
UNIVERSIDAD AUTÓNOMA DE...
51
1 UNIVERSIDAD AUTÓNOMA DE BARCELONA FACULTAD DE BIOCIENCIAS MÁSTER EN GENÉTICA AVANZADA “EVALUACIÓN DEL FUNCIONAMIENTO DE UN VECTOR BICISTRÓNICO EN EL ANÁLISIS DE LA ACTIVIDAD DE SECUENCIAS IRES EN DIFERENTES LÍNEAS CELULARES” 2011 – 2012 MARÍA JOSÉ MUÑOZ
Transcript of UNIVERSIDAD AUTÓNOMA DE...

1
UNIVERSIDAD AUTÓNOMA DE BARCELONA
FACULTAD DE BIOCIENCIAS
MÁSTER EN GENÉTICA AVANZADA
“EVALUACIÓN DEL FUNCIONAMIENTO DE UN VECTOR
BICISTRÓNICO EN EL ANÁLISIS DE LA ACTIVIDAD DE
SECUENCIAS IRES EN DIFERENTES LÍNEAS CELULARES”
2011 – 2012
MARÍA JOSÉ MUÑOZ

2
LABORATORIO DE ANATOMÍA PATOLÓGICA
HOSPITAL VALL D’HEBRON
Certifico que el trabajo para disertación de Máster en Genética Avanzada de la candidata
María José Muñoz Guijarro ha sido concluido de conformidad con las normas establecidas;
por lo tanto, puede ser presentado para la calificación correspondiente.
Dr. Trond Aasen
Director
Barcelona, 11 de Julio 2012

3
Contenido 1 RESUMEN ............................................................................................................................ 5
2 INTRODUCCIÓN ................................................................................................................ 7
2.1 Mecanismos de inicio de traducción ...................................................................................... 7
2.1.1 Características del inicio de la traducción cap-depediente .......................................... 7
2.1.2 Características de la iniciación de la traducción cap-independiente ........................... 8
2.2 Revisión de los sitios de entrada internos del ribosoma (IRES) y las proteínas de unión al
ARN ....................................................................................................................................... 9
2.2.1 Elementos IRES .......................................................................................................... 9
2.2.2 Factores trans-activadores IRES (ITAFs) ................................................................... 9
2.3 IRES celulares ........................................................................................................................ 9
2.3.1 Identificación de elementos IRES en mARN celular .................................................. 9
2.3.2 Datos funcionales e ITAFs importantes para la actividad celular de secuencias IRES
………………………………………………………………………………………………………………………………10
2.4 Vectores bicistrónicos .......................................................................................................... 11
3 OBJETIVOS ....................................................................................................................... 13
3.1 Objetivos generales .............................................................................................................. 13
3.2 Objetivos específicos ............................................................................................................ 13
4 MATERIALES Y MÉTODOS .......................................................................................... 14
4.1 Descripción de la metodología ............................................................................................. 14
4.2 Reacción en cadena de la polimerasa (PCR) ........................................................................ 14
4.3 Purificación de los fragmentos de PCR usando el kit Wizard® SV gel and PCR clean-up
system. .................................................................................................................................. 16
4.4 Digestión con enzimas de restricción ................................................................................... 16
4.5 Formación de moléculas de ADN recombinante .................................................................. 16
4.5.1 Desfosforilación del vector ....................................................................................... 17
4.5.2 Ligación ..................................................................................................................... 17
4.5.3 Transformación ......................................................................................................... 18
4.6 Extracción de ADN plasmídico (mini y midipreps) ............................................................. 18
4.7 Análisis del ADN extraído ................................................................................................... 19
4.8 Test de mycoplasma ............................................................................................................. 19
4.9 Transfección a líneas celulares ............................................................................................. 20

4
4.9.1 Preparación y contaje de células para transfectar ...................................................... 21
4.9.2 Transfección celular usando fosfato de calcio .......................................................... 21
4.9.3 Mecanismo de transfección celular con jetPEITM
(polímero catiónico) .................... 22
4.10 Western blot ......................................................................................................................... 22
4.10.1 Lisado celular ............................................................................................................ 22
4.10.2 Cuantificación de proteínas mediante el kit bio-rad protein assay ............................ 23
4.10.3 Electroforesis en gel de poliacrilamida SDS-PAGE. ................................................ 24
4.10.4 Revelado de la membrana de nitrocelulosa ............................................................... 25
4.11 Cultivo de líneas celulares bajo condiciones de estrés ......................................................... 25
4.12 Construcción de partículas infeccionas con el plásmido pPLCX ......................................... 26
4.12.1 Construcción del plásmido pLPC-SRp20 .................................................................. 27
4.12.2 Construcción del plásmido pPLC-MFR .................................................................... 27
4.13 Producción de partículas virales usando células Phoenix .................................................... 27
5 RESULTADOS ................................................................................................................... 29
5.1 Amplificación mediante PCR de secuencias IRES .............................................................. 29
5.2 Clonación de secuencias IRES en el plásmido reportero eucariota pMFR .......................... 29
5.3 Extracción y análisis de ADN plasmídico (mini y midipreps) ............................................. 30
5.4 Cultivo de líneas celulares en medio DMEM y test de mycoplasma ................................... 31
5.5 Transfección del vector pMFR – IRES en células 293T ...................................................... 32
5.5.1 Análisis de la fluorescencia de las proteínas Cherry y GFP ...................................... 32
5.6 Western blot cuantificación de la expresión de Cherry y GFP ............................................. 33
5.6.1 Cuantificación de proteínas mediante el kit bio-rad protein assay ............................ 33
5.7 Producción viral en células Phoenix ..................................................................................... 37
5.8 Establecimiento de líneas celulares productoras de virus .................................................... 37
5.9 Cultivo de líneas celulares bajo condiciones de estrés ......................................................... 39
5.10 Evaluación de la expresión del plásmido pLPC-SRp20 ....................................................... 40
6 DISCUSIÓN ........................................................................................................................ 42
7 CONCLUSIONES Y FUTURO ........................................................................................ 46
8 BIBLIOGRAFIA ................................................................................................................ 48

5
1 RESUMEN
La traducción del mARN en células eucariotas típicamente ocurre mediante el
mecanismo “cap-dependiente”. Un modelo alternativo “mediado por IRES” es empleado
por varios virus y, recientemente este mecanismo de traducción cap-independiente también
se ha visto que es utilizado por ciertos mARNs eucariotas. La traducción IRES-dependiente
es frecuentemente activada bajo ciertas circunstancias incluyendo el estrés celular como: la
hipoxia, la falta de nutrientes, o durante la mitosis. Sin embargo, es poco conocido el
número de mARNs eucariotas que contienen elementos IRES funcionales, o como estos
son regulados por varios estímulos celulares (estrés), y la función de las denominadas
proteínas ITAF, las cuales modulan su represión traduccional, o activación.
El objetivo de este proyecto, por lo tanto, fue probar un reportero bicistrónico
fluorescente y su utilidad para analizar elementos IRES y como estos pueden ser
influenciados por estrés celular, modulación de las vías de señalización y su regulación por
factores ITAF específicos conocidos.
Se realizó amplificación por PCR de varias secuencias IRES conocidas, las mismas
que fueron clonadas para el análisis de la expresión cap-dependiente (visualizada por la
expresión de la proteína mCherry) y la mediada por IRES (que activa la traducción de
GFP). Una vez obtenidos los constructos, se analizó la expresión de Cherry/EGFP mediante
western blot, microscopía de fluorescencia y con un lector de fluorescencia múltiple. Los
constructos cherry-IRES-GFP fueron clonados en un vector retroviral y transfectados en
células Phoenix. Con este vector se produjeron partículas virales que se utilizaron para
infectar una variedad de líneas celulares como HaCaT, HeLa, A549 y MDA-MB-231.
Además, se evaluó el funcionamiento de los ITAF SRp20 y YB-1 mediante transfección en
líneas celulares que expresaban cherry-IRES-EGFP. Se observó una fuerte activación del
IRES de C-myc en la línea celular de cáncer mamario MDA-MB-468 determinada por
SRp20.
En resumen, en este estudio de prueba, nos hemos concentrado en el uso de un vector
fluorescente para estudiar elementos IRES, conocidos y desconocidos, se ha demostrado la
utilidad del vector, además se utilizó retrovirus para la generación de líneas celulares

6
reporteras estables, y la identificación de cómo el estrés celular, las vías de señalización y
los factores ITAFs, pueden modular la traducción cap-independiente de proteínas.

7
2 INTRODUCCIÓN
La biosíntesis de proteínas es un proceso complejo que consta de tres fases principales:
iniciación, elongación y terminación (Lodish, et al., 2000; Strachan & Read., 1999). Los
organismos eucariotas, tienen varias estrategias para regular la traducción del mARN
(Fitzgerald & Semler, 2009), siendo la iniciación la fase más compleja de la traducción, la
cual puede continuar por diferentes rutas (Sonenberg & Hinnebusch, 2009; Jackson, et al.,
2010). Además del modelo canónico cap-dependiente de reconocimiento y escaneo del
ribosoma, un método alternativo, cap-independiente, de traducción fue descrito por primera
vez en los picornavirus y, posteriormente en un subconjunto creciente de mARNs celulares.
En este mecanismo 5’-independiente de inicio de la traducción, los ribosomas son
reclutados hacia el mARN en un sitio de entrada interna del ribosoma (IRES). Los IRES’s
son elementos estructurales del ARN que funcionan en la unión de los ribosomas al mARN
y modulan la traducción de mARNs celulares especialmente durante el desarrollo, el estrés
(por ejemplo hypoxia en tumores) y la enfermedad, lo que sugiere que estas estructuras del
ARN son importantes para la dirección de la traducción en contextos celulares en los que la
traducción cap-dependiente esta down-regulado (Fitzgerald & Semler, 2009).
2.1 Mecanismos de inicio de traducción
2.1.1 Características del inicio de la traducción cap-depediente
La mayor parte de regulación traduccional se produce durante la fase de iniciación, por
lo tanto se ha puesto mayor énfasis en el estudio de los detalles moleculares de los
mecanismos convencionales y los mecanismos alternativos de inicio (Sonenberg &
Hinnebusch, 2009; Jackson, 2005). La traducción de los mARNs celulares ocurre con
mayor frecuencia a través del mecanismo de iniciación cap-dependiente. Los mARNs
celulares contienen un capuchón de 7-metil guanosina en el extremo 5’del ARN y esta
estructura es reconocida por el factor de inicio eucariota 4F (eIF4F) del complejo de unión
cap. El complejo consta de los factores de iniciación 4A, 4G y 4E, que reclutan al ribosoma
al sitio de inicio. El factor eIF4E es el componente del complejo de unión, mientras eIF4G
es una proteína de andamiaje que se une a eIF4E, eIF4A y al mARN. eIF4A es una helicasa
dependiente de ATP responsable de la relajación de la estructura secundaria y terciaria del
ARN durante el proceso. Esta actividad helicasa es estimulada por el factor asociado,

8
eIF4B. La subunidad 40S ribosomal se une a un complejo proteico que consta de eIF1,
eIF2-GTP-Met-tRNA (complejo ternario), eIF3, y eIF5. El ensamblaje del complejo
proteico, denominado 43S complejo de pre-inicio, se une al mARN en la estructura del cap
mediante la interacción de un dominio central de eIF4G con eIF3. El límite del complejo de
pre-inicio entonces examina la molécula de mARN hasta encontrar un codón de inicio
AUG que es reconocido en un contexto favorable (Fitzgerald & Semler, 2009).
Cuando se encuentra un codón de inicio adecuado, el GTP se hidroliza a GDP en
presencia de eIF5, y varios de los factores de iniciación se disocian. La subunidad
ribosómica grande entonces, se une a la subunidad pequeña para generar la competente
elongación 80S del ribosoma. En este punto se inicia la síntesis de proteínas, y los factores
de inicio se reciclan para posteriores rondas de iniciación. Algunas condiciones celulares,
tales como el estrés o una infección viral puede resultar en una baja regulación de la
traducción cap-dependiente, frecuentemente al interferir con los factores de iniciación
(Fitzgerald & Semler, 2009).
2.1.2 Características de la iniciación de la traducción cap-independiente
En el mecanismo de inicio de traducción cap-independiente, los ribosomas son
reclutados al ARN mediante un mecanismo desconocido. La subunidad 40S ribosomal
reconoce una secuencia de ARN, una estructura o un complejo de ribonucleoproteína
dentro de la región 5’ no codificante (5’ UTR), y el sitio de inicio en el codón de inicio
auténtico puede producirse varios cientos de nucleótidos downstream del extremo 5’. Esta
forma alternativa de inicio de traducción no requiere un cap en el extremo 5’ como un sitio
de reunión para los factores de iniciación, y el reconocimiento del cap de la subunidad
ribosomal 40S a través de la formación del complejo de unión formado por el factor eIF4F
intacto no se produce. El mecanismo de inicio cap-independiente, por lo tanto, involucra
características que se distinguen de la unión al cap canónica, se lleva a cabo por el modelo
de análisis ribosomal (Fitzgerald & Semler, 2009).

9
2.2 Revisión de los sitios de entrada internos del ribosoma (IRES) y las
proteínas de unión al ARN
2.2.1 Elementos IRES
Los poliovirus (PV) y el virus de encefalomiocarditis (EMCV) presentan genomas
virales de ARN sin capuchón y se ha visto que contienen secuencias en sus extremos 5’-
UTR que están mediando la traducción eficiente en células eucariotas a través de la unión
interna de los ribosomas (Pelletier & Sonenberg, 1988; Pelletier, et al., 1988; Jang, et al.,
1988). En casi todas las infecciones por picornavirus, la traducción cap-dependiente se
apaga y el ARN viral utiliza la traducción mediada por IRES para dirigir la síntesis de
proteínas virales (Fitzgerald & Semler, 2009).
2.2.2 Factores trans-activadores IRES (ITAFs)
Los elementos IRES típicamente consisten en largas y altamente estructuradas
secuencias de ARN que funcionan para reclutar ribosomas, aunque el mecanismo de
reclutamiento no ha sido totalmente definido y puede variar entre los diferentes elementos
IRES. Algunos, por ejemplo, requieren por lo menos un subconjunto de eIFs así como
ciertas proteínas de unión al ARN para facilitar la traducción mediada por IRES (Costa-
Mattioli, et al., 2004; Walter, et al., 2002). Estos factores no canónicos se denominan
factores trans-activadores IRES, o ITAFs. Varios ITAFs han demostrado, previamente, que
se unen a los elementos IRES y actúan en la traducción mediada por IRES (Mitchell, et al.,
2005; Holcik, et al., 2003; Schepens, et al., 2007; Paek, et al., 2008).
2.3 IRES celulares
2.3.1 Identificación de elementos IRES en el mARN celular
Ha sido difícil identificar las clases funcionales de IRES celulares porque existe una
secuencia primaria pequeña principal o porque presentan similitudes estructurales entre
ellos, lo que provoca variaciones en su eficiencia. Mientras que algunos IRES virales
pueden tener contextos comunes de síntesis y traducción en el citoplasma de células
cíclicas (dando lugar a clasificaciones estructurales y funcionales), un gran número de
contextos celulares que requieren regulación diferencial podría dar lugar a un número muy

10
grande de clases IRES con secuencia diferente y muchos componentes estructurales (Baird,
et al., 2006). Por lo tanto, los IRES celulares han sido a menudo discutidos en términos de
las condiciones que inducen su actividad o de las condiciones que resultan de su actividad.
Estructuras secundarias se han derivado de varios IRES celulares utilizando la estructura
enzimática y química obtenida de los transcritos, incluyendo c-Myc, Apaf-1, Kv1.4, y FGF-
2, entre otros (Le Quesne, et al., 2001; Mitchell, et al., 2003; Jang, et al., 2004; Bonnal, et
al., 2003).
2.3.2 Datos funcionales e ITAFs importantes para la actividad celular de
secuencias IRES
La identificación de muchos elementos IRES celulares se ha basado en la demostración
de la función de estos elementos y su capacidad para dirigir el inicio interno de la
traducción. El test más común para la actividad IRES es un ensayo de células de mamíferos
utilizando la transfección con vectores bicistrónicos reporteros. La expresión de un segundo
marco de lectura downstream, depende de secuencias IRES que dirigen la entrada interna
del ribosoma y el inicio de la traducción. Aunque es un primer paso importante en la
determinación de si una secuencia de ARN tiene actividad IRES, este enfoque ha sido
criticado debido a que los mARNs generados a partir de células transfectadas con vectores
plasmídicos bicistrónicos de ADN generalmente tienen una actividad IRES mucho mayor
que cuando se transfecta los ARNs correspondientes (Jimenez, et al., 2005). La diferencia
en los niveles de expresión se puede interpretar como un artefacto de la transfección con
ADN que conduce a la expresión reportera que es independiente de la función IRES (Van
Eden, et al., 2004; Kozak, 2005; Bert, et al., 2006). A pesar de esta deficiencia, la
traducción mediada por secuencias IRES se produce por un subconjunto de mARN
celulares.
Un IRES muy estudiado es el elemento IRES de c-myc (Nanbru, et al., 1997). Las
proteínas que pertenecen a la familia Myc tienen un papel en la proliferación y la apoptosis
celular, y la sobre expresión de Myc ha sido asociado con la tumorigénesis (Pelengaris, et
al., 2002). El ARN mensajero de c-myc tiene un cap en el extremo 5’, y puede ser
traducido ya sea mediante la vía cap dependiente o independiente (Stoneley, et al., 2000).
Por lo tanto, es probable que la variación en la abundancia de ITAFs específicos contribuya

11
a esta diferencia en la eficiencia de traducción. Se han identificado varios ITAFs que
estimulan la traducción de c-myc, incluyendo: PCBP2, IRP, unr, DAP5, YB-1, GRSF-1,
PSF, p54nrb (Cobbold, et al., 2008; Evans, et al., 2003; Martínez-Salas, et al., 2001).
Todo esto sugiere que los elementos IRES están regulados diferencialmente en
diversas condiciones fisiológicas. Mientras que las secuencias IRES celulares continúan
siendo identificadas y son añadidas a la creciente lista de mARN que poseen IRES, existen
factores ITAFs que también están siendo investigados por estos IRES. Además, hay
pruebas de que muchos ITAFs que regulan la traducción de secuencias celulares IRES son
proteínas que se encuentran tanto en el núcleo como en el citoplasma (Schepens, et al.,
2007; Graber & Holcik, 2007).
2.4 Vectores bicistrónicos
La expresión de dos secuencias codificantes diferentes bajo el control de un único
promotor permite la identificación del producto codificado por el segundo cistrón, lo que
asegura que el primer cistrón también se ha expresado. Algunos vectores bicistrónicos se
encuentran comercialmente disponibles basados en secuencias IRES a partir del virus
EMCV, en los cuales el codón AUG ha sido modificado (Pelletier, et al., 1988). Además,
los vectores bicistrónicos proveen la posibilidad de infectar células transfectadas tomando
en cuenta la base de la fluorescencia de las proteínas controladas por IRES, así como por la
expresión de la proteína de interés, fusionada a una proteína fluorescente separable
espectralmente (Bonnal, et al., 2003).
Además de revelar el insólito mecanismo de traducción, las secuencias IRES han sido
exitosamente introducidas entre 2 cistrones para la construcción de vectores bicistrónicos,
por ejemplo se ha demostrado la co-expresión simultánea de 2 proteínas, siendo una un
marcador de selección o un gen reportero y la otra el gen de interés (Martin, et al., 2006).
Por lo tanto, la identificación y el desarrollo de elementos IRES útiles para los vectores de
expresión se han vuelto una herramienta muy útil en la biotecnología.
En este estudio, evaluamos varias secuencias IRES que podrían permitir la expresión
génica bicistrónica en cultivos celulares mediante la transfección del plásmido pMFR, el
cual ha sido mejorado en el laboratorio de Anatomía Patológica, por Dr. Trond Aasen

12
(Grupo de Patología Molecular, VHIR). Se trata de una construcción bicistrónica
fluorescente que contiene la expresión de mCherry bajo una regulación cap-dependiente, un
sitio de clonación múltiple para insertar una secuencia IRES de interés, seguida por la
expresión de la proteína EGFP que depende directamente de la actividad de la secuencia
IRES clonada (Figura 1). Después de la clonación de las secuencias IRES candidatas en
este vector, se procederá a evaluar la activación de la traducción a través de estas
secuencias.
Figura 1. Estructura del vector pMFR
Construcción bicistrónica fluorescente, en la cual mCherry está regulado por la vía normal
de traducción, un MSC para insertar secuencias IRES de interés, seguida por la expresión
de la proteína EGFP

13
3 OBJETIVOS
3.1 Objetivos generales
Evaluar el comportamiento de un vector bicistrónico en el análisis de secuencias
IRES y factores ITAFs, mediante la transfección en líneas celulares.
Generación de partículas virales, portadoras de las secuencias IRES, para la
infección de varias líneas celulares y selección de clones estables.
Evaluar el efecto de varios ITAFs y el estrés celular como la falta de suero en la
eficiencia de traducción cap-dependiente Cherry e IRES-dependiente EGFP.
3.2 Objetivos específicos
Introducir secuencias IRES en el en MCS del vector pMFR.
Evaluar la funcionalidad de los constructos mediante la transfección en líneas
celulares.
Clonación del inserto Cherry-MCS-GFP en un plásmido retroviral pLPCX, para
generar partículas virales, e infectar líneas celulares y seleccionar con puromicina
para establecer líneas celulares estables.
Comparar los niveles de expresión de las secuencias IRES al someter a las células a
diferentes mecanismos de estrés o con la modulación de los niveles de ITAFs (como
YB-1 y SRp20).
Evaluar el efecto del ITAF SRp20, en líneas celulares.

14
4 MATERIALES Y MÉTODOS
4.1 Descripción de la metodología
Se analizaron varias secuencias IRES y (posibles ITAFs) de cMyc, Connexin 26,
Connexin 43, EMCV, 19kD (un IRES putativo dentro de la región codificante de connexin
43) relacionados con el proceso de traducción cap-independiente mediante el uso del vector
de expresión pMFR. Para este estudio se determinó las diferencias de expresión de estos
genes utilizando líneas celulares sometidas a situaciones de estrés. Se realizó la Reacción
en Cadena de la Polimerasa (PCR), con el fin de amplificar las secuencias de los genes a
analizar en el estudio. Los fragmentos amplificados fueron identificados en geles de
agarosa al 1%. A continuación, se procedió a realizar el corte de la banda del gel para poder
obtener el ADN amplificado y purificarlo para posteriormente evaluar su concentración
utilizando el Nanodrop 2000. Cada uno de los productos obtenidos, así como el vector,
fueron sometidos a digestión con las enzimas de restricción KpnI y BamHI; los productos
resultantes fueron purificados y ligados. Se procedió a la transformación bacteriana de la
cepa de E. coli DH5, y a la obtención de ADN plasmídico de alta calidad. Finalmente, se
transfectó líneas celulares y se evalúo la expresión proteica mediante Western Blot. Cuando
se obtuvieron los resultados de los tratamientos y se recopilaron todos los datos de las
líneas celulares analizadas.
4.2 Reacción en cadena de la polimerasa (PCR)
La Reacción en Cadena de la Polimerasa es una técnica in vitro que permite la
amplificación exponencial de una región específica del ADN, situada entre dos regiones de
secuencia conocida. Su eficiencia y especificidad son determinadas por muchos factores
como: calidad del ADN templado, diseño de los primers, concentración del MgCl2 y de la
ADN polimerasa (Newton y Graham, 1994).
En este estudio las condiciones de la PCR se estandarizaron tomando en cuenta la
capacidad de síntesis de la ADN polimerasa. Cada tubo de reacción para la amplificación
de la muestra contenía: 32µl de H2O Mili-Q, 5µl de buffer de PCR 10X, 3µl de MgCl2 25
mM, 5µl de dNTPs (dideoxinucleótidos trifosfato) a una concentración de 2 mM, 1,5µl de
cada uno de los primers a una concentración de 10µM, 1µl del ADN templado en una

15
concentración de 50ng/µl, 1µl de KOD Hot Start DNA Polymerase (Novagen®), en un
volumen final de 50µl. Todas las reacciones de amplificación se llevaron a cabo en un
termociclador Veriti® 96-Well Thermal Cycler (Life Technologies, Applied Biosystems
Inc., Van Allen Way, Carlsbad, CA., USA).
Los primes que se utilizaron es este estudio fueron diseñados para la amplificación de
fragmentos de 250, 450, 600 y 700 pares de bases. Las secuencias se muestran en la tabla 1.
Tabla 1. Secuencias de los genes analizados y primers utilizados para su amplificación
Genes Secuencia (forward y reverse)
rCx43 Fw: 5’- GCGACCACGAGGTATCAGCACTTTTC -3’
R: 5’-GCGGGATCCGTCTGGGCACCTCTCTTTCAC -3
Cx26 Fw: 5’- GCGGGTACCCAGGTGTTGCGCCCCGCGCG-3’
R: 5’-GCGGGATCCCTTCTACTCTGGGCGGTTT -3’
19Kd Fw: 5’- GCGGGTACCGGTGACTGGAGCGCCTTAGGC-3’
Rv: 5’- GCGGGATCCGAAGATGATGAAGATGGTTT- 3
Cmyc Fw: 5’- GCGCGTACCAATTCCAGCGAGAGGCAAGG-3’
Rv: 5’- GCGCGTACCAATTCCAGCGAGAGGCAGAGG-3
hCx43 Fw: 5’- CGGGTACCACGAGGTATCAGCACTTTTC -3’
Rv: 5’- GCGGGATCCGTCTGGGCACCTCTCTTTCAC -3’
Las condiciones utilizadas para la amplificación de todos los fragmentos fue:
desnaturalización inicial a 95°C por 2 minutos; seguido de 29 ciclos a 95°C por 20
segundos, 60°C por 30 segundos, con una disminución de 0,5°C por cada ciclo y 70°C por
15 segundos. Finalmente hubo un ciclo de extensión final a 72°C por 10 minutos, estas
condiciones se usaron para amplificar las secuencias de todos los genes descritos.
Después de la PCR, se realizó una electroforesis horizontal en geles de agarosa en
una concentración de 1%, y de este modo se determinó la presencia de los fragmentos
amplificados para cada uno de los genes estudiados. Los geles se prepararon con SYBR
Safe® (Invitrogen) como colorante para su observación en el transiluminador. Estos geles
fueron corridos en una cámara de electroforesis, sumergidos en solución de buffer TAE 1X,
a 70 voltios durante una hora. Para poder determinar el tamaño de los fragmentos
amplificados se utilizó un marcador de peso molecular de 100 pares de bases (pb), de la
casa comercial Fermentas.

16
4.3 Purificación de los fragmentos de PCR usando el kit Wizard® SV gel and
PCR clean-up system.
Se extrajo el fragmento de ADN de interés a partir de un gel de agarosa, usando un
bisturí. El mecanismo de purificación se llevo a cabo siguiendo las indicaciones del
fabricante, de acuerdo al protocolo de la casa comercial.
4.4 Digestión con enzimas de restricción
Los productos de amplificación por PCR, de los diferentes genes, así como del
vector fueron sometidos a la digestión mediante enzimas de restricción; en todos los casos
las enzimas empleadas fueron KpnI y BamHI, las mismas que dan como resultado la
formación de los sitios de reconocimiento para la inserción en el vector.
Cada tubo eppendorf utilizado para la digestión de los productos de PCR y del
vector contenía: 22µl de H2O Mili-Q, 3µl de fast digest buffer 10X, 1,5µl de cada una de
las enzimas (KpnI y BamHI) (Fermentas®), 1µl del ADN (purificado de PCR y del vector)
en una concentración de 1g/µl, en un volumen final de 30µl. Estos tubos fueron incubados
a 37°C durante 1 hora.
La visualización de los productos de las digestiones, fueron llevadas a cabo mediante
una electroforesis horizontal en geles de agarosa en una concentración del 1%. El vector
linealizado y los productos de PCR, obtenidos mediante la digestión con enzimas de
restricción, fueron purificados a partir del corte del gel de agarosa utilizando el mecanismo
descrito previamente.
4.5 Formación de moléculas de ADN recombinante
Los cuatro pasos esenciales de la clonación de ADN basado en células son:
1. Construcción de moléculas de ADN recombinantes mediante una unión covalente in
vitro (ligación) de los fragmentos de ADN deseado (ADN diana) a un replicón
(cualquier secuencia capaz de una replicación independiente del ADN). Este paso es
facilitado por el corte del ADN diana y las moléculas del replicón con

17
endonucleasas de restricción específicas antes de la unión de los diferentes
fragmentos de ADN utilizando la enzima ADN ligasa.
2. Transformación, las moléculas de ADN recombinante son transferidas hacia el
interior de las células hospederas (frecuentemente bacterianas o células de levadura)
en la que el replicón elegido puede ser sometido a la replicación del ADN
independientemente del cromosoma de la célula hospedadora.
3. Propagación selectiva de los clones, que se da en dos pasos. Inicialmente las células
transformadas son cultivadas en placas con agar con el fin de promover el
crecimiento de colonias aisladas. Estos son los clones (poblaciones de células
idénticas, todas descendientes de una sola célula). Subsecuentemente, las colonias
individuales pueden seleccionadas de una placa de cultivo para ampliar su
crecimiento en medio líquido.
4. Aislamiento de clones de ADN recombinantes mediante la cosecha de los cultivos
bacterianos y el aislamiento selectivo del ADN recombinante (Strachan & Read,
1999).
4.5.1 Desfosforilación del vector
El proceso de desfosforilación de extremos 5’ con la enzima FastAPTM
Thermosensitive Alkaline Phosphatase (Fermentas®), se realizó en un volumen final de
20µl de H2O Mili-Q, 2µl de FastAPTM
buffer 10x (100 mM Tris-HCl (pH 8.0 at 37°C), 50
mM MgCl2, 1 MKCl, 0.2% Triton X-100 and 1 mg/ml BSA), 1l de enzima, 1l de vector
lineal en una concentración de 80ng/µl. Los tubos fueron incubados a 37°C durante 10
minutos.
4.5.2 Ligación
Los productos de amplificación por PCR digeridos con las enzimas de restricción, así
como el vector linealizado y desfosforilado, fueron ligados utilizando la enzima T4 DNA
ligasa, la misma que requiere ATP como cofactor. La reacción de ligación se llevo a cabo a
22oC, durante toda la noche. Cada tubo eppendorf contenía, en un volumen total de 20µl:
2µl de T4 DNA ligase buffer 10X (400mM Tris-HCl, 100mM MgCl2, 100mM DTT, 5mM

18
ATP (pH 7.8 at 25°C)), 1l de enzima, 80ng/l de vector lineal, la concentración de trabajo
del inserto se calculó con la siguiente fórmula:
vectorbp
insertobpvectornginserto
.
.*.
4.5.3 Transformación
Para la transformación se utilizaron células competentes de la cepa de bacterias E.
coli DH5, las mismas que se conservan a -80oC, y se mantienen en hielo hasta que se
encuentren totalmente descongeladas. El mecanismo de transformación se llevó a cabo de
la siguiente manera: se colocó 50l de bacterias en un eppendorf y se agregó 5l de
producto de ligación, se mantuvo la mezcla en hielo durante 30 minutos. Se realizó un
choque térmico a 42oC, durante 42 segundos. Posteriormente, se colocaron los tubos en
hielo durante 1 minuto. Y finalmente, se colocó 1ml de medio LB líquido sin antibiótico y
se dejaron las muestras en agitación a 37oC durante 1 hora.
Se procedió a retirar las muestras, se las centrifugó durante 3 minutos a 3000 rpm y
se sembró 100l de pellet en placas de agar LB con el antibiótico kanamicina. Por último,
las placas fueron incubadas toda la noche a 37oC.
Se prepararon además dos controles: 1) Negativo: sólo las bacterias para verificar la
resistencia al antibiótico y 2) Positivo: plásmido sin digerir transformado.
Una vez obtenidas las colonias se procedió a ampliar el crecimiento bacteriano
sembrando colonias individuales en 3 ml de medio LB con kanamicina en concentración
1:200 ml. Los cultivos se colocaron en incubación con agitación a 37oC, durante toda la
noche; se trabajó con 3-4 repeticiones. Al siguiente día se realizó la extracción de ADN.
4.6 Extracción de ADN plasmídico (mini y midipreps)
La extracción de ADN de miniprep se realizó a partir de 3ml medio de cultivo líquido
LB, incubado durante toda la noche a 37oC y con agitación de colonias individuales que
fueron seleccionadas con antibiótico siguiendo las indicaciones del fabricante (Promega®).

19
Así mismo, para la extracción de ADN plasmídico midiprep de gran rendimiento y
pureza se utilizó el kit: Extracción de ADN midiprep con el kit Pure link™ hipure plasmid
filter purification kit (Invitrogen). En este caso se dejó incubando 50 ml de medio líquido
LB con antibiótico y la colonia de bacteria respectiva. Se siguió el protocolo descrito por el
fabricante.
4.7 Análisis del ADN extraído
Un ADN de alto peso molecular principalmente libre de contaminantes es muy
importante para la aplicación de técnicas moleculares, como la clonación de secuencias de
ADN que es la base para el resto del análisis (Manak, 1993; Ausbel et al., 1995).
Para determinar la calidad y cantidad del ADN, se utilizó el espectrofotómetro
Thermo Scientific NanoDrop™ 2000. Para lo cual se calibró el equipo utilizando 1µl del
agua con la cual se diluyó el ADN. Posteriormente se midió la concentración de cada ADN
colocando 1µl de muestra sobre el lector.
Luego de haber determinado la calidad y cantidad de ADN, este material fue diluido
a una concentración final de 1µg/µl.
4.8 Test de mycoplasma
Los cultivos celulares frecuentemente se contaminan con diferentes especies de
mycoplasmas, lo cual altera el crecimiento celular, los patrones enzimáticos, la
composición de la membrana e induce anormalidades cromosómicas. Dichas alteraciones
influyen considerablemente en la toma de decisiones de los resultados obtenidos. Los
mycoplasmas son un grupo de microrganismos que pertenecen a la clase Mollicutes y se
caracterizan por ser los procariotes más pequeños de vida libre hasta ahora descritos.
Carecen de pared celular, el tamaño de su genoma oscila entre 577 y 2220 kb, son exigentes
desde el punto de vista nutricional y difíciles de cultivar (Rivera-Tapia, et al., 2002). Los
métodos para detección de contaminación por mycoplasmas son el cultivo microbiológico,
técnicas de microscopia, métodos bioquímicos y uso de técnicas de biología molecular. El
protocolo usado para la detección de mycoplasma se describe a continuación:

20
Se tomó 1 ml de medio de cultivo de cada línea celular a analizar. Se centrifugó la
muestra durante 1 minuto a 250 rpm. Se retiró el sobrenadante y se volvió a centrifugar a
máxima velocidad durante 10 minutos. Se aspiró el sobrenadante y se eliminó. Se
resuspendió el pellet en 10 l de buffer solution (kit). Se sometió a la muestra a choque
térmico durante 3 minutos a 95oC.
La muestra fue preparada en tubos de 0,2 ml. La reacción contenía 7 l de agua MiliQ
estéril, 2 l de reaction mix (kit), 1 l de la muestra con buffer solution. Se preparó bajo las
mismas condiciones un control positivo, incluido en el kit, y un control negativo que
únicamente contenía agua.
Las condiciones en el termociclador fueron: denaturación inicial a 94oC durante 30
segundos, seguido de 36 ciclos a 94oC 30 segundos, annealing 60
oC durante 2 minutos y
extensión a 72oC durante 1 minuto. Finalmente se realizó una extensión final a 72
oC
durante 5 minutos. Las muestras fueron mantenidas a 16oC. La comprobación se realizó
mediante electroforesis en gel de agarosa al 2%.
4.9 Transfección a líneas celulares
Es un mecanismo mediante el cual se introduce un material genético externo en
células eucariotas mediante plásmidos, vectores víricos (en este caso también se habla de
transducción) u otras herramientas para la transferencia. La transfección de células
animales generalmente se lleva a cabo abriendo poros o "agujeros" transitorios en la
membrana plasmática de las células, lo que permite el paso del material genético. Hay
varios métodos para introducir DNA en una célula eucariota. Se usan muchos materiales
como vehículos para la transfección, divisibles en tres tipos: polímeros (catiónicos),
liposomas y nanopartículas. Uno de los métodos más baratos y óptimos es la transfección
mediante fosfato de calcio (Graham & Van der, 1973). En este mecanismo se utiliza una
solución salina tamponada con HEPES buffer que contiene iones fosfato y se combina con
una solución de cloruro de calcio que contiene el ADN a transfectar. Cuando ambas se
combinan, se forma un precipitado fino formado por el calcio cargado positivamente y el
fosfato cargado negativamente. Esta suspensión se añade a las células que se quieren
transfectar (normalmente un cultivo celular en monocapa). Mediante un proceso no

21
comprendido completamente, las células toman parte del precipitado, y junto con él, el
ADN.
4.9.1 Preparación y contaje de células para transfectar
Se tomó células en cultivo de la línea celular humana HEK-293, las cuales fueron
preparadas de la siguiente manera:
Se realizaron 2 lavados con 10 ml de PBS 1X estéril. Se añadió 1.5ml de tripsina y se
colocó las placas a 37oC en la incubadora durante 2 minutos. Se añadió 3 ml de medio de
cultivo DMEN (Dulbecco‘s modified Minimum Essential Medium) Gibco® suplementado
(5.5 ml de penicilina-estreptomicina, 10% BSA, 5 ml L-Glutamina) para inactivar la
tripsina. Se re suspendió las células y se las transfirió a un tubo falcon de 15ml. Se
centrifugó las células a 1200 rpm durante 5 minutos a temperatura ambiente. Se retiró el
medio y se colocó nuevamente 5 ml de medio y se resuspendió las células.
A continuación, se contó las células para sembrar un número determinado de las
mismas para transfectar. Se colocó 1ml de las células resuspendidas en un tubo eppendorf
de 1,5ml. De esta alícuota, se tomó 10l de células y se colocó sobre la placa Neubauer.
Una vez obtenido el total de células se procedió a sembrar 3 millones de células en placas
p100, y se agregó 10ml de medio. Las placas se colocaron en la incubadora con 5% de CO2
a 37oC.
4.9.2 Transfección celular usando fosfato de calcio
Para la transfección se utilizó el mecanismo del fosfato de calcio, para lo cual se
prepararon dos tubos falcon de 15ml por cada muestra a transfectar. Un tubo contenía 50l
de CaCl 2.5M, 10g de ADN de cada vector en un volumen total de 500l. En el otro tubo
se colocó 500l de HBS. Una vez obtenida la mezcla del vector, se colocó gota a gota sobre
el HBS para permitir la formación de los cristales de ADN. Se mezcló 10 segundos en el
vortex y se dejó reposar durante 15 minutos a temperatura ambiente. Finalmente, la mezcla
se añadió en las células sembradas el día anterior, a las cuales, previamente, se les cambió
el medio. Se dejó incubando durante 24 horas a 37oC con CO2. A las 24 horas de
transfección, se realizó un control de las células y se procedió a cambiar el medio.

22
4.9.3 Mecanismo de transfección celular con jetPEITM
(polímero catiónico)
JetPEI™ es un reactivo de transfección de gran alcance que asegura una efectiva y
reproducible transfección con una baja toxicidad. El reactivo, permite una transfección
superior in vitro en comparación con varios lípidos catiónicos y polímeros. JetPEI ™ ha
demostrado una buena entrega de material genético a diversas líneas celulares establecidas,
así como células primarias (Horbinski, et al., 2001). JetPEI™ compacta el ADN en
partículas con carga positiva, capaces de interactuar con proteoglicanos aniónicos en la
superficie celular, de esta manera ingresa en las por endocitosis; además posee la propiedad
única de actuar como una "esponja de protones" que amortigua el pH endosomal y protege
el ADN de la degradación (jetPEITM
Polypus Transfecction, manual).
4.10 Western blot
Es una técnica analítica que permite detectar proteínas específicas en una muestra
determinada (una mezcla compleja de proteínas, como un extracto tisular). Mediante una
electroforesis, electroblotting, en gel de SDS-poliacarilamida se separan las proteínas de
acuerdo al peso molecular, estructura, hidrofobicidad, etc. Posteriormente, las proteínas son
transferidas a una membrana adsorbente (típicamente de nitrocelulosa o de PVDF) para
poder buscar la proteína de interés con anticuerpos específicos contra ella. Finalmente, se
detecta la unión antígeno-anticuerpo por actividad enzimática, fluorescencia entre otros
métodos. Debido al marcaje, el anticuerpo produce una banda oscura sobre un film de rayos
X al momento de revelado (autorradiograma). De esta forma se puede estudiar la presencia
de la proteína en el extracto y analizar su cantidad relativa respecto a otras proteínas
(Sambrook, et al., 1989; Berg, et al., 2002).
4.10.1 Lisado celular
Los cultivos celulares mantenidos en incubación con CO2 y a 37oC durante 48-72
horas, fueron retirados de la incubadora y mantenidos en hielo para su procesamiento. Las
placas de células primeramente fueron lavadas 2 veces con 5ml de PBS 1x.
Posteriormente se preparó 1ml de buffer NP 40 de lisis (hepes 1M, NaCl 2,5M; NP
40, glicerol, MgCl2 1.5M, EDTA 0.5M), al mismo que se le adicionó 1l de DTT, 10l de

23
PMSE, 20l de Naf, 5l de NaPPi (0.1M), 1l de Na3VO4 y 10l de -glicerofosfato (1M).
Una vez preparado en buffer se colocó entre 30-150l de buffer en cada una de las placas.
Las placas fueron raspadas para obtener el lisado celular, se colocó en eppendorfs
estériles de 1.5ml y se mantuvo en hielo durante 15 minutos. Finalmente se centrifugó
durante 20 minutos a 14.000 rpm a 4oC.
4.10.2 Cuantificación de proteínas mediante el kit bio-rad protein assay
El kit de Bio-Rad, se basa en el método Bradford, el mismo que es un método
simple y preciso para determinar la concentración de proteínas solubilizadas. Requiere la
adición de un tinte acídico a la solución proteica, y la subsecuente medición a 595 nm con
un espectrofotómetro o un lector de microplacas. El ensayo se basa en la fijación de
colorante, en el cual un cambio diferencial de color se produce en respuesta a varias
concentraciones proteína. La absorbancia máxima de la solución acídica del tinte azul
brillante de Coomassie® G-250 cambia de 465 nm a 595 nm cuando la unión a proteínas
ocurre. El colorante azul de Coomassie se une a principalmente residuos de aminoácidos
básicos y ácidos aromáticos, especialmente arginina.
Para medir la concentración de las proteínas, se preparó el reactivo de Bradford
diluyendo 1 volumen de reactivo concentrado (5x) con 4 partes de agua miliQ (1x) y se
mezcló con el vortex. En una placa de 96-well, en cada pocillo, se colocó 200 μl del
reactivo de Bradford, diluido, de acuerdo al número de muestras y a la escalera patrón. Por
último se colocó 1l de muestra, tanto de proteína como de recta patrón y se mezcló por
pipeteo. Se incubó durante 5 minutos a temperatura ambiente y se midió la absorbancia a
595 nm. La regla patrón para la medición de la concentración de proteína se preparó como
se indica el fabricante.
Una vez obtenida la medición de las proteínas, se calculó la concentración de cada una,
y las muestras fueron mantenidas a una concentración de 2g/l, para lo cual se agregó la
cantidad de buffer de lisis adecuada a cada una.

24
4.10.3 Electroforesis en gel de poliacrilamida SDS-PAGE.
Una molécula con una carga neta puede moverse en un campo eléctrico, mediante
electroforesis, lo cual permite la separación de proteínas y otras macromoléculas como el
ADN y el ARN. La velocidad de migración de una proteína (o cualquier molécula) en un
campo eléctrico depende de la intensidad del campo eléctrico, la carga neta de la proteína y
un coeficiente de fricción. Aquellas moléculas que sean más pequeñas que el tamaño del
poro del gel se moverán más rápido a través del gel, mientras que las moléculas mucho más
grandes que los poros son casi inmóviles. Las moléculas de tamaño intermedio se mueven a
través del gel con diferentes grados de facilidad (Sambrook, et al., 1989; Berg, et al., 2002).
Las proteínas se pueden separar en base a su masa por electroforesis bajo
condiciones denaturantes. La mezcla de proteínas es primeramente disuelta en una solución
de dodecil de sodio (SDS), un detergente aniónico que rompe caso todas las interacciones
no covalentes en las proteínas nativas. Mercaptoetanol (2-tioetanol) o ditiotreitol (DTT)
también es añadido para reducir los enlaces disulfuro. Este complejo SDS con una proteína
denaturada posee una gran carga negativa neta que es aproximadamente proporcional a la
masa de la proteína. La carga negativa adquirida con la unión del SDS es usualmente
mucho mayor que la carga de la proteína original, esta carga nativa se vuelve entonces
insignificante. Una vez completada la electroforesis, las proteínas en el gel se pueden
visualizar mediante tinción con plata o con un tinte como el azul de Coomassie, el cual
revela una serie de bandas. El marcaje radiactivo puede ser detectado colocando una hoja
de film de rayos X sobre el gel, mediante auto radiografía (Sambrook, et al., 1989; Berg, et
al., 2002).
Para la electroforesis se preparó un gel de electroforesis cuya concentración fue de
12%, en un volumen de 10m, utilizando 3.3 ml de H2O miliQ, 4 ml de mix de acrilamida
30%, 2.5 ml de Tris 1.5M (pH 8.8), 100 l de SDS 10%, 100 l de persulfato de amonio
10% y 4 l de TEMED. Para su polimerización se cubrieron los vidrios hasta el borde con
Isopropanol. Una vez polimerizado el gel, se retiro el isopropanol y se preparó el gel
Stacking fue a una concentración del 5% en un volumen final de 4 ml, para lo cual e
mezcló 2,7 ml de H2O miliQ, 670 l de mix de acrilamida 30%, 500 l de Tris 1.0 M (pH
6.8), 40 l de SDS 10%, 40 l de persulfato de amonio 10% y 4 l de TEMED (Sambrook,

25
et al., 1989). Una vez polimerizado el gel se procedió a preparar las muestras para la
electroforesis.
Todas las muestras fueron preparadas en un volumen total de 20 l, se adicionó 2,5
l de buffer de carga 5x y 0,2 l de DTT. Se desnaturalizó las muestras previamente
durante 5 minutos a 96oC. La electroforesis se llevo a cabo en una cubeta Bio Rad, durante
3 horas a 80V. El buffer de corrida fue preparado a una concentración 1x. Una vez
terminada la electroforesis se procedió a la transferencia del gel a la membrana de
nitrocelulosa, para lo cual primeramente se activó la membrana colocándola durante 1
minuto en metanol 100%, se lavó con agua destilada y finalmente se la mantuvo en buffer
de transferencia 1x que contenía metanol. La electroforesis de trasferencia se llevó a cabo
durante una hora y media a 300 mA.
4.10.4 Revelado de la membrana de nitrocelulosa
Una vez terminada la electroforesis de transferencia, la membrana fue bloqueada
durante 30 minutos utilizando el buffer de leche (DifcoTM
Skim Milk) (TBS 1x y 0.01%
Tween 20). Posteriormente se procedió a lavar la membrana 3 veces con el buffer TBST
durante 10 minutos cada una. Por último, la membrana fue incubada con agitación durante
una hora con los anticuerpos -His y -GFP a temperatura ambiente. Una vez transcurrido
el tiempo de incubación la membrana fue lavada, nuevamente, 3 veces y cada lavado
durante 10 minutos.
A continuación, se preparo 1 ml de solución de revelado, se colocó la membrana sobre
el casete y se adicionó la solución sobre la membrana. Las condiciones de revelado
dependieron de la intensidad de la proteína pero se mantuvo el film de rayos X entre 10 y
15 segundos para una exposición mínima y de 1 a 3 minutos para obtener una máxima
exposición.
4.11 Cultivo de líneas celulares bajo condiciones de estrés
El uso de sistemas de cultivos celulares para evaluar los efectos de la toxicidad
principalmente, de agentes anticancerígenos empezó 50 años atrás. Existe una gran
variedad de ensayos para evaluar la sensibilidad de las células a situaciones de estrés

26
provocado por cambios en la expresión génica de las mismas ó mediante la adición de
sustancias químicas que pueden alterar el metabolismo celular (Langdon, 2010).
En este ensayo se utilizaron las líneas celulares SW620 y HaCat para probar diferentes
tipos de estrés celular. Para ello se transfectaron células en placas de 96-well con,
aproximadamente, 10.000 células por tratamiento y por línea celular. Los tratamientos se
realizaron por triplicado y las condiciones fueron: medio de cultivo DMEM con 10% FBS
(control), medio DMEM con 0% FBS, PP242 2.5 M, CoCl2 250 μM y CGP 40M. Las
células fueron incubadas a 37oC durante 24 horas con 5% de CO2.
Para la evaluación de la expresión de la proteína Cherry y GFP, transcurridas las 24
horas de incubación, se realizó la medición de la absorbancia de las proteínas. Las
mediciones se realizaron en los intervalos de 485/535 nm para GFP y entre 544/590 nm
para Cherry. Las mediciones fueron realizadas por duplicado, una primera vez con el medio
de cultivo. Para la segunda medición de fluorescencia, primero se realizó 2 lavados de las
placas con 30 l de PBS 1x, posteriormente se colocó 30 l de buffer de lisis NP40 sobre
cada tratamiento y se dejó reposar durante 10 minutos. La medición de la absorbancia se
realizó bajo las mismas condiciones anteriores.
4.12 Construcción de partículas infeccionas con el plásmido pPLCX
Es posible construir vectores retrovirales de replicación competente añadiendo
secuencias a virus existentes, pero más comúnmente el diseño involucra el remplazo de las
secuencias retrovirales para la creación de vectores deficientes de replicación. Además, la
cantidad de ADN externo que debe ser acomodado en los vectores de replicación
competente es mucho más pequeña que la que se debe colocar en los vectores de
replicación defectiva. La expresión de proteínas retrovirales en la mayor parte de retrovirus
oncogénicos que se producen naturalmente es dirigida por un solo promotor en las regiones
5’ LTR (Long Terminal Repeats), y la expresión de múltiples regiones virales codificantes
se alcanza mediante el splicing alternativo. Sin embargo, el diseño del vector no se limita al
uso de un único promotor retroviral con splicing alternativo. Otras estrategias incluyen el
uso de múltiples promotores, inserción de genes en orientación reversa, y la utilización de
secuencias IRES (Coffin, et al., 1997).

27
4.12.1 Construcción del plásmido pLPC-SRp20
A partir del vector pEGFPC1-SRp20 se realizó la extracción de la secuencia SRp20,
que es un factor de splicing, mediante la digestión con las enzimas de restricción EcoRI y
NotI. Al mismo tiempo se trató al vector pLPCX con las mismas endonucleasas como se ha
descrito anteriormente. Por último los fragmentos fueron ligados y transformados para
obtener el vector para transfectar células de carcinoma mamario MDA-MB-468.
4.12.2 Construcción del plásmido pPLC-MFR
A partir del vector pMFR se realizó la extracción de las secuencias IRES EMCV,
19kD (secuencia interna en la región codificante del gen connexin 43) y c-Myc mediante la
digestión con las enzimas de restricción BglII y NotI. El vector pLPCX fue digerido con las
mismas enzimas como se ha descrito. Los fragmentos fueron ligados y transformados para
obtener el vector retroviral para infectar células Phoenix (derivadas a partir de las células
HEK-293).
4.13 Producción de partículas virales usando células Phoenix
Durante las últimas dos décadas, los vectores retrovirales han surgido exclusivamente
como un sistema de transferencia de genes de gran alcance. Las características que dotan a
los retrovirus con su potencial patogénico también los hacen atractivos para la ingeniería de
vectores seguros para transferencia genética. Por ejemplo, los retrovirus se integran
establemente en muchos tipos celulares. Además, la expresión de los retrovirus puede ser
constitutiva o inducible usando una variedad de configuraciones del promotor (Pear, et al.,
1993; Hitoshi, et al., 1998). Para el empaquetamiento del vector se utilizó líneas celulares
Phoenix (empaquetadores de retrovirus), mediante la transfección de las mismas con
fosfato cálcico; para la recolección de las partículas virales se siguió el protocolo descrito a
continuación:
A las 24 horas de la transfección en células Phoenix, se aspiró el medio de cultivo
DMEM (suplementado) que contenía las partículas virales, el mismo que fue pasado a
través de filtro de 0,2 m. Sobre las células Phoenix se añadió 10 ml de medio DMEM y se
las dejó en la incubadora a 37°C. El medio filtrado, que contenía los virus, fue colocado en
tubos falcon de 15 ml y se añadió 10 Polybrene (0.8mg/ml) por cada ml de sobrenadante.

28
La mezcla viral y el polybrene fueron distribuidos sobre las líneas celulares HOP-62,
MDA-MB-231, 293T y A549 para su infección y producción estable del virus.
El proceso de infección sobre líneas celulares se repitió a las 24 horas. Finalmente,
sobre las células infectadas se colocó medio de selección y se las mantuvo en paso normal
de selección durante una semana.

29
5 RESULTADOS
5.1 Amplificación mediante PCR de secuencias IRES
Para amplificar los correspondientes IRES de 19kD, rCx43, hCx43 (Cx43 humano) y
Cx26, se utilizó como base un plásmido que ya tenía el la secuencia clonada. Usando
primers específicos diseñados para introducir las dianas KpnI - BamHI se amplificó los
fragmentos por PCR y se comprobó mediante electroforesis en gel de agarosa (figura 2).
Figura 2. Amplificación mediante PCR de los genes Cx26, eCx43, C-myc y hCx43, gel
de agarosa 1%.
Productos de PCR correspondientes a los fragmentos de los genes Cx26, rCx43 (filas 2, 3-
5, 6) con un tamaño de 200 pares de bases. En la fila 8, el producto de amplificación del
gen c-Myc con un tamaño de 350 pares de bases. Las filas 10 y 11 el producto de
amplificación del gen 19kD con un peso de 750 pares de bases. Además los marcadores de
peso molecular de 1kb, en las fila 1 y de 100 pares de bases en la fila 12. Los fragmentos
de PCR fueron amplificados usando ADN plasmídico.
5.2 Clonación de secuencias IRES en el plásmido reportero eucariota pMFR
Una vez obtenidos los fragmentos amplificados mediante PCR de las secuencias
IRES, se cortaron las bandas correspondientes del gel y se procedió a purificarlas con el kit
de promega. A continuación, el vector pMFR y los productos de PCR purificados, fueron
digeridos con las enzimas de restricción KpnI y BamHI, lo cual permitió lineralizar el
plásmido y generar los sitios de inserción del producto de PCR. Finalmente se realizó la
transformación bacteriana para la obtención de colonias.
1 2 3 4 5 6 7 8 9 10 11 12
Mw Cx26 rCx26 Cmyc 19kD MW 1kb 100pb
200pb
350pb
750pb

30
De cada una de las placas transformadas con las secuencias IRES, en agar con
kanamicina, se escogieron 2-3 colonias y se hizo el cultivo más grande miniprep. Se extrajo
el ADN plasmídico usando el kit de extracción de miniprep y se verificó la presencia del
inserto mediante la digestión con las enzimas KpnI – BamHI, obteniéndose así las bandas
del vector lineal, sin inserto, y los fragmentos de cada uno de los genes amplificados
mediante PCR como se indica en la figura 3.
Figura 3. Verificación mediante digestión con las enzimas de restricción KpnI y
BamHI de la transformación bacteriana, gel de agarosa 1%.
En la primera fila se observa el marcador de peso molecular de 1kb, en las filas 2-3, 5-6 y
13-14 se observa un fragmento de 200 pares de bases de los fragmentos Cx26, rCx26 y
hCx43; en la fila 8 un producto de 350 pares de bases, que corresponden al fragmento c-
Myc. La banda de 6000 pares de bases corresponde al vector pMFR lineal.
5.3 Extracción y análisis de ADN plasmídico (mini y midipreps)
El ADN obtenido mediante extracción de miniprep a partir de 3 ml de cultivos
bacterianos fue de alto peso molecular, así como también el ADN obtenido a partir del
protocolo de Midiprep a partir de 50 ml de medio de cultivo. La concentración obtenida y
la calidad del ADN se muestran en la figura 4, obtenida a partir de la medición de
concentración con el Nanodrop.
MW 1kb
MW 100 pb
Cx26 rCx26 Cmyc 19kD hCx43
200pb
750pb
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
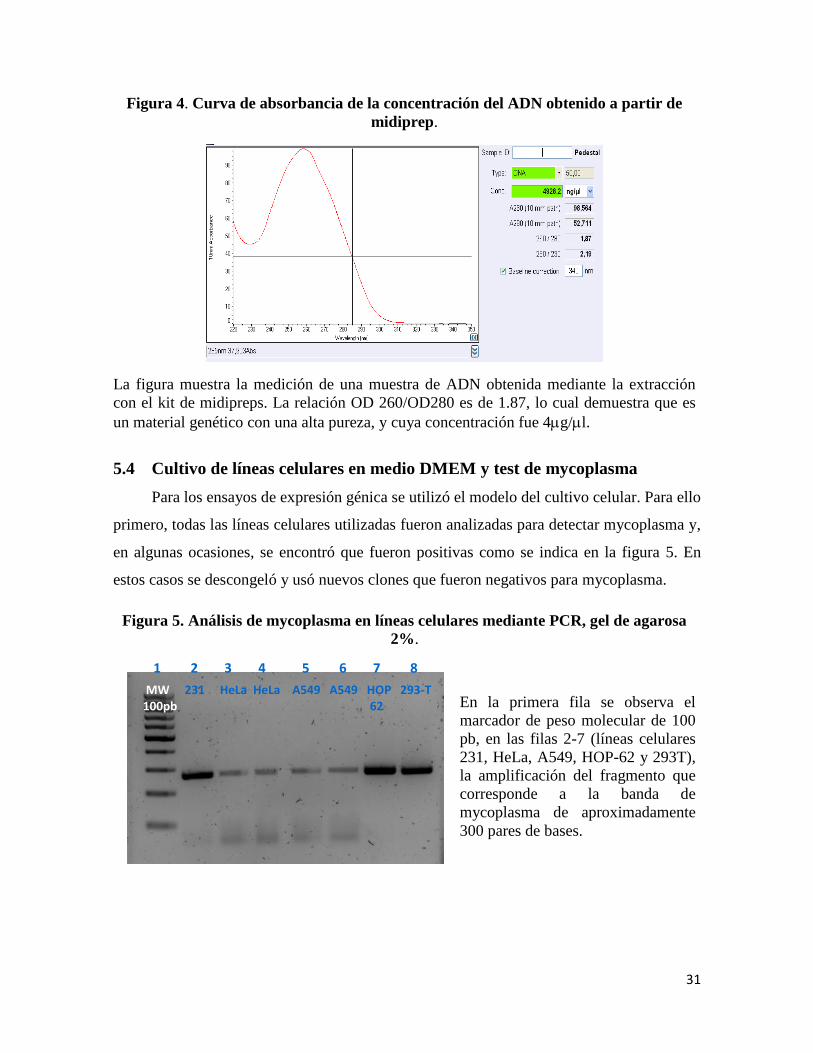
31
Figura 4. Curva de absorbancia de la concentración del ADN obtenido a partir de
midiprep.
La figura muestra la medición de una muestra de ADN obtenida mediante la extracción
con el kit de midipreps. La relación OD 260/OD280 es de 1.87, lo cual demuestra que es
un material genético con una alta pureza, y cuya concentración fue 4g/l.
5.4 Cultivo de líneas celulares en medio DMEM y test de mycoplasma
Para los ensayos de expresión génica se utilizó el modelo del cultivo celular. Para ello
primero, todas las líneas celulares utilizadas fueron analizadas para detectar mycoplasma y,
en algunas ocasiones, se encontró que fueron positivas como se indica en la figura 5. En
estos casos se descongeló y usó nuevos clones que fueron negativos para mycoplasma.
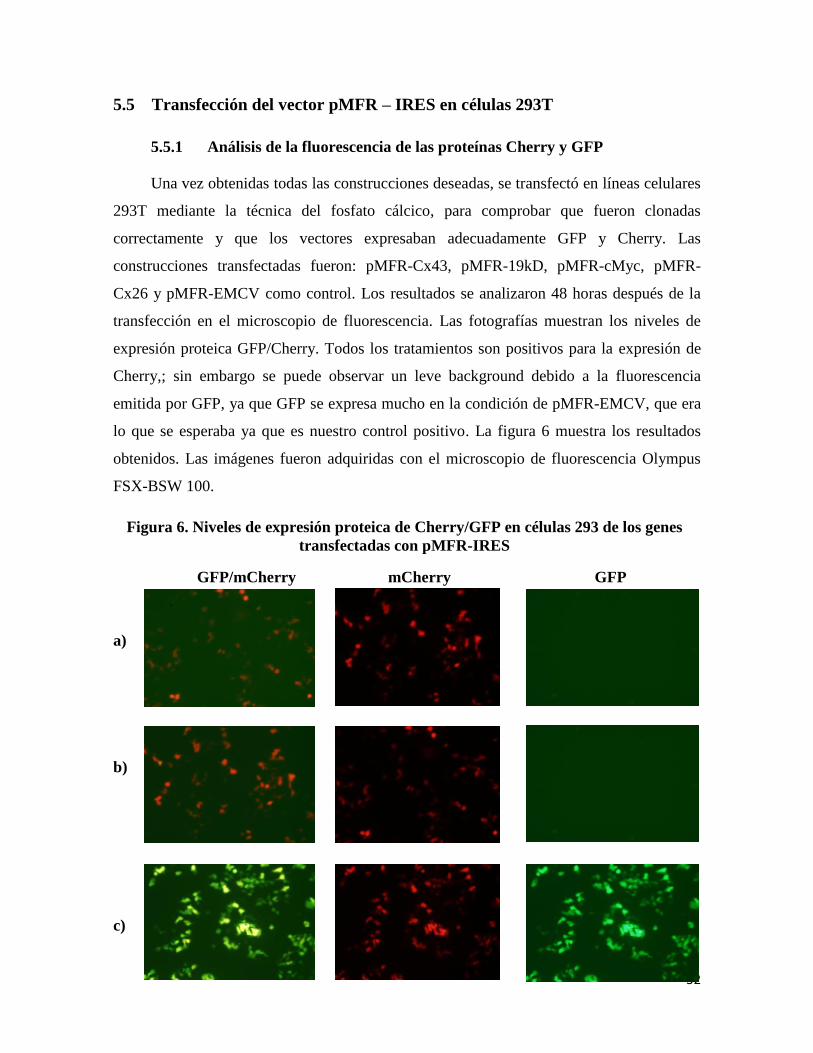
Figura 5. Análisis de mycoplasma en líneas celulares mediante PCR, gel de agarosa
2%.
En la primera fila se observa el
marcador de peso molecular de 100
pb, en las filas 2-7 (líneas celulares
231, HeLa, A549, HOP-62 y 293T),
la amplificación del fragmento que
corresponde a la banda de
mycoplasma de aproximadamente
300 pares de bases.
1 2 3 4 5 6 7 8
MW 231 HeLa HeLa A549 A549 HOP 293-T 100pb 62
32
5.5 Transfección del vector pMFR – IRES en células 293T
5.5.1 Análisis de la fluorescencia de las proteínas Cherry y GFP
Una vez obtenidas todas las construcciones deseadas, se transfectó en líneas celulares
293T mediante la técnica del fosfato cálcico, para comprobar que fueron clonadas
correctamente y que los vectores expresaban adecuadamente GFP y Cherry. Las
construcciones transfectadas fueron: pMFR-Cx43, pMFR-19kD, pMFR-cMyc, pMFR-
Cx26 y pMFR-EMCV como control. Los resultados se analizaron 48 horas después de la
transfección en el microscopio de fluorescencia. Las fotografías muestran los niveles de
expresión proteica GFP/Cherry. Todos los tratamientos son positivos para la expresión de
Cherry,; sin embargo se puede observar un leve background debido a la fluorescencia
emitida por GFP, ya que GFP se expresa mucho en la condición de pMFR-EMCV, que era
lo que se esperaba ya que es nuestro control positivo. La figura 6 muestra los resultados
obtenidos. Las imágenes fueron adquiridas con el microscopio de fluorescencia Olympus
FSX-BSW 100.
Figura 6. Niveles de expresión proteica de Cherry/GFP en células 293 de los genes
transfectadas con pMFR-IRES
GFP/mCherry mCherry GFP
a)
b)
c)
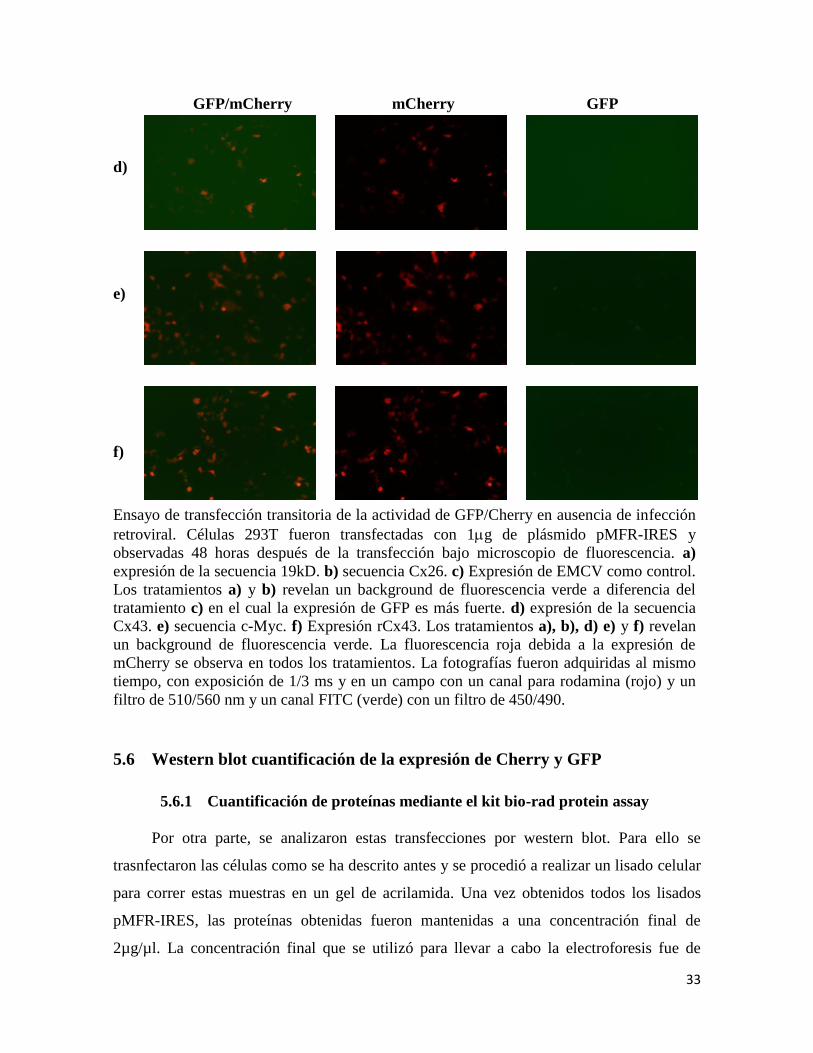
33
GFP/mCherry mCherry GFP
d)
e)
f)
Ensayo de transfección transitoria de la actividad de GFP/Cherry en ausencia de infección
retroviral. Células 293T fueron transfectadas con 1g de plásmido pMFR-IRES y
observadas 48 horas después de la transfección bajo microscopio de fluorescencia. a)
expresión de la secuencia 19kD. b) secuencia Cx26. c) Expresión de EMCV como control.
Los tratamientos a) y b) revelan un background de fluorescencia verde a diferencia del
tratamiento c) en el cual la expresión de GFP es más fuerte. d) expresión de la secuencia
Cx43. e) secuencia c-Myc. f) Expresión rCx43. Los tratamientos a), b), d) e) y f) revelan
un background de fluorescencia verde. La fluorescencia roja debida a la expresión de
mCherry se observa en todos los tratamientos. La fotografías fueron adquiridas al mismo
tiempo, con exposición de 1/3 ms y en un campo con un canal para rodamina (rojo) y un
filtro de 510/560 nm y un canal FITC (verde) con un filtro de 450/490.
5.6 Western blot cuantificación de la expresión de Cherry y GFP
5.6.1 Cuantificación de proteínas mediante el kit bio-rad protein assay
Por otra parte, se analizaron estas transfecciones por western blot. Para ello se
trasnfectaron las células como se ha descrito antes y se procedió a realizar un lisado celular
para correr estas muestras en un gel de acrilamida. Una vez obtenidos todos los lisados
pMFR-IRES, las proteínas obtenidas fueron mantenidas a una concentración final de
2µg/µl. La concentración final que se utilizó para llevar a cabo la electroforesis fue de

34
20µg/µl de cada lisado proteico. Para el cálculo de las concentraciones se utilizó una
regresión, en la cual la curva y la ecuación de la curva nos permitió calcular la cantidad
exacta de buffer de lisis que se debía añadir a los lisados para obtener la concentración
deseada, figura 7.
Figura 7. Recta de regresión para calcular la concentración de proteínas.
En el eje de las X se indica la concentración de la proteína y en el eje Y la absorbancia a
690nm. La recta nos permite identificar una relación entre las proteína obtenidas en cada
línea celular, ya que la cantidad utilizada para la medición de cada una fue 1µl. La
ecuación de la recta nos permitió calcular la concentración de proteína en mg/ml.
Una vez obtenidas las concentraciones adecuadas de los lisados proteicos de las
células que expresaban mCherry-His y EGFP-His bajo la activación de los IRES
conrrespondientes de las proteínas 19kD, Cx26, rCx43, C-myc, Cx43 y EMCV, se cargó 20
g/l de proteína para realizar una electroforesis SDS-PAGE. Una vez terminada la
electroforesis y realizada la transferencia, la membrana fue sometida al anticuerpo -His.
Las células que contenían las construcciones de 19kD, c-Myc y rCx43 casi no expresaban
GFP, es decir, casi no mostraron actividad IRES, mientras que pMFR-EMCV expresó una
mayor cantidad de GFP-His, lo que quiere decir que el IRES está activo figura 8. En
contraste, la expresión de mCherry fue muy similar entre todos los vectores, indicando que
el primer cistrón se expresa de manera similar en todos los constructos.
Recta de regresión
y = 0,1425x + 0,5352
R2 = 0,8464
0
0,2
0,4
0,6
0,8
1
0 0,2 0,4 0,6 0,8 1 1,2 1,4 1,6 1,8 2[mg/ml]
Ab
s (
690)

35
Figura 8. Western Blot del vector pMFR y las secuencias de 5 genes IRES.
La figura indica la expresión de las proteínas Cherry y GFP por análisis de western blot.
Cantidades equivalentes de lisado de proteína fueron cargados. En el carril 6 se muestra el
vector control pMFR-EMCV en el cual se observa claramente la fuerte expresión de GFP.
En los carriles 1, 4 y 5 se distinguen las bandas de 29kD y 27kD correspondientes a Cherry
y GFP, respectivamente.
Por otro lado, también se transfectó los vectores PMFR que contenían las secuencias
de los genes IRES en dos líneas celulares HeLa y MDA-MB-231. Los tratamientos
aplicados en las células fueron: como control el vector pMFR; se realizó dos co-
transfecciones 1) pMFR-cMyc con el plásmido control pLPCX y 2) pMFR-cMyc con un
vector que expresa el ITAF YB-1. pMFR-19kD y el disolvente DMSO como control;
pMFR-19kD con la droga CGP (un inhibidor de Mnk1 kinasa que previene la fosforilación
de eIF4E) y el vector pMFR-EMCV como control.
En este caso, con todos los vectores transfectados y las co-transfecciones se realizó
el análisis de western blot. Las membranas fueron sometidas, igualmente, al anticuerpo -
His. Sorprendentemente, ningún constructo mostró expresión de eGFP y solamente se pudo
detectar la proteína Cherry (figura 9).
1 2 3 4 5 6

36
Figura 9. Western Blot para el análisis de co-transfección en células HeLa.
Efecto de la co-transfección del vector pMFR-IRES. Corrida electroforética SDS-PAGE y
analizada con el anticuerpo -His.
Debido a que mediante western blot no se pudo detectar la proteína GFP en células
HeLa, se realizó mediciones en el lector de fluorescencia Thermo Scientific Appliskan para
determinar la relación de expresión proteica de GFP/mCherry. La figura 10 muestra que
GFP aumenta en pMFR-EMCV (control positivo) tanto en la línea celular HeLa como en
231, pero en las otras construcciones no se observa una expresión transitoria.
Figura 10. Ratio de expresión de las proteínas Cherry-GFP en dos líneas celulares
HeLa y 231.
En la figura se muestra la proporción de expresión de las proteínas GFP/Cherry. La
expresión del vector pMFR se toma como inicial, a partir de la cual se hace un ratio para
cada tratamiento. Se puede observar que la proporción de la expresión en el vector que
posee el gen EMCV es mayor en las dos líneas celulares; así mismo la expresión de c-Myc
se ve incrementada únicamente en la línea celular 231 en el tratamiento con el vector viral.
Sin embargo, al realizar la co-transfección cMyc-Yb1 se observa una mayor expresión en
las células 231. Igualmente sucede con la expresión del gen 19kD, en el tratamiento con
DMSO hay una mayor expresión en las células HeLa, en cambio con el tratamiento de
CGP se incrementa la proporción GFP/Cherry en las células 231.
-202468
10121416
HeLa
231
1 2 3 4 5 6

37
Podemos ver que en la línea celular MDA-MB-231 hay una mayor expresión de los
vectores (cMyc+pPLCX, pMFR-EMCV y pMFR-19kd (CGP)) y no en las células HeLa
excepto por pMFR-EMCV cuya expresión es fuerte en los dos casos. Los resultados fueron
obtenidos con la relación de GFP IRES-dependiente dividido por la expresión de Cherry
cap-dependiente.
5.7 Producción viral en células Phoenix
A partir del vector pMFR se realizó la extracción del fragmento Cherry-MCS-GFP el
cual contiene el MCS de diferentes secuencias IRES, EMCV, 19kD (secuencia interna en la
región codificante del gen connexin 43) y c-Myc mediante la digestión con las enzimas de
restricción BglII y NotI. El vector pLPCX fue digerido con las mismas enzimas como se ha
descrito. Por último los fragmentos fueron ligados usando la ligasa T4 (FERMENTAS®).
Para la transformación, con los productos ligados, se utilizó bacterias competentes de E.
coli de la cepa DH5; las mismas que fueron plaqueadas en agar LB con ampicilina como
antibiótico de selección. Finalmente, se obtuvieron colonias positivas, se realizó la
extracción de ADN plasmídico el mismo que fue utilizado para infectar células Phoenix
(derivadas a partir de las células HEK-293).
5.8 Establecimiento de líneas celulares productoras de virus
Obtenidas las líneas celulares Phoenix transfectadas con los vectores pLPCX: pLPCX-
Cherry-EMCV-GFP, pLPCX-Cherry-c-Myc-GFP, pLPCX-Cherry-19kD-GFP, se recogió
los virus y se los filtró. Se adicionó polibreno al medio y se infectó 4 líneas celulares: 1)
HeLa (cáncer de cérvix), 2) Hop-62 (cáncer de pulmón), 3) MDA-MB-231 (cáncer de
mama) y 4) A549 (línea celular de cáncer de pulmón) para la producción de virus de
manera continua. El proceso de infección se repitió a las 24 y 48 horas de producción viral.
Una vez obtenidas las células infectadas se realizó el cambio de medio de cultivo y se
adicionó puromicina para seleccionar clones positivos. Se mantuvo las células en selección
con antibiótico durante una semana cambiando el medio y añadiendo puromicina cada 48
horas. Posteriormente, se procedió a cambiar las células a medio de cultivo normal, sin
antibiótico) para que se recuperen al tratamiento y así aislar las colonias positivas. Sin
embargo, el tratamiento con el antibiótico afecto la morfología de las células, y la línea

38
celular Hop-62 no sobrevivió ni al tratamiento con virus ni a la selección con puromicina,
confirmado así que la puromicina elimina eficazmente las células no infectadas.
Se logró obtener clones en las las líneas celulares A549, HeLa, y MDA-MB-231 a
partir de la construcción que contiene el IRES de c-Myc figura 11. Además se obtuvieron
clones positivos de las secuencias de los genes EMCV y 19kD en la línea celular 231,
además se obtuvo el vector pMFR como control figura 12.
Figura 11. Clones del gen c-Myc y la relación entre la expresión de las proteínas
Cherry-GFP en líneas celulares HeLa, A549 y 231
Detección de clones positivos de las secuencias del gen c-Myc en 3 líneas celulares
mediante la expresión de eGFP-mCherry. a) se muestra una colonia positiva para la
expresión de mcherry y GFP. b) se muestra una colonia positiva para mCherry y muy
pocas células positivas para GFP. c) En la línea celular 231 únicamente se distinguen
células positivas para mCherry. Todas la fotografías fueron tomadas en el mismo campo
con un canal para rodamina (rojo) con un filtro de 510/560 nm y un canal FITC (verde)
con un filtro de 450/490. Todas las fotografías fueron tomadas con el mismo tiempo de
exposición 1/3 260 ms.
Contraste de fases GFP mCherry
a) HeLa – c-Myc
b) A549 – c-Myc
c) 231 – c-Myc

39
Figura 12. Clones estables en la línea celular 231 de EMCV, 19kD y del vector control
pMFR.
Detección de clones positivos en la línea celular 231 mediante la evaluación de la
expresión de eGFP-mCherry. a) Se muestran colonias positivas para la expresión de
mcherry y GFP control. b) colonias positivas para la expresión de EMCV IRES c) Del
fragmento 19kD únicamente se distinguen colonias positivas para la expresión de mcherry.
5.9 Cultivo de líneas celulares bajo condiciones de estrés
Se utilizaron dos líneas celulares estables, obtenidas a través de la infección viral,
SW620 (línea celular humana de cáncer de colon metastático) y HaCat (línea celular huma
de keratinocitos inmortalizados), para identificar como ciertas sustancias afectan la
traducción de GFP iniciada a partir de cMyc-IRES. En los tratamientos llevados a cabo, se
eliminó el suero fetal bovino del medio de cultivo (FCS 0%). También, se inhibió la
proteína reguladora mTOR mediante el uso de PP242 (muy similar a rapamicina), además
se sometió a las células a estrés por la ausencia de oxígeno usando cloruro de cobalto
(como se ha visto en ciertos tumores). Al someter a las células, en cultivo, a diferentes
condiciones de estrés, se provocó una sobre regulación de la actividad traduccional de Myc-
IRES. La condición de 0% FCS provoca, en las células, estrés debido a la ausencia de la
señal mitogénica del crecimiento; CPG tiene un menor efecto. Se observa que los
a) 231 -
pMFR
b) 231 -
EMCV
GFP/Cherry Cherry GFP Contraste de
fases
c) 231 –
19kD

40
tratamientos aplicados en las líneas celulares incrementan el inicio de la traducción IRES-
dependiente, como se indica en la figura 13.
Figura 13. Condiciones de estrés en células HaCat.
Comparación de la expresión de GFP y mCherry en células HaCat sometidas a 5
tratamientos de estrés celular. Las condiciones 0 y 10% suero fetal bovino (FBS), PP242
(2.5 M), CoCl2 (250 M) y CGP (40M). Las células HaCat utilizadas eran células
estables que expresan el vector pLPCX-Cherry-c-Myc-GFP. A las 24 horas de la
transfección, se evalúo la fluorescencia emitida. Todos los datos fueron normalizados de
acuerdo a la eficiencia de transfección efectuando la proporción GFP/mcherry y
presentada como medidas promedio de los 5 tratamientos.
Se puede observar que bajo las condiciones de estrés hay un incremento en la
expresión del cMyc-IRES (tanto en el tratamiento con 0% FBS como en el del PP242); este
incremento en la expresión puede deberse a que los elementos IRES celulares están
presentes en mARNs que codifican proteínas bajo ciertas condiciones de estrés.
5.10 Evaluación de la expresión del plásmido pLPC-SRp20
Se trabajo en células MDA-MB-468, estables, con los vectores pMFR-EMCV y
pMFR-cMyc. Los cultivos fueron inducidos con diferentes tipos de estrés: 1) 10% FBS, 2)
0% FBS, 3) CoCl2 250 M 3) CoCl2 250 M 4) PP242 2.5 M y 5) rapamicina.
Además se realizó la transfección de la construcción pLPC-SRp20 que es un
conocido ITAF, utilizando el reactivo JetPei como se ha descrito anteriormente.
Transcurridas 24 horas de la transfección se trató las células con 0 y 10% de FBS; 24 horas
después del tratamiento se realizó la medición de la fluorescencia de GFP con respecto a
0
1
2
3
4
5
10% FBS0% FBS PP242 CoCl2 CGP
Fold
Ch
ange
EG
FP
/ m
Ch
erry
41
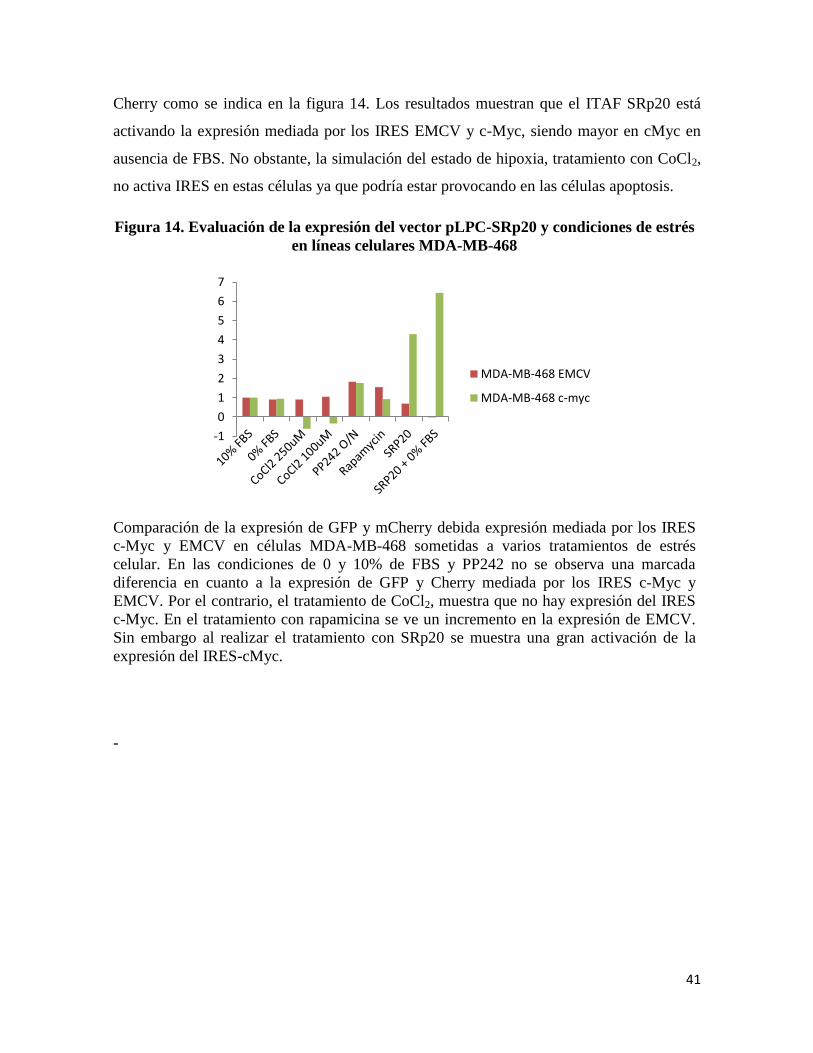
Cherry como se indica en la figura 14. Los resultados muestran que el ITAF SRp20 está
activando la expresión mediada por los IRES EMCV y c-Myc, siendo mayor en cMyc en
ausencia de FBS. No obstante, la simulación del estado de hipoxia, tratamiento con CoCl2,
no activa IRES en estas células ya que podría estar provocando en las células apoptosis.
Figura 14. Evaluación de la expresión del vector pLPC-SRp20 y condiciones de estrés
en líneas celulares MDA-MB-468
Comparación de la expresión de GFP y mCherry debida expresión mediada por los IRES
c-Myc y EMCV en células MDA-MB-468 sometidas a varios tratamientos de estrés
celular. En las condiciones de 0 y 10% de FBS y PP242 no se observa una marcada
diferencia en cuanto a la expresión de GFP y Cherry mediada por los IRES c-Myc y
EMCV. Por el contrario, el tratamiento de CoCl2, muestra que no hay expresión del IRES
c-Myc. En el tratamiento con rapamicina se ve un incremento en la expresión de EMCV.
Sin embargo al realizar el tratamiento con SRp20 se muestra una gran activación de la
expresión del IRES-cMyc.
-
-1
0
1
2
3
4
5
6
7
MDA-MB-468 EMCV
MDA-MB-468 c-myc

42
6 DISCUSIÓN
El control de la traducción de proteínas juega un papel esencial en el crecimiento
celular y es un proceso altamente regulado en la progresión tumoral. En particular, se ha
visto que la traducción mediada por secuencias IRES, aunque aún sea considerada como
una vía “alternativa”, se utiliza con relativa frecuencia tanto en condiciones fisiológicas
normales como en condiciones patológicas o de estrés, y el mecanismo de escaneo cap-
dependiente está comprometido (Komar & Hatzoglou; 2011). Por lo tanto, se piensa que la
vía de traducción cap-independiente podría ser estar contribuyendo a la transformación
tumoral.
Con el fin de evaluar la activación de la traducción mediada por IRES, en este trabajo
se ha puesto a punto el clonaje de diferentes secuencias IRES ya conocidas en un vector
bicistrónico fluorescente denominado pMFR, previamente generado en el laboratorio,
cuyos insertos dan como resultado la expresión constante de Cherry y la expresión
alternativa de GFP como dos especies de proteínas distintas. Para analizar la posibilidad de
que varias proteínas presenten secuencias IRES, se clonaron las secuencias Connexin 26,
Connexin 43, EMCV (el cual no es traducido, pero funciona como una secuencia que
permite el inicio de la traducción downstream del ARN, en este caso hacia GFP), la
secuencia IRES ya conocida de c-Myc y 19kD (un IRES putativo dentro de la región
codificante de connexin 43, ya que las secuencias putativas pueden actuar como promotores
crípticos, sitios de splicing que modulan el fraccionamiento por RNAasa) (Van Eden, et al.,
2004), en un vector bicistrónico, de modo que si las secuencias IRES se activan, se
impulsaría la expresión de GFP en este vector.
Así, en la primera parte de los resultados podemos concluir que hemos clonado con
éxito las todas las secuencias IRES, ya que la posterior comprobación de los clonajes, así lo
demuestra. En una segunda fase, se valoró la actividad mediada por las secuencias IRES y
se evalúo la activación de las mismas mediante transfecciones (figura 9) y co-
transfecciones (figura 10), de las construcciones generadas, en líneas celulares.
El posterior análisis indica que el experimento está bien ejecutado, ya que el control
positivo, el IRES viral EMCV está activando la expresión de GFP. Éste resultado se ha

43
observado con los tres métodos utilizados: microscopía de fluorescencia (figura 6),
cuantificación de la fluorescencia con el fluorímetro (figura 10) y también se confirmó
mediante el análisis de western blot, en el cual se observó una banda correspondiente a la
proteína quimérica GFP-His (figura 10). Los otros IRES no son muy efectivos o en realidad
no son secuencias IRES ya que no se observó una expresión significativa de GFP con
respecto a Cherry, esto fue evaluado en condiciones normales de cultivo celular con 10%
FBS.
Otro mecanismo de evaluación fue a través de infecciones virales, se trabajó con líneas
celulares estables con capacidad de traducir la fluorescencia roja de mCherry a partir de la
traducción cap-dependiente y fluorescencia verde correspondiente a la proteína EGFP
mediada por secuencias IRES (figuras 11 -12). Una vez obtenidas las líneas celulares
estables, que contenían el vector bicistrónico, se comprobó si las secuencias IRES
funcionaba y se indujo la expresión de GFP (figuras 13-14). Se vio que, para estudiar la
activación de IRES es mejor tener líneas estables que transfecciones transitorias, ya que en
estas, a veces se observan muy pocas células transfectadas y no podemos ver el efecto. Por
el contrario, en líneas estables la mayoría ó todas las células, al menos deberían estar
expresando la proteína cherry.
Posteriormente, para determinar si el IRES de c-Myc es activo en nuestro sistema,
planteamos provocar condiciones de estrés en las líneas celulares para activar esta
secuencia. Se observó que con falta de nutrientes se incrementa la fluorescencia
cuantificada con fluorímetro en células no tumorales HaCat (figura 14), pero no sucede lo
mismo en células MDA-MB-468 (carcinoma mamario) (figura 15). Es conocido que con
falta de nutrientes, se inactiva el mecanismo de traducción cap-dependiente y se activa el
sistema mediado por IRES en el 5’UTR de los mRNAs (Fitzgerald & Semler, 2009).
El hecho de que en células tumorales no se active el IRES de c-Myc puede deberse a
que estas células sobre expresan Myc para permitir la proliferación normal de células. En
cuanto al tratamiento con PP242, inhibidor de mTOR, se inhibió la vía cap-dependiente y
consecuentemente se observó vía cap-independiente, mediada por el IRES de c-Myc y la
expresión de GFP en células HaCat (figura 14) y también en células MDA-MB-468 (figura
15). El funcionamiento de mTOR, es un mecanismo crucial para el crecimiento tumoral,

44
por lo que se ha vuelto un blanco popular para los tratamientos terapéuticos (Hoang, et al.,
2012). Con estos resultados, podríamos pensar que se ha identificado una nueva vía de
activación de la sobre regulación del oncogén Myc.
Así mismo, la inducción de hipoxia afectó sólo a las células no tumorales HaCat y se
vio que se activa el IRES de c-Myc, pero no se activó en células MDA-MB-468
posiblemente porque podría estar induciendo apoptosis y no estar favoreciendo el efecto
deseado. De esta manera se puede relacionar al estrés producido con un incremento
endógeno de la proteína Myc (la cual contiene la secuencia myc-IRES en su región
5’UTR). A pesar de que las células responden al estrés fisiológico (hipoxia) y patológico
mediante la fosforilación de la subunidad eIF2 la cual da como resultado la atenuación
global de la síntesis de proteínas (Joachims, et al., 1999).
Además, para analizar el funcionamiento de las secuencias de los genes tranfectados
junto con el vector pMFR-c-Myc se añadió el plásmido YB-1 para determinar si el factor
YB-1 ITAF podría afectar la traducción de GFP desde el sitio del IRES-cMyc. De acuerdo
a los datos obtenidos parece que estaría reprimiendo la traducción en las dos líneas
celulares, HeLa y 231 (figura 11), lo cual es inconsistente con datos reportados
previamente, en los cuales se indica que YB1 se une y regula promotores que no pertenecen
sólo a la caja-Y, entre ellos el receptor del factor de crecimiento epidermal (Stratford, et al.,
2007), aunque YB-1 puede regular positiva o negativemente la transcripción y la
traducción; además activa genes asociados con la porliferacion celular y el cáncer, y
reprime genes asociados con la muerte celular (Wang, & Hirschberg, 2011).
Finalmente, se observó que la expresión de un ITAF (SRp20) en líneas celulares
estables transfectadas con los vectores bicistrónicos de los IRES EMCV y Myc provocan
una activación de la expresión de GFP (figura 15). Probablemente este factor ha sido
implicado en funciones alternativas de expresión génica, posiblemente proporcionando un
vínculo entre el procesamiento y exportación del mARN y la traducción. Sin embargo,
SRp20 ha sido detectado en fracciones de la subunidad ribosomal, pero la sobre expresión
de SRp20 no tiene un efecto estimulatorio en la traducción cap-dependiente (Bedard, et al.,
2007).

45
En definitiva, el sistema combinado de expresión de la proteína GFP y mCherry podría
ser una alternativa interesante en varias circunstancias: 1) una herramienta para la
elaboración de líneas celulares estables (incluyendo el uso para la selección de células
mediante citometría de flujo), 2) creación de ratones transgénicos que expresen genes
reporteros fluorescentes, 3) como un instrumento para la revisión de librerías de shARNi o
la investigación de químicos usando lecturas evaluaciones fluorescentes de intensidad, ya
sea manualmente o mediante robots.

46
7 CONCLUSIONES Y FUTURO
1. Algunas secuencias IRES conocidas (EMCV viral, c-Myc eucariota, Cx26 y Cx43)
así como un IRES putativo no probado (denominado 19kD) fueron exitosamente
amplificados por PCR. Por lo tanto, el enfoque de los estudios futuros en el campo
deben centrarse en el significado fisiológico celular de la traducción mediada por
IRES; ya que esta vía puede permitir el entendimiento de varias hipótesis sobre los
mecanismos de traducción cap-independiente que permitan descubrir nuevos
mARNs candidatos, cuyo mecanismo de traducción cap-independiente mediada por
IRES sea facilitada/impulsado por las secuencias reguladoras ITAFs.
2. Las secuencias IRES fueron clonadas con éxito dentro del sitio de clonaje múltiple
entre Cherry y EGFP para permitir el análisis de la expresión de cherry cap-
dependiente y la expresión de GFP mediada por IRES.
3. Estudios de expresión transitoria confirmaron, como se esperaba, que el IRES viral
EMCV presenta la más fuerte activación traduccional de EGFP. El c-Myc-IRES
posee una actividad más baja, lo cual depende de la línea celular que haya sido
utilizada y, sorprendentemente, parece ser reprimido cuando co-expresa el factor
ITAF YB-1. El IRES putativo 19kDa, aparentemente no presenta actividad o esta es
limitada.
4. La transfección de células Phoenix con este vector permitió la producción de
partículas virales y la infección de una variedad de líneas celulares como HaCaT,
HeLa, A549 y MDA-MB-231. Usando la resistencia a la puromicina, presente en el
vector, estas líneas celulares fueron seleccionadas adecuadamente por una expresión
de fluorescencia estable y actualmente están siendo expandidas para su uso en una
variedad de proyectos.
5. Las líneas celulares han sido, hasta ahora, utilizadas con éxito para investigar la
actividad IRES en células sujetas ya sea a través de estrés o por la manipulación de
las vías de señalización por fármacos. Usando células HaCaT, una fuerte activación
de la actividad del IRES de Myc fue descubierta ya sea a través de la tensión o la
manipulación de las vías de señalización por fármacos, la privación de suero y la
simulación de hipoxia con cloruro de cobalto. Adicionalmente, la represión de la vía

47
de mTOR por PP242, permitió una elevada activación del IRES de cMyc, mientras
que Mnk1, inhibidor CGP, tuvo poco efecto.
6. Finalmente, el conocido ITAF SRp20 fue clonado en un vector de expresión y
transfectado en líneas celulares que ya expresaban establemente cherry-EGFP
conteniendo varios elementos IRES. Una fuerte activación de la actividad del IRES
de myc se observó en la línea celular de cáncer mamario MDA-MB-468
determinada por SRp20. Esta es la primera vez que se observa una asociación entre
SRp20 y la activación de Myc. El análisis de muchos otros ITAFs mediante sobre
expresión de ARNi es actualmente en marcha.
7. En el futuro, se obtendrá una línea celular estable con IRES específicos (por
ejemplo HCV IRES); el screening de ARN (una librería de siARN cubriendo la
mayor parte del genoma. Se podrán realizar transfecciones robóticas en 2 días, y
lecturas automatizada de la proporción de GFP para que bioinformáticos puedan
identificar todos los genes que regulan específicamente la actividad IRES (pero no
la traducción normal de cherry), así mismo se podrá realizar el screening químico de
bibliotecas mediante robots.

48
8 BIBLIOGRAFIA
Ausubel, F., Brent, R., Kingston, R.E., Moore, D.D., Seidman, J.G., Smith, J.A.,
Struhl, K. 1995. Short Protocols in Molecular Biology. John Wiley & Sons, Inc., USA.
Baird, S.D., Turcotte, M., Korneluk, R.G., Holcik, M. 2006. Searching for IRES. RNA;
12(10): 1755-1785.
Bedard, K.M., Daijogo, S., Semler, B.L. 2007. A nucleo-cytoplasmic SR protein
functions in viral IRES-mediated translation initiation. EMBO J; 26(2): 459 - 67.
Berg, J.J., Tymoczko, J.L., Stryer, L. 2002. Biochemistry. 5th
edition. New York: W H
Freeman.
Bert, A.G., Grepin, R., Vadas, M.A., Goodall, G.J. 2006. Assessing IRES activity in
the HIF-1alpha and other cellular 5′ UTRs. RNA; 12(6): 1074-1083.
Bonnal, S., Schaeffer, C., Creancier, L., Clamens, S., Moine, H., Prats, A.C., Vagner,
S. 2003. A single internal ribosome entry site containing a G quartet RNA structure drives
fibroblast growth factor 2 gene expression at four alternative translation initiation codons.
J. Biol. Chem; 278(41): 39330-39336.
Cobbold, L.C., Spriggs, K.A., Haines, S.J., Dobbyn, H.C., Hayes, C., de Moor, C.H.,
Lilley, K.S., Bushell, M., Willis, A.E. 2008. Identification of internal ribosome entry
segment (IRES)-trans-acting factors for the Myc family of IRESs. Mol. Cell Biol; 28(1):
40-49.
Coffin, J.M., Hughes, S.H., Varmus, H.E. 1997. Retroviruses. Cold Spring Harbor
(NY): Cold Spring Harbor Laboratory Press.
Costa-Mattioli, M., Svitkin, Y., Sonenberg, N. 2004. La autoantigen is necessary for
optimal function of the poliovirus and hepatitis C virus internal ribosome entry site in vivo
and in vitro. Mol. Cell Biol; 24(15): 6861-6870.
Evans, J.R., Mitchell, S.A., Spriggs, K.A., Ostrowski, J., Bomsztyk, K., Ostarek, D.,
Willis, A.E. 2003. Members of the poly (rC) binding protein family stimulate the activity of
the c-myc internal ribosome entry segment in vitro and in vivo. Oncogene; 22(39): 8012-
8020.
Fitzgerald, K.D. & Semler, B.L. 2009. Bridging IRES elements in mRNAs to the
eukaryotic translationapparatus. Biochim Biophys Acta; 1789(9-10): 518–528.
Graber, T.E. &Holcik, M. 2007. Cap-independent regulation of gene expression in
apoptosis. Mol. Biosyst; 3(12): 825-834.
Graham, F.L., Van der Eb A.J., 1973. A new technique for the assay of infectivity of
human adenovirus 5 DNA. Virology; 52(2): 456-467.

49
Hitoshi, Y., Lorens, J., Kitada, S.I., Fisher, J., La-Barge, M., Ring, H., Francke, U.,
Reed, J.C., Kinoshita, S., Nolan, G.P. 1998. Toso, a cell surface, specific regulator of Fas-
induced apoptosis in T cells. Immunity; 8(4): 461-471.
Hoang, B., Benavides, A., Shi, Y., Yang, Y., Frost, P., Gera, J., Lichtenstein, A. 2012.
J Biol Chem; 287(26): 21796 – 21805.
Holcik, M., Gordon, B.W., Korneluk, R.G. 2003. The internal ribosome entry site-
mediated translation ofantiapoptotic protein XIAP is modulated by the heterogeneous
nuclear ribonucleoproteins C1 andC2. Mol. Cell Biol; 23(1): 280-288.
Horbinski, C.M., Stachowiak, K., Higgins, D., Finnegan, S.G. 2001 Polyethylenimine
mediated transfection of cultured postmitotic neurons from rat sympathic ganglia and adult
human retina. BMC Neuroscience; 2: 2.
Jackson, R.J., Hellen, C.U., Pestova, T.V. 2010. The mechanism of eukaryotic
translation initiation and principles of its regulation. Nat Rev Mol Cell Biol; 11(2): 113-127.
Jackson, R.J. 2005. Alternative mechanisms of initiating translation of mammalian
mRNAs. Biochem. Soc.Trans; 33(6): 1231-1241.
Jang, G.M., Leong, L.E., Hoang, L.T., Wang, P.H., Gutman, G.A., Semler, B.L, 2004.
Structurally distinct elements mediate internal ribosome entry within the 5′-noncoding
region of a voltage-gated potassium cannel mRNA. J. Biol. Chem; 279(46): 47419-47430.
Jang, S.K., Kräusslich, H.G., Nicklin, M.J., Duke, G.M., Palmenberg, A.C., Wimmer,
E. 1988. A segment of the 5’ nontranslated region of encephalomyocarditis virus RNA
directs internal entry of ribosomes during in vitro translation. J. Virol; 62(8): 2636-2643.
Jimenez, J., Jang, G.M., Semler, B.L., Waterman, M.L. 2005. An internal ribosome
entry site mediates translation of lymphoid enhancer factor-1. RNA; 11(9): 1385-1399.
Joachims, M., Van Breugel, P.C., Lloyd, R.E. 1999. Cleavage of poly(A)-binding
protein by enterovirus proteases concurrent with inhibition of translation in vitro. J. Virol;
73(1): 718-727.
Kozak, M. 2005. A second look at cellular mRNA sequences said to function as
internal ribosome entry sites. Nucleic Acids Res; 33(20): 6593-6602.
Komar, A.A & Hatzoglou, M. 2011. Cellular IRES-mediated translation The war of
ITAFs in pathophysiological states. Cell Cycle; 10(2): 229-240.
Langdon, S.P. 2010. Cancer Cell Culture. Methods and Protocols. Humana Press.
Totowa, New Jersey.
Le Quesne, J.P., Stoneley, M., Fraser, G.A., Willis, A.E. 2001. Derivation of a
structural model for the c-myc IRES. J. Mol. Biol; 310(1): 111-126.
Lodish, H., Berk, A., Zipursky, S.L., Matsudaira, P., Baltimore, D. Darnell, J. 2000.
Molecular Cell Biology, 4th
edition. New York: W. H. Freeman.

50
Manak, M.M. 1993. Sample Preparation. En: DNA probes (G.H. Keller y M.M.
Manak, eds) pp. 27 - 68. Stockton Press, New York, USA.
Martin, P., Albagli, O., Poggi, M.C., Boulukos, K.E., Pognonec, P. 2006. Development
of a new bicistronic retroviral vector with strong IRES activity. BMC Biotechnol: 12; 6:4.
Martínez-Salas, E., Ramos, R., Lafuente, E., López, de Q.S. 2001. Functional
interactions in internal translation initiation directed by viral and cellular IRES elements. J.
Gen. Virol; 82(5): 973-984.
Mitchell, S.A., Spriggs, K.A., Bushell, M., Evans, J.R., Stoneley, M., Le Quesne, J.P.,
Spriggs, R.V., Willis, A.E. 2005. Identification of a motif that mediates polypyrimidine
tract-binding protein-dependent internal ribosome entry. Genes Dev; 19(13): 1556-1571.
Mitchell, S.A., Spriggs, K.A., Coldwell, M.J., Jackson, R.J., Willis, A.E. 2003. The
Apaf-1 internal ribosome entry segment attains the correct structural conformation for
function via interactions with PTB and unr. Mol. Cell; 11(3): 757-771.
Nanbru, C., Lafon, I., Audigier, S., Gensac, M.C., Vagner, S., Huez, G., Prats, A.C.
1997. Alternative translation of the proto-oncogene c-myc by an internal ribosome entry
site. J. Biol. Chem; 272(51): 32061-32066.
Newton, C.R., Graham, A. 1994. PCR. Bios Scientific Publishers, Oxford, UK.
Paek, K.Y., Kim, C.S., Park, S.M., Kim, J.H., Jang, S.K. 2008. RNA-binding protein
hnRNP D modulates internal ribosome entry site-dependent translation of hepatitis C virus
RNA. J. Virol; 82(24): 12082-12093.
Pear, W.S., Nolan, G.P., Scott, M.L., Baltimore,D. 1993. Production of high-titer
helper-free retroviruses by transient transfection. Proc. Natl.Acad.Sci. USA; 90(18): 8392-
8396.
Pelengaris S, Khan, M., Evan, G. 2002. c-MYC: more than just a matter of life and
death. Nat. Rev. Cancer; 2(10): 764-776.
Pelletier, J., Sonenberg, N. 1988. Internal initiation of translation of eukaryotic mRNA
directed by a sequence derived from poliovirus RNA. Nature; 334(6180): 320-325.
Pelletier, J., Kaplan, G., Racaniello, V.R., Sonenberg, N. 1988. Cap-independent
translation of poliovirus mRNA is conferred by sequence elements within the 5′ noncoding
region. Mol. Cell Biol; 8(3): 1103-1112.
Rivera-Tapia, J.A., Cedillo, R.L., Gil, J.C. 2002. Some biological featuresof
Mollicutes. Rev Latinoam Microbiol; 44(2); 53-57.
Sambrook, J., Fritsch, E.F., Maniatis, T. 1989. Molecular Cloning: A Laboratory
Manual. Cold Spring Harbor Laboratory Press, New York, USA.

51
Schepens, B., Tinton, S.A., Bruynooghe, Y., Parthoens, E., Haegman, M., Beyaert, R.,
Cornelis, S. 2007. A role forhnRNP C1/C2 and Unr in internal initiation of translation
during mitosis. EMBO J; 26(1); 158-169.
Sonenberg, N., Hinnebusch, A.G. 2009. Regulation of translation initiation in
eukaryotes: mechanisms and biological targets. Cell; 136(4):731-745.
Strachan, T., Read, A.P., 1999. Human Molecular Genetics, 2nd
edition. New York:
Wiley-Liss. Garland Science.
Stoneley, M., Subkhankulova, T., Le Quesne, J.P., Coldwell, M.J., Jopling, C.L.,
Belsham, G.J., Willis, A.E. 2000. Analysis of the c-myc IRES; a potential role for cell-type
specific trans-acting factors and the nuclear compartment. Nucleic Acids Res; 28(3); 687-
694.
Stratford, A.L., Habibi, G., Astanehe, A., Jiang, H., Hu, K., Park, E., Shadeo, A., Buys,
T.P.H., Lam, W., Pugh, T., Marra, M., Nielsen, T.O., Klinge, U., Mertens, P.R., Aparicio,
S., Dunn, S.E. 2007. Breast Cancer Res; 9(5): R61.1.
Van Eden, M.E., Byrd, M.P., Sherrill, K.W., Lloyd, R.E. 2004. Demonstrating internal
ribosome entry sites in eukaryotic mRNAs using stringent RNA test procedures. RNA;
10(4): 720-730.
Walter, B.L., Parsley, T.B., Ehrenfeld, E., Semler, B.L. 2002. Distinct poly(rC)
binding protein KH domain determinants for poliovirus translation initiation and viral RNA
replication. J. Virol; 76(23): 12008-12022.
Wang, S., Hirschberg, R. 2011. Y-Box Protein-1 is a Transcriptional Regulator of
BMP7. Journal of Cellular Biochemistry; 112(4):1130-1137.


















