
Reglamento Trabajo Final - IIB-INTECH · Maite Lobo*, Giannina Carlevaro*, Luciano Melli*, ......
65
FACULTAD DE INGENIERIA Y CIENCIAS EXACTAS Y NATURALES Licenciatura en Ciencias Biológicas TRABAJO FINAL Expresión, localización y perfil antigénico de proteínas TolT de Trypanosoma cruzi. Comisión Evaluadora: Integrante N°1: Integrante N°2: Integrante N°3: Director del trabajo: Buscaglia Andrés Carlos. Codirectora del trabajo: Virginia Balouz. Alumna: Lobo Mabel Maite. Orientación: Ciencias de la Salud. Fecha: 00-08-17. Firma integrante N°1 de CE Firma integrante N°2 de CE Firma integrante N°3 de CE Firma integrante N°1 de CTF Firma integrante N°2 de CTF
Transcript of Reglamento Trabajo Final - IIB-INTECH · Maite Lobo*, Giannina Carlevaro*, Luciano Melli*, ......

FACULTAD DE INGENIERIA Y CIENCIAS EXACTAS Y NATURALES
Licenciatura en Ciencias Biológicas
TRABAJO FINAL
Expresión, localización y perfil antigénico de proteínas TolT de
Trypanosoma cruzi.
Comisión Evaluadora:
Integrante N°1:
Integrante N°2:
Integrante N°3:
Director del trabajo: Buscaglia Andrés Carlos.
Codirectora del trabajo: Virginia Balouz.
Alumna: Lobo Mabel Maite.
Orientación: Ciencias de la Salud.
Fecha: 00-08-17.
Firma integrante N°1 de CE Firma integrante N°2 de CE
Firma integrante N°3 de CE
Firma integrante N°1 de CTF
Firma integrante N°2 de CTF

Agradecimientos

Parte de los resultados obtenidos en este trabajo han sido incluidos
en:
Expresión, localización y perfil antigénico de proteínas TolT de Trypanosoma
cruzi.
Lobo M, Balouz V, Melli LJ, Ciocchini A, Agüero F, Buscaglia CA.
Reunión Anual de la Sociedad Argentina de Protozoología, Santa Fe, Argentina, 2016.
Molecular characterization of TolT antigens from Trypanosoma cruzi.
Maite Lobo*, Giannina Carlevaro*, Luciano Melli*, Virginia Balouz*, María Eugenia
Cortina, María de los Milagros Cámara, Gaspar E. Cánepa, Santiago Carmona, Oscar
Campetella, Andrés Ciocchini, Fernán Agüero, Juan Mucci, Carlos A. Buscaglia.
*Todos estos autores contribuyeron igualmente al trabajo.
Manuscrito en preparación.

Índice
ABREVIATURAS ............................................................................................................... 1
RESUMEN ........................................................................................................................... 3
ABSTRACT .......................................................................................................................... 5
INTRODUCCIÓN ............................................................................................................... 7
TRYPANOSOMA CRUZI Y ENFERMEDAD DE CHAGAS .......................................................... 7
TRYPANOSOMA CRUZI - FLAGELO ...................................................................................... 8
TOLT ................................................................................................................................. 9
OBJETIVOS ....................................................................................................................... 12
MATERIALES Y MÉTODOS ......................................................................................... 13
ANÁLISIS DE SECUENCIAS ............................................................................................... 13
STOCK DE PARÁSITOS Y LÍNEAS CELULARES .................................................................. 13
PURIFICACIÓN DE PROTEÍNAS ANCLADAS POR GPI ....................................................... 14
ENSAYO DE SECRECIÓN ................................................................................................... 14
DISEÑO Y VALIDACIÓN DE OLIGONUCLEÓTIDOS PARA REAL TIME QPCR ................... 15
EXTRACCIÓN DE ARN Y RETROTRANSCRIPCIÓN ........................................................... 15
CUANTIFICACIÓN DE LA EXPRESIÓN GÉNICA POR REAL-TIME QPCR .......................... 15
EXTRACCIÓN DE ADN Y AMPLIFICACIÓN GÉNICA POR PCR ........................................ 17
CLONADO ......................................................................................................................... 18
SECUENCIACIÓN .............................................................................................................. 19
PREPARACIÓN DE CÉLULAS COMPETENTES ................................................................... 19
CONDICIONES DE CULTIVO BACTERIANO ....................................................................... 19
EXPRESIÓN DE PROTEÍNAS TOLT RECOMBINANTES EN BACTERIAS .............................. 19
TÉCNICAS DE BIOLOGÍA MOLECULAR ............................................................................ 20
PÉPTIDOS ......................................................................................................................... 20
INMUNIZACIÓN ................................................................................................................ 20
DECLARACIÓN DE ÉTICA ................................................................................................. 21
ELECTROFORESIS EN GEL DE POLIACRILAMIDA Y WESTERN BLOT .............................. 21
ENSAYOS DE INMUNOFLUORESCENCIA INDIRECTA (IFI) ............................................... 21
POBLACIÓN EN ESTUDIO ................................................................................................. 22
MICROARREGLO DE PÉPTIDOS ....................................................................................... 22

ELISA ............................................................................................................................. 23
ANÁLISIS ESTADÍSTICO ................................................................................................... 23
RESULTADOS .................................................................................................................. 25
IDENTIFICACIÓN Y CARACTERIZACIÓN IN SILICO DE LA FAMILIA GÉNICA TOLT EN T.
CRUZI ............................................................................................................................... 25
FAMILIA GÉNICA TOLT .......................................................................................................... 25 PROTEÍNAS TOLT ................................................................................................................... 30
EXPRESIÓN DE ARNM Y PRODUCTOS DE TOLT DURANTE EL CICLO DE VIDA DE T.
CRUZI ............................................................................................................................... 35
EXPRESIÓN GÉNICA DE TOLT ................................................................................................ 35 EXPRESIÓN DE LAS PROTEÍNAS TOLT .................................................................................. 36 LOCALIZACIÓN SUBCELULAR DE LAS PROTEÍNAS TOLT .................................................... 38
IDENTIFICACIÓN Y MAPEO DE LOS EPITOPES B DE TOLT .............................................. 39
CONCLUSIONES Y DISCUSIÓN .................................................................................. 45
IDENTIFICACIÓN Y CARACTERIZACIÓN IN SILICO DE LA FAMILIA GÉNICA TOLT EN T.
CRUZI ............................................................................................................................... 45
EXPRESIÓN DE ARNM Y PRODUCTOS DE TOLT DURANTE EL CICLO DE VIDA DE T.
CRUZI ............................................................................................................................... 47
IDENTIFICACIÓN Y MAPEO DE LOS EPITOPES B DE LA FAMILIA TOLT .......................... 48
INFORMACIÓN SUPLEMENTARIA ........................................................................... 49
BIBLIOGRAFÍA ............................................................................................................... 56

Trabajo Final - Lobo Maite Abreviaturas
1
Abreviaturas
aa: Aminoácido/s.
ADN: Ácido desoxirribonucleico.
Ala: Alanina.
Arg: Arginina.
ARN: Ácido ribonucleico.
ARNm: Ácido ribonucleico mensajero.
BLASTN: Basic Local Alignment Search Tool N.
BSA: Albúmina sérica bovina.
Cys: Cisteína.
DAPI: 4’,6-diamino-2-fenilindol.
dNTPs: Desoxirribonucleótidos trifosfato.
ELISA (enzyme-linked immunosorbent assay): Ensayo por inmunoabsorción ligado a
enzimas.
FCS (fetal calf serum): Suero fetal de ternera.
FITC (fluorescein isothiocyanate): Isotiocianato de fluoresceína.
fw: Forward.
Glu: Ácido glutámico.
GPI: Glicosilfosfatidilinositol.
GST: Glutatión-S-Transferasa.
HAI: Hemoaglutinación indirecta.
HRP (horseradish peroxidase): Peroxidasa de rábano.
IFI: Inmunofluorescencia indirecta.
IgG: Inmunoglobulina G.
IPTG: Isopropil-β-D-tiogalactopiranósido.
kDa: KiloDalton.
LB: Medio de cultivo Luria-Bertani.
Leu: Leucina.
Lys: Lisina.
MASPs (mucin-associated surface proteins): Proteínas de superficie asociadas a mucinas.
MCS (multiple cloning site): Sitio de clonado múltiple.
MEM (minimum essential medium): Medio mínimo esencial.
NLS (nuclear localization signal): Señal de localización nuclear.

Trabajo Final - Lobo Maite Abreviaturas
2
nm: Nanómetros.
nt: Nucleótido/s.
OD (optical density): Densidad óptica.
PAGE (polyacrylamide gel electrophoresis): Electroforesis en gel de poliacrilamida.
PBS: Buffer fosfato salino.
PBSA: Albúmina sérica bovina en PBS.
PBST: Tween 20 en PBS.
PCR (polymerase chain reaction): Reacción en cadena de la polimerasa.
PFA: Paraformaldehído.
pI: Punto isoeléctrico.
PI-PLC (phosphoinositol specific phospholipase C): Fosfatidilinositol fosfolipasa tipo C.
PMSF (phenylmethylsulfonyl fluoride): Fluoruro de fenilmetilsulfonilo.
PVDF: Polifluoruro de vinilideno.
RE: Retículo endoplasmático.
rpm: Revoluciones por minuto.
rv: Reverse.
SD: Desvío estándar.
SDS: Dodecil sulfato sódico.
SDS-PAGE: PAGE en condiciones desnaturalizantes.
SP (signal peptide): Péptido señal.
TCA: Tricloro acético.
TcCLB: Trypanosoma cruzi clon CL Brener.
TCDM: Trypanosoma cruzi clon Dm28c.
Tc_MARK: Trypanosoma cruzi marinkellei.
Tc_SYL: Trypanosoma cruzi Sylvio X10/1.
TRSC58: Trypanosoma rangeli SC58.
U.A.: Unidades Arbitrarias.

Trabajo Final - Lobo Maite Resumen
3
Resumen
Trypanosoma cruzi, agente etiológico de la Enfermedad de Chagas, posee un ciclo de
vida digenético, que alterna entre un insecto vector y un mamífero hospedador y consta de
varias formas de desarrollo. A lo largo de su ciclo de vida, el parásito experimenta cambios
en sus caracteres morfológicos, fisiológicos y bioquímicos siendo de los más remarcables
la organización del aparato flagelar. En el año 1990 el grupo de Jerry Manning identificó
una familia de antígenos aparentemente relacionados, con expresión localizada en la
superficie del flagelo en contacto con el cuerpo celular de tripomastigotes y peso
molecular entre 34 y 45 kDa, dependiendo del aislamiento del parásito. El mismo grupo
luego describió un tándem genómico de 3 genes altamente relacionados (>97% de
identidad nucleotídica) como codificantes de estos antígenos. Los productos deducidos de
estos genes mostraron homología con las proteínas TolA de Escherichia coli y
Pseudomonas aeruginosa por lo que se los nombró TolT. Las proteínas TolT, además,
resultaron blanco de la respuesta inmune celular durante la infección. Ante la
disponibilidad de nueva información genética surgida de la secuenciación de múltiples
genomas de distintos aislamientos de T. cruzi y organismos filogenéticamente
emparentados, en esta Tesina de Licenciatura se reevaluó la complejidad de la familia
TolT con el objetivo de llevar a cabo una caracterización detallada a nivel de expresión,
localización sub-celular, y perfil antigénico. Una búsqueda de tipo BLASTN definió 22
secuencias con similitud significativa con TolT1 en el genoma de tripanosomátidos, los
que se agruparon en 3 subfamilias denominadas TolT-A, TolT-B y TolT-C en base a
criterios estructurales. Los miembros de la familia se disponen en tandem en orientación
cabeza-cola de a 3 unidades, poseen un péptido señal clivable en el extremo N-terminal,
seguido por una región madura más conservada y una señal de anclaje por GPI en el
extremo C-terminal y sufren modificaciones post-traduccionales de tipo N-glicosilación.
TolT-A (96-99% identidad nucleotídica entre ellos) y B (76-90% identidad nucleotídica
con TolT-A) presentan una región C-terminal muy conservada y un menor grado de
identidad hacia la región N-terminal, mientras que TolT-C (~47% de identidad
nucleotídica con TolT-A) es mucho más divergente. Se analizó la expresión a nivel de
ARNm mediante Real-Time qPCR observándose, para todos los estadios, una mayor
expresión de TolT-A, seguida por TolT-B y una mínima expresión de TolT-C. Un análisis
comparativo entre estadios indicó que TolT-A presenta mayor expresión en
tripomastigotes, mientras que TolT-B y C resultan más homogéneas. A nivel proteico

Trabajo Final - Lobo Maite Resumen
4
TolT-A se expresa en tripomastigotes y, en mucha menor medida, en amastigotes mientras
que TolT-B lo hace solo en tripomastigotes según ensayos de WB realizados con
anticuerpos específicos para las distintas subfamilias. La localización de TolT-A y B se
restringe al flagelo de tripomastigotes y al extremo posterior o anterior de amastigotes,
respectivamente. No se obtuvieron datos de expresión proteica para TolT-C en ningún
estadio del parásito. Estos resultados muestran una alta correlación entre los niveles de
mRNA y proteína para las distintas subfamilias de TolT. La antigenicidad de TolT está
dirigida mayormente contra el C-terminal compartido por los grupos TolT-B y A, cuya
prevalencia en la población de pacientes seropositivos resultó ser de ~42.9%. En algunos
pocos casos, también encontramos anticuerpos dirigidos contra la región N-terminal
diferencial de TolT-B y contra TolT-C. En conjunto, nuestros resultados indican que TolT
resulta ser una familia multigénica mucho más compleja que la descripta originalmente,
dentro de la cual pueden delinearse subfamilias de proteínas con patrones de expresión y
localización (y probablemente también función) distintos.

Trabajo Final - Lobo Maite Abstract
5
Abstract
Trypanosoma cruzi, etiological agent of Chagas disease, has a digenetic life cycle that
alternates between an insect vector and a vertebrate host, and consists of several
developmental forms. Along these transformations, the parasite undergoes remarkable
morphological, physiological and biological changes, including the organization of its
flagellar apparatus. In 1990, Jerry Manning’s group identified a family of apparently
related antigens that localized to the surface of the flagellum in contact with the cell body
of trypomastigotes. These molecules displayed a relative molecular mass between 34 and
45 kDa, depending on the parasite strain. Then, the same group described a genomic
tandem of 3 highly related genes (>97% nucleotide identity) coding for these antigens. The
predicted products encoded by these genes showed homology with the TolA proteins of
Escherichia coli and Pseudomonas aeruginosa, and they were accordingly designated
TolT. TolT proteins also resulted target antigens of the cellular immune response during T.
cruzi experimental infection. Due to the availability of new genetic information arising
from the sequencing of multiple genomes of different T. cruzi strains and other
phylogenetically related organisms, in this Thesis, we reevaluated the complexity of the
TolT family and provided a detailed characterization of its expression pattern, localization,
and antigenic profile. A BLASTN search defined significant similarity between TolT1 and
22 sequences in the genome of trypanosomatids, which were split into 3 subfamilies
named TolT-A, TolT-B y TolT-C based on structural criteria. Family members are
arranged in head-to-tail tandem of 3 genes in different chromosomes. Their encoded
products possess a cleavable N-terminal signal peptide followed by a more conserved
mature region and a GPI anchor signal at the C-terminal end, and undergo N-
glycosylation. TolT-A (96-99% nucleotide identity among them) and TolT-B (76-90%
nucleotide identity with TolT-A) have a highly conserved C-terminal region and a lower
degree of identity towards the N-terminal region, whereas TolT-C (~47% nucleotide
identity with TolT-A) is much more divergent. mRNA expression levels were quantified
by Real-Time qPCR showing, for all developmental stages, a higher expression of TolT-A,
followed by TolT-B and a minimal expression of TolT-C. A comparative analysis between
stages indicated that TolT-A shows higher expression in trypomastigotes, whereas TolT-B
and C are more homogeneous. At the protein level TolT-A is expressed in trypomastigotes
and, to a lesser extent, in amastigotes whereas TolT-B is restricted to trypomastigotes
according to the WB assays with specific antibodies for the different subfamilies. The

Trabajo Final - Lobo Maite Abstract
6
localization of TolT-A and TolT-B is restricted to the flagellum of trypomastigotes and to
the posterior or anterior end of amastigotes, respectively. No protein expression data were
obtained for TolT-C at any developmental stage of the parasite. These results show a high
correlation between mRNA and protein levels for the different TolT subfamilies. The
humoral immune response of T. cruzi-infected individuals is mainly directed against the C-
terminal end shared by TolT-B and A (seroprevalence 42.9%). In a few cases, we also
detected antibodies directed against the N-terminal differential region of TolT-B and
against TolT-C. Taken together, our results indicate that TolT is a much more complex
multigenic family than originally described, within which protein subfamilies with distinct
expression and localization patterns (and likely function) can be delineated.

Trabajo Final - Lobo Maite Introducción
7
Introducción
Trypanosoma cruzi y enfermedad de Chagas
Los protozoarios son eucariotas unicelulares de morfología y metabolismo complejos.
Son holozoicos, algunos poseen estructuras para la propulsión u otro tipo de movimiento
(pseudópodos, cilios o flagelo) y la mayoría se reproducen asexualmente por fisión binaria
y tienen vida libre, existiendo también comensales y parásitos [1]. Dentro de estos se
encuentra el orden Trypanosomatida, cuyos miembros son todos parásitos que causan
enfermedades en plantas y animales, e incluye especies de Leishmania, causante de
distintas leishmaniasis en todo el mundo, Trypanosoma brucei, responsable de la
enfermedad del sueño o tripanosomiasis africana y Trypanosoma cruzi (Familia
Trypanosomatidae), el agente etiológico de la Enfermedad de Chagas. Esta enfermedad es
endémica de América Latina, y se estima que afecta a unas 6-7 millones de personas [2].
En la actualidad no hay vacunas disponibles y las drogas utilizadas en el tratamiento son
tóxicas y no totalmente efectivas por lo que la enfermedad de Chagas se ha convertido en
un problema emergente de la salud pública global [3].
T. cruzi posee un ciclo de vida digenético, que alterna entre un insecto vector
(Subfamilia: Triatominae, Familia: Reduviidae, Orden: Hemiptera) y un mamífero
hospedador (doméstico o salvaje, incluyendo al hombre) y consta de varias formas de
desarrollo. Los tripomastigotes sanguíneos (Figura I.1.B) son las formas circulantes,
infectivas, no replicativas, presentes en el hospedador mamífero, y consumidas por el
insecto hematófago durante la ingesta de sangre. En el tracto digestivo anterior, se
diferencian extracelularmente a epimastigotes (Figura I.1.C), forma replicativa no
infectiva. Los epimastigotes se adhieren a la pared de la ampolla rectal, donde se
diferencian a tripomastigotes metacíclicos (bioquímica y morfológicamente distintos a los
tripomastigotes sanguíneos) (Figura I.1.B), forma no replicativa que transmite la infección
al hospedador mamífero al depositarse junto con la excreta en la herida resultante de la
picadura o en las mucosas. Los tripomastigotes metacíclicos infectan una amplia variedad
de células nucleadas en el hospedador [4] donde se diferencian intracelularmente a
amastigotes (Figura I.1.A), forma replicativa que tras varias rondas de división binaria se
transforman nuevamente en tripomastigotes sanguíneos. Estos se liberan al medio
extracelular, a la circulación sanguínea y a la circulación linfática siendo capaces de
invadir nuevas células o son eventualmente ingeridos por un insecto, cerrando así el ciclo

Trabajo Final - Lobo Maite Introducción
8
[5]. La repetición de esta ronda de invasión y replicación intracelular desencadena la fase
aguda de la enfermedad, caracterizada por alta parasitemia y parasitismo de tejidos,
mientras que la fase crónica se alcanza 45-60 días después de la infección original, cuando
la primera es controlada pero falla la eliminación completa de los parásitos [6] y se
caracteriza por un espectro de secuelas patológicas que incluyen complicaciones
gastrointestinales y/o cardíacas desde subclínicas hasta mortales.
A lo largo de las transformaciones sufridas por el parásito durante de su complejo ciclo
de vida hay cambios en los caracteres morfológicos, fisiológicos y bioquímicos [7], siendo
de los más remarcables la organización del aparato flagelar.
Figura I.1: Microscopía de contraste de fases donde se observan las formas amastigotes
(A), tripomastigotes (B) y epimastigotes (C) de T. cruzi. Imagen extraída de [8].
Trypanosoma cruzi - Flagelo
Una característica propia de T. cruzi, y tripanosomátidos en general, es la presencia de
un único flagelo. En tripomastigotes, éste se origina en el extremo posterior y posee una
porción que se encuentra anclada a lo largo de todo el cuerpo celular mientras otra,
pequeña, emerge libre al final. Esta disposición flagelar es la base morfológica de la
llamada ‘membrana ondulante’, típica del estadio tripomastigote [7]. En epimastigotes, por
el contrario, el flagelo se origina en el extremo anterior donde una pequeña fracción se
ancla al cuerpo y luego emerge como una organela libre. En amastigotes solo se
encuentran un flagelo incipiente y no funcional en términos de motilidad [7].
El flagelo es una estructura tipo cola que emerge del cuerpo celular a través del bolsillo
flagelar [9] y se encuentra unida lateralmente mediante la zona de anclaje flagelar, una
especialización de microtúbulos compleja conectada al citoesqueleto a través de 2
membranas de localización basal. Consiste en un axonema con configuración de 9 pares de
microtúbulos periféricos y 1 par central (9+2, conservado en eucariotas) y un bastón
paraflagelar [9,10].

Trabajo Final - Lobo Maite Introducción
9
La principal función del flagelo está asociada a la propulsión, aunque también se
demostró que, en tripanosomátidos, juega roles clave en la morfogénesis celular [11], la
interacción con el insecto vector [12], la segregación del ADN mitocondrial [13], la
división celular [14] y los mecanismos de endo- y exocitosis [9]. Existen múltiples
evidencias de que los componentes flagelares son muy antigénicos [15,16] y que se
asociarían a la inducción de respuestas inmunoprotectivas en infecciones experimentales
[17]. También se los ha vinculado al desarrollo de patologías del tipo autoinmune [18]
durante la enfermedad de Chagas.
Figura I.2: Micrografía electrónica de barrido de un epimastigote de T. cruzi. El
flagelo (puntas de flecha), el cual emerge del bolsillo flagelar (flecha), está anclado
al cuerpo celular a lo largo de la zona de anclaje flagelar. Imagen extraída de [19].
TolT
En el año 1990, en un intento de identificar nuevos antígenos del parásito, el grupo de
Jerry Manning produjo hibridomas derivados de células de bazo de ratones infectados con
la cepa Perú de T. cruzi y evaluó la reactividad de los anticuerpos monoclonales
producidos. Uno de estos, denominado mAb20H1, reconoció un antígeno (20H1) cuya
expresión resultó exclusiva del estadio tripomastigote (Figura I.3.A) y su localización
restringida a la superficie del flagelo en contacto con el cuerpo celular (Figura I.4).
Mediante WB se observó que este anticuerpo reconocía varias bandas de peso molecular
aparente similar, entre 34 y 45 kDa dependiendo de la cepa estudiada (Figura I.3.B),
sugiriendo que se trataba de una familia de antígenos relacionados con polimorfismos de
secuencia y/o modificaciones post-traduccionales distintas [16]. Más tarde, el mismo grupo
describió en la cepa Esmeraldo que estos antígenos estaban codificados por al menos 3
genes altamente relacionados (>97% de identidad nucleotídica) en el genoma de T. cruzi y
que las proteínas deducidas a partir de estos genes tenían homología con las proteínas
TolA de Escherichia coli (32% de identidad en una superposición de 229 aminoácidos) y
Pseudomonas aeruginosa (34% de identidad en una superposición de 195 aminoácidos)
por lo que les fue asignado el nombre de TolT [20]. Las proteínas TolT, además, resultaron

Trabajo Final - Lobo Maite Introducción
10
ser blanco de la respuesta inmune celular durante la infección experimental, por lo que se
evaluó su potencial como reactivo vacunal [20]. Posteriormente, un miembro de la familia
TolT, aunque levemente divergente a las proteínas TolT1-3 descriptas originalmente, fue
postulado como potencial reactivo para el diagnóstico serológico de la enfermedad de
Chagas [21].
Figura I.3: Análisis de WB revelados
con el anticuerpo mAb20H1. A: (a)
marker de peso molecular; (b) lisado de
epimastigotes cepa Peru; (c) lisado de
tripomastigotes cepa Peru. B: (a) marker
de peso molecular; (b) lisado de
epimastigotes y lisado de tripomastigotes
cepa CL (c), Esmeraldo (d), Peru (e),
Silvio (f) e Y (g) de T. cruzi. Imagen
extraída de [16].
Trabajo Final - Lobo Maite Introducción
11
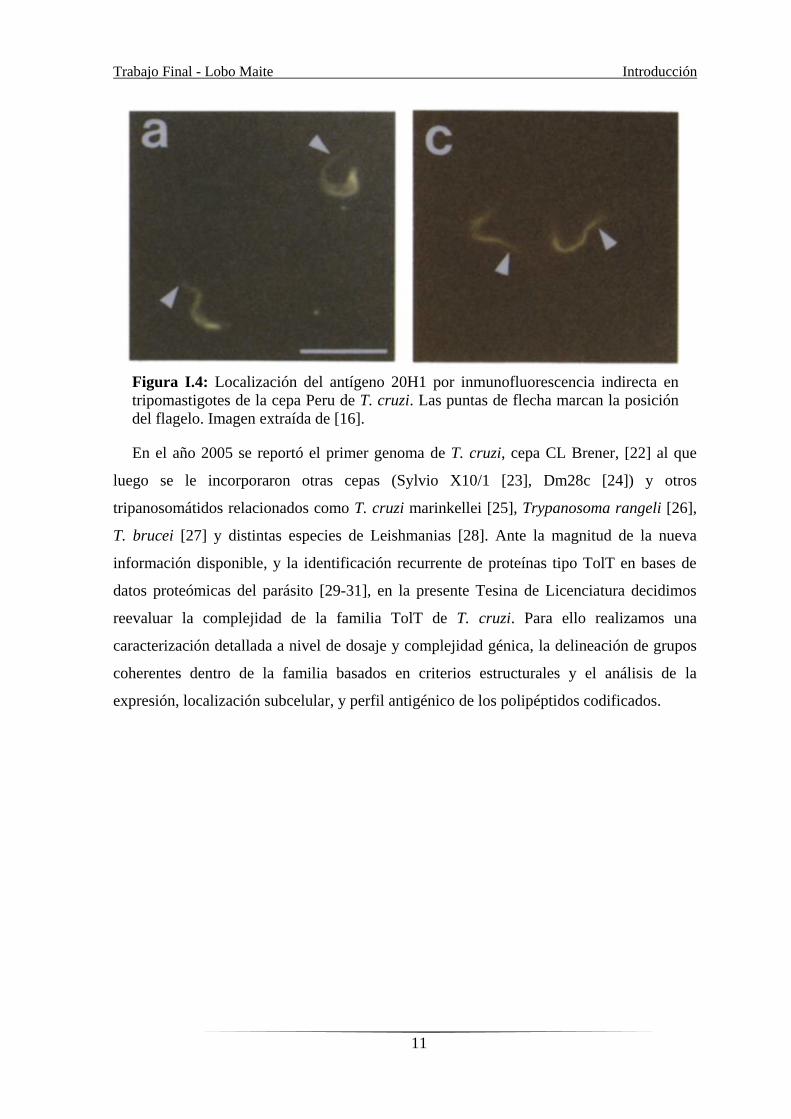
Figura I.4: Localización del antígeno 20H1 por inmunofluorescencia indirecta en
tripomastigotes de la cepa Peru de T. cruzi. Las puntas de flecha marcan la posición
del flagelo. Imagen extraída de [16].
En el año 2005 se reportó el primer genoma de T. cruzi, cepa CL Brener, [22] al que
luego se le incorporaron otras cepas (Sylvio X10/1 [23], Dm28c [24]) y otros
tripanosomátidos relacionados como T. cruzi marinkellei [25], Trypanosoma rangeli [26],
T. brucei [27] y distintas especies de Leishmanias [28]. Ante la magnitud de la nueva
información disponible, y la identificación recurrente de proteínas tipo TolT en bases de
datos proteómicas del parásito [29-31], en la presente Tesina de Licenciatura decidimos
reevaluar la complejidad de la familia TolT de T. cruzi. Para ello realizamos una
caracterización detallada a nivel de dosaje y complejidad génica, la delineación de grupos
coherentes dentro de la familia basados en criterios estructurales y el análisis de la
expresión, localización subcelular, y perfil antigénico de los polipéptidos codificados.

Trabajo Final - Lobo Maite Introducción
12
Objetivos
I. Caracterizar in silico a la familia génica TolT en T. cruzi.
II. Determinar la expresión de ARNm y proteínas TolT durante el ciclo de vida de T.
cruzi.
III. Identificar, mapear y validar los epitopes B de TolT.

Trabajo Final - Lobo Maite Materiales y métodos
13
Materiales y Métodos
Análisis de secuencias
Se llevaron a cabo búsquedas de similitud de secuencias utilizando BLASTN, la cual
explora una base de datos nucleotídica utilizando como señuelo una secuencia nucleotídica
en TriTrypDB (http://tritrypdb.org/tritrypdb/) con la configuración definida por default. En
nuestro caso, usamos como señuelo la secuencia completa del gen TolT1 (GenBank:
AF099099.1) Se definieron como significativos aquellos resultados con un E-value menor
que 10-5. Las secuencias codificantes de TolT en CL Brener fueron descargadas de
TriTrypDB. Con ellas se llevó a cabo un alineamiento múltiple utilizando el programa T-
Coffee [32] y, tras el curado manual, la construcción de un árbol filogenético basado en el
método de distancias Neighbor-Joining [33] con un análisis de bootstrap [34] de 1000
repeticiones lo que permitió la separación de las proteínas TolT en 3 grupos robustos.
Luego se procedió a realizar nuevas búsquedas, utilizando ahora como señuelo un
miembro representativo de cada uno de los grupos definidos (TolT-A: TcCLB.508767.20,
TolT-B: TcCLB.510433.20, TolT-C: TcCLB.506815.20), se agregaron los nuevos matches
significativos al alineamiento (Figura S.1) y se reconstruyó el árbol tal como se describió
previamente. El árbol final obtenido fue visualizado y editado gráficamente para su
presentación con el programa iTOL [35]. Para cada grupo se definió el porcentaje de
identidad con TolT1 y la ubicación en el genoma, así como la masa, el pI y la presencia de
SP a partir de información provista por TriTrypDB, mientras que las señales de anclaje por
GPI se predijeron usando el algoritmo PredGPI [36]. Además se realizaron predicciones de
modificaciones post-traduccionales: fosforilación, N-glicosilación, O-glicosilación y
palmitoilación utilizando los algoritmos NetPhos 3.1 [37], NetNGlyc 1.0, NetOGlyc 4.0
[38] y CSS-Palm 3.0 [39] respectivamente. También se evaluó la homología a las TolA
bacterianas mediante búsquedas de tipo BLASTP desde el servidor UniProt
(http://www.uniprot.org/blast/) utilizando como señuelo cada una de las secuencias de
TolT identificadas, con restricción a la base de datos de Bacteria y la configuración
determinada por default. Para la generación de logos se utilizó WebLogo (disponible en
http://weblogo.berkeley.edu/logo.cgi).
Stock de parásitos y líneas celulares
Diferentes estadios de desarrollo de parásitos de la cepa CL Brener se obtuvieron y
purificaron como se describió previamente [40]. Brevemente, los epimastigotes se

Trabajo Final - Lobo Maite Materiales y métodos
14
crecieron a 28°C en medio BHT suplementado con 10% de FCS (Gibco) inactivado por
calor. Los tripomastigotes y amastigotes derivados de células se cosecharon (a distintos
tiempos post-infección) del sobrenadante de células Vero libres de micoplasmas (ATCC)
crecidas a 37°C y 5% de CO2 en MEM suplementado con 10% de FCS, 0.292 g/L de L-
glutamina, 100 IU/mL de penicilina y 100 µg/mL de estreptomicina (de Laboratorios
Gibco). En todos los casos cada estadio tenía un nivel de pureza ≥ 90%.
Purificación de proteínas ancladas por GPI
Pellets compuestos por 1x108 tripomastigotes de T. cruzi CL Brener fueron
homogeneizados en 2 ml de buffer GPI [Tris/HCl 10 mM pH 7.4, NaCl 150 mM, Triton
X-114 2%, PMSF 1 mM y un cocktail inhibidor de proteasas (Sigma)] en hielo por 1 hora
[41]. El homogenato se centrifugó a 8800 g y 0°C por 10 minutos y el sobrenadante (S1) se
guardó a -20°C durante 24 horas. El pellet (P1) se lavó con 1 ml de buffer A (Tris-HCl 10
mM pH 7.4, NaCl 150 mM, TX-114 0.06%, PMSF 1 mM) y se guardó. El S1 fue
descongelado y sometido a una separación de fases a 37°C por 10 minutos seguida de una
centrifugación a 3000 g por 3 minutos a temperatura ambiente. Se recolectó la fase
superior (S2) y a la fase rica en detergente se le realizó una re-extracción con 1 ml de
buffer A. La fase superior (S3) se recolectó y la fase rica en detergente se sometió a una re-
extracción con 1 ml de buffer A, se homogeneizó, se incubó a 0°C durante 30 minutos y se
centrifugó a 18000 g por 10 minutos a 0°C. El pellet (P2) se lavó con 1 ml de buffer A y se
guardó mientras el sobrenadante (S4) se sometió a una nueva fase de separación. La fase
superior (S5) fue recolectada y la inferior, fase rica en detergente y enriquecida con
proteínas ancladas a GPI, se recolectó como la fracción GPI (GPI). Las distintas fracciones
(P1, S1, S2, S3, S4, S5 y GPI) se analizaron por WB.
Ensayo de secreción
Se cosecharon 3x108 tripomastigotes derivados de células y se eliminó el medio por
centrifugación. Se resuspendieron en 5 ml de PBS suplementado con 2% de glucosa estéril
y se incubaron en estufa a 37°C durante 2 horas. Luego se centrifugó a 7000 rpm durante
10 minutos, se separó el sobrenadante (S) del pellet (P) y el primero se filtró con una
membrana de 2 µm y se precipitó con Ácido Tricloroacético (TCA) [40]. Brevemente, se
agregó 1 volumen (100% p/v) de TCA cada 4 volúmenes de muestra, se incubó 10 minutos
a 4°C, se centrifugó a 14000 rpm durante 5 minutos y se removió el sobrenadante. Se lavó
el pellet con 200 µl de acetona fría, se centrifugó a 14000 rpm durante 5 minutos, dos

Trabajo Final - Lobo Maite Materiales y métodos
15
veces y por último se dejó secar el pellet a 95°C durante 10-15 minutos. Pellet y
sobrenadante se analizaron mediante WB.
Diseño y validación de oligonucleótidos para Real time qPCR
Se diseñaron 3 pares de oligonucleótidos en regiones específicas de TolT-A, TolT-B y
TolT-C. Para el diseño se utilizaron las secuencias TcCLB.506617.10. TcCLB.510433.20
y TcCLB.504277.30 respectivamente, las cuales se alinearon y se buscaron las regiones
más divergentes entre ellas. A su vez, se cargaron las secuencias de los genes completos en
el software PerlPrimer v1.1.21 para el diseño de oligonucleótidos de Real Time. Se
eligieron aquellos que hibridaban en las secuencias de interés para minimizar
amplificación cruzada. Cada par de oligonucleótidos fue utilizado en reacciones de PCR a
punto final para evaluar la amplificación de un solo producto de tamaño esperado en geles
de agarosa y la ausencia de producto en el control sin templado. Los productos obtenidos
en cada reacción fueron clonados en un plásmido pGEM-Teasy (Promega) y secuenciados
usando los primers T7 y SP6, los cuales flanquean al MCS (Figura S.2). Luego de esta
validación se procedió con la realización y cuantificación de la Real-Time qPCR.
Extracción de ARN y retrotranscripción
El ARN total de tripomastigotes, amastigotes extracelulares y epimastigotes de T. cruzi
CL Brener (4x108 de parásitos en cada caso) se extrajo utilizando el reactivo TRIzol
(Invitrogen, USA) y se trató con ADNasa I (Sigma-Aldrich, USA) [40]. La cantidad y
pureza del ARN se estimó mediante espectrofotometría a 260/280 nm utilizando un
espectrofotómetro UV-VIS Micro-volumen (Thermo Scientific™ NanoDrop™). La
integridad se verificó por electroforesis en gel de agarosa 2.5% (p/v). Todas las reacciones
de transcripción reversa partieron de 3 µg de ARN utilizando primer oligo-dT 10 µM,
dNTPs 10 mM y la transcriptasa reversa SuperScript™ II (Sigma), de acuerdo a [40].
Cuantificación de la expresión génica por Real-Time qPCR
El ensayo de Real-Time qPCR se realizó en el Sistema de PCR en tiempo real
7500 de Applied BioSystems, utilizando la Master Mix KAPA SYBR® FAST qPCR
(Kapa Biosystems, USA). Las condiciones para la Real-Time qPCR fueron: tras una pre
incubación de 2 minutos a 50°C, un paso inicial de desnaturalización a 95°C por 10
minutos, seguidos por 40 ciclos de desnaturalización, alineamiento y extensión a 95°C por
15 segundos y 63°C por 1 minuto. Para monitorear la especificidad de los oligonucleótidos

Trabajo Final - Lobo Maite Materiales y métodos
16
se realizaron las curvas de disociación (melting curve) al finalizar cada experimento
(Figura S.3). Las reacciones se hicieron por triplicado utilizando 2 µl de ADNc de
tripomastigotes, amastigotes y epimastigotes como templado en un volumen total de 10 µl.
Los oligonucleótidos utilizados se muestran en la Tabla M.1 La cuantificación relativa de
los genes de interés se realizó considerando la eficiencia de la reacción la cual se calculó
utilizando el approach matemático LinReg PCR V12.17 [42,43]. Éste calcula la eficiencia
de cada curva de fluorescencia vía regresión lineal en la parte exponencial de la misma.
Como control endógeno se usaron los genes Calmodulina y GAPDH de T. cruzi. Para
calcular la relación entre el gen de referencia y el gen target, incluyendo la corrección de la
eficiencia, se utilizó la fórmula modificada de Pfaffl, 2001 [44].
Tabla M.1: Oligonucleótidos utilizados para el análisis de expresión génica de TolT
mediante Real-Time qPCR.
Nombre Gen target
(TcCLB.) Secuencia (5’-3’)
Producto
amplificado
(pb)
TcTolTART Fw
TcTolTART Rv
TolT-A
506617.10
506617.20
CTGGGCTTGAGAAAGAAGCAG
CTGGAATTCCTGCTCCTCCT 229
TcTolTBRT Fw
TcTolTBRT Rv
TolT-B
508767.10
510433.20
AGGCTACAAGGATGAGCAGG
CCAACCTTAGTCTTCTCGTCC 175
TcTolTCRT Fw
TcTolTCRT Rv
TolT-C
504277.30
CCAAGAAATACGCCGAAGAG
TTTGTCAGTCGAATCTGCAGAG 241
TcCalm Fw
TcCalm Rv
TcCalmodulina
506391.10
506391.20
507483.30
507483.39
507483.50
CCCGACGGAGGCGGAGCTGC
GTCCACGTCGGCCTCGCGGA
268

Trabajo Final - Lobo Maite Materiales y métodos
17
Extracción de ADN y amplificación génica por PCR
El ADN genómico de T. cruzi (cepa CL Brener) se purificó por el método de fenol-
cloroformo [45].
Los fragmentos génicos de interés, descriptos en la tabla M.2 y representados en la
Figura R.9, se amplificaron por PCR utilizando 1-10 ng de ADN como templado, Taq
ADN polimerasa de alta fidelidad (Invitrogen) y los oligonucleótidos correspondientes
detallados en la tabla M.3. Los oligonucleótidos se diseñaron tratando de evitar los
polimorfismos existentes entre las distintas secuencias de una misma subfamilia de TolT.
Tabla M.2: Fragmentos de interés para la generación de proteínas recombinantes.
Gen
target Fragmento Desde - Hasta*
Producto
(pb)
GST-Producto
(kDa)
TolT-A TolT-A N-terminal F54 - T174 360 39
TolT-B
TolT-B N-terminal
TolT-B Centro
TolT-B C-terminal
TolT-B Full
Q61 - S103
S97 - L162
G155 - R260
Q61 - R260
126
195
315
597
31
33
38
48
TolT-C TolT-C A83 - D313 690 51
*Se utiliza el código de una letra para definir el aminoácido.
Tabla M.3: Oligonucleótidos utilizados para la amplificación de los fragmentos de interés.
Nombre Descripción* Secuencia nucleotídica (5’ a 3’)**
TolT-A Fw Fw para amplificar
TolT-A desde F54,
con sitio BamHI.
GCggatccTTTGACTGGGCATTCAAG
TolT-A Rv Rv con Stop para
amplificar TolT-A
hasta T174, con
sitio XhoI.
GActcgagCTAAGTATGATTTGCCGCCTT
TolT-B Fw
Fw para amplificar
TolT-B desde Q61,
con sitio BamHI.
CGggatccCAGGAGTACGCTGATGAGGCT
TolT-B Rv Rv para amplificar
TolT-B hasta S103,
CGgaattcGCTCATCCTTGTAGCCTCAGA
TcGAPDH Fw
TcGAPDH Rv
TcGAPDH
506943.50
506943.60
509065.60
509065.70
GTGCGGCTGCTGTCAACA
AAAGACATGCCCGTCAGCTT
178

Trabajo Final - Lobo Maite Materiales y métodos
18
con sitio XhoI.
TolT-B 2 Fw Fw para amplificar
TolT-B desde S97,
con sitio BamHI.
CGggatccTCTGAGGCTACAAGGATGAGC
TolT-B 2 Rv Rv para amplificar
TolT-B hasta L162,
con sitio XhoI.
CGgaattcCAACAGCTCCGGTCCACTTCC
TolT-B 3 Fw Fw para amplificar
TolT-B desde G155,
con sitio BamHI.
CGggatccGGAAGTGGACCGGAGCTGTTG
TolT-B 3 Rv Rv para amplificar
TolT-B hasta A260,
con sitio XhoI.
CGgaattcGCGCTGCGTCATTCCCCTTCT
TolT-C Fw Fw para amplificar
TolT-C desde A83,
con sitio BamHI.
GAggatccGCGACAATGCAATGCATG
TolT-C Rv Rv con Stop para
amplificar TolT-C
hasta D313, con
sitio XhoI.
GActcgagCTAACTCCCACTGCGTCTGACTC
pGEX Fw Universal pGEX Fw GGGCTGGCAAGCCACGTTTGGT
pGEX Rv Universal pGEX Rv CCGGGAGCTGCATGTGTCAGA
T7 Universal T7 Fw AATACGACTCACTATAGGG
SP6 Universal SP6 Rv ATTTAGGTGACACTATAG
*Se utiliza el código de una letra para definir el aminoácido.
** Los sitios blanco de las enzimas de restricción se muestran subrayados.
Clonado
Los amplicones obtenidos fueron purificados de acuerdo al protocolo de desalado
QIAEX II, clonados en el vector pGEM-Teasy y utilizados para transformar células DH5α
F’IQ™. Además del rastreo por color (azul/blanco), las colonias se chequearon por PCR
utilizando oligonucleótidos que se unen a las regiones codificantes de los promotores de
las ARN polimerasas SP6 y T7 (Figura S.2).
Los amplicones clonados en el vector pGEM-Teasy se liberaron tras la digestión con las
enzimas BamHI y XhoI (Fermentas) y se ligaron a un vector pGEX2T con el MCS
modificado con los sitios necesarios para la compatibilidad con los insertos. Los plásmidos

Trabajo Final - Lobo Maite Materiales y métodos
19
de la línea pGEX permiten expresar proteínas como fusión al extremo Carboxilo terminal
de la proteína Glutatión-S-Transferasa de Schistosoma japonicum (GST). Se transformaron
bacterias competentes DH5α F’IQ™ con los distintos plásmidos y se realizó el rastreo de
las colonias por PCR utilizando los oligonucleótidos pGEX Fw y pGEX Rv. Los
plásmidos se purificaron a partir de cultivos de colonias positivas crecidos toda la noche a
37°C en agitación y se analizaron por secuenciación. Para evaluar la especificidad de los
antisueros generados (ver debajo), los constructos también se clonaron en el plásmido
pET28, que provee fusión N-terminal a un tag de 6 residuos histidina en tándem. La
inducción y purificación de estas proteínas se realizó de acuerdo a lo descripto [46].
Secuenciación
La secuenciación de ADN fue realizada por Macrogen, por el método de Sanger.
Preparación de células competentes
Las células competentes de las cepas E. coli DH5α F’IQ™ y E. coli BL21-CodonPlus
(DE3)-RP fueron preparadas por el método Inoue [47].
Condiciones de cultivo bacteriano
Las cepas de E. coli fueron cultivadas a 37ºC en agitador rotatorio (200-250 rpm) o en
placas de Petri con 1.5% de agar en medio LB. Según la cepa, el medio se suplementó con
los siguientes antibióticos: ampicilina 100 μg/mL o cloranfenicol 34 μg/mL.
Expresión de proteínas TolT recombinantes en bacterias
Se transformaron bacterias competentes E. coli BL21-CodonPlus (DE3)-RP con los
plásmidos resultantes. La cepa usada posee genes extra que codifican para los ARN de
transferencia menos representados en el genoma bacteriano, permitiendo mayores niveles
de expresión de proteínas codificadas por genes heterólogos. Se cultivaron hasta llegar a
una OD600 nm de 0.6, luego se indujo la expresión de las proteínas con IPTG (Fermentas)
250 µM y se incubó 16 horas a 28ºC en agitación. La purificación se realizó a partir de la
fracción soluble por cromatografía de afinidad hacia el dominio GST de fusión, utilizando
columnas de Glutatión-Sefarosa, siguiendo las indicaciones del fabricante (Glutathione
Sepharose 4 Fast Flow, GE Healthcare). Las proteínas se dializaron contra PBS 0.5 X en
columna NAP10 y se concentraron en SpeedVac (Savant ISS110) alcanzando
aproximadamente la mitad del volumen de elución. Se verificó la pureza e integridad de
las proteínas obtenidas en geles de poliacrilamida 12% teñidos con Coomassie Blue

Trabajo Final - Lobo Maite Materiales y métodos
20
(Figura S.4). La cuantificación se realizó por Bradford según las indicaciones del
fabricante (BioRad) usando como estándar soluciones de concentraciones conocidas de
BSA [48].
Técnicas de biología molecular
Las técnicas de restricción, ligación y purificación de ADN plasmídico, PCR,
generación y transformación de bacterias competentes por el método Inoue son de uso
corriente en el laboratorio. Se realizaron según se indica en Sambrook, J. & Russell, D. W.
Molecular Cloning: A Laboratory Manual, 2001 y las indicaciones de los fabricantes de los
diferentes kits y/o insumos utilizados para cada caso.
Péptidos
El péptido correspondiente a los residuos K144 a H163 de TolT-C (5’-
©KAAVDADTAALAALLEVLQH-3’) se mandó a sintetizar a GenScript. Se agregó un
residuo de cisteína en el extremo N-terminal para el acoplamiento a la proteína KLH
(maleimide) (Sigma), lo cual se realizó según las instrucciones del fabricante [49].
Inmunización
Para la obtención de antisueros (Tabla M.4), 100 µg de cada proteína GST-TolT o del
péptido-KLH fueron emulsionados en adyuvante completo de Freund e inyectados
intraperitonealmente en ratones BALB/c (de entre 60 y 90 días de edad) [50]. A los 30 y
45 días post inmunización se realizaron boosters usando 50 µg del correspondiente
inmunógeno emulsionado en adyuvante incompleto. Finalizado el protocolo, se sangraron
los ratones a blanco, se preparó el suero y se midió el título contra cada proteína en un
ensayo de ELISA (utilizando proteínas fusionadas a Histidina para no considerar los títulos
de anticuerpos contra la proteína GST).
Tabla M.4.: Antisueros utilizados para el estudio de la expresión a nivel proteico de TolT
y análisis de seroprevalencia.
Nombre Descripción*
α TolT-A N-terminal Antisuero de ratón inmunizado con TolT-A F54 a T174
fusionada a GST
α TolT-B N-terminal Antisuero de ratón inmunizado con TolT-B Q61 a S103
fusionada a GST
α TolT-B Centro Antisuero de ratón inmunizado con TolT-B E97 a L162

Trabajo Final - Lobo Maite Materiales y métodos
21
fusionada a GST
α TolT-B C-terminal Antisuero de ratón inmunizado con TolT-B G155 a A260
fusionada a GST
α TolT-B Full Antisuero de ratón inmunizado con TolT-B Q61 a A260
fusionada a GST
α TolT-C Antisuero de ratón inmunizado con TolT-C K144 a H163
acoplado a KLH: ©KAAVDADTAALAALLEVLQH
*Se utiliza el código de una letra para definir el aminoácido.
Declaración de ética
La inmunización, sangrado y manejo de los animales se llevó a cabo en estricta
conformidad con las recomendaciones del Comité Institucional para el Cuidado y Uso de
Animales de Experimentación (CICUAE-UNSAM). El protocolo fue aprobado por el
mismo y se hizo todo el esfuerzo para minimizar el número y sufrimiento de los animales.
Electroforesis en gel de poliacrilamida y Western blot
Las electroforesis en gel se realizaron utilizando SDS-PAGE 12% donde se corrieron
las muestras de interés. Para WB, la corrida se transfirió a una membrana de PVDF (GE
Healthcare), se la incubó durante toda la noche a 4°C en agitación con el antisuero
correspondiente (dilución 1:500 en PBST 0.05% con 5% leche) seguido por el anticuerpo
secundario, anti IgG de ratón, conjugado a HRP (Sigma) (dilución 1:5000 en PBST 0.05%
con 5% leche) y por último se reveló usando el sustrato quimioluminiscente SuperSignal
West Pico (Pierce).
Ensayos de Inmunofluorescencia indirecta (IFI)
Para los ensayos de IFI, formas extracelulares de parásitos se cosecharon y lavaron en
PBS, se adhirieron 1x106 a cubreobjetos cubiertos con poli-L-lisina (Sigma), se fijaron por
30 minutos en PFA 4% en PBS, se bloquearon por 30 minutos en BSA (Sigma) 4% en
PBS (PBSA) suplementado con saponina al 0.5% y se incubó con el anticuerpo
correspondiente diluido en PBSA [51]. Tras extensivos lavados en PBS, se agregó el
anticuerpo secundario conjugado a Alexa Fluor (Molecular Probes) y DAPI (10 µg/ml)
para la tinción de los núcleos por último se montó sobre un portaobjetos con el reactivo
FluorSave (CalBiochem). Para evaluar la reactividad en estadios intracelulares, 10000
células Vero se colocaron en cubreobjetos y tras incubarlas toda la noche se las infectó con
1x106 tripomastigotes CL Brener por cubreobjeto. Tras 16 horas, las células infectadas se
lavaron con PBS, se fijaron y procesaron para IFI. En ambos casos, previo al bloqueo, se

Trabajo Final - Lobo Maite Materiales y métodos
22
incubó con cloruro de amonio 25 mM en PBS para disminuir la fluorescencia basal. Los
antisueros para TolT-A y TolT-B se utilizaron en diluciones 1:50 y 1:25, respectivamente.
Las imágenes se obtuvieron con un microscopio de epi-fluorescencia Nikon Eclipse 80i
acoplado a una cámara DS-Qi1 CCD y fueron procesadas con el programa ImageJ [51].
Población en estudio
El panel de sueros de pacientes infectados (entre 10 y 65 años de edad, de ambos sexos,
cursando la fase crónica de la enfermedad, sin sintomatología asociada) fue provisto por el
Instituto Nacional de Parasitología “Dr. Mario Fatala Chabén” (Buenos Aires, Argentina)
[50]. Las muestras de sangre se obtuvieron por venipuntura y se analizó la presencia de
anticuerpos específicos contra T. cruzi por dos kits comerciales disponibles: ELISA,
utilizando homogenato total de parásito y HAI (ambos de Wiener lab, Argentina). Los
sueros de individuos no infectados con T. cruzi se obtuvieron de distintos bancos de
sangre: Fundación Hemocentro Buenos Aires (Buenos Aires, Argentina, Hospital de
Enfermedades Infecciosas ‘Dr. Francisco Javier Muñiz’ (Buenos Aires, Argentina),
Hospital Italiano de Buenos Aires (Buenos Aires, Argentina) y Hospital Municipal ‘Dr.
Diego E. Thompson’ (San Martín, Buenos Aires, Argentina).
Microarreglo de péptidos
En el contexto de un proyecto tendiente a la identificación y mapeo de epitopes B en las
proteínas de T. cruzi a gran escala [52], se probó con sueros de pacientes Chagásicos
crónicos la secuencia de distintos miembros de la familia TolT (TolT-A:
TcCLB.506617.10; TolT-B: TcCLB.510433.20 y TolT-C: TcCLB.504277.30) en
microarreglos peptídicos, los cuales presentaban la totalidad de la secuencia anotada de
estas proteínas representada por péptidos de 15 aminoácidos, con un solapamiento parcial
de 14. Cada muestra se conformó por IgG purificadas a partir del suero de 5 individuos
distintos (3 μl de cada uno). Los arreglos de péptidos se ensayaron secuencialmente,
primero con una muestra negativa (IgG purificada a partir de 5 individuos sanos) y luego
con la muestra positiva (IgG purificada a partir de 5 individuos infectados con T. cruzi).
Con este diseño experimental, se obtuvieron 2 conjuntos de datos para cada experimento:
uno correspondiente a la lectura de individuos sanos (control negativo) y uno
correspondiente a la señal acumulada de las muestras negativas + positivas, a partir de los
cuales se determinó la señal “definitiva” por sustracción. Integrando todas las señales del
microarreglo (que incluía controles positivos y negativos) se puntuó la reactividad para

Trabajo Final - Lobo Maite Materiales y métodos
23
cada péptido. Se graficaron los valores medios de fluorescencia obtenidos luego de la
sustracción de cada uno de los péptidos [48].
ELISA
Se realizó la adsorción del antígeno a placas de 96 pocillos (Nunc MaxiSorp) en buffer
bicarbonato 0.05 M pH 9.6 a 4°C durante 16 horas. A continuación se bloqueó por una
hora a 37°C, se lavó y se incubó con el anticuerpo primario una hora a 37ºC. Pasado el
tiempo de incubación, se lavó y se incubó una hora con el anticuerpo secundario
correspondiente. Finalmente se lavó el anticuerpo secundario y se procedió con el revelado
y finalización de la reacción para luego hacer la lectura de absorbancia a 450 nm. En todos
los casos los sueros se analizaron por duplicado. Para el análisis de reactividad de las
proteínas de fusión a GST o de los péptidos sin acoplar en muestras de pacientes con
infección crónica, se usó 0.2 μg de antígeno por pocillo. Se bloqueó con leche descremada
4% en PBS-T 0.05% y se incubaron los sueros diluidos 1:500 en esta misma solución. Se
reveló con anticuerpos anti-IgG humanos acoplados a HRP (Sigma) 1:2500, en solución de
bloqueo. Las soluciones de revelado (1 mg de TMB (Sigma-Aldrich)), 333 μl de DMSO y
120 μl de H2O2 (30 vol) en 10 ml de buffer citrato 0.1 M pH 4.2 y finalización (Ácido
sulfúrico 2 M) fueron idénticas para todos los ensayos de ELISA. La detección
colorimétrica en los ensayos ELISA se realizaron con el sistema FilterMax F5 Multimode
Microplate Reader (Molecular Devices) [48]. Todos los valores de absorbancia se
relativizaron al valor de un suero positivo analizado en cada ensayo.
Criterios de positividad y negatividad en los ensayos de ELISA: en todos los ensayos se
usaron muestras de sueros de pacientes infectados crónicos y se incluyeron sueros
negativos de individuos no infectados con T. cruzi con características similares a los del
muestreo. Los sueros analizados se consideraron:
- positivos para cierto antígeno si la media de la absorbancia de las muestras, menos 3
SD, es mayor a la media del valor obtenido para el mismo antígeno con los sueros
negativos, más 3 SD.
- negativos si la media de la absorbancia de la muestra, menos 3 SD, es menos a la
media del valor obtenido para el mismo antígeno con los sueros negativos, más 3 SD.
Análisis estadístico

Trabajo Final - Lobo Maite Materiales y métodos
24
Los distintos test estadísticos se realizaron con GraphPad Prism versión 5 (GraphPad
software, San Diego, CA). El ensayo de Real-Time qPCR se analizó utilizando el test
estadístico ANOVA de dos vías con corrección de Bonferroni. Los resultados de ELISA se
analizaron con la prueba t-Student no pareado. Para todos los casos se consideró
significativo un valor de p<0.05.

Trabajo Final - Lobo Maite Resultados
25
Resultados
Identificación y caracterización in silico de la familia génica TolT en T. cruzi
Familia génica TolT
Para analizar la complejidad de los genes tipo TolT y sus productos en T. cruzi se
llevaron a cabo búsquedas en la base de datos genómica de kinetoplástidos, TriTrypDB,
utilizando como secuencia señuelo el marco de lectura abierto de TolT1 [20]. Un total de
22 secuencias diferentes mostrando similitud significativa (44-99% de identidad
nucleotídica) con TolT1 se obtuvieron a partir de distintos aislamientos de T. cruzi (17
secuencias), el parásito de murciélagos T. cruzi marinkellei (4 secuencias) y el patógeno
humano Trypanosoma rangeli (1 secuencia). Ninguna secuencia con alta identidad a TolT1
se encontró en los genomas de otros patógenos humanos relevantes como T. brucei o
Leishmanias. Un árbol filogenético basado en el método de Neighbor-Joining definió 3
grupos principales de secuencias cercanas a TolT1, a los cuales definimos como TolT-A,
TolT-B y TolT-C (Figura R.1).
Figura R.1: Árbol filogenético basado en el método de Neighbor-Joining donde se
discrimina por colores los 3 grupos de secuencias asociados a TolT1 definidos como TolT-
A (rojo), TolT-B (verde) y TolT-C (azul) en base a las 3 principales ramas observadas. Se
muestran los valores de Boostrap en cada rama. TcCLB: T. cruzi CL Brener. TCDM: T.
cruzi clon Dm28c. Tc_MARK: T. cruzi clon marinkellei. Tc_SYL: T. cruzi clon Sylvio
X10/1. TRSC58: T. rangeli clon SC58. *Secuencias anotadas como pseudeogén.

Trabajo Final - Lobo Maite Resultados
26
El primer grupo, denominado TolT-A mostró 96-99% de identidad con el marco de
lectura abierto de TolT1 e incluye 5 secuencias completas del clon híbrido de referencia
CL Brener1 [22] (TcCLB.508767.20, TcCLB.506617.10, TcCLB.506617.20,
TcCLB.504157.130 y TcCLB.511109.10) y una secuencia parcial (TcCLB.506617.5) con
el C-terminal truncado aparentemente debido a problemas de ensamblado del genoma
(Figura R.1 y S.1). Como originalmente se describió para los genes TolT1-3 de la cepa
Esmeraldo [20], 5 de los genes putativos de TolT-A están arreglados en tándem en
orientación cabeza-cola de a 3 y 2 unidades en el cromosoma 23 haplotipo No-Esmeraldo
(P) y Esmeraldo (S), respectivamente, mientras que la sexta secuencia también se localiza
en el cromosoma 23 No-Esmeraldo, pero por fuera del cluster conteniendo los 3 genes
antes mencionados (Figura R.2). TcCLB.511109.10 está anotado como pseudogen debido
a la deleción de un nucleótido que origina un corrimiento del marco de lectura dentro del
péptido N-terminal predicho. Río abajo de esta mutación, sin embargo, esta secuencia es
~98% y 96% idéntica a TcCLB.508767.20 a nivel de ADN y proteína respectivamente
(Figura R.3).
1 Distintas tipificaciones bioquímicas y moleculares realizadas sobre múltiples aislamientos o cepas del
parásito han permitido delinear al menos 6 linajes evolutivos o unidades discretas de tipificación (UDT)
dentro de la especie T. cruzi, llamados TcI a TcVI. CL Brener pertenece a la UDT TcVI, la cual es un
híbrido resultante de un cruzamiento entre un parental TcII y otro TcIII.

Trabajo Final - Lobo Maite Resultados
27
Figura R.2: Esquema representativo de la disposición de los miembros de la familia TolT
en el genoma de CL Brener. Se indica en código de colores la subfamilia a la que
pertenece: TolT-A (rojo), TolT-B (verde) y TolT-C (azul). En la parte superior, número de
acceso en GeneDB, abajo, la localización y orientación en el cromosoma y alelo
correspondientes. Las zonas tramadas indican las porciones de Tc.CLB.506815.20 con un
alto porcentaje de identidad nucleotídica con Tc.CLB.504277.20 y Tc.CLB.504277.30.
Las líneas verticales en los extremos indican que el gen se encuentra truncado.

Trabajo Final - Lobo Maite Resultados
28
Figura R.3: Alineamiento de secuencias nucleotídicas y aminoacídicas (corregida) de
TolT-A TcCLB.508767.20 y TcCLB.511109.10 donde se observa un nivel de identidad
del 98% y 96%, respectivamente, río debajo de la deleción del nucleótido 46.
El segundo grupo, denominado TolT-B, tiene 76-90% de identidad con TolT1 y está
compuesto por 4 secuencias en el genoma de CL Brener, 2 completas (TcCLB.510433.20
y TcCLB.504277.20) y 2 secuencias parciales (TcCLB.504277.11 y TcCLB.508767.10),
ambas con el N-terminal truncado probablemente debido a problemas de ensamblado del
genoma (Figura R.1 y S.1). Tres de estas secuencias se localizan en el cromosoma 35, 1 en
el haplotipo No-Esmeraldo y 2 en el Esmeraldo, mientras que TcCLB.508767.10 se ubica
río abajo de TcCLB.508767.20 dentro del cluster genómico de TolT-A en el cromosoma
23 Esmeraldo, antes mencionado (Figura R.2). TcCLB.510433.20 y TcCLB.504277.20 son
secuencias casi idénticas excepto por su C-terminal, donde se vuelven altamente
divergentes (Figura R.4).

Trabajo Final - Lobo Maite Resultados
29
Figura R.4: Alineamiento de secuencias aminoacídicas de TolT-B TcCLB.510433.20 y
TcCLB.504277.20 donde se observa que ambas secuencias son casi idénticas hasta su C-
terminal donde divergen y la segunda carece de su señal de anclaje por GPI (marcado en
verde). ω indica el residuo predicho de adición del GPI en TcCLB.510433.20.
De acuerdo a esta nueva clasificación, los ortólogos de los productos de los genes
TolT1-3 originales [20] formarían parte del grupo TolT-A, mientras que la mezcla utilizada
para serodiagnóstico [21] correspondería a dos miembros de TolT-B (TcCLB.510433.20 y
TcCLB.504277.11) y uno de TolT-A (TcCLB.504157.130).
El tercer grupo, denominado TolT-C está compuesto solamente por 2 alelos
(TcCLB.504277.30 y TcCLB.506815.20) (Figura R.1 y S.1) localizados en los
cromosomas Esmeraldo y No-Esmeraldo dentro del cluster genómico de TolT-B, en el
cromosoma 35 (Figura R.2). TcCLB.506815.20 está anotado como el pseudogen de una
MASP. A pesar de su localización genómica y las características tipo TolT predichas
(péptido señal N-terminal y señal de anclaje por GPI C-terminal y composición
aminoacídica en general), los genes de TolT-C muestran muy baja similitud con TolT1
(~46.8-47.1% de identidad nucleotídica). La Figura R.5 esquematiza las distintas variantes
dentro de las subfamilias de TolT.

Trabajo Final - Lobo Maite Resultados
30
Figura R.5: Esquema representativo de la estructura general de los miembros de la
familia TolT. En código de colores se representa el grado de identidad. TolT-A
completas: TcCLB.508767.20, TcCLB.506617.10, TcCLB.506617.20,
TcCLB.504157.130 y TcCLB.511109.10, TolT-A con C-terminal truncado:
TcCLB.506617.5; TolT-B completas: TcCLB.510433.20 (con señal de anclaje por GPI)
y TcCLB.504277.20 (sin señal de anclaje por GPI), TolT-B con N-terminal truncado:
TcCLB.504277.11 y TcCLB.508767.10; TolT-C: TcCLB.504277.30 y
TcCLB.506815.20. Círculos superiores indican los sitios de N-glicosilación predichos.
A: Alanina, C: Cisteína, R: Arginina, E: Ac. Glutámico.
Proteínas TolT
Las proteínas codificadas por TolT-A, TolT-B y TolT-C contienen 310, 305/284 y 344
aminoácidos, respectivamente, con masas de ~30.9-37.1 kDa y pI de 7.2-8.4. Todas
muestran un péptido señal N-terminal y una señal de anclaje por GPI en el extremo C-
terminal (excepto TcCLB.504277.20 que carece de la última) (Figura R.5 y R.7). Estas
señales serían responsables del tráfico intracelular y localización en membrana plasmática
de las proteínas TolT. El anclaje por GPI fue confirmado experimentalmente por
fraccionamiento en Triton X114 (Figura R.6.A) y además se evaluó la potencial secreción

Trabajo Final - Lobo Maite Resultados
31
espontánea de los productos TolT, la cual no sucede o sucede a niveles no detectables
(Figura R.6.B).
Figura R.6: Purificación de proteínas ancladas por GPI mediante el
método con Tritón X-144 (A) y ensayo de secreción (B). Las muestras de
cada paso de purificación (A), así como el pellet y el sobrenadante filtrado
(en algunos casos, B), se analizaron con el antisuero indicado mediante
WB. P: pellet, S: sobrenadante, Sf: sobrenadante filtrado. Se sembraron
1x106 y 6x106 parásitos y, su equivalente en extracto de secreción,
respectivamente.
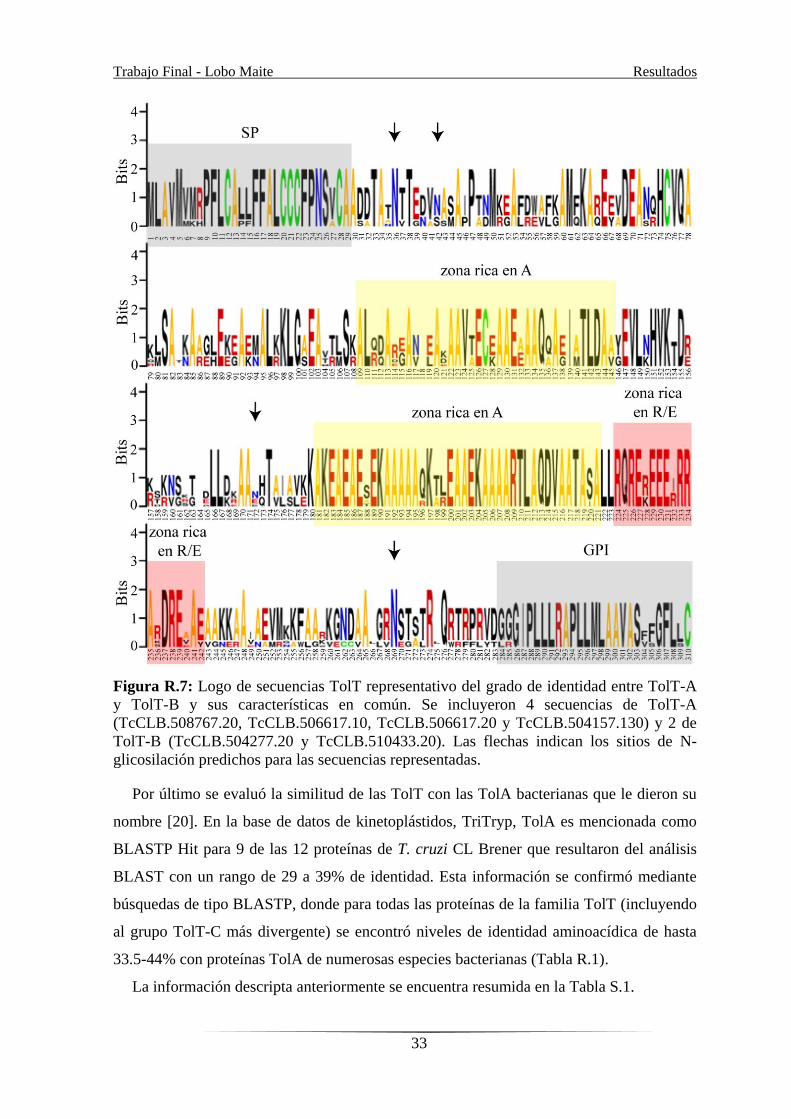
Los polipéptidos presentan un alto contenido de Ala (18.8 a 24.4%), Glu (9.0 a 11.9%),
Leu (8.1 a 9.9%), Lys (6.3 a 10.4%) y Arg (4.9 a 8.1%), con estos residuos no igualmente
distribuidos a lo largo de las proteínas (Figura R.7.).
Dentro de las moléculas de TolT, la única característica común y reconocible fue la
presencia de 2 regiones ricas en Ala con estructura secundaria no definida y la presencia de
varios residuos Cys conservados. Los productos de TolT-A y TolT-B, además, contienen

Trabajo Final - Lobo Maite Resultados
32
un motivo Cys-X7-Cys3 en su SP, así como una región particular hacia su señal de anclaje
por GPI que está enriquecida en residuos de Arg (R) y Glu (E) (Figura R.5 y R.7).
Análisis bioinformáticos sobre las modificaciones post-traduccionales putativas de las
secuencias predichas de TolT, mostró que tendrían también de 17 a 41 sitios potenciales
para fosforilación, de 2 a 5 para N-glicosilación (Figura R.5 y R.7) y de 27 a 36 para O-
glicosilación. Las secuencias de los grupos TolT-A y TolT-B, además, presentan una
cisteína altamente conservada en posición C-terminal (ver Figura R.7) que constituye un
sitio de palmitoilación según los algoritmos utilizados. Este residuo, sin embargo, no se
encontraría en la proteína madura ya que forma parte de la señal clivable de adición de
GPI, por lo que desestimamos la significancia biológica de esta predicción.
La región N-terminal madura de 135 aminoácidos predicha (del residuo 40 al 175)
mostró el menor grado de identidad entre TolT-A y TolT-B (Figura R.7 y S.1).
Predicciones de topología, de acuerdo a otras moléculas de superficie de T. cruzi ancladas
por GPI [53], sugieren que esta región constituye el extremo distal a la membrana de las
moléculas de TolT mientras que el extremo proximal a la membrana sería el C-terminal
altamente conservado.
Trabajo Final - Lobo Maite Resultados
33
Figura R.7: Logo de secuencias TolT representativo del grado de identidad entre TolT-A
y TolT-B y sus características en común. Se incluyeron 4 secuencias de TolT-A
(TcCLB.508767.20, TcCLB.506617.10, TcCLB.506617.20 y TcCLB.504157.130) y 2 de
TolT-B (TcCLB.504277.20 y TcCLB.510433.20). Las flechas indican los sitios de N-
glicosilación predichos para las secuencias representadas.
Por último se evaluó la similitud de las TolT con las TolA bacterianas que le dieron su
nombre [20]. En la base de datos de kinetoplástidos, TriTryp, TolA es mencionada como
BLASTP Hit para 9 de las 12 proteínas de T. cruzi CL Brener que resultaron del análisis
BLAST con un rango de 29 a 39% de identidad. Esta información se confirmó mediante
búsquedas de tipo BLASTP, donde para todas las proteínas de la familia TolT (incluyendo
al grupo TolT-C más divergente) se encontró niveles de identidad aminoacídica de hasta
33.5-44% con proteínas TolA de numerosas especies bacterianas (Tabla R.1).
La información descripta anteriormente se encuentra resumida en la Tabla S.1.

Trabajo Final - Lobo Maite Resultados
34
Tabla R.1: Proteínas bacterianas con máximo porcentaje de identidad, resultantes de una
búsqueda de tipo BLASTP, para cada uno de los miembros de la familia TolT de T. cruzi
CL Brener.
Nro de
acceso
GeneDB
(TcCLB.)
Grupo
TolT
% identidad con TolA (aa)
Especie
508767.20 A
37.4 Proteína TolA
Yersinia enterocolitica subsp. Palearctica serotype
511109.10 33.5 Proteína TolA
Enterobacter cloacae S611
506617.5 41.7 Proteína TolA, proteína de integridad de la envoltura
celular, proteína de integridad de la membrana interna de la
envoltura celular, proteína anclada a membrana en complejo
TolA-TolQ-TolR
Serratia sp.
506617.10 39.2 Proteína anclada a membrana en complejo TolA-TolQ-
TolR
Serratia proteamaculans
506617.20
39.2 Proteína anclada a membrana en complejo TolA-TolQ-
TolR
Serratia proteamaculans
504157.130 39 Proteína anclada a membrana en complejo TolA-TolQ-TolR
Serratia proteamaculans
508767.10 B
36.8 Proteína TolA
Salmonella bongori N268-08
504277.11 40.4 Proteína TolA
Enterobacter cloacae UCICRE 12
510433.20 36.9 Proteína TolA
Paracoccus sp. MKU1
504277.20 37.1 Proteína TolA
Dickeya zeae strain Ech586
506815.20 C 34.5 Familia de proteínas TolA
Obesumbacterium proteus ATCC 12841
504277.30 40.6 Proteína TolA
Edwardsiella piscicida

Trabajo Final - Lobo Maite Resultados
35
Expresión de ARNm y productos de TolT durante el ciclo de vida de T. cruzi
Expresión génica de TolT
De acuerdo a resultados observados en un experimento de hibridación de microarreglos
[54], el nivel de expresión de ARNm de TcCLB.504277.20 (TolT-B) y TcCLB.504277.30
(TolT-C) es mayor en tripomastigotes derivados de células mientras el nivel de expresión
de TcCLB.511109.10 (TolT-A) es mayor en epimastigotes. Estos resultados no son
consistentes con el hecho de que las proteínas TolT originales (TolT-A de acuerdo a
nuestra clasificación) fueron descriptas como moléculas restringidas a la forma
tripomastigote derivado de células [16].
Para abordar esta cuestión, se purificaron muestras de ARN total de los distintos
estadios de T. cruzi y se analizó la expresión génica de miembros representativos de las 3
subfamilias de TolT (A, B y C) mediante Real-time qPCR. Se observa, en general, una
mayor expresión de TolT-A, seguida por TolT-B y una mínima expresión de TolT-C,
obteniendo diferencias estadísticamente significativas solo entre la expresión de TolT-A y
TolT-C en tripomastigotes. Entre estadios, la expresión de TolT-A es claramente mayor en
tripomastigotes, mientras que la de TolT-B y TolT-C resulta más homogénea, siendo
estadísticamente significativa la diferencia de expresión de TolT-A entre tripomastigotes y
epimastigotes (Figura R.8.). En general las curvas de disociación resultaron en un pico
único a la temperatura correspondiente para cada producto (excepto para TolT-B) (Figura
S.3) mientras que no se observó amplificación en los controles sin templado. Los
productos de amplificación fueron secuenciados confirmando su identidad.

Trabajo Final - Lobo Maite Resultados
36
Figura R.8: Análisis de la expresión a nivel de ARN mediante Real-Time qPCR de
cada uno de los grupos de TolT en los principales estadios de T. cruzi. La
cuantificación se relativizó al nivel de expresión de TolT-C en epimastigotes y se
normalizó al de los genes GAPDH y Calmodulina.
Expresión de las proteínas TolT
Respecto a la expresión a nivel proteico, y más allá de los datos originales del grupo de
Jerry Manning [16,20] diferentes sets de datos proteómicos revelaron expresión de
diferentes miembros de TolT, particularmente en estadios que se desarrollan en el
hospedador mamífero (tripomastigotes derivados de células y/o tripomastigotes
sanguíneos) [29-31]. Para obtener más información sobre la expresión y caracterización de
los productos de TolT, se produjeron antisueros en ratón contra proteínas recombinantes
fusionadas a GST, conteniendo regiones particulares de un miembro representativo de los
distintos grupos de TolT definidos (Figura R.1 y S.1) Para minimizar la reacción cruzada
entre TolT-A y TolT-B, el inmunógeno se limitó a la región N-terminal divergente, aunque
también se generaron antisueros contra la región C-terminal común (TolT-B/A). Para
TolT-C, los animales fueron inmunizados con un péptido específico, que surgió como el
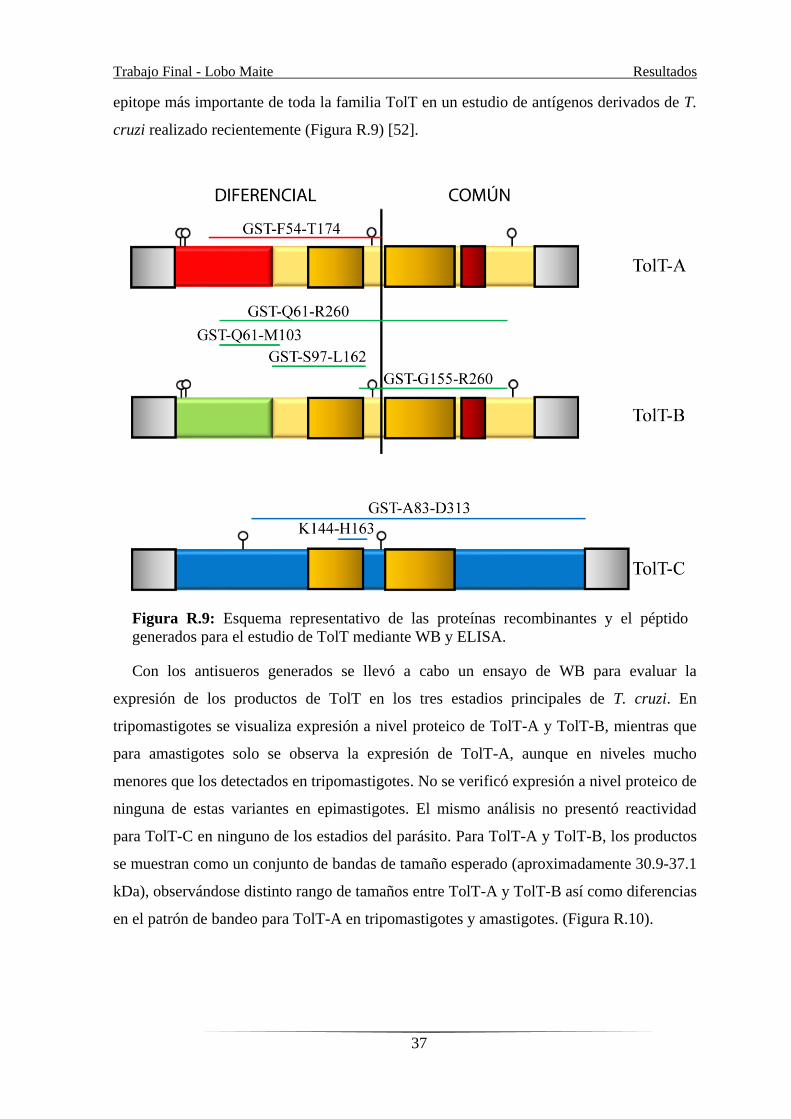
Trabajo Final - Lobo Maite Resultados
37
epitope más importante de toda la familia TolT en un estudio de antígenos derivados de T.
cruzi realizado recientemente (Figura R.9) [52].
Figura R.9: Esquema representativo de las proteínas recombinantes y el péptido
generados para el estudio de TolT mediante WB y ELISA.
Con los antisueros generados se llevó a cabo un ensayo de WB para evaluar la
expresión de los productos de TolT en los tres estadios principales de T. cruzi. En
tripomastigotes se visualiza expresión a nivel proteico de TolT-A y TolT-B, mientras que
para amastigotes solo se observa la expresión de TolT-A, aunque en niveles mucho
menores que los detectados en tripomastigotes. No se verificó expresión a nivel proteico de
ninguna de estas variantes en epimastigotes. El mismo análisis no presentó reactividad
para TolT-C en ninguno de los estadios del parásito. Para TolT-A y TolT-B, los productos
se muestran como un conjunto de bandas de tamaño esperado (aproximadamente 30.9-37.1
kDa), observándose distinto rango de tamaños entre TolT-A y TolT-B así como diferencias
en el patrón de bandeo para TolT-A en tripomastigotes y amastigotes. (Figura R.10).

Trabajo Final - Lobo Maite Resultados
38
Figura R.10: Análisis de la expresión a nivel proteico por
WB de TolT-A y B sobre lisado total de parásitos en cada
uno de los principales estadios de T. cruzi. Se sembraron
2x107 de parásitos de cada estadio por calle. T:
tripomastigote, A: amastigote, E: epimastigote.
Localización subcelular de las proteínas TolT
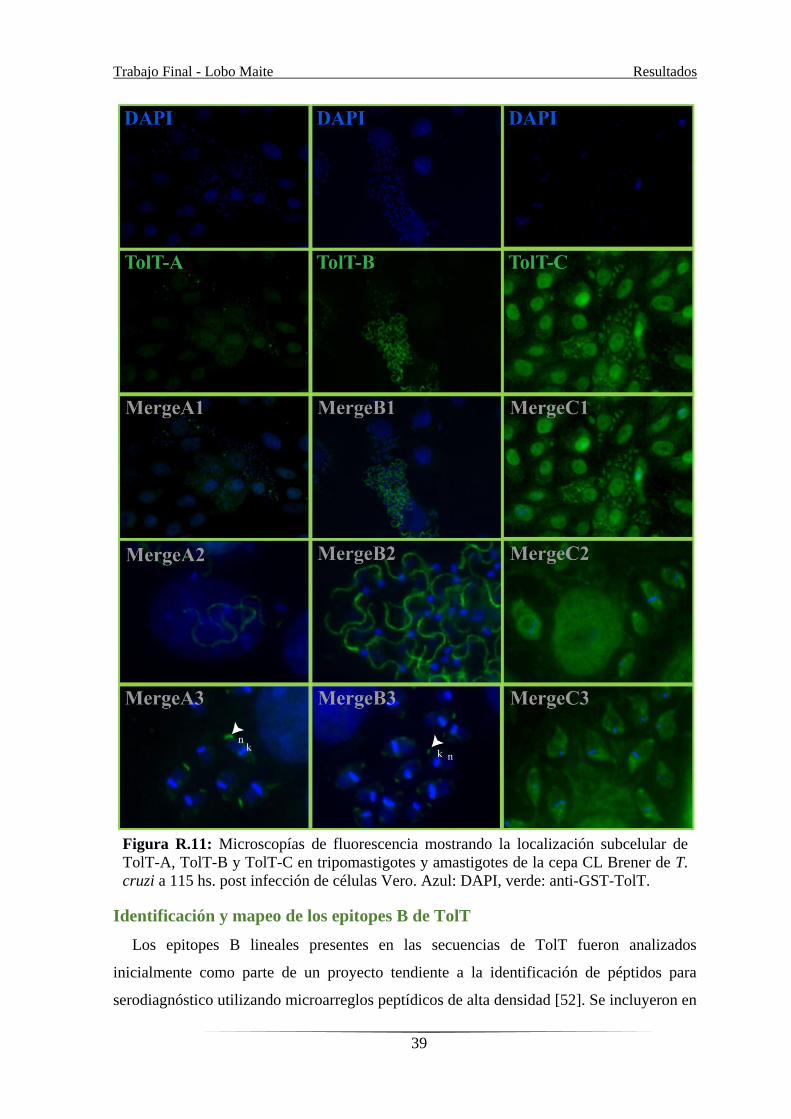
Con el fin de profundizar el estudio de los productos de TolT, se analizó su localización
subcelular mediante Inmunofluorescencia indirecta (IFI) en los distintos estadios de T.
cruzi CL Brener. Las microscopías muestran que TolT-A se localiza en el flagelo de
tripomastigotes derivados de células y en el extremo posterior de amastigotes
intracelulares. TolT-B también tiene localización predominante en flagelo de
tripomastigotes, aunque también puede observarse señal en el extremo anterior de los
amastigotes (indetectable en el WB). Para TolT-C, en cambio, no se observó marca
específica en ninguno de los estadios (el nivel de fluorescencia en los parásitos fue
equivalente al de las células por lo que se consideró inespecífica), coincidente con los
datos de WB. Análisis similares no mostraron señal para ninguno de los antisueros en
epimastigotes (Figura R.11).
Trabajo Final - Lobo Maite Resultados
39
Figura R.11: Microscopías de fluorescencia mostrando la localización subcelular de
TolT-A, TolT-B y TolT-C en tripomastigotes y amastigotes de la cepa CL Brener de T.
cruzi a 115 hs. post infección de células Vero. Azul: DAPI, verde: anti-GST-TolT.
Identificación y mapeo de los epitopes B de TolT
Los epitopes B lineales presentes en las secuencias de TolT fueron analizados
inicialmente como parte de un proyecto tendiente a la identificación de péptidos para
serodiagnóstico utilizando microarreglos peptídicos de alta densidad [52]. Se incluyeron en

Trabajo Final - Lobo Maite Resultados
40
el arreglo 3 productos diferentes de TolT-A (TcCLB.506617.10, TcCLB.504157.130 y
TcCLB.508767.20), 3 de TolT-B (TcCLB.510433.20, TcCLB.504277.20 y
TcCLB.504277.11) y uno de TolT-C (TcCLB.504277.30) (Figura R.1).
En general, los productos de TolT no mostraron reactividad a lo largo de su SP y señal
de GPI, lo que resulta consistente con que estas regiones son procesadas y escindidas
previo a su exposición y anclaje en la superficie del parásito. Dentro de la región de la
proteína madura, las señales obtenidas fueron muy homogéneas para los distintos
miembros de una misma subfamilia, consistente con su altísima identidad aminoacídica,
así que sólo se grafica un miembro de cada una. TolT-A muestra solo un pico de
reactividad débil, localizado cerca de su C-terminal:
261GNDAALGRNSTSTRLQRTRPRVD, secuencia que se encuentra también conservada
en TolT-B: 256GNDAAPGRNSTATRIQRTRPRVD, pero muestra un pico más fuerte,
posiblemente debido a los polimorfismos entre ambas secuencias. TolT-C muestra un
perfil completamente diferente con un pico antigénico único:
141EAEKAAVDADTAALAALLEVLQHSKF (Figura R.12). En términos relativos, esta
secuencia fue la que presentó mayor reactividad de toda la familia en los micro-chips, y
sobre ella se derivó posteriormente el péptido sintético utilizado para generar anticuerpos
anti-TolT-C.

Trabajo Final - Lobo Maite Resultados
41
Figura R.12: Microarreglos de péptidos de alta
densidad de miembros representativos de cada
subfamilia de TolT (se grafica un miembro de
cada subfamilia ya que los perfiles obtenidos
entre miembros de la misma fueron similares). Se
muestra la reactividad de 3 pooles de sueros
distintos (1, 2 y 3).
El perfil antigénico de los productos de TolT durante la infección por T. cruzi fue
además evaluado usando proteínas recombinantes como se describió previamente [48].

Trabajo Final - Lobo Maite Resultados
42
Con ese fin se generaron y purificaron de sistemas bacterianos una serie de constructos. Se
utilizaron proteínas recombinantes fusionadas a GST específicas de cada subfamilia: TolT-
A (desde F54 hasta T174), TolT-B (desde Q61 hasta S103) y TolT-C (desde A83 hasta
D313). Además, se incluyó en el análisis a la proteína TolT-B/A desde G157 hasta A260
(Figura R.9), que tiene un ~80.2% de identidad aminoacídica con TolT-A, para buscar
anticuerpos específicos en pacientes Chagásicos crónicos mediante ELISA. Las 31
muestras de suero fueron heterogéneas, incluyendo individuos que viven en áreas
endémicas y no endémicas, probablemente infectados por distintas cepas de T. cruzi. A su
vez, se incluyeron 19 sueros provenientes de individuos sanos. Se analizó la reactividad de
todos los sueros contra GST como control y la reactividad se normalizó a un suero positivo
incluido en todos los ensayos. Las regiones N-terminales divergentes entre TolT-A y TolT-
B no resultan antigénicas mientras que se observa un reconocimiento, estadísticamente
significativo, por parte de los sueros hacia la región C-terminal conservada (residuos
G157-A260 de TolT-B), (Figura R.13). TolT-C fue únicamente reconocida por 2 sueros
(no significativo). La prevalencia de los anticuerpos dirigidos a la zona conservada entre
TolT-A y B entre sueros argentinos potencialmente chagásicos resultó del 0%, 2.9%,
42.9% y 5.7% para N-terminal de TolT-A, N-terminal de TolT-B, C-terminal de TolT-B/A
y TolT-C, respectivamente. Ninguna muestra de suero de individuos no infectados con T.
cruzi (n=19) resultó reactiva para TolT.
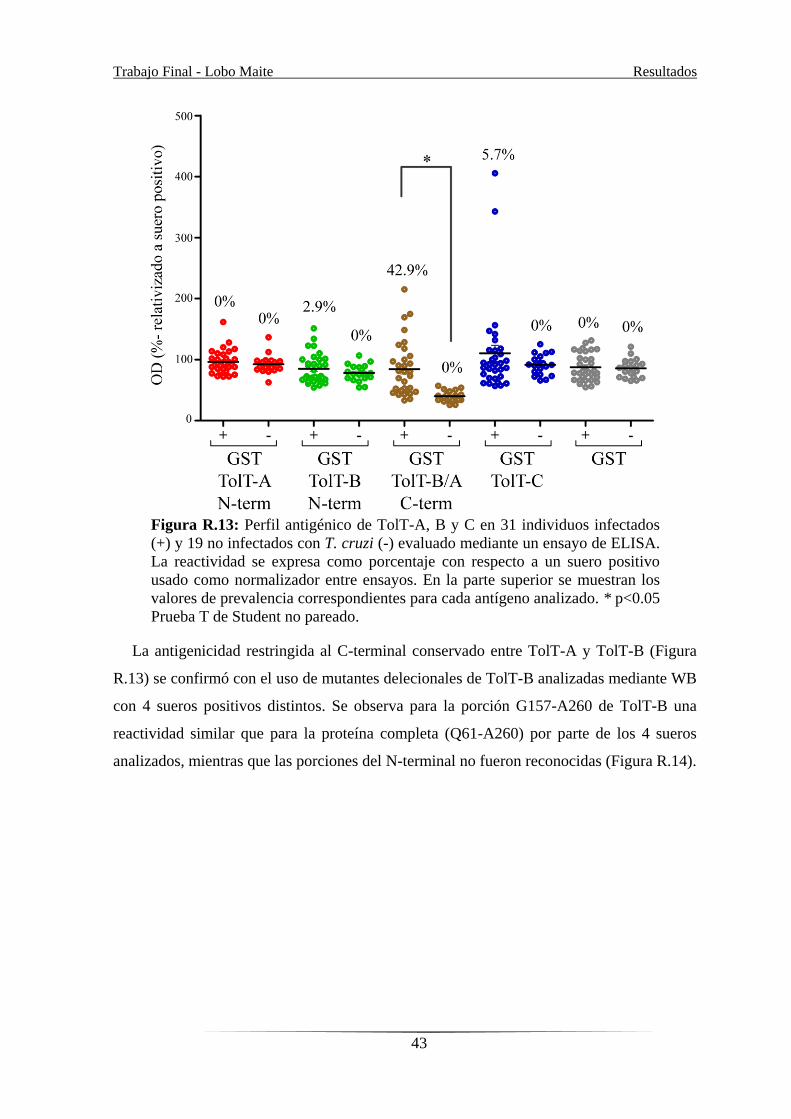
Trabajo Final - Lobo Maite Resultados
43
Figura R.13: Perfil antigénico de TolT-A, B y C en 31 individuos infectados
(+) y 19 no infectados con T. cruzi (-) evaluado mediante un ensayo de ELISA.
La reactividad se expresa como porcentaje con respecto a un suero positivo
usado como normalizador entre ensayos. En la parte superior se muestran los
valores de prevalencia correspondientes para cada antígeno analizado. * p<0.05
Prueba T de Student no pareado.
La antigenicidad restringida al C-terminal conservado entre TolT-A y TolT-B (Figura
R.13) se confirmó con el uso de mutantes delecionales de TolT-B analizadas mediante WB
con 4 sueros positivos distintos. Se observa para la porción G157-A260 de TolT-B una
reactividad similar que para la proteína completa (Q61-A260) por parte de los 4 sueros
analizados, mientras que las porciones del N-terminal no fueron reconocidas (Figura R.14).

Trabajo Final - Lobo Maite Resultados
44
Figura R.14: Mapeo delecional de TolT-B mediante WB con 4 sueros chagásicos
distintos. Se sembraron 0.5 µg de cada proteína recombinante por calle.

Trabajo Final - Lobo Maite Conclusiones y Discusión
45
Conclusiones y Discusión
Identificación y caracterización in silico de la familia génica TolT en T. cruzi
Como paso inicial de esta tesina, se reevaluó la complejidad de la familia génica TolT a
la luz de la disponibilidad de nuevos datos genómicos. Un total de 22 secuencias diferentes
mostraron similitud significativa con TolT1, las cuales se dividieron en 3 grupos
coherentes y robustos, a los que denominamos TolT-A, TolT-B y TolT-C. Los resultados
de esta nueva búsqueda expandieron 5-6 veces el número de secuencias TolT reconocidas
y permitió definir grupos robustos dentro de la familia.
TolT-A, grupo más cercano a TolT1, incluye 5 secuencias completas y una parcial en la
cepa CL Brener. Estas se encuentran arregladas en tándem en orientación cabeza-cola y
localizadas en el cromosoma 23 por lo que serían los ortólogos de TolT1-3 de la cepa
Esmeraldo [20]. Una de las secuencias se encuentra anotada como pseudogen debido a una
deleción en su N-terminal. En general, tras la mutación que origina un pseudogen las
secuencias quedan liberadas de presión selectiva y comienzan a acumular mutaciones; pero
en este caso, la falta de divergencia indicaría un evento de pseudogenización reciente o una
secuencia anotada incorrectamente, problema recurrente en el genoma de T. cruzi. TolT-B
está conformado por 4 secuencias de CL Brener, 2 completas y 2 parciales, todas
localizadas en el cromosoma 35, excepto TcCLB.508767.10 que lo hace en el cromosoma
23. TcCLB.510433.20 y Tc.CLB.504277.20 son secuencias similares, excepto por su C-
terminal donde la segunda carece de su señal de anclaje por GPI, lo cual podría tener
impacto en el tráfico y/o disposición final de las proteínas ya que, en T. cruzi (a diferencia
de mamíferos) el carecimiento de ésta parece promover la retención de las proteínas en el
RE [55]. El tercer grupo, TolT-C, incluye solo dos alelos con muy baja homología a TolT1
ubicados dentro del cluster genómico de TolT-B, uno de ellos anotado como pseudogen de
una MASP. Esto último fue analizado con más profundidad y se determinó que el N-
terminal de esta secuencia contiene una porción con similitud con el extremo C-terminal
de la secuencia TolT-B que carece de la señal de anclaje por GPI mientras que su C-
terminal es casi idéntico al otro alelo de TolT-C. Interesantemente, en cada uno de los
cromosomas 35 (Esmeraldo y No-Esmeraldo) se observa la siguiente disposición: TolT-B
con señal de anclaje por GPI -TcCLB.510433.20/TcCLB.504277.11 (N-terminal
truncado)- + TolT-B sin señal de anclaje por GPI -N-terminal de
TcCLB.506815.20/TcCLB.504277.20- + TolT-C -C-terminal de
TcCLB.506815.20/TcCLB504277.30-, conformando un arreglo en tándem en orientación

Trabajo Final - Lobo Maite Conclusiones y Discusión
46
cabeza-cola de 3 genes tal como las secuencias TolT-A (Figura R.2, regiones con trama).
Todos estos datos nos permiten hipotetizar que esta variedad de secuencias y su
disposición en el genoma es el resultado de duplicaciones génicas seguidas por
diversificación y eventual pseudogenización aunque se requiere de un análisis más
profundo de las regiones intergénicas y flanqueantes de estos genes para poder detallar el
proceso [56].
Respecto a las características de los productos de TolT, se determinó que en general
despliegan SP y señal de anclaje por GPI y que se caracterizan por un alto contenido de
Ala, Glu, Leu, Lys y Arg. Presentan 2 regiones ricas en Ala con estructura secundaria no
definida y varios residuos Cys con posición conservada los que serían importantes para su
plegamiento final. TolT-A y B también contienen una región enriquecida en Arg y Glu
hacia la señal de anclaje por GPI así como un motivo Cys-X7-Cys3 en su SP que también se
encuentra conservado en otras moléculas expresadas en tripomastigotes como TcMUC
[53,57] y varias MASPs [58], por lo que podría estar asociado a la expresión de estos
genes en tripomastigotes.
TolT sufriría además modificaciones post-traduccionales ya que se predijeron sitios
para fosforilación, N-glicosilación y O-glicosilación, aunque solamente se confirmó
experimentalmente la presencia de N-glicanos anclados a proteínas TolT-A y TolT-B [20].
En general, TolT-A y TolT-B son muy parecidas entre sí, diferenciándose de TolT-C.
Estas presentan menor grado de identidad hacia su región N-terminal la cual según
predicciones de topología constituiría el extremo distal a la membrana y estaría
involucrada en mediar interacciones con ligandos y/o servir como señuelo inmune,
mientras que el extremo proximal a la membrana, altamente conservado, provee la señal
para anclaje por GPI participando en la orientación y unión de la proteína a la superficie
del parásito. Este escenario es similar al de otras familias de proteínas ancladas por GPI
como los antígenos tipo mucina TcMUC, donde la parte distal a la membrana de su
secuencia es muy diversa probablemente debido a presión selectiva del sistema inmune del
mamífero [53,59,60].
El nombre de este grupo de proteínas deviene de su identidad de secuencia
aminoacídica y sus características físicas y biológicas en común con las proteínas TolA de
E. coli y P. aeruginosa, de acuerdo a lo reportado por Manning [20]. Si bien en la base de
datos TriTryp TolA aparece como resultado de un análisis BLASTP, esta información se
corroboró realizando un análisis más amplio mediante una búsqueda manual donde se

Trabajo Final - Lobo Maite Conclusiones y Discusión
47
observó, en general, niveles de identidad aminoacídica similares a lo reportado, pero frente
a proteínas TolA de una población más diversa de bacterias (Serratia, Yersinia,
Enterobacter, Salmonella, etc.). Las proteínas TolA son parte del sistema Tol de la
mayoría de las bacterias gram negativas, participan en la secreción de factores y están
involucradas en la preservación de la estabilidad de la membrana bacteriana [61-63].
Confirmar este parecido, sumado a que el bolsillo flagelar cumple un rol clave en los
procesos endo-exocíticos en tripanosomátidos [9], abre camino a la futura búsqueda de una
función relacionada para TolT.
Expresión de ARNm y productos de TolT durante el ciclo de vida de T. cruzi
Como primer paso hacia la caracterización de TolT, se llevó a cabo un análisis de
expresión a nivel de ARNm de las 3 subfamilias de TolT mediante ensayos de Real-time
qPCR. En términos generales, en todos los estadios se observa una mayor expresión de
TolT-A, seguida por TolT-B y una mínima expresión de TolT-C. Entre estadios, la
expresión de TolT-A es evidentemente mayor en tripomastigotes mientras que la de TolT-
B y C es más homogénea. Esta homogeneidad observada la cual se ve acompañada por un
alto nivel de error podría esclarecerse con un mayor número de ensayos. El análisis de las
curvas de disociación demostró, en general, una amplificación específica, determinada por
un único pico estrecho a la temperatura correspondiente para cada producto y ausencia de
amplificación en los controles sin templado. Solo para TolT-B se observa un pico de
menor altura a una temperatura inferior, lo que corresponde con artefactos como dímeros
de primers,
Los resultados obtenidos por Real-Time qPCR concuerdan con la descripción original
de las proteínas TolT y, particularmente TolT-A, como moléculas restringidas a la forma
tripomastigote [16].
En segunda instancia se analizó la expresión de TolT a nivel proteico mediante WB e
IFI donde se obtuvieron algunos resultados discordantes. Por IFI se observa la localización
de TolT-A en el flagelo de la forma tripomastigote tal como lo describió Manning [16] así
como en el extremo posterior de amastigotes intracelulares, mismos estadios en los que se
observa expresión por WB, sin embargo, TolT-B se localizaría, al igual que TolT-A, en el
flagelo de tripomastigotes y en el extremo anterior de amastigotes intracelulares, estadio en
el que no se observa expresión por WB. No se observó expresión de ninguna de las
subfamilias en epimastigotes, lo cual concuerda con lo observado por Manning [16] para
TolT-A (en ese trabajo no se analizan amastigotes). En WB se presentó una banda para

Trabajo Final - Lobo Maite Conclusiones y Discusión
48
TolT-A la cual sería inespecífica dado que la masa predicha para TolTs es de 30.9-37.1
kDa.
Identificación y mapeo de los epitopes B de la familia TolT
Como parte de un proyecto tendiente a la identificación de péptidos para
serodiagnóstico utilizando microarreglos peptídicos de alta densidad [52], se analizaron los
epitopes B lineales presentes en 3 secuencias distintas de TolT. TolT-A presentó un único
pico de reactividad débil hacia su C-terminal el cual es compartido con las secuencias de
TolT-B. Para TolT-C también se identificó un pico antigénico hacia su C-terminal, aunque
con un perfil distinto. Si bien los picos antigénicos obtenidos para TolT no presentan la
magnitud de intensidad (12-15 veces menores) observada para otros antígenos de T. cruzi
validados para el serodiagnóstico (TSSA, MASPs, TcMUC), estos estudios fueron de
utilidad para dar una idea general de la zona antigénica de las proteínas TolT.
Se profundizó el análisis del perfil antigénico de los productos de TolT buscando
anticuerpos específicos en pacientes chagásicos mediante ELISA, donde, si bien el epitope
predicho por el CHIP para TolT-A y TolT-B se incluyó parcialmente (G256-A260) debido
a que se encuentra muy cerca de la señal de anclaje por GPI y su inclusión generaría
problemas para la expresión en bacterias, se confirmó que la inmunogenicidad está dirigida
hacia el C-terminal compartido entre TolT-A y TolT-B y que los anticuerpos dirigidos
hacia esta región tienen una prevalencia del 42.9%. Esto se confirmó además con el
análisis por WB de mutantes delecionales de TolT-B.
En términos generales TolT constituye una familia multigénica mucho más compleja
que la originalmente descripta, la cual se divide en tres subfamilias que a pesar de su alto
grado de similitud de secuencias tendrían distintos perfiles de expresión, localización sub-
celular y, probablemente, función.

Trabajo Final - Lobo Maite Información suplementaria
49
Información suplementaria
Figura S.1: Alineamiento múltiple de las secuencias aminoacídicas de los miembros de la
familia TolT de T. cruzi cepa CL Brener. A la izquierda se muestra en código de colores el
número de acceso en GenDB de cada proteína, indicando la subfamilia a la que pertenece:
TolT-A (rojo), TolT-B (verde) y TolT-C (azul).

Trabajo Final - Lobo Maite Información suplementaria
50
Figura S.2: Mapa del vector pGEM®-T y puntos de referencia de
secuencia. En verde se señalan las regiones codificantes de los
promotores de las ARN polimerasas SP6 y T7, que flanquean al MCS,
y a las cuales se unen los oligonucleótidos utilizados para el chequeo
por PCR de los clonados en este vector.

Trabajo Final - Lobo Maite Información suplementaria
51
Figura S.3: Curvas de disociación de los productos de Real-Time PCR (TolT-A, TolT-B,
TolT-C, TcGAPDH y TcCalmodulina). En el eje Y representa la fluorescencia (derivative
reporter (∆Rn)) mientras que el eje X representa la temperatura (°C). Las figuras muestran
la temperatura de disociación correspondiente para cada gen, con dímeros de primers
detectados solo en TolT-B.

Trabajo Final - Lobo Maite Información suplementaria
52
Figura S.4: SDS-PAGE de proteínas TolT con fusión a GST purificadas.

Trabajo Final - Lobo Maite Información suplementaria
53
Tabla S.1: Resumen detallado de características de los distintos miembros de la familia
TolT en T. cruzi CL Brener.
Nro de
acceso
GeneDB
(TcCLB.)
Grupo
TolT
Cromo-
soma
% ident
TolT-1
(nt-aa)
Características
proteicas
% ident TolA
(aa)
Especie
Anotación
508767.20 A
23 E
99-99.4 SP, GPI, Cys
palmitoilable
en C-term,
hélice
superenrollada,
motivo
C7CCC en SP
37.4
Proteína
TolA
Yersinia
enterocolitica
subsp.
Palearctica
serotype
511109.10 98-73.9 33.5
Proteína
TolA
Enterobacter
cloacae S611
Pseudogen,
deleción
simple de T
en SP N-
term
506617.5 98-95.6 41.7
Proteína
TolA
Proteína de
integridad de
la envoltura
celular
Proteína de
integridad de
la membrana
interna de la
envoltura
celular
Proteína
anclada a
membrana en
complejo
TolA-TolQ-
TolR
Serratia sp.
Secuencia
parcial (C-
term
truncado)
506617.10 23 nE
97-92.9 39.2
Proteína
anclada a

Trabajo Final - Lobo Maite Información suplementaria
54
membrana en
complejo
TolA-TolQ-
TolR
Serratia
proteamacu-
lans
506617.20
96-92.6
39.2
Proteína
anclada a
membrana en
complejo
TolA-TolQ-
TolR
Serratia
proteamacu-
lans
504157.130 96-91.3 39
Proteína
anclada a
membrana en
complejo
TolA-TolQ-
TolR
Serratia
proteamacu-
lans
508767.10 B
23 E
90-77.5 36.8
Proteína
TolA
Salmonella
bongori
N268-08
Secuencia
parcial (N-
term
truncado)
504277.11 35 E
87-76.1 SP, GPI, Cys
palmitoilable,
en C-term,
hélice
supererollada
40.4
Proteína
TolA
Enterobacter
cloacae
UCICRE 12
Secuencia
parcial (N-
term
truncado)
510433.20 35 nE
81-70.6 SP, GPI, Cys
palmitoilable
36.9
Proteína

Trabajo Final - Lobo Maite Información suplementaria
55
en C-term,
hélice
superenrollada,
motivo
CX7CCC en
SP
TolA Para-
coccus sp.
MKU1
504277.20 35 E 76-62.1 SP, hélice
superenrollada,
motivo
CX7CCC en
SP
37.1
Proteína
TolA
Dickeya zeae
strain
Ech586
C-term
altamente
divergente
506815.20 C 35 nE 81
(47)-
28.1
SP, GPI, Cys
palmitoilable
en C-term,
hélice
superenrollada
34.5
Familia de
proteínas
TolA
Obesumbac-
terium
proteus
ATCC 12841
Anotado
como
pseudogen
de MASP
(inserción
en N-term)
504277.30 35 E 47-31 SP, GPI, Cys
palmitoilable
en C-term,
hélice
superenrollada,
SLN bipartita
40.6
Proteína
TolA
Edwardsiella
piscicida

Trabajo Final - Lobo Maite Bibliografía
56
Bibliografía
1. Yaeger RG (1996) Protozoa: Structure, Classification, Growth, and Development. In:
Baron S, editor. Medical Microbiology. 4th ed. Galveston (TX).
2. Stanaway JD, Roth G (2015) The burden of Chagas disease: estimates and challenges.
Glob Heart 10: 139-144.
3. Rassi A, Jr., Rassi A, Marcondes de Rezende J (2012) American trypanosomiasis
(Chagas disease). Infect Dis Clin North Am 26: 275-291.
4. Caradonna KL, Burleigh BA (2011) Mechanisms of host cell invasion by Trypanosoma
cruzi. Adv Parasitol 76: 33-61.
5. Tyler KM, Engman DM (2001) The life cycle of Trypanosoma cruzi revisited. Int J
Parasitol 31: 472-481.
6. Coura JR, Borges-Pereira J (2010) Chagas disease: 100 years after its discovery. A
systemic review. Acta Trop 115: 5-13.
7. de Souza W (1984) Cell biology of Trypanosoma cruzi. Int Rev Cytol 86: 197-283.
8. Cadavid-Restrepo G, Gastardelo TS, Faudry E, de Almeida H, Bastos IM, et al. (2011)
The major leucyl aminopeptidase of Trypanosoma cruzi (LAPTc) assembles into a
homohexamer and belongs to the M17 family of metallopeptidases. BMC Biochem
12: 46.
9. Field MC, Carrington M (2009) The trypanosome flagellar pocket. Nat Rev Microbiol 7:
775-786.
10. Sunter JD, Gull K (2016) The Flagellum Attachment Zone: ‘The Cellular Ruler’ of
Trypanosome Morphology. Trends Parasitol 32: 309-324.
11. Kohl L, Robinson D, Bastin P (2003) Novel roles for the flagellum in cell
morphogenesis and cytokinesis of trypanosomes. EMBO J 22: 5336-5346.
12. Tetley L, Vickerman K (1985) Differentiation in Trypanosoma brucei: host-parasite
cell junctions and their persistence during acquisition of the variable antigen coat. J
Cell Sci 74: 1-19.
13. Robinson DR, Gull K (1991) Basal body movements as a mechanism for mitochondrial
genome segregation in the trypanosome cell cycle. Nature 352: 731-733.
14. Moreira-Leite FF, Sherwin T, Kohl L, Gull K (2001) A trypanosome structure involved
in transmitting cytoplasmic information during cell division. Science 294: 610-612.
15. Paulin JJ, Keith CH, Tarleton RL (1988) A monoclonal antibody to alpha tubulin
recognizes host cell and Trypanosoma cruzi tubulins. J Protozool 35: 123-129.
16. Saborio JL, Wrightsman RA, Kazuko SG, Granger BS, Manning JE (1990)
Trypanosoma cruzi: identification of a surface antigen restricted to the flagellar
region of the infective form of the parasite. Exp Parasitol 70: 411-418.
17. Segura EL, Vazquez C, Bronzina A, Campos JM, Cerisola JA, et al. (1977) Antigens
of the subcellular fractions of Trypanosoma cruzi. II. Flagellar and membrane
fraction. J Protozool 24: 540-543.
18. Van Voorhis WC, Eisen H (1989) Fl-160. A surface antigen of Trypanosoma cruzi that
mimics mammalian nervous tissue. J Exp Med 169: 641-652.
19. Rocha GM, Teixeira DE, Miranda K, Weissmuller G, Bisch PM, et al. (2010)
Structural changes of the paraflagellar rod during flagellar beating in Trypanosoma
cruzi. PLoS One 5: e11407.
20. Quanquin NM, Galaviz C, Fouts DL, Wrightsman RA, Manning JE (1999)
Immunization of mice with a TolA-like surface protein of Trypanosoma cruzi
generates CD4(+) T-cell-dependent parasiticidal activity. Infect Immun 67: 4603-
4612.

Trabajo Final - Lobo Maite Bibliografía
57
21. Cooley G, Etheridge RD, Boehlke C, Bundy B, Weatherly DB, et al. (2008) High
throughput selection of effective serodiagnostics for Trypanosoma cruzi infection.
PLoS Negl Trop Dis 2: e316.
22. El-Sayed NM, Myler PJ, Bartholomeu DC, Nilsson D, Aggarwal G, et al. (2005) The
genome sequence of Trypanosoma cruzi, etiologic agent of Chagas disease.
Science 309: 409-415.
23. Franzen O, Ochaya S, Sherwood E, Lewis MD, Llewellyn MS, et al. (2011) Shotgun
sequencing analysis of Trypanosoma cruzi I Sylvio X10/1 and comparison with T.
cruzi VI CL Brener. PLoS Negl Trop Dis 5: e984.
24. Grisard EC, Teixeira SM, de Almeida LG, Stoco PH, Gerber AL, et al. (2014)
Trypanosoma cruzi Clone Dm28c Draft Genome Sequence. Genome Announc 2.
25. Franzen O, Talavera-Lopez C, Ochaya S, Butler CE, Messenger LA, et al. (2012)
Comparative genomic analysis of human infective Trypanosoma cruzi lineages
with the bat-restricted subspecies T. cruzi marinkellei. BMC Genomics 13: 531.
26. Stoco PH, Wagner G, Talavera-Lopez C, Gerber A, Zaha A, et al. (2014) Genome of
the avirulent human-infective trypanosome—Trypanosoma rangeli. PLoS Negl
Trop Dis 8: e3176.
27. Berriman M, Ghedin E, Hertz-Fowler C, Blandin G, Renauld H, et al. (2005) The
genome of the African trypanosome Trypanosoma brucei. Science 309: 416-422.
28. Ivens AC, Peacock CS, Worthey EA, Murphy L, Aggarwal G, et al. (2005) The
genome of the kinetoplastid parasite, Leishmania major. Science 309: 436-442.
29. Nakayasu ES, Sobreira TJ, Torres R, Jr., Ganiko L, Oliveira PS, et al. (2012) Improved
proteomic approach for the discovery of potential vaccine targets in Trypanosoma
cruzi. J Proteome Res 11: 237-246.
30. Brunoro GV, Caminha MA, Ferreira AT, Leprevost Fda V, Carvalho PC, et al. (2015)
Reevaluating the Trypanosoma cruzi proteomic map: The shotgun description of
bloodstream trypomastigotes. J Proteomics 115: 58-65.
31. Lantos AB, Carlevaro G, Araoz B, Ruiz Diaz P, Camara Mde L, et al. (2016) Sialic
Acid Glycobiology Unveils Trypanosoma cruzi Trypomastigote Membrane
Physiology. PLoS Pathog 12: e1005559.
32. Notredame C, Higgins DG, Heringa J (2000) T-Coffee: A novel method for fast and
accurate multiple sequence alignment. J Mol Biol 302: 205-217.
33. Saitou N, Nei M (1987) The neighbor-joining method: a new method for reconstructing
phylogenetic trees. Mol Biol Evol 4: 406-425.
34. Efron B (2011) The bootstrap and Markov-chain Monte Carlo. J Biopharm Stat 21:
1052-1062.
35. Letunic I, Bork P (2016) Interactive tree of life (iTOL) v3: an online tool for the
display and annotation of phylogenetic and other trees. Nucleic Acids Res 44:
W242-245.
36. Pierleoni A, Martelli PL, Casadio R (2008) PredGPI: a GPI-anchor predictor. BMC
Bioinformatics 9: 392.
37. Blom N, Gammeltoft S, Brunak S (1999) Sequence and structure-based prediction of
eukaryotic protein phosphorylation sites. J Mol Biol 294: 1351-1362.
38. Julenius K, Molgaard A, Gupta R, Brunak S (2005) Prediction, conservation analysis,
and structural characterization of mammalian mucin-type O-glycosylation sites.
Glycobiology 15: 153-164.
39. Zhou F, Xue Y, Yao X, Xu Y (2006) CSS-Palm: palmitoylation site prediction with a
clustering and scoring strategy (CSS). Bioinformatics 22: 894-896.

Trabajo Final - Lobo Maite Bibliografía
58
40. Durante IM, Camara Mde L, Buscaglia CA (2015) A Novel Trypanosoma cruzi Protein
Associated to the Flagellar Pocket of Replicative Stages and Involved in Parasite
Growth. PLoS ONE 10: e0130099.
41. Urban I, Santurio LB, Chidichimo A, Yu H, Chen X, et al. (2011) Molecular diversity
of the Trypanosoma cruzi TcSMUG family of mucin genes and proteins. Biochem
J 438: 303-313.
42. Ruijter JM, Ramakers C, Hoogaars WM, Karlen Y, Bakker O, et al. (2009)
Amplification efficiency: linking baseline and bias in the analysis of quantitative
PCR data. Nucleic Acids Res 37: e45.
43. Tuomi JM, Voorbraak F, Jones DL, Ruijter JM (2010) Bias in the Cq value observed
with hydrolysis probe based quantitative PCR can be corrected with the estimated
PCR efficiency value. Methods 50: 313-322.
44. Pfaffl MW (2001) A new mathematical model for relative quantification in real-time
RT-PCR. Nucleic Acids Res 29: e45.
45. Campo V, Di Noia JM, Buscaglia CA, Aguero F, Sanchez DO, et al. (2004)
Differential accumulation of mutations localized in particular domains of the mucin
genes expressed in the vertebrate host stage of Trypanosoma cruzi. Mol Biochem
Parasitol 133: 81-91.
46. Buscaglia CA, Alfonso J, Campetella O, Frasch AC (1999) Tandem amino acid repeats
from Trypanosoma cruzi shed antigens increase the half-life of proteins in blood.
Blood 93: 2025-2032.
47. Inoue H, Nojima H, Okayama H (1990) High efficiency transformation of Escherichia
coli with plasmids. Gene 96: 23-28.
48. Balouz V, Camara Mde L, Canepa GE, Carmona SJ, Volcovich R, et al. (2015)
Mapping Antigenic Motifs in the Trypomastigote Small Surface Antigen from
Trypanosoma cruzi. Clin Vaccine Immunol 22: 304-312.
49. Campo VA, Buscaglia CA, Di Noia JM, Frasch AC (2006) Immunocharacterization of
the mucin-type proteins from the intracellular stage of Trypanosoma cruzi.
Microbes Infect 8: 401-409.
50. Di Noia JM, Buscaglia CA, De Marchi CR, Almeida IC, Frasch AC (2002) A
Trypanosoma cruzi small surface molecule provides the first immunological
evidence that Chagas’ disease is due to a single parasite lineage. J Exp Med 195:
401-413.
51. Canepa GE, Degese MS, Budu A, Garcia CR, Buscaglia CA (2012) Involvement of
TSSA (trypomastigote small surface antigen) in Trypanosoma cruzi invasion of
mammalian cells. Biochem J 444: 211-218.
52. Carmona SJ, Nielsen M, Schafer-Nielsen C, Mucci J, Altcheh J, et al. (2015) Towards
High-throughput Immunomics for Infectious Diseases: Use of Next-generation
Peptide Microarrays for Rapid Discovery and Mapping of Antigenic Determinants.
Mol Cell Proteomics 14: 1871-1884.
53. Buscaglia CA, Campo VA, Frasch AC, Di Noia JM (2006) Trypanosoma cruzi surface
mucins: host-dependent coat diversity. Nat Rev Microbiol 4: 229-236.
54. Minning TA, Weatherly DB, Atwood J, 3rd, Orlando R, Tarleton RL (2009) The
steady-state transcriptome of the four major life-cycle stages of Trypanosoma cruzi.
BMC Genomics 10: 370.
55. Canepa GE, Mesias AC, Yu H, Chen X, Buscaglia CA (2012) Structural Features
Affecting Trafficking, Processing, and Secretion of Trypanosoma cruzi Mucins. J
Biol Chem 287: 26365-26376.

Trabajo Final - Lobo Maite Bibliografía
59
56. Cerqueira GC, Bartholomeu DC, DaRocha WD, Hou L, Freitas-Silva DM, et al. (2008)
Sequence diversity and evolution of multigene families in Trypanosoma cruzi. Mol
Biochem Parasitol 157: 65-72.
57. Buscaglia CA, Campo VA, Di Noia JM, Torrecilhas AC, De Marchi CR, et al. (2004)
The surface coat of the mammal-dwelling infective trypomastigote stage of
Trypanosoma cruzi is formed by highly diverse immunogenic mucins. J Biol Chem
279: 15860-15869.
58. Bartholomeu DC, Cerqueira GC, Leao AC, daRocha WD, Pais FS, et al. (2009)
Genomic organization and expression profile of the mucin-associated surface
protein (masp) family of the human pathogen Trypanosoma cruzi. Nucleic Acids
Res 37: 3407-3417.
59. Di Noia JM, D’Orso I, Aslund L, Sanchez DO, Frasch AC (1998) The Trypanosoma
cruzi mucin family is transcribed from hundreds of genes having hypervariable
regions. J Biol Chem 273: 10843-10850.
60. Mucci J, Lantos AB, Buscaglia CA, Leguizamon MS, Campetella O (2017) The
Trypanosoma cruzi Surface, a Nanoscale Patchwork Quilt. Trends Parasitol 33:
102-112.
61. Lazzaroni JC, Dubuisson JF, Vianney A (2002) The Tol proteins of Escherichia coli
and their involvement in the translocation of group A colicins. Biochimie 84: 391-
397.
62. Anderluh G, Hong Q, Boetzel R, MacDonald C, Moore GR, et al. (2003) Concerted
folding and binding of a flexible colicin domain to its periplasmic receptor TolA. J
Biol Chem 278: 21860-21868.
63. Goemaere EL, Cascales E, Lloubes R (2007) Mutational analyses define helix
organization and key residues of a bacterial membrane energy-transducing
complex. J Mol Biol 366: 1424-1436.

Trabajo Final - Lobo Maite
Carlos A. Buscaglia
Director
Di
Virginia Balouz
Codirectora
Lobo Maite M.
Alumna


















