
REPÚBLICA BOLIVARIANA DE VENEZUELA - … · Porcentajes de resistencia y sensibilidad de las cepas...
84
1 REPÚBLICA BOLIVARIANA DE VENEZUELA LA UNIVERSIDAD DEL ZULIA FACULTAD EXPERIMENTAL DE CIENCIAS DIVISIÓN DE ESTUDIOS PARA GRADUADOS MAESTRÍA EN MICROBIOLOGÍA Relación entre la Producción de Betalactamasas de Espectro Expandido (BLEE) y la Resistencia a Fluoroquinolonas en Cepas de Enterobacterias Provenientes de Centros Hospitalarios de Venezuela Trabajo de Grado presentado ante la División de Estudios para Graduados para optar al Grado de Magíster Scientiarum en Microbiología Autor: Lic. Daniel Marcano Zamora C.I. 14.775.231 Tutor: Dr. Carlos D. Ramírez C.I. 82.233.808 Co-tutor: Msc. Jorge Guiñez-Ortega C.I. 81.255.143 Maracaibo, Marzo 2011
Transcript of REPÚBLICA BOLIVARIANA DE VENEZUELA - … · Porcentajes de resistencia y sensibilidad de las cepas...

1
REPÚBLICA BOLIVARIANA DE VENEZUELA LA UNIVERSIDAD DEL ZULIA
FACULTAD EXPERIMENTAL DE CIENCIAS DIVISIÓN DE ESTUDIOS PARA GRADUADOS
MAESTRÍA EN MICROBIOLOGÍA
Relación entre la Producción de Betalactamasas de Espectro Expandido (BLEE) y la Resistencia a Fluoroquinolonas en Cepas de Enterobacterias
Provenientes de Centros Hospitalarios de Venezuela
Trabajo de Grado presentado ante la División de Estudios para Graduados para optar al Grado de Magíster Scientiarum en Microbiología
Autor: Lic. Daniel Marcano Zamora C.I. 14.775.231
Tutor: Dr. Carlos D. Ramírez C.I. 82.233.808
Co-tutor: Msc. Jorge Guiñez-Ortega C.I. 81.255.143
Maracaibo, Marzo 2011

4
AGRADECIMIENTOS
A Dios, ya que sin El no estaría aquí A mi novia Andreína, por no sólo motivarme a culminar este trabajo y darme apoyo emocional, sino también por ayudarme en la revisión y corrección del manuscrito. A las profesoras Lorena Atencio, Irene Zabala y Marynés Montiel, por todo su estímulo y paciencia para ayudarnos a culminar con éxito esta estapa académica de nuestras vidas. A todo el personal del Instituto Nacional de Higiene “Rafael Rangel”, en particular al adscrito a la Gerencia de Docencia e Investigación y a la Gerencia Sectorial de Diagnóstico y Epidemiología por su indiscutible apoyo en distintas etapas de esta maestría. A la Licenciada Omaira Da Mata por su ayuda en la optimización de ciertas técnicas moleculares A los Técnicos Eidha Moreno y Samari Torres y los Bachilleres Jeisybeth Luque, Patricia Bohorquez, José Angel Rodríguez y Richard Aparicio por su ayuda en distintos puntos clave de la fase experimental. A todos mis compañeros del Laboratorio de Aislamiento e Identificación Bacteriana por su apoyo y estímulo durante la fase experimental. A la Doctora Clara Martínez y todo su equipo del Laboratorio de Biología Molecular de Protozoarios del Instituto de Medicina Tropical de la UCV por su invaluable apoyo para la secuenciación de los productos de amplificación de este trabajo. Al personal del Servicio de Antimicrobianos del INEI ANLIS “Dr. Carlos Malbrán” de Argentina por su asesoría contínua durante la estandarización de las técnicas moleculares. A mis tutores, que siempre estuvieron allí para orientarme en todas las fases del trabajo.
A todo aquel que no haya mencionado en este espacio tan corto y que me haya ayudado,
Muchas gracias
Daniel Marcano Zamora

5
TABLA DE CONTENIDOS
INDICE DE TABLAS ........................................................................................................ 6
INDICE DE FIGURAS ...................................................................................................... 7
RESUMEN ....................................................................................................................... 8
ABSTRACT ...................................................................................................................... 9
INTRODUCCION ........................................................................................................... 10
JUSTIFICACIÓN ............................................................................................................ 14
OBJETIVO GENERAL ................................................................................................... 15
OBJETIVOS ESPECÍFICOS .......................................................................................... 15
MARCO TEÓRICO ......................................................................................................... 16
Terapia antimicrobiana y resistencia bacteriana ......................................................... 16
Antibióticos B- lactámicos. ........................................................................................... 16
Mecanismo de acción de las Quinolonas: ................................................................... 18
Resistencia enzimática a los antibióticos betalactámicos ........................................... 20
Resistencia a las Quinolonas ...................................................................................... 25
Relevancia de la transmisión de la resistencia bacteriana .......................................... 30
Caracterización de mecanismos de resistencia en el laboratorio de microbiología ..... 35
MARCO METODOLÓGICO ........................................................................................... 38
Tipo de estudio ............................................................................................................ 38
Muestra ....................................................................................................................... 38
Metodología de laboratorio .......................................................................................... 39
RESULTADOS ............................................................................................................... 48
Estudio fenotípico y de susceptibilidad. ....................................................................... 48
Detección fenotípica de la producción de enzimas mediante el método de Hodge. .... 54
Determinación de Punto Isoeléctrico mediante ensayos de Isoelectroenfoque........... 55
Ensayos de Conjugación ............................................................................................. 57
Estudio molecular ........................................................................................................ 58
Estudio de integrones .................................................................................................. 60
Secuenciación ............................................................................................................. 63
DISCUSIÓN ................................................................................................................... 66
CONCLUSIONES ........................................................................................................... 74
RECOMENDACIONES .................................................................................................. 77
REFERENCIAS BIBLIOGRÁFICAS ............................................................................... 78

6
INDICE DE TABLAS
Tabla 1. Clasificación de las Quinolonas ........................................................................ 18
Tabla 2. Esquema de Clasificación de Betalactamasas Bacterianas ............................. 21
Tabla 3. Iniciadores empleados en el estudio ................................................................ 42
Tabla 4. Perfil de Resistencia a Betalactámicos de cepas de Enterobacterias BLEE positivas seleccionadas para el estudio. ........................................................................ 52
Tabla 5. Perfil de Resistencia a Quinolonas de cepas de Enterobacterias BLEE positivas seleccionadas para el estudio. ........................................................................ 52
Tabla 6. Resultados obtenidos al aplicar el método de Hodge comparado con el método de difusión del disco para detección de BLEE ............................................................... 55
Tabla 7. Valores de pI obtenidos para las cepas en estudio. ......................................... 56
Tabla 8. Ensayos de Conjugación en aislados de Enterobacterias BLEE positivas que presentaron integrones de clase 1 ................................................................................. 57
Tabla 9. Resultados de amplificación de genes de betalactamasas. ............................. 59
Tabla 10. Resultados de amplificación de genes codificantes para resistencia plasmídica a quinolonas. ................................................................................................ 60
Tabla 11. Resultados de amplificación por PCR de genes de integrasa de clase 1....... 61
Tabla 12. Resultados de amplificación de la región variable de integrones clase 1 ....... 62
Tabla 13. Tamaño de amplicones obtenidos durante el mapeo de integrones clase 1 con distintas combinaciones de iniciadores. .................................................................. 62
Tabla 14. Detección de mutaciones en gyrA .................................................................. 64
Tabla 15. Variantes de TEM y SHV encontradas en el estudio. ..................................... 64

7
INDICE DE FIGURAS
Figura 1. Anillo Betalactámico ................................................................................... 17
Figura 2. Núcleo central de las Quinolonas ............................................................... 19
Figura 3. Mecanismo de acción de las quinolonas .................................................... 20
Figura 4. Esquema del proceso de Conjugación bacteriana ..................................... 33
Figura 5. Distribución de aislados de Enterobacterias en estudio ............................. 48
Figura 6. Porcentajes de resistencia y sensibilidad de las cepas en estudio productoras de Betalactamasas de espectro expandido ............................................................... 49
Figura 7. Correlación entre los resultados de susceptibilidad para Cefotaxima y Ciprofloxacina de las cepas en estudio ..................................................................... 50
Figura 8. Correlación entre los resultados de susceptibilidad para Cefotaxima y Acido Nalidíxico de las cepas en estudio ............................................................................ 50
Figura 9. Distribución de halos obtenidos para betalactámicos y quinolonas en cepas BLEE positivas y evaluación de la Concentración Mínima Inhibitoria a Ciprofloxacina por la técnica de E-test. ................................................................................................... 53
Figura 10. Algunos perfiles fenotípicos de BLEE clásicas encontrados. ................... 53
Figura 11. Resultados de prueba de Concentración Inhibitoria Mínima a Ciprofloxacina por E-test. .................................................................................................................. 54
Figura 12. Prueba de Hodge de las cepas en estudio. .............................................. 55
Figura 13. Gel de isoelectroenfoque. ........................................................................ 56
Figura 14. Ensayo de conjugación. ........................................................................... 58
Figura 15. Amplificación de genes TEM, CTX-M, SHV y gyrA de las cepas en estudio. .................................................................................................................................. 58
Figura 16. Amplificación de Integrasa clase 1 de las cepas en estudio. ................... 61
Figura 17. Estructura teórica propuesta de la región variable de integrones encontrados en las cepas en estudio. ............................................................................................ 63
Figura 18. Electroferograma de TEM-150 ................................................................. 65

8
Marcano Z, Daniel. Relación entre la producción de Betalactamasas de Espectro Expandido y la resistencia a Fluoroquinolonas en cepas de Enterobacterias provenientes de centros hospitalarios de Venezuela. Proyecto de Trabajo de Grado. La Universidad del Zulia, Facultad Experimental de Ciencias, División de Estudios para Graduados, Maestría en Microbiología, Maracaibo, Venezuela 2011. 86 p
RESUMEN
Existen reportes de cepas en las cuales parece haber asociación entre mecanismos de resistencia a antibióticos, por lo que el objetivo de este estudio fue determinar la relación entre la producción de Betalactamasas de Espectro Expandido (BLEE) y la resistencia a fluoroquinolonas en Enterobacterias provenientes de centros hospitalarios de Venezuela. La identificación de las cepas se realizó mediante pruebas bioquímicas convencionales, y la resistencia se determinó mediante antibiograma y concentración mínima inhibitoria. La caracterización de resistencia se realizó mediante Hodge, isoelectroenfoque, conjugación y PCR. Escherichia coli fue la cepa BLEE aislada con mayor frecuencia. Se encontró alto nivel de resistencia a betalactámicos y quinolonas. Se observó que las cepas BLEE presentaron un resultado Hodge positivo con Cefotaxima o Ceftazidima, y negativo empleando Ciprofloxacina. Se demostró por conjugación la presencia de genes BLEE en elementos genéticos transferibles en 4 de 8 cepas ensayadas. Se detectó mediante PCR la presencia de los genes SHV, TEM, PER, CTX-M grupo 2, QnrB y AAC(6’)Ib-cr. Se demostró la presencia de integrones clase 1, y que los genes BLEE y de resistencia plasmídica a quinolonas se encontraban en estos elementos transferibles. Se propone la coexistencia de genes BLEE y de resistencia plasmídica a quinolonas en el mismo integrón, lo cual explicaría la asociación entre estos mecanismos. La secuenciación del gen gyrA permitió encontrar dos mutaciones codificantes para el aminoácido 83, lo cual es consistente con la CIM para Ciprofloxacina de dichas cepas. Se encontró relación estadísticamente significativa (p<0,05) entre la producción de BLEE y la resistencia a quinolonas; producción de BLEE y presencia de mecanismos cromosomales de resistencia a quinolonas; y producción de BLEE y presencia de mecanismos plasmídicos de resistencia a quinolonas. La asociación encontrada entre los mecanismos de resistencia a betalactámicos y quinolonas refleja la necesidad de explorar alternativas terapéuticas para el tratamiento de este tipo de cepas.
Palabras Clave: Betalactamasas de Espectro Expandido, Fluoroquinolonas, Enterobacterias
Correo electrónico: [email protected]

9
Marcano Z, Daniel. Relación entre la producción de Betalactamasas de Espectro Expandido y la resistencia a Fluoroquinolonas en cepas de Enterobacterias provenientes de centros hospitalarios de Venezuela. Proyecto de Trabajo de Grado. La Universidad del Zulia, Facultad Experimental de Ciencias, División de Estudios para Graduados, Maestría en Microbiología, Maracaibo, Venezuela 2011. 86 p
ABSTRACT
There are reports of strains which apparent association between mechanisms of resistance to antibiotics, sothe objective of this study is to determine the relationship between the production of Extended Spectrum Betalactamases (ESBL) and resistance to fluoroquinolones in enterobacteria from hospitals of Venezuela. Identification was performed using conventional biochemical tests and resistance was determined by minimum inhibitory concentration and antibiogram. Resistance characterization was performed using Hodge, isoelectric focusing, conjugation and PCR. Escherichia coli were the ESBL strain isolated more frequently. We found high levels of beta-lactam and quinolone resistance. ESBL strains presented a positive Hodge result with Cefotaxime or Ceftazidime, and a negative result using Ciprofloxacin. Conjugation demonstrated the presence of ESBL genes in transferable genetic elements in 4 of 8 strains tested. SHV, TEM, PER, Group 2 CTX-M, QnrB and AAC(6') Ib-cr genes were detected by PCR. The presence of class 1 integrons was detected, and the presence of ESBL and quinolone plasmidic resistance genes in these transferable elements. The coexistence of ESBL and quinolone plasmidic resistance genes in a single integron is proposed, which would explain the association between these mechanisms. The sequencing of gyrA enabled the finding of two mutations coding for the amino acid in position 83, which is consistent with the Ciprofloxacin MIC for these strains. We found a statistically significant relationship (p <0.05) between ESLB production and quinolone resistance, ESLB production and chromosomal quinolone resistance mechanisms, and ESLB production and plasmidic quinolone resistance mechanisms. The association found between Beta-lactam and quinolone resistance mechanisms reflects the need to explore alternative therapies for the treatment of this type of strains. Keywords: Extended Spectrum Betalactamases, Fluoroquinolones, Enterobacteria
E-mail: [email protected]

10
INTRODUCCION
Un antimicrobiano es “cualquier agente químico que evita o destruye
microorganismos o inhibe su proliferación”. Un antibiótico puede definirse como “un
fármaco empleado para el tratamiento de las enfermedades infecciosas producidas por
bacterias” (Jehl y col., 2004). Los agentes quimioterapeuticos son compuestos
elaborados en forma total o parcial por síntesis química, ambos compuestos tienen la
capacidad de inhibir o destruir el desarrollo de otros microorganismos a bajas
concentraciones (Joklik y col., 1998).
La clasificación más empleada de los antibióticos está basada en su diferenciación
según el lugar de actuación en la bacteria o los procesos fisiológicos con los que
interactúa; a su vez, dentro de cada uno de estos grupos la clasificación por familias se
basa en la estructura química de las diferentes moléculas y a cada familia le
corresponde un mecanismo molecular específico, encontrando así los siguientes grupos
(Jehl y col., 2004):
- Antibióticos activos sobre la pared bacteriana: betalactámicos, glicopéptidos y
fosfonopéptidos.
- Antibióticos activos sobre la membrana citoplasmática: gramicidinas y
polimixinas.
- Antibióticos activos sobre la síntesis proteica: aminoglicósidos, tetraciclinas,
macrólidos, lincosamidas, estreptograminas, rifampicina, cloramfenicol, ácido
fusídico.
- Antibióticos activos sobre la replicación del ADN: fluoroquinolonas.

11
- Antibióticos inhibidores de vías metabólicas: sulfonamidas,
trimetroprim.Inhibidores de betalactamasas: ácido clavulánico, sulbactam,
tazobactam.
Entre los antibióticos inhibidores de la pared celular bacteriana están los
antibióticos β – lactámicos, los cuales incluyen las penicilinas, cefalosporinas,
carbapenemas, monobactamas y los inhibidores de betalactamasas, y actúan como
agentes bactericidas. La resistencia bacteriana a los antibióticos β – lactámicos se
puede atribuir a cuatro mecanismos diferentes:
a) Inactivación de la droga
b) Alteración del sitio blanco
c) Bloqueo del transporte de la droga al interior de la célula
d) Eflujo activo
De estos, el mecanismo más importante de resistencia a los antibióticos
betalactámicos es la inactivación producida por las β – lactamasas, las cuales abren el
anillo β – lactámico del antibiótico. Las β – lactamasas procedentes de los
microorganismos Gram negativos difieren en muchos aspectos de las enzimas de las
especies Gram positivas. En general son constitutivas y están unidas a la célula, y
pueden estar codificadas en plásmidos o en el cromosoma. Se producen en cantidades
menores que en las Gram positivas y tienen una afinidad mucho más baja por sus
sustratos. Las betalactamasas, dependiendo de su espectro de acción, pueden tener
como sustrato penicilinas, cefalosporinas, cefamicinas, monobactamas e incluso
carbapenemas, pero no son activos contra otras familias de antibióticos (Joklik y col.,
1998).

12
Las β – lactamasas de espectro expandido (BLEE) son enzimas que expanden su
espectro de acción a los β – lactámicos anti β – lactamasas como las oximino –
cefalosporinas, cefamicinas, carbapenemas y monobactámicos, confiriendo resistencia
a una amplia variedad de β–lactámicos (Iyobe, 1997). Estas enzimas fueron descritas
por primera vez en Alemania en 1983 (Knothe y col., 1983), y desde entonces su
número ha aumentado rápidamente y ahora su distribución es mundial (Lucet y
Regnier, 1998); a pesar del amplio uso internacional de los antibióticos betalactámicos,
la distribución de las BLEE no es uniforme en todos los países: algunos hospitales en
los Estados Unidos no presentan BLEE, mientras que otros presentan hasta 40% de
cepas de Klebsiella pneumoniae productoras de estas enzimas. Las BLEE se
encuentran con más frecuencia en especies de Klebsiella pneumoniae y Escherichia
coli, pero se pueden encontrar en otras enterobacterias y bacilos no fermentadores; la
presencia de estas enzimas pareciera ser mayor en Latinoamérica que en Estados
Unidos y Europa, y sus distintos genotipos (SHV, TEM, CTX-M, PER) varían de región a
región (Jacoby y Muñoz-Price, 2005).
En lo que respecta a la epidemiología de estas enzimas hay que tomar en cuenta
que los genes que codifican las BLEE se encuentran en elementos genéticos móviles
(plásmidos, transposones), por lo cual dicha resistencia puede ser transferida a otras
bacterias mediante conjugación o transposición y aumentar el número de cepas
resistentes. En un estudio realizado en 1998 con 502 cepas bacterianas aisladas de
siete hospitales venezolanos se determinó que el 30% de las cepas de K. pneumoniae
aisladas producían BLEE (Pfaller y col., 1998) y al determinar los mecanismos
moleculares y genéticos involucrados en la resistencia antimicrobiana de bacilos Gram-
negativos de origen nosocomial (entre ellos 4 cepas de K. pneumoniae) se observó la
presencia de plásmidos transferibles que codificaban betalactamasas pertenecientes al
grupo TEM. Cabe destacar que la mayoría de estas bacterias multirresistentes se aíslan
en el ambiente intrahospitalario, ya que es en dicho ambiente donde se generan
mecanismos de transmisión tanto vertical como horizontal de genes plasmídicos y
transposones que codifican para determinantes de resistencia antimicrobiana, esto trae
como consecuencia el establecimiento y diseminación de dichos determinantes de
resistencia.

13
Lo anteriormente expuesto demuestra la relevancia epidemiológica de la posible
diseminación intranosocomial de las mencionadas cepas multirresistentes. Según los
datos del Programa Venezolano de Vigilancia de la Resistencia Bacteriana a los
Antimicrobianos (en el cual participan 29 laboratorios de microbiología de distintas
áreas del país, incluida el área metropolitana) en 1998 el 61% de las cepas de E. coli
presentaron resistencia a ampicilina, el 50% a trimetoprim-sulfametoxazol, el 6% a
amikacina y el 10% a gentamicina, y se observa con preocupación el ascenso de la
resistencia a fluoroquinolonas y cefalosporinas de tercera generación; esto último
sugiere que estamos en presencia de cepas productoras de Betalactamasas de
Espectro Expandido.
En el caso de Klebsiella pneumoniae un porcentaje mayor del 90% es resistente
a ampicilina, situación que se reduce a una tercera parte (30%) con la combinación de
ampicilina/sulbactam, lo que hace pensar que la producción de betalactamasas es uno
de los mecanismos de resistencia implicados, pero no el único. Por otra parte, en el año
1999 se aprecia un aumento en la resistencia a cefalosporinas de tercera generación:
29% a ceftazidima y 39% a ceftriaxona, lo que indica la presencia de Betalactamasas
de Espectro Expandido (Comegna y col., 2000).
Es frecuente encontrar en las cepas productoras de Betalactamasas de Espectro
Expandido resistencia asociada a quinolonas como Ciprofloxacina y Norfloxacina
(Marcano, 2002; Martínez y col., 2002), aun cuando estas enzimas no confieren
resistencia a dichas familias de antibióticos; esto podría ser debido a algún tipo de
afinidad de los plásmidos codificantes para Betalactamasas de Espectro Expandido a
cepas con susceptibilidad disminuida a fluoroquinolonas o por diseminación conjunta de
ambos mecanismos.
El presente estudio tiene como objetivo conocer la relación entre la producción
de betalactamasas de espectro expandido y la resistencia a quinolonas en cepas de
enterobacterias aisladas de muestras clínicas provenientes de distintos centros
hospitalarios del área metropolitana de Caracas.

14
JUSTIFICACIÓN
El gran número de especies dentro de la familia de las enterobacterias conlleva a
una gran variabilidad de patrones de sensibilidad natural. Esta diversidad se ve además
incrementada por la posibilidad de adquirir genes de resistencia, tanto de
microorganismos de la misma especie como de otras. . La resistencia antimicrobiana
aparece como parte del progreso de la infección en muchas personas y es letal en
algunos casos, pero no es un trastorno humano, es propio de las poblaciones
bacterianas, las cuales tienden a crear mecanismos de resistencia como una
información genética, luego de que el paciente es sometido por largo tiempo a un
tratamiento antibacteriano. La adquisición de multirresistencia puede llevar a la
ineficacia de la mayoría de los antimicrobianos utilizados en la práctica clínica (Navarro
y col., 2002), y existen cepas en las cuales pareciera existir asociación en la presencia
de diversos mecanismos de resistencia a antibióticos de distintas familias, por ejemplo
en el caso de Klebsiella pneumoniae se ha descrito que aquellas cepas que producen
betalactamasas de espectro expandido (cuyo sustrato son los betalactámicos)
presentan resistencia a fluoroquinolonas con más frecuencia que aquellas cepas de
Klebsiella pneumoniae que no expresan dichas enzimas (Martínez y col., 2002).
La importancia de este estudio radica en la gran diversidad genética que se
presenta en la familia de betalactamasas de espectro expandido y de la posible
asociación de enzimas particulares a otros mecanismos de resistencia a diversas
familias de antibióticos, por lo tanto se considera necesaria la detección y
caracterización de las Betalactamasas encontradas en cepas hospitalarias del área de
Caracas y sus mecanismos de resistencia acompañante. Estos datos aportarán
información epidemiológica valiosa para los profesionales de las ciencias de la salud
acerca de los mecanismos de resistencia y diseminación de estas bacterias en el país y
colaborará en las políticas de tratamiento antimicrobiano, además de alertar sobre la
posibilidad de diseminación horizontal de esta resistencia acompañante en conjunto con
las Betalactamasas de Espectro Expandido.

15
OBJETIVO GENERAL
Determinar la relación existente entre la producción de betalactamasas de
espectro expandido (BLEE) y la resistencia a fluoroquinolonas, en cepas de
enterobacterias aisladas desde enero hasta junio del año 2006, de muestras clínicas
provenientes de distintos centros hospitalarios del área metropolitana de Venezuela.
OBJETIVOS ESPECÍFICOS
Determinar fenotípicamente los perfiles de resistencia in vitro de las cepas en
estudio.
Establecer las propiedades físico-químicas de las Betalactamasas de Espectro
Expandido aisladas.
Identificar las características genéticas de estas Betalactamasas de Espectro
Expandido.
Identificar los mecanismos de resistencia a Fluoroquinolonas en las cepas en
estudio.
Establecer si existe una relación entre la expresión de genes codificantes de
Betalactamasas de Espectro Expandido y la expresión de genes codificantes de
resistencia a Fluoroquinolonas.

16
MARCO TEÓRICO
Terapia antimicrobiana y resistencia bacteriana Uno de los principales logros de la ciencia médica ha sido la casi total erradicación
de muchas enfermedades infecciosas gracias al uso de agentes antimicrobianos
específicos. Entre los descubrimientos de importancia que dan inicio a la “edad de oro”
de la terapéutica antimicrobiana, se encuentra el descubrimiento y el posterior
desarrollo de la penicilina a partir de filtrados de cultivos del hongo Penicillium notatum.
Si bien la penicilina había sido descubierta por Fleming en 1929, fueron Florey, Chain y
sus colaboradores en la Universidad de Oxford quienes en 1940 demostraron y
publicaron un informe sobre su enorme potencia y la posibilidad de su extracción de los
sobrenadantes del cultivo (Joklik y col., 1992).
Antibióticos B- lactámicos.
Los antibióticos betalactámicos son los antibióticos más variados y de uso más
extendido a nivel mundial. Desde 1940, se han descrito numerosos compuestos
naturales y sintéticos basados en el principio activo de la penicilina: el anillo
betalactámico. Los betalactámicos constituyen uno de los antibióticos que son más
usados de forma sistémica. El éxito se debe a la baja toxicidad y la biodisponibilidad de
una gran variedad de ellos. Apartando las micobacterias, los patógenos intracelulares y
las especies deficientes de pared, existen pocos organismos que puedan resistir a la
acción de todos los betalactámicos disponibles actualmente (Lorian, 1996). Los
antibióticos betalactámicos inhiben o detienen el crecimiento bacteriano por unión
covalente a las Proteínas Fijadoras de Penicilina (PBP, Protein Binding Penicillin)
presentes en la membrana citoplasmática. Estas proteínas blanco catalizan la síntesis
de peptidoglicano que forma la pared celular de las bacterias, y alteraciones en ellas
pueden llevar a la resistencia a los antibióticos betalactámicos (Mandell, 2002). Los B-
lactámicos son uno de los grupos de antibióticos más usados en la práctica médica,
poseen una gran eficacia terapéutica y no son tóxicos para el hombre; estas dos

17
características hacen que estos antibióticos sean uno de los mejores grupos
terapéuticos bacterianos.
Los β-lactámicos son agentes bactericidas de efecto lento. La eficacia está
relacionada más con el tiempo de actuación que con las concentraciones del antibiótico
en el medio. Su mecanismo de acción consiste en el bloqueo de la actividad
transpeptidasa, carboxipeptidasa, endopeptidasa o transglucosilasa de las proteínas
fijadoras de penicilina (PBP) con la consiguiente inhibición o disminución de la síntesis
de la capa de peptidoglucano.
Figura 1. Anillo Betalactámico. Tomado de Goodman y Gilman’s. The pharmacological basis of
therapeutics. McGraw-hill
Los antibióticos que poseen en su estructura química el anillo β-lactámico (Figura
1) se comportan como agentes acilantes, que reaccionan para producir derivados
peniciloicos. Se ha propuesto que la penicilina actuaría como análogo del sustrato PBP,
acilando el sitio activo de la enzima con formación de un complejo inactivo bastante
estable. La similitud estereoquímica de estos antibióticos con la secuencia terminal del
pentapéptido de la mureína les permite interaccionar con las PBP situadas en la
superficie de la membrana plasmática bacteriana, moléculas encargadas de modelar la
configuración definitiva de la capa de peptidoglicano, además de guiar su
reorganización durante la división de las bacterias. La inhibición de las reacciones de
biosíntesis por antibióticos β- lactámicos está acompañada por cambios morfológicos
característicos. Las diferencias morfológicas observadas, se deben a la PBP que resulta
afectada. Cuando las bacterias se desarrollan en presencia de penicilina se acumulan
intermediarios de la síntesis de la pared celular, nucleótidos de uridina sin uniones
cruzadas, y las nuevas paredes no se pueden formar, muriendo así la bacteria por
efecto osmótico o digerida por enzimas auto líticas, que se activan como consecuencia

18
del bloqueo de la función de una varias PBP. Sin embargo el mecanismo exacto por el
cual los agentes sensibles son lisados aun no está muy dilucidado.
Mecanismo de acción de las Quinolonas: Las quinolonas actúan inhibiendo rápidamente la síntesis del ADN por inhibición
del ADN girasa y la topoisomerasa IV, siendo éstas el blanco de las quinolonas, lo cual
resulta en muerte bacteriana. Como regla general, la actividad en las bacterias Gram
negativas se correlacionan con la inhibición de la ADN girasa, y su actividad en Gram
positivos se corresponde con la inhibición de la topoisomerasa de ADN tipo IV. Las
Quinolonas penetran a la célula mediante un sistema de difusión pasiva, principalmente
atravesando la membrana, y a través de las porinas. Parece ser que las Quinolonas
mas hidrofóbicas penetrarían a través de la bicapa lipídica, mientras que las Quinolonas
más hidrofílicas utilizarían las porinas como puerta de entrada (Sierra, 2005). Al igual
que las Cefalosporinas, las Quinolonas pueden clasificarse en generaciones (Alós,
2009) según su momento de aparición en la historia (Tabla 1)
Tabla 1. Clasificación de las Quinolonas
GENERACIÓN COMPUESTO ESPECTRO
1ra Generación Ac. Nalidíxico
Ac. Pipemídico
Enterobacterias
2da Generación Norfloxacina
Enterobacterias,
P. aeruginosa
Moderado para
Grampositivos
Determinados patógenos
atípicos
3ra Generación Ciprofloxacina
Levofloxacina
Ofloxacina
Mejor actividad frente a
Grampositivos y patógenos
atípicos
4ta Generación Moxifloxacina Buena actividad frente a
Grampositivos y
Anaerobios

19
Las Topoisomerasas son enzimas que participan en el proceso de síntesis del
ADN, por desenrollamientos y enrollamientos del ADN cromosómico. En
Gramnegativos, la Topoisomerasa que inhiben principalmente es la ADN girasa, que
tiene 2 subunidades A y 2 subunidades B (Alós, 2009) La ADN girasa es la única
enzima capaz de introducir un superenrollamiento negativo en la cadena de ADN,
facilitando la separación de la doble hélice de ADN para mantener la estructura de la
horquilla de replicación, mientras que la Topoisomerasa IV (diana principal en
Grampositivos) es responsable del desencadenamiento de las dos moléculas “hijas” al
finalizarse la replicación del ADN, permitiendo la segregación de dos nuevos
cromosomas bacterianos en dos nuevas células “hijas” (Drlica, 1999). Los genes que
codifican para estas enzimas son GyrA y GyrB (para la subunidad A y B
respectivamente) de la ADN girasa y los genes ParC y ParE (para la subunidad A y B
respectivamente) de la Topoisomerasa IV (Sierra, 2005)
El mecanismo de acción más aceptado en la actualidad es el propuesto por
Palumbo (Palumbo y col., 1993). En el mismo se establece la formación de un complejo
entre la Topoisomerasa – ADN – Quinolona, que se encarga de bloquear la acción de la
enzima, provocando de esta forma la muerte celular.
Figura 2. Núcleo central de las Quinolonas. Tomado de Goodman y Gilman’s. The pharmacological basis of therapeutics. McGraw-hill
En este modelo los radicales en posición 3 y 4 de la molécula de Quinolona
(Figura 2) producirían enlaces con un átomo de Mg2+ y éste a su vez se uniría al ADN
mediante enlaces con los grupos fosfato de los nucleótidos. Además, se produciría un
apilamiento entre una de las bases de la región monocatenaria de la cadena de ADN y
el anillo de la molécula de la Quinolona. También se postula que los radicales situados

20
en posición 1 y 7 son los responsables de la interacción con la ADN girasa o la
Topoisomerasa IV (Palumbo y col., 1993).
Resistencia enzimática a los antibióticos betalactámicos
Las betalactamasas son enzimas que destruyen las penicilinas y cefalosporinas
mediante hidrólisis y son el principal mecanismo de resistencia a estos antimicrobianos.
La mayoría de las betalactamasas actúan por un mecanismo serina-éster, pero algunas
pocas son dependientes de zinc. La diferencia fundamental entre la vía catalítica de una
serinobetalactamasa y una Proteína Fijadora de Penicilina (Protein Binding Peniciline, el
sitio de acción de los betalactámicos) es que el éster formado por la betalactamasa se
hidroliza rápidamente, regenerándose así la enzima, mientras que los que se forman
con la PBP son estables.
Figura 3. Mecanismo de acción de las quinolonas. Tomado de Goodman y Gilman’s. The pharmacological basis of therapeutics. McGraw-hill
Se han hecho muchos intentos para clasificar las betalactamasas, basándose la
mayoría en propiedades fenotípicas como su afinidad por sustratos (cefaloridina vs
bencilpenicilina), perfil de inhibición (inhibición con oxacilina, clavulanato, aztreonam o
Ácido Etilen Diamino Tetraacético, EDTA) y si la enzima está codificada en el
cromosoma o a nivel de un plásmido; sin embargo, la tendencia actual es clasificar las
betalactamasas por secuencia y no por sus propiedades fenotípicas, tal como fue
propuesto por Ambler (Ambler, 1980). Las clasificaciones basadas en secuencia

21
reflejan relaciones fundamentales y, a diferencia de las clasificaciones fenotípicas, no
pueden ser distorsionadas por mutaciones que alteren la especificidad de sustrato o el
perfil de inhibición. Actualmente sólo existen cuatro clases moleculares de
betalactamasas: las clases A, C y D (las cuales son serinoenzimas) y la clase B
(compuesta por enzimas dependientes de zinc) (Lorian, 1996).
Tabla 2. Esquema de Clasificación de Betalactamasas Bacterianas. Tomado de Bush y Jacoby, 2010
Entre los miembros de la familia Enterobacteriaceae, la producción de
Betalactamasas de Espectro Expandido ha emergido como un importante mecanismo
de resistencia a betalactámicos, los cuales constituyen aproximadamente el 50% del
consumo de antibióticos. Estas enzimas son enzimas plasmídicas de clase molecular A
que corresponden al subgrupo 2be de Bush y que son capaces de hidrolizar un amplio
rango de betalactámicos, incluidas las cefalosporinas de más reciente aparición, pero
no son activas frente a las cefamicinas y carbapenemas.

22
Al interpretar el fenotipo de las cepas productoras de BLEE en lo que se refiere a
los Betalactámicos, debe tenerse en cuenta que la resistencia a los antibióticos también
puede deberse a la actividad combinada de una BLEE específica con otras
betalactamasas (por ejemplo AmpC cromosomal). Entre las enterobacterias, las BLEE
se han encontrado principalmente en Klebsiella spp. y E. coli, pero también se han
encontrado en otros géneros como Citrobacter, Enterobacter, Morganella, Proteus,
Providencia, Salmonella, y Serratia. Las infecciones causadas por cepas BLEE
frecuentemente involucran a pacientes inmunocomprometidos, haciendo difícil su
erradicación en servicios de alto riesgo como las unidades de terapia intensiva (Spanu y
col., 2002). Las infecciones causadas por bacterias multirresistentes que expresan
BLEE presentan un reto para los médicos, debido a que son resistentes a la mayoría de
los betalactámicos, las infecciones causadas por estos microorganismos complican la
terapia antimicrobiana al limitar las opciones terapéuticas disponibles. Adicionalmente,
los pacientes infectados con bacterias productoras de BLEE pueden presentar una tasa
de mortalidad más alta y requerir tiempos de hospitalización más prolongados (Ramphal
y col., 2006). Los organismos que producen BLEE son, con frecuencia, resistentes a
otros agentes antimicrobianos como los aminoglicósidos, tetraciclina y trimetoprim-
sulfametoxasol, ya que muchos de estos genes de resistencia están codificados en el
mismo plásmido. La resistencia a fluoroquinolonas, la cual también está asociada
frecuentemente a la producción de BLEE es normalmente codificada a nivel
cromosomal, a diferencia de las otras resistencias asociadas; sin embargo
recientemente se ha descrito resistencia a quinolonas codificada en plásmidos. La
prevalencia de resistencia a fluoroquinolonas entre cepas productoras de BLEE varía
de acuerdo a las regiones geográficas, desde 13,7% en Canadá y 34,2% en Europa a
65,5% en la región del oeste del Pacífico (Branger y col., 2005).
Es posible clasificar las BLEE en tres grupos:
1. Las derivadas de enzimas TEM o SHV: Las enzimas TEM-1, TEM-2 y SHV-1
son enzimas plasmídicas que se encuentran frecuentemente en la
enterobacterias y que confieren resistencia a penicilinas y cefalosporinas de
espectro reducido pero no a las de espectro ampliado; sin embargo, las enzimas

23
derivadas de ellas proveen una resistencia variable a Cefotaxima, Ceftazidima y
Aztreonam. Ellas difieren de las enzimas TEM y SHV originales en cambios de
uno a cuatro aminoácidos por mutaciones puntuales que modifican su
especificidad de sustrato. Hay una gran variedad de estas enzimas (más de 20
tipos en la familia TEM).
2. Las no derivadas de enzimas TEM o SHV: tales como las enzimas MEN-1,
CTX-M1 y CTX-M2.
3. Cefalosporinasas mediadas por plásmidos: tales como FEC-1, MIR-1, CMY-1,
BIL-1, CMY-2, LAT-1, MOX-1. Ellas proveen resistencia a cefamicinas, oximino-
cefalosporinas y Aztreonam, y tienen una fuerte homología con genes ampC y
son de aparición más reciente que las derivadas de enzimas TEM o SHV
(Matsumoto y col., 1988).
En 1995, las combinaciones con inhibidores de betalactamasas reemplazaron las
cefalosporinas de tercera generación como terapia empírica en los Estados Unidos en
un esfuerzo por controlar la resistencia mediada por las BLEE. En un estudio publicado
en el año 2005, el grupo de Graffunder investigó la relación entre el uso de antibióticos
y la aparición de cepas productoras de BLEE desde 1994 hasta el año 2002 usando
análisis molecular y epidemiológico (Graffunder y col., 2005). Se realizó un estudio de
119 pacientes con cepas BLEE (+) y 132 pacientes con cepas BLEE (-). Mediante
análisis multivariado se determinó que el número de días con ventilación mecánica y el
uso previo de aminoglicósidos, cefalosporinas de tercera generación o
trimetoprim/sulfametoxasol estaban significativamente asociados con la producción de
BLEE, mientras que las combinaciones con inhibidores de betalactamasas no
estuvieron asociadas con las BLEE. La hibridación de los extractos plasmídicos
demostró que el 95% de los organismos BLEE (+) portaba int1, un elemento móvil de
ADN con un gen de resistencia a sulfonamidas y otros factores de resistencia. Se
encontraron genes específicos de resistencia a trimetoprim y aminoglicósidos en 26% y
40% de los extractos respectivamente. Estos datos indican que, aparte de los factores
de riesgo del paciente y el tratamiento con cefalosporinas de tercera generación, otros

24
antibióticos pueden proveer presión selectiva en el mantenimiento de cepas BLEE
debido a la presencia de múltiples genes de resistencia en los plásmidos. Las
combinaciones con inhibidores de betalactamasas parecen ser una opción terapéutica
aceptable para sustituir las cefalosporinas de tercera generación en las estrategias de
control de las BLEE (Graffunder y col., 2005).
Mendelson y colaboradores en el año 2005, publicaron un estudio cuyo propósito
fue determinar la prevalencia de BLEE entre cepas de Escherichia coli y Klebsiella
pneumoniae, obtenidas de muestras de orina de residentes de un centro hospitalario de
larga estadía y determinar los factores de riesgo para la adquisición de cepas
productoras de BLEE. La tasa promedio de BLEE entre las cepas de E. coli y K.
pneumoniae de este estudio fue de 25.6%. El análisis multivariado reveló tres factores
de riesgo independientes relacionados con la presencia de BLEE: anemia, sondas
urinarias permanentes y antibioticoterapia previa. El uso de fluoroquinolonas estuvo
fuertemente asociado con la presencia de cepas productoras de BLEE. La prevalencia
de cepas de E. coli y K. pneumoniae en centros hospitalarios de larga estadía fue
inesperadamente alta y corrobora la noción que los centros de larga estadía pudieran
ser reservorios importantes de bacterias resistentes.
En el año 2002 se realizó una encuesta a nivel nacional en Italia para determinar la
prevalencia y susceptibilidad antimicrobiana de miembros de la familia
Enterobacteriaceae que producian BLEE. En un período de 6 meses se obtuvieron
8015 aislamientos provenientes de pacientes hospitalizados y se les determinó la
resistencia a cefalosporinas de espectro expandido y monobactámicos. En base al
efecto sinergístico entre ácido clavulánico y ciertos betalactámicos (ceftazidima,
aztreonam, cefotaxima, cefepima y ceftriaxona) se determinó que 509 aislamientos
(6,3%) eran productores de BLEE. La hibridización de las colonias con sondas de ADN
para blaTEM y blaSHV permitió distinguir 4 genotipos distintos: positivo para TEM, positivo
para SHV, positivo para TEM y SHV y BLEEs que no eran TEM ni SHV. A todas las
cepas se les determinó la concentración mínima inhibitoria de betalactámicos,
combinaciones de betalactámicos con inhibidores, aminoglicósidos y quinolonas
mediante E-test. El 73.6% de los aislamientos correspondían a especies de Klebsiella

25
pneumoniae, Proteus mirabilis, y Escherichia coli. En general, prevalecieron las BLEE
tipo TEM con respecto a las tipo SHV (234 versus 173), mientras que la prevalencia de
las cepas que producían tanto TEM como SHV era similar a las que no producían ni
TEM ni SHV (55 y 38 respectivamente). In vitro, todos menos un aislamiento eran
susceptible a Imipenema. La susceptibilidad a otras drogras era variable:
piperacilina/tazobactam, 91%; amoxicilina/ac.clavulánico, 85%; cefoxitina, 78%;
amikacina, 76%; ampicilina-sulbactam, 61%; ciprofloxacina, 58%; y gentamicina, 56%.
Se observó con más frecuencia resistencia asociada a aminoglicósidos y quinolonas en
las cepas que producian BLEE tipo TEM (Spanu y col., 2002).
Resistencia a las Quinolonas Los mecanismos de resistencia a fluoroquinolonas incluyen mutaciones en las
topoisomerasas II y III. Estas modificaciones en la región determinante de resistencia a
quinolonas (QRDR) de los genes gyrA o gyrB y parC o parE pueden elevar los valores
de las Concentraciones Minimas Inhibitorias (CIM) hasta el rango de resistente; esta
disminución en la susceptibilidad a quinolonas se presenta principalmente en
Latinoamérica y el sur de Europa (Lorian, 1996).
La resistencia a quinolonas puede ser por diferentes mecanismos y esto tiene gran
impacto clínico. Las mutaciones pueden ocurrir durante la antibioticoterapia, y esto
constituye un factor limitante del uso de estos antimicrobianos. La resistencia a estos
antibióticos se ha relacionado con mutaciones en los genes gyrA o gyrB, como primer
paso que le confiere resistencia a ácido nalidíxico y una segunda mutación en el gen
parC o parE (los genes que codifican la topoisomerasa IV) le confieren resistencia a
Fluoroquinolonas. Adicionalmente se han observado bajos niveles de resistencia por
eflujo o impermeabilidad. La presencia de bombas de eflujo, así como alteraciones de
la permeabilidad por parte de la bacteria, normalmente conlleva a una resistencia de
bajo nivel. Ambos mecanismos pueden encontrarse asociados con mutaciones en las
topoisomerasas, incrementando el nivel de resistencia y contribuyendo, a su vez, a la
selección de la resistencia a lo largo del tratamiento (Lorian., 1996).

26
En el caso de las alteraciones del blanco de acción en microorganismos Gram
negativos, la ADN-girasa parece ser la primera diana para todas las quinolonas. Las
alteraciones en la diana se concentran en la región de la enzima denominada QRDR.
Alteraciones en las regiones QRDR, tanto de las dos subunidades de la ADN-girasa
como de la topoisomerasa IV van asociadas a un incremento de la CIM de todas las
quinolonas; de hecho, la resistencia a estos antibióticos parece ser la resultante de
varios escalones en cada uno de los cuales se produce una nueva mutación. De este
modo, la cepa, tras la primera mutación en QRDR (generalmente de gyrA), aparecerá
resistente al ácido nalidixico pero sensible a Fluoroquinolonas (solo incrementa
ligeramente la CIM) y posteriormente mutaciones en ese u otro QRDR harán que la
cepa pase a ser resistente a Fluoroquinolonas (aunque no a todas por igual).
Generalmente estas mutaciones sucesivas se asocian con otros mecanismos como las
bombas de eflujo. Por todo lo anteriormente expuesto se hace necesario informar la
presencia de estas cepas al clínico ya que están asociadas a fracaso terapéutico
(Lorian, 1996).
El aumento de la corresistencia a las fluoroquinolonas ha empezado a disminuir su
efectividad contra los patógenos productores de BLEE. Un estudio prospectivo de
infecciones sistémicas por Klebsiella pneumoniae realizado en 7 países encontró que el
18% de los aislamientos productores de BLEE también eran resistentes a
Ciprofloxacina, y que el 78% de los aislamientos de Klebsiella pneumoniae resistente a
Ciprofloxacina que causaban bacteriemia nosocomial también producían BLEE. En un
estudio realizado en Italia con enterobacterias productoras de BLEE sólo el 58%
resultaron susceptibles a Ciprofloxacina. La tasa de resistencia a Ciprofloxacina en Asia
entre cepas productoras de BLEE es muy alta; en Taiwán se ha encontrado resistencia
a Ciprofloxacina en casi el 20% de los aislamientos de Klebsiella pneumoniae
productoras de BLEE. Por otro lado, en el año 1999 en Brooklyn, Nueva York, un grupo
de 15 hospitales reportó que 34% de sus aislamientos de Klebsiella pneumoniae eran
posibles productores de BLEE, y que de estos sólo el 42% eran susceptibles a
Ciprofloxacina. Los factores de riesgo identificados para la resistencia a

27
fluoroquinolonas en E. coli y K. Pneumoniae productoras de BLEE incluyen el uso de
fluoroquinolonas, aminoglicósidos y una hospitalización larga (Ramphal y col.. 2006).
Recientemente se ha descrito que la resistencia a las Quinolonas también tiene un
origen plasmídico, contrario a lo que anteriormente se creía. En 1998 en un aislamiento
de K. pneumoniae en USA se identificó el primer gen de resistencia a las Quinolonas, el
gen QnrA transferible por plásmidos, el cual codifica para la proteína QnrA (Jiang y col.,
2008) Esta proteína interfiere con la acción de las Quinolonas sobre sus dianas,
confiriendo bajos niveles de resistencia a las Quinolonas (Minarini y col., 2008). Otros
dos genes plasmídicos han sido identificados como QnrB y QnrS cuyas proteínas solo
difieren en un 41% y un 60% de aminoácidos con respecto a QnrA respectivamente
(Cattoir y col., 2007) y que también codifican para proteínas protectoras de las dianas.
La expresión de estos genes induce a un bajo nivel de resistencia, observándose cepas
que presentan la proteína Qnr y son Ácido Nalidíxico sensibles, esto las hace reservorio
oculto para la propagación de genes plasmídicos tipo Qnr. Igualmente se ha descrito la
presencia de mecanismos de inactivación enzimática relacionados con una variante de
la enzima Acetilasa de Aminoglicósidos denominada AAC(6’)-Ib-cr, capaz de reducir los
halos de sensibilidad para Ciprofloxacina y Norfloxacina (Pitout y col., 2008).
Desde el punto de vista epidemiológico, la presencia de estos cinco genes
plasmídicos que confieren resistencia a las Quinolonas es alarmante, debido a la
posible transmisión horizontal a través de elementos génicos móviles a cepas
originalmente sensibles a las Quinolonas, hecho que en un futuro podría determinar la
aparición de brotes epidémicos difíciles de controlar. La presencia de mecanismos
plasmídicos de resistencia a las Quinolonas ligado a una mutación cromosómica
favorecería la aparición de cepas altamente resistentes a las Fluoroquinolonas.
La resistencia debida a Proteínas Qnr ha sido identificada en K. pneumoniae, K.
oxytoca, E. coli, C. freundii, C. koseri, E. cloacae, S. flexneri y Salmonella sp. (Szabó y
col., 2008; Cavaco y col., 2008; Rodríguez-Martínez y col., 2008) Esta proteína se une a
la DNA-girasa y se postula que esta unión ocurre antes del bloqueo de la enzima por
parte de la Quinolona, evitando que la misma se una a la DNA-girasa inactivándola.

28
(Sierra, 2005) La presencia de este gen de resistencia se ha descrito en diferentes
partes del mundo: China, USA, Alemania, Francia, Japón y Turquía, aunque con una
baja prevalencia (Jonas y col., 2005; Mammeri y col., 2005; Wang y col., 2003; Wang y
col., 2004). Este mecanismo de resistencia aumenta la CIM de las cepas que la
presentan de 4 a 8 veces (Rodríguez-Martínez, 2005; Tran y col., 2005) y complementa
a otros mecanismos de resistencia como la adquisición de mutaciones o a los sistemas
de expulsión activa (Tran y Jacoby., 2002). Se cree que estos mecanismos producen un
bajo nivel de resistencia a las Quinolonas (Nordmann y Poirel., 2005; Chen y col., 2006;
Wang y col., 2004; Alós, 2009), pero estudios realizados en Sevilla, España, revelaron
que la presencia de mecanismos de resistencia plasmídicos tipo QnrA1 junto a
mecanismos adicionales de resistencia (Alteración de las dianas, déficit de porinas o
bombas de eflujo) aumentan de forma significativa la CIM de Ciprofloxacina y
Levofloxacina (Rodríguez-Martínez y col., 2008) lo cual conduciría probablemente al
fracaso terapéutico cuando el tratamiento de elección son las quinolonas.
Las proteínas Qnr pertenecen a una familia de pentapéptidos que para la fecha
tiene 90 miembros conocidos, y que han sido bien descritos en las Cianobacterias,
donde forman parte de las membranas y están vinculadas al citoplasma. Ellas tienen
una estructura en alfa-hélice en su circunferencia externa y hojas beta-plegadas en su
circunferencia interna, característica que la hace una estructura apropiada para la
interacción con proteínas (Nordmann y Poirel., 2005).
El mecanismo de acción de las proteínas Qnr se basa en la unión de esta
proteína a la ADN girasa; esta unión no requiere la presencia del complejo ADN girasa-
Quinolona-ADN para su formación. Se postula que la formación del complejo Qnr-ADN
girasa ocurre antes del bloqueo de la ADN girasa por parte de la Quinolona. Además se
ha observado una disminución en la formación del complejo en presencia de Qnr (Tran
y Jacoby., 2002).
La proteína QnrA (la primera de estas proteínas en ser caracterizada) está
formada por 218 aminoácidos, y fue detectada en un aislado de K. pneumoniae en
USA en el año 1998 (Martínez-Martínez y col., 1998) A partir de esa fecha, hasta la

29
actualidad han sido descritas 6 variantes de la misma (QnrA1-QnrA6). Otras 2
proteínas, las QnrB y QnrS hasta el momento presentan 19 variantes y 3 variantes
respectivamente (Minarini y col., 2008) y difieren aproximadamente en un 41% (QnrB) y
un 60% (QnrS) de aminoácidos con respecto a QnrA (Cattoir y col., 2007).
La proteína QnrA y QnrB han sido descritas principalmente en especies como K.
pneumoniae, K. oxytoca, E. coli, C. freundii, C. koseri y E. cloacae, mientras que la
proteína QNRS fue descrita por primera vez en 2005 en un aislado de S. flexneri en
Japón (Hata y col., 2005).
Se cree que estos mecanismos producen un bajo nivel de resistencia a las
Quinolonas, pudiendo incrementar las CIMs de 8 a 12 veces para estos antibióticos
(Jiang y col., 2008). Algunos autores sugieren que las proteínas Qnr confieren
resistencia al Ácido Nalidíxico más no a las Fluoroquinolonas (Nordmann y Poirel.,
2005), mientras que otros autores, van más allá y aseguran que estas proteínas brindan
un bajo nivel de resistencia a las Fluoroquinolonas, efecto que se ve pronunciado si se
adiciona la presencia de alguno de los mecanismos clásicos de resistencia a
Quinolonas, como lo son las alteraciones de las dianas por mutaciones cromosómicas
(Rodríguez-Martínez, 2005). Igualmente se ha descrito que estos genes se transmiten
de forma conjunta, en la mayoría de los casos, con genes de resistencia codificantes
para BLEE (Chen y col., 2006; Wang y col., 2004).
En los procesos de adaptación de resistencia para los Aminoglicósidos, las
enzimas modificadoras de la estructura de la molécula estas sufrieron variaciones y una
de esas modificaciones condujo a la formación de la enzima aac(6´)-Ib-cr, la cual es
capaz de inactivar las Fluoroquinolonas que poseen un grupo piperacínico como radical
sustituyente, tal es el caso de Norfloxacina y Ciprofloxacina. Además, es capaz de
inactivar los Aminoglicósidos: Kanamicina, Tobramicina y Amikacina (Pitout y col.,
2008). Esto las hace reconocibles en los Antibiogramas. Estas enzimas son capaces de
reducir la actividad de Ciprofloxacina y Norfloxacina, lo cual aumentaría las CIM para
estos antibióticos. Por ser un mecanismo enzimático, depende del inóculo, a mayor
inóculo, mayor producción de la enzima, por lo tanto, la cantidad de antibiótico

30
inactivado será mayor. Se piensa que el gen aac(6´)-Ib-cr que codifica para esta
enzima, se transmite de forma conjunta con los genes Qnr y genes codificantes para
BLEE (Jiang y col., 2008).
Relevancia de la transmisión de la resistencia bacteriana La terapéutica de las infecciones nosocomiales y las adquiridas en la comunidad
está seriamente afectada por la evolución continua de la resistencia antimicrobiana. Se
estima que más del 60% de los dos millones de infecciones nosocomiales que ocurren
en los Estados Unidos cada año son causadas por bacterias resistentes a los
antibióticos, lo que resulta en un aumento de la morbilidad y mortalidad y altos costos
de salud. La emergencia y diseminación de los determinantes de resistencia
antimicrobiana continúa siendo un reto para el tratamiento de infecciones serias. A lo
largo de la última década, hemos sido testigos del feroz aumento de la resistencia
antimicrobiana en los ambientes nosocomiales y comunitarios; aun cuando en los
últimos años se han diseñado nuevos antibióticos para tratar los microorganismos
multirresistentes, la resistencia ha continuado emergiendo y diseminándose. Hemos
aprendido mucho en la última década sobre los mecanismos por los cuales estas
bacterias se hacen resistentes a múltiples antibióticos, y se espera que este
conocimiento sirva para implementar nuevas estrategias para el desarrollo de drogas y
a la vez limitar la morbilidad y mortalidad asociada de forma más eficiente.
La mayoría de los antibióticos usados en la actualidad son productos derivados de
hongos y bacterias; por lo tanto, es razonable pensar que aquella bacteria que
produzca antibióticos debe a su vez poseer mecanismos por los cuales pueda evitar la
acción letal del mismo. Por otro lado, en vista de que es raro encontrar en la naturaleza
cultivos puros, también es lógico razonar que las bacterias que comparten un nicho
ecológico con microorganismos productores de antibióticos deberían desarrollar
también mecanismos para sobrevivir en dicho ambiente, siendo el mecanismo más
directo en muchos casos la adquisición de los genes de resistencia que poseen los
microorganismos productores de antibióticos (Lorian, 1996). Es interesante señalar que

31
el genoma bacteriano es una entidad dinámica y cambiante, que no ha dejado de
evolucionar aunque mantiene un tamaño reducido y consiste en un mosaico de genes
con diferentes historias. Se ha sugerido que al menos 17% de los genes de E. coli son
adquisiciones foráneas relativamente recientes, con una tasa de transferencia
horizontal de aproximadamente 16kb cada millón de años.
Una notable importancia del genoma bacteriano consiste en que la información
genética esencial de la célula bacteriana está contenida en una única molécula de ADN
de doble cadena con un tamaño típico de unos pocos millones de bases; el cromosoma
de las bacterias contiene, en esencia, toda la información necesaria para la regulación
de la expresión de las vías metabólicas, para la duplicación de todos los componentes
celulares durante el crecimiento y para la diferenciación estructural que se presenta
durante procesos tales como la esporulación (Joklik y col., 1992). En vista de la
complejidad funcional del genoma bacteriano y de la capacidad de diseminación
horizontal de genes de resistencia entre las bacterias, muchos investigadores han
buscado estudiar dichas características genéticas y su interrelación; por ejemplo, en el
año 2005 el grupo de Branger y colaboradores (Branger y col., 2005), para determinar
la implicación del “trasfondo” genético de cepas de Escherichia coli en la emergencia de
BLEE y su capacidad de diseminación, estudiaron 55 cepas clínicas con BLEE tipo
TEM, 52 con BLEE tipo CTX-M y 22 con BLEE tipo SHV provenientes de muestras
extraintestinales en lo que respecta a sus grupos filogenéticos, contenido de factores de
virulencia (genes pap, sfa/foc, hly y aer) y resistencia a fluoroquinolonas. Un análisis
factorial de correspondencia demostró que las BLEE tipo SHV, y en menor grado las
TEM, se observaban principalmente en las cepas del grupo filogenético B2, las cuales
presentaban numerosos factores de virulencia pero eran susceptibles a
fluoroquinolonas, mientras que el tipo CTX-M estaba asociado con el grupo filogenético
D, que era deficiente- en factores de virulencia pero era resistente a fluoroquinolonas;
esto indica un claro intercambio entre la expresión de factores de virulencia y la
resistencia a fluoroquinolonas. Es importante destacar que el alto nivel de resistencia a
quinolonas en las cepas con CTX-M y su bajo nivel de virulencia les provee una gran
ventaja selectiva (en particular en ambientes donde se usen las fluoroquinolonas como

32
terapéutica) lo cual podría explicar la gran capacidad de diseminación que han
demostrado las cepas con este genotipo en los últimos años.
Por otro lado, la investigación por Reacción en Cadena de la Polimerasa (PCR) de
los genes plasmídicos de resistencia a quinolonas qnr resultó negativa, lo cual permitió
deducir que la asociación entre ambas resistencias no fue debida a que los genes de
BLEE y la resistencia a quinolonas estuviesen codificados en el mismo elemento móvil;
en este estudio se determinó que existía una relación causal entre el uso de
fluoroquinolonas y la emergencia de BLEE. El grupo concluyó que las BLEE no se
distribuyen al azar entre la gran diversidad genética de las distintas E. coli, sino que su
captación, expresión y mantenimiento depende del tipo de BLEE, el trasfondo
filogenético de la cepa, la virulencia intrínseca de la misma y la presencia de resistencia
asociada a fluoroquinolonas, lo cual refleja la diversidad de los nichos ecológicos con
diferentes presiones selectivas (Branger y col., 2005). Es importante destacar que la
transferencia de genes entre especies bacterianas que no son naturalmente
competentes está mediada principalmente por elementos genéticos móviles como son
los plásmidos a través del proceso de conjugación, y éstos pueden a su vez contener
elementos como los integrones.
Los plásmidos son elementos genéticos compuestos de ADN que se replican
independientemente del cromosoma del hospedador, y que a diferencia de los virus no
tienen forma extracelular. Estos elementos pueden codificar fenotipos de resistencia a
varios antibióticos, varios iones metálicos (mercurio), radiación ultravioleta y bromuro de
etidio. Es posible diferenciar entre un plásmido y un cromosoma en los procariotas
porque los plásmidos no llevan genes que sean requeridos por el hospedador en todas
las condiciones (no transportan genes indispensables); sin embargo, esta característica
a veces es difícil de demostrar y por tanto, a veces es complicado distinguir entre
cromosomas y plásmidos muy grandes. Los plásmidos tienen la capacidad de
diseminarse epidémicamente de modo "horizontal" (es decir, entre células distintas de
la misma especie o de distintas especies) y pueden ser transferibles por sí mismos
(plásmidos conjugativos) o requerir de otros plásmidos para movilizarse (plásmidos no
conjugativos); cuando no existe presión selectiva, los plásmidos de resistencia pueden

33
perderse de la mayor parte de las bacterias de una determinada población, pero su
modo de transmisión "epidémica" los capacita para diseminarse rápidamente a la
mayoría de la población cuando la situación lo requiere (Madigan, 1998).
La conjugación es el intercambio unidireccional de información genética mediada
por plásmidos desde una bacteria donante a otra receptora mediante un contacto real
(Figura 4). La capacidad de conjugación depende de la presencia en la bacteria de
plásmidos conjugativos que contienen los genes necesarios para tal proceso. Los
genes que controlan la conjugación se encuentran localizados en la región tra del
plásmido, los cuales están relacionados con la síntesis de una estructura superficial
conocida como pilus sexual, que permite y establece el contacto físico entre la bacteria
donante y la receptora.
Figura 4. Esquema del proceso de Conjugación bacteriana. Tomado de
http://blog.evolutionibus.info

34
Se distinguen en la conjugación dos fases:
Primera fase (contactos entre células): Las células donantes de 1 a 10 pili. El Pili
interacciona con un receptor de la célula receptora. El pili se va despolimerizando
desde su base, lo cual provoca la apariencia de que se va retrayendo. En ese
movimiento de retracción, la cédula receptora se va acercando a la donante. Cuando el
Pili se termina de desintegrar, las células se ponen en contacto directo pared-pared. Se
forma un puente conjugativo que pone en contacto los citoplasmas de ambas cédulas y
se produce el paso de una hebra de ADN. Tras cierto tiempo de conjugación, el
agregado se desagrega de forma activa.
Segunda fase (transferencia de ADN): Esta transferencia representa un proceso
especial de replicación asimétrica por círculo rodante. Una de las dos cadenas
parentales del plásmidos pasa a la célula receptora, replicándose en ella. La otra
cadena parental se queda en el donador, sirviendo a su vez como molde para la
síntesis de una nueva cadena complementaria (Koneman y col., 1999).
La conjugación es uno de los procesos de transferencia de material genético mas
sofisticado, complejo y de mayor rango de acción, superando la barrera no solo de las
especies, sino también de los reinos, además de ser uno de los medios de
transferencia de genes de resistencia más efectivo y especifico que se conoce. La
conjugación es esencialmente un proceso de replicación, que permite que un plásmido
se replique más frecuentemente que los genes cromosomales, y además habilita la
transferencia alternativa de un hospedador bacteriano. Por consiguiente, una gran
proporción de varios tipos de plásmidos aislados de diferentes géneros bacterianos son
conjugativos (Madigan, 1998).
Los integrones son trozos de ADN de doble hebra que capturan genes de
resistencia del citoplasma celular y que contiene promotores fuertes para expresar esos
genes. Los genes que captan se denominan genes “casettes” y se insertan entre los

35
extremos 5’CS y 3’CS (secuencias conservadas) a través de una integrasa codificada
por el gen int. Cabe destacar que los genes que captan tienen en su extremo final una
secuencia de 59 pb, a través de las cuales se produce su inserción. Se han detectado
en integrones genes de resistencia a los aminoglucósidos (acetiltransferasas,
nucleotidiltransferasas o fosfotransferasas), genes de resistencia a trimetoprima,
cloranfenicol y betalactámicos. Los integrones mejor estudiados se sitúan sobre
transposones y requieren incorporarse a material genético capaz de replicarse para
poder dividirse y diseminarse (Madigan M., 1998).
Caracterización de mecanismos de resistencia en el laboratorio de microbiología
El papel primordial del laboratorio de microbiología clínica consiste en brindar
información con la cual los médicos puedan diagnosticar y tratar enfermedades
infecciosas, siendo una de las piezas de información más importante la que arroja un
cultivo, en cuanto a qué antimicrobiano servirá para un tratamiento adecuado y a cuales
será resistente la bacteria, siendo importante además determinar si dicha resistencia es
intrínseca de la bacteria o si por el contrario es adquirida y potencialmente transmisible
a otras cepas. A nivel del Laboratorio de Microbiología se pueden caracterizar los
mecanismos de resistencia de los antibióticos mediante el uso de técnicas accesibles,
como son:
Técnica de Hodge: Es un método microbiológico cualitativo y sensible que permite
determinar si la resistencia bacteriana es de naturaleza enzimática, el cual ha sido
aceptado como una herramienta útil en el Laboratorio.
Conjugación plasmídica: El método de conjugación permite detectar la transferencia
de un plásmido seleccionado que posee resistencia a un determinado antibiótico.
Para llevar a cabo tal experimento se requiere de una célula donadora (denominado
macho), que contiene un tipo particular de plásmido conjugativo que permite
transferir una copia de sí mismo a una célula receptora que carece de él

36
(denominado hembra), cuyo proceso de transferencia es llevado a cabo por el pili
sexual que posee la célula donadora.
Isoelectroenfoque: Esta técnica permite determinar el punto isoeléctrico de la enzima
o enzimas en estudio y es de gran ayuda para confirmar la interpretación fenotípica,
por cuanto permite detectar la presencia de una o más enzimas de resistencia y
aporta información fisicoquímica sobre su naturaleza. Esta técnica se basa en que
las cargas que tienen las proteínas dependen del número e hidratación de los
residuos de aminoácidos con cadenas laterales ionizables; si estos grupos están
cargados o no depende del pH. Si las proteínas se colocan en un gradiente de pH
generado eléctricamente ellas migrarán de acuerdo a su carga, y su carga variará
mientras se mueven a regiones de distinto pH; cuando la enzima alcance su punto
isoeléctrico su carga neta será 0, dejará de migrar y formará una banda definida en el
gel (Lorian, 1996).
PCR: es una técnica altamente sensible por medio de la cual secuencias de ADN que
se encuentran en cantidades muy pequeñas se pueden amplificar enzimáticamente
de un modo tal que es posible disponer de una cantidad suficiente para alcanzar una
señal umbral detectable. La técnica puede ser usada para detectar cantidades muy
pequeñas de ácido nucleico específico en muestras clínicas en las que se piensa que
agentes bacterianos, fúngicos o virales tienen un papel causal. La base fundamental
de esta tecnología es que cada agente infeccioso causante de una enfermedad
posee una secuencia característica propia en su composición de ADN o ARN por
medio de la cual puede ser identificado (Koneman y col., 1999). El PCR es un
proceso altamente sensible; aun cuando la mayoría de los análisis bioquímicos
(incluida la detección de ácidos nucleicos con radioisótopos) requieren el uso de
cantidades significativas de material biológico, para la PCR la cantidad de muestra
requerida es muy poca. Esta característica hace que la técnica sea extremadamente
útil, no sólo en investigación básica, sino también en usos comerciales como pruebas
de identificación genética, métodos forenses, control de calidad industrial y
diagnóstico in vitro. Tal como fue desarrollado originalmente (Mullis y Faloona, 1987),
el PCR amplifica segmentos cortos (de aproximadamente 100 a 500 pares de bases)

37
de una molécula de ADN más grande. Una amplificación típica incluye la muestra de
ADN blanco, una polimerasa de ADN termoestable, dos “iniciadores”,
desoxinucleótidos trifosfato (dNTPs), buffer de reacción, magnesio y aditivos
opcionales. Los componentes de esta reacción son mezclados y la reacción toma
lugar en un termociclador, el cual es un instrumento automatizado que lleva la
reacción a través de una serie de temperaturas diferentes por tiempos variables. Esta
serie de temperaturas y ajustes de tiempo son lo que se conocen como un ciclo de
amplificación. Cada ciclo de PCR teóricamente dobla la cantidad de ADN blanco
(amplicón) presente en la reacción. Diez ciclos teóricamente multiplican el amplicón
por un factor de mil; 20 ciclos por un factor de más de un millón en cuestión de horas.
Cada ciclo de amplificación por PCR consiste en un número de pasos que producen
dos hebras de ADN de cadena simple cebados por los “iniciadores”, inician la
reacción de polimerización, y sintetizan una copia de cada hebra del ADN blanco.
Estos pasos deben ser optimizados para cada combinación de iniciadores-ADN
blanco (Doyle, 1996).
Secuenciación: La técnica de secuenciación más utilizada es la desarrollada por
Frederick Sanger en 1975. En ella, se emplean dideoxinucleótidos trifosfato (ddNTP)
en la síntesis del ADN. Son moléculas que parecen nucleótidos normales excepto
por carecer de un grupo hidroxilo en posición 3’. Pueden añadirse al extremo en
crecimiento de la cadena, pero finalizan la síntesis catalizada por ADN polimerasa
porque no pueden añadirse más nucleótidos para ampliar la cadena. Se mezcla una
cadena sencilla del ADN que se va a secuenciar con un cebador, la ADN polimerasa
I, nucleósidos trifosfato normales (NTP) y una pequeña cantidad de los
desoxinucleótidos que han sido marcados con un colorante fluorescente (cada
ddNTP se marca con un colorante de distinto color). La síntesis del ADN se inicia con
el cebador y finaliza cuando se incorpora ddNTP en lugar de un NTP normal. El
resultado en una serie de fragmentos de longitudes diferentes. Se producen cuatro
reacciones, cada una con un ddNTP fluorescente diferente. La mezcla con ddATP
produce fragmentos con un A terminal, la mezcla con ddCTP produce fragmentos
con un C terminal y así sucesivamente. Los fragmentos fluorescentes se separan del
molde de ADN y se someten a electroforesis en gel de poliacrilamida para separarse

38
unos de otros según su tamaño. En los sistemas automatizados, los productos de las
cuatro reacciones se mezclan y se someten juntos a electroforesis; como cada
ddNTP presenta una fluorescencia de diferente color, puede comprobarse el gel con
un detector y determinar rápidamente la secuencia según el orden de los colores que
aparecen en las bandas (Prescott y col., 1999).
MARCO METODOLÓGICO
Tipo de estudio Según la metodología empleada este trabajo se inscribe dentro de la clasificación
de trabajo de campo experimental, ya que en él se determinó experimentalmente la
sensibilidad o resistencia de ciertas cepas de Enterobacterias frente a algunos
antibióticos predeterminados. Según los objetivos el trabajo es de tipo descriptivo, ya
que caracterizó el fenómeno de la producción de BLEE en las cepas de
Enterobacterias aisladas.
Muestra La muestra estuvo conformada por 50 cepas de Enterobacterias con
susceptibilidad disminuida a cefalosporinas aisladas de distintas fuentes biológicas.
El muestreo se obtuvo a partir de cepas con sospecha fenotípica de producción de
BLEE aisladas de los laboratorios de bacteriología desde enero hasta junio del año
2006 del Hospital Universitario de Caracas, Hospital J. M. de los Ríos, Hospital Dr.
Ricardo Baquero, Centro Policlínico Valencia, Laboratorio Cesar Sánchez Font,
Policlínica Santiago de León, Hospital Central Antonio María Pineda e Instituto
Nacional de Higiene, aceptándose cepas hasta alcanzar el número planteado. De
estas cepas, sólo se caracterizaron aquellas que fueron confirmadas como
productoras de Betalactamasas de Espectro Expandido.

39
Metodología de laboratorio Se realizó la identificación a nivel de género y especie de las cepas en estudio
mediante pruebas bioquímicas convencionales, evaluándose los siguientes
parámetros:
Fermentación de Glucosa y Lactosa a partir de Agar Kligler.
Determinación de la vía de utilización de la glucosa en caldo MRVP.
Utilización del Malonato como fuente de carbono en Caldo Malonato.
Utilización del Citrato como fuente de carbono en Agar Citrato.
Descarboxilacion de los Aminoácidos Lisina, Arginina y Ornitina en Caldos
Moeller Descarboxilasa.
Metabolismo de la Urea en agar Urea de Christensen.
Determinación de la presencia de la enzima Ortonitrofenilgalactosidasa
mediante el uso de taxo ONPG.
Fermentación de Arabinosa, Ramnosa e Inositol en Caldo Andrade.
Metabolismo de Fenilalanina en Agar Fenilalanina.
Determinación de motilidad en agar semisólido.

40
Para determinar el patrón fenotípico de resistencia de las enterobacterias se
usó la metodología de Kirby-Bauer recomendada por la CLSI 2010 y se determinó la
susceptibilidad a los siguientes antimicrobianos: Ceftazidima 30 µg, Cefotaxima 30
µg, Amoxicilina/Ac. Clavulánico 20/10 µg, Ciprofloxacina 5 µg, Levofloxacina 5 µg,
Ofloxacina 5 µg, Ac. Nalidíxico 30 µg, Trimetoprim/Sulfametoxasol, Cefepima 30 µg
y Aztreonam 30 µg de las cepas en estudio. Todos los discos empleados fueron
marca Oxoid. Adicionalmente se determinó la Concentración Mínima Inhibitoria para
Ciprofloxacina por el método de E-test (Biomerieux)
La detección presuntiva las cepas productoras de BLEE fue determinada por dos
métodos:
1) El método de difusión en disco de Kirby-Bauer empleando en conjunción discos
de Amoxicilina/ácido Clavulánico con discos de Ceftazidima, Cefotaxima,
Cefepima y Aztreonam separados entre sí por 25 mm (difusión doble del disco),
en el cual una prolongación de la zona de inhibición mayor o igual a 4 mm
alrededor de un disco de un oximino -lactámico en dirección al disco de ácido
clavulánico sugirió la presencia de enzimas betalactamasas de espectro
expandido.
2) El método de difusión en disco de Kirby-Bauer comparando el halo de inhibición
(obtenido en un antibiograma realizado por el método de Kirby-Bauer) alrededor
de un disco de una cefalosporina de tercera generación (Cefpodozima,
Ceftazidima, Cefotaxima) con el halo de inhibición alrededor de un disco que
contiene una combinación de la misma Cefalosporina más Acido Clavulánico
(Ceftazidima/Clavulánico, Cefotaxima/Clavulánico); en este caso se consideró la
presencia de BLEE cuando existe una diferencia mayor o igual a 5 mm entre el
halo de inhibición de la cefalosporina y el de la cefalosporina más clavulánico.
La confirmación genotípica de la producción de las BLEE y resistencia a
quinolonas se realizó mediante la determinación de los genes que codifican para
dicha enzima al realizar ensayos de Reacción en Cadena de la Polimerasa (PCR)
del tipo PCR simple usando iniciadores específicos para los genes que codifican

41
dichas enzimas (TEM, SHV, PER, CTX-M, gyrA, qnrA, qnrB, qnrS, aac(6’)ib-cr).
La mezcla de reacción empleada fue la misma para todos los genes: Vol. final: 50
μl; Buffer ADN Taq polimerasa: 1X; Primers: 50 pmoles de c/u.; ADN Taq
polimerasa: 1 U; dNTPs: 25 μM de cada uno; ADN molde: 5 μl; MgCl2: 1,5 mM. El
programa empleado en el ciclador térmico contempló un paso de
desnaturalización inicial de 5 min. a 94ºC, y la amplificación se llevó a cabo por 30
ciclos, con pasos de desnaturalización (30 seg. a 94ºC), alineación (30 seg a 55
ºC para TEM, CTX-M, GyrA y PER, a 54 ºC para Aac(Ib)cr y QnrB, a 57 ºC para
QnrA y QnrS y a 59 ºC para SHV) y extensión (30 seg. a 72ºC), y un paso final de
extensión de 5 min. a 72ºC (Galas y col., 2004). Se corrieron controles positivos
de extracción de ADN a cada cepa en estudio paralelamente a la amplificación de
detección, empleando iniciadores específicos para el cluster génico codificante
del ARNr 16S, un control positivo del gen a amplificar y un control negativo sin
ADN. Los productos finales de amplificación fueron detectados en geles de
agarosa al 1,2 %, teñidos con bromuro de etidio 0,5 μg/mL y se visualizaron bajo
luz ultravioleta mediante el equipo Gel Doc 2000 (Bio-Rad), empleando un
marcador de peso molecular de 100 pares de bases. Los iniciadores y los
controles positivos que se emplearon para el PCR se especifican en la tabla 3.
Una vez obtenidos los productos de amplificación, aquellos cuyos iniciadores
no permiten discriminar entre las múltiples variantes del gen o en los que se
requerían la detección de mutaciones (TEM, SHV, gyrA) se remitieron al servicio
de secuenciación del Laboratorio de Biología Molecular de Protozoarios del
Instituto de Medicina Tropical de la UCV, donde se utilizó un secuenciador de
ácidos nucléicos modelo ABI 3130XL (Applied Biosystems) de cuatro capilares.
En esta metodología, los fragmentos de ADN se marcan con cuatro fluoróforos,
de manera que cada nucleótido tiene un color diferente. Una vez obtenida la
secuencia se determinó, mediante la comparación de la secuencia obtenida con
la base de datos GeneBank empleando el programa BLAST de los Institutos
Nacionales de Salud de los Estados Unidos (NCBI/NIH), la variante del gen o la
mutación que se encuentraba presente en la cepa.

42
Tabla 3. Iniciadores empleados en el estudio
ENZIMA SECUENCIA AMPLICON
(pb) REFERENCIA
CONTROL POSITIVO
16S F
16S R
5’-AGGAGGTGATCCAACCGCA-3’
5’-AACTGGAGGAAGGTGGGGAT-3’ 370 Greisen, 1994
Escherichia coli ATCC
35218
PER F
PER R
5’-GTAGTATCAGCCCAATCCCC-3’
5’-CCAATAAAGGCCGCCGTCCATCA-3’ 740
Melano R, 2005. Comunicación personal.
Instituto “Dr. Carlos Malbrán”. Argentina
Klebsiella pneumoniae INH 77915
OS(SHV) F OS(SHV) R
5’-TCGGGCCGCGTAGGCATGAT-3’ 5’- GCAGGGCGACAATCCCGCG-3’
620 Arlet y Philippon, 1991 Klebsiella
pneumoniae INH 77915
OT(TEM) F OT(TEM) R
5’-TTGGGTGCACGAGTGGGTTA-3’ 5’-TAATTGTTGCCGGGAAGCTA-3’
502 Arlet y Philippon, 1991 Klebsiella
pneumoniae INH 77915
CTX-M-2 F CTX-M-2 R
5’-CGGAATTCATGATGACTCAGAGCATTCG-3’ 5’-GCTCTAGATTATTGCATCAGAAACCGTG-3’
850
Melano R, 2005. Comunicación personal.
Instituto “Dr. Carlos Malbrán”. Argentina
Klebsiella pneumoniae INH 77915
GyrA 5´- CAACGAAATCGACCGTCTCTT-3´ 5´- ACGTATTGGGCAAYTACTGCA-3´
282
Melano R, 2005. Comunicación personal.
Instituto “Dr. Carlos Malbrán”. Argentina
Escherichia coli ATCC
35218
QnrA 5’- AGAGGATTTCTCACGCCAGG-3’ 5’- TGCCAGGCACAGATCTTGAC-3’
579 Cattoir y col., 2007 -
QnrB 5’-GATCGTGAAAGCCAGAAAGG-3’ 5’-ACGATGCCTGGTAGTTGTCC-3’
469 Pitout y col., 2008 Klebsiella
pneumoniae EB 083
QnrS 5’-GCAAGTTCATTGAACAGGGT-3’
5’-TCTAAACCGTCGAGTTCGGCG-3’ 427 Cattoir y col., 2007
Klebsiella pneumoniae
EB 024
Aac(Ib)cr 5’-TTGCGATGCTCTATGAGTGGCTA-3’
5’-CTCGAATGCCTGGCGTGTTT-3’ 482 Minarini y col., 2008
Klebsiella pneumoniae
EB 092
5’CS 3’CS
5’-GGCATCCAAGCAGCAAG-3’ 5’-AAGCAGACTTGACCTGA-3’
variable
Melano R, 2005. Comunicación personal.
Instituto “Dr. Carlos Malbrán”. Argentina
Klebsiella pneumoniae
KP13
Int1 5´- ATCATCGTCGTAGAGACGTCGG -3´ 5´- GTCAAGGTTCTGGACCAGTTGC -3´
893 Reyes y col, 2003
Salmonella sp. EB 2757/09
La caracterización fenotípica de la resistencia enzimática se efectuó mediante la
técnica de Hodge, la cual determina si la célula produce alguna enzima capaz de
hidrolizar el antibiótico en estudio (Sandra y col., 2009). Para realizar la técnica de
Hodge se hisopa una suspensión 0,5 McFarland de una cepa de referencia (E. coli
ATCC 25922) según la técnica de Kirby y Bauer en agar Mueller Hinton, se coloca un
taxo del antibiótico a evaluar y dentro del diámetro del halo de susceptibilidad esperado
para esa cepa de referencia/antibiótico se coloca una estría de la cepa en estudio
perpendicular al halo de susceptibilidad esperado sobrepasándolo. Una prueba positiva
está constituida por el crecimiento confluente de la cepa de referencia hacia la estría de
la cepa problema lo cual conlleva a la deformación del halo de susceptibilidad.
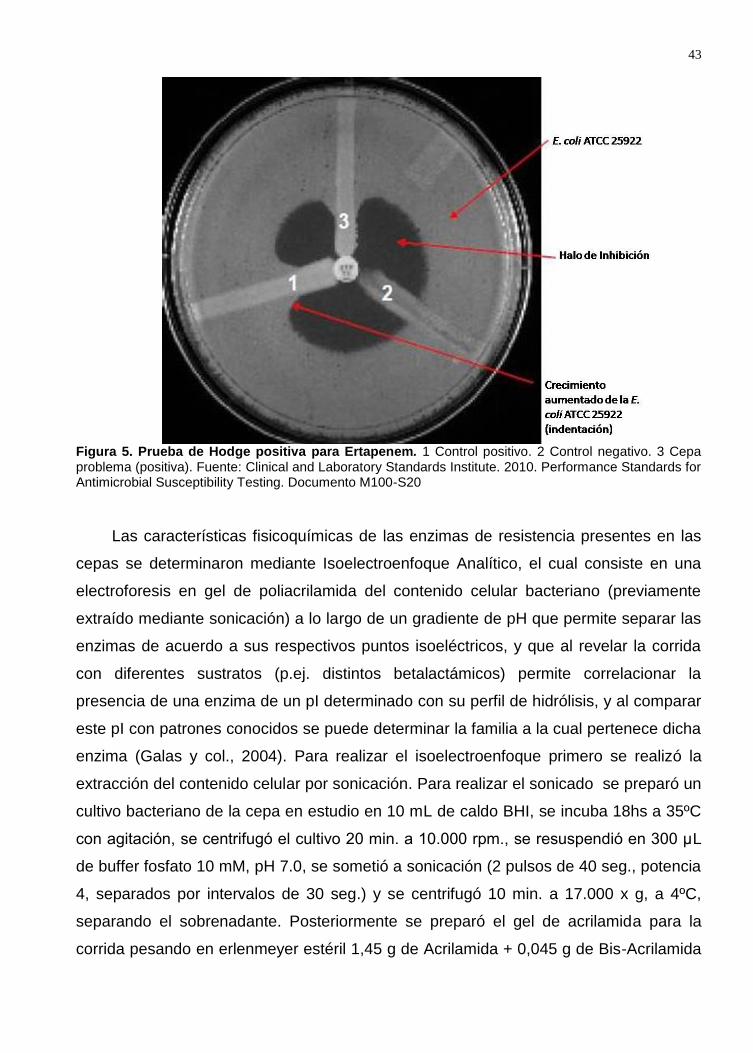
43
Figura 5. Prueba de Hodge positiva para Ertapenem. 1 Control positivo. 2 Control negativo. 3 Cepa problema (positiva). Fuente: Clinical and Laboratory Standards Institute. 2010. Performance Standards for Antimicrobial Susceptibility Testing. Documento M100-S20
Las características fisicoquímicas de las enzimas de resistencia presentes en las
cepas se determinaron mediante Isoelectroenfoque Analítico, el cual consiste en una
electroforesis en gel de poliacrilamida del contenido celular bacteriano (previamente
extraído mediante sonicación) a lo largo de un gradiente de pH que permite separar las
enzimas de acuerdo a sus respectivos puntos isoeléctricos, y que al revelar la corrida
con diferentes sustratos (p.ej. distintos betalactámicos) permite correlacionar la
presencia de una enzima de un pI determinado con su perfil de hidrólisis, y al comparar
este pI con patrones conocidos se puede determinar la familia a la cual pertenece dicha
enzima (Galas y col., 2004). Para realizar el isoelectroenfoque primero se realizó la
extracción del contenido celular por sonicación. Para realizar el sonicado se preparó un
cultivo bacteriano de la cepa en estudio en 10 mL de caldo BHI, se incuba 18hs a 35ºC
con agitación, se centrifugó el cultivo 20 min. a 10.000 rpm., se resuspendió en 300 μL
de buffer fosfato 10 mM, pH 7.0, se sometió a sonicación (2 pulsos de 40 seg., potencia
4, separados por intervalos de 30 seg.) y se centrifugó 10 min. a 17.000 x g, a 4ºC,
separando el sobrenadante. Posteriormente se preparó el gel de acrilamida para la
corrida pesando en erlenmeyer estéril 1,45 g de Acrilamida + 0,045 g de Bis-Acrilamida

44
y agregando 28,3 ml de agua destilada estéril, 1.75 ml de anfolitos, 36 microlitros de
persulfato y 176 microlitros de TEMED (Tetrametiletilendiamina). Inmediatamente
después del agregado de los reactivos, se volcó en el molde y se selló con butanol. Una
vez preparado el gel se colocó el mismo en la cámara electroforética refrigerada a 4˚C,
se impregnaron los electrodos de papel con H3PO4 1 M (electrodo positivo) y NaOH 1M
(electrodo negativo) y se colocaron encima del gel. Se colocaron los sembradores en el
gel y se sembraron 15 μL de las muestras y los patrones. Se corrió el gel con
condiciones iniciales de 15’ W= 6 W ( V=3000 V; Amp= >80 A y posteriormente a 120’
W=25 W. A la hora de corrida se sacaron los sembradores, y se secaron bien los
alambres de los electrodos. Una vez finalizada la corrida se removió el exceso de sales
con agua de chorro. El revelado de las ß-lactamasas se realizó cubriendo el gel de pI,
una vez finalizada la corrida, con un “sobregel” con el sistema yodo-almidón, que
inicialmente es azul. Para diferenciar entre las bandas producidas por actividades de
Betalactamasas de espectro ampliado (BLEA) o Betalactamasas de espectro extendido
(BLEE), se realizaron revelados diferenciales suplementando el sobregel de agar yodo-
almidón con los siguientes antibióticos como substratos: Penicilina (0,1 mg/mL) +
Cefaloridina (1mg/mL), que revela actividades de BLEA Y BLEE, y Ceftazidima
(1mg/mL) + Ceftriaxona (1mg/mL), que revela actividad de BLEE. La presencia de una
banda con actividad de ß-lactamasa quedó de manifiesto como una zona de
decoloración del sobregel al ser hidrolizado el antibiótico incorporado a él (Galas y col.,
2004).
Para determinar la capacidad de diseminación de los genes de resistencia se
efectuó conjugación de las cepas en estudio hacia una cepa de Salmonella sensible a
los antibióticos probados. El ensayo se llevó a cabo entre las cepas Donantes (CD)
resistentes a betalactámicos, lactosa positivo y negativas para la producción de
hidrógeno sulfurado, y se conjugaron con una cepa de Salmonella receptoras (CR)
sensible a todos los betalactámicos, lactosa negativo y productora de hidrógeno
sulfurado. Este proceso se llevó a cabo según el siguiente protocolo (modificado de
Lorian, 1996):

45
1. Aislamiento y purificación de cepas Donantes y Receptoras en placas de agar Mc
Conkey.
2. Se inoculó 4mL de caldo LB con la cepa receptora y donante respectivamente a
37 °C de 18-20h.
3. Se prepararon las siguientes mezclas en tubos separados:
a. Mezcla Test: 0,250 mL de la célula receptora, 0,250 mL de la cepa donante
en 2 mL de caldo LB
b. Control célula receptora: 0,500 mL de la célula receptora en 2 mL de caldo.
c. Control célula donante: 0,500 mL de la célula donante en 2 mL de caldo.
4. Se filtró la mezcla a través de un filtro estéril de 0,45µm. El filtro permite la
formación de pares estables. Se colocaron los filtros en placas de agar Mueller-
Hinton y se incubaron a 37 °C durante 18-20h.
5. Se colocó el filtro en 3 mL de PBS estéril a pH 7.2 y se resuspendió por agitación
en un vortex.
6. Se inocularon placas de agar SS conteniendo el agente selectivo con 100 µL de
diluciones 10-1 y 10-2 de la mezcla y controles. Las placas contenían Cefotaxima
a una concentración de 30 µg/mL de concentración (al cual la cepa donante es
resistente).
7. Se incubaron las placas a 37 °C durante 24-48 horas.

46
8. Se seleccionaron las colonias lactosa negativa con producción de hidrógeno
sulfurado (Salmonellas transconjugantes).
9. Se realizaron pruebas de identificación bioquímica y determinación del perfil de
resistencia de la cepa transconjugante.
Para determinar si un gen de resistencia se encuentra presente en un integrón se
determinó la presencia de las enzimas tipo Integrasa de clases 1 mediante PCR con
el iniciador Int1 (Reyes y col., 2003). Una vez que se determinó cuales cepas
presentaban integrones de clase 1, se procedió a amplificar por PCR la región
variable de éstos, y posteriormente se amplificaron los genes detectados
previamente en estas cepas empleando como templado únicamente la región
variable del integrón correspondiente. Posteriormente se efectuó un mapa genético
de la región variable mediante PCR con combinaciones de los iniciadores Int, 5’CS,
3’CS y los iniciadores de los genes detectados; el tamaño del producto de
amplificación obtenido permitió discernir la posición relativa del gen en estudio con
respecto a las regiones conservadas del integrón y por ende su posición relativa
dentro del mismo. La mezcla de reacción empleada fue: Vol. final: 50 μl; Buffer ADN
Taq polimerasa: 1X; Primers: 50 pmoles de c/u.; ADN Taq polimerasa: 1 U; dNTPs:
25 μM de cada uno; ADN molde: 5 μl; MgCl2: 1,5 mM. El programa empleado en el
termociclador contempló un paso de desnaturalización inicial de 5 min. a 94ºC, y la
amplificación se llevó a cabo por 35 ciclos, con pasos de desnaturalización (30 seg. a
94ºC), alineación (30 seg a 55 ºC para TEM, CTX-M, y PER y a 59 ºC para SHV) y
extensión (1 minuto 30 seg. a 72 ºC), y un paso final de extensión de 10 min. a 72ºC
(Galas y col., 2004). Los productos finales de amplificación fueron detectados en
geles de agarosa al 1,2 %, teñidos con bromuro de etidio 0,5 μg/mL y se visualizaron
bajo luz ultravioleta mediante el equipo Gel Doc 2000 (Bio-Rad), empleando un
marcador de peso molecular de 100 pares de bases.
Para determinar si existía una relación estadísticamente significativa entre la
producción de betalactamasas de espectro expandido y la resistencia a quinolonas,

47
así como con los diferentes genotipos de BLEE y los mecanismos de resistencia a
quinolonas hallados en las cepas se empleó del programa SPSS 12.0, usando como
herramienta estadística el método de chi-cuadrado. Mediante esta prueba no
paramétrica se midió la discrepancia entre una distribución observada y otra teórica,
indicando en qué medida las diferencias existentes entre ambas, se deben al azar en
el contraste de hipótesis (nos permite determinar si existe una relación entr dos
variables categóricas). Es necesario resaltar que esta prueba nos indica si existe o
no una relación entre las variables, pero no indica el grado o el tipo de relación; es
decir, no indica el porentaje de influencia de una variable sobre la otra o la variable
que causa la influencia. De esta forma se pudo contrastar si las frecuencias
observadas en cada una de las clases de las variables categóricas establecidas
varían de forma significativa de las frecuencias que se esperaría encontrar si la
muestra hubiese sido extraída de una población con una determinada distribución de
frecuencias.

48
RESULTADOS
Estudio fenotípico y de susceptibilidad. Se recolectaron 50 aislados de Enterobacterias provenientes de los centros
asistenciales Hospital Universitario de Caracas, Hospital J. M. de los Ríos, Hospital Dr.
Ricardo Baquero, Centro Policlínico Valencia, Laboratorio Cesar Sánchez Font,
Policlínica Santiago de León, Hospital Central Antonio María Pineda e Instituto Nacional
de Higiene “Rafael Rangel” durante el período Enero - Junio 2006, los cuales
presentaban susceptibilidad disminuída a cefalosporinas. Se les realizó confirmación
bioquímica de Género y Especie obteniéndose 19 (38%) aislados de E. cloacae, 16
(32%) aislados de E. aerogenes, 7 (14%) aislados de E. coli; 6 (12%) aislados de K.
pneumoniae y 2 (4%) aislados de K. oxytoca (Figura 5).
Figura 6. Distribución de aislados de Enterobacterias en estudio
De las 50 cepas de Enterobacterias con susceptibilidad disminuida a
cefalosporinas aisladas de distintas fuentes biológicas, sólo 20 fueron confirmadas
fenotípicamente como productoras de BLEE, cumpliendo realmente con los criterios de
selección establecidos para esta investigación. Aun cuando se realizó la confirmación
bioquímica de género y especie y detección de BLEE a las 50 cepas muestreadas, sólo
N=50
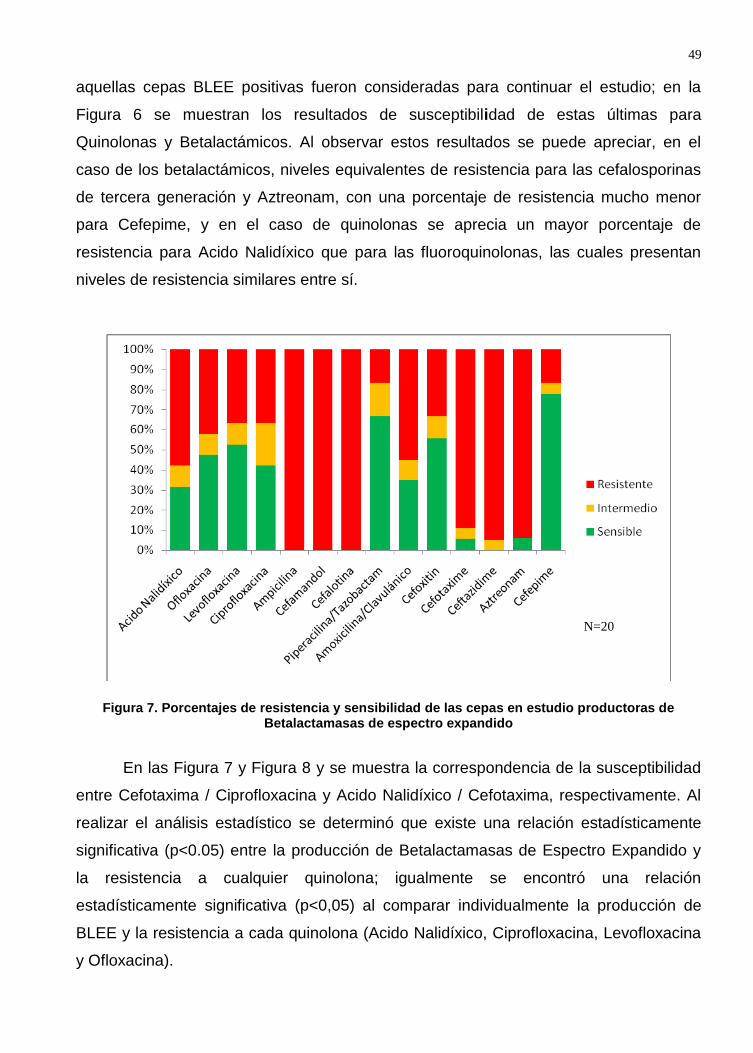
49
aquellas cepas BLEE positivas fueron consideradas para continuar el estudio; en la
Figura 6 se muestran los resultados de susceptibilidad de estas últimas para
Quinolonas y Betalactámicos. Al observar estos resultados se puede apreciar, en el
caso de los betalactámicos, niveles equivalentes de resistencia para las cefalosporinas
de tercera generación y Aztreonam, con una porcentaje de resistencia mucho menor
para Cefepime, y en el caso de quinolonas se aprecia un mayor porcentaje de
resistencia para Acido Nalidíxico que para las fluoroquinolonas, las cuales presentan
niveles de resistencia similares entre sí.
Figura 7. Porcentajes de resistencia y sensibilidad de las cepas en estudio productoras de Betalactamasas de espectro expandido
En las Figura 7 y Figura 8 y se muestra la correspondencia de la susceptibilidad
entre Cefotaxima / Ciprofloxacina y Acido Nalidíxico / Cefotaxima, respectivamente. Al
realizar el análisis estadístico se determinó que existe una relación estadísticamente
significativa (p<0.05) entre la producción de Betalactamasas de Espectro Expandido y
la resistencia a cualquier quinolona; igualmente se encontró una relación
estadísticamente significativa (p<0,05) al comparar individualmente la producción de
BLEE y la resistencia a cada quinolona (Acido Nalidíxico, Ciprofloxacina, Levofloxacina
y Ofloxacina).
N=20
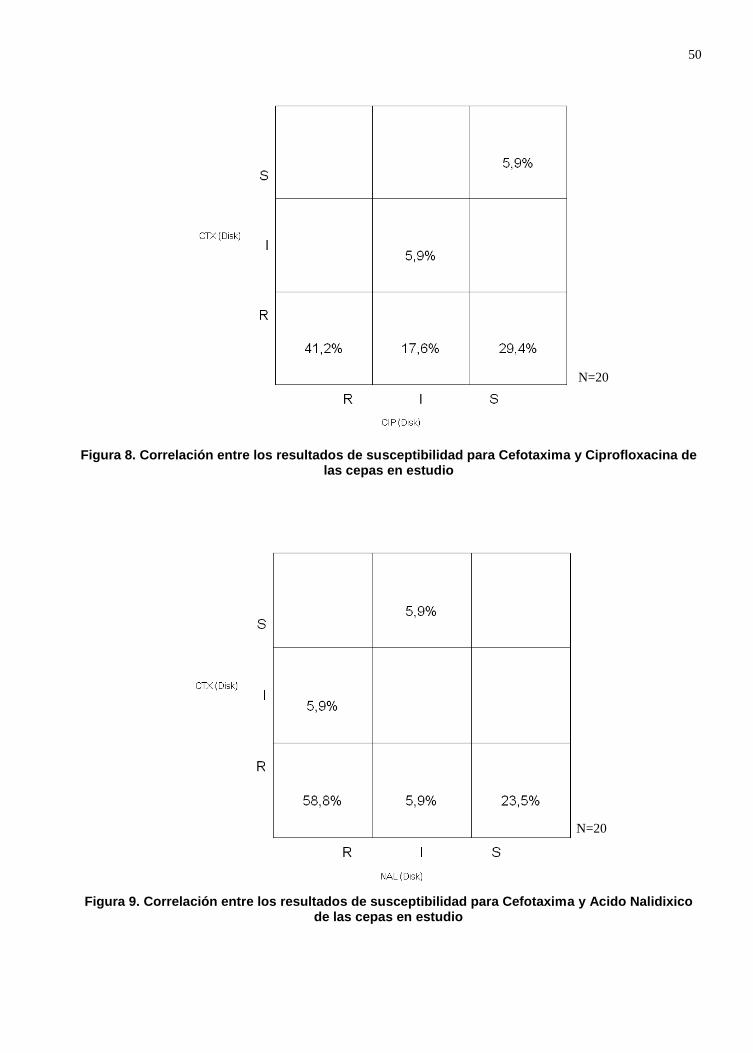
50
Figura 8. Correlación entre los resultados de susceptibilidad para Cefotaxima y Ciprofloxacina de las cepas en estudio
Figura 9. Correlación entre los resultados de susceptibilidad para Cefotaxima y Acido Nalidixico
de las cepas en estudio
N=20
N=20

51
En la Tabla 4 y la Figura 10 se pueden observar los perfiles de resistencia
individuales a betalactámicos de cada cepa BLEE positiva. Vale la pena destacar la
presencia de cepas con un típico perfil cefotaximasa, en las cuales se aprecia reducción
significativa tanto de los halos de Cefotaxima como de Cefepima, cepas con perfil
ceftazidimasa, donde hay una marcada reducción de Ceftazidima en comparación a
Cefotaxima y Cefepime, y cepas donde puede evidenciarse fenotípicamente la
presencia simultánea de varias betalactamasas, ya que se ven afectados de forma
similar los tres antibióticos.
Al observar la distribución poblacional de los halos para Cefotaxima y
Ceftazidima (Figura 9), se puede apreciar que en ambos casos, los valores obtenidos
en la mayoría de las cepas pertenecen a la categoría resistente, independientemente
de cual perfil de resistencia expresaron, y adicionalmente se observa una población
importante que presenta halos completamente cerrados (6 mm) para Ceftazidima.
Con respecto a las quinolonas, en la Tabla 5 se puede observar los perfiles de
resistencia a quinolonas y betalactámicos de las mismas cepas; en este caso se
pueden apreciar igualmente varios perfiles de resistencia marcados: alto nivel de
resistencia (halos de 6 mm para todas las quinolonas), nivel intermedio de resistencia
(halos disminuidos para todas las quinolonas), alto nivel de resistencia al Acido
nalidíxico con halos disminuidos para el resto de las quinolonas, y el perfil sensible
(halos amplios en todos los antibióticos).
Al analizar la distribución poblacional de Ciprofloxacina (Figura 9) se puede
observar claramente la presencia de tres poblaciones: un importante número de cepas
con halos completamente cerrados (6 mm) o valores de MIC muy elevados (≥32 µg/ml),
una población en la categoría intermedia (correspondería a los perfiles de nivel
intermedio de resistencia), y la población sensible; ejemplos de estos perfiles se pueden
observar en la figura 11.

52
Tabla 4. Perfil de Resistencia a Betalactámicos de cepas de Enterobacterias BLEE positivas seleccionadas para el estudio.
Cepa Género-
Especie
CTX
(mm)
CAZ
(mm)
FEP
(mm)
FOX
(mm)
230477 E. cloacae 6 6 23 6 229268 K. pneumoniae 6 17 13 26 230056 E. coli 6 11 14 13 227131 E. coli 16 6 20 22 226102 E. coli 6 20 12 23 225377 E. aerogenes 8 6 14 6 223274 E. cloacae 28 12 26 6 222515 K. pneumoniae 23 6 26 24 222308 K. pneumoniae 11 6 16 24 220664 K. pneumoniae 15 6 22 23 220624 K. pneumoniae 18 6 23 23 219712 K. oxytoca 14 14 26 15 220666 E. coli 10 9 27 17 220667 E cloacae 20 13 27 13 220663 E. cloacae 12 6 26 6 221297 E. coli 11 14 28 6 221302 E. coli 16 12 22 23 222211 K. oxytoca 22 6 28 28 222220 E. coli 13 6 20 23 222448 K. pneumoniae 18 6 24 22
CTX = Cefotaxima; CAZ = Ceftazidima; FEP = Cefepime; FOX = Cefoxitin
Tabla 5. Perfil de Resistencia a Quinolonas de cepas de Enterobacterias BLEE positivas seleccionadas para el estudio.
Cepa Género-Especie NA
(mm)
CIP
(mm)
CIP
(ug/ml)
OFL
(mm)
LEV
(mm)
230477 E. cloacae 26 32 0,023 27 30 229268 K. pneumoniae 6 6 >32 6 6 230056 E. coli 6 6 >32 6 9 227131 E. coli 6 6 >32 6 9 226102 E. coli 26 34 0,008 29 30 225377 E. aerogenes 10 18 1 12 16 223274 E. cloacae 16 25 0,19 23 25 222515 K. pneumoniae 6 16 1 14 17 222308 K. pneumoniae 6 6 4 6 6 220664 K. pneumoniae 6 6 >32 6 6 220624 K. pneumoniae 6 6 >32 6 6 219712 K. oxytoca 24 24 0,012 25 26 220666 E. coli 27 35 0,12 33 35 220667 E cloacae 16 25 0,19 21 22 220663 E. cloacae 26 34 0,008 29 30 221297 E. coli 6 19 0,75 16 19 221302 E. coli 27 35 0,006 30 35 222211 K. oxytoca 25 29 0,19 26 28 222220 E. coli 6 6 32 6 6 222448 K. pneumoniae 6 16 1 14 16
NA = Acido Nalidíxico; CIP = Ciprofloxacina; OFL = Ofloxacina; LEV = Levofloxacina
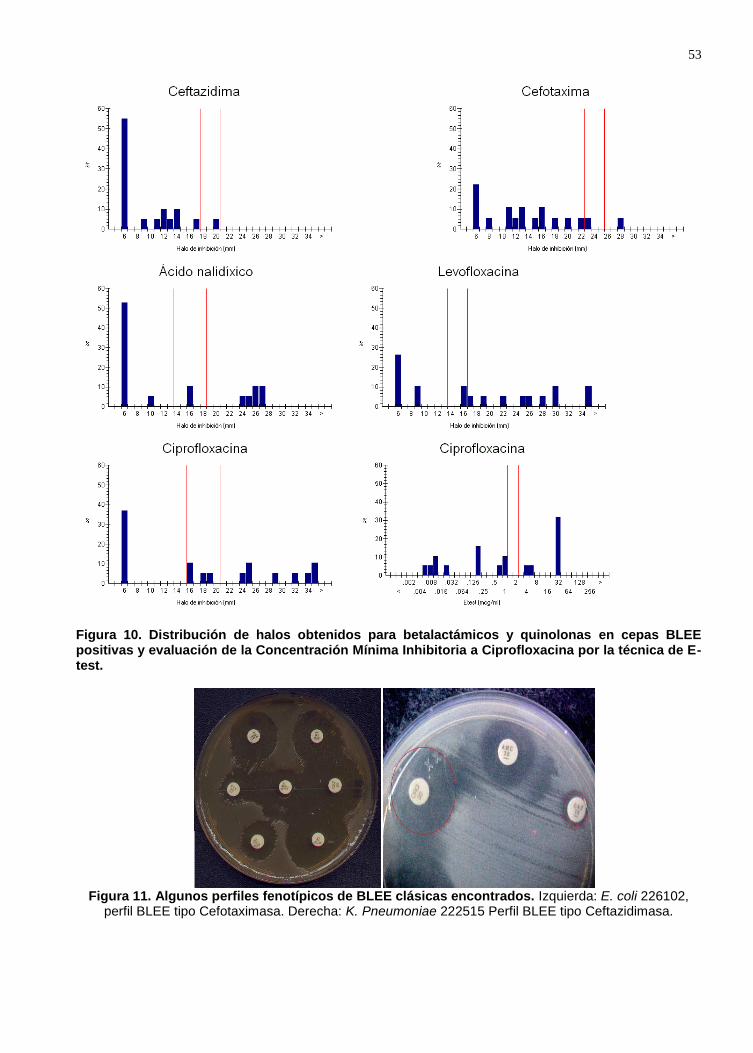
53
Figura 10. Distribución de halos obtenidos para betalactámicos y quinolonas en cepas BLEE positivas y evaluación de la Concentración Mínima Inhibitoria a Ciprofloxacina por la técnica de E-test.
Figura 11. Algunos perfiles fenotípicos de BLEE clásicas encontrados. Izquierda: E. coli 226102,
perfil BLEE tipo Cefotaximasa. Derecha: K. Pneumoniae 222515 Perfil BLEE tipo Ceftazidimasa.

54
Figura 12. Resultados de prueba de Concentración Inhibitoria Mínima a Ciprofloxacina por E-test. Izquierda: E. coli 226102. Cepa sensible (CIM = 0.008 µg/mL). Centro: E. aerogenes 225377. Nivel intermedio de resistencia a quinolonas (CIM = 1 µg/mL). Derecha: K. pneumoniae 226268. Alto nivel de resistencia a quinolonas (CIM ≥ 32 µg/mL)
Detección fenotípica de la producción de enzimas mediante el método de Hodge.
Simultáneamente se realizó la determinación de la naturaleza enzimática de la
resistencia a un antibiótico mediante la pruebas de Hodge para las 50 cepas, y se
compararon los resultados obtenidos con el método de difusión del disco para detección
y confirmación de BLEE propuesto por Jarlier y col., 1988.
Del total de cepas estudiadas, 20 (40%) dieron positivas a la detección fenotípica
de BLEE por el método de difusión del disco y el 60% negativas, mientras que por el
método Hodge para betalactámicos la positividad fue del 60% y la negatividad del 40%,
indicando la presencia de un 20% de cepas con betalactamasas no inhibibles por ácido
clavulánico. Los resultados positivos por el método Hodge dieron un 70,0% para
Cefotaxima y un 85,0% para Ceftazidima. Todas las cepas con resistencia a quinolonas
arrojaron un resultado negativo para el método de Hodge empleando Ciprofloxacina,
indicando la ausencia de enzimas que hidrolicen quinolonas. En la figura 12 se pueden
apreciar los resultados de esta prueba.

55
Figura 13. Prueba de Hodge de las cepas en estudio. Izquierda: Resultados positivos para Cefotaxima (CTX) y Ceftazidima (CAZ). Derecha: Resultados negativos para Ciprofloxacina.
De los 20 resultados positivos por el método de difusión del disco, 10 cepas
(50%) presentaron efecto de sinergia con ácido clavulánico con Cefotaxima; 5 con
Ceftazidima (25%) y 5 con ambos antibióticos (25%). En los resultados negativos del
método de difusión del disco, hubo un 66% de coincidencia con los resultados
negativos del método Hodge; esto nos confirma nuevamente que el mejor antibiótico
detector resultó ser la Cefotaxima, tal como se observa en la tabla 6. Se estableció que
hubo una asociación significativa (P < 0,05) entre los resultados obtenidos por ambos
métodos (Difusión del disco para detección de BLEE y Hodge).
Tabla 6. Resultados obtenidos al aplicar el método de Hodge comparado con el método de difusión del disco para detección de BLEE
RESULTADOS DE HODGE
RESULTADOS DE BLEE
DIFUSIÓN DEL DISCO
CEPAS N
NEGATIVO N
CTX N
CAZ N
CTX / CAZ N
POSITIVA 20 30 10 5 5 NEGATIVA 30 20 0 0 10
CTX = Cefotaxima; CAZ = Ceftazidima.
Determinación de Punto Isoeléctrico mediante ensayos de Isoelectroenfoque.
Los resultados del Punto Isoeléctrico de las cepas BLEE positivas se reflejan en
la Tabla 7, donde se puede observar que, de las veinte cepas que se les detectó β-
lactamasas de espectro expandido (BLEE) por los otros métodos fenotípicos, sólo 15
(75%) dieron resultado por isoelectroenfoque. Los valores de pI se determinaron por

56
comparación con extractos de β-lactamasas de pI conocidos provenientes de las mismas
cepas controles empleadas para la caracterización de genes de betalactamasas por
PCR, y dependiendo del sustrato que hidrolizaba la enzima (penicilina + cefaloridina del
lado derecho del gel o ceftazidima + ceftriaxona del lado izquierdo del mismo) se
clasificó la banda como BLEA o BLEE (Figura 13). Al observar los datos de pI de las
bandas BLEE se puede apreciar que los valores predominantes son 5.6 y 7.6,
correspondientes a BLEE tipo TEM y SHV, respectivamente; las bandas de pI 5.4
corresponderían a BLEE tipo PER-2 y las de 7.9 a CTX-M-2.
Tabla 7. Valores de pI obtenidos para las cepas en estudio.
Cepa Género-Especie pI
BLEA
pI
BLEE 230477 E. cloacae 5.4 229268 K. pneumoniae 7.6 5.4; 7.6; 7.9 230056 E. coli 5.4 227131 E. coli 5.4 5.4; 8.2 226102 E. coli 5.4 5.4; 7.9 225377 E. aerogenes 5.6; 7.9; 8.2 223274 E. cloacae 5.6 222515 K. pneumoniae 5.6 222308 K. pneumoniae 7.6 5.6; 7.6 220624 K. pneumoniae 7.6 5.6; 7.6 220663 E. cloacae 8.2 221302 E. coli 5.4 5.4; 8.2 222211 K. oxytoca 5.6; 8.2 222220 E. coli 5.4 5.4; 8.2 222448 K. pneumoniae 5.6; 7.6
Figura 14. Gel de isoelectroenfoque. Obsérvese en el lado izquierdo de la foto las bandas correspondientes a enzimas tipo BLEE y en la parte derecha las correspondientes a BLEA.

57
Los valores de pI obtenidos en este estudio sugieren la producción de β-
lactamasas tipo TEM, SHV, PER-2 y CTX-M-2. Estos resultados guardan similitud con
los obtenidos por García y colaboradores en el 2005, quienes realizaron la
caracterización molecular de aislamientos de Enterobacter cloacae multirresistentes
productores de β-lactamasas provenientes de pacientes de un hospital de tercer nivel de
Bogotá, encontrando que todos los aislamientos fueron productores de β-lactamasas con
puntos isoeléctricos que podrían corresponder a BLEE del tipo SHV y TEM, que son las
β-lactamasas más frecuentemente informadas en enterobacterias (García y col., 2005).
Ensayos de Conjugación
Se realizó ensayos de conjugación para determinar la capacidad de transferencia
de resistencia a betalactámicos a los 8 aislados positivos para la presencia de genes
BLEE y que mostraron la presencia de integrones, aumentando de esta manera la
probabilidad de su transferencia debido a la alta frecuencia con la cual se encuentran
los integrones dentro de plásmidos (Alvarez-Fernández y col., 2003).
Los resultados del presente estudio señalan que, entre los 8 aislados utilizados
como donantes, 4 aislados fueron capaces de transferir plásmidos (Tabla 8),
representando el 50% de los aislados estudiados. En la figura 15 se puede apreciar la
placa selectora empleada y la transferencia del fenotipo de resistencia a
betalactámicos.
Tabla 8. Ensayos de Conjugación en aislados de Enterobacterias BLEE positivas que presentaron integrones de clase 1
Donante Resultado de Conjugación
Cepa Donante
Cepa Receptora
229268 + K. pneumoniae Salmonella sp. INH 604834 230056 - E. coli Salmonella sp. INH 604834
227131 - E. coli Salmonella sp. INH 604834
226102 + E. coli Salmonella sp. INH 604834
225377 + E. aerogenes Salmonella sp. INH 604834
223274 - E. cloacae Salmonella sp. INH 604834
220624 + K. pneumoniae Salmonella sp. INH 604834
220667 - E. cloacae Salmonella sp. INH 604834

58
Figura 15. Ensayo de conjugación. Izquierda: Placa selectora con 30 µg/ml de Cefotaxima. Las
colonias con punto negro son las Salmonellas transconjugantes. Derecha: Antibiograma de cepa transconjugante. Obsérvese como se tranfirió la resistencia a Amoxicilina/Acido Clavulánico (AMC), Cefotaxima (CTX), Ceftazidima (CAZ) y Cefepime (FEP).
Estudio molecular La caracterización de los fenotipos de betalactamasas observados dio como
resultado la presencia de 11 cepas con genotipo SHV (5 de ellas presuntamente SHV
tipo BLEA), 11 cepas TEM (4 de ellas presuntamente TEM tipo BLEA), 6 cepas tipo
PER y 3 cepas tipo CTX-M del grupo 2; de éstas, 5 cepas presentaron más de un
genotipo BLEE simultáneamente (cepas 229268, 227131, 226102, 225377 y 221302).
En la Figura 15 se muestra el gel correspondiente a algunos de estos hallazgos.
Figura 16. Amplificación de genes TEM, CTX-M, SHV y gyrA de las cepas en estudio. 1: Control positivo TEM; 2: 227131; 3: Control positivo CTX-M; 4: 229268; 5: Control positivo SHV; 6: 227131; 7: Control positivo gyrA; 8: 227131; 9: 226102; 10: 220624; 11: Control negativo; 12: DNA ladder 100 pb

59
Al evaluar cada uno de los mecanismos enzimáticos detectados en relación al
género y especie de los aislados (Tabla 9), se observa que la betalactamasa tipo SHV
se presenta con mayor frecuencia en Klebsiella, mientras que la betalactamasa tipo
TEM se encontró en la misma proporción entre Klebsiella y E. coli; sin embargo, al
analizar estos datos, hay que tomar en cuenta la presencia de BLEAs tipo SHV-1
asociadas con Klebsiella pneumoniae y TEM-1 asociadas con E. coli. Al realizar el
análisis estadístico entre la resistencia a quinolonas y la presencia de cada uno de los
genes BLEE por separado (Quinolona R / TEM; Quinolona R / SHV; Quinolona R / CTX-
M-2; Quinolona R / PER-2) se determinó que no existe una relación estadísticamente
significativa (p>0,05).
Tabla 9. Resultados de amplificación de genes de betalactamasas.
Cepa Género-Especie TEM SHV CTX-M-2 PER 230477 E. cloacae - - - + 229268 K. pneumoniae - + + + 230056 E. coli - - - + 227131 E. coli + + - + 226102 E. coli + - + + 225377 E. aerogenes + + + - 223274 E. cloacae + - - - 222515 K. pneumoniae + - - - 222308 K. pneumoniae + + - - 220664 K. pneumoniae - + - - 220624 K. pneumoniae + + - - 219712 K. oxytoca - - - - 220666 E. coli - - - - 220667 E cloacae - - - - 220663 E. cloacae - + - - 221297 E. coli - - - - 221302 E. coli + + - + 222211 K. oxytoca + + - - 222220 E. coli + + - - 222448 K. pneumoniae + + - -
Al analizar la Tabla 10 se puede observar que no se encontraron qnr de los tipos
A y S, y se encontraron 4 cepas con genotipo QnrB y 3 con Aac(6’)ibcr; una de las
cepas presentó simultáneamente ambos genes (220667). Al realizar el análisis
estadístico se determinó que existe una relación estadísticamente significativa (p<0,05)
entre la producción de Betalactamasas de Espectro Expandido y la presencia de

60
mecanismos plasmídicos de resistencia a quinolonas; sin embargo, no se encontró una
relación estadísticamente significativa (p>0,05) entre la producción de BLEE y la
presencia de cada uno de los mecanismos plasmídicos encontrados por separado
(BLEE / Aac(6’)ibcr o BLEE / QnrB).
Tabla 10. Resultados de amplificación de genes codificantes para resistencia plasmídica a
quinolonas.
Cepa Género-
Especie QnrA QnrB QnrS Aac(6’)ibcr
230477 E. cloacae - - - - 229268 K. pneumoniae - - - - 230056 E. coli - - - + 227131 E. coli - - - - 226102 E. coli - - - - 225377 E. aerogenes - + - - 223274 E. cloacae - - - - 222515 K. pneumoniae - - - + 222308 K. pneumoniae - + - - 220664 K. pneumoniae - + - - 220624 K. pneumoniae - - - - 219712 K. oxytoca - - - + 220666 E. coli - - - - 220667 E cloacae - + - + 220663 E. cloacae - - - - 221297 E. coli - - - - 221302 E. coli - - - - 222211 K. oxytoca - - - - 222220 E. coli - - - - 222448 K. pneumoniae - - - -
Estudio de integrones De las 20 cepas analizadas, 8 mostraron la presencia de integrones clase 1
(Tabla 11). La mayoría de los integrones clase 1 se encontrarion en E. coli, y en menor
número en K. pneumoniae. En la Figura 16 se puede observar el gel donde se
evidencia la amplificación de las integrasas clase 1, el cual fue el parámetro empleado
para la detección de esta clase de integrones.

61
Tabla 11. Resultados de amplificación por PCR de genes de integrasa de clase 1.
Cepa Género-Especie Integron Clase 1 230477 E. cloacae - 229268 K. pneumoniae + 230056 E. coli + 227131 E. coli + 226102 E. coli + 225377 E. aerogenes + 223274 E. cloacae + 222515 K. pneumoniae - 222308 K. pneumoniae - 220664 K. pneumoniae - 220624 K. pneumoniae + 219712 K. oxytoca - 220666 E. coli - 220667 E cloacae + 220663 E. cloacae - 221297 E. coli - 221302 E. coli - 222211 K. oxytoca - 222220 E. coli - 222448 K. pneumoniae -
Figura 17. Amplificación de Integrasa clase 1 de las cepas en estudio. 1: DNA ladder 100 pb; 2: 230477; 3: 229268; 4: 230056; 5: 227131; 6: 226102; 7: 225377; 8: 223274; 9:220624; 10: 222515; 11: 220667; 12: Control positivo; 13: Control negativo.
Una vez que se determinó cuales cepas presentaban integrones de clase 1, se
procedió a amplificar por PCR la región variable de éstos (cepas 229268, 230056,
227131, 226102, 225377 y 223274), y posteriormente se amplificaron los genes
detectados previamente en estas cepas empleando como templado únicamente la
región variable del integrón correspondiente. Los resultados de ambas amplificaciones
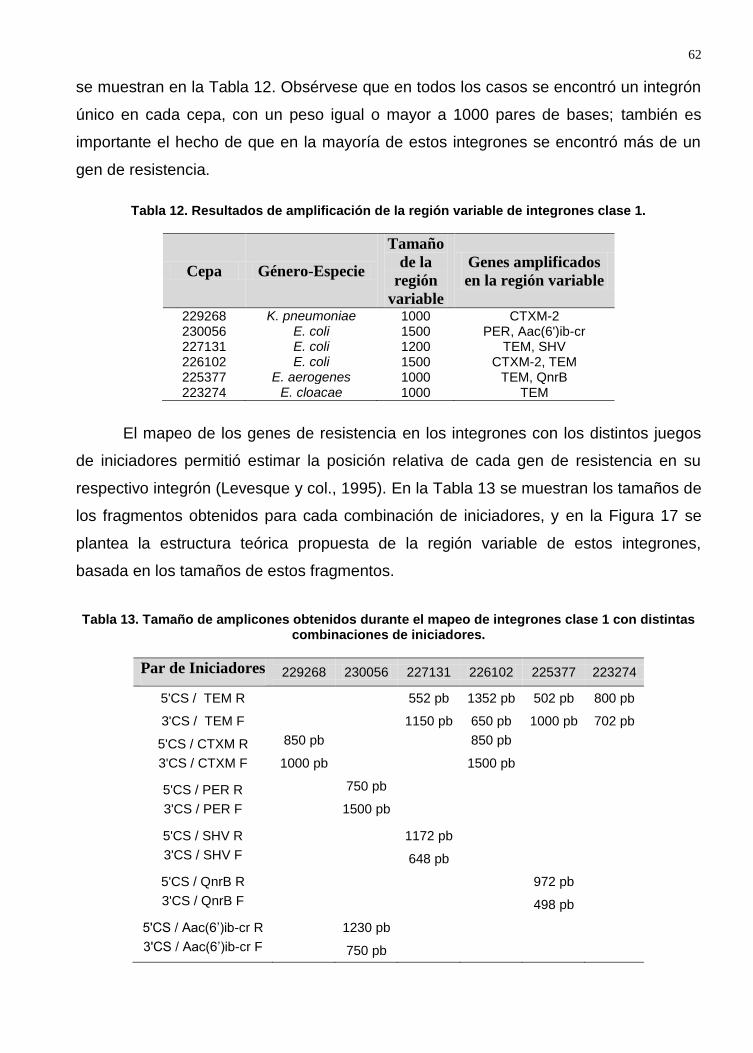
62
se muestran en la Tabla 12. Obsérvese que en todos los casos se encontró un integrón
único en cada cepa, con un peso igual o mayor a 1000 pares de bases; también es
importante el hecho de que en la mayoría de estos integrones se encontró más de un
gen de resistencia.
Tabla 12. Resultados de amplificación de la región variable de integrones clase 1.
Cepa Género-Especie
Tamaño
de la
región
variable
Genes amplificados
en la región variable
229268 K. pneumoniae 1000 CTXM-2 230056 E. coli 1500 PER, Aac(6')ib-cr 227131 E. coli 1200 TEM, SHV 226102 E. coli 1500 CTXM-2, TEM 225377 E. aerogenes 1000 TEM, QnrB 223274 E. cloacae 1000 TEM
El mapeo de los genes de resistencia en los integrones con los distintos juegos
de iniciadores permitió estimar la posición relativa de cada gen de resistencia en su
respectivo integrón (Levesque y col., 1995). En la Tabla 13 se muestran los tamaños de
los fragmentos obtenidos para cada combinación de iniciadores, y en la Figura 17 se
plantea la estructura teórica propuesta de la región variable de estos integrones,
basada en los tamaños de estos fragmentos.
Tabla 13. Tamaño de amplicones obtenidos durante el mapeo de integrones clase 1 con distintas combinaciones de iniciadores.
Par de Iniciadores 229268 230056 227131 226102 225377 223274
5'CS / TEM R 552 pb 1352 pb 502 pb 800 pb
3'CS / TEM F 1150 pb 650 pb 1000 pb 702 pb
5'CS / CTXM R 850 pb 850 pb
3'CS / CTXM F 1000 pb 1500 pb
5'CS / PER R 750 pb
3'CS / PER F 1500 pb
5'CS / SHV R 1172 pb
3'CS / SHV F 648 pb
5'CS / QnrB R 972 pb
3'CS / QnrB F 498 pb
5'CS / Aac(6’)ib-cr R 1230 pb
3'CS / Aac(6’)ib-cr F 750 pb

63
Figura 18. Estructura teórica propuesta de la región variable de integrones encontrados en las cepas en estudio.
Secuenciación
Se secuenciaron los productos de amplificación de gyrA de aquellas cepas que
presentaron resistencia a quinolonas, se compararon las secuencias obtenidas con
secuencias de cepas con mutaciones conocidas en gyrA (Números de acceso
Genebank EU430280 a EU430289) y con la de una cepa susceptible patrón K.
pneumoniae ATCC 13883 (Número de acceso Genebank DQ673325) empleando el
programa BLAST de los Institutos Nacionales de Salud de los Estados Unidos, y se
dedujeron las secuencias de aminoácidos de las regiones mutadas (Fu y col.2008). En
la Tabla 14 se presentan las variaciones de aminoácidos deducidas respecto a la
secuencia tipo, encontrándose en la mayoría de las cepas mutaciones codificantes para
el aminoácido 83, observándose que el 15% de las cepas BLEE presentaron un cambio
de serina a leucina y el 35% de serina a isoleucina. Al realizar el análisis estadístico se
determinó que existe una relación estadísticamente significativa (p<0,05) entre la
producción de Betalactamasas de Espectro Expandido y la presencia de mecanismos
cromosomales de resistencia a quinolonas.
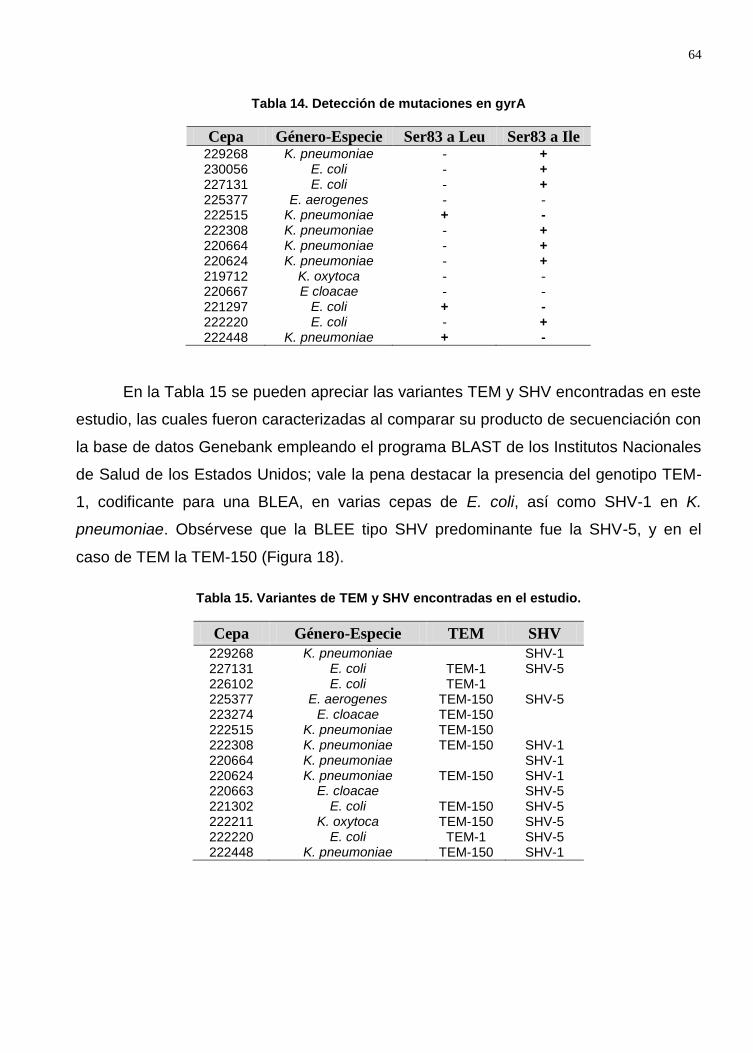
64
Tabla 14. Detección de mutaciones en gyrA
Cepa Género-Especie Ser83 a Leu Ser83 a Ile 229268 K. pneumoniae - + 230056 E. coli - + 227131 E. coli - + 225377 E. aerogenes - -
222515 K. pneumoniae + - 222308 K. pneumoniae - + 220664 K. pneumoniae - + 220624 K. pneumoniae - + 219712 K. oxytoca - -
220667 E cloacae - -
221297 E. coli + - 222220 E. coli - + 222448 K. pneumoniae + -
En la Tabla 15 se pueden apreciar las variantes TEM y SHV encontradas en este
estudio, las cuales fueron caracterizadas al comparar su producto de secuenciación con
la base de datos Genebank empleando el programa BLAST de los Institutos Nacionales
de Salud de los Estados Unidos; vale la pena destacar la presencia del genotipo TEM-
1, codificante para una BLEA, en varias cepas de E. coli, así como SHV-1 en K.
pneumoniae. Obsérvese que la BLEE tipo SHV predominante fue la SHV-5, y en el
caso de TEM la TEM-150 (Figura 18).
Tabla 15. Variantes de TEM y SHV encontradas en el estudio.
Cepa Género-Especie TEM SHV
229268 K. pneumoniae SHV-1 227131 E. coli TEM-1 SHV-5 226102 E. coli TEM-1 225377 E. aerogenes TEM-150 SHV-5 223274 E. cloacae TEM-150 222515 K. pneumoniae TEM-150 222308 K. pneumoniae TEM-150 SHV-1 220664 K. pneumoniae SHV-1 220624 K. pneumoniae TEM-150 SHV-1 220663 E. cloacae SHV-5 221302 E. coli TEM-150 SHV-5 222211 K. oxytoca TEM-150 SHV-5 222220 E. coli TEM-1 SHV-5 222448 K. pneumoniae TEM-150 SHV-1
65
Figura 19. Electroferograma de TEM-150

66
DISCUSIÓN
Los antibióticos betalactámicos son las drogas más usadas para el tratamiento
de infecciones bacterianas tanto a nivel de la comunidad como hospitalario (Walsh,
2003). El amplio espectro, la baja toxicidad y la actividad fuertemente bactericida sobre
la mayoría de los microorganismos son algunas de las causas de su amplia utilización;
sin embargo, prácticamente todos los miembros de la familia Enterobacteriaceae se
caracterizan por poseer mecanismos enzimáticos naturales de resistencia a los
antibióticos betalactámicos, que les confieren a las enterobacterias fenotipos
particulares que en muchos casos son útiles como ayuda de identificación, y
adicionalmente, como resultado de la captación de los elementos de ADN móviles
(plásmidos, transposones, etc.) o a través de mutaciones cromosómicas, se han
presentado microorganismos multirresistentes.a esta familia (Pfaller y col., 2000). Con
respecto a las Quinolonas, éstas han sido usadas desde hace años para el tratamiento
de infecciones bacterianas, teniendo una utilidad clínica muy extensa, lo que se basa en
su efecto bactericida, su amplio espectro de actividad, buena absorción por vía
digestiva, amplia distribución en los tejidos, prolongada vida media y prolongado efecto
postantibiótico, de lo que resultan planes terapéuticos eficaces y cómodos (Errecalde,
2004); sin embargo, al igual que con los betalactámicos, en los últimos años se ha visto
una emergencia importante de cepas resistentes a estos antibióticos (Walsh, 2003), lo
que se traduce en fallo terapéutico.
Al realizar la identificación de las cepas con susceptibilidad disminuida a
cefalosporinas, llama la atención que las dos especies encontradas con mayor
frecuencia fueron Enterobacter cloacae y Enterobacter aerogenes, en vista de que, aun
cuando este género puede presentar Betalactamasas de Espectro Expandido,
tradicionalmente es Escherichia coli la descrita como la que presenta BLEE con mayor
frecuencia (Diestra y col., 2009). Sin embargo, al realizar la detección de BLEE a las
cepas que conformaron la muestra, se pudo apreciar que la mayoría (aun cuando no
todas) las cepas correspondientes al género Enterobacter fueron BLEE negativas, por

67
lo cual se podría explicar la disminución de la susceptibilidad a las cefalosporinas en
estas cepas por otro tipo de betalactamasas, como por ejemplo AmpC derreprimido o
cefalosporinasas no inhibibles por ácido clavulánico (Bush y Jacoby, 2010). Una vez
excluidas aquellas cepas BLEE negativas, se observó que Escherichia coli fue la
principal representante en el estudio, lo cual es compatible con la alta frecuencia con la
que se encuentran tradicionalmente este tipo de genes de resistencia en esta especie
(Diestra y col., 2009), y en segundo lugar se encuentra Klebsiella pneumoniae como
productor de BLEE, la cual también es considerada como una de las principales
productoras de este tipo de enzimas (Perozo y Castellano, 2009), hasta el punto que
hasta el año 2004 el manual de la CLSI recomendaba realizar el tamizaje de BLEE solo
para estos dos géneros de bacterias.
Al observar los resultados de las pruebas de susceptibilidad de las cepas BLEE
positivas, se puede apreciar un porcentaje alto (entre el 40 y el 60%, según el
antibiótico) de resistencia a quinolonas, en virtud de que las BLEE no afectan a las
quinolonas, y el mecanismo de resistencia de estas últimas es principalmente
cromosomal. La resistencia de las Enterobacterias a la Ciprofloxacina ha aumentado
aproximadamente en un 17% entre los años 2000 y 2008 según los datos
proporcionados por el Programa Venezolano de Vigilancia de la Resistencia Bacteriana
a los Antimicrobianos (PROVENRA). En este Estudio se observó una alta tasa de cepas
resistentes a las Quinolonas, siendo el Ácido Nalidíxico el representante que mayor
resistencia presentó en las pruebas de susceptibilidad. El análisis estadístico verificó la
presencia de una relación estadísticamente significativa (p<0.05) entre la producción de
BLEE y la resistencia a cualquier quinolona, así como entre la producción de BLEE y la
resistencia a cada quinolona individual, lo cual demuestra que debe existir alguna
asociación a nivel molecular entre los mecanismos que confieren resistencia a ambas
familias de antibióticos. Otro punto importante respecto a la susceptibilidad a las
quinolonas es la presencia de varios perfiles de resistencia varios perfiles de resistencia
marcados con poblaciones distintivas respecto a los halos de susceptibilidad y valores
de CIM para Ciprofloxacina, lo cual evidencia que la resistencia a esta familia de
antibióticos esta determinada por diferentes mecanismos moleculares en estas cepas.

68
En lo que respecta a los niveles de resistencia a los betalactámicos, se
encuentran niveles equivalentes de resistencia para las cefalosporinas de tercera
generación y Aztreonam, y niveles de resistencia menores para Cefepime. Este
hallazgo a primera vista pareciera demostrar la presencia de un genotipo de resistencia
que afectara de forma homogénea a Cefotaxima, Ceftazidima y Aztreonam,
hidrolizando de forma pobre a Cefepime; sin embargo, al analizar los perfiles de forma
individual por cepa, se encontraron perfiles tipo Ceftazidimasa, perfiles tipo
Cefotaximasa y perfiles de alto nivel de hidrólisis para ambas cefalosporinas. La
discrepancia entre el nivel de resistencia encontrado a Cefepime y a las otras
cefalosporinas podría ser explicado en parte por la baja eficiencia de ciertas enzimas
BLEE (como por ejemplo algunas variantes SHV y TEM) para hidrolizar este antibiótico
(Biedenbach y col., 1999), y en parte por el cambio en los lineamientos de reporte de
Betalactámicos para las cepas BLEE efectuado por la CLSI en el año 2010, ya que que
aun cuando varían los puntos de corte para Cefotaxime, Ceftazidime y Aztreonam,
mantienen los mismos puntos para el resto de los Betalactámicos (entre ellos Cefepime)
y recomiendan no hacer cambios de interpretación (CLSI M100-S20, 2010), lo cual
resulta en discrepancias en el reporte de betalactámicos como Cefepime en cepas
BLEE positivas.
La gran variabilidad observada en los perfiles fenotípicos obtenidos en el trabajo
puede ser debida a que la afinidad de hidrólisis por los antibióticos es diferente para
cada enzima betalactamasa, inclusive pueden observarse variaciones en el perfil
fenotípico dentro de subvariedades o subtipos de la misma enzima; esto se debe
principalmente a que microorganismos no relacionados genotípicamente pueden
producir la misma BLEE mediante transferencia plasmídica, y la misma BLEE puede ser
mediada por plásmidos distintos (Garza-Ramos y col., 2007).
Con respecto a la prueba de Hodge, se observa que todas las cepas
determinadas como BLEE positivas presentaron un resultado positivo empleando una
cefalosporina de tercera generación, y algunas de las cepas BLEE negativas (excluidas
del estudio) también presentaron una reacción positiva; esto es debido a que la prueba
de Hodge sólo permite determinar la presencia de una enzima que hidrolice el

69
antibiótico que se está evaluando, pero no es capaz de discriminar la naturaleza de esa
enzima (BLEE, AmpC u otro tipo de enzima). También es importante destacar que el
resultado negativo de la prueba de Hodge para Ciprofloxacina en todas las cepas es
debido a que las cepas no presentaron ningún mecanismo que hidrolizara esta familia
de antibióticos, sino que le conferían resistencia por otros mecanimos (Sandra y col.,
2009). Tradicionalmente la resistencia a las Quinolonas era debida a mutaciones en los
genes que codifican la ADN girasa y la Topoisomerasa IV dando lugar a las QRDR (del
inglés “Quinolone Resistance-Determining Region (Hata y col., 2005). Recientemente,
se han descrito mecanismos de resistencia plasmídicos a las Quinolonas (Wang y col.,
2004) que involucran la síntesis de proteínas protectoras de las dianas y enzimas
modificadoras de Quinolonas.
Respecto al Isoelectroenfoque (IEF), esta técnica permite conocer el punto
isoelectrico (pI) de las proteínas y fue un método de gran utilidad cuando existían pocas
BLEE descritas. En la actualidad, se utiliza poco debido a la descripción de diferentes
BLEE que comparten idéntico pI. Sin embargo, es muy útil si se combina con el análisis
del fenotipo de sensibilidad ya que puede orientar el tipo de BLEE para un análisis
posterior del perfil de substrato o estudio molecular. Las BLEE de tipo TEM descritas
hasta el momento tienen valores de pI que oscilan entre 5,4 y 6,4, las de tipo SHV entre
7,0 y 8,2 y las CTX-M entre 7,6 y 9,0. El perfil de sustrato es imprescindible para la
caracterización final de las BLEE (Oliver y Cantón, 2008).
Al observar los pI de las bandas BLEE obtenidas se puede apreciar que los
valores predominantes son 5.6 y 7.6, sugerentes de BLEE tipo TEM y SHV,
respectivamente; las bandas de pI 5.4 sugieren a BLEE tipo PER-2 y las de 7.9 a CTX-
M-2. Tomando en cuenta los resultados obtenidos en esta investigación, se puede
observar que la detección del Punto Isoeléctrico es una técnica sensible más no
específica, ya que muchas enzimas pueden tener el mismo punto isoeléctrico y esto
dificulta su clasificación y ubicación. Por lo tanto esta es una técnica que se puede
emplear como auxiliar o prueba piloto para tener una idea previa de cuales pueden ser
las enzimas que están presentes y de esta manera escoger los iniciadores que van a
ser usados para realizar la detección molecular. También es importante resaltar que la

70
detección del pI permite una comparación visual de los patrones de bandas de
diferentes enzimas, y que al ser una comparación visual, la lectura de las bandas puede
llevar implícito la subjetividad del observador.
Respecto a los ensayos de conjugación, se seleccionaron aquellas cepas
positivas para la presencia de genes BLEE y que mostraron la presencia de integrones,
aumentando de esta manera la probabilidad de su transferencia debido a la alta
frecuencia con la cual se encuentran los integrones dentro de plásmidos (Alvarez-
Fernández y col., 2003), sin embargo, se obtuvo transferencia del fenotipo de
resistencia sólo en la mitad de las cepas ensayadas; esto puede ser debido a
diferencias en las características de los plásmidos en los cuales se sospecha se
presentaban estos integrones (por ejemplo, plásmidos no conjugativos) o que
simplemente en esas cepas los integrones no se encontraran en plásmidos (Garza-
Ramos y col., 2007). Por otro lado, en las cepas en las que se obtuvo transferencia del
fenotipo de resistencia por conjugación se demostró la presencia de los genes
codificantes para BLEE en elementos genéticos transferibles.
En los ensayos moleculares se observa mediante la amplificación por PCR la
presencia de los genotipos SHV, TEM, PER y CTX-M del grupo 2, e incluso la presencia
de varios genotipos en la misma cepa; sin embargo, en el caso de SHV y TEM, hay que
tomar en cuenta la presencia de BLEAs en las cepas de K. pneumoniae y E. coli,
respectivamente. Para poder discriminar en estos casos las BLEA SHV y TEM de las
BLEE, deben interpretarse los resultados de IEF y secuenciación de dichas cepas. Al
realizar el análisis estadístico entre la resistencia a quinolonas y la presencia de cada
uno de los genes BLEE por separado (Quinolona R / TEM; Quinolona R / SHV;
Quinolona R / CTX-M-2; Quinolona R / PER-2) se determinó que no existe una relación
estadísticamente significativa (p>0,05).
En lo que respecta a la determinación de genes plasmídicos de resistencia a
quinolonas, se logró observar no solo la presencia individual del gen qnrB y aac(6’)Ib-cr
en el 20% de las cepas, sino que también se obtuvieron aislados con 2 mecanismos
plasmídicos (cepa 220667); además, se pudo observar que el género en el que se

71
presentaron con mayor frecuencia estos mecanismos fue Klebsiella sp (20%). Estudios
realizados en China indicaron, al igual que la presente investigación, que la prevalencia
de proteínas Qnr es más alta en aislados de K. pneumoniae que en otras
enterobacterias (Jiang y col., 2008), y se ha descrito que estos genes se transmiten de
forma conjunta, en la mayoría de los casos, con genes de resistencia codificantes para
BLEE (Pitout y col., 2008; Rodríguez-Martínez y col., 2008; Bouchakour y col., 2010). A
diferencia de investigaciones realizadas en Brasil, en donde solo se logró encontrar 6
aislados con determinantes tipo qnrB (Minarini y col., 2008) en Venezuela se comprobó
adicionalmente la presencia del determinante aac(6’)Ib-cr; pero al igual que el
mencionado estudio, no se lograron encontrar aislados que presentaran qnrA, a
diferencia de países como Canadá, China y Francia, en donde la mayor prevalencia la
tiene qnrA (Nordmann y Poirel, 2005). Cabe destacar que, de todas la aac (6’) descritas
la aac (6’)-Ib es la más prevalente entre los diversos microorganismos Gram negativos.
El gen de la aac (6’)-Ib se ha detectado en transposones e integrones. Es razonable
pensar que la ubicación del gen que codifica para la aac (6’)-Ib en elementos
genéticos móviles ha facilitado su rápida difusión entre una amplia gama de
microorganismos (Vakulenko y Mobashery, 2003), y a finales del año 2005 se publicó la
aac (6’)-Ib-cr, capaz de afectar la actividad de ciprofloxacina mediante la N-acetilación
del nitrógeno del grupo piperazinil. El gen de esta enzima se ha localizado en
plásmidos al igual que qnrA, y confiere bajo nivel de resistencia a quinolonas
(Rodríguez, 2006).
Al realizar el análisis estadístico se determinó que existe una relación
estadísticamente significativa (p<0,05) entre la producción de Betalactamasas de
Espectro Expandido y la presencia de mecanismos plasmídicos de resistencia a
quinolonas; sin embargo, no se encontró una relación estadísticamente significativa
(p>0,05) entre la producción de BLEE y la presencia de cada uno de los mecanismos
plasmídicos de resistencia a quinolonas encontrados por separado. Esto puede ser
debido a la presencia habitual de cassettes de resistencia plasmídica a quinolonas en
los mismos integrones y/o plásmidos que presentan genes codificantes para BLEE ya
que le proporciona una ventaja adicional para la supervivencia de la bacteria en
ambientes con alta concentración de antibióticos como lo son los pacientes

72
hospitalizados, pero sin embargo sólo pareciera estar asociada la presencia de ambos
tipos de resistencia transferible pero sin existir asociación con un genotipo específico.
Al estudiar los integrones, la amplificación del gen de integrasa clase 1 permitió
demostrar la presencia de estos elementos en 8 de las 20 cepas (40%), lo cual
representa un porcentaje importante. La amplificación de la región variable de los
integrones y la subsecuente amplificación de los genes de resistencia previamente
detectados en estas cepas empleando como templado únicamente la región variable
del integrón correspondiente permitió, por un lado, determinar en todos los casos la
presencia de un integrón único en cada cepa, con regiones variables con pesos de
1000, 1200 o 1500 pb, lo cual es consistente con reportes previos donde demuestran
que las regiones variables de integrones clase 1 encontrados con mayor frecuencia en
enterobacterias presentan pesos de 800, 1000 y 500 pares de bases (Fernández y col.,
2003), y por otro lado demostró que, en aquellas cepas que presentaron estos
integrones, los genes codificantes para BLEE y/o resistencia plasmídica a quinolonas
detectados se encontraban en estos elementos transferibles.
El mapeo de los genes de resistencia en los integrones con los distintos juegos
de iniciadores permitió estimar la posición relativa de cada gen de resistencia en su
respectivo integrón, y en base al tamaño de los amplicones obtenidos se propuso la
estructura teórica de la región variable de los mismos (Levesque y col., 1995). Un
hallazgo importante es que se propone, en algunos casos, la presencia de múltiples
cassettes de resistencia en el mismo integrón, existiendo algunos casos donde
coexistirían un gen BLEE y uno de resistencia plasmídica a quinolonas, lo cual
explicaría la relación estadística encontrada entre estos mecanismos de resistencia.
Los resultados de secuenciación de los productos de amplificación de SHV y
TEM permitió determinar la presencia del genotipo TEM-1, codificante para una BLEA,
en varias cepas de E. coli, así como SHV-1 en K. pneumoniae, verificando así los
resultados obtenidos por IEF. Con respecto a los genotipos BLEE encontrados, la
BLEE tipo SHV predominante fue la SHV-5, y en el caso de TEM la TEM-150. Los
resultados de la secuenciación de los productos de amplificación de gyrA de aquellas

73
cepas que presentaron resistencia a quinolonas, se compararon con las secuencias
patrones conocidas y se dedujeron las secuencias de aminoácidos de las regiones
mutadas, encontrándose en la mayoría de las cepas mutaciones codificantes para el
aminoácidos 83, el cual (serina) fue sustituído en algunos casos por leucina y otros por
isoleucina; cabe destacar que la CIM para Ciprofloxacina aumentó más en las cepas
donde la serina fue sustituída por isoleucina que en las que se sustituyó por leucina, lo
cual coincide con lo determinado por otros autores (Morgan-Linnell y Zechiedrich, 2007;
Fu y col., 2008). Al realizar el análisis estadístico se determinó que existe una relación
estadísticamente significativa (p<0.05) entre la producción de Betalactamasas de
Espectro Expandido y la presencia de mecanismos cromosomales de resistencia a
quinolonas, tal como se ha descrito previamente (Lagacé-Wiens y col., 2007). Esta
asociación podría ser explicada por el desarrollo de mutaciones que confieren alto nivel
de resistencia a quinolonas bajo presión selectiva debido al incremento de la
concentración de prevención de mutantes por algún elemento presente en el plásmido
en el cual se encuentra el gen codificante para la BLEE, o por otro lado pudiera deberse
a que las cepas con mutaciones que confieren alto nivel de resistencia a quinolonas
presenten alguna característica que permita captar de forma más eficiente los
plásmidos con BLEE bajo presión selectiva con respecto a otras células bacterianas.

74
CONCLUSIONES
- Escherichia coli fue la principal representante en el estudio, lo cual es compatible
con la alta frecuencia con la que se encuentran tradicionalmente este tipo de
genes de resistencia en esta especie, y en segundo lugar se encuentra Klebsiella
pneumoniae.
- Se puede apreciar un porcentaje alto de resistencia a quinolonas.
- El análisis estadístico verificó la presencia de una relación estadísticamente
significativa (p<0,05) entre la producción de BLEE y la resistencia a cualquier
quinolona, así como entre la producción de BLEE y la resistencia a cada
quinolona individual, lo cual demuestra que debe existir alguna asociación a nivel
molecular entre los mecanismos que confieren resistencia a ambas familias de
antibióticos.
- Se encontraron niveles equivalentes de resistencia para las cefalosporinas de
tercera generación y Aztreonam, y niveles de resistencia menores para
Cefepima, discrepancia que podría ser explicada por la diferencia en la eficiencia
de ciertas enzimas BLEE respecto a estos antibióticos y al cambio en los
lineamientos de reporte de Betalactámicos para las cepas BLEE efectuado por la
CLSI en el año 2010.
- Con respecto a la prueba de Hodge, se concluye que todas las cepas
determinadas como BLEE positivas presentaron un resultado positivo empleando
una cefalosporina de tercera generación, y el resultado negativo de esta prueba
para Ciprofloxacina en todas las cepas es debido a que no presentaron ningún
mecanismo que hidrolizara quinolonas.

75
- La detección del Punto Isoeléctrico es una técnica sensible más no específica, ya
que muchas enzimas pueden tener el mismo punto isoeléctrico y esto dificulta su
clasificación y ubicación.
- En las cepas en las que se obtuvo transferencia del fenotipo de resistencia por
conjugación se demostró la presencia de los genes codificantes para BLEE en
elementos genéticos transferibles.
- Se detectó la presencia de los genotipos SHV, TEM, PER y CTX-M del grupo 2, y
se determinó que no existe una relación estadísticamente significativa entre la
resistencia a quinolonas y la presencia de cada uno de los genes BLEE por
separado (p>0.05).
- Respecto a la determinación de genes plasmídicos de resistencia a quinolonas,
se logró observar no solo la presencia individual del gen qnrB y aac(6’)Ib-cr, sino
que también se obtuvieron aislados con 2 mecanismos plasmídicos.
- Al realizar el análisis estadístico se determinó que existe una relación
estadísticamente significativa (p<0.05) entre la producción de Betalactamasas de
Espectro Expandido y la presencia de mecanismos plasmídicos de resistencia a
quinolonas; sin embargo, no se encontró una relación estadísticamente
significativa (p>0.05) entre la producción de BLEE y la presencia de cada uno de
los mecanismos plasmídicos de resistencia a quinolonas encontrados por
separado.
- Se demostró la presencia de integrones de clase 1 en las cepas en estudio y la
presencia de genes codificantes para BLEE y/o resistencia plasmídica a
quinolonas en estos elementos transferibles.
- Se propone, en algunos casos, la presencia de múltiples cassettes de resistencia
en el mismo integrón, existiendo algunos casos donde coexistirían un gen BLEE

76
y uno de resistencia plasmídica a quinolonas, lo cual explicaría la relación
estadística encontrada entre estos mecanismos de resistencia.
- Los resultados de secuenciación de los productos de amplificación de SHV y
TEM permitió determinar la presencia del genotipo TEM-1, codificante para una
BLEA, en varias cepas de E. coli, así como SHV-1 en K. pneumoniae, verificando
así los resultados obtenidos por IEF. La BLEE tipo SHV predominante fue la
SHV-5, y en el caso de TEM la TEM-150.
- Se encontró en la mayoría de las cepas mutaciones en gyrA codificantes para el
aminoácidos 83, el cual (serina) fue sustituído en algunos casos por leucina y
otros por isoleucina; cabe destacar que la CIM para Ciprofloxacina aumentó más
en las cepas donde la serina fue sustituída por isoleucina que en las que se
sustituyó por leucina, lo cual coincide con lo determinado por otros autores
- Al realizar el análisis estadístico se determinó que existe una relación
estadísticamente significativa (p<0,05) entre la producción de Betalactamasas de
Espectro Expandido y la presencia de mecanismos cromosomales de resistencia
a quinolonas.
- La asociación encontrada entre los mecanismos de resistencia a betalactámicos
y quinolonas refleja la necesidad de explorar otras alternativas terapéuticas para
el tratamiento de cepas en el cual se le detecte alguno de estos mecanismos de
resistencia.

77
RECOMENDACIONES
- Ampliar el estudio a otras áreas geográficas del país para establecer una
comparación entre los mecanismos de resistencia observados en Caracas con
otras ciudades.
- Establecer por centro la participación del personal del laboratorio de
microbiología para seguir realizando estudios en esta área para mantener una
evaluación continua sobre la situación de cada uno de estos mecanismos de
resistencia en nuestro país.
- Se recomienda continuar trabajos de vigilancia epidemiológica, en los que se
pueda realizar un seguimiento al comportamiento de la resistencia bacteriana a
betalactámicos en los diferentes centros de Caracas, para así establecer
medidas de prevención de propagación de dichos mecanismos.
- Se invita a realizar un trabajo que incluya a otros miembros del área de la salud
como los médicos, para establecer una relación entre los patrones de resistencia
observados en cada centro y la antibioterapia utilizada en los mismos, y así
establecer medidas de control y prevención para reducir el desarrollo y
propagación de nuevas resistencias.

78
REFERENCIAS BIBLIOGRÁFICAS
Alós J. 2009. Quinolonas. Enferm Infec Microbiol Clin; 27(5): 290-297.
Alvarez-Fernández M.; Rodríguez-Sousa T.; Brey-Fernández E.; López-
Meléndez C.; Piñeiro L. 2003. Asociación entre Integrones de clase con
Resistencia a multiples antimicrobianos y plásmidos conjugativos en
Enterobacteriaceae. Rev. Esp. Quimioterap. 16(4):394-397.
Ambler, R. 1980. The structure of b-lactamases. Philos. Trans. R. Soc. Lond. (Biol.)
289:321–331.
Arlet G.; Philippon A. 1991. Construction by polymerase chain reaction and use of
intragenic DNA probes for three main types of transferable beta-lactamases (TEM,
SHV, CARB). FEMS Microbiol Lett. 15; 66(1):19–25.
Biedenbach D.; Johnson D.; Jones R. In vitro evaluation of cefepime and other
broad-spectrum beta-lactams in eight medical centers in Thailand. 1999. The
Thailand Antimicrobial Resistance Study Group. Diagn Microbiol Infect Dis; 35: 325-
331.
Bouchakour M.; Zerouali K.; Perrier J.; Claude G.; Amarouch H.; El Mdaghri N.;
Courvalin P.; Timinouni M. 2010. Plasmid-mediated quinolone resistance in
expanded spectrum beta lactamase producing enterobacteriaceae in Morocco. J
Infect Dev Ctries; 4(12):799-803.
Branger C.; Zamfir O.; Geoffroy S.; Laurans G.; Arlet G.; Thien H.; Gouriou S.;
Picard B.; Denamur E. 2005. Genetic background of Escherichia coli and Extended-
Spectrum Beta-Lactamase Type. Emerg Infect Dis. 11 (1): 54-61.
Bush K.; Jacoby G. 2010. Updated functional classification of β-lactamases.
Antimicrob Agents Chemother; 54:969-76.

79
Cattoir V.; Poirel L.; Rotimi V.; Soussy C.; Nordmann P. 2007. Multiplex PCR for
detection of plasmid-mediated quinolone resistance qnr genes in ESBL-producing
enterobacterial isolate. J Antimicrob Chemother; 60:394-397.
Cavaco L.; Korsgaard H.; Sorensen G.; Aarestrup F. 2008. Plasmid-mediated
quinolone resistance due to qnrB5 and qnrS1 genes in Salmonella enterica serovars
Newport, Hadar and Saintpaul isolated from Turkey meat in Denmark. J Antimicrob
Chemother 62(3): 632-634.
Chen Y.; Shu H.; Li L.; Liao T.; Wu K.; Shiau Y. 2006. Complete Nucleotide
Sequence of pk245, a 98-Kilobase Plasmid Conferring Quinolone Resistance and
Extended-Spectrum β-lactamase Activity in a Clinical Klebsiella pneumoniae isolate.
Antimicrob Agents Chemother 50(11): 3861-3866.
Clinical and Laboratory Standards Institute. 2010. Performance Standards for
Antimicrobial Susceptibility Testing. Documento M100-S20.
Comegna M.; Guzmán M.; Carmona O.; Molina M.; Grupo colaborativo del
Grupo Venezolano de Resistencia Bacteriana. 2000. Resistencia bacteriana a los
antimicrobianos en Venezuela-Nuevos hallazgos. Boletín SVM 20(1): 58-63
Diestra K.; Juan C.; Curiao T.; Moyá B.; Miró E.; Oteo J.; Coque T.; Pérez-
Vázquez M.; Campos J.; Cantón R.; Oliver A.; Navarro F.; Red Española de
Investigación en Patología Infecciosa. 2009. Characterization of plasmids
encoding blaESBL and surrounding genes in Spanish clinical isolates of Escherichia
coli and Klebsiella pneumoniae. J Antimicrob Chemother.63(1):60-6
Doyle, K. 1996. The source for discovery: protocols and applications guide. Catálogo
Promega Biologicals Research Products.
Drlica K. 1999. Mechanism of fluoroquinolone action. Curr Opin Microbio; 2: 504-
508.

80
Errecalde J. 2004. Uso de Antimicrobianos en Animales de consumo: Incidencia del
desarrollo de resistencias en Salud Pública. Estudios FAO Producción y Sanidad
Animal, p71.
Fernández A., Sousa R., Fernández B., Melendez L., Piñeiro L. 2003. Asociación
entre Integrones de clase I con Resistencia a multiples antimicrobianos y plásmidos
conjugativos en Enterobacteriaceae. Rev. Esp. Quimioterap. 16(4): 394-397
Fu Y.; Guo L.; Xu Y.; Zhang W.; Gu J.; Xu J.; Chen X.; Zhao Y.; Ma J.; Liu X.;
Zhang F. 2008. Alteration of GyrA amino acid required for Ciprofloxacin resistance in
Klebsiella pneumoniae isolates in China. Antimicrob Agents Chemother. 52(8): 2980-
2983
Galas M.; Melano R.; Petroni A.; Pasterán F.; Faccone D. 2004. Manual de
Procedimientos de Biología Molecular y Proteínas. Servicio de Antimicrobianos INEI
ANLIS “Dr. Carlos G. Malbrán”, Argentina.
García I.; Valenzuela E.; Saavedra C.; Leal A.; Eslava J.; Mantilla J. 2005.
Caracterización Molecular de aislamientos de Enterobacter cloacae multirresistentes
productores de B-lactamasas provenientes de pacientes de un hospital de tercer
nivel de Bogotá. Rev Fac Med Univ Nac Colombia.; 53(3): 148-159.
Garza-Ramos U.; Martínez-Romero E.; Silva-Sánchez J. 2007. SHV-type
extended-spectrum B-lactamase (ESBL) are encoded in related plasmids from
enterobacteria clinical isolates from Mexico. Salud pública Méx. 49(6): 415-421.
Goodman y Gilman. The pharmacological basis of therapeutics. McGraw-Hill. New
York. Chicago. 2001
Graffunder E.; Preston K.; Evans A.; Venezia R. 2005. Risk factors associated
with extended-spectrum beta-lactamase-producing organisms at a tertiary care
hospital. J Antimicrob Chemother; 56(1):139-45.

81
Greisen K.; Loeffelholz M.; Purohit A.; Leong D. 1994. PCR primers and probes
for the 16S rRNA gene of most species of pathogenic bacteria, including bacteria
found in cerebrospinal fluid. JCM 32: 335-351;
Grinsted, J.; Bennett, P. 1998. Plasmid Technology. 2nd Edition. Academic Press
Limited.
Hata M.; Suzuki M.; Matsumoto M.; Takahashi M.; Sato K.; Ibe S.; Sakae K.
2005. Cloning of a novel gene for quinolone resistance from a transferable plasmid in
Shigella flexneri 2b. Antimicrob. Agents Chemother; 49: 801-803.
Iyobe, S. 1997. Appearance of extended-spectrum beta-lactamases. Nippon Rinsho
55(5):1219-24
Jacoby, G.; Muñoz-Price, S. 2005. The new betalactamases. N. Eng J Med.
352(4):380-391.
Jarlier V., Nicolas M., Fournier G., Philippon A. 1988. Extended broad-spectrum
β-lactamases conferring transferable resistance to newer β-lactam agents in
Enterobacteriaceae: prevalence and susceptibility patterns. Rev Infect Dis 10: 867-
78.
Jehl, F.; Chomarat, M.; Weber, M.; Gérard, A. 2004. Del Antibiograma a la
prescripción. Éditions Biomériux. Segunda edición.
Jiang Y.; Zhou Z.; Qian Y.; Wei Z.; Yu Y.; Hu S. 2008. Plasmid-mediated quinolone
resistance determinants qnr and aac (6’)-Ib-cr in extended-spectrum β-lactamase-
producing Escherichia coli and Klebsiella pneumoniae in China. J Antimicrob
Chemother; 61:1003-1006
Joklik, W.; Willet, H.; Amos, D.; Wilfert, C. 1998. Zinsser Microbiología. Editorial
Médica Panamericana. 20 Ed.

82
Jonas D.; Biehler K.; Hartung D.; Spitzmuller B.; Daschner F. 2005. Plasmid-
mediated quinolone resistance in isolates obtained in german intensive care units.
Antimicrob Agents Chemother; 49:773-775.
Knothe, A.; Shah, P.; Kremery, V.; Antal, M.; Mitsuhashi, S. 1983. Transferable
resistance to Cefotaxima, cefoxitin, cefamandole and cefuroxime in clinical isolates
of Klebsiella pneumoniae and Serratia marcescens. Infection 11:315-317
Koneman, E.; Allen, S.; Dowell, V.; Janda, W.; Sommers, H.; Winn, W. 1999.
Diagnóstico Microbiológico texto y atlas color. Editorial Médica Panamericana. 3era
edición.
Lagacé-Wiens P.; Nichol K.; Nicolle L.; DeCorby M.; McCracken M.; Alfa M.;
Mulvey M.; Zhanel G. 2007. ESBL genotypes in fluoroquinolone-resistant and
fluoroquinolone-susceptible ESBL-producing Escherichia coli urinary isolates in
Manitoba. Can J Infect Dis Med Microbiol. 18(2): 133–137.
Levesque C.; Piche L.; Larose Ch.; Roy P. 1995. PCR mapping of integrons
reveals several novel combinations of resistance genes. Antimicrob Agents
Chemother. 39(1): 185-191.
Lorian, V. 1996. Antibiotics in Laboratory Medicine. 4th Ed. Williams and Wilkins.
USA.
Lucet, J.; Regnier, B. 1998. Enterobacteria producing extended spectrum beta-
lactamases. Pathol Biol 46(4):235-43.
Madigan, M. 1998. Biología de los Microorganismos. 2da edición.
Mammeri H.; Van De L.; Poirel L.; Martinez-Martinez L.; Nordmann P. 2005.
Emergence of plasmid-mediated quinolone resistance in Escherichia coli in Europe.
Antimicrob Agents Chemother; 49:71-76.
Mandell, D. 2002. Enfermedades Infecciosas. 5ta edición. Editorial Panamericana.

83
Marcano, D. 2002. Detección de Betalactamasas de Espectro Expandido en cepas
de Enterobacterias aisladas de pacientes del Hospital Universitario de Caracas.
Trabajo Especial de Grado para optar al Título de Licenciado en Bioanálisis.
Universidad Central De Venezuela, Escuela de Bioanálisis. 97 p
Martínez, L.; Pascual, A.; Conejo, M.; García, I.; Joyanes, P.; Domenech, A.;
Benedi, J. 2002. Energy-Dependent Accumulation of Norfloxacin and Porin
Expression in Clinical Isolates of Klebsiella pneumoniae and Relationship to
Extended-Spectrum Betalactamase Production. Antimicrob Agents Chemother.
46(12): 3926-3932.
Martinez-Martinez L.; Pascual A.; Jacoby G. 1998. Quinolone resistance from a
transferable plasmid. Lancet 351:797-799.
Matsumoto, Y.; Ikeda, F.; Kanimura, T.; Yokota, Y.; Mine, Y. 1988. Novel plasmid-
mediated -lactamase from Escherichia coli that inactivates oxymino-cephalosporins.
Antimicrob Agents Chemother. 32:1243-1246.
Mendelson, G.; Hait, V.; Ben-Israel, J.; Gronich, D.; Granot, E.; Raz, R.
2005. Prevalence and risk factors of extended-spectrum beta-lactamase-producing
Escherichia coli and Klebsiella pneumoniae in an Israeli long-term care facility. Eur J
Clin Microbiol Infect Dis;24(1):17-22.
Minarini L.; Poirel L.; Cattoir V.; Darini A.; Nordmann P. 2008. Plasmid-mediated
quinolone resistance determinants among enterobacterial isolates from outpatients in
Brazil. J Antimicrob Chemother; 62: 474-478.
Morgan-Linnell S.; Zechiedrich L. 2007. Contributions of the Combined Effects of
Topoisomerase Mutations toward Fluoroquinolone Resistance in Escherichia coli.
Antimicrob Agents Chemother. 51(11): 4205-4208.
Mullis K.; Faloona F. 1987. Specific synthesis of DNA in vitro via a polymerase-
catalyzed chain reaction. Meth. Enzymol 155,335-350.

84
Navarro F.; Miró E.; Mirelis B. 2002. Lectura interpretada del antibiograma de
enterobacterias. Enferm. Infecc. Microbiol. Clin 20(5):225-234.
Nordmann P.; Poirel L. 2005. Emergence of plasmid-mediated resistance to
quinolones in Enterobacteriaceae. J Antimicrob Chemother 56: 463-469.
Oliver A.; Cantón R. 2008. Enterobacterias productoras de ß-lactamasas
plasmídicas de espectro extendido. Revista de Control de Calidad SEIMC.
Palumbo M.; Gatto B.; Zagotto G.; Palu G. 1993. On the mechanism of action of
quinolone drugs. Trends Microbiol; 1: 232-235.
Perozo A.; Castellano M. 2009. Detección de Betalactamasas de Espectro
Extendido en cepas de la familia Enterobacteriaceae. Kasmera 37(1):.25-37.
Pfaller M.; Jones R.; Doern G.; Sader H.; Messer S.; Houston A.; Coffman S.;
Hollis R. 2000. Bloodstream infections due to Candida species: SENTRY
antimicrobial surveillance program in North America and Latin America, 1997-1998.
Antimicrob Agents Chemother. 44(3):747-51.
Pfaller, M.; Jones, R.; Doern, G. 1998. Multicenter evaluation of the antimicrobial
activity for six broad-spectrum beta-lactams in Venezuela using the Etest method.
The Venezuelan Antimicrobial Resistance Study Group. Diagn Microbiol Infect Dis
30(1):45-52
Pitout J, Wei Y, Church D, Gregson D. 2008. Surveillance for plasmid-mediated
quinolone resistance determinants in Enterobacteriaceae within the Calgary Health
Region, Canada: the emergence of aac(6’)-Ib-cr. J Antimicrob Chemother; 61:999-
1002.
Prescot, L.; Harley, J.; Klein, D. 1999. Microbiología. Editorial McGraw-Hill.
Ramphal, R.; Ambrose, P. 2006. Extended Spectrum Betalactamases and Clinical
Outcomes: Current Data. Clin. Infect. Dis. 42:S164-172.

85
Reyes A.; Bello H.; Dominguez M.; Mella S.; Zemelman R.; González G. 2003.
Prevalence and types of class 1 integrons in aminoglycoside-resistant
Enterobacteriaceae from several Chilean hospitals. J. Antimicrob. Chemother. 51:
317-321.
Rodríguez J. 2006. Resistencia mediada por plásmidos en Enterobacteriaceae.
Tesis doctoral, Universidad de Sevilla, España.
Rodriguez-Martinez J. 2005. Mechanisms of plasmid-mediated resistance to
quinolones. Enferm Infecc Microbiol Clin; 23:25-31.
Rodríguez-Martínez M.; Pichardo C.; García I.; Pachón-Ibañez M.; Docobo-
Pérez F.; Pascual A. 2008. Activity of ciprofloxacin and levofloxacin in experimental
pneumonia caused by Klebsiella pneumoniae deficient in porins, expressing active
efflux and producing QnrA1. Clin Microbiol Infect. 14: 691-697.
Sierra J. 2005. Bases Moleculares de la Resistencia a Quinolonas en
Staphylococcus aureus, Streptococcus pneumoniae y Corynebacterium spp [Tesis
Doctoral] España: Universidad de Barcelona.
Spanu, T.; Luzzaro, F.; Perilli, M.; Amicosante, G.; Toniolo, A.; Fadda, G. 2002.
Occurrence of Extended-Spectrum ß-Lactamases in Members of the Family
Enterobacteriaceae in Italy: Implications for Resistance to ß-Lactams and Other
Antimicrobial Drugs. Antimicrob Agents Chemother 46(1): 196-202.
Speldooren V.; Heymn B.; Labia R.; Nicolas-Chanoine MH. 1998. Discriminatory
Detection of Inhibitor-Resistant B-Lactamases in Escherichia coli by Single-Strand
Conformation Polymorphism-PCR. Antimicrob Agents Chemother 42(4): 879 – 884.
Szabó D, Kocsi B, Rókusz L, Szentandrássy J, Katona K, Kristóf K. 2008. First
detection of plasmad-mediated, quinolone resistance determinants qnrA, qnrB, qnrS
and aac(6’)-Ib-cr in extended spectrum β-lactamase (ESBL)-producing
Enterobacteriaceae in Budapest, Hungary. J Antimicrob Chemother 62(3): 630-632.

86
Tran J, Jacoby G, Hooper D. 2005. Interaction of the plasmid encoded quinolone
resistance protein Qnr with Escherichia coli DNA gyrase. Antimicrob Agents
Chemother; 49:118-125.
Tran J, Jacoby G. 2002. Mechanism of plasmid-mediated quinolone resistance.
Proc Natl Acad Sci USA; 99:5638-5642.
Vakulenko S., Mobashery S. 2003. Versatility of Aminoglycosides and Prospects for
Their Future. Clinical Microbiology Reviews, 16: 430-450.
Walsh, C. Antibiotics: Actions, origins, resistance. 2003. ASM press. 1era ed.
Washington DC. p 6-15 (335p)
Wang M.; Sahm D.; Jacoby G.; Hooper D. 2004. Emerging plasmid-mediated
quinolone resistance associated with the qnr gene in Klebsiella pneumoniae clinical
isolates in the United States. Antimicrob Agents Chemother; 48:1295-1299.
Wang M.; Tran J.; Jacoby G.; Zhang Y.; Wang F.; Hooper D. 2003. Plasmid-
mediated quinolone resistance in clinical isolates of Escherichia coli from Shanghai,
China. Antimicrob Agents Chemother; 47:2242-2248.


















