Preferencia y eficacia de Notonecta indica y Luis A ... · por ser vectores de enfermedades...
30
Preferencia y eficacia de Notonecta indica (Notonectidae) para diferentes instares larvales de Aedes aegypti (Culicidae) Cristina Zapata-Maspoli * y Luis A. Guerra* * Estudiantes de Biología, Universidad Nacional de Colombia, sede Caribe; pasantes Entomología médica del Laboratorio de Salud Pública, Secretaria de Salud departamental de San Andrés Islas, Colombia. Correo electrónico: [email protected], [email protected] RESUMEN Con el fin de establecer las habilidades de predación a partir de sus preferencias y eficacias tróficas hacia larvas de diferentes instares de Aedes aegypti, se empleó el insecto Notonecta indica, predador de pequeños invertebrados en comunidades acuáticas. Esta especie se encontró en cuerpos de agua en la isla de San Andrés, Caribe Colombiano. Se separaron las pruebas según el tipo de instar de N. indica. Se contó siempre el número de larvas predadas por los diferentes instares del predador. En los bioensayos se encontró que N. indica tiene preferencias para predar larvas de Ae. aegypti de instares de mayor tamaño. Además, se obtuvo que N. indica preda en mayor número larvas de Ae. aegypti de instares iniciales cuando no encuentra más opciones. Con este trabajo se avanzó hacia la consolidación de una línea base que permitan la elaboración de planes de control biológico de mosquitos en la isla de San Andrés y sistemas semejantes. Palabras Claves: Biocontrol, Mosquitos, relación predador-presa, preferencia, eficacia del predador.
Transcript of Preferencia y eficacia de Notonecta indica y Luis A ... · por ser vectores de enfermedades...

Preferencia y eficacia de Notonecta indica (Notonectidae) para diferentes instares larvales de Aedes aegypti (Culicidae)
Cristina Zapata-Maspoli* y Luis A. Guerra*
* Estudiantes de Biología, Universidad Nacional de Colombia, sede Caribe; pasantes Entomología médica del Laboratorio de Salud Pública, Secretaria de Salud
departamental de San Andrés Islas, Colombia.
Correo electrónico: [email protected], [email protected]
RESUMEN
Con el fin de establecer las habilidades de predación a partir de sus preferencias y eficacias
tróficas hacia larvas de diferentes instares de Aedes aegypti, se empleó el insecto Notonecta
indica, predador de pequeños invertebrados en comunidades acuáticas. Esta especie se
encontró en cuerpos de agua en la isla de San Andrés, Caribe Colombiano. Se separaron las
pruebas según el tipo de instar de N. indica. Se contó siempre el número de larvas predadas
por los diferentes instares del predador. En los bioensayos se encontró que N. indica tiene
preferencias para predar larvas de Ae. aegypti de instares de mayor tamaño. Además, se
obtuvo que N. indica preda en mayor número larvas de Ae. aegypti de instares iniciales
cuando no encuentra más opciones. Con este trabajo se avanzó hacia la consolidación de
una línea base que permitan la elaboración de planes de control biológico de mosquitos en
la isla de San Andrés y sistemas semejantes.
Palabras Claves: Biocontrol, Mosquitos, relación predador-presa, preferencia, eficacia del
predador.

2
ABSTRACT
We used the water-bug Notonecta indica, a little invertebrate predator in aquatic
community, to know their predator abilities since the preference and predator efficacy in
different larval instars of Aedes aegypti. We found this predator in water bodies at San
Andrés Island, Caribbean of Colombia. We carry out each one tests beginning from
different instars of N. indica. We ever count up the number of larval Ae. aegypti predated
by each instars of N. indica in any test. The bioassays show N. indica have preferences to
prey late larval instars of Ae. aegypti. We found N. indica prey more initial than late larval
instars of Ae. aegypti when it have no more options to choose. This work was carried out by
us to give a baseline to develop researches and projects of mosquito biocontrol at the San
Andrés Island and related systems.
Key Words: Biocontrol, Mosquitoes, predator-prey relationship, preference, predator
efficacy.

3
INTRODUCCIÓN
Algunas especies de los géneros Culex, Aedes, Anopheles y otros representantes de la
familia Culicidae, principalmente distribuidos en el trópico (Lane, 1953), se caracterizan
por ser vectores de enfermedades tropicales como la fiebre amarrilla, dengue, filaríasis,
malaria, entre otras muchas de importancia médica y biológica (Lane, 1953). Estas
enfermedades representan una constante amenaza para la salud pública del Caribe y de
otras zonas tropicales, por lo que es justificable adelantar trabajos que propicien el control
de estos agentes transmisores y sus fuentes de infección.
Las especies de Aedes se encuentran principalmente asociadas a asentamientos humanos.
Regularmente se hallan en recipientes domiciliarios, en cuerpos de agua potable hasta en
aguas turbias y ricas en materia orgánica. Aedes aegypti Linnaeus 1762, se considera vector
de enfermedades epidemiológicas importantes como la fiebre amarilla y el dengue
(Consoli y De Oliveira, 1998). Por ello es importante el control de estos insectos vectores
para evitar la propagación de cualquier enfermedad importada. En el caso de las islas del
gran Caribe, en las que se desarrolla una economía principalmente turística y comercial, los
puertos, cabeceras urbanas y hoteles de mayor capacidad de ocupación se encuentran muy
cercanos a zonas de humedal, depósitos de agua abandonados, lagunas o charcas no
permanentes.
Hasta el momento se han implementado diversos métodos de erradicación de estos
vectores. En el Caribe se ha empleado el uso de insecticidas que han contribuido al
aumento de la resistencia de los mosquitos a estos agentes químicos (Rawlins, 1998). Éstos
en algunos casos son poco fructíferos, obligando a repetirlos periódicamente con el fin de

4
disminuir las posibilidades de brotes de las enfermedades transmitidas por estos (Nelson,
1986).
Las estrategias empleadas hasta el momento para el control de mosquitos vectores
van desde el uso de agentes químicos hasta el uso de agentes biológicos. Estos últimos son
organismos con la capacidad de parasitar, predar al insecto vector, entre otras (Chapman,
1974). Sin embargo, muchas de estas estrategias generan impactos que pueden ser
invaluables para comunidades biológicas y humanas. Pues en general, muchas de las
especies empleadas hasta el momento para el control biológico de organismo son extrañas
al hábitat en donde se han introducido, generando a su llegada desequilibrios entre
poblaciones de los sitios intervenidos (Murdoch et al., 1985; Lawler et al., 1999). Muchas
de estas especies controladoras son consideradas amables en comparación con los
controladores químicos. En diferentes países tropicales se han empleado estrategias de
control biológico de mosquito. Brasil, por ejemplo, se ha empleado hasta el momento
Bacillus thuringiensis var. israelensis como biocontrolador de mosquito (Lima et al.,
2005). También se registran estudios de control de mosquito con especies animales como
peces (Bence, 1998; Lee, 2000; Hurst et al., 2004), ranas (Willems et al., 2005), planarias
(Perich et al., 1990), decápodos (Mkoji et al., 1999), libélulas (Quiroz-Martínez et al.,
2005), larvas de otros mosquitos (Mercer et al, 2005) e incluso algunas especies del género
Notonecta que han sido empleadas en conjunto con bacterias (Neri-Barbosa et al., 1997).
Debido a la situación biogeográfica de la isla de San Andrés, reconocida por sus
ecosistemas costeros marinos, fue declarada el diez de noviembre del 2000 como Reserva
de la Biosfera SEAFLOWER por UNESCO. Esto propicia el desarrollo de planes que

5
conserven la diversidad biológica de la isla, que puede afectarse por la introducción de
especies foráneas. Para evitar la introducción de especies se sugiere la implementación de
programas de control con organismos nativos para disminuir posibles impactos sobre los
frágiles ecosistemas de la isla.
Algunas especies de Notonecta, también llamados backswimmer, se han
identificado como predadores de larvas de mosquitos y otros invertebrados planctónicos
(Blaustein, 1998; Chesson, 1989; Sih, 1986; Murdoch y Sih, 1978). La importancia de estos
organismos radica en sus hábitos alimentarios y su natural disponibilidad en zonas de esta
isla, como la nativa Notonecta indica Linnaeus 1771. Se ha encontrado para especies de
Notonecta que existen preferencias hacia el tamaño de sus presas (Scott y Murdoch, 1983).
Las especies de Notonecta presentan seis instares que dependen de los cambios en tamaño y
muda, de los cuales el último corresponde al adulto (Mc Ardle y Lawton, 1979; Scott y
Murdoch, 1983; Neri-Barbosa et al., 1997). Murdoch y Sih (1978) han sugerido para
Notonecta hoffmanni que en una población con diferentes clases de edades existe
interferencia por parte de las formas adultas sobre las juveniles en cuanto a su tasa de
predación o número de larvas predadas en el tiempo.
Con el presente estudio se desea conocer las habilidades predadoras de N. indica
hacia los diferentes instares larvales de mosquitos transmisores de enfermedades tropicales
como Ae. aegypti, a partir de su eficacia trófica. Esta eficacia se evalúa a partir del número
de larvas predadas en un tiempo estipulado. Además se pretende estudiar los fenómenos de
preferencia por parte de N. indica hacia algún o algunos instares larvales de Aedes aegypti y
cómo se relacionan estos fenómenos de preferencia con el grado de desarrollo de N. indica.

6
MATERIALES Y MÉTODOS
AREA DE ESTUDIO
El presente estudio se realizó entre el 6 de Febrero de 2008 y el 24 de Mayo de 2008 en la
isla de San Andrés ubicada en el Caribe Colombiano entre los 12º28’ - 12º35’ N y 72º29’ -
81º44’. En esta isla se presentan dos picos de lluvias al año, entre mayo-junio y octubre-
noviembre, y la temperatura promedio es de 27.4º C, precipitación media anual de 1900
mm, su superficie es de 25 km2. Su actividad económica se orienta hacia el sector turístico
y comercial (Aguado, 2004).
MATERIAL BIOLÓGICO
El material biológico utilizado en el presente estudio provino de material recolectado en
campo compuesto principalmente por N. indica y de material suministrado por el
laboratorio de salud pública de la isla que corresponde a larvas de Ae. aegypti.
El predador del mosquito
Para las diferentes pruebas realizadas se utilizaron individuos de N. indica que fueron
recolectados en campo el día 25 de abril del 2008, en horas de la mañana, en dos piscinas
abandonadas al oeste de la isla (12º 33’ 50.2’’ N, 81º 43’ 30.3’’ W). La de mayor tamaño
tenía un cuerpo de agua de no más de 80 cm de profundidad, un ancho de 9.60m y una
longitud de 17.60m, pH de 8.00, 4.85 mg/ L de oxigeno disuelto, conductividad de 2250
µS/ cm y temperatura de 32º C. La menor de las piscinas tenía un cuerpo de agua de 20 cm
de profundidad, un ancho de 3.70m, una longitud de 5.80m, pH de 8.65, 4.94 mg/ L de
oxigeno disuelto, conductividad de 2450 µS/ cm y temperatura de 28º C. Estas variables

7
se midieron empleando sondas multiparámetros WTW (Wissenschaftich-Technische
Werstätten) ref. 330i/340i para medición de pH, Oxigeno disuelto, Conductividad y
temperatura.
Se recolectaron N. indica de sexto instar (67 individuos), de quinto instar (75 individuos),
de cuarto instar (52 individuos) y de tercer instar (tan solo cinco individuos). Para el
proceso de recolección se emplearon coladores plásticos de 21 cm de diámetro. Los
individuos recolectados pudieron mantener su número e incluso cambiar de instar a lo largo
del periodo de estudios en agua tratada de llave. Después de los experimentos se encontró
que este organismo podía reproducirse en el laboratorio. Se obtuvieron 431 huevos de los
cuales eclosionaron 284, sin embargo, solo nueve N. indica lograron mantenerse hasta el
último día de estudios.
Se han empleado para los experimentos instares avanzados de N. indica. Estos van entre el
cuarto instar hasta el sexto y se han nombrado como N4, N5 y N6. En Fig. 1 se expone una
muestra de N. indica ordenada según su instar. Se omite el segundo instar por falta
individuos recolectados.
El mosquito
En el laboratorio se contó con material vivo de Ae. aegypti en todas sus fases. Se empleó
las cantidades disponibles para cada instar larval del mosquito. Se ha nombrado del primer
al cuarto instar larval del mosquito como L1, L2, L3 y L4 (Fig. 2). Estas larvas se
obtuvieron a partir de un proceso de producción de huevos de mosquito que son
almacenados en condiciones aisladas no totalmente deshidratadas donde los huevos se

8
conservaban en estado quiescente. Estos huevos bajo ciertas condiciones pueden
mantenerse viables y eclosionar después de un año (Consoli y de Oliveira, 1998). Estos
huevos que corresponden a cuarta generación (F4) de larvas capturadas en campo pueden
ser reactivados en condiciones óptimas de humedad siempre y cuando no superen los cuatro
meses de almacenamiento.
ACLIMATACIÓN DEL MATERIAL BIOLÓGICO
Las N. indica se trasladaron a laboratorio. Se ubicaron en recipientes de plástico con agua
de llave con un pH de 7.7, 4.81 mg de O2/ mL, y 654 µS/ cm de conductividad, con una
temperatura de 26º C. Para las pruebas en que se utilizó este organismo se les negó
alimentación por lo menos de un día para asegurarse que no se sacien durante el
experimento como lo sugiere McArdle (1979). Con el fin de disminuir el estrés por
disturbios en su traslado a los recipientes de prueba se les dejó reposar 15 min antes de
iniciar cada bioensayo.
BIOENSAYOS EN CONDICIONES SEMICONTROLADAS
Prueba de densidades predador-presa
El experimento se llevó a cabo para determinar si la tasa de predación por las diferencias en
número de las poblaciones tanto del predador como de la presa. Para esto se propusieron
tres tratamientos que conservaron la densidad del predador y presa en un volumen de agua,
manteniendo entre los organismos siempre una relación 1: 5 (N. indica: larva de Ae.
aegypti). En la prueba se utilizó N. indica en su quinto instar (N5) y larvas de Ae. aegypti
en su segundo instar (L2). Para los tres tratamientos: T1, T2 y T3, fue constante la
luminosidad y temperatura del agua (28ºC). En el primer tratamiento (T1) se emplearon

9
dos N. indica por diez larvas de Ae. aegypti en 37.5 mL de agua, para el siguiente
tratamiento (T2) se emplearon ocho N. indica por 40 larvas de Ae. aegypti en 150 mL de
agua y por último, en el tercer tratamiento (T3), se emplearon 16 N. indica por 80 larvas de
Ae. aegypti en 350 mL de agua. Para todos los casos se utilizó recipientes transparentes.
Prueba de eficacia trófica según el tipo de larva
Este experimento se llevó a cabo con el fin de comparar la eficacia trófica de N. indica para
predar, según el instar utilizado (N4, N5 y N6), cada instar larval de Ae. aegypti. La
eficacia se ha definido como el número de presas predadas en un tiempo determinado. En
las pruebas de eficacia se utilizaron siempre grupos de tres N. indica. Cada instar de este
predador se enfrentó a cada tipo de instar larval de Ae. aegypti obteniéndose cuatro
tratamientos para cada instar de N. indica. Para cada tratamiento se utilizó un volumen
constante de 380 mL de agua a 24ºC, con una profundidad de 8.7 cm, luminosidad
permanente y un número de 20 larvas por réplica. Se contó el número de larvas que fueron
predadas por N. indica en 30 minutos de tiempo. Se emplearon beakers de 400 mL como
recipientes, cubiertos con papel aluminio en sus paredes laterales. Cada tratamiento vario
en el número de replicas entre tres y siete debido a la disponibilidad en número de cada
instar larval de Ae. aegypti y el número de N. indica recolectadas.
Prueba de preferencia trófica por estadio de larva
Se realizó este experimento con el fin de determinar si N. indica presenta preferencias
tróficas hacia alguno o algunos de los instares larvales de Ae. aegypti. En la prueba de
preferencias se utilizaron los tres últimos instares de N. indica (N4, N5 y N6) y los cuatro
instares larvales de Ae. aegypti (L1, L2, L3 y L4). Para cada instar de N. indica se usaron

10
cinco beakers de 400 mL con 380 mL de agua, con una profundidad de 8.7 cm, a una
temperatura de 24º C, con tres N. indica/ beaker. En cada recipiente se suministró diez
larvas de cada instar larval (40 larvas/ beaker). Finalizado el experimento, se procedió a
separar las larvas muertas, se clasificaron y contaron según el instar larval.
ANALISIS ESTADISTICO
Se realizaron análisis de varianzas para todas las pruebas en las que se aceptó la
distribución normal de los datos según el estadístico prueba de Ryan-Joiner (RJ) similar a
Shapiro-Wilk con valor de probabilidad mayor al grado de significancia. Las medias
encontradas para los diferentes tratamientos estudiados se reportan con su respectivo error
estándar (SE) y desviación estándar (St. Dev). Se presentan intervalos de confianza del
95% (CI).
Para aquellas pruebas en las que se realizó el análisis de varianza, y se rechazó la hipótesis
de inexistencia de diferencias significativas entre los efectos de los tratamientos (P< 0.05;
Fcalculado> Ftabulado), se prosiguió con la prueba de comparaciones múltiples pareadas de
Tukey que compara entre las diferencias mínimas entre las medias de los tratamientos. Los
resultados de las pruebas, tablas y las gráficas reportadas se han adaptado de los arrojados
por el software estadístico MINITAB 15. 30day-trial.
RESULTADOS
PRUEBA DE DENSIDADES PREDADOR-PRESA
En esta prueba se registraron los tiempos requeridos en cada tratamiento para que las N.
indica predaran la totalidad de larvas dispuestas por recipiente. Los datos de estos tiempos

11
medidos en minutos se reportan en Tabla 1. Se encontró que no hay diferencias
significativas entre los tratamientos (Tabla 2), lo que sugiere que no hay cambios o
diferencias en la tasa de predación cuando se mantiene la densidad de predador-presa
constante y la relación numérica entre predador y presa (1: 5).
PRUEBA DE EFICACIA TRÓFICA SEGÚN EL TIPO DE LARVA
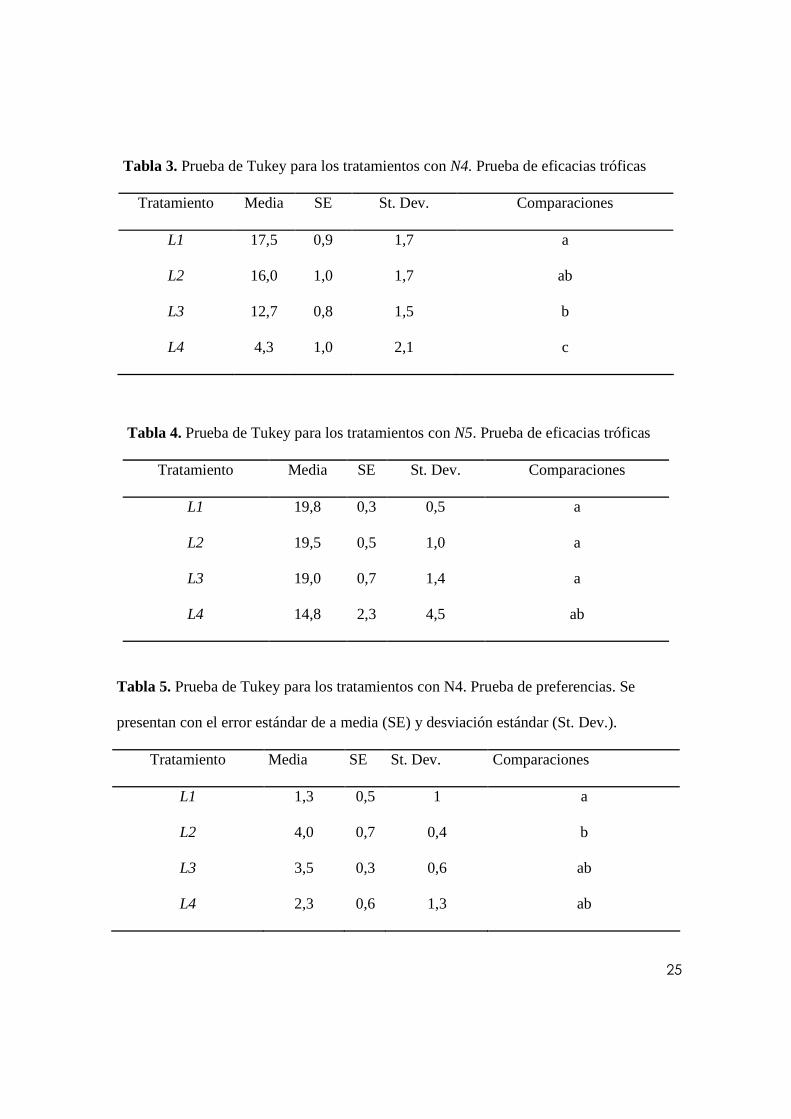
Se separaron los resultados según el tipo de N. indica empleada para cada tratamiento. El
número promedio de larvas predadas por cada uno de los tres instares de N. indica se
presentan en la Fig. 3. Se realizó un análisis de varianza para los tratamientos con N4 y N5
para probar si existían diferencias significativas entre el número de larvas predadas en 30
minutos para cada tratamiento. Para el caso de N6 no se hace dicho análisis, puesto que los
datos son muy explícitos, y sugieren notables diferencias entre el número de larvas de
cuarto instar (L4) predadas respecto a las otras formas larvales de Ae. aegypti. Tanto para
N4 (α= 0.05, p< 0.000) como para N5 (α= 0.05, p< 0.041) se encontraron diferencias
significativas. Se realizó un procedimiento de comparaciones múltiples de Tukey a partir de
la premisa de que los datos son normales a partir de la prueba de Ryan-Joiner (RJ) para N4
(RJ= 0.981, p> 0.100) y N5 (RJ= 0.954, p>0.100) y que presentan diferencias significativas
entre los tratamientos asignados según el tipo de larva. Los resultados de este análisis se
reportan en las tablas 3 y 4.
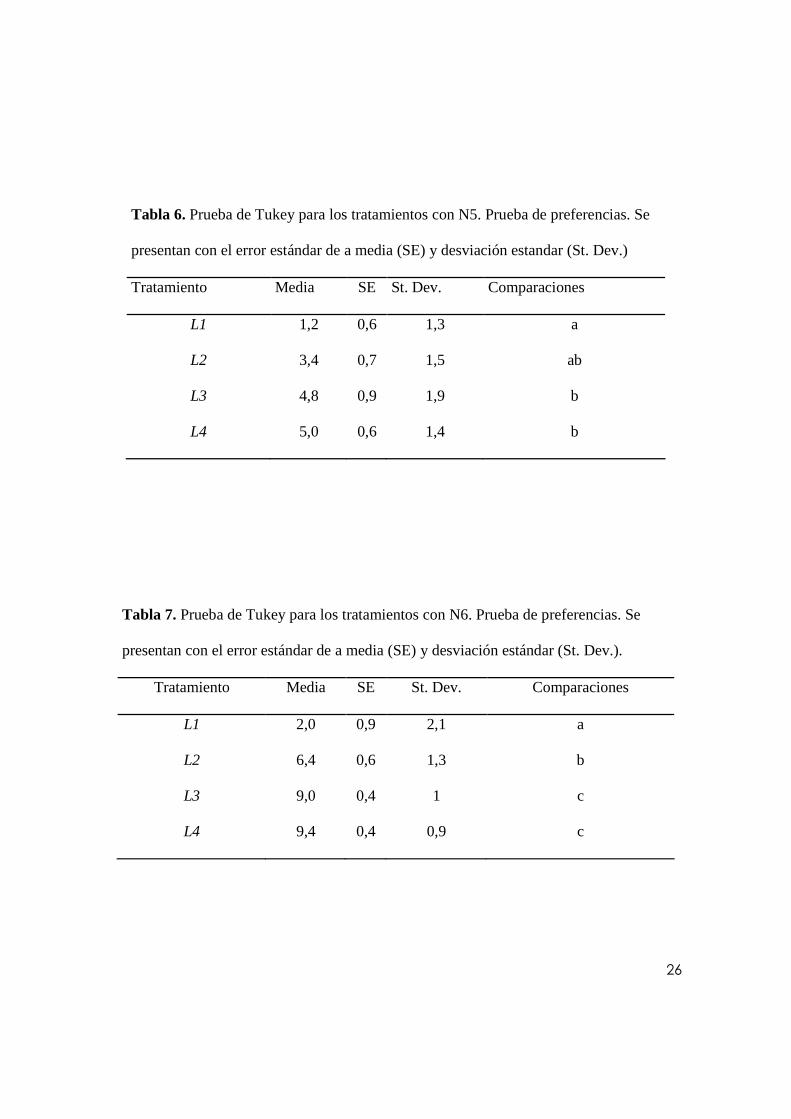
PRUEBA DE PREFERENCIA TRÓFICAS POR ESTADIO DE LARVA
Se asumió que cada tratamiento corresponde al número de larvas de un mismo instar que
fueron predadas por un instar particular de N. indica. El promedio de larvas predadas para
cada instar larval del mosquito se han agrupado según el instar del predador (Fig. 4). Cabe

12
anotar que este experimento se diferenció del anterior en cuanto a que cada recipiente
contenía los cuatro tipos de larva de mosquito en conjunto. Para cada grupo de datos
asociado a un tipo de predador se ha realizado un análisis de varianza (α= 0.05) para probar
si existían diferencias significativas entre los tratamientos. Se obtuvo tanto para N4 (p=
0.017) como en N5 (p= 0.005) y N6 (p= 0.000) que por lo menos uno de los tipos larvales
presentaba diferencias respecto a los demás en cuanto a la proporción absoluta o número de
larvas predadas. Se corroboró que cada grupo de datos se distribuye de manera normal (N4:
RJ= 0.98, p>0.10; N5: RJ= 0.97, p>0.10; N6: RJ= 0.98, p>0.10). A partir de estos
resultados se ha optado por realizar una prueba Tukey para cada grupo de datos analizado.
Los resultados de esta prueba se reportan en las tablas 5, 6 y 7. Se ha asociado la media de
cada tratamiento a un grupo nombrado con una letra del alfabeto; los tratamientos que
guardan semejanzas entre sí comparten la misma letra o grupo.
DISCUSIÓN
Murdoch y Sih (1978) encontraron fuertes relaciones entre diferentes densidades del
predador N. hoffmanni y de su presa (larvas de mosquito). Esta relación interviene en el
número de larvas consumidas finalmente por el instar del predador. Instares maduros de
Notonecta interfirieren en la tasa de alimentación de los instares juveniles de este predador.
Por esto se ha decidido separar las pruebas según el instar de Notonecta.
En la primera prueba, manteniendo siempre la misma relación de abundancia de N. indica y
de su presa, se pretendió evaluar si al variar el número del predador se afectaba la tasa de
predación de este. En este bioensayo no se evidenció diferencias en cuanto a la tasa de
larvas predadas. Se determinó que no hay diferencias significativas que demuestren una

13
dependencia de la tasa de predación respecto al número de predadores presentes en los
medios empleados. Se pensó que estos resultados podrían soportar la hipótesis de que es lo
mismo trabajar con un número bajo de predadores que trabajar con un mayor número de
estos.
Pese a que el experimento sugiere que pueden aplicarse pruebas con un bajo número de N.
indica, se decidió no continuar con esta metodología pues no todos los factores pueden ser
controlados por esta. En el momento de hacer el experimento las N. indica empleadas
asumían comportamientos anormales para el experimento evidenciados por el posible estrés
asociado a choques que estas sufrían contra las paredes translucidas de los recipientes, el
desinterés de predar durante todo el experimento e incluso el abandono del recipiente
disminuían la competencia por la presa entre los predadores aun presentes. Incluso, la
dificultad de predar en cuerpos de agua muy poco profundos puede ser también un factor de
estrés. En este bioensayo, la mayor altura que podía tener un cuerpo de agua mínima fue de
no más de ocho mm y la máxima fue de 30 mm muy por debajo de lo empleado por
McArdle y Lawton (1979).
La forma de tomar los tiempos fue crucial para la obtención de resultados. En la prueba de
densidades se esperó que cada grupo del predador agotara la totalidad de las presas. Esto no
permite excluir o minimizar el efecto de la saciedad del predador. Esto incluye un gran
error al experimento pues esto no permitiría obtener datos controlados acerca de la tasa de
predación (McArdle y Lawton, 1979). Se considera por parte de los autores como adecuado
tomar datos para la tasa de predación (Número de larvas predadas/ min) por debajo del
punto de saciedad. Para el caso de N. indica no se conoce este punto de saciedad

14
relacionado con el número de presas consumidas. Para evitar el efecto de la saciedad del
predador, se tomó el experimento de densidades como exploratorio para los siguientes
experimentos. En este se observó que dos, ocho e incluso 16 N. indica aun después de una
hora continuaban alimentándose de larvas de mosquitos del instar cuarto. Por esto se
decidió que para posteriores pruebas se evaluaría el número de larvas consumidas en un
tiempo no mayor a 30 min. La optimización o mejora de este experimento no se logró
concluir puesto que no se contó con el material biológico necesario para rehacer dichas
pruebas.
Se supone que este predador como posible control biológico de larvas de mosquito debe
operar de forma colectiva en cuerpos de agua y no de manera individual o sin compañía. En
campo este predador tendrá que someterse al efecto de la competencia intra-especifica e
incluso ínter-especifica. Esto último, pues existen otras especies que se han encontrado
compartiendo hábitat con este organismo y que también predan larvas de mosquito, tal es el
caso de individuos (instares inmaduros) recolectados de la familia Libellulidae, algunos
especímenes del género Buenoa (Notonectidae), entre otros. De estos grupos se
recolectaron algunos especímenes vivos tomados en campo en el momento de la captura de
las N. indica. Se pudo constatar en campo y en laboratorio que tanto Buenoa sp. como los
nayares de libélulas recolectadas predan a N. indica y larvas de mosquitos de la familia
Culicidae como Ae. aegypti y Culex quinquefasciatus.
McArdle y Lawton (1979) emplearon Notonecta para hallar los efectos predador-presa y el
instar de este predador en un organismo presa como Daphnia con diferentes clases de
tamaño. Para este experimento McArdle y Lawton utilizaron una profundidad constante de

15
cinco cm y empleando un individuo del predador contra diferentes abundancias de la presa.
En este estudio encontró que los dos últimos instares juveniles de Notonecta tienen una tasa
de ataque o predación mayor que la del último instar de Notonecta (Adulto). En nuestro
trabajo se encontró que la tasa de predación cambiaba según el experimento analizado.
Estos se explican a continuación:
En el experimento de eficacia según tipo de larva, las mayores tasas de predación del
predador, que pueden definirse como el número medio de larvas predadas en 30 min,
corresponden a aquellos instares de N. indica que predaban las larvas de menor tamaño (L1,
L2 y L3). Para esta prueba no se permitía que el predador eligiese sus presas según el
tamaño o volumen de fluidos corporales que pudiese ser aprovechado de estos
invertebrados. Es probable que estos resultados se asocien a ventajas debidas a las
adaptaciones de alguno de los organismos estudiados. En el caso del predador es posible
que posea ventajas para atacar y atrapar a presas de menor tamaño o que estas por sí
mismas sean más vulnerables que las formas más desarrolladas al ataque del predador. En
el caso de la larva de mayor tamaño (L4) se obtuvo que cada una de las diferentes N. indica
predaban un menor número de esta presa. Es posible que L4 tenga ventajas para huir, poner
sobre aviso a sus compañeras, secretar sustancias o simplemente suplir más rápidamente los
requerimientos energéticos del predador (Sih, 1986). La N. indica de quinto instar en
general fue más eficaz que los otros instares, pues predó un mayor número de larvas de
cada instar. Este fenómeno puede requerir mayor aproximación, pero indudablemente esta
eficacia puede deberse a la necesidad de aumentar de tamaño y continuar hacia su instar
maduro en el que requerirá muchas reservas para su reproducción (Murdoch y Sih, 1978).

16
Si el medio carece de alimento de gran provecho nutricional, N. indica debe orientar sus
esfuerzos hacia la captura de presas que puedan suplir estas necesidades. Para ello debe con
el paso del tiempo ganar experiencia que signifique afinamiento y mayor especialización de
sus habilidades predadoras hacia su presa.
En cuanto al ensayo de preferencias tróficas por estadio de larva se obtuvo que los
diferentes instares de N. indica se alimentan preferiblemente de larvas de mosquito de
mayor desarrollo (L3 y L4). Se presentaron casos en los que el número de las larvas de
mosquito L1 podían mantenerse intactos al final del experimento. Se presentan
comportamientos particulares en cuanto preferencias para cada instar de N. indica hacia
cada instar larval de Ae. aegypti. Sus esfuerzos, que son proporcionales a la tasa de ataque
o predación, parecen correlacionarse al tamaño de la presa y la destreza que se adquiere con
el cambio de instar en el tiempo. Para el caso de N4 y de N5 se presentan semejanzas entre
el número de larvas predadas de los tres últimos instares. Sin embargo, N4 orienta de forma
más dispersa sus esfuerzos a la captura de larvas de instares medios como L2. El predador
tipo N5 orienta de manera muy semejante sus esfuerzos a las dos clases larvales de mayor
talla. La N. indica de mayor desarrollo (N6) evidentemente distribuye sus mayores
esfuerzos de manera casi equitativa hacia las larvas de mayor tamaño (L3 y L4). Las
comparaciones hechas sobre las medias de los números de larvas predadas por las
diferentes clases de N. indica muestran que estas tienden a alimentarse de las larvas de
mayor tamaño, destacándose su predación sobre las presas del tipo L3 y L4 (Tabla 8).
En una prueba relacionada, hecha por Scott y Murdoch (1983), se evaluó la preferencia de
una especie de Notonecta. En este trabajo se encontró que la preferencia de Notonecta hacia

17
clases de Daphnia disminuía con el tamaño de la presa. En general, la mayor de las presas
evaluadas no tenía más de dos mm de longitud. En el caso de N. indica, la mayor de sus
presas podían superar los siete mm de longitud (Fig. 2), y sus preferencias hacia su presa
aumentaron proporcionalmente con el tamaño de la larva de Ae. Aegypti (Fig. 5).
Para los últimos tres instares de N. indica, el aumento de tamaño se relaciona directamente
con su instar de desarrollo y muda, de la misma manera este aumento se relaciona con la
cantidad y calidad de alimento necesario para lograr estos cambios. Se considera más
apropiado para la N. indica invertir sus esfuerzos de captura hacia presas de mayor
provecho energético-nutricional antes que invertir en presas que representen valores bajos
de satisfacción.
Ya que los experimentos se adelantaron en condiciones semejantes a los reservorios
comunes de agua potable utilizados en San Andrés Isla se consideran estos resultados como
prometedores para el control de larvas de mosquito en este tipo de reservorio. A partir de
los resultados obtenidos se considera que Notonecta podría ser empleada como
biocontrolador de larvas de mosquito en la isla de San Andrés y Providencia. Puesto que N.
indica se alimenta de larvas del mosquito Aedes aegypti de avanzado desarrollo,
disminuyendo el número de potenciales adultos que pueden ser transmisores de
enfermedades como el dengue. Inclusive, las larvas de menor tamaño, que en algún
momento crecerán, se encuentran en riesgo de ser predadas, debido a que N. indica
demuestra ser eficaz alimentándose de este tipo de larvas cuando no hay más opción. Esto
parece ser más ventajoso que los métodos tradicionales como el uso de insecticidas para el
control de Ae. aegypti que actualmente se emplean en el Caribe, los cuales podrían

18
contribuir al aumento de la resistencia de los mosquitos hacia estos insecticidas. Estudios
consultados sugieren que el frecuente uso de estos químicos aumenta la resistencia de los
mosquitos de la familia Culicidae (Reyes-Villanueva et al., 1992; Rawlins, 1998; Poopathi
et al. 2001).
Con miras a la aplicabilidad de planes de control biológico de mosquito, es necesario
obtener informaciones relativas al efecto de la predación de N. indica en otras especies de
mosquito vectores de enfermedades, producción de huevos de mosquitos y cantidad de
alimento requerido para el mantenimiento de N. indica en laboratorio y según su instar,
punto o nivel de saciedad de N. indica de acuerdo con el número de larvas consumidas en el
tiempo, crecimiento de poblaciones requeridas en laboratorio para mantenimiento de
cohortes de N. indica, viabilidad de los huevos de N. indica obtenidos en laboratorio,
resistencia del predador a agentes químicos como cloro (por ejemplo: donde se recolectó N.
indica se cloraba el cuerpo de agua, esto según los habitantes del lugar) e insecticidas,
relación entre la cantidad de alimento disponible y cambios de instar del predador. También
se propone adelantar estudios en los que se conozca el desempeño de N. indica como
predador de larvas de mosquito en medios con variaciones en salinidad, turbidez, riqueza
de materia orgánica, entre otras variables ambientales. Todo lo anterior se sugiere con el fin
de optimizar los planes de manejo de N. indica en condiciones insulares con el fin de
permitir su producción y uso como biocontrolador.
AGRADECIMIENTOS
Ofrecemos el mayor de los agradecimientos al Profesor PhD. Bryan McArdle por su valiosa
colaboración científica. Muchas gracias ofrecemos al Biólogo Fabio Costa del INPA por

19
facilitarnos literatura acerca de Culicidae. A su vez agradecemos al profesor PhD. Ernesto
Mancera y por su contribución en la revisión de este trabajo.
Agradecemos de manera muy atenta a la secretaría de salud del departamento archipiélago
de San Andrés Isla, quienes muy amablemente nos ofrecieron espacio físico para adelantar
nuestras investigaciones y la oportunidad de intercambiar experiencias y conocimientos.
Del mismo modo agradecemos al Jardín Botánico de la sede Caribe de la Universidad
Nacional por su valiosa colaboración en la divulgación de nuestro trabajo y por prestarnos
su equipo de microscopia. También extendemos nuestros agradecimientos al biólogo Fáber
González por su colaboración en asesoría científica y recolección de datos. No menos
importantes son nuestros agradecimientos a los jóvenes del colegio Luis Amigó: Daniela,
Sandra y Sebastián por su valioso apoyo y colaboración incondicional. También
agradecemos a todas aquellas personas que consideraron, consideran y consideraran
siempre apoyar toda iniciativa para la resolución de problemáticas que impactan
directamente la salud pública.

20
BIBLIOGRAFÍA
AGUADO JP. Propuesta metodológica para la valoración del manejo de agua para uso
domestico de San Andrés Isla Colombia [tesis de pregrado]. Palmira: Facultad de
Ingeniería y Administración, Universidad Nacional de Colombia; 2004.
BENCE J R. 1998. Indirect Effects and Biological Control of Mosquitoes by Mosquitofish.
The Journal of Applied Ecology. 25 (2): 505-521.
BLAUSTEIN L. 1998. Inflence of the Predatory Backswimmer, Notonecta maculata, on
Invetebrate Community Structure. Ecological Entomology. 23: 246-252.
CHAPMAN H. C. 1974. Biological Control of Mosquito Larvae. Annu. Rev. Entomol. 19:
33-59.
CHESSON J. 1989. The Effect of Alternative Prey on the Functional Response of
Notonecta Hoffmani. Ecology. 70 (5): 1227-1235.
CONSOLI R, DE OLIVEIRA RL. 1998. Principais mosquitos de importancia sanitária no
Brasil. Fiocruz. Rio de Janeiro.
HURST TP, BROWN MD, KAY BH. 2004. Laboratory evaluation of the predation
efficacy of native australian fish on Culex annulirostris (Diptera: culicidae). Journal of the
American Mosquito Control Association. 20 (3): 286-291.
LANE J. 1953. Neotropical Culicidae. University of Sao Paulo. Sao Paulo.
LAWLER S P, DRITZ D, STRANGE T, HOLYOAK M. 1999. Effect of Introduced
Mosquitofish and Bullfrogs on Threatened California Red-Legged Frog. Conservation
Biology. 13 (3): 613-622.

21
LEE DK. 2000. Predation efficacy of the fish muddy loach Misgurnus mizolepis, against
Aedes and Culex mosquitoes in laboratory and small rice plots. Journal of the American
Mosquito Control Association. 16 (3): 258-261.
LIMA JB, DE MELO NV, VALLE D. 2005. Residual effect of two Bacillus thuringiensis
var. israelensis products assayed against Aedes aegypti (Diptera: Culicidae) in laboratory
and outdoors at Rio de Janeiro, Brazil. Rev Inst Med Trop. 47 (3):125-130.
MCARDLE BH, LAWTON JH. 1979. Effects of prey-size and predator-instar on the
predation of Daphnia by Notonecta. Ecological Entomology. 4: 267-275.
MERCER DR, WETTACH GR, SMITH JL. 2005. Effects of larval density and predation
by Toxorhynchites amboinensis on Aedes polynesiensis (Diptera: Culicidae) developing in
coconuts. Journal of the American Mosquito Control Association. 21 (4): 425-431.
MKOJI GM, BOYCE TG, MUNGAI BN, COPELAND RS, HOFKIN BV, LOKER ES.
1999. Predation of aquatic stages of Anopheles gambiae by the Louisiana red swamp
crawfish (Procambarus clarkii). Journal of the American Mosquito Control Association. 15
(1): 69-71.
MURDOCH WW, CHESSON J, CHESSON PL. 1985. Biological Control in Theory and
Practice. Am. Nat. 125 (3): 344-366.
MURDOCH WW, SIH. A. 1978. Age-Dependent Interference in a Predatory Insect.
Journal of Animal Ecology. 47: 581-592.
NELSON MJ. 1986. Aedes aegypti: Biología y ecología. Organización panamericana de la
salud. Washington DC.

22
NERI-BARBOSA J F, QUIROZ-MARTINEZ I H, RODRIGUEZ-TOVAR M L, TEJADA
LO, BADII MH. 1997. Use of Bactimos briquets (B.t.i formulation) combined with the
backswimmer Notonecta irrorata (Hemiptera: Notonectidae) for control of mosquito
larvae. Journal of the American Mosquito Control Association. l3 (1):87-89.
PERICH MJ, CLAIR PM, BOOBAR LR. 1990. Integrated use of planaria (Dugesia
dorotocephala) and Bacillus thuringiensis var. israelensis against Aedes taeniorhynchus: A
laboratory bioassay. Journal of American Mosquito Control Association. 6 (4): 667-671.
POOPATHI S, ARUNACHALAM N, GOPALAN N, BASKARAN G, MANI TR. 2001.
Resistance to Malathion in Culex quinquefasciatus say (Diptera: Culicidae) from Madurai,
south India. Insect Sci. Applic. 21 (3): 251-255.
QUIROZ-MARTINEZ H, RODRIGUEZ-CASTRO VA, SOLIS-ROJAS C,
MALDONADO-BLANCO MG. 2005. Predatory capacity and prey selectivity of nymphs
of the dragonfly Pantala hymenaea. Journal of the American Mosquito Control
Association. 2l (3): 328-330.
RAWLINS SC. 1998. Spatial distribution of insecticide resistance in Caribbean populations
of Aedes aegypti and its significance. Pan. Am. J. Public. Health. 4: 243-251.
REYES-VILLANUEVA F, GARZA-GARZA H, FLOREZ-LEAL JA. 1992. Efecto de
concentraciones subletales de Abate sobre algunos parámetros biológicos de Aedes aegypti.
Salud Publica Mex. 34: 406-412.
SCOTT MA, MURDOCH WW. 1983. Selective predation by the backswimmer,
Notonecta. Limnol. Oceanogr. 28( 2): 352-366.

23
SIH A. 1986. Antipredator Responses and the Perception of Danger by Mosquito Larvae.
Ecology. 67 (2): 434-441.
WILLEMS KJ, WEBB CE, RUSSELL RC. 2005. Tadpoles of four common australian
frogs are not effective predators of the common pest and vector mosquito Culex
annulirostris. Journal of the American Mosquito Control Association. 21 (4): 492-494.

24
Tabla 1. Prueba de densidades. En esta tabla se presentan los datos obtenidos del
tiempo en minutos que requirió N. indica (N5) para alimentarse de la totalidad de
larvas (L2). Cada T representa el tipo de tratamiento aplicado.
T1 T2 T3
65 65 85
147 74 47
40 77 88
95 82 110
150 85 42
Tabla 2. Análisis de varianza para los datos obtenidos a partir del experimento de
densidades.
Fuente G L S C P C F Probabilidad
Tratamientos 2 1916 958 0,87 0,442
Error 12 13164 1097
Total 14 15080
25
Tabla 3. Prueba de Tukey para los tratamientos con N4. Prueba de eficacias tróficas
Tratamiento Media SE St. Dev. Comparaciones
L1 17,5 0,9 1,7 a
L2 16,0 1,0 1,7 ab
L3 12,7 0,8 1,5 b
L4 4,3 1,0 2,1 c
Tabla 4. Prueba de Tukey para los tratamientos con N5. Prueba de eficacias tróficas
Tratamiento Media SE St. Dev. Comparaciones
L1 19,8 0,3 0,5 a
L2 19,5 0,5 1,0 a
L3 19,0 0,7 1,4 a
L4 14,8 2,3 4,5 ab
Tabla 5. Prueba de Tukey para los tratamientos con N4. Prueba de preferencias. Se
presentan con el error estándar de a media (SE) y desviación estándar (St. Dev.).
Tratamiento Media SE St. Dev. Comparaciones
L1 1,3 0,5 1 a
L2 4,0 0,7 0,4 b
L3 3,5 0,3 0,6 ab
L4 2,3 0,6 1,3 ab
26
Tabla 6. Prueba de Tukey para los tratamientos con N5. Prueba de preferencias. Se
presentan con el error estándar de a media (SE) y desviación estandar (St. Dev.)
Tratamiento Media SE St. Dev. Comparaciones
L1 1,2 0,6 1,3 a
L2 3,4 0,7 1,5 ab
L3 4,8 0,9 1,9 b
L4 5,0 0,6 1,4 b
Tabla 7. Prueba de Tukey para los tratamientos con N6. Prueba de preferencias. Se
presentan con el error estándar de a media (SE) y desviación estándar (St. Dev.).
Tratamiento Media SE St. Dev. Comparaciones
L1 2,0 0,9 2,1 a
L2 6,4 0,6 1,3 b
L3 9,0 0,4 1 c
L4 9,4 0,4 0,9 c

27
Tabla 8. Resultados prueba de Tukey para los tratamientos con todas las Notonecta
indica. Prueba de preferencias. Se presentan con el error estándar de a media (SE) y
desviación estándar (St. Dev.).
Tratamiento Media SE St. Dev. Comparaciones
L1 1,5 0,4 1,5 a
L2 4,6 0,5 1,9 b
L3 5,9 0,7 2,7 b
L4 5,8 0,9 3,2 b
Fig. 1 Notonecta indica corresponde a sus instares seis (N6), cinco (N5), cuatro (N4), tres
(N3) y uno (N1) presentado en orden de izquierda a derecha. La escala ubicada a la derecha
corresponde a milímetros.

28
Fig. 2 Larvas del mosquito Aedes aegypti. Las larvas corresponden a los instares primero
(L1), segundo (L2), tercero (L3) y cuarto (L4), presentados en dicho orden de izquierda a
derecha. La escala empleada a la izquierda corresponde a milímetros.
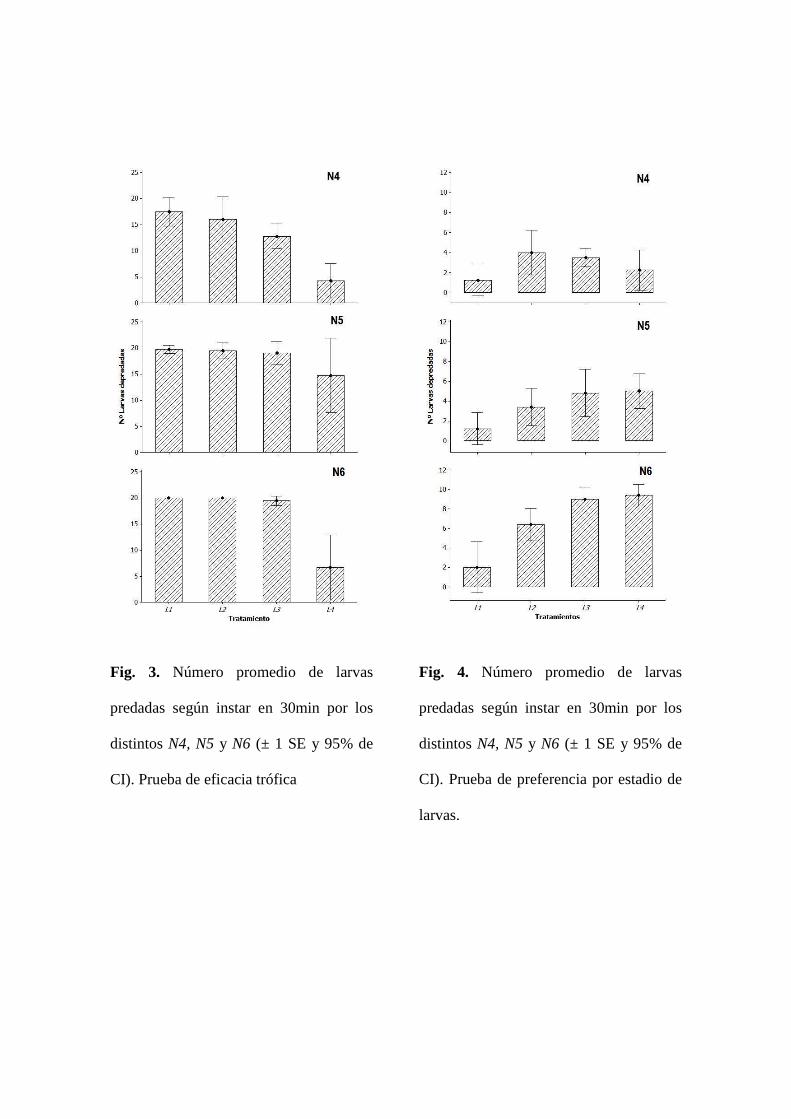
Fig. 3. Número promedio de larvas
predadas según instar en 30min por los
distintos N4, N5 y N6 (± 1 SE y 95% de
CI). Prueba de eficacia trófica
Fig. 4. Número promedio de larvas
predadas según instar en 30min por los
distintos N4, N5 y N6 (± 1 SE y 95% de
CI). Prueba de preferencia por estadio de
larvas.

N vs L4N vs L3N vs L2N vs L1
8
7
6
5
4
3
2
1
0
Tratamientos
Nº Larvas depredadas
Fig. 5 Promedio del número de larvas predadas según su instar en 30min por los distintos
N4, N5 y N6 en conjunto (± 1 SE y 95% de CI). Prueba de preferencias analizadas en
conjunto.