
Tema 13. Microbiología -...
50
Germán Tenorio Biología NS-Diploma BI Tema 13. Microbiología 13.1 Generalidades: Virus y bacterias PAU Idea Fundamental: Los microorganismos son un grupo heterogéneos de seres vivos que se caracterizan por su tamaño microscópico.
Transcript of Tema 13. Microbiología -...

Germán Tenorio
Biología NS-Diploma BI
Tema 13. Microbiología13.1 Generalidades: Virus y bacteriasPAU
Idea Fundamental: Los microorganismosson un grupo heterogéneos de seres vivosque se caracterizan por su tamañomicroscópico.

Diversidad de microbiosDP/PAU
Los microbios son sorpendentes,entre otras cosas por:
- Constituyen el 50% de la biomasaexistente en el planeta Tierra.
- Reciclan toneladas de materiaorgánica.
- Utilizan distintas formas denutrición.
- Junto con sus enzimas estánimplicados en la elaboración de unagran cantidad de productoscotidianos.
- Son responsables de la mayoríade las enfermedades infecciosas.

Concepto de microorganismoDP/PAU
La Microbiología estudia unenorme y diverso grupo demicroorganismos formado porvirus y otras partículasinfectivas subvirales,bacterias, arqueobacterias,protistas (algas y protozoos) yhongos.
Los microorganismos omicrobios son un variadogrupo de seres vivos quetienen como característicacomún su pequeño tamaño,no pudiendo ser percibidos porel ojo humano sin la ayuda deun microscopio.
Son diversos en apariencia,metabolismo, fisiología ygenética.
1 mm = 103 µm = 106 nm = 107 Å

Características de los microorganismosPAU
Su pequeño tamaño determinan varias de propiedades biológicasimportantes:
- El pequeño volumen de su citoplasma mantiene una gran superficiede contacto con el entorno, facilitando el intercambio de sustanciascon el medio. A medida que disminuye el tamaño, aumenta larelación superficie/volumen (hombre 0.3 cm2/g versus ameba 400cm2/g).
- Su pequeño tamaño hace que todos los puntos estén próximos, porlo que las pérdidas por dilución se reducen al máximo y lasreacciones metabólicas ocurren a gran velocidad.
- Debido a su rápido metabolismo, originan productos de desecho alexterior. Por tanto, alteran en poco tiempo el medio en el queviven.
- Debido a su elevada tasa metabólica, se multiplican con granrapidez.

Característica Virus Bacterias Algas Protozoos Hongos
Presencia de estructura
celular
Noacelulares
Si Si Si Si
Tipo de estructura
celular
- Procariota Eucariota Eucariota Eucariota
Número de células
- unicelular Unicelular/pluricelular
unicelular Unicelular/pluricelular
Tipo de nutrición
- Autótrofa/heterótrofa
Autótrofa Heterótrofa Heterótrofa
Tipo de División celular
- Bipartición Mitosis Mitosis Mitosis
Tamaño 0.01-0.3 µm
1 µm 1µm-metros
10-100 µm 1µm-metros
Criterios para clasificar los microorganismosPAU
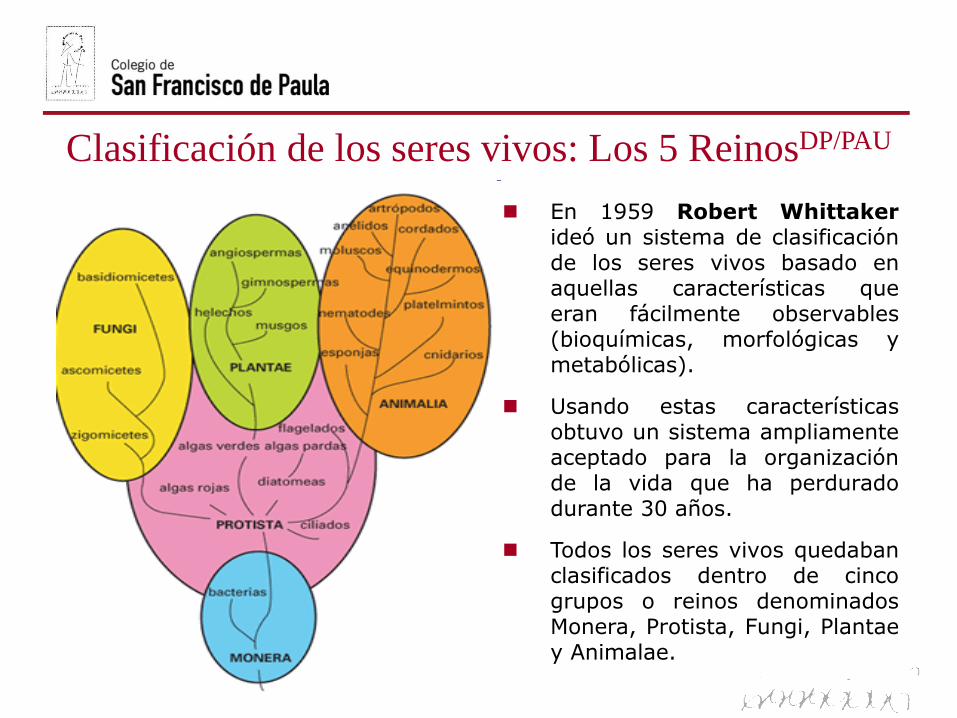
Clasificación de los seres vivos: Los 5 ReinosDP/PAU
En 1959 Robert Whittakerideó un sistema de clasificaciónde los seres vivos basado enaquellas características queeran fácilmente observables(bioquímicas, morfológicas ymetabólicas).
Usando estas característicasobtuvo un sistema ampliamenteaceptado para la organizaciónde la vida que ha perduradodurante 30 años.
Todos los seres vivos quedabanclasificados dentro de cincogrupos o reinos denominadosMonera, Protista, Fungi, Plantaey Animalae.

Los tres dominiosDP/PAU
Otro científico, Carl Woese,intentó 30 años más tardemejorar la precisión del sistemade clasificación a partir de losnuevos conocimientos obtenidosde la biología molecular.
Observó que dentro de los propiosprocariotas existían grandesdiferencias en su ARNr 16S, yque estas diferencias eran tangrandes como las existentes entreprocariotas y eucariotas.
Woese dividió entonces losprocariotas en dos grupos,Eubacteria y Archaea, pasandoa tener el nuevo modelo declasificación de los seres vivos 3dominios: Eukarya, Eubacteria yArchaea.
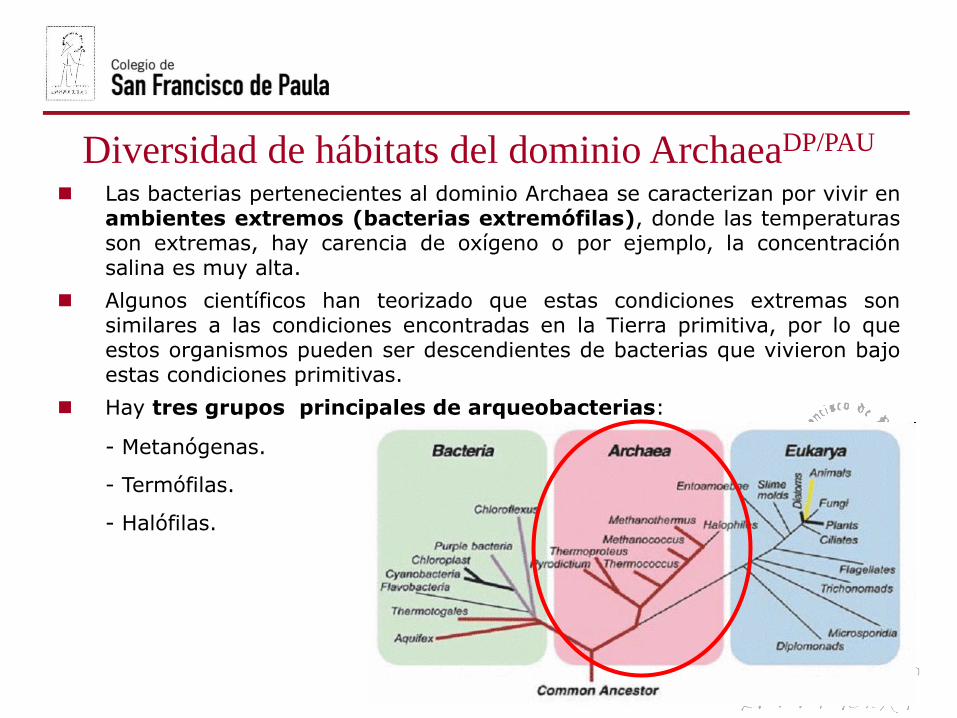
Diversidad de hábitats del dominio ArchaeaDP/PAU
Las bacterias pertenecientes al dominio Archaea se caracterizan por vivir enambientes extremos (bacterias extremófilas), donde las temperaturasson extremas, hay carencia de oxígeno o por ejemplo, la concentraciónsalina es muy alta.
Algunos científicos han teorizado que estas condiciones extremas sonsimilares a las condiciones encontradas en la Tierra primitiva, por lo queestos organismos pueden ser descendientes de bacterias que vivieron bajoestas condiciones primitivas.
Hay tres grupos principales de arqueobacterias:
- Metanógenas.
- Termófilas.
- Halófilas.
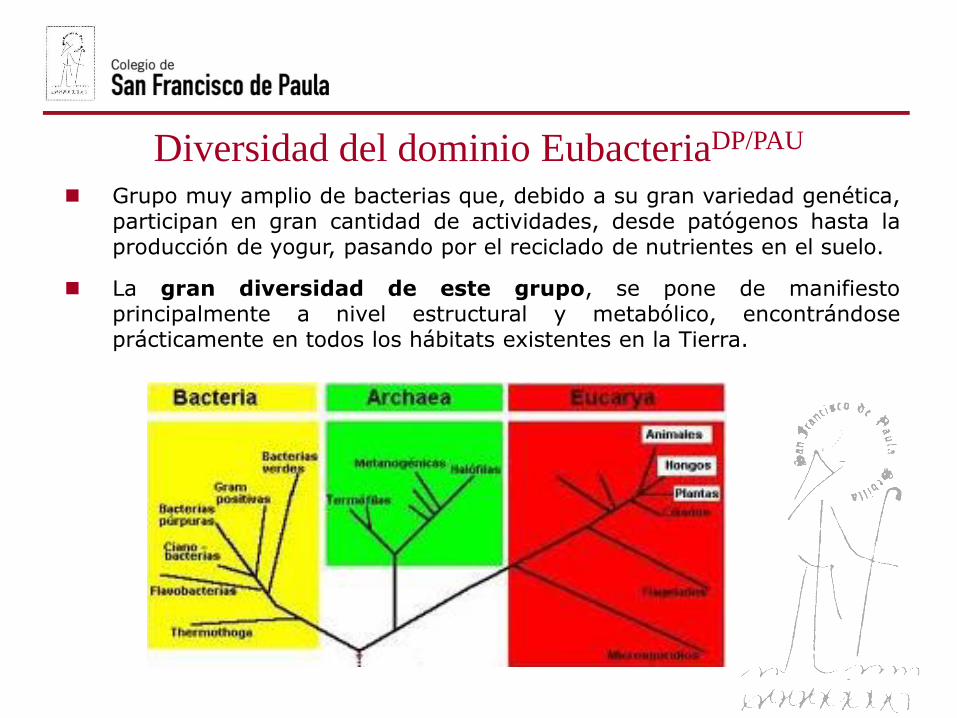
Diversidad del dominio EubacteriaDP/PAU
Grupo muy amplio de bacterias que, debido a su gran variedad genética,participan en gran cantidad de actividades, desde patógenos hasta laproducción de yogur, pasando por el reciclado de nutrientes en el suelo.
La gran diversidad de este grupo, se pone de manifiestoprincipalmente a nivel estructural y metabólico, encontrándoseprácticamente en todos los hábitats existentes en la Tierra.

Diversidad del dominio EubacteriaDP/PAU
A nivel estructural, las eubacterias presentan cinco tipos de morfologíasprincipales, esféricos aislados o en grupo (cocos), alargados y cilíndricos(bacilos), bacilo ovalado (cocobacilos), helicoidales (espirilos), y cortos ycurvados con forma de coma (vibrios).
Web1
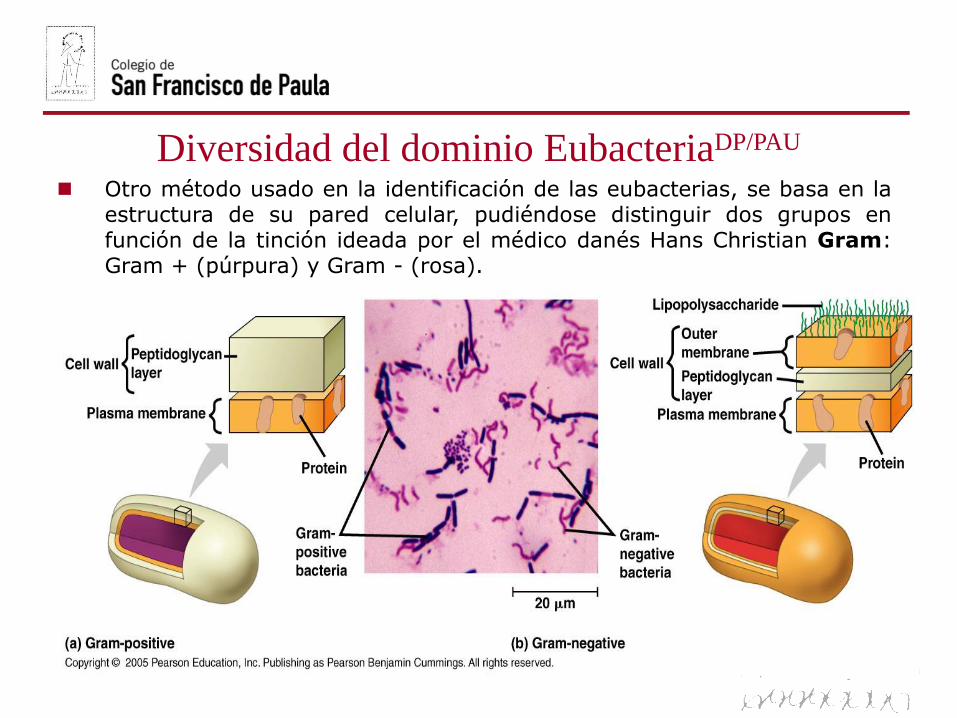
Diversidad del dominio EubacteriaDP/PAU
Otro método usado en la identificación de las eubacterias, se basa en laestructura de su pared celular, pudiéndose distinguir dos grupos enfunción de la tinción ideada por el médico danés Hans Christian Gram:Gram + (púrpura) y Gram - (rosa).

Características estructurales del dominio bacteriaDP/PAU
Las bacterias verdaderas son microorganismos unicelularesprocariotas, cuyo tamaño varía entre 1-10 micrómetros, adaptadasa vivir en cualquier ambiente y que poseen todos los tipos denutrición: autótrofa (fotosintética y quimiosintética) y heterótrofas(saprófitas, simbióticas y parásitas).
Esta diversidad las hace indispensables para el mantenimiento delequilibrio ecológico, al intervenir en los ciclos biogeoquímicos quereciclan la materia en la biosfera.
La ultraestructura de las bacterias sólo es visible al microscopioelectrónico, distinguiéndose hacia el interior: Cápsula o glucocálix,pared cellular, membrana plasmática….
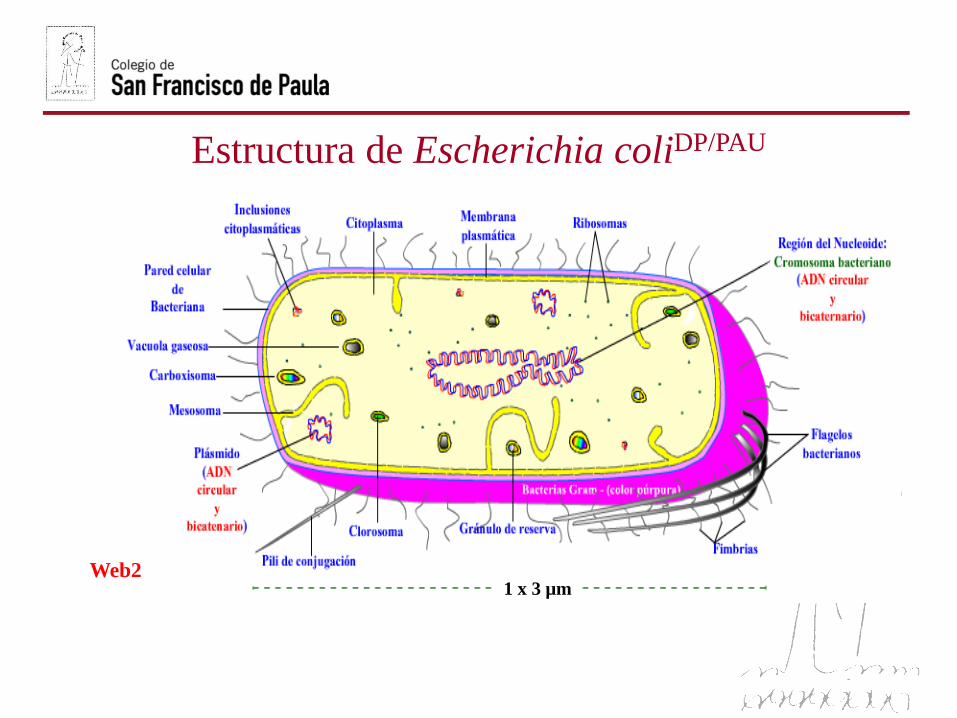
Estructura de Escherichia coliDP/PAU
1 x 3 µmWeb2

Función de reproducción del dominio bacteriaDP/PAU
Se reproducen asexualmente porbipartición o fisión binaria, queocurre tras la replicación del ADN.
La membrana plasmática seinvagina y la pared bacteriana crecehasta formar un tabique transversalque divide a las dos bacterias.
En condiciones normales puedendividirse cada 20 minutos, lo queles permite adaptárse rápidamentea cambios en el ambiente.

Además, las bacterias presentan mecanismos parasexuales deintercambio de ADN que no implican aumento del número de individuos:
- Transformación: Se transfiere un fragmento de ADN libre procedentede una bacteria donadora hasta la receptora. No es necesario elcontacto entre ambas.
- Conjugación: Se transfieren plásmidos conjugativos a través de pilisexuales. Se requiere contacto.
- Transducción: Se transfieren fragmentos de ADN desde la bacteriadonadora a la receptora a través de un virus lisogénico.
Función de reproducción del dominio bacteriaDP/PAU
Web1-2

Función de reproducción del dominio bacteriaDP/PAU

Función de nutrición del dominio bacteriaDP/PAU
La diversidad del dominio Eubacteria también se pone de manifiestoobservando la amplia diversidad nutricional que presentan.
Independientemente del tipo de nutrición, las bacterias pueden necesitarO2 (bacterias aerobias) o no (anaerobias). Para algunas bacterias el O2 esletal (anaerobias estrictas), mientras que otras lo utilizan cuando estápresente, aunque pueden vivir sin él (anaerobias facultativas).

Tipos de nutrición bacterianaDP/PAU
Las bacterias, grupo muy heterogéneo, pueden colonizar todos losambientes, ya que presentan todos los tipos de metabolismoconocido, y algunas son capaces de cambiar de metabolismo en funcióndel tipo de nutrientes que encuentran en el medio.
La bacterias se pueden clasificar en función de:
- Si la fuente de energía es la de la luz (fotótrofas) o la de compuestosquímicos (quimiótrofas).
- Si la fuente de carbono son moléculas inorgánicas como el CO2
(autótrofas) o moléculas orgánicas sencillas (heterótrofas).
- Si la fuente de protones y electrones son moléculas inorgánicassimples (litótrofos) o moléculas orgánicas complejas (organótrofos).

Tipos de nutrición bacterianaDP/PAU
Bacterias púrpuras no sulfurosas
Cianobacterias (fotosintéticas)
Bacterias verdes y púrpuras sulfurosas
Bacterias quimiosintéticas
Bacterias saprófitas
(descomponedores)
FOTOAUTÓTROFOS
O
QUIMIOAUTÓTROFOS
O
FOTOHETERÓTROFOS
O
QUIMIOHETERÓTROFOS
O
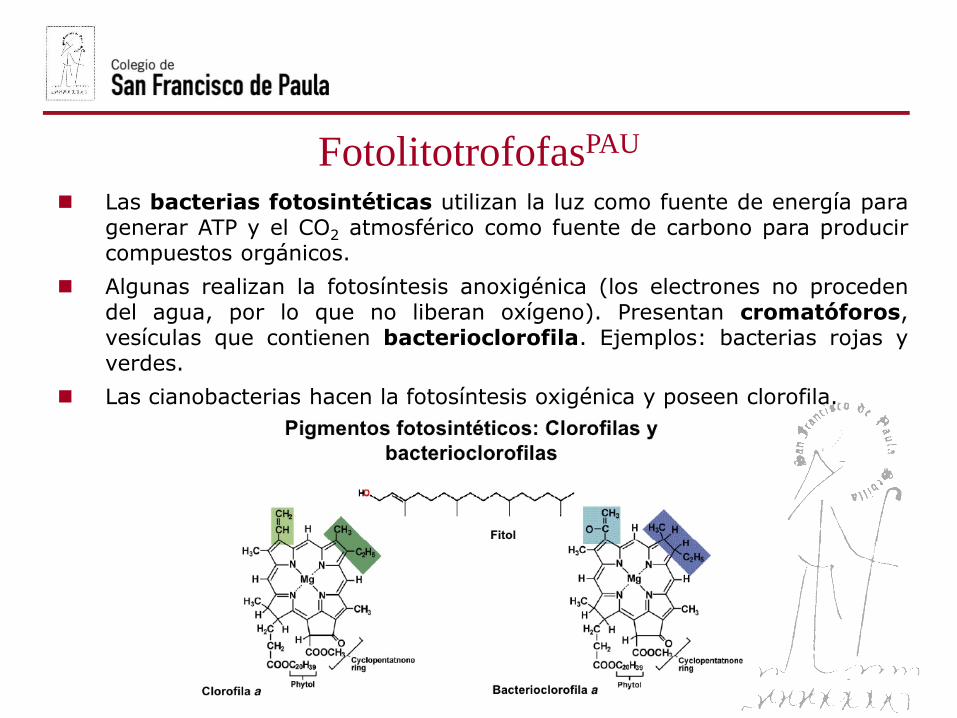
FotolitotrofofasPAU
Las bacterias fotosintéticas utilizan la luz como fuente de energía paragenerar ATP y el CO2 atmosférico como fuente de carbono para producircompuestos orgánicos.
Algunas realizan la fotosíntesis anoxigénica (los electrones no procedendel agua, por lo que no liberan oxígeno). Presentan cromatóforos,vesículas que contienen bacterioclorofila. Ejemplos: bacterias rojas yverdes.
Las cianobacterias hacen la fotosíntesis oxigénica y poseen clorofila.

QuimiolitotrofasPAU
Las bacterias quimiosintéticas usan energía de reacciones químicascomo fuente de energía para generar ATP y el CO2 atmosférico comofuente de carbono para producir compuestos orgánicos.
Obtienen energía oxidandosustratos inorgánicos (H2,NH3, H2 S, S, etc.), con laque obtienen ATP yNADPH (o NADH), queluego emplean para laasimilación reductora delcarbono, mediante el ciclode Calvin, como lasplantas y del nitrógeno.

QuimiolitotrofasPAU
Son importantísimas en los ciclos biogeoquímicos, pues mineralizan lamateria orgánica.
Participan en laasimilación reductoradel nitrógeno. La mayoríade las bacterias debenreducir los nitratos anitritos y éstos aamoniaco, con el fin defabricar grupos amino.Algunas bacterias, sinembargo, pueden fijar(reducir) directamente elnitrógeno atmosférico yreducirlo a nitrógenoorgánico. Por ejemplo:Rhizobium legominosarum.

OrganotrofasPAU
Las bacterias heterótrofas, se nutren a expensas de compuestosorgánicos elaborados por otros seres vivos, que descomponen mediante ladigestión (hidrólisis) realizada por enzimas liberados por exocitosis.
Las saprófitas realizan fermentaciones y putrefacciones de gran interésecológico e industrial. Las simbióticas se encargan, por ejemplo, dedegradar la celulosa en los rumiantes, de fijar N2 atmosférico en losnódulos de las leguminosas, etc. Las parásitas producen enfermedades.
Pueden ser fotoorganotrofas,que usan la luz como fuente deenergía o quimioheterótrofos,si usan las energía dereacciones químicas comofuente de energía para generarATP, pero en ambos casos,usan compuestos orgánicos deotros organismos como fuentede carbono.

Comparación tipos de nutriciónDP/PAU
Fuente de carbono
Compuestosinorgánicos como CO2
Compuestos orgánicos
Fuente de energía
Luz Fotoautótrofo
(bacterias verdes sulfurosas: Chlorobium)
Fotoheterótrofo
(bacterias púrpuras no sulfurosas:
Rhodospirillum)
Química Quimioautótrofo
(bacterias oxidantes del hierro: Thiobacillus
ferrooxidans)
Quimioheterótrofo
(La mayoría de bacterias:
E. coli)

CianobacteriasPAU
Son un grupo de bacterias muy particulares. Son procariotas autótrofosque tienen casi la misma forma y tamaño que las algas unicelulares. Igualque ellas hacen la fotosíntesis oxigénica.
Tienen la pared como las Gramnegativas y muchas producenenvueltas mucilaginosas que permitenla formación de colonias.
Internamente poseen un complejosistema multilaminar de membranascon las sustancias necesarias para lafotosíntesis oxigénica (pigmentos,enzimas, etc.). Todas poseen losfotosistemas I y II.
Algunas cianobacterias, como lacianobacteria filamentosa Anabaena,pueden fijar el nitrógeno atmosféricoen unas células especializadas(incoloras y redondeadas) llamadasheterocistos.

CianobacteriasPAU
Su nutrición es muy simple: al ser fotosintéticas, obtienen los compuestosorgánicos a partir de inorgánicos y energía solar. Utilizan nitratos o amoníacocomo fuente de N. Otras, como Anabaena, pueden fijar el N2.
No se mueven (carecen de flagelos). Se reproducen por fisión binaria,gemación, fragmentación y por fisión múltiple. Viven aisladas o formandocolonias. Se encuentran en casi todos los ambientes (resistencia a ladesecación y a las altas temperaturas).
Algunas viven en simbiosiscon plantas. La cianobacteriafija el nitrógeno atmosférico(es innecesario agregarfertilizantes nitrogenadospara la planta) y la planta leproporciona un ambienteprotegido.
Algunas especies puedenenvenenar a animalespróximos ya que producentoxinas.

Los virus como microorganismosDP/PAU
Los virus se clasifican por su tamaño, dentro del grupo de losmicroorganismos, sin embargo, presentan una serie de característicasúnicas que los diferencian del resto de microorganismos:
1. Estructura acelular.
2. Carencia de metabolismopropio.
3. Genoma de ARN enalgunos tipos de virus.
4. Presencia de uno, peronunca de los dos tipos deácidos nucleicos.

¿Cómo clasificamos a los virus? DP/PAU
Existe gran controversia respecto a laconsideración de los virus como seresvivos.
Dado que los virus son formas acelularesy no pueden reproducirse sin utilizar lamaquinaria celular de otro organismo, nose consideran seres vivos, sino parásitosintracelulares obligados.
Virus influenza Los virus presentan dos estados:
- Extracelular metabólicamente inerte,denominado virión o partícula vírica.
- Intracelular, que se adquiere cuando elvirus infecta a una célula huésped.
Según el hospedador al que parasitan,hay virus bacterianos (bacteriófagos ofagos), virus vegetales y virus animales.

Diversidad estructural en los virusPAU
La estructura básica de un virus está formada por un ácido nucleico(material genético), varias enzimas y una cubierta proteica (cápsida)que puede estar desnuda (bacteriófagos) o cubierta por una envolturalipoproteica (VIH).
- El material genético puede ser ADN (monocatenario o bicatenario) oARN (monocatenario o bicatenario). Cada virus sólo posee un tipo deácido nucleico. La cantidad de información genética contenida va desde3-4 genes en virus pequeños hasta centenares de genes en los grandes.
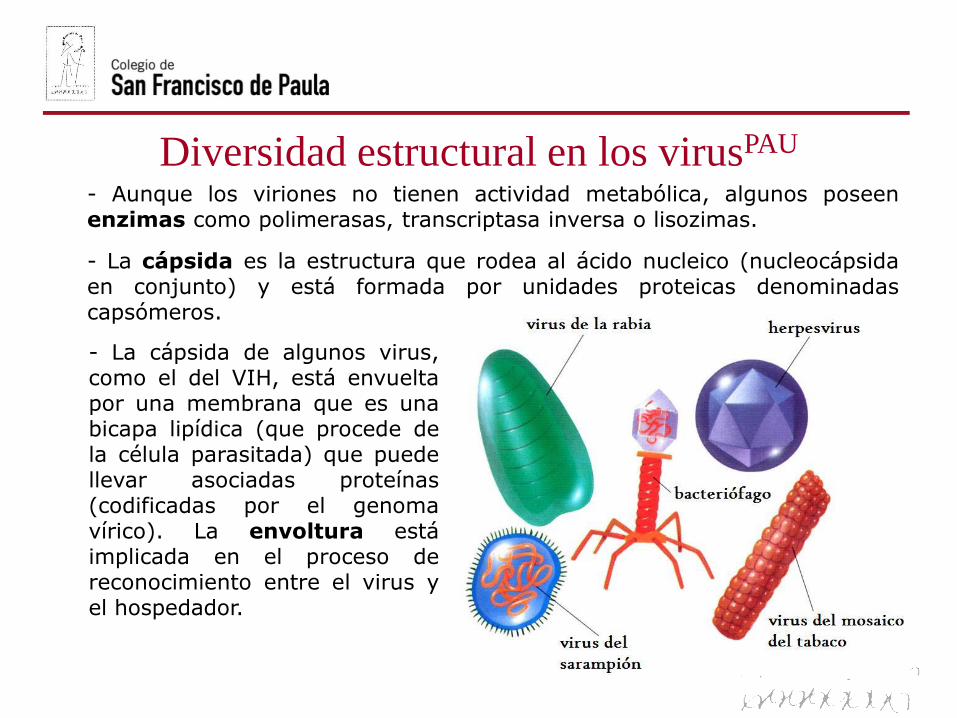
Diversidad estructural en los virusPAU
- Aunque los viriones no tienen actividad metabólica, algunos poseenenzimas como polimerasas, transcriptasa inversa o lisozimas.
- La cápsida es la estructura que rodea al ácido nucleico (nucleocápsidaen conjunto) y está formada por unidades proteicas denominadascapsómeros.
- La cápsida de algunos virus,como el del VIH, está envueltapor una membrana que es unabicapa lipídica (que procede dela célula parasitada) que puedellevar asociadas proteínas(codificadas por el genomavírico). La envoltura estáimplicada en el proceso dereconocimiento entre el virus yel hospedador.

Según la simetría de su cápsida los virus pueden ser fundamentalmente:
- Helicoidales o cilíndricos, como el virus del mosaico del tabaco (VMT) oel de la rabia.
- Poliédricos, siendo los más simples los icosaédricos con 20 carastriángulos equiláteros, como los adenovirus o el virus de la polio.
Diversidad estructural en los virusPAU

Estructura del bacteriófago T4PAU
- Complejos, como los bacteriófagos, que son el resultado de combinarambas estructuras, siendo la porción poliédrica la cabeza y la helicoidal lacola, separados por un cuello formado de capsómeros distintos.
La cabeza está formada por lacápsida proteica conteniendo elmaterial genético (ADN)
El cuello formado de uncollar de capsómeros entrela cabeza y la cola.
La cola es una vaina contráctil parainyectar el ADN, con una placa basalen su extremo que posee unospuntos de anclaje que sirven parafijar el virus a la membrana celular.De la placa salen también unas fibrasproteicas que ayudan a la fijacióndel virus sobre la célula hospedadora.
(1)
(2)
(1)
(3)
(4)(5)
(5)
(6)
(7)
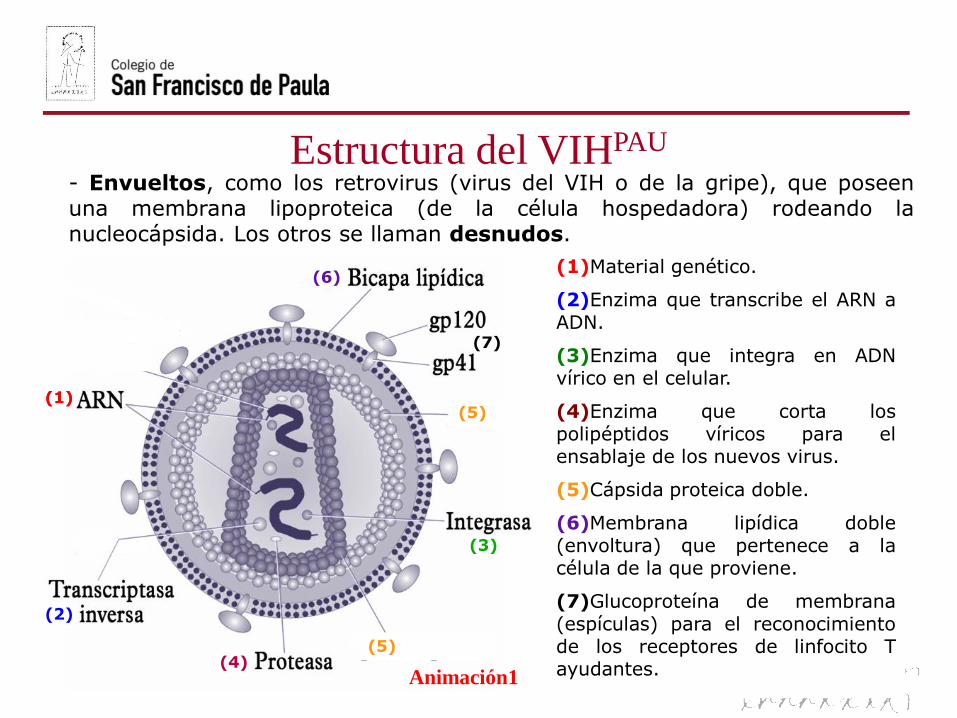
Estructura del VIHPAU
Animación1
(1)Material genético.
(2)Enzima que transcribe el ARN aADN.
(3)Enzima que integra en ADNvírico en el celular.
(4)Enzima que corta lospolipéptidos víricos para elensablaje de los nuevos virus.
(5)Cápsida proteica doble.
(6)Membrana lipídica doble(envoltura) que pertenece a lacélula de la que proviene.
(7)Glucoproteína de membrana(espículas) para el reconocimientode los receptores de linfocito Tayudantes.
- Envueltos, como los retrovirus (virus del VIH o de la gripe), que poseenuna membrana lipoproteica (de la célula hospedadora) rodeando lanucleocápsida. Los otros se llaman desnudos.
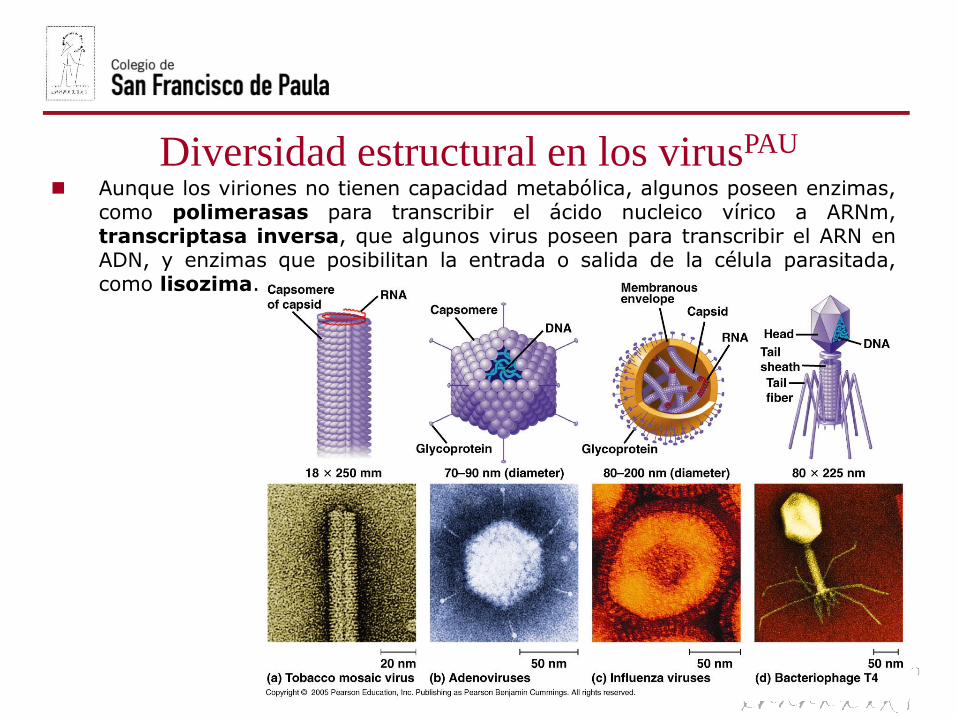
Diversidad estructural en los virusPAU
Aunque los viriones no tienen capacidad metabólica, algunos poseen enzimas,como polimerasas para transcribir el ácido nucleico vírico a ARNm,transcriptasa inversa, que algunos virus poseen para transcribir el ARN enADN, y enzimas que posibilitan la entrada o salida de la célula parasitada,como lisozima.

Ciclo de vida de un virusPAU
Los virus son formas acelulares e inanimadas en estado extracelular. Enel estado de virión "solamente" esperan encontrar una célulahospedadora donde poder realizar su único objetivo, la producción denuevas partículas víricas.
Los virus utilizan a la célula huéspedpara su replicación, logrando que éstasintetice todos sus componentes paragenerar nuevas partículas víricas. Paraello, penetran en la célulahospedadora, utilizan su maquinariareplicativa, y al abandoner la célulahuésped, ésta termina lisándose(muriendo). Este proceso recibe elnombre de ciclo lítico.
Sin embargo, no siempre se produce la lisiso rotura de la célula. Así, algunos viruspenetran en las células hospedadoras ypermanecen en ellas sin producir nuevaspartículas víricas completas. Estos virussiguen un ciclo lisogénico.
IMAGEN: cienciaybiologia.com
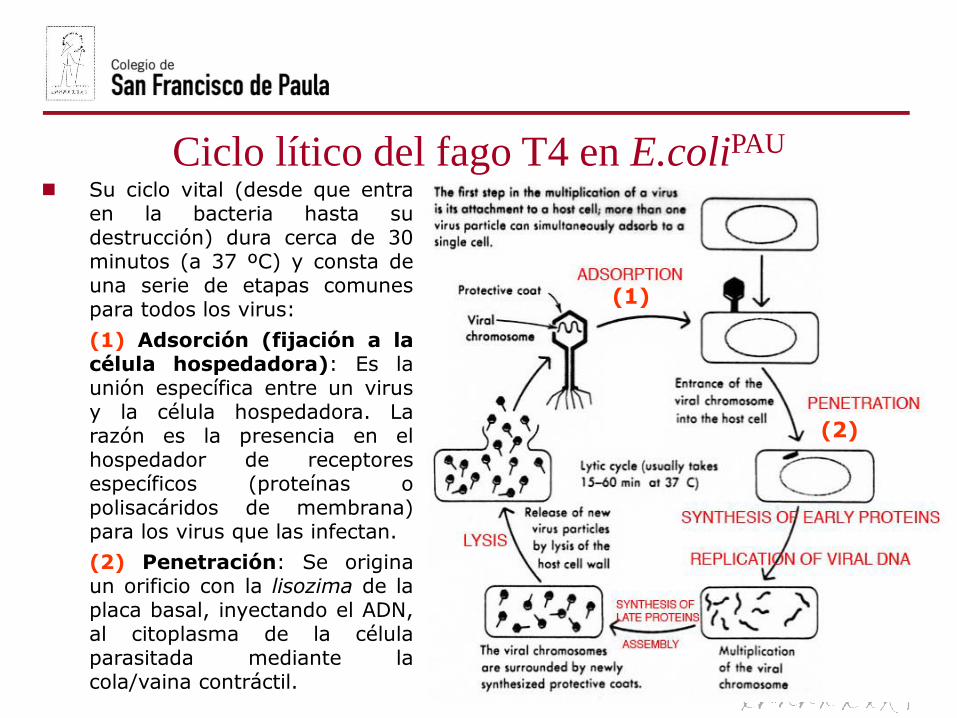
Ciclo lítico del fago T4 en E.coliPAU
Su ciclo vital (desde que entraen la bacteria hasta sudestrucción) dura cerca de 30minutos (a 37 ºC) y consta deuna serie de etapas comunespara todos los virus:
(1) Adsorción (fijación a lacélula hospedadora): Es launión específica entre un virusy la célula hospedadora. Larazón es la presencia en elhospedador de receptoresespecíficos (proteínas opolisacáridos de membrana)para los virus que las infectan.
(2) Penetración: Se originaun orificio con la lisozima de laplaca basal, inyectando el ADN,al citoplasma de la célulaparasitada mediante lacola/vaina contráctil.
(1)
(2)
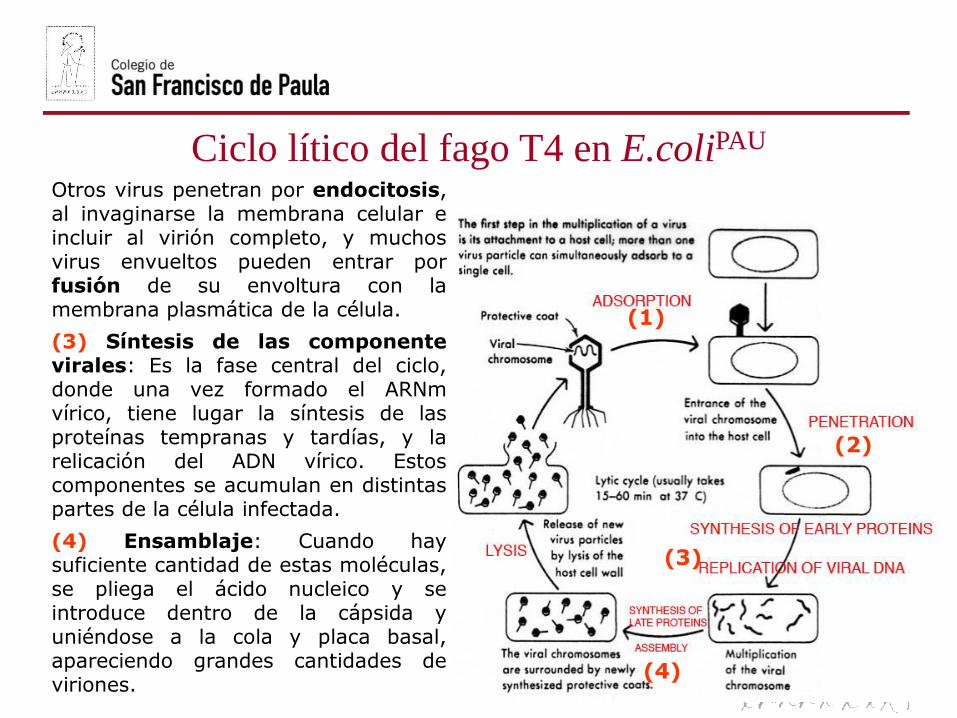
Ciclo lítico del fago T4 en E.coliPAU
Otros virus penetran por endocitosis,al invaginarse la membrana celular eincluir al virión completo, y muchosvirus envueltos pueden entrar porfusión de su envoltura con lamembrana plasmática de la célula.
(3) Síntesis de las componentevirales: Es la fase central del ciclo,donde una vez formado el ARNmvírico, tiene lugar la síntesis de lasproteínas tempranas y tardías, y larelicación del ADN vírico. Estoscomponentes se acumulan en distintaspartes de la célula infectada.
(4) Ensamblaje: Cuando haysuficiente cantidad de estas moléculas,se pliega el ácido nucleico y seintroduce dentro de la cápsida yuniéndose a la cola y placa basal,apareciendo grandes cantidades deviriones.
(4)
(1)
(2)
(3)

Ciclo lítico del fago T4 en E.coliPAU
(5) Liberación: Salen de lacélula las partículas víricas oviriones por diferentesprocedimientos:
- Lisis o desintegración de lacélula infectada, es el másfrecuente y está provocada porenzimas víricos que rompen lamembrana celular.
- Gemación, consistente en unproceso inverso a la penetraciónpor fusión. Es típico de virusenvueltos.
Animación2
(3)
(4)
(1)
(2)
(5)

Ciclo lisogénico del fago λ en E.coliPAU
La mayoría del los virus bacterianos son virulentos,es decir, siempre que penetran en una bacteriarealizan el ciclo lítico.
Sin embargo, hay una serie de virus, llamadosatemperados, que son capaces de permanecer enestado latente en la célula que parasitan, gracias a laintegración de su ADN vírico en el ADN celular. En estecaso el ciclo se llama lisogénico.
La forma integrada del virus en el ADN celular sedenomina provirus o profago.
El fago λ tras la adsorción y penetración y una vezdentro el ADN vírico, éste puede seguir un ciclo lítico olisógeno en función de la concentración dedeterminadas proteínas represoras, que regulan laexpresión del ácido nucleico del virus.
Si son abundantes, impiden la expresión de proteínasvíricas a excepción de la enzima integrasa.

Ciclo lisogénico del fago λ en E.coliPAU
En general, el genoma del virus queda silenciado y nose expresa, replicándose con el ADN bacterianogeneración tras generación.
Las bacterias que tienen integrado un profago soninmunes a la infección por fagos similares (inmunidada la superinfección).
Bajo determinados circunstancias (luz UV), se induceel ciclo lítico, soltándose el profago del ADN bacterianopara expresar sus genes y culminar el ciclo lítico.
Algunas veces, al soltarse el profago, puede quitarlealgunos genes a la bacteria a la que va a destruir einsertárselos a la próxima a la que le haga el ciclolisogénico (transducción).
Animación3

Ciclo lisogénico del fago λ en E.coliPAU

Ciclo de infección del VIHPAU
1.Adsorción y penetración. Los receptores gp120 del virus se unen a losreceptores CD4 del linfocito Th (se necesitan, además, otros correceptores de lamembrana del linfocito). Esto provoca la fusión de las dos bicapas de lípidos,penetrando el virus en la célula. Ahora se desprende de la cápsida proteica yquedan libres las dos moléculas de ARN y las dos enzimas transcriptasas inversas.
Animación4
2.Retrotranscripción. Cadauna de las retrotranscriptasasutiliza una cadena de ARN comomolde para fabricar una cadenacomplementaria de ADN(ADNc). La misma enzimafabrica otra cadena de ADNcomplementaria a la de ADNc.
3.Migración y 4.Integración.Se forma una doble hélice deADN vírico que se integra, porun determinado punto, en elgenoma del Th mediante laenzima integrasa. Se convierte,por tanto, en provirus.
IMAGEN: maph49.galeon.com/
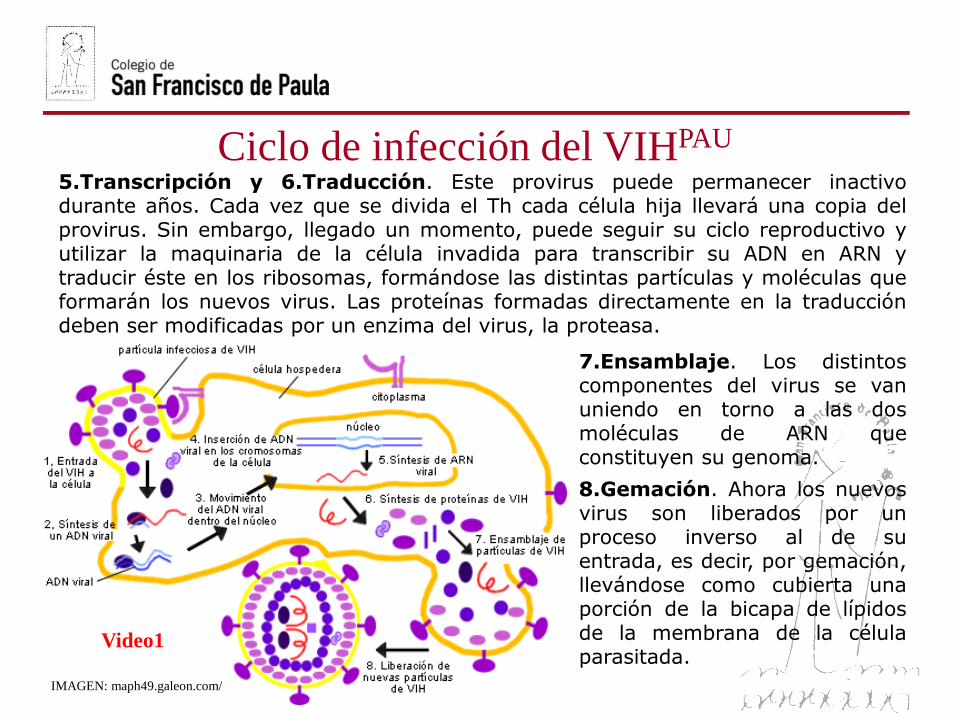
Ciclo de infección del VIHPAU
5.Transcripción y 6.Traducción. Este provirus puede permanecer inactivodurante años. Cada vez que se divida el Th cada célula hija llevará una copia delprovirus. Sin embargo, llegado un momento, puede seguir su ciclo reproductivo yutilizar la maquinaria de la célula invadida para transcribir su ADN en ARN ytraducir éste en los ribosomas, formándose las distintas partículas y moléculas queformarán los nuevos virus. Las proteínas formadas directamente en la traduccióndeben ser modificadas por un enzima del virus, la proteasa.
7.Ensamblaje. Los distintoscomponentes del virus se vanuniendo en torno a las dosmoléculas de ARN queconstituyen su genoma.
8.Gemación. Ahora los nuevosvirus son liberados por unproceso inverso al de suentrada, es decir, por gemación,llevándose como cubierta unaporción de la bicapa de lípidosde la membrana de la célulaparasitada.
Video1
IMAGEN: maph49.galeon.com/
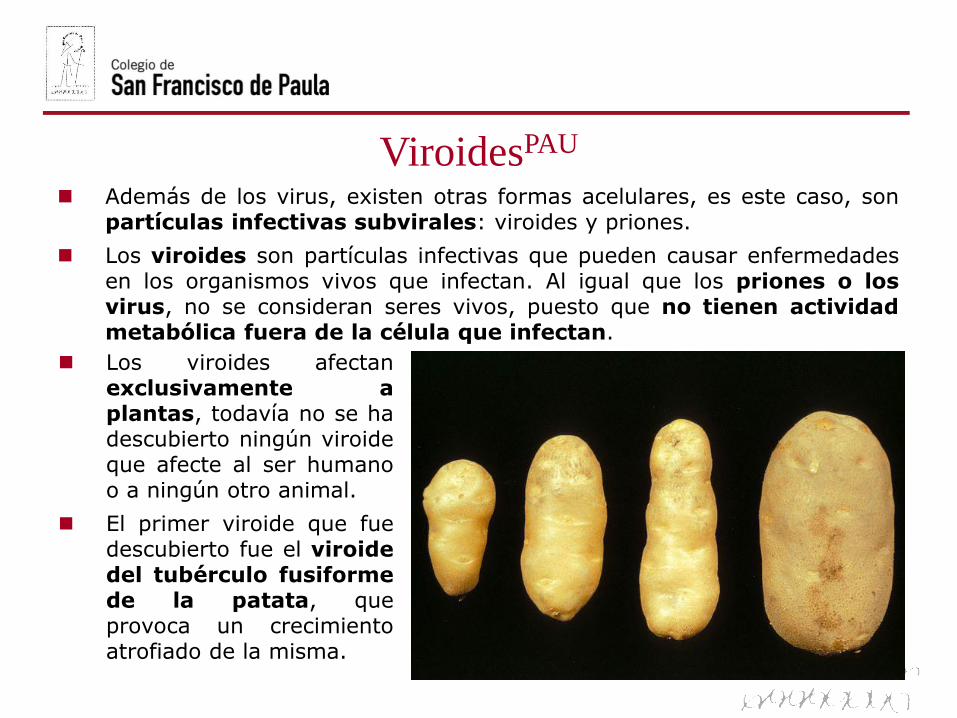
ViroidesPAU
Además de los virus, existen otras formas acelulares, es este caso, sonpartículas infectivas subvirales: viroides y priones.
Los viroides son partículas infectivas que pueden causar enfermedadesen los organismos vivos que infectan. Al igual que los priones o losvirus, no se consideran seres vivos, puesto que no tienen actividadmetabólica fuera de la célula que infectan.
Los viroides afectanexclusivamente aplantas, todavía no se hadescubierto ningún viroideque afecte al ser humanoo a ningún otro animal.
El primer viroide que fuedescubierto fue el viroidedel tubérculo fusiformede la patata, queprovoca un crecimientoatrofiado de la misma.

ViroidesPAU
Los viroides están formados exclusivamente por una cadena circularde ARN monocatenario de pequeño tamaño pero de altaestructura secundaria y capacidad autocatalítica.
El viroide no codifica proteínas de ninguna clase. Esa es la principaldiferencia con los viriones y los virus. Por otro lado, el hecho de poseermaterial hereditario es lo que los diferencia de los priones.
Hay muchas cuestiones de los viroides que aún se desconocen. No sesabe, por ejemplo, cómo logran replicarse, aunque sí se advierte quesuelen alojarse en los núcleos de las células.

PrionesPAU
Como se ha dicho anteriormente, además de los viroides, existen otraspartículas infectivas subvirales denominadas priones, formasacelulares y de menor tamaño que un virus.
Los priones son partículas infectivas formadas de unaproteína denominada priónica capaz de formar agregados molecularesaberrantes.
Los priones producen las encefalopatías espongiformestransmisibles, un grupo de enfermedades neurológicas degenerativastales como la enfermedad de Creutzfeldt-Jakob y la encefalopatíaespongiforme bovina.
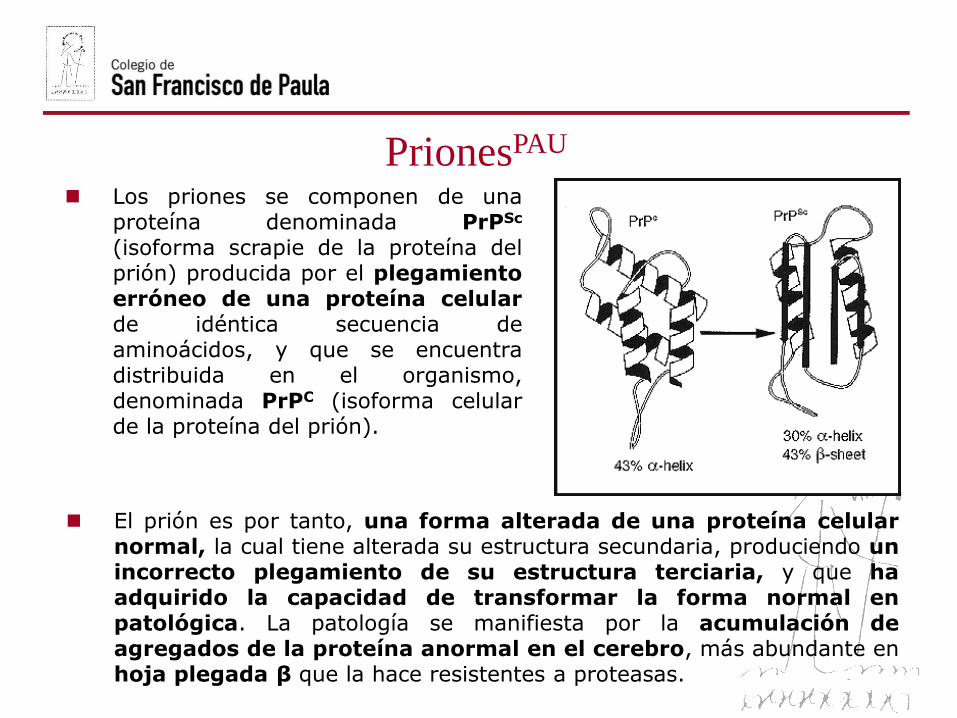
PrionesPAU
Los priones se componen de unaproteína denominada PrPSc
(isoforma scrapie de la proteína delprión) producida por el plegamientoerróneo de una proteína celularde idéntica secuencia deaminoácidos, y que se encuentradistribuida en el organismo,denominada PrPC (isoforma celularde la proteína del prión).
El prión es por tanto, una forma alterada de una proteína celularnormal, la cual tiene alterada su estructura secundaria, produciendo unincorrecto plegamiento de su estructura terciaria, y que haadquirido la capacidad de transformar la forma normal enpatológica. La patología se manifiesta por la acumulación deagregados de la proteína anormal en el cerebro, más abundante enhoja plegada β que la hace resistentes a proteasas.

PrionesPAU
A partir de tejido cerebral procedente de un individuo infectado sepueden extraer vesículas membranosas ricas en PrPSc. La proteólisisparcial de estas vesículas da lugar a partículas con forma de varilla,visibles al microscopio electrónico.
Esta forma anormal (PrPSc) puede trasmitirse mediante el consumode animales infectados y por contacto directo con fluidos corporales oincluso instrumentos, ya que los priones son resisten a la esterilización.No existe cura para la enfermedad.
Web4

PrionesPAU
El plegamiento erróneo de la Prpc a PrpSc confiere a la PrpSc dospropiedades que la diferencian de la Prpc: la resistencia parcial a ladigestión por proteasas y su insolubilidad. Estas dos propiedades hacenque la PrpSc sea estable y la capacitan para poder formar agregadosproteicos responsables de la acumulación de PrpSc en forma de placasamiloides en el tejido nervioso.
Las principales diferencias entre la forma normal del prion, Prpc, y laforma patógena, PrpSc son las siguientes:
PrPC PrPSC
Estructura alfa-hélice Estructura hoja plegada beta
Susceptible a proteasas Resistente a proteasas
Proteína monomérica Agregados proteicos
Resistencia normal Altísima resistencia a la radiación ydisolventes

PrionesPAU
Una proteína PrpSc actúa de agente infeccioso al contactar con unaproteína PrP normal, haciendo que adquiera estructura plana en forma delámina beta, es decir, pasará a ser una PrpSc. Como consecuencia de estecambio conformacional, la nueva proteína no puede ser degradada yactúa como agente infeccioso sobre otros Prpc provocando el malplegamiento de una manera exponencial.
Este hecho es el causantede la acumulación deagregados de PrpSc enforma de placas amiloides,agregados proteicospatógenos que se acumulanen forma de fibrasinsolubles y que matan lasneuronas produciendoagujeros en el cerebro.









![[XLS] · Web view4.95. 4.95. 5. 5. 5. 5. 5. 5. 4.95. 5. 5. 4.95. 4.95. 5. 4.95. 5. 5. 4.95. 4.95. 5. 5. 5. 5. 5. 5. 4.95. 5. 4.95. 4.95. 4.95. 4.95. 5. 5. 4.95. 4.95. 5. 5. 4.95.](https://static.fdocuments.ec/doc/165x107/5ba3371b09d3f2cc2e8da3f6/xls-web-view495-495-5-5-5-5-5-5-495-5-5-495-495-5-495.jpg)








