
Resultados de Avances de Investigación en … · Prolina se determinó por el método de Bates et...
38
Resultados de Avances de Investigación en Ecofisiología y Nutrición de Cítricos Programa Nacional en Producción 10 de Abril de 2013 y Sustentabilidad Ambiental Serie Actividades Programa Nacional de Citricultura de Difusión N° 708
Transcript of Resultados de Avances de Investigación en … · Prolina se determinó por el método de Bates et...

Resultados de Avances
de Investigación en Ecofisiología y Nutrición
de Cítricos
Programa Nacional en Producción 10 de Abril de 2013 y Sustentabilidad Ambiental Serie Actividades Programa Nacional de Citricultura de Difusión N° 708

Programa Nacional de Investigación en Producción y Sustentabilidad Ambiental Programa Nacional de Investigación en Citricultura
INIA Salto Grande 1
TABLA DE CONTENIDO Estudio de los cambios fisiológicos y metabólicos provocados por el anegamiento en mandarinas Satsumas y Clemenules…………………..……2 Estudios de la influencia del anegamiento y la temperatura del suelo sobre el desempeño de los portainjertos más usados en Uruguay…....…16 Viabilidad del Índice de Estrés Hídrico de los Cultivos en árboles cítricos………………………………………………………………………………..26 La distribución espacial de los árboles cítricos afecta el muestreo foliar y la interpretación del análisis foliar……………………………………..………..32 Fondo Concursable Interno FCI_02 : “Bases Fisiológicas para la Mitigación de la Vulnerabilidad de los Sistemas Productivos Agrícolas (arroz, cebada, cítricos, trigo, forestales) ante Estreses Abióticos Causados por el Cambio Climático en Uruguay”.

Programa Nacional de Investigación en Producción y Sustentabilidad Ambiental Programa Nacional de Investigación en Citricultura
INIA Salto Grande 2
Estudio de los cambios fisiológicos y metabólicos provocados por el anegamiento en mandarinas Satsumas y Clemenules.
Carmen Goñi1 y Alvaro Otero1 2
1Programa Nacional de Investigación en Producción Citrícola, 2 Programa Nacional de Producción y Sustentabilidad Ambiental. Instituto Nacional de Investigación Agropecuaria. [email protected]; [email protected]
Introducción:
La respuesta de los vegetales a los distintos tipos de estreses está condicionada a las diferentes estrategias que han desarrollado para sobreponerse a las mismas, que en forma general podemos englobar, según su génesis, en bióticos o abióticos. La variabilidad de las condiciones climáticas de Uruguay ocasiona periodos de estrés hídrico (déficit y excesos de agua) que condicionan los volúmenes y la calidad de la fruta para exportación.
El aumento de la intensidad en las precipitaciones puede provocar, dependiendo del tipo de suelo y sistema de plantación, situaciones temporales de falta de oxígeno en el suelo, debido al anegamiento del suelo.
En cítricos estas situaciones ambientales pueden tener consecuencias negativas importantes. Iwasaki (1975) señala el efecto depresivo que tiene la disminución del nivel de oxígeno en el suelo en los porcentajes de cuajado y desarrollo del fruto en mandarina Satsuma. Existen registros para la región citrícola del litoral noroeste del Uruguay que evidencian, como consecuencia de precipitaciones intensas en el periodo de octubre y noviembre (Otero et al., 2000), drásticas caídas de frutos pequeños, inmediatamente luego de caída de pétalos (Fig.1).
Figura 1. Relación entre la precipitación acumulada (mm) para el periodo setiembre y octubre, y el porcentaje de cuajado de frutos en la mandarina Satsuma Owari, (Otero et al., 2000).
Además, existen evidencias en Uruguay sobre el cambio actual del régimen
climático (Giménez et al., 2008), donde se aprecia la existencia de alteraciones en la distribución de las precipitaciones ocurridas para los meses de enero, febrero y marzo. Se registra un aumento real del orden de 15 a 35% en las precipitaciones a nivel del país para las tres últimas décadas estudiadas (1970-2000) en comparación al período 1930-1960 (fig.2).
Período Setiembre - Octubre
Precipitacion acumulada (mm)
140 150 160 170 180 190 200 210 220
Por
cent
aje
de C
uaja
do d
e Fr
uto
6
8
10
12
14
16
18
20
22
% CuajadoY = 45.382 - 0.176 Xr2=0.998

Programa Nacional de Investigación en Producción y Sustentabilidad Ambiental Programa Nacional de Investigación en Citricultura
INIA Salto Grande 3
Figura 2. Cambios actuales en la distribución de la pluviométrica para los meses de enero, febrero y marzo. Diferencia en los registros de precipitaciones (mm) entre los períodos 1930-960 y 1970-2000, (Giménez et al., 2008).
Aunque las condiciones de anegamiento afectan mucho a los frutales tropicales
y subtropicales (Phung et al., 1976), éstos difieren en su capacidad de tolerancia al anegamiento. Los cítricos están clasificados como un grupo medianamente tolerante (Ford, 1968). El anegamiento puede ocurrir en cualquier etapa crítica del desarrollo fenológico de los cultivos, pero los efectos suelen ser más severos cuando se dan en condiciones de crecimiento que en periodos de dormancia (Salesses et al., 1970). La intensidad y duración de las condiciones de anegamiento puede causar condiciones de anoxia o de hipoxia. Anoxia es la carencia de oxigeno en el suelo, mientras que hipoxia es la falta temporal de oxigeno en el suelo, es decir cuando el contenido de oxígeno del suelo es inferior al óptimo (Tadeo et al., 2008).
Los efectos del anegamiento afectan la fisiología de toda la planta, incluyendo
la fotosíntesis, transpiración, conductancia estomática (Davies y Flore, 1986), la traslocación del carbono y la absorción de nutrientes (Syvertsen, et al., 1983; Schaffer, 1991). La respuesta de la planta al anegamiento durante la estación de crecimiento incluye la inhibición del crecimiento vegetativo y reproductivo, cambios en la anatomía, senescencia temprana e incluso la muerte (Ericsson, 1989). La respuesta específica de los árboles a las condiciones de anegamiento varían según la época en la que se produce, la duración del estrés y también con la edad de los árboles (Kolozlowsky, 2002). Según varios autores la tolerancia de los cítricos al anegamiento está determinada por el portainjerto y no por la copa (Rowe y Beardsell, 1973; Shaffer y Moon, 1990; Shaffer et al., 1992). La floración y el cuajado de frutos son componentes importantes del rendimiento (Guardiola, 1988) y dado que la reducción del nivel de oxígeno disminuye el cuajado en la mandarina Satsuma (Iwasaki, 1975), parecía oportuno la evaluación de este estrés en nuestras condiciones. El objetivo de este es conocer los cambios fisiológicos ocurridos a nivel de la planta en el campo debidos al estrés hídrico por anegamiento durante la floración y cuajado del fruto en la mandarina Satsuma Owari y Clemenules al final del invierno y comienzo de la primavera.
La hipótesis de trabajo formulada fue que las condiciones de anoxia en el suelo aumentan la caída de frutitos en ambas variedades por encima de lo normal.

Programa Nacional de Investigación en Producción y Sustentabilidad Ambiental Programa Nacional de Investigación en Citricultura
INIA Salto Grande 4
Materiales y Métodos
Los experimentos se instalaron en el campo experimental de INIA-Salto Grande en montes adultos, durante la primavera 2010, en Satsuma Owari sobre P. trifoliata, y en la primavera 2012 en Clemenules sobre P. trifoliata evaluándose el efecto fisiológico del anegamiento provocado por inundación del suelo.
Las condiciones de anegamiento se indujeron en el suelo a través de la aplicación de una lámina permanente de agua sobre el nivel del suelo, desde el inicio de la brotación hasta el final de la floración y desarrollo inicial del frutito.
Con este fin se confinó el sistema radicular de las plantas mediante la colocación de un anillo de chapa (30 cm por encima del suelo y hasta una profundidad de 40-50 cm) permitiendo solamente la percolación de agua en profundidad pero no la lateral (Fig. 3). De este modo se logró mantener el nivel del agua a unos 15-20 cm por encima del suelo, en forma continua, sobre el sistema radicular de la planta, logrando condiciones de anoxia persistentes (Fig.4). La naturaleza del suelo con la presencia de un horizonte Bt a los 40-50 cm permitió reducir la percolación del agua en profundidad. En el tratamiento testigo (sin anegar), las plantas fueron regadas en forma convencional. El riego se aplicó cada vez que el contenido del agua del suelo llegaba a un potencial de 30 cbar, evaluado mediante tensiómetros ubicados a dos profundidades.
Oxígeno en el suelo, estado hídrico de las plantas y Prolina. El contenido
de agua del suelo se midió a dos profundidades 0-20 y 20-40 cm usando los sensores de humedad de suelo (10HS Decagon). El oxigeno del suelo (Fig.5) se determinó con medidas de la tasa de difusión de oxigeno (ODR) usando cuatro electrodos de platino y uno de referencia (DIK 5100, Japón), siguiendo los cálculos propuestos por Poel (1960) y el potencial redox del suelo (ORP) con un electrodo combinado de platino (Horiba, Japón) expresado en mv. Las medidas de ODR y del potencial redox (ORP) se tomaron a los 10 cm de profundidad del suelo y se repitieron tres veces por árbol con una frecuencia de dos veces por semana.
El potencial xilemático de tallo (Ψtallo) fue medido semanalmente, al mediodía en tres hojas por árbol con una cámara de Scholander, tipo DIK 700 Japón (Scholander et al., 1965). Las hojas fueron embolsadas en sobres de nylon negro y cubiertas con papel de aluminio, dejadas estabilizar durante tres horas previo a las lecturas de forma de permitir el equilibrio del Ψhoja con el tallo.
Los niveles de Prolina y carbohidratos se determinaron en muestras de 10 hojas por árbol. Las hojas se congelaron en nitrógeno líquido y se liofilizaron. La Prolina se determinó por el método de Bates et al. (1973) y los carbohidratos por el procedimiento de Mehouachi et al. (1995).
Al finalizar cada experimento de anegamiento se tomaban muestras de suelo alrededor de cada árbol de 0-20 y 20-30 cm con un taladro de raíces con un diámetro de 12 cm. Las muestras de suelo con raíces se refrigeraron en forma inmediata y posteriormente fueron lavadas para separar las raíces del suelo con un lavador de raíces (DeltaT) y posteriormente liofilizadas. El contenido de Prolina también se determinó en hojas.
Medidas de intercambio de gases y daño del fotosistema II en las hojas.
En los tratamientos se midió la asimilación neta de CO2 (Aco2), la conductancia estomática (gs) y se calculó la tasa de transpiración (E) en hojas individuales con un sistema portable de fotosíntesis (CIRAS-1, PP Systems, Haverhill, MA). Todas las medidas fueron hechas entre las 8:30 y 10:30 am cuando la radiación fotosintéticamente activa fue superior a 800 µmol m-2 s-1. Las medidas se realizaron

Programa Nacional de Investigación en Producción y Sustentabilidad Ambiental Programa Nacional de Investigación en Citricultura
INIA Salto Grande 5
cada cuatro días, en 3 hojas de 6 meses de edad en cada uno de los árboles de los experimentos, durante el periodo que duró cada uno.
El daño del fotosistema II fue evaluado a través de la medición de la emisión de la fluorescencia de la clorofila de la hoja usando un fluorómetro de pulso-modulado (OS5-FL, OptiScience, USA), operando con el test Fv/Fm en hojas adaptadas a la oscuridad por lo menos por 20 min (van Kooten y Snel, 1990). Se realizaron cada cuatro días, tres medidas en diferentes partes del árbol en cada uno de los árboles del experimento.
Tasa relativa de caída de fruta. En seis ramas previamente marcadas y
numeradas por árbol, de aproximadamente 50 cm de largo se siguió semanalmente la evolución del número de frutitos según la metodología de Agustí et al. (1992) junto al número de nudos y flores. La tasa relativa (Hunt, 1990) de caída de fruta se calculó como el cociente entre:
Ln (Frutitos 1) – Ln (Frutitos 2) Tasa Relativa de Caída de Frutitos = (Tiempo 2-Tiempo 1) La intensidad y largo de los brotes vegetativos se midió con un marco de 1 m2
(Albrigo et al., 1975) ubicado en las cuatro direcciones cardinales de cada árbol. Se utilizaron 9 plantas para cada tratamiento, en tres parcelas de tres plantas
cada una sorteadas al azar en un diseño de parcelas al azar. Se utilizaron los procedimientos clásicos estadísticos siguiendo el procedimiento GLM del SAS (SAS Institute, Cary, NC) y las medias fueron separadas por el test de rango múltiple Duncan a p<0,05. Resultados y Discusión
Figura 3. Anillo de chapa y plantas anegadas de Clemenules sobre P. trifoliata.

Programa Nacional de Investigación en Producción y Sustentabilidad Ambiental Programa Nacional de Investigación en Citricultura
INIA Salto Grande 6
Figura 4. Condiciones de anegamiento en las parcelas instaladas.
Figura 5. Vista de los electrodos de platino para contenido de oxigeno (ODR) izquierda y potencial de oxido reducción (ORP) derecha del suelo.
0
2
4
6
8
10
04-oct 09-oct 14-oct 19-oct 24-oct 29-oct
µg O
2 cm
-2 m
in-1
AnegadoNo Anegado
‐150
‐100
‐50
0
50
100
150
200
250
300
350
400
3‐oct 8‐oct 13‐oct 18‐oct 23‐oct 28‐oct 2‐nov
Potencial O
xido
Red
uccción de
l Sue
lo
(mv) Anegado
No Anegado
Figura 6. Tasa de Difusión de Oxigeno en el suelo para Owari (izquierda) y Potencial de Oxido Reducción en Clemenules (derecha).
0.00
0.02
0.04
0.06
0.08
0.10
0.12
0.14
07‐oct 17‐oct 27‐oct 06‐nov 16‐nov 26‐nov 06‐dic
Tasa Relativa de
caída
de frutos Anegado
No Anegado
Programa Nacional de Investigación en Producción y Sustentabilidad Ambiental Programa Nacional de Investigación en Citricultura
INIA Salto Grande 7
0.00
0.02
0.04
0.06
0.08
0.10
0.12
0.14
5‐oct 25‐oct 14‐nov 4‐dic 24‐dic
Tasa Relativa de
Caída
de Frutos
Anegado
No_Anegado
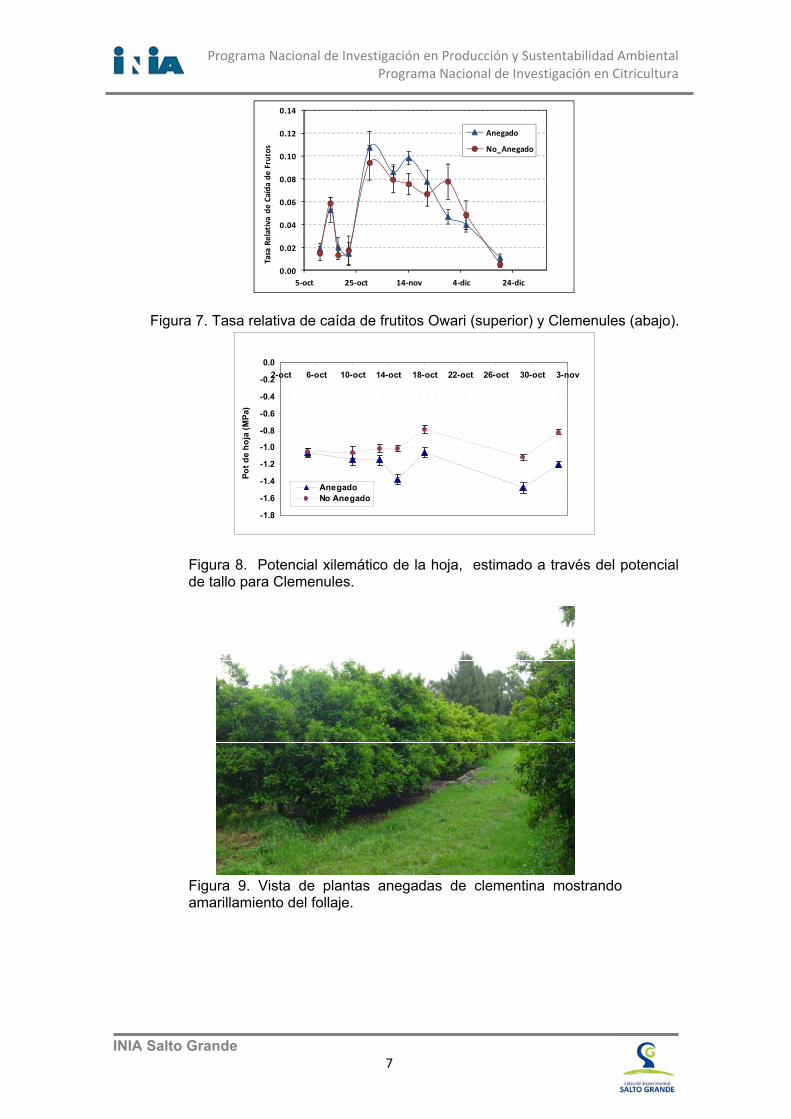
Figura 7. Tasa relativa de caída de frutitos Owari (superior) y Clemenules (abajo).
-1.8
-1.6
-1.4
-1.2
-1.0
-0.8
-0.6
-0.4
-0.2
0.02-oct 6-oct 10-oct 14-oct 18-oct 22-oct 26-oct 30-oct 3-nov
Pot d
e ho
ja (M
Pa)
AnegadoNo Anegado
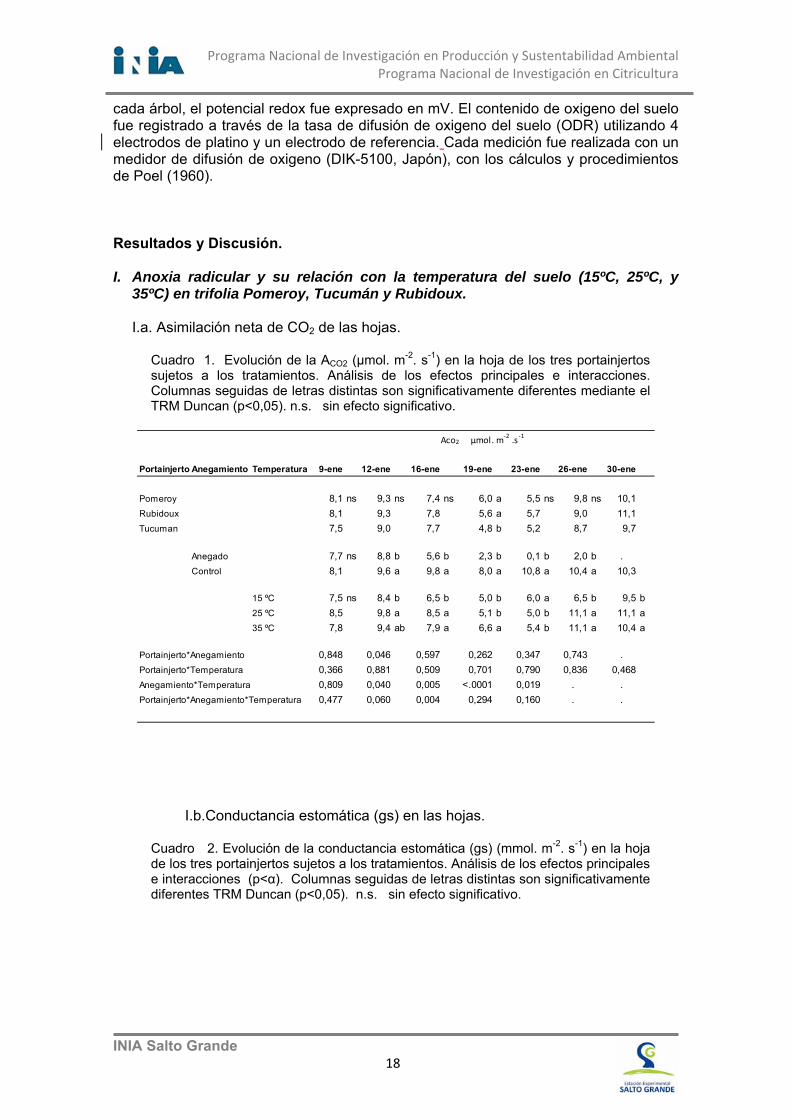
Figura 8. Potencial xilemático de la hoja, estimado a través del potencial de tallo para Clemenules.
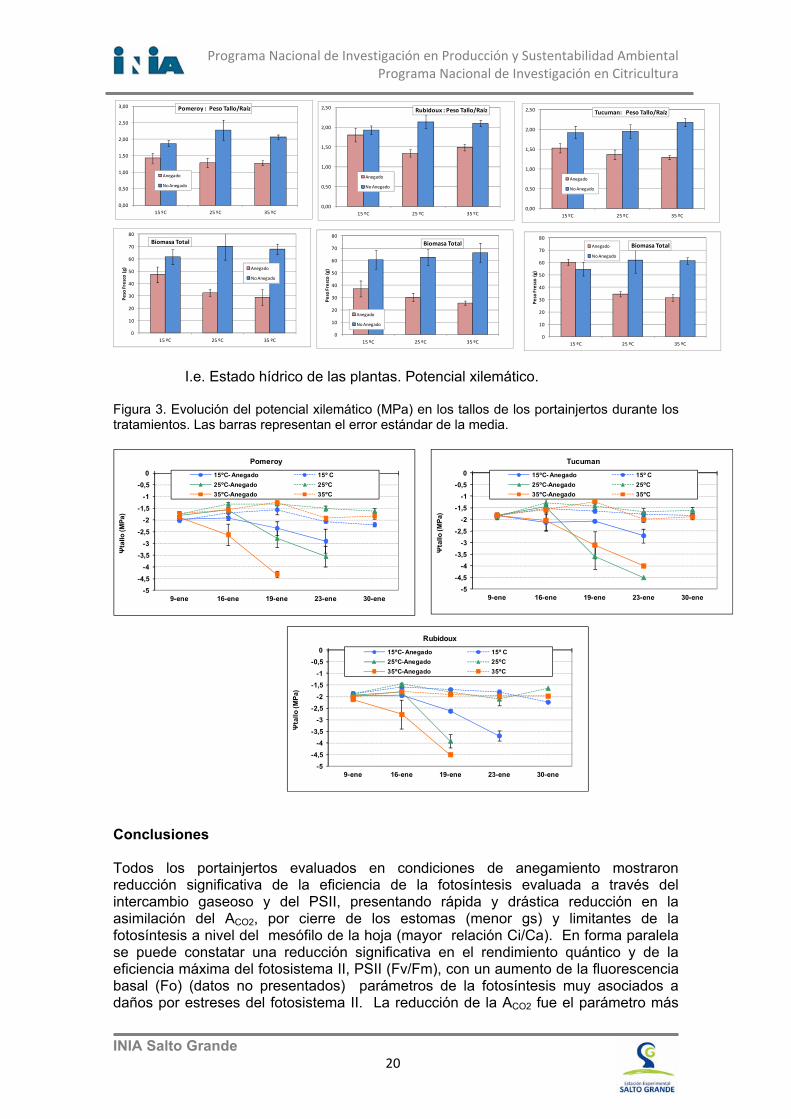
Figura 9. Vista de plantas anegadas de clementina mostrando amarillamiento del follaje.

Programa Nacional de Investigación en Producción y Sustentabilidad Ambiental Programa Nacional de Investigación en Citricultura
INIA Salto Grande 8
Clementina
40424446485052545658
5-oct 9-oct 13-oct 17-oct 21-oct 25-oct 29-oct 2-nov
Indi
ce S
PAD
AnegadoNo Anegado
Figura 10. Comparación de los niveles estimados de clorofilas (índice SPAD) entre plantas anegadas y no anegadas).
0
10
20
30
40
50
60
70
Anegamiento Testigo
N° Brotes / m2
Longitud del Brote (cm)
Figura 11. Efecto del anegamiento en la brotación de primavera. Las barras verticales representan el error estándar de la media (n=9).

Programa Nacional de Investigación en Producción y Sustentabilidad Ambiental Programa Nacional de Investigación en Citricultura
INIA Salto Grande 9
Satsuma Owari Clementina Clemenules
ACO2
0.0
1.0
2.0
3.0
4.0
5.0
6.0
7.0
4-oct 9-oct 14-oct 19-oct 24-oct 29-oct 3-nov
µmol
m-2
s-1
AnegadoNo Anegado
2
6
10
14
3-oct 8-oct 13-oct 18-oct 23-oct 28-oct 2-nov
(µm
ol. c
m-2
. s-1
)
AnegadoNo Anegado
Gs
0
20
40
60
80
100
4-oct 9-oct 14-oct 19-oct 24-oct 29-oct 3-nov
mm
ol m
-2 s
-1
Anegado
No Anegado
50
150
250
350
450
550
3-oct 8-oct 13-oct 18-oct 23-oct 28-oct 2-nov
(mm
ol. c
m-2
.s-1
)AnegadoNo Anegado
Ci/Ca
0.20
0.30
0.40
0.50
0.60
0.70
0.80
4-oct 9-oct 14-oct 19-oct 24-oct 29-oct 3-nov
Ci/C
a
Anegado
No Anegado
0.72
0.76
0.80
0.84
0.88
0.92
3-oct 8-oct 13-oct 18-oct 23-oct 28-oct 2-nov
Ci/C
a
Anegado
No Anegado
Figura 12. Comparación de la evolución de la fotosíntesis entre plantas anegadas y no anegadas. Asimilación neta del CO2 (ACO2), la conductancia estomática (gs) y relación CO2 interno/ CO2 del aire (Ci/Ca) para las mandarinas Owari y Clemenules.

Programa Nacional de Investigación en Producción y Sustentabilidad Ambiental Programa Nacional de Investigación en Citricultura
INIA Salto Grande 10
0.400
0.450
0.500
0.550
0.600
0.650
0.700
0.750
5-oct 9-oct 13-oct 17-oct 21-oct 25-oct 29-oct 2-nov
Qua
ntum
Yie
ld
AnegadoNo Anegado
0.600
0.700
0.800
0.900
5-oct 9-oct 13-oct 17-oct 21-oct 25-oct 29-oct 2-nov
Fv_F
m
AnegadoControl
60
80
100
120
140
160
180
5-oct 9-oct 13-oct 17-oct 21-oct 25-oct 29-oct 2-nov
Fo
AnegadoNo Anegado
Figura 13. Evolución de los parámetros de funcionamiento del Fotosistema II (PSII): eficiencia máxima relación (Fv/Fm), fluorescencia basal de la hoja (Fo) y el rendimiento quántico (Yield) entre plantas anegadas y no anegadas.

Programa Nacional de Investigación en Producción y Sustentabilidad Ambiental Programa Nacional de Investigación en Citricultura
INIA Salto Grande 11
0
10
20
30
40
Anegado No Anegado
μM P
rolin
a /g
MS
Raíces
50
60
70
80
90
100
5-oct 9-oct 13-oct 17-oct 21-oct 25-oct 29-oct 2-nov
µM P
rolin
a/g
MS
Anegado No Anegado
Hojas
Figura 14. Diferencias entre los contenidos de Prolina en raíces (izquierda) a 20 cm de profundidad y los contenidos en hojas (derecha) para Satsuma Owari.
Figura 15. Vista de la floración brotación 2011 de Satsuma Owari entre plantas no anegadas (frente) y las anegadas al fondo.
0
40
80
120
160
Flores en inflorescencias sin hoja
Flores en inflorescencias con hoja
Nº d
e flo
res/
100
nudo
s Anegado
No Anegado
Figura 16. Intensidad de la floración al año siguiente del anegamiento de las plantas de Satsuma Owari.

Programa Nacional de Investigación en Producción y Sustentabilidad Ambiental Programa Nacional de Investigación en Citricultura
INIA Salto Grande 12
Figura 17. Comparación de la Floración 2011 de plantas de Satsuma Owari anegadas (izquierda) y no anegadas (derecha).
A los pocos días de comenzados los experimentos en Owari y Clemenules fue posible observar diferencias en el contenido de oxigeno del suelo y en el potencial de oxido reducción (Fig. 6), estas diferencias entre los tratamientos indican que se dieron condiciones de anoxia. Bajo estas condiciones, las raíces de las plantas cambian su metabolismo a condiciones bioquímicas de aclimatación a la falta de oxígeno (anoxia). El primer síntoma observado es la reducción de la fotosíntesis (Fig. 12) seguido por la deshidratación (Fig. 8) y la caída de frutitos (Fig. 7). La deshidratación de las plantas está acompasada por la reducción en el potencial xilemático de la hoja, que es estimado a través del potencial de tallo (Fig. 8). Los árboles anegados experimentan una progresiva reducción del potencial Ψhoja a valores cercanos a -1,4 MPa, mientras que las plantas no anegadas permanecen estables entre -0,8 y -1,1 MPa.
En paralelo ocurre la caída de las estructuras florales y de pequeños frutitos.
Esta caída podría estar relacionada a las concentraciones 1-aminociclopropano 1 carboxílico (ACC) o al acido abscísico (ABA) como señales originadas en las raíces y transportadas hacia las hojas. El ACC es un precursor del etileno y responsable de la caída de los frutitos (Arbona y Gómez Cadenas, 2008). La anoxia también redujo el crecimiento vegetativo del flujo de primavera; en el caso de Satsuma Owari con una reducción del número de brotes y no del largo de los mismos (Fig.11), en clementinas las plantas no brotan.
El anegamiento redujo la fotosíntesis, la asimilación neta de CO2 (ACO2), la
conductancia estomática (gs) y el rendimiento quántico (Yield) del fotosistema II (PSII) en las dos variedades (Fig.12). El cierre estomático en condiciones de anegamiento ocurre en muchas especies leñosas (Kozlowski, 2002) y está asociado a una disminución de la conductividad hidráulica en las raíces (Syvertsen et al., 1983; Davies y Flore, 1986). Estos mecanismos pueden ser de restricción del agua o respuestas a cambios hormonales (Arbona y Gómez Cadenas, 2008).
La reducción de la conductancia estomática (gs) estuvo correlacionada con los
descensos en la asimilación neta de CO2 (ACO2). Esto no es claramente evidente al comienzo de los experimentos, cuando aún la diferencia del contenido de oxigeno en el suelo no es muy pronunciada entre las plantas. Cuando se instalan las condiciones de anoxia las diferencias en el funcionamiento del sistema fotosintético de las hojas se hace muy evidente entre plantas anegadas y no anegadas (Fig..6).

Programa Nacional de Investigación en Producción y Sustentabilidad Ambiental Programa Nacional de Investigación en Citricultura
INIA Salto Grande 13
En Satsuma se dieron cambios menores en la concentración interna de CO2, lo que sugiere que los factores estomáticos fueron más importantes que los factores no estomáticos en limitar ACO2 (García- Sánchez et al., 2007) y no hubieron cambios en la eficiencia quántica máxima del PSII (Fv/Fm) y en la fluorescencia basal (Fo). En Clemenules se dan cambios similares en la asimilación neta de CO2 (ACO2), pero a diferencia de Satsuma Owari se reduce en forma significativa (Fig.13) la eficiencia quántica (Fv/Fm) y aumenta la florescencia basal (Fo).
En Satsuma se observaron cambios significativos en los niveles de Prolina de
las raíces de las plantas anegadas, pero no a nivel de las hojas (Fig. 14). La Prolina es un osmolito compatible que contribuye a incrementar el turgor de las plantas (Correa Molinari et al., 2004) y a reducir el estrés oxidativo de las mismas. Además, se dio un incremento en el contenido de almidón de las hojas pero no en los azúcares reductores (datos no mostrados). En Clemenules aún no disponemos de esa información.
Conclusiones
Como se estudió el efecto del anegamiento del suelo sobre la caída de frutitos en experimentos similares, en Satsuma Owari durante (2010-11) y en Clemenules (2012), podemos hacer algunas comparaciones del comportamiento entre ambas mandarinas. Las condiciones de anegamiento tanto en Satsuma Owari como en Clemenules aumentaron significativamente la caída de estructuras florales y frutitos (Fig.7). Esta caída de frutitos y la tasa relativa de caída de los mismos, ha sido significativamente superior en los árboles anegados.
En las plantas anegadas de Satsuma Owari y Clemenules se produjeron
reducciones importantes del contenido de agua en las hojas y tallos, evidenciada por la reducción del potencial xilemático (Fig. 8), con una deshidratación y marchitamiento significativo de las plantas.
En Satsuma Owari, la deshidratación y abarquillamiento de las hojas no se
producen con cambios en el color de follaje (nivel de clorofilas e índice SPAD), sin embargo en los árboles de Clemenules (Fig.9) los cambios en el color del follaje comienzan rápidamente, las plantas anegadas “amarillean” muy rápido (Fig. 10). En ambas variedades se produjeron cambios significativos en la brotación de primavera, mientras dura el estrés y posteriormente al mismo. En Satsuma Owari se redujo el número y no el largo de los brotes (Fig. 11); mientras que las plantas anegadas de Clemenules no brotan.
En ambas variedades, el anegamiento redujo la fotosíntesis (Fig.12), la ACO2,
gs y el rendimiento quántico (Yield) del fotosistema II (PSII). Esto no es claramente evidente al comienzo del experimento, cuando aún la diferencia del contenido de oxígeno en el suelo no es muy pronunciada entre las plantas, mientras que cuando se instalan las condiciones de bajos potenciales redox (anoxia) las diferencias en el funcionamiento del sistema fotosintético de las hojas se hacen más evidente entre plantas anegadas y no anegadas (Fig.6 y 12). En Satsuma se dieron cambios menores en la concentración interna de CO2 y sin cambios en la eficiencia quántica máxima del PSII (Fv/Fm) y en la fluorescencia basal (Fo). Esto sugiere que factores estomáticos fueron más importantes que los factores no estomáticos en limitar la ACO2. En Clemenules (Fig.13) a diferencia de Satsuma Owari se reduce en forma significativa la eficiencia quántica (Fv/Fm) y aumenta la florescencia basal (Fo).

Programa Nacional de Investigación en Producción y Sustentabilidad Ambiental Programa Nacional de Investigación en Citricultura
INIA Salto Grande 14
En las plantas anegadas de Satsuma el contenido de Prolina fue más alto en las raíces que en las hojas (Fig.14) y se da una acumulación significativa del contenido de almidón en hojas, pero no en el contenido de azúcares solubles (fructosa, glucosa, sacarosa) y finalmente se dan cambios del patrón de floración del siguiente año (Fig.15, 16 y 17). En clementinas aún no disponemos de esa información.
De la comparación de ambos experimentos, podemos concluir que la mandarina Clementina de nules sería más sensible a la asfixia radicular que la mandarina Satsuma durante el período de floración y desarrollo inicial del fruto. Bibliografía Agustí M., Almela, V. y J. Pons. 1992. Effects of girdling on alternate bearing in citrus.
J. Hortic. Sci. 67: 203-207. Albrigo, L.G., Anderson, G.J., Edwards, G.J., Bistline, F.W., Hepburn, W.J. y T. Cary.
1975. Yield estimates of Valencia orange research plots and groves. Proc. Fla. State Hortic. Soc. 88:44-49.
Armstrong, W. 1978. Root aeration in the wetland condition. In: Plant life in anaerobic environment. Ed: Hook, DD, Cradford. Amer. Arbor. Sci., Ann Arbor. Michigan, 269-297.
Arbona, V. y A. Gómez Cadenas. 2008. Hormonal modulation of citrus responses to flooding. J. Plant. Growth Regul. 27:241-250.
Arbona, V., Hossain, Z., López-Climent, M.F., Pérez-Clemente, R.M. y A. Gómez-Cadenas. 2008. Antioxidant enzymatic activity is linked to waterlogging stress tolerance in citrus. Physiologia Plantarum 132:452-466.
Bates, L.S., Waldren, R.P. y I.D. Teare. 1973. Rapid determination of free proline for water stress studies. Plant Soil 39: 205-207.
Correa Molinari, H B., Marur, C.J., Bespalhok (Jr), J.C, Kobayashi, A.K. Pileggi, M., Pereira Leites (Jr), R., Protasio Pereira, L. y L.G. Esteves Vieira. 2004. Osmotic adjustment in transgenic citrus rootstock Carrizo citrange (Citrus sinensis Osb. X Poncirus trifoliata L. Raf.) overproducing proline. Plant Science 167 (6):1375-1381.
Davies, F.S. y J.A. Flore. 1986. Flooding, gas exchange and hydraulic conductivity of highbush blueberry. Phydiol. Plant. 67: 545-551.
Erikson, M.E. 1989. Survival of plant materials established on a floodplain in central Oklahoma. Wildl. Sci. Bull 17:63-65.
Ford, H.W. 1968. Water management of wetland Citrus in Florida. Proc. 1st. Int. Citrus Symp. Vol 3: 1759-1770.
García-Sánchez, F., Syvertsen, J.P., Gimeno, V., Botía, P. y J.G. Perez-Perez. 2007. Responses to flooding and drought stress by two citrus rootstocks seedlings with different water use efficiency. Physiologia Plantarum.130:532-542.
Giménez A., Castaño J.P., Baethgen, W. y B. Lanfranco. 2008 Cambio Climático en Uruguay, posibles impactos y medidas de adaptación en el Sector Agropecuario. INIA. Serie Técnica 178.
Guardiola, J.L. 1988. Factors limiting productivity in citrus. A physiological approach. Proc. Sixth Citrus Cong.1: 381-394.
Hunt, R. 1990. Basic Growth Analysis. Unwin Hyman Ltd. London. UK. pp 111. Iwasaki, K. 1975. Flowering, Fruit Set Development, Size and Quality of Satsuma
Mandarin as Affected by Oxygen Content of Soil Atmosphere. J. Japan Soc. Hort. Sci. 44(3) 228-234.

Programa Nacional de Investigación en Producción y Sustentabilidad Ambiental Programa Nacional de Investigación en Citricultura
INIA Salto Grande 15
Kozlowski, T.T, 2002. Aclimatation and adaptative responses of woody plants to environmental stresses. The Botanical Review. 68(2):270-234.
Maxwell, K. y G.N. Johnson. 2000. Chlorophyll fluorescence –a practical guide. J Exp Bot 51: 659-668.
Mehouachi, J., Serna, D., Zaragoza, S., Agusti, M., Talon, M. y E. Primo-Millo. 1995. Defoliation increases fruit abscission and reduces carbohydrates levels in developing fruit and woody tissues of Citrus unshiu. Plant Sci. 107: 189-197.
Pezeshki, S.R. y J.L.Chamber. 1985. Stomatal and photosynthetic response of sweet gum (Liquidambar styraciflua) to flooding. Can. J. For. Res. 5:371-375.
Poel, L.W. 1960. The estimation of oxygen diffusion rates in soils. Jour of Ecology Vol 48:1 165-173.
Ortuño, M.F., Alarcón, J.J., Nicolas, E. y A.Torrecillas. 2007. Water status indicators of lemon trees in response to flooding and recovery. Biologia Plantarum 51(2) 292-296.
Otero, A., Ogata, R., Ishiakawa, K. y T. Kihara. 2000. Predicción del rendimiento a través de la evaluación de la floración en citrus. Resultado de Investigación Protección de árboles frutales 1995/2000. Proyecto INIA-JICA. 39-42.
Phung, H.T. y E.B. Knipling. 1976. Photosynthesis and transpiration of citrus seedlings under flooded conditions. HortSci. 11:131-133.
Rowe R.N. y D.V. Beardsell. 1973. Waterlogging of fruit trees. Hort.Abstr.43:534-543. Schaffer, B., P.C. Andersen, y R.C. Ploetz. 1992. Responses of fruit crops to flooding,
p. 257-313. In. J. Janick (ed.). Horticultural reviews. vol. 13.Wiley, New York. Salesses, G., Saunier, R. y A. Bonnet. 1970. Lásphyxie radiculare chez les arbres
frutiers. Bull. Tech. Inf. 251. p 1-13. Schaffer, B. y P.A. Moon. 1990. Influence of rootstock on flood tolerance of Tahiti lime
trees. Proc. Fla. State Hort. Soc. 103:318-321. Schaffer, B. 1991. Flood tolerance of Tahiti lime rootstocks in south Florida soil. Proc.
Fla. State Hort. Soc. 104:31-32. Syvertsen, J.P., R.M. Zablotowicz, y M.L. Smith. 1983. Soil temperature and flooding
affects two species of citrus. I. Plant growth and hydraulic conductivity. Plant Soil 72:3-12.
Scholander, P.E., Hammel, H.T., Bradstreet, E..D. y E.A. Hemmingsen. 1965. Sap pressure in vascular plants. Science 148:339-346.
Tadeo, FR, Gomez-Cadenas, A. Fisiología de las plantas y el estrés. En: Azcón-Bieto J, Talón M (eds.). Fundamentos de Fisiología Vegetal, 2 ed. Madrid, McGraw-Hill/Interamericana, 2008.
van Kooten, O. y J.F.H. Snell. 1990. The use of chlorophyll fluorescence nomenclature and plant stress physiology. Photosynth. Res. 25: 147-150.

Programa Nacional de Investigación en Producción y Sustentabilidad Ambiental Programa Nacional de Investigación en Citricultura
INIA Salto Grande 16
Estudios de la influencia del anegamiento y la temperatura del suelo sobre el desempeño de los portainjertos más usados en Uruguay.
Alvaro Otero 1; Carmen Goñi 1
1 Programa Nacional de Investigación en Producción Citrícola. Instituto Nacional de Investigación Agropecuaria. INIA Salto Grande. Uruguay. [email protected]; [email protected] Objetivo. Determinar la respuesta fisiológica a la temperatura y al anegamiento del suelo en los portainjertos más usados en la citricultura uruguaya y explorar el efecto de la copa en los cambios fisiológicos manifestados. La hipótesis a probar es si la interacción de las condiciones de temperatura y anegamiento del suelo inciden en forma distinta en el comportamiento fisiológico observado en estos portainjertos. Estrategia. Se diseñaron e implementaron 4 grupos de experimentos bajo invernáculo, algunos de los cuales se realizaron 2 o 3 veces para comprobar su repetibilidad.
• Interacción del anegamiento y la temperatura del suelo (15⁰C, 25⁰C y 35⁰C) en Citrange Carrizo, Troyer y Trifolia (CT33).
• Interacción del anegamiento y la temperatura del suelo (15⁰C, 25⁰C y 35⁰C) en Poncirus trifoliata: Pomeroy, Tucumán y Rubidoux.
• Tolerancia al anegamiento con temperatura del suelo constante (25ºC) en Citrange Carrizo, Citrumelo, CT33, Rubidoux, Troyer y Tucumán.
• Tolerancia al anegamiento en combinaciones de copa y portainjerto: naranja navel Fukumoto, naranja Salustiana y mandarina satsuma Okitsu, injertadas sobre Citrange Carrizo y Trifolia (CT33).
Se van a presentar dos de los cuatro grupos de experimentos planificados. Materiales y Métodos. Se seleccionaron plantas de portainjertos cítricos de 11-12 meses de edad; de portainjertos emparentados genéticamente pero con diferente comportamiento durante el invierno e inicio de primavera, respecto a la brotación, madurez de la fruta y calidad, tamaño de la canopia. Se trabajó en condiciones controladas de invernáculo definiéndose tres niveles de temperatura del suelo (15⁰C, 25⁰C y 35⁰C) y dos condiciones de oxigenación del mismo: condiciones normales y condiciones de restricción de O2 en el suelo por medio del anegamiento. Cada tratamiento estuvo compuesto de nueve plantas para cada portainjerto (3 plantas/parcela con 3 repeticiones) Los tratamientos de temperatura del suelo se realizaron poniendo las macetas en recipientes más grandes (bandejas de 1 x 2 m), donde por pasaje de agua caliente en tuberías finas de cobre en el suelo, se calentaban hasta la temperatura deseada. Para condiciones de temperatura más bajas de la ambiental se hizo circular una corriente de agua fría que, por transmisión de energía, bajaba la temperatura del suelo. El

Programa Nacional de Investigación en Producción y Sustentabilidad Ambiental Programa Nacional de Investigación en Citricultura
INIA Salto Grande 17
encendido y apagado del sistema fue automáticamente regulado ubicando sensores de temperatura en la zona radicular de las plantas Los tratamientos de anegamiento se realizaron anegando las macetas hasta saturación (Figura 1) y manteniendo el nivel del agua en forma constante, 2-3 cm sobre la superficie del suelo.
Figura 1. Vista de una maceta anegada y otra no anegada Los parámetros evaluados fueron:
• Biomasa radicular, tallos y hojas. • Contenido de clorofila y SPAD. Potencial hídrico en tallo (Ψtallo). • Contenido relativo de agua en la hoja. • Evolución del intercambio de gases (Pn): Aco2, gs, , Ci/Ca. • Actividad del Fotosistema II: Fo, Fv/Fm, Quantum Yield. • Evolución del contenido de oxígeno en el suelo (ODR) y potencial redox (ORP). • Condiciones ambientales (temperatura y HR%) del invernáculo y de las
macetas.
Potencial xilemático de tallo (Ψtallo). Se midió en 2 hojas por planta en todas las plantas de los tratamientos. Se utilizaron hojas maduras de la parte media del tallo. A cada hoja se la envolvió en un sobre de plástico negro, que a su vez fue recubierto por una hoja de papel aluminio. Luego de 3 h a cada hoja se le midió el potencial hídrico con una cámara de presión (Scholander, 1965) asumiendo que las hojas estaban en equilibrio con el Ψtallo (Barry et al. 2004).
Medidas de intercambio de gases y daño del fotosistema II en las hojas.
Durante los tratamientos se evaluó la asimilación neta de CO2 (Aco2), la conductancia estomática (gs) y la tasa de transpiración (E) en hojas individuales con un sistema portable de fotosíntesis (CIRAS-1, PP Systems, Haverhill, MA). Todas las medidas fueron hechas entre las 8:00 y 10:00 am cuando la radiación fotosintéticamente activa era superior a 800 µmol m-2 s-1, o con suplemento de luz artificial. El daño del fotosistema II fue evaluado a través de la medición de la emisión de la fluorescencia de la clorofila de la hoja usando un fluorómetro de pulso-modulado (OS5-FL, OptiScience, USA), operando con el test Fv/Fm en hojas adaptadas a la oscuridad por lo menos por 20 min (van Kooten y Snel, 1990). Se realizaron tres medidas en diferentes partes del árbol.
Medidas del contenido de oxigeno y potencial redox en el suelo. En cada
maceta se midió el potencial redox (ORP) del suelo a los 10 cm con un electrodo combinado de platino (Horiba, Japón), la medida bajo la copa se repitió tres veces en
Programa Nacional de Investigación en Producción y Sustentabilidad Ambiental Programa Nacional de Investigación en Citricultura
INIA Salto Grande 18
cada árbol, el potencial redox fue expresado en mV. El contenido de oxigeno del suelo fue registrado a través de la tasa de difusión de oxigeno del suelo (ODR) utilizando 4 electrodos de platino y un electrodo de referencia. Cada medición fue realizada con un medidor de difusión de oxigeno (DIK-5100, Japón), con los cálculos y procedimientos de Poel (1960).
Resultados y Discusión. I. Anoxia radicular y su relación con la temperatura del suelo (15ºC, 25ºC, y
35ºC) en trifolia Pomeroy, Tucumán y Rubidoux.
I.a. Asimilación neta de CO2 de las hojas.
Cuadro 1. Evolución de la ACO2 (µmol. m-2. s-1) en la hoja de los tres portainjertos sujetos a los tratamientos. Análisis de los efectos principales e interacciones. Columnas seguidas de letras distintas son significativamente diferentes mediante el TRM Duncan (p<0,05). n.s. sin efecto significativo.
Portainjerto Anegamiento Temperatura 9-ene 12-ene 16-ene 19-ene 23-ene 26-ene 30-ene
Pomeroy 8,1 ns 9,3 ns 7,4 ns 6,0 a 5,5 ns 9,8 ns 10,1Rubidoux 8,1 9,3 7,8 5,6 a 5,7 9,0 11,1Tucuman 7,5 9,0 7,7 4,8 b 5,2 8,7 9,7
Anegado 7,7 ns 8,8 b 5,6 b 2,3 b 0,1 b 2,0 b .Control 8,1 9,6 a 9,8 a 8,0 a 10,8 a 10,4 a 10,3
15 ºC 7,5 ns 8,4 b 6,5 b 5,0 b 6,0 a 6,5 b 9,5 b25 ºC 8,5 9,8 a 8,5 a 5,1 b 5,0 b 11,1 a 11,1 a35 ºC 7,8 9,4 ab 7,9 a 6,6 a 5,4 b 11,1 a 10,4 a
Portainjerto*Anegamiento 0,848 0,046 0,597 0,262 0,347 0,743 .Portainjerto*Temperatura 0,366 0,881 0,509 0,701 0,790 0,836 0,468Anegamiento*Temperatura 0,809 0,040 0,005 <.0001 0,019 . .Portainjerto*Anegamiento*Temperatura 0,477 0,060 0,004 0,294 0,160 . .
Aco2 µmol. m‐2 .s ‐1
I.b.Conductancia estomática (gs) en las hojas.
Cuadro 2. Evolución de la conductancia estomática (gs) (mmol. m-2. s-1) en la hoja de los tres portainjertos sujetos a los tratamientos. Análisis de los efectos principales e interacciones (p<α). Columnas seguidas de letras distintas son significativamente diferentes TRM Duncan (p<0,05). n.s. sin efecto significativo.

Programa Nacional de Investigación en Producción y Sustentabilidad Ambiental Programa Nacional de Investigación en Citricultura
INIA Salto Grande 19
Portainjerto Anegamiento Temperatura 9-ene 12-ene 16-ene 19-ene 23-ene 26-ene 30-ene
Pomeroy 173,8 ns 299,0 ns 153,8 ns 152,6 a 120,4 ns 284,2 ns 206,5 nsRubidoux 157,7 301,5 163,7 143,9 a 154,2 228,1 243,9Tucuman 153,1 276,4 154,3 108,4 b 131,2 233,8 197,9
Anegado 157,4 ns 283,4 ns 96,2 b 57,9 b 25,7 b 40,7 bControl 165,9 301,9 220,2 a 195,7 a 241,3 a 284,4 a 216,1
15 ºC 159,6 ns 253,8 b 116,1 b 104,0 130,2 b 174,6 b 195,4 b25 ºC 166,0 302,2 a 179,2 a 135,1 124,2 c 303,0 a 235,7 a35 ºC 159,2 330,6 a 175,0 a 176,3 151,9 a 302,5 a 217,3 a
Portainjerto*Anegamiento 0,836 0,397 0,679 0,025 0,060 0,786 .Portainjerto*Temperatura 0,191 0,654 0,379 0,163 0,498 0,695 0,828Anegamiento*Temperatura 0,088 0,024 0,010 0,000 0,143 . .Portainjerto*Anegamiento*Temperatura 0,394 0,155 0,004 0,226 0,283 . .
Gs mmol. m-2 .s-1
I.c. Eficiencia quántica del PSII en las hojas.
Cuadro 3. Evolución de la eficiencia cuántica del PSII del mesófilo de las hojas de los portainjertos bajo los tratamientos. Análisis de los efectos principales e interacciones (p<α). Letras distintas en una misma columna representan diferencias significativas a través del TRM Duncan (p<0,05). n.s. sin efecto significativo
Portainjerto Anegamiento Temperatura 9-ene 12-ene 16-ene 19-ene 23-ene 26-ene 30-ene
Pomeroy 0,70 ns 0,73 ns 0,62 ns 0,58 ns 0,64 ns 0,69 ns 0,63 nsTucuman 0,69 0,74 0,60 0,59 0,69 0,70 0,65Rubidoux 0,70 0,73 0,61 0,56 0,63 0,68 0,63
Anegado 0,70 ns 0,73 ns 0,59 b 0,51 b 0,56 b 0,61 b 0,50 bTestigo 0,70 0,73 0,64 a 0,65 a 0,67 a 0,70 a 0,66 a
15ºC 0,70 0,72 b 0,63 a 0,66 a 0,61 b 0,66 c 0,61 b25ºC 0,70 0,74 a 0,62 a 0,59 b 0,65 a 0,70 b 0,65 a35ºC 0,69 0,73 a 0,59 b 0,49 c 0,67 a 0,72 a 0,67 a
Portainjerto*Anegamiento ns ns ns ns ns ns <0.0001Portainjerto*Anegamiento ns ns 0,048 ns ns ns nsAnegamiento*Temperatura ns ns ns <0.0001 ns ns nsPortainjerto*Anegamiento *Temperatura ns ns 0,004 ns ns ns ns
Rendimiento Cuántico PSII
I.d. Biomasa al final del experimento. Figura 2. Biomasa de los tres portainjertos: peso fresco (g) y relación peso tallo/peso raíz al final de los tratamientos. Barras corresponden al error estándar de la media. 18 plantas. 3 repeticiones por tratamiento.
Programa Nacional de Investigación en Producción y Sustentabilidad Ambiental Programa Nacional de Investigación en Citricultura
INIA Salto Grande 20
0,00
0,50
1,00
1,50
2,00
2,50
3,00
15 ºC 25 ºC 35 ºC
Pomeroy : Peso Tallo/Raíz
Anegado
No Anegado
0
10
20
30
40
50
60
70
80
15 ºC 25 ºC 35 ºC
Peso Fresco (g)
Biomasa Total
Anegado
No Anegado
0,00
0,50
1,00
1,50
2,00
2,50
15 ºC 25 ºC 35 ºC
Rubidoux : Peso Tallo/Raíz
Anegado
No Anegado
0
10
20
30
40
50
60
70
80
15 ºC 25 ºC 35 ºC
Peso Fresco (g)
Biomasa Total
Anegado
No Anegado
0,00
0,50
1,00
1,50
2,00
2,50
15 ºC 25 ºC 35 ºC
Tucuman: Peso Tallo/Raíz
Anegado
No Anegado
0
10
20
30
40
50
60
70
80
15 ºC 25 ºC 35 ºC
Peso Fresco (g)
Biomasa TotalAnegado
No Anegado
I.e. Estado hídrico de las plantas. Potencial xilemático. Figura 3. Evolución del potencial xilemático (MPa) en los tallos de los portainjertos durante los tratamientos. Las barras representan el error estándar de la media.
-5-4,5
-4-3,5
-3-2,5
-2-1,5
-1-0,5
0
9-ene 16-ene 19-ene 23-ene 30-ene
Ψta
llo (M
Pa)
Pomeroy
15ºC- Anegado 15º C25ºC-Anegado 25ºC35ºC-Anegado 35ºC
-5-4,5
-4-3,5
-3-2,5
-2-1,5
-1-0,5
0
9-ene 16-ene 19-ene 23-ene 30-ene
Ψta
llo (M
Pa)
Tucuman
15ºC- Anegado 15º C25ºC-Anegado 25ºC35ºC-Anegado 35ºC
-5-4,5
-4-3,5
-3-2,5
-2-1,5
-1-0,5
0
9-ene 16-ene 19-ene 23-ene 30-ene
Ψta
llo (M
Pa)
Rubidoux
15ºC- Anegado 15º C25ºC-Anegado 25ºC35ºC-Anegado 35ºC
Conclusiones Todos los portainjertos evaluados en condiciones de anegamiento mostraron reducción significativa de la eficiencia de la fotosíntesis evaluada a través del intercambio gaseoso y del PSII, presentando rápida y drástica reducción en la asimilación del ACO2, por cierre de los estomas (menor gs) y limitantes de la fotosíntesis a nivel del mesófilo de la hoja (mayor relación Ci/Ca). En forma paralela se puede constatar una reducción significativa en el rendimiento quántico y de la eficiencia máxima del fotosistema II, PSII (Fv/Fm), con un aumento de la fluorescencia basal (Fo) (datos no presentados) parámetros de la fotosíntesis muy asociados a daños por estreses del fotosistema II. La reducción de la ACO2 fue el parámetro más

Programa Nacional de Investigación en Producción y Sustentabilidad Ambiental Programa Nacional de Investigación en Citricultura
INIA Salto Grande 21
sensible a la anoxia y a la temperatura del suelo; a los 4 días de comenzado el experimento, estos parámetros ya fueron afectados significativamente por los factores ambientales. La anoxia provocó en éstos además una reducción importante del potencial xilemático (Ψtallo), existiendo marchitez y muerte de las plantas. Se produjeron cambios significativos en la biomasa con modificaciones en el crecimiento entre tallos y raíces entre los portainjertos. Fue constatado una reducción de la relación Tallos/Raíces y también en el contenido de las clorofilas (SPAD) (datos no presentados). Por otro lado, la temperatura del suelo y su interacción con el anegamiento ha jugado un papel importante y significativo en el desempeño fisiológico de estos portainjertos. Cuando se analiza solamente el efecto del genotipo se puede apreciar que las diferencias en los parámetros fisiológicos evaluados entre los tres tipos de Trifolia no fueron significativas, pero si fue significativa la interacción del tipo de portainjerto, con el anegamiento y la temperatura del suelo,. Desde el punto de vista práctico esto es importante ya que en este caso la respuesta al anegamiento y temperatura dependería del tipo de Trifolia. También es de destacar la interacción del anegamiento con la temperatura; en condiciones de anegamiento, la velocidad de la reducción de los parámetros de la fotosíntesis, como expresión del daño de todo el sistema de producción de carbohidratos, aumenta a medida que aumenta la temperatura del suelo. En la práctica a nivel de campo y particularmente en el área del litoral norte, es posible sugerir que no es lo mismo tener anegamiento del suelo en pleno verano, que tenerlo en invierno con temperaturas más bajas del suelo. En verano las condiciones de anoxia, como consecuencia del anegamiento temporal o permanente, se instalaría más rápidamente en los genotipos estudiados, no solo por la mayor actividad de los portainjertos en este período, sino también porque la actividad de la microflora es mayor con temperatura estivales altas, contribuyendo a reducir más rápidamente el oxigeno del suelo (Reichardt, W. et al. 2000). A los pocos días de instalada la anoxia, el potencial xilemático es menor en los portainjertos anegados, pero decrece más en relación a la temperatura. Las plantas a 35ºC se presentaban más deshidratadas que las plantas a 25ºC y a su vez que a 15ºC. Se evidenció una reducción importante en el crecimiento vegetativo de los Trifolias bajo anegamiento. En condiciones de crecimiento sin anegar, la temperatura del suelo es importante en el crecimiento vegetativo. Tucumán y Rubidoux mostraron un crecimiento constante para las tres temperaturas del suelo, mientras que Pomeroy presentó una leve pero significativa reducción del crecimiento a temperaturas bajas (15ºC). La diferencia de crecimiento entre plantas anegadas y no anegadas de los tres genotipos teniendo en cuenta la temperatura fue diferente. Rubidoux fue el portainjerto de mayor diferencia en el crecimiento a la temperatura de 15ºC (Figura 2).

Programa Nacional de Investigación en Producción y Sustentabilidad Ambiental Programa Nacional de Investigación en Citricultura
INIA Salto Grande 22
II. Anoxia radicular por anegamiento en combinaciones de portainjertos y copas de plantas de cítricos.
A los efectos de determinar la respuesta fisiológica de tres cultivares cítricos
(Fukumoto, Okitsu y Salustiana) al anegamiento del suelo en los dos portainjertos (C. Carrizo y P. trifoliata) más usados en la citricultura uruguaya. Para ello se evaluó la interacción de la copa (variedad) y el portainjerto en relación a la tolerancia al anegamiento y se comparó este comportamiento con el portainjerto sin injertar.
‐300
‐200
‐100
0
100
200
300
400
08/Sep 13/Sep 18/Sep 23/Sep 28/Sep
Potencial Red
ox del sue
lo (mV)
Anegada
Sin Anegar
Figura 4. Evolución del potencial redox de las macetas (mV). Las barras corresponden al desvío estándar de todas las medidas para cada fecha.
-4.0
-3.5
-3.0
-2.5
-2.0
-1.5
-1.0
-0.5
0.08-sep 12-sep 16-sep 20-sep 24-sep 28-sep
Pote
ncia
l de
tallo
(MPa
)
Fukumoto
Carrizo-Anegado Carrizo-s/AnegarTrifolia- Anegado Trifolia-s/Anegar
-4.0
-3.5
-3.0
-2.5
-2.0
-1.5
-1.0
-0.5
0.08-sep 12-sep 16-sep 20-sep 24-sep 28-sep
Pote
ncia
l de
tallo
(MPa
)
Okitsu
Carrizo-Anegado Carrizo-s/AnegarTrifolia -Anegado Trifolia-s/Anegar
Programa Nacional de Investigación en Producción y Sustentabilidad Ambiental Programa Nacional de Investigación en Citricultura
INIA Salto Grande 23
-4.0
-3.5
-3.0
-2.5
-2.0
-1.5
-1.0
-0.5
0.08-sep 12-sep 16-sep 20-sep 24-sep 28-sep
Pote
ncia
l de
Tallo
(MP
a)
Salustiana
Carrizo-Anegado Carrizo-s/AnegarTrifolia-Anegado Trifolia-s/Anegar
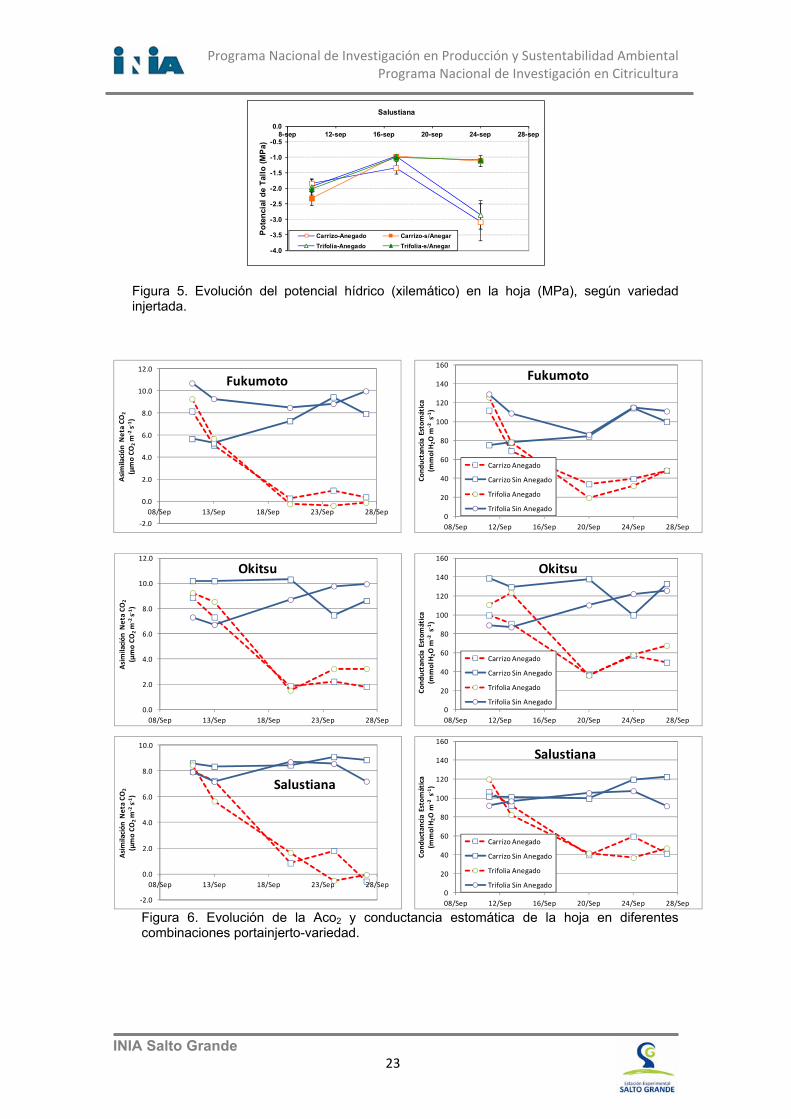
Figura 5. Evolución del potencial hídrico (xilemático) en la hoja (MPa), según variedad injertada.
‐2.0
0.0
2.0
4.0
6.0
8.0
10.0
12.0
08/Sep 13/Sep 18/Sep 23/Sep 28/Sep
Asimilación
Neta CO
2
(µmo CO
2m
‐2s‐1 )
Fukumoto
0
20
40
60
80
100
120
140
160
08/Sep 12/Sep 16/Sep 20/Sep 24/Sep 28/Sep
Cond
uctancia Estom
ática
(mmol H
2O m
‐2s‐1 )
Fukumoto
Carrizo Anegado
Carrizo Sin Anegado
Trifolia Anegado
Trifolia Sin Anegado
0.0
2.0
4.0
6.0
8.0
10.0
12.0
08/Sep 13/Sep 18/Sep 23/Sep 28/Sep
Asimilación
Neta CO
2
(µmo CO
2m
‐2s‐1 )
Okitsu
‐2.0
0.0
2.0
4.0
6.0
8.0
10.0
08/Sep 13/Sep 18/Sep 23/Sep 28/Sep
Asimilación
Neta CO
2
(µmo CO
2m
‐2s‐1 )
Salustiana
0
20
40
60
80
100
120
140
160
08/Sep 12/Sep 16/Sep 20/Sep 24/Sep 28/Sep
Cond
uctancia Estom
ática
(mmol H
2O m
‐2s‐1 )
Okitsu
Carrizo Anegado
Carrizo Sin Anegado
Trifolia Anegado
Trifolia Sin Anegado
0
20
40
60
80
100
120
140
160
08/Sep 12/Sep 16/Sep 20/Sep 24/Sep 28/Sep
Cond
uctancia Estom
ática
(mmol H
2O m
‐2s‐1 )
Salustiana
Carrizo Anegado
Carrizo Sin Anegado
Trifolia Anegado
Trifolia Sin Anegado
Figura 6. Evolución de la Aco2 y conductancia estomática de la hoja en diferentes combinaciones portainjerto-variedad.

Programa Nacional de Investigación en Producción y Sustentabilidad Ambiental Programa Nacional de Investigación en Citricultura
INIA Salto Grande 24
Discusión Las condiciones de anoxia, evaluadas a través del potencial oxido reductor del suelo, se expresaron en las macetas a partir de los 8 días de comenzar el experimento; la diferencia en el potencial oxido reducción entre las macetas anegadas y las no anegadas es altamente significativa (P<0.05) (Figura 4). A los 15 días de instalado el experimento, el Ψtallo de la hoja fue significativamente distinto entre plantas anegadas y no anegadas. La deshidratación de las hojas, evidenciada por la reducción del potencial xilemático, fue mayor en plantas anegadas. Esta diferencia es muy marcada en Salustiana, seguida de Okitsu y menos en Fukumoto. Además de la respuesta diferencial entre las variedades de copa, no se encontraron efectos significativos entre los portainjertos en Okitsu y Salustiana en condiciones de anegamiento; en el caso de las plantas injertadas con Fukumoto, estas fueron más tolerantes al anegamiento cuando estaban injertadas en Carrizo en comparación con Trifolia (Figura 5) al final del experimento. Es de destacar la velocidad de la deshidratación de las hojas de los cultivares; en poco más de 8 días desde que se instaló la falta de oxígeno en el suelo, los valores de las plantas bajo anoxia llegaron a ser críticos, hasta valores de no recuperación de la hidratación de las hojas y en consecuencia de muerte del tejido foliar. Por otro lado, la curva de reducción de la ACO2 y de Gs de las plantas anegadas sigue bastante ajustadamente la evolución del potencial redox del suelo, a los pocos días del comienzo del experimento. Esta relación no es tan evidente cuando se compara con la reducción de la fotosíntesis en portainjertos (Otero, et al. 2013). Conclusiones El sistema fotosintético de la hoja es altamente influenciado por las condiciones de anoxia en las macetas. La actividad de intercambio gaseoso de las hojas, debida mayormente a la fotosíntesis y a la transpiración, ha demostrado ser uno de los parámetros más sensibles al cambio de las condiciones de anoxia en el suelo. Casi inmediatamente de comenzar el anegamiento de las plantas, aun cuando las condiciones de reducción del oxigeno en el suelo se estaban desarrollando, los parámetros del intercambio gaseoso de la hoja fueron reducidos significativamente en las plantas anegadas; aun cuando otros parámetros fisiológicos relacionados con la fotosíntesis no lo manifestaban al comienzo (Figura 6). De acuerdo a la velocidad de reducción de los parámetros de la fotosíntesis y en comparación con los experimentos del comportamiento de los portainjertos sin injerto de copa.. Las plantas injertadas manifiestan más rápidamente los síntomas provocados por la anoxia, que los portainjertos solos. Muy pocos días de anegamiento dañan temporal o permanentemente el sistema fotosintético y deshidratan la planta injertada.

Programa Nacional de Investigación en Producción y Sustentabilidad Ambiental Programa Nacional de Investigación en Citricultura
INIA Salto Grande 25
Bibliografía Barry, G.H., W.S. Castel y F.S. Davies. 2004. Rootstocks and plant water relations
affect sugar accumulation of citrus fruit via osmotic adjustment. J. Amer. Soc. Hort. Sci. 129(6):881-889.
Otero, A.; C. Goñi y J. Syvertsen. 2013. Flooding and Soil Temperature Affect Water Relations and Photosynthesis of Citrus Rootstock Leaves. International Citrus Congress. Valencia. España.
Poel, L.W. 1960. The estimation of oxygen diffusion rate in soils. Journal of Ecology. 48 (1) 165-173.
Reichardt, W., Inubushi, K. y Tiedje, J. 2000. Microbiological processes in C and N dynamics. In. Carbon and Nitrogen Dynamics in Flooded Soils, Eds. G.J.D Kifk and D.C Olk. IRRI (International Rice Research Institute). Makati City. Philippines.188 p.
Scholander, P.F., H.T. Hammel, E.D. Bradstreet, y E.A. Hemmingsen. 1965. Sap pressure in vascular plants. Science 148:339-346.
van Kooten, O y J.F.H. Snel. 1990. The use of fluorescence nomenclature in plant stress physiology. Photosyn. Res. 25:147-150.

Programa Nacional de Investigación en Producción y Sustentabilidad Ambiental Programa Nacional de Investigación en Citricultura
INIA Salto Grande 26
Viabilidad del Índice de Estrés Hídrico de los Cultivos en árboles cítricos
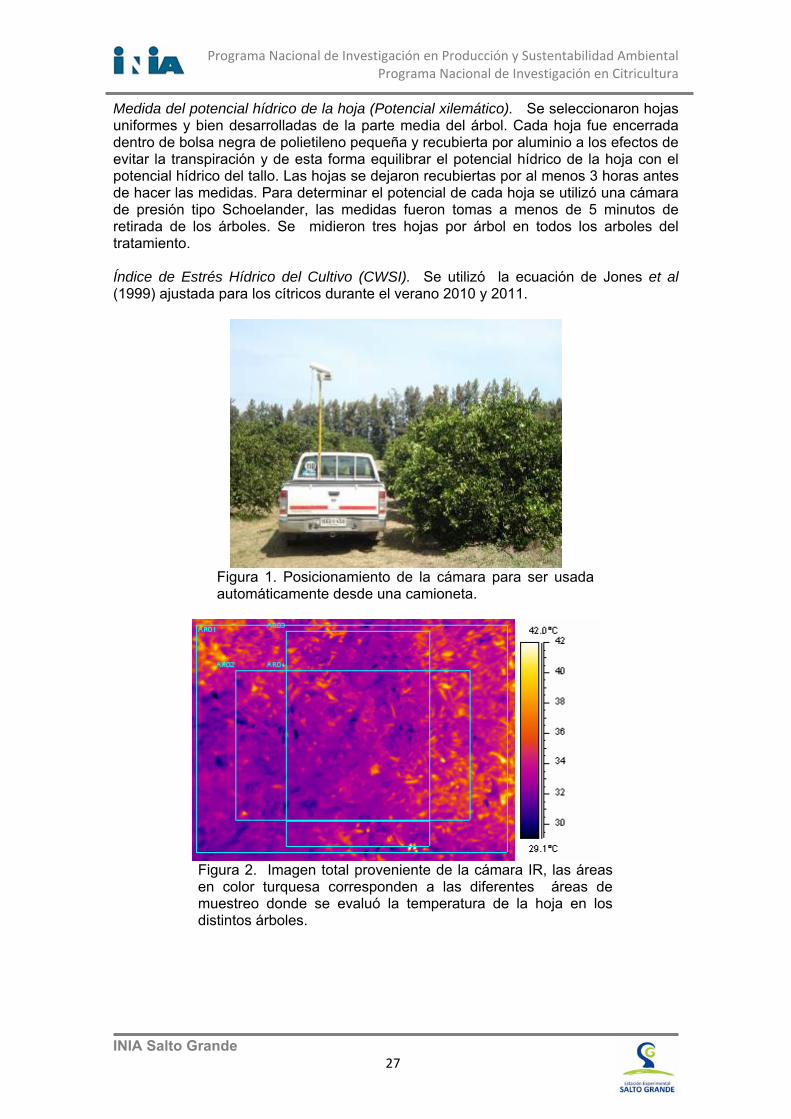
Alvaro Otero 1; Carmen Goñi 2; Andres Berger3 1 Programa Nacional de Investigación en Producción y Sustentabilidad Ambiental. 2 Programa Nacional de Investigación en Producción Citrícola. Instituto Nacional de Investigación Agropecuaria. INIA Salto Grande. Uruguay. 3 Programa Nacional de Cultivos de Verano [email protected]; [email protected] ; [email protected] Uno de los mecanismos más importantes que tienen las plantas para evitar el sobre calentamiento de la canopia es el aumento de la transpiración, principalmente realizado a través del aumento de la conductancia estomática de las hojas. En este sentido, la canopia de los cultivos necesita continuamente transpirar agua por sus estomas para mantenerse a pocos grados centígrados en torno a la temperatura ambiental. Cuando se dan condiciones de reducción del agua en el suelo, a niveles que son sensibles para la deshidratación de la planta, los estomas se cierran y en consecuencia, la temperatura de las hojas aumenta, independientemente de la temperatura del aire. Este fenómeno ya fue descripto y cuantificado por Idso (1981, 1982), con el fin de poder ser usado como índice del estado hídrico de la planta en los programas de riego y para evaluar el efecto de la sequía (Jackson et al., 1981). La ecuación desarrollada (Crop Water Stress Index, CWSI) relaciona la diferencia entre la temperatura aire y la temperatura de la canopia de las plantas, y tiene un efecto regional muy importante, que requiere habitualmente una validación regional para los distintos cultivos. Objetivo. Evaluar si la metodología desarrollada por Idso (1981) y Jackson (1981), CWSI, es apropiada para estimar el estrés hídrico en los cítricos a través de imágenes infrarrojas, con posibilidades de monitorizar la distribución espacial del estado hídrico de los cítricos. Metodología (Resumen). Tratamientos. A los efectos de lograr variaciones importantes en el estado hídrico de las plantas, se utilizaron plantas adultas de naranja Spring Navel de 12 años de edad. Las plantas estaban todas en un cuadro experimental con tratamientos en secano, bajo riego y con mulch de plástico en la superficie a los efectos de que no llegue agua de la lluvia a las mismas (Figura 3). Los tratamientos se instalaron a principios del mes de enero de 2013. Medida de la temperatura de la copa del árbol. Para medir la temperatura de la superficie de la copa de los árboles cítricos, se realizaron varias tomas con una cámara térmica, que registra imágenes en el infrarrojo térmico (cámara FLIR A615) montada en una camioneta (Figura 1), en condiciones de riego, secano, y mulch; y se analizaron los pixeles de cada imagen, de acuerdo al protocolo desarrollado para las mismas. Todas la imágenes fueron tomadas con la máxima diferencia térmica diaria, entre la temperatura del aire y de la copa de los árboles, entre las 13.00 y 15.00 horas. Las imágenes fueron tomadas del lado de mayor exposición al sol, en este caso la exposición noroeste. Para calcular la temperatura de la copa cada imagen, se utilizó el procedimiento previamente validado (Figura 2) sub muestreando cada imagen y tomando el promedio y desvío estándar de los pixeles de cada sub muestra.
Programa Nacional de Investigación en Producción y Sustentabilidad Ambiental Programa Nacional de Investigación en Citricultura
INIA Salto Grande 27
Medida del potencial hídrico de la hoja (Potencial xilemático). Se seleccionaron hojas uniformes y bien desarrolladas de la parte media del árbol. Cada hoja fue encerrada dentro de bolsa negra de polietileno pequeña y recubierta por aluminio a los efectos de evitar la transpiración y de esta forma equilibrar el potencial hídrico de la hoja con el potencial hídrico del tallo. Las hojas se dejaron recubiertas por al menos 3 horas antes de hacer las medidas. Para determinar el potencial de cada hoja se utilizó una cámara de presión tipo Schoelander, las medidas fueron tomas a menos de 5 minutos de retirada de los árboles. Se midieron tres hojas por árbol en todos los arboles del tratamiento. Índice de Estrés Hídrico del Cultivo (CWSI). Se utilizó la ecuación de Jones et al (1999) ajustada para los cítricos durante el verano 2010 y 2011.
Figura 1. Posicionamiento de la cámara para ser usada automáticamente desde una camioneta.
Figura 2. Imagen total proveniente de la cámara IR, las áreas en color turquesa corresponden a las diferentes áreas de muestreo donde se evaluó la temperatura de la hoja en los distintos árboles.

Programa Nacional de Investigación en Producción y Sustentabilidad Ambiental Programa Nacional de Investigación en Citricultura
INIA Salto Grande 28
Figura 3. Plantas con cubierta de mulch de polietileno en el suelo, impermeable.
Resultados y Discusión. La ubicación de la cámara nuevamente resultó muy favorable para este tipo de estudios, y validó la facilidad de colocar estos instrumentos en vehículos; que luego en forma automatizada y en conexión con un DGPS, pueden ser utilizadas por una sola persona, y recoger datos de cuadros de cítricos enteros en plantaciones, y luego procesar sus valores en forma de distribución espacial (Figura 1). Los tratamientos fueron efectivos para lograr diferencias significativas en la temperatura de la copa, aunque no en todos los días evaluados. La temperatura de la copa aumentó en los tratamientos que tenían menos agua en el suelo: secano y mulch (Cuadro 1), por otro lado, la diferencia entre la temperatura del aire y la temperatura de la copa también está directamente relacionada con el contenido de agua del suelo, secano y mulch, los tratamientos con menor cantidad de agua en el suelo fueron los que tuvieron mayor diferencia en la temperatura de la copa (Cuadro 2) respecto al aire.
Cuadro 1. Temperatura de la copa de los árboles obtenida del análisis de las imágenes de las imágenes IR (ºC).
Tratamiento 28‐Ene 30‐Ene 04‐Feb 12‐Feb 06‐MarRiego 35,0 ns 37,1 b 28,5 b 37,0 b 27,8 nsMulch 36,4 39,2 a 29,5 b 39,3 a 28,1Secano 35,9 38,6 ab 31,4 a 40,3 a 28,6
Temp Aire (ºC) 34,1 36,5 28,4 36,4 27,8
Temperatura de la copa del árbol (ºC)
Medias en las columnas seguidas de letras diferentes son significativamente distintas TRM Duncan (p<0.05)
Programa Nacional de Investigación en Producción y Sustentabilidad Ambiental Programa Nacional de Investigación en Citricultura
INIA Salto Grande 29
Cuadro 2. Diferencia entre la temperatura del aire y la temperatura de la copa (ºC).
Tratamiento 28‐Ene 30‐Ene 04‐Feb 12‐Feb 06‐MarRiego ‐0,9 a ‐0,6 a ‐0,1 a ‐0,6 a 0,0 nsMulch ‐2,3 b ‐2,7 b ‐1,1 a ‐2,9 b ‐0,3Secano ‐1,8 ab ‐2,1 b ‐3,0 b ‐3,9 b ‐0,8
Temperatura del Aire ‐ Temperatura la Copa (ºC)
Medias en las columnas seguidas de letras diferentes son significativamente distintas TRM Duncan (p<0.05)
El potencial hídrico de la hoja, evaluado a través del método de la cámara de presión de Schoelander, es un método muy preciso de evaluar el status hídrico de la planta, pero tiene el gran inconveniente de que lleva mucho tiempo, y en consecuencia es poco aplicable en grandes extensiones en el campo. Por esta razón, otros parámetros fisiológicos de las plantas, asociados al cambio hídrico en la planta se están desarrollando desde hace tiempo, uno de estos parámetros es la diferencia entre la temperatura del aire y la temperatura de la canopia de las plantas. Los tratamientos que tenían menor cantidad de agua en el suelo presentaron menor potencial hídrico en la hoja (Cuadro 3), evidenciando la sensibilidad de este parámetro fisiológico; que se puede relacionar significativamente con la diferencia de temperaturas. A menor potencial hídrico de la hoja (hoja más deshidratada) la diferencia entre la temperatura del aire y la temperatura de la copa aumenta significativamente (Figura 4).
Cuadro 3. Potencial hídrico de la hoja (MPa)
28‐Ene 04‐Feb 12‐Feb 06‐Mar
Riego ‐2,0 a ‐2,0 a ‐2,3 a ‐1,8 aMulch ‐2,5 c ‐2,9 c ‐3,0 c ‐2,0 bSecano ‐2,1 b ‐2,6 b ‐2,9 b ‐1,9 a
Potencial Hidrico en Hoja (MPa)
Medias en las columnas seguidas de letras diferentes son significativamente distintas TRM Duncan (p<0.05)

Programa Nacional de Investigación en Producción y Sustentabilidad Ambiental Programa Nacional de Investigación en Citricultura
INIA Salto Grande 30
y = 2.2789x + 3.7275R² = 0.617
‐4,5
‐4,0
‐3,5
‐3,0
‐2,5
‐2,0
‐1,5
‐1,0
‐0,5
0,0‐3,5‐3,0‐2,5‐2,0‐1,5‐1,0‐0,50,0
Temp Aire ‐Tem
p Co
pa (ºC)
Potencial Xilematico de la Hoja (Mpa)
Todos Mulch
Riego Secano
Riego_N
Figura 4. Relación entre el potencial hídrico (xilemático) en la hoja y la diferencia entre la temperatura del aire y temperatura de la copa de los árboles.
El índice de estrés hídrico del cultivo (CWSI) se mostró bastante sensible a las condiciones de riego del cultivo, permitiendo distinguir con bastante facilidad las condiciones en la planta (Cuadro 4), a pesar de ello no fue consistente entre todos los días evaluados. La mejor hora para realizar estas medidas es luego de las 12:00 am; las variaciones de viento durante las medidas podrían afectar las evaluaciones relativas entre ellas. El déficit de presión de vapor (DPV) es la fuerza motriz de la evapotranspiración del cultivo (Idso 1982, Jackson et al. 1981). El índice CWSI toma en cuenta las condiciones ambientales en las cuales no hay limitaciones de agua para el cultivo, en este caso, a medida de aumenta el DPV (demanda atmosférica), la diferencia de temperatura entre el aire y la copa no debería de cambiar; por otro lado, a medida que falta agua en el suelo y la planta entra en condiciones de déficit hídrico, y la diferencia entre la temperatura del aire y de la copa aumenta a medida que aumenta el DPV. Cuadro 4. Índice de Estrés Hídrico del Cultivo (Idso, 1981) en las plantas. Déficit de presión de Vapor (DPV, kPa)
Tratamiento 28‐Ene 30‐Ene 04‐Feb 12‐Feb 06‐MarRiego 0,28 b 0,32 b 0,16 b 0,31 b 0,15 nsMulch 0,38 a 0,50 a 0,21 b 0,51 a 0,16Secano 0,35 a 0,44 a 0,31 a 0,59 a 0,19
VPD (kPa) 2,21 2,38 1,53 2,32 1,39Temp Aire (ºC) 34,1 36,5 28,4 36,4 27,8
Indice de Estrés Hidrico del Cultivo (CWSI)
Medias en las columnas seguidas de letras diferentes son significativamente distintas TRM Duncan (p<0.05)

Programa Nacional de Investigación en Producción y Sustentabilidad Ambiental Programa Nacional de Investigación en Citricultura
INIA Salto Grande 31
El índice de estrés hídrico de los cultivos fue originalmente desarrollado para regiones áridas y semiáridas, en consecuencia muchas de las aplicaciones prácticas están desarrolladas para cultivos en este tipo de ambientes. En nuestras condiciones templadas, esta tecnología aún no está del todo pronta y requiere de mayor validación entre otros cultivares cítricos e incluso en años de mayor demanda atmosférica. Sin embargo los datos obtenidos sobre la sensibilidad del índice a las condiciones diarias de demanda atmosférica y disponibilidad de agua, permiten apreciar su potencial utilidad, especialmente si tenemos en cuenta la posibilidad de usar este índice con referencia al posicionamiento espacial de las plantas. Bibliografía Idso SB, Jackson RD, Pinter Jr PJ, Reginato RJ, Hatfield JL. 1981. Normalizing the
stress-degree-day parameter for environmental variability. Agricultural Meteorology 24, 45–55.
Idso SB. 1982. Non-water-stressed baselines: a key to measuring and interpreting plant water stress. Agricultural Meteorology 27, 59–70.
Jackson RD, Idso SB, Reginato RJ, Pinter Jr PJ. 1981. Canopy temperature as a drought stress indicator. Water Resources Research 17, 1133–1138.
Jones HG. 1999. Use of infrared thermometry for estimation of stomatal conductance as a possible aid to irrigation scheduling. Agricultural and Forest Meteorology 95, 139–149.

Programa Nacional de Investigación en Producción y Sustentabilidad Ambiental Programa Nacional de Investigación en Citricultura
INIA Salto Grande 32
La distribución espacial de los árboles cítricos afecta el muestreo foliar y la interpretación del análisis foliar.
Alvaro Otero 1; Carmen Goñi 2
1 Programa Nacional de Producción y Sustentabilidad Ambiental. 2 Programa Nacional de Investigación en Producción Citrícola. Instituto Nacional de Investigación Agropecuaria. INIA Salto Grande. Uruguay. [email protected]; [email protected] El análisis foliar ha demostrado, a lo largo de los años, ser una herramienta de gran valor en el diagnóstico del estado nutricional de las plantaciones cítricas. Es una técnica ampliamente adoptada por los productores, y que podría ser adaptada a las innovadoras y dinámicas tecnologías de agricultura de precisión y sensoramiento remoto en continuo desarrollo en la citricultura. El muestreo foliar de un lote de producción está fundado en una distribución espacial aleatoria del estado nutricional de los árboles y por tanto, que no haya una fuerte dependencia espacial entre los nutrientes en la planta y la posición de los árboles. En este sentido, el desarrollo de metodologías de agricultura de precisión, permite interpretar y analizar con mayor precisión las variaciones espaciales de muchas variables de producción, tanto estructurales como no estructurales, entre éstas últimas las nutricionales. Se realizó el siguiente experimento a los efectos de determinar la variabilidad espacial del nivel foliar de nitrógeno y potasio en plantas cítricas y ajustar el correcto muestreo foliar. Metodología (Resumen). Descripción del sitio. Se seleccionaron cuatro cuadros de producción comercial de Clementina Fina (Citrus clementina hort. Ex Tanaka) y Clementina de Nules (Citrus clementina hort. Ex Tanaka) injertadas sobre trifolia (Poncirus trifoliata (L) Raf) de 16 años de edad, en la ribera del río Arapey en la zona de Constitución, predio de la empresa Coraler S.A. con un marco de plantación de 5 x 3,5 m en secano. Cada lote tenía una superficie mínima de 2,8 has. Los dos suelos donde se encuentran los cuadros están clasificados como un Inceptisol Ócrico Arenoso franco y un Inceptisol Melánico Limo Arcilloso (Acosta et al. 1985). Los cuadros fueron seleccionados por la excelente sanidad de sus plantas y por la uniformidad en la conducción y poda de las mismas. Muestreo de la planta y el suelo. En cada lote se seleccionaron al menos 56 plantas distribuidas en forma de grilla (30 x 30 m aproximadamente) a los efectos de facilitar la interpolación estadística y la delimitación de zonas homogéneas. Todas las plantas fueron georreferenciadas. En cada una de las plantas se tomo una muestra de 20 hojas fructíferas de los brotes de la primavera en la zona ecuatorial del árbol. El muestreo se realizó en el mes de abril. En cada muestra se determinó el contenido de nitrógeno (%), y potasio (%) en base a peso seco de la hoja. Los análisis fueron realizados en el laboratorio de suelos de INIA LE. La fertilización de cobertura previa a inicio del ciclo reproductivo se realizó en el mes de noviembre con 500 g por planta de urea y 400 g de cloruro de potasio.

Programa Nacional de Investigación en Producción y Sustentabilidad Ambiental Programa Nacional de Investigación en Citricultura
INIA Salto Grande 33
Mapa topográfico. La elevación del terreno fue determinada mediante el uso de un GPS (Leica GS20) con corrección diferencial usando otro GPS (Leica GS20) como antena. Se recorrió toda el área de trabajo, tomado las medidas en el centro de las filas de plantación. Los mapas topográficos fueron generados mediante modelos digitales de elevación (DEM) utilizando el software ArcGis (9.2) como sistema de información geográfica (GIS), posteriormente fue calculada la pendiente del terreno y se confeccionó el mapa altimétrico a un nivel de detalle de 1 metro de intervalo vertical. Análisis estadístico y espacial. Las capas de información de las características del suelo, del posicionamiento de los árboles y de los datos de los análisis foliares, fueron cargadas en el ArcGis en un sistema de coordenada común (ROUUSAMS). Para cada variable (contenido foliar de nitrógeno y potasio) se realizó la evaluación de su dependencia espacial, como una forma de determinar el grado de agregación de la variable, a tal efecto se utilizó el software GS+. Aquellas variables de planta y del suelo con mayor dependencia espacial fueron interpoladas para los cuadros de producción en base a una cuadrícula de 5 x 5 m. A los efectos de determinar el número de plantas que se deben muestrear se utilizó la ecuación 1 de muestreo básico de una población normal, para un error absoluto (D) de 0.10 y dentro de un intervalo de confianza (α) del 95% a dos colas. Ecuación 1 [Zα/2/D]2 * [S2/Media2] Siendo: S, desvío estándar de las muestras.
Media de las muestras. Zα/2 Valor estandarizado de la distribución normal. D error absoluto aceptado.
Resultados y Discusión El estudio de las frecuencias de los niveles foliares (nitrógeno y potasio) en cada uno de los muestreos es mostrada en las figuras 1 y 2. La dispersión de los valores muestréales es grande y depende bastante de la variedad y del nutriente en estudio.
0
5
10
15
20
25
1,4 1,6 1,8 2,0 2,2 2,4 2,6
Frecue
ncia
N% foliar
NitrogenoClementina Fina
Clementina de Nules
Figura 1. Frecuencia de distribución del contenido foliar de nitrógeno.
0
2
4
6
8
10
12
14
16
0,25 0,35 0,45 0,55 0,65 0,75 0,85 0,95 1,05
Frecuencia
K% Foliar
PotasioClementina Fina
Clementina de Nules
Figura 2. Frecuencia de distribución del contenido foliar de potasio.

Programa Nacional de Investigación en Producción y Sustentabilidad Ambiental Programa Nacional de Investigación en Citricultura
INIA Salto Grande 34
La dependencia espacial de una variable puede evidenciarse a través del ajuste de modelos entre la varianza de los puntos a distintas distancias entre sí (Variogramas). Esta relación es de gran utilidad práctica, ya que en situaciones de variables espacialmente agregadas, los puntos más cercanos son los más parecidos. La distribución espacial del nitrógeno foliar de las plantas evidenció una dependencia espacial significativa en Clementina de Nules y en Fina (Figuras 4 y 5), mientras que para potasio se encontró una mayor dependencia espacial (grado de agregación) sólo en el caso de Nules (Figuras 6). Podemos ver la baja dependencia espacial del potasio en Clementina Fina a través del bajo coeficiente R2, como una medida de la precisión del ajuste entre el modelo entre la varianza de los datos y la distancia entre ellos (semivariograma) (Figura 7).
Figura 3. Distribución de las plantas muestreadas y valores del contenido de nitrógeno (puntos verdes, derecha) y potasio (puntos amarillos, izquierda) foliar (%).
Figura 4. Semivariograma del contendio foliar de nitrógeno en Clementina Fina. Co=nugget o pepita; Co+C_ sill o silla; Ao=rango efectivo. Elaborado con GS+.
0.0000
0.0041
0.0082
0.0122
0.0163
0.00000 42.51892 85.03785 127.55677
Semivariance
Separation Distance (h)
N: Isotropic Variogram
Gaussian model (Co = 0.00007000; Co + C = 0.01514000; Ao = 20.50000; r2 = 0.753)

Programa Nacional de Investigación en Producción y Sustentabilidad Ambiental Programa Nacional de Investigación en Citricultura
INIA Salto Grande 35
Figura 5. Semivariograma del contendio foliar de nitrógeno en Clementina de Nules. Co=nugget o pepita; Co+C_ sill o silla; Ao=rango efectivo. Elaborado con GS+.
Figura 6. Semivariograma del contendio foliar de potasio en Clementina de Nules. Co=nugget o pepita; Co+C_ sill o silla; Ao=rango efectivo. Elaborado con GS+.
0.0000
0.0063
0.0125
0.0188
0.0250
0.00000 67.33333 134.66667 202.00000
Semivariance
Separation Distance (h)
K: Isotropic Variogram
Exponential model (Co = 0.00196000; Co + C = 0.02112000; Ao = 3.99960; r2 = 0.766)
0.0000
0.0199
0.0398
0.0596
0.0795
0.00000 67.33333 134.66667 202.00000
Semivariance
Separation Distance (h)
N: Isotropic Variogram
Exponential model (Co = 0.01450000; Co + C = 0.06640000; Ao = 28.60000; r2 = 0.761)

Programa Nacional de Investigación en Producción y Sustentabilidad Ambiental Programa Nacional de Investigación en Citricultura
INIA Salto Grande 36
Figura 7. Semivariograma del contendio foliar de potasio en Clementina Fina. Co=nugget o pepita; Co+C_ sill o silla; Ao=rango efectivo. Elaborado con GS+.
La dispersión estadística de los análisis del contenido foliar cambia entre años consecutivos, hasta un 50% de variación entre años (Cuadro 1). Esto es importante teniendo en cuenta que este desvío de la población que se quiere muestrear, es el criterio más importante cuando diseña un sistema de de muestreo y se debe de elegir el número de árboles a muestrear en un cuadro de producción. En condiciones de producción, el resultado analítico que proveniente de un cuadro de producción, está compuesto por un solo dato por cada nutriente: “el resultado del análisis”; que representa la media, el valor promedio del contenido del nutriente en todas las hojas muestreadas. Muchas veces este promedio puede no reflejar el estado nutricional real de todo el lote de producción, tal como lo evidencia este trabajo, sino se hace un correcto muestreo de los árboles, especialmente si procuramos adaptar la fertilización a las distintas áreas del cuadro.
Cuadro 1. Promedios y desvío de los nutrientes foliares y el número de plantas estimado a muestrear.
Año 1 Año 2Clementina Fina Promedio Desv Est Promedio Desv EstFosforo (%) 1,19 0,11 19Nitrogeno (%) 2,08 0,12 2,11 0,14 24 33Potasio (%) 0,68 0,12 0,97 0,18 23 53
Clementina NulesFosforo (%) 1,31 0,21 67Nitrogeno (%) 1,89 0,25 2,31 0,16 100 43Potasio (%) 0,54 0,15 1,13 0,16 35 39
Error Aceptable 0,10Intervalo Confianza 0,05
Año 2Año 1Analisis FoliarAnalisis Foliar Nº plantas a muestrear
A los efectos de reducir este desvío muestreal, convendría la identificación y delimitación más precisa de zonas homogéneas, ya sea por la floración, por la carga de fruta o cualquier otro factor que afecte el nivel nutricional de los árboles.
0.0000
0.0038
0.0076
0.0113
0.0151
0.00000 42.51892 85.03785 127.55677
Semivariance
Separation Distance (h)
K: Isotropic Variogram
Exponential model (Co = 0.00001000; Co + C = 0.01262000; Ao = 10.50000; r2 = 0.103)

Programa Nacional de Investigación en Producción y Sustentabilidad Ambiental Programa Nacional de Investigación en Citricultura
INIA Salto Grande 37
Conclusiones En nuestros cuadros de producción citrícola, puede darse el caso de que la variación de los nutrientes en la planta no tenga la dispersión de valores inicialmente prevista para un determinado muestreo. Esto puede ser ocasionado por distintos factores que afectan el estado nutricional de cada árbol. El estado nutricional de una planta no solo es afectado por el tipo de suelo donde se encuentra y por la fertilización que tuvo, sino también por las condiciones de producción, como ser variaciones en el rendimiento, riego, manejo del suelo, sanidad, etc. Esta variación de la dispersión (mayor o menor) nos va a afectar la interpretación del análisis foliar a los efectos de hacer una buena recomendación. Si en nuestro cuadro de producción tenemos una variabilidad entre los árboles mayor a la esperada, corremos el riesgo de que al considerar solo la media del valor del análisis foliar, tengamos arboles en zonas de deficiencia o exceso del nutrientes que no los estemos considerando. En este sentido, es altamente aconsejable evaluar con más precisión, las zonas “homogéneas” de muestreo, ya que podemos asumir semejanzas entre las plantas que en la realidad no lo son y de esta forma cometemos imprecisiones importantes al realizar correctamente la dosificación de los fertilizantes.


















