajo y destacaron que el agua es el componente
esencial de las células de las plantas, que casi todos los procesos metabólicos
dependen de su presencia. Además, se requiere para el mantenimiento de la presión
de turgencia, la difusión de solutos en y entre las células, y suministra el hidrógeno y
oxígeno que están involucrados durante el proceso fotosintético, lo que permite
reafirmar las altas correlaciones con las variables, tanto de rendimiento como de
composición química.
Las variables químicas mostraron de forma general altos coeficientes para la
temperatura afuera, lo que evidenció la relación, dado en entre otros aspectos por la
influencia que ejerce la temperatura en el metabolismo de las plantas altas
productoras de forrajes.

Al tener en cuenta las correlaciones establecidas entre las variables climáticas, con
las de rendimiento y la composición química, se establecieron modelos líneales
simples, para conocer la relación entre el clima, el rendimiento y la calidad. En todos
los casos es necesario destacar que se utilizó la edad como variable independiente
(tablas 20 y 21).
Para el método 1 a excepción de la FB y el P, los coeficientes fueron altos, con bajos
errores estándar y cuadrados medios del error, lo que demostró la influencia de las
variables independientes sobre las dependientes, dándoles confiabilidad a los
modelos (Chacin 1998). En el método 2 los ajustes fueron mayores incluyendo el
fósforo (tabla 21), lo que pudo estar dado por un mejor comportamiento de las
variables medidas, influyendo la distancia y número de estacas empleadas en la
plantación, entre otros aspectos.
El objetivo del análisis de regresión, es predecir, sobre la base de la información
conocida y valores fijos de las variables independientes, el comportamiento de la
variable dependiente en relación a las primeras; y cuando el objetivo no es predecir
el valor de las observaciones, sino estudiar el efecto de las variables explicativas,
entonces se utiliza para indagar la importancia relativa de cada una de las variables
explicativas en el modelo (Chacin 1998).
La literatura reflejó la utilización de ecuaciones matemáticas para predecir la
producción de biomasa para bosques tropicales, templados y algunos tipos de
vegetación semiárida (Schroeder et al. 1997, Ter-Mikaelian y Korzukin 1997, Brown et
al. 1989 y Návar et al. 2001).
En la actualidad el número de trabajos donde se emplean los factores del clima en
expresiones matemáticas para explicar el comportamiento de la producción y de otros
indicadores de los pastos, no es amplio en la literatura publicada en el Ecuador. Sin
embargo, en otros países de América Latina, como es el caso específico de Cuba se
han desarrollo diferentes modelos con el objetivo de relacionar las variables
climáticas con el rendimiento y la calidad de especies de gramíneas y leguminosas.

Los trabajos de Santana y Pérez (2009) confirmaron la utilización de expresiones
matemáticas para predecir la composición química de los pastos. Estos autores
encontraron estrecha relación de la proteína bruta, la PC, la lignina y el contenido
celular con la edad, obteniendo ecuaciones de regresiones con coeficientes de
correlación de 0,81 y 0,93 para la proteína bruta digestible y la energía metabolizable,
respectivamente, al explicar la disminución del valor nutritivo con el aumento del
rendimiento y la edad.
En trabajos mas recientes de Ramírez et al. (2012) en la especie Pennsiteum
purpureum vc CT 169 se encontraron altos coeficientes de regresión (superiores a
0,93) al emplear ecuaciones lineales múltiples, donde se relacionaron el rendimiento
y la composición química con las variables climáticas. Estos autores utilizaron
fundamentalmente las precipitaciones y la humedad relativa en sus modelos, aunque
las condiciones experimentales fueron diferentes, lo que reafirma la alta relación entre
estas variables y avala los resultados de este trabajo.
Por otra parte, Verdecia (2015) estableció ecuaciones lineales múltiples para
relacionar las variables climáticas y la edad, con el rendimiento y la composición
química de especies de leguminosas, este autor logró altos coeficientes y grandes
ajustes cuando emplearon la humedad y las precipitaciones en sus modelos. Similar
comportamiento al obtenido en este trabajo. Estudios recientes Uvidia et al. (2015b)
reportaron valores similares a los de esta investigación a lo que ajuste se refiere.
Los resultados presentados en el presente capítulo revisten especial importancia para
la Amazonía Ecuatoriana, ya que anterioridad no se habían realizados estudios
similares. Sin embargo, como toda investigación científica al dar respuesta a un grupo
de problemas se crean otros, que debe ser solucionados con posterioridad.
En este capítulo se desarrollaron un grupo de principios básicos, formulados a través
de expresiones matemáticas, que al ser bien comprendidos y utilizados pueden ser
de extrema utilidad para la evaluación, selección, producción y manejo de nuevas
variedades que requieran ser introducidas o de aquellas que se utilizan
tradicionalmente.

Estas expresiones propician relacionar algunos factores del clima con importantes
indicadores de los pastos y permiten, con cierto grado de precisión, explicar su
comportamiento. Además, pudieran ser las bases para desarrollar modelos para
predecir su respuesta a los factores climáticos.

CONCLUSIONES
1. Se inventariaron y describieron las especies que forman la sucesión vegetal
secundaria de la provincia Pastaza en la Amazonia Ecuatoriana, la familia de
mayor aparición lo constituyó la fabácea.
2. Los indicadores morfológicos, de rendimientos y químicos se afectaron al
disminuir la distancia de plantación, y aumentar el número de estaca.
3. El rendimiento se incrementó con la edad de la planta, disminuyendo la
calidad en especial la proteína bruta. La relación funcional entre estas
variables mostró grandes ajustes para los indicadores de rendimiento
principalmente al plantar la especie con una estaca, a un metro de distancia.
4. Se establecieron ecuaciones de regresión múltiple lineal que permiten calcular
para la especie evaluada en los dos métodos de siembra empleados RMVH,
RMVT, RTMV, RMSH, RMST, RTMS, PB, FB, Ca y P a partir de las
precipitaciones, la humedad y la sensación térmica.

RECOMENDACIONES
1. Emplear las ecuaciones matemáticas que expresan la relación de los diferentes
indicadores del rendimiento y de calidad con la edad de rebrote y los factores
climáticos, como elementos básicos que contribuyan al mejor manejo de la
variedad evaluada.
2. Emplear el método de plantación de una estaca acostada, a un metro de distancia
para la especie estudiada en las condiciones de la Provincia Pastaza.
3. Emplear los resultados de esta tesis en la docencia de pre y postgrado de las
ciencias agropecuarias.
REFERENCIAS BIBLIOGRÁFICAS
1. Aguirre, Z. y Yaguana, N. 2012. Guía práctica para realizar estudios de
comunidades vegetales. Herbario Loja # 5. Departamento de Botánica y
Ecología de la Universidad Nacional de Loja. Loja – Ec. 30 p.
2. Alltech, 2015. La ganadería y sus desafíos en América Latina y el Caribe.
Disponible en http://es.alltech.com/blog/posts/la-ganaderia-y-sus-desafios-en-
america-latina-y-el-caribe (Consultado el 22-04-2015).
3. Alzamora, E. 2011. Evaluación del comportamiento productivo forrajero del
Pennisetum purpureum vc. Maralfalfa, bajo la aplicación diferentes niveles de
humus. Tesis de grado. Riobamba. Ecuador. 131p.
4. Andrade, D. 2009. Evaluación de dos sistemas y tres distancias de siembra del
pasto Maralfalfa (Pennisetum sp) en la localidad de Chalguayacu, cantón
Cumanda, provincia de Chimborazo. Riobamba, Ecuador. Escuela Superior
Politécnica de Chimborazo. 90 p.
5. Angiosperm Phylogeny Group (APG). 2003. The linear (LAPG) III: a linear
sequence of the familias in APG (III). Botanical Journal of the Linnean Society
161: 128-131.

6. Antolin, M.C. y Sánchez-Díaz, M., 1993. Effects of temporally droughts on
photosynthesis of alfalfa plants. Journal of Experimental Botany, 44 (265), 1341 -
1349.
7. AOAC. 1995. Official Methods of Analysis. 16th Ed. Ass. Off. Anal. Chem.
Washington, D.C.
8. Ayala, J. 1990. Plantación y establecimiento. En: King grass. Plantación,
establecimiento y manejo en Cuba. En Pennisetum purpureum para la ganadería
tropical Ed. Edica, Instituto de Ciencia Animal. Mayabeque-Cuba.
9. Ayala, J.R., Sistachs, M. y Herrera, R.S. 1989. Methodology to determine planting
time in king grass (Pennisetum sp.) using historical climate indices. Proceedings
XVI International Grassland Congress. Nice, France
10. Balcazar, M.P., Rangel, J.O. y Linares, E.L. 2000. Diversidad florística de la
serranía de las quinchas, Magdalena medio (Colombia). Caldasia. 22(2):191-224.
11. Barkley, Theodore M., Luc Brouillet, John L. Strother. 2008. Flora of North
America. Disponible en
http://www.efloras.org/florataxon.aspx?flora_id=1&taxon_id=10074 (Consultado el
22-03-2015).
12. Baruch, Z. 1994a. Response to drought and flooding in tropical forage grasses. II.
Leaf water potencial, photosynthesis rate and alcohol dehydrogenase activity.
Plant and Soil, 164, 97 - 105.
13. Baruch, Z. 1994b. Response to drought and flooding in tropical forage grass. I.
Production and allocation of biomass leaf growth and mineral nutrients. Plant and
Soil, 164, 87 - 96.
14. Baruch, Z. y Fisher, M.J.1991. Factores climáticos de competencia que afectan el
desarrollo de la planta en el crecimiento. En: Establecimiento y renovación de
pasturas. Conceptos, experiencia y enfoques de la investigación, Red de
Investigación y Evaluación de Pastos Tropicales. CIAT. Colombia p. 103.
15. Bayble, T., Solomon, M. y Prasad, N. K. 2007. Effects of cutting dates on nutritive
value of Napier (Pennisetum purpureum) grass planted sole and in association
with Desmodium (Desmodium intortum) or Lablab (Lablab purpureus). Livestock
Research for Rural Development 19(1).

16. Beliuchenko, I. S. y Febles, G. 1980. Factores que afectan la estructura de pastos
puros de gramíneas. Influencia de la relación hoja-tallo y contenido químico del
tallo. Rev. Cubana Cienc. Agríc. 14(2):167-174.
17. Benitez, A. 1980. Pastos y Forrajes. Editorial Universitaria Quito-Ecuador. 9, 35-
39p.
18. Bernal, J. 1984. Manual de pastos y forrajes para Colombia. Cuarta edición.
Colombia. 170 p.
19. Bernal, R., S.R. Gradstein y M. 2015. Catálogo de plantas y líquenes de
Colombia. Ediciones Celis. Instituto de Ciencias Naturales, Universidad Nacional
de Colombia, Bogotá. http://catalogoplantascolombia.unal.edu.co.
20. Blanco, F., 1995. Dinámica de crecimiento y variación de las reservas en Panicum
maximun. En: Informe final de etapa. Método de manejo de pastizales (Estudio
del Pastoreo Racional Voisin), 1-35. Ed. Eepf. Indio Hatuey. Matanzas (Cuba).
21. Bonet, A. 2004. Secondary succession of semi - arid Mediterranean old - fields in
south - eastern Spain: insights for conservation and restoration of degraded
lands. Journal of Arid Environments 56: 213-233.
22. Brown, S., Gillespie, A. J., y Lugo, A. E. 1989. Biomass estimation methods for
tropical forest with applications to forest inventory data. For. Sci. 35(4): 881-902.
23. Buestán, D. 2013. Evaluación de tres distancias y dos sistemas de siembra del
pasto maralfalfa (Pennisetum sp.) en el Centro de Investigación y postgrado de la
conservación de la biodiversidad Amazónica. Tesis de Ingeniero Agropecuario.
Universidad Estatal Amazónica. Escuela de Ingeniería Agropecuaria. Pastaza-
Ecuador.
24. Bustamante, M.M., O-Cardona, J., Loderman, J., Castañeda, J.A. y Eduardo, M.
2009. Biodiversidad de plantas en el borde amazónico putuyamanse: I.
inventario. CienciaAgro. 1(4): 130-138.
25. Buxton, D.R. y Fales, S.L. 1994. Plant environment and quality. En: Forage
Quality Evaluation and Utilization. Edited by American Society Agronomy Crop
Science Soc. of American, Madison, USA: p. 155-199.

26. Caamal, A. y Armendariz, I. 2002. La sucesión secundaria en los ecosistemas y
agroecosistemas tropicales – El henequén (Agave fourcroydes) en el contexto de
la diversificación. Tropical and Subtropical Agroecosystems. 1(1): 28-32.
27. Cáceres, O. y González, E. 2000. Metodología para la determinación del valor
nutritivo de los forrajes tropicales. Pastos y Forrajes, Vol. 23, Núm. 2:23-27.
28. Calzada, M., Enríquez, F., Hernández, A., Ortega, E. y Mendoza, S. 2014.
Análisis de crecimiento del pasto maralfalfa (Pennisetum sp.) en clima cálido
subhúmedo. Rev Mex Cienc Pecu. 5(2):247 - 260.
29. Cárdenas, L.R., Pinto-Ruiz, R., Medina, F.J., Guevara, F., Gómez, H.,
Hernández, A. y Carmona, J. 2012. Producción y calidad del pasto Maralfalfa
(Pennisetum sp) durante la época seca. Quehacer Científico en Chiapas. 1 (13):
38-46.
30. Castro, H.S. Galán, R. 2003 Conocimiento y manejo del bosque. Los Uiotots
en la Amazonia Colombiana. Biodiversidad. p 120.
31. Cerón, M.C. 1993. Manual de botánica ecuatoriana, sistemática y métodos de
estudio. Ediciones Abya – Ayala. Quito, Ecuador. 315 p.
32. Chacin, F. 1998. Análisis de regresión y superficie de respuesta. Universidad
Central de Venezuela. Primera Edición. Editorial Aragua. Venezuela. p 274.
33. Chacón-Moreno, E.; Nada, F; Sarmiento, G., 1995. Intercambio gaseoso,
nitrógeno foliar y optimización en el manejo del Panicum maximun (Tipo común)
sometido a diferentes frecuencias de cortes. Turrialba, 45(1-2), 19-26.
34. Chapin, F.S., Zavaleta, E.S., Eviner, V.T., Naylor, R.L., Vitousek, P.M., Reynolds,
H.L., Hooper, D.U., Lavorel, S., Sala, O.E., Hobbie, S.E., Mack, M.C. y Diaz, S.
2000. Consequences of changing biodiversity [Review]. Nature, 405, 234-242.
35. Choloquinga, M. y Cunuhayg, J. 2011. Evaluación de la adaptación del pasto
Maralfalfa (Pennisetum sp), en dos pisos altitudinales con tres distancias de
siembras en el Campus Juan Lunardis y Naste del Cantón paute. Tesis de
Ingeniería Agropecuaria Industrial. Cuenca. Ecuador.230p.
36. Clark, D. B., and Clark, D. A. 1993. Comparative analysis of microhabitat
utilization by saplings of nine tree species in Neotropical rain forest. Biotropica
25: 397-407.

37. Clavero, T. y Razz, R. 2009. Valor nutritivo del pasto Maralfalfa (Pennisetum
purpureum x Pennisetum glaucum) en condiciones de defoliación. Rev. Fac.
Agron. (Luz). 26: 78-87.
38. Corbea, L. y Martínez, L. 1982. Influencia de la densidad de siembra en el
establecimiento del King grass. Pastos y forraje: 178-183.
39. Correa, H. 2006. Calidad nutricional del pasto Maralfalfa (Pennisetum sp.)
cosechado a dos edades de rebrote. Livestock Research for Rural Development.
18(4). Disponible en: http:/www.irrd.org/irrd1816/corr18084. (Consultado 17-04-
2015)
40. Correa, H. J. C., Evangelista, C. J. y Pabón, R. 2009. Valor nutricional del pasto
kikuyo (Pennisetum clandestinum Hoechst Ex Chiov.) para la producción de leche
en Colombia (Una revisión): I. Composición química y digestibilidad ruminal y
posruminal. Disponible en: http://www.veterinaria. unal.edu.co /inv/
nutricion/kikuyo%202.pdf [Consultado 12-10-2013].
41. Croat, T. 1983. A Revision of the Genus Anthurium (Araceae) of Mexico and
Central America, Part I. Mexico and Middle America; Part II: Panama. MBG Press.
42. Cuatrecasas, G. (1958). Plantas Fanerógamas de Colombia. Instituto Alexander
von Humboldt, Instituto de Ciencias Naturales-Universidad Nacional de Colombia,
Ministerio del Medio Ambiente., Bogotá, Colombia.
43. Cuatrecasas, G. 1958. Plantas Fanerógamas de Colombia. Instituto Alexander
von Humboldt, Instituto de Ciencias Naturales-Universidad Nacional de
Colombia, Ministerio del Medio Ambiente., Bogotá, Colombia.
44. D'Atri, P. 2007. Pastizales del mundo. Novedades de Biodiversidad en América
Latina Nº 160. IPS (Inter Press Service) e IFEJ (Federación Internacional de
Periodistas Ambientales).
45. Dawson JE and Hatch ST. 2002. A world wide web key to the grass genera of
Texas. S.M. Tracy Herbarium, Department of Rangeland Ecology and
Management, Texas A&M University. Disponible en:
http://www.csdl.tamu.edu/FLORA/taes/tracy/610/index.html (Consultado el 15-04-
2015).

46. De Marzi, Valeria, 2006. Cien Plantas Argentinas. Editorial Albatros. Buenos
Aires-Argentina. p 32.
47. Del Pozo, P. 1998. Análisis del crecimiento del pasto Estrella (Cynodon
nlemfuensis) bajo condiciones de corte y pastoreo. Tesis de Doctorado. Instituto
de Ciencia Animal. Universidad Agraria de la Habana, 105pp.
48. Del Pozo, P. 2002. Bases ecofisiológicas para el manejo de los pastos
Tropicales. Pastos, XXXII (2), 109 – 137. San José de las Lajas. Mayabeque,
Cuba.
49. Del Pozo, P. 2004. Bases ecofisiológicas para el manejo de los pastos tropicales.
Sitio Argentino de Producción Animal. Disponible en http://www.produccion-
animal.com.ar/ (Consultado 16-04-2015).
50. Del Pozo, P. y Jerez, I. 1999. Experiencias en la implementación de un módulo
silvopastoril en una unidad de producción de leche en Cuba. En: Primer Congreso
Latino Americano de Agroforestería para la Producción Animal Sostenible. 25 al
27 de Octubre 1999. Cali – Colombia.
51. Del Pozo, P.P., Herrera, R., García, M., Cruz, A. y Romero, A. 2001. Análisis del
crecimiento y desarrollo del pasto estrella con y sin adición de fertilizante
nitrogenado. Rev. Cubana Cienc. Agríc. 35(1):51-58.
52. Delnum, B. 1966. Climate, nitrogen and Grass. Research inti the influnce of light
intensity, temperature, water supply and nitrogen on the production and chemical
composition of Grass. Medod. Landboowhogeschool, Wagenigen, Holanda.
53. Delnum, B. 1976. Photosynthesis and sink size: na explination for the low
production of Grass ewards in automn. Neth. J. agric. Sci. 24: 238.
54. Didham, R. and Lawton, J. 1999. Edge structure determines the magnitude of
changes in microclimate and vegetation structure in tropical forest fragments.
Biotropica 31: 17-30.
55. Dirven, J. 1976. Tropical rooghage. Int. Course on dairy cattle hosbandry. Int.
Agric. Centre, Wageningen, Holanda.
56. Echeverry, R. 2011. Plantas nativas en el jardín botánico Alejandro Von Humboldt
de la Universidad de Tolima-Ibagué. Colombia. Disponible en:

http://www.colombia.travel/es/a-donde-ir/andina/ibague/actividades/conoce-el-
jardin-botanico-alejandro-von-humboldt. (Consultado 23-05-2015).
57. Ecología Áreas verdes. 2015. Disponible en.
http://www.academia.edu/4939737/PROYECTO_AULA._Ecologia._areas_verdes.
(Consultado 23-04-2015).
58. Ecuador en Cifras. 2013. Disponible en http://www.ecuadorencifras.com/cifras-
inec/main.html. (Consultado el 10-03-2015).
59. Fabian, W. 2009. Evaluación de clones versus el CT-115 de Pennisetum
purpureum para la producción de biomasa. Riobamba, Ecuador. 90p.
60. Fales, S.L.; Muller, L.D.; Ford, S.A.; O'sallivan, M.; Hoover, R.J.; Holden, L.A;
Lanyon, L.E.; Buckmaster, D.R., 1995. Stocking rate affects production and
profitability in a rotationally grazed Pasture System. Journal Production
Agriculture, 8 (1), 88-96.
61. FAO 2009. Organización Mundial de las Naciones Unidas para la Agricultura y la
Alimentación Disponible en http://www.fao.org/docrep/012/i0876s/i0876s00.HTM
(Consultado el 22-04-2015)
62. FAO 2011. Organización Mundial de las Naciones Unidas para la Agricultura y la
Alimentación. Disponible en http://www.fao.org/docrep/016/i2373s/i2373s01.pdf
(Consultado 22-2015).
63. FAO 2012. Organización Mundial de las Naciones Unidas para la Agricultura y la
Alimentación. Disponible en http://www.fao.org/publications/sofa/2013/es/
(Consultado 22-04-2015)
64. FAO 2013. Organización Mundial de las Naciones Unidas para la Agricultura y la
Alimentación. Ganadería sostenible y cambio climático en América Latina y el
Caribe Disponible en http://www.fao.org/americas/perspectivas/ganaderia-
sostenible/es (Consultado 22-04-2015).
65. FAO 2013ª. Organización Mundial de las Naciones Unidas para la Agricultura y la
Alimentación. Disponible en http://www.fao.org/news/story/es/item/198166/icode
(Consultado 22-04-2015).
66. FAO 2014. El estado actual de los bosques del mundo. Potenciar los beneficios
socioeconómicos del bosque. Roma, 132p.

67. FAO 2014. Organización Mundial de las Naciones Unidas para la Agricultura y la
Alimentación. Capítulo 2.2 Ganadería. Disponible en
http://www.iica.int/Esp/organizacion/LTGC/ForosTecnicos/Documents/Foro6-
2013/Perspectivas2014/2.2Ganaderia.pdf (Consultado 22-03-2015).
68. Faría-Mármol, J., González B., Chirinos, Z., Alvarez, R. 2007ª. Efecto de la
frecuencia de corte sobre la producción forrajera y valor nutritivo de cuatro
cultivares de Pennisetum Purpureum . En: Memorias XX Reunión de la
Asociación Latinoamericana de Producción Animal (ALPA). Cuzco, Perú.
69. Flora Ibérica, 2012. Plantas vasculares de la Península Ibérica e Islas Baleares.
Real Jardín Botánico. Madrid-España.
70. Flores, J. 1986. Manual de Alimentación Animal. Editorial Limusa. Primera
edición. Tomo I. México. 232 p.
71. Foster, B.L. 2002. Constraints on colonization and species richness along a
grassland productivity gradient: the role of propagule availability. Ecology Letters
4: 530-535.
72. Gentry, A.H. 1982. Increasing turnover through time in tropical forests. Science
263:954– 58
73. Gentry, A.H. 1988. Changes in Plant Community Diversity and Floristic
composition on Environmental and Geographical Gradients. Ann. Missouri Bot.
Gard. 75(1): 2-34
74. Gentry, A.H. 1995. Patterns of diversity and floristic composition in Neotropical
montane forest, pp 103-126. En: Churchil, S,P, Balslev, H., Forrero E y Luteyn,
J.L. (ed). Biodiversity and Conservation of Neotropical montane forest. The New
York Botanical Garden, New York.
75. Gliessman, S. 1997. Agroecology. Ecological processes in sustainable
agriculture. Ann Arbor Press. USA. 357 p.
76. Gold, K. 1968. Grass systematics. Mc. Grow-Hill Book Company.
77. González, R y Caballero, H. 1989. Informe técnico final primera fase (1983-
1988). Programa de producción animal. Estación experimental Napo-Payamino.
INIAP, Quito-Ecuador.

78. Grime, J. P. 2001. Plant strategies, vegetation process and ecosystem
properties. Wiley & Sons, Chichester.
79. Grubben, G. 2004. Plant Resources of Tropical Africa: Vegetables (en inglés).
PROTA. p. 184.
80. Guamanquishpe, L. 2012. Evaluación de la productividad del pasto Maralfalfa
(Pennisetum sp), mediante dos tipos de multiplicación asexual y dos abonos
orgánicos en Cunchibamba, provincia de Tungurahua. Tesis de Ingeniero
Agrónomo. Universidad Estatal de Bolívar. Facultad de Ciencias Agropecuarias
Recursos Naturales y Del Ambiente.
81. Guirado, J. y Mendoza, R. 2000. La regeneración del medio forestal almeriense.
En Desertificación en Almería. Eds. Carmona, M. y Sánchez, M. (Grupo
Ecologista Mediterráneo, Almería, España) p. 101-110.
82. Häfliger E and Scholz H. 1980. Grass weeds 1: weeds of the subfamily.
83. Hernandez, G., McClellan, F., Branson, A. 2013. Mexico: Livestock and products
i-annual: pork sector appears promising as beef sector faces challenges. GAIN
Report MX3016, Foreign Agriculture Service, U.S. Department of Agriculture,
Washington, D.C., March 4. Disponible en http://gain.fas.usda.gov/Recent GAIN
Publications/Livestock and Products Semi-annual_Mexico City_Mexico_3-4-2013.
84. Herrera, R. 2006. Fisiología, Calidad y muestreos. En: Fisiología,
producción de biomasa y sistemas silvopastoriles en pastos tropicales. Abono
orgánico y biogás. Eds. Herrera, R., Rodríguez I. y Febles, G. Ed. Edica p.1.
Mayabeque-Cuba
85. Herrera, R. 2008. Principios básicos de Fisiología Vegetal. En: Pastos tropicales,
principios generales agrotecnia y producción de materia seca. Ed. Instituto de
Ciencia Animal y FIRA, México. p 1.
86. Herrera, R. 1990. Introducción y características botánicas En: King grass.
Plantación, establecimiento y manejo en Cuba. Ed. Edica. Instituto de Ciencia
Animal, Mayabeque-Cuba.
87. Herrera, R. 2006. Fotosíntesis En: Pastos tropicales, contribución a la Fisiología,
establecimiento, rendimiento de biomasa, producción de biomasa, producción de
semillas y reciclaje de nutrientes, Ed. EDICA. ICA, La Habana. p. 37.

88. Herrera, R. y Ramos, N. 2006. Factores que influyen en la producción de biomasa
y calidad. En Pennisetum purpureum para la ganadería tropical. EDICA, La
Habana. p. 79.
89. Herrera, R., Del Pozo, P., Blanco, F. 2006.b Bases ecofisiológicas del crecimiento
de los pastos. Cuba.
90. Herrera, R., García, M., Cruz, A. y Romero, A. 2012. Evaluación de clones de
Pennisetum purpureum obtenidos por cultivo de tejidos in vitro. Revista Cubana
de Ciencia Agrícola, 46(4):427-433.
91. Hobbs, R.J., Arico, S., Aronson, J., Baron, J.S., Bridgewater, P. and Cramer, V.A.
2006. Novel ecosystems: theoretical and management aspects of the new
ecological world order. Global Ecology and Biogeography. 15: 1-7.
92. Huston, M.A. 1994. Biological Diversity. Cambridge University Press. 681p.
93. IDEAM-FAO. 2002. Colombia: VIII. Estado actual de la información sobre
recursos forestales y cambio en el uso de la tierra (Instituto de Hidrología,
Meteorología y Estudios ambientales-(IDEAM), Consultores. Depósito de
Documentos de la FAO. P 90.
94. INAMHI 2006. Anuario meteorológico del Ecuador. Quito. Ecuador.
95. Instituto Nacional Autónomo de Investigaciones Agropecuarias (INIAP), 2011.
Disponible en
http://www.iniap.gob.ec/sitio/index.php?option=com_content&view=article&id=452
&Itemid=250 (Consultado el 22-04-2015).
96. Instituto Nacional Autónomo de Investigaciones Agropecuarias (INIAP), 2002.
Proyecto: Rehabilitación de pastizales degradados y mejoramiento de la
sostenibilidad de la ganadería en zonas intervenidas de la Amazonía Ecuatoriana.
Estación Experimental Napo Payamino. Programa Silvopastoril. Napo-Ecuador.
97. Intergovermental Panel of Climate Change (IPCC), 2002. Climate Change and
Biodiversity. Disponible en https://www.ipcc.ch/pdf/technical-papers/climate-
changes-biodiversity-en.pdf (Consultado el 22-03-2015).
98. Jardín Botánico de Acapulco. 2014. Disponible en
http://www.acapulcobotanico.org/?page_id=210 (Consultado el 22-03-2015).

99. Jardines, S., Hernández, R., Hondal, T., Moro, A. y García, R. 2000. Nota
Técnica: Caracterización de leguminosas naturales en áreas de pastoreo.
Alysicarpus vaginalis L. DC. Revista Pastos y Forrajes. 23(3): 1-6.
100. Jeffrey, C. 2007. Compositae: Introduction with key to tribes. Pages 61-87 in
Families and Genera of Vascular Plants, vol. VIII, Flowering Plants, Eudicots,
Asterales (J. W. Kadereit and C. Jeffrey, eds.). Springer-Verlag, Berlin.
101. Jones, R.J and Sandland, R. L. 1974. The relation between animal gain and
stocking rate. Derivation of the relation from the resultais of grazing triais. Journal.
Agricultural. Science, Cambrige, 83: 335-342
102. Jorgensen, P. y Leon, Y.S. 1999. Catalogue of vascular Plants of Ecuador.
Monographys in Sistematic Botany from the Missouri Botanical Garden. pp 75, 93.
103. Keels, S., Gentry, A. y Spinzi, L. 1997. Using vegetation analysis to facilitate
the selection of conservation sites in eastern Paraguay. (Biodiversity measuring
and monitoring certification training, V (2). Washington: SI/MAB.
104. Langdon, R. 2001. The Bamboo Raft as a Key to the Introduction of the Sweet
Potato in Prehistoric Polynesia, The Journal of Pacific History', Vol. 36, No. 1.
105. Longman, K. A. and Jenik, J. 1987. Tropical forest and its environment. 2nd.
Ed. John Wiley and Sons. New York Longman Scientific and Technical. USA. p.
65-69.
106. Lösch, R. 1995. Plant water relations. En: Physiology. Progress in Botany. V
56 Springer Forlag Berlin 56: 55- 96.
107. Lozano, P. 2002. Los tipos de bosque en el sur de Ecuador. Botánica
Austroecuatoriana. Estudios sobre los recursos vegetales en las provincial de El
Oro, Loja y Zamora-Chinchipe. Abya-Yala, Quito, 29-49.
108. Lozano, P. 2002. Los tipos de bosque en el sur de Ecuador. Bótanica
Austroecuatoriana. Estudios sobre los recursos vegetales en las provincial de El
Oro, Loja y Zamora-Chinchipe. Abya-Yala, Quito, 29-49.
109. MAE. 2011. Ministerio del Ambiente del Ecuador. Manejo forestal sustentable.
Norma técnica Bosque Tropical. Oficina Técnica Forestal Napo. Ecuador. 30p.

110. Maraschin, G.E. 1996. Manejo de Coast-cross-1 SOB pastejo. En: Anais do
workshop sobre o potencial forrajeiro do género Cynodon, 93-107.
EMBRAPA/CNPGL (Brazil).
111. Marcial C. Rendimiento y composición química del Pennisetum purpureum vc.
Cuba CT-115 en los Llanos Occidentales de Venezuela. 2014.[ Tesis MSc.
Universidad de Granma]. Cuba. 98.
112. Márquez, F., Sánchez, J., Urbano, D. y Dávila, C. 2007. Evaluación de la
frecuencia de corte y tipos de fertilización sobre tres genotipos de pasto elefante
(Pennisetum purpureum).1.Rendimiento y contenido de proteína. Zootecnia
Trop., 25(4): 253-259.
113. Martí, H. R., Corbino, G. B. y Chludil, H. D. 2011. La batata, el
redescubrimiento de un cultivo. Disponible en
http://www.cienciahoy.org.ar/ln/hoy121/index.htm (Consultado el 22-03-2015).
114. Martínez, R. 2001. Cómo guardar comida para la seca. Hierba Elefante CT
115. Insitituto de Ciencia Animal. Editorial EDICA. p.7.
115. Martínez, R., Herrera, R., Cruz, R. y Torres, V. 1996. Cultivo de tejidos y
fitotecnia de las mutaciones en pastos tropicales. Pennisetum purpureum: otro
ejemplo para la obtención de nuevos clones. Revista Cubana de Ciencias
Agrícolas. 31 (1): 1-10.
116. Maya, G. y Durán, C. 2006. Valor nutritivo del pasto estrella solo y en
asociación con Leucaena a diferentes edades de corte durante el año. Disponible
en: http://www.biblioteca.palmira.unal.edu.co. [Consultado 8-05-214].
117. Meneses, E. 2011. Conviértase en empresario ganadero. Grupo Latino. D´vinni
S.A. Colombia. P 271
118. Ministerio Coordinador de Producción Empleo y Competitividad (MCPEC).
2011. Disponible en http://www.produccion.gob.ec/2011-resolucionescomex/
(Consultado 20-04-2015).
119. Ministerio de la coordinación de la productividad, empleo y competitividad.
2011. Agenda para la transformación productiva territorial: Provincia de Pastaza.
Disponible en http://www.produccion.gob.ec/. (Consultado 12-03-2015).

120. Miranda, J.D., Padilla, F.M. y Pugnaire, F.I. 2004. Sucesión y restauración en
ambientes semiáridos. Ecosistemas. 13(1):55-58.
121. Mislevy, R. y Pate, F. 1996. Establishment, management and utilization of
Cynodon grasses in Florida. En: Anais do workshop sobre o potential forrajeiro do
Género Cynodon, 127 - 138. EMBRAPA/CNPEL. Brazil.
122. Missouri Botanical Garden. 2014. National Historic Landmark summary listing.
National Park Service.
http://tps.cr.nps.gov/nhl/detail.cfm?ResourceId=1187&ResourceType=District.
Retrieved 2015-02-26.(Consultado 10-04-2014).
123. Mitsch, W. and Gosselink, J. 2000. Wetlands. John Wiley and Sons. New York.
124. Mittermeier, R., Mittermeier, C. y Robles, P. 1997. Megadiversity: Earth’s
biologically wealthiest nations. CEMEX, México, D.F.
125. Mouquet, N., Munguia, P., Kneitel, J. M. and Miller, T. E. 2003. Community
assembly time and the relationship between local and regional species richness.
Oikos 103: 618-626.
126. Murillo, R., Chacón, E., Ramírez de la Ribera, J., Álvarez, G., Álvarez, P.,
Plúa, K. y Álava, A. 2015 (Rendimiento y calidad de dos especies del género
Pennisetum en Ecuador REDVET Rev. Electrón. vet. Volumen 16( 8): 1-3.
Disponible en: http://www.veterinaria.org/revistas/redvet/n080815.html.
Consultado (15-08-2015).
127. Myers, N., Mittermeier, R., Mittermeier, C., Da Fonseca, B. y Kent, J. 2000.
Biodiversity hotspots for conservation priorities. Nature 853-858.
128. Návar, J., Nájera, A., Domínguez, P. A. y Jurado, E. 2001. Biomass
estimation equations in the Tamaulipan thomscrub of northeastem México. In
Press in Journal of Arid Environments.
129. Navas J., Nava C., Gutiérrez E., Zavala, F., García, E., Treviño, E., Bernal H. y
Herrera R. 2013. Establecimiento del pasto 'CT-115' (Pennisetum purpureum) en
una zona semiárida del noreste de México. Rev. fitotec. mex .36(3): 239-244.
130. Neill, D. 2012. ¿Cuántas especies nativas de plantas vasculares hay en
Ecuador? Revista Amazónica Ciencia y Tecnología. 1:70-83.

131. Neill, D. 1999. Vegetación. En Catálogo de las plantas vasculares del Ecuador,
editado por P. Møller Jørgensen y S. León. Missouri: Missouri Botanical Garden
Press.
132. Neill, D. 1999. Vegetación. En Catálogo de las plantas vasculares del Ecuador,
editado por P. Møller Jørgensen y S. León. Missouri: Missouri Botanical Garden
Press.
133. Neill, D. 2012. ¿Cuántas especies nativas de plantas vasculares hay en
Ecuador? Revista Amazónica Ciencia y Tecnología. 1:70-83.
134. Neill, D.A. y Ulloa-Ulloa, C. 2011. Adiciones a la Flora del Ecuador: Segundo
suplemento, 2005–2010. Fundación Jatun Sacha, Quito, 202 pp.
135. Nelson, C.J. y Moser, L.E. 1994. Plant Factors Affecting Forage Quality. In:
Farey Jr., G.C., Ed., Forage Quality, Evaluation, and Utilization, America Society
of Agronomy, Wisconsin, 59-114.
136. Olmstead, R. G., Sweere, J. A., Spangler, R. E., Bohs, L. y Palmer, J. D. 1999.
Phylogeny and provisional classification of the Solanaceae based on chloroplast
DNA. Pp. 111-137.
137. Ortiz, B. 2008. Evaluación Morfológica y Agronómica de los Pastos OM-22 y
CT-115 en Zona Henequenera del Sureste de México. Instituto Tecnologico de
Conkal.
138. Padilla, C. et al 2004. Efecto del momento de inicio de la preparación del
suelo en el establecimiento de hierba elefante Cuba CT-115 (Pennisetum
purpureum) y el control de espartillo (Sporobolus indicus L R. Br.). Revista
Cubana de Ciencias Agrícolas. 40:123: 85-87.
139. Padilla, C., Sardiñas, Y., Cino, D., Curbelo, F. 2005. Momento de inicio de la
preparación del suelo y su repercusión económica en el establecimiento del
Pennisetum purpureum vc Shul (hierba elefante Cuba CT 115) y Sporobolus
indicus L.BR (espartillo). Revista Cubana de Ciencia Agrícola. 39(4): 643-650.
140. Paéz, A., González, M.E., Yrausquin, X., Salazar, A., Casanova, A. 1995.
Water stress and clipping management effects on guinea grass. I Growth and
biomass Allocation. Agronomy Journal, 87, 698-706.

141. Pérez, M. 2013. Cyperacea. Disponible en
http://www.botanicayjardines.com/cyperus-rotundus/ (Consultado el 22-03-2015).
142. Pezo, D. e Ibrahim, M. 1999. Sistemas silvopastoriles 2da ed. Turrialba, Costa
Rica CATIE. Proyecto Agroforestal CATIE/GTZ. 276p.
143. Phillips, O. y Miller, J.S. 2002. Global patterns ok plant diversity: Alwayn H.
Gentry’s forest transsect date set. Monographs in Systematic Botany from the
Missoun Botanical Garden. 89:164-199.
144. Phillips, O. y Miller, J.S. 2002. Global patterns ok plant diversity: Alwayn H.
Gentry’s forest transsect date set. Monographs in Systematic Botany from the
Missoun Botanical Garden. 89:164-199.
145. Pinto, R., Ramírez, L., Kú Vera, J.C. y Ortega, L. 2002. Especies arbóreas y
herbáceas forrajeras del sureste de México. Revista Pasto y Forrajes. 25(3):171-
180.
146. Pirela, M.F. 2005. Valor nutritivo de los pastos tropicales. Manual de
ganadería doble propósito. Disponible en. http://avpa.vc/doc pdf s/libroson
line/Manual de ganadería/sección 3/articulo 6.pdf. [Consultado 9-10-2012].
147. Ramírez J. L., Leonard, I., Verdecia, D., Pérez, Y., Arseo, Y., Álvarez, Y. 2014.
Relación de dos minerales con la edad y los elementos del clima en un pasto
tropical. REDVET 15(5):1-8. Disponible en:
www.veterinaria.org/revistas/redvet/n155.html. [Consultado 22-03- 2015].
148. Ramírez N., Izquierdo, C., Paladino, F. 1996. Producción y utilización de
pastizales, cinco zonas agroecológicas del Ecuador MAG, REPAAN. 1996.
149. Ramírez, A., Benítez, D. G., Guevara, O., Díaz, M. y Guerra, J. R. 2004.
Impacto de la actividad ganadera en los macizos montañosos cubanos. En:
Memorias del Congreso Internacional de Agricultura en Ecosistemas Frágiles y
Degradados. Granma, Cuba: IIA Jorge Dimitrov.
150. Ramírez, J. L. 2006. Rendimiento y componentes del valor nutritivo en cuatro
pastos tropicales en las actuales condiciones edafoclimáticas del Valle del Cauto.
Tesis MSc. Universidad de Granma.p.82

151. Ramírez, J. L. 2010. Rendimiento y calidad de cinco gramíneas en el Valle del
Cauto. Tesis presentada en opción al grado Científico de Doctor en Ciencias
Veterinarias. Universidad de Granma. Cuba. p.100.
152. Ramírez, J. L., Herrera, R. S., Leonard, I., Cisneros, M., Verdecia, D., Álvarez,
Y. López, B. 2012. Relationship between quality indicators and age indicators in
Pennisetum purpureum vc. Cuba CT-169 in Cauto Valley, Cuba. Cuban Journal of
Agricultural Science. 46(3): 315-319.
153. Ramos, N., Herrera, R.S., Padilla, C., Barrientos, A., Aguilera, J. M. 1987.
Pasto estrella mejorado (Cynodon nlemfuensis) su establecimiento y utilización en
Cuba. Ed. Edica, 153 pp. Mayabeque, Cuba.
154. Rodríguez, L., Larduet, R., Martínez, R., Torres, V., Herrera, M., Medina Y.,
Noda, A. 2013. Modelación de la dinámica de acumulación de biomasa en
Pennisetum purpureum vc. king grass en el occidente de Cuba. Revista Cubana
de Ciencia Agrícola. 47(2): 119-124.
155. Romero, C. S., Medina, R. y Flores, R. 1998. Efecto de la fertilización
nitrogenada sobre los componentes morfológicos del pasto estrella (Cynodon
plectostachyus) en la zona de bajo Tocuyo, Estado Falcon. Rev. Zootecnia
Tropical. 16: 41.
156. Samaniego, E., García, Y. y Neill, D. 2015. Estructura y composición del
bosque húmedo tropical premontano de la estribación oriental del Parque
Nacional Llanganates. Revista Amazónica Ciencia y Tecnología, 3(3): 231-247.
157. Samaniego, E., Vargas, J. y Uvidia, H. 2014. Establecimiento de un banco de
germoplasma de productos forestales no maderables. Revista Forestal Baracoa,
33: 265-274.
158. Sánchez, J. 2009. Utilización eficiente de las pasturas tropicales en la
alimentación del ganado lechero. XI Seminario de manejo y utilización de los
Pastos y Forrajes en Sistemas de Producción Animal. Disponible:
http://avpa.ula.ve/eventos/xi_seminario/Conferencias/Articulo-2.pdf. (Consultado
23-03-2013).
159. Santana, P. y Pérez, L. 2009. Determinación de la edad óptima de corte del
Napier (Pennisetum purpureum) a partir de la variación de su valor nutritivo y

rendimiento en la época lluviosa. Disponible en: www.ilustrados.com [Consultado
20-09-2012].
160. Scheffer, M. and Carpenter, S. 2003. Catastrophic regime shifts in ecosystems:
linking theory to observation. Trends in Ecology and Evolution 18: 648 - 656.
161. Schroeder, P., Brown, S., Mo, J., Birdsey, R. y Cieszewski, C. 1997. Biomass
estimation for temperate broadleaf forest of the United States using inventory
data. For. Sci. 43 (3): 424-434.
162. Sere, C., y Steinfeld, H. 1996. World livestock production systems: current
status, issues and trends. Animal production and health paper No127. FAO.
Rome.
163. Sierra, R. 1999. Propuesta Preliminar de un Sistema de Clasificación de
Vegetación para el Ecuador Continental. Proyecto INEFAN/GEF-BIRG &
EcoCiencia, Quito. 194 pp.
164. Sinoquet, H. y Cruz, P. 1995. Ecophysiology of tropical intercropping. INRA,
France, pp. 425.
165. Sistachs, M. 1986. Malas hierbas. En: Los Pastos en Cuba. Tomo 1.
Producción. EDICA. La Habana, Cuba. p. 579.
166. Soto, A y Castro, J. 2011. Biodiversidad de Costa Rica. Witheringia
solanácea. Disponible en http://atta2.inbio.ac.cr/neoportal-
web/species/witheringia%20solanacea (Consultado el 22-03-2015).
167. Soto, A. 2015. Biodiversidad de Costa Rica. Disponible en:
http://atta2.inbio.ac.cr/neoportal-web/species/witheringia%20solanacea.
(Consultado 15-03-2015).
168. Stevens, P. F. 2012. Asteraceae. Angiosperm Phylogeny Website. Cornell
University.
169. Tergar, L.E., Velez, S., Saldanas, D. 1988. Production of grazed tropical
grassesing different agroecosystem in Puerto Rico Humid Mountain. The Journal
of Agriculture of University of Puerto Rico, 72(1), 99 - 108.
170. Ter-Mikaelian, M. T. y Korzukhin. 1997. Biomass equations for sixty five North
American tree species. Forest Ecology and Management 97: p.1-24.

171. Tierra pastos y ganado. 2012. Memorias Seminario Internacional Campo Suelo
Ecológico – Pasto verde – Carne verde – Leche verde. Universidad Estatal
Amazónica. Puyo – Ecuador.
172. Tilman, D. and C. Lehman, 2001. Human-caused environmental Change:
Impacts on plant diversity and evolution. Proceedings of the National Academy of
Sciences of the United States of America. 98, 5433-5440.
173. Trejo, R., Canul, J. y Duarte, F. 2007. Producción de tres variedades de
Pennisetum purpureum fertilizadas con dos diferentes fuentes nitrogenadas en
Yucatán, México. Revista Bio Ciencias; 2(2): 60-68.
174. Turner, C.L. Y Seastedt, T.R. 1993. Maximization of above ground grassland
production the role of defoliation frecuency intensity and history. Ecological
Aplications, 3(1), 175 - 186.
175. Universidad de Bayreuth. 2013. The Genera of Asclepiadoideae,
Secamonoideae and Periplocoideae (Apocynaceae): Descriptions, Illustrations,
Identification, and Information Retrieval.Disponible en:http/www.uniapart.de.
(Consultado 15-02-2014).
176. Urbano, D., Dávila, C. y Castro, F. 2005. Efecto de la frecuencia de corte sobre
cinco variedades de Pennisetum en zona alta del estado Mérida, Venezuela.
Biotam Nueva Serie. 2: 460-463.
177. USDA, ARS, National Genetic Resources Program. 2013. Germplasm
Resources Information Network - (GRIN) [Online Database]. National Germplasm
Resources Laboratory, Beltsville, Maryland. Disponible en http://www.ars-
grin.gov/cgi-bin/npgs/html/gnlist.pl?1773 (Consultado el 20-03-2015).
178. Uvidia, H., Ramírez, J., Vargas, J., Leonard, I. y Sucoshañay, J. 2015b.
Relación del clima con el rendimiento y la calidad del Pennisetum purpureum en
la Amazonia Ecuatoriana. REDVET Rev. Electrón. vet. 16 (6) Disponible en: -
http://www.veterinaria.org/revistas/redvet/n060615.html . Consultado (20-07-
2015).
179. Uvidia, H., Ramírez, J., Vargas, J., Leonard, I. y Sucoshañay, J. 2015a.
Rendimiento y calidad del Pennisetum purpureum vc Maralfalfa en la Amazonía
ecuatoriana -REDVET Rev. Electrón. vet. 16 (6): 1-3. Disponible en:

http://www.veterinaria.org/revistas/redvet/n060615.html. Consultado (12-07-
2015).
180. Valencia, R., Pitman, N., León, Y.S. y Jorgensen, P.M. 2000. Libro Rojo de las
plantas endémicas del Ecuador. Quito: Herbario QCA, Pontificia Universidad
Católica del Ecuador.
181. Valenciaga, D., Chongo, B., Herrera, R. S., Torres, V., Oramas, A., Cairo, J.G.
y Herrera, M. 2009 .Efecto de la edad de rebrote en la composición química del
Pennisetum purpureum vc. Cuba CT 115. Rev. Cubana Cienc. Agríc. 43(1): 73-
79.
182. Van der Valk, A. 1985. Vegetation dynamics of praire glacial marshes. In:
White, J. (ed). The Population Structure of Vegetation. Dr. W. Junk Publishers,
Dordrecht. Pp.297-312.
183. Vandermeer, J., Brenner, A. and Granzow, I. 1998. Growth rates of tree height
six years after hurricane damage at four localities in eastern Nicaragua.
Biotropica 30: 502-509.
184. Vandermeer, J., Granzow, I. and Boucher, D. 1997. Contrasting growth rate
patterns in eighteen tree species from a post. Hurricane forest in Nicaragua.
Biotropica 29: 151-161.
185. Vargas, R. 1988. Fertilización y distancias de siembra en el pasto King grass
(Pennisetum purpureum x Pennisetum typhoides Staff y Hubbaard) en la
parroquia Pedro Vicente Maldonado. Tesis de Ingeniero Agrónomo. Quito.
Universidad Central del Ecuador. Facultad de ciencias Agrícola. 56p.
186. Vázquez, G.J.A. y Givnish, T.J. 1998 Altitudinal gradients in tropical forest
composition, structure, and diversity in the Sierra de Mazatlán, Jalisco, México.
Journal of Ecology. 86: 999 -1020.
187. Vélez, G.A. 2000. Alianza entre la guerra biológica y la bioprospección en la
Amazonía colombiana. Bogotá, p 44.
188. Verdecia, D. 2015. Composición química y metabolitos secundarios en seis
variedades de árboles, arbustos y leguminosas volubles en el Valle del Cauto.

Tesis en opción al grado Científico de Doctor en Ciencias. Universidad de
Granma. Cuba.p.100.
189. Villarreal H., Álvarez, M., Córdoba, S., Escobar, F., Fagua, G., Gast, F.,
Mendoza, H., Ospina, M. y Umaña, A.M. 2006. Manual de métodos para el
desarrollo de inventarios de biodiversidad. Programa de Inventarios de
Biodiversidad. Instituto de Investigación de Recursos Biológicos Alexander von
Humboldt. Segunda edición. Bogotá-Colombia. 236 p.
190. Wassenaar, T., Gerber, P., Verburg, P.H., Rosales, M., Ibrahim, M., Steinfeld,
H. 2007. Projecting land use changes in the Neotropics: the geography of pasture
expansion into forest. Global Environmental Change 17:86-104.
191. Watson, L. y Dallwitz M. J. 2014. The families of flowering plants Apocynaceae.
192. Watson, L. y Dallwitz, M. J. 2008. The Families of Flowering Plants. Interactive
Identification and Information Retrieval. Nordic Journal of Botany. Volume 14,
Issue 5: 486-490.
193. Wilson, J. R., Denium, B. y Engels, F. M. 1991. Temperature effects on
anatomy and digestibility of leaf and stem of tropical and temperate forage
species. Netherlands. Journal of Agricultural Science, 39:31-48.
194. Wilson, J.R. 1993. Shade effect on growth ofold established pastures in project
AM - 25. Improving sown grass productivity. Ed. Csiro, 62 pp. Australia.
195. Yaguano, C., Lozano, D., Neill, D. y Asanza, M. 2012. Diversidad florística y
estructural del bosque nublado del río Numbala, Zamora – Chinchipe, Ecuador:
“El bosque gigante” de Podocarpaceae adyacente al Parque Nacional
Podocarpus. Amazónica. Ciencia y Técnología 1(3): 226-247.


















