
Transferencia de Embriones en El Ganado Bovino
67
Manual de Transferencia de Embriones en el Ganado B Octubre de 2006
-
Upload
cristian-exequiel-carbajal -
Category
Documents
-
view
166 -
download
5
Transcript of Transferencia de Embriones en El Ganado Bovino

Manual de Transferencia de Embriones en el Ganado B
Octubre de 2006

Gobierno del Estado de Chiapas
Lic. Pablo Salazar Mendiguchia Gobernador Constitucional del Estado
Ing. Javier Antonio Ruiz Morales Secretario de Desarrollo Rural
Dr. Fidelfo Rodriguez Fernandez Subsecretario de Ganaderia
MVZ. Rafael Gonzalez Castellanos Director de Fomento Ganadero
MVZ. Oscar Moguel Delarbre Jefe de Departamento de Vinculaci6n
Ing. Napoleon Orantes Solorzano Coordinador General D.P.A.I.
Secretaria de Agricultura, Ganaderia, Desarrollo Rural, Pesca y Alimentacion
MVZ. Jose Angel Del Valle Molina Delegado Estatal
Ing. Israel de Jes6s G6mez Torres Subdelegado Agropecuario
Ing. Carlos Antonio Toledo Culebro Jefe de Fomento Pecuario
Fundacidn Produce Chiapas A.C.
Sr. Hipolito Pedrero Alegria Presidente Ejecutivo
Dr. Jorge Luis Zuarth Macias Secretario
C.P. Araceli Ramirez Martinez Tesorera

Universidad Aut6noma de Chiapas
MC. Jorge Ordofiez Ruiz Rector
Dr. Hugo GuillCn Trujillo Secretario General
Dr. Carlos Eugenio Ruiz Hernandez Secretario Academico
Dr. Jos6 Alfredo Medina Melendez Director General de Extension
MVZ. Adizain Alegria C6rdova Director de la FMVZ
MVZ. Alfredo Castellanos Coutifio Secretario Acaddmico
MC. Roberto R. Coutifio Ruiz Director de la Facultad de Ciencias Agron6micas
Dr. Ramiro Ruiz Najera Secretario Academico
MVZ. EPA Oscar Leon Velasco Coordinador General de Vinculacibn
y Extensibn Universitaria
Coordinadores y Responsables del Curso Cuerpo AcadCmico
"BIOTECNOLOG~A Y MEJORAMIENTO GENETICO ANIMAL"
Dr. Horacio Lebn Velasco MC. Horacio Ruiz Hernandez MVZ. Alfonso Ruiz Moreno
MVZ. Alfonso Villalobos Enciso
Colaboradores en la elaboraci6n del material didactic0
MVZ. Oscar Miguel Dominguez Galdimez Juan Luis Orantes Arrazate
William Esponda Hernandez

I.. INTRODUCCION .................................................................................... 1
........... 11.. ANTECEDENTES DE LA TRANSFERENCIA DE EMBRIONES 3 2.1 .. Fisiologia de la reproduccion ....................................................... 6 2.2.. Mecanismo de selection ......................................................... 13 2.3.. Agentes superovulatorios .................................................... 16 2.4.. Metodos de sincronizacion de estros ......................................... 17 2.5.- Tecnicas de Superovulacion .................................................... 21 2.6.- Deteccion de Estro .................................................................... 24
............ 2.7.. Insemination de las vacas donadoras y superovuladas 24
Ill.. MEDIOS PARA LA COLECCION DE EMBRIONES .............................. 26 3.1 .. Congelacion ............................................................................... 27 3.1 . 1 .- Molaridad .................................................................................. 28 3.1.2.- Funcion de 10s crioprotectores ................................................. 29 3.1.3.- Velocidad de enfriamiento ....................................................... 30
......................................... 3.1.4.- Clasificacion de 10s crioprotectores 31 .................................................................. 3.1.5.- Funcion del glicerol 33
............................................................ 3.1.6.- Funcion del etilenglicol 33 ................................... 3.1.7.- Congelacion de embriones con glicerol 34
.............................. 3.1 .8.- Congelacion de embriones con etilenglicol 36 .... ..................... 3.1.9.- Descongelacion de embriones con glicerol .. 38
...................... 3.1.10.- Descongelacion de embriones con etilenglicol 41 .......................................................... 3.1 . 11 .- Preparacion de medios 42
IV.. COLECCION DE EMBRIONES .......................................................... 43 .............................................................. 4.1 .. Evaluacion Embrionaria 44
4.1 . 1 .. Embriones transferibles ........................................................... 45 ............................................................ 4.1.2.. Lavado de 10s embriones 47
........................................................ 4.1.3.- Transferencia de embriones 47 4.1.4.- Bipartition de embriones ............................................................. 49
......................................................................... 4.1.5.- Micromanipulador 49 ............................................... 4.1.6.- Division artificial de 10s embriones 49
...................................... 4.1.7.- Ventajas de la biparticion de embriones 51 ........................................................................................ 4.1 .8.- Mellizos 52
.......................................................... 4.1.9.- Mecanismo de gemelacion 52 .................................. 4.1.10.- Tasas de gemelacion en ganado lechero 53
4.1 . 11 .- Freemartinismo .......................................................................... 54
V.. LITERATURA CITADA .............................................................................. 57

La reproduccion en 10s animales domesticos se ha incrementado su desarrollo gracias
a la manipulacion de biotecnologias como la inseminacion artificial (IA) y la
transferencia de embriones (TE). La eficiencia de estos procedimientos y de la
posibilidad de mantener viables las celulas germinativas desde el momento de la
coleccion hasta su aplicacion, desde el punto de vista cientifico cobra gran
importancia. En este sentido, la congelacion de estas celulas ofrece una solucion
practica para el almacenamiento prolongado de las mismas sin perder su viabilidad.
La criobiologia tiene como objetivo el estudio de las causas de dafios celular durante la
congelacion y descongelacion, asi como la implementacion de metodos disefiados
para reducir o evitar ese dafio; de tal forma, que las celulas Sean capaces de continuar
en forma normal su desarrollo completo despues de la descongelacion.
La transferencia de embriones es una herramienta para el mejoramiento genetico del
ganado y tiene como objetivo incrementar la tasa reproductiva de las hembras de alto
valor genetico. La transferencia de embriones consiste en inducir un embrion en etapa
de preimplantacion en el utero de la hembra denominada receptora, la cual se
encargara de gestarlo y llevarlo al nacimiento. El embrion transferido puede ser fresco
o congelado. Esta tecnica consiste en un tratamiento hormonal que se aplica a las
hembras donadoras para inducir la maduracion y ovulation de un gran numero de
ovulos (superovulacion).
La congelacion de celulas embrionarias tiene por objeto interrumpir el metabolismo de
forma reversible permitiendo de ese modo, su conservation durante un tiempo
indefinido en estado solido, manteniendose la viabilidad del embrion despues de la
descongelacion. La supervivencia depende esencialmente de las condiciones de
cambio de fase del agua celular y de la eficacia de 10s crioprotectores.
Con el uso de crioprotectores como el etilenglicol y el glicerol se podran definir nuevas
estrategias, tanto en la congelacion de embriones como en el programa de
transferencia de embriones en el ganado bovino.

OBJETIVOS.
1 .- Actualizar y capacitar a 10s profesionales y estudiantes de la Medicina Veterinaria en
Biotecnologia Reproductiva especificamente en transferencia de embriones en el
ganado bovino como una herramienta para el mejoramiento genetic0 de la ganaderia.
2.-Promover la transferencia de tecnologia en materia de transferencia de embriones a
10s productores pecuarios y vincular la Universidad Autonoma de Chiapas con el
subsector agropecuario del pais.

11.-ANTECEDENTES DE LA TRANSFERENCIA DE EMBRIONES
La historia de la transferencia de embriones se remonta a 1890 cuando el ingles Walter Heape, realizo con exito la primera transferencia embrionaria en conejos. A partir de entonces se han informado transferencias embrionarias exitosas en todo tip0 de animales de granja. La comercializacion de tecnologia para la transferencia embrionaria
comenzo en America del Norte.
Una hembra (donadora) puede aumentar su numero de crias durante su vida al concebir de manera repetida y mediante la recuperacion de embriones al inicio de la gestacion y
la transferencia de estos a 10s aparatos reproductores de otras hembras (receptoras) para completar la gestacion. Este procedimiento depende por completo de la disponibilidad de una fuente de embriones de calidad adecuada y el medio uterino propicio de la receptora al momento de la transferencia embrionaria (sincronia) (Hafez, 2000). El principal objetivo del transplante de embriones es incrementar la tasa reproductiva de una hembra de excelentes caracteristicas, fecundada con un tor0 de
alta genetica (Aspron, 1992).
La transferencia embrionaria se ha aplicado muy extensamente en la vaca; en
consecuencia, la tecnologia ha avanzado con mas rapidez en esta especie. En 10s arios
60, la anestesia general y la laparotomia eran necesarias para la coleccion y
transferencia de embriones bovinos, y en el Reino Unido y America del Norte esta tecnica se utilizo sobre todo para la rapida multiplication razas exoticas de carne importadas (Palomino eta/., 1998).
La utilization de embriones junto a la inseminacion artificial en 10s programas de mejora
genetica es una practica que ha venido difundiendose en forma creciente en 10s ultimos
20 aiios. Sin embargo, esta tecnologia que tiene un enorme potencial de mejora
genetica, no ha tenido la difusion masiva que debiera en funcion de tal potencialidad.
Las limitantes han sido tanto 10s altos costos de la misma, como la enorme variabilidad
en la respuesta de 10s animales destinados a producir tales embriones. Lo anterior
parece marcar limites economicos y biologicos en la tecnologia que no han podido ser
superados en 10s ultimos 20 atios y que limitan su desarrollo.
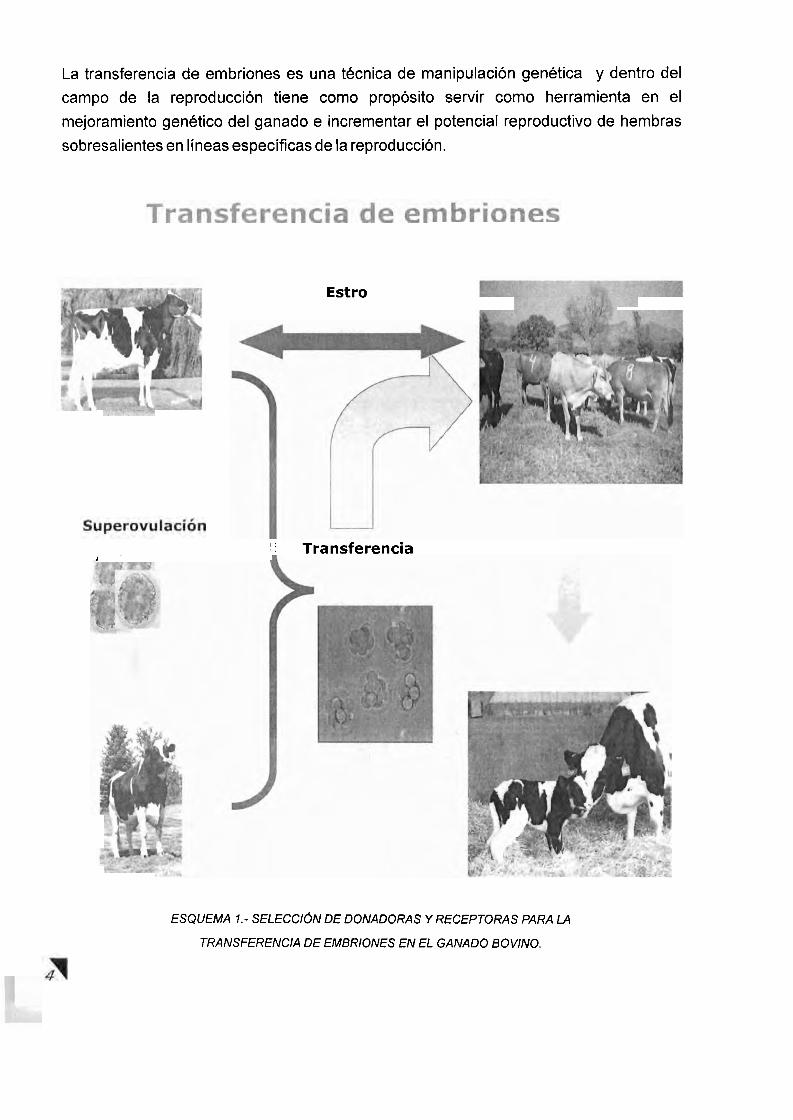
La transferencia de embriones es una tecnica de rnanipulacion genetica y dentro del
campo de la reproduccion tiene como proposito servir como herramienta en el
mejoramiento genetic0 del ganado e incrementar el potencial reproductivo de hembras
sobresalientes en lineas especificas de la reproduccion.
Estro
I 1 1 Transferencia
ESQUEMA 1.- SELECCI~N DE DONADORAS Y RECEPTORAS PARA LA
TRANSFERENCIA DE EMBRIONES EN EL GANADO BOVINO.

El utilizar embriones congelados permite aprovechar al maximo el potencial de una hembra; en virtud, que es posible congelar todos 10s embriones de excelente y buena calidad, ser transportados a cualquier pace del mundo en forma facil y economica. Por esto, la TE ha tenido un impact0 considerable sobre 10s programas de reproduccion en paises desarrollados desde 10s aAos 70's, esta influencia ha sido principalmente dentro del sector de razas puras con el objeto de aumentar el potencial reproductivo de hembras superiores y donde el alto valor comercial de sus crias ha justificado el gasto involucrado.
Existen tres metodos de coleccion de embriones, dos quirurgicos y uno no quirurgico, el
primer0 requiere de medicos, anestesistas y cirujanos, lo cual hace de esta tecnica algo
impractico quedando en desuso. El segundo metodo consiste en una laparotomia por uno
de 10s flancos usando anestesia regional y local. El tercer metodo es el no quirljrgico o
transcervical, que actualmente es el mas usado. Con el desarrollo del metodo de coleccion de embriones no quirurgico, existe la posibilidad de la dispersion de esta tecnologia dentro
de paises desarrollados. Una de las principales razones para el uso de la TE es el mejoramiento genetico y se puede superar algunas formas de infertilidad que puede surgir
a consecuencia de fallas de fertilization debido a altas temperaturas. Este proceso puede
ser usado para establecer bancos de embriones congelados y permitir la importancia de
10s mismos y por lo tanto la introduccion y transferencia del nuevo material genetico.
LAVADO DE VACAS DONADORAS PARA LA TRANSFERENCIA DE EMBRIONES (METODO NO QUIRURGICO 0
TRANSCERVICAL).

Ciclo estral del ganado bovino.
El ciclo estral es una serie de eventos hormonales que ocurren desde un period0 de estro
hasta la manifestacion del siguiente. El estro es comunmente referido con el dia cero en el ciclo estral de la vaca, momento en el cual la vaca es sexualmente receptiva al toro. El ciclo
estral en el ganado bovino tiene una duracion aproximada de 21 dias, con un interval0 de 17 a 24 dias para que sea considerado como normal. En el caso de las vaquillas el ciclo
estral dura menos que en las vacas.
El ciclo estral puede ser dividido en dos fases: la primera, llamada fase folicular que se
caracteriza por que durante este tiempo ocurren las fases finales de desarrollo folicular.
Esta fase es relativamente corta y dura aproximadamente de 4 a 5 dias, iniciandose con la
regresion del cuerpo luteo (CL) y culminando con la ovulacion. La segunda o fase Iutea, es
caracterizada por el desarrollo del CLy tiene una duracion de 16 a 17 dias.
Los cambios anatomicos y fisiologicos que sufren 10s organos reproductivos durante estas
fases son regulados por hormonas entre las que se encuentran.
a) GnRH. Hormona liberadora de gonadotropinas que es producida a nivel
hipotalamico.
b) LH y FSH. Hormona luteinizante y foliculo estimulante respectivamente sintetizadas a nivel hipofisiario.
c) Inhibina, estrogenos y progesterona que son producidos por el ovario. d) PGF, . Prostaglandina F, alfa-sintetizados en el endometrio.
e) IGF-I. Factor de crecimiento parecido a la insulina numero 1 sintetizado en el higado.
La duracion del estro en el ganado cebu es alrededor de 10 horas (h) per0 tiene un amplio
rango de variacion que va de 2.2 a 18.4 h. Es conveniente mencionar que la duracion del ciclo estral puede verse alterada debido a ciertos factores como son; 10s cuerpos luteos de
vida corta que son precedidos por foliculos deficientes en su desarrollo, asi como a las fallas que ocurren por muerte embrionaria temprana.
Caracteristicas de la conducta estral
Ademas de 10s tratamientos y tecnicas utilizados en la sincronizacion de estros, es
importante considerar tambien que existen problemas para detectar con precision la

manifestacion de conducta estral, debido a la composicion, tamafio y orden social que se
tenga en un grupo de hembras bovinas.
Se ha observado que existe una relacion entre conducta estral y jerarquia en las hembras, cuando se encuentran en estro dentro de un grupo. Ademas del nljmero de estas que presentan estro al mismo tiempo. En efecto, una o varias hembras pueden presentar una conducta de dominancia sexual ante el grupo; es decir, presentar comportamiento activo, que consiste en conductas estrales hacia otras hembras.
En un hato con vacas Indubrasil, se observo que el 60% de montas fueron realizadas por hembras que presentaban mayor jerarquia; sin embargo, aunque en menor proporcion, las hembras dominantes pueden ser montadas por hembras consideradas como
inferiores dentro del orden social.
De acuerdo a investigaciones realizadas, se ha observado que la vaca que monta,
generalmente domina a la que es montada, encontrando que se pueden presentar 5.3
montas por una hora en vacas que se encuentran en pastoreo; en contraste, con 8.7
montas en promedio que presentaron hembras que se encontraban en condiciones de estabulacion. Esto puede ser un efecto de que al tener una alteracion de las condiciones ambientales en hembras que se encuentran en pastoreo, estas pueden suprimir la manifestacion de conducta estral.
Asimismo, se ha observado que las hembras con una mayor dominancia dentro del grupo,
inhiben el comportamiento sexual de sus compaAeras, dando como resultado que las
primeras presenten mayor indice de comportamiento estral y social; ademas de
interactuar con sus compafieras que son racialmente semejantes.
Las variaciones que se pueden presentar en el comportamiento social y jerarquico,
dependen de la hora en que se aplique el tratamiento para sincronizar, la hora en que se
manifieste la mayor actividad sexual y de la observacion de estro. Ademas, existen efectos que controlan la forrnacion de grupos homosexuales, en donde una hembra podria dejarse montar por una de sus compafieras y no por la presencia de un macho.
Otra variacion que se puede presentar en ganado Cebll cuando se encuentra en etapa estral, es un comportamiento de agresividad en algunas vacas, manifestando con topeteo, el cual puede ser de tipo no agresivo con la presentacion de caracteristicas como

bramido, lamer los genitales, roce corporal u oler otras regiones corporales de su compaAera que en ese momento tambien presenta estro. Ademas, las hembras tienden a
presentar signos como inflamacion de vulva, hiperemia del vestibulo vaginal, eliminacion de moco filamentoso y arborizacion del moco cervical.
Por otra parte, algunos investigadores tambien describen un efecto de bioestimulacion,
dada por feromonas que se encuentran en el moco cervico-vaginal de las hembras que se
encuentran en estro y que pueden estimular a otras hembras anestricas. Esto se corrobora
con un posterior estudio realizado por Wright et a/. (1992) quienes demuestran que al
manifestarse estro un grupo de hembras sincronizadas, estas pueden estimular la
actividad ovarica de hembras no tratadas, acortando la duracion de anestro posparto.
Efecto ambiental en la conducta de estro.
Se ha observado que se presenta un alto indice de estros en hembras lndubrasil durante el
transcurso de horas oscuridad, que en las primeras horas de luz. Sin embargo, la presencia de comportamiento estral en Bos taurus, es mas frecuente durante las horas de
oscuridad y las primeras horas de luz, entre 1.30 y 7:00 horas.
Por otra parte, las condiciones climatologicas elevadas al tropico, se pueden presentar en
el ciclo estral con mayor regularidad en primavera que en otras estaciones del aAo,
observando que la determinacion de estros, solo puede presentarse alrededor del 30%. Esto se confirma con lo observado por Lozano et a/. (1 987) quienes indican un porcentaje
similar en la deteccion de estros en 10s meses en que se incrementa el fotoperiodo.
Particularmente en el ganado de tip0 Bos indicus y Bos taurus, se ha observado una
actividad reproductiva continua durante todo el aAo; aunque 10s cambios fisiologicos
parecen alterarse, al efecto climatic0 de la estacion y fotoperiodo del aAo. En cambio, Rice
(1 988) menciona que vaquillas de la raza 60s indicus, son mas afectadas por el clima que
el ganado de tip0 Bos taurus. Ciclos estrales cortos

Al reanudar la actividad ciclica despues del parto, 10s animales presentan uno o dos ciclos cortos, 10s cuales contribuyen a problemas de infertilidad durante 30 a 40 dias despues del
parto. Cuando 10s ciclos estrales cortos ocurren, el cuerpo luteo regresa erroneamente antes de que se pueda enviar la seiial de que la gestacion ha comenzado. Esta regresion
prematura del cuerpo luteo, aparentemente se debe a que durante este tiempo se
presentan niveles altos de prostaglandinas F, alfa (PGF, ) producidos por el ljtero como
parte de 10s mecanismos normales de la involuciisn uterina, por lo que la presencia de ciclos cortos y la involution uterina se encuentran intimamente ligados.
Desarrollo folicular durante el ciclo estral.
Dentro de un ovario, 10s foliculos se encuentran en diferentes estadios de desarrollo y han
sido clasificados acorde al nllmero de capas de celulas que rodean al oocito, asi como por la presencia del antro.
La clasificacion mas comljn divide a 10s foliculos en:
a) Foliculo primordial. Estos foliculos se encuentran en gran numero en el ovario antes del nacimiento. En estos foliculos el oocito se encuentra rodeado por u n a
capa de celulas foliculares aplanadas, llamadas tambien celulas de epitelio
folicular escamoso. El diametro de 10s oocitos en la vaca varia entre 79 y 120
micrometros.
b) Foliculo primario. El oocito se encuentra rodeado por una simple capa de celula
cuboidales o poliedricas. Este foliculo, tiene un diametro aproximado de 150 micras.
c) Foliculo secundario. El oocito esta rodeado por varias capas de celulas
poliedricas o cuboidales, momento en el cual, las celulas de la granulosa
desarrollan receptores para FSH y estrogenos. d) Foliculo terciario. El oocito esta rodeado por varias capas de celulas granulosas y
el antro folicular esta formado. En vacas, la formacion antral inicia cuando el foliculo
tiene un diametro aproximado de 0.5 mm.
e) Foliculo de Graff. Este tip0 de foliculo se forma como resultado de la acumulacion
de fluido folicular y hace que se extienda el antro, el oocito se ubica en la periferia
del foliculo y se rodea por una acumulacion de celulas de la granulosa. El foliculo
preovulatorio corresponde a esta clasificacion folicular. Debido a su talla, el foliculo
grafiano maduro, emerge desde el ovario a la superficie en forma de ampula.

Etapas del ciclo estral
Estro. En esta etapa la hembra acepta la monta del macho o de una compariera de hato. Esta
conducta es determinada por un increment0 significativo de las concentraciones de estradiol, que es consecuencia del desarrollo de un foliculo y por la ausencia de un cuerpo
Iuteo. En este momento las concentraciones de progesterona se encuentran en niveles basales ya que tuvo que haber ocurrido la regresion del cuerpo Iuteo.
Durante el estro la hembra esta inquieta e interactua con sus comparieras (conducta homosexual). Por efecto de 10s estrogenos 10s genitales externos se edematizan y hay
produccion de moco cervical. La duracion del estro es variable fluctuando de 8 a 18 h y es afectada por el tip0 de ganado y por las condiciones ambientales. El ganado tipo Bos taurus la conducta estral es muy repetible; sin embargo, factores ambientales tales como
instalaciones con pisos de cement0 provocan que el estro se acorte y sea menos intenso.
En contraste el piso de tierra permite una mayor actividad estral. En el ganado tip0 Bos
indicus es mas variable la conducta y la duracion del estro.
SIGNOS Y CONDUCTA ESTRAL EN EL GANADO BOVINO
Metaestro
Durante del metaestro ocurre la ovulation y se desarrolla el cuerpo luteo pasando por el
estadio intermedio conocido como cuerpo hemorragico, el cual es un estado de transicion
entre el foliculo recien ovulado y el cuerpo Iuteo. En esta etapa las concentraciones de progesterona comienzan a incrementarse hasta que alcanzan concentraciones mayores de 1 nglml, momento a partir del cual se considera que termina el metaestro y comienza el diestro. La duracion del metaestro es de 4 - 5 dias. Un evento hormonal que se destaca en este period0 consiste en la presentacion del segundo pic0 de secrecion de FSH que 1 10
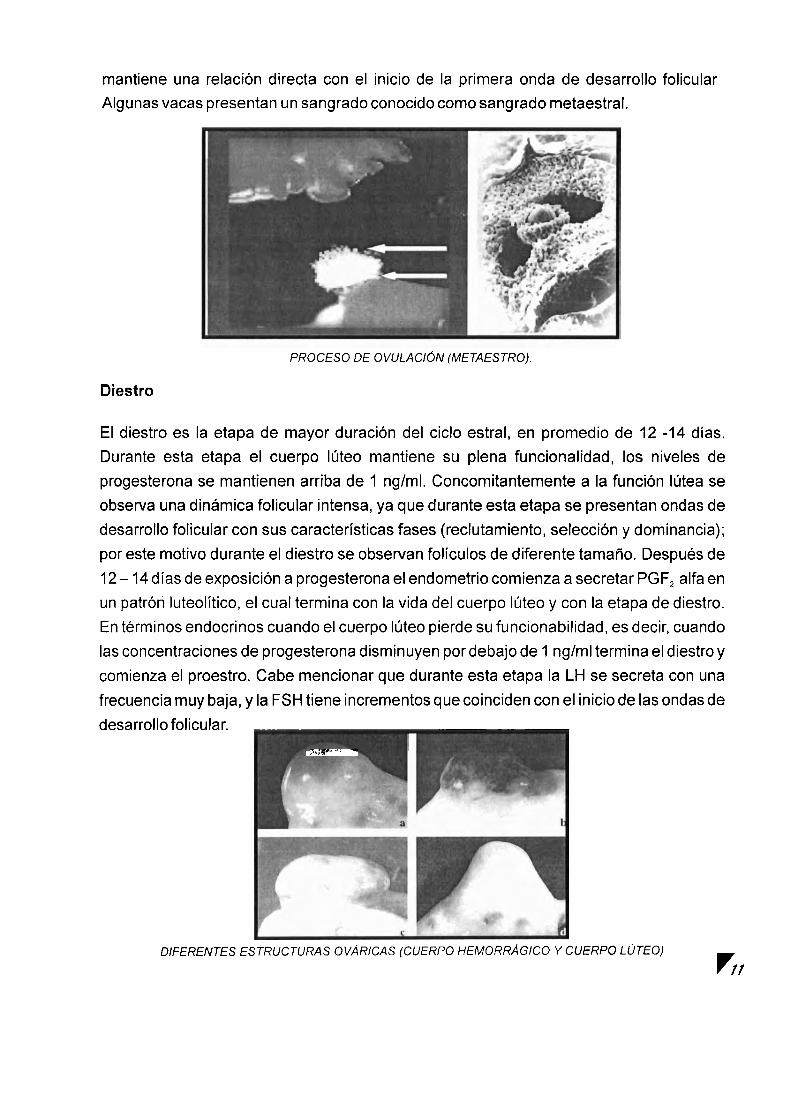
mantiene una relacion directa con el inicio de la primera onda de desarrollo folicular Algunas vacas presentan un sangrado conocido como sangrado metaestral.
PROCESO DE O V U L A C I ~ N (METAESTRO).
Diestro
El diestro es la etapa de mayor duracion del ciclo estral, en promedio de 12 -14 dias.
Durante esta etapa el cuerpo lljteo mantiene su plena funcionalidad, 10s niveles de
progesterona se mantienen arriba de 1 nglml. Concomitantemente a la funcion lljtea se
observa una dinamica folicular intensa, ya que durante esta etapa se presentan ondas de
desarrollo folicular con sus caracteristicas fases (reclutamiento, selection y dominancia);
por este motivo durante el diestro se observan foliculos de diferente tamafio. Despues de
12 - 14 dias de exposicion a progesterona el endometrio comienza a secretar PGF, alfa en
un patrori luteolitico, el cual termina con la vida del cuerpo luteo y con la etapa de diestro.
En terminos endocrinos cuando el cuerpo luteo pierde su funcionabilidad, es decir, cuando
las concentraciones de progesterona disminuyen por debajo de 1 nglml termina el diestro y
comienza el proestro. Cabe mencionar que durante esta etapa la LH se secreta con una
frecuencia muy baja, y la FSH tiene incrementos que coinciden con el inicio de las ondas de
desarrollo folicular. :.,;e". , -
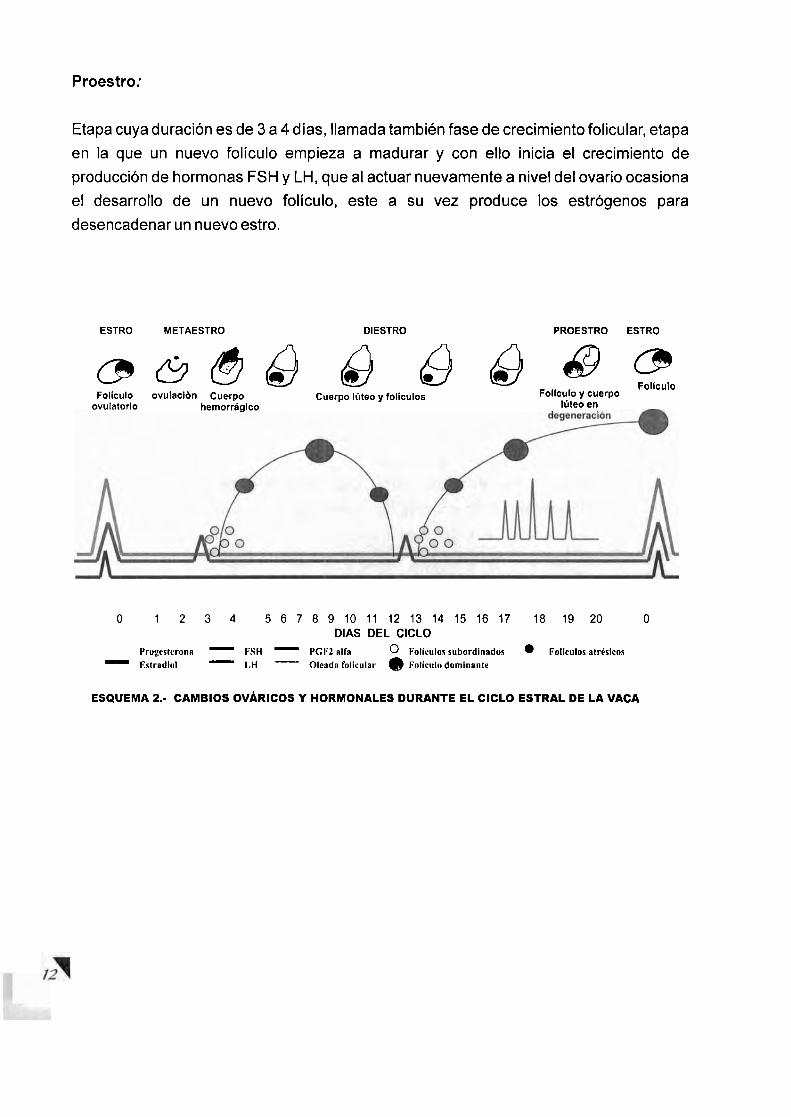
DIFERENTES ESTRUCTURAS OVARICAS (CUERPO HEMORRAGICO Y CUERPO LUTEO)
El
Proestro:
Etapa cuya duracion es de 3 a 4 dias, llamada tambien fase de crecimiento folicular, etapa
en la que un nuevo foliculo empieza a madurar y con ello inicia el crecimiento de
produccion de hormonas FSH y LH, que al actuar nuevamente a nivel del ovario ocasiona
el desarrollo de un nuevo foliculo, este a su vez produce 10s estrogenos para
desencadenar un nuevo estro.
ESTRO METAESTRO DIESTRO PROESTRO ESTRO
0 , 6 & Q Q 8 B @ @ Foliculo
Foliculo ovulaclon Cuwpo Cuerpo luteo y foliculos Foliculo y cuerpo ovulatorio hernorragico luteo en
0 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 0 DlAS DEL ClCLO
Progcstcrona - FSH - PCF2 alfa 0 Foliculos subordinadus Foliculos atr6sicos - Estradiul - 1.H - Olcada folicular a) Foliculo don~inante
ESQUEMA 2.- CAMBlOS OVARICOS Y HORMONALES DURANTE EL ClCLO ESTRAL DE LA VACA

2.2.- SELECCION DONADORAS
Es el principal paso a dar para el inicio del programa de transferencia de embriones y para
esto se utilizan dos criterios intimamente ligados. El primero, el cual desea el productor es
la eficiencia productiva del animal seglln su finalidad zootecnica y ligado a su capacidad genetica; per0 el mas importante es el criterio elegido por el tecnico el cual consiste en que el animal donante provenga de lineas fertiles, tenga un aparato reproductor y ciclo estral
normal, sin antecedentes de partos distocicos y retenciones placentarias, no mas de dos servicios por concepcion y tener una edad de 3 a 10 atios (Elsden, 1980).
En el aspect0 sanitario deberan de estar libres de enfermedades, especialmente aquellas
que afectan la funcion reproductiva; segun la region deberan estar sometidas a un estricto
programa de vacunacion y desparasitacion para garantizar un buen estado de salud.
En el caso de las receptoras, practicamente seguimos 10s mismos puntos que en las
donadoras, per0 se pueden emplear animales de cualquier raza o cruza (el cruzamiento
aumenta la fertilidad), sin importar su procedencia, per0 teniendo en cuenta que se traten
de animales jovenes, sanos con buen desarrollo corporal, especialmente la pelvis (ya que
muchas veces tendran que parir becerros muy pesados), en perfecto estado nutricional,
dociles y faciles de manejar y con una produccion de leche suficiente para criar hasta el
destete su becerro (Aspron, 1992).
La donadora puede ser una vaca o vaquilla, si se escoge una vaca esta debe de tener por
lo menos 3 a 4 meses posparto. Si la futura donadora no se encuentra ciclando, se le
puede inducir al estro con un implante de Norgestomet por 7 -1 0 dias y aplicandole400 U.I.
de gonadotropina serica de la yegua pretiada (eCG) al momento del retiro del implante. El
primer ciclo de este tratamiento generalmente es de corta duracion, per0 el siguiente ciclo
es de duracion normal. Una buena decision es que la vaca tenga por lo menos dos ciclos
normales antes de comenzar el programa de superovulacion.
En el caso de que se desee superovular una vaquilla, es necesario que esta tenga un peso
vivo por lo menos de 350 kg, y que ya este ciclando normalmente. No es recomendable
utilizar implantes de norgestomet en vaquillas de por lo menos de 350 kg. que aun no
hayan empezado a ciclar, ya que cabe la posibilidad de que entre en anestro prolongado
(Canseco, 1999).

C; ,
S E L E C C I ~ N DE DONADORAS
Seleccion y manejo de Receptoras
Las hembras receptoras deben de cumplir con determinadas caracteristicas sanitarias,
nutricionales y reproductivas. Para la selection de esta se requiere de un balance entre las
caracteristicas raciales y de factibilidad economica. La raza parece no tener una consideracibn significativa aunque 10s animales cruzados son mas fertiles (Elsden et a/.,
1 986).
La receptora debe ser una vaca con uno a tres partos o vaquillas bien desarrolladas de la
raza y tip0 que se disponga, fertilidad probada capacidad materna, preferentemente de
buena talla y con una historia de partos normales, de facil manejo, con condiciones de
desarrollo corporal aceptables, deben de estar ciclando y por lo menos de 80 dias despues
del parto (Reagan, 1993), control de sanidad y programas de concentraciones de
receptoras de diversas explotaciones; es necesario establecer una cuarentena o en
programas en particular en una explotacion es necesario examinar cada animal para algunas enfermedades como la Tuberculosis, Brucelosis, Anaplasmosis, Lengua Azul, y
vacunar 10s animales nuevos contra Rinotraqueitis lnfecciosa Bovina, Diarrea Viral
bovina, Leptospirosis y Clostridiasis. Ademas de controles de parasitos internos y
externos (Elsden eta/. , 1986).

En el aspect0 nutricional las receptoras deberan estar sometidas a un regimen alimenticio que permita el increment0 gradual de 500 a 900 g al dia y sales minerales a libre acceso (
Elsden eta/., 1986; Reagan, 1993).
Las vacas receptoras preseleccionadas se determina su estado de salud y reproductiva, deben para lo cual es necesario una identificacion visible y permanente en el animal (arete,
fierro marcador o tatuaje). Para posteriormente levantar controles por medio de tarjetas individuates de sus eventos reproductivos (Reagan, 1993).
La palpacion rectal de las hembras se realiza para determinar su estado reproductivo, preferentemente anormalidades del cervix, cuernos uterinos, ovarios y diagnosticos de
gestaciones tempranas y tomar decisiones del metodo para implementar el programa.
En resumen Elsden y Seidel(1986), Reagan (1 993) citan que una receptora debe cumplir
con 10s siguientes criterios. Buena condicion corporal. Vacas jovenes de 1 a 3 partos 6 vaquillas bien
desarrolladas. Cruzas de Bos indicus y Bos taurus (FI), esta ultima de preferencia de razas
lecheras. Fertilidad probada y el cervix debe ser pasable facilmente.
Habilidad materna y que tengan un diagnostico previo de no preAez.
Suficiente aporte de nutrientes, minerales y vitaminas liposolubles principalmente
ADE. Deben ser identificadas con arete, fierro quemador o tatuaje.
Programa de vacunacion y de desparasitacion con fundamento a 10s resultados del
laboratorio de diagnostico.
Libre de enfermedades infectocontagiosas Brucelosis, Tuberculosis,
Leptospirosis, Tricomoniasis, Campilobacteriosis, Rinotraqueitis lnfecciosa Bovina
(IBR) y DiarreaViral Bovina (DVB).
.; 1
'"hr' 4% (c. I
I
f f
A . - /
\
:i k- i
S E L E C C I ~ N DE RECEPTORAS E5

2.3.-Agentes superovulatorios
La superovulacion es la induction de ovulaciones multiples mediante la disminucion de la
atresia folicular con el uso de hormonas. Existen varios productos comerciales que se
pueden usar en 10s programas de superovulacion, la mayoria estan basados en la de
estimulacion folicular (Mombiela, 2000).
En 10s bovinos la superovulacion puede ser provocada por diferentes preparados de actividad gonadotropica siendo 10s principales: la gonadotropina corionica equina (eCG) y
la hormona foliculo estimulante de origen porcino (FSH-P) (Caral eta/., 1986).
Segun Elsden (1 980) el tratamiento mas utilizado en la actualidad es el de la aplicacion
decreciente de FSH-P durante cuatro dias a una dosis de 280 mg. a 400 mg tomando
como primer dia, el decimo a partir de la aparicion de estro y aplicando prostaglandinas
sinteticas (PGF, alfa) el tercer dia de superovulacion (Cuadro 1).
CUADRO 1.- SlSTEMA DE SUPEROVULACI~N CON FSH EN DOSlS DE CRECIENTE
Dia
0 = ESTRO
10
11
12
13
14
14
Actividades
am Pm
ESTRO
FSH FSH
FSH FSH
FSH + PGF2x FSH + PGIS:!
FSH FSH
ESTRO ESTRO
I .A I .A
La hormona FSH-P induce una respuesta superovulatoria mas uniforme y preferible que la lograda con la eCG(Elsden, 1978).

2.4.- Metodos de sincronizacion de estros
Como se menciono las inyecciones exogenas de FSH (eCG o FSH) tiene uso difundido en 10s programas de superovulacion multiple y transferencia embrionaria para incrementar la
disponibilidad de embriones provenientes de animales de merito genetic0 superior. Las inyecciones subcutaneas o intramusculares de eCG o FSH por lo general estimula el
crecimiento adicional de foliculos 10s cuales ovularan de manera espontanea sin
necesidad de LH o hCG exogena en el ganado bovino.
En virtud de que la FSH tiene una vida media mas corta (2 a 2.5 horas) que la eCG (24-72
horas). Por lo general es necesario dividir la dosis total, inyectar a intervalos de 12 horas
durante tres a cuatro dias para estimular la misma cantidad de crecimiento folicular que
resultaria de una sola inyeccion de eCG. Pese a la superioridad de la FSH, la eCG
continlja utilizandose sola o combinada con antisueros, 10s cuales se unen y bloquean a
cualquier exceso de eCG en la circulacion despues del inicio del estro. Sin embargo, el
suero anti-eCG no tuvo ningun efecto importante en las cantidades de cuerpos luteos ode
embriones (Hafez, 2000).
Durante 10s ljltimos 10 aAos la Prostaglandina F, alfa y su analogo el Cloprostenol han
contribuido de manera importante a la superovulacion, ya que la PGF, alfa no solo
aumenta la flexibilidad para programar la superovulacion sino tambien representa un
tratamiento excelente para la produccion de gran nljmero de embriones normales. El
tratamiento superovulatorio puede iniciarse en cualquier momento entre el dia 6 del ciclo
estral y la regresion natural del cuerpo lirteo; sin embargo, el mejor momento para el
tratamiento en el ganado bovino es entre 10s dias 8 y 12 del ciclo estral (Hafez y Hafez,
2004).
Prostaglandinas
Las prostaglandinas derivan de 10s acidos grasos esenciales por oxidacion catalizada por enzimas, son agentes paracrinos de vida corta cuyas acciones estan mediadas por receptores especificos de la membrana plasmatica. Estas hormonas median la secrecion
de GnRH, ovulacion, regresion del cuerpo Iuteo, maduracion del cuello uterino y el parto.
Las prostaglandinas derivan del acido araquidonico, presentan 20 atomos de carbon0 y un
anillo ciclopentano, las que se encuentran mas relacionadas con la reproduccion es la
PGF2a y prostaglandina E,. Son transportadas en la sangre para actuar en un tejido
blanco lejos del lugar de su produccion, ademas es el agente luteolitico natural que finaliza

la fase lutea del ciclo estral y permite el inicio de un nuevo ciclo estral en ausencia de fertilization.
PROSTAGLANDINA NATURAL (DINOPROST TROMETAMINA)
El proceso de luteolisis inicia con 10s estrogenos, estos preparan receptores para oxitocina
en la mucosa del endometrio, evento que se da en la etapa del del diestro del ciclo, el cuerpo luteo (CL) en el dia 16 y 17 comienza a liberar oxitocina, esta se dirige a la
membrana celular localiza su receptor y se lleva a cab0 el proceso de reacciones enzimaticas como la fosfolipasa A, para la sintesis de PGF,a e inician 10s pulsos que van de
6 a 8, 10s cuales llegan por via local o general y destruyen (lisis) al CL disminuyendo la concentracion de progesterona hasta hacerlo desaparecer.
Existen prostaglandinas naturales y sintetica, las naturales son aquellas que tienen su
estructura quimica y biologica parecidas a las que se producen en un ser vivo, ejemplo de
esto se tiene (Lutalyse, Lab. Pfizer) las sinteticas su estructura quimica y biologica son
modificadas a las que se produce en el organism0 animal Luprostiol (Prosolvin, Lab,
Intervet). Esta hormona solo funciona cuando existe un cuerpo luteo funcional. Es
importante realizar un diagnostic0 por palpacion o ultrasonografia para detectar la estructura y poder aplicar la PGF,a.
LUPROSTIOL (ANALOGO SINTETlCO DE LA PROSTAGLANDINA F,ALFA)

Progesterona natural y progestagenos
Ireland y Roche (1982), citado por Montiel (1996) indican que la progesterona y 10s progestagenos sinteticos suprimen el estro y la ovulacion, actuando a traves de un mecanismo de retroalimentacion negativa sobre la liberacion de la hormona luteinizante (LH), por lo mismo probablemente se reduce la frecuencia de 10s pulsos de esta hormona y se impide que algljn foliculo ovarico complete su desarrollo y ovule. Al retirar la hormona 10s foliculos de todas aquellas vacas tratadas completaran su desarrollo al mismo tiempo lo que provoca el estro sincronizado.
lnicialmente la progesterona se aplicaba en inyecciones diarias haciendo poco practico el sistema. Mas tarde, Christian y Casida (1980) demostraron que las inyecciones de
progesterona prolongaban la etapa del diestro por tanto tiempo como fueran aplicadas, al
termino las hembras entraban en estro y ovulaban en forma sincronizada. Posteriormente
se descubre 10s progestagenos via oral administrados en el alimento, el inconveniente es
que no existe control precis0 del consumo de las dosis diaria para cada animal, lo que
ocasiona respuestas variables en la sincronizacion.
Aplicacion via oral
El uso de progestagenos en el alimento como acetato de flurogestona (FGA): 30 6 45 mg
por dosis total, acetato de melengestrol (MGA): 11 a 60 mgldia, simplifica la
administracion de la hormona, pudiendo controlar el final del tratamiento mediante el retiro
del progestageno de la mezcla del alimento. La duracion puede ser de 7 a 14 dias de
aplicacion. La desventaja seria que no todos 10s animales comen la misma cantidad de
alimento y como consecuencia de la hormona.
Dispositivos intravaginales
En el mercado se encuentran dos productos: uno de ellos es el dispositivo en forma de espiral de acero inoxidable de liberacion controlada (PRID) con el 9% de progesterona del
laboratorio Sanofi Animal Helt y el dispositivo liberador de droga interna (CIDR) con 1.9 g de
progesterona del laboratorio Pfizer., ambos se aplican por periodos de 9 a 10 dias.

PROGESTERONA NATURAL - CIDR. LAB-PFIZER
El porcentaje de vacas que entran en estro despues del tratamiento con el CIDR, depende
de la etapa del ciclo estral en que se encuentre. Asi, el porcentaje que entra en estro es
mayor si el animal se encuentra en el diestro tardio y mas bajo en el diestro temprano ya
que en el retiro del CIDR algunas vacas todavia presentan un cuerpo lljteo funcional.
lmplantes auriculares
Se coloca debajo de la piel de la oreja, entre cartilago y piel en forma subcutanea durante 9
dias. El principio activo es el Norgestomet de 3 mg por dosis total, mas valerato de
estradiol: 5 mg intramuscular (Crestar, Lab. Intervet), este llltimo se modifica, en algunos
casos se aplica en otros no.
El norgestomet ejerce un efecto sobre el aparato reproductivo y el hipotalamo, suprime la
conducta estral y la ovulacion, Al retiro del implante se presenta la conducta estral y la
ovulacion. Los animales presentan estro dentro de las 100 horas posteriores al retiro del implante.
NORGESTOMET MAS VALERATO DE ESTRADIOL

El mecanismo de accion del progestageno junto con el estradiol es el de suprimir el estro
por medio del implante de norgestomet y la inyeccion con valerato de estradiol. El valerato
de estradiol causa atresia del foliculo antral, regresion del cuerpo lute0 y provoca el
reclutamiento de un nuevo grupo de foliculos de 4-5 dias de la aplicacion del tratamiento, el
norgestomet por si solo no afecta la forrnacion y funcion del cuerpo Iuteo natural ni la
gestacion.
2.5.- Tecnicas de Superovulacion
La respuesta superovulatoria depende de muchos factores, como son: raza, condicion
corporal, epoca del aAo, produccion lactea, edad, numero de parto, hormona, dosis,
regimen de colecta, dia del ciclo, superovulaciones previas y factor de individualidad del
animal. Algunos factores pueden ser controlados, aunque parcialmente, en tanto otros no,
lo que dificulta asignar una dosis adecuada a cada animal y predecir la respuesta.
GANADO CON BUENA CONDIC16N CORPORAL PARA LA SUPEROVULACI~N.
La superovulacion consiste en inducir el crecimiento extra de foliculos ovaricos mediante
la aplicacion de gonadotropinas. La aplicacion de este tipo de hormonas permite rescatar
de la atresia a un numero determinado de foliculos ovaricos de manera que estos puedan
seguir su crecimiento y llegar a la ovulacion.
Las gonadotropinas mas utilizadas son la hormona foliculo estimulante (FSH) y la
gonadotropina corionica equina (eCG). Tambien se han utilizado en forma experimental
extractos hipofisiarios de equino (HAP).

GONADOTROPINA CORldNlCA EQUINA (eCG) CON ACTIVIDAD DE LA HORMONA FOL~CULO ESTIMULANTE (FSH).
La FSH esta compuesta por glucoproteinas, dos subunidades diferentes que contienen
carbohidratos y que se hallan asociadas por uniones no covalentes, estas subunidades se
denominan a y p, esta ultima le confiere la actividad biologica a esta hormona. La
expresion de las gonadotropinas es modulada por factores hipotalamicos como la
hormona liberadora de las gonadotropinas (GnRH), por factores intrahipofisiarios . . _ I . _ L - I _ _ .- <.-*!-I _ _ - _A!. .!.-A . . I-I!-L-L!-- . . - - - --*---a: ---- I--::- 1-1 L - - 1 - -I- prlnclpalrnenre 10s peprlaos acrlvlna y rollsrarlna y por rerroallrnenraclon gonaaal ranro ae
hormonas esteroides como de peptidos. Los receptores de las gonadotropinas son m i m m h r n o A n I- f -mil;- A n r n r r n n t n r n e 3 r r n n l n A n e r r n n In nrntninn f2 I n hinein+neie A n I-e
suDunlaaaes ae las gonaaorroplnas proceae ae la rraauccron ae las suDunlaaaes ae 10s
RNArn, de modificaciones postraduccionales de las subunidades precursoras y del
plegamiento y la combinacion de las subunidades, del empaquetamiento de la hormona
madura y la secrecion de la hormona (Yen et a/., 2001). La hormona tiene un peso
molecular de 33,000 daltons, actua en las celulas de la granulosa, celulas de Sertoli e
hipotalamo, su funcion es el crecimiento folicular, mitosis de las celulas de la granulosa e
interviene en la maduracion del foliculo y del ovocito.
La FSH ha sido uno de las mas hormonas ensayadas para superovular y con las que se
tienen mejores resultados. En general, se trata de extractos hipofisiarios proveniente de ovino y porcino parcialmente purificados, tiene una actividad biologica corta en el torrente
circulatorio, no mayor de 3 horas, por lo que para que ejerza su efecto se tiene que aplicar cada 12 horas durante4 6 5 dias. El Folltropin-V (Lab. Vetrepharm-Canada) es un product0
comercial que tiene una purificacion en proporcion de LHlFSH de 1 :5, con dosis de 280 a 400 mg dependiendo la talla del animal ha demostrado proporcionar buenos resultados. La FSH producida por ingenieria genetica esta exenta de contaminacion con LH y no varia
dependiendo del lote. La dosis total de FSH se divide en 8 fracciones en forma
decreciente. 77

.- u - m r - r
'OLLTROP rr 4 PA.&.
HORMONA FOLICULO ESTIMULANTE (FSH). NIH-FSH-PI
I I I I I 1 I I I I 9.10 11 0 8 9 10 11 12 13 0 0 6 7
nun plqmmpqq 00 A A A t 3 i I i i i ; I 1 i i
00 1 ( 1 1 .- l~~. F S H' F S H F S H F s H 9
53 11% mq 30 mg, 20 ma
ESTRO
i 1 -7
PGFZs
C 4 - 4 C
SIWCRONILACION S U P E R O V U ~ ~ C ~ ~ N IA COLECCI~N
DE EMBRIONES
- - UI
yh. 1111 Prescntacion dcl astro
m .-. I Duracion dcl ostro 1
1 No do cuerpo~ lr~teos I NO do embrlonos y ~ v u ~ o s I
' I l l !\L I c.1 ll 11\11 > \ I ) I l l I
l'li/i*r I
- -
ESQUEMA 3. DIAGRAMA DE LA SINCRONIZAC~~N, SUPEROVULACI~N Y COLECCldN
DE EMBRIONES EN LAS VAQUILLAS DONADORAS.
La eCG antes conocida como PMSG (Gonadotropina serica de la yegua preriada), tambien es una de las hormonas mas investigadas para superovular, esta hormona tiene
una actividad biologica larga (48 a 72 horas), por su contenido de acido sialico es metabolizado lentamente y por lo mismo se aplica una sola vez. Su dosis varia de 1,500 a
3,000 U.I. y se aplica via intramuscular durante cualquiera de 10s dias del noveno y
doceavo del ciclo estral. Esta hormona tiene la desventaja de que despues de haber
causado la superovulacion sigue promoviendo el desarrollo de foliculos ovaricos, lo que causa una elevation exagerada de estrogenos que afecta la coleccion y calidad de 10s embriones.

2.6.- Deteccion de estro
En un programa de transferencia de embriones, la deteccion de estros es uno de 10s aspectos mas importantes. Al empezar el programa podria ser suficiente observar calores de 3 6 4 veces a1 dia; sin embargo, al finalizar la superovulacion se debe realizar las 24 horas del dia, tanto para inseminar a tiempo a las donadoras como para poder seleccionar las receptoras que entraron en estro en forma mas sincronizadas con las donadoras.
MANIFESTACION DE ESTRO EN EL GANADO BOVINO.
2.7.- Inserninacion de las vacas donadoras y superovuladas
La recomendacion mas comun es inseminar las hembras donadoras 12 horas despues
del inicio del estro y reinseminar 12 horas mas tarde. Esto se debe a que el animal
superovulado las primeras ovulaciones ocurren mas temprano que en las hembras no
superovuladas y se requieren de la presencia de semen viable durante el period0 en el que
ovulan el resto de 10s foliculos ovaricos. El lapso entre la primera y la ljltima ovulation generalmente es de 12 horas.
En algunos protocolos se han usado 2 dosis de semen, en la primera o segunda inseminacion artificial con el objeto de tratar de aumentar la fertilization de 10s ovocitos, aunque frecuentemente el precio de las dosis del semen es limitante. Rivera, ( I 987) indica
tres inseminaciones con dosis dobles a interval0 de 12 horas despues de iniciado el estro. Coelho y Acevedo, (1991) mencionan utilizar en vacas superovuladas dos inseminaciones 12 y 24 horas despues de iniciando el estro

INSEMINACI~N ARTIFICIAL EN EL GANADO BOVINO

Ill.- MEDlOS PARA LA COLECCION DE EMBRIONES
La manipulacion de 10s embriones fuera del ambiente uterino significa un gran riesgo, para evitar esto, se busca eliminar 10s cambios bruscos en la constitucion de las sustancias que
entran en contact0 con 10s embriones, pretendiendo igualar a las sustancias y secreciones existentes en el ambiente uterino. Las caracteristicas mas importantes en las soluciones utilizadas para este fin son: osmolaridad, pH, esterilidad y toxicidad.
La osmolaridad es la medida de la concentracion total de sales y compuestos de peso
molecular bajo en la solucion. Debe estar entre 270 a 310 miliosmoles por kilogramo,
desviaciones a mayor o menor concentracion pueden causar dafios irreversibles al
embrion. El pH debera mantenerse entre 7.2 y 7.6, aunque 10s embriones bovinos toleran
desviaciones sustanciales del rango optimo durante cortos periodos de tiempo. La
esterilidad de la solucion es muy importante, ya que la gestacion puede verse
drasticamente disminuida dependiendo del tipo de microorganismos presentes. La
toxicidad de algunos compuestos quimicos organicos o inorganicos puede dafiar y hasta destruir 10s embriones como la presencia de pequefias cantidades de ciertos metales,
como plomo y mercurio.
MEDlOS PARA LA C O L E C C I ~ N DE EMBRIONES
El metabolismo de microorganismos contaminantes pueden alterar el medio y volverlo
toxico para 10s embriones. La filtration de millipore (0.22 - 0.45 micras), constituye el
metodo idoneo para la esterilizacion de 10s medios, utilizando tecnicas y recipientes
esteriles, ademas es importante la adicion de antibioticos y antimicoticos.
Para lograr el pH fisiologico, algunos medios son amortiguados con carbonatos, es
necesaria una atmosfera del5% de CO, para mantener la conservation de carbonatos y
con ellos el pH. La solucion Hartmann modificada y solucion buferada fosfatada (PBS) Dulbecco, son las soluciones que constituyen 10s medios mas practicos para su uso en la
transferencia de embriones en bovinos.

3.1 .- Congelacion
El agua es el constituyente principal de 10s fluidos biologicos, responsable del transporte interno de las sustancias quimicas esenciales. El agua pura se congela y forma cristales a
O°C, mientras que esta misma que contiene iones y otras sustancias en solucion lo hace a
temperaturas mas bajas, dependiendo de la concentracion de tales sustancias. Conforme
el agua de una solucion se congela, 10s cristales de agua pura que se forman van dejando mayores concentraciones liquidas de aquellas sustancias que estan en solucion. Este
hecho aumenta la presion osmotica del resto del soluto, lo que puede determinar las lesiones de las celulas.
Las principales consecuencias fisicas y quimicas de la congelacion son la separacion del
agua pura de la solucion para formar hielo y la mayor concentracion de soluto resultante en el liquido residual. Estos hechos y sus efectos sobre las celulas se hallan influidos por el
nivel y 10s tipos de agentes crioprotectores, por la osmolaridad y el pH del diluyente y por la
rapidez de congelacion.
Desde que Whittingham et a/. (1972) y Wilmut (1972) quienes usaron escalas de
velocidad de enfriamiento lento (0, l-0.3°C/min) desde temperatura ambiente hasta -60 o - 80°C, se han hecho grandes modificaciones en las curvas de enfriamiento. En casi todos
10s trabajos publicados ultimamente se colocan 10s embriones directamente a -7"C,
permanecen 10 minutos, se enfrian a 0.5-0.6"CImin hasta -35°C y se sumergen en
nitrogen0 liquido. Este avance hoy dia es aplicado por todas las empresas e
investigadores en el proceso de congelamiento de embriones el cual reduce el tiempo de congelacion a no mas de una hora.
ESQUEMA DE LOS EVENTOS F~SICOS OCURRIDOS EN EL CONGELAMIENTO (HEXAGONOS REPRESENTAN CRISTALES DE HlELO DE DIFERENTE TAMAflO SEGUN LA VELOCIDAD DE ENFRIAMIENTO).

El mol, es la unidad de medida basica del sistema internacional de unidades, definida como la cantidad de una sustancia que contiene tantas entidades elementales (atomos,
moleculas, iones, electrones u otras particulas) como atomos hay en'0.012 kg (12 g) de
carbono 12. Esa cantidad de particulas es aproximadamente de 6.0221 X 1 o * ~ , el llamado
nimero de Avogadro. Por lo tanto, un mol es la cantidad de cualquier sustancia cuya masa expresada en gramos es numericamente igual a la masa atornica de dicha
sustancia.
La molaridad se define como la cantidad de sustancia de soluto, expresada en moles,
contenida en un cierto volumen de disolucion, expresado en litros, es decir: M = nlv. El
nljmero de moles de soluto equivale a1 cociente entre la masa de soluto y la masa de un
mol (masa molar) de soluto. Por ejemplo, para conocer la molaridad de una disolucion
que se ha preparado disolviendo 70 g de cloruro de sodio (NaCI) hasta obtener 2 litros
de disolucion, hay que calcular el nirmero de moles de NaCI; como la masa molar del
cloruro de sodio es la suma de las masas atomicas de sus elementos, es decir, 23 +
35.5 = 58.5 glmol, el nljmero de moles sera 70158.5 = 1.2 por tanto, M = 1.2/2= 0.6 M.
La molaridad se define por la siguiente ecuacion:
M = Molaridad = moles de soluto
litros de disolucion
Una disolucion 1.46 molar de glucosa, escrita como 1.46 M contiene I .46 moles de
soluto de glucosa en un litro de la disolucion, una disolucion de 500 ml que contiene
0.730 moles de glucosa tambien tiene una concentracion de 1.46 M. La unidad de
molaridad es moles por litro, por lo que una disolucion de 500 ml que contiene 0.730 moles de glucosa equivale a 1.46 mollLo 1.46 M.
La disoluciones son las mezclas homogeneas de dos o mas sustancias, la sustancia
presente en mayor cantidad suele recibir el nombre de disolvente y a la de menor
cantidad se le llama soluto y es la sustancia disuelta. El soluto puede ser un gas, un
liquido o un solido y el disolvente puede ser tambien un gas, un liquido o un solido. El
agua con gases un ejemplo de un gas (dioxido de carbono) disuelto en un liquido (agua). Existen distintas formas de expresar la concentracion de una disolucion, per0 las dos mas utilizadas son: gramos por litro (gll) y molaridad (M). Los gramos por litro indican la

masa de soluto expresada en gramos, contenida en un determinado volumen de
disolucion, expresado en litros. Asi, una disolucion de cloruro de sodio con una
concentracion de 40 g/l contiene 40 g de cloruro de sodio en un litro de disolucion.
3.1.2.- Funcion de 10s crioprotectores
Estos compuestos muy hidrofilos (en general polialcoholes) tienen la capacidad de
penetrar facilmente en la celula por simple difusion (osmosis), sus moleculas retrasan la
formacion de cristales de hielo por el descenso del punto de congelacion. Por otra parte,
limitan 10s efectos de la solucion, eliminando una parte del agua intracelular por
elevacion de la presion osmotica extracelular.
En 1973 se dio a conocer la primera gestacion en bovinos producida con un embrion
congelado/descongelado. Desde entonces se han publicado miles de articulos sobre
experimentos que tratan diferentes aspectos de la criobiologia embrionaria, que han
conducido, no solo a una mejora sustancial de 10s procedimientos para la congelacion
embrionaria, sin0 aun mejor entendimiento de 10s fundamentos de la respuesta de 10s
embriones a 10s procesos de congelacion y descongelacion de embriones.
La criopreservacion y el almacenamiento de embriones a bajas temperaturas (nitrogen0
liquido) presentan numerosas ventajas, tanto desde el punto de vista biologico como
desde el comercial.
TERM0 DE NITR~GENO LIQUID0 PARA CONGELAMIENTO
YALMACENAMIENTO DE EMBRIONES.
Permite una reduccion de costos derivados de la aplicacion de las tecnologias
reproductivas.
Facilita una disociacion de todas estas tecnicas de la actividad reproductiva ciclica
que presentan las especies mamiferas de interes, consiguiendose una

independencia temporal del estado fisiologico de 10s animales (hembras
receptoras).
Elimina las patologias que normalmente se asocian a1 mantenimiento de animales
vivos.
Posibilita la conservacion de razas o especies en riesgo de extincion mediante la
creacion de bancos de embriones congelados, manteniendo intact0 el patrimonio
genetico.
La criopreservacion esta influida por un elevado nljmero de variables, de forma que
ninguna aproximacion que contemple solo uno o parte de 10s aspectos implicados
garantiza una eficacia total. A pesar de esto y con independencia del sistema utilizado, 10s
principios basicos persiguen una proteccion de las celulas frente a 10s principales efectos
perjudiciales del proceso, como la formacion de hielo intracelular, deshidratacion y efectos
toxicos de 10s crioprotectores.
3.1.3.- Velocidad de enfriamiento.
El agua atraviesa con facilidad las membranas para diluir las soluciones, per0 usualmente
es mas dificil que las sales atraviesen las membranas. El medio usado para la estancia de
10s embriones tiene una concentracion de sales de 270 a 320 mOsmIkg y la mayoria es
cloruro de sodio. A esta concentracion, las sales fuera de las celulas balancean a las
proteinas del interior, que son demasiado grandes para pasar por las membranas
semipermeables. De igual manera, las moleculas grandes de algunos aditivos
crioprotectores, incluyendo 10s azljcares, son incapaces de atravesar las membranas
celulares.
L),wio ce1111il tl111.111te el C O I I ~ J ~ ~ ~ I I I I I ~ I I ~ I ~
A
L .-.:A f , 8 % , ,a:.,
muy lenta dpilma : muy r5plda
+ Velociclacl cle e~rfrin~niemto
CURVAS DE SOBREVIVENCIA DE C~LULAS SEGUN LA VELOCIDAD DE CONGELACI~N.

Si el enfriamiento es demasiado rapido, el agua intracelular no puede salir en cantidad
suficiente, formandose en medida considerable cristales de hielo en el interior de las
celulas, cristales que aumentaran de tamaAo, durante la congelacion y la descongelacion,
con perjuicio de las estructuras celulares. Si por el contrario, el enfriamiento es muy lento, la
deshidratacion progresiva de las celulas determina una elevacion sensible de la
concentracion de electrolitos en el medio celular, modificando sus propiedades fisico-
quimicas (efecto de solucion) fenomeno que comporta la perdida de viabilidad de las
celulas.
3.1.4.- Clasificacion de 10s crioprotectores.
Los crioprotectores se clasifican en penetrantes y no penetrantes, dentro de 10s
compuestos penetrantes se encuentra el glicerol, dimetilsulfoxido (DMSO), propilenglicol,
polivinilpirrolidona, etanol, etilenglicol y otros alcoholes. En 10s no penetrantes se
encuentran la sacarosa, g lucosa, trialosa y rafinosa.
Penetrantes.
Protegen a las celulas de 10s efectos dariinos de la alta concentracion de soluto producido
en el medio exterior durante la congelacion. La cinetica de entrada y salida de estos
crioprotectores esta caracterizado por algunos factores como cociente de permeabilidad,
cociente de temperatura y cociente de reflexion.
Durante la adicion de un crioprotector penetrante las celulas se reducen transitoriamente
hasta un nivel que depende de 10s factores antes mencionados, ademas de la velocidad de
adicion. Si el crioprotector es eliminado despues de la descongelacion por dilucion, las
celulas se hidratan transitoriamente hasta un nivel que depende de la velocidad de dilucion.
La adicion al crioprotector consiste en pasar el embrion por diferentes concentraciones
crecientes de esta sustancia, partiendo de la solucion buferada fosfatada (PBS) hasta una
concentracion final del crioconservador de I .4 M mas PBS (Solano, 1988). El glicerol a 1.4
M permite obtener un elevado porcentaje de viabilidad cuando 10s embriones son
congelados a una velocidad de 0.3"Cl minuto hasta -30 o -35°C; sin embargo, tiene la
desventaja que al exponer el embrion a la solucion de glicerol para que este penetre y
tambien despues de la descongelacion para su elimination, se necesita varios pases que
consume mucho tiempo.

En un principio se empleaban 4 pases incrementando la concentracion del crioprotector, con intervalos de 10 minutos cada uno, este metodo era tan prolongado que afectaba la viabilidad de 10s embriones debido a la constante manipulacion. Posteriormente se
redujeron a 3 pases, hasta que en 1982 se demostro que 10s embriones pueden ser congelados en un solo pase al crioprotector.
Algunos estudios han descrito ventajas de 10s glicoles en la crioconservacion de
embriones porque sus moleculas son mucho mas permeables. Se utilizan de forma
exitosa, tambien en soluciones vitrificantes, debido a que su peso molecular es menor
que el del glicerol, propilenglicol y DMSO. De esta forma, la suficiente penetracion del
etilenglicol en el embrion tiene el lugar en corto tiempo y es eliminado rapidamente
durante la dilucion, resultando en una disminucion de la toxicidad embrionaria.
No penetrantes.
Son agentes que mantienen una presion osmotica alta en el medio extracelular durante
la eliminacion del crioprotector. Esto evita 10s choques osmoticos debido a la difusion del
glicerol hacia fuera despues de la descongelacion. Si se aiiade sacarosa a la solucion de
glicerol, despues de equilibrados 10s embriones, el tamatio de 10s mismos se reduce en
un 30 a 4O0/0 en comparacion con 10s equilibrados solo en glicerol.
Cuando se descongelan embriones en presencia de sacarosa, se limita el movimiento
del agua a traves de las membranas y asi se evita la lisis de las celulas embrionarias
durante la eliminacion del glicerol. Este beneficio de la sacarosa se debe a sus
propiedades osmoticas, estando involucrado tambien el transporte activo de iones a
traves de la membrana. Este proceso es controlado por el sistema Na+ y K ATPasa, el
cual es inhibido en presencia del glicerol. Si esto se aplica a embriones bovinos, la utilizacion de sacarosa en el medio de congelacion que contiene glicerol, permite restaurar el equilibrio quimico durante la congelacion.
Actualmente hay una mayor tendencia hacia la combinacion de 10s crioprotectores
penetrantes con 10s no penetrantes, brindando la oportunidad de congelar 10s embriones
en la misma explotacion ganadera y dejarlos a la disposicion del tecnico quienes pueden
transferirlos a las vacas en el momento apropiado.

3.1 5.- Funcion del glicerol
En general el glicerol, en la congelacion de embriones continua siendo el patron de
comparacion para cualquier agente crioprotector (Elsden et a/., 1986; Nljriez y Solano,
1996). Se dice que el glicerol actlja mediante un mecanismo de tamponamiento salino
(Lovelock, 1953). El glicerol se une con el agua y disminuye acentuadamente el punto de
congelacion de las soluciones, se forma en su presencia menos hielo a cualquier
temperatura dada (Stein, 1962; Farrant, 1964). En consecuencia, la concentracion de
solutos en el liquido residual se reduce correspondientemente. La influencia perjudicial de
10s solutos concentrados parece depender de la temperatura de aqui que el glicerol, al
reducir la temperatura al que se obtienen tales concentraciones de soluto, disminuye sus
efectos perjudiciales. Con base a esto, el nivel optimo de glicerol de un diluyente aumenta
conforme incrementa la tonicidad del mismo.
GLICOLES PARA LA CRlOPRESERVACldN (GLICEROL)
3.1 -6.- Funcion del etilenglicol
En algunos trabajos han descrito ventajas de 10s glicoles en la criopreservacion de
embriones porque sus moleculas son mucho mas permeables. Se utilizan de forma
exitosa tambien en soluciones vitrificantes, debido a que su peso molecular es menor que
el del glicerol, propilenglicol y DMSO. De esta forma el etilenglicol es sumamente
permeable en corto tiempo y es eliminado rapidamente durante la dilucion, resultando en
una disminucion de la toxicidad embrionaria.

GLICOLES PARA LA CRIOPRESERVACI~N (ETILENGLICOL)
3.1.7.- Congelacion de embriones con glicerol
En la actualidad existen diferentes maquinas congeladoras de embriones. Las de tipo manual o de campo y automaticas programadas para la congelacion. Tambien se
disponen de cuatro procedimientos de congelacion: 1) metodo convencional de
congelacion lenta y controlada, 2) congelacion ultrarrapida, 3) congelacion a un paso y 4) vitrification.
La congeladora de tip0 manual denominada Peter Elsden, consiste en un barril de acero donde se colocan las pajillas que contienen 10s embriones, conectado a un termocoplex
para el control de la temperatura. El embrion evaluado y lavado se pasa a una solucion que contiene PBS + alblimina serica bovina (ASB) + 10% glicerol (1.4 M), mientras tanto se
rotulan las pajillas francesas de 0.25 ml con 10s datos de la donadora y el semental, con la
ayuda de una jeringa de 1 ml se acopla a la pajilla francesa se succionan tres o cuatro
columnas de medio separadas por burbuja de aire, de manera que el embrion quede colocado en la ultima columna, inmediatamente se coloca en la congeladora de embriones
a temperatura de 0°C , se baja gradualmente 1°C por minuto hasta 10s -7°C donde se
efectua la cristalizacion, que consiste en inducir la formacion de cristales en el medio
interior de la pajilla. Esto se realiza facilmente tocando la pajilla con una pinza enfriada en
nitrogen0 liquid0 en la columna donde se encuentra el embrion. La formacion progresiva
de cristales de hielo en el medio que rodea a 10s embriones produce un aumento de la concentracion de 10s electrolitos en la fraccion todavia liquida de la solucion. A
consecuencia del aumento de la presion osmotica se produce la salida por difusion de una parte del agua intracelular y el embrion comienza a deshidratarse. Despues de la cristalizacion, el embrion permanece 10 minutos a -7 "C. A partir de esta temperatura el
enfriamiento se producira a un ritmo de 0.5 "Clminuto hasta 10s -30" C, llegando a esta
1 34

temperatura las pajillas con 10s embriones se sumer'gen directamente en el nitrogeno liquido.
TECNICA DE CONGELAMIENTO (PETER ELSDEN)
En otros estudios N~li iez y Solano (1996) mencionan que una vez colocados 10s embriones en las pajillas en un equipo programable que alcanza una temperatura de -5 a
-7 "C a una velocidad de 1 a 5"CIminuto. En este equipo se induce la formacion de
cristales de hielo en la parte superior de la pajilla durante 5 minutos, posteriormente la
temperatura es descendida a 0.3"CIminuto hasta -30°C o -35°C temperatura a la que se
sumergen las pajillas en nitrogeno liquido para su almacenamiento
Algodon + Polivinilico Aire Aire
, I I *
A
Sello Glicerol Glicerol
+ Embrion
LLENADO DE PAJILLA FRANCESAS DE 0.25 ML CON EL SlSTEMA DE GLICEROL

CUADRO 2. EMBRIONES CONGELADOS CON EL SISTEMA DE GLICEROL (ADAPTACI~N DE PETER ELSDEN, 1977).
3.1.8.- Congelacion de embriones con etilenglicol
I "C x I min. 4
1 "C x 2 min?
i , lnmediato 4
En la ultima decada del siglo XX se hizo una importante innovacion con base al
descubrimiento de que se puede congelar embriones con ciertos aditivos crioprotectores
que no necesitan extraerse al descongelamiento y permiten la transferencia directa del
embrion a la vaca receptora. El metodo utilizando es conocido como transferencia
directa, iniciado porvoelkel y Hu, 1992, haciendo que la transferencia sea muy parecida a
la inseminacion artificial.
La transferencia directa de embriones es el resultado de un avance tecnologico en la criopreservacion, que permite que la transferencia de un embrion sea como una inseminacion artificial. Esto es posible porque el agente crioprotector que se usa en 10s
embriones de transferencia directa permite que el embrion sea rehidratado dentro de la
receptora eliminando asi la necesidad del microscopio y el embriologo que se usan cuando se descongelan embriones congelados de manera tradicional.
Observaciones realizadas por varios investigadores sugieren que la permeabilidad de 10s embriones ovinos y bovinos (estadios de morulas y blastocistos) al etilenglicol y a1
0°C
-7°C
-32°C
-1960C
Se colocan las pajillas en la congeladora a 0°C
Se inicia el connelado gradual 1°C x minu
- Cristalizacion (Seeding) durante 10 minutos
Se continira el congelado gradual con escala 1 "C x 2 minutos hasta -32°C
Termina el congelado gradual
Se sumerge la pajilla a nitrogen0 liquid0 en forma inmediata a -1 96°C

propilenglicol es mayor que la permeabilidad al glicerol. Otros han referido metodos de
transferencia directa que utilizan como crioprotectores al glicerol, propilenglicol y el
etilenglicol mezclados o no con sacarosa.
Massip y Zwalmen (1 984) quienes trabajaron con glicerol 1.5 M y sacarosa 0.25 M como crioprotectores utilizando el procedimiento de un paso y transfiriendo directamente 10s
embriones en el ljtero a la receptora. Suzuki et a/. (1990) quienes utilizaron propilenglicol
1.6 M lo cual que permite que el embrion se rehidrate directamente en el medio materno,
encontrandose resultados satisfactorios de porcentajes de prefiez. No obstante, Cseh et a/. (1994) sugieren que la transferencia directa de embriones con el uso de sacarosa en la
pajilla puede ser aplicada en la practica con buenos resultados especialmente para
transferir embriones de baja calidad.
Los embriones se exponen a una solucion de etilenglicol al 1.5 M por 5 minutos a
temperatura ambiente (25 a 30°C), para estabilizarlos y se envasan en pajillas francesas
de 0.25 ml de la siguiente manera: se realizan 3 columnas de medio de mantenimiento
(PBS + 10% de ASB) de 2.5 cm cada uno, mas una columna de etilenglicol.de 2 cm donde
se coloca el embrion, las columnas van separados con espacios de aire. lnmediatamente
se coloca la pajilla francesa con el embrion en la congeladora manual a - 6"C, donde se
esperan 5 minutos para realizar la cristalizacion donde el embrion permanece 10 minutos a
esa temperatura, para luego realizar el enfriamiento a un ritmo de 0.5"C por minuto hasta
10s -32°C. Al alcanzar esta temperatura se sumerge la pajilla directamente en el nitrogen0
liquido a -1 96OC
Sol. Hartmann Sol. Hartmann
Etilenglicol + Embridn
LLENADO DE PAJlLLA FRANCESAS DE 0.25 ML CON EL METODO DE ETlLENGLlCOL

CUADRO 3. CONGELACION DE EMBRIONES BOVINOS CON EL METODO DE
ETILENGLICOL
an las F
?pose, c ultima
despue! pajilla
3.1.9.- Descongelacion de embriones con glicerol
- 6°C S ngeladora a
- Se deja
hat
Durante el proceso de congelamiento de 10s embriones a -1 96°C se forma inevitablemente
una cierta cantidad de hielo en las celulas. Para evitar un aumento destructivo del tamaAo de 10s pequefios cristales por aglomeraci6n durante la descongelacion, el proceso de recalentamiento debe ser tambien lo mas rapido posible (del orden de 2000°C/minuto).
Las pajillas extraidas del nitrogeno liquido se descongelan en agua de baAo maria a 37°C
durante 15 a 20 segundos.
lnmediatamente despues de la descongelacion es indispensable eliminar el crioprotector
para la supervivencia del embrion. No obstante, las celulas que presentan una
conservacion elevada de crioprotectores no deben ser colocadas directamente en un
medio isotonico. En tal caso, las diferencias de presion osmotica provocaria una entrada
Cristalizacion (Seeding) durante 10 minutos
Se continlia el congelado gradual con escala 1°C x 2 minutos hasta -32°C
Termina' el congelado gradual
Se sumerge la pajilla a nitrogeno liquido en forma inmediata a -1 96°C
-7°C
I "C x 2 min. 4
-
-32°C P
lnmediato
-1 96°C

demasiada rapida de agua en la celula con el riesgo de reventar. Es necesario proceder a
la eliminacion progresiva del crioprotector, de manera que 10s flujos de salida de
crioprotector y de entrada de agua Sean compatibles con la permeabilidad de la membrana
plasmatica.
La eliminacion del crioprotector por efecto de la sacarosa es por principio acelerar el flujo
de salida de este. La sacarosa de peso molecular elevado no penetra en la celula por simple difusion. Su presencia en el medio crea un aumento de la presion osmotica del
medio extracelular, favoreciendo la difusion masiva del crioprotector en el cornpartimiento
celular.
Pozo 2 I
I Sacarosa + I
I 50% Medio de .r I Transferencia ?
I
Pozo 3 Pozo 4 Medio de Se sube el
Transferencia embrion en la _. . I , Z 1 !
E L ~ M ~ N A C ~ ~ N DEL GLICEROL E H ~ D R A T A C ~ ~ N DEL E M B R ~ ~ N UTlLlZANDO LA CAJA MULTIWELDISCH.
El glicerol puede ser removido de varias formas:
1). Pasar al embrion por cuatro soluciones permitiendo una estancia de 5 minutos por
solution. a) PBS mas 10% de suero fetal bovino (SFB), con 6% de glicerol, mas 10% de sacarosa
(0.3 M).
b) PBS mas 10% SFB, con 3% de glicerol mas 10% de sacarosa (0.3 M).
c) PBS mas 10% SFB, con 0% de glicerol mas 10% de sacarosa (0.3 M).
d) PBS mas 10% SFB, sin glicerol sin sacarosa.
2). Eliminacion del crioprotector en forma inversa, se pasan 10s embriones por bafios de
concentracion decreciente, ejemplo: etilenglicol 1.5 M, despues 1.0 M, 0.5 M y por ultimo I;,

0 M. La duracion de cada baiio es de 5 a 10 minutos.
3). Retiro del crioprotector en tres pasos, por el metodo decreciente utilizando glicerol: 1.4 M, 0.93 M y 0.45 MI posteriormente medios para hidratar y transferir el embrion PBS
+ 10% SFB.
4). Saito (1994) recomienda una combinacion de 10% de sacarosa (0.3 M) con solucion
de PBS mas 20% de caseina serica bovina (CSB), con reposo de 5 a 10 minutos cada
paso. Este mismo autor realiza un proceso decreciente para esto utiliza glicerol: 10,
6.6 y 3.3% con un reposo de 5 minutos en cada paso, en el ljltimo paso PBS + 20% CSB
con un reposo de 10 minutos.
5). Concentraciones decrecientes de sacarosa: 10% de sacarosa + PBS con un reposo
de 10 minutos, 5% de sacarosa + PBS + 10% SFB con reposo de 5 minutos, 2.5% de sacarosa + PBS + 10 % SFB 5 minutos y el ultimo paso PBS + 10% SFB, quedando listo
para realizar la transferencia. Tambien hace referencia del retiro del crioprotector de un
paso utilizando soluciones de sacarosa al 10% durante 5 minutos, luego se hidrata en
PBS + 10% SFB y se trasfiere el embrion a la receptora.
Aire Aire
Medio de Embrion + Medio de Tran sferencia Medio de Transferencia
Transferencia
LLENADO DE PAJILLA FRANCESA DE 0.25 MLPARA REALIZAR LA TRANSFERENCIA DE EMBRIONES.

3.1 .lo.- Descongelacion de embriones con etilenglicol
En 1972, 10s cientificos descubrieron un nuevo crioprotector, el etilenglicol al 1.5 M, este
tiene una gran ventaja sobre el glicerol y es el de ser sumamente permeable. Una de las
mayores ventajas de 10s embriones congelados con etilenglicol es el de poder ser
transferidos de manera directa al utero de las receptoras, sin la necesidad de utilizar
diluyentes ni el microscopio para observar 10s embriones.
Esto representa por si misma una ventaja para la industria ganadera. Los veterinarios,
tecnicos inseminadores con experiencia son ahora candidatos para transferir embriones
en forma directa al ser estos congelados con etilenglicol. Las receptoras seran animales
sincronizados o buen trasferidos despues de la presentacion natural del estro. Todo esto
considerado dentro de la industria ganadera da un nuevo mercado para la transferencia
embrionaria representa un gran avance para el mundo cientifico en beneficio de la
ganaderia.
El protocolo para descongelar y transferir embriones de transferencia directa es el
siguiente:
La receptora debe haber sido detectada en estro siete dias antes de la
transferencia.
Debe existir un buen cuerpo luteo funcional.
Aplicar anestesia por via epidural de 4 a 6 ml de lidocaina al 2% a la altura de la
ultima vertebra sacra y la primera coccigea alrededor del cordon medular. ldentificar el embrion leyendo el rotulado en la parte superior. Sacar la pajilla del
termo de nitrogen0 Iiquido y mantenerlo al aire por 12 segundos. Sumergir la pajilla en agua a 35°C por 20 segundos.
Sacar la pajilla del agua, secarlo con servilleta, realizar el corte en la parte donde
fue sellada hermeticamente a calor o alcohol polivinilico. Colocar la pajilla en el aplicador de transferencia de embriones.
Montar la funda azul sobre la pistola, apretar firmemente para asegurarse que la
pajilla este bien colocada, usar el anillo de plastic0 para fijar la funda sobre la
pistola.
Pistola yfunda se protege dentro de la camisa sanitaria.
Una vez que la pistola este cargada, debe mantenerse en posicion horizontal. Esto
hace que el embrion se quede en el centro de la columna de liquido. Para transferir el embrion pasar la pistola suavemente por el cervix hasta el cuerno

uterino correspondiente a lado del ovario que contiene el cuerpo lirteo. El embrion se coloca a 10s 13 a 18 cm dentro del cuerno uterino. Si esto no fuera
posible, depositar el embrion lo mas profundo posibleen el cuerno.
3.1 .I1 .- Preparacion de medios
Durante la tecnica de coleccion de embriones se utilizan dos tipos de medios que son el
medio de coleccion y el medio de mantenimiento y de transferencia de embriones. El medio de coleccion es adicionado con 1% de suero sanguineo o proteina serica con la finalidad de lograr un siliconizado evitando con ello que 10s embriones se adhieran al
material con que entran en contact0 durante su coleccion, debido a la presencia de
macromoleculas como es la albljmina serica bovina.
+..? . . .-.tx . . . . . . . . . . . : . . . . , . " , t " I . ,, , ,. .; . .: . ' .
, 2;; : - , , - ., .- - ;, ! .... , , . . . . . . . - . .".;. :. : . . ':. ' .. ;.
. . . -,;..!. .".. ,, .: ,., ,.,; 8 <,& , .
, .' . ?'" . : , ..,.,r, .. \. ,':.. ;.,: ,: $ .,,, ..;.;: . :.!'.; ;; ,-. -. . . ,.,.;I:... .<. . t , * 'T ,it .',': - 3 . . . , , .< ; .. . . . . .
'. . . ,,.. ,::., , r,, . ,i>'+; .+.,!.j;*. .: f . , .>:...: <, -. I - + . , ., 6<,;,'<.-:
. . . . . - , :,. ,.,, :..:. t- - .-. . , 1 , : , , j;.: .,l,:".. ' :. , C ' i.? ', 3; .... :.. ;; I . . . . l ? . i
: j.il~, .i
, ,, . ! : ., .i', .. . . ,'. k I.... :. ..:.: . . . ,,.. ' . #, -.- .,.,,,, :.-~~.; . ." , . . ;.;, " .;.*;. t., ' . I(.':' i; + . 'f:;,,.. ., ; + , I ; *.<.; t;, 7.:. y , . ,-;; ,, ,4* ;:.it, . . . . . ,.. . ,. ! .. , ,;: ..,v, , > , . ,. i i
; 4 . . . . . .... .. .,, , .. '.;:.;::. # ." w., 2% r.?,;!:!:.:
~ S A Fr, Vi: . - ; . . - enydrate * - : S:ore 2
..,
Para el medio de mantenimiento se adiciona el 10% de suero sanguineo o proteina serica
para dotarlo de algunos factores nutricionales, ademas de evitar con ello que 10s
embriones floten o se adhieran, pueden emplearse suero fetal bovino, suero de oveja
menor de un atio, suero de becerro recien nacido, suero de novillo de 185 a 205 dias de
edad o bien alblimina serica bovina. Si el suero se obtiene de animales sanos y procesado
en forma adecuada 10s resultados seran sirnilares con cualquiera de ellos siempre y
cuando Sean tratados con calor (56OC130 minutos) para termoinactivar o destruir el
complemento, de no ser asi daiiaria a 10s embriones ya que es embriotoxico, ademas se
debe esterilizar porfiltracion con poros de 0.45 micras.

Actualmente la tecnica. mas usada en bovinos en la coleccion no quirurgica
(transcervical). Esta consiste en la lavar varias veces el cuerno uterino con el medio de
coleccion. Esto se consigue introduciendo el cateter de Foley a traves del cervix hasta la
curvatura mayor del cuerno, lugar donde se fija inflando el globo del cateter, el cual se une
a la manguera tygon. Por un lado de la bifurcacion entra el medio al utero, cada vez que se
llena este se le da un masaje, posteriormente se extrae el medio el cual sale hacia el filtro
colector de embriones. Una vez lavado un cuerno, se saca el cateter y se repite la
operacion en el cuerno opuesto. El momento indicado para realizar la coleccion es entre
el dia 6.5 a 7 de iniciado el estro debido al tipo de estructuras que se encuentran en estos
dias considerando de que 10s embriones pasan del oviduct0 al cuerno uterino en el cuarto
o quinto dia. Despues de la coleccion embrionaria las donadoras reciben una inyeccion de
prostaglandinas F2 a (25 mg de Dinoprost trometamina) con el objeto de inducir la
regresion de 10s cuerpos Iuteos, evitar una posible gestacion, incluso multiple y acortar el
period0 entre colecciones.
LAVADO DE LAS VACAS DONADORAS Y DE COLECCI~N DE EMBRIONES.
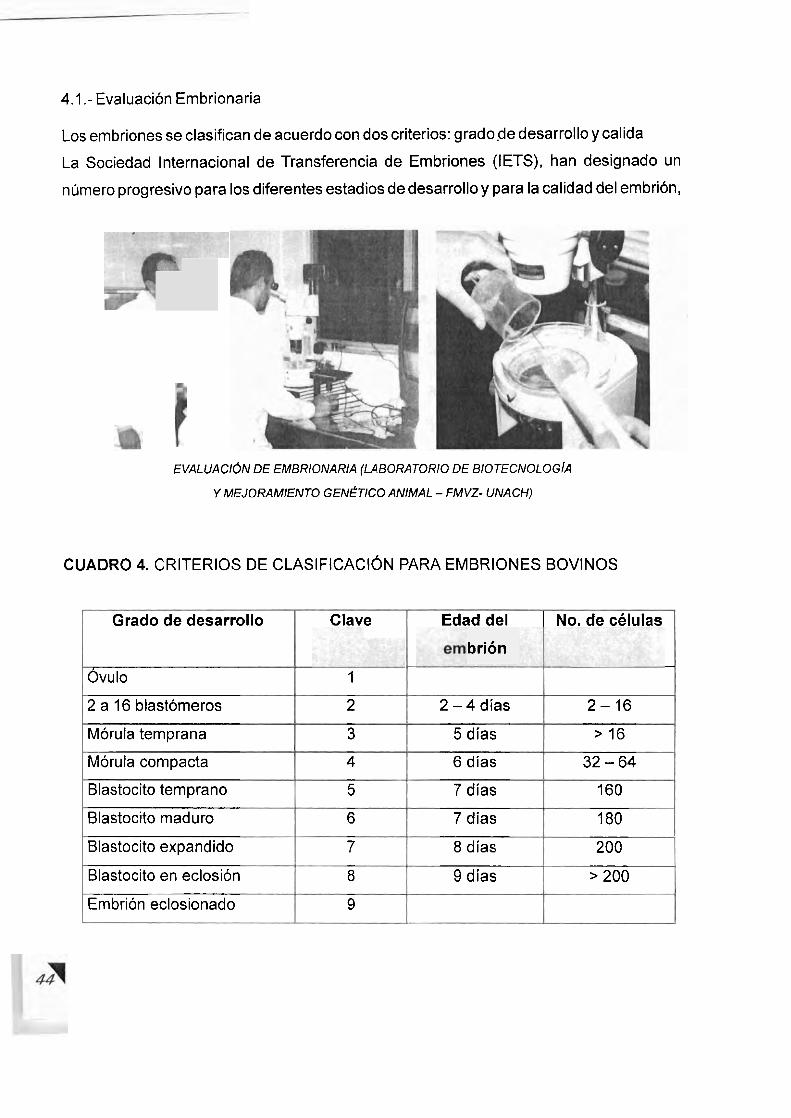
4.1 .- Evaluacion Embrionaria
Los embriones se clasifican de acuerdo con dos criterios: gradode desarrollo y calida
La Sociedad lnternacional de Transferencia de Embriones (IETS), han designado un
numero progresivo para 10s diferentes estadios de desarrollo y para la calidad del embrion,
EVALUACI~N DE EMBRlONARlA (LABORATORIO DE B~OTECNOLOG~A
Y MEJORAMIENTO GEN~TICO ANlMAL - FMVZ- UNACH)
CUADRO 4. CRlTERlOS DE CLASIFICACION PARA EMBRIONES BOVINOS
bridn
Grado de desarrollo
6vu10
2 a 16 blastomeros
Morula temprana
Morula compacta
Blastocito temprano
Blastocito maduro
Blastocito expandido
Blastocito en eclosion
Embrion eclosionado
Clave
1
2
3
4
5
6
7
8
9
Edad del I No. de celulas
2 - 4 dias
5 dias
6 dias
7 dias
7 dias
8 dias
9 dias
-
2-16
> 16
32 - 64
160
180
200
> 200

4.1.1 .- Embriones transferibles
Con base a sus caracteristicas morfologicas, cada embrion recibe una clasificacion por su
edad y calidad.
a) Morula compacta. Debido a la compactacion, las uniones de 10s blastomeros han formado una masa celular, de manera que ya no se aprecian 10s blastomeros individuates. El embrion ocupa del60 al70% del espacio perivitelino.
MORULA COMPACTA
b) Blastocito temprano. En la masa celular aparece una pequeiia cavidad llena de
liquid0 llamado blastocele. Aqui las celulas empiezan a diferenciarse. El embrion ocupa
del 70 al 8O0/0 del espacio perivitelino. La diferencia visual entre el trofoblasto y la masa
celular es apreciable.
BLASTOCITO TEMPRANO
E5

c) Blastocito maduro. Se observa las celulas del tabique embrionario (disco embrionario), que originan al embrion y a las celulas del trofoblasto. El embrion tiene apariencia de anillo
y ocupa el 90% del espacio perivitelino.
BLASTOCITO MADURO
d) Blastocito expandido. El diametro del embrion aumenta 1.2 a 1.5 veces y la zona
pelucida sufre un adelgazamiento. Estos embriones con frecuencia se colapsan sin que
se aprecie el blastocele; sin embargo, la zona pelucida permanece adelgazada.
BLASTOClTO EXPANDIDO
e) Blastocito en eclosion. El embrion tiene fisurada la zona pelljcida por donde eclosiona
paulatinamente hasta salir totalmente de ella, el embrion puede tener ya sea la forma
esferica de un blastocito expandido o bien estar colapsado.
BLASTOClTO EN ECLOSI~N
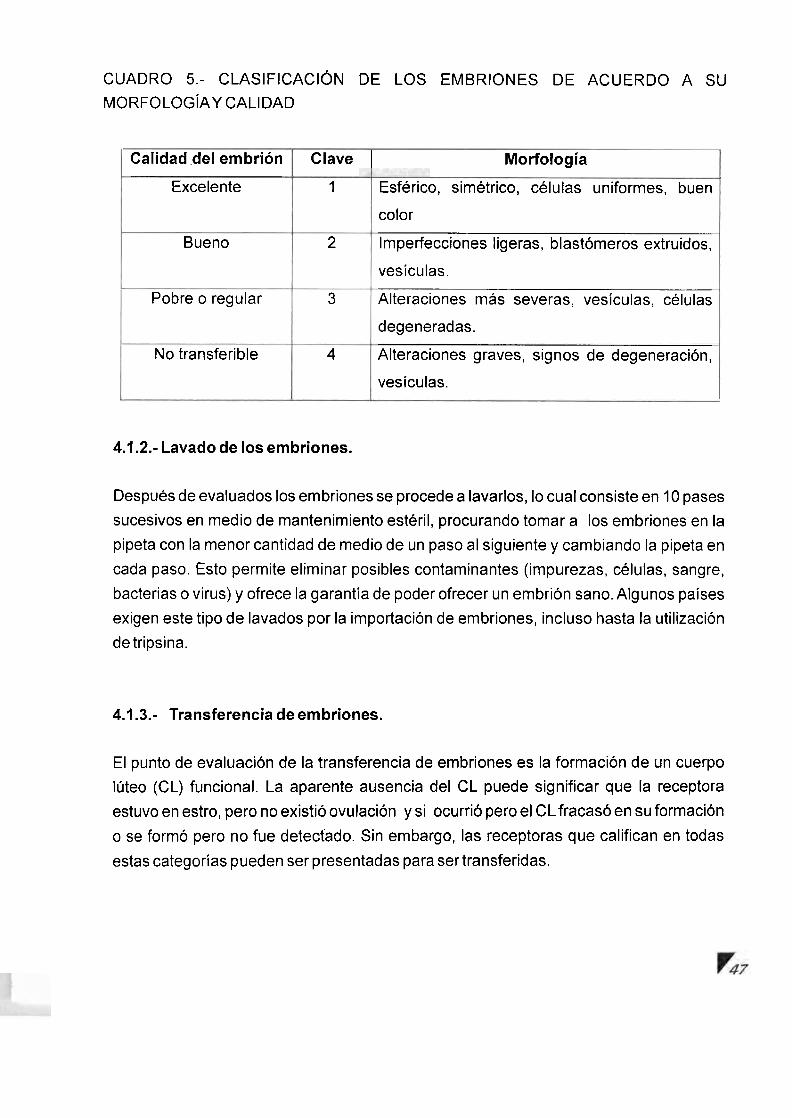
CUADRO 5.- CLASIFICACI~N DE LOS EMBRIONES DE ACUERDO A SU MORFOLOG~AY CALIDAD
4.1.2.- Lavado de 10s embriones.
Calidad del embrion
Excelente
Bueno
Pobre o regular
No transferible
Despues de evaluados 10s embriones se procede a lavarlos, lo cual consiste en 10 pases
sucesivos en medio de mantenimiento esteril, procurando tomar a 10s embriones en la
pipeta con la menor cantidad de medio de un paso al siguiente y cambiando la pipeta en
cada paso. Esto permite eliminar posibles contaminantes (impurezas, celulas, sangre,
bacterias o virus) y ofrece la garantia de poder ofrecer un embrion sano. Algunos paises
exigen este tip0 de lavados por la importacion de embriones, incluso hasta la utilizacion
de tripsina.
4.1.3.- Transferencia de embriones.
Clave ' Morfologia
El punto de evaluation de la transferencia de embriones es la formacion de un cuerpo
lllteo (CL) funcional. La aparente ausencia del CL puede significar que la receptora
estuvo en estro, per0 no existio ovulation y si ocurrio per0 el CLfracaso en su formacion
o se form6 per0 no fue detecfado. Sin embargo, las receptoras que califican en todas
estas categorias pueden ser presentadas para ser transferidas.
1
2
3
4
Esferico, simetrico, celulas uniformes, buen
color
lmperfecciones ligeras, blastomeros extruidos,
vesiculas.
Alteraciones mas severas, vesiculas, celulas
degeneradas.
Alteraciones graves, signos de degeneracion,
vesiculas.

TRANSFERENCIA DE EMBRIONES POR VIA TRANSCERVICAL
El metodo de transferencia de embriones no quirurgica es un procedimiento rapido y
relativamente barato. Es el metodo de mayor eleccion en gran parte de 10s programas de
TE en explotaciones comerciales. La evolution mas reciente en relacion a la TE, es el
almacenar embriones en pajillas congeladas con etilenglicol, que permite su transferencia
en forma directa despues del descongelamiento, haciendo este metodo como si fuera una
inseminacion artificial y es cada vez mas atractivo para 10s tecnicos y 10s productores
pecuarios. El factor determinante sobre la actividad de este metodo, es la destreza y
experiencia del profesional para determinar el sitio de la colocacion del embrion dentro del
cuerno uterino, como tambien el grado de lesion implicado durante la transferencia de
embriones.

4.1.4.- Biparticion de embriones.
La habilidad de producir muchas copias a partir de un solo individuo es algo que interesa a
10s productores pecuarios, porque constituye una poderosa herramienta de seleccion y
propagacion. Si se logran producir grandes cantidades de individuos geneticamente
identicos, se podrian usar como base en la produccion de carne, leche y en la seleccion
fenotipica que puede permitir un rapido cambio en caracteristicas especificas y muy
seleccionadas. Esta tecnologia combinada con la fertilizacion in vitro podria utilizarse para
producir grandes cantidades de embriones de alta calidad que despues serian congelados
o transplantados.
4.1 5.- Micromanipulador.
La biseccion de embriones es un procedimiento que da como resultado la produccion de
gemelos identicos al dividir el embrion usando tecnicas de microcirugia. El metodo mas
sencillo y eficiente para producir gemelos identicos es obtener embriones de 6 a 7 dias de
edad, cortarlos por la mitad y transplantar las dos mitades a igual nljmero de receptoras.
4.1.6.- Division artificial de 10s embriones.
En trabajos realizados por Leibo (1986) y Takeda (1 986) las tasas de pretiez obtenidas con
la transferencia de hemiembriones son muy variables con una media que varia de un 45 a
50 %. Asu vez la ausencia de la zona pelljcida en 10s miembros no afecta a 10s porcentajes
de pretiez para embriones transferidos frescos.
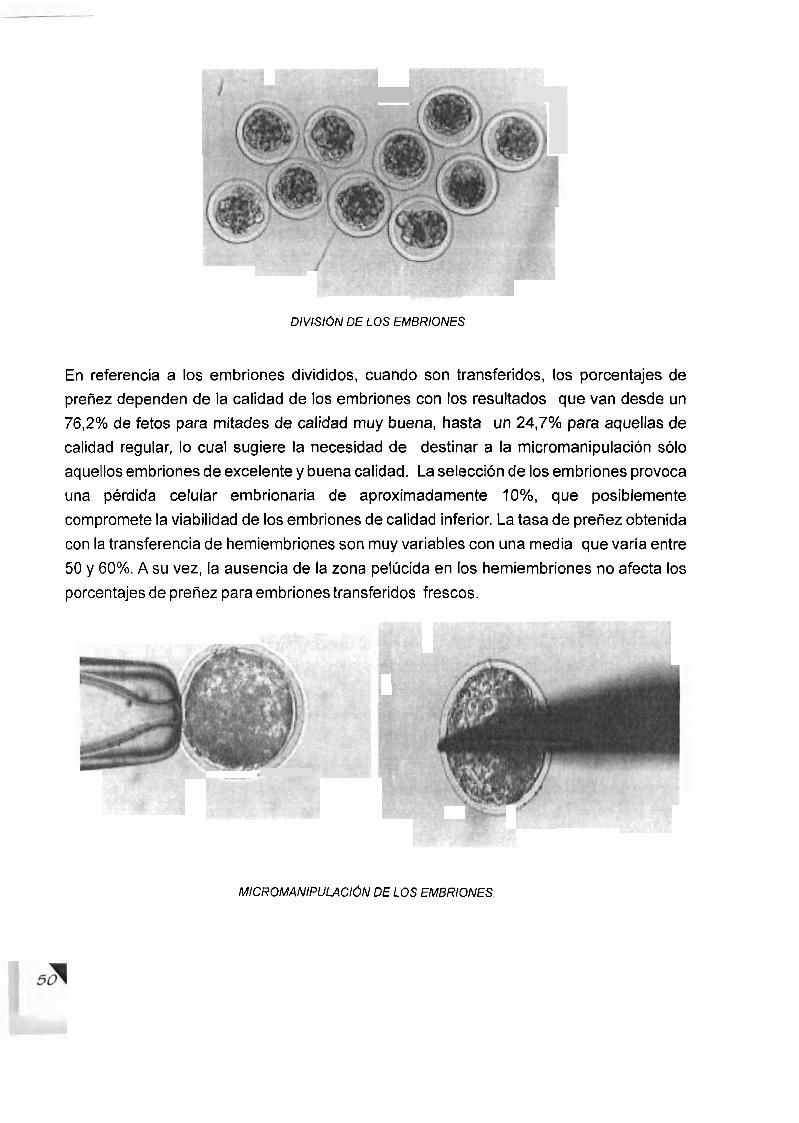
DlVlSl6N DE LOS EMBRIONES
En referencia a 10s embriones divididos, cuando son transferidos, 10s porcentajes de
preAez dependen de la calidad de 10s embriones con 10s resultados que van desde un
76,2% de fetos para mitades de calidad muy buena, hasta un 24,7% para aquellas de
calidad regular, lo cual sugiere la necesidad de destinar a la micromanipulacion solo
aquellos embriones de excelente y buena calidad. La selecci6n de 10s embriones provoca
una perdida celular embrionaria de aproximadamente lo%, que posiblemente
compromete la viabilidad de 10s embriones de calidad inferior. La tasa de preiiez obtenida
con la transferencia de hemiembriones son muy variables con una media que varia entre
50 y 60%. A su vez, la ausencia de la zona pelucida en 10s hemiembriones no afecta 10s
porcentajes de preiiez para embriones transferidos frescos.
M~CROMANIPULACI~N DE LOS EMBRIONES.

La division de embriones ha sido utilizada en diversos proyectos con la finalidad de
incrementar la eficiencia de programas de transferencia de embriones. La tecnica permite
incrementar el numero de crias obtenido por donadoras y generar pares de gemelos
identicos. El impact0 potencial de la tecnica en programas de mejoramiento genetic0 en
diversas especies de interes zootecnico ha sido ampliamente demostrado en diversos
paises del mundo. Asi mismo, diversos investigadores han reconocido 10s beneficios de
generar pares e gemelos identicos en la realizacion de investigaciones en ciencia animal.
4.1.7.-Ventajas de la bipartition de embriones
Actualmente 10s embriones bovinos son micromanipulados con el fin de obtener algunas
celulas y proceder a la identificacion del sex0 previo a su transferencia o para ser divididos
en dos partes iguales (hemiembriones) y producir mellizos identicos, lo que permite la
disponibilidad de animales identicos en 10s programas de transferencia embrionaria.
Con referencia a aquellos embriones que son destinados a la identificacion del sexo, 10s
blastomeros son abordados penetrando la zona pelucida y aspirando de 4 a 6 celulas o
cortando la zona y 10s blastomeros con una navaja. En trabajos realizados con embriones
sexados y transferidos frescos, la tasa de prefiez fue de alrededor de 45%, observandose
ademas que de 10s resultados dependieron de la calidad del embrion.

4.1.8.- Mellizos
El tener mellizos es el resultado inevitable de produccion en ganado lechero y algo
indeseable en la ganaderia lechera puesto que reduce la rentabilidad total y la eficiencia
reproductiva. Los mellizos reducen el resultado reproductivo al aumentar 10s dias abiertos
y servicios por concepcion durante la subsiguiente lactancia.
Las vacas que tienen mellizos presentan altos riesgos de nacer muertos, tendencia a
retencion placentaria, metritis y desplazamiento de abomaso. Las incidencias de aborto van desde un 12% vs. 29 %, la mortalidad neonatal de terneros 3.0 % vs. 15 %, teniendo un
bajo peso al nacer de 31 vs. 41 kg.; retencion placentaria que varia de 7 % vs. 34 %.
4.1.9.- Mecanismo de Gemelacion
El ganado es una especie monotoca, lo que significa que una preriez resulta en el nacimiento de un solo ternero. El mecanismo fisiologico responsable de regular el nllmero
de foliculos que se hacen dominantes dentro de cada curva folicular usualmente resulta
en la seleccion de un solo foliculo dominante capaz de ovular.
Sin embargo, se seleccionan dos foliculos para que continOen con su crecimiento entre el grupo de foliculos en crecimiento en cada curva folicular lo que resulta en un fenomeno que se denomina codominancia, cuando el estimulo se presenta en la ovulation ya sea en la forma natural o inducida, 10s foliculos codominantes, liberan dos oocitos uno de cada
7 foliculo. Si posteriormente son fertilizados por un esperma reciben el nombre de cigoto.
52

Por lo tanto 10s resultados de la ovulacion y fertilizacion de 10s dos oocitos se llaman mellizos dicigotos.
GEMELOS POR BIPARTICI~N DE EMBRIONES
La mayoria de 10s mellizos en el ganado son de tipo dicigoto. Los mellizos son dicigotos
pueden ser del mismo sex0 u opuesto y no son parecidos geneticamente ni
fenotipicamente que a 10s hijos nacidos de 10s mismos padres durante gestaciones
diferentes. Los mellizos que resulten de la ovulacion y fertilizacion de un solo oocito
reciben el nombre de mellizos monocigotos, el mecanismo por el cual ocurre la gemelacion
monocigota puede ser considerado una clonacion natural del cigoto original en vivo.
4.1 .lo.- Tasas de gemelacion en ganado lechero
Los factores de riesgo para la gemelacion en ganado incluyen efectos del tipo de raza
lecheras, el porcentaje de gemelos nacidos tambien varia entre las estaciones del at70 con
una tendencia hacia mas nacimientos de mellizos durante 10s meses de verano, este
efecto de las estaciones en la gemelacion ha sido atribuida a un increment0 en el plan de
nutricion durante el otofio cuando las vacas en paricion durante el verano habrian
concebido, un descenso del period0 de luz, y un descenso en la viabilidad de 10s embriones en fases tempranas concebidos durante 10s meses de verano comparados con 10s
concebidos durante 10s meses mas frios del otot7o. La alta produccion acumulativa de
leche y la gemelacion previa son factores adicionales que aumentan el riesgo de este.

En general la tasa de gemelacion para la mayoria de ganado de carne es menor de l l %. El efecto de paridad en la tasa de gemelacion no se extiende con claridad per0 se puede explicar por una habilidad incrementada devacas mayores para soportar gemelos durante la gestacion, un aumento en la tasa de doble ovulacion o una interaccion de ambos factores. Ademas, la incidencia de la doble ovulacion en las vacas lecheras en lactancia es de aproximadamente el 14 %. Tambien se sugiere que el proporcionar dietas de mayor energia metabolizable a las vacas de alta produccion puede incrementar la incidencia de doble ovulacion y por lo tanto la tasa de gemelacion. Este efecto nutricional es similar a la
Las tasas generales de gemelacion reportadas para vacas lecheras en estudios recientes
son (3%) mayores que las reportadas en muchos reportes previos, indicando que la tasa
de gemelacion puede estar aumentando en el tiempo en la poblacion de ganado lechero
como un todo. Si la gemelacion se relaciona con la nutricion ylo produccion de leche, este
aumento en gemelacion no seria inesperado considerando las tendencias recientes en
las practicas de alimentacion y 10s incrementos anuales en la produccion de leche por
vaca.
NAClMlENTO GEMELAR
4.1 .I1 .- Freemartinismo
El freemartinismo en vaquillas resulta de la gemelacion cuando las membranas
embrionicas de 10s fetos masculine y femenino se fusionan durante la gestacion lo que
causa un intercambio de sangre entre 10s fetos macho y hembra, 10s factores endocrinos o
las celulas del ternero macho causan un desarrollo anormal de 10s organos reproductivos
de la ternera hembra que causa infertilidad. El freemartinismo se presenta en
aproximadamente el 92 % de las vaquillas nacidas como resultado de preAez gemelar

heterosexual. Por lo tanto, aproximadamente 8 % de las vaquillas de prefiez gemelar
heterosexual sera fertil presumiblemente porque las membranas fetales no se fusionaron
o porque la fusion de la membrana se present6 despues del period0 critic0 de la
diferenciacion sexual (organos reproductores). El desarrollo temprano de anormalidades
del tracto reproductor femenino que resulta en freemartinismo se presenta entre los dias
49 a 52 posfertilizacion.

LITERATURA CITADA.
Aguilar M., Galina C., Merchant H., Montiel F.,Canseco R., Marquez YC. 2002.
Comparasion of stereoscopy , ligh microscopy and ultraestructural methods for evaluation of bovine embryos. Reprod. Dom. Anim. 37: 1-6.
Alonso, P. F. 2003. Teoria de costos dentro de la economia. Curso de actualizacion en
economia, administracion y comercializacion de productos ganaderos. Departamento de
administracion y economia, FMVZ-UNAM. 28-42
Alonso, P. FA., Bachtold, G. E.,Aguilar, V.A., Juarez G. J., Casas, P. VM., 1989. Economia
Zootecnica. 2a edicion, Edit. Limusa, Mexico D.F. 751.
~ lvarez, N. R. y T. R. De Armas. 1996. Manual practico, preparacion de medios de cultivo.
Centro de investigacion para el mejoramiento animal, ciudad de la Habana, Cuba. 1-27.
~ l v a r e z , S. F. 2004. Pr inc ip ios de economia agropecuaria htt://mx.geocities.com/feralvarezsiman.
Allen. R.L., Bondioli, K.R. and Wright, W. A Jr. 1982. The ability of fetal calf serum, new-born calf serum and normal steer serum to promote the in vitro develop-ment of bovine morulae. Theriogenology. 1 8: 1 85-1 89.
Amann. R.P., and J.O.. Almquist. 1957. Freezing of bovine semen. Effect of milk solids level, glycerol level, and fructose on freezability of bull spermatozoa in reconstituted and fresh skim-milk diluents. J. Dairy Sci. 40:1542-1549.
Arroyo, A.; Prat, M.1994. Direccion Financiera. 20a edicion, Edit. Deusto, Madrid, Esparia.
Aspron P. M. A. 1999. La tecnologia de transferencia directa de embriones. The World's source for bovine genetics.
Aspron, P. M. A. 1995b. Evaluacion de dos metodos de remocion del crioprotector (Glicerol) en embriones bovinos congelados. Comision Nacional para el Mejoramiento Genetico y la Reproduccion Animal, A.C.
Aspron, P .M.A. 1985. Transplante de embriones, seleccion y manejo (Transferencia de embriones en ganado bovino). Direccion General de Normatividad Pecuaria. SARH; Mexico, D.F.
Astiazaran J.R.and J. Longoria. 1988. Hartmann's solution as a medium for colletion and transfer of rat and cow embryos. Theriogenology. 29: 176-1 8 1
r;,

Austin,C.R. and Short, R.V. 1972. Reproduccion in mammals. Germ Cells and fertilitation. Cameridge University Press.
~ v i l a Garcia J. 1999. Resultados de la tecnica de congelacion y descongelacion de embriones en un solo paso. Vlll Curso lnternacional de Reproduccion Bovina. FMVZ- UNAN. 151 - 154.
~ v i l a , Garcia J. 2002. Nuevas estrategias aplicadas a la superovulacion y transferencia de embriones. "IX Curso Internacional de Reproduccion Bovina". FMVZ-UNAM. 71-75.
Bazer, F.W. 1983. Pregnancy an Parturition, J. Anim Sci., 57 2, pp. 425-460.
Betteridge, K. J. 1997. Embryo transfer in far0 animals. A review of techniques and applications. Ontorio: Canada Departament ofAgriculture.
Boland, M.P., Crosby, T.F. and Gordon, 1. 1989. Birth of twin calves following a single transcervical non-surgical egg transfertechnique. Vet. Rec. 99:274-275.
Bonilla, J.M. 1993. La competitividad de la economia espatiola: una aproximacion macroeconornica, cuadernos de informacion economica, fundacion FIES, 11 -1 9.
Brad K.S., D. 1990. Embryo transfer. Veterinary learning Co. Vol. 5 No. 5.
Broadbent, P.S. Stewart M. and Dolman, D.F. 1991. Recipients Management and Embryo Transfer. Theriogenology 35: 125-1 39.
Canals, J. 1991. Competitividad internacional y estrategia de la empresa, Ariel, Barcelona, Espatia.
Carpenter, J.F. y J.H. Crowe. 1988. The mechanism of cryoprotection of proteins by solutes, Ctyobiology25: 244-255 Cartelier, Jean. 1986. Excedente y reproduccion: la formacibn de la economia politica clasica, Mexico: Fondo de cultura economica.
Case, Karl E. Fair, Ray C. 1992. Fundamentos de economia. Edit. Prentice-Hall. Mexico, D.F
Chang Raymond y College Williams. 2003. Quimica. 7a edicion. Editorial McGraw- Hill, M6xicoD.F. 69-131.
CIBA. 1977. The freezing of mammalian embryos. Ciba Found. Symp. 52 (new series). Elseiver, Excerpta Medica, North-Holland,Amsterdam. 330 pp. Coelho, E. and Acevedo, N.A. 1991. Embryo transfer in zebu cattle. IX Congreso Brasileiro de Reproducao Animal; Belo Horizonte, Brasil; Vol. 2: 33-45.
Cseh, S., J. Seregi and L. Solti. 1994. Practical experience with direct transferred.' Theriogenology. 41 : 185-1 90.

Coss, B. R. 1999. Analisis y evaluacion de proyectos de inversion. edicion, Editorial Limusa. Mexico D.F. 91.
De Armas, T. R. 1996. Micromanipulacion de embriones. Centro de investigacion para el mejoramientoanimal, Habana-Cuba. 1-36. De la Fuente J., Fuentes S. y Hernandez F. 1999. Transferencia de embriones en programas de mejora genetica de vacuno de leche. Ed.: Luzan 5. ISSN: 11 30- 4804. Octubre de 1999. No 90. 55-65.
Dobrinsky JR, Pursel VG, Long CR, Johnson LA. 1997. Birth of normal piglets after cytoskeletal stabilization of embryos and cryopreservation by vitrification. Theriogenology. 49 (1): 345 abstr.
Dobrinsky JR. 1996. Cellular approach to cryopreservation of embryos. Theriogenology. 45 (1): 17-26.
Elsden, R.P. 1977. Embryo Transfer by surgical method. Embryo transfer in farm animal. Edited by K.J. Betteridge. Agriculture Canada. Monogradph 16:27-28.
Elsden, R.P., and G.E. Seidel, Jr. 1982. Magnament of Recipients im: embryo transfer
procedures for catle C.S.U. fort Collins Cd. USA.
Elsden, R.P., and Seidel G.E. 1982. Embryo transfer Procedures ford cattle, Colorado
state university.
Elsden, R.P. y Seidel. G.E. 1986. Manual de procedimientos para recoleccion, division y transferencia de embriones en bovinos. Colorado State University. Fort Collins Co.
Fahy GM, MacFarlane DR, Angell CA, Meryman HT. 1987. Vitrification as an approach to cryopreservation. Cryobiology. 2 1 : 407-426. Farrand, G.D. 1981. Identification, classification and handling of embryos. Short Course on EmbryoTransfer in Beef Cattle. Colorado State University, Ft, Collins Colorado.
Farrant. J. 1964. Pharmacological actions and toxicity of dimethyl sulphoxide and other compounds which proteet smooth musele during freezing and thawing. J. Pharm. Pharmacol. 16:472-483.
Fuentes, S. 1. 2002. Utilization de tratamientos hormonales para la sincronizacion de
receptoras de embriones, Aberekin, S.A.
Gallopin, G. C. 1996a. Environmental and sustainability indicators and the concept of situational indicators. Asystems approach, Environmental Modelling & Systems 1 (3 ) : 101 - 117.
Garcia., E. 1988. Modificaciones al sistema de clasificacion de Koppen. lnstituto de

Geografia. Universidad Nacional Autonoma de Mexico. Mexico, D.F.
Giussanim, P. 1993. La determinacibn de 10s precios de produccibn, politica y sociedad, n1 1411 5: 235-244.
Gonzalez, R. Velarde, J. C. Zambrano, S. 2000. Produccion y transplante de embriones congelados de bovinos criollo limonero. lnseminacion Artificial y Transplante de
Embriones, C.A. Universidad de Zulia. Facultad de Ciencias Veterinarias. Caracas,
Venezuela.
Goto K., Kajihara Y., Kosaka S., Koba M., Nakanishi Y., and Ogawa K. 1988. Pregnancies after co-culture of cljmulus cells with bovine embryos derived from in-vitro fetilization of in- vitro matured follicular oocytes. J ReprodFerfil. 83: 753-758.
Hafez, E. S. E.; Hafez, B. 2000. Reproduccion e lnseminacion Artificial en Animales. 7a Edicion, Editorial Mc Graw Hill Interamericana, Mexico, D. F.
Hasler, J. F. McCauley, A.D. Lathrop. W.F. y Foote R.H. 1987. Effect of donor-ernbryo- recipient interactions on pregnancy rate in a large-scale bovine embryo transfer program. Theriogenology. 27: 1 39.
Hernandez, C. J. 1999a. Ciclo estral en la hembra bovino. Mejoramiento animal, reproduccion bovino. SUA, FMVZ-UNAM. 23-31
Hernandez, C. J. 1999b. Sincronizacion de estros utilizando prostaglandina F2a ylo progestagenos. Mejoramiento animal, reproduccion Bovinos. SUA, FMVZ-UNAM. 11 3- 121.
Iglesias, C., Solano, R., Caral, J., Renard, S.T., Dzil, J.P. y Testart, J. 1981. Produccibn, recoleccion y transplante de embriones de la raza Charolais, bajo condiciones climatologicas tropicales. Rev. Cub. Reprod. Anim. Cuba. Vol. 7. No. 2.
INEGI .2000. lnstituto Nacional de Estadistica Geografica e lnformatica.
Ireland, J.J., and Roche, J.F. 1982. Development of antral follicles in cattle after prostaglandin-induced luteolysis. Changes in serum hormones, steroids in follicular fluid, and gonadotropin receptors. Endocrinology. 11 1 :2077.
Kay, R. D. 1986. Administracion Agricola y Ganadera. Planeacion, Control e Implementaci6n. Ed. CECSA.
Kesler, D. J. y Troxel, T.R. 1995. Estrus synchronization in cattle. Veterinary profesional topics. Cattle. 9: 1-6.
Kruger Heinze G. 1987. Preparacion de Equipo para Colecci6n y Transferencia de Embriones. Curso de Transferencia de Embriones ESSPA. Queretaro, Mexico.

Kumm, K. L. 2000. sustainability of organic meat production under swedish conditions. Agriculture, Ecosystems and Environment 88: 95-1 01.
Leibo, S. P. 1982. A one-step method for direct non-surgical transfer of frozen-thawed bovine embryos. En: Proc. Int. Congr. EmbryoTransfer in Mammals.Annecy, France. 97.
Leibo, S. P. 1983. Embryo transfer method and apparatus. Rio Vista Int., Inc., assignee. Us Pat No. 4:380,997.
Leibo, S. P. 1986. Commercial production of pregnancies from one-step diluted frozen- thawed bovine embryos. Theriogenology 25: 166.
Leon, V. H., Hernandez C.J., Keisler D.H. y Gutierrez C.G. 2003. Relacion entre 10s cambios de la condicion corporal y las concentraciones plasmaticas de Leptina, IGF-I e insulina en vaquillas de doble proposito. FMVZ-UNAM, Mexico, D.F.
Li, R., Cameron, A.W.N., Batt, P.A. and Trounson, A.O. 1990. Maximum survival of frozen goat embryos is attained at the expanded, hatching and hatched blastocyst stages of development. Reprod. Fertil Dev2: 345-350.
Lindner, G.M. and Wright, R.W. 1983. Bovine embryo morphology and evaluation. Theriogenology. 20:407-4 1 6.
Lovelock, J.E. 1953. The mechanism of the protective action of glycerol against haemolysis by freezing and thawing. Biochim. Biophys . Acta. 11 :28-36.
Mac lntosh and L.N. Hazeleger. 1994. The use of ethyleneglycol for freezing bovine embryos. Theriogenology. 41 :253.
Macmillan, K.L., and Burke CR. 1991. Effects of oestrus cycle on reproductive efficiency. Anim Reprod Sci. 42:3007:-20.
Mahmoudzadeh, A.R., A. Van Soom, I. Van Vlaenderen and A. De Kruif. A. 1994. Comparative study of onestep addition of different vitrification solutions on in vitro survival of vitrified bovine embryos. Theriogenology. 39: 1291 -1 302.
Mahon, G.D. y Rawle, J.E. 1987. The export of deep frozen bovine embryos. Theriogenology. 27 :2 1 -35.
Manzur, P. 1977. The role of intercellular freezing in the death of cells cooled at supraoptimal rates. Crybiology. 14:251-272.
Marquez, YC, Galina CS, Castilla B, Leon H, Moreno MN. 2004. Evidence of Damage in Cryopreserved and Fresh Bovine Embryos Using the Tunel Technique. Reprod Dom Anim. 39:141-145.
Massip. A. and Der Zwalmen. 1984. Direct transfer of frozen cow embryos in glycerol- sucrose. Vet. Rec. 11 5: 327-328.

Megahed G. A. 1995. The influence of season and embryos stage on pregnancy rate of
frozen thawed cow embryos. Indian Vef. J. 72: 146-149.
Meryman, H.T. 1966. Cryobiology. Academic Press, London and New York. 1-114 Montiel, P.F. 1996. Irnplementacion de un programa de transferencia de embriones Holstein - Cebu, criopreservados, sexados y fertilizados in vitro con productores del tropic0 hljmedo. Tesis de Maestria FMVZ, UNAM.
Noriega S. R. Martinez B. S. y Flores C. E. 1995. Tecnicas de procesamiento de embriones para la transferencia en bovinos, UNAM, Mexico, D.F.
Nljiiez, B. J., y Solano F. R. 1996. Congelacion de ovocitos y embriones en animales domesticos. Centro de Investigacion para el Mejoramiento Animal (CIMA), La Habana Cuba, 45.
NRC. 1988. Nutrient Requeriments of Dairy Cattle. Sixth Edition. National Academy Press. Washington, D.C.
Odde, K.G. 1990. A Review of synchronization of estrus in postpartum cattle. J. Amim. Sci. 68 :817.
Pollard JW, Leibo SP. 1994. Chilling sensitivity of mammalian embryos.Theriogenology. 41:lOl-106. Porras. A. A. 2000. Pubertad y estacionalidad reproductiva en ovinos. Departamento de reproduccion. FMVZ - UNAM. 8 - 18
Rangel, S. R. 1998. Aplicacion de tecnicas biotecnologias en la reproduccion de ovinos y
caprinos. Programa de Maestria en Reproduccion Animal, Chapingo, Mexico. 1-1 94
Reagan Broks. 1993. Transferencia de Embriones. Aplicaciones para un programa exitoso. Texas ARM. University Collage Station, Texas, EE.UU.
Reglas de Operacion Alianza Contigo. 2003. Subsecretaria de Desarrollo Rural (SAGARPA). www. Sagarpa.gob. mxlsdrl-26 k. DOF 25/07/03. 44,45,46.
Renard, J.P., Heyman, Y. and Ozil, J.P. 1982. Congelation de I'embryon bovin: une nouvelle methode de decongelation pour le transfert cervical d' embryons conditionnes une seule fois en paillettes. Ann. Med. Vet. 126 : 23-32.
Rivera S. R. 1987. Fertilizacion y desarrollo embrionario. Curso de Transferencia de Embriones ESSPA. Queretaro, Mexico.
Rowe, F.R. 1982. Nonsurgical transfer of bovine embryons. Proceeding Owners Managens Wonrkshop. Vlll th Ann. Meet Ing. Embryo Transfer Soc. 57-61, Denver, Colorado.

Ruiz, M. A. 1999. Analisis retrospectivo de la fertilidad de embriones descongelados en el ganado bovino, bajo condiciones de tropico. Tesis de Licenciatura, FMVZ, UNACH.
Saito N. DVM. 1994. Manual of embryo transfer & in vitro fertilization in cattle. National Livestock Breeding Center, MAFF, Japan.
Sabino Carlos. 1991. Diccionario de Economia y Finanzas, Economia de Escala. Edit. Panapo, Caracas Venezuela.
Salisbury, G. W.; van Demark, N.L.; Lodge, J.R. 1978. Fisiologia de la reproduccion e inseminacion artificial de 10s bovinos, 2Oedicion. EditorialAcriba, Zaragoza, Espaiia. 831.
Sanchez Sanz R. 2000. Enciclopedia Microsoft Encarta. Masa molecular y disoluciones.
Saroff. J., and J.P. Mixner. 1955. The relationship of egg yolk and glycerol content of diluters and glycerol equilibration time to survival of bull spermatozoa after low temperature freezing. J. Dairy Sci. 38:292-297.
Seidel, G.E., Jr, Seidel, S.M. and Bowen, E. A. 1980. Bovine Embryo Transfer Procedures. C.S. U. Exp. Sxp. Stat. and Reprod. Lap. General Series 975, Fortdallins, Colorado.
Seidel, G. E., Jr. 1977. Short-term maintenance and Culture of Embryos. Embryo transfer in Farm animals. Edited by K. J. Betteridge. Agriculture Canada Mohograph. 16:20-24.
Seidel, G. E., Jr. 1985. Principles of cryopreservation of mammalian embryos. Short couse techniques freezing mammalian embryos, Colorado States University.
Solano, R. 1988. Congelacion de embriones bovinos I. Efecto de la calidad y estadio de desarrollo durante la congelacion y descongelacion. Rev. Cub. Ciencia Vet. 19(3): 183-1 92.
Sreenan, J.M. y Diskin, M.G. 1987. Factors affecting pregnancy rates following embryo transfer in the cow. Theriogenology. 27:99- 11 3.
Steelflorrie. 1998. Bioestadistica. Principios y procedimientos. 2a edicion, Edit. Mc GrawHill. Mexico, D. F. 458-464.
Stein, W. D. 1962. Spontaneous and enzyme-induced dimmer formation and its role in membrane permeability. The mechanism of movement of glycerol across the human erythrocyte membrane. Biochim. Biophys. Acta. 59:47-65.
Steinbach. J. and R.H. Foote. 1967. Osmotic pressure and pH effects on survival of frozen bovine spermatozoa. J. Dairy Sci. 50:205-213.
Suzuki, T., M. Yamamoto, M. Ooe,A. Sakata M. Matsuoka, Y. Nishikata, K. Okamato. 1990. Effect of sucrose concentration used for one-step dilution upon in vitro and in vivo survival of bovine embryos refrigerated in glycerol and 1,2-propanediel. Theriogenology. 34: 1051 - 1057.

Thompson JG. 1996. Defining the requirements for bovine embryo culture. Theriogenology.
45: 27-40.
Trueta, S. R. 2003. Los cocientes entre precios. Curso de actualizacion en economia,
administracion y comercializacion de productos ganaderos. Departamento de administracion y economia, FMVZ-UNAM. 18-26
Valencia. 1999.Transferencia de embriones en bovinos. Mejoramiento Animal, Reproduccion Bovinos. SUA. FMVZ- UNAM. 205-21 8
Vivanco, H.W. 1998. Transferencia embrionaria en ovinos y caprinos. Centro de
lnvestigacion en Ruakuna, Nueva Zelanda.
Voelkel, S.A., Arntson-Morcan, K.C. and Godke, R.A. 1981. Heat inactivation of lamb, newborn calf and steer sera for potential use in embryo transplant procedure. Theriogenology. 1 5: 1 25-1 28.
Voelkel, S.A.and Hu, Y.X. 1992. Direct transfer of Frozen-Thawed bovine embryos. Theriogenology. 23 - 37.
Watson PF, Morris GJ. 1987. Cold shock injury in animal cells. In: Temperature and Animal Cells (K. Bowler and B.J. Fuller, Eds.). 311-340.
Whittingham, D.G.; Leibo, S.P. and Manzur, P. 1972. Survival of mouse embryos frozen to -196 "C and -269 "C, Science 178: 411-414 Wildman,E.E., Jones, G.M. and Wagner, P.E. 1982. Dairy body condition scoring system and its relationship to selected production characteristics. J. Dairy Sci. 65: 495.
Wilmut, 1. 1972. The effect of cooling rate, warming rate, cryoprotective agent and stage of development on survival of mouse embryo during freezing and thawing, Life Sci. 11 part 2: 1071 -1 079 Wiltbank, J.N. 1969. Short breeding periods with the aid estrus synchronization. Proceeding of the Range Beef Cow. ASymposium on Production. Chadron Nebraska.
Wright, J.M., 1981. Non-surgical embryo transfer in cattle embryo-recipient interactions. Theriogenology. 1 5:43-56.
Yen, S.C.; Jaffe, B.R.; Barbieri, L. R. 2001. Endocrinologia de la Reproduccion. Fisiologia, fisiopatologia y manejo clinico, Editorial Panamericana, 91 1


















