
Trabajo de Grado para optar por el Título de LICENCIADO EN ...

i
UNIVERSIDAD NACIONAL PEDRO RUIZ GALLO
FACULTAD DE CIENCIAS BIOLÓGICAS
Caracterización de la variabilidad genética e identificación de
marcadores genéticos en accesiones silvestres del Jatropha curcas
de la costa norte del Perú.
Tesis
Para optar el título Profesional de:
Licenciado en Biología
Autor
LUIS MIGUEL SERQUÉN LÓPEZ
Asesor
PEDRO CHIMOY EFFIO, Ph.D.
Lambayeque-Perú, 2010

ii
UNIVERSIDAD NACIONAL PEDRO RUIZ GALLO
FACULTAD DE CIENCIAS BIOLÓGICAS
TESIS
(CARÁTULA)
Titulo: Caracterización de la variabilidad genética e
identificación de marcadores genéticos en
accesiones silvestres del Jatropha curcas de la
costa norte del Perú.
Autor: LUIS MIGUEL SERQUÉN LÓPEZ
Patrocinador: PEDRO CHIMOY EFFIO, Ph.D.
Co-Patrocinador: LUIS DESTÉFANO BELTRÁN, Ph.D.
Aprobado por:
……………………………….. ……………………………………
PRESIDENTE DEL JURADO MIEMBRO DEL JURADO
………………………………… ……………………………………..
MIEMBRO VOCAL JEFE CENTRO INVESTIGACIÓN
…………………………………. ………………………………….......
JEFE DEPARTAMENTO DECANO
Lambayeque,….de…………….del…...

iii
La ciencia y tecnología ayuda a construir un país con esperanza; puesta en un grupo de personas, acaso dignas, del progreso y desarrollo común. Hay mucho que hacer, ya que los avances científicos llegan tarde para muchos, justo a tiempo para pocos y temprano para nadie
Mientras más grande la prueba Más glorioso es el triunfo
AGRADECIMIENTOS

iv
Índice
A una hermosa Mujer a quien todos llaman Betty
López Vásquez -M y que tengo el gran privilegio
de decirle MAMÁ
A mi Papá Pedro, mi Hermana Jessica, y mis Sobrinos
Jonathan y Gabriel, que Junto a María Angélica me soportan y
dan vida a cambio de tiempo.
A mis amigos de la UNPRG y SJO que si comienzo a
nombrarlos temería olvidarme de un solo nombre, pero que
saben de mi respeto y estima.
Al Dr.Pedro Chimoy, Luis Destefano y Betty Millán
”Mentores”, por la confianza puesta en mi y todo su apoyo
para lograr la culminación de esta Tesis.
Financiamiento
La financiación del Proyecto de la Tesis está dada por un convenio entre la Universidad
Peruana Cayetano Heredia (UPCH) e INCAGRO mediante el subproyecto:
Caracterización de la variabilidad genética y del contenido de aceite del la semilla del
piñón blanco (Jatropha curcas L) con fines de producción de biodiesel en la costa norte y
en la región San Martín y Ucayali.
A JHWH por darme la vida y su amor;
Porque la fe ilumina la ciencia,
don del espíritu de Dios
De manera especial a: Mónica, Cecilia, Marianella, Paty,
Diana, Julio, Jerisf, Antoine, Savina, German, integrantes
del (GRU), por motivarme a seguir en todo momento y hacer
especial mi estadía en el LID-UPCH
A Giankarlos Vásquez , Ricardo Plaza Alexander
Huaman y Miguel Machahua, amigos y compañeros de
largas y entretenidas tertulias,
¡A todos ellos éxitos y bendiciones a lo largo de toda su vida!

v
I. Resumen 1
II. Introducción 2
III. Antecedentes 3
3.1 Biodiesel como fuente de energía renovable. 3
3.2 Jatropha curcas L. ―Piñón‖ 7
3.2.1 Posición taxonómica de Jatropha curcas. 9
3.2.2 Usos de Jatropha curcas L. 10
3.2.3 Toxicidad de J. curcas 10
3.2.4 Jatropha curcas como biodiesel 11
3.3 Variabilidad Genética y Caracterización de las especies vegetales 11
3.3.1 Fuentes de variabilidad genética. 12
3.3.2 Perdida de la variabilidad genética. 13
3.3.3 Polimorfismo. 14
3.3.3.1 Polimorfismo en regiones génicas codificantes. 15
3.3.3.2 Polimorfismo en regiones génicas no codificantes. 15
3.3.3.3 Polimorfismo en regiones no génicas. 16
3.4 Marcadores genéticos . 16
3.4 .1 Definición. 16
3.4.2 Clases de Marcadores genéticos. 16
3.4.3 Marcadores moleculares. 18
3.4.3.1 Ventajas de los marcadores moleculares. 18
3.4.3.2 Marcadores basados en la hibridación de sondas. 19
3.4.3.3 Marcadores basados en PCR (reacción en cadena
de la polimerasa). 19
3.4.3.4 Clasificación de los marcadores moleculares basados
en PCR. 20
3.3.4 Marcadores Inter-Secuencias Simples Repetitivas (ISSRs). 20
3.3.5 Random Amplified polimorphic DNA (RAPDS). 21
3.5 Variabilidad genética y filogenia del genero Jatropha. 22
3.6 Diversidad genética intraespecífica en J. curcas. 23
3.7 Marcadores de variedades tóxico y no tóxico de J. curcas. 24
IV. Objetivos 24
4.1 Objetivos Generales. 24
4.2 Objetivos específicos. 24
4.3 Lugar de ejecución del proyecto. 24
V. Materiales y Metodología 25
5.1 Materiales. 25

vi
5.1.1 Material biológico. 25
5.1.2 Equipos. 25
5.1.3 Reactivos. 25
5.1.4 Otros accesorios. 26
5.2 Metodología. 26
5.2.1 Colecta de las accesiones. 26
5.2.2 Extracción de ADN. 27
5.2.3 Calidad y concentración del ADN. 28
5.2.4 Selección de los individuos para el estudio. 28
5.2.5 Pre-selección de iniciadores ISSR y RAPD. 28
5.2.6 PCR con los 60 individuos del estudio. 29
5.2.6.1 Condiciones PCR – ISSR. 29
5.2.6.2 Condiciones PCR – RAPD. 29
5.2.7 Electroforesis en gel de agarosa. 30
5.2.8 Construcción de la Matriz Básica de Datos y determinación
de valores descriptivos. 30
5.2.9 Análisis Multivariado. 31
5.2.10 Validación Interna. 32
VI. Resultados 33
6.1 Colecta y conservación de los genotipos de Jatropha curcas L. 33
6.2 Extracción y Calidad de ADN. 33
6.3 Elección de los individuos. 33
6.4 Elección de los iniciadores. 35
6.5 Amplificación con los marcadores ISSR y RAPDs. 35
6.6 Análisis Multivariado. 37
6.6.1 Agrupamiento UPGMA. 37
6.6.2 Coeficiente de correlación (r). 38
VII. Discusión 39
7.1 Elección de los genotipos. 39
7.2 Condiciones del ADN y la PCR 39
7.3 Valores descriptivos de los marcadores 40
7.4 Agrupamiento 41
7.5 Validación interna 43
7.6 Análisis Poblacional 43
7.6.1 Porcentaje de loci polimórficos. 43
VIII. Conclusiones 44
IX. Recomendaciones 45
X. Referencias Bibliográficas 46
XI. Anexos 50

vii
Anexo N°1 Datos de colecta realizada en distintas localidades
de la región Lambayeque, Piura y Tumbes. 50
Anexo N° 2 Electroforesis del ADN de muestras de las regiones
Piura, Tumbes y Lambayeque. 58
Anexo N° 3 Lista de los 53 Iniciadores ISSR y 12 RAPD. 62
Anexo N° 4 Datos Binarios (presencia: 1 ó ausencia: 0) de 93 loci
(caracteres) para 60 genotipos (OTUs). 63
Anexo N° 5 Matriz de similitud de la comparación uno a uno
entre las 60 OTUs. 67
Anexo N° 6.
6.1. Fenograma UPGMA generado con los 60 individuos
usando los marcadores ISSR y el coeficiente
Simple Matching. 72
6.2. Fenograma UPGMA generado con los 60 individuos
usando los marcadores RAPD y el coeficiente
de similitud Simple Matching. 73
Anexo N° 7. 74
7.1.- Representación gráfica de la correlación entre la
Matriz de Similitud y la Matriz cofenética para
los marcadores RAPD. 74
7.2.- Representación gráfica de la correlación entre la
Matriz de Similitud y la Matriz cofenética para los
marcadores ISSR. 75
Anexo N°8. Extracción de ADN por el método cetil trimetil amonio
bromuro (CTAB). 76
LISTA DE ABREVIATURAS

viii
A Adenina
ADN Ácido desoxirribonucleico
AFLP Polimorfismo de longitud de fragmentos amplificados (Amplified Fragment
Lenght Polymorphism)
CAPS Secuencia polimórfica amplificada y cortada. (Cut Amplified Polymorphic
Sequence)
cM centimorgan
CO2 Dióxido de carbono
CONAM Consejo Nacional del Ambiente
CTAB Bromuro de hexadeciltrimetilamonio
dATP 2’-desoxiadenosina-5’-trifosfato
dCTP 2’-desoxicitidina-5’-trifosfato
dGTP 2’-desoxiguanosina-5’-trifosfato
dTTP 2’-desoxitimidina-5’-trifosfato
dNTP 2’-desoxirribonucleósido-trifosfato
EDTA Ácido etilendiaminotetraacético
G Guanina
Gg Giga galones
GPS Sistema de Posicionamiento global (Global Positioning System)
ISSR Inter- microsatelites (Inter Simple Sequence Repeat)
MAS Selección asistida (Marker Assisted Selection)
Min Minutos
MJ Megajulio equivalente a 239 Kilo Calorias
mM Mili Molar
NOX óxidos de nitrógeno
ng Nanogramos
OTU Unidades taxonómicas operacionales (Operational taxonomic units)
PAA Poliacrilamida
pb Pares de bases
PCR Reacción en cadena de la polimerasa (Polymerase Chain Reaction)
PIC Índice de contenido polimórfico (Polymorphic Index Content)
PVP Polivinilpirrolidona
QTL Locus de un carácter cuantitativo
RAPD Amplificación de fragmentos de ADN usando partidores de diseño
aleatorio (Random Amplified Polymorphic DNA)
RF Frecuencia de recombinación
RFLP Polimorfismo en la longitud de fragmentos de restricción. (Restriction
Fragment Length Polymorphism)
RNA Ácido ribonucleico
RNasa RNA nucleasa
Rpm Revoluciones por minuto
SAMPL Amplificación selectiva de loci polimórficos de microsatélites. (Selective
Amplification of Microsatellite Polymorphic Loci)
SCARs Regiones amplificadas de secuencias caracterizadas (Sequence
characterized amplified region)
SNP Polimorfismo de un solo nucleótido (Single-nucleotide polymorphism)
SSR Microsatélites (Simple Sequence Repeat)
T Timina
Tris-HCl Hidrocloruro de tris(hidroximetil)-aminometano
TAE TRIS-HCl + Acido Acético Glacial + EDTA (Tris base, acetic acid and EDTA)

ix
U Unidades enzimáticas
ul Micro litros
UPGMA Sin ponderar Grupo Par Método con media aritmética (Unweighted Pair Group
Method with Arithmetic Mean)
UV Ultra violeta
GLOSARIO

x
ADN (Acido desoxirribonucleico). Base molecular de la herencia. Una molécula de ADN es
un polímero de desoxinucleótidos ordenados en una doble hélice (dos hebras) de azucares
fosfatados con bases purinas y pirimidinas proyectándose al centro. Cada hebra se mantiene
unida por enlaces fosfo diester entre los carbonos 3' y 5' de nucleótidos adyacentes. De
acuerdo al modelo de Watson y Crack, el ADN forma una doble hélice que es mantenida
unida por enlaces de hidrogeno entre pares de bases (doble enlace entre timina y adenina y
triple enlace entre citosina y guanina), por tanto cada hebra en la doble hélice es
complementaria en su secuencia.
ADN genómico. ADN que incluye toda la secuencia de nucleótidos contenida en el genoma
de un individuo. Término utilizado para referirse al ADN extraído mediante el use de
proteinasa K, detergentes (SDS) y solventes (fenol). Con este método se obtienen fragmentos
de 100 a 150 kb adecuados para la producción de genotecas mediante una digestión previa
con enzimas de restricción. A nivel práctico el ADN extraído incluirá el ADN nuclear
(cromosomal) y ADN mitocondrial.
AFLP- amplified fragment-length polymorphism. Polimorfismo de la longitud de
fragmentos amplificados. Es una variante de la técnica de la huella genética (DNA
fingerprinting) que se basa en la amplificación selectiva, mediante una reacción en cadena de
la polimerasa (PCR), de fragmentos procedentes de la digestión de un ADN genómico con un
par de enzimas de restricción.
Alelo. Cada una de las posibles formas en las que existe un gen a consecuencia de una o más
mutaciones.
Bioinformática. Informática aplicada a la recolección, al almacenamiento y al análisis de
datos biológicos
Biotecnología. Utilización de organismos vivos (usualmente microorganismos) o de algunos
de sus constituyentes (generalmente enzimas) con fines industriales; ello incluye desde las
tradicionales técnicas de fermentación industrial hasta, en los últimos tiempos, la utilización
de plantas y animales transgénicos para producir proteínas recombinadas con fines
alimentarios o terapéuticos.
Clonación de ADN. Técnica que permite que secuencias individuales de ADN en mezclas
complejas sean aisladas y copiadas permitiendo su manipulación y análisis detallado. El ADN
a clonar es recombinado con ADN de un vector e introducido en una célula hospedera donde
será copiado.

xi
Desnaturalización del ADN. Separación de las dos hebras que constituyen al ADN por
rompimiento de los enlaces de hidrogeno entre bases complementarias. El ADN se
desnaturaliza a temperaturas superiores a 90° C. Primera etapa en la PCR.
dNTPs (desoxinueleotidos). Nucleótidos cuya pentosa es una desoxirribosa y por ende son
los monómeros del ácido desoxirribonucleico (ADN).
Endogamia. Apareamiento de organismos relacionados entre si, que trae consigo un aumento
en la homocigosidad y posible expresión de genes deletéreos.
Enzima de restricción. Enzimas de origen bacteriano que cortan moléculas de ADN extraído
en sitios específicos de reconocimiento. Su nomenclatura se basa en una abreviatura de la
bacteria donde fueron aisladas, seguida por una numeración romana que da el orden
cronológico de su descubrimiento.
Extensión del ADN. Tercera etapa en la PCR. La temperatura es elevada a 72° C para
permitir que una ADN polimerasa inicie la síntesis a partir del iniciador agregando dNTPs
libres sobre la hebra que sirve de templado.
Genética. Rama de la Biología en la que se estudia la herencia y variación de los caracteres,
así coma la estructura y función del material genético (ADN).
Genética cuantitativa. Rama de la Genética en la que se analizan las características
fenotípicas que pueden ser medidas en una escala de manera continua, y que son determinadas
por la acción aditiva de múltiples genes.
Genética de poblaciones. Rama de la Genética en la que se estudia la variabilidad de las
poblaciones naturales, expresada en frecuencias genotípicas y alélicas de ciertos marcadores
moleculares, así como los mecanismos que la hacen variar en generaciones consecutivas.
Gen. Unidad hereditaria que contiene información genético (ADN) que es transcrita a ARN y
traducida a una cadena polipeptidica (proteína). En el genoma de un organismo, una secuencia
de nucleótidos a la que se le puede asignar una función especifica.
GenBank (Banco génico o genómico). Base de datos de libre acceso de secuencias
nucleotidicas y polipeptidicas, administrada por el "National Center for Biotechnology
Information" (NCBI). www.ncbi.nlm.nih.gov/Genbank!GenbankSearch.html
Genoma. Término utilizado para referirse a todos los genes (información genético) contenida
en un solo gameto, es decir un representativo de cada cromosoma.
Genoteca. Colección de fragmentos de ADN de una determinada especie insertados al azar en
vectores (tales como plásmidos o fagos lambda) y clonados en hospederos (ej. bacterias). La
colección debe de ser lo suficientemente grande para incluir toda la secuencia nucleotidica del
genoma.

xii
Genoteca parcial o librería subgenomica. Colección incompleta de ADN de un determinado
individuo la cual se origina al seleccionar fragmentos de ADN de un específico rango de
tamaños.
Genotipo. La constitución genética de un organismo, que estará dada por la información
heredada por ambos padres. Específicamente para un solo locus, el genotipo estará dado por
los alelos heredados por ambos padres (ver homocigoto, heterocigoto).
Haplotipo. Contracción de la expresión «genotipo haploide»; se refiere a la constitución
genética de un cromosoma individual. En el caso de organismos diploides, el haplotipo
contendrá un miembro de la pareja de alelos para cada locus. Puede referirse a un conjunto de
marcadores (p. ej., polimorfismos de un solo nucleótido − SNP) que estadísticamente están
asociados a un único cromosoma. Sabido esto, se cree que la identificación de unos cuantos
alelos de un bloque de haplotipo puede identificar de manera inequívoca el resto de loci
polimórficos de la región. Dicha información es de gran utilidad para investigar la genética de
los caracteres complejos.
Heterocigosis. Proporción de loci heterocigóticos en animales cruzados comparados con los
valores dentro de cada raza.
Heterosis. Ventaja de los animales cruzados sobre los animales de raza pura (ver sección
Razas, cruzas y VGEs de animales cruzados).
Información genómica. La información genómica se obtiene directamente de análisis de la
estructura del material genético de los animales (ADN).
Heterocigosidad. La proporción de individuos heterocigotos para un locus o en promedio
para varios loci. Es un estimador de variabilidad genética en poblaciones naturales o
domesticadas.
Heterocigosidad esperada. Proporción de organismos heterocigotos calculada a partir del
equilibrio de Hardy-Weinberg. Su cálculo implica la obtención de las frecuencias genotípicas
a partir de las frecuencias alélicas siguiendo un binomio al cuadrado (p + q)2, en donde p y q
son las frecuencias alélicas y 2pq corresponderá a la frecuencia de heterocigotos.
Heterocigosidad observada. Proporción de organismos heterocigotos calculada a partir de los
genotipos observados en una muestra poblacional.
Heterocigoto. Un individuo diploide o poliploide que ha heredado diferentes alelos en uno o
más de sus loci.
Heteroduplex. ADN de doble hebra en el cual cada cadena tiene un origen diferente y par
tanto no son completamente complementarias.

xiii
Homocigoto. Un individuo que ha heredado el mismo alelo de ambos padres para un
determinado locus.
Iniciador (del inglés Primer). Oligonucleótido (alrededor de 20 pb) de una sola hebra que se
alinea (homologa) para iniciar la replicación del ADN mediante una polimerasa. Se requieren
dos iniciadores (uno para cada hebra de ADN), los cuales flanquearan la región por
amplificar.
Loci. Plural de locus.
Locus. Lugar específico del cromosoma en el que se ubica un gene.
Mapa genético. Arreglo lineal de sitios mutables (polimórficos) sobre un cromosoma,
deducidos de experimentos de recombinación genética. Implica un ordenamiento de
marcadores y genes estableciendo distancias genéticas entre los mismos.
Marcador genético: un polimorfismo del ADN que se puede detectar fácilmente mediante
análisis fenotípico o molecular. El marcador puede hallarse dentro de un gen o en un ADN sin
función conocida. Dado que los segmentos de ADN que se encuentran próximos entre sí en
un cromosoma tienden a heredarse juntos, los marcadores se suelen utilizar como maneras
indirectas de seguir la pista del patrón de herencia de un gen que aún no se ha identificado,
pero cuya localización aproximada sí es conocida.
Marcador molecular. Genes, proteínas o fragmentos de ADN polimórficos que permiten
distinguir entre individuos, familias, poblaciones o especies.
MAS. (del inglés Marker Assisted Selection). Selección asistida por marcadores. Estrategia
para auxiliar en el mejoramiento genético seleccionando a los reproductores o progenies en
los primeros estadios de crecimiento en base a un marcador molecular que esté relacionado
con un mayor desempeño. En estos casos se eliminarán los organismos que porten marcadores
que actúen en detrimento de la producción (los mas pequeños, los menos resistentes), la
ventaja del MAS comparado con el mejoramiento tradicional es que no será necesario crecer a
los organismos para darse cuenta de si tienen buen o mal desempeño sino desde un principio
con un análisis de ADN serán seleccionados.
Microsatélite. Regiones de ADN con secuencias cortas de una a seis pares de bases (pb)
repetidas en tándem, es decir la misma secuencia una tras otra, llegando a medir toda la región
hasta 100 pb. Se analizan mediante amplificaciones por PCR, para lo cual es necesario diseñar
iniciadores flanqueantes, lo cual representa una identificaci6n previa del microsatélite en la
especie en cuestión implicando un proceso de clonación y secuenciado. También son
conocidos como SSRs (del inglés Simple Sequence Repeats), STRs (del inglés Short Tandem

xiv
Repeats) 0 VNTRs (del inglés Variable Number Tandem Repeats), esté Último término
incluyendo a los minisatélites.
Minisatelite. Los minisatelites son secuencias de 10 a 100 pb repetidas en tandem en el
genoma. El número de repeticiones normalmente es inferior a 1000, abarcando una región de
102 a 10' pb. También pueden ser referidos como VNTR (del inglés Variable Number of
Tandem Repeats). Los minisatélites usualmente son analizados al digerir el ADN con enzimas
de restricción, para posteriormente transferir a una membrana (Southern blot) y realizar una
hibridación con la sonda minisatélite marcada.
QTL: Loci de efecto cuantitativo, son genes con efectos mayores en características afectadas
por muchos loci con efectos generalmente pequeños (poligenes).
PCR (del inglés Polymerase Chain Reaction). Reacción en cadena de la polimerasa. Técnica
con la que se copian las hebras complementarias de una molécula de ADN blanco
simultáneamente a través de una serie de ciclos, que incluyen una desnaturalización del ADN,
alineamiento de iniciadores y extensión (síntesis) mediante una polimerasa (comúnmente Taq
polimerasa). 30 ciclos producirán un factor de amplificación de 100 millones. Técnica
desarrollada por Mullis y col. en 1985.
Plásmido. ADN bacteriano extracromosomal en el que se codifican genes que le confieren a
la bacteria ciertas propiedades (resistencia, fertilidad, virulencia, etc.). Mantienen una
replicación independiente al ADN cromosomal.
Población. Grupo de organismos de la misma especie que habitan una determinada área
geográfica y que por ende se entrecruzan y comparten un acervo común de genes. Ver stock.
Polimorfismo: Es la digestión de un segmento de ADN (o el genoma completo del ADNmt)
por enzimas de restricción que producen fragmentos de una longitud particular, dependiendo
de la colocación del sitio de restricción (p. ej., la enzima Msel corta todos los sitios
'TTAA').Una mutación a un sitio de restricción, impediría la digestión de la enzima y de esta
manera se generarían fragmentos de longitud diferente.
RAPD (del inglés Random Amplified Polymorphic DNA). ADN Polimórfico Amplificado al
Azar. Técnica basada en el uso de PCR, en donde un solo iniciador (de alrededor de 10 pb)
diseñado arbitrariamente puede amplificar fragmentos polimórficos de ADN flanqueados por
dos sitios complementarios de alineamiento en orientación invertida, es decir el mismo
iniciador se une a ambas hebras de ADN.
RFLPs (del ingles Restriction Fragment Length polymorphism). Polimorfismo de la
longitud de Fragmentos de Restricción. Técnica en la que originalmente se digería el ADN
genómico con enzimas de restricción para posteriormente transferir a unamembrana (Southern

xv
blotting) e hibridar con una sonda conocida (ej. minisatelite). Actualmente, existe una
variante en la que primeramente se amplifican por PCR regiones conocidas, comúnmente de
ADN mitocondrial, y posteriormente se digieren con enzimas de restricción, evaluando el
polimorfismo en cuanto al largo de los fragmentos generados.
Satélite. Grandes regiones de ADN (103 a 107 pb) con segmentos de más de 100 pb repetidos
en tándem.
Secuenciación. Establecimiento del orden de las bases nucleotidicas (secuencia) de 5' a 3' en
un fragmento de ADN de interés.
VNTR (del inglés Variable Number of Tandem Repeats). Número variable de repeticiones
en tándem. Término genérico para denotar a las secuencias repetidas en tándem de 1 a 100 pb,
incluye las regiones microsatelite y minisatelite.

1
I. Resumen La dependencia de combustibles fósiles, el aumento sostenido del precio del barril de petróleo
en el mercado internacional junto a la preocupación por los efectos del cambio climático que
ya se viene percibiendo en los últimos años ha hecho viable la propuesta de implementar el
cultivo de Jatropha curcas L. ―piñón blanco‖ en el Perú como alternativa para diversificar el
abastecimiento energético. Se propone que su cultivo contribuirá además a la recuperación de
zonas marginales, deforestadas y a la reducción de la emisión de gases con efecto
invernadero, ayudando de esa manera a mitigar el impacto del cambio climático global. Este
interés se debe a las características deseables del piñón blanco, basándose en su variabilidad
genética, que le permite resistir condiciones diversas y exigentes, sin embargo, esta cualidad,
muy probablemente se presente como dificultad cuando se desee obtener cierta uniformidad y
homogeneidad en grandes extensiones de cultivo. Para esto es necesario realizar estudios
moleculares que ayuden a establecer la situación actual de la estructura genética en las
poblaciones distribuidas en el país, identificando marcadores genéticos asociados a la calidad
del aceite del ―Piñón blanco‖, y así seleccionar genotipos específicos. La presente
investigación ha sido realizada para evaluar la medida de la variabilidad genética en un
conjunto representativo de 60 accesiones de J. curcas proveniente de la costa norte del Perú.
Como resultado se obtuvo un polimorfismo molecular de 48,35% usando 15 primers ISSR y
47,11% con 08 iniciadores RAPDs que indica niveles modestos de variabilidad genética en
las accesiones colectadas en la costa norte del Perú, también se evaluó la capacidad
informativa de estos marcadores mediante el índice de contenido polimórfico (PIC). Además
se construyó un fenograma con los datos obtenidos utilizando el coeficiente de similaridad
simple matching y el agrupamiento tipo UPGMA no encontrándose marcadores específicos
que ayuden a diferenciar las poblaciones en estudio.

2
II. Introducción El cambio climático a nivel mundial se ha visto acelerado por el incremento del efecto
invernadero, influenciado por el actual modelo energético, que es un sistema abierto, en la que
el hombre adiciona a la atmósfera elevadas cantidades de dióxido de carbono CO2 (30 mil Gg
de CO2 al año en el Perú)(CONAM 2001)debido básicamente al cambio en el uso del suelo
(principalmente por la deforestación) y a las emisiones de los combustibles fósiles,
representando el 66% de la fuente de energía primaria donde más de las ¾ partes está
sustentado por el uso de petróleo3.
Entre los años 2006-20091 el Perú importó 126.91 millones de barriles de petróleo y 24.21
millones de barriles de diesel 2 para abastecer el mercado interno, existiendo una dependencia
directa del precio del barril de petróleo en el mercado internacional75
. Esto ha llevado a un
aumento sostenido en el precio del barril de petróleo llegando a cifras record por encima de
los 140 dólares por barril en junio del 2008, manteniéndose por encima de los 80 dólares en
octubre del 201076
. Este sostenido incremento en el precio del barril de petróleo, así como el
hecho de que todos los días por 4 barriles de petróleo que se consume se descubre uno, ha
hecho viable la propuesta de producir combustibles alternativos como gasol y biodiesel para
incrementar la diversidad del abastecimiento energético, contribuyendo además a la
recuperación de zonas marginales, deforestadas y a la reducción de la emisión de gases con
efecto invernadero y mitigar así el impacto del cambio climático disminuyendo la polución
del aire en grandes ciudades afectadas por la contaminación producida por el transporte
automotor.
La producción mundial de biodiesel en el 2008 fue liderada por Alemania con una
participación del 22,2%, seguido por EE UU (18,3% del total), Francia (14,3%), Brasil (8,1%)
y Argentina (7,6%).77
En esta producción sobresalen los cultivos de soya, canola y palma
aceitera impulsados por políticas nacionales que respaldan su desarrollo. En el 2004 Brasil
autorizó la mezcla de 2% de biodiesel en el diésel convencional, la que a partir del 2008 será
obligatoria. Del mismo modo a partir del 2013, la mezcla de 5% de biodiesel en el diésel (B5)
será obligatoria.
Varios gobiernos han anunciado metas ambiciosas de producción de biodiesel en sus países,
entre los que destacan el de la India (4.500 millones de litros/año de biodiesel de piñón),
Brasil (2000 millones de litros/año de biodiesel, principalmente de palma, ricino y soya, hacia
el 2012) e Indonesia (4.700 millones de litros/año de biodiesel de palma hasta el 2025).78
En
1 Información de enero-mayo.

3
el Perú por Decreto Supremo Nº 016-2008-AG se declaró al cultivo de la higuerilla y el piñón
de interés nacional , Esto ayudará a reducir las importaciones que en el 2009 ascendieron a
469 mil barriles de biodiesel sumando US$ 52,4 millones.76
Para lograr este abastecimiento
interno y obtener una sostenibilidad en la producción de aceites vegetales para la obtención de
biodiesel es importante realizar estudios previos que permitan estimar, mantener y aumentar
la variabilidad genética del cultivo a implementar, de manera que se logren seleccionar
aquellos individuos o poblaciones más adecuados, logrando determinar en gran medida la
evolución futura de las poblaciones, su adaptación al medio y su conservación.
Bajo estos criterios, el cultivo de Jatropha curcas L. ―Piñón blanco‖ presenta grandes
ventajas debido a características deseables que se basarían en su variabilidad genética que le
permite resistir condiciones tan diversas y exigentes, como las de la costa desértica o la selva
lluviosa tropical. Sin embargo, esta cualidad, muy probablemente se presente como dificultad
cuando se desee obtener cierta uniformidad y homogeneidad en grandes extensiones de
cultivo. Para esto el Instituto Nacional de Innovación Agraria (INIA) ha venido desarrollando
diversos trabajos de investigación en los que se contempla la selección de germoplasma y el
manejo agronómico. Para el primero es, necesario realizar estudios moleculares que ayuden a
establecer la situación actual de la estructura genética de las poblaciones distribuidas a lo
largo del país, identificando marcadores genéticos asociados a la calidad del aceite del ―Piñón
blanco‖, y de ese modo seleccionar genotipos específicos.
Este estudio utilizó herramientas moleculares para el análisis de la variabilidad genética
natural de Jatropha curcas L. ―Piñon blanco‖ distribuidas en la costa norte del Perú, las
cuales permitirán identificar marcadores que ayuden a la conservación de su acervo genético y
su utilización en programas de mejoramiento genético escogiendo las variedades más
apropiadas para la producción de aceite a gran escala.
III. Antecedentes
3.1 Biodiesel como fuente de energía renovable.
En los últimos años el mundo se ha enfrentado a una crisis debido al agotamiento de los
recursos energéticos y al aumento de los problemas ambientales. Esta situación ha llevado a la
búsqueda de un combustible alternativo, cuya producción no sólo sea sostenible, sino también
―amigable‖ con el medio ambiente. Para los países en desarrollo, los combustibles de origen
biológico, tales como el alcohol, aceites vegetales, biomasa, biogás, combustibles sintéticos,
etc., se han convertido cada vez más en piezas importantes dentro de las estrategias
energéticas a implementar.1

4
La idea de usar aceites vegetales como combustible para los motores de combustión interna
data de 1895 cuando el Dr. Rudolf Diesel desarrollaba su motor de combustión interna. En la
presentación del motor diesel en la Exposición Mundial de París en 1900; el Ing. Diesel usó
aceite de maní como combustible. En esta presentación señaló «el uso de los aceites
vegetales como combustibles para los motores puede parecer insignificante hoy en día,
pero con el transcurso del tiempo puede ser tan importante como los derivados del petróleo y
el carbón en la actualidad» (Shay, 1993)2.citado por
3
Hoy en día con el perfeccionamiento de los motores diesel y el uso de destilados de petróleo
que poseen menor viscosidad no podría usarse aceites vegetales directamente, debido
precisamente, por su mayor viscosidad., Para solucionar este problema se usa un proceso
químico denominado transesterificación, la cual consiste en reemplazar el glicerol (alcohol
trivalente) por un alcohol monovalente («más ligero») usualmente metanol o etanol,
formando moléculas más pequeñas (ésteres monoalquílicos), a los cuales se les denomina
Biodiesel haciendo referencia a su origen biológico. Este proceso se realiza bajo la acción de
un catalizador que puede ser de origen acido o básico4.
Fig.1. Proceso de transesterificación para la producción de biodiesel. Sacado de Arpel Iica 2009
Aunque difícilmente el biodiesel pueda reemplazar totalmente al combustible diesel basado
en petróleo, hay por lo menos cinco razones que justifican su desarrollo:
1. La apertura del mercado para el exceso de aceites vegetales y grasas animales.
2. Disminuye, aunque no elimina, la dependencia del país del petróleo importado.
3. El biodiesel es renovable y no contribuye al calentamiento global debido a su cierre
del ciclo de carbono. Un análisis del ciclo de vida del biodiesel mostró que, en general
las emisiones de CO2 se redujeron en un 78% en comparación con el combustible
diesel basado en petróleo.

5
4. Las emisiones de monóxido de carbono y las emisiones de partículas del biodiesel
son más bajos que con el combustible diesel regular. Desafortunadamente, la mayoría
de las pruebas de emisiones han mostrado un ligero incremento en óxidos
de nitrógeno (NOx).
5. Cuando se añade al combustible diesel regular en una cantidad igual al 2.1%, se
puede convertir el combustible con propiedades lubricantes pobres, como el
combustible diesel ultra-moderna de bajo contenido en azufre, en un combustible
aceptable5.
Otras características importantes del biodiesel es que contiene 10-11% de oxígeno (w/w),
mejorando así el proceso de combustión en un motor. También se ha reportado que el uso de
aminas grasas y amidas terciarias pueden ser eficaces en la mejora de la calidad de ignición
del biodiesel sin tener ningún efecto negativo sobre sus propiedades de flujo en frío. Sin
embargo, persisten problemas de arranque en frío. Además, el biodiesel tiene valores bajos de
calentamiento volumétrico (alrededor del 12%), un alto índice de cetano y un alto punto de
inflamación, siendo los puntos de inflamación del biodiesel entre 15-25 °C más altas que las
del diesel (ver tabla N° 1)1.
Tabla Nº 1: Propiedades de biodiesel provenientes de diferentes fuentes vegetales1.
Ésteres metílicos de aceites
vegetales metílico (biodiesel)
La viscosidad cinemática (mm2/ s)
Cetano.
Menor poder
calorífico (MJ / kg)
Punto de
nube (°C)
El punto
de fluidez
(°C)
Punto de inflamación
(°C)
Densidad (kg / l)
Piñon 4,9252 53 47.8 0 170 0.86
Maní 4.9 54 33.6 5 - 176 0.883
soja 4.5 45 33.5 1 -7 178 0.885
Palma 5.7 62 33.5 13 - 164 0.880
Girasol 4.6 49 33.5 1 - 183 0.860
Diesel 3.06 50 43.8 - -16 76 0.855
20% de mezcla de biodiesel
3.2 51 43.2 - -16 128 0.859
Dependiendo de las condiciones climáticas y del suelo, los distintos países están buscando
diferentes tipos de aceites vegetales como sustitutos de los combustibles diesel. Por ejemplo,
el aceite de soja en los EE.UU., la colza y el girasol en Europa, el aceite de palma en el
Sudeste de Asia (principalmente Malasia e Indonesia) y el aceite de coco en las Filipinas. La
producción de semillas oleaginosas a nivel mundial y el porcentaje de recuperación de aceite
por semilla se indica en la Tabla N° 2 , lo que indica que el uso de aceites vegetales como
fuentes de biodiesel, requieren más esfuerzos para aumentar la producción de semillas

6
oleaginosas y para desarrollar nuevas especies de plantas más productivas y con alto
rendimiento de aceite.1,6
Tabla Nº 2: Producción mundial de semillas oleaginosas y porcentaje de recuperación de aceite en semilla.
Semillas oleaginosas
Producción Mundial (millones de toneladas)
% Recuperación
Soya 123.2 17 Semilla de algodón 34.3 11
Maní 19.3 40 Girasol 25.2 35 Colza 34.7 33
Sésamo 2.5 – Palma 4.8 – Copra 4.9 65 Linaza 2.6 43 Piñón 1.3 42 Níger 0.8 30
Salvado de arroz
–
15
Total 253.6 –
En Sudamérica, Brasil tienen una larga experiencia en producción de etanol a partir de caña
de azúcar para su uso como combustible alternativo a los combustibles fósiles, incluyendo el
desarrollo de motores adaptados para el uso de etanol puro. En el caso del biodiésel, en el
2004 se lanzó el Programa Nacional de Producción y Uso del Biodiésel, que busca impulsar a
este combustible como una opción para el desarrollo del agro en las zonas más pobres de ese
país. Este programa consta de un marco regulatorio, de metas físicas de uso del biodiésel, y de
una planificación de cultivos oleaginosos en todo el país. Por este motivo en el 2004 se
autorizó la mezcla de 2% de biodiésel en el diésel convencional y amparado bajo una ley, a
partir del 2008 este porcentaje de mezcla será obligatorio. Asimismo, a partir del 2013, se
promoverá la mezcla del 5% de biodiesel en el diésel (B5), destinándose como meta la
producción de 2.000 millones de litros/año de biodiesel hacia el 2012, principalmente de
palma, ricino, soya y piñón. Por su parte, el Perú estableció la mezcla obligatoria ―B2‖ en el
2009 y la del ―B5‖ a partir del 2011 por lo que se prevé una demanda nacional de biodiesel
de 215 millones de litros para el 2011 y de 250 millones para el 20167. (Ver cuadro N° 3)
Tabla Nº 3: Demanda nacional de biodiesel (en millones de litros) hasta el año 2016. Combustible (millones de
litros)
2006 2007 2008 2009 2010 2011 2012 2013 2014 2015 2016
Diesel 3482.1 3540.1 3656.2 4004.4 4120.4 4294.5 4410.6 4584.7 4758.8 4874.9 4991.0
Biodiesel (2%)
80.09 82.41
Biodiesel (5%)
214.73 220.5 229.2 237.9 243.7 249.5

7
De esta manera el Perú ha puesto gran interés al cultivo de piñón para la producción de
biodiesel, impulsado particularmente por tres características de la planta que son importantes
para nuestra realidad: a) rusticidad y adaptabilidad, b) Toxicidad que le permite no competir
con otros aceites comestibles, c) Recuperación de tierras marginadas, deforestadas o utilizadas
en cultivos ilegales.
3.2 Jatropha curcas L. “Piñón”
El género Jatropha pertenece a la tribu Joannesieae de la familia Euphorbiaceae y contiene
aproximadamente 170 especies conocidas. Linneo en 1753 fue el primero en nombrar el
género Jatropha dentro de su obra "Species Plantarum", y esto sigue siendo válido hasta
hoy. El nombre del género Jatropha deriva de la palabra griega ―jatrós‖ (médico) y ―trophe‖
(alimentos), que indica los usos medicinales atribuidos8.
El piñón blanco es un árbol pequeño o arbusto grande, de hasta 5.7 m de altura, con una
madera suave y una vida útil de hasta 50 años. La planta tiene su área de distribución natural
desde México en América Central hasta Brasil, Bolivia, Perú, Argentina y Paraguay en
Sudamérica, aunque hoy en día tiene una distribución pantropical con diferentes procedencias
de semilla. La planta desarrolla una raíz pivotante profunda y en un principio cuatro raíces
laterales poco profundas 9. La raíz principal puede estabilizar el suelo contra deslizamientos
de tierra, mientras que las raíces poco profundas se distribuyen para prevenir y controlar la
erosión del suelo causada por el viento o el agua. Las hojas son lisas, con 4-6 lóbulos y de 10-
15 cm de longitud. La planta es monoica y las inflorescencias terminales contienen flores
unisexuales. La proporción de flores masculinas y flores femeninas esta en el rango de 13:01
a 29:1 10
y disminuye con la edad de la planta. Normalmente las flores de Jatropha curcas
brotan solo una vez al año durante la temporada de lluvias. En las regiones húmedas o bajo
condiciones de riego, las flores brotan de forma permanente casi todo el año. Después de la
polinización, las inflorescencias forman un racimo de frutos verdes dispuestos de manera
elipsoidal. Las semillas negruzcas de la mayoría de las procedencias contienen toxinas, como
esteres de forbol , curcina, inhibidores de tripsina, lectinas y fitatos, a niveles tales que las
semillas, el aceite y la torta de semillas no son comestibles sin una previa desintoxicación 11,12
y 13.

8
Fig.2. Partes importantes de piñón: Rama con flores (a); Corteza (b); Nervadura de la hoja (c); Pistilos de la flor
(d); Estambres de la flor (e); Corte transversal del fruto inmaduro (f); Fruto (g); Corte longitudinal del
fruto (h); semillas (i) sacado de Joachim Heller 1996.
Dependiendo de la variedad, las semillas peladas contienen un 40-60% de aceite.14 y 15
Los
productos químicos aislados de diferentes partes de la planta se muestran en la Tabla Nº4;
Estos productos químicos pueden ser utilizados en diversas aplicaciones industriales pero es
su uso tradicional ampliamente desarrollado por los pobladores donde habita esta especie
atribuyéndole desde propiedades afrodisiacas hasta anticancerígenas.79

9
Tabla Nº4: Composición química de diferentes partes de Jatropha curcas L.8
Partes de la
planta Composición química Referencias
partes aéreas Ácidos orgánicos (o y p-cumárico ácido, p-OH del ácido
benzoico, ácido protocatéquico, ácido resorsilico,
saponinas y los taninos
Hemalatha y Radhakrishnaiah
(1993)
Corteza del
tallo β-amirina, β-sitosterol y taraxerol
Mitra et al. (1970)
Hojas Triterpenos cíclicos estigmasterol, stigmast-5-en-3β, 7 β-
diol, stigmast-5-en-3β, 7α-diol, colest-5-en-3β, 7β-diol,
colest-5-en-3FL, 7α -diol, campesterol, β-sitosterol, 7-
ceto-β-sitosterol, así como el β-d-glucósido de β-
sitosterol. Los flavonoides apigenina, vitexina, isovitexin
Mitra et al. (1970), Khafagy et
al. (1977), Hufford y
Oguntimein (1987)
Las hojas también contienen el dímero de un alcohol
triterpeno (C63H117O9) y dos glucósidos flavonoidal
Khafagy et al. (1977)
Látex Curcacyclina A, un octapéptido cíclico
Van den Berg et al. (1995)
Curcina (una proteasa) Nath y Dutta (1991)
Semillas Curcina, (una lectina) Stirpe et al. (1976)
Esteres de Forbol
Adolf et al. (1984), Makkar et
al. (1997)
Esterasas (JEA) y lipasa (JEB)
Staubmann et al. (1999)
Torta de prensa
Fitatos, saponinas y un inhibidor de la tripsina
Aregheore et
al. (1997), Makkar y Becker
(1997), Wink et al. (1997)
Raíces β-sitosterol y su β-d-glucósido, marmesin, propacin, el A
y B curculathyranes y el curcusones A-D. diterpenoides
jatrophol y jatrofolona A y B, la tomentin cumarina, el-
lignanos jatrophin coumarino así como taraxerol
(Naengchomnong et al., 1986)
y (Naengchomnong et al.,
1994)
3.2.1 Posición taxonómica de Jatropha curcas.
Reino Plantae Plantas, vegetales
Subreino Tracheobionta - plantas vasculares
División Magnoliophyta Angiospermas, plantas con flores, fanerógamas.
Clase Magnoliopsida dicotiledóneas.
Subclase Rosidae
Orden Euphorbiales
Familia Euphorbiaceae - Euphorbes,
Género Jatropha L. Jatropha (jatros) médico, (trofes) alimentos.
Especies Jatropha curcas L. – Piñón blanco pagina web

10
3.2.2 Usos de Jatropha curcas L
a) En la medicina.- Los extractos de J. curcas se utilizan en remedios caseros para una
serie de desórdenes fisiológicos19
. El jugo de la hoja se utiliza como una aplicación
externa para las hemorroides, y la decocción de las hojas en la enfermedad de la
artritis y venéreas. El té de hoja se utiliza para el marasmo y la ictericia18
. El látex de
J. curcas contiene alcaloides como la jatrophine, jatropham, jatrophone y curcina la
cual se le atribuye propiedades anti-cancerígenas. El látex se utiliza también para
cubrir las llagas, úlceras y la lengua inflamada. Las semillas se utilizan para la
hidropesía, la gota, parálisis y enfermedades de la piel16
. Las raíces se utilizan en
infusiones como enjuague bucal para el sangrado de las encías y dolor de muelas.
También se han aislado compuestos antibacterianos en el tejido del tallo de J.
curcas18
. En el sur de Sudán, las semillas y los frutos se utilizan como un método
anticonceptivo17
.
b) Como fertilizante.- La torta de semilla procedente de la extracción del aceite contiene
principios tóxicos como los esteres de forbol y curcina, lo que lo hace inadecuado para
la alimentación animal. Sin embargo, las aplicaciones potenciales como fertilizante o
en la producción de biogás han sido informado.20, 21 y 22
Por ser rico en nitrógeno, la
torta de semillas es una excelente fuente de nutrientes para las plantas y también ayuda
en el mejoramiento del suelo. La aplicación de la biomasa residual exedente del
proceso de elaboración del biodiesel ha incrementado el rendimiento de muchos
cultivos demostrándose su aplicabilidad como fertilizante, además puede ser un
recurso valioso en la producción de bajo costo de muchas enzimas industriales.
c) Otros usos.- Además de su valor medicinal y como fertilizante, el aceite ha sido
utilizado para la iluminación como aceite en lámparas, y en la fabricación de jabón y
velas23
. Las nueces recogidas de una variedad mexicana que no son tóxicas se asan y
se consumen. El látex es un potente inhibidor del virus mosaico de la sandia24
, y la
corteza se utiliza como veneno para peces23
.
3.2.3 Toxicidad de J. curcas
Como ya hemos mencionado, la presencia de esteres de forbol hace a la semilla de piñón no
aptos para el consumo humano y ganado, a pesar del contenido alto en proteínas y favorable
composición de otros nutrientes25
. Los intentos para inactivar los principios tóxicos han sido
variados desde la utilización de procesos hidro-térmicos, extracción con solventes,
tratamiento con NaHCO3 y radiación ionizantes dando como resultado la reducción del

11
principio tóxico, pero estos métodos son costosos en tiempo y dinero.26
En México y América
central se reportó una variedad no tóxica de J. curcas que no contienen o sólo contienen baja
cantidad de esteres de forbol. El cultivo de variedades no tóxicas aportaría un valor añadido a
los cultivos mediante la utilización de la torta de semillas desaceitada para la alimentación
animal. Sin embargo, no hay diferencia morfológica entre las variedades tóxicas y no tóxicas.
De allí que es importante la identificación de marcadores ligados a las variedades tóxicas o no
tóxicas para la búsqueda de nuevas accesiones con características no tóxicas, esto agregaría
valor al cultivo, posibilitando su cultivo para consumo humano. 26 y 27
3.2.4 Jatropha curcas como biodiesel
La semilla de Piñón contiene un 30-40% de aceite de las cuales el 21% son ácidos grasos
saturados y el 79% son ácidos grasos insaturados. El reconocimiento de su potencialidad para
producir un biodiesel de alta calidad ha llevado a una oleada de interés en todo el mundo; más
aún, ya que debido a su toxicidad, evita el dilema "alimentos versus combustible.28, 29 y 30
Además de biodiesel, a partir de la semilla se puede obtener la torta de prensa que contiene
19.0% de proteínas, 17.0% de carbohidratos y 16.0% de fibra. La torta desaceitada es similar
al estiércol de pollo y se puede utilizar como un excelente abono orgánico.25
Su alto punto de
inflamación (160º-170°C) y el número de cetano (55-58) hace del diesel de Jatropha más
ecológico que los productos petroquímicos convencionales.28, 17
El rendimiento por hectárea
por año va desde 1,5 hasta 7,5 toneladas de semillas secas con un porcentaje de aceite de
34,4%. Partiendo de un promedio de 4,5 toneladas de semillas secas por hectárea por año, el
rendimiento es de unos 1.500 litros de aceite de piñón. Bajo estos criterios el área necesaria
para satisfacer la demanda nacional de B2 y B5 sería de 84.000 - 400.000 hectáreas con un
promedio de 144.000 hectáreas para el 2011. Siendo todo un reto que se debe asumir de
manera multisectorial para cumplir con las metas trazadas7.
3.3 Variabilidad Genética y Caracterización de las especies vegetales
La población que conforma una especie vegetal está generalmente correlacionada con la
variación que se encuentra en los factores del medio ambiente de su área de distribución
natural o de plantación en la que crece esta población32
, para ello cada especie adapta la
información contenida en el genoma de acuerdo con las necesidades de sobrevivir en su
entorno. El resultado de esta interacción adaptativa se traduce en la acumulación de la
información genética que a manera de variantes las especies van guardando entre los
miembros de su población, y que se va transmitiendo en las subsiguientes generaciones a
través del tiempo. De esta manera, aunque la población de individuos en una especie

12
comparten características comunes y se pueden cruzar entre ellos, también es cierto que en
cada uno existen muchas variantes individuales. La suma de todos los individuos con sus
respectivas variantes es de lo que se conoce como variabilidad genética de una especie, la cual
permite a dicha especie adaptarse a los cambios que se pueden presentar en su entorno32
.
Como resultado de la variabilidad producida en los procesos antes mencionados la
variabilidad genética se almacena en el genoma y pueden o no expresarse en características
que permitan ser identificadas, por lo tanto desde el punto de vista de su expresión, la
variabilidad contenida en el genoma de una especie puede ser agrupada en dos grandes clases
(1) la que se expresa en características visibles y que conforman el fenotipo, y (2) la que no se
expresa en características visibles y que en general se refiere a los procesos o productos de la
planta. Distinguir entre lo que puede o no ser expresado en forma visual es necesario para
precisar que porción de la variabilidad total de la especie se está analizando en la
caracterización. Así en la caracterización de una especie se estima la variabilidad existente en
el genoma de la población de individuos que la conforman, esas variantes se van acumulando
entre los diferentes miembros de la especie y la suma de todos los efectos de los genes y sus
variantes es lo que se denomina variabilidad genética de una especie.
Dado a las características visibles y no visibles que expresa el genoma, la caracterización
podrá medir o describir el nivel de variabilidad, utilizando diversas herramientas o métodos
estadísticos para analizar los datos resultantes de un estudio de caracterización.
La caracterización de la variabilidad detectable visualmente se pueden enfocar en las
características responsables de la morfología y la arquitectura de la planta utilizadas en la
clasificación taxonómica, las características relacionadas con aspectos de manejo agronómico
y las características que se expresan como reacción a estímulos medioambientales, este tipo
de caracterización se denomina de ―evaluación‖ y para su correcta cuantificación,
generalmente, se requieren diseños experimentales separados de la caracterización
morfológica y agronómica.33
Por otro lado la caracterización de la variabilidad no detectable visiblemente utiliza técnicas
sensibles que permiten medir y evaluar un conjunto de marcadores moleculares que reflejan la
variabilidad genética, a este tipo de caracterización se le denomina caracterización molecular.
3.3.1 Fuentes de variabilidad genética
Son diversas las causas o fuentes que pueden aumentar o disminuir la variabilidad genética las
cuales podemos resumir de la siguiente manera.
a) Fuente evolutiva.- Es producida durante los procesos evolutivos de especiación por la
que haya pasado la población, principalmente durante las etapas de aislamiento

13
reproductivo, así como a la dinámica que la especie ha tenido y sigue teniendo en
condiciones naturales. En este aspecto Ford-Lloyd y Jackson consideran que los
patrones de diversidad genética de las plantas cultivadas resultan de la interacción de
los factores como mutación, migración, recombinación, Selección y deriva genética.
En esta interacción entra a jugar un papel importante la biología reproductiva,
autógama o alógama, con sus respectivas variantes que desarrolle la especie esperando
una mayor variabilidad en las alógamas que en las autógamas34
.
b) Geográfica.- Se encarga de proporcionar características importantes como aislamiento
y distribución para crear variantes genéticas de adaptación como respuesta a
variaciones en los componentes ambientales.
c) Domesticación.- Durante el proceso de domesticación de las especies cultivadas el
hombre ha ejercido una fuerte presión de selección que ha permitido la preservación
de muchas variantes las cuales, posiblemente, hubieran desaparecido en condiciones
naturales. De la misma manera, el hombre también indujo la producción de nuevas
variantes, tanto para facilitar el manejo agronómico como para incrementar la
producción.33
3.3.2 Pérdida de la variabilidad genética
La variabilidad genética es importante porque permite a las poblaciones adaptarse a un medio
cambiante. Los individuos con ciertos alelos o ciertas combinaciones de alelos pueden poseer
precisamente aquellas características adecuadas para sobrevivir y reproducirse bajo las nuevas
condiciones. En una población, ciertos alelos pueden variar en frecuencia desde muy raros
hasta muy comunes. En las poblaciones pequeñas, las frecuencias alélicas pueden cambiar de
una generación a otra simplemente debido al azar, dependiendo de qué individuos se apareen
y produzcan descendencia, un proceso conocido como deriva genética. Cuando un alelo se
encuentra en una población con frecuencia baja tiene una probabilidad significativa de
desaparecer por azar en la siguiente generación. Para el caso teórico de una población aislada
en la que existan dos alelos por gen, Wright (1931) propuso una fórmula que expresa la
proporción de heterozigocidad (individuos que poseen los dos alelos diferentes de un mismo
gen) inicial que queda después de cada generación (H) para una población N, de adultos en
edad reproductora:
H= 1- 1/2Ne
Según esta ecuación, una población de 50 individuos retendría un 99% de su heterozigosidad
inicial al cabo de una generación debido a la perdida de alelos raros, y poseería todavía un

14
90% al cabo de 10 generaciones. En cambio una población de 10 individuos retendría tan sólo
95% de la heterozigosidad inicial al cabo de una generación y sólo el 60 % al cabo de 10
generaciones36
.
Esta fórmula demuestra que en las poblaciones pequeñas aisladas, en particular las que se
ubican en islas y paisajes fragmentados, se pueden producir pérdidas notables de variabilidad
genética. No obstante, la migración de individuos entre poblaciones y la mutación normal de
genes tienden a aumentar la variabilidad genética en una población contrarrestándose así los
efectos de la deriva genética. Basta una frecuencia baja de migración de individuos entre
poblaciones para minimizar la pérdida de la variabilidad genética asociada al pequeño tamaño
de la población.37 y 38
Es este flujo genético lo que al parecer evita la pérdida de la variabilidad
genética en poblaciones pequeñas39
y les permite adaptarse rápidamente a cambios
ambientales e incluso catastrófico. Aunque las tasa de mutación que se da en la naturaleza es
de 1 por cada 1000 y 1 por cada 10000 bases nucleotídicas por gen y generación, es suficiente
para contrarrestar la pérdida de alelos en poblaciones grandes, siendo suficientes para evitar la
deriva genética en poblaciones pequeñas de unos 100 individuos o menos.36
Hay dos tipos de variación genética que son importantes cuando se trata de poblaciones
pequeñas uno es el grado de homocigosis en los individuos en una población, es decir, la
proporción de cada uno de los loci que contienen un par de alelos homocigotos en lugar de
alelos heterocigotos y el otro es el grado de monomorfismo y polimorfismo en una población,
es decir, el número de alelos diferentes del mismo gen existente en el acervo genético de una
población. El polimorfismo puede ser particularmente importante en los loci implicados en la
respuesta a la adaptación a los cambios en el ambiente, de allí su amplio uso en el estudio de
caracterización de poblaciones.40
3.3.3 Polimorfismo
Como hemos mencionado la variabilidad genética se debe a variaciones en la secuencia del
genoma; por tanto en un sentido amplio el concepto de variabilidad genética se hace sinónimo
de polimorfismo genético.
Dentro de la cadena de ADN el polimorfismo afecta tanto a regiones codificantes del genoma
como a regiones no codificantes, en ambos casos puede consistir en la variación de un solo
par de bases del ADN o en millones de pares de bases, el primer caso se conoce como
polimorfismo de un solo nucleótido o SNP (single-nucleotide polymorphism; ―snip‖).
Cualquier variación se puede dar en células germinales o reproductoras con lo que se trasmite
a la descendencia (polimorfismo hereditario), o bien en células somaticas no reproductoras en
cuyo caso no se trasmite (polimorfismo no hereditario).41
Inicialmente la definición de

15
polimorfismo se refería a las proteínas; y luego a sus genes. Hoy en día, se debe definir como
la ―existencia simultánea en una población de genomas con distintos alelos para un locus
determinado‖.
El grado de polimorfismo en una población es tanto mayor cuanto más individuos contenga,
viene determinado por el número de alelos distintos existentes para un locus concreto y refleja
el grado de heterozigosis o proporción de individuos heterocigóticos que forman parte de la
población, lógicamente, la causa última de la existencia de polimorfismo es la mutación del
ADN. La distinción entre el uso de los términos de mutación y polimorfismo no es clara ni
unánime, pero en general se asocia el primero con situaciones excepcionales, en especial
patológicas, mientas que se habla de polimorfismo cuando la presencia de la variación
genética es razonablemente común en la población y, por tanto estable y nada o poco
perjudicial. Por convenio se dice que un locus es polimórfico cuando la variabilidad genética
afecta a más del 1% de la población42
.
3.3.3.1 Polimorfismo en regiones génicas codificantes.
Los polimorfismos en regiones codificantes reciben el nombre de «polimorfismos génicos».
Esta clase de polimorfismos pueden tener o no un efecto sobre el fenotipo. Los polimorfismos
génicos, sin efecto fenotípico, son los más comunes y son los responsables de la diversidad
genética normal entre individuos (p. ej., los polimorfismos existentes en proteínas).
Pero cuando un polimorfismo génico (es decir, que afecta a una región del ADN codificante)
da como resultado una alteración fenotípica, en la mayoría de los casos es perjudicial, ya que
puede modificar las características bioquímicas, fisiológicas e incluso morfológicas de la
célula, pudiendo originar procesos patológicos. Sólo en casos excepcionales, esta variación o
mutación puede ser beneficiosa, dando lugar a una ventaja adaptativa al individuo, siendo éste
el motor de la evolución de las especies.
Los polimorfismos con alteración del fenotipo, pero que no influyen a la susceptibilidad a
enfermedades, determinan las características diferenciales entre los individuos de una misma
especie. Los polimorfismos que generan variaciones fenotípicas pueden influir de forma leve
en la susceptibilidad a distintas enfermedades.
3.3.3.2 Polimorfismo en regiones génicas no codificantes.
Teniendo en cuenta la definición más completa del gen, se establece con un criterio funcional
existen regiones del ADN que forman parte del gen, pero no codifican su producto génico.
Tal es el caso de las regiones reguladoras (promotores), los intrones y las regiones 3’ y 5’ que
no son traducidas. Se debe considerar por ello la posibilidad de polimorfismo en estas
regiones génicas no codificantes. Evidentemente, aunque tal polimorfismo no afectara la

16
secuencia de la proteína, puede afectar su expresión, dependiendo de la función que ejerza las
secuencias no codificantes que lo sufre. Muchos polimorfismos en los intrones no tendrán
efecto alguno, y serán equivalentes al polimorfismo en las regiones no génicas, estudiado a
continuación. Otros podrán afectar al corte y empalme, resultando en un polimorfismo
proteico. Las variaciones en secuencias promotoras probablemente perturben la expresión del
gen. Por tanto, el polimorfismo en regiones génicas no codificantes pueden manifestarse o no
en un efecto fenotípico.
11.3.3.3 Polimorfismo en regiones no génicas
La mayor parte de las mutaciones del genoma ocurren en las regiones del ADN que no
codifican ningún producto, debido a su abundancia en el genoma humano y de otros
eucariotas. Por lo tanto los cambios de las bases en el ADN no tienen efecto fenotípico alguno
de esta manera no se altera la secuencia, estructura o la función de la proteína. Son, sin
embargo, polimorfismo de gran interés aplicado: principalmente, para la búsqueda de genes
relacionados con enfermedades y para la identificación genética de individuos.
La elevada cantidad de loci polimórficos que se encuentran el ADN no codificante, junto a la
falta de trascendencia funcional de sus alteraciones (que permiten que se perpetué libremente
en sucesivas generaciones), hacen que sea esta región del ADN la que más difiera de uno a
otros individuos.42
3.4 Marcadores genéticos
3.4.1 Definición
Es un fragmento de ADN, o el producto de su expresión, que presenta polimorfismo y tiene
una pauta de herencia contrastada cuyo patrón de transmisión de generación en generación
Pueden ser monitoreados, sin importar la técnica que se utilice para ser detectada. Los
marcadores se utilizan a modo de ―etiquetas‖ de una unidad genómica concreta (gen, región
cromosómica, cromosoma, complemento cromosómico) en un individuo, población o especie.
Puede ser descrito como una variante (que pueden surgir debido a la mutación o alteración en
el locus genómico) que se pueden observar.43
3.4.2 Clases de Marcadores genéticos
Los marcadores genéticos pueden dividirse en tres clases: los marcadores morfológicos, los
marcadores bioquímicos y los marcadores basados en ADN o moleculares; los dos últimos
han sido los más utilizados en estudios de mejoramiento genético forestal. Los marcadores
bioquímicos pueden ser compuestos orgánicos de los organismos (por ejemplo terpenos,
alcaloides, etc.) o bien, pueden ser proteínas (isoenzimas), las cuales son las más ampliamente

17
utilizadas en estudios de especies forestales. Los marcadores bioquímicos examinan los
productos de los genes. Este análisis puede complicarse por efectos de la expresión génica,
epistasis (interacción con otros genes) y redundancia del código genético. El surgimiento de
las enzimas de restricción y de la tecnología de la Reacción en Cadena de la Polimerasa (PCR
por sus siglas en inglés), ha permitido investigar a los organismos a través del ADN.44
a) Marcadores morfológicos
Los marcadores morfológicos son aquellos cuya expresión fenotípica es observada a
simple vista siendo generalmente asociada a caracteres como colores, formas o
dimensiones expresadas en los individuos; son caracteres influenciados por el ambiente y
por interacción génica (epistasis) por lo que es difícil diferenciar los efectos genéticos de
los efectos ambientales .Para utilizar adecuadamente estos marcadores se debe tener una
información exacta del pedigrí del organismo en estudio.
Los primeros trabajos con este tipo de marcadores fueron los realizados por mendel en
guisantes en él año 1880 y en drosophila por Morgan en 191045
, Siendo este último quien
los uso para obtener mapas.
b) Marcadores Bioquímicos
Son aquellos cuya expresión fenotípica es observada con el uso de técnicas bioquímicas
mayormente realizadas en el laboratorio por ejemplo las isoenzima, que vienen a ser
enzimas que llevan a cabo la misma función, pero tienen una diferente composición de
aminoácidos y pueden diferir en sus propiedades cinéticas. Sometidas a electroforesis y
visualizadas mediante diferentes métodos de tinción, estas bandas suministrar información
como marcadores co-dominantes. Sin embargo, las desventajas que presentan estos
marcadores son las modificaciones que sufren en los procesos post- trascripciónales, el
escaso número de isoenzimas presentes en un organismo, la necesidad de tener un tejido
fresco o debidamente conservado para obtener actividad enzimática y la poca resolución
que se obtiene.
c) Marcadores moleculares.
Son aquellos que no necesitan de expresión fenotípica, si no, usan directamente el
ADN como fuente de variabilidad y polimorfismo dentro de las poblaciones en
estudio, tienen como mayor ventaja el no estar influenciadas bajo el medio ambiente
ni epistasis, Su uso es muy extendido por sus distintas cualidades y ventajas
desarrollándose en un gran número de poblaciones en diversas especies.46

18
Tabla Nº 5 Características importantes de los diversos marcadores utilizados en la caracterización de las
especies46
Característica Proteínas Isoenzima RFLP RAPD VNTR AFLP SSR Polimorfismo Alto Bajo Bajo-alto Medio-alto Medio-alto Medio-
alto Alto
Estabilidad ambiental
Alta Moderada Alta Alta Alta Alta Alta
Número de loci Bajo Medio Alto Alto Alto Alto Alto Reproducibilidad Alta Moderada-
alta Alta Moderada-
alta Alta Alta Alta
Aplicación Rápida-barata
Rápida-barata
Lenta- cara
Rápida- cara
Intermedia Lenta- cara
Lenta- cara
RFLP: Fragmentos de restricción polimórficos
RAPD: Amplificación de ADN al azar
VNTR: Número variable de repeticiones en tandem
AFLP: Amplificación de fragmentos polimórficos
SSR: Secuencias simples repetidas
3.4.3 Marcadores moleculares.
Un marcador molecular es cualquier fenotipo molecular oriundo de la expresión de un gen
o de segmentos específicos de ADN, que pueden ser detectados y su herencia
monitoreada.45
El polimorfismo basado en proteínas ha sido de gran utilidad en las investigaciones
realizadas en plantas, pero con el desarrollo de las tecnologías basadas en ADN, la
investigación en esta área se ha visto favorecida con la disponibilidad de una mayor
cantidad de marcadores, aquellos basados en Fragmentos de Restricción Polimórficos
(RFLP) y en la Reacción en Cadena de la Polimerasa (PCR). Ambas técnicas han
derivado en múltiples técnicas como son la Amplificación de ADN al Azar (RAPD),
Fragmentos Polimórficos de ADN Amplificados (AFLP), minisatélites (VNTR) y
microsatélites (SSR) e inter-microsatelites (ISSR) entre otros. 47
Los avances en el campo de la biología molecular proporciona muchas herramientas para
estudiar la diversidad genética en el genoma con el fin de investigar las relaciones
filogenéticas entre diferentes especies. Herramientas moleculares basadas en el análisis
del ADN son ideales para la caracterización del germoplasma y estudios filogenéticos.
En los años recientes el uso de marcadores moleculares ha facilitado significativamente el
análisis genético, ya que ha sido posible mapear un grupo de genes relacionados a control
de caracteres cuantitativos.

19
3.4.3.1 Ventajas de los marcadores moleculares
La característica que hacen del uso de los marcadores moleculares más conveniente que el
de los marcadores morfológicos son los siguientes:
a) Neutralidad fenotípica: A diferencia de los marcadores morfológicos, la marcadores
moleculares no presentan influencia fenotípica, esto significa que la interacción entre
el medio ambiente y las regiones de ADN, empleadas como marcadores, es nula, ya
que ésta no son codificantes. Se puede hacer el estudio del fenotipo sin interferencia
alguna de locus marcador.
b) Polimorfismo: Esta característica es muy importante, ya que depende directamente de
la variación alélica del marcador, recombinación genética, mutaciones puntuales,
insertos/delecciones (indel), entre otros; además los métodos para detectar el
polimorfismo en los marcadores moleculares son más sensibles y objetivos que los
empleados para marcadores morfológicos
c) Abundancia: Esta referida a la ocurrencia de variaciones alélica, en los marcadores
moleculares, a lo largo de todo del genoma de una especie. Teóricamente es posible
cubrir todo el genoma en detectar todos los poligenes de una especie.
d) Co dominancia: En cualquier generación segregante se puede detectar los diferentes
alelos de un mismo loci sin importar el carácter dominante o recesivo que este posea.
La mayoría de estos marcadores moleculares son de naturaleza co-dominante.45
3.4.3.2 Marcadores basados en la hibridación de sondas.
Utiliza sondas marcadas que se unen a regiones especificas en fragmentos productos de
enzimas de restricción; por ejemplo tenemos los RFLP (Restriction Fragment Length
Polymorphism), polimorfismo en la longitud de los fragmentos de Restricción, se basan en la
hibridación diferencial de una sonda como una muestra de ADN genómico digerida con una
enzima de Restricción, la cual reconoce secuencias nucleotidicas específicas (sitios de
Restricción)
El Procedimiento es el siguiente: Los fragmentos de ADN digeridos con una enzima de
Restricción son separados mediante electroforesis en un gel de Agarosa y luego transferidos a
un soporte solido, el cual puede ser una membrana de nylon o de nitrocelulosa (southern
blotting). El ADN es fijado en la membrana por acción de luz UV o calor y luego hibridizado
por una sonda de ADN marcada con un isótopo radiactivo o con una sustancia
quimioluminicente luego la detección de las bandas RFLP se puede hacer mediante auto
radiografía o quimioluminicencia.

20
Los RFLP son marcadores de naturaleza co-dominante, es decir, pueden observarse a los dos
alelos del marcador cuando están presentes en un individuo heterocigoto.48
3.4.3.3 Marcadores basados en PCR (reacción en cadena de la polimerasa)
La reacción en cadena de la polimerasa (PCR), desarrollada por Faloona y Mullis, (1987), es
un método enzimático que permite generar millones de copias de fragmentos de ADN a partir
de unas pocas copias. Este procedimiento costa de tres etapas:
a) Denaturación del ADN: Ruptura de los enlaces de hidrógeno que mantienen la
estructura de doble hélice del ADN a altas temperaturas (92-95)ºC
b) Alineamiento o apareamiento de los iniciadores: Apareamiento de los iniciadores
con un fragmento de ADN cuya secuencia es complementaria a (35-68)ºC, dicha
temperatura es crítica.
c) La extensión de las cadenas de ADN: Elongación de la cadena de ADN resultante
del apareamiento del iniciador, añadiendo nucleotididos al extremo 3’ mediante la
acción de una ADN polimerasa resistente a altas temperaturas. La cantidad de ADN
molde se duplica luego de cada ciclo, es decir, luego de 35 ciclo de PCR se obtendrán
235 copias del molde del ADN original.49
3.4.3.4 Clasificación de los marcadores moleculares basados en PCR
En los últimos años, una variedad de técnicas basadas en el ADN (por ejemplo, RAPD,
AFLP, SSR y CAPS, entre otros) se vienen desarrollando para estudiar la variación genética
en diferentes especies (Strofar, 1996). Las cuales podemos clasificar en:
a) Técnicas basadas en PCR que usan iniciadores arbitrarios o amplifican multi-
locus: Tienen la característica de no necesitar mayor información de la secuencia del
genoma en la que se está investigándose. Se puede mencionar a los RAPDs (Random
Amplified Polymorphic DNA que presentan iniciadores arbitrarios; AFLPs (Amplified
Fragment Lengh Polymorphism), los ISSR que utilizan microsatélites como
iniciadores para amplificar las secuencias que se encuentran flanqueadas por estos
microsatélites.
b) Técnicas Basadas en PCR de secuencia específica: La cual es necesario el
conocimiento de las zonas flanqueantes para la obtención de los iniciadores, dentro de
estos tenemos a los CAPS (Cleaved Amplified Polymorphic Sequence), los
microsatélites (SSR) y los SCAR (sequence characterized amplified regions).50 y 51

21
3.3.4 Marcadores Inter-Secuencias Simples Repetitivas (ISSRs)
Los marcadores ISSR es una técnica basada que utiliza el PCR mediante cebadores de
secuencias repetitivas no específicos (GCAGCAGCAGCAn) y con uno a tres nucleótidos
adiciones en el extremo 5' o 3' que anclan a uno de los extremos de las regiones inter-SSR.
Los ISSR son altamente polimórficos, determinan la diversidad genética y pueden ser
utilizados en trabajos de huella digital en individuos dentro de una población.52
Para los
estudios de población, los marcadores ISSR presentan de 3 a 5 veces mayor variabilidad que
los RAPDs y están disponibles para todos los grupos de plantas independientemente de la
disponibilidad previa de las secuencias del genoma.53
Además, los iniciadores ISSRs en
comparación con los RAPDs, permiten temperaturas más estrictas, que proporcionan una
mayor reproducibilidad de las bandas.
El procedimiento fue aplicado por primera vez en hongos55
, luego en animales.54
Utilizándose
para encontrar polimorfismo. El método proporciona resultados altamente reproducibles y
genera abundantes polimorfismos en muchos sistemas. Los amplicones son de tamaños
diferentes, desde los 100 pb hasta los 4000 pb; los productos de PCR ser observados en gel de
agarosa teñidos con bromuro de etidio56, 57
; en gel de poliacrilamida empleando tinción de
plata, o usando la detección isotópica.58
y 59
Figura 3. Amplificación con un iniciador (CA)n anclado en el extremo 5’con tres nucleótidos extras. Se
amplifica el segmento intermedio entre dos secuencias de microsatélite en orientación invertida.60

22
3.3.5 Random Amplified polimorphic DNA (RAPDS)
ADN polimórfico amplificado aleatoriamente (RAPD) son marcadores que provienen de la
modificación de la técnica de PCR convencional utilizando ―iniciadores‖ de secuencias
arbitrarias las cuales facilitan y aceleran los estudios de organismos de secuencias
desconocidas, combinando la utilización de un solo primer en vez de un par de iniciadores y
el corto tamaño que este posee (8 a 10 nucleotidos de longitud) siendo su secuencia blanco
desconocida.
Para que se genere un fragmento RAPD es necesario que las dos hebras del ADN en estudio
presenten sitios de hibridación con el oligonucleótido en orientaciones opuestas
suficientemente cercanas (menos de 3000bp) como para permitir la amplificación. La
secuencia del oligonucleótido es aleatoria al igual que los sitios de hibridación, por lo que la
secuencia amplificada es desconocida. El polimorfismo que se observa entre distintos
individuos consiste en la presencia o ausencia de fragmentos de ADN amplificado.
Como producto de esta amplificación se obtienen bandas de diferentes tamaños que son
visualizados en gel de electroforesis ya sea teñidos con bromuro de etidio en gel de agarosa y
en presencia de luz UV, o con tinción en nitrato de plata en gel de poliacrilamida de mas alta
resolución. La usencia de banda puede indicar una mutación puntual que evita el
apareamiento del primer observándose de esta manera polimorfismo en la población.61 y 62
3.5 Variabilidad genética y filogenia del genero Jatropha
Estudios limitados se han llevado a cabo sobre la diversidad genética y filogenia del género
Jatropha. Puangpaka y Thaya (2003)64
estudiaron el núcleo de cinco especies de Jatropha
mediante la tinción de los cromosomas de microsporocitos, e informó, que la mayoría de las
especies tenían un número cromosómico de 2n = 22. Este estudio también señalo que J. auras
y J. multifida fueron cromosómicamente similares.
La cariología de J. gossypifolia se determina a partir de las primeras células en anafase
observándose en los cromosomas separados una distribución 11:11. Así, según Puangpaka y
Thaya (2003)63
, J.curcas, J. ultifida, y J. gossypifolia parecen estar estrechamente
relacionados entre sí, en función de su configuración meiótica y la similitud
morfológica. Carvalho et al. (2008)65
estudiaron el tamaño del genoma, composición de bases
nucleotidicas y el cariotipo de J. curcas. Los resultados mostraron que el valor 2C del
genoma J. curcas fue de 0,85 pg, con una composición de bases GC promedio del 38,7%. El
cariotipo de J. curcas está formado por 22 cromosomas metacéntricos y submetacéntricos
relativamente pequeñas, cuyo tamaño varía desde 1,71 hasta 1,24 um. Con base en estudios

23
citológicos e isoenzima peroxidasa, Prabakaran y Sujatha (1999)66
informó que J. tanjorensis
es un híbrido natural interespecífico de J. curcas y J. gossypifolia.
La diversidad genética y el análisis filogenético usando RAPD, AFLP y análisis nrDNA ITS
sugirió divergencia genética interespecífica de 97,74% (RAPD), 97,25% (AFLP) y 0.419 por
nrDNA ITS secuencias67
. Arboles filogenéticos construidos usando RAPD, AFLP y datos de
secuencias nrDNA ITS apoyó la mayor similitud genética entre J. curcas y J. integerrima. La
distribución geográfica de J. glandulifera es más amplia, y esta tiene morfológicamente rasgos
distintivos que separan clados mayores y menores. La afirmación de que J. tanjorensises es
un híbrido espontáneo entre J. curcas y J. gossypifolia66
no pudo ser apoyado por datos
moleculares67
. El análisis RAPD reveló que la diversidad genética entre las diferentes
especies de Jatropha es 80,2%, y confirmó la distinción con J. glandulifera. Tambien se
reporto un conjunto de marcadores RAPD podrían ser utilizados para identificar las especies
de Jatropha de cualquier población mixta y para la caracterización de hibridos
intraespecificos. 68
3.6 Diversidad genética intraespecífica en J. curcas
Los ensayos sistemáticos para la evaluación de la procedencia de J. curcas son
limitados. Accesiones provenientes de Nicaragua y Cabo Verde provenientes de diferentes
sitios en el norte de Nicaragua demostró que el genotipo de Nicaragua tiene menos ramas,
hojas más grandes y semillas de mayor tamaño, mientras que la que proviene de Cabo Verde
presenta un alto rendimiento de semilla. En el germoplasma de Nicaragua se observó una
planta macho estéril que produce más fruto que otros tipos de hermafroditas.30
La evaluación
en el rendimiento de 13 accesiones provenientes de distintos localidades de Senegal y Cabo
Verde se observaron diferencias significativas en el crecimiento vegetativo.
Los estudios genéticos similitud de las variedades tóxicas y no tóxicas mediante RAPD indicó
96.3% de similaridad.26
En otro estudio, se reportó 84,91% y 83,59% de similitud usando
RAPD y AFLP, respectivamente63
. Estudios de inter e intrapoblaciones usando RAPD junto a
ISSR (inter secuencias simples repetitivas) donde se analizo 42 accesiones de germoplasmas
de J. curcas recogidos en diferentes regiones de la India, junto con un genotipo no tóxico de
México mostró un 42% y 37,4% de polimorfismos usando RAPD e ISSR, respectivamente. 26
Ganesh Ram et al. (2008) caracterizo cinco accesiones de una región usando 18 cebadores
RAPD68
, y Ranade et al. (2008) analizaron 22 accesiones de seis regiones usando 7 RAPD y
cuatro ―Dirigida amplificación de minisatelites de ADN‖ (DAMD) iniciadores69
. Marcadores

24
específicos para poblaciones se aislaron26
, utilizando marcadores SCAR que permiten la
identificación de poblaciones particulares mediante PCR de un solo paso. Análisis de la
diversidad intraespecífica mediante RAPD y AFLP de J. curcas recogidos de diferentes
regiones geográficas de la India indicó la existencia de baja diversidad genética, el análisis de
microsatélites dentro de una población local de J. curcas mostró un muy bajo equilibrio
Hardy-Weinberg.27
3.7 Marcadores de variedades tóxico y no tóxico de J. curcas
Una variedad no toxica de Jatropha curcas proveniente de México fue reportado, en la que
sus semillas pueden utilizarse para el consumo humano después de la torrefacción. El cultivo
de variedades no tóxicas podría proporcionar aceite para el biodiesel y la torta de semilla
servir como alimento para el ganado, y así dar valor agregado a la cosecha. No hubo
diferencias significativas morfológicas, cualitativas ni cuantitativas observadas entre
variedades tóxicas y no tóxicas, excepto por el contenido de éster de forbol.70
Los marcadores
moleculares específicos de variedades tóxicas y no tóxicas, se identificaron utilizando los
iniciadores selectivos en una única reacción de PCR utilizando las técnicas de RAPD y
AFLP. Los iniciadores IDT E-18 y OPL14, y la combinación de cebadores AFLP selectiva
E-ACC/M-CAC resultaron marcadores polimórficos para ambas variedades tóxicas y no
tóxicas. Nuevos marcadores microsatélites también fueron probados por su polimorfismo en
las variedades tóxicas y no tóxicas, y 7 de los 12 marcadores a conseguido demostrar
polimorfismo.27
Marcadores SCAR aislados confirman la especificidad y reproducibilidad en
la discriminación de genotipos tóxicos y no tóxicos.26
Estos marcadores específicos para
distinguir variedades no tóxicos a partir de variedades tóxicas de Jatropha curcas podrán ser
utilizados en la selección asistida por marcadores (MAS), análisis de caracteres cuantitativos
(QTL) y otros estudios de mejoramiento asistido con marcadores moleculares.17
IV. Objetivos 4.1 Objetivos Generales:
Caracterizar la variabilidad genética del piñón blanco, J. curcas, en la costa norte del Perú.
4.2 Objetivos específicos:
Colectar accesiones de J. curcas en la costa norte (Tumbes, Piura y Lambayeque)
Identificar marcadores genéticos que permitan diferenciar a las distintas poblaciones
naturales de J. curcas presentes en la costa norte.

25
4.3 Lugar de ejecución del proyecto: La investigación se ejecutó en la Universidad
Peruana Cayetano Heredia, en la Unidad de Genómica ubicada en la Facultad de Ciencias y
Filosofía.
V. Materiales y método 5.1 Materiales
5.1.1 Material biológico
Muestras foliares de Jatropha curcas L., extrayendo las hojas jóvenes ubicadas en los
ápices de las ramas y que no presenten daños mecánicos ni causados por insectos.
5.2 Metodología
5.2.1 Colecta de las accesiones
Las poblaciones que se utilizaron para realizar este estudio se localizan en la costa norte del
Perú en los departamentos de Piura, Tumbes y Lambayeque. De cada población se colecto
tejido foliar de 50 individuos distribuidos en 5 localidades para la región Tumbes, 100
individuos distribuidos en 10 localidades para la región Piura y 50 individuos provenientes de
4 localidades de la región Lambayeque, se tomó en cuenta que las accesiones colectadas estén
a una distancia mínima de 100 m entre ellas, los puntos fueron georeferenciados con la ayuda
de un GPS (Garmin). Se tomó de cada planta las hojas más jóvenes, evitando que estén
maltratadas ya sea por daños mecánicos o causados por insectos, virus u otros. Se colocaron
las hojas frescas en un sobre rotulado y luego se introdujo en una bolsa ziploc conteniendo
silica gel para deshidratar rápidamente la muestra y evitar la acción de polifenoles que puedan
dañar el ADN de la muestra. Se realizó el mismo procedimiento para cada muestra colectada
en las distintas localidades.

26
Figura 4. Mapa de la distribución geográfica de las poblaciones analizadas de J. curcas L.
5.2.2 Extracción de ADN
La extracción de DNA se basó en una modificación al método de Doyle y Doyle (1987).74
Se
pesó la cantidad aproximada de 100 mg de tejido (completamente deshidratados con silica
gel) para cada una de las muestras. Cada una de éstas se colocaron en un microtubo nuevo y
estéril de 2ml se lisaron las paredes celulares por medio de la pulverización con nitrógeno
líquido en un mortero y la adición de buffers de extracción CTAB 2X con mercaptoetanol al
2% y CTAB 10X que sirven como detergentes para solubilizar la membrana y eliminar
azúcares, también se añade RNAsa para digerir al RNA. Con la adición de cloroformo-octanol
se eliminan restos celulares, compuestos fenólicos y otras moléculas que permanezcan en
solución; posteriormente el ADN es precipitado con la adición de isopropanol frío, seguido de
una limpieza con etanol al 70% y finalmente es re-hidratado en agua ultra pura añadiendo
200 ul por cada muestra. Además se agregó PVP (polivinil pirrolidona) para evitar aún más la
acción de polifenoles que puedan dañar la calidad del ADN de la muestra. (Para ver
detalladamente el protocolo de extracción ver anexo N°8).

27
5.2.3 Calidad y concentración del ADN
La evaluación de la calidad e integridad del ADN se llevó a cabo mediante electroforesis en
gel de agarosa cuya concentración fue de 1% en buffer TAE. Se diluyo la agarosa en buffer
TAE para luego colocarlo al microondas hasta casi hervir, se dejó enfriar hasta
aproximadamente 60°C y luego se agregaron 2 μl de bromuro de etidio (2mg/ml) para 100 ml
de buffer. Se vertió en un molde con los peines esperando hasta que gelifique; el gel listo fue
colocado en la cámara de electroforesis conteniendo buffer TAE 1X. En los pocillos del gel se
cargaron 2 μl de la muestra previamente mezclados con 8 μl de buffer de carga. Se utilizó un
ADN control y las muestras de los genotipos se corrieron por duplicado. Se aplicó corriente a
80 voltios por una hora, transcurrido este tiempo se visualizó y capturó la imagen del ADN
con la ayuda de un fotodocumentador Fotodine ®.
La concentración fue determinado usando 3ul de muestra mediante un espectrofotómetro
Nanodroop® el mismo que proporcionó directamente la relación A 260 /A 280 (tasa de
absorbancias a 260 nm sobre 280 nm), proporcionando directamente la concentración de la
muestra (ng/μl).
5.2.4 Selección de los individuos para el estudio.
De los 200 individuos colectados en los departamentos de Piura Tumbes y Lambayeque
(Anexo Nº1) se escogieron 3 individuos por cada localidad obteniéndose de esta manera 15
individuos provenientes de 5 localidades de la región tumbes, 30 individuos provenientes de
10 localidades de la región Piura y 15 individuos provenientes de 4 localidades de la región
Lambayeque las cuales fueron seleccionados de acuerdo a la uniformidad de la calidad
concentración y distribución geográfica de la colecta realizada.
5.2.5 Pre-selección de iniciadores ISSR y RAPD
Con la finalidad de elegir los iniciadores que proporcionen una mejor respuesta en términos
de amplificación de bandas nítidas y polimórficas; se realizó una preselección de un total de
53 iniciador evaluados los cuales 15 iniciadores amplificaron con ADN genómico de Jatropha
curcas utilizando las temperaturas trabajadas por Basha et al. 2007 (comunicación personal).
Además se seleccionaron 08 iniciadores RAPD (Tabla Nº 7). Se amplificaron los ADN de 3
accesiones de piñón, cada uno con su duplicado de ADN aislado independientemente (réplica
biológica).

28
5.2.6 PCR con los 60 individuos del estudio
A partir de los resultados de la pre-selección del apartado anterior, se seleccionó un grupo de
iniciadores con los cuales se realizó la amplificación del ADN de las 60 accesiones
(previamente aisladas y purificadas)
5.2.6.1 Condiciones PCR – ISSR
Para realizar las reacciones en cadena de la polimerasa (PCR), se procedieron a diluir las
muestras de ADN hasta una concentración de 10 ng/μl. Se llevó a cabo en un volumen final
de 10 μl, usando las siguientes cantidades por tubo de reacción: 20 ng de ADN; 1X PCR
buffer (10 mM Tris pH 9.0, 50 mM KCl, 1.5 mM MgCl2); 0.2 mM dNTPs; 2 mM MgCl2;
0.4 μM iniciador y 0.5 U Taq polimerasa tabla Nº6. Las reacciones se llevaron a cabo en un
termociclador My Cycler (BIO RAD®). El programa de amplificación se muestra
esquemáticamente en la Figura Nº5.
Fig.5. Programa de temperatura ingresada en el termociclador My Cycler (BIO RAD).
5.2.6.2 Condiciones PCR – RAPD
Las amplificación de los marcadores RAPD fueron realizados mediante el método de
Williams et al. 1990 descrito en el trabajo de Basha 2007. La cual se realizó en un volumen
final de 10 ul, conteniendo 2.5 ng de ADN, 1X PCR buffer (10 mM Tris pH 9.0, 50 mM KCl,
1.5 mM MgCl2), 100 uM de cada uno de los dNTPs, 0.4 uM del iniciador RAPD y 0.5 U de
Taq polimerasa. Las reacciones se llevaron a cabo en un termociclador My Cycler (BIO
RAD®), con una denaturación inicial de 94ºC durante 3 min seguido por 45 ciclos de 94ºC por
45s, 36ºC durante 30 s y 72ºC por 2 min con una extensión final de 72ºC por 7 min.

29
5.2.7 Electroforesis en gel de agarosa
Los productos que se obtuvieron del PCR con los diferentes iniciadores fueron separados por
medio de electroforesis en geles de agarosa al 2%, utilizando TAE 1X como buffer de corrida,
los geles se tiñeron con bromuro de etidio para poder ser visualizados bajo luz ultravioleta.
En cada corrida de los geles se colocó además de los individuos un marcador de peso
molecular (ladder) de 100 pares de bases marca Invitrogen de 250 µg (1.0 µg/µl) con el
objetivo de poder calcular el tamaño (en pares de bases) de los diferentes loci amplificados.
Cada gel fue fotografiado con un fotodocumentador FOTODINE® y las imágenes fueron
digitalizadas usando el programa FOTODINE 3.0.
5.2.8 Análisis Diversidad de la población.
Para un análisis real de la diversidad se tiene que tomar en cuenta el número de alelos
detectados y la frecuencia que estos presenten, por consiguiente un población que presente
un alto grado de diversidad será aquella donde la presencia y ausencia de alelos se
distribuyan equitativamente, es decir que la población presente un gran número de alelos de
frecuencias similares.
Construcción de la Matriz Básica de Datos
Se procedió a realizar la lectura de manera manual; esto es, identificando los patrones de los
diferentes loci amplificados y ordenándolos de acuerdo al tamaño en pares de bases que estos
presentaron y así poder armar una matriz con valores binarios de presencia (1) y ausencia (0)
de bandas para cada individuo y para cada iniciador. A los fragmentos con amplificación
tenue, se les asignó un valor de 9, consignándolos como datos perdidos o missing data. Esta
información de bandas se ingresó en hojas de cálculo de Microsoft Excel 97-2003 (Anexo Nº
4). En esta misma hoja de cálculo, se determinó el porcentaje de datos perdidos por cada
locus ingresado.71
Determinación de valores descriptivos.
Se calcularon las frecuencias de presencias (p) para cada loci. Se dividió la cantidad de
fragmentos amplificados (1) entre el total de individuos sin considerar los datos perdidos. La
frecuencia de ausencias (q) se obtuvo restando 1 menos el valor de p. Los loci con una
frecuencia de presencia de banda (p) menores a 0.05 se consideraron alelos raros. Los loci
con una frecuencia de presencia de banda (p) mayor e igual a 0.95 son considerados
monomórficos. Para conocer la utilidad de los marcadores moleculares, se cuentan con tres
medidas: riqueza de alelos, Polymorphic Index Content (PIC) y el poder discriminatorio de
los marcadores. Para este trabajo se calculó el PIC para cada locus con la siguiente fórmula72
:

30
fi: frecuencia de banda del i-ésimo alelo.
p: frecuencia de presencia de banda.
q: frecuencia de ausencia de banda.
Finalmente, se sumaron todos los valores PIC para cada iniciador ISSR, generando sus
respectivos índices informativos ISSR.
5.2.9 Análisis Multivariado
Con la finalidad de entender la diversidad molecular entre todos los miembros de la colección
estudiada se realizó un análisis multivariado de 93 loci (caracteres) y 60 OTUs (taxa),
ordenados en una ―matriz básica‖, siendo las filas los loci y las columnas las OTUs (Anexo
N° 5). Se utilizó el paquete estadístico NTSYS v2.1p (Rohlf, 2000). Mediante este software se
realizaron tres análisis: agrupamiento por UPGMA. En primer lugar, se calculó la similitud
entre cada par de genotipos usando los métodos para calcular la diversidad basada en
diferencias cualitativas.
Gracias a esto se logra la construcción de una ―Matriz de Similitud‖. En la comparación de
OTU i vs. OTU j, se pueden dar las siguientes cuatro posibilidades (Crisci y López, 1983) citado
en 60: coincidencia del carácter presencia (1,1) en ambos genotipos, presencia en el genotipo i y
no en el j (1,0); ausencia en el genotipo i y no en el j (0,1) y coincidencia de ausencias en
ambos genotipos (0,0). Esto se resume en la siguiente matriz:
Fig. Nº 6. Representación de los valores entre dos OTUs en la construcción de la matriz de similitud
Para obtener la matriz de similitud se recurren a diferentes coeficientes de asociación (o
coeficientes de similitud), que expresan de manera cuantitativa el parecido entre distintas
OTUs, existen varios coeficientes de similitud (Jaccard, Dice, Simple Matching, entre otros),
que permiten calcular las similitudes o las diferencias a través de operaciones matemáticas.
(Crisci y López, 1983; Rohlf, 2000).
SM o Simple matching: (a+d)/(a+b+c+d).
DICE: 2a/(2a+b+c) (Dice (1945), Nei and Li, 1979).
JACCARD: a /(a+b+c), Jaccard (1908).

31
Estos coeficientes pueden variar entre un valor mínimo de 0 (cuando el par de individuos no
tienen bandas en común) y un valor máximo de 1 (cuando todas las bandas presentes en un
individuo están presentes en el otro). Dependiendo de cuál de estos coeficientes se utilice se
acentúan distintos aspectos de la similitud, se ordenaron estas comparaciones uno a uno entre
las OTUs, y se generó una Matriz de Similitud (Anexo Nº 5). Obtenida ésta, se llevó a cabo
un agrupamiento UPGMA (Unweighted Pair Group Method using Arithmetic Averages), que
es un algoritmo jerárquico que agrupa a los taxones en grupos similares, teniendo en cuenta
las distancias geométricas entre OTUs. El resultado de este procedimiento se grafica en un
dendograma o árbol fenético. Se dibujaron dos árboles, utilizando el coeficiente de asociación
Simple matching (SM) para cada uno de los tipos de marcadores ISSR y RAPD.
5.2.10 Validación Interna.
Con la finalidad de comprobar la independencia de la matriz y el nivel de confianza,
se realizó una validación interna, mediante la elaboración de las matrices de distancia y
cofenética, El método de ligamiento que se emplea para agrupar los individuos puede generar
distorsiones en el fenograma respecto de los datos de similitud originales. La cuantificación
de dicha distorsión se realiza mediante el cálculo de un coeficiente de correlación denominado
―cofenético‖, que consiste en correlacionar la Matriz de similitud original con una Matriz
cofenética, que surge de las similitudes observadas entre los pares de individuos comparados,
en este caso, a partir de los datos del fenograma. Así se determinó el coeficiente de
correlación cofenética o de Mantel (r). Esta es conocida como Prueba de Mantel.73

32
VI. Resultados
6.1 Colecta y conservación de los genotipos de Jatropha curcas L.
Se colectaron 200 nuestras provenientes de 20 localidades a lo largo de la costa norte del Perú
en los departamentos de Lambayeque Piura y Tumbes, entre los 3º 31´48.90´´ hasta los 6º
51´22.50´´ latitud sur y entre los 80º 44´ 12.20´´ hasta los 79º 39´ 46.10´´ longitud oeste.
Los puntos geográficos y las localidades donde se realizaron las colectas se observan en el
anexo Nº 1 donde también detallan las localidades por departamento.
En los tres departamentos donde se realizaron las colectas, las accesiones fueron encontradas
mayormente en los cercos de cultivos como plátano, arroz, Maíz, Algodón, entre otros; junto
a canales y acequias. No lográndose encontrar genotipos ―silvestres‖ en ninguna de las
localidades visitadas.
6.2 Extracción y Calidad de ADN
La extracción de ADN que se basó en una modificación al método de Doyle y Doyle (1987),
proporciono los resultados que se ven en el anexo N° 8 en la que se observan los valores de la
concentración de ADN en ng/ul de las 200 muestras extraídas provenientes de los
departamentos de Piura, Tumbes y Lambayeque, la calidad del ADN extraído se determinó
mediante corrida electroforética del ADN extraído previamente teñido con bromuro de etidio
utilizando un marcador de peso molecular en la que se determino la presencia y degradación
del ADN extraído (ver Anexo N 2). Se observo que los valores varían desde 78.2 hasta los
3740 ng/ul en su concentración, además se encontró presencia de acido ribonucleico ARN el
cual se degrado utilizando enzima ribonucleasa. La digestión con esta enzima se realizó
posteriormente de la purificación con todas las 200 muestras de ADN, y se comprobó su
efectividad mediante corridas electroforéticas.
6.3 Elección de los individuos
De las 200 muestras colectadas en los departamentos de Piura Tumbes y Lambayeque Se
seleccionaron 3 individuos por cada localidad donde se realizaron las colectas, obteniéndose
60 individuos como se muestra en la tabla N° 6. Estos individuos serán empleados para los
estudios moleculares planteados en este trabajo.

33
Tabla Nº 6. Genotipos seleccionados para el estudio con los marcadores ISSR y RAPD.
Individuos seleccionados departamento de Lambayeque
Individuos seleccionados departamento de Piura
N° Código Localidad
N° Código Localidad
1 L1-1 Túcume
16 P3-3 Sullana
2 L9-5 Túcume
17 P7-7 Sullana
3 L14-10 Olmos
18 P8-8 Sullana
4 L15-11 Olmos
19 P13-13 Sullana
5 L16-12 Olmos
20 P16-16 Sullana
6 L21-17 Olmos
21 P19-19 Sullana
7 L31-23 Motupe
22 P22-22 Sullana
8 L34-25 Motupe
23 P29-29 Sullana
9 L36-27 Motupe
24 P30-30 Sullana
10 L39-29 Motupe
25 P34-34 Buenos Aires
11 L46-36 Motupe
26 P36-35 Buenos Aires
12 L47-37 Motupe
27 P40-39 Buenos Aires
13 L57-45 Reque
28 P43-42 Morropón
14 L58-46 Reque
29 P48-47 Morropón
15 L60-48 Reque
30 P49-48 Morropón
Individuos seleccionados departamento de Tumbes
31 P52-51 Morropón
32 P54-53 Morropón
N° Código Localidad
33 P58-57 Morropón
46 T3 Zorritos
34 P62-61 Chulucanas
47 T6 Corrales
35 P66-65 Chulucanas
48 T9 Corrales
36 P69-68 Chulucanas
49 T11 Corrales
37 P73-72 Chulucanas
50 T17 Corrales
38 P78-77 Chulucanas
51 T19 Corrales
39 P80-79 Chulucanas
52 T23 Corrales
40 P85-84 Chulucanas
53 T25 Corrales
41 P87-86 Chulucanas
54 T29 Corrales
42 P89-88 Chulucanas
55 T33 San Jacinto
43 P94-93 Tambo Grande
56 T35 San Jacinto
44 P96-95 Tambo Grande
57 T37 San Jacinto
45 P97-96 Tambo Grande
58 T41 Tumbes
59 T45 Corrales
60 T49 Corrales
42 45 47 50 53 55 58 60 100bp
Fig. Nº 7. Electroforesis en gel de agarosa al 1.7% de producto de PCR
en la que se evaluó el gradiente de temperatura del iniciador 807.

34
6.4 Elección de los iniciadores
Las pruebas de amplificación de los 53 iniciadores ISSR y 12 RAPD con 3 accesiones de
prueba; lograron determinar una lista de iniciadores cuyo patrón de bandas fue satisfactorio y
reproducible. Asimismo, las pruebas de gradiente de temperatura resultaron con una
considerable cantidad de bandas polimórficas (Tabla Nº7).
Tabla Nº 7. Lista de los 15 Iniciadores ISSR y 8 RAPD que dieron
buena respuesta en la amplificación de ADN en Piñón.
Marcadores Moleculares
Marcadores ISSR Marcadores RAPD
UBC807 OPG18
UBC808 OPC 18
UBC809 OPP- 3
UBC810 OPW-17
UBC812 OPV-08
UBC816 OPQ-15
UBC823 OPO-19
UBC830 OPL- 14
UBC834
UBC835
UBC836
UBC840
UBC841
UBC842
UBC861
Con el resto de iniciadores no se logró una buena amplificación (Lista de códigos no resaltado
en el Anexo N° 3); sin embargo, no se pueden descartar en un 100%, ya que no se realizaron
adicionales pruebas de amplificación PCR.
6.5 Amplificación con los marcadores ISSR y RAPDs
Las amplificaciones del ADN con todos los individuos se realizaron utilizando 15 de los
iniciadores ISSR y 8 de los iniciadores RAPD que dieron mejor respuesta en el proceso de
preselección (Tabla Nº 7).
100bp JcL1 JcL2 JcL3 JcL4 JcL5 JcL6 JcL7 JcL8 JcL9 JcJ10 JcL11 JcL12 JcL13 JcL14 JcL15 PJc1 PJc2 PJc3
Primer UBC 807

35
100bp JcL1 JcL2 JcL3 JcL4 JcL5 JcL6 JcL7 JcL8 JcL9 JcJ10 JcL11 JcL12 JcL13 JcL14 JcL15 PJc1 PJc2 PJc3 PJc4
Figura Nº 8. Electroforesis en gel de Agarosa al 1.8% mostrando los patrones de amplificación del iniciador
ISSR UBC 807 (a) y el iniciador RAPD OPW-17 con los genotipos cuyos códigos se muestran en el
lado superior del carril, el marcador de peso molecular utilizado es de 100 bp.
6.6 Análisis Diversidad de la población.
Para un análisis real de la diversidad se tiene que tomar en cuenta el número de alelos
detectados y la frecuencia que estos presenten, por consiguiente un población que presente un
alto grado de diversidad será aquella donde la presencia y ausencia de alelos se distribuyan
equitativamente, es decir que la población presente un gran número de alelos de frecuencias
similares.
Dentro de la matriz binaria que se construyó para la cuantificación de la presencia y ausencia
de bandas, se calculó la frecuencia o ausencia de los alelos, es decir presencia el numero de
(p) se dividió entre el número total de individuos, a su vez la frecuencia de ausencia de
bandas (q) se cálculo de la misma manera, sin tomar en cuenta los valores perdidos o missing
data (9). Cabe resaltar que para el análisis solo se tomaron en cuenta solo se tomaron en
cuenta todos aquellos loci que tenían un índice de (p) a partir de 0.05
Se originaron patrones de bandas (ver figura Nº 8) a las que luego se les dio los valores de (1)
presencia y (0) ausencia para construir la matriz de similitud (anexo Nº 5). En la figura Nº 8
se muestran la cantidad de loci totales amplificados por iniciador; además la cantidad de loci
polimórficos, el ―Índice de Iniciador o Índice ISSR/RAPD‖. Estos valores fueron obtenidos
después del proceso de colección de datos y exclusión de los datos perdidos. Así
mismo también se estimó el nivel de capacidad de los marcadores moleculares para detectar el
grado de polimorfismo de la población en estudio observándose que las amplificaciones con
los iniciadores ISSR (UBC-835) y RAPD (OPO-19) presentaron un índice mayor 3.56 y 4.8
respectivamente obtenidas con las 60 accesiones seleccionadas.
Primer OPW 17
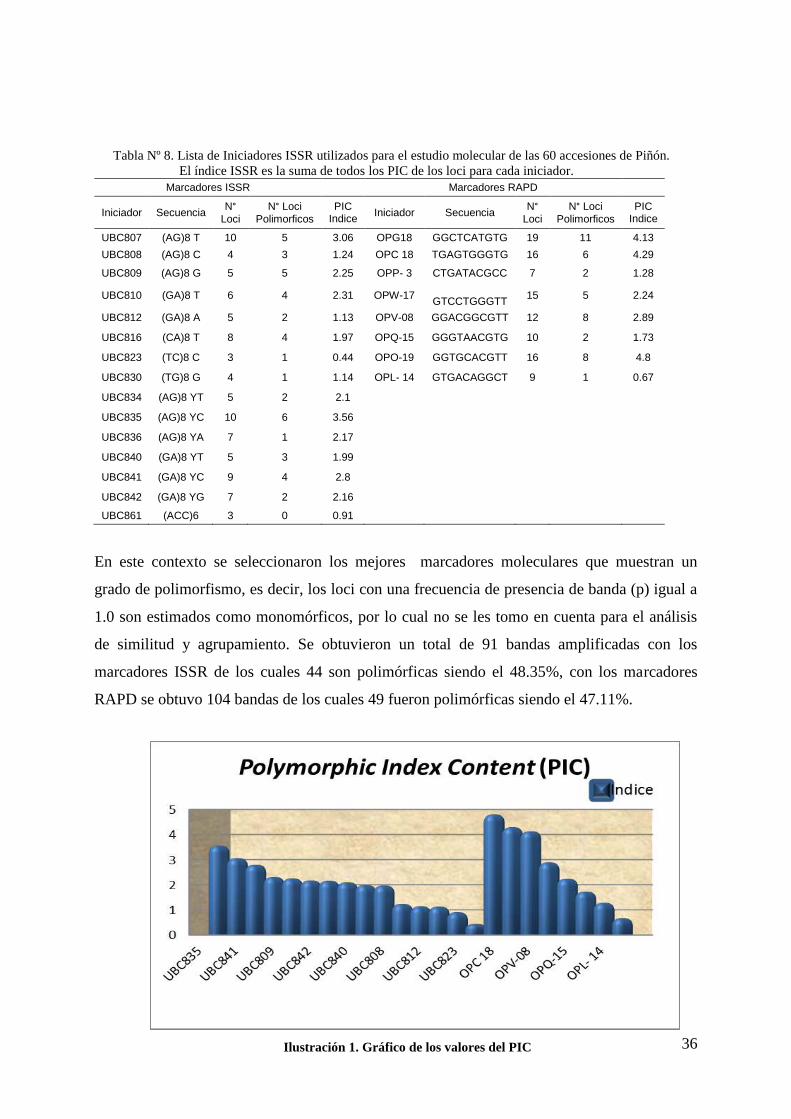
36
Tabla Nº 8. Lista de Iniciadores ISSR utilizados para el estudio molecular de las 60 accesiones de Piñón.
El índice ISSR es la suma de todos los PIC de los loci para cada iniciador.
Marcadores ISSR Marcadores RAPD
Iniciador Secuencia N°
Loci
N° Loci
Polimorficos
PIC Indice
Iniciador Secuencia N°
Loci
N° Loci
Polimorficos
PIC Indice
UBC807 (AG)8 T 10 5 3.06 OPG18 GGCTCATGTG 19 11 4.13
UBC808 (AG)8 C 4 3 1.24 OPC 18 TGAGTGGGTG 16 6 4.29
UBC809 (AG)8 G 5 5 2.25 OPP- 3 CTGATACGCC 7 2 1.28
UBC810 (GA)8 T 6 4 2.31 OPW-17
GTCCTGGGTT 15 5 2.24
UBC812 (GA)8 A 5 2 1.13 OPV-08 GGACGGCGTT 12 8 2.89
UBC816 (CA)8 T 8 4 1.97 OPQ-15 GGGTAACGTG 10 2 1.73
UBC823 (TC)8 C 3 1 0.44 OPO-19 GGTGCACGTT 16 8 4.8
UBC830 (TG)8 G 4 1 1.14 OPL- 14 GTGACAGGCT 9 1 0.67
UBC834 (AG)8 YT 5 2 2.1
UBC835 (AG)8 YC 10 6 3.56
UBC836 (AG)8 YA 7 1 2.17
UBC840 (GA)8 YT 5 3 1.99
UBC841 (GA)8 YC 9 4 2.8
UBC842 (GA)8 YG 7 2 2.16
UBC861 (ACC)6 3 0 0.91
En este contexto se seleccionaron los mejores marcadores moleculares que muestran un
grado de polimorfismo, es decir, los loci con una frecuencia de presencia de banda (p) igual a
1.0 son estimados como monomórficos, por lo cual no se les tomo en cuenta para el análisis
de similitud y agrupamiento. Se obtuvieron un total de 91 bandas amplificadas con los
marcadores ISSR de los cuales 44 son polimórficas siendo el 48.35%, con los marcadores
RAPD se obtuvo 104 bandas de los cuales 49 fueron polimórficas siendo el 47.11%.
Ilustración 1. Gráfico de los valores del PIC

37
6.7 Análisis Multivariado
6.7.1 Agrupamiento UPGMA
Sub Cluster 1.1
Sub Cluster 1.2
Sub Cluster 1.3
Sub Cluster 1.4
Sub Cluster 1.5
Sub Cluster 2.1
Sub Cluster 2.2
Sub Cluster 2.3
Sub Cluster 2.4
Coeficiente
Figura N° 10. Fenograma generado con el coeficiente de asociación Simple Matching usando
marcadores ISSR+RAPD. A un valor de 0,67 de similitud, se muestran 02 clusters en
diferentes colores. PJc1-PJc15 (Lambayeque);PJc16-PJc45 (Piura); PJc46-PJc60 (Tumbes).

38
Se generaron tres fenogramas el primero usando los marcadores ISSR, el segundo los
marcadores RAPD (Ver anexo Nº 6) y el tercero combinado los dos tipos de marcadores
ISSR+RAPD (ver figura Nº 9) para las 60 accesiones de Piñón usando el programa NTSYSpc
v2.1 y el coeficiente de asociación o similitud Simple Matching (SM).
En el fenograma construido con los marcadores ISSR+RAPD (figura Nº 9) fue el que
presento mayor coherencia en la formación de los cluster, se escogió un valor de similitud de
0.67 para resolver 02 agrupamientos o ―clusters‖; los cuales se incluyen con colores diferente
a la vez se formaron sub cluster dentro de cada agrupación.
6.7.2 Coeficiente de correlación (r)
Se obtuvo el coeficiente de correlación (r) con los marcadores ISSR+ RAPD obteniéndose el
valor de 0.7182, indicando una pobre correlación entre la matriz de similitud original y la
matriz obtenida del agrupamiento UPGMA realizado. El mayor valor obtenido es de 0.786
(ver anexo N° 7) utilizando los marcadores RAPD para los 60 genotipos. La correlación
observada con los marcadores ISSR es de 0.698 es la mas baja de los tres coeficientes de
correlación obtenidos.
Matriz
Cofenética
X Label
0.47 0.59 0.70 0.82 0.94
Y Label
0.66
0.73
0.80
0.87
0.94
Matriz de distancias
(r) = 0.71820
Figura N° 11. Comparación de las dos matrices: cofenética y de distancia de los 60 genotipos
utilizando los marcadores ISSR+RAPD.

39
VII. Discusión 7.1 Elección de los genotipos.
Las 60 muestras que se escogieron del total de las accesiones colectadas fueron seleccionadas
a base de dos criterios:
Por la calidad de ADN obtenido del proceso de extracción y purificación.
Por la distancia entre las localidades de colecta de las accesiones.
Suponiendo con estos criterios que se tomaría una muestra representativa de todas las
localidades colectadas; evitando problemas en la amplificación por la calidad de ADN.
No se encontraron diferencias morfológicas entre las accesiones colectadas, que sugieran
mayor divergencia genética, molecular y que proporcionen mayor polimorfismo en los
patrones de amplificación.
El tamaño de la muestra es significativo tomando en cuenta trabajos de caracterización
molecular en la que se consideraron 43 accesiones de Jatropha curcas L. (Basha y Sujatha
2007).26
7.2 Condiciones del ADN y la PCR
La extracción del ADN es un proceso fundamental, ya que es uno de los factores que influyen
en la reproducibilidad de los ensayos. En esta tesis, se siguió un protocolo para la extracción
del ADN basado en la metodología desarrollada por Doyle y Doyle (1987).74
Las
concentraciones de ADN (ng/μl) obtenidas variaron desde 78.2 hasta 3740 ng/μl resultando en
una amplia gama de concentraciones, pero de valores aceptables, dado que el PCR puede
utilizar una cantidad muy pequeña de ADN por reacción. Se comprobó en electroforesis en
agarosa (Anexo Nº 2) la integridad del ADN total extraído y se verificó que era necesario un
tratamiento enzimático con ribonucleasa.
En relación a las condiciones del PCR, se probaron los iniciadores a temperaturas de
amplificación trabajadas por Basha y Sujatha (Obtenidas mediante conversación personal),
los iniciadores que no mostraron un patrón definido fueron evaluados mediante gradiente de
temperatura. De esta manera se logro temperaturas óptimas de los iniciadores ISSR y RAPD
presentados en este estudio.
De las pruebas de preselección con los 53 iniciadores ISSR y 12 RAPD, se escogió un grupo
de los mejores iniciadores 15 ISSR y 8 RAPD para realizar las reacciones definitivas de
amplificación con los 60 genotipos.
De la lista de iniciadores ISSR con mejor amplificación (Tabla N°8), catorce de ellos son
iniciadores anclados y uno es no anclados. Los primeros tienen una secuencia que permite

40
el alineamiento estable al extremo 3’ o 5’ de las secuencias microsatélites la cual demostró
mayor reproducibilidad.
La electroforesis en gel de agarosa al 1.8% demostró baja resolución entre bandas con poca
diferencia en su peso molecular. Consecuentemente, la asignación de las bandas con los
valores binarios (1,0) se realizó minuciosamente y las bandas dudosas se consideraron como
datos perdidos, representados con el valor 9.
7.3 Valores descriptivos de los marcadores
La confiabilidad de la asignación de datos y la reproducibilidad de las observaciones son
cruciales para un sólido estudio de diversidad genética.
Por ello, se vio en la necesidad de considerar los datos perdidos o ―missing data‖
considerando que se deben excluir loci con más del 10% de datos perdidos, para nuestro caso
ninguno de los loci presento más del 10% de datos perdidos.
Se obtuvieron un total de 91 bandas amplificadas con los marcadores ISSR de los cuales 44
son polimórficas siendo el 48.35%, con los marcadores RAPD se obtuvo 104 bandas de los
cuales 49 fueron polimórficas siendo el 47.11%. Comparado con el estudio realizado por
Basha y Sujatha 2007 en la que trabajaron con 43 accesiones de Jatropha curcas, 100
iniciadores ISSR y 400 RAPD con los que lograron un 42% y 33.5% de polimorfismo
respectivamente, los resultados obtenidos en esta tesis es ligeramente superior.
Es posible medir la riqueza de los alelos para cada marcador, o la información que cada
marcador imparte al estudio este puede considerarse como la medida de la utilidad de cada
marcador en distinguir un individuo de otro, de esta manera el valor ―PIC‖ sirve para obtener
una noción del valor de la utilidad de los iniciadores utilizados para diferenciar los taxas.
Se sumaron los PIC de todos los loci analizados para cada iniciador, proporcionando el Índice
de Marcador para cada iniciador ISSR y RAPD. Los índices de iniciador más altos
correspondieron a los iniciadores con códigos: UBC835, UBC807 y UBC841; cuyos índices
fueron 3,56; 3,06 y 2,8 respectivamente para ISSR. Junto a los iniciadores OPO19, OPC18 y
OPG18; cuyos índices fueron 4.8, 4.29 y 4.13 respectivamente para RAPD. Lo cual señala su
valor potencial para un uso futuro en la caracterización molecular de piñón usando estos
marcadores ISSR y RAPD.
Conjuntamente con otros marcadores moleculares, estos resultados permitirían una
identificación directa y reconocimiento de ciertos genotipos valiosos.
Entonces, sería posible correlacionar las características de interés y marcadores moleculares
asociados a estos, con estudios más detallados y prolongados en segregación y herencia de los
diversos genotipos de Jatropha para comprobar la estabilidad de su herencia en varias

41
generaciones, y asegurar su utilidad como asistente molecular en un Programa de
Mejoramiento.
7.4 Agrupamiento
Con el objetivo de definir las relaciones de similitud entre los 60 genotipos de piñón
estudiados y encontrar entre ellos conjuntos consistentes y significativos de individuos, se
realizó un análisis de agrupamiento utilizando el algoritmo UPGMA. Éste se deriva a partir
de una matriz de distancias, calculada para nuestro caso con un coeficiente de asociación
(llamado también de similitud). Los cálculos de la matriz de distancia el coeficiente
simple matching ―SM‖ debido a la naturaleza dominante de los marcadores ISSR y RAPD, en
la que asumimos que tanto las presencias como ausencias en un loci determinado tienen un
mismo origen para todo el sistema.
Se examinaron tres fenogramas obtenidos con el coeficiente de similaridad Simple Matching
―SM‖ con la finalidad de lograr agrupaciones que expliquen de una manera más acertada las
relaciones de similitud y las razones de su vinculación al correspondiente cluster. Entonces, al
comparar los fenogramas generados (ISSR+RAPD: Figura N°10; ISSR y RAPD: Anexo
N°6), se observó un mejor agrupamiento en el fenograma generado por los marcadores
ISSR+RAPD. Esto significa que este último agrupamiento muestra una mejor disposición de
los cluster que correlaciona con la estructuración geográfica de las accesiones colectadas.
Entonces, bajo estos criterios, se eligió el fenograma mostrado en el Figura Nº, a un valor de
0,67 de similitud con el coeficiente Simple Matching, se distinguieron 02 clusters, en la que
los genotipos que la conforman se sobreponen geográficamente en el departamento de Piura
observándose que ningún genotipo Lambayecano se agrupa con algún genotipo Proveniente
de Tumbes, los genotipos colectados en el departamento de Piura se agrupan tanto con los
genotipos de Tumbes como de Lambayeque aunque forman sub cluster que las diferencia,
estas agrupaciones se detallan a continuación:
Cluster 1: Se encuentra todas las accesiones provenientes del departamento de Lambayeque
además de las accesiones provenientes de las localidades de Querecotillo, Salitral, Mallaritos,
El ingenio, Pampa la Hacienda, Morropón, Yapatera, Chulucanas, en el departamento de
Piura indicando que las accesiones colectadas en estas zonas tendrían un origen genético en
común.
Sub Cluster 1.1: Conformado, en su mayoría, por accesiones provenientes del
departamento de Lambayeque en las localidades de Motupe, Olmos, Tucume y Reque.
Denota coherencia con la distribución geográfica debido a la barrera geográfica que
presenta como es el desierto de Sechura. Aunque también se encuentra dos accesiones

42
provenientes de las localidades de Morropón y en el departamento de Piura debido a
que probablemente compartan un origen genético común detectado por los
marcadores.
Sub cluster 1.2: Se encuentran accesiones colectadas exclusivamente en el
departamento de Piura, observándose influencia de la distribución geográfica que los
separa de los genotipos Lambayecanos.
Sub cluster 1.3: Aquí se han agrupado accesiones provenientes de la localidad de
motupe en el departamento de Lambayeque sugiriendo diferencias que las separa del
rsto colectados en el mismo departamento.
Sub cluster 1.4: Singularmente este genotipo proveniente de Olmos (Lambayeque),
en la que las accesiones colectadas están formando parte del cluster 1.1 y 1.3; sin
embargo, se separa del resto de genotipos.
Sub cluster 1.5: Formado por las accesiones PJc17, PJc18, PJc19 ubicadas en las
localidades cercanas de Salitral y Querocotillo siendo las accesiones colectadas en la
zona mas septentrional del cluster 1.
Cluster 2: Se encuentran todas las accesiones colectadas en el departamento de Tumbes
además de las accesiones provenientes de las localidades de Chulucanas, y Tambo grande en
el departamento de Piura. Dentro de este agrupamiento se observan 4 sub cluster que se
detallan a continuación:
Sub Cluster 2.1: Conformado por accesiones provenientes del departamento de Piura
a excepción del genotipo PJc48 de la localidad de corrales en la zona sur del
departamento de Tumbes.
Sub cluster 2.2: Los miembros de este cluster provienen de colectas en zonas muy
cercanas entre si, en las localidades de Corrales y El Oidor, ubicadas en la parte mas
meridional de las colectas realizadas en el departamento de tumbes.
Sub cluster 2.3: Formado por dos accesiones PJc46 y PJc47 provenientes de Tumbes
y una accesión proveniente de Piura PJc45 siendo geográficamente diferentes al igual
que la accesión PJc48 en el cluster 2.1 lo que sugiere un flujo que podría deberse a que
los genotipos en un pasado hayan sido trasladados de un departamento a otro.
Sub cluster 2.4: Formado por los genotipos PJc48, PJc49 y PJc50 Es interesante ver
que geográficamente están localizadas cerca de las accesiones del cluster 2.2 pero
forman un cluster separado pudiéndose deber a características morfológicas no
determinadas por lo que las accesiones provienen de una colecta reciente.

43
Estas Agrupaciones sugieren una distribución longitudinal de dos poblaciones que se
interceptan en el departamento de Piura encontrándose en cada cluster que las accesiones
colectadas mas hacia el norte muestran para cada caso una agrupación que difieren con las
accesiones restantes de lo cual se puede inferir que existe una distribución proveniente de
norte sur. Esta es una hipótesis que quedaría comprobarla mediante estudios de flujo génico.
7.5 Validación interna
Con el fin de demostrar la solidez de los gráficos de agrupamiento generados, se utilizó la
prueba conocida como índice de correlación de Mantel de dos vías, en el software NTSYSpc
v2.1p, mediante la cual se puede demostrar, comparando la matriz de similitud versus la
matriz cofenética, que hay una adecuada representación de las similitudes genéticas por parte
del fenograma.
Se generaron los gráficos que se muestran en la figura Nº 10 y anexo Nº 7 los cuales señalan
la relación entre las matrices de similitud y cofenética, que surge de las similitudes
observadas entre los pares de individuos comparados. En los fenogramas generados con las 60
accesiones los coeficientes de correlación cofenética o ―r‖, resultaron siendo entre 0.69, 0.71,
0.78 para los fenogramas generado con los marcadores ISSR, ISSR+RAPD y RAPD
respectivamente. Esto indica que la técnica de agrupamiento UPGMA origina una
considerable distorsión, o dicho en otras palabras, que los agrupamientos en los fenogramas
representan modestamente los valores de similitudes genéticas entre las 60 accesiones de
Jatropha curcas L.
7.6 Análisis Poblacional
7.6.1 Porcentaje de loci polimórficos.
Se calculó el porcentaje de loci polimórficos directamente de los datos (conteo de presencias
y de ausencias) para el total de las accesiones colectadas. Observando estos resultados, el
polimorfismo es de 48.35% con los marcadores ISSR y 47.11% para RAPD. Estos valores
estarían indicando modestos niveles de variabilidad genética en la población analizada. Es
necesario continuar con los estudios con mayor número de marcadores o con marcadores mas
resolutivos como AFLP o Microsatelites.

44
VIII. CONCLUSIONES
1. Los marcadores moleculares ISSR y RAPDs demostraron ser eficientes en la
caracterización de la variabilidad genética de Jatropha curcas, obteniéndose 15
iniciadores ISSR y 8 RAPDs cuyo índice de contenido polimórfico reveló su alto valor
informativo.
2. Se observó una modesta variabilidad genética en la población analizada conformada
por 60 accesiones colectadas en los departamentos de Tumbes Piura y Lambayeque,
además los fenogramas construidos muestran una ―pobre‖ correlación entre la matriz
de similitud original con la Matriz cofenética.
3. Se logró encontrar la formación de dos clusters, en la que los genotipos que la
conforman se interceptan geográficamente en el departamento de Piura observándose
que ningún genotipo Lambayecano se agrupa con algún genotipo Proveniente del
departamento de Tumbes
4. Todas las accesiones se encontraron asociadas a otros cultivos como arroz, algodón,
plátano, entre otros; formando cercos vivos para la separación entre fincas, al borde de
caminos o carreteras en los departamentos de Tumbes, Piura y Lambayeque en la que
se realizaron las colectas.

45
IX. Recomendaciones
1. Ensayar con marcadores mas resolutivos como AFLP o microsatelites, afín de validar
los resultados obtenidos en esta tesis.
2. Utilizar la colección completa de 200 accesiones en estudios con los mismos y otros
marcadores para mayor comprensión de la dinámica en la población.
3. Las representaciones fenéticas (por agrupamiento) de los 60 individuos de Piñon se
pueden considerar como un criterio más a considerar en los programas de
mejoramiento genético o de selección asistido por marcadores (MAS).
4. Incluir en posteriores estudios accesiones colectadas a nivel nacional y accesiones
internacionales para un mayor entendimiento de la distribución y evolución de la
especie.

46
X. Referencias Bibliográficas
1. B.K. Barnwal & M.P. Sharma, Prospects of biodiesel production from vegetables oils
in India,Renew Sustain Energy Rev 9 (2005), pp. 363–378.
2. SHAY E.G. (1993). «Diesel fuel from vegetable oil: status and opportunities».
Biomass Bioenergy 4(4): 227–42.
3. Castro,Paula, Opciones para la producción y uso del biodiésel en el Perú Lima:
Soluciones Prácticas-ITDG; 2007
4. F. Ma & M.A. Hanna, Biodiesel production: a review. Bioresour. Technol. 70 (1999),
pp. 1–15.
5. VAN GERPEN J.«Biodiesel processing and production». Fuel Processing
Technology 86. (2005): 1097–1107.
6. Goering CE, Schwab AW, Daugherty MJ, Pryde EH, Heakin AJ. Fuel properties of
eleven vegetable oils.Trans ASAE 1982;85:1472–8
7. Baukje Bruinsma.(2009). Producción de biodiesel de palma aceitera y jatropha en la
Amazona del Perú y el impacto para la sostenibilidad, Un Análisis Sostenible del
Ciclo de Vida.
http://dspace.ou.nl/bitstream/1820/3116/2/NWBBruinsma%20spaanse%20versie.pdf
revisado el 12-08-2010.
8. A. Kumar & S. Sharma, An evaluation of multipurpose oil seed crop for industrial
uses (Jatropha curcas L): a review, Ind Crop Prod 28 (2008), pp. 1–10.
9. Heller J. Physic nut. Jatropha curcas L. Promoting the conservation and use of
underutilized and neglected crops. 1. PhD dissertation, Institute of Plant Genetic and
Crop Plant Research, Gatersleben, Germany, and International Plant Genetic Resource
Institute, Rome, Italy, 1996
10. D.N. Tewari, Jatropha and biodiesel (1st ed), Ocean Books Ltd, New Delhi (2007).
11. A.J.S. Raju & V. Ezradanam, Pollination ecology and fruiting behaviour in a
monoecious species, Jatropha curcasL. (Euphorbiaceae), Current Science 83 (2002),
pp. 1395–1398
12. A.R. Prakash, J.S. Patolia, J. Chikara & G.N. Boricha, Floral biology and flowering
behaviour of Jatropha curcas,FACT seminar on Jatropha curcas L. agronomy and
genetics, Wageningen, The Netherlands, March 26–28, FACT Foundation,
Wageningen (2007) Article no. 2
13. W.M.J. Achten, L. Verchot, Y.J. Franken, E. Mathijs, V.P. Singh & R. Aerts et al.,
Jatropha bio-diesel production and use, Biomass Bioenergy 32 (2008), pp. 1063–1084.
14. Wink, M., Koschmieder, C., Sauerwein, M & Sporer, F., 1997.Phorbol esters of J.
curcas—biological activities and potential applications. In: Gubitz, G.M., Mittelbach,
M., Trabi, M. (Eds.),Biofuels and Industrial Products from Jatropha curcas. DBV
Graz, pp. 160–166.
15. Makkar, H.P.S., Becker, K., Sporer, F. & Wink, M., 1997. Studies on nutritive
potential and toxic constituents of different provenanaces of Jatropha curcas. J Agric.
Food Chem. 45,3152–3157.

47
16. JM. Watt & Brandwijk MG Breyer, (1962) las plantas medicinales y venenosas de
África meridional y oriental. , E & S Livingstone, Edinburgh (1962)
17. K. G. Ramawat.(2010).Desert Plants: Biology and Biotechnology Springer BerlinCY.
BerlinY1.
18. Morton Jr (1981) Atlas of medicinal plants of middle America. Development study of
Jatropha curcas L. (Sabu Dum) oil as a substitute for diesel engine oil in Thailand. 1
Agric Assoc China 120:14
19. Duke JA & Wain KK (1991) Medicinal plants of the world. USDA Computer listing
of folk medicine of the world.
20. Staubmann R. Foidl G, Foidl N, Gubitz GM. Lafferty RM, Arbizu VM & Steiner W
(1997) Biogass production from Jatropha (arras cake. Appl Biochem Biotechnol
63:457-467
21. Karve AD (2005) Compact biogas plant-compact. low-cost digcstcr for biogas from
wastc starch. Mimeo.
22. Visser J. & Adriaans T. (2007) Anaerobic digestion of Jatropha curcas press cake.
Report produced for FACT, Ingenia, Eindhoven
23. Wan IM. Breyer-Brandwijk MG (1962) The medicinal and poisonous plants of
Southern and Eastern Africa. 2nd cdn. Livingstone. Edinburgh
24. Tewari JP. & Shukla K (1982) inhibition of infectivity of two strains of watermelon
mosaic virus by latex of some angiosperms. Geobios 9:124-126
25. Makkar HPS & Becker K (1997) Potential of Jatropha seed cake as protein
supplement in livestock feed and constraints to its utilization. In: Proceedings of the
first international symposium on biofuel and industrial products from .Iatropha curcas
and other tropical oil seed plants. 23-27 February 1997. Managua, Nicaragua, pp 23-
27
26. Basha SD & Sujatha M (2007) Inter- and intra-population variability of J. curcas (L.)
characterized by RAPD and ISSR markers and development of population-specific
SCAR markers. Euphyilea 56:375-386
27. Sudheer PDVN. Singh S. Shaik MG. Patel J. & Reddy MP (2009) Molecular
characterization and identification of markers for toxic and non-toxic varieties of
Jatropha curcas L. using RAPD. AFLP and SSR markers. Mol Riol Rep 36:1357-
1364. doi 10.1007/s11033-008- 9320-6
28. Mandpe S. Kadlaskar S. Degen W. Keppeler S (2005) On road testing of advanced
common rail diesel vehicles with biodicscl from the Jatropha curcas plant. Soc
Automat Eng 26: 356-364
29. Ghost, A, Chaudhary DR. Reddy MR Rao SN, Chikara J. Pandya 1B, Patolia IS,
Gandhi MR. Adimunhy S. Vaghela N, Mishra S. Rathod MR. Prakash AR. Shethia
BD. Upadhyay SC, Balakrishna V. Prakash CHR, Ghosh PK (2007) Prospects for
Jatropha methyl ester (biodiesel) in India. Int 1 Environ Stud 64:659-674
30. Heller J (1996) Physic nut. Jatropha curcas L. Prmoting the conservation and use of
underutilized and neglected crops. International Plant Genetic Resources Institute,
Rome, Indy.
http://books.google.com.pe/books?id=NgrKy17M6bgC&lpg=PA1&dq=Physic%20nut

48
%2C%20Jatropha%20curcas%20L.&hl=es&pg=PA1#v=onepage&q&f=false,
Revisado el 18-08-2010.
31. Claudio Donoso Zegers. 2004 Variación intraespecífica en las especies arbóreas de los
bosques templados de Chile y Argentina 1era ediccion. Santiago de Chile. Editorial
Universitaria. 2004.
32. BAUDOIN, J.P. Genetic resources, domestication and evolution of lima bean,
Phaseolus lunatus. Kluwer Academic Publishers. Dordrecht, NL; Boston, MA, USA;
London, GB. 1988. p. 393-407
33. Franco, T. L. e Hidalgo, R. (eds.). 2003. Anilisis Estadistico de Datos de
Caracterización Morfológica de Recursos Fitogenéticos. Boletin tecnico no. 8,
Institute internacional de Recursos Fitogenéticos (IPGRI), Cali, Colombia. 89 p.
34. Ford-Lloyd, B. & Jackson, M. 1986. Plant genetic resources An introduction to their
conservation and use. Edward Arnold, Publ. Baltimore. 152. p.
35. John Ramsay MacKinnon. Manejo de áreas protegidas en los trópicos. 1era edición.
1990. IUCN.314.p.
36. Richard B. Primack & Joandomènec Ros. Introducción a la biología de la
conservación. 1era edición Editorial ARIEL. España. 2002. 470.p.
37. Mills, LS & Allendorf, FW (1996).Restoration Ecology: A Synthetic Approach to
Ecological Research. Cambridge University Press, Cambridge. 205-220.
38. Bryant, D.; Burke, L.; McManus) & Spalding. M. (199S): Reefs at Risk: A Map.Based
Indicator of Threats to the World's Coral Reefs. World Resources Institute,
Washington. D. C.
39. Grant, PR & Grant, BR (1992): ''Darwin's finches: Genetically effective population
sizes'', Ecology, 73: 766-784.
40. Laurence Martin Cook. Genética de poblaciones.1era edición. Edición Omega.
Barcelona. España 1979. 91.p.
41. Passarge. Color atlas of genetics. 3era edición. Editorial Medica Panamericana.
Madrid. España. 2007. 545.p.
42. José Luque Cabrera. Texto ilustrado de biología molecular e ingeniería genetic. 1era
edición. Editorial Harcourt. España. 2001. 467.p.
43. Daniel L. Hartl,Elizabeth W. Jones. Genetics: analysis of genes and genomes. 7ma
Edition, McGraw-Hill. 2009. 763. P.
44. Glaubitz, JC. & Moran, GF. 2000. Genetic tools: the use of biochemical and molecular
markers. In Young, A; Boshier, D; Boyle, T (Eds). Forest Conservation Genetics,
Principles and Practice. Collingwood, AU. CSIRO Publishing. 352 p.
45. Ferreira, M. & Grattapaglia, D. 1998. Introducción al uso de marcadores moleculares
en el analisis genetico. EMBRAPA. 218 p.
46. Becerra, V. & Paredes, M. 2000. Use of biochemical and molecular markers in genetic
diversity studies.]-Agr. tec. 60: 270–281
47. Araya, E; Murillo, O; Aguilar, G & Rocha, O. 2005. A ADN extraction protocol and
initial primer screening in Hyeronima alchorneoides Fr. All. for AFLP applications.
Foresta Veracruzana 7:1-4.

49
48. Andrew Kalinski. Application of Rflp & Rapd Molecular Technologies to Plant
Breeding: A Project Listing & Bibliography. Diane Pub Co. Baltimore. North
America. 1994. 166.p.
49. Franco Alirio Vallejo Cabrera. Mejoramiento genético de plantas. Universidad
nacional de Colombia. Colombia 2002.
50. Antonio M. de Ron Pereira. Actas de mejora genética vegetal. Universidade de
Santiago de Compostela. Servizo de Publicacións e Intercambio Científico. 2000.
191.p.
51. Caetano-Anolles, G. & Gresshoff, PM. 1997. DNA markers protocols, applications,
and overviews. Wiley-liss, Inc. New York. 364 p
52. Zehdi S, Sakka H, Rhouma A, Old Mohamed Salem A, Marrakchi M, Trii M. African
Journal of Biotechnology 2004; 3(4): 215
53. Zietkiewicz, E.; A. Rafalski; D. Labuda. 1994. Genome finger-printing by Simple
Sequence Repeat (SSR)-anchored polymerase chain reaction amplification. Genomics
20:176-183.
54. Gupta AK, Sauder DN, Shear NH (1994) Antifungal agents: An overview. Part I. J
Am Acad Dermatol 30: 677-698.
55. Meyer, W., T.G. Mitchell, E.Z. Freedman & R. Vilgays, 1993. Hybridization probes
for conventional DNA fingerprinting used as single primers in the polymerase chain
reaction to distinguish strains of Cryptococcus neoformans. J Clin Microbiol 31:
2274–2280
56. Joshi, S.P., V.S. Gupta, R.K. Aggarwal, P.K. Ranjekar & D.S. Brar,2000. Genetic
diversity and phylogenetic relationship as revealed by inter-simple sequence repeat
(ISSR) polymorphism in the genus Oryza. Theor Appl Genet 100: 1311–1320.
57. Nagaoka, T. & Y. Ogihara, 1997. Applicability of inter-simple sequence repeat
polymorphisms in wheat for use as DNA markers in comparison to RFLP and RAPD
markers. Theor Apppl Genet 94: 597–60
58. Blair, M.W., O. Panaud & S.R. McCouch, 1999. Inter-simple sequence repeats (ISSR)
amplification for analysis of microsatellite motif frequency and fingerprinting in rice
(Oryza sativa L). Theor Appl Genet 98: 780–792
59. Ashwani Kumar. Plant Tissue Culture And Molecular Markers: Their Role In
Improving Crop. 1era edicion. I.K. International. New Delhi. India. 2009. 688.p.
60. Chia,J. 2009. Caracterización molecular mediante marcadores ISSR de una colección
de 50 árboles clonales e híbridos de cacao (Theobroma cacao L.) de la UNAS-Tingo
María. Tesis de Maestria. Universidad Nacional Mayor De San Marcos.118.p.
61. Caetano-Anollés, G.; Bassam, B.J.; Gresshoff, P.M. 1991. DNA amplification
fingerprinting: a strategy for genome analysis. Plant Mol. Biol. Rep. 9: 294-307
62. Williams J G K, A E Kubelik, K J Levak & S. C. Tingey (1990) DNA polymorphisms
amplified by arbitrary primers are useful as genetic markers. Nucleic Acids Res
18:6531-6535.
63. Sudheer PDVN. Singh S. Shaik MG. & Patel Reddy MP (2009b) Molecular
characterization and identification of markers for toxic and non—toxic varieties of
Jatropha curcas L. using RAPD. AFLP and SSR markers. Mel Rio! Rep 36:1357-1364

50
64. Puangpaka SS & Thaya (2003) Karyology of Jatropha (Euphorhiaceae) in Thailand.
Thai For Bull (Bot) 31:105-112
65. Carvalho CR, Clarindo W, Praca MM, Araujo FS. & Cards N (2008) Genome size,
base composition and karyotype of /atrophy tiaras L.. an important bloke! plant. Plant
Sci 174:613-617
66. Prabakaran AJ. & Sujatha M (1999) Jatropha tanjorensis Ellis & Saroja: a natural
interspecilic hybrid occurring in Tamil Nadu, India. Genet Resour Crop Evol 46:213-
218
67. Sudheer DVNP, Pandya N. Reddy MP. & Krishnan TR (2009c) Comparative study of
interspecific genetic divergence and phylogenic analysis of genus Jatropha by RAPD
and AFLP. Mot Biol Rep 36:901-907, doi: 10.1007/s11033-008-9261-0
68. Ganesh Ram S, Parthiban KT, Senthil KR, Thimvengadam V. & Paramathma M
(2008) Genetic diversity among Jatropha species as reveled by RAPD markers. Genet
Resour Crop Evol 55:803409
69. Ranade SA. Srivastava AP. Rana IS. Srivastava J. & Tuli R (2008) Easy assessment of
diversity in Jatropha curcas L. plants using two single-primer amplification reaction
(SPAR) methods. Biomass Bioenergy 32:533-540
70. Makkar HPS & Becker K (1997) Potential of Jatropha seed cake as protein
supplement in livestock feed and constraints to its utilization. In: Proceedings of the
first international symposium on biofuel and industrial products from .Jatropha tiaras
and other tropical oil seed plants. 23-27 February 1997. Managua. Nicaragua. pp 23-
27
71. WARBURTON, M. & CROSSA, J. 2002. Data Analysis in the CIMMYT Applied
Biotechnology Center. For Fingerprinting and Genetic Diversity Studies,
http://ufdc.ufl.edu/UF00077469/00001 visitado el 27-07-2010.
72. GHISLAIN, M.; ZHANG, D.; FAJARDO, D.; HUAMAN, Z. & HIJMANS, R.J.
1999. Marker assisted sampling of the cultivated Andean potato Solanum phureja
collection using RAPD markers. Genetic Resources and Crop Evolution 46: 547-555.
73. ROHLF, F.J. 2000. NTSYSpc Numerical Taxonomy and Multivariate Analysis
System, Version 2.1. User Guide. Department of Ecology and Evolution. State
University of New York, Stony Brook, NY.
74. DOYLE, J.J. & DOYLE, J.L.1987. A rapid DNA isolation procedure for small
quantities of fresh leaf tissue. Phytochemical Bulletin 19:11-15.
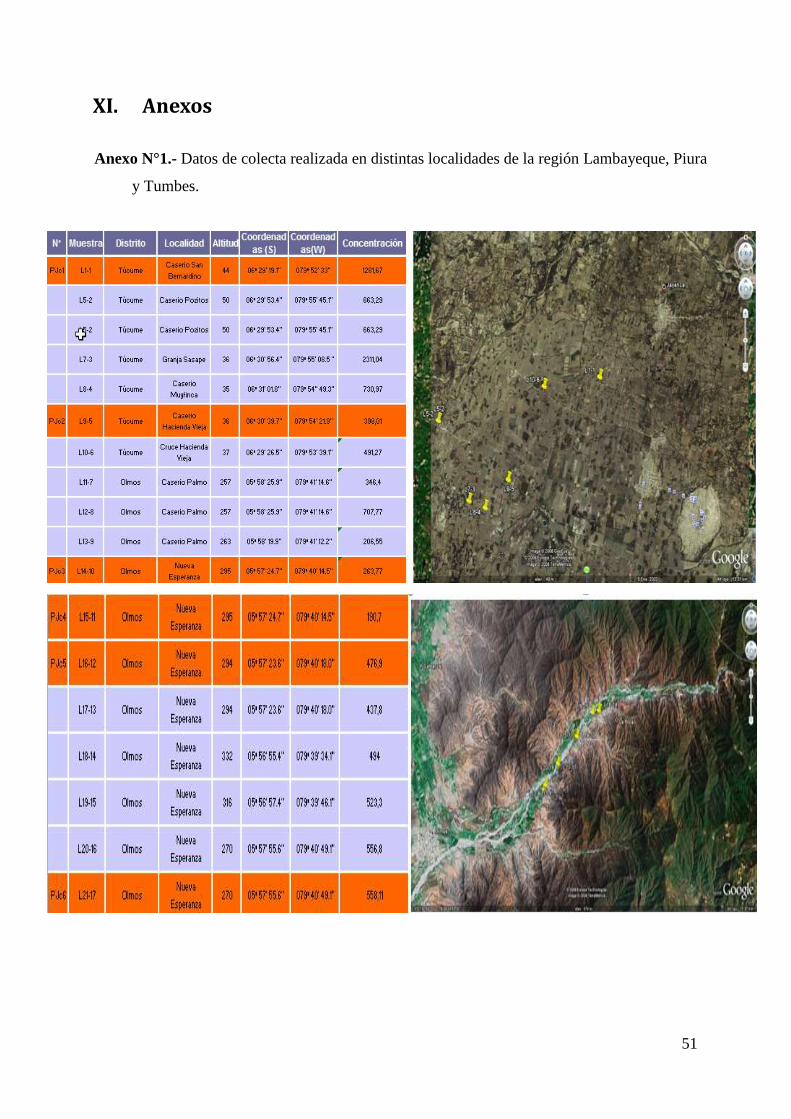
51
XI. Anexos
Anexo N°1.- Datos de colecta realizada en distintas localidades de la región Lambayeque, Piura
y Tumbes.

52

53

54

55

56

57

58

59
Anexo 2.- Electroforesis del ADN de muestras de las regiones Piura, Tumbes y
Lambayeque
GEL 15-10(M)Fila 1
100
1281,67
663,29
2311,04
730,97
398,01 346,4
707,77
206,55 190,7
467
100
476,9437,8
494523,3
558,11
556,8491,27
263,77
0
500
1000
1500
2000
2500
ML1-1 L5-2
L7-3 L8-4 L9-5
L10-6
L11-7
L12-8
L13-9
L14-1
0
L15-1
1
L16-1
2
L17-1
3
L18-1
4
L19-1
5
L20-1
6
L21-1
7
L27-1
9 M
GEL 15-10 (M)Fila 2
431,1
100
648,77
415,1463,3
375,3429,25
202,8249,4
287,5
417,44364,3380,9
248
99,15
190,2244
100
196,65
102,35
0
100
200
300
400
500
600
700
L28-20 M
L29-21
L30-22
L34-25
L35-26
L36-27
L38-28
L39-29
L44-34
L46-36
L50-39
P2-2P4-4
P13-13
P14-14
P16-16
P18-18
P19-19 M

60
GEL 14-10 (T)
100
1013,6
1347
970,7
636,7
1228,6
583,6
888,4
1792,8
1251,2
840,7
1503,9
2151,1
1180,4
1863,9
621,4
1054,3
720,6
100
1915,5
1235,41266,7
1379,2
860,3805,3
770,4668,8
658,4
892,2
948,1
0
500
1000
1500
2000
2500
M
P40-39
P41-40
L55-43
L42-32P7-7
P11-11
P9-9
L52-41P5-5
L32-24P6-6
L60-48
L51-40
L61-49
L58-46
L31-23
L62-50
L47-37P3-3
L41-31
L45-35
L57-45
P42-41
L59-47
L22-18P8-8
P1-1
P10-10 M
GEL 16-10(M)Fila 1
176,1
674,7
168
539,9
100
1347
457,9
158,6
679,4802,2
1450,4
100325.6
304245
148,2
948,11012,7
1135,61013,6
0
200
400
600
800
1000
1200
1400
1600
P21-21
P22-22
P27-27
P29-29
P30-30
P31-31
P32-32
P34-34
P40-39 M
P41-40
P42-41
P43-42
P44-43
P45-44
P46-45
P47-46
P48-47
P49-48 M
GEL 16-10 (M)Fila 2
100
667,2
87,3
1362,8
2128,3
898,1
1436,3
1977
1090,7
481,3
100
15011519,1
1789,8
204,9229,3
233,8269,4
338,7468,4
0
500
1000
1500
2000
2500
M
P50-49
P51-50
P52-51
P53-52
P54-53
P55-54
P56-55
P57-56
P58-57
P59-58
P60-59
P61-60
P62-61
P63-62
P64-63
P65-64
P66-65
P67-66 M

61
GEL 17-10(M)Fila 2
100
860 955,6
400,26
1988,2
191,5
1490,7
2214,2
272,2100
0
500
1000
1500
2000
2500
M
T32
-32
T33
-33
T34
-34
T35
-35
T36
-36
T37
-37
T38
-38
T39
-39 M
GEL 16-10 (T)Fila1
100
471,7
160,6
334,4
910,25
462,6
256,2
800,3
363,2
78,2
199,3
1111,1
491,9
573,7
358,8369,5
100120,3
676,6725,1
0
200
400
600
800
1000
1200
M
P69-68
P70-69
P73-72
P75-74
P78-77
P79-78
P80-79
P82-81
P84-83
P85-84
P86-85
P87-86
P88-87
P89-88
P90-89
P91-90
P92-91
P93-92 M
GEL 16-10(T)Fila 2
100
888,4
3740
1368,41713,9
2929,8
639,5 773,2436,4
475,3
315,9 100151,2238,3309,5
231,5491,17
751,3480,2
836
0500
1000150020002500300035004000
M
P94-93
P95-94
P96-95
P97-96
P98-97
P99-98
P100-99
P101-100
T1-1T2-2
T3-3T4-4
T6-6T7-7
T8-8T9-9
T10-10
T11-11 M
GEL 17-10(M)Fila 1
100157,5 155 210,4
519,7
373,9 398
1079,2
398,4
705
520,3
278,4
620,3
258356,9 364
824,9
350,1
512,4
100
0
200
400
600
800
1000
1200
M
T13-13
T15-15
T16-16
T17-17
T18-18
T19-19
T20-20
T21-21
T22-22
T23-23
T24-24
T25-25
T26-26
T27-27
T28-28
T29-29
T30-30
T31-31 M

62
GEL 17-10(T)Fila 1
100
1542,2
215,3108
1066
165,9318,05
1061,65
145,99335,8
100
1467,5 1498,7
1001,1
1139,61060,3
471,5638,5
837,6937,6
0200400600800
10001200140016001800
M
T40-40
T41-41
T42-42
T43-43
T44-44
T45-45
T46-46
T47-47
T48-48
T49-49
T50-50
R.P17-1
7
R.P36-3
5
R.P68-6
7
R.P76-7
5
R.P77-7
6
R.T12-1
2
R.P20-2
0 M
GEL 17-10(T)Fila 2
100
559,74
668 659,4
248,51
762,16
638,29
270,36
1142,03 1131,08
100
0
200
400
600
800
1000
1200
M
R.P24
-24
R.P25
-25
R.P28
-28
R.P33
-33
R.P37
-36
R.P72
-71
R.P81
-80
R.P82
-83
R.P72
-71 M

63
Anexo 3.- Lista de los 53 Iniciadores ISSR y 12 RAPD. Los códigos ISSR corresponden a
iniciadores de la Universidad de British Columbia. Los códigos resaltados pertenecen
a los 15 iniciadores ISSR y 08 RAPD cuyos perfiles de amplificación fueron más
consistentes que del resto.
Iniciadores ISSR
UBC802 ATA TAT ATA TAT ATA TG UBC851 GTG TGT GTG TGT GTG TYG
UBC807 AGA GAG AGA GAG AGA GT UBC855 ACA CAC ACA CAC ACA CYT
UBC808 AGA GAG AGA GAG AGA GC UBC856 ACA CAC ACA CAC ACA CYA
UBC809 AGA GAG AGA GAG AGA GG UBC860 TGT GTG TGT GTG TGT GRA
UBC810 GAG AGA GAG AGA GAG AT UBC861 ACC ACC ACC ACC ACC ACC
UBC812 GAG AGA GAG AGA GAG AA UBC862 AGC AGC AGC AGC AGC AGC
UBC813 CTC TCT CTC TCT CTC TT UBC864 ATG ATG ATG ATG ATG ATG
UBC815 CTC TCT CTC TCT CTC TG UBC865 CCG CCG CCG CCG CCG CCG
UBC816 CAC ACA CAC ACA CAC AT UBC866 CTC CTC CTC CTC CTC CTC
UBC817 CAC ACA CAC ACA CAC AA UBC867 GGC GGC GGC GGC GGC GGC
UBC818 CAC ACA CAC ACA CAC AG UBC868 GAA GAA GAA GAA GAA GAA
UBC820 GTG TGT GTG TGT GTG TC UBC873 GAC AGA CAG ACA GAC A
UBC821 GTG TGT GTG TGT GTG TT UBC880 GGA GAG GAG AGG AGA
UBC822 TCT CTC TCT CTC TCT CA UBC881 GGG TGG GGT GGG GTG
UBC823 TCT CTC TCT CTC TCT CC UBC885 BHB GAG AGA GAG AGA GA
UBC824 TCT CTC TCT CTC TCT CG UBC886 VDV CTC TCT CTC TCT CT
UBC825 ACA CAC ACA CAC ACA CT UBC887 DVD TCT CTC TCT CTC TC
UBC828 TGT GTG TGT GTG TGT GA UBC888 BDB CAC ACA CAC ACA CA
UBC829 TGT GTG TGT GTG TGT GC UBC889 DBD ACA CAC ACA CAC AC
UBC830 TGT GTG TGT GTG TGT GG UBC891 HVH TGT GTG TGT GTG TG
UBC834 AGA GAG AGA GAG AGA GYT Iniciadores RAPD
UBC835 AGA GAG AGA GAG AGA GYC OPG18 GGCTCATGTG
UBC836 AGA GAG AGA GAG AGA GYA OPV17 ACCGGCTTGT
UBC840 GAG AGA GAG AGA GAG AYT OPC 18 TGAGTGGGTG
UBC841 GAG AGA GAG AGA GAG AYC OPG 17 ACGACCGACA
UBC842 GAG AGA GAG AGA GAG AYG OPK-01 CATTCGAGCC
UBC843 CTC TCT CTC TCT CTC TRA OPP- 3 CTGATACGCC
UBC844 CTC TCT CTC TCT CTC TRC OPW-17 GTCCTGGGTT
UBC846 CAC ACA CAC ACA CAC ART OPV-08 GGACGGCGTT
UBC847 CAC ACA CAC ACA CAC ARC OPQ15 GGGTAACGTG
UBC848 CAC ACA CAC ACA CAC ARG OPO19 GGTGCACGTT
UBC849 GTG TGT GTG TGT GTG TYA OPL 14 GTGACAGGCT
UBC850 GTG TGT GTG TGT GTG TYC OPR 8 CCCGTTGCCT

64
PJc
1
PJc
2
PJc
3
PJc
4
PJc
5
PJc
6
PJc
7 P
Jc8
PJc
9
PJc
10
PJc
11
PJc
12
PJc
13
PJc
14
PJc
15
PJc
16
PJc
17
PJc
18
PJc
19
PJc
20
PJc
21
PJc
22
PJc
23
PJc
24
PJc
25
PJc
26
PJc
27
PJc
28
PJc
29
PJc
30
PJc
31
PJc
32
PJc
33
PJc
34
PJc
35
PJc
36
PJc
37
PJc
38
PJc
39
PJc
40
PJc
41
PJc
42
PJc
43
PJc
44
PJc
45
PJc
46
PJc
47
PJc
48
PJc
49
PJc
50
PJc
51
PJc
52
PJc
53
PJc
54
PJc
55
PJc
56
PJc
57
PJc
58
PJc
59
PJc
60
807-1 1 1 1 0 1 1 1 1 1 0 1 1 1 1 1 1 1 1 1 1 0 0 0 0 1 0 0 1 0 0 1 0 0 0 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
807-3 1 1 1 1 1 1 1 0 1 1 1 0 0 1 1 0 0 0 0 1 1 0 0 1 1 1 1 1 0 1 1 1 1 1 0 1 1 1 1 1 0 1 1 1 1 1 1 1 0 1 1 1 1 0 0 0 0 1 1 1
807-5 1 1 1 1 1 1 0 0 0 0 1 1 1 0 1 1 0 0 0 1 1 1 1 1 1 1 1 1 0 1 1 1 1 1 1 1 1 1 0 1 0 0 0 0 1 0 0 0 0 0 0 1 0 1 1 0 0 1 1 1
807-8 1 0 0 1 1 1 1 1 1 1 1 1 1 1 1 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 1 1 1 1 0 0 0 0 1 1 1 1 1 0 1 1
807-9 0 0 1 1 1 1 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
808-1 1 1 0 0 0 0 0 1 0 0 0 1 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
808-2 1 0 1 1 1 0 0 0 0 1 0 0 0 0 1 0 0 0 0 0 1 0 0 0 0 1 0 1 0 0 0 0 1 0 0 1 1 0 1 1 1 1 1 1 0 1 1 1 1 1 0 0 0 0 0 0 1 0 0 0
808-3 1 1 1 1 1 1 1 1 0 1 1 1 1 1 1 1 0 1 0 1 1 0 1 1 1 1 1 1 0 0 1 1 1 1 1 1 1 1 1 1 1 1 0 1 1 1 0 1 1 1 1 1 1 1 1 1 1 1 1 1
809-1 1 1 1 0 1 1 1 1 1 0 0 0 1 1 1 1 1 1 0 1 1 0 0 1 1 0 1 1 1 1 1 1 1 1 1 1 1 1 0 1 1 1 1 1 1 1 1 1 1 1 0 1 1 1 1 1 1 1 0 1
809-2 1 1 1 0 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 1 1 1 1 1 1 1 1 1 1 1 0 1 1 1 1 1 1 1 1 1
809-3 1 1 1 0 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 1 1 0 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 1 1 1 1 1 1 1 1 1 1 1 0 1 1 1 1 1 1 1 1 1
809-4 1 1 1 0 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 1 1 0 0 0 1 1 1 1 1 1 1 1 1 1 1 1 0 1 0 1 1 1 1 1 1 1 1 1 1 0 0 1 1 1 0 1 1 1 1 1
809-5 1 1 1 0 1 1 1 1 1 1 1 0 1 1 1 1 0 0 0 1 1 1 1 1 1 1 1 1 1 1 1 0 1 1 1 0 0 1 0 9 1 1 1 1 1 1 1 1 1 9 0 1 1 0 1 1 1 1 1 1
810-1 1 0 1 0 1 0 1 0 1 1 1 1 1 9 1 9 9 0 1 1 1 1 1 1 1 0 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
810-2 1 0 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 0 0 1 1 1 1 1 1 1 1 9 1 1 0 1 1 1 1 1 1 1 1 1 1
810-4 0 1 0 0 0 0 0 0 0 1 1 0 0 0 0 0 0 0 1 1 1 1 0 0 1 1 0 0 0 1 1 1 1 0 0 0 0 0 1 1 1 1 1 1 0 1 1 1 0 1 0 0 1 1 1 1 1 0 0 1
810-6 1 0 1 1 1 1 1 1 1 1 1 1 1 0 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 1 1 0 1 1 1 1 1 1 1 1 1 9 1 1 9 1 1 1 1 1 1 1 1 1 1
812-3 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 1 0 0 0 1 1 1 0 1 1 1
812-5 1 1 1 1 1 0 1 0 1 1 1 0 1 1 1 1 1 1 1 1 0 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 1 1 1 1 1 0 1 1 1 0 1 1 1
816-1 0 1 1 0 1 1 1 1 1 1 1 1 1 1 0 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 1 1 1 1 1 1 1 1 1 1 1 1 1 1
816-2 0 1 1 0 1 1 1 1 1 1 1 0 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 0 1 1 1 1 1 0 1 1 1 1 1 0 0 1 0 1 1 1 1 1
816-4 0 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 1 1 1 1 1 1 1 1 1 1 0 1 1 0 0 1 1 1 1 0 1 1 0 1 1
816-6 0 1 1 1 1 0 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 1 1 1 1 1 1 1 1 1 0 0 1 1 1 1 1 1 1 1 1 1 1 1 0 1 1 1
823-1 0 1 1 1 1 9 9 1 0 1 1 1 1 1 1 1 9 1 1 1 1 1 1 1 1 1 1 1 1 0 1 1 1 1 1 1 1 0 1 1 1 1 1 1 1 1 1 1 0 9 1 1 1 1 1 1 1 1 1 1
830-1 1 1 1 0 1 1 1 1 0 0 1 1 0 0 1 1 0 0 0 1 1 1 1 1 0 0 1 1 1 0 0 1 1 0 1 0 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 1 1 1 1
834-2 0 0 1 1 1 1 1 1 1 0 1 1 1 1 1 1 1 1 1 0 1 0 0 1 0 1 0 0 1 1 1 1 1 0 1 0 0 0 1 1 1 1 1 1 1 1 1 1 1 0 1 0 1 1 1 1 1 1 0 1
834-3 1 1 1 1 1 1 1 1 0 1 1 1 1 1 1 1 1 1 0 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 1 1 1 0 0 1 1 1 0 1
Anexo 4.- Datos Binarios (presencia: 1 ó ausencia: 0) de 93 loci (caracteres) para 60 genotipos (OTUs). El código 9 es el asignado a los
datos perdidos o ―missing data‖. Se muestran sólo los loci con un porcentaje de polimorfismo mayor al mayor 5%.

65
834-4 0 0 1 1 1 1 1 1 1 0 1 1 1 1 0 1 0 0 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
835-2 1 1 1 0 1 0 1 1 1 1 1 1 1 1 1 1 0 0 1 1 1 1 1 1 1 0 1 0 1 1 0 0 1 0 1 1 1 0 1 1 1 0 1 0 1 1 1 1 1 1 1 1 1 1 0 0 1 1 1 1
835-3 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 0 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 0 1 1 1 1
835-4 0 1 0 0 1 0 1 1 0 0 1 0 1 0 1 1 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 1 0 0 1 0 1 1 0 0 1 0 1 0 1 1 0 0 0 0 1 0
835-5 1 1 1 1 1 0 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 1 1 1 1 1 1 1 1 1 1 1 0 1 1 1 1
835-7 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 0 0 1 1 1 1 1 1 1 1 1 1 0 1 1 1 1 1 1 1 0 1 1 1 0 1 1 1 1 1 1 1 1 1 1 1 1 0 0 0 1 1 1
835-9 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 0 1 1 1 1 1 1 0 1 1 1 1 1 1 1 1 1 1 1 0 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 1 1 1 1
836-3 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 0 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 1
840-1 1 1 1 1 1 0 1 0 1 1 1 1 0 1 1 1 1 1 1 1 1 1 1 1 0 1 0 1 1 1 1 0 1 1 1 1 1 1 1 0 1 1 1 1 1 0 1 1 1 1 1 1 0 1 1 1 1 1 1 1
840-2 1 1 1 1 1 1 1 0 1 1 1 1 0 1 1 1 1 1 1 1 1 1 1 1 1 1 0 1 1 1 1 0 1 1 1 1 1 1 1 0 1 1 1 1 1 0 1 1 1 1 1 1 0 0 1 1 1 1 1 1
840-4 1 1 1 1 1 0 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 1 1 1 1 1 1 1 1 1 1 1 0 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 1 1 1 1 1 1 1
841-3 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 0 0 0 0 0 0 0 0 0 1 1 1 1 1 1 1 1 1 1 0 1 1 0 1 1 0 1 1 1 1 1 1 1 0 1 0 0 0 1 1 1
841-6 1 1 1 1 1 0 0 0 0 0 1 0 0 0 0 1 1 1 0 1 1 1 1 0 1 1 1 1 1 1 1 1 1 0 0 0 0 0 0 0 1 1 0 1 1 1 0 0 1 0 1 0 1 0 1 1 1 0 0 0
841-7 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 1 1 1 1 1 1 1 1 0 1 1 0 1 1 1 1 1 1 1 1 1 1
841-8 1 1 1 1 1 1 0 1 1 0 0 1 1 1 1 1 1 1 0 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 0 0 1 0 0 0 0 0 0 0 1 0 1 1 1
842-1 0 0 1 1 1 1 1 0 1 1 1 1 1 1 1 0 1 1 1 1 1 1 1 1 1 1 0 0 0 0 1 1 0 1 1 1 1 1 1 1 1 1 1 1 1 0 0 0 0 1 1 0 1 1 1 1 1 0 1 1
842-2 0 0 1 1 1 0 1 1 1 1 1 1 1 0 0 1 1 1 1 1 1 1 1 0 0 1 0 0 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 0 1 0 0 1 1 1 1 1 1 1 1 1 1 1 1 1
OPC18-3 1 1 1 1 1 1 1 1 1 1 1 1 0 0 1 1 1 1 1 1 1 1 1 1 1 1 0 0 0 0 1 1 1 1 1 1 1 1 1 1 1 0 0 0 9 0 0 0 0 0 0 1 0 0 0 9 1 1 1 1
OPC18-4 1 1 1 1 1 1 1 1 1 1 1 1 9 1 1 1 1 1 1 1 1 1 1 1 9 1 1 1 1 0 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 1 1 1 1 9 0 0 1 1 1 1
OPC18-8 1 1 1 1 1 1 1 1 1 1 1 0 0 9 1 1 1 9 0 1 1 1 1 1 1 0 0 0 1 1 1 1 0 0 1 9 0 1 0 0 0 0 0 0 0 0 0 0 1 1 1 1 1 0 0 0 9 1 1 0
OPC18-9 1 1 1 1 1 1 0 1 1 1 1 1 1 0 1 1 0 1 1 1 1 1 1 1 0 1 1 1 1 1 1 1 1 1 1 0 0 9 0 1 1 1 1 1 1 0 9 0 0 0 0 1 1 1 1 1 1 1 1 1
OPC18-11 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 9 0 0 0 1 0 0 0 1 0 1 1 1 1 1 0 1 0 0 1 1 1 9 1 1 0 1 1 0 0 1 1 1 1 1 1 1 1 1 1 1 1
OPC18-13 1 1 1 1 1 1 1 1 9 1 1 0 0 1 1 1 1 9 1 1 1 1 1 0 1 1 1 0 1 1 1 1 1 0 1 0 0 0 0 0 0 0 0 9 9 9 0 0 0 0 1 1 0 0 1 0 0 0 0 0
OPG18-1 0 0 0 0 0 0 0 9 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 9 0 0 0 0 0 0 1 0 9 9 0 1 1 1 1 1 1 1 0 0 0 0 0 1 0 0
OPG18-2 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 9 9 1 1 9 1 0 0 0 0 0 9 0 0 0 0 0 0 1 0 0
OPG18-3 0 0 0 0 0 0 0 9 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 9 9 1 9 1 1 1 0 0 0 0 0 0 0 0 0 1 1 0
OPG18-4 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 1 0 1 0 0 0 0 0 0 0 0 0 1 0 9 9

66
OPG18-6 0 0 0 0 1 0 1 1 1 0 1 1 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 9 1 1 1 1 0 0 1 0
OPG18-7 1 1 1 0 1 1 1 1 1 1 9 1 1 1 1 9 1 9 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 1 0 1 1 1 0 1 1 0 0 0 0 0 0 0 0 1 9
OPG18-13 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 9 9 9 1 9 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 9 0 0 0 0 0 0 0 0 0 0 1 0
OPG18-14 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 1 1 1 1 1 1 1 1 1 1 1 1 1 9 9 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 1 1 0
OPG18-15 0 0 0 0 9 1 1 1 0 0 0 0 1 1 0 0 1 0 0 0 0 0 0 0 0 0 0 9 9 0 1 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 9
OPG18-17 1 0 1 1 1 1 1 1 1 1 1 1 1 1 0 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 9 0 9 0
OPG18-18 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 9 0 0
OPG18-19 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 9 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 9 1 1 1 1 1 1 1 1 1 1 1 1 0 1 0 0
OPL14-2 1 0 0 0 1 1 1 1 1 0 1 1 1 1 1 1 9 0 0 0 0 0 0 9 0 0 0 0 0 0 0 0 0 1 0 1 1 0 0 1 1 0 0 0 9 9 0 0 0 0 0 0 0 0 0 0 0 0 0 0
OPO19-1 1 1 1 1 1 1 1 0 0 1 1 0 1 1 1 0 0 0 1 1 1 1 1 1 1 1 1 1 1 1 0 1 1 1 1 1 1 1 1 1 1 9 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
OPO19-2 0 9 9 9 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 1 0 0 0 0 0 0 0 0 0 0
OPO19-3 1 0 0 1 1 0 0 0 0 0 1 0 9 9 1 9 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
OPO19-4 1 1 1 1 1 1 1 0 0 9 1 9 1 1 1 0 1 0 1 1 1 1 1 1 1 0 1 1 1 1 0 1 1 1 1 1 0 0 1 0 1 9 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
OPO19-5 1 1 1 1 1 1 1 0 0 0 1 0 1 1 1 1 0 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 1 1 1 1 1 1 0 1 1 1 1 1 1 1 1 1 1 1
OPO19-6 1 1 1 1 1 1 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 9 9 9
OPO19-9 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 1 1 1 0 0 0 0 0 0 0 0 0 0 0 1 1 0
OPO19-10 1 1 1 1 1 1 1 0 0 0 1 0 1 1 1 1 0 1 1 0 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 1 1 1 0 1 9 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
OPO19-11 1 1 1 1 1 1 1 0 0 0 1 0 1 1 1 1 1 0 1 1 0 1 1 1 1 1 1 1 1 1 1 1 1 1 0 0 1 1 1 1 1 0 1 1 1 1 1 1 1 0 1 1 1 1 1 1 1 1 1 1
OPO19-12 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 1 1 0 1 1 1 1 1 0 0 1 1 1 0 0 0 0 0 0 0 1
OPO19-13 1 1 1 1 1 1 1 1 0 1 1 0 0 1 0 0 0 0 0 1 1 1 1 1 1 0 1 1 1 1 1 1 1 1 0 0 1 1 1 1 1 0 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
OPO19-14 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 9 9 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
OPO19-16 1 1 1 1 1 1 1 0 1 1 1 0 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 1 1 0 0 0 0 0 0 0 0 9 9 0 0 0 0 0 0 0 0 0 0 0 0 9 0
OPP3-5 0 0 0 0 0 0 0 0 9 9 1 1 1 1 0 0 9 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
OPP3-7 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 0 1 1 0 0 1 1 9 1 1 1 1 1 0 1 1 0 1 1 1 1 1 1 1 1 1 1 1 0 1 1 1 1 1 1 1 1 1 1 1 1 1

67
OPQ15-6 1 1 0 0 1 1 1 1 1 0 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 0 1 1 1 1 1 0 0
OPQ15-9 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 1 1 1 1 0 1 1 1 0 1 1 1 1 0 0 0 0
OPV8-1 1 1 1 1 1 1 0 0 9 9 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 1 1 1 1 1 1 1 1 1 1 1 0 0 0 1
OPV8-2 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 9 9 9 0 0 0 0 0 0 0 0 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 1 1
OPV8-3 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 9 9 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 1 1 1 1 1 1 1 0 1 1 1 1 0 0 1 1 0 0 9 0 0
OPV8-6 1 1 1 1 1 1 1 1 0 0 0 1 1 1 0 1 1 1 1 1 1 1 1 0 0 1 0 0 0 0 0 0 0 0 0 1 1 0 1 1 1 0 1 0 0 0 0 1 1 1 1 1 1 1 1 1 1 1 1 1
OPV8-7 0 0 0 0 0 0 0 0 1 1 0 0 0 0 1 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 1 0 0 0 0 1 0 0 0 1 0 1 1 1 1 0 0 0 0 0 0 0 0 0 0 0 0 9
OPV8-8 0 0 1 0 0 0 1 1 0 0 0 0 0 0 0 1 0 0 0 1 1 1 1 0 1 1 1 1 1 1 1 1 0 1 1 1 1 0 0 0 0 0 0 0 0 0 1 1 0 0 0 0 0 0 0 0 0 0 9 0
OPV8-9 1 0 0 0 0 0 0 0 0 1 1 0 1 1 1 9 0 9 9 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 0 0
OPV8-11 0 0 0 9 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 1 1 0 1 0 0 1 1 1 1 1 1 1 1 1 9 0 0
OPW17-1 0 0 0 1 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0
OPW17-3 0 0 9 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 1 1 1 0 0 0 0 0 9 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 1 1 0 0 0
OPW17-9 0 0 0 0 0 0 0 0 0 1 0 1 0 0 1 0 0 0 0 0 0 0 0 0 9 9 9 0 0 0 1 1 0 0 0 0 0 1 9 0 0 0 0 0 0 1 0 0 0 0 9 0 0 0 0 0 0 1 1 1
OPW17-11 0 0 0 0 0 1 0 1 1 0 1 1 1 0 9 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
OPW17-15 0 0 0 0 0 0 9 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 1 0 0 0 0 0 0 1 9 9 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0

Anexo 5.- Matriz de similitud de la comparación uno a uno entre las 60 OTUs. Coeficiente de
Asociación simple matching ―SM‖.
PJc1 PJc2 PJc3 PJc4 PJc5 PJc6 PJc7 PJc8
-------------------------------------------------------------------------
PJc1 | 1.0000
PJc2 | 0.8172 1.0000
PJc3 | 0.7957 0.8280 1.0000
PJc4 | 0.7312 0.6989 0.8065 1.0000
PJc5 | 0.8387 0.8065 0.8925 0.7849 1.0000
PJc6 | 0.7742 0.7527 0.7957 0.7097 0.8280 1.0000
PJc7 | 0.7312 0.7312 0.7957 0.6667 0.8495 0.7849 1.0000
PJc8 | 0.6452 0.6774 0.6774 0.5484 0.7312 0.7312 0.7527 1.0000
PJc9 | 0.6667 0.6344 0.6989 0.5914 0.7419 0.6989 0.7312 0.7527
PJc10 | 0.6989 0.6882 0.7312 0.6667 0.7097 0.6344 0.6882 0.6452
PJc11 | 0.7527 0.7419 0.7634 0.6882 0.8495 0.7312 0.7957 0.6989
PJc12 | 0.6667 0.6344 0.6774 0.6129 0.7204 0.6882 0.6882 0.7957
PJc13 | 0.7097 0.6882 0.7097 0.6129 0.7742 0.7419 0.7419 0.7527
PJc14 | 0.7527 0.7419 0.7527 0.6667 0.7957 0.7742 0.8065 0.6989
PJc15 | 0.8065 0.7527 0.7742 0.6882 0.8387 0.7527 0.7419 0.6559
PJc16 | 0.7634 0.7634 0.7957 0.6774 0.8280 0.7312 0.7849 0.7849
PJc17 | 0.6022 0.6237 0.6559 0.5806 0.6774 0.6667 0.6989 0.6774
PJc18 | 0.6237 0.6667 0.6882 0.6344 0.6667 0.6344 0.6344 0.6452
PJc19 | 0.6022 0.6452 0.6882 0.6667 0.6667 0.6129 0.6774 0.5806
PJc20 | 0.7312 0.7849 0.8280 0.6774 0.7849 0.7097 0.7527 0.6774
PJc21 | 0.7204 0.7527 0.8495 0.7097 0.7957 0.7204 0.7419 0.6882
PJc22 | 0.6667 0.7204 0.7742 0.6989 0.7204 0.6452 0.6882 0.6129
PJc23 | 0.6989 0.7312 0.8065 0.7097 0.7527 0.6774 0.7204 0.6452
PJc24 | 0.6667 0.7097 0.7419 0.6452 0.7419 0.6882 0.7204 0.6129
PJc25 | 0.6882 0.7419 0.7634 0.6344 0.7204 0.7097 0.7097 0.6129
PJc26 | 0.6129 0.6667 0.7312 0.6989 0.6882 0.6344 0.6452 0.5806
PJc27 | 0.7312 0.7634 0.7849 0.6559 0.7419 0.7097 0.7097 0.6774
PJc28 | 0.7097 0.7204 0.7634 0.6344 0.7419 0.6989 0.6774 0.6022
PJc29 | 0.6989 0.7312 0.7957 0.6667 0.7742 0.6882 0.7527 0.6989
PJc30 | 0.6882 0.7204 0.7849 0.6559 0.7204 0.6559 0.6989 0.6129
PJc31 | 0.6774 0.7312 0.7957 0.6882 0.7634 0.7204 0.7419 0.6882
PJc32 | 0.6559 0.7097 0.7742 0.6882 0.7419 0.7204 0.7204 0.6667
PJc33 | 0.7742 0.7849 0.8280 0.7204 0.8065 0.7312 0.7419 0.6559
PJc34 | 0.6774 0.7097 0.7312 0.6452 0.7097 0.6774 0.7204 0.6452
PJc35 | 0.7097 0.7419 0.8065 0.6559 0.7849 0.7097 0.7957 0.7204
PJc36 | 0.6989 0.7097 0.7742 0.6667 0.7527 0.6774 0.7419 0.6667
PJc37 | 0.6882 0.7204 0.7634 0.6559 0.7419 0.6882 0.7312 0.6559
PJc38 | 0.6882 0.7204 0.7527 0.6129 0.7204 0.6989 0.7097 0.6344
PJc39 | 0.6452 0.6344 0.6989 0.7204 0.6989 0.6237 0.6882 0.5914
PJc40 | 0.6774 0.6882 0.7312 0.6344 0.7742 0.6774 0.7204 0.7312
PJc41 | 0.7097 0.6989 0.7634 0.6344 0.7634 0.6667 0.7097 0.6774
PJc42 | 0.5699 0.5806 0.6452 0.5484 0.6237 0.5484 0.5806 0.5914
PJc43 | 0.6667 0.6774 0.6989 0.5591 0.6989 0.6452 0.6774 0.6344
PJc44 | 0.6344 0.6237 0.6667 0.5699 0.6452 0.6344 0.5914 0.5484
PJc45 | 0.6774 0.6774 0.7204 0.6129 0.7527 0.6559 0.7204 0.6129
PJc46 | 0.6882 0.6237 0.6237 0.5699 0.6559 0.5806 0.6452 0.5806
PJc47 | 0.6237 0.6452 0.6667 0.5161 0.6344 0.6022 0.6344 0.5914
PJc48 | 0.6774 0.6667 0.7312 0.6022 0.7097 0.6344 0.6989 0.6452
PJc49 | 0.7097 0.6774 0.7204 0.5806 0.7204 0.6129 0.6989 0.6452
PJc50 | 0.6559 0.6774 0.6882 0.5806 0.6559 0.6022 0.6667 0.5591
PJc51 | 0.6344 0.6667 0.7097 0.6989 0.7097 0.6129 0.6989 0.5914
PJc52 | 0.7527 0.7419 0.7849 0.6882 0.7527 0.6882 0.7204 0.6559
PJc53 | 0.6559 0.6667 0.7097 0.6344 0.7312 0.6774 0.6989 0.6667
PJc54 | 0.6452 0.6774 0.6989 0.6237 0.7419 0.6452 0.7097 0.6559
PJc55 | 0.6559 0.6237 0.6774 0.6344 0.6882 0.6129 0.6344 0.5591

PJc56 | 0.6237 0.6344 0.6882 0.6237 0.6989 0.6452 0.6452 0.5914
PJc57 | 0.6237 0.6022 0.6667 0.5591 0.6559 0.6022 0.6237 0.5699
PJc58 | 0.6774 0.7097 0.7312 0.6237 0.7097 0.6559 0.6774 0.6344
PJc59 | 0.6129 0.6559 0.6989 0.5806 0.7097 0.6129 0.6882 0.6022
PJc60 | 0.6559 0.7097 0.7527 0.6344 0.7527 0.6882 0.6882 0.6022
PJc9 PJc10 PJc11 PJc12 PJc13 PJc14 PJc15 PJc16
-------------------------------------------------------------------------
PJc9 | 1.0000
PJc10 | 0.7849 1.0000
PJc11 | 0.7419 0.7527 1.0000
PJc12 | 0.7849 0.7097 0.7419 1.0000
PJc13 | 0.7204 0.6559 0.7849 0.7742 1.0000
PJc14 | 0.7312 0.7204 0.7849 0.6989 0.8387 1.0000
PJc15 | 0.7312 0.7527 0.8065 0.6989 0.7634 0.7742 1.0000
PJc16 | 0.7419 0.6667 0.8172 0.7419 0.7957 0.7957 0.7849 1.0000
PJc17 | 0.7527 0.6344 0.6774 0.6989 0.6667 0.7097 0.6237 0.6989
PJc18 | 0.7097 0.6559 0.6774 0.6989 0.6882 0.7204 0.6452 0.7742
PJc19 | 0.6774 0.6774 0.6989 0.6667 0.6882 0.6989 0.6452 0.6989
PJc20 | 0.6882 0.7204 0.7742 0.6774 0.6882 0.7312 0.7097 0.7849
PJc21 | 0.6774 0.7312 0.7634 0.6882 0.6774 0.7204 0.7204 0.7742
PJc22 | 0.6237 0.6774 0.7312 0.6344 0.6452 0.6667 0.6452 0.7419
PJc23 | 0.6344 0.6882 0.7419 0.6667 0.6774 0.6989 0.6774 0.7742
PJc24 | 0.6774 0.6667 0.7312 0.6344 0.6882 0.7097 0.7527 0.7204
PJc25 | 0.6452 0.6882 0.7312 0.6022 0.6882 0.7312 0.6989 0.7204
PJc26 | 0.6129 0.6667 0.6774 0.6237 0.6344 0.6559 0.6452 0.7204
PJc27 | 0.6452 0.6882 0.7312 0.6452 0.7312 0.7419 0.7204 0.7634
PJc28 | 0.6129 0.6344 0.6989 0.6129 0.6774 0.7097 0.6882 0.7204
PJc29 | 0.6989 0.6882 0.7527 0.6452 0.7097 0.7527 0.6989 0.7957
PJc30 | 0.6882 0.6774 0.7419 0.5914 0.6774 0.7097 0.6667 0.7419
PJc31 | 0.7204 0.7204 0.7849 0.6774 0.6989 0.7419 0.7204 0.7957
PJc32 | 0.6344 0.6882 0.7527 0.6452 0.6989 0.6989 0.6989 0.7312
PJc33 | 0.7097 0.7634 0.7957 0.6774 0.7097 0.7419 0.7742 0.7849
PJc34 | 0.6882 0.6882 0.6989 0.6667 0.6989 0.7312 0.6989 0.7097
PJc35 | 0.7312 0.6989 0.7742 0.7204 0.7527 0.7312 0.7742 0.8280
PJc36 | 0.7312 0.7097 0.6989 0.7419 0.7312 0.7634 0.7204 0.7527
PJc37 | 0.6559 0.6452 0.6882 0.7097 0.6882 0.7419 0.6882 0.7419
PJc38 | 0.7204 0.7097 0.6989 0.6882 0.6667 0.6989 0.7312 0.7097
PJc39 | 0.6129 0.6667 0.6882 0.7097 0.6452 0.6989 0.6559 0.6559
PJc40 | 0.6882 0.6559 0.7419 0.7527 0.7419 0.6882 0.6989 0.7312
PJc41 | 0.6559 0.6774 0.7527 0.6989 0.7312 0.7419 0.7097 0.7527
PJc42 | 0.6559 0.6774 0.6344 0.6452 0.6129 0.6237 0.6344 0.6344
PJc43 | 0.6129 0.6344 0.6774 0.6129 0.6882 0.7204 0.6882 0.6559
PJc44 | 0.5914 0.6237 0.6559 0.5484 0.6022 0.6667 0.6559 0.6129
PJc45 | 0.6667 0.6667 0.7527 0.6237 0.6989 0.6989 0.7204 0.7097
PJc46 | 0.5699 0.6344 0.6559 0.5484 0.6344 0.6559 0.6882 0.6129
PJc47 | 0.6022 0.6452 0.6344 0.5699 0.6022 0.6774 0.6452 0.6129
PJc48 | 0.6129 0.6559 0.6882 0.6237 0.6774 0.7312 0.6559 0.6989
PJc49 | 0.6237 0.6452 0.6774 0.6129 0.6774 0.6882 0.6452 0.6989
PJc50 | 0.6129 0.6667 0.6344 0.5806 0.6022 0.6774 0.6022 0.5914
PJc51 | 0.5806 0.6452 0.7204 0.6129 0.6344 0.6989 0.6452 0.6774
PJc52 | 0.6452 0.7097 0.7419 0.6667 0.6667 0.7097 0.6882 0.7312
PJc53 | 0.6129 0.6559 0.7419 0.6452 0.6989 0.6774 0.6344 0.6774
PJc54 | 0.6452 0.6452 0.7527 0.6989 0.7742 0.6989 0.6667 0.7097
PJc55 | 0.5806 0.5914 0.6989 0.6237 0.6452 0.6452 0.5914 0.6559
PJc56 | 0.6237 0.6237 0.6882 0.6344 0.6882 0.6882 0.6022 0.6667
PJc57 | 0.5484 0.6129 0.6667 0.6022 0.6129 0.6452 0.6129 0.6452
PJc58 | 0.6129 0.6129 0.6667 0.6559 0.6452 0.6667 0.6882 0.6989
PJc59 | 0.6129 0.6344 0.6989 0.6452 0.6237 0.6237 0.6559 0.6559
PJc60 | 0.6344 0.6774 0.6989 0.6774 0.6774 0.6882 0.6989 0.6882

PJc17 PJc18 PJc19 PJc20 PJc21 PJc22 PJc23 PJc24
-------------------------------------------------------------------------
PJc17 | 1.0000
PJc18 | 0.7527 1.0000
PJc19 | 0.6667 0.7634 1.0000
PJc20 | 0.6774 0.6989 0.7204 1.0000
PJc21 | 0.6237 0.7097 0.7097 0.9032 1.0000
PJc22 | 0.6129 0.6774 0.7849 0.8710 0.8817 1.0000
PJc23 | 0.6237 0.6882 0.7419 0.8602 0.8710 0.9355 1.0000
PJc24 | 0.5806 0.6344 0.6667 0.7957 0.8280 0.8065 0.8280 1.0000
PJc25 | 0.6129 0.6559 0.6774 0.8710 0.8387 0.8065 0.7957 0.7957
PJc26 | 0.5914 0.7312 0.7204 0.7957 0.8280 0.7849 0.7742 0.7204
PJc27 | 0.5914 0.6452 0.6667 0.8495 0.8387 0.8065 0.8387 0.7957
PJc28 | 0.5914 0.6559 0.6344 0.8280 0.8172 0.7634 0.7742 0.7742
PJc29 | 0.6882 0.6882 0.6989 0.8172 0.8280 0.8172 0.8280 0.7634
PJc30 | 0.6559 0.6559 0.6882 0.8065 0.8280 0.7957 0.7849 0.7419
PJc31 | 0.7204 0.7312 0.6774 0.8387 0.8280 0.7742 0.7849 0.7419
PJc32 | 0.6667 0.6882 0.6774 0.8172 0.8280 0.7957 0.8065 0.7419
PJc33 | 0.6344 0.6882 0.7097 0.8280 0.8602 0.7849 0.7957 0.8172
PJc34 | 0.6129 0.6882 0.6667 0.7957 0.7849 0.7527 0.7634 0.7849
PJc35 | 0.6559 0.7419 0.7312 0.8280 0.8387 0.8065 0.8387 0.8065
PJc36 | 0.6774 0.7419 0.6989 0.8387 0.8280 0.7527 0.7634 0.7527
PJc37 | 0.6022 0.6774 0.6774 0.8065 0.7957 0.7634 0.7742 0.7634
PJc38 | 0.6344 0.6989 0.6667 0.7634 0.7527 0.7204 0.7527 0.7742
PJc39 | 0.5699 0.6129 0.7527 0.7204 0.7312 0.7419 0.7204 0.6989
PJc40 | 0.6022 0.6344 0.6452 0.7527 0.7419 0.6882 0.6882 0.7204
PJc41 | 0.6237 0.6989 0.7204 0.7742 0.7849 0.7527 0.7419 0.7204
PJc42 | 0.5806 0.6559 0.6237 0.6774 0.6989 0.6237 0.6237 0.6452
PJc43 | 0.5699 0.5914 0.6559 0.6989 0.6882 0.6559 0.6559 0.6774
PJc44 | 0.5376 0.6237 0.5914 0.7312 0.6989 0.6452 0.6237 0.6774
PJc45 | 0.5699 0.6022 0.6344 0.7204 0.6882 0.6559 0.6774 0.7312
PJc46 | 0.4946 0.5054 0.5376 0.6022 0.5914 0.5376 0.5376 0.6129
PJc47 | 0.4731 0.5484 0.6237 0.6559 0.6882 0.6344 0.6022 0.6452
PJc48 | 0.5484 0.6129 0.6344 0.7312 0.7419 0.6882 0.6882 0.6667
PJc49 | 0.6022 0.6022 0.6129 0.6559 0.6667 0.6344 0.6559 0.6344
PJc50 | 0.5376 0.5699 0.6452 0.6774 0.6882 0.6559 0.6559 0.6344
PJc51 | 0.5806 0.6129 0.7097 0.6882 0.6774 0.7097 0.7097 0.6667
PJc52 | 0.5806 0.6237 0.6559 0.7634 0.7312 0.6989 0.7527 0.6882
PJc53 | 0.5376 0.5806 0.6237 0.7097 0.7204 0.6452 0.6559 0.6559
PJc54 | 0.5699 0.6237 0.6989 0.6989 0.6882 0.6774 0.6774 0.6774
PJc55 | 0.5806 0.6237 0.6882 0.6882 0.6989 0.7097 0.7097 0.6452
PJc56 | 0.6344 0.7097 0.6989 0.6989 0.7097 0.6774 0.6882 0.6452
PJc57 | 0.5484 0.6129 0.6452 0.6882 0.7312 0.6559 0.6882 0.6237
PJc58 | 0.5699 0.6022 0.6022 0.7419 0.7097 0.6667 0.7312 0.7097
PJc59 | 0.5484 0.5269 0.5806 0.6989 0.6344 0.6237 0.6774 0.6452
PJc60 | 0.5591 0.6022 0.6237 0.7204 0.6989 0.6452 0.6774 0.6667
PJc25 PJc26 PJc27 PJc28 PJc29 PJc30 PJc31 PJc32
-------------------------------------------------------------------------
PJc25 | 1.0000
PJc26 | 0.7742 1.0000
PJc27 | 0.8710 0.7742 1.0000
PJc28 | 0.8065 0.7527 0.8710 1.0000
PJc29 | 0.7742 0.7204 0.8602 0.8602 1.0000
PJc30 | 0.7957 0.7312 0.8280 0.8065 0.9032 1.0000
PJc31 | 0.8280 0.7957 0.8065 0.7957 0.8280 0.8280 1.0000
PJc32 | 0.8065 0.7634 0.8495 0.7849 0.8387 0.8172 0.9032 1.0000
PJc33 | 0.7849 0.7957 0.8495 0.8065 0.8387 0.8280 0.8172 0.8172
PJc34 | 0.7742 0.7527 0.7957 0.7957 0.7634 0.7527 0.7849 0.7634

PJc35 | 0.7849 0.7527 0.8065 0.7634 0.8387 0.7849 0.8172 0.8172
PJc36 | 0.7957 0.7527 0.7634 0.7849 0.7419 0.7312 0.7634 0.7419
PJc37 | 0.7849 0.7312 0.7634 0.7849 0.7312 0.6989 0.7527 0.7312
PJc38 | 0.7419 0.6989 0.7527 0.7419 0.7312 0.7527 0.7957 0.7527
PJc39 | 0.6989 0.6882 0.6989 0.6882 0.6774 0.6452 0.6774 0.6774
PJc40 | 0.6882 0.6774 0.7312 0.6989 0.6882 0.6774 0.7204 0.7312
PJc41 | 0.7097 0.6989 0.7204 0.7419 0.7634 0.7097 0.7312 0.7097
PJc42 | 0.6237 0.6667 0.6667 0.6989 0.6667 0.6559 0.6774 0.6344
PJc43 | 0.6344 0.6237 0.6774 0.6882 0.6989 0.6667 0.6452 0.6237
PJc44 | 0.6667 0.6559 0.6882 0.7527 0.6882 0.6559 0.6774 0.6559
PJc45 | 0.6559 0.6022 0.7097 0.6882 0.6882 0.6559 0.6667 0.6452
PJc46 | 0.6129 0.5376 0.6774 0.6237 0.6237 0.5914 0.5914 0.6129
PJc47 | 0.6452 0.5914 0.6774 0.7097 0.6989 0.6667 0.6237 0.6237
PJc48 | 0.6882 0.6559 0.7097 0.7634 0.7527 0.7204 0.6882 0.6774
PJc49 | 0.6237 0.6022 0.6559 0.6667 0.7204 0.6882 0.6237 0.6022
PJc50 | 0.6344 0.5806 0.6344 0.6452 0.6559 0.6344 0.6237 0.6129
PJc51 | 0.6667 0.6559 0.6882 0.6344 0.7097 0.6559 0.6667 0.6667
PJc52 | 0.6989 0.6667 0.7419 0.7097 0.7419 0.7097 0.7097 0.6882
PJc53 | 0.6882 0.6344 0.7097 0.6559 0.6882 0.6559 0.6559 0.6774
PJc54 | 0.6559 0.6237 0.6989 0.6667 0.6989 0.6774 0.6667 0.6882
PJc55 | 0.6344 0.6344 0.6667 0.6559 0.6667 0.6882 0.6559 0.6344
PJc56 | 0.6667 0.6882 0.6667 0.6989 0.6882 0.7097 0.6882 0.6452
PJc57 | 0.6129 0.6344 0.6237 0.6344 0.6559 0.6344 0.6344 0.6129
PJc58 | 0.6667 0.6237 0.7097 0.6774 0.7097 0.6774 0.7097 0.6882
PJc59 | 0.6344 0.5591 0.6344 0.6237 0.6344 0.6022 0.6559 0.6237
PJc60 | 0.6559 0.6237 0.6774 0.6774 0.6882 0.6559 0.7204 0.6989
PJc33 PJc34 PJc35 PJc36 PJc37 PJc38 PJc39 PJc40
-------------------------------------------------------------------------
PJc33 | 1.0000
PJc34 | 0.7849 1.0000
PJc35 | 0.8172 0.7957 1.0000
PJc36 | 0.7634 0.8172 0.8172 1.0000
PJc37 | 0.7634 0.8387 0.7634 0.8925 1.0000
PJc38 | 0.7634 0.7742 0.7634 0.7634 0.7957 1.0000
PJc39 | 0.7419 0.6667 0.6774 0.7204 0.7634 0.6774 1.0000
PJc40 | 0.7742 0.7204 0.7312 0.7634 0.7849 0.7097 0.7957 1.0000
PJc41 | 0.8065 0.7527 0.7419 0.7527 0.7742 0.6989 0.7957 0.8387
PJc42 | 0.7312 0.6344 0.6667 0.6559 0.6344 0.6774 0.7312 0.7097
PJc43 | 0.7419 0.6667 0.6774 0.6559 0.6774 0.6559 0.7419 0.7634
PJc44 | 0.7742 0.6882 0.6452 0.6344 0.6559 0.6559 0.6882 0.7097
PJc45 | 0.7419 0.6559 0.6989 0.6344 0.6452 0.6774 0.6989 0.7097
PJc46 | 0.6882 0.5484 0.6022 0.5591 0.5699 0.5914 0.6559 0.6882
PJc47 | 0.6989 0.6667 0.6559 0.6559 0.6882 0.6667 0.6452 0.6237
PJc48 | 0.7527 0.7097 0.6882 0.7312 0.7527 0.6559 0.7957 0.7419
PJc49 | 0.6989 0.6129 0.6989 0.6452 0.6559 0.6667 0.6989 0.6882
PJc50 | 0.6559 0.6667 0.6774 0.7204 0.7419 0.6989 0.7097 0.6882
PJc51 | 0.6882 0.6237 0.6882 0.6559 0.6882 0.6452 0.8065 0.7097
PJc52 | 0.7634 0.7204 0.7419 0.7097 0.7204 0.7204 0.6989 0.7419
PJc53 | 0.6882 0.6129 0.6667 0.6129 0.6452 0.6237 0.7312 0.7634
PJc54 | 0.7204 0.6882 0.7204 0.6774 0.6989 0.6559 0.7634 0.8065
PJc55 | 0.6882 0.6344 0.6667 0.5806 0.6237 0.6452 0.7312 0.6989
PJc56 | 0.6989 0.6667 0.6559 0.6344 0.6344 0.6989 0.7204 0.6882
PJc57 | 0.7097 0.5914 0.6344 0.6237 0.6237 0.6344 0.7097 0.6559
PJc58 | 0.7419 0.6989 0.7419 0.6882 0.6989 0.7204 0.7204 0.7097
PJc59 | 0.6344 0.6452 0.6667 0.6344 0.6344 0.6667 0.6559 0.6559
PJc60 | 0.7419 0.6774 0.6882 0.6667 0.6882 0.7097 0.7204 0.7204

PJc41 PJc42 PJc43 PJc44 PJc45 PJc46 PJc47 PJc48
-------------------------------------------------------------------------
PJc41 | 1.0000
PJc42 | 0.7742 1.0000
PJc43 | 0.8172 0.7742 1.0000
PJc44 | 0.8280 0.7742 0.8280 1.0000
PJc45 | 0.7527 0.7204 0.7527 0.7527 1.0000
PJc46 | 0.7204 0.6452 0.7312 0.7419 0.7957 1.0000
PJc47 | 0.7312 0.6344 0.7204 0.7312 0.6989 0.7204 1.0000
PJc48 | 0.8495 0.7312 0.7957 0.7634 0.7097 0.7419 0.8065 1.0000
PJc49 | 0.7849 0.7097 0.7527 0.6882 0.7097 0.7527 0.6882 0.7957
PJc50 | 0.7419 0.6667 0.7312 0.6667 0.6452 0.6882 0.7419 0.7527
PJc51 | 0.7742 0.6667 0.7312 0.7097 0.7204 0.6989 0.6344 0.7419
PJc52 | 0.7634 0.6452 0.7312 0.6774 0.7097 0.6774 0.6774 0.7742
PJc53 | 0.7312 0.6989 0.7097 0.6667 0.7204 0.7204 0.6667 0.7419
PJc54 | 0.7849 0.7097 0.7634 0.6989 0.7527 0.7097 0.6774 0.7957
PJc55 | 0.7312 0.7204 0.6667 0.6774 0.7097 0.6667 0.6022 0.7204
PJc56 | 0.7634 0.7419 0.6882 0.7097 0.6989 0.6129 0.6237 0.7419
PJc57 | 0.7527 0.6989 0.7097 0.6989 0.6667 0.5914 0.6452 0.7204
PJc58 | 0.7634 0.6559 0.7419 0.6774 0.6774 0.6452 0.6129 0.7634
PJc59 | 0.6452 0.5376 0.6559 0.5806 0.6667 0.5699 0.5591 0.6774
PJc60 | 0.7312 0.6237 0.6989 0.6559 0.6989 0.6129 0.6559 0.7419
PJc49 PJc50 PJc51 PJc52 PJc53 PJc54 PJc55 PJc56
-------------------------------------------------------------------------
PJc49 | 1.0000
PJc50 | 0.7957 1.0000
PJc51 | 0.8172 0.7634 1.0000
PJc52 | 0.7849 0.7419 0.7957 1.0000
PJc53 | 0.7742 0.6882 0.7634 0.8065 1.0000
PJc54 | 0.7957 0.7527 0.7742 0.7742 0.8387 1.0000
PJc55 | 0.7527 0.7097 0.7204 0.7419 0.7849 0.8495 1.0000
PJc56 | 0.7312 0.6452 0.6774 0.7097 0.7849 0.8065 0.8710 1.0000
PJc57 | 0.7204 0.6344 0.6667 0.7204 0.7742 0.7312 0.7419 0.7742
PJc58 | 0.7204 0.6559 0.6882 0.7742 0.6559 0.6989 0.6452 0.6452
PJc59 | 0.6022 0.5806 0.6022 0.6989 0.6559 0.6882 0.5914 0.6022
PJc60 | 0.6344 0.6344 0.6452 0.7527 0.6989 0.7312 0.6559 0.6882
PJc57 PJc58 PJc59 PJc60
-------------------------------------
PJc57 | 1.0000
PJc58 | 0.6667 1.0000*
PJc59 | 0.6559 0.7527 1.0000
PJc60 | 0.7419 0.7634 0.7849 1.0000

\\
Coefficient
Anexo N° 6.
6.1.- Fenograma UPGMA generado con los 60 individuos analizados, utilizando el coeficiente
de similitud Simple Matching.

Coefficient
6.2.- Fenograma UPGMA generado con los 60 individuos analizados, utilizando el coeficiente de
similitud Simple Matching.

Anexo N° 7.
7.1 Comparación de las dos matrices: cofenética y de distancia de los 60 genotipos
utilizando los marcadores RAPD. Se muestran los coeficientes de correlación cofenética para
cada gráfico.
(r) = 0.78405
X Label
0.39 0.53 0.66 0.80 0.94
Y Label
0.58
0.67
0.76
0.85
0.94
Matriz de distancias
Matriz
Cofenetica

7.2 Representación gráfica de la correlación entre la Matriz de Similitud y la Matriz
cofenética para los marcadores ISSR.
RAPD 60 SM
(r) = 0.69835
ISSR 60 Tesis NTSYS edit SM.NTS
0.36 0.51 0.66 0.81 0.95
ISSR 60 Tesis NTSYS edit SM UPGMA coph.NTS
0.63
0.71
0.79
0.87
0.95
RAPD 60 SM
UPGMA coph

Anexo N°8.- Extracción de ADN por el metodo cetil trimetil amonio bromuro (CTAB)
1. Obtener 5g de tejido, triturarlo con nitrógeno liquido
2. Transferir el polvo a tubos de 50ml /conteniendo 20ml de buffer de extracción y 200ul
de 2- mercaptocthanol,min 98%.
3. Incubar a 65ºC por 20 min. , mezclar cada 5 minutos.
4. Enfriar a temperatura ambiente por 10 minutos añadir un volumen igual al contenido
en el tubo falcon de cloroformo alcoholisoamilico (24:1), mezclar suavemente por 5
min.
5. Centrifugar a 4000rpm por 20 min. a 4ºC.
6. Transferir la fase superior acuosa (sobrenadante) a un nuevo tubo de 50 ml.
7. Añadir un volumen igual al sobrenadante de cloroformo alcoholaisoamilico (24:1)
mezclar suavemente por 5 min.
8. Centrifugar a 4000rpm por 20 min. a 4ºC.
9. Transferir el sobrenadante a un nuevo tubo de 50 ml conteniendo 2/3 del volumen del
sobrenadante , de isopropanol (15ml) mezclar suavemente.
Opcion A:
Dejar el tubo falcon a temperatura ambiente.
Colectar el ADN por pesca con una pipeta pasteur
Transferir el ADN a un nuevo tubo empendolf conteniendo 300 ul de Wash 1
buffer dejarlo a temperatura ambiente por 30 min.
Transferir el ADN a un nuevo tubo empendolf conteniendo 300 ul de Wash 2
por 5 segundos.
Transferir el ADN a un nuevo tubo empendolf y secar a temperatura ambiente
por 30 min.
Disolver el ADN en 500-1000 ul de T10 E1 (x tamaño del pellet)
Añadir RNAsa (10ng/ul) incubar a 37ºC por una hora.
Opcion B:
Dejar a 20ºC al menos una hora centrifugar a 4000 rpm por 30 min.
Dejar escurrir el isopropanol y secar a temperatura ambiente los tubos
destapados e invertidos.
Disolver el pellet de ADN en 500-1000ul de T10 E1 y 5-10 ul de RNAsa libre
de DNAsa (10ng/ml) incubar a 37ºC por una hora.
Almacenar a 4ºC.
Extracción de ADN Por el método CTAB (Para hojas secas)
1. En un tubo de 2ml agregar 1ml de buffer de extracción CTAB 2 X y 2ul de b-
mercaptoetanol.
2. Moler con nitrógeno líquido aproximadamente 100 mg de tejido vegetal.
3. Vaciar el polvo al tubo con el Buffer de extracción.
4. Incubar a 65ºC por 45 minutos. Agitar tres veces durante los primeros 5 minutos,
luego agitarlos dos veces cada 20 minutos.
5. Dejar enfriar a temperatura ambiente.
6. Agregar 1 ml de cloroformo:isoalmilico (24:1)
7. Agitar ligeramente hasta homogenizar el contenido.
8. Centrifugar a 14,000 rpm por 5 minutos.
9. transferir el sobrenadante a tubos de 1.5ml (usar tips de punta cortada).

10. Agregar 75 ul de CTAB 10X (65ºC) y agitar hasta homogenizar.
11. Agregar 650 ul de cloroformo:isoamilico (24:1) y agitar hasta homogenizar
completamente.
12. Centrifugar a 14,000 rpm x 5 minutos.
13. Transferir el sobrenadante (a tubos de 1.5 ml) esteriles.
14. Agregar 500 ul de isopropanol frio y agitar suavemente por espacion de 10 segundos –
usar repetitivo.
15. Incubar a -20ºC x 30 minutos ( puede detenerse el procedimiento hasta el dia
siguiente)
16. Centrifugar a 14,000 rpm por 20 minutos.
17. Decantar el isopropanol con cuidado de no perder el pellet.
18. Dejar los tubos invertidos por 1 minuto sobre papel toalla.
19. Agregar 300 ul de Wash 1 por 10 minutos.
20. Decantar con cuidado de no perder el pellet.
21. Dejar los tubos invertidos sobre papel toalla por 30 segundos.
22. Enjuagar con 300 ul deWash 2.
23. Decantar con cuidado.
24. Dejar secar los tubos por 30 minutos.
25. ( Opcional: secar en liofilizador por 10 minutos, opción médium)
26. Resuspender cada pellet en 75 ul de T10E1
(Opcional: Dejar overnight a 4ºC para una dilución total del pellet)
27. Agregar 2 ul de RNAsa A
28. Dejar por 30 minutos a 37ºC
CORRIDA ELECTROFORETICA
29. Tomar 9 ul de loading buffer
30. adicionar 1 ul de la muestra de ADN
31. Mezclar
32. Cargar en un gel de agarosa 1% y correr a 80-90 Volt x 30 minutos
33. (Nota.- Se usó buffer TBE)


















