
REVISTA CUBANA DE INVESTIGACIONES PESQUERAS 29(1) 2012_… · absorción de agua. Con el empleo del...
81
REVISTA CUBANA DE INVESTIGACIONES PESQUERAS ENERO-DICIEMBRE, 2012, vol. 29, NO. 1 EDITADA POR EL CENTRO DE INVESTIGACIONES PESQUERAS
Transcript of REVISTA CUBANA DE INVESTIGACIONES PESQUERAS 29(1) 2012_… · absorción de agua. Con el empleo del...

REVISTA CUBANADE INVESTIGACIONES PESQUERAS
ENERO-DICIEMBRE, 2012, vol. 29, NO. 1
EDITADA POR EL CENTRO DE INVESTIGACIONES PESQUERAS

Edición, diseño interior y diagramación
Ing. Isis Pérez Hernández
D. R. © Centro de Investigaciones Pesqueras5ta. Ave. y calle 246, Santa Fe,Ciudad de La Habana, Cuba.Teléfono: (537) 209-7875ISSN 0138-8452RNPS 0485
Revista Cubana de Investigaciones Pesqueras es una publicación semestral iniciada en 1953 bajo el título deContribuciones. En 1974 cambió su título por Revista de Investigaciones Pesqueras y en 1978 por el actual. Es editadapor el Centro de Investigaciones Pesqueras perteneciente al Ministerio de la Industria Alimentaria. Los artículos queaquí aparecen son de investigación científica en el campo de las ciencias marinas, tecnológicas, cultivo de organismosacuáticos y medio ambiente. La Revista Cubana de Investigaciones Pesqueras está certificada por el CITMA comoPublicación Seriada Científico-Tecnológica con el código: 1243111. Se encuentra registrada en el Catálogo Nacionalde Publicaciones Seriadas de Cuba. Está indizada en la base de datos Aquatic Science and Fisheries Abstract (ASFA)de la FAO, y está colocada en el repositorio digital OCEANDOCS, con acceso abierto a texto completo. La impresión deeste número corresponde al vol. 29, No. 1, enero-diciembre de 2012.
Comité Científico
M. Sc. Eduardo Raúl Flores Gutiérrez (Editor científico)
Dra. C. María Estela de León GonzálezDra. C. Raquel Silveira Coffigny
Dra. C. J. Susana Alvarez CapoteDr. C. Gustavo Arencibia CarballoDr. C. Barbarito Jaime CeballosM. Sc. Norberto Capetillo Piñar
Revista Cubana de Investigaciones PesquerasISSN 0138-8452 RNPS 0485
Director General
Dr. C. Rafael A. Tizol Correa
Enero-diciembre, 2012, vol. 29, No. 1

ACUICULTURA
Evaluación del concentrado de pectina de cítrico comoaglutinante en el alimento para camarón / 1
Iliana Fraga Castro y Barabarito Jaime Ceballo
Alternativa de biofertilizante como medio de cultivopara el crecimiento poblacional de dos microalgasmarinas empleadas en la camaronicultura / 6
Missael Guerra Aznay, Lourdes Pérez Jar, Sylvia LealLorenzo, Barabarito Jaime Ceballos, Redney JiménezCabrera, Sunney Pérez Díaz y Jorge Bobadilla González
Efecto de diferentes niveles de alimentación paraClarias gariepinus en preceba / 12
José Llanes Iglesias y José Toledo Pérez
Crecimiento poblacional del rotifero nativo Brachionussp. “Cayman”, al evaluar diferentes microalgas comoalimento / 18
Rosario Cisneros
Tecnología cubana de cultivo de tilapia y carpa comúnimplementada en la región sur de Namibia / 24
Noris Millares Dorado y Mabel Navarro Jam
MEDIO AMBIENTE
El efecto de la temporada ciclónica 2005 sobre elfitoplancton de la Bahía de Cienfuegos, Cuba / 30
Ángel Moreira, Augusto Comas, Mabel Seisdedoy Sinai Barcia
Características físico-químicas de las aguas y delfitoplancton en zonas de baño de la Bahía deCienfuegos, Cuba (2008-2009) / 38
Mabel Seisdedo, Ángel R. Moreira y Gustavo Arencibia
PRODUCTOS PESQUEROS
Composición y calidad de la fracción lipídica de losensilajes de residuos de tilapias / 44
José E. Llanes Iglesias, José Toledo Pérez, Lourdes SavónValdés y Odilia Gutiérrez Borroto
Hidrocarburos aromáticos policíclicos (HAPs) en mejillónverde (Perna viridis) de la Bahía de Cienfuegos / 49
Luisa Vega Bolaños, Roberto Castelo Baez, José A. AriasVerdes y Ramón Sersa Espinosa
ECOLOGÍA
Éxito reproductivo de tortugas marinas en playa ElGuanal (2008-2010), Isla de la Juventud / 54
Yosvani Medina Cruz, Félix G. Moncada Gavilán y GonzaloNodarse Andreu
Composición y abundancia de dinoflageladosepibentónicos tecados potencialmente tóxicos de laEnsenada Guajimico, región Centro-Sur de Cuba / 59
Rosely Peraza Escarrá y Ángel R. Moreira González
Dispersión de larvas de langosta en el Caribe basadaen simulaciones numéricas / 67
Alina Rita Gutiérrez Delgado, Julio Baisre Álvarezy Amaury Álvarez Cruz
NOTA CIENTÍFICA
Capturado el camarón tigre Penaeus monodon(Fabricius, 1798) en las costas de Cuba / 77
INSTRUCCIONES PARA LOS AUTORES / 81
OFERTA DE CURSOS DE POSTGRADO 2013 / 84
Tabla de contenidos
Enero-diciembre, 2012, vol. 29, No. 1

1
Revista Cubana de Investigaciones PesquerasEnero-junio, 2012, vol. 29, NO. 1, ISSN 0138-8452, pp. 1-5
INTRODUCCIÓN
La industria citrícola comenzó a desarrollarse en Cubadesde el año 1968 a lo largo de todo el territorio en diezprovincias, alcanzando producciones de hasta 500 000 tde cítricos al año (Pino, 2008). Posteriormente se crearonlos combinados industriales para la extracción de jugos,aceites esenciales y producción de conservas.
Evaluación del concentrado de pectina de cítrico como aglutinanteen el alimento para camarón
Binding effect of citrus pectin concentrate in shrimp food
Iliana Fraga Castro y Barbarito Jaime Ceballos Centro de Investigaciones Pesqueras. Ave. 5ta. y calle 246, Santa Fe, Playa,
La Habana, Cuba, CP 19100, E-mail: [email protected],[email protected]
RESUMEN
Con el objetivo de evaluar el efecto aglutinante del concentrado de pectina de cítrico (CPC) en el alimento paracamarón, se desarrolló un diseño experimental completamente aleatorizado con cuatro tratamientos y tres réplicaspor cada uno, en condiciones controladas. Las dietas se elaboraron con 0; 2; 5 y 7 % de CPC. Las pruebas deestabilidad del alimento se realizaron introduciendo 1 g de cada alimento, en bolsas de 0,1 mm de luz de mallapreviamente pesada y se colocaron en agua de mar (35 ups y 27 ºC), durante 1; 12 y 24 h. Al final de cadaintervalo de tiempo, los alimentos se secaron y pesaron para calcular peso seco, pérdida de materia seca yabsorción de agua. Con el empleo del CPC, los alimentos alcanzaron una estabilidad superior al 80 % despuésde 24 h de inmersión en agua, superior a la lograda cuando no se incluyó el aglutinante. Al correlacionar lapérdida de materia seca de las dietas con los tiempos de exposición al agua, se alcanzaron coeficientes significativosde R2 = 0,944 2; R2 = 0,972 5; R2 = 0,911 8 y R2 = 0,851 4 respectivamente. El porcentaje de absorciónde agua del alimento varió en dependencia del tiempo de inmersión y fue similar entre tratamientos transcurridas12 h. A las 24 h, la dieta que incluyó 0 y 2 % de aglutinante, alcanzó valores de absorción significativamentemayores (p < 0,05) que el resto de las fórmulas.
Palabras clave: alimento, concentrado de pectina de cítrico, aglutinante, estabilidad en agua, pérdida de materiaseca.
ABSTRACT
In order to evaluate the binding effect of citrus pectin concentrate (CPC) in crustacean food, a completelyrandomized experimental design was developed, with four treatments and three replicates each, under controlledconditions. Diets were prepared with 0; 2; 5 and 7 % of CPC. Food stability tests were carried out in seawater(35 ups and 27 º C), where remained for intervals of 1; 12 and 24 h. At the end of each period, dry weight, drymatter loss and water absorption was calculated. Food stability achieved with CPC, was over 80 % after 24 himmersion in sea water, superior to that achieved when binder was not included. Significant correlation coefficientwere achieved between loss dry matter of diets and water exposure times, R2 = 0,944 2; R2 = 0,972 5;R2 = 0,911 8 y R2 = 0,851 4 respectively. The percentage of food water absorption varied depending on theimmersion time and was similar between treatments within 12 h. At 24 h, diets without CPC and with 2 %binder, reached significantly higher water absorption (p < 0,05) than other.
Keywords: food, citrus pectin concentrate, binding, water stability, loss of dry matter.
En el proceso industrial de los cítricos, se generansubproductos como la harina de cáscara y semilla, hollejopara pectina y concentrado de pectina que en totalrepresentan entre 50 y 60 % de la producción total decítricos (Gohl, 1978).
Estos desechos han sido evaluados para la alimentaciónde diferentes especies con buenos resultados nutricionalesy económicos (García et al., 1992; Moreno et al., 2000;González et al., 2007; Sotto et al., 2009).

2
La pectina es una mezcla compleja de polisacáridosque constituye la pared celular de los vegetales y suprincipal componente es el ácido galacturónico unido porenlaces glicosídicos. Su alto poder gelificante y emul-sionante, lo convierte en un insumo esencial en laindustria alimentaria (Mesbahi et al., 2005). El concen-trado de pectina de cítricos (CPC) se obtiene a partir dela cáscara, membranas y vesículas del jugo, sometidosa un proceso de acidificación y ajuste de pH, parasolubilizar la pectina. La misma se presenta como unlíquido viscoso de color verdoso, de bajo costo (8 USD/kg),que se almacena en refrigeración para su conservación. Elsecado se realiza a temperaturas elevadas y se obtenieneun polvo dorado que se comercializa entre 35 y 50 USD/kg(Rojas et al., 2009).
La estabilidad del alimento en el agua es un factor atener en cuenta en las especies acuáticas y es crítico enaquellas con hábitos bentónicos, como los camaronespor ser comedores lentos (Galindo, 2007) y para suconsumo eficientemente es necesario que mantenga suintegridad en el agua y reducir la contaminación ambiental(Akiyama & Chang, 1993). Con la inclusión de unaglutinante en la fórmula, se logra la mayor estabilidaddel granulado (Compabadal & Celis, 1999). Aunqueexisten otros métodos que aseguran la hidroestabilidad,como el molinado fino de los ingredientes y temperaturasmayores de 90 ºC (Molina-Poveda, 2008), se hacomprobado que el costo puede elevarse expo-nencialmente, requiriendo 2,3 kwat/t para obtener untamaño de partícula de 272 μm (Obaldo et al., 1998;Obaldo & Tacon, 2001). De ahí que los aglutinantes sonrequeridos para reducir el costo de fabricación debalanceados al retardar la desintegración del granulado,reducir la lixiviación de nutrientes, disminuir la producciónde finos durante el embalaje y transporte y minimizar elriesgo de contaminación ambiental (Compabadal & Celis,1999).
Esta investigación se realizó con el objetivo de conocerel efecto aglutinante del concentrado de pectina de cítricoen la elaboración del alimento para camarón.
MATERIALES Y MÉTODOS
Un diseño experimental completamente aleatorizado, concuatro tratamientos y tres réplicas por cada uno, sedesarrolló en el Laboratorio de Nutrición del Centro deInvestigaciones Pesqueras, del Municipio Playa, en laprovincia La Habana, Cuba. Para evaluar el efecto aglutinantedel concentrado de pectina de cítrico en el alimento paracamarón, se elaboraron dietas con 0; 2; 5 y 7 % de CPClíquido (TABLA 1), producido en la Industria Procesadora deCítricos de Jagüey Grande, provincia Matanzas, siguiendola metodología descrita por Galindo et al. (2009).
Las pruebas de estabilidad de los alimentos, se realizaronintroduciendo 1 g de alimento de cada fórmula, en bolsasrectangulares de malla (4 x 3 cm) de 0,1 mm de luz demalla, previamente pesadas en una balanza Sartorio de0,001 g de precisión. Las bolsas suspendidas de una varillametálica, se sumergieron en cubetas plásticas con 40 L deagua de mar (35 ups y 27 ºC) con aireación continua, dondepermanecieron durante diferentes intervalos de tiempo(1; 12 y 24 h). Al final de cada intervalo, las bolsas sepesaron y pusieron a secar en una estufa con recirculaciónde aire forzado, a 60 ºC durante 6 h. Posteriormente sepesaron para determinar el peso seco final.
La pérdida de materia seca (PMS) de los alimentospara los diferentes intervalos de tiempo y niveles de CPCse determinó por la fórmula:
PMSi = [1 – (PSAi/PSAf)] x 100)
Donde:
PMSi: porcentaje de pérdida de materia seca en eltiempo i
PSAi: peso seco del alimento (g) después desumergirlo en el agua en el tiempo i
PSAf: peso seco del alimento antes de sumergirlo enel agua (g)
La absorción de agua de cada dieta (AAi) durante lainmersión en los intervalos de tiempo i se calculó por lafórmula:
AAi = [(PHAi – PSAf)/PSAf] x 100
Donde:
PHAi: peso del alimento después de sumergido en elagua de mar durante el tiempo i
Se realizaron análisis de regresión y correlación entretiempos de inmersión y pérdida de materia seca y absorciónde agua de los alimentos para los diferentes niveles deinclusión de CPC, ajustándolos a una ecuación polinomialcuadrática (Shearer, 2000; Hernández-Llamas, 2009).
TABLA 1. Composición porcentual (g/100 g de dieta) delos alimentos ensayados en el experimento con
diferentes niveles de concentrado de pectina de cítrico(CPC) como aglutinante
Ingredientes Patrón CP CPC CPC 2 % 5 % 7 %
Harina de pescado 30 30 30 30
Harina de carne 10 10 10 10
Harina de soya 24 24 24 24
Trigo entero molido 14 14 14 14
Levadura torula 5 5 5 5

3
RESULTADOS Y DISCUSIÓN
Durante la primera hora de inmersión en agua de mar,todos los alimentos presentaron porcentajes de pérdidade materia seca (PMSi) estadísticamente similares(p > 0,05) con valores entre 4,5 y 6 %, siendo mayoren el granulado Patrón que no incluyó CPC (Fig. 1).Resultados similares alcanzaron Carvalho & Nunes (2006)al evaluar una dieta comercial para Litopenaeus vannamei.Cruz-Suárez et al. (2002) al evaluar diferentes alimentoscomerciales alcanzaron pérdidas de materia seca entre1,8 y 6,3 luego de permanecer 1 h sumergido en agua demar con 34-35 UPS y consideraron como normal, del 5 %en balanceados comerciales. Otros autores han consignadopérdidas del 12 % a las 4 h de sumergir el granulado(Carvahlo & Nunes, 2006; Smith et al., 2002).
Los alimentos con 5 y 7 % de CPC no presentarondiferencias significativas en PMSi para los diferentesintervalos de tiempo (p < 0,05) con valores entre 4-5 %,5-6 % y 9,8-10 %, al permanecer sumergidos durante1; 2 y 12 h respectivamente. La mayor PMSi se alcanzóa las 24 h de inmersión con valores de 12,5 y 19 %,siendo significativamente menores en las dietas con5 y 7 % de CPC.
Estos resultados demostraron el efecto aglutinantedel CPC líquido en los alimentos para crustáceos y laconveniencia de su empleo a un nivel de inclusión del 5 %,ya que posibilita garantizar el tiempo necesario para quelos crustáceos lo ingieran, sin pérdidas importantes denutrientes, durante las dos primeras horas de añadido.
Al correlacionar la pérdida de materia seca de losalimentos, Patrón (0 %), CPC 2 %, CPC 5 % y CPC 7 %,con los tiempos de exposición al agua (1; 12 y 24 h), seobtuvieron coeficientes significativos de R2 = 0,976 93;R2 = 0,923 2; R2 = 0,911 8 y R2 = 0,931 8,respectivamente (Fig. 1). Los resultados de este estudiodemostraron que el CPC incluido entre 5 y 7 % reducesignificativamente las PMSi durante períodos deinmersión prolongados (> 12 h). García et al. (1992), alemplear el CPC al 5 % de inclusión en la dieta paraposlarvas de camarón, obtuvieron crecimiento ysupervivencias superiores a los obtenidos con goma guar.
Premezcla devitaminas y minerales 5 5 5 5
Aceite de pescado 2 2 2 2
Colesterol 1 1 1 1
Lecitina de soya 2 2 2 2
CPC 0 2 5 7
Alfa celulosa 7 5 2 0
Durante la primera hora, el porcentaje de absorciónde agua (AAi) del alimento varió significativamente(p < 0,05) entre los balanceados, siendo menor para ladieta que incluyó 7 % de CPC (AAi = 50 %), seguida dela dieta con 5 % de CPC (AAi = 55 %) y varió endependencia del tiempo de exposición al agua de mar.A partir de las 12 h la AAi alcanzó valores superiores al80 % en todos los tratamientos (Fig. 2). Carvalho & Nunes(2006), observaron un porcentaje de absorción del 61 %durante la primera hora de sumergido el alimento, similaral obtenido en este experimento con la dieta Patrón (0 %CPC) y la que incluyó 2 % del aglutinante.
El análisis de regresión mostró una relación signifi-cativa entre el porcentaje de absorción de agua de lasdietas con diferentes niveles de inclusión de CPC, en losdiferentes intervalos de tiempo de inmersión en aguade mar con valores de R2 = 0,907 4; R2 = 0,901 6;R2 = 0,908 7; R2 = 0,917 4 (Fig. 2). Este indicador esde suma importancia debido a las implicaciones que tieneen la pérdida de nutrientes por lixiviación. Según observa-ciones de Smith et al. (2002), los camarones permanecenalimentándose mientras el balanceado mantiene suintegridad hasta 4 h después de añadido.
Fig. 1 Porcentaje de pérdida de materia seca de dietaselaboradas con 2; 5 y 7 % de inclusión de CPC
sumergidas en agua de mar durante 1; 12 y 24 h
0 5 10 15 20 25 30Tiempo (h)
Patrón CPC2% ----- x CPC7%CPC5%
y (Patrón) = –0,0171x2 + 0,9499x + 6,0672, R2 = 0,97693y (CPC2%) = –0,0233x2 + 0,9576x + 4,8657, R2 = 0,92320y (CPC5%) = –0,0198x2 + 0,8353x + 4,3346, R2 = 0,91182y (CPC7%) = –0,0203x2 + 0,8542x + 3,666, R2 = 0,93182
12
8
642
0
10
18
16
14
20
PMSi a
ab
c
Fig. 2 Variación de la absorción de agua de los alimentoscon diferentes niveles de inclusión de CPC sumergidas en
agua de mar durante 1; 12 y 24 h
Tiempo de inmersión en agua (h)
Patrón CPC2% CPC7%CPC5%
100
80
70
60
50
40
90
y (Patrón) = –0,0962x2 + 3,7958x + 61,3, R2 = 0,90740y (CPC2%) = –0,112x2 + 4,365x + 55,747, R2 = 0,90167y (CPC5%) = –0,1163x2 + 4,6024x +50,514, R2 = 0,90871y (CPC7%) = –0,1126x2 + 4,5553x + 46,557, R2 = 0,91745
0 12 24
AA
i (%
)
a
cb
c

4
CONCLUSIONES Y RECOMENDACIONES
Los resultados indican la conveniencia de emplearniveles de inclusión de CPC de 5-7 % para garantizarun margen amplio en la estabilidad del alimentocuando se pone en contacto con el agua. Las dietasalcanzan una estabilidad entre 94 y 96 % durante laprimera hora de inmersión y superior al 80 % a las24 h. La pérdida de materia seca de los alimentos enrelación con los tiempos de exposición al agua,mostraron coeficientes significativos de R2 = 0,976 9;R2 = 0,923 2; R2 = 0,911 8 y R2 = 0,931 8,respectivamente.
Los alimentos con 5 y 7 % de concentrado de pectinade cítrico presentaron un porcentaje de absorción deagua menor de 50 y 55 % durante la primera hora.
REFERENCIAS
Akiyama, D. & Chiang, N. (1993). Requerimientosnutricionales del camarón y manejo del alimento. EnL . E. Cruz-Suárez, D. Ricque Marie & R. Mendoza(Eds.), Memorias Primer Simposium Internacional deNutrición y Tecnología de Alimentos Acuáticos.Asociación Americana de la Soya y Facultad deCiencias Biológicas, Universidad Autónoma de NuevoLeón, México, pp. 479-491.
Carvalho, E. A. & Nunes, A. J. P. (2006). Effect offeeding frequency on feed leaching loss and grow-out patterns of the white shrimp Litopenaeusvannamei feed under a diurnal feeding regime in pondenclosures. Aquaculture, 252, 494-502.
Campabadal, C. & Celis, A. (1999). Factores que afectanla calidad de los alimentos acuícolas. En: L. E. Cruz-Suárez, D., Ricque Marie & R. Mendoza (Eds.),Avances en Nutrición Acuícola III. Memorias delTercer Simposium Internacional de Nutrición Acuícola,11-13 de noviembre de 1996. Monterrey, NuevoLeón, México, pp. 532-540.
Cruz-Suárez, L. E., Ricque-Marie, D., Tapia-Salazar, M.,Marín-Zaldivar, L. F., Guajardo-Barbosa et al. (2002).Historia y estatus actual de la digestibilidad y algunascaracterísticas físico-químicas de los alimentoscomerciales para camarón usados en México. En M.Salazar, G. Gaxiola & N. Simoes (Eds.), Avances enNutrición Acuícola VI. Memorias del VI SimposiumInternacional de Nutrición Acuícola, Cancún, QuintanaRoo, México, pp. 516-526.
Galindo, J. (2007). Aglutinantes. En T. García-Galano,H. Villarreal-Colmenares & J. L. Fenucci (Eds.), Manualde ingredientes proteicos y aditivos empleados en laformulación de alimentos balanceados para camarones
peneidos. FUDEM, ISBN: 978-987-1371-02-0,pp. 211-215.
Galindo, J., Jaime, B., Fraga, I. & Alvarez, J. S. (2009).Empleo de subproductos de la caña de azúcar parala alimentación del camarón del Caribe. Disponibleen: REDVET. Rev.electrón.vet. http://www.veterinaria.org/revistas/redvet – http://revista.veterinaria.org, vol. 10, No. 7, julio/2009 – http://www.veterinaria.org/revistas/redvet/n070709.html
García, T., Jaime, B. & García, V. (1992). Crecimiento depostlarvas de camarón blanco Penaeus schmitti,utilizando diferentes aglutinantes. Rev. Inv. Mar., 13 (1),87-91.
Göhl, B. (1978). Los subproductos de los cítricos parala alimentación del ganado. Revista Mundial deZootecnia, 12, 30-40.
González, A., Ojeda, F. & Mora, M. (2007). Constituciónfísica del bagazo de naranja (Citrus sinensis var.Valencia) y características organolépticas y enrique-cimiento proteico de este material sometido aamonificación. En M. F. Díaz, R. S. Herrera &T. Ruiz (Eds.), II Congreso de Producción AnimalTropical. San José de las Lajas, pp. 76-77.
Hernández-Llama, A. (2009). Conventional andalternative dose-response model to estimate nutrientrequirements of aquaculture species. Aquaculture,292, 207-213.
Mesbahi, G., Jamalian, J. & Farahnaky, A. (2005). Acomparative study on functional properties of beetand citrus pectins in food systems. Food Hydrocolloid,19 (4), 731-738.
Molina-Poveda, C. (2008). Alimento Artificial. En C.Molina-Poveda & H. Villarreal-Colmenares (Eds.),Estrategias de alimentación en la etapa de engordedel camarón. La Paz, B.C.S.; CIDNOR, S. A., CYTEDy PRONACA, pp. 31-87.
Moreno, M. J., Hernández, J. G., Rovero, R., Tablante,A. & Rangel, L. (2000). Alimentación de tilapia conraciones parciales de cáscara de naranja. Ciencia yTecnología Alimentaria, 3 (001), 29-33.
Obaldo, L., Dominy, W. C., Terpstra, J., Cody, J. &Behnke, K. C. (1998). The effect of ingredientparticle size on shrimp feed. Journal of AppliedAquaculture, 8, 55-66.
Obaldo, L. & Tacon, A. (2001). Manufacturing differentdiet sizes and its effect on pellet water stability andgrowth of three size classes of pacific white shrimpLitopenaeus vannamei. Journal of AppliedAquaculture, 11, 57-66.
Pino, B. N. (2008). Estudio de la utilización de hollejo decítrico para la ceba de toros en la Empresa CitrícolaVictoria de Girón. Tesis presentada en opción alTítulo de Máster en Pastos y Forrajes. Universidadde Matanzas Camilo Cienfuegos, Cuba, 94 pp.

5
Rojas, J., Jennifer, P., Perera, V. & Rstashenko, E. (2009).Obtención de aceites esenciales y pectinas a partirde subproductos de jugos cítricos. Revista de laFacultad de Química Farmacéutica, 16 (1), 110-115.
Shearer, K. (2000). Experimental design, statisticalanalysis and modeling of dietary nutrient requirementstudies for fish: a critical review. Aquacult. Nutr.,11, 263-272.
Smith, D., Burford, M., Tabrett, S., Irvin, S. & Ward, L.(2002). The effect of feeding frequency on waterquality and growth of the black tiger shrimp (Penaeusmonodon). Aquaculture, 207, 125-136.
Sotto, V., Brito, M., López, B. Pérez, E. & Velásquez, F.(2009). Empleo de la harina de cítricos en la alimentaciónde cerdas durante la gestación y la lactancia. RevistaComputadorizada de Producción Porcina, 6 (1), 5 pp.

Revista Cubana de Investigaciones PesquerasEnero-diciembre, 2012, vol. 29, NO. 1, ISSN 0138-8452, pp.
6
Alternativa de biofertilizante como medio de cultivo para elcrecimiento poblacional de dos microalgas marinas empleadas en la
camaronicultura
Alternative bio-fertilizer as culture medium for the population growth of two marinemicroalgae used in shrimp farming
Missael Guerra Aznay,1 Lourdes Pérez Jar,1 Sylvia Leal Lorenzo,2 Bárbarito Jaime Ceballos,1 Redney Jiménez Cabrera,1
Sunney Pérez Díaz1 y Jorge Bobadilla González1
1 Centro de Investigaciones Pesqueras. Ave. 5ta. y calle 246, Santa Fe, Playa, La Habana,Cuba, CP 19100, Teléfono: (537) 209-7852, E-mail:[email protected]
2 Centro de Investigaciones Marinas. Calle 16 No. 114 entre 1ra. y 3ra., Miramar, La Habana, Cuba, CP 11300
RESUMEN
Con el propósito de evaluar el humus de lombriz roja californiana Eisenia foetida como biofertilizante para elcrecimiento poblacional de dos microalgas marinas, fueron cultivadas Tetraselmis tetrathele y Chaetoceros muellerien agua de mar enriquecida con cuatro diluciones (50; 200; 350 y 500 mL) de extracto líquido de humus. Laespecie T. tetrathele, cultivada en la dilución de 200 mL, no mostró diferencias con el patrón medio f/2 Guillardpara los parámetros: concentración celular, velocidad de crecimiento, tiempo de duplicación y productividad.C. muelleri cultivado en el mismo biofertilizante a las diluciones antes descritas, mostró un comportamientoproductivo inferior al patrón, por lo que fueron evaluadas dos nuevas diluciones (550 y 650 mL). Lasconcentraciones celulares alcanzadas con 550 y 650 mL del fertilizante orgánico y el patrón, no mostrarondiferencias (p > 0,05) hasta el tercer día del cultivo, sin embargo, al evaluar el indicador productividad seobtuvieron los mejores valores con 550 mL del extracto líquido de humus. Por los resultados obtenidos en esteestudio con el biofertilizante y su bajo costo de producción se concluyó que resulta una alternativa viable parala producción de las microalgas C. muelleri y T. tetrathele hasta volúmenes de 2 L.
Palabras clave: humus, biofertilizante, Chaetoceros muelleri, Tetraselmis tetrathele.
ABSTRACT
To evaluate the humus of of red worm californian Eisenia foetida were cultured microalgaes Tetraselmis tetratheleand Chaetoceros muelleri in seawater enriched with four dilutions (50; 200; 350 and 500 mL) of liquid extractof humus of red worm californian Eisenia foetida. The T. tetrathele specie, grown at a dilution of 200 mL,showed no differences with the standard f/2 Guillard for the parameters: cell concentration, growth rate, doublingtime and productivity. C. muelleri grown in the same biofertilizer to dilutions as described above, showed alower productive performance pattern, so that two further dilutions were evaluated (550 and 650 mL). The cellconcentrations reached with 550 and 650 mL of organic fertilizer and the pattern did not differ (p > 0,05) untilthe third day of culture, however, in assessing the productivity indicator the best values were obtained with 550 mLof liquid extract humus. From the results obtained in this study with the alternative means and low cost ofproduction is concluded that it is a viable alternative for the production of microalgaes C. muelleri andT. tetrathele up to volumes of 2 L.
Keywords: humus, biofertilizer, Chaetoceros muelleri, Tetraselmis tetrathele.
INTRODUCCIÓN
Las microalgas son importantes en la acuicultura, ya queconstituyen el primer alimento para las fases tempranasde desarrollo de casi todos los organismos cultivados,
6-11
principalmente moluscos, peces y crustáceos, así comopara algunas especies intermedias como son copépodos,rotíferos y artemias, siendo altamente nutritivas y fácilesde ingerir debido al tamaño que poseen (Ritar et al., 2004).
Dos de las especies de microalgas más comúnmenteutilizadas en el maricultivo son Chaetoceros sp. y

7
Tetraselmis sp. por su excelente aceptación y elevadocontenido proteico y de ácidos grasos poliinsaturados(Brown, 1991; Piña et al., 2006), con las cuales a suvez, se han obtenido altas tasas de sobrevivencia en laslarvas de camarón (Valenzuela et al., 1999; Pacheco-Vega,2003; López-Elías, et al., 2005). Por este motivo, en todoslos laboratorios productores de larvas de camarón seencuentra una sección especial, diseñada y operada conel fin exclusivo de producir una o varias especies demicroalgas a nivel masivo. El mantenimiento de estasinstalaciones unido al empleo de variantes del medio f/2(Guillard, 1975) que contiene sales nutritivas (reactivospuros para laboratorios) para el cultivo de las algas, hacenque se incrementen notablemente los costos de producciónen la camaronicultura (Martínez-Córdova, 1999; López-Elías,2002).
Con el propósito de reducir los costos de operaciónpor concepto de reactivos en las instalaciones queproducen fitoplancton, diversos autores han evaluado elcrecimiento de microalgas en medios de cultivoalternativos preparados con fertilizantes agrícolas(López-Elías & Voltolina, 1993; Nieves et al., 1994;Valenzuela-Espinoza et al., 1999; Simental-Trinidad &Sánchez-Saavedra, 2003). Otros han investigado cómovaría la composición bioquímica de diferentes especiesde microalgas al cultivarlas en medios preparados confertilizantes orgánicos (Simental-Trinidad et al., 2001).Estos estudios mencionan que la cantidad y calidad de labiomasa producida no se afecta por el uso de estas fuentesde nutrientes.
Recientemente Ruiz (2009), recomendó la aplicaciónen estanques de engorde, antes de la siembra de poslarvasde camarón, un fertilizante orgánico llamado vermicompostfabricado a base de líquido de humus de lombriz rojacaliforniana Eisenia foetida (té de humus), cuyo elevadocontenido y producción de hormonas estimulan el creci-miento y funciones vitales de microalgas, fundamental-mente diatomeas.
El humus de lombriz no es más que los excrementosde las mismas que transforman los residuos orgánicoscomo restos de cosechas, excrementos de animales decría (vacas, cerdos, gallinas, entre otros), restos decocina o papelería en desuso. Entre los componentesfundamentales del humus de lombriz están el nitrógeno,fósforo, potasio y el calcio. Esta composición de losnutrientes es altamente variable, dependiendo de losfactores bióticos y abióticos (Caro, 2004).
En Cuba los reactivos puros de laboratorio requeridospara el cultivo progresivo de las microalgas marinas sonen su gran mayoría de importación y de difícil adquisición,por lo que teniendo en cuenta los antecedentes sobre lautilización de fertilizantes orgánicos en la acuicultura, elpropósito del presente trabajo fue la evaluación de unextracto líquido del humus de lombriz roja californianaEisenia foetida como biofertilizante, para determinar su
efecto en el crecimiento poblacional de células de lasmicroalgas marinas Chaetoceros muelleri y Tetraselmistetrathele.
MATERIALES Y MÉTODOS
El presente estudio se desarrolló en el Centro Genéticode Camarón, Municipio Mariel, perteneciente al Centrode Investigaciones Pesqueras (CIP), MINAL. El diseñoconsistió en la obtención de un extracto líquido a partirde humus de lombriz roja californiana Eisenia foetidacultivada con vacaza, para probarlo como medio decultivo de dos microalgas marinas.
Obtención del extracto líquido de humusde lombriz
La obtención del extracto se hizo a partir de unamodificación de la metodología de preparación debiofertilizantes líquidos a partir de humus de lombriz,según lo describen Casco & Iglesias (2005). Se tomó unfrasco de 1 L de capacidad que contenía agua de marpasada por filtros de: 20; 10; 5; 1 y 0,32 µm, yposteriormente por lámpara UV. Una vez pesado el humusde lombriz se añadió al frasco con agua de mar filtrada,se mezcló vigorosamente por unos segundos. Se utilizóla proporción de: 50 g de humus por cada litro de aguade mar.
El frasco con la mezcla se colocó en una autoclavecon capacidad de 3 L durante 1½ h a 120 °C y 1,5 ATM,para su esterilización (las bacterias contaminan losinóculos de microalgas impidiendo su crecimiento celular)y promover la extracción de las sales nutrientes contenidasen el humus. Terminado este proceso, se dejó reposar lamezcla durante 24 h para que sedimentara el materialsólido. Se extrajo el frasco de la autoclave y con cuidadose separó el extracto filtrándolo por una malla de 100 µmLa preparación del medio de cultivo Guillard f/2 se realizósegún lo descrito por Guillard (1975).
Preparación de los inóculosde las dos microalgas
Las cepas de las microalgas utilizadas, Chaetocerosmuelleri y Tetraselmis tetrathele, provinieron del algariodel Centro de Investigaciones Pesqueras, donde fueronmantenidas con medio Guillard f/2 según está descritopor Guillard (1975).
Los experimentos de cultivo de microalgas sedesarrollaron en el algario del Centro Genético de Camarónen Mariel, ubicado en un local cerrado con iluminaciónfluorescente y temperatura controlada (2000 Lux y 23 °C

8
respectivamente), donde los frascos fueron mantenidos conaireación constante.
Se prepararon previamente los inóculos de cadamicroalga a evaluar ya adaptadas a las condiciones delalgario, las mismas fueron utilizadas cuando alcanzaron lafase de crecimiento exponencial (2-3 días). Se les realizóconteo a cada frasco preparado con el medio de cultivocorrespondiente (Guillard f/2 o el extracto líquido de humus)para calcular la cantidad de mililitros que debían emplearsepara el inicio del ensayo experimental. La concentracióncelular inicial fue similar en todos los frascos experimen-tales. Fueron empleados para los conteos un microscopiobiológico y una cámara de Neubauer (hematocitómetrode 0,1 mm de profundidad).
Ensayo experimental. Cultivo de microalgas
Para los cultivos la temperatura se mantuvo controladaen 24 °C, la salinidad del agua se ajustó a 35 ups y lailuminación fue constante proveniente de cuatro lámparasfluorescentes de luz blanca fría de 40 W, en estantespreparados para el mantenimiento y crecimiento progre-sivo de microalgas.
Se emplearon cuatro diluciones del extracto líquidode humus (EH50, EH200, EH350, y EH500 mL) paraevaluar el crecimiento celular de las microalgas en estudio(C. muelleri y T. tetrathele). Como patrón en amboscultivos monoalgales fue utilizado el medio Guillard f/2(MG). Para los ensayos experimentales se usaronrecipientes de cristal transparente de 2 L de capacidad.Se trabajó con tres réplicas por cada tratamiento. Losconteos de crecimiento poblacional se realizaron cada24 h para determinar posteriormente los indicadorespoblacionales velocidad de crecimiento (K), tasa deproducción diaria (PD) y tiempo de duplicación (TD).
En el caso específico de C. muelleri cultivado conextracto líquido de humus, se añadió 1 mL/L deNa2SiO39H2O según propuesta de Guillard (1975) parael desarrollo de las diatomeas.
Se realizó un segundo bioensayo para evaluar otrasdiluciones del extracto líquido de humus en caso de queno se alcanzaran concentraciones de células cercanasal patrón. Este estudio tuvo lugar con la microalgaC. muelleri, por lo cual se aplicó el mismo diseñoexperimental probando dos nuevas diluciones (EH550 yEH650 mL del extracto).
Todos los datos fueron analizados estadísticamentemediante el Programa SigmaStat 3.5.2.
RESULTADOS Y DISCUSIÓN
El desarrollo eficiente de un cultivo microalgal en ellaboratorio depende dentro de otros factores, de las
condiciones físico-químicas del medio en que sedesarrollan, al igual que en el medio natural (Leal &Bonaechea, 1994). Las variables físico-químicas en esteestudio se mantuvieron controladas dentro de lo reportadocomo óptimos para el cultivo de estas especies, lo cualfavoreció que las dos microalgas, bajo estas condiciones,crecieran dentro del intervalo esperado para su especieen número de células y similar al reportado por otrosautores (Pacheco & Sánchez-Saavedra, 2003; López-Elíaset al., 2008).
Al analizar el comportamiento de las curvas decrecimiento exponencial de cada tratamiento, en cadamicroalga evaluada, se sugiere que la cinética algal pudohaber estado influenciada por la composición del mediode cultivo. En el primer bioensayo realizado con ladiatomea C. muelleri, la baja densidad celular obtenidacon las cuatro diluciones del extracto líquido de humus(EH50, EH200, EH350 y EH500 mL), repercutió en laimplementación de un nuevo ensayo con dos dilucionesmás concentradas (EH550 y EH650 mL), lográndose conlas mismas un crecimiento celular que no difiere (p > 0,05)del patrón hasta el tercer día de cultivo, superándose los600 x 104 de cel.mL-1 (Fig. 1), valores similares a los quese obtienen a nivel comercial en la Empresa de Desove deCamarones YAGUACAM, Cienfuegos.
Las altas concentraciones celulares alcanzadas enla fase exponencial de los cultivos de C. muelleri dondefueron evaluadas las diluciones EH550 y EH650 mL delbiofertilizante (Fig. 1), provocaron en el tercer día decultivo un salto hacia la fase de decaimiento sin pasarpor la fase estacionaria, como bien se observa en eltratamiento con medio f/2 Guillard. Este comportamientopudo estar inducido por un efecto sombra (densidad celularalta + ligera turbidez provocada por el extracto líquidode humus) que impidió la penetración homogénea de laluz, inhibiendo el proceso de fotosíntesis y acelerando lallegada del cultivo a la fase de muerte celular.
Los indicadores poblacionales evaluados, velocidadde crecimiento celular (K), producción diaria (PD) y tiempode duplicación (TD) que describen el comportamiento delos cultivos de C. muelleri (TABLA 1), muestran laexistencia de diferencias significativas (p < 0,05) entrelos tratamientos, lográndose los mejores resultados conla dilución EH550 mL.
Para T. tetrathele (Fig. 2) no se observarondiferencias significativas (p > 0,05) en las concen-traciones celulares de EH50 mL, EH200 mL conrespecto al patrón durante los tres primeros días delcultivo, tendencia que se mantuvo solamente para ladilución EH200 mL hasta el séptimo día de cultivodonde se lograron concentraciones celulares de173 ± 5,8 x 104 cel.mL-1 y de 175 ± 1,0 x 104 cel.mL-1
con Guillard. Igualmente no difirieron los parámetrospoblacionales (TABLA 1) entre ambos tratamientos (p > 0,05).

9
N
Fig. 1 Concentración celular acumulada de Chaetocerosmuelleri cultivada con el medio Guillard f/2 (patrón) yen dos diluciones concentradas del extracto de humus
de lombriz Eisenia foetida (EH550 y EH650 mL).N: No. de células x 104 cel.mL-1
Fig. 2 Concentración celular acumulada de Tetraselmistetrathele cultivada con el medio Guillard f/2 (MG) y endiferentes diluciones del extracto de humus de lombrizEisenia foetida (EH50, EH200, EH350 y EH500 mL).
N: No. de células x 104 cel.mL-1
0 1 2 3 4 5
Días
Patrón 550 mL 650 mL
TABLA 1. Valores promedios de la velocidad de crecimiento celular (K) acumulada, producción diaria (PD) y tiempode duplicación (TD) de las especies de microalgas cultivadas en el medio Guillard (MG) y en diferentes diluciones
del extracto líquido de humus de lombriz Eisenia foetida
Las otras dos diluciones evaluadas EH350 y EH500 mLmantuvieron prácticamente durante todo el tiempo una bajaconcentración celular. Este comportamiento se pudo atribuira un marcado efecto de autosombreado, pero esta vez
causado por la coloración del medio que impidió la penetra-ción y distribución homogénea de la luz, inhibiéndose así elproceso de fotosíntesis y por consiguiente el desarrollodel cultivo.
Estos resultados son similares a los reportados porGodínez (2000), al evaluar el crecimiento de las microalgasC. muelleri y T. suecica en doce extractos líquidos defertilizantes orgánicos, lográndose concentraciones celulares3,3 % superiores para la diatomea cultivada en el medioorgánico y en el caso de la flagelada valores que no difierena los del control. Este autor alcanzó concentraciones celularesinferiores a los obtenidos en este trabajo, lo cual puededeberse, entre otros factores, a la composición química delos biofertilizantes empleados. En este estudio las altasconcentraciones reportadas de nitrito (TABLA 2) al evaluar lacomposición química de la dilución EH550 mL del extractolíquido de humus, pudo haber favorecido el rápido crecimientode C. muelleri.
El comportamiento en el crecimiento celular de lasmicroalgas cuando el nitrógeno está disponible en excesoen cualquiera de las formas asimilables, fue observadopor García & Jiménez (1990), cuando evaluaron unamezcla de fertilizantes agrícolas para el cultivo deIsochrysis galbana, al usar tres concentraciones defertilizantes basados en urea, para dar valores de 60,75;27,0 y 6,75 mg/L de nitrógeno. Su tratamiento controlfue una modificación del medio f, con 6,75 mg/L de N.Estos autores reportaron que el medio que mejorresultados alcanzó fue el que contenía la mayor concen-tración de nitrógeno y que las dosis menores de esteelemento eran limitantes para el crecimiento de lamicroalga evaluada.
N
0 1 2 3 4 5 6 7
Días
EH(550 mL) EH(200 mL) EH(350 mL) EH(500 mL)MF
200
160
140
120
100
80
180
60
40
20
0
600
400
300
200
1000
500
700
Especies Parámetro MG EH (50 mL) EH (200 mL) EH (350 mL) EH (500 mL) EH (550 mL) EH (650 mL)
K 3,1a 3,0a 3,1a 2,5b 2,5b
Tetraselmis PD 80,6a 5,5b 80,9a 6,7b 32,8ctetrathele
TD 0,34a 0,37a 0,30a 0,42b 1,34c
Chaetoceros K 0,5a 0,84b 0,76cmuelleri
PD 41,24a 115,4b 82,3c
TD 4,74a 2,58b 2,28c

10
TABLA 2. Concentraciones de nitrito, amonio y fósforo presentes en el extracto líquido de humus de la lombrizEisenia foetida y el medio Guillard (f/2)
Especies Parámetro MG EH(50 mL) EH(200mL) EH(350 mL) EH(500 mL) EH(550 mL) EH(650 mL)
K 3,1a 3,0a 3,1a 2,5b 2,5b
Tetraselmis PD 80,6a 5,5b 80,9a 6,7b 32,8ctetrathele
TD 0,34a 0,37a 0,30a 0,42b 1,34c
Chaetoceros K 0,5a 0,84b 0,76cmuelleri
PD 41,24a 115,4b 82,3c
TD 4,74a 2,58b 2,28c
Cook & Clifford (1998) plantearon que es impor-tante mantener el balance correcto entre los nutrientesy que la cantidad relativa de N y P disponible en elagua ejerce una influencia sobre el tipo de alga quedomina. Cuando la tasa de estos dos elementos esmuy cercana (N:P < 5:1) favorece el crecimiento delos dinoflagelados y flagelados, cuando es alta15-20:1 es promovido el crecimiento de diatomeas, loque coincide con el trabajo de Lourenco et al. (1997)cuando probaron un fertilizante con proporción N:P de5,5:1, que modificaron adicionando nitrógeno paraobtener una relación similar al medio Conway (9,2:1)utilizado como control, encontrando un mayor crecimientocelular de C. gracilis en los dos medios con la razón N: Pmás elevada.
Otro factor reportado recientemente por Ruiz (2009),al trabajar con lombricompost como biofertilizante enestanques de tierra y que pudo incidir en los favorablesresultados obtenidos en cuanto a concentración celu-lar en las algas cultivadas con el extracto líquido dehumus, es el elevado porcentaje de ácidos húmicos,fúlvicos y de fitohormonas como el ácido indolacéticoy giberelico que posee este fertilizante orgánico y queestimulan el crecimiento y funciones vitales de lasalgas.
De acuerdo con los resultados obtenidos en estetrabajo, se concluyó que el extracto líquido de humusevaluado como medio de cultivo alternativo en el CentroGenético de Camarón en Mariel, fue fácilmente asimiladopor las dos especies de microalgas estudiadas, lográndoseconcentraciones celulares para el tercer día de cultivode C. muelleri con la dilución EH550 mL y para el séptimodía de cultivo de T. tetrathele con la dilución EH200 mLcomparables con las concentraciones alcanzadas con eltratamiento control.
REFERENCIAS
Brown, M. R. (1991). The amino-acid and sugarcomposition of 16 species of microalgae used inmariculture. J. Exp. Mar. Biol. Ecol., 145, 79-99.
Caro, I. (2004). Caracterización de algunos parámetrosquímico-físicos del Liplant, humus líquido obtenido apartir del vermicompost de estiércol vacuno. Tesispresentada en opción al grado académico de Másteren Química aplicada a la Agricultura, UNAH, LaHabana, Cuba, p. 90.
Casco, C. & Iglesias, M. C. (2005). Producción debiofertilizantes líquidos a base de lombricompuesto.Comunicaciones Científicas y Tecnológicas.Universidad Nacional del Nordeste, Argentina.Resumen, A-063, 4 pp.
Cook, C. I. & Clifford, H. C. (1998). Fertilization of shrimpponds and nursery tanks. Aquaculture Magazine,24 (3), 52-62.
García-Rodríguez, E. & Jiménez, S. (1999). The use ofsome agricultural fertilizer for the mass productionof marine algae. Aquaculture, 36, 245-256.
Godínez, D. E. (2000). Evaluación de dos fertilizantesorgánicos como medios alternativos para laproducción de Tetraselmis suecica y Chaetocerosmuelleri. Tesis de Maestría. Postgrado Interinsti-tucional de Ciencias Pecuarias. Universidad deColima, Tecomán, México, 65 pp.
Guillard, R. R. L. (1975). Culture of phytoplankton forfeeding marine invertebrates. In L. Smith & M. H.Chanley (Eds.), Culture marine invertebrate animals,New York, pp. 29-59.

11
Leal, S. & Bonaechea, I. (1994). Concentración óptimade nutrientes de tres especies de microalgas marinasen cultivo. Rev. Inv. Mar., 15 (1), 73-79.
López-Elías, J. & Voltolina, D. (1993). Cultivossemicontinuos de cuatro especies de microalgas conun medio no convencional. Ciencias Marinas, 19 (2),168-180.
Lourenco, S. A., Barbarino, E., Lanfer, U. M. & Aidar, E.(1997). Distribution of intracellular nitrogen in marinemicroalgae: basis for calculation of specific nitrogen-to-protein conversion factors. J. Phycol., 34, 798-811.
López-Elías, J. A. (2002). Evaluación cuantitativa ycualitativa de los sistemas de producción demicroalgas de seis laboratorios comerciales delNoroeste de México. Tesis Doctoral. PostgradoInterinstitucional de Ciencias Pecuarias. Universidadde Colima, Tecomán, México, 117 pp.
López-Elías, J. A., Voltolina, D., Ávila-Mercado, I. S.,Nieves, M. & Cordero-Esquivel, B. (2005). Growth,composition and biomass yields of Chaetocerosmuelleri mass cultures with different routines andtank depths. Rev. Inv. Mar., 26 (1), 67-72.
López-Elías, J. A., Enríquez-Ocaña, F., Pablos-Mitre, M.,Huerta-Aldaz, N., Leal, S., Miranda-Baeza et al.(2008). Growth and biomasa production ofChaetoceros muelleri in mass outdoor cultures: effectof the hours of the inoculation, size of the inoculumand culture medium. Rev. Inv. Mar., 29 (2), 171-177.
Martínez-Córdova, L. R. (1999). Cultivo de CamaronesPeneidos. Principios y Prácticas. AGT Editores, S. A.,México, 283 pp.
Nieves, M., Vega, P. (1994). Tasa de crecimiento,biomasa y costo de producción de Tetraselmissuecica (Chlarophyceae) y Chaetoceros sp.
(Bacillariophyceae), cultivadas con el medio f y tresmedios alternativos. Rev. Ciencias del Mar, UAS,Época I, 13, 39-53.
Piña, P., Voltolina, D., Nieves, M. & Robles, M. (2006).Survival, development and growth of the Pacific whitshrimp Litopenaeus vannamei protozoea larvae, fedwith monoalgal and mixed diets. Aquaculture, 253,523-530.
Pacheco-Vega, J. M. (2003). Evaluación del valornutrimental de la microalga Chaetoceros muellericultivada en un medio no convencional para alimentara larvas de camarón blanco (Litopenaeus vannamei).Tesis de Maestría en Ciencias. Centro de Investi-gación Científica y de Educación Superior deEnsenada, Ensenada Baja California, México, 85 pp.
Ruiz, M. (2009). Camaronicultura Orgánica. Disponibleen http://www.cesasin.com.mx
Ritar, A. J., Dunstan, G. A., Nelson, M. M., Brown, M.R., Nichols P. D., Thomas, C. W. et al. (2004).Nutritional and bacterial profiles of juvenile Artemiafed different enrichments and during starvation.Aquaculture, 239, (1-4), 351-373.Simental-Trinidad, J. A. & Sánchez-Saavedra, M. P.
(2003). The effect of agricultural fertilizer on growth rateof benthic diatoms. Aquac. Engin., 27, 265-272.
Simental-Trinidad, J. A., Sánchez-Saavedra, M. P. &Correa-Reyes, J. G. (2001), Biochemical composition ofbenthic marine diatoms using as culture medium a commonagricultural fertilizer. J. Shellfish Res., 20, 611-617.
Valenzuela-Espinoza, E., Gendrop-Funes, V., PérezCastañeda, R. & Wilburn-González, J. G. (1999).Supervivencia de larvas de Litopenaeus vannamei (Boone)alimentadas con Chaetoceros muelleri producido confertilizantes agrícolas. Ciencias Marinas, 25 (3), 423.

Revista Cubana de Investigaciones PesquerasEnero-diciembre, 2012, vol. 29, NO. 1, ISSN 0138-8452, pp.
12
12-17
Efecto de diferentes niveles de alimentación para Clarias gariepinusen preceba
Effect of different levels of feeding for Clarias gariepinus in the pre-fattening stage
José Llanes Iglesias y José Toledo Pérez
Empresa Desarrollo Tecnologías Acuícolas. Carretera Central km 20 ½,Loma de Tierra, Cotorro, La Habana, Cuba,
E-mail: [email protected]
RESUMEN
Se evaluaron cuatro niveles de alimentación en el comportamiento productivo de Clarias gariepinus (8,0 + 0,01 g depeso medio inicial) en la etapa de preceba, mediante un diseño completamente aleatorizado. Los tratamientosfueron: TI- Pienso vegetal al 5 % del peso corporal; TII- Pienso vegetal al 3 % y desechos pesqueros al 2 % delpeso corporal; TIII- Pienso vegetal al 3 % y desechos pesqueros al 2 % en base seca (que representó el 6 % enbase húmeda) del peso corporal y TIV- Pienso vegetal y desechos pesqueros según Tabla del Laboratorio deNutrición. Los mejores crecimientos (p < 0,001) se encontraron con los tratamientos TIII y TIV (111,2 y 116 gde pesos finales, respectivamente), los que se correspondieron con los niveles de alimentación más altos. Laeficiencia alimentaria fue más favorable con TII (conversión del alimento de 1,2 y eficiencia proteica de 3,8), laque difirió (p < 0,01) del resto de las dietas. La supervivencia no presentó diferencias significativas (p > 0,05),sin embargo, la productividad fue mayor en TIV (11,4 kg/m3). El análisis económico mostró la mayor gananciapara TIV con un monto de $ 9 493,7 en menor tiempo de cultivo (50 días).
Palabras clave: desechos pesqueros, clarias, niveles de alimentación.
ABSTRACT
Four levels of feeding in the productive behaviour of Clarias gariepinus (8,0 + 0,01 g of average weight) in thepre-fattening period, according to a complete randomized experimental design were evaluated. The treatmentswere: TI- Vegetable feed at 5 % of body weight; TII- Vegetable feed at 3 % of body weight and fishing wasteat 2 % of body weight; TIII- Vegetable feed at 3 % and fishing waste at 2 % in dry base (that represented 6 %humid basis) of body weight; TIV- Vegetable feed and fishing waste according to the table of the NutritionLaboratory. The better growths (p < 0,001) were found in TIII and TIV (with 111, 2 and 116 g of final weight,respectively), which were associated with the highest feeding levels. The most favorable feeding efficience waswith TII (feeding conversion of 1,2 and protein efficience of 3,8), this one differed (p < 0,01) from the rest ofthe treatments. The survival rate didn’t presented significatives differences (p > 0,05), however the productivitywas highest in TIV (11,4 kg/m3). The economical analysis showed that the major gain was for the TIV with$ 9 493,7 and the fewest culture time (50 days).
Keywords: fishing waste, clarias, levels of feeding.
INTRODUCCIÓN
Uno de los problemas a los que se enfrentan los acuicul-tores es la necesidad de obtener un equilibrio entre elrápido crecimiento del pez y el uso eficaz de la dieta quese proporciona; prácticas inadecuadas de alimentaciónpueden llevar a un gasto inapropiado del alimento oinsuficiencia del mismo, que resulta en altos costos deproducción y contaminación del medio acuático.
En Cuba, el engorde del bagre africano Clarias gariepinusse realiza en dos etapas: una preceba de 10-100 g de
peso promedio en sesenta días y la ceba hasta alcanzarpeso comercial (600-700 g). Se utilizan los desechosfrescos del procesamiento pesquero y pienso comercialelaborado con materias primas de origen vegetal(fundamentalmente harina de soya). Los niveles dealimentación se rigen por una tabla (porcentaje del pesocorporal por día), incluida en los ProcedimientosOperacionales de Trabajo (POT) para el cultivo intensivodel pez gato africano (Elizarde et al., 2010), pero existela dificultad que las cantidades de alimentos a suministrarvarían en dependencia de las disponibilidades de los

13
mismos en las diferentes provincias del país, lo cualpropicia diferencias en los rendimientos de los cultivos.
El objetivo de este trabajo fue evaluar los efectos decuatro niveles de alimentación en el comportamientoproductivo de Clarias gariepinus en preceba.
MATERIALES Y MÉTODOS
Los experimentos se realizaron en la Planta de Producciónde Alimentos no Convencionales de la Empresa DesarrolloTecnologías Acuícolas (EDTA). Mediante un diseñocompletamente aleatorizado se distribuyeron 420 alevinesde Clarias gariepinus de 8,0 + 0,01 g de peso medio inicialen 12 piscinas rectangulares de cemento de 0,31 m3 decapacidad (tres piscinas por tratamiento), a una densidadde 0,9 kg/m3 (35 peces por acuatorio). El flujo de agua seestandarizó a 1,2 L/min y diariamente se midieron latemperatura y el dioxígeno disuelto con un OxímetroHANNA y el pH con un peachímetro de igual marca.
Los peces se mantuvieron una semana antes decomenzar el bioensayo para su adaptación a lascondiciones experimentales y se alimentaron con unadieta balanceada comercial (32 % de proteína bruta)hasta la saciedad. Transcurrido este tiempo, se suminis-traron los alimentos experimentales (pienso comercialvegetal y desechos frescos del fileteado de clarias) endos raciones al día; por la mañana una porción de pienso(50 %) y por la tarde el resto del mismo y los desechosde pescado (DP), para simular el proceso de alimentaciónen las granjas, según lo establecido por el POT. Elexperimento duró 60 días y cada 15 días se realizaronlos muestreos para el ajuste de la ración.
Del proceso industrial se recogieron los DP en unacaja limpia y se molieron en un molino de carne JAVAR 32a un tamaño de 1 cm congelándose (–10oC) en recipientesplásticos para su distribución diaria. La composiciónproximal de los alimentos experimentales se muestra enlas TABLAS 1 y 2.
TABLA 1. Composición porcentual y proximal delalimento comercial vegetal
TABLA 2. Composición proximal de los desechosfrescos de clarias
TABLA 3. Niveles de alimentación y suscorrespondientes gramos de proteína bruta/kg de pesovivo según Procedimientos Operacionales de Trabajo
para el cultivo intensivo del pez gato africano
TABLA 4. Niveles de alimentación y suscorrespondientes gramos de proteína bruta/kg de pesovivo según Laboratorio de Nutrición (tratamiento IV)
Intervalos de % peso Gramos de proteínapeso medio (g) corporal/día bruta/kg de peso vivo
10-15 10,5 2,2 16-35 9,6 2,0 36-60 8,5 1,75 61-90 7,2 1,52
Intervalos de % peso Gramos depeso medio (g) corporal/día proteína bruta/kg de peso vivo
TI TII TIII
10-75 5 1,30 1,10 1,74 76-115 4,5 1,17 1,00 1,57 118-180 3,5 0,91 0,77 1,22
Ingredientes g/100 gHarina de soya 50Maíz 20Salvado de trigo 23Aceite de soya 3Fosfato dicálcico 3Mezcla vit-mineral 1Humedad 11,2Proteína bruta 26,1Energía digestible (kcal/kg) 2 300
Los tratamientos experimentales fueron:
I. Pienso vegetal al 5 % del peso corporal, según tabla dealimentación del POT de Cultivo intensivo de Clarias(TABLA 3).
II. Pienso vegetal al 3 % y DP al 2 % (en base húmeda)del peso corporal (5 % en total).
III. Pienso vegetal al 3 % y DP al 2 % en base seca (querepresentó el 6 % en base húmeda) del peso corporal.
IV. Pienso vegetal y DP, según tabla del laboratorio denutrición que disminuye los intervalos de peso mediopara el ajuste de la ración y ofrece mayores contenidosde proteína bruta/kg de peso vivo (TABLA 4).
Indicadores g/100 gHumedad 68,5Proteína bruta 16,2Extracto etéreo 8,2Cenizas 4,8
La composición bromatológica de las muestras depienso y desechos de pescado se hicieron triplicadas yde acuerdo con los procedimientos de la AOAC (1995).
Al final del experimento se pesaron individualmentetodos los animales para calcular los siguientesindicadores nutricionales: Peso final (Pf); Ganancia

14
TABLA 5. Resultados de los indicadores productivos de Clarias gariepinus con diferentes niveles de alimentación
diaria = Pf – Pi/días de cultivo; Factor de Conversióndel Alimento (FCA) = Alimento añadido/Ganancia enpeso; Supervivencia = No. Animales finales/No.Animales iniciales x 102; Tasa de eficiencia proteica(TEP) = Ganancia en peso/Proteína suministrada;Productividad = Biomasa final/Área de cultivo.
Se comprobó la normalidad de los datos por la pruebade Kolmogorov-Smirnov y la homocedasticidad de varianzapor la prueba de Bartlett. Posteriormente, se realizó unanálisis de varianza de clasificación simple y lacomparación de medias se docimó según Duncan, pormedio del software estadístico INFOSTAT versión 1.0(Balzarini et al., 2001).
La evaluación económica se realizó a partir de losprecios que brindó el Dpto. de Economía de la EDTA:$ 721,00/tonelada de pienso vegetal; $ 32,00/toneladade desechos de pescado y $ 2 689,5/tonelada depescado entero congelado (clarias). El costo dealimentación se consideró como el 70 % de los costostotales (Toledo et al., 2009) y se calcularon lossiguientes indicadores económicos: Cantidad depienso por tonelada de pescado = FCA x 1 000/% depienso respecto al total de alimento; Cantidad dedesechos por tonelada de pescado = FCA x 1 000/% de
Indicadores TI TII TIII TIV ±±±±± EE sign
Peso final (g) 65,0 c 85,4 b 111,2 a 116,0 a ***± 3,35 ± 3,11 ± 2,95 ± 3,01
Ganancia diaria (g/día) 0,9 c 1,3 b 1,7 a 1,8 a 0,05 ***
Conversión alimentaria 1,5 b 1,2 a 1,8 c 1,5 b 0,08 **
Eficiencia proteica 2,5 c 3,8 a 2,9 bc 3,1 b 0,14 ** Supervivencia (%) 71,4 79,0 76,2 87,6 3,51
Productividad (kg/m3) 5,3 d 7,6 c 9,5 b 11,4 a 0,5 ***
Letras diferentes en la misma fila, difieren estadísticamente para p < 0,05 según Duncan± EE Error estándar** p <0,01 *** p <0,001
La productividad presentó diferencias significativas(p < 0,01) influenciadas por los niveles de alimentacióna favor del TIV, a pesar que no se encontraron diferen-cias (p > 0,05) entre los valores de supervivencias(TABLA 5), lo que indicó que los tratamientos evaluadosno son promotores de mortalidades y los tenores que se
lograron son similares a los obtenidos a escala productivasegún el técnico de la EDTA (Oliva, 2012, Comunicaciónpersonal), los que se atribuyen al canibalismo de la especieen esta etapa de cultivo.
Se demostró que los indicadores productivos de Clariasgariepinus (TABLA 5), se afectaron significativamente por los
I. Pienso vegetal al 5 % del peso corporal, según tabla de alimentación del POT de Cultivo intensivo de Clarias (TABLA 3).II. Pienso vegetal al 3 % y DP al 2 % (en base húmeda) del peso corporal (5 % en total).III. Pienso vegetal al 3 % y DP al 2 % en base seca (que representó el 6 % en base húmeda) del peso corporal.IV. Pienso vegetal y DP, según tabla del laboratorio de nutrición que disminuye los intervalos de peso medio para el
ajuste de la ración y ofrece mayores contenidos de proteína bruta/kg de peso vivo (TABLA 4).
desecho respecto al total de alimento; Costo portonelada de pescado = Precio del alimento x FCA x 0,7;Ganancia por tonelada de pescado = Precio de venta deuna tonelada de pescado – Costo por tonelada de pescado.
RESULTADOS Y DISCUSIÓN
Durante los días de experimentación, la concentración dedioxígeno del agua fue 5 y 7 mg/L, la temperatura varióentre 26-28 oC y el pH de 7,5-8,0, los que se encuentrandentro de los rangos convenientes para el buencomportamiento productivo de la especie, según De Graafy Janssen (1996).
Los indicadores nutricionales evaluados al final delbioensayo entre los juveniles de Clarias gariepinus(TABLA 5), presentaron diferencias significativas, donde losmayores crecimientos (p < 0,001) se obtuvieron con TIII yTIV, los que se corresponden con los niveles de alimentaciónmás altos. Sin embargo, la eficiencia alimentaria (conversióndel alimento y eficiencia proteica) fue más favorable(p < 0,01) para el TII, con la menor adición de gramos deproteína por kilogramo de peso vivo (PV).

15
niveles de alimentación evaluados y repercutieron directa-mente sobre la ingestión y asimilación de los alimentos.
En el tratamiento TII donde se aplicó igual nivel dealimentación que en TI, pero con la sustitución de 40 %del alimento vegetal por DP, se encontró mayor tasa decrecimiento y eficiencia alimentaria a pesar que los pecesrecibieron menores cantidades de materia seca (por losaltos porcentajes de humedad de los DP) y gramos deproteína por kilogramo de PV (TABLA 3). Estos resultadosse corresponden con las conclusiones del experimentorealizado por Llanes et al. (2011), que consignaron elalto valor nutricional que adquieren las dietas vegetalescuando se suplementan con subproductos pesqueros enla alimentación de Clarias gariepinus.
Esto reafirma aún más, el efecto positivo de los DPcuando se cultivan bagres africanos con raciones vegetales,debido a una suplementación de aminoácidos esenciales(principalmente los azufrados), que favorecen la calidadde la proteína dietética. De hecho, la metionina es el primeraminoácido limitante de varias proteínas vegetalesincluyendo la soya (NRC, 1993). De ahí, que al utilizar elalimento vegetal (TI) compuesto por esta única fuenteproteica fue difícil cubrir los requerimientos de eseaminoácido (24 g de metionina/kg proteína, reportado porUnprasert, 1994) para el crecimiento óptimo de las clarias.
Según Fagbenro & Davies (2001), la sustituciónde más de 50 % de harina de pescado (HP) por soyaen la dieta de Clarias gariepinus solo fue posible conla suplementación de metionina. Además, losresultados que informaron Toko et al. (2008)mostraron que la sustitución parcial de la HP por soyay algodón en raciones de la misma especie dismi-nuyeron el crecimiento y los contenidos de calcio,fósforo y zinc en las canales y los filetes de los pecesexperimentales, lo que asociaron al ácido fítico,aspecto importante ya que la absorción de calcio delagua por los peces es alta, por tanto son menosdependientes del calcio dietético, no así el fósforoque su disponibilidad en agua y absorción por los peceses baja, según Hepher (1988).
De hecho, al utilizar los subproductos pesqueros sedisminuyeron las concentraciones de factores antinutri-cionales (cantidad por día), como los inhibidores deproteasas que afectan la actividad proteolítica de losaparatos enzimáticos del tracto digestivo de los peces,presentes en la soya (principal proteína del alimentocomercial) y el ácido fítico que reduce las cantidades defósforo y zinc, lo que tiene una relevancia especial puesestos minerales son críticos para el crecimiento, laregulación del equilibrio ácido-base, el metabolismo deaminoácidos, lípidos, carbohidratos y como cofactor decomponentes biológicos importantes como hormonas yenzimas.
Con el tratamiento TII el FCA mejoró significati-vamente en 300 g menos de alimento por cada kilogramo
de incremento en peso vivo, a pesar de los contenidosde humedad que proporcionó los DP que se refleja eneste indicador y se puede atribuir al alto valor nutricionalde la ración por la suplementación de aminoácidosesenciales y un nivel de energía procedente de los aceitescontenidos en los subproductos pesqueros (ricos enácidos grasos insaturados), que permiten una mejorutilización de la proteína para el crecimiento (efecto deahorro de proteína).
Sin embargo, cuando comparamos este mismotratamiento en relación con TIII y TIV se necesitaría 600y 300 g más de alimento por kg de peso, respectivamente,pero los crecimientos resultaron mayores (p < 0,001) y sedebe tener en cuenta que en estos dos tratamientos seutilizaron altas cantidades de DP que repercuten en los FCApor los contenidos de humedad (TABLA 2), lo que evidencióque los niveles de alimentación de los TI y TII (5 %) nofueron suficientes para cubrir las necesidades diarias paraun máximo rendimiento productivo de la especie.
En los TIII y TIV se encontraron iguales crecimientos(p > 0,05), pero con diferencias significativas para laconversión alimentaria y la productividad (kg/m3), quese puede explicar por un mejor aprovechamiento de lasraciones con las cantidades de alimentos que sesuministraron durante las etapas de muestreos en elTIV. De hecho, con el TIII no se varió el porcentaje deadición del alimento durante los 60 días de bioensayo,ya que en el tercer muestreo (45 días) los pecespresentaron un peso promedio de 56,8 g, cifra menor ala estipulada (70 g) para el cambio del porcentaje deración (TABLA 3). Esto sugiere que al cultivar bagresafricanos con alimentos alternativos es convenienteutilizar tablas con intervalos de peso menores y sumi-nistrar mayores contenidos de proteína por kilogramo depeso vivo (TABLA 4).
Cuando se analizó el esquema de alimentación parael cultivo de bagres que propone la compañía internacionalALLER AQUA GROUP (www.aller-aqua.com) para lapreceba (TABLA 6), se observó que los requerimientos degramos de PB/kg PV fueron similares a los que seutilizaron en el TIV (TABLA 4).
TABLA 6. Esquema de alimentación de la compañíainternacional ALLER AQUA GROUP para el cultivo de
bagres africanos en Europa
Intervalos de Proteína bruta/ Gramos de peso (g) Energía digestible proteína bruta/ (%/MJ) kg de peso vivo
10-20 45/17,3 2,1
20-50 45/17,3 1,4 50-100 37/15,3 1,5
16
El análisis económico (TABLA 7) mostró que el mayorcosto para la producción de una tonelada de pescado fuecon el alimento vegetal (TI), debido a su precio y a lapobre calidad de la proteína vegetal, condiciones que seagravaron por los bajos niveles de alimentación que se
aplicaron. Sin embargo, cuando se complementó estepienso con DP (TII; TIII, TIV), este indicador económicose favoreció por los bajos precios de este subproducto yla alta calidad de su proteína para la especie (Llanes et al.,2011).
TABLA 7. Análisis económico (pesos)
Indicadores TI TII TIII TIV
Cantidad de pienso por tonelada de pescado (kg) 1 500 720 600 720
Cantidad de desechos por tonelada de pescado (kg) - 480 1 200 780
Costos por tonelada de pescado ($) 1 546,4 764,2 673,4 777,8
Ganancia por tonelada de pescado ($) 1 143,1 1 925,3 2 064,1 1 911,7
Diferencias con T-I ($) - 782,2 873,0 768,6
Duración de la preceba (días) 100 70 53 50
Diferencias con T-I (días) - 30 47 50
I. Pienso vegetal al 5 % del peso corporal, según tabla de alimentación del POT de cultivo intensivo de clarias(TABLA 3).
II. Pienso vegetal al 3 % y DP al 2 % (en base húmeda) del peso corporal (5 % en total).III. Pienso vegetal al 3 % y DP al 2 % en base seca (que representó el 6 % en base húmeda) del peso corporal.IV. Pienso vegetal y DP, según tabla del laboratorio de nutrición que disminuye los intervalos de peso medio
para el ajuste de la ración y ofrece mayores contenidos de proteína bruta/kg de peso vivo (TABLA 4).
Los tratamientos TIII y TIV mostraron ganancias/toneladas de pescado similares a TII, pero con unareducción de 20 días de cultivo que representan unaganancia adicional en mano de obra, riesgos a pérdidaspor diferentes índoles, así como tiempo que permitiríarealizar más ciclos de cultivo por año, lo que se afectóen TII por la baja cantidad de DP que se adicionó, lo
cual genera mayor tiempo para alcanzar el pesofinal.
Si se extrapolaran estos resultados a niveles produc-tivos en estanques de 600 m3 (TABLA 8), los mejoresindicadores económicos resultarían con el TIV con un montode ganancias de $ 1 606,0, dado a un mayor rendimientoen kg/m3 respecto a TIII.
TABLA 8. Análisis económico de la preceba de Clarias gariepinus en un estanque de 600 m3 según los resultadosexperimentales (pesos)
Indicadores TI TII TIII TIV
Días de cultivo 100 70 53 50Biomasa final (kg) 3 174 4 584 5 712 6 864Costos totales ($) 4 908,3 3 503,1 3 846,5 5 338,8Ingresos/ventas ($) 8 536,5 12 328,7 15 362,4 18 460,7Ganancias con TI ($) - 5 197,4 7 887,7 9 493,7

17
Es importante destacar que las cantidades de DP quese utilizaron en el TIII (6 % del peso corporal/día) sondifíciles de adquirir diariamente a niveles productivos porla escasez de materias primas en algunas provincias yno se ha logrado una infraestructura de equipamientoque permita el aprovechamiento en su totalidad de todoel subproducto diario que genera la actividad pesqueraen el país.
De ahí, que una solución rápida es la conservación deestos subproductos por técnicas simples y relativamenteeconómica como los ensilajes. Referente a esto, seinformó que la inversión de una Planta de alimentosemihúmedo a base de ensilajes pesqueros con capacidadde producción de 312 t/año de ensilaje de pescado y 780 tde alimento/año para el cultivo de tilapias tuvo un costode inversión de 85 138,57 CUC y un período derecuperación de 2,9 años, los cuales hacían factible lainversión (Toledo et al., 2010).
Un estudio de Toledo et al. (2009), demostró losefectos positivos en el comportamiento productivo y lafactibilidad económica del uso de los ensilajes de residuospesqueros para la alimentación de bagres africanos.
Este estudio fue de especial importancia para el paíspor tratarse de una especie con alto valor comercial ymedioambiental, que requiere de información sobre losrequerimientos cualitativos y cuantitativos de nutrientespara su crecimiento óptimo en el menor tiempo posible,dado que el alimento es uno de los factores más limitanteen la acuicultura, por lo que se estima que estos resul-tados son un aporte para avanzar en el conocimiento de laalimentación de Clarias gariepinus a niveles productivoscon alimentos no convencionales.
CONCLUSIONES
1. La aplicación del 5 % de pienso vegetal (TI), así comoel fraccionamiento con desechos pesqueros (TII),demostraron bajos niveles de alimentación en lapreceba de Clarias gariepinus.
2. La adición de desechos de pescado en base seca(TIII) y la disminución de los intervalos de peso parael ajuste de la ración con mayor adición de gramosde proteína por kilogramo de peso vivo (TIV),mostraron los indicadores productivos y económicosmás favorables, los que se corresponden con losniveles más altos de alimentación.
3. La disminución de los intervalos de peso para el ajustede la ración con mayor adición de gramos de proteínapor kilogramo de peso vivo (TIV), propició los mayoresrendimientos productivos (kg/m3) y monto deganancias.
REFERENCIAS
AOAC (1995). Official Methods of Analysis (16th ed.).Washington, D.C.: Association of Official AnalyticalChemists.
Balzarini, M., Casanoves, F., Di Rienzo, J. A., González,I. A., Robledo, C. W. & Tablada, M. E. (2001).Software estadístico INFOSTAT. Manual de usuario(1a ed.). Córdoba, Argentina.
De Graaf, G. & Janssen, H. (1996). Manual dereproducción artificial y cultivo en estanques del pezgato africano Clarias gariepinus en ÁfricaSubsahariana. Documento Técnico FAO, No. 362,FAO, Roma.
Elizarde, S., Gutiérrez, D., Díaz, G. & Riera, J. (2010).Procedimientos operacionales de trabajo para elcultivo intensivo del pez gato africano (2ª ed.). LaHabana: Ministerio de la industria Alimentaria.
Fagbenro, O. A. & Davies, S. J. (2001). Use of soybeanflour (dehulled, solventextracted soybean) as a fishmeal substitute in practical diets for African catfish,Clarias gariepinus (Burchell, 1822): growth, feedutilization and digestibility. J. Appl. Ichthyology, 17,64-69.
Hepher, B. (1988). Nutrition of pond fishes. New York:Cambridge Univ. Press.
Llanes, J., Toledo, J. & Lazo de la Vega, J. M. (2011).Efectos de dos alimentos en el desempeño productivode Clarias gariepinus en tanques de cementos. Rev.Cub. Inv. Pesq., 28 (1), 24-29.
NRC (National Research Council) (1993). NutrientRequirement of Fish. Committee on AnimalNutrition, Board of Agriculture, National ResearchCouncil. Washington, D.C.: National AcademicPress.
Toko, I. I., Fiogbe, E. D. & Kestemont, P. (2008). Mineralstatus of African catfish (Clarias gariepinus) fed dietscontaining graded levels of soybean or cottonseedmeals. Aquaculture, 275, 298-305.
Toledo, J., Botello, A. & Llanes, J. (2009). Evaluación delos ensilajes químicos de pescado en la alimentaciónde Clarias gariepinus. Rev. Cub. Inv. Pesq., 26 (1),14-18.
Toledo, J., Llanes, J., Aguiar, O. & Lazo de la Vega, J. M.(2010). Factibilidad económica de una planta dealimento semihúmedo a partir de ensilajes desubproductos pesqueros. ACUACUBA, 12 (2), 24-27.
Unprasert, N. G. (1994). An evaluation of the use of“ideal” protein concept to estimate essentialamino acid requirements of the Clarias hybrid(Clarias macrocephalus × C. gariepinus). PhDDissertation, Mississippi State University, MississippiState, M.S.

Revista Cubana de Investigaciones PesquerasEnero-diciembre, 2012, vol. 29, NO. 1, ISSN 0138-8452, pp.
18
18-23
Crecimiento poblacional del rotifero nativo Brachionus sp.“Cayman”, al evaluar diferentes microalgas como alimento
Population growth of the native rotifer Brachionus sp. “Cayman”, with differentsmicroalgae as food
Rosario Cisneros
Unidad de Investigaciones en Acuicultura,Instituto del Mar del Perú
Apartado 22, Chucuito, Callao,[email protected]
RESUMEN
Se estudió el crecimiento poblacional de la cepa Brachionus sp. “Cayman”, cepa Chilca, Perú, en respuesta adiferentes tratamientos con microalgas: Isochrysis galbana (T1), Chaetoceros gracilis (T2), Nannochloropsisoculata (T3), Isochrysis galbana + Chaetoceros gracilis + Nannochloris maculata (T4), Isochrysis galbana +Nannochloris maculata + Nannochloropsis oculata (T5) y Nannochloris maculata (T6). Se evaluó la densidadfinal de la población (DF), tasa de crecimiento (TC) y productividad (R). Los resultados mostraron diferenciasestadísticas entre los tratamientos, encontrándose que T5 presentó mejores resultados en cuanto a DF(190 ± 5,51 indiv.mL-1), TC (0,40 ± 0,01 día-1) y R (38 080 ± 1,53 rot.L-1.día-1).
Palabras clave: rotifero, Brachionus sp., tasa de crecimiento, productividad, microalgas.
ABSTRACT
We study the population growth of Brachionus sp. “Cayman” strain Chilca, Peru, in response to differentmicroalgae, : Isochrysis galbana (T1), Chaetoceros gracilis (T2), Nannochloropsis oculata (T3), Isochrysis galbana+ Chaetoceros gracilis + Nannochloris maculata (T4), Isochrysis galbana + Nannochloris maculata +Nannochloropsis oculata (T5) y Nannochloris maculata (T6). Evaluating the final population density (D), populationgrowth rate (TC) and productivity (R). The results showed statistical differences between treatments, T5 showedthe best results in D (190 ± 5,51 indiv.mL-1), TC (0,40 ± 0,01 día-1) and R (38 080 ± 1,53 rot.L-1.día-1) .
Keywords: rotifer, Brachionus sp., growth rate, productivity, microalgae.
INTRODUCCIÓN
El complejo de rotíferos Brachionus plicatilis agrupavarias especies cosmopolitas, partenogenéticas cíclicasque aparecen regularmente en lagos salados y lagunascosteras. Es el único rotífero con importancia comercialy aplicada a través de su uso como alimento vivo paralarvas de peces y crustáceos (Yoshimatzu et al., 2008;Prol-García et al., 2010; Malekzdeh Viayeh et al., 2010;Haché & Plante, 2011). Debido a su corto tiempo degeneración y a su fácil cultivo en laboratorio ha sidoobjeto de muchas investigaciones básicas en ecología yfisiología. Se han descrito tres especies que muestrandiferencias morfológicas y ecológicas, las cuales hansido descritas o redescritas como B. plicatilis sensu
stricto (L), B. rotundiformis (SM) y B. ibericus (SS)(Gómez et al., 2002; Hagiwara et al., 2007; Papakostaset al., 2009). Muchas especies de rotíferos y candidatosa especies son usualmente referidos como linajes/biotipos y han sido introducidas bajo la designación de“cripticas”. En las cepas de “hatcheries”, estudiosmoleculares han revelado la presencia de cinco especiescripticas/biotipos llamados B. plicatilis s.s., B. sp.“Austria”, B. sp. “Manjacavas”, B. sp. “Nevada” y B. sp.“Cayman” (Ciros-Pérez et al., 2001; Dooms et al., 2007;Vasileiadou et al., 2009).
Una de las preocupaciones recientes sobre la biologíade los rotíferos es la diversidad de características delcomplejo de especies B. plicatilis y sus cepas originadasen diferentes regiones geográficas (Gómez et al., 2002).Con este fin, en este trabajo, se estudió el crecimiento

19
poblacional de Brachionus sp. “Cayman”, cepa Chilca,Perú, en respuesta a diferentes microalgas comoalimento, las que son usadas comúnmente en su cultivo(Kostopoulou & Vadstein, 2007; Yin & Zhao, 2008;MalekzadeH Viayeh et al., 2010), evaluándose la tasade crecimiento y densidad final de la población.
MATERIALES Y MÉTODOS
El trabajo se realizó en el laboratorio de alimento vivodel Centro Acuícola Alexander Von Humboldt, Institutodel Mar del Perú. La cepa de rotíferos utilizada perteneceal complejo Brachionus plicatilis, biotipo Brachionus sp.“Cayman” (Romero, 2008).
Recolecta y aislamiento de la cepa
La cepa fue aislada de muestras de agua recolectadasusando una malla de plancton de 53 mm, en la lagunasalina “La Encantada” (12º 32’ 34,61’’ LS y 76º 43’38,26’’ LW), Chilca, Lima, entre abril y noviembre de2008. Se tomó en cuenta el tipo de microalgapredominante, así como la temperatura, salinidad y pH,durante el período de colecta (TABLA 1). La cepa aisladafue climatizada gradualmente a salinidad de 35 ups ytemperatura de 25 ºC (Cisneros, 2011). Se trabajó conestos parámetros debido a que los organismos marinos,tales como microalgas y peces planos, entre otros, quese cultivan en nuestro laboratorio se realizan a 35 ups,además en otros experimentos con la misma cepa derotífero, Cisneros (2011) observó una buena adaptaciónde esta a la salinidad mencionada, comprobándoseademás que una temperatura de 25°C (temperaturaambiente promedio de la sala de cultivo de rotíferos)proporcionó buen crecimiento poblacional durante laclimatación.
Luego se estableció un cultivo clonal en vasos deprecipitación de 500 mL, con agua de mar filtrada a0,22 μm y autoclaveada, manteniéndose en una cámaraclimática a 25 ºC, utilizando la microalga Nannochlorismaculata como alimento. Las renovaciones del mediode cultivo se realizaron una vez por semana. Se midió latalla de los rotíferos, así como la longitud del huevopartenogenético, tomando como referencia el eje mayor(Ogata et al., 2011), en un microscopio biológicocompuesto, a magnificación de 20X (Fig. 1).
Protocolo experimental
El cultivo masivo de B. sp. “Cayman”, se realizó enrecipientes de 10 L, que contenían agua de mar a 35 upsde salinidad. Por ser los recipientes pequeños no se pudo
utilizar termostatos para elevar la temperatura, seconsiguió mantenerla en 24,5 ºC, al utilizar iluminaciónpermanente para incrementar la temperatura delambiente. El alimento consistió en diferentes especiesde algas (seis tratamientos) durante cuatro días: Isochrysisgalbana (T1), Chaetoceros gracilis (T2), Nannochloropsisoculata (T3), Isochrysis galbana + Chaetoceros gracilis+ Nannochloris maculata (T4), Isochrysis galbana +Nannochloris maculata + Nannochloropsis oculata (T5)y Nannochloris maculata (T6). Las concentracionesalgales fueron de 3,7 x 106 cel.mL-1 (T1), 4 x 106 cel.mL-1
(T2), 11 x 106 cel.mL-1 (T3), 3,6 x 106 cel.mL-1 (T4),11 x 106 cel.mL-1 (T5) y 4 x 106 cel.mL-1 (T6), las mismasque se determinaron mediante conteo en una cámara deNeubauer. El recuento del número de rotíferos se realizódiariamente, utilizando una cámara de Sedgwick-Rafter.La densidad inicial de la población fue de 38 a40 indiv.mL-1. Se determinó la densidad final (DF)(indiv.mL-1) obtenida con cada tratamiento, así como latasa de crecimiento poblacional (TC) (día-1), mediante lasiguiente fórmula (Suantika et al., 2000; Tinh et al.,2006; Rioboo et al., 2007):
TC = (ln Nt – ln No)/t
La productividad (R) en rot.L-1.día-1 (Verginelli et al.,1994), aplicando:
R = Nt – No/t
Donde:No: densidad inicial de rotíferos (indiv.mL-1)Nt: densidad de rotíferos (indiv.mL-1) al tiempo tt: período de cultivo (días)
Análisis estadístico
Todos los tratamientos fueron realizados por triplicado.Los datos de productividad (R) fueron transformados aescala logarítmica con el fin de aplicar el análisisestadístico. La normalidad de los datos y homogeneidadde varianzas, se determinaron mediante las pruebasde Shapiro-Wilkins y Levene respectivamente. Paradeterminar diferencias significativas entre lostratamientos, se aplicó un Análisis de Varianza de unaVía (ANOVA) a TC y R, seguido de la prueba de Tukey(p < 0,05). Todos los análisis se realizaron con elprograma SPSS versión 19.0.
RESULTADOS Y DISCUSIÓN
En la TABLA 1 se muestran los parámetros físico-químicosregistrados en “La Encantada” durante los meses derecolecta, observándose menor abundancia entre mayo

20
y junio (otoño), ausencia entre julio y agosto (invierno),aumentando la abundancia a partir de septiembre(primavera). En este sentido Gómez et al. (2002)mencionan que las poblaciones de rotíferos son confrecuencia estacionales y la reproducción sexual esinducida bajo condiciones ambientales adversas,produciéndose huevos sexuales en estado de
latencia, estos huevos son resistentes a la dese-cación y permanecen en el sedimento hasta que lascondiciones favorables vuelvan a aparecer, laestrategia es la de maximizar la supervivencia de laspoblaciones silvestres, a través de estos bancos dehuevos y reducir el impacto de las variacionesestacionales.
TABLA 1. Parámetros físico-químicos, rotíferos y microalgas registrados en la laguna “La Encantada”, Chilca, Lima,durante los meses de recolecta de Brachionus sp. “Cayman”. Abril-noviembre, 2008
Mes Temperatura Salinidad pH Rotíferos Microalgas (oC) (ups)
Abril 27,0 50 7 ++++ Skeletonema costatum
Mayo 18,0 58 9 + Nannocloropsis sp.Nannochloris sp.
Junio 18,0 46 8 + Nannocloropsis sp.
Julio 19,5 47 8 – Nannochloris sp.Dunaliella salina
Agosto 20,0 45 10 – Nannocloropsis sp.Nannochloris sp.Tetraselmis sp.
Septiembre 21,0 46 10 ++ Nannochloris sp.Tetraselmis sp.
Octubre 25,5 46 10 ++ Nannochloris sp.Nannocloropsis sp.
Noviembre 26,0 46 10 ++ Nannochloris sp.Nannocloropsis sp.
(+) Densidad poblacional muy baja(++) Densidad poblacional baja(+++) Densidad poblacional media(++++) Densidad poblacional alta
En la TABLA 2 se observan las medidas promedio delongitud de la lórica y eje mayor del huevo partenogenéticode B. sp. “Cayman”. Las hembras maduras generalmentellevan de 1 a 4 huevos partenogenéticos (Fig. 1),
ocasionalmente cinco (eje mayor, 84,1 ± 16,1 μm;n = 34). Así mismo se hace una comparación conparámetros morfológicos de las tres especies del complejoB. plicatilis.
Especie/biotipo Longitud lórica (n) Eje mayor huevo partenogenético (n) Referencia
B. sp. “Cayman” 171 ± 22 (40) 84,1 ± 16,1 (34) Presente estudioB. plicatilis (tipo SS) 179 ± 4 Ogata et al., 2011B. plicatilis (tipo S) 212 ± 20 Ogata et al., 2011B. plicatilis (tipo L) 275 ± 24 Ogata et al., 2011
TABLA 2. Comparación de parámetros morfológicos ( ± DS) entre B. sp. “Cayman” cepa Chilca, Perú y tresespecies del complejo B. plicatilis sp. (los valores están en micras)
x

21
De acuerdo con lo observado, la cepa estudiada en estetrabajo presenta características similares a B. ibericus encuanto a su morfología (descrita por Gómez et al., 2002) ypreferencia de parámetros abióticos. Ciros-Pérez et al.(2001) describen que B. ibericus tiene una ocurrencia a altassalinidades (8 a 50 ups) y a altas temperaturas (> 15 ºC),similar a lo encontrado con nuestra cepa.
En cuanto a características morfológicas, la cepaChilca presenta cuerpo muy pequeño, redondeado, conpocas espinas pequeñas, la longitud de la lórica (171 ±22 μm) es mucho más pequeña que el tipo-S del complejoB. plicatilis y un poco menos que el tipo-SS (TABLA 2).Estudios realizados sobre el uso de rotíferos pequeñosen larvicultura, fueron reportados por Hagiwara et al.(2007) (en Ogata et al., 2011) y Wullur et al. (2009);estos estudios fueron realizados con Brachionus tipo-SS(longitud de lórica 176 ± 4 μm) y Proales similis (longitudtotal 109 ± 15 μm).
Fig. 1. Brachionus sp. “Cayman”, aislado de una lagunasalina en la localidad de Chilca, Lima, Perú
El tamaño pequeño de la cepa Chilca resulta ventajosopara alimentar larvas con tamaño de boca pequeño, yaque el uso de este tipo de rotíferos como alimento, mejorasignificativamente el rendimiento y aumenta la tasa desupervivencia de las larvas (Ogata et al., 2011).
Tasa de crecimiento (TC) y productividad (R)
La temperatura mostró tener una gran influencia sobre elcrecimiento de la cepa estudiada, habiéndose realizadoanteriormente su cultivo masivo entre 25 y 27 °C (Cisneros,2011). En este sentido se ha demostrado que el aumentode temperatura incrementa la reproducción de los rotíferos,como resultado de una aceleración del metabolismo y
desarrollo, así mismo la temperatura óptima es diferentepara las diferentes cepas de rotíferos (Hagiwara et al., 2007;Wullur et al., 2009; Vasileiadou et al., 2009).
La DF de la población al término del experimento fuemayor en T5 (190 ± 5,51 indiv. mL-1) (Fig. 2). En relacióncon TC, este parámetro es considerado como una variableindicador en la dinámica de poblaciones de los rotíferos,así como para evaluar la producción en sistemas de cultivomasivo (Malekzadeh Viayeh et al., 2010).
Densidad inincialDensidad final
Tratamientos
Den
sida
d (in
div.
mL–
1
T1 T2 T3 T4 T5 T6
250
200
150
100
50
0
Fig. 2. Densidad inicial y final ( ± DS) (indiv.mL-1)obtenidas en el cultivo del rotífero Brachionus sp.“Cayman” cepa Chilca, Perú, utilizando diferentes
algas (p < 0,05)
Los resultados del ANOVA para TC y R mostrarondiferencias estadísticas entre los tratamientos(p < 0,05), T5 presentó mayor TC (0,40 ± 0,01 día-1)(TABLA 3), como consecuencia, esta mezcla de algastambién proporcionó mejor R (38 080 ± 1,53 rot.L-1. día-1)(Fig. 3).
Tratamientos
Prod
uctivi
dad
(rot
. L–1.d
ía–1
T1 T2 T3 T4 T5 T6
50000
40000
30000
20000
10000
0
Fig. 3. Productividad ( ± DS) (rot. mL-1. día-1) obtenidaen el cultivo del rotífero Brachionus sp. “Cayman” cepa
Chilca, Perú utilizando diferentes algas (p < 0,05)
x
x

22
En relación con este parámetro, nuestros resultadosfueron mayores a los de Abu-Rezq & James (2005),quienes encontraron un R de 12 130 ± 1,89 rot.L-1.día-1
alimentando rotíferos con Chlorella + levadura, adicio-nando bacterias lácticas, así como 6 640 rot.L-1.día-1 conla misma dieta pero sin bacterias.
TABLA 3. Estimados ( ± DS) de la tasa de crecimiento (TC) (día-1) obtenida en el cultivo del rotífero Brachionussp. “Cayman” cepa Chilca, Perú, con diferentes algas
x
Tratamientos T1 T2 T3 T4 T5 T6
TC 0,32 ± 0,03 0,26 ± 0,02 0,31 ± 0,01 0,21 ± 0,06 0,40 ± 0,01 0,28 ± 0,02
En el presente trabajo, T5 proporcionó mejor ren-dimiento poblacional por su contenido de Nannochloropsis,la cual está entre una de las principales algas usadas enla producción en masa de rotíferos, por su alta tasa decrecimiento, su pequeño tamaño y contenido de ácidosgrasos altamente insaturados de la serie ω-3 (20:5ω-3),los cuales son esenciales para el crecimiento en larvasde peces (King et al., 2002; Cabrera et al., 2005).Además T5 contenía Isochrysis galbana, microalga quepresenta niveles altos de 22:6ω-3, en este sentido Kinget al. (2002), mencionan que rotíferos alimentados conuna mezcla de Isochrysis galbana y Nannochloropsisgaditana por 24 h, presentaron niveles altos de ácidosgrasos como el 20:4ω-6 y 20:5ω-3. Malekzadeh Viayehet al. (2010) refieren que el alimento es el componentemás importante en un cultivo de rotíferos y el tipo dealgas usadas afecta directamente su reproducción yconsecuentemente la tasa de crecimiento poblacional.Estos mismos autores encontraron valores de TC entre0,62 y 0,82 día-1 en cultivo de B. plicatilis conNannochloropsis. Sin embargo, los valores de TCobtenidos en el presente trabajo, están dentro del rangoobtenido por otros autores, tales como Cabrera, T. et al.(2005), quienes evaluaron los efectos de diferentesmicroalgas y la salinidad sobre el crecimiento de trestipos de B. plicatilis, encontrando con Nannochloropsisoculata, valores de 0,34 a 0,59 día-1 para B. plicatilis,0,42 a 0,64 día-1 para B. rotundiformis y 0,35 a 0,38 día-1
para una cepa SS aislada en zonas tropicales, los valoresmás altos correspondieron a cultivos realizados asalinidad de 32 ups. En este sentido, los valores de TCpueden variar con el tipo de rotífero, parámetros comosalinidad y temperatura, así como las condiciones decultivo y forma de cosecha de la microalga (Ferreiraet al., 2008). Así mismo Cabrera, T. et al. (2005), refierenque rotíferos originados en aguas costeras, se esperaque tengan una TC superior en cultivos con salinidad másalta y nuestros resultados coinciden con otros estudiosque han reportado buen crecimiento de rotíferos atemperaturas alrededor de 25 ºC y 35 ups de salinidad,particularmente al utilizar Nannochloropsis como alimento.
En este sentido Yin & Zhao (2008) en un trabajo realizadocon diferentes cepas de Brachionus plicatilis, encontraronvalores de 0,4 a 0,5 a 18 ºC, incrementándose la TC alaumentar la temperatura hasta 18 ºC. Así mismo, losvalores de TC obtenidos en el presente trabajo están dentrodel rango encontrado en otros estudios, con otras microalgas,como el de Cabrera, M. I. (2008), en un experimento decultivo con Brachionus rotundiformis, utlizando Chlorella,quien encontró valores de 0,37 a 1,96 día-1, dependiendode la densidad algal.
Determinar las condiciones óptimas de cultivo y límitespara el crecimiento poblacional del alimento vivo utilizadoen larvicultura de peces, es un requerimiento para suproducción en masa (Wullur et al., 2009), de ahí laimportancia de conocer las preferencias de alimento y elrango de factores ambientales tolerados por una especie,por ello sería recomendable realizar más investiga-ciones con esta cepa, utilizando diferentes parámetrosambientales, con el fin de optimizar las condiciones decultivo para obtener un máximo de producción.
REFERENCIAS
Abu-Rezq, T. & James, C. (2005). Application ofprobiotics in rotifer production systems for marinefish hatcheries. Marine finfish Aquaculture Network,July-September.
Cabrera, M. I. (2008). Tasa de crecimiento ploblacionaldel rotífero Brachionus rotundiformis (Rotifera:Brachionidae) en un quimiostato de dos cámaras. Rev.Biol. Trop., 56, 1149-1157.
Cabrera, T., Bae, J. H., Bai, S. C. & Hur, S. B. (2005).Effects of microalgae and salinity on the growth ofthree types of the rotifer Brachionus plicatilis. J. Fish.Sci. Technol., 8 (2), 70-75.
Ciros-Perez, J., Gómez, A. & Serra, M. (2001). On thetaxonomy of three sympatric sibling species of theBrachionus plicatilis (Rotifera) complex from Spain,with the description of B. ibericus n. sp. Journal ofplankton research, 23 (12), 1311-1328.

23
Cisneros, R. (2011). Rendimiento poblacional del rotíferonativo Brachionus sp. “Cayman”, utilizandodiferentes enriquecedores. Ecol. apl., 10 (2), 99-105.
Dooms, S., Papakostas, S., Hoffman, S., Delbare, D.,Dierckens, K., Triantafyllidis et al. (2007). DenaturingGradient Gel Electrophoresis (DGGE) as a tool forthe characterisation of Brachionus sp. Strains.Aquaculture, 262, 29-40.
Ferreira, M., Maseda, A., Fabregas, J. & Otero, A.(2008). Enriching rotifers with “premium”microalgae. Isochrysis aff. galbana clone T-ISO.Aquaculture, 279, 126-130.
Gómez, A., Serra, M., Carvalho, G. & Lunt, D. H. (2002).Speciation in ancient cryptic species complexes:evidence from the molecular phylogeny of Brachionusplicatilis (rotifera). Evolution, 56 (7), 1431-1444.
Haché, R. & Plante, S. (2011). The relationship betweenenrichment, fatty acid profiles and bacterial load incultured rotifers (Brachionus plicatilis L-strain) andArtemia (Artemia salina strain Franciscana).Aquaculture, 311, 201-208.
Hagiwara, A., Suga, K., Akazawa, A., Kotani, T. &Sakakura, Y. (2007). Development of rotifer strainswith useful traits for rearing fish larvae. Aquaculture,268, 44-52.
King, J., Xiaoming, L. & Rusch, K. (2002). Nutritionalproperties of the marine rotifer Brachionus plicatilisfed the fresh water microalgae Selenastrumcapricornutum. World Aquaculture Society, 33,478-488.
Kostopoulou, V. & Vadstein, O. (2007). Growthperformance of the rotifers Brachionus plicatilis, B.“Nevada” and B. “Cayman” under different foodconcentrations. Aquaculture, 273, 449-458.
Malekzdeh Viayeh, R., Mohammadi, H. & Banj Shafiei,A. (2010). Population growth of six IranianBrachionus rotifer strains in response to salinityand food type. Internat. Rev. Hydrobiol, 95 (6),461-470.
Ogata, Y., Tokue, Y., Yoshikawa, T., Hagiwara, A. &Kurokura, H. (2011). A loation strain of the rotiferBrachionus angularis holds promise as a food sourcefor small-mouthed larvae of freshwater fish inaquaculture. Aquaculture, 312, 72-76.
Papakostas, S., Triantafyllidis, A., Kappas, I. &Abatzopoulos, T. J. (2009). Clonal composition ofBrachionus plicatilis s.s and B. sp. Austria Hatchery
strains based on microsatellite data. Aquaculture,296, 15-20.
Prol-García, M. J., Planas, M. & Pintado, J. (2010).Different colonization and residence time of Listonellaanguilarum and Vibrio splendidus in the rotiferBrachionus plicatilis determined by real-real time PCRand DGGE. Aquaculture, 302 (2010), 26-35.
Rioboo, C., Prado, R., Herrero, C. & Cid, A. (2007).Population growth study of the rotifer Brachionussp. fed whit triazine-exposed microalgae. AquaticToxicology, 83, 247-253.
Romero, L. (2008). Caracterización morfométrica yaspectos filogenéticos de cepas de rotíferos del grupoBrachionus plicatilis (Rotifera: Brachionidae) utilizadosen la acuicultura peruana. Tesis para optar por eltítulo profesional de Biólogo, Facultad de CienciasBiologicas, Universidad Nacional Mayor de SanMarcos, Lima, Perú.
Suantika, G., Dhert, P., Rombaut, G., Vandenbergue, J.,De Wolf, T. & Sorgeloos, P. (2002). The use of ozonein a high density recirculation system for rotifers.Aquaculture, 201, 35-49.
Tinh, N., Phuoc, N., Dierckens, K., Sorgeloos, P. &Bossier, P. (2006). Gnotobiotically grown rotiferBrachionus plicatilis sensu strictu as a tool forevaluation of microbial functions and nutritional valueof different food types. Aquaculture, 253, 421-432.
Vasileiadou, K., Papakostas, S., Triantafyllidis, A.,Kappas, I. & Abatzopoulos, T. (2009). A multiplexPCR method for rapid identification of Brachionusrotifers. Mar biotechnol, 11, 53-61.
Verginelli, R., Marín, N. & Lodeiros, C. (1994). Cultivomasivo del rotífero Brachionus plicatilis con tresdietas de microalgas. Rev. Lat. Acui. Lima-Perú, 43,68-71.
Wullur, S., Sakakura, Y. & Hagiwara, A. (2009). Theminute monogonont rotifer Proales similis deBeauchamp: Culture and feeding to small mouthmarine fish larvae. Aquaculture, 293, 62-67.
Yin, X. W. & Zhao, W. (2008). Studies on life historycharacteristics of Brachionus plicatilis O.F. Müller(Rotifera) in relation to temperature, salinity and foodalgae. Aquatic ecol., 42, 165-176.
Yoshimatsu, T., Higuchi, T., Hamasaki, Y. & Tanaka, K.(2008). Preliminary Trials on the Effects of Lightingfor the Populations Growth of the Rotifer, Brachionusplicatilis. JARQ, 42, 131-136.

Revista Cubana de Investigaciones PesquerasEnero-diciembre, 2012, vol. 29, NO. 1, ISSN 0138-8452, pp.
24
Technology Cuban tilapia and common carp implemented in southern Namibia
Noris Millares Dorado1 y Mabel Navarro Jam2
1 Centro de Investigaciones Pesqueras. Ave. 5ta. y calle 246, Santa Fe, Playa,La Habana, Cuba, CP 19100, Teléfono: (537) 209-7875,
E-mail: [email protected] Estación de Alevinaje Pavón. Encrucijada. Villa Clara. Cuba
RESUMEN
En el Centro de Acuicultura Continental Hardap se implementó un programa de desarrollo de la acuicultura apartir de la formación de tres bancos de reproductores, dos de tilapia mossambica, uno tomado de la presaHARDAP (246 y 216 , y el otro (250 y 215 a partir de alevines seleccionados del segundo ciclode desove de esta especie logrado en dicho centro. El tercer banco de reproductores formado fue de carpacomún (200 y 236 capturado del medio natural. Se estableció un flujo productivo de ambas especies porciclos que permitió obtener alevines destinados a las granjas de peces estatales, privadas, y a las comunidadessituadas en las regiones de: Karas, khomas, Omaheke y Hardap. En el primer cuatrimestre del año 2011 sellevaron a crecimiento en tierra 135 650 alevines, se inicio la siembra de 14 350 ejemplares a densidadesde 3 ej./m2 en los 48 acuatorios previamente seleccionados en condiciones de monocultivo, o en policultivo enuna proporción de tilapia al 60 % y 40 % de carpas. La cosecha se realizó entre los 9-11 meses de cultivo conpesos entre 200-300 g y supervivencia de 70-85 %. Miembros de la comunidad previamente capacitados, seincorporaron a la atención del cultivo integrado de peces con vegetales (zanahoria, remolacha, cebolla) y animalesdomésticos (cabras, chivos y carneros). Para promocionar el consumo de pescado y métodos de preparación seimpartió un taller a la comunidad de Hardap. Se desarrolló una labor divulgativa sobre la actividad acuícola convista a orientar el perfil vocacional de los alumnos del último año de secundaria de las regiones de Mariental yRehoboth. Al término de la asistencia técnica en Hardap se evaluó en los técnicos los conocimientos piscícolasy la aplicación de las habilidades adquiridas en los entrenamientos.
Palabras clave: cultivo, carpa común, tilapia.
ABSTRACT
In the Hardap Continental Aquaculture Center, was implemented a program of aquaculture development from theformation of three groups of parents, two of the tilapia mossambica, one taken from Hardap Dam (246 and216 , and other (250 and 215 selected fingerlings from the second spawning cycle of this speciesmade in the center. The third groups of parents was formed of common carp (200 and 236 captured fromthe wild. A production flow was established that yielded fingerlings of both species for fish farming state,private, and communities in the regions of: Karas, Khomas, Omaheke and Hardap. In the first quarter of 2011were carried fattening 135 650 fingerlings; began stocking 14 350 fingerlings at densities of 3 fish/m2 in the48 previously selected reservoirs in monoculture, polyculture was used in the ratio for tilapia 60 % and 40 %of carp. The harvest took place between 9-11 months of cultivation weighing between 200-300 g and survivalof 70-85 %. Community members previously trained, joined the growing attention of integrated fish farmingvegetables (carrots, beets, onions) and domestic animals (goats, sheep and goats). Fish consumption andpreparation methods were promoted in a workshop to the community of Hardap. An informative work wasdeveloped on aquaculture to guide the vocational profile of students in their final year of secondary regionsMariental and Rehoboth. Fish knowledge and application of skills learned in training were evaluated at the endof technical assistance in Hardap
Keywords: culture, common carp, tilapia.
24-29
Tecnología cubana de cultivo de tilapia y carpa común implementadaen la región sur de Namibia
))
)
)
) )

25
INTRODUCCIÓN
La acuicultura en los países de bajos ingresos comoNamibia gana cada día más importancia, se mejora laalimentación de la población con productos sanos, ricosen proteína como el pescado y se generan fuentes deempleo (OLDEPESCA, 2012). La República de Namibialimita al Norte con Angola y Zambia, al Este con Botswanay Sudáfrica, al Sur con Sudáfrica, y al Oeste con elocéano Atlántico.
Tiene 824 269 km² de superficie. El clima es cálido yseco. La población es escasa, de 2 063 927 hab. en2008, y tiene una de las más bajas densidades depoblación del mundo 2,5 hab./km2.
Existe un potencial acuícola provenientes de los ríosOkavango (1), Kunene (2), Zambezi (3), Orange (4), asícomo lagos y presas (Fig. 1).
2 1 3
4
Fig. 1
La política de desarrollo del Ministerio de Pesqueríasy Recursos Marinos (MFRM) de Namibia está dirigida areducir la pobreza del país a partir de mejorar la alimen-tación, aumentar los empleos, el nivel educacional y losconocimientos científico-técnicos.
El objetivo del trabajo fue implementar un sistemade producción de alevines de tilapia y carpa destinados agarantizar el cultivo en el centro Hardap, el suministrode peces a granjas privadas y proyectos comunitarios;además de capacitar y entrenar a técnicos y obreros enhabilidades piscícolas.
MATERIALES Y MÉTODOS
El trabajo se realizó en el Centro de AcuiculturaContinental Hardap durante dos años a partir de abrilde 2009 (Fig. 2). Para la formación del banco de repro-
ductores, se procedió a probar todos los estanques detierra, con la finalidad de preparar (chapeo, encalado)aquellos que reunían las condiciones para el cultivo, elresto se cuantificó para ser sometidos a reparación.
Fig. 2 Centro de Acuicultura Continental Hardap
Se desarrolló la técnica del cultivo de la tilapiamossambica (Oreochromis mossambicus) y la carpacomún (Cyprinus carpio) (Fig. 3), por ser especies nativasresistentes, soportar temperaturas entre 24-30 oC, conhábitos alimentarios diferentes, alta fecundidad, buencrecimiento aceptando el alimento natural y artificial, yencontrarse entre las preferidas en el mercado local(Alamilla, 2002).
La presa Hardap situada aproximadamente a unadistancia de 500 m es la que abastece de agua a laestación, esta se caracteriza por presentar muchamateria orgánica en suspensión lo que limita la actividadde fertilización de los estanques. Los reproductores detilapia y carpa fueron capturados de dicha presa dosmeses antes de la etapa de reproducción para suaclimatación a las condiciones de cautiverio.
La producción de alevines de ambas especies fuerealizada en la tilapia a partir de la reproducción naturalestablecida por ciclos, y en la carpa común por elmétodo natural y semiinducido aplicando una dosisúnica de 0,5 mL/kg de Ovaprim (Arias & Hernández,2009).
Las larvas obtenidas recibieron alimento artificial enpolvo con 50 % de Proteína Bruta al 30 % de la biomasahasta la talla de 0,5 g; los alevines se alimentaron conun alimento extrusado (32 % PB) al 15-10 % de labiomasa total, en el engorde este se suministró al 2 %hasta la talla de cosecha. A partir del resultado de los
Fig. 3 Tilapia mossambica (Oreochromis mossambicus)y carpa común (Cyprinus carpio)

26
muestreos mensuales la ración fue reajustada basadaen el peso y la tasa de crecimiento. Al término del cultivose calculó la supervivencia (%) (López, 2011).
Los alevines obtenidos en el centro fueron destinadosa granjas de peces estatales, privadas, y como parte deun programa de desarrollo de acuicultura comunitaria alas regiones de: Karas, khomas, Omaheke y Hardap dondea partir de una valoración visual se seleccionaron48 acuatorios que fueron sembrados con 14 350 alevinesde 10 g de peso promedio de tilapia mossambica y carpacomún, en densidad de 3 ej./m2 en monocultivo opolicultivo (60 % tilapia y 40 % carpas) hasta la talla demercado. Conjuntamente se realizó una labor de capacita-ción con los miembros de la comunidad en prácticaspiscícolas (destacando el uso del agua desechada) y suintegración con el cultivo de vegetales (zanahoria,remolacha, cebolla) y animales de crianza (cabras, chivosy carneros).
Se llevó a cabo un chequeo diario de las variablesfísico-químicas: la temperatura (oC) y el dioxígeno disuelto(mg/L), determinados con un equipo marca HANNA y latransparencia con el disco Secchi; el pH fue valoradouna vez a la semana con un equipo de igual marca. Endías alternos se repuso el agua perdida por la evaporación.
La implementación de la disciplina tecnológica decultivo se apoyó en un programa de capacitación yentrenamiento a técnicos y trabajadores del centro y lagranja privada Ecofish, con temas generales depiscicultura (preparación de estanques, fertilización,sistemas de cultivo, precría, alevinaje, ceba, parámetrosfísico-químicos, entre otros), con especial interés seimpartió el cultivo de tilapia y carpa, prácticas de sexado,muestreos, alimentación, aplicación de hormonas en eldesove semiinducido de la carpa e incubación de huevos.
En las escuelas de Mariental y Rehoboth sedesarrolló una labor divulgativa dirigida a los alumnosdel último año de secundaria, abordando la importanciade la acuicultura con el objetivo de ampliar su perfilvocacional.
Se efectuó como promoción un taller dirigido a dar aconocer los atributos del pescado y las diferentes formasde prepararlo en la comunidad Hardap, ya que el mismono se encuentra entre las carnes preferidas por lapoblación, quizás por el alto precio.
RESULTADOS Y DISCUSIÓN
Namibia es un país que muestra un constante crecimientoaunque la producción de las regiones es todavía marginalcomparada con su potencial de desarrollo (Halwart &Dam, 2010). La población existente puede apenasmantenerse con la actual producción agrícola por lo quese impone integrarla con otros cultivos como los pecesy animales domésticos.
Resultados logrados en los diferentes pasos delProcedimiento Operacional de Trabajo para el cultivo detilapia y carpa:
1. Selección y preparación de estanques de tierra paraestablecer la producción de peces.
ANTES
DESPUÉS
De la batería de 20 estanques de tierra en desusodel Centro de Acuicultura Continental Hardap, solose pudo preparar el 35 % de los estanques paraimplementar la reproducción y el cultivo de alevinesde tilapia y carpa.
2. Fomento del banco de reproductores a partir de lacolecta de los peces de la presa Hardap.
a) Se seleccionaron de la presa Hardap 246 hembrasy 216 machos de tilapia mossambica con pesosmayores de 200 g y de la carpa común 200 hembrasy 236 machos con pesos superiores a 300 g.

27
b) Se ubicaron las hembras de las diferentesespecies juntas en un mismo estanqueseparadas de los machos que fueron colocadosunidos en otro estanque. Gupta & Acosta(2004), refieren que si se trabaja con reproduc-tores tomados directamente del medio debenser capturados antes de la etapa reproductivapara que puedan adaptarse a las condicionesde cautiverio.
3. Inicio del primer ciclo reproductivo a partir de losreproductores capturados del medio natural.En Namibia las estaciones de primavera-verano sepresentan desde octubre hasta marzo. Los mesesmás fríos son junio, julio y agosto. En la región surse alcanzan temperaturas mínimas entre 0-8 oC, porlo que en este tiempo recesan las actividades decultivo.En las visitas a las granjas de cultivo de peces enla región norte (Rundu y Katima Murilo) y sur(Hardap), se observó que la reproducción de latilapia se realizaba en un ciclo con una duración de3-4 meses.Las larvas no eran pescadas diariamente, por loque se unían con los reproductores, larvas reciénnacidas, postlarvas de diferentes tallas y alevines(10-20 g). Esta práctica no permitía efectuar unpronóstico de la cantidad de alevines a obtener deacuerdo con el número de hembras en desove, lasupervivencia estimada oscilaba entre 10-20 %,afectada por el canibalismo entre los animales dediferentes tallas.En el mes de enero de 2010 se comenzó lareproducción de la tilapia, seguido de la carpa.Se aplicó el método de trabajo establecido porciclos que coincide con lo reportado por El-Sayed(2006); Lim & Webster (2006). Los pasos a seguirson:
• Sexado, selección de hembras maduras y machoscon esperma.
• Remoción de huevos o larvas de la cavidad bucalde las hembras.
• Pesca diaria de larvas para ser sembradas enprecría.
De los reproductores maduros de tilapia salvajes seobtuvo un total de 10 000 alevines con 40 % desupervivencia.De acuerdo con la capacidad de las instalaciones apartir de estos alevines se formó por selección masalel segundo banco de reproductores de: 250 hembrasy 215 machos. Estos se incorporaron a la reproduc-ción en el segundo ciclo del año 2011.
4. Inicio del primer y segundo ciclo reproductivo con elbanco de reproductores salvajes y los formados apartir de la progenie.En el primer cuatrimestre de 2011 se establecióel flujo productivo de la tilapia con dos bancos dereproductores con un sistema alterno de trabajo(mientras un grupo está en reproducción, el otrose prepara para el siguiente ciclo), en la carpa solose emplea un banco por ser la cantidad de alevinesa obtener menor. Se llevaron a crecimiento entierra 135 650 alevines y fueron trasladados14 350 alevines para el engorde (de estos 5 740de carpas, el 40 % de la proporción en el policultivocon la tilapia) a las granjas de peces y comunidadesde las regiones de Karas, khomas, Omaheke yHardap.Se determinó aplicar técnicas sencillas de produccióncon el mínimo manejo posible, baja densidad desiembra y bajo recambio de agua, con la intención deque grupos familiares pudieran desarrollarlo mediantesistemas de cría extensivos o semiintensivos,integrados con otros cultivos en áreas adyacentespreparadas para el cultivo de cebolla, zanahoria,remolacha, maíz y el pastoreo de animalesdomésticos, lo que proporciona un uso al aguadesechada rica en nutrientes de los intercambios,cosechas parciales o totales para el autoconsumo ola comercialización parcial (Saavedra, 2006; Yang &Chen, 2006).

28
Prein & Dey (2010) señalan que la integración delos cultivos es una alternativa que atenúa la pobrezay la escasez de agua, lo que obliga a utilizar ladisponible de la forma más racional y económicaposible e incluso reutilizarla. En estas regiones deNamibia este método de obtener el agua para elriego de las cosechas y abasto para los animales,reduce los riesgos de pérdidas por la extremavariabilidad de las lluvias, pero, aun así es difícil deimplementar debido al carácter impredecible delagua disponible.En el engorde en las áreas comunitarias losanimales mostraron un incremento diario en pesoentre 0,87-0,7 g/día; a los 9-11 meses se realizóla cosecha de los peces con tallas entre 200-300 gy supervivencia de 70-85 %. Se alcanzaron produc-ciones de 21-155 kg de acuerdo con los parámetrosantes señalados.
5. Promoción de la acuicultura en las escuelas ycomunidades.Se realizó una labor de promoción a la jovengeneración en las escuelas de Mariental y Rehobothcon vista a enriquecer su perfil vocacional y en lascomunidades se divulgó los objetivos y la importanciade la acuicultura como medio de obtener empleos ymejorar la situación económica y alimentaria de lasfamilias.
6. Promoción sobre el uso del pescado y formas deprepararlo.Se dieron a conocer diferentes recetas para lapreparación del pescado combinado con vegetales,arroz, frutas, entre otros, con buena aceptación.
CONCLUSIONES
1. Se implementó un sistema de producción de alevinesde tilapia y carpa destinados a garantizar el cultivoen el centro Hardap, el suministro a granjas de pecesestatales, privadas y proyectos comunitarios.
2. Se estableció para el cultivo de la tilapia el métodode trabajo por ciclos, logrando una produccióncontinua de larvas y alevines a partir de dos bancosde reproductores con un sistema alterno de trabajo(un grupo en reproducción, el otro preparándose parael siguiente ciclo).
3. Se estableció para el cultivo de la carpa el desovenatural y semiinducido con métodos para la incuba-ción de huevos.
4. En las comunidades se integró el cultivo de peces aotras actividades para mejorar el nivel de vida y laalimentación.
5. Se capacitó a técnicos, obreros y personas de lascomunidades en el cultivo de peces.
6. Se realizó un trabajo de promoción con la jovengeneración, en escuelas y comunidades divulgandolos objetivos de la acuicultura, las temáticas quetrabaja y las ofertas de empleos.
RECOMENDACIÓN
Existen tecnologías adecuadas como la acuicultura quepueden ponerse a disposición del desarrollo de lapoblación pobre evaluando previamente el entorno demanera adecuada para evitar un impacto negativo en laintegridad del sistema.
REFERENCIAS
Alamilla, H. (2002). Cultivo de Tilapias. ZOETecnoCampo, México, 16 pp.
Arias, J. J. & Hernández, J. L. (2009). Efectos delextracto hipofisiario de carpa común y el análogo dela GnRH sobre la maduración final del oocito y eldesove de la cachama negra (C. macropomum).Revista Científica, FCV-Luz, vol. XIX, 5, 486-494,Universidad de Antioquia.
El-Sayed, A. F. M. (2006).Tilapia Culture. CABI Publisher,Wallingford, Oxfordshire, UK.
Gupta, M. V. & Acosta, B. O. (2004). A review of globaltilapia farming practices. Aquacult. Asia, 10 (1), 7-16.
Halwart, M. M. & Dam, A. A. (2010). Integración yacuicultura en África occidental. Conceptos, prác-ticas y potencial. Roma, FAO, 193 pp.

29
Lim, C. & Webster, C. (2006). Tilapia: Biology, Culture,and Nutrition. Hawthorn Press.
López, J. A. (2011). Acuicultura integrada. Cultivo depeces y vegetales. Consultado el 20/2/2012 en http://www.buenastareas.com/ensayos
OLDEPESCA (2012). Diagnóstico de la AcuiculturaMarina en la región de América Latina y el Caribe.XXII Conferencia de Ministros, La Habana, 17-18de mayo de 2012.
Prein, M. & Dey, M. M. (2010). Piscicultura comunitariaen llanuras inundables estaciónales. En M. M.
Halwart, M. M. & Dam, A. A. (Eds.), Integración yacuicultura en África occidental. Conceptos, prácticasy potencial (pp. 17-26). Roma, FAO.
Saavedra, M. A. (2006). Manejo del cultivo de tilapia.Departamento de Tecnología y Arquitectura. Facultadde Ciencia, Tecnología y Ambiente. UniversidadCentroamericana. Managua, Nicaragua, marzo, 2006.
Yang, H. Y., Fang, Y. & Chen, Z. (2006). Agro-acuiculturaintegrada. Manual básico. Departamento de Pesca.Depósito de documentos de la FAO. Consultado enhttp://www.fao.org/docrep/006/y1187s05.htm

Revista Cubana de Investigaciones PesquerasEnero-diciembre, 2012, vol. 29, NO. 1, ISSN 0138-8452, pp.
30
30-37
The effect of the cyclonic season 2005 on the phytoplanktonfrom Cienfuegos Bay, Cuba
Ángel Moreira,1 Augusto Comas,1 Mabel Seisdedo1 y Sinai Barcia2
1 Centro de Estudios Ambientales de Cienfuegos (CEAC). Calle 17 esq. Ave. 46 s/n,Reparto Reina, Cienfuegos, Cuba, CP 55100
2 Centro Meteorológico Provincial de Cienfuegos. Calle 52 A, e/ 24 y 26, s/n, CP 55100, Cienfuegos, Cuba
RESUMEN
El fitoplancton es uno de los componentes más abundantes de la biodiversidad de la Bahía de Cienfuegos. Se estudióel impacto de la temporada ciclónica 2005 sobre la composición y abundancia de estos organismos en el período demarzo-octubre. La composición específica de microalgas planctónicas de la bahía en marzo estuvo dominada pordiatomeas y dinoflagelados marino-estuarinos como Nitzschia longissima, Thalassionema nitzschioides y Prorocentrummicans. Al final de la temporada ciclónica, los cambios en la estructura de la comunidad fitoplanctónica fueron drásticos,prácticamente no se registró fitoplancton marino en la bahía. Las microalgas dulciacuícolas de los géneros Chlorella,Monoraphidium, Cyclotella, Aulacoseira, Pseudoanabaena, Planktothrix, Dolichospermum, Microcystis y Cryptomonaspredominaron con altas concentraciones, lo cual constituye un indicador de aporte de agua dulce a este ecosistema.
Palabras clave: Bahía de Cienfuegos, fitoplancton, huracán Dennis, microalgas tóxicas.
ABSTRACT
Phytoplankton is one of the most abundant components of biodiversity of Cienfuegos Bay. The impact of thecyclonic season 2005 on the composition and abundance of these organisms was studied from the period March toOctober. The specific composition of planktic microalgae of the bay in March was dominated by diatoms anddinoflagellates marine-estuarines such as Nitzschia longissima, Thalassionema nitzschioides and Prorocentrum micans.At the end of the cyclonic season, changes in the structure of phytoplanktic community were drastic; practically themarine phytoplankton was not registered at the bay. Freshwater microalgae of genus Chlorella, Monoraphidium,Cyclotella, Aulacoseira, Pseudoanabaena, Planktothrix, Dolichospermum, Microcystis and Cryptomonas predominatedin high concentrations, which constitute an indicator of freshwater inflow to this ecosystem.
Keywords: Cienfuegos Bay, phytoplankton, Dennis hurricane, toxic microalgae.
El efecto de la temporada ciclónica 2005 sobre el fitoplanctonde la Bahía de Cienfuegos, Cuba
INTRODUCCIÓN
Las microalgas, macroalgas y fanerógamas, constituyenlos eslabones primarios de la gran cadena alimentaria delmundo acuático y por ello están considerados de alto valorecológico. Se estima que el 90 % del total de lafotosíntesis y del aporte de oxígeno a la atmósfera tienelugar en los océanos producto a la actividad delfitoplancton. Estos organismos desempeñan además unpapel importante en la alimentación de diversidad depeces, moluscos, larvas de crustáceos y otros organismosde interés comercial. Algunos animales marinos mayores,como ballenas, tiburones gigantes y la raya diablo, la utilizandirectamente como alimento.
Los ecosistemas marinos en Centroamérica y el Caribeson afectados anualmente por el paso de varios huracanes.
Varios estudios han evidenciado que después del paso dehuracanes se produce un desequilibrio en la estructura delos ecosistemas, lo cual se manifiesta en un cambiodrástico en la estructura de las comunidades planctónicasy bentónicas. Especialmente, los sistemas estuarinosreciben un incremento en la carga orgánica y biomasa defitoplancton, sufriendo este último cambios en sucomposición taxonómica (Walker et al., 2005; Paerl et al.,2005).
La Bahía de Cienfuegos es uno de los recursosnaturales más preciados de la provincia de igual nombre,se encuentra ubicada en el centro-sur de Cuba y formaparte del complejo estuarino costero el cual, según Baisre(1992), está limitado a unas pocas porciones de laplataforma cubana. Entre los recursos más abundantesde este acuatorio se encuentra el fitoplancton marino

31
(Moreira et. al., 2007). El presente trabajo tiene comoobjetivo estudiar el impacto producido por la temporadaciclónica 2005 sobre la composición y abundancia de lascomunidades del fitoplancton en la Bahía de Cienfuegos,Cuba.
MATERIALES Y MÉTODOS
Área de estudio
La Bahía de Cienfuegos (Fig. 1) se encuentra ubicada enlos 22o 9' L. N. y 80o 27' L. O. en la región sur-centralde Cuba. Es una bahía semicerrada con un áreasuperficial de 90 km² y profundidad media de 14 m,conectada al Mar Caribe por un estrecho canal deaproximadamente 3 km de largo y una profundidad de30 m. La celda norte recibe la mayor parte del impactode las descargas de residuales urbanos de la ciudad deCienfuegos (Villasol et al., 1990), mientras que el lóbulosur recibe un menor impacto antrópico, aportado por losríos Caonao y Arimao.
La bahía presenta características estuarinas basadasen la estratificación vertical como resultado del aportede los ríos y a la baja energía mareal. Durante la estaciónde lluvia (mayo-octubre) la bahía muestra bajos valoresde salinidad en la capa superficial (16-20 ‰), mientrasque en la capa de fondo mantiene salinidades oceánicas.El resto del año muestra salinidades de 30-32 ‰(Seisdedo & Muñoz, 2004).
Fig. 1 Red de monitoreo hidrológico en la Bahía deCienfuegos
Mediciones físico-químicas
Se tomaron muestras en superficie y fondo para lamedición de la temperatura, la salinidad y nutrientes como
fosfato (P-PO43-), nitrito (N-NO2
-) y silicato (Si-SiO32-).
La salinidad y la temperatura se determinaron in situmediante una sonda digital modelo YSI-30. El silicato sedeterminó por la técnica del ácido silicomolíbdico y elfosfato por el método de Murphy y Riley, ambos segúnla UNEP (1991). Para la cuantificación del nitrito se utilizóla metodología de la UNESCO (1983).
Se tomaron los valores de las precipitaciones antes,durante y después del paso del huracán, así como lavelocidad del viento durante el paso del eventometeorológico. Estos datos fueron ofrecidos por elInstituto de Recursos Hidráulicos y Centro MetereológicoProvincial de Cienfuegos.
Muestreo y procesamiento de muestras
De los 16 puntos que conforman la red de monitoreohidroquímico (Seisdedo et al., 2004) de la Bahía deCienfuegos se seleccionaron 10 estaciones, teniendo encuenta para sus localizaciones los principales rasgosmorfológicos de este ecosistema, sus diferentes usos,las fuentes contaminantes presentes, entre otras(Fig. 1). Las estaciones 2; 14 y 15 corresponden al LóbuloSur de la bahía, mientras que las estaciones 7; 8; 10;11 y 12 corresponden al Lóbulo Norte.
Las muestras fueron colectadas durante los mesesde marzo (seca), y octubre (lluvia) de 2005. La colectase realizó al nivel subsuperficial (0,2 m de profundidad)durante las horas claras del día, con la ayuda de unabotella Niskin. El volumen de muestra total fue de 1 L yla fijación se efectuó inmediatamente con lugol ácido. Seempleó el método de sedimentación. Para el análisis delas muestras se utilizó una cámara de conteo Rigosha yun microscopio biológico OLYMPUS BH-2, con aumentoocular de 12x y objetivos de 16x, 40x y 100x. Laconcentración de los organismos se expresó en célulaspor litro. Para la identificación de las especies seconsultaron: Balech, 1988; Hallegraeff et al., 1995; Hasle& Syverstsen, 1997).
Con el objetivo de comparar la variación de los valoresde densidad microalgal entre estaciones y meses seutilizó un análisis de varianza bifactorial (p < 0,05). Elanálisis de los datos se realizó a través del paqueteestadístico STATISTICA 5.0.
RESULTADOS Y DISCUSIÓN
Condiciones hidrometeorológicas
Los valores de precipitación entre los meses de enero yabril fueron muy bajos. En mayo y junio se produjo unpequeño aumento de las lluvias, no obstante, losacumulados se quedaron muy por debajo de la media

32
histórica para este bimestre, el más lluvioso del añohistóricamente (Barcia & Orbe, 2006) (Fig. 2). Losvalores de salinidad de la bahía en este períodoevidenciaron marcadas condiciones oceánicas, con valorpromedio de 32 ppt en superficie (TABLA 1).
Las intensas lluvias asociadas al huracán Dennis (julio,2005) contribuyeron a que el acumulado en el mes dejulio estuviera al 265 % con respecto a la media histórica(Fig. 2). En relación con la velocidad del viento, la estaciónmeteorológica de Cienfuegos registró al paso del meteorouna racha máxima de 158 km/h de región nordeste, lo
que ubicaba a la zona de la bahía en el radio de vientoshuracanados de Categoría 2 (Escala Saffir-Simpson)(Barcia & Orbe, 2006; Alfonso et al., 2005).
La salinidad, después del paso del huracán, descendióbruscamente en toda la bahía. Un día posterior al pasode Dennis, la salinidad registrada cerca del canal deentrada a la bahía fue de 7,3 ppt. Posteriormente, en elmes de octubre el valor promedio de salinidad fue de10,8 ppt en superficie. Los valores de algunos nutrientescomo nitrato, nitrito, fosfato y silicato se incrementaronen este período lluvioso (TABLA 1).
TABLA 1. Resultados de los indicadores hidroquímicos antes del huracán (seca, marzo/2005) y al finalizar latemporada ciclónica (lluvia, octubre/2005)
Fig. 2 Comportamiento mensual de las precipitaciones con respecto al promedio histórico. Provincia Cienfuegos, 2005(porcentaje con respecto al valor histórico)
Real
Real
Histórico
Histórico
E F M A M J J A S O N D
38 2635 47 66 177 257 184 214 230 186 50
500450400350250200150100500
milí
met
ros
Fuente: Instituto de Recursos Hidráulicos.
Bahía de Cienfuegos Seca Lluvia
Indicadores Unidad Superf. Fondo Superf. Fondo
Salinidad ppt 32,61 32,41 10,84 22,81Temperatura oC 26,16 26,06 29,76 29,76 pH u 8,02 8,04 8,24 8,01Transp. (%) % 49,56 11,26 OD mg/L 6,79 6,95 7,65 5,30Sólidos susp. mg/L 6,06 7,79 10,71 6,44 N-NH3 umol/L 0,49 0,015 N-NO2 umol/L 0,060 0,063 0,61 0,46 N-NO3 umol/L 0,41 0,40 1,63 1,32 Si umol/L 29,81 28,65 143,00 95,06 P-PO4 umol/L 0,36 0,49 1,59 1,07 DBO5 mg/L 1,23 3,99 Grasas y aceites mg/L 7,59 12,96

33
Composición y abundancia del fitoplancton
Antes del huracán Dennis (marzo, 2005) se registraronvalores mesotróficos de concentración fitoplanctónica.El valor promedio de densidad celular en toda la bahíafue de 2,82 x 105 cél./L. Estos resultados concuerdancon los obtenidos por Moreira et al. (2007), y evidencianel carácter mesotrófico del ecosistema Bahía deCienfuegos para el mes de marzo del período de seca.Las estaciones 11; 12 y 14 fueron las que presen-taron mayores concentraciones fitoplanctónicas,estas estaciones presentan influencia de aportesantropogénicos (residuales domésticos) e influenciafluvial.
La composición específica de microalgas planctónicasde la bahía estuvo representada por microalgas estuarino-marinas, mientras que las microalgas dulciacuícolas noestuvieron representadas. Se registraron un total de 36especies: 19 diatomeas (1 Amphora, 1 Bacteriastrum,1 Biddulphia, 4 Chaetoceros, 1 Cocconeis, 1 Cyclotella,1 Navicula, 3 Nitzschia, 1 Pleurosigma, 1 Pseudonitzschia,1 Thalassionema, 1 Thalassiosira, 1 Thalassiothrix y 1Rhizosolenia), 11 dinoflagelados (1 Ceratium, 1 Dinophysis,1 Gymnodinium, 1 Hermesinum, 1 Pyrophacus, 3
Prorocentrum y 1 Protoperidinium), 1 crisofícea(Dictyocha fibula), 2 cianofíceas (1 Oscillatoria y 1Merismopedia), 1 euglenofícea (Eutreptiella marina) y 1criptofícea (Cryptomonas sp.).
Las diatomeas fueron los organismos más abundantesdurante el mes de marzo en todas las estaciones, exceptoen las estaciones 11 y 13, donde dominaron losdinoflagelados (Fig. 3). Entre las diatomeas, las especiesNitzschia longissima, Nitzschia sp. y Thalassionemanitzschioides fueron las especies más abundantes en lamayoría de las estaciones (TABLA 2). Como se mencionóanteriormente, los dinoflagelados fueron abundantes enlas estaciones 11 y 12, donde la especie cosmopolitaformadora de mareas rojas, Prorocentrum micans yProrocentrum compressum fueron las especies domi-nantes (Tabla 2). Dinophysis caudata (2,2 x 103 cél./L),potencialmente productora de toxinas diarreicas seregistró en la estación 12, mientras que Gymnodiniumcatenatum (1,6 x 102 cél./L) potencialmente productorade toxinas paralizantes se registró en la estación 14. Ladiatomea Pseudonitzschia sp. potencialmente productorade toxina amnésica presentó una mayor distribuciónespacial, registrándose en la mayoría de las estaciones(TABLA 2 y Fig. 4).
ChlorophyceaeChrysophyceaeCryptophyceaeCyanophyceaeEuglenophyceaeDinophyceaeBacillariophyceae
100
90
80
70
60
50
40
30
20
10
0
Porc
enta
je
Antes del huracán
100
90
80
70
60
50
40
30
20
10
0
Porc
enta
je
ChlorophyceaeXantophyceaeCryptophyceaeCyanophyceaeDinophyceaeBacillariophyceae
Efecto del huracán
Fig. 3 Variación del porcentaje de abundancia de los principales grupos del fitoplancton antes del huracán(marzo de 2005) y al finalizar la temporada ciclónica (octubre de 2005)
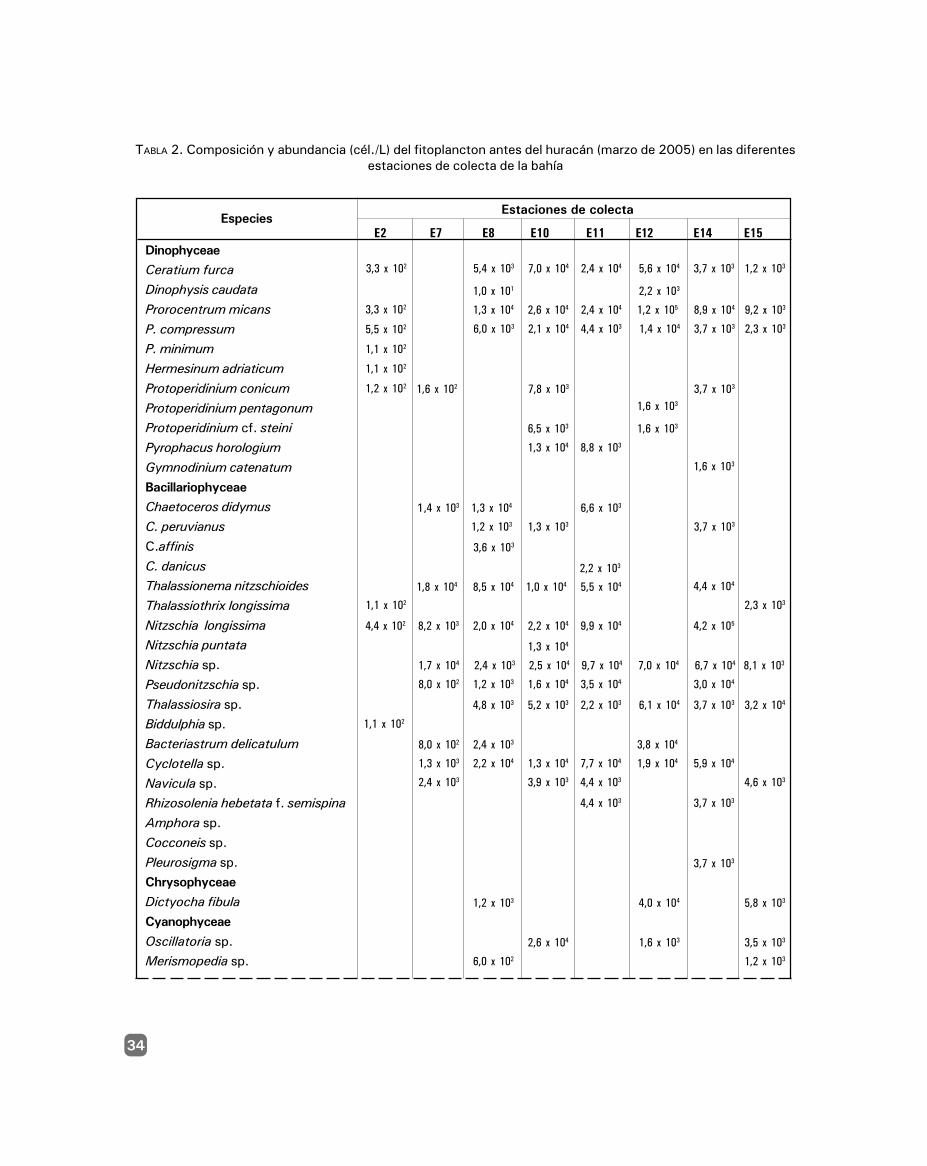
34
TABLA 2. Composición y abundancia (cél./L) del fitoplancton antes del huracán (marzo de 2005) en las diferentesestaciones de colecta de la bahía
Estaciones de colecta
E2 E7 E8 E10 E11 E12 E14 E15DinophyceaeCeratium furcaDinophysis caudataProrocentrum micansP. compressumP. minimumHermesinum adriaticumProtoperidinium conicumProtoperidinium pentagonumProtoperidinium cf. steiniPyrophacus horologiumGymnodinium catenatumBacillariophyceaeChaetoceros didymusC. peruvianusC.affinisC. danicusThalassionema nitzschioidesThalassiothrix longissimaNitzschia longissimaNitzschia puntataNitzschia sp.Pseudonitzschia sp.Thalassiosira sp.Biddulphia sp.Bacteriastrum delicatulumCyclotella sp.Navicula sp.Rhizosolenia hebetata f. semispinaAmphora sp.Cocconeis sp.Pleurosigma sp.ChrysophyceaeDictyocha fibulaCyanophyceaeOscillatoria sp.Merismopedia sp.
Especies
3,3 x 102
5,5 x 102
1,1 x 102
1,1 x 102
1,2 x 102
1,1 x 102
4,4 x 102
1,1 x 102
7,0 x 1043,3 x 102
1,6 x 102
1,4 x 103
2,2 x 103
8,2 x 103
1,7 x 104
8,0 x 102
8,0 x 102
1,3 x 103
2,4 x 103
5,4 x 103 2,4 x 104 5,6 x 104 3,7 x 103 1,2 x 103
2,2 x 1031,0 x 101
1,3 x 104 1,2 x 1052,6 x 104 2,4 x 104 8,9 x 104 9,2 x 103
6,0 x 103 2,1 x 104 4,4 x 103 1,4 x 104 3,7 x 103 2,3 x 103
7,8 x 103 3,7 x 103
1,6 x 103
6,5 x 103 1,6 x 103
1,3 x 104 8,8 x 103
1,6 x 103
1,3 x 104 6,6 x 103
1,2 x 103 1,3 x 103 3,7 x 103
3,6 x 103
1,8 x 104 8,5 x 104 1,0 x 104 5,5 x 104 4,4 x 104
2,3 x 103
2,0 x 104 2,2 x 104 9,9 x 104 4,2 x 105
1,3 x 104
2,4 x 103 2,5 x 104 9,7 x 104 7,0 x 104 6,7 x 104 8,1 x 103
1,2 x 103 1,6 x 104 3,5 x 104 3,0 x 104
4,8 x 103 5,2 x 103 2,2 x 103 6,1 x 104 3,7 x 103 3,2 x 104
2,4 x 103 3,8 x 104
2,2 x 104 1,3 x 104 7,7 x 104 1,9 x 104 5,9 x 104
3,9 x 103 4,4 x 103 4,6 x 103
4,4 x 103 3,7 x 103
3,7 x 103
1,2 x 103 4,0 x 104 5,8 x 103
2,6 x 104 1,6 x 103 3,5 x 103
6,0 x 102 1,2 x 103

35
Estaciones de colecta
E2 E7 E8 E10 E11 E12 E14 E15ChlorophyceaeChlorophyceaeo EustigmatophyceaeMonoraphidium sp.BacillariophyceaeAulacoseira granulataThalassiosira sp.Cyclotella meneghiniana
Fig. 4 Especies potencialmente tóxicas presentes durante elestudio: A-C: Antes del huracán, A. Dinophysis caudata, B.Gymnodinium catenatum, C. Pseudonitzschia sp. D-F: Final
de la temporada ciclónica, D. Microcystis aeruginosa, E.Dolichospermum solitarium, F. Planktothrix isothrix
Después del paso del huracán y el resto de latemporada ciclónica que incluyó altas precipitacionesdurante el mes de octubre, la composición específica demicroalgas cambió totalmente, comportándose la bahíacomo un acuatorio dulciacuícola, registrándose altasconcentraciones de microalgas de agua dulce (Fig. 3),las cuales debieron ser arrastradas hacia la bahía a través
de los ríos por la enorme influencia fluvial asociada a latemporada ciclónica.
Es de notar que después del paso del huracánprácticamente no se registraron organismos marinoscomo los dinoflagelados, los cuales son abundantes enla Bahía de Cienfuegos. Ceratium furca, dinoflageladotípico de la bahía solo se encontró cerca del canal deentrada. Las especies dulciacuícolas dominantes fueronlas clorofíceas como Monoraphidium sp., diatomeascentrales como Cyclotella meneghiniana y Aulacoseiragranulata; cianofíceas como Pseudoanabaena sp.,Planktothrix isothrix, Dolichospermum solitarium yMicrocystis aeruginosa; y flagelados como Cryptomonasmarssonii. Es de destacar que la pequeña especie similara Chlorella (Chlorophyceae o Eustigmatophyceae, de 1-2 μmde diámetro) registró concentraciones muy elevadas,superiores a 107 en la mayoría de las estaciones (TABLA 3y Fig. 4). Es de notar que la mayoría de las cianofíceasregistradas en altas concentraciones son potencialmenteproductoras de toxinas neurodigestivas. Similar alpresente resultado, durante el mes de junio de 1988, seobservaron florecimientos de Microcystis aeruginosa enplayas de la Bahía de Cienfuegos producto a altasprecipitaciones registradas en la provincia de Cienfuegos(A. Comas, comunicación personal).
TABLA 3. Composición y abundancia del fitoplancton después del efecto del huracán y la temporada ciclónica(octubre de 2005)
Especies
ChlorophyceaeChlorophyceaeo Eustigmatophyceae 6,5 x 103
EuglenophyceaeEutreptiella marina 6,0 x 102 9,1 x 103 1,5 x 104 1,6 x 103 3,7 x 104
CryptophyceaeCryptomonas sp. 1,0 x 103
1,6 x 102
3,2 x 103
3,9 x 107 5,0 x 106 2,4 x 106 4,1 x 107 2,5 x 107 1,2 x 107 1,4 x 107 3,2 x 107
2,0 x 105 1,6 x 108 7,9 x 104 1,6 x 105 2,8 x 105
3,5 x 105 2,2 x 104 1,8 x 104 3,6 x 103
3,2 x 105 9,3 x 106 4,4 x 105 1,3 x 106 1,6 x 105 2,8 x 105
36
TABLA 3 (continuación)
Estaciones de colecta
E2 E7 E8 E10 E11 E12 E14 E15Especies
XantophyceaeCentritratus cf. belenophorusChryptophyceaeCryptomonas cf. marssoniiCyanophyceaePseudoanabaena sp.Planktothrix isothrixDolichospermum solitariumMicrocystis aeruginosaDinophyceaeCeratium furca
Finalizando la temporada ciclónica 2005, todas lasestaciones registraron elevadas concentraciones demicroalgas de agua dulce, sin embargo, contradic-toriamente, las estaciones de mayor influencia fluvialcomo la 7 y la 8 no presentaron las mayores concen-traciones, lo cual pudiera estar relacionado con lapropia influencia de las corrientes fluviales quearrastran el fitoplancton hacia la salida de la bahía.El incremento de la concentración fitoplanctónica podríaestar asociado al incremento de los valores de algunosnutrientes como nitrato, nitrito, fosfato y silicato, quese incrementaron en este período lluvioso debido a lainfluencia de los ríos y arrastres hacia la bahía (TABLA 1).
Al comienzo del período seco 2005-2006 (noviembrede 2005), siguiendo posibles eventos de florecimientoso mareas rojas de microalgas, pudo observarse en unárea próxima a la termoeléctrica (E10), que el agua seencontraba de color rojo intenso y esa misma coloraciónse repetía en la zona de Punta Gorda (Estación 12).Esto estuvo asociado a la presencia de florecimiento dedinoflagelados no tóxicos, predominando la especieProrocentrum compressum. Si bien pudiera resultarcontradictorio la ocurrencia de estos eventos en unaetapa del año donde existen buenas condicioneshidroquímicas, esto puede justificarse con una respuestano inmediata de los organismos fitoplanctónicos antelas condiciones que aproximadamente un mes antespudieron registrarse. Dichas condiciones comprendíanmayores valores de nutrientes y materia orgánica, loque resulta favorable para el desarrollo del fitoplancton.Resultados similares han sido observados en estuariosdel sur de la Florida después del paso de huracanes ytormentas tropicales (Walker et al., 2005; Paerl et al.,2005).
En otro sentido, la renovación de la microflora alginade la Bahía de Cienfuegos estudiada en el presente trabajopudiera ser beneficiosa para este ecosistema. Se hademostrado que el incremento del aporte fluvial en lossistemas acúaticos disminuye el tiempo de renovaciónde las aguas en los acuatorios, y ayuda a eliminar o disiparla acumulación de nutrientes proveniente de residualesurbanos, así como los focos de microalgas (mareas rojas),incluyendo las especies tóxicas (Reynolds & Lund, 1988).
CONCLUSIONES
• Se registraron intensas lluvias asociadas al HuracánDennis y durante la temporada ciclónica 2005 engeneral, que contribuyeron a un brusco descenso delos valores de salinidad y a un ligero incremento enla concentración de nutrientes en la Bahía deCienfuegos.
• Debido a las altas precipitaciones, la microflora alginade la Bahía de Cienfuegos sufrió una renovación total.Prácticamente no se registraron organismos marinos,mientras que las especies dulciacuícolas se regis-traron en altas concentraciones.
REFERENCIAS
Alfonso, G., Regueira, V., Santana, O. & Barcia, S. (2005).Dennis; el gran huracán de julio 2005 en Cienfuegos.Lecciones. Memorias en soporte magnético del IIICongreso Cubano de Meteorología, La Habana, 5-9 dediciembre, 31 pp.
4,0 x 104
1,0 x 105 6,7 x 105 5,5 x 105 5,5 x 105 1,6 x 106 1,4 x 106 2,8 x 105
1,3 x 106 1,2 x 105 2,1 x 106 1,3 x 105 1,4 x 106 1,3 x 106 1,6 x 106
1,8 x 105 6,8 x 105 1,6 x 106
1,3 x 103 2,2 x 103 1,2 x 103 2,1 x 103 1,3 x 103 1,4 x 103 1,3 x 103 1,6 x 103
1,7 x 103 1,1 x 103 2,1 x 103
1,8 x 103

37
Baisre, J. S. (1992). Marine Resources of the Antilles.FAO. Fish. Tech. Pap., 326, 182-235.
Balech, E. (1988). Los dinoflagelados del AtlánticoSudoccidental. Publ. Espec. Inst. Esp. Oceanogr., 1,1-310.
Barcia, S. & Orbe, G. (2006): Resumen Climático Anual2005. Centro Meteorológico Provincial de Cienfuegos(inédito), 16 pp.
Hallegraeff, G. M., Anderson, D. M., & Cembella, A. D.(1995). Manual on Harmful Marine Microalgae. IOCManuals and Guides No. 33, UNESCO, 551 pp.
Hasle, G. R. & Syverstsen, E. E. (1997). Marine Diatoms.In C. R. Tomas (Ed.), Identyifying Marine Diatomsand Dinoflagellates (pp. 5-385). California: AcademicPress.
Moreira, A., Seisdedo, M., Leal, S., Comas, A.,Delgado, G., Regadera, R. et al. (2007).Composición y abundancia del fitoplancton de labahía de Cienfuegos, Cuba. Rev. Invest. Mar., 28(2), 97-109.
Paerl, H., Valdés, L., Joyner, A., Peierls, B., Piehler, M.,Riggs, S. et al. (2006). Ecological Response toHurricane Events in the Pamlico Sound System, NorthCarolina, and Implications for Assessment andManagement in a Regime of Increased Frequency.Estuaries and Coasts, 29, 1033-1045.
Reynolds, C. S. & Lund, J. W. G. (1988). Thephytoplankton of an enriched, soft-water lake subject
to intermittent hydraulic flushing (Grasmere,English Lake District). Freshwater Biology, 19,379-404.
Seisdedo, M. & Muñoz, A. (2004). Influencia de laestacionalidad en el comportamiento fisicoquímicode las aguas de la Bahía de Cienfuegos. Rev. Cub.Inv. Pesq. Número especial. III Taller InternacionalCONyMA, 2004.
Seisdedo, M, Muñoz, A. & Castellanos, M. E. (2004).Propuesta de actualización del Programa de Moni-toreo Hidrológico de la Bahía de Cienfuegos. Rev.Cub. Inv. Pesq., 1, Edición Especial, 1-16.
UNEP (1991). Standard chemical methods for marineenvironmental monitoring. Reference Methods forMarine Pollution Studies. No. 50.
UNESCO (1983). Chemical Methods for use in MarineEnviromental Monitoring. Manual and Guides. IOC.No. 12.
Villasol, A., Jaime, N., Mederos, R., Quintana, H.,Martínez, J., Fernández, I. et al. (1990). Estudios dela Contaminación en la Bahía de Cienfuegos. Informefinal del proyecto. Ministerio del Transporte, Inst.Invest. Transp., 66 pp.
Walker, N., Leben, R. & Balasubramanian, S. (2005).Hurricane-forced upwelling and chlorophylla enhancement within cold-core cyclones in the Gulfof Mexico. Geophysical Research Letters, 32,10-16.

Revista Cubana de Investigaciones PesquerasEnero-diciembre, 2012, vol. 29, NO. 1, ISSN 0138-8452, pp.
38
38-43
Physical-chemical characteristics of the waters and phytoplankton in bathroomareas of Cienfuegos Bay, Cuba (2008-2009)
Mabel Seisdedo1, Ángel R. Moreira1 y Gustavo Arencibia2
1 Centro de Estudios Ambientales de Cienfuegos (CEAC), Calle 17 esq. Ave. 46 s/n.Reparto Reina, Cienfuegos, CP 55100, Cuba, E-mail: [email protected]
2 Centro de Investigaciones Pesqueras. Ave. 5ta. y calle 246, Santa Fe, Playa,La Habana, Cuba, CP 19100, Teléfono: (537) 209-7852
RESUMEN
Se estudian las características físico-químicas de las aguas y el fitoplancton en zonas de baño de la Bahía deCienfuegos, Cuba, para identificar los problemas relacionados con la calidad físico-química de las aguas y con lapresencia de fitoplancton tóxico. Se presentan los resultados de salinidad, pH, nutrientes, sólidos suspendidos,demanda bioquímica de oxígeno (DBO5) y saturación de oxígeno, obtenidos en 14 zonas de baño, en cuatrocampañas de muestreos realizadas durante el período 2008-2009. Se obtuvo que al menos uno de los criteriosde calidad físico-químicos establecidos por la NC.22: (1999) fue violado en 11 de las 14 zonas de bañoestudiadas. En el período de estudio la comunidad microfitobéntica estuvo dominada por diatomeas, lo cual estáen correspondencia con la obtención de mayores niveles tróficos promedio para el silicio (Si), respecto a losobtenidos para el nitrógeno inorgánico disuelto (NID) y el fósforo inorgánico (PI). También fueron registradasespecies tóxicas en algunas de las zonas estudiadas, las cuales incrementaron sus concentraciones al final de latemporada poco lluviosa y principio de la temporada lluviosa.
Palabras clave: playas, calidad de agua, fitoplancton, Bahía de Cienfuegos.
ABSTRACT
The physical-chemical characteristics of waters and phytoplankton in bathroom areas of Cienfuegos Bay, Cubaare studied in order to identify the problems related to the physical-chemical quality of waters and with thepresence of toxic phytoplankton. The results of salinity, pH, nutrients, suspended solids, biochemical oxygendemand (BOD5) and oxygen saturation, obtained in 14 bathroom areas, in four campaigns of samplings carriedout during the period 2008-2009, were considered. It was obtained that at least one of the physical-chemicalcriteria settled down by NC.22: (1999) was violated in 11 of the 14 studied bathroom areas. In the studyperiod, the microphytobentic community was dominated by diatoms, which is in correspondence with theobtaining of higher average trophic levels for silicon (Si), regarding the obtained levels for inorganic nitrogen(DIN) and inorganic phosphorus (IP). Toxic species were also registered in some of the studied areas, whichincreased their concentrations at the end of the not very rainy season and at the beginning of the rainy season.
Keywords: beaches, water quality, phytoplankton, Cienfuegos Bay.
Características físico-químicas de las aguas y del fitoplanctonen zonas de baño de la Bahía de Cienfuegos, Cuba (2008-2009)
INTRODUCCIÓN
Actualmente, en el mundo la mayoría de los grandesnúcleos de población mundial son costeros. Se consideraque entre el 50 y 70 % de la población actual vive enáreas costeras y se calcula que para el 2025 el 75 %de la población mundial va a vivir a menos de 60 km delmar (Moreno, 2007). Con el aumento de la población enestas zonas, se han incrementado el número de obras y
actividades realizadas en función de su bienestar, entrelas que se encuentran: marítimo-portuarias, urbanas,pesqueras, industriales y turístico-recreativas.
Pese a que las playas ubicadas dentro de las ensenadasy bahías de Cuba no figuran en el grupo de las mejorespotenciadas para el turismo, estas constituyen unos delos sitios más concurridos por la población de los territoriosdonde se ubican (Tristá et al., 2004). Por ello, el presentetrabajo tiene como objetivo estudiar las característicasfísico-químicas de las aguas y el fitoplancton en zonas de

39
baño de la Bahía de Cienfuegos. Esto permite identificarlos problemas relacionados con la calidad físico-químicade las aguas y la presencia de fitoplancton tóxico. Elestudio de la respuesta de la comunidad de fitoplancton avariables físicas y químicas es útil en la interpretación devariaciones hidrológicas en zonas costeras (Troccoli et al.,2004), debido a que la composición del fitoplancton seconsidera un bioindicador natural que responde de formarápida y compleja a fluctuaciones de las condicionesambientales (Livingston, 2001).
MATERIALES Y MÉTODOS
Se realizaron cuatro campañas de muestreo, dos de ellascorrespondientes a la temporada poco lluviosa (abril/2008 y noviembre/2009) y las restantes, corres-pondientes a la temporada lluviosa (octubre/2008 ymayo/2009). Se estableció una red de 14 estacioneslocalizadas en áreas utilizadas por la población para elbaño (Fig. 1).
Fig. 1 Red de zonas de baño muestreadas de la Bahía de Cienfuegos, Cuba
9
E1 - Playa Punta las CuevasE2 - Playa Junco SurE3 - Playa AlegreE4 - Playa Laguna del CuraE5 - Playa del MellaE6 - Playa el CuritaE7 - Playa la PuntaE8 - Playa Círculo JuvenilE9 - Playa Club CienfuegosE10 - Playa ReinaE11 - Playa ObourkeE12 - Playa Castillo-PerchéE13 - Playa Rancho ClubE14 - Playa Tetas de Tomaza
kilómetros
0 1,5 3 6
13
12
14
11
10
87 6 5
4
3 21
9
N
Las colectas de las muestras de agua se realizaronen el nivel superficial de acuerdo con las especificacionesde APHA (1998) y se utilizó para ello botellas Niskin de5 L de capacidad.
Las determinaciones de los indicadores hidroquímicosevaluados se realizaron mediante diversos métodos. Eloxígeno disuelto se analizó mediante el método Winkler,modificado por Carriet & Carpenter (1966); mientras quepara la determinación de la DBO5 se empleó el métodode incubación a 20 °C durante cinco días. Los nitratosse determinaron por espectrofotometría por reduccióncon hidracina, mientras que los nitritos y el amonio, deacuerdo con las metodologías de la UNESCO (1983). Elanálisis del ortofosfato disuelto y el silicio se realizarona partir de métodos propuestos en UNEP (1991). Lossólidos en suspensión se determinaron por gravimetría auna temperatura de 121 °C. La salinidad y la temperaturase obtuvieron in situ empleando una sonda digital modeloYSI-30, para el análisis del pH se utilizó un pH-metrodigital. La saturación de oxígeno (en porcentaje) se obtuvoaplicando la ecuación empírica propuesta por Weiss(1970):
sat. OD (%) = ODmáx./OD (1)
Para la evaluación de calidad del agua se utilizaronlos criterios establecidos en NC.22: (1999). “Lugaresde Baño en Costas y Masas de Aguas Interiores. RequisitosHigiénicos Sanitarios”. Teniendo en cuenta que esta normaestablece que los indicadores nitrogenados y de fósforo nodeben estar en niveles eutróficos, se utiliza el índice deeutrofización (IE) propuesto por Karydis et al. (1983), elcual permitió determinar el IE para el nitrógeno inorgánicodisuelto (NID) (N-NH4
+ + N-NO2– + N-NO3
–), el fósforoinorgánico (PI) (PO4
3–) y el silicio, basado en la siguienteecuación:
AxC
CIEi
loglog–
+= (2)
donde:IE: índice de eutrofización por nutriente de cada estaciónde muestreo, por campaña.A: número de estaciones de muestreo durante el períodode estudio (en este trabajo, 14).C: logaritmo de la concentración total del nutrientedurante el período de estudio, es decir, es la suma delas concentraciones xij del nutriente obtenidas en cadauna de las Ai estaciones por campaña.

40
La escala de clasificación es la siguiente: si EI < 3indica estado oligotrófico, para 3 ≤ IE ≥ 5 es mesotróficoy para IE > 5 es eutrófico.
Por la gran abundancia de las comunidades macrofito-bentónicas en las playas, se estudiaron las microalgasbentónicas, haciendo énfasis en las especies tóxicas.Se utilizaron las macroalgas como sustrato para desprenderlas microalgas bentónicas epífitas. Las macroalgas sesuspendieron en agua de mar y se agitaron fuertementepara conseguir que se desprendieran las microalgasadheridas. El extracto obtenido se filtró por una serie defiltros de abertura de malla de 250 y 150 μm para eliminardetritus y organismos de gran tamaño. Las muestras sefijaron con lugol ácido para el proceso de recuento de lasmicroalgas. Para sedimentar el material microalgal, lasmuestras se dejaron reposar por 5 días. Posteriormente,se tomó una alícuota (0,04 mL) del material sedimentado yse contaron las especies a través de una cámara de conteoRigosha y un microscopio biológico OLYMPUS BH-2 con
TABLA 1. Valores promedios e intervalos de los indicadores hidrológicos por temporada climática
aumentos de 20X y 40X. La concentración final de lasmicroalgas se expresó en número de células/g de macroalgahúmeda.
Además, para la identificación de las microalgastóxicas, las muestras se estudiaron en vivo y fijadas conformaldehído al 3 % y se utilizó el Manual microalgastóxicas de la UNESCO (Hallegraeff et al., 2003).
RESULTADOS Y DISCUSIÓN
El pH presentó valores entre 7,40 y 8,51 unidades y unvalor medio de 8,22 unidades en la temporada pocolluviosa; mientras que en la temporada lluviosa se obtuvouna variación entre 6,53 y 8,37 unidades y un valor mediode 7,86 unidades (TABLA 1). Este indicador cumple conel criterio de aceptable (6,1-8,9) establecido por laNC.22: (1999).
Indicadores Valores Intervalo Magnitud promedios
Salinidad en t. lluviosa 27,5 7,30-34,7 upsSalinidad en t. poco lluviosa 34,6 33,6-35,0 upsTemperatura en t. lluviosa 28,4 25,2-31,2 ºCTemperatura en t. poco lluviosa 26,9 25,6-28,7 ºCpH en t. lluviosa 7,86 6,53-8,37 upH en t. poco lluviosa 8,22 7,40-8,51 uOxígeno disuelto en t. lluviosa 5,95 4,87-7,31 mg.L-1
Oxígeno disuelto en t. poco lluviosa 6,46 4,66-8,86 mg.L-1
Sat. de oxígeno en t. lluviosa 89,8 72,7-117,8 %Sat. de oxígeno en t. poco lluviosa 98,8 69,7-137,1 %DBO5 en t. lluviosa 2,74 1,60-3,60 mg.L-1
DBO5 en t. poco lluviosa 2,16 0,20-4,00 mg.L-1
Fosfato en t. lluviosa 1,78 0,32-9,84 μmol L-1
Fosfato en t. poco lluviosa 3,5 0,5-35,1 μmol L-1
N-amoniacal en t. lluviosa 6,44 < 3,21-84,1 μmol L-1
N-amoniacal en t. poco lluviosa 3,42 < 3,21-7,57 μmol L-1
N-nitrito en t. lluviosa 0,26 0,07-0,43 μmol L-1
N-nitrito en t. poco lluviosa 0,14 0,07-0,29 μmol L-1
N-nitrato en t. lluviosa 1,99 < 0,43-22,4 μmol L-1
N-nitrato en t. poco lluviosa 1,11 < 0,43-10,8 μmol L-1
Si-SiO2 en t. lluviosa 49,0 4,47-127 μmol L-1
Si-SiO2 en t. poco lluviosa 15,1 3,4-33,1 μmol L-1
Sólidos suspendidos en t. lluviosa 16,1 4,0-93,7 mg.L-1
Sólidos suspendidos en t. poco lluviosa 6,15 1,70-29,90 mg.L-1

41
La temperatura de las aguas osciló entre 25,2y 31,2 oC durante la temporada lluviosa, la cual corres-ponde a los meses más cálidos; sin embargo, en latemporada poco lluviosa el intervalo de valores fueligeramente menor (25,6-28,7 oC).
La salinidad, mostró en la temporada poco lluviosa,un valor medio de 34,59 ups y concentraciones con unintervalo entre 33,6 y 35,0 ups, mientras que en latemporada lluviosa, el valor medio fue de 27,54 ups y unintervalo entre 7,3 y 34,7 ups (TABLA 1). Los valoresmínimos correspondieron a la estación próxima a lasTetas de Tomaza, en la campaña de octubre de 2008. Sibien, para este indicador, la NC.22: (1999) establece quesus valores no deben ser inferiores de 36 ups, es válidotener en cuenta que las áreas evaluadas se encuentrandentro de una bahía semicerrada con característicasestuarinas según Tomczak & García (1975), la cualpresenta diferencias ecológicas con respecto a sistemasabiertos, y la falta de normativas para sistemas estuarinospuede estar propiciando una evaluación más rigurosa.
Las concentraciones de oxígeno disuelto obtenidasestuvieron entre 4,66 y 8,86 mg.L–1 en la temporada poco
lluviosa, mientras que en la temporada lluviosa estuvieronentre 4,87 y 7,31 mg.L–1. La evaluación de los nivelesde calidad de las aguas según los criterios para losporcentajes de saturación de oxígeno (≥ 70 %), mostrócalidad aceptable, excepto en la zona de baño de O’Bourke,en la campaña de noviembre de 2009, con un valor mínimode 69,7 %.
La demanda bioquímica de oxígeno (DBO5), comoindicador de los niveles de materia orgánica, presentóen la temporada poco lluviosa un valor medio 2,16 mg.L–1
y una variación entre 0,2 y 4,0 mg.L–1, cumpliéndoselas condiciones requeridas para las zonas de baño(≤ 3,0 mg.L–1) en el 90 % de las estaciones mues-treadas e incumpliéndose para las zonas de PlayaAlegre, Playa Laguna del Cura y el Curita. Sinembargo, en la temporada lluviosa, se obtuvo un valormedio 2,74 mg.L–1 y una variación entre 1,6 y 3,6 mg.L–1,obteniéndose en cuatro de las estaciones valoressuperiores al límite aceptable, estas fueron: PlayaLaguna del Cura, Playa del Círculo Juvenil, así comolas zonas de baño próximas al Castillo-Perché y a lasTetas de Tomaza (TABLA 2).
TABLA 2. Estaciones con valores superiores a los recomendados por la NC.22: (1999) para NI, PI, % sat. O, DBO5 y pH
Los niveles de nitrógeno inorgánico (NID) mos-traron valores promedios de IE correspondientes aniveles oligotróficos en ambas campañas de latemporada poco lluviosa; mientras que en la temporadalluviosa, estas fueron oligo y mesotróficos (IE < 5).
Solo se vulneró el criterio normado de calidad en laestación de Reina, al obtenerse condiciones eutróficasen base al NID en la campaña de mayo/2009,reflejadas a través de los valores máximos regis-trados en dicha campaña (Fig. 2).
Estaciones IE (NI) IE (PI) % Sat. O DBO5 pH
E1 – Playa Punta las Cuevas
E2 – Playa Junco Sur
E3 – Playa Alegre x
E4 – Playa Laguna del Cura x
E5 – Playa del Mella x
E6 – Playa el Curita x
E7 – Playa la Punta x
E8 – Playa Círculo Juvenil x
E9 – Playa Club Cienfuegos
E10 – Playa Reina x
E11 – Playa O’Bourke x
E12 – Playa Castillo-Perché x
E13 – Playa Rancho Club
E14 – Playa Tetas de Tomaza x
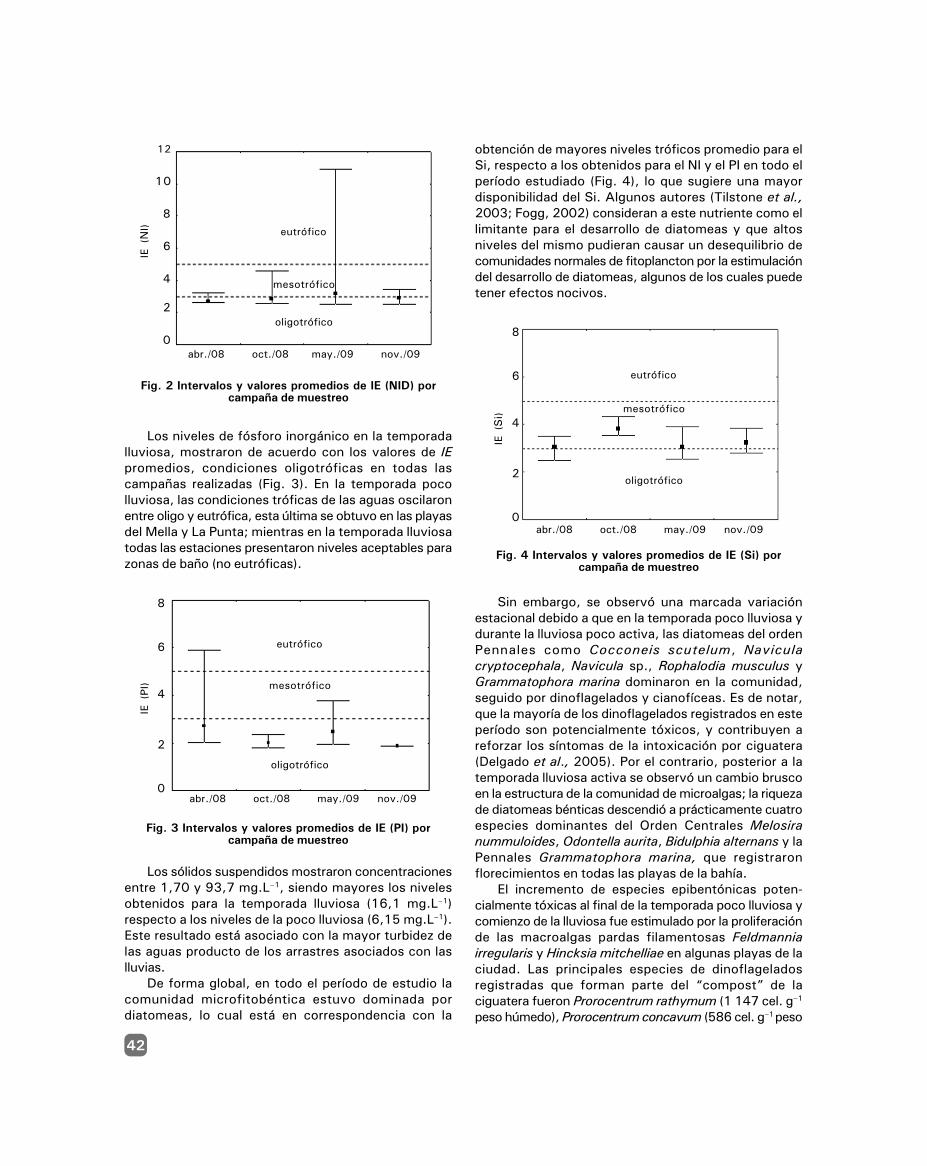
42
Fig. 2 Intervalos y valores promedios de IE (NID) porcampaña de muestreo
Los niveles de fósforo inorgánico en la temporadalluviosa, mostraron de acuerdo con los valores de IEpromedios, condiciones oligotróficas en todas lascampañas realizadas (Fig. 3). En la temporada pocolluviosa, las condiciones tróficas de las aguas oscilaronentre oligo y eutrófica, esta última se obtuvo en las playasdel Mella y La Punta; mientras en la temporada lluviosatodas las estaciones presentaron niveles aceptables parazonas de baño (no eutróficas).
Fig. 3 Intervalos y valores promedios de IE (PI) porcampaña de muestreo
eutrófico
8
6
4
2
0
IE (
PI)
abr./08 oct./08 may./09 nov./09
mesotrófico
oligotrófico
Los sólidos suspendidos mostraron concentracionesentre 1,70 y 93,7 mg.L–1, siendo mayores los nivelesobtenidos para la temporada lluviosa (16,1 mg.L–1)respecto a los niveles de la poco lluviosa (6,15 mg.L–1).Este resultado está asociado con la mayor turbidez delas aguas producto de los arrastres asociados con laslluvias.
De forma global, en todo el período de estudio lacomunidad microfitobéntica estuvo dominada pordiatomeas, lo cual está en correspondencia con la
obtención de mayores niveles tróficos promedio para elSi, respecto a los obtenidos para el NI y el PI en todo elperíodo estudiado (Fig. 4), lo que sugiere una mayordisponibilidad del Si. Algunos autores (Tilstone et al.,2003; Fogg, 2002) consideran a este nutriente como ellimitante para el desarrollo de diatomeas y que altosniveles del mismo pudieran causar un desequilibrio decomunidades normales de fitoplancton por la estimulacióndel desarrollo de diatomeas, algunos de los cuales puedetener efectos nocivos.
Fig. 4 Intervalos y valores promedios de IE (Si) porcampaña de muestreo
Sin embargo, se observó una marcada variaciónestacional debido a que en la temporada poco lluviosa ydurante la lluviosa poco activa, las diatomeas del ordenPennales como Cocconeis scutelum, Naviculacryptocephala, Navicula sp., Rophalodia musculus yGrammatophora marina dominaron en la comunidad,seguido por dinoflagelados y cianofíceas. Es de notar,que la mayoría de los dinoflagelados registrados en esteperíodo son potencialmente tóxicos, y contribuyen areforzar los síntomas de la intoxicación por ciguatera(Delgado et al., 2005). Por el contrario, posterior a latemporada lluviosa activa se observó un cambio bruscoen la estructura de la comunidad de microalgas; la riquezade diatomeas bénticas descendió a prácticamente cuatroespecies dominantes del Orden Centrales Melosiranummuloides, Odontella aurita, Bidulphia alternans y laPennales Grammatophora marina, que registraronflorecimientos en todas las playas de la bahía.
El incremento de especies epibentónicas poten-cialmente tóxicas al final de la temporada poco lluviosa ycomienzo de la lluviosa fue estimulado por la proliferaciónde las macroalgas pardas filamentosas Feldmanniairregularis y Hincksia mitchelliae en algunas playas de laciudad. Las principales especies de dinoflageladosregistradas que forman parte del “compost” de laciguatera fueron Prorocentrum rathymum (1 147 cel. g–1
peso húmedo), Prorocentrum concavum (586 cel. g–1 peso
eutrófico
8
6
4
2
0
IE (
Si)
abr./08 oct./08 may./09 nov./09
mesotrófico
oligotrófico
12
10
8
6
4
2
0
IE (
NI)
abr./08 oct./08 may./09 nov./09
eutrófico
mesotrófico
oligotrófico

43
húmedo), Prorocentrum lima (290 cel. g–1 peso húmedo).Otras especies de dinoflagelados tóxicos registrados enbajas concentraciones fueron Prorocentrum belizeanum,Prorocentrum emarginatum y Ostreopsis ovata. Laespecie Gambierdiscus cf. caribaeus, potencial agentede la ciguatera, solo se registró en la playa Rancho Club,en las afueras de la bahía y en bajas concentraciones. Esde significar, la relativa abundancia en este período deciertas cianofíceas tóxicas como Lyngbya majuscula(1 583 cel. g–1 peso húmedo) y Anabaena sp. (75 cel g–1
peso húmedo). De estas dos especies, la primera producetoxinas neurotóxicas (Nogle et al., 2001) y derma-totóxicas por contacto directo, y pudiera ocasionar dañosa bañistas en las mencionadas playas; mientras que lasegunda produce toxinas hepatotóxicas.
CONCLUSIONES
De manera general, se encontró, al menos, una violaciónde los criterios de calidad físico-químicos normados,en 11 de las 14 zonas de baño muestreadas.
En todo el período de estudio la comunidadmicrofitobéntica estuvo dominada por diatomeas, lo cualestá en correspondencia con la obtención de mayoresniveles tróficos promedio para el Si, respecto a losobtenidos para el NI y el PI. También fueron registradasespecies tóxicas en algunas de las zonas estudiadas, lascuales incrementaron sus concentraciones al final de latemporada poco lluviosa y principio de la temporadalluviosa.
AGRADECIMIENTOS
A los especialistas y técnicos del laboratorio del CEACque contribuyeron a este estudio mediante la realizaciónde los muestreos y ensayos analíticos. Al ProyectoRegional del OIEA RLA/7/-014 (Diseño e implementaciónde un sistema de alerta temprana y evaluación detoxicidad de FAN en la región del Caribe, aplicandotécnicas nucleares avanzadas, evaluaciones radioecotoxi-cológicas y bioensayos).
REFERENCIAS
APHA (1998). Standard methods for the examination ofwater and wastewater. American Public HealthAssociation (20th edition, 210 pp.).
Carriet, D. E. & Carpenter, J. H. (1966). Comparisonand evaluation of the Winkler method for determining
dissolved oxygen in sea water. Jour. Mar. Res., 24(3), 286-318.
Delgado, G., Popowski, G., García, C., Lagos, N. &Lechuga, C. (2005). Presence of DSP-Toxins inProrocentrum lima (Ehrenberg) Dodge in Cuba. Rev.Inv. Mar., 26 (3), 229-234.
Fogg, G. E. (2002). Harmful algae—a perspective.Harmful algae 1, pp. 1-4.
Hallegraeff, G., Anderson, D. & Cembella, D. (2003).Manual on Harmful Marine Microalgae. IOC Manualsand Guides No. 33, UNESCO.
Karydis, M., Ignatiades, L. & Moschopoulou, N. (1983).An Index associated with Nutrient Eutrophication inthe Marine Environment. Estuarine, Coastal and ShelfScience, 16, 339-344.
Livingston, R. J. (2001). Eutrophication processes incoastal systems: origin and sucession of planktonblooms and effects on secondary production inGulf Coast estuaries). USA: Center for AquaticResearch and Resource Management. Florida StateUniversity.
Moreno, I. (2007). Manejo integral costero. Por una costamás ecológica, productiva y sostenible (1ra. ed.).Universitat de les Illes Balears.
Nogle, L., Okino, T. & Gerwick, W. (2001). Antillatoxin B, aneurotoxic lipopeptide from the marine cyanobacteriumLyngbya majuscula. J. Nat. Prod., 59, 27-30.
NC.22: (1999). “Lugares de Baño en Costas y Masas deAguas Interiores. Requisitos Higiénicos Sanitarios”.Oficina Nacional de Normalización. Ciudad de La Habana.
Tilstone, G. H., Figueiras, F., Lorenzo, L. & Arbones, B.(2003). Phytoplankton composition, photosynthesisand primary production during different hydrographicconditions at the Northwest Iberian upwellingsystem. Marine Ecology Progress Series, 252,89-104.
Tomczak, M. & García, C. (1975). A numerical model ofcirculation in Cienfuegos Bay, Cuba. Estuarine andCoastal Marine Science, 3, 391-412.
Tristá, E., Juanes, J. L., Salazar, H. & Hernández, K.(2004). Evaluación de las actuaciones costerasaplicadas en las playas interiores de Cuba. Rev. Inv.Pesq., No. 1, Edición Especial.
Troccoli, G. L., Herrera-Silveira, J. A. & Comín, F. A.(2004). Structural variations of phytoplankton in thecoastal seas of Yucatan, Mexico. Hidrobiología, 519,85-102.
UNEP (1991). Standard chemical methods for marineenvironmental monitoring. Manual and Guides,Intergovernmental Oceanographic Committee, No. 12.
Weiss, R. (1970). The solubility of nitrogen, oxygen, andargon in water and seawater. Deep-Sea Res., 17,721-735.

Revista Cubana de Investigaciones PesquerasEnero-diciembre, 2012, vol. 29, NO. 1, ISSN 0138-8452, pp.
44
Composition and quality of lipid fraction of waste tilapia silage
José E. Llanes Iglesias,1 José Toledo Pérez,1
Lourdes Savón Valdés2 y Odilia Gutiérrez Borroto2
1 Empresa de Desarrollo de Tecnologías Acuícolas. Carretera Central km 20 ½,Loma de Tierra, Cotorro, La Habana, Cuba
2 Instituto de Ciencia Animal. Apartado Postal 24, San José de las Lajas, Mayabeque. Cuba
RESUMEN
El objetivo del presente trabajo fue determinar la composición y los índices de identificación y calidad de lafracción lipídica de los ensilajes (químico y biológico) de residuos del fileteado de tilapias. El ensilaje químico sepreparó con 2 % de ácido sulfúrico 98 % y 1 % de ácido fórmico (p/v) y el biológico con 15 % de miel de cañay 3 % de yogurt (Lactobacillus acidophillus) (p/p) y se almacenaron por 30 días. Los resultados mostraron queambos ensilajes poseen todos los ácidos grasos presentes en la materia prima y el 60 % fueron insaturados. Eloleico (18:1ω9) fue el de mayor concentración (30,2 %) seguido del palmitico (18:0) (27,4 %) y linoleico(18:2ω6) (16,8 %). Los índices de iodo (878 mg/g), saponificación (194,5 mg KOH/g) y ácidos grasos libres(3,4 %) fueron compatibles con los datos de la literatura. Los valores de peróxidos resultaron bajos en losensilajes e indicaron una menor tendencia a la oxidación en los residuos fermentados.
Palabras clave: ácidos grasos, ensilajes de pescado, fracción lipídica.
ABSTRACT
To determine the composition and indexes of identification and quality the lipid fraction of the silage (chemicaland biological) from tilapia filleting waste was the aim of the present work. The chemical silage was preparedwith 2 % of sulfuric acid 98 % and 1 % formic acid (w/v) and the biologycal silage with 15 % of sugar canemolasses and 3 % of yogurt (Lactobacillus acidophilus) (w/w) and stored for 30 days. The results showed thatboth silages have oll fatty acids from raw material, with 60 % value of unsaturated acids. The oleic acid(18:1ω9) was higher concentration (30,2 %) followed by the palmitic acid (18:0) (27,4 %) and linoleic acid(18:2ω6) (16,8 %). The indexs of iodo (878 mg/g), saponification (194,5 mg KOH/g) and free fatty acids (3,4 %)were compatibles with the data in the literature. The value of peroxid resulted low and indicated fermented wastepresented fewer tendencies to oxidize.
Keywords: fatty acid, fish silage, lipid fraction.
Composición y calidad de la fracción lipídica de los ensilajesde residuos de tilapias
44-48
INTRODUCCIÓN
El deseo de producir dietas con proteína de origen animalpara el cultivo de peces condujo al desarrollo de diferentestecnologías que permitieron reciclar los desechos bioló-gicos. El aprovechamiento de los residuos pesqueros portecnologías simples y relativamente económicas como elensilaje (conservación en medio ácido), fue la opción quepermitió mejorar la calidad de las dietas vegetales para lapiscicultura de agua dulce en Cuba.
Según la NRC (1993), la composición química es uno delos indicadores fundamentales para evaluar el valornutricional de ingredientes o dietas. Diferentes trabajos(Gerón et al., 2007; Llanes et al., 2011) consignaron quelos ensilajes de residuos del fileteado de tilapias presentaronaltos niveles de proteínas (hasta 45,9 g/100 g materiaseca) con una calidad nutricional acorde con los reque-rimientos de aminoácidos que estableció la NRC (1993)para tilapia del Nilo Oreochromis niloticus.
Por esto, el objetivo de este trabajo fue determinarla composición y los índices de identificación y calidad

45
de la fracción lipídica de los ensilajes (químico y biológico)de residuos del fileteado de tilapias.
MATERIALES Y MÉTODOS
El estudio se realizó en el Laboratorio de Nutrición de laEmpresa de Desarrollo de Tecnologías Acuícolas en LaHabana. Se utilizaron residuos del fileteado de tilapias,los que se molieron en una máquina de carne JAVAR 32.Para preparar el ensilaje químico (EQ) se utilizó 2 % deácido sulfúrico 98 % (p/v) y 1 % de ácido fórmico (p/v)adquiridos en la firma comercial MERCK y el ensilajebiológico (EBL) con 15 % de miel final (p/p) y 3 % deyogurt, Lactobacillus acidophilus (1 x 107 UFC/g) comocultivo de bacterias ácido-lácticas (Llanes et al., 2011).Los silos se realizaron por triplicados y se almacenaronen tanquetas plásticas con tapas por 30 días.
Los aceites de los residuos frescos y ensilajes seextrajeron por el método de Bligh & Dyer (1959). Losácidos grasos (AG) se determinaron en un cromatógrafode gases Hewlett Packard modelo HP6890 GC System,equipado con un detector FID y una columna SP2380(30 m de largo, 0,25 mm diámetro interno, 0,20 μm
espesor de film), con helio como gas transportador,después de la esterificación en metanol/ácido sulfúrico,según Hartman & Lago (1973).
El valor peróxido se determinó iodométricamente porla técnica 965.33, el contenido de ácidos grasos libresse expresó en porcentaje de ácido oleico y se determinópor la técnica 940.28, el índice de Iodo por la 920.159 yel índice de saponificación por la 920.160, descritas enlos procedimientos de la AOAC (1995).
Los valores promedios de los indicadores de identi-ficación y calidad de los aceites se analizaron medianteun análisis de varianza de clasificación simple por mediodel software estadístico INFOSTAT versión 1.0 (Balzariniet al., 2001).
RESULTADOS Y DISCUSIÓN
La composición de ácidos grasos (TABLA 1) reveló quelos silos de residuos del fileteado de tilapia poseen todoslos AG prevalecientes en la materia prima y muestranla buena calidad de la fracción lipídica aunado por lasaltas concentraciones de ácidos grasos insaturados,importante en la nutrición de peces.
TABLA 1. Composición de ácidos grasos (g/100 g de grasa) de los residuos de tilapias y sus ensilajes
Nombre Fórmula Residuos Ensilaje Ensilaje frescos químico biológico
Ácido mirístico C14:0 3,32 3,56 3,26Ácido n-pentadecanoico C15:0 0,66 0,59 0,51Ácido palmítico C16:0 26,52 29,19 25,53Ácido palmitoleico C16:1 8,46 8,94 8,42Ácido margárico C17:0 0,72 0,56 0,90Ácido esteárico C18:0 5,93 7,01 5,82Ácido oleico C18:1 (n-9) 31,15 28,96 31,49Ácido linoleico C18:2 (n-6) 15,20 16,32 17,38
α Ácido linolénico C18:3 (n-3) 2,11 2,12 2,76
Ácido linolénico C18:3 (n-6) 0,62 0,71 0,80Ácido araquídico C20:0 1,12 1,30 0,98Ácido di-homolinolenico C20:3 (n-3) 0,58 0,24 0,79Ácido eicosapentanoico C20:5 (n-3) < 0,01 < 0,01 < 0,01Ácido docosahexaenoico C22:6 (n-3) 0,26 0,12 0,23
Ácidos grasos saturados - 38,27 42,21 37,00Ácidos grasos insaturados - 58,39 57,42 62,63
Nota: Los valores presentados corresponden a una muestra/ingrediente.

46
Se identificaron 14 AG y los principales fueron el oleico(30,22 %), palmítico (27,36 %), linoleico (16,85 %) ypalmítoleico (8,68 %), los cuales representaron el 83 %del total.
Las concentraciones de AG insaturados fueron altas(60 %) en ambos ensilajes y el material crudo, destacán-dose el ácido oleico, que es muy abundante en el aceitede oliva (hasta 75 %), pero igual que en otros aceites deorigen animal nunca excede del 40 %.
De los AG saturados, el palmítico presentó el valormás alto similar a lo reportado en otros ensilajes deresiduos de tilapias (Gerón et al., 2007; Vidotti et al.,2011). Este AG está ampliamente distribuido en lanaturaleza y presente a lo largo de toda la cadenaalimentaria. Además, forma parte de la composición denumerosos aceites de animales y plantas acuáticas,actuando como precursor de AG insaturados y saturadosde cadenas largas.
La proporción de AG saturados e insaturados difierensegún la materia prima que se utilice en su elaboración,de tal forma que al evaluar el ensilaje químico de faunaacompañante Espíndola-Filho (2002) encontró mayorcantidad de AG saturados que insaturados y lo atribuyóa la presencia de camarones Farfantepenaeus brasiliensisen el material que ensiló. Resultados que coincidieroncon Vidotti et al. (2008) con residuos de ranas. Sinembargo, con vísceras de tilapias estos mismos autoresreportaron un comportamiento inverso, similar a loobtenido por Santana-Delgado et al. (2008) con macarelas(Scomberomorus maculatus) fuera de talla comercial.
En relación con los AG de la serie linolénica (ω-3) ylinoleica (ω-6), se encontró mayores concentraciones deω-6 que se puede relacionar con la composición de lasdietas de las tilapias que contienen ingredientes derivadosde oleaginosas (harina y aceite de soya), ricos en esteácido graso. Mientras, los ω-3 permiten un grado mayorde insaturación, requisito indispensable para una mayorfluidez, flexibilidad y permeabilidad de las membranas abajas temperaturas, por lo que se encuentran en mayorconcentración en peces marinos y de aguas frías por lasbajas temperaturas del medio (Bureau, 2004).
De ahí que los aceites extraídos de peces marinos,hígado de bacalao y sardinas fueron fuentes ricas enAG ω-3; cuyos contenidos en ácidos eicosapentaenoico(20:5 ω-3, EPA) y docosahexanoico (22:6 ω-3, DHA)representaron más del 20 % del total de AG (Bureau,2004). Así mismo, Vidotti et al. (2002) reportaron quelos silos de pescados marinos presentaron elevadosniveles de ácido erúcico 22:1ω-9 (6,7 %) y DHA (11,1 %)en relación con los de agua dulce (0,3 y 1,3 % respecti-vamente), resultados que se correspondieron con este
trabajo al utilizar residuos de tilapias con apenas trazas(menor que 0,3 %) para los ácidos grasos poliinsaturados.
La esencialidad de los ω-6 para tilapia se estudió enmuchas especies (NRC, 1993), pero aún está por deter-minar la importancia de los ácidos grasos ω-3 en la dietade estos peces. Un trabajo de Lim et al. (2008) evidencióque las tilapias del Nilo tienen requerimientos para ambos,aunque los ω-6 parecieron tener mejor efecto promotordel crecimiento que los ω-3.
En peces se demostró la competencia entre losAG ω-6 y ω-3 como sustrato de varias enzimasimplicadas en el metabolismo de los lípidos, de ahí laimportancia en el equilibrio ω-6/ω-3 en las dietas deengorde para tilapias. Además, la disminución de lasconcentraciones dietéticas de estos AG puede traermalformaciones y alteraciones en la calidad de las ovasy las larvas (Bureau, 2004), baja respuesta del sistemainmunológico (Lim et al., 2008) y afectar el valornutricional del pescado para la alimentación humana.
La compañía internacional Aller Aqua Group(www.aller-aqua.com) que produce alimentos comercia-les para los cultivos intensivos de peces en Europa,incluye aceite de pescado (rico en ω-3) en las dietas detilapias. Por tanto, al incluirse el ensilaje de residuos detilapia cuyo contenido de ácido linoleico (C18:2 ω6) fue16,85 %, se puede garantizar el suministro de lasnecesidades diarias (0,5-1 %) que estableció la NRC(1993) para tilapias del Nilo, así como el aporte de unapequeña cantidad de ω-3.
Los aceites de soya y girasol son normalmente losque se adicionan a las raciones de los peces omnívoroscon la finalidad de aumentar el valor calórico de la ración.Así, el aceite de los ensilajes de pescado se podría utilizaren sustitución de estos aceites principalmente por el bajocosto y contenido de AG insaturados disminuyendo lacompetencia con el consumo humano. De hecho, Herteset al. (2010) reportaron una digestibilidad de 76,30 %para el aceite de soya en juveniles de tilapias del Nilo, queresultó inferior a la que informaron Llanes et al. (2011)para la fracción lipídica de ensilajes de residuos de tilapia(87,1 %) en alevines de tilapias rojas (Oreochromis sp.).
El iodo y la saponificación de la fracción lipídica (TABLA 2)como índices del grado de insaturación de los AG y materiasaponificable respectivamente, presentaron composicio-nes afines para ambos silos, dado que se utilizó la mismamateria prima y no hubo influencia de las metodologías deensilaje. Los valores encontrados en este estudio soncaracterísticos de peces de agua dulce y coinciden conlos informados por Vidotti & Sampaio (2006) en aceitesextraídos de silos de residuos de tilapias (880 mg/g deiodo y 197 mg KOH/g de saponificación).

47
TABLA 2. Índices de identificación y calidad del aceite de los ensilajes de residuos de tilapias
Índices de calidad Ensilaje químico Ensilaje biológico EE(±) Sig
Índice de iodo (mg/g) 871 887 ± 0,02Índice de saponificación (mg KOH/g) 195 194 ± 0,02Ácidos grasos libres (g/100 g) 2,8 3,9 ± 0,07 *Índice de peróxido (meq O2/kg) 4,9 0,9 ± 0,84 **Olor No rancio No rancio-
**p < 0,01 *p < 0,05
Los índices de calidad (ácidos grasos libres, AGL yperóxido) variaron en función del procesamiento (TABLA 2).Los residuos fermentados sufrieron un proceso hidroliticomás intenso que resultó con un mayor contenido de AGL yse puede relacionar con una ruptura más efectiva de lascadenas de triglicéridos por la acción de los microorganismosdel medio que libera grandes cantidades de AG que estándispersos en toda la fracción lipídica, mientras los acidi-ficados (EQ) a pesar que hubo una mayor ruptura de lasestructuras celulares y micelas lipídicas por la adición directade los ácidos, los triglicéridos se preservaron mejor por elbajo valor del pH.
Según Bureau (2004), los aceites de pescado debuena calidad deben tener valores de peróxidos menoresque 10 meq O2/kg de aceite. De ahí que los valores quese encontraron en este trabajo (TABLA 2) fueronsatisfactorios y se puede atribuir a que las grasas enlos silos están inmersas en el agua y por tanto se reduceel contacto con el dioxígeno del aire. Las diferenciassignificativas que se encontraron (p < 0,01) entre lossilos evidencia que las condiciones de anaerobiosis delmedio (EBL) retardan el proceso oxidativo de los AGpoliinsaturados. Estos resultados sugieren que losensilajes por fermentación láctica son menossusceptibles a los procesos oxidativos que por acidi-ficación.
CONCLUSIONES
1. Los ensilajes químico y biológico de residuos delfileteado de tilapias presentaron todos los ácidosgrasos prevalecientes en la materia prima, predomi-nando los insaturados (60 %), importantes para lanutrición de peces.
2. Los ácidos grasos de mayor concentración fueron eloleico (C18:1ω9), palmítico (C16:0) y linoleicoC18:2ω6).
3. Los índices de calidad del aceite de los silos deresiduos de tilapia almacenados durante 30 díasindicaron que están aptos para la alimentación depeces comerciales.
4. Los aceites de los residuos pesqueros fermentadospresentaron menor riesgo de procesos oxidativosque los acidificados.
REFERENCIAS
AOAC (1995). Official Methods of Analysis (15 ed.).Washington. Association of Official AnalyticalChemists. Official Method 940.28. Fatty Acids (Free)in Crude and Refined Oils. Titration Method.
AOAC (1995). Official Method 965.33. Peroxide Valueof Oils and Fats. Titration Method.
AOAC (1995). Official Method 920.159. IodineAbsortion Number of Oils and Fats. Wijs Method.
AOAC (1995). Official Method 920.160. SaponificationNumber (Koettstorfer Number) of Oils and Fats.Titratimetric Method.
Balzarini, M., Casanoves, F., Di Rienzo, J. A., González,I. A., Robledo, C. W. & Tablada, M. E. (2001).Software estadístico INFOSTAT. Manual de usuario,Versión 1. Córdoba. Argentina.
Bligh, E. G. & Dyer, W. J. (1959). A rapid method oftotal lipid extraction and pirification. Can. J. Biochem.Physiology, 37 (8), 911-917.
Bureau, D. P. (2004). Animal fats as aquaculture feedingredients. International AQUAFEED, 7 (1), 33-35.
Espíndola Filho, A. (2002, agosto). Aprovechamientosde residuos sólidos de peces, camarones y bivalvoscomo ingredientes de raciones para acuicultura.Ponencia presentada en el XVIII Congreso Brasileirode Ciencias y Tecnologías de Alimentos. Portoalegre-RS, Brasil.
Gerón, L, J., Zeoula, L., Meire, R., Matsushira, M.,Kasama, R., Ferreira, S. & Fereli, F. (2007). Chemical

48
characterization, dry matter and crude proteinruminal degradability and in vitro intestinal digestionof acid and fermented silage from tilapia filletingresidue. Animal Feed Science and Technology, 136,226-239.
Hartman, L. & Lago, R. A. (1973). A rapid Preparationof fatty acid ethy esters from lipids. LaboratoryPractice, 22 (8), 475-476.
Hertes, D., Rogério, W., Anne, L., Andressa, T., Wolff,G. & Feiden, A. (2010, septiembre). Energía digestibledel aceite de soya para tilapia del Nilo (Oreochromisniloticus) en fase de terminación. Ponenciapresentada en el II Simposio Nacional de Ingenieríade Pesca y XII Semana Académica de Ingeniería dePesca. Sao Paolo, Brasil.
Llanes, J., Toledo, J., Savón, L. & Gutiérrez, O.(2011). Evaluación nutricional de ensilajes deresiduos pesqueros para la alimentación de tilapiasrojas (Oreochromis mossambicus x O. niloticus).Revista Cubana de Investigaciones Pesqueras, 28(2), 10-14.
Lim, C., Yildirim-Aksoy, M., Allen Davis, D. & Klesius,P. (2008). Effects of varied dietary lipid sourcestested in tilapia study. GLOBAL AQUACULTUREADVOCATE, May/June, 68-70.
NRC (1993). Nutrient Requirement of Fish. Committeeon Animal Nutrition, Board of Agriculture, National
Research Council. Washington, D.C.: NationalAcademic Press.
Santana-Delgado, H., Avila, E. & Sotelo, A. (2008).Preparation silage from Spanish mackerel(Scomberomorus maculates) and its evaluation inbroiler diets. Animal Feed Science and Tecnology,141, 129-132.
Vidotti, R. M., Macedo, E. M. & Carneiro, D. J. (2002,agosto): Producción y caracterización de la fracciónlipídica de ensilajes de pescados. Ponencia presentadaen el XVIII Congreso Brasileiro de Ciencias yTecnologías de Alimentos. Porto alegre, Brasil.
Vidotti, R. M. & Sampaio, G. S. (2006). Producáo ecaracterizacáo de silagem, farinha e oleo de tilapia edua utilizacao na alimentacao animal. Disponible en:www.pesca.sp.gov.br
Vidotti, R. M., Sampaio, G., Verardino, M., Alves deSouza, M. & Santa Rosa, M. J. (2008, Julho).Produção e caracterização de silagens ácidas deresíduos do processamento de rãs e vísceras detilápias. Ponencia presentada en la 45a Reuniao Anualda Sociedade Brasileira de Zootecnia. Lavras, Brasil(CD-ROM).
Vidotti, R. M., Bertoldo, M. T. & Sampaio, G. (2011).Characterization of the oils present in acid andfermented silages produced from Tilapia filletingresidue. Rev. Bras. Zootec, 40 (2), 240-244.

49
Revista Cubana de Investigaciones PesquerasEnero-diciembre, 2012, vol. 29, NO. 1, ISSN 0138-8452, pp. 49-53
Polycyclic aromatic hydrocarbons (PAHs) in green mussel of the Bay of Cienfuegos
Luisa Vega Bolaños,1 Roberto Castelo Baez,2 José A. Arias Verdes3 y Ramón Sersa Espinosa1
1 Instituto de Nutrición e Higiene de los Alimentos (INHA),E-mail: [email protected]
2 Centro de Investigaciones Pesqueras. Ave. 5ta. y calle 246, Santa Fe, Playa,La Habana, Cuba, CP 19100, Teléfono: (537) 209-7852
3 Centro de Ingieneria y Manejo Ambiental de Bahías y Costas (CIMAB)
RESUMEN
Los hidrocarburos aromáticos policíclicos (HAPs) son contaminantes alimentarios y su toxicidad se debe a quealgunos de ellos son conocidos cancerígenos humanos, clasificados en el grupo dos de la Agencia Internacionalde Investigación sobre el Cáncer (IARC). La determinación de los niveles de estas sustancias en los alimentos,especialmente en productos de la pesca, se ha incrementado en los últimos años por diversas causas. El objetivode este estudio fue la determinación de los hidrocarburos aromáticos policíclicos (HAPs) en mejillones (Pernaviridis) de la Bahía de Cienfuegos, 50 especímenes de mejillón con dimensiones entre 80-100 mm de largoantero-posterior de concha, se homogeneizaron y se tomó una muestra de 5 g de peso fresco, se trataron conn-hexano y saponificaron con KOH-metanol, las fracciones obtenidas fueron concentradas y purificadas encolumna de sílice-alúmina; y analizadas mediante Cromatografía Gaseosa con detector de ionización de llama,utilizando surrogados y estándar interno isotópicos para la cuantificación. Se detectaron 14 HAPs y laconcentración total de HAPs en los mejillones fue de 1 171,90 μg/kg (peso fresco), correspondiéndole losvalores más altos a: fenantreno (264,01 μg/kg), fluoranteno (246,59 μg/kg) y criseno (148,19 μg/kg), entanto los valores de benzo(a)pireno fueron de 25,88 µg/kg, valor este muy superior al límite máximo permitidode este compuesto en moluscos bivalvos (10 μg/kg peso fresco). Estos resultados indican una alta contaminaciónen HAPs en la zona de estudio, lo cual debe estar influenciado por la urbanización/industrialización de las costasde esta bahía.
Palabras clave: HAPs, mejillón verde, fenantreno, fluoranteno, criseno y benzo(a)pireno.
ABSTRACT
Polycyclic Aromatic hydrocarbons (PAHs) are polluting alimentary and their toxicity is due to that some of themare known cancerigenic human, classified in two groups of the International Agency of Investigation on theCancer (IARC). The determination of the levels of these substances in foods, especially in products of thefishing, it has been increased in the last years by diverse causes. The objective of this study was the determinationof the PAHs in mussels of Bay Cienfuegos, 50 mussels specimens with dimensions among 80-100 mm of long,they were homogenized and I take a sample of 5 g of fresh weight they talked to n-hexano and saponificationwith KOH-methanol, the obtained fractions were concentrated and purified in silica-alumina column; and determinedby gas chromatography with detecting of flame ionization, using surrogates and standard internal isotopic forthe quantification. 14 PAHs and the total concentration of PAHs were detected in the mussels it wasof 1 171,90 μg/kg, corresponding him the highest values to; phenanthrene (264,01 μg/kg), fluoranthene(246,59 µg/kg) and chrysene (148,19 µg/kg), as long as the benzo(a)pyrene values were of 25,88 μg/kg,value this very superior one to the allowed maximum limit of this compound in bivalve molluscs (10 μg/kg freshweight). These results indicate a high contamination with PAHs, which should be influenced by the urbanization/industrialization of the costs of this bay.
Keywords: PAHs, green mussel, phenanthrene, fluoranthene, chrysene and benzo(a)pyrene.
Hidrocarburos aromáticos policíclicos (HAPs) en mejillón verde(Perna viridis) de la Bahía de Cienfuegos

50
INTRODUCCIÓN
Los hidrocarburos aromáticos policíclicos (HAPs) sonmoléculas de dos o más anillos aromáticos fusionados,de baja solubilidad en agua y elevada en disolventesorgánicos y con reconocida capacidad mutagénica ycarcinogénica, siendo el benzo(a)pireno (BaP) el másconocido y estudiado (Marce & Borrull, 2000). Su presenciaen la atmósfera, suelo, sedimentos, plantas y animalesmarinos hace que la exposición del hombre a los mismossea inevitable (Bories, 1990; Iniesta & Blanco, 2005). Suamplia distribución en el medio ambiente y la utilizaciónde ciertas técnicas durante el procesado y la conservaciónde alimentos, hace que estos contaminantes esténpresentes en los alimentos a niveles traza (Lijinsky, 1991),siendo su extracción, purificación y análisis, dificultosos(Guillen & Sopelana, 2004).
Las fuentes naturales y antropogénicas másfrecuentes de HAPs son: la combustión espontánea oprovocada (incendios), las erupciones volcánicas, laproducción industrial de energía, las calefaccionesdomésticas, el tráfico, la conservación de la madera, ladestrucción de la paja tras la cosecha de los cereales,etc. (Antón & Lizaso, 2001; Bosveld et al., 2000).
Los HAPs son considerados cancerígenos alimentariospotenciales desde 1950 cuando se observó una elevadaincidencia de cáncer de estómago en poblaciones queconsumían apreciables cantidades de productos ahumados.Desde entonces, en la población general no fumadora, laingesta dietética ha sido identificada como la principal rutade exposición a los HAPs (Ramesh, 2004). Trazas de HAPshan sido halladas en muchos alimentos (vegetales, aceitesvegetales, frutas, cereales, alimentos marinos, carne ala brasa, alimentos ahumados, café y té (Germuska &Michalski, 2000).
El uso de organismos como bioindicadores de conta-minantes químicos tales como los HAPs radica en elhecho de que la contaminación presente en la columnade agua puede no hallarse bien registrada en el sedimento.El análisis de las concentraciones de HAPs en sedimentoy agua puede dar información acerca de los niveles decontaminación, pero no de la fracción biodisponible,además los bivalvos pueden acumular y por lo tantoconcentrar contaminantes a niveles muy por encima delos del agua en el que desarrollan su ciclo vital. Entreestos organismos el mejillón está considerado entre losmejores centinelas en el seguimiento de la contaminaciónmarina, ya que son organismos sedentarios, filtradoresy poseen una limitada capacidad de metabolización deHAPs (Bodin, 2004; Baumard, 1998).
En el 2005 se reportó por primera vez en nuestropaís la presencia de Perna viridis en la Bahía deCienfuegos, especie oriunda del Indo Pacífico, la cual enpoco tiempo ha colonizado los sustratos duros naturales
y artificiales presentes en esta zona, lo que ha provocadoserios inconvenientes a la economía del país. Una posiblerespuesta a ello pudiera ser la explotación comercial deeste recurso, lo que disminuiría la presencia de estosorganismos a niveles permisibles. La Bahía de Cienfuegosreporta valores de contaminación en sedimentos que lasitúan entre las dos más contaminadas en nuestro país(Martinez et al., 2010), desde el 2002 la pesca estatal eneste ecosistema está prohibida, es por ello que el objetivode este trabajo fue evaluar la contaminación por HAPs delmejillón Perna viridis de la Bahía de Cienfuegos, teniendoen cuenta que en este muelle se encuentran las mayoresconcentraciones de estos organismos dentro de la bahía.
MATERIALES Y MÉTODOS
La muestra estuvo formada por 50 especímenes delMuelle Pablo Guzmán de la Bahía de Cienfuegos, contallas entre 8 y 10 cm de largo antero-posterior de laconcha, los cuales fueron congelados y mantenidos encongelación a –18 ºC hasta su análisis, para lo cualfueron descongelados, luego se les extrajo la masa, semolió y homogenizó para obtener el pool que fue poste-riormente analizado.
La metodología analítica empleada fue la descrita enEPA 8100 (1990), pesándose dos muestras de 10 g delpool, las cuales se trataron en soxhlet con una mezclade hexano/diclorometano durante 8 h, después de estetiempo se realizó la saponificación con una mezclaKOH/metanol y se extrajo con n-hexano, posteriormentela extracción obtenida se purificó en columnas con sílicay alúmina previamente tratadas y se eluyó con una mezclade hexano/diclorometano 1-1.
La mezcla del estándar analítico utilizada fuesuministrada por la firma ULTRA SCIENTIFIC, la cualcontenía los siguientes hidrocarburos aromáticos:acenafteno, acenaftileno, antraceno, benzo(a)antraceno,benzo(a)pireno, benzo(b)fluoranteno, benzo(e)pireno,benzo(g,h,i)perileno, benzo(k)fluoranteno, bifenil, criseno,dibenzo(a,h)antraceno, dibenzotiofeno, fluoranteno,fluoreno, indeno(1,2,3-cd)pireno, naftaleno, 1-metilnaftaleno, 2-metil naftaleno, 2,6-dimetil naftaleno,2,3,5-trimetil naftaleno, perileno, fenantreno y 1-metilfenantreno, pireno.
Para los análisis se utilizó un cromatógrafo de gasesShimadzu modelo 2014 con una columna capilar BP5(5 % Phenyl y 95 % dimethyl polysiloxane) de30 m x 0,25 mm de DI y 0,25 µm de espesor depelícula. La temperatura del inyector fue de 300 ºC, latemperatura del detector 320 ºC y un programa detemperatura para la columna con una temperaturainicial de 45 ºC durante 5 min, seguido de rampas detemperaturas hasta 310 ºC.

51
Los resultados de contenido de cada uno de los HAPspresentes en ambas muestras fueron promediados yreportados cada HAPs como los valores de contaminaciónpresente en los mejillones en masa fresca.
RESULTADOS Y DISCUSIÓN
En la figura 1 se muestran las concentraciones promediosde las dos determinaciones de cada uno de los HAPspresentes en las muestras de mejillón evaluadas.
300
250
200
150
100
50
0
μg/k
g
44,821,63 15,22
61,28
264,01
28,06
246,59
25,88
12,5148,19
62,87
26,04 43,9457,6
Naftale
no
Acenaft
eno
Fluore
no
Dibenzo
tiofen
o
Fenant
reno
Antrace
no
Fluora
nteno
Pireno
Benzo(a
) antra
cenoCrise
no
Perilen
o
Benzo(e
) piren
o
Benzo(g
,h,i) an
tracen
o
Benzo(a
) perile
no
Fig. 1 Concentración promedio en µg/kg de cada uno delos hidrocarburos aromáticos policíclicos encontrados en
el pool de mejillones
Fig. 2 Porcentaje promedio de los HAPs encontrados en elpool de mejillones
TABLA 1. Relaciones entre HAPs más estudiadas y valor de estas relaciones encontradasen el pool de mejillones analizados
En la figura 2, se representa el porcentaje de cadaunos de los HAPs con respecto a la cantidad total deHAPs detectados. Las mayores contribuciones dehidrocarburos aromáticos estuvieron representadas porel fenantreno, fluoranteno, criseno y pireno con el 22,5;
Relación entre HAP Petrogénico Pirogénico Valor encontrado
Fenantreno/antraceno ≥ 10 < 10 9,41
Fluoranteno/pireno < 1 ≥ 1 1,97
Fluoranteno/fluoranteno + pireno ≤ 0,5 > 0,5 0,66
21,0; 12,7; y 10,7 % respectivamente del total de loshidrocarburos encontrados.
Cada fuente de contaminación genera una distribucióncaracterística entre los diferentes HAPs, para determinarel posible origen se evaluaron las relaciones de HAPs másestudiadas que son; fenantreno/antraceno, fluoranteno/pireno y fluoranteno/fluoranteno + pireno (Aatmeeyata,2010; Fernández et al., 1999; Page, 1999; Soriano, 2006).Estos criterios se basan en peculiaridades en la composicióny patrón de distribución de HAPs en función de la fuente deemisión. Cuando la relación fenantreno/antraceno es < 10como en nuestro caso, la contaminación tiene un origenproducto de procesos de combustión incompleta decombustibles fósiles, algo similar a lo ocurrido con el restode las relaciones evaluadas, en que en todos los casosindicaron que las fuentes de contaminación encontradasfueron pirogénicas (TABLA 1).
En la figura 3 se muestran los porcentajes de losHAPs cancerígenos o posibles cancerígenos, resultó del20,33 % (benzo(a)pireno, benzo(a)antraceno y el criseno)del total HAPs. De acuerdo con estudios realizados porla Agencia Internacional de Investigaciones del Cáncerde Lyon Reglamento (UE Nº 835/2011), el benzo(a)pirenoha sido considerado como agente carcinógeno humano
de categoría I, en tanto los dos restantes benzo(a)antracenoy criseno como posibles carcinógenos humanos con cate-goría II B.
La figura 4, muestra el contenido de benzo(a)pirenoencontrado en el pool de mejillones, cuyo valor sobrepasóen más del doble el límite máximo de residuo permitido(Reglamento UE, Nº 835/2011).

52
Fig. 3 Valores promedios de porcentajes de los HAPscancerígenos en relación con el total de HAPs,
encontrados en el pool de mejillones
Fig. 4 Contenido real promedio encontrado en el pool demejillones y límite máximo de residuo para el
benzo(a)pireno
120,0100,0
80,060,0
40,0
20,00,0μg
/kg
peso
húm
edo
Benzo(a)antraceno
CrisenoBenzo(a)pireno
Totalcancerígenos
Total HAPs
HAPs cancerígenos y totales
% de HAPs cancerígenos en el total de HAPs
30
25
20
15
10
5
0Valor real LM
μg/k
g
Fig. 5 Sumatoria de las concentraciones debenzo(a)pireno, benzo(a)antraceno, benzo(a)fluorantenoy criseno, y límite máximo permitido para estos HAPs
cancerígenos o posibles cancerígenos
Totalcancerígenos
250
200
150
100
50
0μg/k
g de
HA
Ps e
n pe
sohú
med
o
Límitemáximo
Los resultados discutidos anteriormente eximen almejillón verde de la Bahía de Cienfuegos como un posiblealimento con destino al consumo humano.
CONCLUSIONES
Los mejillones estudiados están contaminados con 14HAPs, de los cuales dos están clasificados como posiblescarcinógenos humanos y uno como agente carcinógenohumano, el benzo(a)pireno, cuyo contenido en el músculode este organismo superó el límite máximo permitido deeste contaminante, por lo que su consumo constituyeun serio riesgo para la salud humana.
La principal fuente de contaminación de acuerdo conlas relaciones de HAPs evaluados fue de carácterpirogénico, determinado por la gran urbanización eindustrialización de las zonas adyacentes a la bahía.
REFERENCIAS
Aatmeeyata, M. S. (2010). Aromatic hydrocarbons,elemental and organic carbon emissions fromtire-wear. Science of the total environmental., 408(20), 4563-68.
Antón, A. & Lizaso, J. (2001). Hidrocarburos aromáticospolicíclicos. Fundación Ibérica para la SeguridadAlimentaria. Madrid.
Baumard, P. (1998). Origin and bioavailability of PHAsin the Mediterranean Sea from mussels and sedimentrecords. Estuar. Coast. Shelt Sci., 47 (5), 415-439.
Bodin., T., Burgeot, J. Y., Stanisière, G., Bocquené, D.,Menard, C., Minier, I., et al. (2004). Seasonalvariations of a battery of biomarkers and physiologicalindices for the mussel Mytilus galloprovincialistransplanted into the northwest Mediterranean Sea.Comp. Biochem. Physiol., 138C, 411-427.
Bories, G. (1990). Toxicidad de los hidrocarburosaromáticos policíclicos y de los productos de pirólisis.En J. Derache (Ed.), Toxicología (pp. 295-318).Omega, Barcelona.
Bosveld, A. T. C., de Bie, P. A. F., van den Brink, N. W.,Jongepier, H. F. & Klomp, A. V. (2002). In vitro ERODinduction equivalency factors for the 10 PAHsgenerally monitored in risk assessment studies inthe Netherlands. Chemosphere, 49, 75-83.
Fernandez, P., Vilanova, R. M. & Grimalt, J. O. (1999).Sediment fluxes of polycyclic aromatic hydrocarbonsin European high altitude mountain lakes. Environ.Sci. Technol., 33 (21), 3716-3722.
EPA 8100 (1990). Determination of polycyclic aromaticcompounds for Chromatography.
Germuska, R. & Michalski, P. (2000). Aplication of gelpermation chromatography and HPLC method withfluorescence detector to determination of benzo(a)pyrenein food samples. Cent Eur Public Health & Suppl., 92-93.

53
Guillen, M. D. & Sopelana, P. (2004). Load of polycyclicaromatic hydrocarbons in edible vegetable Oliz:importance of alkylated derivatives. J. Food Protect.,67 (9), 1904-1913.
Iniesta, R. & Blanco, J. (2005). Bioacumulación dehidrocarburos y metales asociados a vertidosaccidentales en especies de interés comercial deGalicia. Recursos Mariños (Monog.), 2, 200 pp.
Lijinsky, W. (1991). The formation and occurrence ofpolynuclear aromatic hydrocarbons associated whithfood. Mut. Res., 259 (3-4), 251-261.
Page, D. S., Boehm, P. D., Douglas, G. S., Bence, A. E.,Burns, W. A. & Mankiewicz, P. G. (1999). Pyrogenicpolyciclic aromatic hydrocarbons in sediments recordpast human activity: A case study in Prince WilliamSound, Alaska. Mar Pollut Bull., 38, 247-260.
Marce, R. M. & Borrull, F. (2000). Solid-phase extractionof polycyclic aromatic compounds. J. Chromatogr.,885, 273-290.
Martínez, R. R., Torres, R. I., Suárez, M. Y. & Beltrán G. J.(2011) (en prensa). Nivel de contaminación porhidrocarburos del petróleo en zonas costeras de Cuba.Centro de Ingeniería y Manejo Ambiental de Bahías yCostas (CIMAB).
Ramesh, A., Walker, S. A., Hood, D. B., Guille, M. D.,Schneider, K. F. & Weyand E. H. (2004).Bioavailability and risk assessment of orally ingestedpolycyclic aromatic hydrocarbons. Int. J. Toxicol.,23 (5), 301-33.
Reglamento (UE) Nº 835/2011. Comisión del 19 de agosto2011. Que modifica el Reglamento (CE) Nº 1881/2006 por lo que respecta al contenido máximo dehidrocarburos aromáticos policíclicos en productosalimenticios.
Soriano, S. J. (2006). Datos preliminares de HAPs enmejillón silvestre de la costa Cantábrica (España)después del vertido del prestige. Ciencias Marinas,32 (2B), 457-463.

Revista Cubana de Investigaciones PesquerasEnero-diciembre, 2012, vol. 29, NO. 1, ISSN 0138-8452, pp.
54
54-58
Reproductive success of sea turtle 2008-2010 at Playa El Guanal,Isla de la Juventud, Cuba
Yosvani Medina Cruz, Félix G. Moncada Gavilán y Gonzalo Nodarse Andreu
Centro de Investigaciones Pesqueras. Ave. 5ta. y calle 246, Santa Fe, Playa,La Habana, Cuba, CP 19100, Teléfono: (537) 209-7852,
E-mail: [email protected]
RESUMEN
Se determinó el éxito de los nidos para las tortugas verde (Chelonia mydas) y caguama (Caretta caretta), a partirde monitoreos realizados en playa El Guanal (Sur de la Isla de la Juventud), durante las temporadas reproductivas2008, 2009 y 2010. Se tomaron como indicadores, Éxito de Emergencia (EE), Neonatos Eclosionados Muertos(NEM), Huevos con Embriones Muertos (HEM) y Huevos Sin Desarrollo Aparente (HSDA). En tortuga caguama,ninguno de los indicadores medidos mostró diferencias significativas en todo el período de estudio, p > 0,05.En cuanto a la tortuga verde los valores medios del EE se mantuvieron entre 71,6 y 88,2 %, disminuyendosignificativamente en las temporadas 2009 y 2010 (p < 0,05), mientras el indicador HEM aumentósignificativamente en 2009 y 2010 (p < 0,05), no resultando así para los indicadores NEM y HSDA, loscuales no encontraron diferencias significativas en todo el período de estudio, p > 0,05. Se estimó que un totalde 61 498 crías de las dos especies lograron llegar al mar, de las cuales 10 452 resultaron ser tortugascaguamas y las restantes 51 046, tortugas verdes.
Palabras clave: anidación, tortuga verde, Chelonia mydas, caguama, Caretta caretta, éxito de los nidos.
ABSTRACT
The success of nesting green turtles (Chelonia mydas) and loggerhead turtle (Caretta caretta), during 2008,2009 and 2010 breading season in El Guanal beach was determined. The hatching Emergence Success, DeadHatching in the Nest , Embryonic Death and Eggs Without Development was taken like indicators to comparethe breading seasons. In the loggerhead turtle, anything indicator show significant statistical different for allstudy period, p > 0,05, By the other hand in the green turtle, the Hatching Emergent Success ranged between71,6 and 88,2 %, decreasing significantly for season 2009 and 2010 (p < 0,05) and the indicator EmbryonicDeath increased significantly in 2009 and 2010 (p < 0,05), beside, the Dead Hatching in the Nest and EggsWithout development not showers differences significant, p > 0,05 between breading season. For the studyperiod a total of 61498 hatching reached the sea, of which 10 452 (loggerhead) and 51 046 (green) wereestimated.
Keyword: nesting, green turtle, Chelonia mydas, loggerhead, Caretta caretta, success nests.
Éxito reproductivo de tortugas marinas en playa El Guanal(2008-2010), Isla de la Juventud, Cuba
INTRODUCCIÓN
El ambiente de las playas de anidación proporciona unaoportunidad para estudiar la biología de la reproducción yde la anidación; por otra parte ha constituido la fuente másconfiable de información sobre el tamaño de las poblacionesy sus tendencias a largo plazo (Richardson, 2000).
La determinación del tamaño de la nidada y los éxitosde eclosión y emergencia, provee información fundamentalpara la conservación y manejo de las tortugas marinas.
Estos datos son esenciales porque ayudan a comprenderla salud general de la población anidadora (Miller, 2000).
Se cree que el éxito del nido, el cual lo define Miller (2000)como el número de crías que eclosionan de acuerdo con elnúmero de huevos que fueron incubados; está influenciadopor un número de factores ecológicos interactuando entre sí(Miller, 1985; Frazier, 1999; Garmestani et al., 2000; Jakshaet al., 2005; Medina et al., 2010). Sin embargo, la tendenciapor parte de las tortugas marinas, a poner varias puestassucesivas y ampliamente separadas, en toda una temporadade anidación (Carr & Stancyk, 1985), hace muy difíciles los

55
estudios sobre el éxito reproductivo de estas especies(Mortimer, 1990); no obstante, el trabajo tiene comoobjetivo analizar el éxito reproductivo de la tortuga verde yla tortuga caguama en playa El Guanal al Sur de la Isla de laJuventud, para las temporadas reproductivas 2008, 2009y 2010.
Fig. 1 Ubicación geográfica de playa El Guanal
MATERIALES Y MÉTODOS
La playa El Guanal se encuentra localizada en la costaSur de la Isla de la Juventud dentro del archipiélago delos Canarreos (Fig. 1).
El monitoreo se llevó a cabo durante los meses deanidación de la tortuga vede y la caguama (mayo-sep-tiembre), para las temporadas reproductivas 2008, 2009y 2010.
El examen del nido se realizó una vez eclosionado elmismo. El número de huevos por nidos se determinó porconteo directo de los cascarones (todos los que superanel 50 % del tamaño del huevo), sumado a los huevos noeclosionados que quedan como resto del nido, los cualesse subdividen en dos categorías Huevos Sin DesarrolloAparente (HSDA) y Huevos con Embriones Muertos(HEM).
Total de huevos = # de cascarones + HSDA + HEM
Los huevos sin eclosionar (HSDA, HEM) y losNeonatos Eclosionados Muertos (NEM) que quedan comoresto del nido son considerados al igual que el Éxito deEmergencia (EE), indicadores del éxito de los nidos. Esteúltimo se calculó según la fórmula propuestas por Miller(2000).
Éxito deemergencia
(%)100x
huevos de totalM)y(Vneonatos–cascaronesde#
=
Donde:
V: neonatos vivos dentro del nidoM: neonatos muertos dentro del nido
Los demás indicadores se calcularán según lasfórmulas siguientes:
100.xhuevosdeTotalNEMdeTotal
(%)NEM
100,xhuevosdeTotalHEMdeTotal
(%)HEM
100,xhuevosdeTotalHSDAdeTotal
(%)HSDA
=
=
=
Se analizó el comportamiento temporal de estosindicadores y no se tubo en cuenta el éxito de eclosiónya que este difiere muy poco del éxito de emergencia(EE), además este último es el de mayor peso porquerepresenta lo que en realidad aporta un área determinadaal medio en cuanto a número de crías.
Para la comparación temporal de los indicadores deléxito de los nidos se utilizó una prueba F de comparaciónde medias, analizando previamente la normalidad yhomogeneidad de los datos. Todos los análisis serealizaron con un nivel de confianza de p ≤ 0,05, medianteel paquete estadístico Statistica Six Sigma 7.0.
RESULTADOS Y DISCUSIÓN
Los valores medios de los Éxitos de Emergencia para latortuga (EE) caguama oscilaron entre el 74 y 82 %(Fig. 2), no obstante, no se encontraron diferenciassignificativas entre las temporadas reproductivasp > 0,05. El indicador Neonatos Eclosionados Muertos
N
S
W E
Golfo de Batabanó
San FelipeIsla de laJuventud
Cayos deDiego Pérez
CayoRosarioCayo
CantilesCayoLargo
Golfo deCazones
CUBA

56
(NEM) afecta directamente el Éxito de Emergencia delas crías y aunque se ha observado un ligero incremento
en el tiempo (Fig. 2), este no resulta significativop > 0,05.
Fig. 2 Indicadores del éxito de los nidos para la tortuga caguama
F (2; 65) = 0,6; p = 0,6 Media Media ± SD F (2; 65) = 1,19; p = 0,31 Media Media ± SD
F (2; 65) = 2,9; p = 0,06 Media Media ± SD F (2; 63) = 1,4; p = 0,3 Media Media ± SD
100
90
80
70
60
50
50
40
30
20
10
0
10
8
6
4
2
0
1412
1086420
–2
HEM
(%)
EE (%
)
NEM
(%)
HSD
A (%
)
2008 2009 2010 2008 2009 2010
2008 2009 2010 2008 2009 2010
Años Años
Años Años
Muy similar a los indicadores analizados, el indicadorHuevos Sin Desarrollo Aparente (HSDA), no mostródiferencias significativas en el tiempo (p > 0,05), comomuestra la figura 2, resultado contrario al encontradopor Medina et al. (2010) en el caso de la tortuga careyen Jardines de la Reina, donde los autores encuentran unaumento progresivo en el tiempo respecto a dichoindicador, por otra parte coincide con los valoresencontrados en tortuga carey por Hitchins et al. (2004)y con los de Arzola-Gonzáles (2007) para la tortugaGolfina (Lepidochelys olivácea).
El indicador que recoge los embriones muertos (HEM),afecta directamente también el Éxito de Emergencia delnido. La muerte de estos embriones, generalmente estádada por las variaciones de humedad en el microambientedonde se encuentra el mismo. No obstante a esto, no seencontraron diferencias significativas en cuanto a dichacategoría (p > 0,05).
Los valores medios del EE para la tortuga verde en elárea de estudio, oscilaron entre 71,6 y 88,2 % (Fig. 3); yaunque entre los dos últimos años no se hallarondiferencias significativas respecto a dicho indicador, seencontró una disminución estadísticamente significativade estos dos últimos respecto al 2008 (p < 0,05). Sinembargo, los valores estuvieron por encima de 70 % el
cual es el indispensable para mantener una poblaciónanidadora en condiciones saludables (Mortimer, 2000).
Según los resultados obtenidos, todo indica que laprincipal causa de la disminución del EE para la tortugaverde en los dos últimos años de estudio respecto al2008, fue el aumento del indicador HEM debido a losvalores de p < 0,05 (Fig. 3). No ocurriendo así para losindicadores HSDA y NEM en los cuales no se registraronaumentos significativos en los dos últimos años res-pecto al 2008 (p > 0,05), como muestra la figura 3.No obstante a que en los dos últimos años de estudio nose reportaron huracanes para el área de estudio, eventosque afectan drásticamente el indicador HEM; puesto quecuando un nido es bañado por el oleaje, en dependenciadel tiempo de exposición los embriones detienen sudesarrollo (Miller, 1996; 1999); las intensas lluviastambién tienen un efecto negativo sobre el desarrollo delos embriones, las cuales resultaron intensas a finalesde las temporadas reproductivas 2009 y 2010. Lasmismas reducen la temperatura de incubación y aumentanconsiderablemente los niveles de humedad, la cual segúnNaranjo (1989), es extremadamente perjudicial para unbuen desarrollo de los embriones.
Lo más importante de la anidación es la contribucióndel área en cuanto al número de neonatos que son

57
capaces de llegar al medio natural. En la mayoría delos casos es imposible realizarle el análisis a todos losnidos que son incubados en un área dado a lo prolongadaque resulta la temporada de anidación y lo distantesque quedan las áreas. Sin embargo, a partir del númerode nidos examinados se pueden hacer estimacionesen cuanto a número de huevos que fueron incubados
en una temporada y el número de neonatos que lograronentrar al medio natural. De esta forma se pudocuantificar un total de 61 498 neonatos para las trestemporadas de estudio en playa El Guanal. De lasmismas 10 452 fueron de tortuga caguama y lasrestantes 51 046 resultaron tortuga verde, comomuestra la TABLA 1.
Fig. 3 Indicadores del éxito de los nidos para la tortuga verde
HEM
(%)
2008 2009 2010 2008 2009 2010Años Años
F (2; 110) = 4,9; p = 0,001 Media Media ± SD Media Media ± SDF (2; 110) = 0,6; p = 0,6
2008 2009 2010 2008 2009 2010Años Años
F (2; 110) = 6,1; p = 0,003 Media Media ± SD Media Media ± SDF (2; 110) = 2,9; p = 0,06
20
18
16
1412
108
6
HSD
A (%
)
100
90
80
70
60
50
EE (%
)
1412
1086
4
20
10
8
6
4
2
0
NEM
(%)
TABLA 1. Número de neonatos estimados para la tortuga verde y la tortuga caguama por temporada de anidación
Sp Años Total de Total de Promedio de Estimado de Total por nidos nidos neonatos neonatos especie
examinados por nidos
Tortuga caguama 2008 42 29 103 4 326 10 4522009 46 30 81 3 7262010 30 21 80 2 400
Tortuga verde 2008 261 43 106 27 666 51 0462009 32 15 77 2 4642010 249 59 84 20 916
Total 660 197 61 498
Es muy evidente que la tortuga verde resultó laespecie con mayor contribución al medio en cuanto alnúmero estimado de neonatos que lograron llegar al mar,muy acorde esto con la proporción del número deanidaciones registrada por la especie respecto a la
caguama. Por otra parte, la tortuga verde es la másrepresentativa en las playas de anidación dentro denuestro archipiélago, causa probable de la recuperaciónde dicha especie en toda la región del Gran Caribe(Seminoff et al., 2007).

58
CONCLUSIONES
• Los éxitos de emergencia en los nidos analizadosestuvieron por encima del 70 % para la tortuga verdey la caguama en las tres temporadas de estudio.
• La tortuga verde resultó la especie con mayorcontribución al medio en cuanto al número estimadode neonatos que lograron llegar al mar, muy acordeesto con la proporción del número de anidacionesregistrada por la especie respecto a la caguama.
REFERENCIAS
Arzola-González, J. F. (2007). Humedad y temperaturaen nidos naturales y artificiales de tortuga golfinaLepidochelys olivacea (Eschssholtz, 1829). Revistade Biología Marina y Oceanografía, 42 (3), 377-383.
Carr, A. & Stancyk, S. (1985). Observation on theecology and survival Outlook of the hawksbill seaturtle. Biol. Cons., 8, 161-172.
Frazier, F. (1999). Generalidades de la historia de vidade las tortugas marinas. En K. L. Echert & F. A.Abreu Grobois (Eds.), Conservación de tortugasmarinas en la región del Gran Caribe: Un diálogo parael manejo regional efectivo (pp. 3-18).
Garmestani, A. S., Percival, H. F., Potier, K. & Rice, K.(2000). Nest-site selection by the loggerhead seaturtle in Florida’s Ten Thousand Islands. Jour. Herp,30 (4), 504-510.
Hitchins, P. M., Bourquin, O. & Hitchins, S. (2004).Nesting success of hawksbill turtle (Eretmochelysimbricata) on Cousine Island, Seychelle. Journal ofZoology, 264 (4), 383-389.
Jaksha, A. P., Foote, J. & Mueller, T. (2005). Affectsof incubation temperature on the hatching successof loggerhead sea turtle (Caretta caretta) nests onSiesta key, Florida. In M. S. Coyne & R. D. Clark(Eds.), Proceeding of Twenty-First Annual Symposiumon Sea Turtle Biology and Conservation (pp.196-197).NOAA Technical Memorandum NMFS-SEFSC-528
Medina, Y., Moncada, F. G. & Nodarse, G. (2010).Selección del sitio de anidación y éxito de eclosiónen nidos de tortuga carey (Eretmochelys imbricata).Revista Cubana de Investigaciones Pesqueras, 27 (1),60-65.
Miller, J. D. (1985). Embryology of marine turtle. In C.Gans, F. Billet & P. F. A. Maderson (Eds.), Biology of
the reptilian (vol. 14a), New York: Development. JohnWiley and Sons.
Miller, J. D. (1996). Reproduction in sea turtles. In Lutz &Musick (Eds.), The biology of sea turtles (pp.51-81).New York: CRC Press.
Miller, J. D. (1999). Determining clutch size and hatchingsuccess. In K. L. Echert, F. A. Abreu Grobois & M.Donnelly (Eds.), Research and managementtechniques for the conservation of sea turtles (pp.124-129). Marine turtle specialist group publicationNo. 4: IUCN/SSC.
Miller, J. D. (2000). Determinación del tamaño de lanidada y el éxito de eclosión. En K. L. Ecker, K. A.Bjorndal, F. A. Abreu-Grobois, & M. Donnelly (Eds.),Técnicas de Investigación y Manejo para laConservación de las Tortugas marinas. Grupo deEspecialistas en Tortugas Marinas. Publicación No. 4,2000 (Traducción al español): UICN/CSE
Mortimer, J. A. (1990). The influence of beach sandcharacteristics on nesting behavior and clutchsurvival of green turtle (Chelonia mydas). Copeia,802-817.
Mortimer, J. A. (2000). Reducción de las amenazas alos huevos y a las crías: los viveros. En. K. L. Eckert,K. A. Bjorndal, F. A. Abreu-Grobois & M. Donnelly(Eds.), Técnicas de Investigación y Manejo para laconservación de las Tortugas Marinas GrupoEspecialista en Tortugas Marinas. Publicación No. 4,2000 (Traducción al español): UICN/CSE.
Naranjo, A. R. (1989). Características del ambiente deincubación natural y su influencia en la supervivenciade los nidos de tortuga negra (Chelonia agassizi:Chelonidae) en las playas de Colola y Maruata,Michoacan. En R. Pérez (Ed.), Memorias del VEncuentro Internacional Universitario sobre TortugasMarinas. Escuela de Biología: UMSNH.
Richardson, J. I. (2000). Prioridades para los estudiossobre la biología de la reproducción y de la anidación.En K. L. Ecker, K. A. Bjorndal, F. A. Abreu-Grobois,& M. Donnelly (Eds.), Técnicas de Investigación yManejo para la Conservación de las Tortugas marinas.Grupo de Especialistas en Tortugas: UICN/CSE.
Seminoff, J., Schroeder, B., MacPherson, S., Possardt,E. & Bibb, K. (2007). 5-Years Review Green SeaTurtle/Chelonia mydas. National Marine FisheriesService Office of Protect Resource Silver SprintMaryland and U. S. Fish and Wildlife ServiceSoutheast Region Jacksonville Ecological Service FieldOffice Jacksonville Florida: 70FR20734.

59
Revista Cubana de Investigaciones PesquerasEnero-diciembre, 2012, vol. 29, NO. 1, ISSN 0138-8452, pp. 59-66
Composition and abundance of potentially toxic epibenthic thecate dinoflagellatesfrom Guajimico Cove, southern-central region of Cuba
Rosely Peraza Escarrá y Ángel R. Moreira GonzálezCentro de Estudios Ambientales de Cienfuegos (CEAC). Calle 17 esq. Ave. 46 s/n,
Reparto Reina, Cienfuegos, Cuba, CP 55100, E-mail: [email protected],[email protected]
RESUMEN
La mayoría de las comunidades de dinoflagelados epibentónicos asociadas con macroalgas, sedimentos, coralesy fanerógamas, son productoras de toxinas que causan síndromes de intoxicación en regiones tropicales ysubtropicales. En Cuba, y especialmente en Cienfuegos, los estudios acerca de estas microalgas han sidoescasos. Este trabajo estuvo dirigido a determinar la composición y abundancia de dinoflagelados epibentónicostecados potencialmente tóxicos en la Ensenada Guajimico, Cienfuegos, y su relación con algunos factoresambientales. Las muestras se tomaron desde mayo de 2011 hasta marzo de 2012, con una frecuencia mensual.Se midió la temperatura, la salinidad, y el pH del agua; así como de manera semestral los nutrientes (N-NO2
–,N-NO3
–, N-NH4+, P-PO4
–3), y clorofila a. Seis especies fueron identificadas: Gambierdiscus caribaeus, Ostreopsislenticularis, Prorocentrum belizeanum, P. concavum, P. lima y P. rhathymum. G. caribaeus constituye un nuevohallazgo para las aguas cubanas. O. lenticularis, G. caribaeus y P. belizeanum fueron las especies de mayorfrecuencia de aparición. La abundancia relativa total varió desde 1,15 x 104 hasta 5,89 x 105 cél./g de macroalgahúmeda, en los meses de octubre y febrero, respectivamente. O. lenticularis fue la especie de mayor densidaddurante todo el período, y definió la abundancia relativa total de especies. Se confirmó la presencia de palitoxinaen el área. El análisis estadístico mostró que los factores temperatura y salinidad, no influyeron en las fluctuacionesde la abundancia de las especies.
Palabras clave: Abundancia, Cienfuegos, dinoflagelados epibentónicos, factores ambientales, toxicidad.
ABSTRACT
The majority communities of epibenthic dinoflagellates associated with macroalgae, sediments, corals andseagrasses, are toxins producers that cause poisoning syndromes in tropical and subtropical regions. In Cuba,especially in Cienfuegos, studies on these microalgae have been scarce. This work was aimed to determine thecomposition and abundance of epibenthic thecate dinoflagellates producing toxins in Guajimico Cove, Cienfuegos,and their relationship with some environmental factors. Samples were taken from May 2011 to March 2012,with a monthly frequency. Temperature, salinity, and pH of the water were measured; the nutrients (N-NO2
–,N-NO3
–, N-NH4+, P-PO4
–3), and chlorophyll a, with a semester frequency were also determined. Six species wereidentified: Gambierdiscus caribaeus, Ostreopsis lenticularis, Prorocentrum belizeanum, P. concavum, P. lima,and P. rhathymum. G. caribaeus is a new report from Cuban waters. O. lenticularis, G. caribaeus and P. belizeanumwere the most frequent species. The total relative abundance ranged from 1,15 x 104 to 5,89 x 105 cell/gseaweed wet weight, in October and February, respectively. O. lenticularis was the higher density species overthe entire period and defined the total relative abundance of species. The presence of palytoxin was confirmedin the area. Statistical analysis showed that temperature and salinity did not affect the abundance and compositionof the species.
Keywords: Abundance, Cienfuegos, environmental factors, epibenthic dinoflagellates, toxicity.
Composición y abundancia de dinoflagelados epibentónicos tecadospotencialmente tóxicos de la Ensenada Guajimico,
región Centro-Sur de Cuba

60
INTRODUCCIÓN
Las comunidades de dinoflagelados epibentónicos seencuentran comúnmente asociadas con macroalgas,sedimentos, corales y fanerógamas. La mayoría de estosdinoflagelados son potenciales productores de toxinas,responsables de síndromes de intoxicación en lascadenas alimenticias, entre los que se encuentran laciguatera, el clupeotoxismo (Riobó, 2008), y la intoxi-cación diarreica por mariscos (FAO, 2004).
La ciguatera, principal síndrome asociado adinoflagelados epibentónicos, es causada por especiesdel género Gambierdiscus Adachi et Fukuyo. Lasciguatotoxinas se acumulan en el cuerpo de peces dearrecifes coralinos.
El clupeotoxismo y la intoxicación diarreica pormariscos, son ocasionados por otras toxinas que sonproducto del metabolismo de especies de los génerosOstreopsis Schmidt y Prorocentrum Ehrenberg, respecti-vamente (Riobó, 2008).
Los estudios acerca de dinoflagelados epibentónicostóxicos en Cuba se han realizado principalmente en lacosta norte de La Habana. La provincia de Cienfuegos, aligual que otras regiones del país, es afectada anualmentepor casos de ciguatera. En esta provincia, los estudios
sobre dinoflagelados epibentónicos potencialmentetóxicos han sido escasos (Moreira, 2009, 2010; Pombo,2000), y se han desarrollado fundamentalmente en laBahía de Cienfuegos, donde estos organismos son pocoabundantes. En prospecciones realizadas en el litoralabierto de la provincia de Cienfuegos, específicamenteen la Ensenada Guajimico, se ha evidenciado la presenciade abundantes poblaciones de dinoflagelados epibentó-nicos potencialmente productores de toxinas, que no hansido identificados. En esta ensenada se realizan actividadesde baño, buceo y pesca, y debido a la importancia queimplica la presencia de estos microorganismos en la zona,se hace necesario emprender un particular estudiotaxonómico-ecológico.
El objetivo de este trabajo fue determinar lacomposición y abundancia de dinoflagelados epi-bentónicos tecados potencialmente tóxicos en laEnsenada Guajimico, y su relación con algunos factoresambientales.
MATERIALES Y MÉTODOS
La Ensenada Guajimico (21º 55´ L. N. y 80º 18´ L. O.)presenta un área superficial de 0,2 km2 y una profundidadmedia de 3 m (Fig. 1).
Fig. 1 Ubicación del área de estudio (Ensenada Guajimico)
El muestreo se desarrolló con frecuencia mensual,en el período comprendido desde mayo de 2011 hastamarzo de 2012, con la excepción del mes de agosto queno pudo ser muestreado.
Se midió de manera mensual la temperatura, la sali-nidad y el pH del agua, utilizando una sonda digital
multiparamétrica modelo YSI-30. Con el propósito deevaluar el estado del agua, se midieron nutrientes (N-NO2
–,N-NO3
–, N-NH4+, P-PO4
–3) y la clorofila a, con unafrecuencia semestral, para lo cual se tomó una mues-tra de agua en un mes distintivo de la época lluviosa(septiembre), y otra de la época seca (enero). Los ensayos
CUBALitoral Oriental
EnsenadaGuajimico
LEYENDARíosProvincia Cienfuegos
0 1 2 4km
CEAC. Lab SIG
Bahía de Cienfuegos
EnsenadaGuajimico
isis
Resaltado
isis
Resaltado
isis
Resaltado

61
analíticos se realizaron en el Laboratorio de EnsayosAmbientales (LEA) del Centro de Estudios Ambientalesde Cienfuegos (CEAC), según los métodos descritos enel manual de procedimientos de esta instalación, y parael análisis de la clorofila a se siguió la metodología descritapor APHA (1998).
Por su abundancia en la zona, el sustrato escogidopara determinar la abundancia de los dinoflageladostóxicos, fueron especies de macroalgas del géneroDyctiota. Mediante buceo autónomo se colectaron al azaraproximadamente 50 g de macroalgas, las que seintrodujeron en bolsas de nailon. Se tomaron en untransecto horizontal a la costa, de aproximadamente10 m de longitud, a una profundidad entre 0,5 y 1 m.
La clasificación de los organismos se realizó deacuerdo con los criterios de Faust & Gulledge (2002) yLitaker et al. (2009).
La identidad de las especies de Ostreopsis yGambierdiscus fue corroborada por análisis filogenéticode ácidos nucleicos, en colaboración con el CentroOceanográfico de Vigo, España. Por otra parte sedeterminó la presencia de palitoxina (PLTX) en unamuestra natural de extracto microalgal, mediante elanálisis de actividad hemolítica en eritrocitos humanos,bioensayos en ratones, y cromatografía líquida condetección de fluorescencia, según la metodología descritapor Riobó & Franco (2011).
Para el recuento celular se usaron cámaras desedimentación según el método de Utermöhl modificadopor Edler & Elbrachter (2010). Se realizó el conteo detres submuestras tomadas de la muestra mensual total.La abundancia de los dinoflagelados se expresó en formade densidad celular (cél./g de macroalga húmeda).
Utilizando el paquete estadístico STATISTICA 8.0,se realizó un análisis ANOVA por rangos de Kruskal-Wallis (p < 0,01), para determinar diferencias en lariqueza de especies y en los valores de abundancia deestos microorganismos durante los meses muestreados.Cuando se detectaron diferencias, se empleó el test dela U de Mann-Whitney (p < 0,05), y una comparación
múltiple de medias no paramétrica (p < 0,01), respec-tivamente. Con el propósito de analizar la posible relaciónentre la abundancia de los dinoflagelados con la tem-peratura, y salinidad del agua, se desarrolló un análisis decorrelación por rangos de Spearman (p < 0,05).
RESULTADOS Y DISCUSIÓN
Análisis de parámetros físico-químicosdel agua
El máximo de temperatura se registró en el mes deseptiembre (30,7 ºC), mientras que el mínimo fue enenero (25,2 ºC). La temperatura del agua puede serrelacionada con la temperatura atmosférica, lo quepermite comprender que en el período lluvioso los valoresde temperatura resultaran ser superiores que los regis-trados para el período seco.
Los valores de salinidad no variaron prácticamentedurante el período de estudio. En la época lluviosa estososcilaron entre 35,3 y 35,7 ups, con un valor medio yuna desviación estándar de 35,52 ups y 0,2 ups,respectivamente. En cambio, en la seca se registraronentre 35,4 y 35,7 ups, con una media 35,56 ups, ydesviación estándar de 0,1 ups, lo que se correspondecon las características del área de estudio, que es unazona poco lluviosa, y posee un pequeño aporte fluvial delarroyo La Jutía.
Los valores de pH presentaron poca variación (desde8,06 en los meses de junio y julio hasta 8,11 en el mesde enero). Se detectaron bajas concentraciones denutrientes y de clorofila a (TABLA 1). Según los rangosestablecidos en la NC 25 (1999), para los nutrientes, elárea presenta aguas de buena calidad, lo que demuestrael poco impacto antropogénico sobre la misma. A partirde la escala de clasificación de las aguas para evaluar elestado trófico dada por Contreras et al. (1994), seconfirmó que en la zona de estudio son oligotróficas.
TABLA 1. Resultados de indicadores hidrológicos por período climático
Temporada N-NO2– N-NO3
– N-NH4+ P-PO4–3 Clorofila a
μμμμμmol/L μ μ μ μ μmol/L μ μ μ μ μmol/L μ μ μ μ μmol/L ug/L
Seca < 0,014 < 0,114 < 0,614 < 0,142 0,11 Lluvia 0,35 < 0,114 1,428 < 0,142 0,16
Identificación de los dinoflageladosy distribución
Se identificaron tres géneros y seis especies de dinofla-gelados epibentónicos tecados potencialmente tóxicos:
Gambierdiscus caribaeus, Ostreopsis lenticularis,Prorocentrum belizeanum, P. concavum, P. lima y P.rhathymum.
G. caribaeus se reporta por primera vez para Cuba, yeste resultado fue corroborado por análisis filogenético

62
de ácidos nucleicos en el Centro Oceanográfico de Vigo,España. El resto de las especies habían sido halladas enla Bahía de Cienfuegos.
En el género Gambierdiscus las diferencias morfoló-gicas entre especies son muy sutiles, lo que significaque la identificación de estos organismos utilizando comoúnico método la microscopía de luz, no es suficiente; senecesitan otros métodos más avanzados, entre ellos lamicroscopía electrónica, y el análisis filogenético deácidos nucleicos. La talla y la morfología de las placas deG. caribaeus, estuvo comprendida dentro del rango de laespecie descrito por Litaker et al. (2009). Debido a suamplia distribución geográfica (Litaker et al., 2010), estaespecie podría ser el agente causante del síndrome de laciguatera en la provincia de Cienfuegos y otras regionesdel país.
La morfología descrita para la especie O. lenticularisse ajustó a la descripción taxonómica ofrecida por Fukuyo(1981) y Faust et al. (1996). Esta especie ha sidoencontrada en áreas someras tropicales asociadas aarrecifes (Rhodes, 2011), fundamentalmente en aguasdel Océano Atlántico y el Caribe (Ashton et al., 2003;Delgado et al., 2006; Moreira, 2009).
Las dimensiones de P. belizeanum coinciden con ladescripción original para la especie ofrecida por Faust(1993). Se localiza comúnmente en aguas tropicales(Steidinger & Tangen, 1996). En costas cubanas la hanidentificado tanto en el litoral de La Habana (Delgadoet al., 2002; Delgado et al., 2006), como en el deCienfuegos (Moreira, 2009).
Las dimensiones de P. concavum fueron ligeramentesuperiores en comparación con la descripción original dadapor Fukuyo (1981), mientras que se corresponden con elrango ofrecido por Faust & Gulledge (2002), Delgadoet al. (2002), y el de la especie encontrada en Malasiapor Mohammad-Noor et al. (2007). Es una especie tropicaly nerítica (Steidinger & Tangen, 1996), lo que indica lanaturalidad de encontrarla en la zona de estudio.
P. lima presentó una talla comprendida dentro delrango dado por Dodge (1975), Delgado et al. (2000) yMohammad-Noor et al. (2007). Esta especie es de ampliadistribución mundial, nerítica, de ambientes estuarinos(Steidinger & Tangen, 1996), lo que coincide con supresencia en el área. Delgado et al. (2000), Delgadoet al. (2006) y Moreira (2009), la detectaron con anterio-ridad en nuestro país.
Las dimensiones de P. rhathymum se correspondencon las ofrecidas por Fukuyo (1981), y son superiores alas dadas por Mohammad-Noor et al. (2007). Variosautores (Faust et al., 1999; Hansen et al., 2001; Faust& Gulledge, 2002) consideraron a P. rhathymum comosinónimo de P. mexicanum Tafall; sin embargo, Cortés-Altamirano & Sierra-Beltrán (2003), se basaron en laforma celular, la ornamentación de la superficie valvar,el hábitat y la biogeografía para demostrar que estas
son especies diferentes. Teniendo en consideración queP. mexicanum es una especie planctónica (Cortés-Altamirano & Sierra-Beltrán, 2003), se puede plantearque los informes de este dinoflagelado para aguas cubanascomo epífito sobre macroalgas (Delgado et al., 2002;Delgado et al., 2006; Moreira, 2009), se correspondenrealmente con la especie epibentónica P. rhathymum. Esposible asumir que P. rhathymum es cosmopolita, debidoa que se ha encontrado en ambos hemisferios (34º N-17º S)en el Océano Pacífico y el Caribe (Cortés-Altamirano &Sierra-Beltrán, 2003).
Conteo y estimación de la densidadde dinoflagelados. Relación de la abundanciade las especies con la temperaturay salinidad del agua
El número de especies detectadas mensualmente variódesde dos, en el mes de octubre, hasta cinco, en losmeses de junio, julio, noviembre, diciembre y enero (Fig. 2).G. caribaeus, O. lenticularis y P. belizeanum fueron lasespecies de mayor frecuencia de aparición. G. caribaeus,y O. lenticularis fueron las únicas especies observadastodos los meses.
Fig. 2 Número de especies detectadas en los mesesmuestreados
La composición específica de dinoflagelados epibentó-nicos tecados potencialmente tóxicos en la ensenada deestudio, es típica de ambientes tropicales saludables. Enambientes similares varios autores (Fukuyo, 1981; Faust,1995; Mohammad-Noor et al., 2007), han encontradocoexistiendo a especies de los géneros Gambierdiscus,Ostreopsis, Prorocentrum, Coolia Meunier y AmphidiniumClaperède et Lachmann. La riqueza total de especies fue
Núm
ero
de e
spec
ies
7
6
5
4
3
2
1
0
May. Jun. Jul. Sep. Oct. Nov. Dic. Ene. Feb. Mar.
2012Meses
2011

63
superior a la detectada por Pombo (2000) en la Bahía deCienfuegos, quien observó el predominio de P. lima, unaespecie tolerante a cambios bruscos de las condicionesecológicas. Resultados diferentes fueron observados porDelgado et al. (2006) en el litoral habanero, pues encontraronsiete especies de dinoflagelados tóxicos, con el predominiode P. lima y G. toxicus.
La concentración total de dinoflagelados varió desde1,15 x 104 hasta 5,89 x 105 cél./g de macroalga húmeda, en
los meses de octubre y febrero, respectivamente. Lasespecies predominantes, en orden de importancia, fueronO. lenticularis, P. belizeanum, y G. caribaeus (Fig. 3).O. lenticularis fue tan abundante que definió laabundancia relativa total de especies; sus valores deabundancia relativa siempre fueron superiores al 85 %.P. concavum, P. lima, y P. rhathymum presentaronbajos valores de abundancia relativa, que no superaronel 1 %.
Fig. 3 Valores de abundancia relativa por especie (a: Gambierdiscus caribaeus; b: Ostreopsis lenticularis; c: Prorocentrumbelizeanum; d: P. concavum; e: P. lima; f: P. rhathymum) con respecto al tiempo
My Jn Jl S O N D E F Mr My Jn Jl S O N D E F Mr My Jn Jl S O N D E F Mr
My Jn Jl S O N D E F Mr My Jn Jl S O N D E F Mr My Jn Jl S O N D E F Mr
Abu
ndan
cia
(cél
./m
acro
alg
a hú
med
a
3,5 x 103
3 x 103
2,5 x 103
2 x 103
1,5 x 103
1 x 103
5 x 102
0
–5 x 102
Meses
7 x 105
6 x 105
5 x 105
4 x 105
3 x 105
2 x 105
1 x 105
0
–1 x 105
7 x 104
6 x 104
5 x 104
4 x 104
3 x 104
2 x 104
1 x 104
0
–1 x 104
2 x 102
1,6 x 102
1,2 x 102
8 x 101
4 x 101
0
–4 x 101
–8 x 101
6 x 102
5 x 102
4 x 102
3 x 102
2 x 102
1 x 102
0
–1 x 102
5 x 102
4 x 102
3 x 102
2 x 102
1 x 102
0
–1 x 102
En los meses de septiembre y octubre, se observóuna disminución de la abundancia relativa de macroalgasy un incremento de la abundancia relativa de cianobac-terias oportunistas (Oscillatoria margaritifera Gomont),a lo que puede asociarse que en estos meses se detec-taran los menores valores de riqueza de especies dedinoflagelados.
El análisis estadístico para los valores de abundanciaentre meses (KW, p< 0,01), indicó que existen diferen-cias significativas para todas la especies.
El análisis de correlación por rangos de Spearman(p < 0,05) no mostró relación entre la abundancia de lasespecies y el factor temperatura, mientras que sí lo hizopara la salinidad, aunque fueron correlaciones débiles.Las especies con correlación entre su abundancia y lasalinidad fueron O. lenticularis (r = 0,53), P. belizeanum(r = 0,62), y P. rhathymum (r = 0,47).
Los valores de densidad obtenidos para G. caribaeusse encuentran en el rango estimado por Litaker et al.(2010) para el género, en los Océanos Atlántico yPacífico. Se conoce que esta especie crece bien encondiciones de laboratorio entre 22-33 ºC, y óptimamenteentre 29-33 ºC (Litaker et al., 2009), y a pesar de que elanálisis estadístico no mostró correlación entre laabundancia y el factor temperatura, la temperatura delagua pudo favorecer la constancia de sus concentra-ciones durante el período de estudio.
Se ha sugerido que además de la temperatura delagua y la salinidad, otros factores determinan el desarrollode Gambierdiscus spp., entre los que se encuentran laconcentración de nutrientes, la competencia con otrosmicroorganismos y factores de crecimiento endógenos(Chateau-Degat et al., 2005), lo cual es aplicable a todaslas especies encontradas en la zona de estudio.
isis
Resaltado

64
El máximo valor de abundancia de O. lenticularis enel mes de febrero estuvo cercano al máximo encontradopara Ostreopsis spp. (5,9 x 105 cél./g de macroalgahúmeda) en el Mediterráneo, que es uno de los valoresmás altos registrados para especies de dinoflageladosbentónicos (Vila et al., 2001). Pudiera ser que latemperatura para el crecimiento óptimo de esta especiese encuentre cercana a los 26 ºC, lo que explicaría losmayores picos de abundancia en los meses de febrero ymarzo. Sin embargo, se debe brindar especial énfasis enel futuro al estudio de micronutrientes presentes en elentorno de las macroalgas, debido a que el incrementode la disponibilidad de nutrientes como el fósforo, podríaestar relacionado con la proliferación de O. lenticularis.
P. belizeanum no se detectó en el mes de octubre, loque podría deberse a que en los meses de septiembre ynoviembre se observaron los valores de abundancia másbajos. Es interesante señalar que Delgado et al. (2002)hallaron este dinoflagelado en las costas cienfueguerassolo en el mes de febrero, que coincide con el mes demayor abundancia de la especie según los resultados deeste trabajo.
En cuanto a la especie P. concavum, los resultadosobtenidos en esta investigación no son suficientes paraser discutidos a profundidad y compararlos con losobtenidos por otros autores, no obstante, es válidodestacar que Delgado et al. (2002) detectaron solo en elmes de mayo a esta especie como epífita de macroalgasen costas cienfuegueras, con una concentración de6,51 x 102 cél./g de macroalga húmeda.
Delgado et al. (2000), registraron densidades de P.lima entre 20-32 cél./g de macroalga húmeda, queresultan menores en comparación con las concen-traciones detectadas en la ensenada (de 40 a2,62 x 102 cél./g de macroalga); mientras que en otrasinvestigaciones (Pombo, 2000; Delgado et al., 2006)detectaron a la especie como la predominante. Moreira(2009) observó que P. lima era abundante en la épocalluviosa (93 cél./g de macroalga), y muy poco abundanteen la seca (9 cél./g de macroalga), lo cual no se correspondecon el presente resultado. Según Morton et al. (1992),el crecimiento óptimo de P. lima se desarrolla a tempera-turas entre 26-28 ºC, y con una salinidad por debajode 35 ups.
P. rhathymum se observó en bajas concentracionesa pesar de que Morton et al. (1992), indicaron que estaespecie (citada como P. mexicanum) mostraba uncrecimiento óptimo a temperaturas entre 26-28°C, y asalinidades oceánicas típicas (~36 ups). Delgado et al.(2006), encontraron a la especie (citada como P.mexicanum) en densidades inferiores a 103 cél./g demacroalga húmeda, lo cual se corresponde con losresultados obtenidos. Por el contrario, Moreira (2009)detectó un florecimiento de 3,1 x 103 cél./g de macroalga,en la Bahía de Cienfuegos.
Análisis de palitoxina en una muestranatural de extracto microalgal
El extracto microalgal de O. lenticularis fue tóxico, debidoa que mostró actividad hemolítica por la presencia dePLTX. Los ratones inyectados con el extracto de estaespecie, murieron entre 4 y 24 h después de la inyección,manifestando los síntomas típicos relacionados con latoxina. El contenido de toxina en las células se estimóen 0,12 pg/cél., mediante el método cromatrográfico ybiológico.
En el Caribe (Puerto Rico e Islas Vírgenes), inves-tigaciones con cultivos de O. lenticularis han demostradola toxicidad de esta especie solo a través de bioensayosen ratones (Tindall et al., 1990; Ashton et al., 2003).
La presencia de abundantes poblaciones de O.lenticularis en la zona de estudio, señala una potencialamenaza a la salud pública. La PLTX producida por unflorecimiento de Ostreopsis spp., ha sido asociada aintoxicaciones por diferentes rutas de exposición,como por ejemplo, la ingestión de mariscos contami-nados, la inhalación, o la exposición cutánea a aguasmarinas que contengan aerosol de esta toxina (Tubaroet al., 2011). Varios autores plantean la posibleimplicación de Ostreopsis spp. en el síndrome deciguatera (Vila et al., 2001; Ashton et al., 2003;Riobó, 2008).
CONCLUSIONES
1. Se identificaron tres géneros y seis especies dedinoflagelados epibentónicos tecados potencialmentetóxicos: G. caribaeus, O. lenticularis, P. belizeanum,P. concavum, P. lima y P. rhathymum. G. caribaeusconstituye un nuevo registro para las aguas cubanas.
2. Las especies O. lenticularis, G. caribaeus y P.belizeanum fueron las de mayor frecuencia deaparición y predominantes por su abundancia. O.lenticularis fue la especie predominante en todos losmeses de colecta, sus valores porcentuales deabundancia relativa fueron muy superiores al restode las especies, siempre por encima del 85 %.
3. En cuanto a los factores ambientales que puedanrelacionarse con la abundancia de los dinoflageladosestudiados, se puede plantear que la temperatura yla salinidad no influyeron en la densidad de lasespecies.
4. Se demostró la presencia de PLTX en el extractomicroalgal de O. lenticularis. El presente estudiopodría ser el primer informe de PLTX en esta especiedeterminada tanto por métodos biológicos yquímicos.

65
AGRADECIMIENTOS
Al Laboratorio de Ensayos Ambientales del CEAC.Esta investigación se desarrolló con el apoyo delProyecto de cooperación técnica del OrganismoInternacional de Energía Atómica “Diseño e implemen-tación de sistemas de alerta temprana y evaluaciónde la toxicidad de los florecimientos algales nocivos(FANs) en la región del Caribe, aplicando técnicasnucleares” (ARCAL RLA, 7/014).
REFERENCIAS
APHA-WPCF-AWWA (1998). Standard Methods for theexamination of Water and Wastewater (20th ed.,210 pp.). American Public Health Association.
Ashton, M., Tosteson, T. & Tosteson, C. (2003). Theeffect of elevated temperature on the toxicity of thelaboratory cultured dinoflagellate Ostreopsislenticularis (Dinophyceae). Rev. Biol. Trop., 51, 1-6.
Chateau-Degat, M. L., Chinain, M., Cerf, N., Gingras,S., Hubert, B. & Dewailly, E. (2005). Seawatertemperature, Gambierdiscus spp. variability andincidence of ciguatera poisoning in French Polynesia.Harmful Algae, 4, 1053-1062.
Contreras, F., Castañeda, O. & García, A. (1994). Laclorofila a como base para un índice trófico en lagunascosteras mexicanas. Ann. Inst. Cienc. Mar. Limnol.,21 (1-2), 55-66.
Cortés-Altamirano, R. & Sierra-Beltrán, A. (2003).Morphology and taxonomy of Prorocentrummexicanum and reinstatement of Prorocentrumrhathymum (Dinophyceae). J. Phycol., 39, 221-225.
Delgado, G., Popowski, G. & Nodar, R. (2000). Primerreporte de Prorocentrum lima (EHR) Dodge 1975, enaguas cubanas. Rev. Invest. Mar., 21 (1-3), 103-105.
Delgado, G., Popowski, G. & Pombo, M. C. (2002). Nuevosregistros de dinoflagelados tóxicos epibénticos paraCuba. Rev. Invest. Mar., 23 (3), 229-232.
Delgado, G., Lechuga-Devéze, C. H., Popowski, G.,Troccoli, L. & Salinas, C. A. (2006). Epiphyticdinoflagellates associated with ciguatera in thenorthwestern coast of Cuba. Rev. Biol. Trop., 54 (2),299-310.
Dodge, J. D. (1975). The Prorocentrales (Dinophyceae).II. Revision of the taxonomy within the genusProrocentrum. Dodge Bot. J. Linn. Soc., 71, 103-125.
Edler, L. & Elbrachter, M. (2010). The Utermöhl methodfor quantitative phytoplankton analysis. In B. Karlson,C. Cusack & E. Bresnan (Eds.), Microscopic andmolecular methods for quantitative phytoplanktonanalysis (110 pp). IOC Manuals and Guides 55,UNESCO, Paris.
FAO (2004). Marine biotoxins. FAO Food and NutritionPaper 80, 278 pp.
Faust, M. A. (1993). Prorocentrurm belizeanum,Prorocentrurm elegans and Prorocentrumcaribbaeum, three new benthic species(Dinophyceae) from a mangrove island Twin Cays,Belize. J. Phycol., 29, 100-107.
Faust, M. A. (1995). Observation of sand-dwelling toxicdinoflagellates (Dinophyceae) from widely differingsites, including two new species. J. Phycol., 31,996-1003.
Faust, M. A., Morton, S. L. & Quod, J. P. (1996). FurtherSEM study of the marine dinoflagellates: the genusOstreopsis (Dinophyceae). J. Phycol., 32, 1053-1065.
Faust, M. A., Larsen, J. & Moestrup, Ø. (1999).Potentially toxic phytoplankton. 3. GenusProrocentrum (Dinophyceae). In J. A. Lindley (Ed.),ICES Identification Leaflets for Plankton (pp. 1-23).International Council for the Exploration of the Sea,Copenhagen.
Faust, M. & Gulledge, R. (2002). Identifying HarmfulMarine Dinoflagellates. Smithsonian Contributionsfrom the United States National Herbarium, vol. 42,144 pp.
Fukuyo, Y. (1981). Taxonomical study on benthicdinoflagellates collected in coral reefs. B. Jpn. Soc.Sci. Fish., 47, 967-78.
Hansen, G., Turquet, J., Quod, J. P., Ten-Hage, L.,Lugomela, C., Kyewalyanga, M., Hurbungs, M. et al.(2001). Potentially Harmful Microalgae of the WesternIndian Ocean – a guide base on preliminary survey.IOC Manuals and Guides No. 41. IntergovernmentalOceanographic Commision of UNESCO, 105 pp.
Litaker, R. W., Vandersea, M. W., Faust, M. A., Kibler,S. R., Chinain, M., Holmes M. J., Holland, W. C. et al.(2009). Taxonomy of Gambierdiscus including fournew species, Gambierdiscus caribaeus,Gambierdiscus carolinianus, Gambierdiscus carpenteriand Gambierdiscus ruetzleri (Gonyaulacales,Dinophyceae). Phycol., 48, 344-390.
Litaker, R. W., Vandersea, M. W., Faust, M. A., Kibler,S. R., Naua, A. W., Holland, W. C., Chinain, M. et al.(2010). Global distribution of ciguatera causingdinoflagellates in the genus Gambierdiscus. Toxicon.,56, 711-730.
Mohammad-Noor, N., Daugbjerg, N., Moestrup, Ø. &Anton, A. (2007). Marine epibenthic dinoflagellatesfrom Malaysia - a study of live cultures and preservedsamples based on light and scanning electronmicroscopy. Nord. J. Bot., 24, 629-690.
Moreira, A. (2009). New toxic benthic dinoflagellates fromsouth central Cuba. Harmful Algae News, 39, 15-16.
Moreira, A. (2010). Dinoflagellate blooms in eutrophiczones of Cienfuegos Bay, Cuba. Harmful Algae News,41, 10-11.
isis
Resaltado
isis
Resaltado
isis
Resaltado

66
Morton, S. L., Norris, D. R. & Bomber, J. W. (1992).Effect of temperature, salinity and light intensity onthe growth and seasonality of toxic dinoflagellatesassociated with ciguatera. J. Exp. Mar. Biol. Ecol.,157 (1), 79-90.
NC 25 (1999). Evaluación de los objetos hídricos de usopesquero. Sistema de Normas para la Protección delMedio Ambiente. Hidrosfera. República de Cuba,9 pp.
Pombo, M. (2000). Dinoflagelados tóxicos bentónicosasociados con la ciguatera en dos localidades dellitoral cubano. Tesis presentada en opción al TítuloAcadémico de Máster en Biología Marina conMención en Ecología Marina. Universidad de LaHabana, 52 pp.
Rhodes, L. (2011). World-wide occurrence of the toxicdinoflagellate genus Ostreopsis Schmidt. Toxicon.,57, 400-407.
Riobó, P. (2008). Palitoxinas, ensayos biológicos ymétodos químicos para su determinación en
organismos marinos. Tesis de doctorado. Universidadde Vigo, 104 pp.
Riobó, P. & Franco, J. M. (2011). Palytoxins: Biologicaland chemical determination. Toxicon., 57, 368-375.
Steidinger, K. & Tangen, K. (1996). Dinoflagellates. InC. Tomas (Ed.), Identifying Marine Diatoms andDinoflagellates (pp. 387-584). Academic Press, Inc.
Tindall, D. R., Miller, D. M. & Tindall, P. M. (1990). Toxicityof Ostreopsis lenticularis from the British and UnitedStates Virgin Islands. In E. Graneli, B. Sundstrom, L.Edler & D. M. Anderson, (Eds.), Toxic MarinePhytoplankton (pp. 424-429). Elsevier, New York.
Tubaro, A., Durando, P., Del Favero, G., Ansaldi, F.,Icardi, G., Deeds, J. R. & Sosa, S. (2011). Casedefinitions for human poisonings postulated topalytoxins exposure. Toxicon., 57, 478-495.
Vila, M., Garcés, E. & Masó, M. (2001). Potentially toxicepiphytic dinoflagellate assemblages on macroalgaein the NW Mediterranean. Aquat. Microb. Ecol., 26,51-60.

67
Revista Cubana de Investigaciones PesquerasEnero-diciembre, 2012, vol. 29, NO. 1, ISSN 0138-8452, pp. 67-76
Dispersion of lobster larvae in the Caribbean based on numerical simulations
Alina Rita Gutiérrez Delgado,1 Julio Baisre Alvarez2 y Amaury Alvarez Cruz1
1 Instituto de Oceanología. Calle 184 entre 1ra. y 3ra., Rpto. Flores,Playa, La Habana, Cuba, teléfono: 271-6008,
E-mail: [email protected], [email protected], [email protected] Acuario Nacional de Cuba. Ave. 1ra. y 60, Playa, La Habana, Cuba
RESUMEN
En Cuba, la langosta espinosa Panurilus argus es el recurso marino de mayor importancia comercial. Lamayor abundancia de larvas de la especie se colecta en las aguas oceánicas próximas a la región suroccidental.En esta región también se producen las mayores pesquerías de todo el archipielago cubano. Este trabajotiene el objetivo de esclarecer cuáles son los mecanismos físicos que propician este hecho, cuantificar losporcentajes potenciales de larvas que se retienen en la plataforma cubana y los aportes de las poblacionesvecinas de Campeche e Isla Caimán Grande. Se utiliza un modelo estocástico lagrangiano para estimar ladispersión de las larvas y se acoplan características del comportamiento de la especie como tiempo deduración larval, sitios históricos de desove, mortalidad y pico de desove. Las simulaciones evidencian queel desove producido en la región suroccidental de Cuba se retiene en gran cuantía en la plataforma cubana.Regiones como la costa NE de la Florida y Jamaica se benfician con estos desoves. Las larvas producidasal suroeste de Cuba llegan hasta regiones lejanas de América Central y la costa norte de América del sur. Losdesoves de Campeche y de la Isla Caimán Grande no deben sustentar la población de langosta en Cuba,pero contribuyen significativamente al mantenimiento genético de la especie. Se concluye que en el Caribela población de langosta está conectada. Principalmente, Cuba se conecta con el Caribe occidental. LascaracterÍsticas del sistema de corrientes y del largo período de duración larval de la especie contribuyen alograr conexiones entre regiones lejanas.
Palabras clave: conexión, langosta, modelo, lagrangiano.
ABSTRACT
In Cuba, the spiny lobster Panurilus argus is the marine resource of greater commercial importance. Thehighest abundance of larvae of the species is collected in ocean waters near the southwestern region. Theregion also produced the largest fisheries of the Cuban archipelago. This paper aims to clarify the physicalmechanisms to promote this fact, also to quantify the potential percentages of larvae that are retained inthe Cuban platform and the contribution of Campeche and Grand Cayman Island populations. We used aLagrangian stochastic model of larval dispersal, coupling behavior of the species as larvae duration,historical spawning sites, mortality and peak of the spawning season. The simulations showed thatspawning occurred in the SW region of Cuba is retained in large amounts in the Cuban platform. Regionssuch as the NE coast of Florida and Jamaica benefit with these spawns. However larvae produced southwestof Cuba reach remote regions of Central America and the northern coast of South America. The spawningfrom Campeche and Grand Cayman Island should not sustain the lobster population in Cuba, but contributesignificantly to the genetic maintenance of the species. We conclude that the lobster population is connectedin the Caribbean. Cuba is connected with the western Caribbean, mainly. The characteristics of thecurrent system and the long period of larval duration of the species contribute to achieving connectionsbetween distant regions.
Keywords: connection, lobster, model, lagrangian.
Dispersión de larvas de langosta en el Caribe basadaen simulaciones numéricas

68
INTRODUCCIÓN
En Cuba, la langosta espinosa Panurilus argus es elrecurso marino de mayor importancia comercial. Por estemotivo numerosas investigaciones científicas tienencomo objetivo conocer las características de la especiey la relación con su hábitat. Estudios precursores indicanque las larvas permanecen en el océano aproximadamentede seis a ocho meses, durante ese período estánexpuestas a la temperatura, presencia o no de alimento,profundidad de la termoclina, luminosidad y a ladepredación (Alfonso et al., 2000).
Los trabajos de Cruz et al. (1991), reportan que eldesove principal de esta especie se realiza entre febreroy mayo, el pico importante de entrada de puerulus a laplataforma es entre septiembre y noviembre, mientrasque el pico de reclutamiento al área de pesca es entremarzo y mayo. En Cuba, el mayor reclutamiento depuerulus se localiza al SW de Cuba (Cruz et al., 1991a).En concordancia con esta característica, la mayorabundancia de larvas de langosta se colecta en las aguasoceánicas próximas a la plataforma de esta región(Alfonso, et al., 1991).
Este trabajo persigue esclarecer los mecanismosfísicos que propician este hecho, así como cuantificar elporcentaje potencial de larvas que se autorecluta en laplataforma cubana y los aportes que pudieran realizarlas poblaciones vecinas de langostas de Campeche e IslaCaimán Grande. Además de conocer las trayectorias másprobables de los desoves de langosta que se producen alSW de Cuba.
Los estudios de conectividad oceánica entre lasregiones del Caribe han estado en el centro de los debatesdurante las últimas décadas (Paris et al., 2005; Cowen,2009). En el proceso de conexión y transporte tienenrelevante importancia los remolinos oceánicos comomecanismos físicos que intervienen en la retención(Guerrero, 2005; Jouanno et al., 2008). Los sistemasde circulación cerrada pueden organizarse en vórticesde larga vida que dominan la dinámica del flujo. Se conoceque estos sistemas pueden actuar como trampas decomunidades planctónicas y nutrientes (Group, 1981;Yeung & Lee, 2002), por lo que constituyen escenariosfavorables para la retención de larvas a lo largo de lasplataformas (Lee & Williams, 2000).
El sistema de corrientes oceánicas al suroeste de Cubase caracteriza por mecanismos que actúan como trampasde partículas. Se han identificado contracorrientes,transporte de Ekman efectivo (Hernández et al., 1995) yremolinos oceánicos cercanos a la plataforma (Siam, 1988).Según, Baisre et al. (1978) y García et al. (1991) estascondiciones pudieran retener larvas originadas en Cuba,así como atrapar larvas provenientes de otras localidades
del Caribe. En esta región el enlace de los remolinosoceánicos permite la conexión e intercambio entre aguaslocalizadas a grandes distancias. La relación de losremolinos oceánicos con la mayor abundancia de larvasde langosta, nutrientes, clorofila a, fitoplancton y bacteriasen la región suroccidental de Cuba muestra que estosmecanismos deben ser el factor principal que propicia laretención al SW de Cuba, por sus características deestabilidad en la región de interacción océano-plataforma(Gutiérrez, et al., 2009).
En esta investigación se simulan las trayectorias delarvas virtuales de langosta liberadas en localidades delsuroeste de Cuba, Yucatán e Isla Caimán Grande. Lasimulación numérica de las trayectorias de larvasvirtuales se logra con un modelo biofísico que incorporael campo de corrientes entre los años 1995 y 2000,proveniente de una serie de datos de altimetría, unesquema estocástico lagrangiano y comportamiento delas fases larva-adulto de la langosta que incluye morta-lidad de las larvas, período de duración larval (PDL), sitiosy principales períodos de desove. En una primera aproxima-ción la simulación de las trayectorias no incluye la difusiónvertical.
Las simulaciones realizadas demuestran que laslarvas de langosta liberadas al SW de Cuba tienen elmayor reclutamiento en Cuba, local o regional. La mayorretención observada en la región suroccidental de Cubaes consistente con los mayores recalos de puerulus ycon los reportes de grandes pesquerías en la región (Puga,et al., 1991).
La simulación de las trayectorias de larvas virtualesde langosta liberadas en Yucatán indica que el mayorporcentaje de las larvas se mantiene dentro del Golfo deMéxico por los movimientos cíclicos de las corrientes.Otra significante cantidad es arrastrada por la corrientede la Florida hacia el NE de la península de la Florida. Laregión noroccidental de Cuba es subsidiada con el 2,8 %del total liberado. Este porcentaje, sino contribuye alaumento de la población de langosta en Cuba, podríamantener la conexión genética entre Cuba y México.
Las simulaciones de las trayectorias de larvasliberadas en la Isla Caimán Grande, sugieren que lapoblación de langosta de esta isla pudiera abastecer laspoblaciones de Jamaica, República Dominicana-Haití. Losrecalos que llegan a Cuba de la Isla Caimán Grandebenefician en mayor cuantía la región NW de Cuba, conel 10 % del total desovado.
El conocimiento de las trayectorias de las larvas delangosta en el Caribe ofrece una perspectiva en elentendimiento de la conectividad entre las poblacionesde la plataforma cubana y áreas vecinas, es útil paramejorar los diseños de las redes de reservas marinas enla región y sugiere la conservación de los hábitas en elCaribe.

69
MATERIALES Y MÉTODOS
Área de estudio
El Mar Intra-Americano es el escenario seleccionado paralas simulaciones de las trayectorias de larvas de langosta,
comprendido en la cuadrícula (8°-30° N y 60°-92° W).En dos polígonos más pequeños, localizados al NW(22°-24° N y 80°50'-86° W) y SW (19°-22° N y80°50'-86° W) de Cuba, se determinan característicasestadísticas del campo de corrientes y del campo deremolinos.
Fuente: Atlas GEBCO, www.ngdc.noaa.gov/mgg/gebco.
Fig. 1 Ubicación geográfica de Cuba en el Mar Intra-Americano
El Mar Intra-Americano comprende las aguas del Golfode México y del Mar Caribe (Mooers & Maul, 1998). Envirtud de su localización también es conocido como MarMediterraneo. El archipiélago cubano ocupa la posicióncentral de este mar. Bordeando la plataforma noroccidentalde Cuba fluyen fuertes corrientes oceánicas como la delLazo y la corriente de la Florida, las cuales modifican elpatrón de circulación inmediato a esta plataforma, mientrasla frontera sur del archipiélago cubano la limitan débilesflujos del Mar Caribe.
La topografía submarina al NW de Cuba muestraamplias llanuras abrasivo-acumulativas que se prolonganpor el estrecho de la Florida y se conectan al Mar Caribepor el estrecho de Yucatán. En el Mar Caribe existencinco cuencas principales. De estas, las próximas a laregión suroccidental de Cuba son la cuenca de Caimán(15o-20o N y 75o-80o W), con una profundidad máximade 7 100 m y la cuenca de Yucatán con una profundidadpromedio de 5 000 m. Los rasgos notorios de la topografíasubmarina del Mar Caribe como dorsales y pasosangostos entre islas o entre plataformas submarinas,pueden originar remolinos y meandros de mesoescala(Molinari et al., 1981). Los pasos y estrechos que rodean
a Cuba facilitan el paso de estos remolinos de mesoes-cala, que pueden trasladarse en el flujo principal desderegiones lejanas como el norte de Brasil (Johns et al., 2003).
Datos
Los datos de hidrodinámica provienen de una serie decorrientes entre los años 1995 y 2000 obtenida de lossatélites ERS1/2. Esta serie de corrientes se utilizó enla modelación de las trayectorias de larvas virtuales. Ladata de 18 cruceros oceanográficos realizados en aguascubanas entre los años 1966 y 2000, se utilizó paracaracterizar el campo de vórtices de la región SW deCuba (Gutiérrez et al., 2009).
Método lagrangiano
El cálculo de las trayectorias de las larvas virtuales serealizó mediante la ecuación de Langevin (Sawford,2001):
dx/dt = u = u* + ud, (1)dy/dt = v = v* + vd, (2)

70
donde:(x, y): posición de la partícula.(u*, v*): velocidad de cada posición de la partícula.(ud, vd): son seleccionadas de manera que llevan unadistribución normal.
Para calcular la posición de la partícula después deun tiempo Δt, [x(t + Δt), y(t +Δt)] partiendo del punto[x(t), y(t)] se usa el método Runge-Kutta de segundoorden:
x(t + Δt) = x(t) + k2, (3)
donde:
k1 = Δt u[t, x(t)] y k2 = Δt u[t + Δt/2, x(t) + k1/2].
De forma similar se calcula y(t + Δt) usando lavelocidad v. El tiempo t se mide en minutos y el campode corrientes (u, v) se mantiene constante durante cincodías. Para calcular la velocidad en los puntos de latrayectoria de la partícula se utiliza la interpolación bilinearde la velocidad euleriana con el método inverso de ladistancia, que toma cuatro puntos de la rejilla entre loscuales está situada la partícula.
Selección de la componente aleatoria
La generación de registros de velocidades aleatorias(ud, vd) se obtiene mediante el método de Monte Carlosujeto a ciertas restricciones estadísticas. El ruidoaleatorio fue generado usando procesos estocásticos deOrnstein-Uhlenbeck (Numerical Recipes, 1985), el cualse expresa de la siguiente forma:
ud = u* – (Δt/Tu) u* + (Δt/Tu σou w)1/2, (4)vd = v* – (Δt/Tv) v* + (Δt/Tv σov w)1/2, (5)
donde:
Tu, Tv: tiempo lagrangiano integral.σou, σov: denotan las desviaciones estándar de lascomponentes horizontales u y v de la velocidad.w: ruido gaussiano con media cero y desviación estándaruno.
En el proceso Ornstein-Uhlenbeck las velocidadesaleatorias varían en cada punto entre determinadoslímites, que dependen del coeficiente de difusión variable.
Simulación de las trayectorias de larvas
Las simulaciones de larvas virtuales están basadas enun modelo biofísico el cual integra datos de corrientescada cinco días, entre los años 1995 y 2000. El campode circulación se acopla a un modelo estocásticolagrangiano (ver ecuaciones 3; 4 y 5) para simular la
trayectoria de larvas virtuales liberadas en la regiónsuroccidental de Cuba, en Campeche (Yucatán) y en laIsla Caimán Grande. Las trayectorias fueron calculadasintegrando en el tiempo la velocidad horizontal de lasseries de corrientes.
Para determinar la resolución espacial del modelo fuenecesario conocer las características de los diámetrosde los vórtices más frecuentes. La dimensión de unvórtice coherente está dada por el radio de deformaciónde Rossby (Rd). Este radio oscila entre 37 y 38 km en laregión SW de Cuba y entre 22 y 23 km en la región NWde Cuba (Gutiérrez & Álvarez, 2008). Para representarun vórtice, Franks (2008) sugiere que se tome al menoscinco puntos de rejilla sobre el Rd, lo cual implica que eltamaño de la rejilla Δx debe variar entre 4 y 7 km, siconocemos que el Rd está entre 22 y 38 km. Oey et al.(2003) toman el criterio empírico Δx/Rd ≈ 1/3 o menospara la selección del tamaño Δx, aunque este criterio esraramente satisfecho en los modelos, en los quegeneralmente se utiliza Δx = Rd.
La producción del desove se simuló con la liberaciónde 26 912 larvas, cada cinco días en los meses de máximodesove durante seis años consecutivos (1995-2000). Seliberaron larvas virtuales en 58 sitios de la región SW deCuba (Fig. 2). Estos sitios corresponden a lugares dondese conoce la dinámica poblacional de la langosta (Cruzet al., 1991a). Entre los 58 puntos seleccionados aparecendiez puntos de registros históricos de abundancia de larvas(Alfonso et al., 1991).
–85 –84,5 –84 –83,5 –83 –82,5 –82 –81,5
Longitud
Latit
ud
23
22,5
22
21,5
21
Fig. 2 Distribución espacial de las 58 localidadesseleccionadas para el desove virtual en la región
suroccidental de Cuba
Para conocer el aporte de larvas de Yucatán e IslaCaimán Grande a la población de langosta en Cuba, sesimuló la liberación de larvas virtuales desde diezlocalidades de estas regiones (TABLA 1).

71
TABLA 1. Coordenadas de puntos de liberación de larvas virtuales en Cuba, Yucatán e Isla Caimán
Cuba Longitud Latitud Yucatán Longitud Latitud Caimán Longitud Latitud
Cayo Cantiles –82,002 21,609 1 –86,970 20,253 1 –81,399 19,222Cayo Cantiles 2 –82,120 21,650 2 –86,834 20,409 2 –81,262 19,247Cayo Avalos –82,168 21,519 3 –86,732 20,597 3 –81,160 19,272Hicacos –82,370 21,564 4 –86,680 20,875 4 –81,109 19,296Hicacos 2 –82,410 21,564 5 –86,578 21,128 5 –81,041 19,321Matías –82,444 21,562 6 –86,663 21,431 6 –81,109 19,394Cayo Los Indios –83,244 21,833 7 –86,663 21,807 7 –81,194 19,394Cayo Los Indios 2 –83,344 21,840 8 –87,055 21,979 8 –81,330 19,402Cayo Campos –82,404 21,602 9 –87,856 22,094 9 –81,484 19,402Cayo del Rosario –82,801 21,700 10 –89,645 22,028 10 –81,484 19,312
Al modelo biofísico se le incorporaron las caracterís-ticas biológicas de la langosta P. argus como es el períodode duración larval (PDL). Las simulaciones comienzan enlos meses de máximo desove de la especie (febrero-marzo-abril-mayo) y finalizan cuando concluye el PDL. Losniveles de reclutamiento fueron escalados con mortalidadnatural de 0,9d–1 (Baisre, 2008, comunicación personal).
Las larvas que arriban a algún sustrato sólido antesque finalice el PDL son cuantificadas, mientras que laslarvas que quedan fuera después que finaliza el PDL tienenuna mortalidad de 100 %. El autoreclutamiento incluyeel reclutamiento local y el de otras regiones de Cuba. Elreclutamiento exportado es aquel que sale fuera de Cuba.
RESULTADOS Y DISCUSIÓN
Simulación de las trayectorias de larvasde langosta liberadas al SW de Cuba
Las larvas virtuales de langosta liberadas al SW de Cubatienen el mayor reclutamiento en Cuba, local o regional(Fig. 3). A la región suroccidental del archipiélagocubano arriba el 27,97 % del total de larvas liberadas.Estos resultados concuerdan con estudios que exponenun buen grado de acople entre la producción de larvasy el autoreclutamiento (Lee & Williams, 2000). Lamayor retención observada en la región SW de Cubaes consistente con los mayores recalos de puerulus(Cruz, et al., 1991) y con los reportes de grandespesquerías (Puga et al., 1991). La región NW de Cubaes la segunda en importancia, con el 16,8 % reclutadodel total liberado. En el SE de Cuba se retiene el 5,9 %,
mientras a la región NE de Cuba arriban cantidades máspequeñas en comparación con el resto de las regionesestudiadas.
Las simulaciones realizadas ofrecen cuantíaspotenciales del reclutamiento, sujetas a las variablesfísicas y biológicas empleadas en el modelo. Noobstante, para lograr el reclutamiento exitoso yposteriormente abundantes pesquerías, numerososfactores deben de acoplarse en el medio. El reclu-tamiento efectivo en cada región del Caribe tienelimitaciones particulares y entre ellas, una de gran pesoes la conservación de los hábitats.
La dinámica de los fluidos a que están expuestaslas larvas liberadas en el SW de Cuba favorece que noentren en su totalidad dentro del flujo principal quefluye hacia el canal de Yucatán. El seguimiento de lastrayectorias de partículas ejemplifica trayectoriascirculares y caóticas al SW de Cuba, indicando que losremolinos de esta región deben dominar sobre el flujomedio (Fig. 4). El SW de Cuba es una zona deintercambio y de mezcla, beneficiada por el tiempo depermanencia de las partículas en el área (diez meses)(Richardson, 2005). Este período es tiempo suficientepara que las larvas de langosta Panurilus argus y demuchas especies tropicales concluyan su PDL(Linderman et al., 2001).
En la figura 4 se observan las trayectorias de larvasvirtuales en un período de ocho meses. Las trayectoriasdescriben la abundancia de remolinos oceánicos alnoroeste y sur de Cuba. Dentro de los remolinos laslarvas permanecen cercanas, mientras que fuera de losvórtices las trayectorias son caóticas después quevencen el tiempo lagrangiano integral (Lacorata et al.,2001).

72
Fig. 3 Distribución en polígonos de los porcentajes de larvas virtuales que arriban al Caribe. Las larvas se liberaronen la región SW de Cuba. Retención en orden de importancia: Región SW de Cuba 27,97 %, NE de la península
de La Florida 23,2 %, Región NW de Cuba 16,8 %, Jamaica 7,9 %, Golfo de México 7 %,Región SE de Cuba 5,9 %, Yucatán 2 %
Fig. 4 Trayectorias lagrangianas de larvas virtuales de langosta liberadas al SW de Cuba en marzo de 1995
El campo de remolinos al SW de Cuba se caracterizapor la mayor cantidad de estas estructuras por encima dela latitud de 21° (área de interacción océano-plataforma)(Gutiérrez & Álvarez, 2008). Esta característica debeestar relacionada con la fricción que ejercen la topografíasubmarina y los cayos que bordean la plataforma. El efectode la fricción propicia la formación de pequeños vórticespor el fraccionamiento de los vórtices grandes en otrosmás pequeños.
Los vórtices más frecuentes en el área de interacciónocéano-plataforma tienen escalas entre 24 y 41 km. El radiode deformación de Rossby (Rd) concuerda con el de losremolinos más estables, que son los más frecuentes en laregión de interacción océano-plataforma. Esta particularidades favorable para la permanencia y el desarrollo de la faselarvaria de la langosta. El análisis de EOF realizado a una
serie de remolinos oceánicos adyacentes a la regiónsuroccidental de Cuba durante los años 1995-2000, nomuestra un patrón temporal que implique que el campo devórtices más frecuentes responda a una época climáticaespecífica (Gutiérrez & Álvarez, 2012). Por lo que ademásde la retención que pueden ejercer los remolinos oceánicos,otros factores y mecanismos deben de estar actuando enmúltiples escalas de manera simultánea.
Las simulaciones numéricas corroboran que lacontracorriente cubana que fluye al NW de Cuba, traenuevamente a la región SW de Cuba las larvas que fueronadvectadas hacia el N de Cuba por la corriente de Yucatán.La contracorriente exparse las larvas por el borde de laplataforma suroccidental cubana y también las traslada aotras islas en el Caribe. La comunicación oceánica entrela región NW y SW de Cuba, observada en las simulaciones
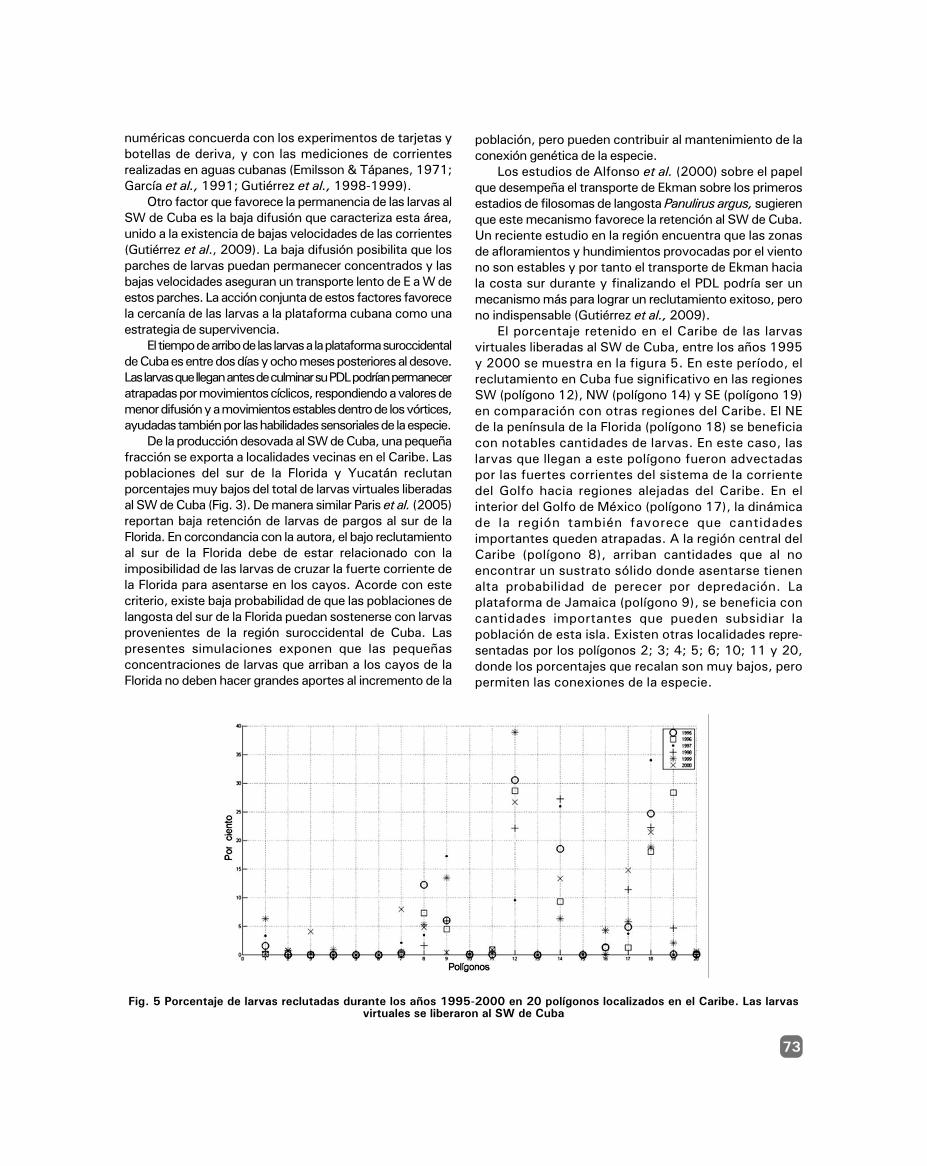
73
población, pero pueden contribuir al mantenimiento de laconexión genética de la especie.
Los estudios de Alfonso et al. (2000) sobre el papelque desempeña el transporte de Ekman sobre los primerosestadios de filosomas de langosta Panulirus argus, sugierenque este mecanismo favorece la retención al SW de Cuba.Un reciente estudio en la región encuentra que las zonasde afloramientos y hundimientos provocadas por el vientono son estables y por tanto el transporte de Ekman haciala costa sur durante y finalizando el PDL podría ser unmecanismo más para lograr un reclutamiento exitoso, perono indispensable (Gutiérrez et al., 2009).
El porcentaje retenido en el Caribe de las larvasvirtuales liberadas al SW de Cuba, entre los años 1995y 2000 se muestra en la figura 5. En este período, elreclutamiento en Cuba fue significativo en las regionesSW (polígono 12), NW (polígono 14) y SE (polígono 19)en comparación con otras regiones del Caribe. El NEde la península de la Florida (polígono 18) se beneficiacon notables cantidades de larvas. En este caso, laslarvas que llegan a este polígono fueron advectadaspor las fuertes corrientes del sistema de la corrientedel Golfo hacia regiones alejadas del Caribe. En elinterior del Golfo de México (polígono 17), la dinámicade la región también favorece que cantidadesimportantes queden atrapadas. A la región central delCaribe (polígono 8), arriban cantidades que al noencontrar un sustrato sólido donde asentarse tienenalta probabilidad de perecer por depredación. Laplataforma de Jamaica (polígono 9), se beneficia concantidades importantes que pueden subsidiar lapoblación de esta isla. Existen otras localidades repre-sentadas por los polígonos 2; 3; 4; 5; 6; 10; 11 y 20,donde los porcentajes que recalan son muy bajos, peropermiten las conexiones de la especie.
numéricas concuerda con los experimentos de tarjetas ybotellas de deriva, y con las mediciones de corrientesrealizadas en aguas cubanas (Emilsson & Tápanes, 1971;García et al., 1991; Gutiérrez et al., 1998-1999).
Otro factor que favorece la permanencia de las larvas alSW de Cuba es la baja difusión que caracteriza esta área,unido a la existencia de bajas velocidades de las corrientes(Gutiérrez et al., 2009). La baja difusión posibilita que losparches de larvas puedan permanecer concentrados y lasbajas velocidades aseguran un transporte lento de E a W deestos parches. La acción conjunta de estos factores favorecela cercanía de las larvas a la plataforma cubana como unaestrategia de supervivencia.
El tiempo de arribo de las larvas a la plataforma suroccidentalde Cuba es entre dos días y ocho meses posteriores al desove.Las larvas que llegan antes de culminar su PDL podrían permaneceratrapadas por movimientos cíclicos, respondiendo a valores demenor difusión y a movimientos estables dentro de los vórtices,ayudadas también por las habilidades sensoriales de la especie.
De la producción desovada al SW de Cuba, una pequeñafracción se exporta a localidades vecinas en el Caribe. Laspoblaciones del sur de la Florida y Yucatán reclutanporcentajes muy bajos del total de larvas virtuales liberadasal SW de Cuba (Fig. 3). De manera similar Paris et al. (2005)reportan baja retención de larvas de pargos al sur de laFlorida. En corcondancia con la autora, el bajo reclutamientoal sur de la Florida debe de estar relacionado con laimposibilidad de las larvas de cruzar la fuerte corriente dela Florida para asentarse en los cayos. Acorde con estecriterio, existe baja probabilidad de que las poblaciones delangosta del sur de la Florida puedan sostenerse con larvasprovenientes de la región suroccidental de Cuba. Laspresentes simulaciones exponen que las pequeñasconcentraciones de larvas que arriban a los cayos de laFlorida no deben hacer grandes aportes al incremento de la
Fig. 5 Porcentaje de larvas reclutadas durante los años 1995-2000 en 20 polígonos localizados en el Caribe. Las larvasvirtuales se liberaron al SW de Cuba

74
Larvas liberadas en Yucatán y en la IslaCaimán Grande
Para conocer el aporte de las poblaciones de langostacercanas a Cuba se simula la liberación de larvas virtualesen diez localidades de Campeche-Yucatán, durante losaños (1995-2000). De manera similar se liberan larvasvirtuales en la Isla Caimán Grande, en el mismo períodode estudio. La simulación de las larvas liberadas enYucatán muestra que el mayor porcentaje de las
partículas (61,9 %) es arrastrado por la corriente delGolfo y mantenido dentro del Golfo de México por losmovimientos cíclicos de las corrientes. Otra significantecantidad (33 %) es arrastrada hacia el NE de la penínsulade la Florida, siguiendo el fuerte flujo que mueve casi latotalidad de las partículas. La región noroccidental deCuba se subsidia con 2,8 % del total liberado, mientrasque regiones como la costa sur de la Florida recibencontribuciones aún menores (0,17 %), que no sustentanla población, pero ayudarían al mantenimiento de laconexión genética y demográfica de la especie (TABLA 2).
TABLA 2. Porcentaje del recalo de las larvas virtuales de langosta en el Caribe. Larvas virtuales liberadasen Yucatán y en la Isla Caimán Grande
Salida de Yucatán. Porcentaje Salida de la Isla PorcentajeRegiones de arribo del recalo Caimán Grande. del recalo
Regiones de arribo
Golfo de México 61,89 % Jamaica 56,54 %Costa NE de la Florida 33,17 % La Española 20,41 %Región NW de Cuba 2,83 % Región NW de Cuba 10,10 %Península de Yucatán 1,94 % Costa NE de la Florida 7,91 %Sur de la Florida 0,17 % Región SE de Cuba 1,66 %
Sur de la Florida 1,27 %Mar Caribe central 0,49 %Belice 0,39 %Golfo de México 0,29 %Región SW de Cuba 0,20 %Península de Yucatán 0,20 %Panamá-Colombia 0,20 %
Las larvas liberadas en Yucatán arriban a la regiónNW de Cuba en aproximadamente tres meses, cuandoaún no ha concluido el PDL. Pero, si se considera queestas larvas puedan permaner atrapadas en remolinoscoherentes hasta que culmine su PDL, entonces este oparte de este porcentaje podría ser considerado dentrode los subsidios de regiones vecinas, lo que seríabeneficioso para compensar las exportaciones que realizala población de Cuba a otras localidades en el Caribe.
La distribución en el Caribe de las larvas liberadas enla Isla Caimán Grande sugiere que esta isla ocupa unaposición estratégica en el Caribe (ver TABLA 2). El desoveproducido en la isla puede abastecer las poblaciones delangosta de Jamaica, República Dominicana-Haití. A estasregiones arriban significativos recalos de 57 y 21 %,respectivamente, lo que manifiesta una fuerte conexiónde la población de langosta entre estas islas.
De los recalos que llegan a Cuba, la región NW es lamás beneficiada con cerca del 10 % del desove de laIsla Caimán Grande, a continuación le sigue la región SEde Cuba con ∼ 2 % de recalos y con un porcentaje máspequeño se favorece la región SW de Cuba. En ladistribución de los porcentajes que arriban a Cuba influyela variabilidad del flujo principal del Caribe. Las intensascorrientes que fluyen muy cerca de la Isla Caimán Grandepueden advectar importantes concentraciones que seretienen al NW de Cuba, mientras que los pequeñosporcentajes que se contienen al SW de Cuba deben serfavorecidos por la presencia de remolinos oceánicos. Lascantidades significativas de larvas que exporta la IslaCaimán Grande, distribuidas principalmente en la regiónoccidental del Caribe demuestran las conexiones genéti-cas de la especie Panurilus argus, propuestas en el estudiode Espinosa et al. (1991).

75
CONCLUSIONES
1. Las larvas virtuales de langosta liberadas al SW deCuba tienen el mayor reclutamiento en Cuba, local oregional. A la región SW del archipiélago cubano arribael 27,97 % del total de larvas liberadas, lo quedemuestra un buen grado de acople entre la produc-ción y el autoreclutamiento.
2. En la región suroccidental de Cuba existen condicionesfísicas que propician la retención de las larvas delangosta. En la zona de interacción océano-plataforma,existe mayor cantidad de remolinos y son los másestables. El largo período de residencia de losremolinos oceánicos favorece el tiempo de perma-nencia de las larvas en el área. La baja difusión quecaracteriza la región posibilita que las concentracionesde larvas permanezcan cercanas y las bajas veloci-dades aseguran un transporte lento de E a W. La acciónconjunta de estos factores favorece la cercanía de laslarvas a la plataforma cubana lográndose una estrategiade retención.
3. El tiempo de arribo de las larvas a la plataformasuroccidental de Cuba es entre dos días y ocho mesesposteriores al desove. Las larvas que llegan antesde culminar su PDL permanecen atrapadas enmovimientos cíclicos, respondiendo a valores demenor difusión y a movimientos estables dentro delos vórtices, ayudadas también por las habilidadessensoriales de la especie.
4. Las poblaciones del sur de la Florida y Yucatánreclutan porcentajes muy bajos de las concentra-ciones liberadas al SW de Cuba. Los bajos nivelesde reclutamiento en estos lugares no deben hacergrandes aportes al incremento de la población, perocontribuyen al mantenimiento de la conexión genéticay demográfica de la especie.
5. Las trayectorias de las larvas liberadas en Yucatánmuestran que el mayor porcentaje (61,9 %) semantiene dentro del Golfo de México, por losmovimientos cíclicos de las corrientes. Otra cantidadsignificativa (33 %), es arrastrada hacia el NE de lapenínsula de la Florida, siguiendo el fuerte flujo quemueve casi la totalidad de las partículas. La regiónnoroccidental de Cuba se subsidia con 2,8 % deltotal liberado, mientras que regiones como la costasur de la Florida reciben contribuciones aún menores(0,17 %), que no sustentan las poblaciones, peroayudan a conectarlas.
6. Las larvas desovadas en la Isla Caimán Grandepueden abastecer las poblaciones de langosta deJamaica y República Dominicana-Haití. A estasregiones arriban significativos recalos que manifiestanuna fuerte conexión de la población de langosta entre
estas islas. De los recalos que llegan a Cuba, la regiónNW es la más beneficiada con cerca del 10 % deldesove de la Isla Caimán Grande.
AGRADECIMIENTOS
Los autores de este estudio agradecen la colaboracióny ayuda del doctor Julio Candela, del CICESE BajaCalifornia, México, por ceder parte de los códigos quese utilizaron en el modelo matemático. También agrade-cemos las valiosas recomendaciones de la doctoraConsuelo Valle, de la UABC. Con gran respeto agrade-cemos el trabajo desarrollado por las investigadorasMáster en Ciencias Pilar Frías e Irma Alfonso, en elestudio de la fase larvaria de especies que habitan lasaguas del Mar Caribe.
REFERENCIAS
Alfonso, I., Frías, M. P., Baisre, J. & Campos, A. (1991).Distribución y abundancia de larvas de la langostaPanulirus argus en aguas oceánicas alrededor deCuba. Rev. Inv. Mar., 12 (1-3), 5-19.
Alfonso, I., Frías, M. P. & Baisre, J. (2000). Distribuciónlarval de la langosta comercial Panulirus argus, enrelación con algunos factores hidrometeorológicos,al Sur de Cuba. Rev. Inv. Mar., 21 (1-3), 23-32.
Baisre, J. A., Blanco, W., Alfonso, I. & Ruiz de Quevedo,M. E. (1978). Distribución y abundancia relativa delas larvas de langosta (Panurilus argus) en el MarCaribe y las Bahamas. Rev. Cub. Inv. Pesq., 3 (1), 1-21.
Candela, J., Tanahara, S., Crepon, M., Barnier, B. &Sheinbaum, J. (2003). Yucatán Channel flow:Observations versus CLIPPER ATL6 and MERCATORPAM models. J. Geophys. Res., 108 (C12).
Cowen, R. & Sponaugle, S. (2009). Larval dispersal andmarine population connectivity. Annu. Rev. Mar. Sci.,1, 443-66.
Cowen, R. K., Paris, C. B. & Srinivasan, A. (2006). Scalingof connectivity in marine populations. SCIENCE, 311,522-527.
Cruz, R. & León, M. E. de (1991). Dinámica reproductivade la langosta Panurilus argus en el archipiélagocubano. Rev. Inv. Mar., 12 (1-3), 234-245.
Cruz, R., León, M. E. de, Díaz, E., Brito, R. & Puga, R.(1991a). Reclutamiento de puerulus de langostaPanurilus argus a la plataforma cubana. Rev. Inv.Mar., 12 (1-3), 66-75.
Emilsson, I. & Tápanes, J. J. (1971). Contribución a lahidrología de la plataforma sur de Cuba. SerieOceanológica, 9 (32).

76
Espinosa, G., Díaz, R. & Berovides, V. (1991). AnálisisGenético Bioquímico de Larvas de Panulirus argusde la ZEE del sur de Cuba. Rev. Inv. Mar., 12, 20-28.
Franks, P. (2008). Real-time coastal observing systemsfor marine ecosystem dynamics and hamful algalblooms. Theory, instrumentation and modelling. Paris,Francia: UNESCO.
García, C., Chirino, A. & Rodríguez, J. (1991). Corrientesgeostróficas en la Z.E.E al sur de Cuba. Rev. Cub.Inv. Pesq., 12 (1-3), 29-38.
Group, R. (1981). Gulf Stream cold core rings: theirphysics, chemistry and biology. Science, 212,1091-1100.
Guerrero, L. (2005): Identificación y seguimiento deremolinos en el Mar Caribe. Tesis de Maestría,CICESE. Baja California. México.
Gutiérrez, A. & Álvarez, A. (2008). Los remolinosoceánicos: mecanismos físicos de retención departículas. Contribución a la Educación y laProtección Ambiental, 8 (1), 185-196.
Gutiérrez, A. & Álvarez, A. (2012). Variabilidad espacialy temporal de los remolinos oceánicos en la regiónsuroccidental de Cuba. Serie Oceanológica, No. 11(Número Especial), ISSN 2072-800x.
Gutiérrez, A. et al. (2009). Proyecto: Los procesosoceanográficos y su relación con especies marinasde interés comercial. Importancia regional ytrascendencia extrarregional. Programa Ramal deCiencia e Innovación Tecnológica: Protección delMedio Ambiente y el Desarrollo Sostenible Cubano(1999-2009). ISBN 978-959-300-06-2
Gutiérrez, A., Pérez, J., Barbeito, R. & Moreau, P.(1998-1999). Estudio de las características hidroló-gicas y de la circulación de las aguas oceánicasalrededor de Cuba durante septiembre de 1998 ymayo de 1999. Informes C-T, GEOCUBA EstudiosMarinos, La Habana.
Hernández, B., Puga, R., Piñeiro, R., Peñate, A., Alfonso,I. & Baisre, J. (1995). Transport, turbulence andhypothetical ocean-shelf coupling mechanism in Puntadel Este, southwestern Cuba. Rev. Cub. Inv. Pesq.,19 (1), 67-72.
Jouanno, J., Sheinbaum, J., Barnier, B., Molines, J.,Debreu, L. & Lemarie, F. (2008). The mesoscalevariability in the Caribbean Sea. Part I: Simulationsand characteristics with an embedded model. OceanModelling, 23, 82-101.
Lacorata, G., Aurell, E. & Vulpiani, A. (2001). Drifterdispersion in the Adriatic Sea: Lagrangian data andchaotic model. Annales Geophysicae, 19, 121-129.
Lee, M. M. & Williams, R. G. (2000). The role of eddiesin the isopycnic transfer of nutrients and their impactof biological production. Journal of Marine Research,58, 895-917.
Linderman, K. C., Lee, T., Wilson, W. D., Claro, R. &Ault, J. S. (2001). Transport of larvae originating insouthwest Cuba and Dry Tortugas: Evidence forparcial retention in grunts and snappers. Proccedingsof the Gulf and Caribbean Fisheries Institute, 1 (52),732-747.
Mooers, C.N. K. & Maul, A. (1998). Intra-americas seacirculation. The Sea, 11, 183-197.
Molinari, R. L., Spillane, M., Brooks, I., Atwood, D. &Duckett, C. (1981). Surface currents in the CaribbeanSea deduced from Lagrangian observations. J.Geophys. Res., 86, 653-654.
Numerical Recipes (1985). The art of scientific computing.Cambridge University Press.
Oey, L. Y., Lee, H. C. & Schmitz, W. J. (2003). Efectsof winds and Caribbean eddies on the frequency ofLoop Current eddy shedding: A numerical modelstudy. J. Geophys. Res., 108, 22-25.
Paris, C. B., Cowen, R. K., Claro, R. & Linderman, K. C.(2005). Larval transport pathways from Cubansnapper (lutjanidae) spawning aggregations based onbiophysical modeling. Marine Ecology Progress Series,296, 93-106.
Puga, R., León, M. E. de & Cruz, R. (1991). Evaluaciónde la pesquería de langosta espinosa Pannulirus argusen Cuba. Rev. Inv. Mar., 12 (1-3), 286-292.
Richardson, P. L. (2005). Caribbean Current and eddiesas observed by surface drifters. Deep-Sea Research,52, 429-463.
Sheng, J. & Tang, L. (2003). A numerical study ofcirculation in the western Caribbean Sea. J. Phys.Oceanogr., 33, 2049-2069.
Sawford, B. (2001). Turbulent Relative Dispersion. J.Fluid. Mech., 33, 289-317.
Siam, C. L. (1988). Corrientes superficiales alrededorde Cuba. Rev. Cub. Inv. Pesq., 13 (1-2), 98-107.
Yeung, C. & Lee, T. N. (2002). Larval transport andretention of the spiny lobster Panulirus argus in thecoastal zone of the Florida Keys USA. Fish.Oceanogr., 11 (5), 286-309.

77
Revista Cubana de Investigaciones PesquerasEnero-diciembre, 2012, vol. 29, NO. 1, ISSN 0138-8452, pp. 77-79
Captured the shrimp tiger Penaeus monodon (Fabricius, 1798) in the Cuban’s costs
Enrique Giménez Hurtado,1 Lourdes Pérez Jar,1 Barbarito Jaime Ceballos,1 Ileana Fraga Castro,1
Redney Jiménez Cabrera,1 Dayamí Cabrera Vilaon2 y Ángela Moreno Urquiza3
1 Centro de Investigaciones Pesqueras. Ave. 5ta. y calle 246, Santa Fe, Playa,La Habana, Cuba, CP 19100, Teléfono: (537) (537) 209-8066,
E-mail: [email protected] 2 Empresa Pesquera Industrial. Ciego de Ávila
3 Empresa de desove de camarón YAGUACAM. Cienfuegos
RESUMEN
Se identificó una especie de camarón exótico como Penaeus monodon, camarón tigre gigante, mediante tresejemplares hembras capturados en lagunas costeras de Tunas de Zaza, golfo de Ana María, Cuba. La talla varióentre 270-290 mm de longitud total (Lt), con un peso entre 140-230 g. La presencia de la especie significa unaadaptación a las aguas cubanas y constituye una perspectiva positiva para su cultivo.
Palabras clave: especie exótica, Penaeus monodon, golfo de Ana María, Cuba.
ABSTRACT
It was identified an exotic shrimp species as Penaeus monodon, the giant tiger shrimp, by means of threefemales captured in coastal lagoons of Júcaro, golf of Ana María, Cuba. The length was from 270-290 mm totallength (Lt) weigh between 140-230 g. The presences of the species represented an adaptation to Cuban watersand constitute a positive perspective for shrimp culture.
Keysword: exotic species, Penaeus monodon, Ana María gulf, Cuba.
Capturado el camarón tigre Penaeus monodon (Fabricius, 1798)en las costas de Cuba
INTRODUCCIÓN
Penaeus monodon, el langostino jumbo, como tambiénse le conoce, es un crustáceo, decápodo marino criadocomercialmente para la alimentación. Su área dedistribución natural es de la región Indo-Pacífica, desdelas costas orientales de África, Arabia, alcanzando elsudeste asiático y el Mar de Japón. También se puedeencontrar en Australia, y un pequeño número hacolonizado el mar Mediterráneo, vía el Canal de Suez.Además se conoce de poblaciones que han invadidoHawai y el Océano Atlántico en las costas de EstadosUnidos (Florida, Georgia y Carolina del Sur).
Ambos sexos alcanzan unos 36 cm de longitud, ylas hembras pueden alcanzar un peso de 650 g,constituyendo el camarón más grande del mundo (Fig. 1).
En Cuba durante los años 2009-2011 se comienza aconocer que se han realizando capturas esporádicas tantoen lagunas costeras como en áreas de pesca comercial
de camarones de grandes proporciones, mayores a lasespecies presentes en nuestra plataforma, que hicieronpensar en la presencia de especies no reportadas ennuestras aguas. Con la finalidad de esclarecer estapresencia durante el mes de noviembre de 2011 serecibieron en el Centro de Investigaciones Pesqueras tresejemplares de la especie y se procedió a colectar informaciónrelacionada con su presencia en zonas de pesca.
Fig. 1 Ejemplar del camarón tigre gigante Penaeusmonodon

78
El siguiente reporte tiene como objetivo mostrar losejemplares capturados en dichas zonas, constituyendola primera evidencia documental de la presencia de unanueva especie en la plataforma cubana, discutiéndose laposible procedencia y movimiento migratorio de la misma.
MATERIAL EXAMINADO
Los camarones recibidos fueron capturados en lagunasdel poblado de Tunas de Zaza, un pueblo situado en lacosta del golfo de Ana María en la plataforma surorientalde Cuba. Los ejemplares fueron clasificados, medidos ysexados. Para la identificación taxonómica, se corroboraronlos caracteres morfológicos externos propuestos porPérez-Farfante & Kensley (1997), para la separación defamilias y géneros de camarones peneidos y sergestoideosdel mundo.
La identificación taxonómica evidenció que se tratabadel camarón tigre, P. monodon (Fabricius, 1798), organismooriundo de las costas de la región Indo-Pacífica (Fig. 2).
Fig. 2 Camarones recibidos en el mes de noviembre de2011 en el Centro de Investigaciones Pesqueras, Cuba
Las medidas morfométricas de los ejemplares recibidosmuestran que son tres hembras con telicum bien desarrolladoy característico de la especie (Fig. 3), presentan tallas entre27-29 cm de largo total con pesos entre 125-230 g.
Fig. 3 Telicum de una de las hembras mostrando unaprotuberancia bien desarrollada
TABLA 1. Biometría de camarones de la especiePenaeus monodon capturados en el poblado de Tunas
de Zaza, Cuba
Ejemplar Peso Largo Total Sexo Largo Total (LT) (cm)* cefalotórax (PT) (g) (LC)(cm)**
1 140,0 27,3 11,0
2 125,0 27,0 11,0
3 230,0 29,0 12,5
LT: (desde punta del rostrum hasta punta de la espina deltelson)LC: (desde la punta del rostrum hasta el extremo dorsalposterior del cefalotórax)
Motoh (1981) reporta que los machos silvestres deesta especie estarían sexualmente maduros a un LC de37 mm y PT de 35 g, mientras en las hembras lo estaríana 47 mm LC y 67,7 g PT, por lo que es de esperarse que lashembras capturadas hayan realizado desoves en el área.
Las encuestas realizadas en la zona de pesca comercialmostraron que se habían capturado dos ejemplares durantemarzo de 2010, una hembra de 20 cm frente al pobladode Júcaro y un macho de 18 cm en la zona cercana a ladesembocadura del río Zaza. Además se capturó unejemplar en la zona de pesca de Cabañones en la cayeríadel borde exterior del golfo de Ana María.
El camarón tigre P. monodon fue introducido porprimera vez en Cuba durante el año 1986 junto a lasespecies Litopenaeus vannamei (Boone, 1931) y L.stylirostris (Stimpson, 1874), para labores en acuiculturacomercial en granjas camaroneras. Su introducción serealizó a partir de postlarvas con tallas de 1-1,5 cm y semantuvieron en cuarentena durante 45 días en el AcuarioNacional de Cuba (Formoso el at., 1990). Durante lacuarentena se monitoreó la supervivencia en las condi-ciones climáticas de Cuba, siendo posteriormenteintroducidos ejemplares en un estanque de tierra de lagranja camaronera CULTIZAZA en el poblado de Tunasde Zaza, con el propósito de lograr tallas comerciales ycorroborar la posibilidad de su cultivo en el país (Formoso,comun. pers.). Según lo describen cultivadores de esepoblado, en el momento de su cosecha final solo secapturó un ejemplar adulto de la especie. Todo hacepensar que se produjo un escape de ejemplares al medionatural encontrando un ambiente apropiado de supervi-vencia en la laguna costera donde desembocan las aguasde la empresa camaronera mencionada y en la actualidadconstituye una especie exótica en las aguas cubanas. Elhecho que individuos adultos de camarón tigre se hayancapturado frente a las costas de Tunas de Zaza y marafuera, evidencia que la especie está reproduciéndose enel área y completando su ciclo biológico enteramente en


















