![ANATOMÍA: Sistema Linfático. [BÁSICO]](https://static.fdocuments.ec/doc/165x107/55c6d0a7bb61eb920a8b4614/anatomia-sistema-linfatico-basico.jpg)
Sistema Linfático
13
Sistema Linfático 2.3 Los tejidos: el sistema Linfoide Los órganos linfoides se pueden clasificar en: órganos linfoides primarios o centrales y secundarios o periféricos (desde un punto de vista funcional) y encapsulados y difusos (desde un punto de vistaanatómico-estructural). En los órganos linfoides primarios es donde se produce la diferenciación de linfocitos (linfopoyesis) T y B. La de linfocitos B ocurre en hígado fetal y médula ósea. La de linfocitos T sucede en el timo. En los órganos linfoides secundarios se presentan los antígenos y se monta la respuesta inmune específica (ganglios linfáticos, bazo, MALT [tejido linfoide asociado a mucosas]) Los conductos linfáticos se distribuyen por todo el organismo, llegan a todas las zonas y tienen cadenas de ganglios intercalados. Destacan las cadenas ganglionares localizadas en la zona inguinal, axilar y amigdalar. El punto de conexión entre vasos linfáticos y vasos sanguíneos es el llamado Ducto (o conducto) torácico: la linfa se vuelca en la vena subclavia. No hay que confundir el concepto de “ganglio linfático” con el de folículo linfoide. Estos últimos no son otra cosa que acumulaciones de linfocitos que adquieren forma esférica. Es un modo, pues, de organización de tejidos linfoides. Existen folículos linfoides en todos los órganos linfoides encapsulados: ganglios, bazo, timo. Además, en los órganos linfoides difusos (como el MALT) se han observado la presencia de folículos linfoides en unas estructuras denominadas Placas de Peyer, pero no en el resto del tejido. 2.4 Órganos Linfoides Primarios. a) Médula Ósea: La médula ósea está formada por islotes de células hematopoyéticas situados en el interior de los huesos. Todas las células del sistema inmune se originan a partir de las células hematopoyéticas primordiales pluripotentes (células stem) de la médula ósea a través de los linajes mieloide y linfoide. Durante la edad fetal estas funciones se realizan por el hígado, que abandona esta actividad después del nacimiento. Además, la médula ósea actúa como órgano
-
Upload
jairo-ignacio-cz -
Category
Documents
-
view
558 -
download
5
Transcript of Sistema Linfático

Sistema Linfático
2.3 Los tejidos: el sistema Linfoide
Los órganos linfoides se pueden clasificar en: órganos linfoides primarios o centrales y secundarios o periféricos (desde un punto de vista funcional) y encapsulados y difusos (desde un punto de vistaanatómico-estructural).
En los órganos linfoides primarios es donde se produce la diferenciación de linfocitos (linfopoyesis) T y B. La de linfocitos B ocurre en hígado fetal y médula ósea. La de linfocitos T sucede en el timo.
En los órganos linfoides secundarios se presentan los antígenos y se monta la respuesta inmune específica (ganglios linfáticos, bazo, MALT [tejido linfoide asociado a mucosas])
Los conductos linfáticos se distribuyen por todo el organismo, llegan a todas las zonas y tienen cadenas de ganglios intercalados. Destacan las cadenas ganglionares localizadas en la zona inguinal, axilar y amigdalar. El punto de conexión entre vasos linfáticos y vasos sanguíneos es el llamado Ducto (o conducto) torácico: la linfa se vuelca en la vena subclavia.
No hay que confundir el concepto de “ganglio linfático” con el de folículo linfoide. Estos últimos no son otra cosa que acumulaciones de linfocitos que adquieren forma esférica. Es un modo, pues, de organización de tejidos linfoides. Existen folículos linfoides en todos los órganos linfoides encapsulados: ganglios, bazo, timo. Además, en los órganos linfoides difusos (como el MALT) se han observado la presencia de folículos linfoides en unas estructuras denominadas Placas de Peyer, pero no en el resto del tejido.
2.4 Órganos Linfoides Primarios.
a) Médula Ósea:
La médula ósea está formada por islotes de células hematopoyéticas situados en el interior de los huesos. Todas las células del sistema inmune se originan a partir de las células hematopoyéticas primordiales pluripotentes (células stem) de la médula ósea a través de los linajes mieloide y linfoide. Durante la edad fetal estas funciones se realizan por el hígado, que abandona esta actividad después del nacimiento. Además, la médula ósea actúa como órgano linfoide secundario (diferenciación final de células B a células plasmáticas).
b) Timo:
Los precursores de los Linfocitos T llegan por vía arterial llegan a la corteza y a través de los capilares pasan a la médula. De la médula salen por los capilares venosos. Los linfocitos se diferencian en el trayecto de la corteza a la médula. La diferenciación consiste en la presentación por parte de las células epiteliales de sus proteínas HLA sucediendo la llamada selección positiva. Después las células dendríticas y los macrófagos enseñan a los timocitos los antígenos HLA con péptidos propios en su hendiduda (selección negativa).Con esta selección se eliminan el 95 % de los posibles linfocitos T. La selección positiva (elimina linfocitos T con receptores poco apropiados) se realiza en la corteza y en la selección negativa (médula ) se eliminan los linfocitos que reconocen elementos propios del organismo.

2.5 Órganos Linfoides secundarios:
a) Ganglio Linfático:
Presenta dos vías; las de entrada son conductos linfáticos aferentes , venas postcapilares y arterias postcapilares. La de salida es un conducto linfático eferente. Existen tres zonas estructuralmente distinguibles:
-corteza , en esta zona existen células B y folículos linfoides. Estos folículos pueden ser primarios (presentan células B vírgenes en reposo) o secundarios (presentan centros germinales con Linfocitos B activados tras la presentación de antígenos)
-paracorteza, muy rica en linfocitos T.
-médula , en esta zona se encuentran los linfocitos maduros que están listos para salir del ganglio.
b) Bazo:
En la pulpa blanca se realiza la presentación de antígenos. Los linfocitos llegan por la arteria esplénica y capilares arteriales y salen por las venas y vasos linfáticos eferentes. En la pulpa blanca existen folículos linfoides.
c) Tejido Linfoide Asociado a Mucosas (MALT):
Son agrupaciones de tejido linfoide no encapsulado, situado en la lámina propia y áreas submucosas de los tractos gastro-intestinal (GALT), respiratorio (BALT) y tracto génito-urinario. Tiene particular interés (dada su extensión) el tejido asociado a la mucosa gastro-intestinal o GALT.
En las microvellosidades de los enterocitos existen redes capilares y vénulas además de un conducto linfático que recibe el nombre de “lacteal”. Los linfocitos están dispersos (tejido difuso) en todo el tejido, salvo en las placas de Peyer, donde existen folículos linfoides no encapsulados pero que aparecen agrupados.
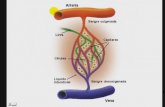
2.6 Recirculación de linfocitos en el organismo:Hay 2 sistemas circulatorios en el cuerpo: la sangre y la linfa. La sangre llega hasta todos los tejidos a través de arterias, arteriolas y capilares arteriales. Parte del fluido sanguíneo de los tejidos drena y entra en los conductos linfáticos eferentes. Así los canales linfáticos forman una red, cuando confluyen varios canales se constituyen los núdulos linfáticos a los que llegan varios conductos aferentes (de entrada), y que drenan por un único eferente (de salida). Finalmente, la linfa encuentra el camino hacia el llamado Ducto torácico que es donde la linfa se vuelca a la sangre (el ducto torácico se funde con la vena subclavia). Una característica única de los linfocitos es que pueden cruzar el cuerpo a través de la sangre y la linfa. Este tráfico de sangre a linfa se denomina “recirculación linfocitaria”. Los linfocitos abandonan los tejidos infectados hacia los ganglios linfáticos regionales. Allí, son activados tras encontrar células presentadoras de antígeno. Una vez activados, vía conductos linfáticos se vuelcan en el ducto torácico a la circulación sanguínea. Y por último, a través de la circulación vuelven al tejido infectado para ejercer su función.

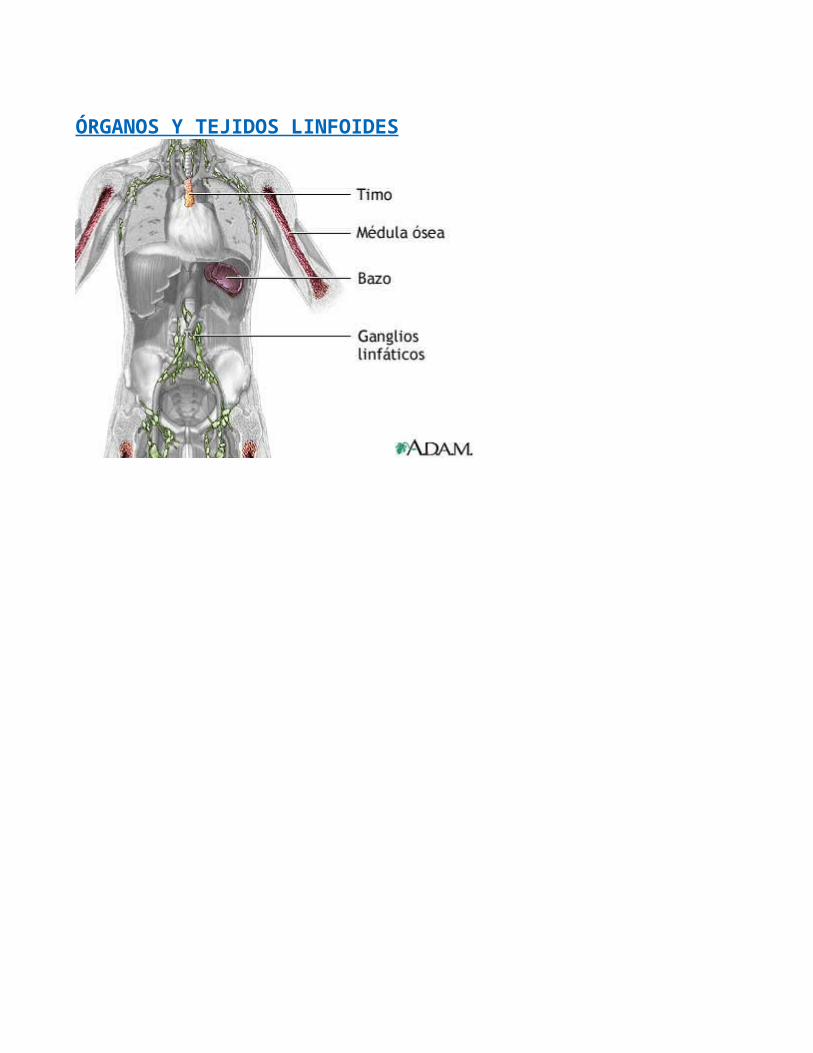
ÓRGANOS Y TEJIDOS LINFOIDES
Los órganos y tejidos del sistema linfático, distribuidos ampliamente en todo el cuerpo, se
clasifican en dos grupos con base en sus funciones. Los órganos linfáticos
primarios constituyen el ambiente idóneo para la división de las células madre y su
maduración en células B y T, o sea, los linfocitos que se encargan de las respuestas
inmunitarias. Dichos órganos son la médula ósea roja, que se encuentra en los huesos
planos y en las epífisis de los huesos largos de adultos, y el timo. Las células madre
pluripotenciales de la médula ósea roja son el origen de las células B maduras y las células
pre-T, de las cuales estas últimas emigran al timo y maduran en él. Los órganosy tejidos
linfáticos secundarios, donde tiene lugar gran parte de las respuestas inmunitarias,
comprenden los ganglios y folículos linfáticos y el bazo. Se considera que el timo, los
ganglios y el bazo son órganos porque los rodea una cápsula de tejido conectivo, mientras
que los folículos no lo son por carecer de ella.
Timo
Esta glándula por lo regular consta de dos lóbulos y se localiza en el mediastino, detrás del esternón. Una capa de tejido conectivo envuelve y mantiene unidos los dos lóbulos tímicos; mientras que una cápsula de tejido conectivo delimita por separado cada lóbulo. Las trabéculas son prolongaciones de la cápsula que penetran en los lóbulos y los dividen enlobulillos. Cada uno consta de corteza externa, que se tiñe de color oscuro, y médula, de color claro a la tinción. La corteza se compone de linfocitos estrechamente apiñados, células epiteliales denominadas epiteliales reticulares que rodean a grupos de linfocitos, y macrófagos. La médula contiene ante todo células epiteliales reticulares, además de linfocitos muy dispersos. Aunque se conocen sólo algunas de sus funciones, las células reticulares producen hormonas tímicas, que, según se piensa, contribuyen a la maduración de las células T. Además, en la médula existen los corpúsculos del timo ( o de Hassall) característicos, los cuales son capas concéntricas de células epiteliales reticulares aplanadas y llenas de gránulos de queratohialina y queratina.
Ganglios linfáticos
Se llama ganglios linfáticos a casi 600 órganos en forma de frijol dispuestos a lo largo de los vasos linfáticos. Están dispersos en todo el cuerpo, tanto en capas superficiales como profundas, usualmente en grupos. Los ganglios son abundantes cerca de las glándulas mamarias y en las axilas e ingles.
Bazo
El bazo es un órgano oval y la masa más grande de tejido linfático del cuerpo. Se situa en el hipocondrio izquierdo, entre el estómago y el diafragma. Su cara superior, lisa y convexa, se conforma a la superficie cóncava del diafragma. Los órganos adyacentes producen depresiones en la cara visceral del bazo: la impresión gástrica para el estómago, la renal para el riñón y la cólica para el ángulo esplénico del colon. Al igual que los ganglios linfáticos, el bazo posee un líquido, que cruzan la arteria y vena esplénicas, así como vasos linfáticos eferentes.
Una cápsula de tejido conectivo denso envuelve el bazo. Las trabéculas penetran desde la cápsula, que a su vez tiene el recubrimiento de una membrana serosa, el peritoneo visceral. Cápsula, trabéculas, fibras reticulares y fibroblastos constituyen el estroma del bazo, mientras que su parénquima consiste en dos tipos de tejido, las pulpas blanca y roja. La pulpa blancaconsta de tejido linfático, principalmente linfocitos y macrófagos, dispuestos alrededor de ramas de la arteria esplénica, llamadas arterias centrales. Lapulpa roja está formada por senos venosos llenos de sangre y cordones de tejido esplénico, llamados cordones esplénicos ( de la pulpa roja o de Billroth). Éstos se componen de eritrocitos, macrófagos, linfocitos, células plasmáticas y granulocitos. Las venas guardan relación estrecha con la pulpa roja.
La sangre que fluye hacia el bazo por la arteria esplénica llega a las arterias centrales de la pulpa blanca. En ésta, las células T y B llevan a cabo funciones inmunitarias, al tiempo que los macrófagos destruyen los

microbios patógenos provenientes de la sangre por fagocitosis. La pulpa roja del bazo se encarga de tres funciones relacionadas con los elementos formes de la sangre:
1) Eliminación de células sanguíneas y plaquetas defectuosas, que efectúan los macrófagos.
2) Almacenamiento de plaquetas, que puede ser hasta un tercio del total.
3) Producción de células sanguíneas (hemopoyesis) durante la vida fetal.
El bazo es el órgano afectado con mayor frecuencia en caso de traumatismo abdominal. Su rotura produce hemorragia intraperitoneal grave y choque. La extirpación pronta del órgano (esplenectomía) es necesaria para evitar que el paciente se desangre hasta morir. Otras estructuras, en particular la médula ósea roja y el hígado, pueden encargarse de las funciones que normalmente realiza el bazo.
Folículos linfáticos Son concentraciones ovales de tejido linfático que no están rodeadas por una cápsula. Debido a que se encuentran dispersos en la lámina propia (tejido conectivo) de la mucosa que reviste los aparatos digestivo, urinario, reproductor y respiratorio, los folículos linfáticos también se denominan tejido linfoide relacionado con mucosas (TLRM).
Aunque muchos ganglios linfáticos son pequeños y están aislados, otros forman grandes agregados en partes específicas del cuerpo. Entre éstos se incluyen las amígdalas en la región faríngea y las placas de Peyer en el íleon. También se encuentran tales cúmulos en el apéndice. Por lo general hay cincoamígdalas en la unión de la boca con la bucofaringe y de la nariz con la nasofaringe. Así pues, las amígdalas están dispuestas estratégicamente para participar en respuestas inmunitarias contra sustancias extrañas inhaladas o ingeridas. La amígdala faríngea o tejido adenoide está incluido en la pared posterior de la nasofaringe. Las dos amígdalas palatinas se localizan en la parte posterior de la boca, una en cada lado, y son las que habitualmente se extraen en la amigdalectomía. El par de amígdalas linguales, situadas en la base de la lengua, también se extirpan a veces en dicha intervención quirúrgica.

Sistema Inmune Innato en vertebrados
En el sistema inmune innato de vertebrados intervienen principalmente dos componentes, el celular y el
humoral.
Celular inespecífico. Los animales vertebrados mantienen el sistema inmune presente en invertebrados,
con células fagocíticas inespecíficas, pero incrementan la diversidad de este tipo celular, y están mejor
caracterizadas. De esta manera se encuentran no sólo macrófagos, sino tambiénneutrófilos, que van a jugar
un papel fundamental también como células fagocíticas, junto a eosinófilos, basófilos y mastocitos. Junto a
los macrófagos, como células fagocíticas y células presentadoras de antígeno, los vertebrados poseen otras
células accesorias como son las células dendríticas interdigitantes y las células dendríticas foliculares. Dentro
de la citotoxicidad natural, los animales vertebrados poseen células “natural killer” (NK).
Humoral inespecífico. Con respecto al complemento, a partir de los ciclóstomos (que sólo poseen C3 y sólo
pueden activar la cascada por la vía alternativa), se han caracterizado otros factores que participan en la
activación de la cascada clásica en tiburones, peces, anfibios, aves y mamíferos. Los vertebrados también
poseen otras sustancias con capacidad opsonizante como las proteínas de fase aguda (proteína C reactiva),
similares a las descritas en invertebrados, lectinas solubles, y gran cantidad de factores con actividad
antimicrobiana.
Dentro del Sistema Inmune Innato de los vertebrados, se han caracterizado distintas familias proteicas que
intervienen en el reconocimiento de patógenos, interactuando con sustancias presentes en una gran
variedad de patógenos, lo que se ha denominado como “Reconocimiento de patrones” (Tabla 2). También se
han encontrado en vertebrados, sobre todo en ratones y humanos, gran cantidad de citocinas como
las interleucinas y las quimiocinas, producidas en su mayoría por linfocitos y macrófagos, que participan en
las respuestas inmunitarias (induciendo diferenciación celular, proliferación, inhibición, actividad
quimiotáctica, etc.), y también sobre otros sistemas (hematológicos, nervioso, endocrino).
Sistema Inmune Específico o Adquirido

Los vertebrados son los únicos animales que poseen el sistema inmune adquirido o específico. Excepto los
agnatos (como la lamprea), el resto de animales vertebrados a partir de los peces cartilaginosos
(representados por el tiburón) poseen linfocitos T, B, expresan moléculas de clase I y II del Complejo
Principal de Histocompatibilidad y producen anticuerpos.
Componente celular específico. Los animales vertebrados presentan elementos celulares más
evolucionados (linfocitos T y B), con receptores específicos en su membrana, que van a permitir el
reconocimiento de una gran diversidad de estructuras antigénicas, con capacidad para recordar exposiciones
previas (memoria) y de responder de forma más efectiva en segundas o posteriores exposiciones al mismo
antígeno (maduración de la respuesta).
Evolución de órganos linfoides. La existencia de linfocitos T y B en los animales vertebrados, está asociado
con la presencia de órganos linfoides dedicados a la producción o diferenciación de estas células.
Sin embargo, aunque se encuentran tejidos y órganos linfoides en todos los vertebrados, éstos sufren una
gran evolución desde los peces hasta los mamíferos, tendiendo hacia un mayor grado de complejidad.
Mientras que se han encontrado agregados linfoides asociados al intestino en todos los animales vertebrados,
el timo se encuentra únicamente a partir de los tiburones, y se mantiene ya a lo largo de toda la escala
filogenética como órgano linfoide primario de linfocitos T.
Lo mismo ocurre con el bazo,
que se encuentra en todos los
animales vertebrados, excepto
en los agnatos (Figura 6). Con
respecto a las células
progenitoras, se ha visto que se
necesita una correcta estructura
ósea para el desarrollo de la
médula ósea y ésta no apareció
hasta que los vertebrados se
adaptaron al medio terrestre.
Así los peces no tienen médula
ósea, mientras que los anfibios
tienen ya restos de tejido
linfoide en sus huesos, siendo la médula ya totalmente funcional en anfibios (anuros), reptiles, aves y
mamíferos.
El riñón, en la porción del pronefros o riñón anterior, se comporta como órgano linfoide primario en peces y
algunos anfibios, supliendo la carencia de médula ósea y nódulos linfoides en estos animales. La médula ósea
también ejerce de órgano linfoide primario de linfocitos B en gran parte de los animales vertebrados. Como
excepciones se encuentran las aves, cuyo órgano linfoide primario para células B es la Bolsa de Fabricio (área
especializada de tejido linfoide intestinal), y en ovejas y cerdos, donde además de la médula ósea, se han
implicado a las placas de Peyer del intestino.
Como estructuras linfoides más organizadas aparecen más tarde en la escala filogenética los ganglios
linfáticos, constituidos por folículos primarios de células B, rodeadas de linfocitos T. Aparecen ya bien
definidos a partir de los anfibios anuros, y se encuentran en el resto de animales vertebrados, aunque con

ciertas diferencias entre ellos. Mientras que los anfibios presentan linfocitos alrededor de capilares
sinusoides, los reptiles tienen primitivos nódulos linfoides alrededor de grandes vasos como aorta y cava, y
las aves presentan unas formaciones linfoides equivalentes a los nódulos pero que carecen de cápsula.
Únicamente en aves y mamíferos, animales homeotermos de sangre caliente, surgen los centros
germinales o folículos linfoides secundarios en los órganos linfoides (Figura 6). Como bacterias y virus se
multiplican bien a esas temperaturas, es probable que haya sido necesario el desarrollo de una estructura
que permita la eliminación más efectiva y rápida de dichos patógenos en estos animales.
En estos centros germinales se produce una correcta cooperación entre linfocitos T y B, la generación de
linfocitos de memoria, y la incorporación de mutaciones en los genes de las inmunoglobulinas por un
mecanismo denominado Hipermutación Somática, que incrementa la afinidad del anticuerpo por el antígeno.
En la (Tabla 3) se muestra la afinidad de los anticuerpos por un antígeno específico, comparando aquellos
animales vertebrados de sangre fría (con afinidad baja) con los de sangre caliente (que incrementan la
afinidad de sus anticuerpos en 100 ó 1000 veces).
Componente humoral específico. Los animales vertebrados son los únicos animales capaces de generar
anticuerpos específicos frente a una gran diversidad de patógenos. Con respecto al número y tipo de
inmunoglobulinas que se encuentran en ellos, se observa una evolución hacia una mayor variedad y
complejidad de los anticuerpos en la escala filogenética, siendo los mamíferos los que mayor variedad de
clases de inmunoglobulinas poseen (Figura 7).
Los peces agnatos, los más primitivos
en la escala filogenética, no
presentan anticuerpos como los
descritos para el resto de vertebrados
(con dos cadenas pesadas y dos
cadenas ligeras iguales dos a dos),
sino que presentan anticuerpos
atípicos con cadena pesada m sin
puentes disulfuro (tienen enlaces

tioéster), y carecen de cadena ligera. Esto también se ha descrito para algunas subclases de Igs de camélidos
(camellos) que carecen también de cadena ligera.
El resto de vertebrados presentan todos ellos inmunoglobulina M, siendo en muchos de ellos la única
inmunoglobulina que producen. La inmunoglobulina Y, que aparece en peces pulmonados y se mantiene en
anfibios, reptiles y aves, se considera precursora de las IgE e IgG de los mamíferos por compartir algunas de
sus funciones como su participación en infecciones parasitarias y en la memoria antigénica, respectivamente.
Carece de región bisagra y aparece tanto de forma íntegra como truncada (sin porción Fc). La IgA es
producida por vez primera en las aves y se mantiene también en mamíferos. El mayor número de Igs
diferentes aparecen ya en mamíferos, donde surgen otras clases como la IgG, la IgE y la IgD, con algunas
excepciones (conejos y cerdos carecen de IgD).
Genes de Igs, del TCR y del CM
La manera en que las moléculas de inmunoglobulinas y el receptor de la célula T están codificadas por
segmentos génicos, así como el tipo y estructura de estos segmentos, se encuentran conservados en los
vertebrados en toda la escala filogenética. Los vertebrados (excepto los agnatos) presentan segmentos
génicos V, (D), J y C, que deben de reagruparse para dar lugar finalmente al gen de la cadena pesada (Figura
8) o ligera de las inmunoglobulinas, o a cada una de las cadenas del receptor del linfocito T.
El gran salto cualitativo en la
escala filogenética, con respecto a
los anticuerpos y a su organización
genómica, se produce en los peces
cartilaginosos, donde ya aparecen
los anticuerpos y el receptor del
linfocito T, aunque la estructura
genómica de ambos se organiza
en múltiples clusters del tipo VDJC-
VDJC-VDJC- a lo largo de diversos
cromosomas (Figura 8). Los
reptiles parece que también tienen
una organización génica de los
anticuerpos en grupos, de forma similar a los tiburones.
Sin embargo, los peces óseos ya presentan una mayor diversificación de los genes variables, permitiendo un
mayor repertorio de anticuerpos distintos, para posteriormente diversificarse también los genes constantes.
Los peces óseos, anfibios y mamíferos presentan una distribución de los segmentos en un único cromosoma
para cada una de las cadenas pesadas y ligeras de las inmunoglobulinas y para cada una de las cadenas del
receptor del linfocito T. La distribución de los segmentos agrupa a los genes V, seguidos de un grupo de
genes D, luego de genes J, y seguido por los segmentos constantes (V1-n, D1-n, J1-n, C). Las aves en cambio
presentan un único gen V funcional y diversos pseudogenes, que se reagrupan por el mecanismo de
conversión génica para aumentar la diversidad de los anticuerpos.
El siguiente salto, que va a permitir un incremento de la afinidad de los anticuerpos, es la aparición en aves y
mamíferos de centros germinales en los órganos linfoides secundarios. Los mamíferos son los animales

vertebrados que presentan una mayor variedad de clases de anticuerpos distintos y los que pueden realizar
mejor una adecuada maduración de las respuestas inmunitarias (Figura 9).
Moléculas de Histocompatibilidad
Los genes que codifican las moléculas del complejo principal de histocompatibilidad (CPH) se han encontrado
en todos los vertebrados, sugiriendo que evolucionaron antes de la emergencia de los peces cartilaginosos.
Se ha conservado la estructura básica de las moléculas del CPH, así como la organización de sus genes, tanto
para las Moléculas de Histocompatibilidad de Clase I, II como para las de clase III (factores de
complemento). Gen ancestral común.
El grado de conservación de la estructura en
segmentos génicos V, D, J, C, en los animales
vertebrados a lo largo de la escala filogenética, así
como la existencia de diversas proteínas que
comparten homología estructural con el dominio
plegado de las inmunoglobulinas (forman parte de la
“superfamilia de las inmunoglobulinas”), sugieren la
existencia de un gen ancestral que pudiera dar
origen a los diversos miembros de la superfamilia.
En cuanto al origen de los genes de los anticuerpos
y del receptor del linfocito T, podrían haberse
originado a partir de la segmentación de
un exón que codificaba un único dominio V, en
elementos separados V y J para dar lugar a una proteína tipo cadena ligera (Figura 10). Ésto pudo estar
seguido por una segunda segmentación que originase las regiones D, y posteriores diversificaciones y
participación de trasposones o mecanismos de recombinación intragénica, permitiría originar la variedad de
segmentos génicos de las Igs y de los receptores de los linfocitos T.