
Una especie nueva de Grundulus (Teleostei: Ostariophysi ... 51-72.pdf · 52 Una especie nueva de...
22
Memoria de la Fundación La Salle de Ciencias Naturales 2003 (“2001”), 155: 51-72 Una especie nueva de Grundulus (Teleostei: Ostariophysi: Characidae) para Colombia y redescripción de Grundulus bogotensis (Humboldt y Valenciennes, 1833) César Román-Valencia, Hans-Joachim Paepke y Fernando Pantoja Resumen. Se describe una nueva especie de Grundulus (Characiformes, Characidae) proveniente de la Laguna de La Cocha, cuenca del alto Putumayo en los Andes al sur de Colombia. El nuevo taxón se distingue de G. bogotensis por el número mayor de dientes en el dentario (10 vs 8-9), en el maxilar (7-10 vs 4-7), de supraneurales (8 vs 7). En la aleta anal el primer pterigióforo está asociado con los dos primeros radios anales y se observa ancho, mientras que en G. bogotensis se observa delgado. Se redescribe a G. bogotensis, endémico de la sabana de Bogotá, con base en material fresco y topotípico y la designación del neotipo. Se amplia la distribución geográfica del género y ambas especies se encontraron en alopatría. Se incluyen datos originales sobre las características de hábitat y dieta de los taxones, además, de consideraciones sobre la biogeografía histórica del grupo. Palabras clave. Grundulus. Characidae. Nueva especie. Sistemática. Colombia. A new species of Grundulus (Teleostei: Ostariophysi: Characidae) for Colombia and a redescription of Grundulus bogotensis (Humboldt and Valenciennes, 1833). Abstract. This paper describes a new species of Grundulus from Laguna de la Cocha in the upper Putumayo Basin in the Andes of southern of Colombia. The new taxon can be distinguished from G. bogotensis by: a greater number of teeth on the dentary (10 vs. 8-9) and on the maxila (7-10 vs 4-7); a greater number of supraneurals (8 vs 7) and a wide vs narrow first pterygiophore of the anal fin. Grundulus bogotensis, endemic to the Sabana de Bogota, is redescribed based on fresh, topotypical material and the designation of a neotype. The geographic distribution of the genus is extended and the two species are seen as allopatric. Data on habitat, feeding and historical biogeography of both species are included. Key Words. Grundulus. Characidae. Tropical Fishes. New Species. Systematics. Colombia. Introducción El género Grundulus se distribuye en los Andes de Colombia. Se ha descrito una especie: G. bogotensis (Humboldt y Valenciennes, 1833) endémica de la Sabana de Bogotá (Cala y Román-Valencia 1994) y se incluye dentro de la subfamilia Cheirodontinae (Eigenmann 1915, 1922, Dahl 1971, Miles 1971, Géry 1977), aunque el género carece de los caracteres diagnósticos de la subfamilia Cheirodontinae (Malabarba 1998, Weitzman y Malabarba 1999, este trabajo). Su relación filogenética dentro de los Characidae es desconocida y el género es de posición incierta dentro de
Transcript of Una especie nueva de Grundulus (Teleostei: Ostariophysi ... 51-72.pdf · 52 Una especie nueva de...

Memoria de la Fundación La Salle de Ciencias Naturales 2003 (“2001”), 155: 51-72
Una especie nueva de Grundulus (Teleostei: Ostariophysi:Characidae) para Colombia y redescripción de Grundulusbogotensis (Humboldt y Valenciennes, 1833)
César Román-Valencia, Hans-Joachim Paepke y Fernando Pantoja
Resumen. Se describe una nueva especie de Grundulus (Characiformes, Characidae) proveniente dela Laguna de La Cocha, cuenca del alto Putumayo en los Andes al sur de Colombia. El nuevo taxónse distingue de G. bogotensis por el número mayor de dientes en el dentario (10 vs 8-9), en el maxilar(7-10 vs 4-7), de supraneurales (8 vs 7). En la aleta anal el primer pterigióforo está asociado con los dosprimeros radios anales y se observa ancho, mientras que en G. bogotensis se observa delgado. Seredescribe a G. bogotensis, endémico de la sabana de Bogotá, con base en material fresco y topotípicoy la designación del neotipo. Se amplia la distribución geográfica del género y ambas especies seencontraron en alopatría. Se incluyen datos originales sobre las características de hábitat y dieta de lostaxones, además, de consideraciones sobre la biogeografía histórica del grupo.
Palabras clave. Grundulus. Characidae. Nueva especie. Sistemática. Colombia.
A new species of Grundulus (Teleostei: Ostariophysi: Characidae) for Colombia and aredescription of Grundulus bogotensis (Humboldt and Valenciennes, 1833).
Abstract. This paper describes a new species of Grundulus from Laguna de la Cocha in the upperPutumayo Basin in the Andes of southern of Colombia. The new taxon can be distinguished from G.bogotensis by: a greater number of teeth on the dentary (10 vs. 8-9) and on the maxila (7-10 vs 4-7); agreater number of supraneurals (8 vs 7) and a wide vs narrow first pterygiophore of the anal fin.Grundulus bogotensis, endemic to the Sabana de Bogota, is redescribed based on fresh, topotypicalmaterial and the designation of a neotype. The geographic distribution of the genus is extended andthe two species are seen as allopatric. Data on habitat, feeding and historical biogeography of bothspecies are included.
Key Words. Grundulus. Characidae. Tropical Fishes. New Species. Systematics. Colombia.
Introducción
El género Grundulus se distribuye en los Andes de Colombia. Se ha descrito unaespecie: G. bogotensis (Humboldt y Valenciennes, 1833) endémica de la Sabana deBogotá (Cala y Román-Valencia 1994) y se incluye dentro de la subfamiliaCheirodontinae (Eigenmann 1915, 1922, Dahl 1971, Miles 1971, Géry 1977), aunque elgénero carece de los caracteres diagnósticos de la subfamilia Cheirodontinae(Malabarba 1998, Weitzman y Malabarba 1999, este trabajo). Su relación filogenéticadentro de los Characidae es desconocida y el género es de posición incierta dentro de

52 Una especie nueva de Grundulus (Teleostei: Ostariophysi: Characidae)
la familia (Malabarba 1998, Weitzman y Malabarba 1999). La captura de ejemplares de Grundulus en la Laguna de La Cocha en los Andes
al sur de Colombia, mostró que pertenecen a una especie no descrita. Por con-siguiente, el propósito del presente artículo es describir la nueva especie, redescribir aG. bogotensis con base en material fresco y topotipo y la designación del neotipo deesta especie del material colectados durante la época en vida de Alexander VonHumboldt
Materiales y Métodos
Se realizaron colecciones en diversos hábitats de la Laguna de La Cocha yRepresa de Neusa. Se registró la coloración del material en vivo y se pasaron a bolsasde polietileno con formol al 10% para su fijación. El material fue preservadoposteriormente en alcohol etílico al 70%, se depositó en el laboratorio de Ictiología delDepartamento de Biología, Universidad del Quindio, Armenia (IUQ) y en el InstitutoSmithsoniano de Investigaciones Tropicales, Panamá (STRI).
Igualmente, se examinó material depositado en la Unidad de Ictiología, Institutode Ciencias Naturales, Universidad Nacional de Colombia, Bogotá (ICNMNH) y enel Instituto de Investigación de Recursos Biológicos “Alexander Von Humboldt” enVilla de Leyva, Boyacá, Colombia (IAvH) y del Museum fuer Naturkunde derHumboldt-Universitaet zu Berlin (ZMB). Los nombres originales correspondientes aG. bogotensis (tal como Poecilia y Bogotensis) citadas por Humboldt y Valenciennes(1833), Cuvier y Valenciennes (1846) se copiaron tal y como aparecen en estaspublicaciones.
Con los métodos de frecuencia de ocurrencia o aparición y numérica (Hyslop1980), se realizó el análisis de los organismos presentes en 34 estómagos con contenido,correspondientes a 120 ejemplares (18-62 mm y promedio 34,6 mm de largo estándar),capturados entre septiembre de 1999 y septiembre del 2000.
Las medidas de los ejemplares se tomaron con un calibrador digital, hastacentésimas de milímetro. Todas las medidas fueron obtenidas punto a punto, i.e. noortogonal al eje principal del cuerpo. Los recuentos de radios, escamas y dientesempleando estereoscopio, aguja de disección y coloreando en algunos casos con azulde metileno. Las medidas y recuentos (Tabla 1) se realizaron sobre el lado izquierdo delos ejemplares, excepto cuando éstos estaban deteriorados en tal lado. Medidas yconteos sigue a Hubbs y Lagler (1958) y a Weitzman y Malabarba (1999).
Las observaciones de las estructuras óseas y cartílagos se hicieron sobre ejemplaresclareados y teñidos de acuerdo a modificaciones al método descrito por Taylor y VanDyke (1985) y Song y Parenti (1995). La nomenclatura de huesos se basó en Weitzman(1962) y Vari (1995). El conteo y las determinaciones de vértebras se hizo con materialdiafanizado y de radiografias (rayos-x) y sigue a Helfman et al. (1997).

53Mem. Fund. La Salle de Cien. Nat. 155
Resultados
Género: Grundulus Cuvier y Valenciennes, 1846
Grundulus Cuvier y Valenciennes, 1846: 216 (Descripción, para Santafé de Bogotá).
Especie tipo por monotípico: Poecilia bogotensis Humboldt y Valenciennes enHumboldt y Bonpland, 1833: 154-156, 159.
Etimología. El género Grundulus nombrado por Cuvier y Valenciennes (1846) notiene un significado conocido. Aunque es posible que este derive del reemplazo de laF por la Gr en el género Fundulus (del latin fundulus, que significa fondo), que Cuviery Valenciennes (1846) citaron, aunque no lo justifican.
Sinonimia. Poecilia Humboldt y Valenciennes en Humboldt y Bonpland 1833: 154-156, 159, pl 45 fig. 1 (descripción: Sabana de Bogotá).
Ctenocharax Regan, 1907, 402 (especie tipo: Ctenocharax bogotensis sp nov.)
Diagnósis. El género Grundulus se puede distinguir por los siguientes caracteresderivados: Ausencia de aleta adiposa, número reducido de escamas perforadas en lalínea lateral (0-8); radios en la aleta anal (iv-v, 11-12), presencia de un supraorbital;orbitoesfenoides y rinoesfenoides fusionados y completamente óseos; hueso pélvicodelgado; maxilar con una pequeña muesca en el borde posterior; dentario con una odos pequeñas fenestras; premaxilar y el dentario sólo con dientes cónicos y elsupraoccipital forma el extremo posterior de la fontanela craneal.
Comentarios. Véase además a Malabarba (1998) y Weitzman y Malabarba (1999)para una nueva definición de Cheirodontinae y el estado sistemático de Grundulus.De acuerdo al análisis efectuado en el presente trabajo, las sinapomorfias observadasen el género Grundulus, excepto la ausencia de aleta adiposa, corresponden a las ano-tadas en su diagnosis.
En el presente trabajo no se observa a los pterigióforos cortos, no alargado y que seubican en la parte anterior en Grundulus como se citó en Weitzman y Malabarba(1999).
Grundulus cochae sp nov. Tabla 1, Figuras 1, 3a-6a.
Holotipo. IUQ 453, recolectado en Colombia, Nariño, Pasto, vereda Romerillo,Laguna de la Cocha (01°05’22”N-77°09’50”O, 2809 m s.n.m.), el 17 de julio del 2000,por C. Román-V., F. Pantoja y C. Pantoja.
Paratipos. IUQ 454, Colombia Nariño: 66 ejemplares colectados con el holotipo.STRI 1369: un ejemplar colectado con el holotipo. IUQ 455: 51 ejemplares,Colombia, Nariño, Pasto, vereda. Romerillo, Laguna de la Cocha en canales de

54 Una especie nueva de Grundulus (Teleostei: Ostariophysi: Characidae)
embarcación, colectados por F. Pantoja y C. Pantoja, el 15 de agosto de 2000. IUQ456: seis ejemplares, Colombia, Nariño, Pasto, Laguna de la Cocha, colectados por F.Pantoja y C. Pantoja, el 20 mayo de 2000.
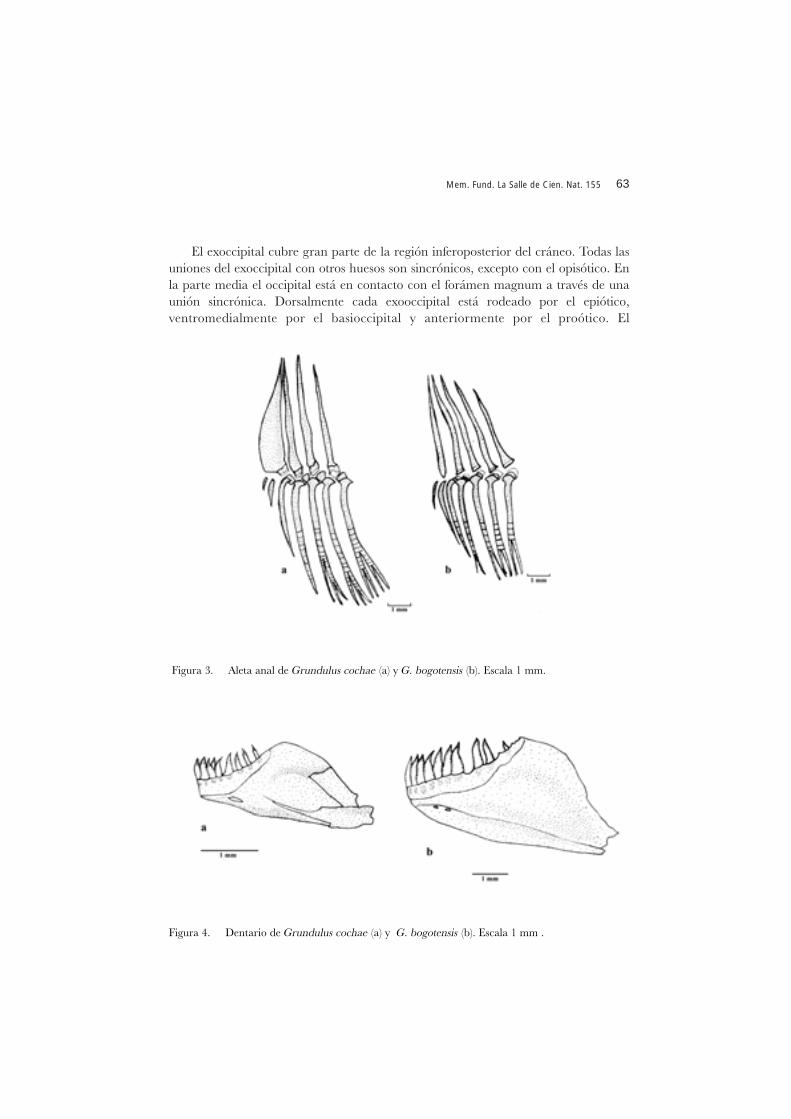
Diagnosis. La especie se puede distinguir de G. bogotensis por el mayor número dedientes en el dentario (10 vs 8-9), en el maxilar (7-10 vs 4-7), de supraneurales (8 vs 7)y de radios simples en las aletas pectorales (iii vs ii) por la forma ancha del primerpterigióforo en la aleta anal (delgado en G. bogotensis) (Figuras 3a y b).
Descripción. Cuerpo corto y robusto en todos los ejemplares. Perfil dorsal de lacabeza en línea curva área superficial en el área de las órbitas convexa. Perfil dorsaldel cuerpo curvo desde el borde posterior de la cabeza a nivel del supraoccipital hasta
G. cochae G. cochae G. bogotensis G. bogotensis G. bogotensis Holotipo Paratipos Neotipo Topotipos Topotipos ZMB
Longitud estándar 54,5 24,4-61,4 (40,2) 50,7 13,8-79,0 (36,1) 38,9-50,7 (45,6)Longitud total 63,3 27,0-65,3 (48,1) 60 18,1-93,0 (44,0) 47,1-59,8 (45,6)
Morfometría Porcentaje de la longitud estándar
1. Profundidad del cuerpo 33,9 25,6-36,4 (29,2) 39,1 18,7-37,9 (28,2) 32,7-39,4 (36,3) 2. Longitud hocico-aleta dorsal 61,4 54,1-69,1 (60,1) 63,9 53,4-64,4 (59,1) 59,9-64,1 (62,6) 3. Longitud hocico-aleta pectoral 28,4 26,0-30,7 (28,6) 29,8 24,3-33,0 (28,3) 28,0-33,7 (30,8) 4. Longitud hocico-aleta pélvica 53,3 51,2-60,2 (55,7) 57,6 46,8-63,9 (53,9) 55,0-62,6 (58,8) 5. Longitud hocico-aleta anal 68 65,2-72,8 (69,4) 73,4 63,2-79,0 (68,3) 73,1-77,6 (75,2) 6. Longitud aleta dorsal-hipurales 42,3 38,8-48,2 (43,9) 41,8 37,8-49,0 (43,6) 37,0-43,4 (41,0) 7. Longitud aleta dorsal 20,7 16,8-24,4 (20,5) 18,9 16,3-29,2 (21,9) 16,1-19,8 (18,5) 8. Longitud aleta pectoral 15,2 10,4-17,4 (14,1) 17,8 8,0-18,7 (13,8) 14,9-18,3 (16,6) 9. Longitud aleta pélvica 13,7 9,1-18,4 (13,1) 15,2 10,5-20,0 (14,2) 11,4-17,5 (15,3) 10. Longitud aleta anal 14,1 11,1-18,6 (14,7) 13,2 10,3-23,8 (16,9) 12,0-18,1 (15,3) 11. Longitud aleta dorsal-aleta anal 34 24,4-35,1 (30,2) 34,3 23,3-38,2 (30,0) 28,4-37,2 (32,4) 12. Longitud aleta dorsal-aletas pectorales 43 36,4-48,1 (42,0) 45 34,2-48,2 (40,9) 33,00-45,0 (39,6) 13. Profundidad del pedúnculo caudal 13,7 10,4-14,5 (12,5) 15 8,4-14,8 (12,1) 13,4-15,9 (14,7) 14. Longitud del pedúnculo caudal 11,2 7,0-15,4 (11,6) 15,2 10,0-19,3 (13,7) 15,2-19,3 (16,9) 15. Longitud de la cabeza 26,1 24,5-30,0 (2646) 26,8 20,2-31,2 (26,1) 26,8-29,9 (28,2)
Porcentaje de la longitud cabeza
16. Longitud hocico 29,9 23,3-31,5 (27,9) 23,5 17,6-37,4(27,4) 16,7-23,5 (19,8)17. Diámetro del ojo 24,1 22,2-35,5 (29,0) 22,8 23,9-41,9 (31,7) 22,1-29,0 (24,6)18. Longitud postorbital de la cabeza 41,3 30,8-53,0 (43,4) 51,5 35,6-57,1 (42,6) 47,8-59,0 (54,7)19. Longitud del hueso maxilar 21,5 16,2-31,1 (22,8) 26,5 15,2-30,7 (22,2) 24,6-32,3 (28,4)20. Ancho interorbital 33,8 27,3-41,1 (33,1) 33,1 25,3-48,4 (34,9) 25,4-33,1 (30,0)21. Longitud mandíbula superior 32 21,4-36,3 (27,7) 36,8 19,8-36,6 (27,3) 29,0-44,4 (35,4)
Merística Número de escamas en la línea lateral 0 0-8 3 0-8 1-4Número de radios en la aleta anal iv,12 iv,11-12 15 iv-v,11-12 14-16
Número de radios en la aleta pélvica ii,6 i-ii,6 7 ii,6 7 - 8
Número de radios en las aletas pectorales iii,10 iii,10 13 ii,13 13-17Número de radios en la aleta dorsal ii,9 ii,8-9 10 ii-iii,10-13 8-11
Tabla 1. Datos morfométricos y merísticos de Grundulus cochae sp nov. y G. bogotensis.Longitudes estándar y total en mm. Promedios se dan entre paréntesis.

55Mem. Fund. La Salle de Cien. Nat. 155
el origen de la aleta dorsal, en línea oblicua desde la parte posterior de la aleta dorsalhasta la base de la aleta caudal. Perfil ventral del hocico hasta la base de la aleta analcurvo, más pronunciado en la parte posterior de las aletas pectorales. La mayorprofundidad del cuerpo se localiza en la parte anterior del origen de la aleta dorsal.Pedúnculo caudal ligeramente comprimido en todos los ejemplares. Área predorsal sinescamas. No se observa dimorfismo sexual. Cabeza y hocico cortos; mandíbulasiguales, boca terminal con los labios blandos y flexibles, que cubren totalmente lahilera de dientes de los premaxilares.
Osteología. Presenta tres huesos infraorbitales osificados, sin segmento alguno de lalínea lateral. El primer infraorbital es un hueso corto y es el más pequeño, cubriendoparcialmente la parte dorso posterior del maxilar. El segundo infraorbital es dos vecesmás grande que el primero y cubre la región postero dorsal del articular. El tercerinfraorbital tiene su borde ventral y posterior en contacto con la superficie delpreopérculo. Supraorbital presente.
Opérculo largo, plano y ubicado en la parte lateral de la mejilla, debajo de laregión posterior del hiomandibular. El subopérculo es una leve hoja de huesoparcialmente ampliada a lo largo del borde interno e inferior del opérculo. Elinteropérculo es delgado y se ubica anterior al subopérculo e inferior al preopérculo.El extremo anterior del preopérculo encaja en una ranura sobre la región mediainferior del cuadrado, mientras que la dorsal se sitúa entre el infraorbital 2 grande y elopérculo. La superficie lateral del preopérculo no contiene canales laterosensoriales.
Mandíbula inferior con una serie de diez dientes de diferente tamaño a cada ladode los dentarios y en línea recta, luego se observa dos dientes muy cercanos entre sí ymás delgados, separados por un espacio sin dientes del resto (Figura 4a). Todos los
Figura 1. Grundulus cochae, Holotipo, IUQ 453: 54,5 mm LE; Colombia: Laguna de La Cocha.

56 Una especie nueva de Grundulus (Teleostei: Ostariophysi: Characidae)
dientes cónicos. Dentario unido a un articular a través de dos apófisis, presenta dospequeñas fenestras en el borde antero ventral (Figura 4a).
El premaxilar presenta un proceso lateral corto con una hilera de dientes cónicossobre cada uno. Siete a ocho dientes en cada lado, orientados en línea recta. El maxilares corto y el extremo posterior se extiende hasta el extremo inferior de los dentarios.Maxilar con siete a diez dientes cónicos de diferente longitud, ubicados en el bordeanterior y dorsal. Maxilar con un proceso doblado en su extremo superior y con unamuesca en forma de saco en su región posterior (Figura 5a).
El hueso etmoide es estrecho, se proyecta hacia adelante y frente de los frontales.Anteriormente el etmoide termina en dos procesos agudos a cada lado, forma unacavidad donde se aloja los procesos laterales de los premaxilares. El prevomer carecede dientes y es plano. Las dos proyecciones del prevomer se ubican junto al procesoespinoso del extremo posterior de los premaxilares. El etmoide lateral forma la paredantero superior de cada orbita.
La superficie dorsal del cráneo está cubierto, en parte, por dos frontales. Losfrontales no están fusionados. Estos huesos están separados por la fontanela craneana.Fontanela angosta a nivel del extremo anterior, ancha en la parte posterior de losfrontales en el ámbito de los parietales y en unión con el supraoccipital. Los dosparietales son grandes y cubren la base de la espina del supraoccipital. Estos parietalesestán limitados por el canal parietal en los lados y por el canal supratemporal en elextremo anterior. Un extraescapular en contacto y unido con el extremo posterior delparietal.
El supraoccipital forma el extremo posterior de la fontanela craneal. Lateral yanteriormente este hueso tiene un arco hacia la parte media posterior del parietal.Ventral y lateralmente el resto del supraoccipital se ubicó sobre el epiótico y elexoccipital por medio de una unión sincrónica. La superficie dorsal del supraoccipitalcontiene una ranura ancha, el canal supraoccipital, que pasa atrás del borde posteriorde la fontanela dorsal del cráneo hasta cerca al borde posterior de la espina delsupraoccipital.
El exoccipital cubre gran parte de la región inferoposterior del cráneo. Todas lasuniones del exooccipital con otros huesos son sincrónica excepto con el opistótico. Laparte media el occipital está en contacto con el forámen magnum a través de unaunión sincrónica. Dorsalmente cada exoccipital está rodeado por el epiótico, ventro-medialmente por el basioccipital y anteriormente por el proótico. El basioccipital estárodeado por el proótico anteriormente, el paraesfenoides ventromedialmente y elexooccipital dorsalmente. Todas las uniones, excepto con el paraesfenoides, son sin-crónicas. El epiótico forma con el extraescapular un espacio tubular para un canalsemicircular. Medial y ventralmente cada epiótico está rodeado por el exoccipital, ydorso medialmente por el supraoccipital. La región antero inferior del epiótico está encontacto con el pterótico, mientras un proceso ancho y despuntado del epiótico seproyecta alrededor de la fosa postemporal en contacto con el parietal arriba y elpterótico abajo. La parte ventral de una vertical a través del canal semicircular cubre

57Mem. Fund. La Salle de Cien. Nat. 155
la región del exoccipital debajo del epiótico. Dorsalmente el pterótico está cubierto porsu respectivo parietal y frontal y es completamente óseo. Un gran forámen presenteentre el esfenótico y el pterótico. El esfenótico se localiza debajo de los frontales y laparte superior de los pteróticos. El frontal y el pterótico cubren el esfenótico formandouna red dilatada.
La región postero superior de la órbita esta formada por el pteroesfenoide con elorbitoesfenoide. Lateralmente el pteroesfenoide está rodeado por el frontal, pos-teriormente por el esfenótico, ventralmente por el proótico y anteriormente por elorbitoesfenoides en unión sincronal. Se observa un pequeño forámen entre elpterosfenoide y el orbitosfenoide. El borde dorsal del orbitoesfenoides está cubierto porlos frontales. En vista lateral el orbitoesfenoides es de forma triangular. Elrinoesfenoides es ventral al orbitoesfenoides, se proyecta hacia la parte anterior delparaesfenoides. El rinoesfenoides se observa completamente óseo, en unión sincronalcon el orbitoesfenoides y está dividido por una sutura. Presentó dos pequeñosforámenes en su parte anterior uno arriba y otro abajo y dos pequeños forámenes enla parte posterior, adyacentes entre sí. No se observan apófisis en ambas estructuras.El paraesfenoides es delgado y no se encuentra dividido. La parte anterior cubre lasuperficie antero dorsal del prevomer; la parte posterior cae bajo y en contacto con elproótico, así como en la parte lateral, ventral e interna del cráneo. Se observan huesosnasales, son alargados, tubulares y su extremo anterior alcanza el extremo dorsal delmaxilar; la parte posterior del nasal apenas alcanza el extremo anterior delsupraorbital.
La parte superior del hiomandibular es de forma aplanada. Su brazo inferior escorto, termina justo en frente del ángulo formado por la punta del borde orbital delpreopérculo. El cuadrado se ubica cubriendo la superficie dorsal del preopérculo.Posee tres brazos. Uno superior que se proyecta hacia el metapterigoide y es óseo. Losdos brazos posteriores son delgados se proyectan arriba y abajo del simpléctico.
El arco palatino se localiza arriba del cuadrado y el simpléctico. El borde posteriordel metapterigoide en contacto con la superficie dorso posterior del simpléctico y laparte anterior del hiomandibular. El borde anterior del metapterigoide en contactocon el cuadrado y el mesopterigoide. El mesopterigoide y ectopterigoide sin dientes, seproyectan en la parte superior del maxilar en la región del lateroetmoide. El extremoanterior del palatino se observa cartilaginoso y alcanza el ángulo formado por laapófisis del maxilar, justo debajo del hueso nasal.
En el aparato de Weber, el centro de la primera vértebra es un disco simple condos depresiones dorsolaterales para la recepción del proceso articular ventral de cadaescafium. La segunda vértebra tiene un proceso lateral alargado. El pedículo del arconeural de la tercera vértebra no está fusionado al centro de la tercera vértebra. Lacuarta vértebra tiene un proceso lateral similar a una paleta, estrecho en su base yancho en el resto de su superficie, el cual se extiende fuera de la parte media. Elcomplejo neural arriba de las primeras cuatro vértebras es un hueso sólido, tiene una

58 Una especie nueva de Grundulus (Teleostei: Ostariophysi: Characidae)
sutura suave en su superficie medio ventral. La parte anterior del complejo neuraltermina en el extremo posterior del cráneo. Cada clastrom osificado se ubica debajode la parte antero lateral inferior del complejo neural a lo largo del borde dorsoposterior interno de cada escafium. El opistótico tiene un brazo lateral largo sinproceso alguno. El tripos es un hueso complejo con el extremo anterior unido alligamento interóseo. Tiene un proceso largo, ancho y en forma de paleta.
Las vértebras precaudales no poseen espinas hemales, se observan en número de16 e incluyen las cuatro vértebras del aparato de Weber. De la quinta hasta la 16 sonmuy parecidas. Cada una de estas vértebras tiene dos precigoapófisis neurales, un arconeural y una espina neural. No poseen postcigoapófisis hemales. Posee 16 vértebrascaudales. Las vértebras 19 a 32 son muy semejantes en su estructura. La vértebra 18posee una espina hemal más corta y alargada, terminando en punta. Dorsalmentecada vértebra posee una espina neural, un arco neural y dos precigoapófisis neuralesy dos postcigoapófisis neurales. Ventralmente cada vértebra posee una espina hemal yun arco hemal. Los pre y postcigo apófisis se notan en todas las vértebras. Las últimastres vértebras caudales sostienen la aleta caudal. Se observan siete hipurales asociadoscon la última vértebra y el urostilo. Entre los extremos anteriores de los hipurales 5-7se forma un foramen redondo. Un segundo forámen rectangular se observó en el sextohipural. El extremo distal alcanza los radios medios de la caudal. Hay 11/11 radiosprincipales. El urostilo se extiende al extremo de la parte superior del abanico dehipulares, el cual es cubierto por los uroneurales. En la parte postero lateral de losurostilos se localiza los uroneurales. En el dorso de los urostilos hay tres epurales, eltercero se halla fusionado. No hay espinas hipurales.
Las proyecciones epineurales se proyectan hacia abajo y hacia arriba de cada arcoy espina neural. Hay ocho a nueve epineurales, cinco a seis anterires son simples, losdos a tres posteriores son ramificados. Se observan diez epipleurales no divididos entrelos músculos hipaxiales, proyectados hacia arriba y hacia abajo de la espina y el arcohemal.
Borde de la aleta dorsal redondeado. El componente proximal y medial de losprimeros pterigióforos insertados entre las espinas 14-19. Todos los radiales proximalescon quilla. Hay ocho supraneurales entre los músculos epaxiales anterior a la aletadorsal, con cartílago en sus extremos superior e inferior y se proyectan entre las espinasneurales. No se observa aleta adiposa.
La cintura pectoral presenta un proceso dorsal del cleitro puntiagudo, que alcanza1/3 partes del supracleitro. El cleitro es un hueso corto y ancho, se ubica en la partemedia posterior del opérculo. El ala anterior está muy desarrollada, observándosecomo una extensa o sobresaliente paleta, la cara media es la superficie para larecepción de la parte lateral de la escápula. El borde anterior del cleitro se localizajusto en la parte lateral del coracoides y sus bordes que se observan unidos porcartílago. Hay dos supracleitro, se localizan en el borde postero inferior del cleitro. Elcoracoides es un hueso largo y achatado, su superficie antero mediana está en contacto

59Mem. Fund. La Salle de Cien. Nat. 155
con su lado opuesto y hay un pequeño espacio entre estos. En la superficie delcoracoides hay uno o dos grandes foramenes. La región dorsal del coracoides seextiende lateralmente el cleitro. Del extremo anterior del coracoides se ubica el cleitroy del lado anterior la escápula. Hay seis radiales proximales. El extremo proximal delradial lateroanterior tiene un proceso articular extendido por debajo de la superficieventral de la escápula. Arriba del cleitro se ubica el supracleitro, que es elongado. Estetiene una fosa relativamente mediana para la recepción del proceso espinoso dorsal delcleitro. El extraescapular esta libre cerca de la parte postero lateral del cráneo.
Borde de las aletas pélvicas redondeado, alcanza el origen de la aleta anal. Elhueso pélvico es una estructura alargada, que se ubica paralelo al vientre. La regiónanterior está en contacto con los radios y es ancho presentando un proceso largo ydespuntado en su parte lateral. La región anterior es puntiaguda y no posee cartílago.El proceso ísquico es alargado con un pequeño forámen en su extremo anterior yextendida por detrás del borde postero medial (Figura 6a).
En la aleta anal el primer pterigióforo está asociado con los dos primeros radiosanales y es ancho (Figura 3a). Radios procurrentes de la aleta caudal 10/9, tanto dorsalcomo ventralmente. Número total de vértebras: 31-32.
Coloración en vivo. Area dorsal y lateral oscura, menos pronunciada en la partelateral del cuerpo; área ventral amarillo claro; aletas oscuras, excepto la aleta caudalque se observó rosada-oscura.
Una mancha humeral oscura alargada verticalmente. No se observó manchapeduncular. Aleta caudal con una banda oscura verticalmente alargada visible enejemplares claros, no visible en individuos más oscuros.
Distribución. Conocida sólo de su localidad típica.
Aspectos bioecológicos. La especie habita en las riberas y pequeños afluentes de laLaguna de la Cocha que presentaron abundantes macrófitas subacuáticas conocidascomo Berros (Nasturtium officinale, Crusifera, Brasicaceae) y se ubica en la superficiede la columna de agua. Sustrato conformado por lodo y abundante material orgánicode origen vegetal en descomposición.
Temperatura superficial 12,8-14 °C; temperatura del aire 11,0 °C; pH alrededorde 7; oxígeno: 0,5-0.7 ppm y 8% de saturación; alcalinidad 9.1 mg/l; conductividad:37,7 µs/cm y sólidos suspendidos 3,1 mg/l; CO2 25 mg/l; profundidad 0,75 m y aguasde color oscuro.
La dieta es poco diversa con predominio de crustáceos. Se observa que elcontenido estómacal está constituido por copépodos (Diaptomus sp), CladócerosBosmina sp y Chydorus sp. También por moscas adultas y larvas (Chironomidae) ycaracoles Pisidium sp (Tabla 2).
En la Laguna de la Cocha también se encuentran el bagre trocomícteridoEremophilus sp y la trucha Oncorhynchus mykiss. Es muy probable que esta segundaespecie introducida sea un depredador de Grundulus cochae, pues los habitantes de la

60 Una especie nueva de Grundulus (Teleostei: Ostariophysi: Characidae)
laguna han hallado ejemplares de G. cochae en el estómago de O. mykiss. Nuestrasobservaciones indican que G. cochae y O. mykiss coexisten en diferentes hábitats.Oncorhynchus mykiss se localiza en la zona limnética, mientras G. cochae se ubica enla zona litoral con condiciones físicas y químicas inaccesibles para la sobrevivencia deO. mykiss.
Malformaciones. Se observó un ejemplar de 61,4 mm de largo estándar que no teníauna aleta pélvica. En su lugar se registró escamas. Dos ejemplares con 31,9 y 34,3 mmde largo estándar presentaron el pedúnculo caudal torcido.
Etimología. Epíteto específico alusivo a la localidad típica del nuevo taxon: la Lagunade la Cocha al sur de los Andes de Colombia.
Comentarios. La especie es parecida y relacionada con G. bogotensis. Sin embargo,el análisis de caracteres osteológicos (Figuras 3-6) y merísticos (Tabla 1) indican queambos taxones son diferentes y que están en alopatría. De esta manera, se amplía ladistribución geográfica del género Grundulus. Con base en trabajos similares que llevóa cabo el primer autor en la región (Román-Valencia 2000a, b) se concluye que G.cochae es una especie endémica.
Grundulus cochae y G. bogotensis registraron una línea lateral reducida o ausente(Tabla 1). Sin embargo, Weitzman y Malabarba (1999) citaron para Grundulus decinco a seis escamas perforadas en la línea lateral. Estos autores plantearon que lareducción o ausencia de la línea lateral se derivó independientemente.
Grundulus bogotensis (Humboldt y Valenciennes, 1833) Tabla 1, Figuras 2, 3b-6b
Poecilia bogotensis Humboldt y Valenciennes 1833 en Humboldt y Bonpland: 154-156,
159, pl. 45, fig. 1 (descripción: Sabana de Bogotá).
Organismos Cantidad Frecuencia (%) (%)
Diaptomus sp 56 49,3Bosmina sp 24 26,1Chydorus sp 19 21,7Pisidium sp 0,4 1,5Chironomidae 0,3 1,5
Tabla 2. Contenido estomacal de Grundulus cochae sp.nov., laguna de la Cocha, Colombia.

61Mem. Fund. La Salle de Cien. Nat. 155
Grundulus bogotensis. Cuvier y Valenciennes 1846: 216-220 (especie tipo de Grundulus).
Grundulus bogotensis.- Eigenmann, 1922: 128 (Sabana de Bogotá y quebrada Suaita en
Santander); Dahl, 1971: 133 (Sabana de Bogotá y Santander); Miles, 1971: 143 (Cita); Géry,
1977: 607 (Cita en clave); Cala y Román-Valencia, 1994: 19 (Cita); Weitzman y Malabarba,
1999: 4-40 (Cita y análisis filogenético).
Ctenocharax Regan, 1907: 402-403 (Especie tipo: Ctenocharax bogotensis sp nov.)
Diagnosis. La especie se puede distinguir de G. cochae por el número menor dedientes en el dentario (ocho a nueve vs diez), en el maxilar (cuatro a siete vs siete adiez), de supraneurales (siete vs ocho); el primer pterogióforo de la aleta anal en G.bogotensis es delgado mientras que en G. cochae es ancho (Figuras 3a y b)
Descripción. Cuerpo corto y robusto. Perfil dorsal de la cabeza en línea cóncavo;área superficial en el área de las órbitas convexo. Perfil dorsal del cuerpo curvo desdeel borde posterior de la cabeza a nivel del supraoccipital hasta el origen de la aletadorsal, en línea oblicua desde la parte posterior de la aleta dorsal hasta la base de laaleta caudal. Perfil ventral del hocico hasta la base de la aleta anal curvo, más pro-nunciado en la parte posterior de las aletas pectorales. La mayor profundidad delcuerpo se localiza en la parte anterior del origen de la aleta dorsal. Pedúnculo caudalligeramente comprimido en todos los ejemplares. Cabeza y hocico cortos; mandíbulasiguales, boca terminal; labios blandos, f lexibles, que cubren totalmente la hilera dedientes de los premaxilares. Área predorsal sin escamas. No se observa dimorfismosexual.
Osteología. Presenta tres huesos infraorbitales osificados, sin segmento alguno de lalínea lateral. El primer infraorbital es un hueso corto y es el más pequeño, cubre par-cialmente la parte dorso-posterior del maxilar. El segundo infraorbital es dos veces másgrande que el primero y cubre la región postero-dorsal del articular. Tercer
Figura 2. Grundulus bogotensis. Neotipo, ZMB 33306: 50.7 mm LE; Colombia, Cundinamarca,Sabana de Bogotá.

62 Una especie nueva de Grundulus (Teleostei: Ostariophysi: Characidae)
infraorbital un poco más grande que el primero y de forma triangular, con el bordeventral y posterior en contacto con la superficie posterior del segundo infraorbital y suparte latero inferior en contacto con el preopérculo. Un supraorbital.
El opérculo es un hueso largo, plano y ubicado en la parte lateral de la mejilla ydebajo de la región posterior del hiomandibular. El subopérculo parcialmenteampliado a lo largo del borde interno e inferior del opérculo. El interopérculo, que esdelgado se ubicó anterior al subopérculo e inferior al preopérculo. El extremo anteriordel preopérculo encaja en una ranura sobre la región media inferior del cuadrado,mientras que la dorsal se sitúa entre el infraorbital 2 grande y el opérculo. La superficielateral del preopérculo no contiene canales latero-sensoriales.
Mandíbula inferior con una serie de ocho a nueve dientes de igual tamaño a cadalado de los dentarios y orientados en forma irregular (Figura 4b). Todos los dientescónicos. Dentario unido a un articular a través de dos apófisis y con una pequeñafenestra alargada en su borde antero ventral (Figura 4b).
Premaxilar con un proceso lateral corto y una hilera de dientes cónicos sobre cadauno. Ocho dientes a cada lado, orientados en línea recta. El maxilar es corto y elextremo posterior apenas alcanzó el extremo inferior del dentario. Maxilar con cuatroa siete dientes cónicos, ubicados en el borde anterior y dorsal. Maxilar con un procesolargo y oblicuo y una pequeña muesca en el borde antero medial (Figura 5b).
El hueso etmoide es estrecho, se proyecta hacia delante y frente a los frontales.Anteriormente el etmoide termina en dos procesos agudos a cada lado, forma unacavidad donde se aloja los procesor laterales de los premaxilares. El prevomer carecede dientes y es plano. Las dos proyecciones del prevomer se ubican junto al procesoespinoso del proceso posterior de los premaxilares. El etmoide lateral forma la paredantero superior de cada órbita. Los frontales no están fusionados. Estos huesos estánseparados por la fontanela craneana. Fontanela corta a nivel de los frontales, ancha enel ámbito de los parietales y en unión con el supraoccipital.
La superficie dorsal del cráneo está cubierta, en parte, por dos frontales. Losfrontales no están fusionados. Estos huesos están separados por la fontanela craneana.Fontanela angosta a nivel del extremo anterior, ancha en la parte posterior de losfrontales, en el ámbito de los parietales y en unión con el supraoccipital. Los dosparietales son grandes y cubren la base de la espina del supraoccipital. Estos parietalesestán limitados por el canal parietal en los lados y por el canal supratemporal en elextremo anterior. Un extraescapular en contacto y unido con el extremo posterior delparietal. El supraoccipital forma el extremo posterior de la fontanela craneal. Lateraly anteriormente este hueso proyecta un arco hacia la parte medio posterior delparietal. Ventral y lateralmente el resto del supraoccipital se ubica sobre el epiótico yel exoccipital por medio de una unión sincrónica. La superficie dorsal delsupraoccipital contiene una ranura ancha, el canal supraoccipital, que pasa atrás delborde posterior de la fontanela dorsal del cráneo hasta cerca al borde posterior de laespina del supraoccipital.
63Mem. Fund. La Salle de Cien. Nat. 155
El exoccipital cubre gran parte de la región inferoposterior del cráneo. Todas lasuniones del exoccipital con otros huesos son sincrónicos, excepto con el opisótico. Enla parte media el occipital está en contacto con el forámen magnum a través de unaunión sincrónica. Dorsalmente cada exooccipital está rodeado por el epiótico,ventromedialmente por el basioccipital y anteriormente por el proótico. El
Figura 4. Dentario de Grundulus cochae (a) y G. bogotensis (b). Escala 1 mm .
Figura 3. Aleta anal de Grundulus cochae (a) y G. bogotensis (b). Escala 1 mm.

64 Una especie nueva de Grundulus (Teleostei: Ostariophysi: Characidae)
basioccipital está rodeado por el proótico anteriormente, el paraesfenoidesventromedialmente y el exoccipital dorsalmente. Todas las uniones, excepto con elparaesfenoides, son sincrónicas. El epiótico forma con el extraescapular un espaciotubular para un canal semicircular. Medial y ventralmente cada epiótico está rodeadopor el exoccipital y dorsomedialmente por el supraoccipital. La región antero-inferiordel epiótico en contacto con el pterótico, mientras un proceso ancho y despuntado delepiótico se proyecta de la fosa postemporal en contacto con el parietal arriba y elpterótico abajo. La parte ventral de una vertical a través del canal semicircular cubrela región del exoccipital debajo del epiótico. Dorsalmente el pterótico está cubierto porsu respectivo parietal y frontal y es completamente óseo. Un gran forámen presenteentre el esfenótico y el pterótico. El esfenótico se localiza debajo de los frontales y laparte superior de los pteróticos. El frontal y el pterótico cubren el esfenótico formandouna red dilatada.
La región postero superior de la órbita está formada por el pteroesfenoide con elorbitoesfenoide. Lateralmente el pteroesfenoide está rodeado por el frontal, poste-riormente por el esfenótico, ventralmente por el proótico y anteriormente por elorbitoesfenoides en unión sincronal. No se observa foramen alguno entre elpteroesfenoide y el orbitoesfenoide. El borde dorsal del orbitoesfenoides está cubiertopor los frontales. En vista lateral el orbitoesfenoides es de forma redondeada. El rino-esfenoides se observa completamente óseo, en unión sincronal con el orbitoesfenoidesy está dividido por una sutura. En algunos casos se observó un pequeño forámen en suparte posterior. No se observan apófisis en ambas estructuras. El paraesfenoides es unhueso delgado y no se observa dividido. La parte anterior cubre la superficie antero
Figura 5. Maxilar de Grundulus cochae (a) y G. bogotensis (b). Escala 1 mm .

65Mem. Fund. La Salle de Cien. Nat. 155
dorsal del prevomer; la parte posterior cae bajo y en contacto con el proótico, así comoen la parte lateral, ventral e interna del cráneo. Se observan huesos nasales, sonalargados con el extremo anterior alcanzando el extremo anterior del supraorbital.
La parte superior del hiomandibular es de forma plana. Su brazo inferior es corto,termina justo frente del ángulo formado por la punta del borde orbital del preopérculo.El cuadrado se ubica cubriendo la superficie dorsal del preopérculo. El cuadradoposee tres brazos. Uno superior que se proyecta hacia el metapterigoide y es óseo. Losdos brazos posteriores son delgados, se proyectan arriba y abajo del simpléctico. Elborde posterior del metapterigoide está en contacto con la superficie dorso-posteriordel simpléctico y la parte anterior del hiomandibular. El borde anterior delmetapterigoide en contacto con el cuadrado y el mesopterigoide. El mesopterigoide yectopetrigoide sin dientes, se proyectan en la parte anterior del maxilar en la región dellateroetmoide. El extremo anterior del palatino es cartilaginoso y alcanza el ánguloformado por la apófisis del maxilar, justo debajo del hueso nasal.
En el aparato de Weber el centro de la primera vértebra es un disco simple condos depresiones dorso laterales para la recepción del proceso articular ventral de cadaescafium. La segunda vértebra tiene un proceso lateral alargado. El pedículo del arconeural de la tercera vértebra no está fusionado al centro de la vértebra. La cuartavértebra tiene un proceso lateral similar a una paleta: estrecho en su base y ancho enel resto de su superficie, la cual se extiende fuera de la parte media. El complejo neuralarriba de las primeras cuatro vértebras es un hueso sólido, tiene una sutura en lasuperficie medio ventral y dos pequeños forámenes en la superficie dorsal. El extremoantero dorsal termina con una apófisis ancha y despuntada y en el extremo posteriordel cráneo. Cada clastrom osificado se ubica debajo de la parte antero-lateral inferiordel complejo neural y a lo largo del borde posterior interno de cada escafium. Elopistótico tiene un brazo lateral largo sin proceso alguno. El tripos es un huesocomplejo con el extremo anterior unido al ligamento interóseo. Tiene un procesolargo, ancho y punteado.
Las vértebras precaudales no poseen espinas hemales y se hallan en número de 16e incluyen las cuatro vértebras del aparato de Weber. De la quinta hasta la 17 son muyparecidas. Cada una de las vértebras tiene dos postcigoapófisis neurales, un arconeural y una espina neural.
Poseen hemales postcigoapófisis. Posee 16 vértebras caudales. Las vértebras 19 a33 son muy semejantes en su estructura. La vértebra 18 posee una espina hemal cortay ancha, con un par de apófisis en su borde postero lateral. Dorsalmente cada vértebraposee una espina neural, un arco neural, dos precigoapófisis neurales y dospostcigoapófisis neurales. Ventralmente cada vértebra posee una espina hemal y unarco hemal. Los pre y postcigoapófisis se encuentran en todas las vértebras. Lasúltimas tres vértebras caudales sostienen la aleta caudal. Se observan siete hipuralesasociados con la última vértebra y el urostilo. Entre los extremos anteriores de loshipurales 5-7 se forma un forámen redondo. Además, se observan pequeños forámenesen la superficie de estos hipurales. Hay 11/11 radios principales. El urostilo se extiende

66 Una especie nueva de Grundulus (Teleostei: Ostariophysi: Characidae)
al extremo de la parte superior del abanico de hipurales, el cual está cubierto por losuroneurales. En la parte postero lateral de los urostilos se localizó los uroneurales. Enel dorso de los urostilos hay uno o dos epurales. No hay espinas hipurales.
Las proyecciones epineurales se proyectan hacia abajo y hacia arriba de cada arcoy espina neural. Hay ocho a nueve epineurales, cuatro a cinco son simples, el resto sonramificados. Se observan diez epipleurales no divididos entre los músculos hipaxialesque se proyectan hacia arriba y hacia abajo de la espina y el arco hemal.
Borde de la aleta dorsal redondeado. El componente proximal y medial de losprimeros pterigióforos insertados entre las espinas 14-19. Todos los radiales proximalescon quilla. Hay siete supraneurales entre los músculos epaxiales anteriores a la aletadorsal. El extremo inferior de estos huesos se proyecta entre las espinas neurales. No seobserva aleta adiposa.
La cintura pectoral presenta un proceso dorsal del cleitro puntiagudo, que alcanzala tercera parte del supracleitro. El cleitro es largo, ubicado en la parte media posteriordel opérculo. El ala anterior está muy desarrollada, observándose como una extensa osobresaliente paleta, la cara media es la superficie para la recepción de la parte lateralde la escápula. Borde anterior del cleitro localizado justo en la parte lateral delcoracoides y sus bordes no se observan unidos por cartílago. Hay dos supracleitro, selocalizan en el borde postero inferior del cleitro. El coracoides es un hueso largo yachatado, su superficie antero mediana está en contacto con su lado opuesto y hay unpequeño espacio entre estos. En la superficie del coracoides hay uno o dos grandesforámenes. La región dorsal del coracoides se extiende lateralmente el cleitro. Hay seisradiales proximales. El extremo proximal del radial latero anterior tiene un procesoarticular extendido por debajo de la superficie ventral de la escápula. Arriba del cleitrose ubica el supracleitro que es alargado. Este tiene una fosa relativamente medianapara la recepción del proceso espinoso dorsal del cleitro. El extraescapular está librecerca de la parte postero lateral del cráneo.
El hueso pélvico es una estructura delgada, que se ubica paralelo al vientre. Elextremo anterior es despuntado. El proceso isquico se extiende por detrás del bordepostero medial, se observa una apófisis corta y doblada (Figura 6b). En la aleta anal elprimer pterigióforo está asociado con los dos primeros radios anales y es ancho (Figura3b). Radios procurrentes de la aleta caudal 9/9 tanto dorsal como ventralmente.Número total de vértebras: 31-34.
Coloración en vivo. Area dorsal café oscuro, con una banda lateral gris oscura. Areaventral blanco plateada. Aletas pectorales café-amarillenta, caudal y dorsal rosado-claro, pélvicas y anal hialinas. Una banda lateral café-amarillenta a nivel delpedúnculo lateral.
Distribución. Sabana de Bogotá, cuenca del río Magdalena, Colombia.
Aspectos bioecológicos. De esta especie se conoce relativamente bien su ecología

67Mem. Fund. La Salle de Cien. Nat. 155
trófica y reproductiva. Grundulus bogotensis se reproduce entre enero-abril yseptiembre-noviembre, de fecundidad alta. Su dieta la conformaron Calanoides,amfípodos, cladóceros así como larvas de Chironomidae, pupas de Diptera y el caracolPisidium (Mora et al. 1992). Se estableció su número cromosómico básico como 2n=50(25 pares) y se encontró que no existe dimorfismo sexual cromosómico en esta especie(González et al. 1992). Los siguientes datos y observaciones diurnas, fueron obtenidosen agosto 9 del 2000 en la Represa Neusa, vereda Llano Grande, municipio de Tause,Cundinamarca: temperatura superficial, 14,5 °C; temperatura del aire, 12 °C; oxígenodisuelto, 6,5 ppm con 71% de saturación; pH alrededor de 7; sustrato confor-mado pordetritos a una profundidad promedio de 0,3 m. Los ejemplares se colectaron en orillade la represa, con abundantes macrófitas.
Comentarios. Ctenocharax es un género monotípico considerado por Eigenmann(1915: 17) y en este trabajo como un sinónimo de Grundulus. La especie tipo deCtenocharax, C. bogotensis Regan (1907) es un homónimo junior de la especie tipo deGrundulus, Poecilia bogotensis de Humboldt y Valenciennes (1833).
Parte del material examinado por Alexander von Humbold se encuentra en elMuseum fuer Naturkunde der Humboldt-Universitaet zu Berlin, Institut fuerSystematische Zoologie (ZMB) que fue donado por él en agosto 15 de 1815 pordisposición de A. Valenciennes, Paris. Este material estuvo en las manos deValenciennes así como de A. v. Humboldt. En la actualidad no existen indicaciones oreferencias acerca de los ejemplares tipo de este taxon en algún otro museo del mundo.Sobre G. bogotensis existen nueve ejemplares en ZMB. Todo el material fue enviadopor Valenciennes. No hay designación del holotipo en la descripción original. Aunqueaparentemente Humboldt y Valenciennes (1833) no seleccionaron o indicaron si este
Figura 6. Cintura pélvica de Grundulus cochae (a) y G. bogotensis (b). Escala 1mm.

68 Una especie nueva de Grundulus (Teleostei: Ostariophysi: Characidae)
es el material tipo de G. bogotensisNo se descarta la posibilidad de que los peces depositados en ZMB sea parte del
material tipo , ya que Humboldt anota en su descripción que el tuvo un gran númerode ejemplares disponibles.
Probablemente el material original se perdió o no se aseguró durante laexpedición, y posiblemente el material citado y examinado en ZMB fue colectado ydonado mas tarde por el profesor de Física Lewy en la misma localidad. Es muyprobable que Valenciennes nunca vió un ejemplar de G. bogotensis antes de que losnueve peces fueran donados por Humboldt al museo de Zoología de Berlin en 1851(Peter Bartsch 2001 com. pers.).
En conexión con la redescripción de la especie G. bogotensis, es necesaria ladesignación de un neotipo, por que el género monotípico Grundulus incluye ahora dosespecies (las cuales no son fáciles de distinguir). El neotipo fue colectado en la localidadtípica de G. bogotensis y se separó de una muestra confirmada de los autoresHumboldt y Valenciennes como pertenecientes a la especie G. bogotensis.
Biogeografía. Geología histórica y paleogeografía (Hoffstetter 1971, Lundberg et al.1986, 1988, Duque-Caro 1990, Lundberg y Chernoff 1992, Hoorn 1994, Diaz deGamero 1996, Lundberg y Marshall 1997, Lundberg 1997, 1998, Monsch 1998,Gregory-Wodzicki 2000) demostraron la conexión histórica entre las cuencas del nortede Suramérica. En especial la cuenca de los ríos Magdalena y Amazonas. El hallazgode un nuevo taxon en la parte Cisandina de Colombia, en evidente alopatría con laespecie descrita por Humboldt y Valenciennes (1833) localizada en la partetransandina, confirmó la hipótesis actual del protoamazonas y protomagdalena, en elsentido de que antes del levantamiento de los Andes (en este caso la cordillera centralde Colombia) el género Grundulus podría registrar una distribución más amplia ycontinua en los drenajes mencionados. Un caso similar se registró dentro del géneroCallichthys tradicionalmente con distribución en el oriente de Suramérica, se halla enla pendiente occidental de los Andes (Román-Valencia et al. 1999). Lo anteriorcoincide con lo anotado por Lynch (1999) en el sentido de que la alopatría es elprincipal factor de especiación.
Es muy probable que la distribución disyunta para las especies actuales deGrundulus: G. bogotensis para la sabana de Bogotá en la cuenca del río Magdalena yde G. cochae para la Laguna de La Cocha en la cuenca del Amazonas, se debió aprocesos de extinción que ocurrieron en las partes bajas de la cuenca del Magdalena,ligado al levantamiento de la cordillera oriental (~15 ma). Este es un caso de un eventovicariante para el endemismo de las especies de Grundulus.
Los levantamientos orogénicos ocasionaron la aparición de ambientes nuevos conclimas isomesotérmicos, isomicrotérmicos e isooligotérmicos (inclusive de nievesperpetuas), que ofrecieron hábitats “vacíos” para ser ocupados por la biota de tierrasbajas que había venido diversificándose progresivamente hacia los climas de altamontaña (Hernández-Camacho et al. 1992). Sin embargo, Lynch (1999) afirma que los

69Mem. Fund. La Salle de Cien. Nat. 155
grupos de anfibios en tierras bajas no tienen ningún pariente en los Andes y la mayoríade los grupos andinos no posee ninguna relación de parentesco en las tierras bajas.Sugiere, además, que los eventos iniciales de especiación fueron pre-andinos, tal vezapareciendo en las colinas o en los arcos de islas y que los eventos posteriorescorrespondieron al surgimiento de las cadenas montañosas durante las orogeniasandinas. Ambas especies de Grundulus se ubicaron a más de 3000 m de altura sobreel nivel del mar, lo cual podría sugerir que estos hábitats constituyeron refugiospleistocénicos.
Material Examinado
Grundulus bogotensis: ZMB 3505, seis ejemplares (sintipos): Colombia,Cundinamarca, Sabana de Bogotá, 1815, Lewy. ZMB 33306, un ejemplar(neotipo): Colombia, Cundinamarca, Sabana de Bogotá, 1815, Lewy. ZMB31499, dos ejemplares (sintipos): Colombia, Cundinamarca, Sabana deBogotá, 1815, Lewy. IUQ 456, 40 ejemplares: Colombia, Cundinamarca,Teusa, vereda. Llano Grande, Represa de Neusa (5°09’51”N-73°57’33”O)3027 m s.n.m. ICNMNH 364, nueve ejemplares: Sabana de Bogotá, Lagunade Hernández. ICNMNH 563, dos ejemplares: Sabana de Bogotá, ríoTunjuelito; ICNMNH 586, nueve ejemplares: cuenca río Magdalena, represade Neusa. ICNMNH 672, ocho ejemplares: cuenca río Magdalena, represa deNeusa. ICNMNH 1815, 15 ejemplares: cuenca río Magdalena, represa deNeusa. ICNMNH 672, 18 ejemplares: sistema río Bogotá, río Madrid.ICNMNH 687, siete ejemplares: sistema del río Suarez, laguna de Fúquene.ICNMNH 4922, un ejemplar: sistema río Suárez, laguna de Fúquene. IAvH2169, 20 ejemplares: Represa de Neusa. IAvH 1660, 54 ejemplares: Represade Neusa.
Agradecimientos. El Instituto Smithsonian de Investigaciones Tropicales, Panamá (STRI) através de Eldredge Bermingham financiaron el trabajo de campo. IDEA WILD por la donaciónde equipo necesario para la toma de datos de campo y laboratorio, Lara Gasser y Walter D. VanSickle III hicieron posible esta ayuda. Luiz R. Malabarba (MCP) y un revisor anónimo leyeronel manuscrito y realizaron valiosas correcciones y sugerencias. Stanley H. Weitzman (USNM)amablemente suministró bibliografía. Los autores están agradecidos con Peter Bartsch (ZMB)por su ayuda con información útil sobre los ejemplares de Grundulus bogotensis de la colección,su historia y por el suministro de algunas placas de raxos-x para análisis. Germán Galvis e IvánMojica (ICNMNH) y Yaneth Muñoz-Saba (IAvH) facilitaron en préstamo material de G.bogotensis. Carlos Pantoja y Antonio Navarro (IUQ) colaboraron durante el trabajo de campo.Raquel I. Ruiz (IUQ) elaboró con admirable paciencia y entusiasmo de sardina las figuras 3 a6.

70 Una especie nueva de Grundulus (Teleostei: Ostariophysi: Characidae)
Bibliografía.
CALA, P. Y C. ROMÁN-VALENCIA. 1994. Lista y distribución geográfica de las especies de lafamilia Characidae (Pisces, Characiformes) en Colombia. Biología y Educación 4: 15-23.
CUVIER, M. LE B. Y M. A. VALENCIENNES. 1846. Histoiré naturelle des poissons. Suite du livredix huitiéme Cyprinoides. Livre dix neuviéme. Des Esoces ou licioides 18: 216-220.
DAHL, G. 1971. Los peces del norte de Colombia. Inderena, Bogotá. 391 pp.
DÍAZ DE GAMERO, M. L. 1996. The changing course of the Orinoco River during the Neogene:a review. Palaeogeography Palaeoclimatology 123: 385-402.
DUQUE-CARO, H. 1990. Neogene stratigraphy, paleoceanography and paleobiogeography innorthwest South America and evolution of the Panama Seway. PalaeogeographyPaleoclimatology 77: 203-234.
EIGENMANN, C. H. 1915. The Cheirodontinae, a subfamily of minute characid fishes of SouthAmerica. Memoirs Carnegie Museum 7: 1-99.
EIGENMANN, C. H. 1922. The fishes of the Northwestern South America including Colombia,Panama and Pacific slopes of Ecuador and Peru, together with appendix upon the fishes ofthe rio Meta in Colombia. Memoirs Carnegie Museum 9: 1-348.
GÉRY, J. 1977. Characoids of the world. T.F.H. Publ. Neptune City. 672 pp.
GONZÁLEZ A. J., M. L. BUENO Y J. E. FORERO. 1992. Caracterización cromosómica de dosespecies ícticas nativas; Guapucha, (Grundulus bogotensis) y Capitán, (Eremophilus mutisii),de la sabana de Bogotá. Acta Biológica Colombiana 7-8: 45-54.
GREGORY-WODZICKI, M. K. 2000. Uplift history of the Central and Northern Andes: A review.GSA Bulletin 112: 1091-1105.
HELFMAN, G. S., B. B. COLLETTE Y D. E. FACEY. 1997. The Diversity of Fishes. BlackwellScience, Malden, Massachussets.
HERNÁNDEZ-CAMACHO, A. HURTADO GUERRA, R. ORTIZ QUIJANO Y T. WALSCHBURGER. 1992.Centros de endemismo en Colombia. Pp. 175-190. En: Haffter, G. (Comp.). DiversidadBiológica de Iberoamérica I. Acta Zoología Mejicana (Volumen especial).
HOFFSTETTER, R. 1971. Los vertebrados cenozoicos de Colombia: yacimientos, faunas,problemas planteados. Geología Colombiana 8: 37-62.
HOORN, C. 1994. An environmental reconstruction of the palaeo-Amazon River system(Middle-Late Miocene, NW Amazonia). Palaeogeography Palaeoclimatology 112: 187-238.
HYSLOP, E. J. 1980. Stomach contents analysis. A review of methods and theirs application.Journal Fisheries Biology 17: 411-429.
HUBBS C. L. Y K. F. LAGLER. 1958. Fishes of the Great Lake Region. Revised edition, CranbookInstitute of Science, Bulletin 26, Bloomfield Hills, Michigan.
HUMBOLDT, F. H. A. VON Y A. VALENCIENNES. 1833. Reserches sur les poissons fluviatiles del’xmerique equinoxiale En: Humboldt F.H. A. et A. Bonpland, d’ observations de zoologie etd’anatomie Comparèe, Paris. 2: 154-156, 159 pl 45, fig. 1.
LUNDBERG, J. G. 1997. Freshwater Fishes and their Paleobiotic Implications. Pp. 67-91. En: R.F. Kay, R. H. Madden, R. L. Cifelli y J. J. Flynn (Ed.) Vertebrate Paleontology in theNeotropics The Miocene Fauna of La Venta, Colombia. Smithsonian Institution Press,Washington y London.
LUNDBERG, J. G. 1998. The Temporal Context for the Diversification of Neotropical Fishes. Pp.49-68. En: L. R Malabarba, R. E. Reis, R. P. Vari, Z. M. S de Lucena y C. A. S. Lucena (ed.).Phylogeny and clasification of neotropical fishes. Edipucrs, Porto Alegre, Brazil.
LUNDBERG, J. G. Y B. CHERNOFF. 1992. A Miocene Fossil of the Amazonian Fish Arapaima(Teleostei, Arapaimidae) from the Magdalena River region of Colombia- Biogeographic and

71Mem. Fund. La Salle de Cien. Nat. 155
Evolutionary implications. Biotropica 24: 2-14.
LUNDBERG J. G. Y L. G. MARSHALL. 1997. A history of tropical South American rivers: 89 mato recent. International Symposium on Phylogeny and Classification of Neotropical Fishesand Neotropical Ichthyological Association 9 th Annual Meeting, Porto Alegre-RS- Brasil p.80 (Resumen).
LUNDBERG, J. G. A. MACHADO-ALLISON Y F. KAY. 1986. Miocene characid fishes fromColombia: evolutionary stasis and extirpation. Science 234: 208-209.
LUNDBERG J. G., O. J. LINARES, M. E. ANTONIO Y P. NASS. 1988. Phractocephalus hemiliopterus(Pimelodidae, Siluriformes) from the upper miocene Urumaco formation, Venezuela: afurther case of evolutionary stasis and local extinction among south american fishes. JournalVertebrate Paleontology 8: 131-138.
LYNCH, J. 1999. Ranas pequeñas, la geometría de evolución y la especiación en los AndesColombianos. Revista Academia Colombiana Ciencias 23: 143-159.
MALABARBA, L. R. 1998. Monophyly of the Cheirodontinae: Characters and major clades(Teleostei: Characidae). Pp. 193-233. En: L. R Malabarba, R. E. Reis, R. P. Vari, Z. M. S deLucena y C. A. S. Lucena (Ed.). Phylogeny and clasification of neotropical fishes. Edipucrs,Porto Alegre, Brazil.
MILES, C. 1971. Los peces del río Magdalena. U. T. Ibagué, Edic. (Reimpresión), 214 pp.
MORA G., L. S. TÉLLEZ, P. CALA Y G. GUILLOT. 1992. Estudio ecológico de la ictiofauna del lagode Tota (Boyacá-Colombia), con énfasis en la Trucha Arco iris, Oncorhynchus mykiss. RevAcademia Colombiana Ciencias 18: 409-422.
MONSCH, A. K. 1998. Miocene fish faunas from the northwestern Amazonia basin (Colombia,Peru, Brazil) with evidence of marine incursions. Palaeogeography Palaeoclimatology 143(1998): 31-50.
REGAN, C. T. 1907. Description of two new characinid fishes South America. Annales MagazineNatural History, ser. 7, 20 (119): 402-403.
Román-Valencia, C. 2000a. Tres nuevas especies de Bryconamericus (Ostariophysi:Characidae) de Colombia y diagnóstico del género. Revista Biología Tropical 48 (2/3): 449-464.
ROMÁN-VALENCIA, C. 2000b. Revisión sistemática de las especies del género Bryconamericus(Teleostei: Characidae) para Centroamérica y el Norte de Suramérica. Tesis DoctoralUniversidad del Valle, Facultad de Ciencias, Departamento de Biología, Cali, Colombia. 182pp.
ROMÁN-VALENCIA C., P. LEHMANN Y A. MUÑOZ. 1999. Presencia del género Callichthys(Siluriformes: Callichthyidae) en Colombia y descripción de una nueva especie para el altoRío Cauca. Dahlia 3: 53-62.
SONG, J. Y L. R. PARENTI. 1995. Clearing and staining whole fish specimens for simultaneousdemostration of bone, cartilage and nerves. Copeia 1995: 114-118.
TAYLOR, W. R. Y G. C. VAN DYKE. 1985. Revised procedures for staining and clearing smallfishes and other vertebrates for bone and cartilage study. Cybium 9: 107-119.
VARI R. P. 1995. The neotropical fish family Ctenoluciidae (Teleostei: Ostariophysi:Characiformes) supra and intrafamilial phylogenetic relationships, with a revisionary study.Smithsonian Contribution Zoology 564: 1-96.
WEITZMAN, S. H. 1962. The osteology of Brycon meeki, a generalized characid fish, with anosteological definition of the family. Stanford Icthyological Bulletin 8: 1-77.
WEITZMAN, S. H. Y L. R. MALABARBA. 1999. Systematics of Spintherobolus (Teleostei:Characidae: Cheirodontinae) from eastern Brazil. Ichthyological Exploration Freshwaters 10:1-43.

72 Una especie nueva de Grundulus (Teleostei: Ostariophysi: Characidae)
Recibido: 19/09/2001Aprobado: 15/05/2002
César Román-Valencia1, Hans-Joachim Paepke2, y Fernando Pantoja3
1 Universidad del Quindío, Departamento de Biología, A. A. 460, Armenia, Quindío, Colombia.Correo electrónico: [email protected].
2 Museum fuer Naturkunde, der Humboldt-Universitaet zu Berlin, Institut fuer SystematischeZoologie, Invalidenstr. 43, D 10099, Berlin, Alemania.
3 Universidad del Quindío, Departamento de Biología, Armenia, Quindío, Colombia. Correoelectrónico: [email protected].


















