Replicacion Patogenia y Mecanismos
19
Replicación, patogenia y mecanismos REPLICACION VIRAL. La expresión "replicación viral" se aplica en este capítulo al proceso de proliferación a "nivel celular" más que a nivel del individuo. La condición de parásitos intracelulares estrictos que define a los virus está precisamente determinada por el hecho de que es en las células donde se fabrica la progenie viral descendiente de un virión parental. ETAPAS DE LA REPLICACIÓN Se han definido cinco etapas indispensables en el proceso de replicación viral: Adsorción: Lo que define al proceso de adsorción es la interacción específica entre el virión y una célula, de tal forma que permita la retención de la partícula en la superficie celular. Dicha retención ocurre porque existe una alta afinidad entre una molécula de la superficie del virus (el ligando) y otra molécula de la célula (el receptor); ambas contrapartes son proteínas, frec;uentemente glicosiladas, con algunos dominios involucrados en esa interacción. Evidentemente, en los virus con manto el ligando es una proteína de membrana, mientras que en los virus desnudos el ligando puede tener también una función estructural, lo que le da estabilidad a la partícula. En ambos casos, el ligando puede estar constituido por más de una cadena polipeptídica, que pueden ser idénticas, La especificidad de la interacción ligando/receptor delimita la posibilidad de que un virus interactúe con un determinado tipo de célula y de tejido, puesto que si un tipo celular no expresa el receptor adecuado, el virus no podrá adsorberse a él. El término tropismo tiene un significado más amplio, pues se refiere a la preferencia de un virus por infectar determinados tejidos, y depende tanto de la presencia de receptores como de la permisividad,
-
Upload
johannaazucenabohorquezmacias -
Category
Documents
-
view
42 -
download
1
description
Replicacion Patogenia y Mecanismos
Transcript of Replicacion Patogenia y Mecanismos

Replicación, patogenia y mecanismos
REPLICACION VIRAL.
La expresión "replicación viral" se aplica en este capítulo al proceso de proliferación a "nivel celular" más que a nivel del individuo. La condición de parásitos intracelulares estrictos que define a los virus está precisamente determinada por el hecho de que es en las células donde se fabrica la progenie viral descendiente de un virión parental.
ETAPAS DE LA REPLICACIÓN
Se han definido cinco etapas indispensables en el proceso de replicación viral:
Adsorción: Lo que define al proceso de adsorción es la interacción específica entre el virión y una célula, de tal forma que permita la retención de la partícula en la superficie celular. Dicha retención ocurre porque existe una alta afinidad entre una molécula de la superficie del virus (el ligando) y otra molécula de la célula (el receptor); ambas contrapartes son proteínas, frec;uentemente glicosiladas, con algunos dominios involucrados en esa interacción.
Evidentemente, en los virus con manto el ligando es una proteína de membrana, mientras que en los virus desnudos el ligando puede tener también una función estructural, lo que le da estabilidad a la partícula. En ambos casos, el ligando puede estar constituido por más de una cadena polipeptídica, que pueden ser idénticas,
La especificidad de la interacción ligando/receptor delimita la posibilidad de que un virus interactúe con un determinado tipo de célula y de tejido, puesto que si un tipo celular no expresa el receptor adecuado, el virus no podrá adsorberse a él.
El término tropismo tiene un significado más amplio, pues se refiere a la preferencia de un virus por infectar determinados tejidos, y depende tanto de la presencia de receptores como de la permisividad, es decir, de su capacidad de replicarse una vez que ha ingresado a la célula.
Penetración Dado que para replicar los virus deben acceder a la maquinaria y a recursos intracelulares, su estructura está adaptada para que, luego de la adsorción, la partícula o parte de ella, sea internalizada. En este proceso, una gran estructura macromolecular -en este caso

al menos una cápside con genoma en su interior- traspasa una estructura delimitante, la membrana citoplasmática celular.
mecanismos de penetración:
Penetración por fusión. Es utilizado por algunos virus con envoltura. Se ba a en la reorganización molecular entre los lípidos del manto viral y los de la membrana celular para formar una sola superficie entre ambos.
Debido a que los lípidos virales y celulares forman una sola lámina, los antígenos virales de superficie quedan retenidos en la membrana celular y expuestos hacia el exterior; sin embargo, no es evidente que dichas proteínas puedan tener alguna función (inmunogénica o señalizadora), pues en algunos casos debido al reciclaje normal de las proteínas de membrana su duración es relativamente efímera.
Penetración por endocitosis (viropexia). Este proceso evidencia la capacidad de los virus de usufructuar de la mecánica celular, pues la mayoría de los tipos celulares está programada para captar moléculas o complejos moleculares del espacio extracelular por medio de la formación de endosomas, o vesículas intracelulares. Notablemente, este proceso es mecánicamente inverso a la fusión, pues consiste en la separación de una parte de la lámina lipídica para la formación del endosoma, sin romper la continuidad de la membrana citoplasmática.
Una vez formada la vesícula endocítica, su destino depende de sus interacciones y señalizaciones con el citoesqueleto; en algunos casos el virus requiere que su material genético a. b. c. MC permanezca en el citoplasma, mientras que en otros debe ser transportado hasta el núcleo. Las células normalmente someten a sus endosomas a un proceso de acidificación por medio de bombas de protones instaladas en la membrana, y frecuentemente, ese cambio de pH induce modificaciones en proteínas virales que son necesarias para continuar con el ciclo replicativo.
así como para otros virus con manto, la liberación de la nucleocápside requiere de la fusión de la envoltura viral con la membrana del endosoma, lo que ocurre después de la acidificación del endosoma, pues con esa señal el ligando viral expone el dominio fusogénico. Aunque este proceso involucra la fusión entre dos membranas, el mecanismo de "penetración por fusión" se refiere únicamente al descrito previamente, donde no hay formación de endosoma.
En los virus desnudos se visualizan dos destinos posibles luego de la endocitosis: algunos endosomas pueden disolverse, liberando con ello su contenido en el citosol, mientras que en otros casos se han observado cápsides virales al interior de cisternas del retículo endoplasmático, lo que sugiere que algunos endosomas se funden con este organelo, liberando contenido en el interior de él

Modelo general de un ciclo replicativo viral. Se grafica la replicación de un virus desnudo con replicación exclusivamente citoplasmática. Se detallan los componentes (a-g) y procesos (1-7). El virión (a) hace contacto con el receptor (b) durante el proceso de adsorción (1 ). Después de la etapa de penetración (2), se inicia la biosíntesis de macromoléculas con la transcripción (3), que permite obtener ARN mensajero vira l (e); la traducción (4), por la que se fabrican las diversas proteínas virales (d) y la replicación genómica (5), en la que participan tanto el genoma viral (e) como enzimas virales (d), obteniéndose así múltiples copias nuevas de genoma viral (f). El ensamblaje (6) ocurre por la interacción de proteínas estructurales (d) y genomas virales (f). Finalmente, la liberación (7) de la progenie (g) ocurre por lisis celular
Síntesis de macromoléculas
El virus debe garantizar la síntesis de dos tipos de macromoléculas: proteínas virales y ácidos nucleicos virales. Aunque los virus con manto deben incluir lípidos en su estructura, su síntesis no es especial, pues los mantos virales derivan directamente de membranas celulares. Muchos virus también tienen la capacidad de modificar el patrón de expresión tanto de ácidos nucleicos como de proteínas celulares.
Transcripción viral. La primera manifestación de actividad biosin~ética viral es la obtención de ARN mensajero viral, en un proceso conocido como transcripción. Estas moléculas, al igual que los mensajeros celulares, son intermediarias mediante las cuales la información fluye desde el genoma hacia la síntesis de proteínas en los ribosomas celulares. Todos los mensajeros son sintetizados necesariamente por una ARN polimerasa, aunque el molde que dicha enzima debe leer para fabricar el mensaje varía dependiendo del tipo de virus.

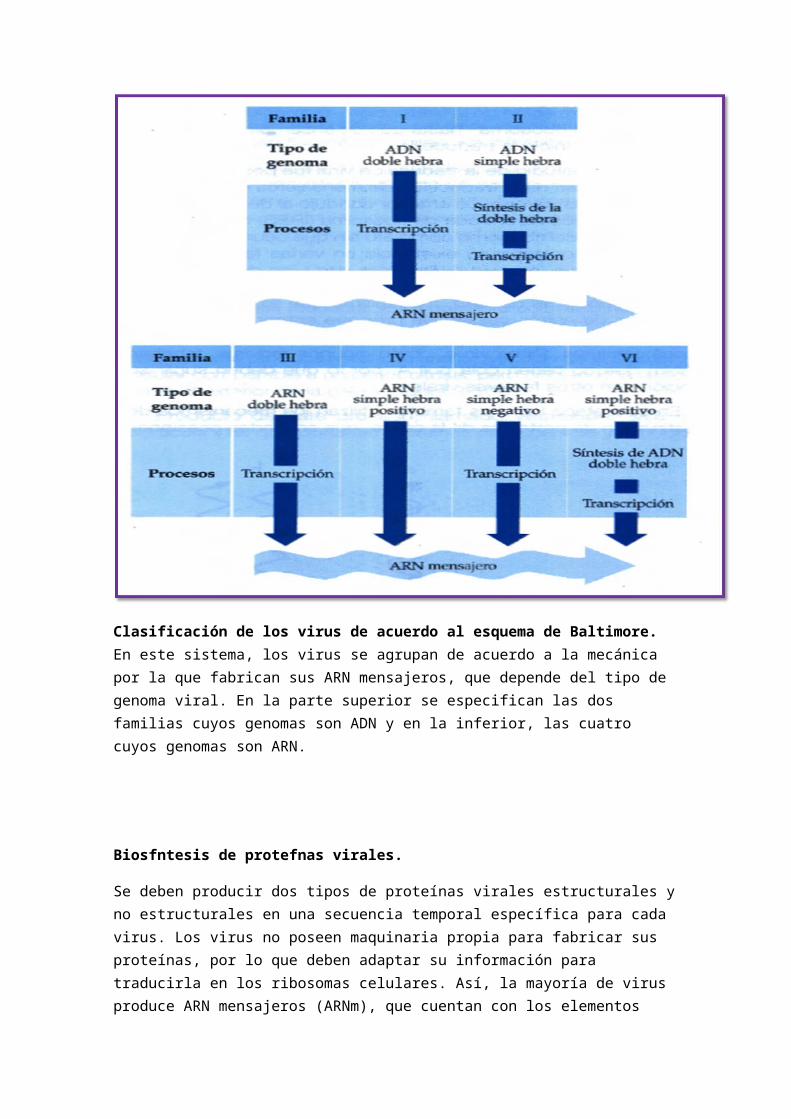
Clasificación de los virus de acuerdo al esquema de Baltimore. En este sistema, los virus se agrupan de acuerdo a la mecánica por la que fabrican sus ARN mensajeros, que depende del tipo de genoma viral. En la parte superior se especifican las dos familias cuyos genomas son ADN y en la inferior, las cuatro cuyos genomas son ARN.
Biosfntesis de protefnas virales.
Se deben producir dos tipos de proteínas virales estructurales y no estructurales en una secuencia temporal específica para cada virus. Los virus no poseen maquinaria propia para fabricar sus proteínas, por lo que deben adaptar su información para traducirla en los ribosomas celulares. Así, la mayoría de virus produce ARN mensajeros (ARNm), que cuentan con los elementos típicos de un mensajero celular: una estructura cap, un marco de lectura abierto que porta la información codificante efectiva y una señal de poliadenilación , o en su defecto, directamente una cola poli A. Esta molécula es sometida a scanning por el complejo de iniciación de la traducción -que incluye la subunidad menor del ribosoma- hasta que se ensambla el ribosoma completo y se inicia la traducción.
Ensamblaje
Producto de esta etapa, es posible observar al interior de la célula las primeras partículas virales o, en el caso de los virus con manto, las primeras nucleocápsides. Las propiedades intrínsecas de las proteínas estructurales de los virus frecuentemente impiden separar la etapa de ensamblaje con la de biosíntesis de proteínas, pues una vez terminada la traducción, las proteínas que forman las cápsides virales empiezan a agregarse en su arquitectura definitiva. De hecho, esta tendencia a autoestructurarse ha permitido usar proteínas recombinantes como inmunógenos, como en el caso de las vacunas contra el papiloma virus, en las que la sola expresión de una proteína de cápside induce la formación de "pseudo-partículas" (VLPs o víral-líke partícles), que tienen propiedades antigénicas como las de una partícula.
Liberación
Se estima que producto del ensamblaje pueden acumularse entre cien y diez mil partículas virales al interior de una célula, dependiendo del virus. Para reiniciar un nuevo ciclo, estas partículas deben salir de la célula, que para ese momento puede ya estar en colapso fisiológico debido a la infección viral. La mayoría de los virus desnudos sale de la célula una vez que la membrana citoplasmática se rompe, liberando todo el contenido interno, en un proceso conocido como lisis celular. Dado que las partículas deben estar listas para infectar una nueva célula, la morfogénesis de estos virus concluye antes de la liberación.
En contraposición, diversos virus con manto concluyen su morfogénesis durante la liberación, pues en esa etapa adquieren el manto que los recubrirá Para que esto ocurra, la nucleocápside debe ser transportada hacia la membrana citoplasmática, donde será reconocida por proteínas virales asociadas a ella; en esta zona se produce entonces una proyección de la membrana celular comparable a una endocitosis, pero dirigiendo la vesícula hacia el exterior de la célula.
Producto de este proceso, llamado gemación, sale una partícula viral que porta las proteínas de superficie y la nucleocápside viral. Es evidente que la membrana celular tiene una capacidad finita para efectuar este proceso, pues la escasez de componentes lipídicos (fosfolípidos y colesterol) termina por acabar con su integridad, por lo que no es raro que algunos virus modulen la liberación de partículas para alargar el proceso de replicación. De esta forma, muchos virus con manto no destruyen á su célula hospedera al concluir su ciclo replicativo. En algunos virus envueltos se ha descrito que una vez fuera de la célula, procesan internamente sus proteínas, en lo que se denomina maduración.
PATOGENIA VIRAL
Es la forma en que un virus invade un organismo y genera una infección con o sin producción de lesión o enfermedad.
La patogenia incluye las etapas sucesivas, que comprenden la fuente de origen de la infección, la entrada del virus al hospedero, la replicación dentro de las

células, las vías de diseminación dentro del cuerpo, las interacciones con la respuesta inmune del huésped y la evolución final de la infección.
En la patogenia de una virosis intervienen tres grupos de factores que interactúan entre sí y que son múltiples y cambiantes
Dependientes del virus. Son aquellos inherentes a la biología del virus. La relación del ligando viral con el receptor celular determina el tropismo del virus por especies y tejidos.
Dependientes del ambiente. Las condiciones locales de temperatura, humedad, salinidad, pH, luz ultravioleta, aireación y otras pueden influir en la viabilidad del virus y afectar su capacidad infectiva.
Dependientes del hospedero. Ciertos factores innatos, como la especie, la raza, la edad, el sexo y otros, a través de receptores celulares específicos determinan la susceptibilidad a los virus. Los mecanismos de defensa adquiridos (vía transplacentaria, infecciones naturales, vacunas, etc.), la actividad y condiciones de vida, y otros factores también variables, influyen el riesgo de infectarse.
A nivel del individuo, el destino final de una infección viral está determinado por la interacción de los factores mencionados, que pueden ser distintos de una célula a otra, de una persona a otra y de una comunidad a otra.
La interacción de factores puede originar cuatro modelos de infecciones virales:
1. Ausencia de infección o infección abortiva2. Infección aguda, con o sin síntomas, que conduce a la recuperación o a
la muerte, con sus estados intermedios3. Infección persistente, ya sea de tipo latente, crónica o lenta4. Infección transformante, que origina un tumor benigno o Maligno
PATOGENIA A NIVEL CELULAR
Todas las alteraciones o efectos citopáticos, pueden ser tan características, que permiten diagnosticar e identificar el virus.
Las células que permiten la replicación viral se llaman permisivas, y las infecciones de estas células, que son usualmente productivas y citolíticas, generan progenie viral y muerte celular. Las infecciones en células no permisivas producen progenies virales no infectivas, llamadas infecciones abortivas.
A continuación se dan ejemplos de efectos y consecuencias de la patogenia de la infección a nivel celular.

Efectos citopáticosDiversas alteraciones que los virus inducen al infectar las células, denominadas efecto citopático (ECP).
Los principales ECP inducidos por virus son: Cambios morfológicos Lisis o muerte celular. Expresión de proteínas y antígenos Fusión celular (entre la envoltura viral y la membrana celular)
Mecanismos de lesión celular
El daño en las células infectadas, que afecta a tejidos y órganos, es uno de los elementos esenciales y determinantes en la patogenia. Este daño puede ser por lesión directa por la acción de los virus o por lesión indirecta, debido a la acción de mecanismos derivados de la respuesta inmune.
La apoptosis es un proceso altamente regulado que permite que la célula se autodegrade para que el organismo elimine aquellas no deseadas o disfuncionales.
PATOGENIA A NIVEL DEL INDIVIDUO
Etapas de la infección viral
Fuentes de contagio. Fómites
Mecanismos de contagio. Directos al tener contacto físico por ejemplo un beso, contacto de
piel o contacto sexual, favorecen la transferencia del agente por alteraciones en las barreras mecánicas que son la puerta de entrada en este caso serían mucosas o piel.

Sin contacto físico: Cuando las virosis se contagian a través de secreciones eliminadas por los individuos infectados. Por ejemplo las partículas menores de 5um que emiten al hablar, estornudar o toser. Como frecuentemente ocurre con los virus exantemáticos (caracterizadas por la aparición de exantemas (erupciones o manchas rojizas en la piel), muy frecuentes en la infancia y que, aunque generalmente no producen secuelas importantes, en ocasiones pueden complicarse con cuadros muy severos y hasta fatales. Como: Virus de la Rubéola, Virus del Sarampión y Parvovirus B19.) Y Virus respiratorios.
Indirectos Este método implica un elemento intermediario inerte (vehículo) como el agua o los alimentos y en ellos se puede encontrar el ROTAVIRUS DEBIDO A QUE PUEDE PERMANECER VIABLE EN EL MEDIO AMBIENTE o un elemento vivo (vector) como insectos que pueden ser mecánicos si se transmite por las patas o probóscide; y biológicos si el agente se multiplica y desarrolla parte del ciclo reproductivo en ellos como ZANCUDOS DEL DENGUE(GÉNERO FLAVIVIRUS) .
Puertas de entrada. ( SIEMPRE CUANDO HAYAN RECEPTORES EN ESTOS SITIOS)-naturales
Piel Debe estar lesionada para permitir el ingreso de la infección viral MucosasAlgunas permiten la adosrción viral a las células epiteliales
(repiratorias, otras requieren lesiones que favorezcan la penetración viral (sexuales)
transplacentaria.
-artificial parenteralJeringas, transfusiones
DISEMINACIÓN EN EL ORGANISMO Localizada: El virus permanece en el sitio de entrada o se disemina
localmente por vecindad ( INFECCIONES RESPIRATORIAS, DIARREAS,HERPES SIMPLE).

Periodo de incubación corto, estimulan preferentemente la inmunidad local y pueden repetirse.
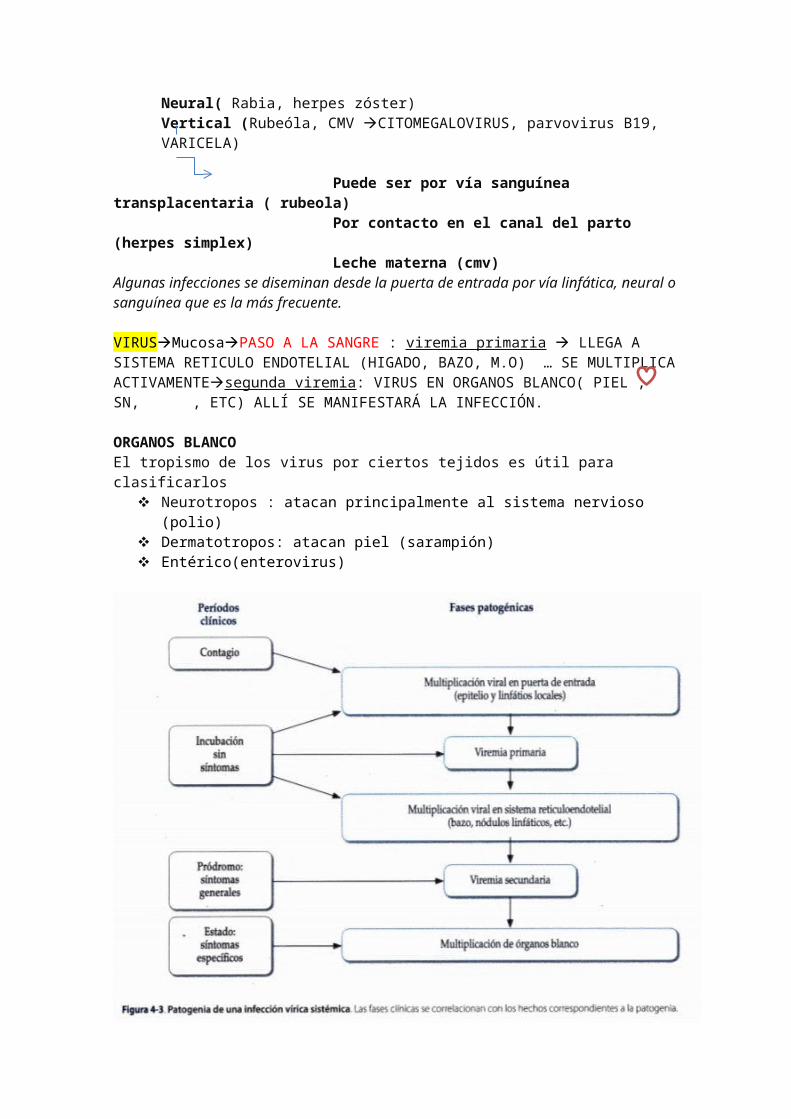
Diseminación/ SITÉMICA/ generlizada: Depende del virus y de su tropismo. Así mismo alteraciones de las respuesta inmune favorecen a ello (inmunospuprimidos) (ENFERMEDDADES EXANTEMÁTICAS, HEPATITIS, ENCEFALITIS)Periodo de incubación largo, al multiplicarse en varios sitios estimulan la respuesta inmune a todo nivel . confieren inmunidad de larga duración.Neural( Rabia, herpes zóster) Vertical (Rubeóla, CMV CITOMEGALOVIRUS, parvovirus B19, VARICELA)
Puede ser por vía sanguínea transplacentaria ( rubeola) Por contacto en el canal del parto (herpes simplex) Leche materna (cmv) Algunas infecciones se diseminan desde la puerta de entrada por vía linfática, neural o sanguínea que es la más frecuente.
VIRUSMucosaPASO A LA SANGRE : viremia primaria LLEGA A SISTEMA RETICULO ENDOTELIAL (HIGADO, BAZO, M.O) … SE MULTIPLICA ACTIVAMENTEsegunda viremia: VIRUS EN ORGANOS BLANCO( PIEL , SN, , ETC) ALLÍ SE MANIFESTARÁ LA INFECCIÓN.
ORGANOS BLANCOEl tropismo de los virus por ciertos tejidos es útil para clasificarlos
Neurotropos : atacan principalmente al sistema nervioso (polio) Dermatotropos: atacan piel (sarampión) Entérico(enterovirus)

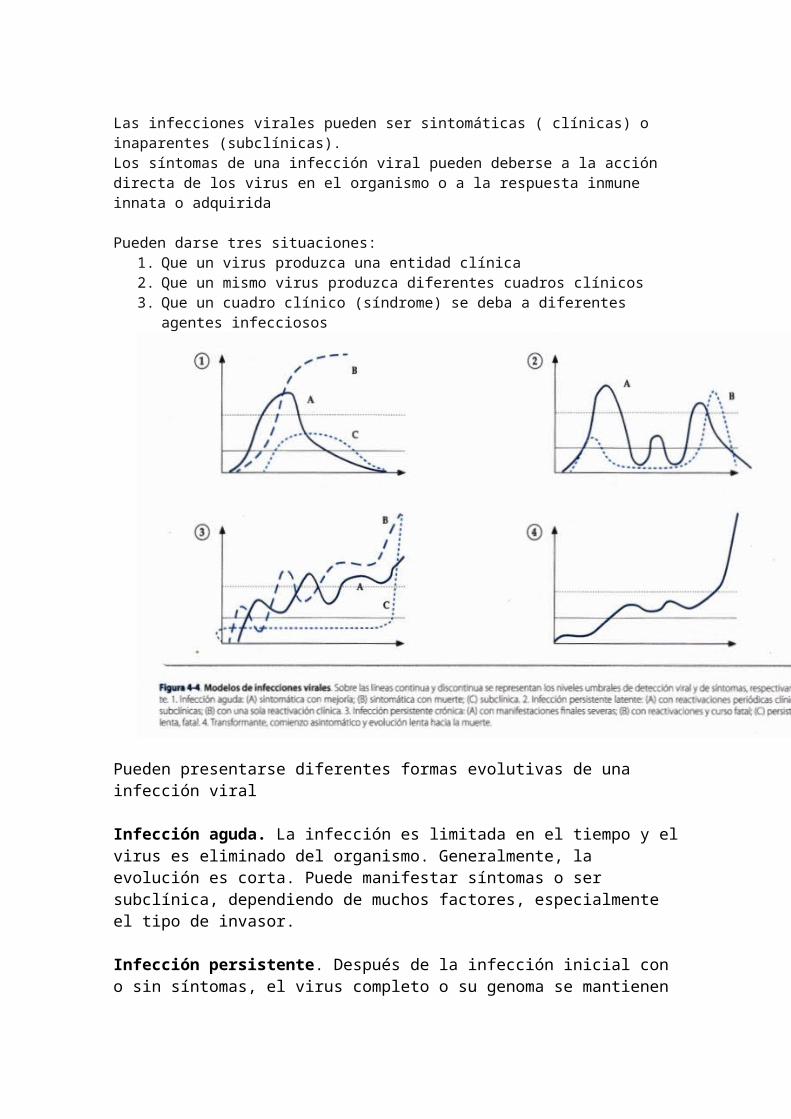
Las infecciones virales pueden ser sintomáticas ( clínicas) o inaparentes (subclínicas).Los síntomas de una infección viral pueden deberse a la acción directa de los virus en el organismo o a la respuesta inmune innata o adquirida
Pueden darse tres situaciones:1. Que un virus produzca una entidad clínica2. Que un mismo virus produzca diferentes cuadros clínicos3. Que un cuadro clínico (síndrome) se deba a diferentes agentes infecciosos
Pueden presentarse diferentes formas evolutivas de una infección viral
Infección aguda. La infección es limitada en el tiempo y el virus es eliminado del organismo. Generalmente, la evolución es corta. Puede manifestar síntomas o ser subclínica, dependiendo de muchos factores, especialmente el tipo de invasor.
Infección persistente. Después de la infección inicial con o sin síntomas, el virus completo o su genoma se mantienen en el organismo por tiempo prolongado meses, años o de por vida con o sin manifestaciones clínicas.
Las infecciones persistentes se pueden clasificar en:
Latente. El virus permanece oculto por tiempo variable luego de la primoinfección, pudiendo reactivarse una o más veces, con o sin manifestaciones clínicas
Crónica. El virus infecta en forma clínica o subclínica, y permanece en multiplicación continua aparente o inaparente, lo cual puede evidenciarse por técnicas de laboratorio.

Lenta. La primoinfección generalmente es asintomática, el virus no es detectable y muchos años después se manifiesta como un cuadro severo, progresivo, que en meses lleva a la muerte
Infección transformante. En este tipo de infección, el virus es capaz de infectar y permanecer en el hospedero sin necesariamente producir partículas virales en forma significativa. En algunos casos el genoma viral está presente en la célula, ya sea integrado o bien episomal (no integrado al DNA celular) y sólo parte de sus genes se traduce en proteínas virales.
MECANISMOS DE DEFENSA ANTIVIRAL
Todos los organismos vivos se defienden de las infecciones mediante procesos b1olog1cos que en su conjunto conforman la respuesta inmune (RI), capaz de detectar virus, bacterias, hongos y parásitos, de reconocerlos como ajenos, e idealmente, de eliminarlos. La respuesta inmune también puede causar daño si es desequilibrada o exagerada.La inmunidad innata, que es la primera defensa contra agentes extraños, actúa como barrera inespecífica o parcialmente específica. Es una respuesta rápida, pues sus componentes están preformados y presentes desde el nacimiento; en muchos casos es suficiente para eliminar el patógeno. Entre ellos se incluyen las barreras mecánicas, algunas células, citoquinas y el sistema del complemento.
Barreras mecánicasLa piel y las barreras mucosas (gastrointestinal, respiratoria, cilios, secreciones glandulares, conjuntiva) conforman el primer obstáculo que deben sobrepasar los agentes infecciosos para ingresar al hospedero. la entrada de los virus se facilita cuando se alteran estas barreras, lo que puede deberse al efecto de agentes químicos como el tabaco o a lesiones producidas por traumatismos, inyecciones, mordeduras de animales o picaduras de mosquitos.
SecrecionesOtros elementos efectores de la respuesta inmune innata son las defensinas, péptidos antimicrobianos producidos por las células epiteliales y leucocitos infectados presentes en las barreras epiteliales de bacterias, plantas y animales vertebrados e invertebrados. Muchas de ellas se expresan en forma constitutiva en las células y se almacenan en gránulos secretorios, mientras que otras se producen en respuesta a un estímulo inflamatorio.
Componente celularEstá constituido por los neutrófilos, los macrófagos, las células dendríticas (CD) y las células asesinas naturales (natural killers, NK).
NEUTROFILOS:- Conforman la mayor proporción de los polimorfonucleares circulantes, fagocitan a los agentes extracelulares, los atrapan en vesículas intracelulares unidas a la membrana (fagosomas) y los degradan mediante enzimas (proteasas, lisozimas, colagenasas) contenidas en gránulos citoplasmáticos.
-Procesan las partículas antigénicas y las presentan -en conjunto con las moléculas del complejo mayor de histocompatibilidad (CMH) celular- a los linfocitos T y/o B.
-Los neutrófilos son activados por el factor de necrosis tumoral {TNF) a y leucotrienos producidos por las células cebadas y otros neutrófilos.
MONOCITOS: -Circulantes migran a los tejidos transformándose en macrófagos, que producen componentes del sistema del complemento, prostaglandinas, interferones (IFN), interleuquinas (IL)1, IL -12, IL-6 y TNF a. A su vez, son estimulados por citoquinas liberadas por linfocitos T, como eiiFN y.
NK: -Son linfocitos sin receptores de tipo B o T, capaces de destruir células infectadas por un mecanismo similar al de los linfocitos T (LT) citotóxicos, liberando gránulos con perforinas que lesionan la membrana plasmática celular, permitiendo el ingreso de mediadores de la apoptosis como las granzimas, pero que a diferencia de éstos, no necesita el reconocimiento específico del antígeno.
-A través de su receptor CD16 actuan como citotoxicidad de células dependientes de anticuerpos, que se une a los anticuerpos que recubren una célula. Habitualmente, su actividad está controlada por una señal inhibitoria desencadenada por la interacción entre el CMH de clase 1 de las células normales y sus receptores KIR (ki!ler inhibítory receptors), por lo que se activan ante la ausencia de esta señal
CITOQUINAS
Sistema del complementoLa principal función del complemento es reconocer y destruir los agentes que infectan al hospedero. Este sistema incluye más de cuarenta moléculas plasmáticas o de membranas biológicas que se activan en cascada. Esta activación, fuertemente regulada, puede ocurrir por tres rutas: clásica, en la cual los complejos inmunes activan a C1 , alterna, con la activación de C3 mediante los factores 8, O y properdina, y de las lectinas, donde la unión de la lectina a la manosa (M8L) presente en el manto viral activa C4.La activación del complemento determina:• La lisis de células agresoras, alterando su permeabilidad por medio de canales creados por la inserción de los complejos de ataques de membrana (MAC) en membranas fosfolipídicas.• La opsonización de antígenos mediante la unión de C3b y C4b a la superficie de los agresores, haciéndolos más "apetecibles" para las células fagocíticas, que los destruirán.• La activación de linfocitos, al unirse C3d y C4d asociados a antígenos a los receptores de los LB.• La solubilización de complejos inmunes mediante el depósito de C3b y C4b en ellos.• La inflamación mediada por el aumento de la permeabilidad vascular por C3a, C4a y C5a.
La inflamación: Es un eficiente mecanismo defensivo a las infecciones, iniciado por la respuesta inmune innata, que alerta a la respuesta adaptativa del peligro. En ella participan células de distinto tipo -endoteliales, mastocitos, neutrófilos, monocitos y linfocitos- y mediadores solubles tales como el sistema del complemento, de la coagulación y citoquinas proinflamatorias como TNF-a e IL -1. ·

RESPUESTA INMUNE ADQUIRIDA
La inmunidad propiamente dicha juega un papel muy importante en el control de las infecciones primarias y secundarias. Su característica mas importante es que es especifica de antígeno y genera memoria inmunológica. La inmunidad específica puede ser de tipo humoral o celular.
Respuesta inmune adquirida humoral
La activación de los linfocitos B por antígenos proteicos
Figura 5·3. Acción del interferón. La célula infectada o por contacto con un virus secretará IFN, el que se unirá a su receptor específico en la superficie de una célula vecinano infectada. Esta sintetizará tres enzimas que inhibirán la replicación de los virus, creando un estado de resistencia antiviral.
Figura 5·4. Esquema de la respuesta inmune adaptativa humoral. (A) El antígeno viral es reconocido por el receptor del linfocito B (inmunoglobulina), lo que desencadena(B) su multiplicación (selección clona!) y (C) su maduración hacia células plasmáticas idénticas (D), las que sintetiza rán anticuerpos específicos contra el antígeno. (E)Algunos linfocitos B originan células de memoria que perduran y permiten una respuesta rápida en los contactos futu ros con el mismo antígeno.

ocurre en tres etapas.Reconocimiento. Es la interacción específica entre el antígeno y el receptor del LB -lgM o lgD- presente en la membrana. Este complejo antígeno-anticuerpo se internaliza, la proteína se degrada y los péptidos obtenidos se unen a moléculas del CMH clase 1, expresándose en la membrana del LB, que puede actuar como célula presentadora de antígeno (CPA) para los LT ayudadores (LTCD4+ o LTh). Este proceso ocurre entre tres a siete días después del ingreso del antígeno.Activación. El LB aumenta la expresión de sus moléculas coestimuladoras, las que se unen a CD28 del LTh, constituyendo la segunda señal necesaria para la activación del linfocito ayudador. Este linfocito activado expresa CD4QL –que interactúa con CD40 del LB- y secreta IL-2, la que a su vez induce al linfocito B a expresar más moléculas coestimuladoras, a proliferar y a diferenciarse, con la consiguiente expansión clonal del linfocito específico para el antígeno.Diferenciación. Se seleccionan los LB que producen inmunoglobulinas de alta afinidad frente a un antígeno y en el bazo.
Respuesta humoral
Respuesta inmune adquirida celularEn esta respuesta participan los linfocitos T CD8+ (LT cito tóxicos,LTc) que destruyen la célula infectada por virus, y los LTCD4+.LT citotóxico. La célula infectada procesa el antígeno viral y lo presenta al LTc, en conjunto con el CMH de clase 1 de la célula, puesto que el receptor específico del linfocito, LT citotóxico. La célula infectada procesa el antígeno viraly lo presenta al LTc, en conjunto con el CMH de clase 1 dela célula, puesto que el receptor específico del linfocito
LT ayudador. El LT ayudador es activado por células presentadoras de antígenos tales como macrófagos y LB, para lo cual es necesario que se unan el CMH de clase 11 de estas células con TCR, CD28 y CD40L del linfocito. Este prolifera, se diferencia y secreta citoquinas que varían según el tipo de antígeno y de citoquinas preponderantes en el medio
LT regulador. El TGF-~, el ácido retinoico e IL-2 estimulan la diferenciación del LT a linfocito regulador, el cual controla la respuesta inflamatoria liberando citoquinas como IL-1 O y TGF-~, que inhiben a los LT citotóxicos, LT ayudadores y a las CPA. A su vez, estos LTr son suprimidos por la IL-6.
La respuesta inmune antiviral se clasifica en inmunidad innata y adaptativa, ambas estrechamenterelacionadas y reguladas entre sí.• El conocimiento actual sobre la -respuesta inmune ha permitido identificar nuevas moléculas intermediarias:(1) los receptores que reconocen patrones moleculares (PRR), presentes en macrófagos,neutrófilos, linfocitos, células endoteliales, células dendríticas y NK, y (2) diversas citoquinas
Figura S-6. Respuestas inmune humoral primaria y secundaria. Ante al primer contacto, luego de una latencia, responde con lgM y luego lgG. La respuesta al segundo estímulo tiene una latencia muy corta, es alta, de mayor duración y en base a lgG.

que regulan las respuestas innata y adquirida.• La respuesta innata actúa rápidamente desde el inicio de la infección e incluye barreras epiteliales,células fagocíticas y mediadores circulantes como las proteínas del complemento y las citoquinas(IFN tipo 1).• La inmunidad adaptativa responde en forma específica al agente, entre siete a diez días postexposicióny comprende una respuesta humoral y una celular.• La respuesta humoral consiste en la producción de anticuerpos -inmunoglobulinas de distintasclases- por los linfocitos B maduros (células plasmáticas), que neutralizan y previenen la reinfección;además, dejan células de memoria para enfrentar futuras invasiones extracelulares del mismoagente.• La respuesta celular -mediada por linfocitos TCD8 y TCD4- es responsable del control y la eliminaciónde la infección intracelular.• Los virus han desarrollado diversos mecanismos de evasión de la respuesta inmune que les permitensobrevivir como especie y en algunos casos establecer infecciones persistentes.• Se han descrito muchos mecanismos mediante los cuales los virus pueden desencadenar fenómenosde autoinmunidad.