
PONTIFICIA UNIVERSIDAD CATÓLICA DEL ECUADOR FACULTAD DE ... · La zona de reproducción de...
117
PONTIFICIA UNIVERSIDAD CATÓLICA DEL ECUADOR FACULTAD DE CIENCIAS EXACTAS Y NATURALES DEPARTAMENTO DE CIENCIAS BIOLÓGICAS “Desplazamientos de las ballenas jorobadas (Megaptera novaeangliae, Borowski, 1781) (Cetacea: Balaenopteridae) entre zonas de reproducción en el Ecuador” Disertación previa a la obtención del Título de Licenciada en Ciencias Biológicas María José Barragán - Paladines Quito 2003
Transcript of PONTIFICIA UNIVERSIDAD CATÓLICA DEL ECUADOR FACULTAD DE ... · La zona de reproducción de...

PONTIFICIA UNIVERSIDAD CATÓLICA DEL ECUADOR
FACULTAD DE CIENCIAS EXACTAS Y NATURALES
DEPARTAMENTO DE CIENCIAS BIOLÓGICAS
“Desplazamientos de las ballenas jorobadas (Megaptera novaeangliae, Borowski, 1781)
(Cetacea: Balaenopteridae) entre zonas de reproducción en el Ecuador”
Disertación previa a la obtención del Título de Licenciada
en Ciencias Biológicas
María José Barragán - Paladines
Quito 2003

96
CERTIFICACIÓN
Certifico que la disertación de la Licenciatura en Ciencias Biológicas de la
candidata María José Barragán Paladines ha sido concluida de conformidad con las
normas establecidas, por lo tanto puede ser presentada para la calificación correspondiente.
__________________________________
Lic. Santiago F. Burneo
Director de Disertación
Fecha:

97
“Los océanos son los últimos grandes espacios salvajes
del planeta: la última frontera del ser humano
y quizás también la última oportunidad
que nos queda para probar que somos
seres razonables”
John L. Culliney

98
iv
A todos quienes se conmueven
con la grandeza de la vida en el mar...

99
v
AGRADECIMIENTOS
A Yaqu Pacha por financiar este estudio y permitirme cumplir un anhelo, al CDC-
Ecuador y a Jatun Sacha por apoyar esta investigación, a mis padres por introducir en mí el
amor al mar y todo lo que en él habita, a mis hermanos y familia por entender mis eternas
ausencias, al Parque Nacional Machalilla por permitir la realización de este trabajo, a Sol
Damerval (Jefa de Área 2001-2002) por su colaboración, apoyo, ayuda y buen ánimo, a
Jorge Cedeño y la señora Nora, los Guardaparques (Mercedes, Pedro, James, Wilder,
Richard, Paul, Samuel, Ulbio, Jorge M., Jorge A., Marcos) por su colaboración y apoyo, a
Rubén, María, Patricia y Diana por su amistad y cariño, a los dueños y/o administradores de
las Operadoras Turísticas del PNM por su invalorable ayuda (Bosque Marino, Luz de Luna,
Machalilla Tours, Ballenas de la Plata, Sercapez, Piqueros Patas Azules, Ecuador Amazing,
Costatour, Exploratour, Pacífico y Mantarraya). A las tripulaciones de los botes: Luis y
Pedrito (Explora I), José (Popeye II), Flavio y Winston (Lobo Marino), Chale (Luz de
Luna), Anibal y Antonio (Divino Niño), Carlos y Richard (Mantarraya II), Mogo y Goyo
(Ana María II), Polo, Edy y Pedro (Costatour), Winston (Mantarraya III), Jaime (Pacífico
III), Wellington (Tahiry II), Julio y Carlos (Amazing I), Wacho y Luis (Ballena de la
Plata), Marco Antonio (Piqueros Patas Azules) y a los compañeros pescadores que hicieron
observación de ballenas en los botes autorizados. A las encargadas y encargados de las
agencias operadoras de Puerto López y Puerto Cayo por su buena voluntad y apoyo.
A todos los y las Guías Naturalistas del Parque Nacional Machalilla por su
colaboración, participación y ayuda. Al CISP, PMRC, Comité Zonal, por el apoyo.

100
vi
A mis amigos del Cantón Puerto López (nativos y adoptados): Diana, Ricardo,
Rosy, Liviston, Jacqueline, Paola A., Andrés B., Andrés V., Michel, Juan, John, Manfred
por su apoyo y amistad. A Claudia por su trabajo y buena disposición. A mis compadres
Wilmar y Priscilla y a mi ahijada Yorleni Sabrina, por anclarme a Puerto López para
siempre.
A los jóvenes del barrio por su compañía y humor. A Julio, María Eugenia, Bachita,
Señora Hortensia, Tico, Jimmy, Soraya, por la solidaridad y cariño. A Radio Ritmo Azul
por la colaboración y buena disposición. A mi asistente estrella: María Moreno de los Ríos
“Guarrita”, por el excelente trabajo, por su buena voluntad y por su amistad. A Christoph
por compartir mis sueños. A Paty Brtnik, colega y amiga, por sus comentarios, sugerencias
y por compartir su información y análisis de datos. A Judith, César Alberto, Drea, Lotta,
Andrea, Alex y Marcela por la ayuda, críticas y colaboración en la recopilación de datos. A
los compañeros pescadores artesanales por su participación en las actividades de
investigación y educación. A los niños de la playa por alegrar las mañanas y las frías tardes
con su sonrisa. Y, a mis queridas jorobadas por venir de tan lejos, saltar, respirar junto a
mi y regalarme los instantes más conmovedores de mi vida.
Muchas Gracias a todos amigos, amigas, conocidos y conocidas, y mil disculpas si
pasé por alto a alguien.....

101
vii
TABLA DE CONTENIDOS
DEDICATORIA ......................................................................................................... iv
AGRADECIMIENTOS .............................................................................................. v
TABLA DE CONTENIDOS ...................................................................................... vii
LISTA DE FIGURAS ................................................................................................. x
LISTA DE TABLAS .................................................................................................. xiv
LISTA DE ANEXOS ................................................................................................. xvii
1. RESUMEN ............................................................................................................. 1
2. ABSTRACT ........................................................................................................... 3
3. INTRODUCCIÓN .................................................................................................. 5
3.1 Aspectos históricos en las migraciones de las ballenas .................................. 5
3.2 La presencia de las ballenas jorobadas en el Ecuador .................................... 7
3.3 Las áreas de reproducción de Megaptera novaeangliae en Ecuador ............. 9
3.4 Estado de conservación de los cetáceos .......................................................... 10
3.5 Uso del recurso ballena jorobada en el área de estudio .................................. 12
3.6 Aspectos biológicos y reproductivos de Megaptera novaeangliae ................ 12
3.7 Objetivo General ............................................................................................. 18
3.8 Objetivos Específicos ..................................................................................... 18
4. MATERIALES Y MÉTODOS................................................................................ 19
4.1 Área de estudio ............................................................................................... 19
4.2 Registro de datos en el campo ........................................................................ 22

102
4.2.1 Fotografías ............................................................................................. 22
4.2.2 Equipo .................................................................................................... 23
4.2.3 Transectos .............................................................................................. 24
4.2.4 Datos ...................................................................................................... 25
4.2.4.1 Temporales ................................................................................ 25
4.2.4.2 Geográficos ................................................................................ 25
4.2.4.3 Climáticos .................................................................................. 25
4.2.3.3 Biológicos y etológicos .............................................................. 26
4.2.4.5 Fotográficos ............................................................................... 26
4.2.5 Ingreso de datos ..................................................................................... 26
4.2.5.1 Formulario de Fotoidentificación .............................................. 26
4.2.5.2 Formulario de Esfuerzo ............................................................. 26
4.2.5.3 Formulario de Registros ............................................................ 27
4.2.5.4 Formulario de Fotografías ......................................................... 27
4.2.6 Fotoidentificación .................................................................................. 27
4.2.7 Análisis .................................................................................................. 29
4.2.7.1 Registros fotográficos ................................................................ 29
4.2.7.2 Calidad de la imagen ................................................................. 30
4.2.7.3 Patrón de las aletas caudales según la coloración ...................... 30
4.2.7.4 Patrón de las aletas caudales según marcas ............................... 31
4.2.7.5 Patrón de las aletas caudales según la forma de la hendidura
media ..........................................................................................
31
4.2.7.6 Análisis espacial y temporal de la distribución de ballenas en
las zonas de reproducción ..........................................................

103
las zonas de reproducción .......................................................... 32
4.2.7.7 Desplazamiento entre las zonas de reproducción en el Ecuador 32
4.2.7.8 Estadístico .................................................................................. 33
4.2.8 Términos y relaciones ............................................................................ 33
4.2.8.1 Población ................................................................................... 33
4.2.8.2 Stock .......................................................................................... 34
4.2.8.3 Agregación ................................................................................. 34
4.2.8.4 Grupo ......................................................................................... 35
4.2.9 Intercambio de individuos .................................................................... 36
4.2.10 Interacciones con otros cetáceos .......................................................... 37
5. RESULTADOS Y DISCUSIÓN............................................................................. 38
5.1 Aspectos climáticos y factores físicos en las dos zonas de estudio ................ 38
5.2 Datos obtenidos .............................................................................................. 39
5.2.1 Frecuencia de individuos en las dos zonas de estudio ........................... 40
5.2.2 Comparación de los registros de M. novaeangliae del área A con los
del área B ...............................................................................................
42
5.2.2.1 Patrón de clasificación por la presencia de marcas, cicatrices y
huellas ........................................................................................
42
5.2.2.2 Patrón de clasificación por la muesca media entre los lóbulos
de las aletas caudales .................................................................
42
5.2.2.3 Categorías en la calidad de las fotografías analizadas ............... 43
5.2.3 Identificación de individuos en las dos zonas de estudio ...................... 43
5.2.4 Patrones de distribución espacial y temporal de M. novaeangliae ........ 44

104
5.2.4.1 Composición grupal en las zonas de reproducción del Ecuador 45
5.2.5 Desplazamiento de las ballenas jorobadas entre las zonas de
reproducción del Ecuador ......................................................................
47
5.2.6 Interacciones con otros cetáceos en las áreas de reproducción ............. 51
5.2.6.1 Con delfines manchados (Stenella attenuata) ........................... 51
5.2.6.2 Con orcas (Orcinus orca) .......................................................... 52
6. RECOMENDACIONES.......................................................................................... 56
6.1 Metodologías para caracterización de ballenas jorobadas .............................. 56
6.1.1 Cantos .................................................................................................... 56
6.1.2 Análisis genético de individuos ............................................................. 57
6.2 Turismo de observación de ballenas en las zonas de reproducción ................ 57
7. BIBLIOGRAFÍA CITADA .................................................................................... 60
8. FIGURAS ............................................................................................................... 77
9. TABLAS ............................................................................................................... 88
10. ANEXOS .............................................................................................................. 95

105
x
LISTA DE FIGURAS
1. Mapa de la zona de estudio A: costas frente a Esmeraldas; zona de estudio B; área
marina del Parque Nacional Machalilla .........................................................................
78
2a. Gráfico de temperatura superficial del mar (Julio) de las dos zonas de estudio ...... 79
2b. Gráfico de temperatura superficial del mar (Agosto) de las dos zonas de estudio .. 79
2c. Gráfico de temperatura superficial del mar (Septiembre) de las dos zonas de
estudio ......................................................................................................................
80
3. Análisis comparativo de las medias de las temperaturas superficiales del mar
semanales) .................................................................................................................
81
4. Bote de observación de ballenas ................................................................................ 81
5. Esquema de análisis de la cobertura del cielo ........................................................... 82
6. Esquema de los patrones de coloración en aletas caudales para identificación ........ 82
7. Esquema de los patrones de clasificación de aletas caudales por la hendidura
media ..........................................................................................................................
83
8a. Esquema de relaciones entre ballenas ...................................................................... 83
8b. Términos que califican a las ballenas jorobadas en su distribución mundial .......... 84
9. Esfuerzo quincenal de muestreo aplicado en las zonas de estudio A y B ................. 84
10. Frecuencia de ballenas jorobadas en las zonas de estudio A y B ............................ 85
11. Gráfico de frecuencia y esfuerzo aplicado en estudios preliminares, año 2000 ...... 85
12. Frecuencia comparativa de ballenas en la zona de estudio A en las temporadas
2000 y 2001..............................................................................................................
86
13. Frecuencia comparativa de ballenas en la zona de estudio B en las temporadas
2000 y 2001 .........................................................................................................

106
2000 y 2001 ......................................................................................................... 86
14. Esquema para contar individuos en grupos de delfines ........................................... 87
15. Esquema de observación del ataque de O. orca sobre una ballena jorobada .......... 87

107
xi
LISTA DE TABLAS
1. Comparación de las temperaturas superficiales del mar, promedio, máximas y
mínimas para las zonas de estudio A y B durante la temporada de estudio .............
89
2. Códigos de identificación de aletas caudales basados en la coloración .................... 89
3. Códigos de marcas y señales de identificación para aletas caudales (Mizroch, et al.
1990) ..........................................................................................................................
90
4. Código de clasificación de aletas caudales según la hendidura media ...................... 90
5. Cambios en la frecuencia de ballenas jorobadas en las dos zonas de estudio A y B
en las temporadas 2000 y 2001 ..................................................................................
91
6. Esfuerzo de muestreo y frecuencia de ballenas jorobadas en la temporada 2001 ..... 91
7. Reavistamientos de ballenas en Colombia, Panamá, Perú, Ecuador. Fechas y
localidades de avistamientos de ballenas jorobadas identificadas en Panamá,
Colombia, Ecuador y Perú ....................................................... ................................
92
8. Resumen de valores de trabajo de campo en la temporada 2001 en las dos zonas de
estudio ........................................................................................................................
93
9. Aletas caudales registradas y agrupadas según la coloración .................................... 93
10. Aletas caudales registradas y agrupadas según la forma de la hendidura media ..... 94
11. Individuos identificados en las dos zonas de estudio .............................................. 94

108
xii
LISTA DE ANEXOS
1. Categorías de comportamiento utilizadas en el estudio ............................................. 96
2. Formulario de campo utilizado en el registro de campo (PhotoID) .......................... 97
3. Formulario de campo utilizado en el registro de campo (Esfuerzo) .......................... 98
4. Formulario de campo utilizado en el registro de campo (Registros) ......................... 99
5. Formulario de campo utilizado en el registro de campo (Fotos y Rollos) ................. 100
6. Fotos de recaptura (reavistamientos de aletas caudales) en temporada 2000 y 2001 101
7. Interacciones de ballenas jorobadas y otros cetáceos (O.orca) ................................. 102

109
1. RESUMEN
Las zonas de permanencia de las ballenas jorobadas fueron identificadas en décadas
anteriores en diferentes regiones del mundo y fueron clasificadas como zonas de
alimentación y zonas de reproducción, las últimas generalmente relacionadas con regiones
tropicales del planeta.
La zona de reproducción de Megaptera novaeangliae en Manabí ha sido estudiada
desde el año 1996, cuando se la identificó como un centro de reproducción para esta
especie. En el año 2000, las costas de Esmeraldas fueron identificadas como otro centro de
reproducción de ballenas jorobadas. El presente estudio de monitoreo de las ballenas
jorobadas en las dos zonas en el año 2001 ha permitido iniciar la caracterización de las
ballenas y de las dos zonas durante su permanencia en la temporada de reproducción, que
va de mayo a septiembre.
Los registros de ballenas en ambas zonas (número total de individuos; n = 1 039
ballenas) permitieron compararlos entre ellas y establecer que aparentemente no existen
patrones de desplazamiento entre ambos centros de reproducción, aún cuando otros autores
anteriormente han comprobado ya que sí existen desplazamientos de individuos entre las
zonas de reproducción de Ecuador y Colombia.
Con el análisis de los datos de campo se establece que, tanto la zona de Esmeraldas
como la de Manabí constituyen áreas de reproducción con permanencia temporal de
ballenas durante su etapa reproductiva, que incluye cortejo, apareamiento y nacimiento de
crías.
Se ha podido comprobar que la abundancia de individuos en las dos zonas no es
igual, que hubo un incremento en la abundancia de individuos en la zona de Esmeraldas

110
mientras que hubo una disminución de la abundancia de individuos en la zona de Manabí,
durante el mismo período.
Los resultados obtenidos permiten también establecer, que los grupos de ballenas
localizadas en la zona de Esmeraldas constituyen una agregación reproductiva diferente a la
agregación reproductiva que está presente en Manabí.
Este antecedente tiene implicaciones directas en el manejo y conservación del
recurso ballena jorobada a mediano y largo plazo. Tomando en cuenta que la actividad de
turismo de observación de ballenas se incrementa año a año y en determinado momento
puede convertirse en una seria amenaza para las ballenas, es necesario tomar medidas para
utilizar este recurso con responsabilidad y ecuanimidad.
El manejo de la especie debe estar directamente relacionado con el concepto de
agregaciones reproductivas en cada una de las zonas, pues cada agregación recibe diferente
nivel de presión debido a la actividad turística. Esta presión se diferencia y no es
equivalente; la cantidad de botes, turistas y demanda es menor en Esmeraldas que en
Manabí. Debido a que no existe intercambio de individuos entre las zonas, cada agregación
de ballenas está sometida al mismo estrés por la actividad turística durante su permanencia
allí, y debido a que no hay movimiento de individuos entre las dos zonas, no será posible
atenuar el impacto del turismo sobre los individuos.
Es necesario profundizar estudios sobre las dos agregaciones de Ecuador para
conocer a detalle sus posibles relaciones.
Palabras clave: agregaciones, ballenas jorobadas, desplazamientos, Ecuador,
fotoidentificación, Megaptera novaeangliae, zonas de reproducción.

111
2. ABSTRACT
The areas where the humpback whales are staying were identified decades ago
around the world. These were classified as feeding and breeding grounds, the latter are
often related with tropical areas of the Earth.
The breeding ground of Megaptera novaeangliae in Manabí province has been
studied since 1996, when it was identified as a breeding center for humpback whales. In
2000, Esmeraldas coastal line was also identified as another humpback whale breeding
center. The monitoring study in 2001 has let us start the characterization of whales from
both areas during their staying in the breeding season, between May and September.
The whale records in both areas (the number of recorded individuals; n = 1039
whales) let us compare them and establish the criteria about the lacking of movement
patterns between the two breeding grounds in Ecuador. In despite of this, some authors
have already proved that there actually are individual movements between breeding
grounds from Ecuador and Colombia.
Field data show us that Esmeraldas and Manabí are actually breeding grounds with
a seasonal permanence of humpback whales. This permanence for reproduction include
courtships, mating and calves birth and nursing.
It has been proved that the whales abundance is not the same for both areas. There is
an increasing number in Esmeraldas while there is a decreasing number of individuals in
Manabí, at the same periods.
This fact has important management implications for the conservation of this
species in a mid and long term period. The whale watching tourist activities is increasing

112
from one year to the next and it could be a threat for the whales if the correct policies are
not taken to use the resource in a responsible and respectful way.
The management of this species has to be related with the aggregation concept in
both areas because each one of them is having different tourism stress. This stress is
unequal, the number of boats, tourists and demand of these activities are less strong in
Esmeraldas than in Manabí. Because there is not whales interchanging between both areas,
each whale aggregation is under the same tourism stress during their staying, and the lack
of individuals movements will not make possible decrease the tourism impact over the
whales.
It is also necessary to continue with the study of the two humpback whales
aggregations to determine the possible relationships between them.
Key words: aggregations, breeding grounds, Ecuador, humpback whale, Megaptera
novaeangliae, movements, photo-identification.

113
3. INTRODUCCIÓN
3.1 ASPECTOS HISTÓRICOS EN LAS MIGRACIONES DE LAS BALLENAS
JOROBADAS
El extinto orden terrestre Condylarthra originó a dos grupos de mamíferos:
Ungulata, que incluyen a los actuales Artiodactyla y Archaeoceti, que evolucionaron en el
Paleoceno tardío y originaron a los actuales cetáceos (Schaeff, 1997). Evolutivamente el
origen de los cetáceos es común al de los artiodactilos; ambos grupos comparten como
tronco original a los penungulados (Harvey-Pough, 1999).
El orden Cetacea comprende dos subordenes vivientes y un suborden extinto. Los
subordenes existentes son Mysticeti (ballenas con barbas) y Odontoceti (ballenas con
dientes) y el suborden extinto es Archaeoceti (ballenas antiguas). Posteriormente en el
Eoceno temprano y medio (hace 50 millones de años) los arqueocetos colonizaron el mar y
originaron a los subordenes de cetáceos conocidos actualmente (Schaeff, 1997). Los
cetáceos primitivos se distribuyeron principalmente en África y en las costas orientales de
Norteamérica (Harvey-Pough, 1999).
El fósil más antiguo de los cetáceos (Pakicetidae: Pakicetus sp.), fue encontrado
recientemente en Pakistán y muestra que vivió en el Eoceno temprano (Bajpai et al., 1998).
Estuvo provisto de dientes y fue un formidable depredador en las aguas cálidas y poco
profundas de donde evolucionó a las formas más recientes (Marine Institute, 2003).
Registros de los primeros arqueocetos (Pakicetidae: Himalayacethus subathuensis)
muestran evidencias de que estos animales permanecieron en ambientes de agua dulce,

114
estuarios y aguas marinas poco profundas, movilizándose entre ellos en busca de alimento,
preferentemente peces (Bajpai et al., 1998); esto demuestra que los cetáceos primitivos
tenían tendencias más bien sedentarias y de movilización en cortos rangos de distancias.
El Eoceno medio se caracterizó por un calentamiento climático en las latitudes altas,
situación que fue cambiando en el Eoceno tardío, cuando las temperaturas medias anuales
empezaron a decaer y se inició el período del frío en el Cenozoico (Harvey-Pough, 1999).
Durante este tiempo en que el agua del mar tenía una temperatura promedio menor, la
distribución espacial y temporal de los cetáceos era uniforme y con pocas migraciones
(SALVAT, 1975). Con el calentamiento posterior de las aguas del mar en el Mioceno
temprano, la abundancia de alimento para los cetáceos disminuyó.
Bajpai et al. (1998) se refieren a las distribuciones de los cetáceos primitivos en
áreas cercanas entre sí, lo que demuestra que las migraciones de largas distancias que
presentan los cetáceos actualmente son mecanismos de supervivencia que no han formado
parte de la historia natural ancestral de este orden. Entre las hipótesis que explican las
migraciones se incluyen criterios sobre la búsqueda y disponibilidad de alimento para
cubrir sus requerimientos energéticos en las zonas frías y la selección de parejas (Valsecchi
et al. 2002).
Asimismo, esta original y uniforme distribución temporal y espacial de los cetáceos
hacía más factible que las crías sean más susceptibles a ataques de depredadores en zonas
en las que las condiciones geográficas facilitaban su ataque (aguas muy profundas por
ejemplo). La actual distribución de algunos misticetos les ha permitido disminuir las
posibilidades de ser presa de sus depredadores naturales, debido a que en aguas tropicales,
donde normalmente se desarrollan las actividades de nacimiento y crianza de los ballenatos,

115
la posibilidad de encuentros con estos grandes depredadores es menor que en zonas de
aguas frías.
En este caso hubo una preferencia por zonas de reproducción con profundidades
bajas, cercanas a las costas, aguas cálidas y poca probabilidad de presencia de
depredadores, en vez de zonas con abundancia de alimento para la reproducción y crianza.
3.2 PRESENCIA DE BALLENAS JOROBADAS EN EL ECUADOR
Ancestralmente el ser humano ha relacionado a los animales marinos y en especial a
las ballenas con seres mágicos, por su apariencia y forma de vida poco conocida; esto fue
motivo para la creación de fábulas y cuentos en los cuales se les atribuía características de
seres espectaculares. Las referencias de su presencia en el Ecuador datan de períodos
prehistóricos (Castro et al., 1999) y su registro en aguas oceánicas ecuatorianas, insulares y
continentales en tiempos remotos se prueba por algunas evidencias de estos cetáceos en
restos arqueológicos de culturas prehispánicas.
Cada año desde finales de mayo, las ballenas jorobadas (Megaptera novaeangliae)
inician su llegada a aguas ecuatorianas provenientes de la Antártida y permanecen en la
zona del Área Marina del Parque Nacional Machalilla (AMPNM) y frente a las costas de
Esmeraldas hasta mediados o finales de octubre, con el objetivo de cumplir una parte de su
ciclo biológico.
En el año 1996 el área correspondiente al AMPNM fue descrita como una zona de
reproducción para esta especie (Denkinger et al.,1997; Scheidat et al., 2000) debido al
registro y comprobación de comportamientos que evidenciaron actividades de
reproducción, entre ellos: cortejos, cantos, crías recién nacidas, demostraciones de

116
competencia, etc. Esta zona está comprendida entre el límite costero de la zona continental
del Parque Nacional Machalilla (PNM) y la Isla de la Plata (zona insular, Figura 1).
En la zona comprendida entre Atacames y Cabo San Francisco, Esmeraldas, se han
realizado observaciones de ballenas jorobadas en actividades de reproducción y registros de
cantos, crías recién nacidas y comportamiento reproductor durante los años 1999, 2000 y
2001 (Denkinger; Brtnik, comunicación personal) probándose que es otra zona de
reproducción de las ballenas jorobadas en Ecuador.
Castro et al. (2002) mencionan que en el AMPNM, la observación de crías recién
nacidas, comportamientos reproductivos, reobservación de animales identificados y registro
de composición grupal son evidencias que la zona de estudio B es la única de apareamiento
y reproducción de ballenas jorobadas que ha sido determinada hasta el momento para el
Ecuador. Sin embargo, Denkinger y Brtnik (en prep.) indican que la zona de las costas
frente a la provincia de Esmeraldas son también parte de una zona de reproducción de
ballenas jorobadas debido al registro de los mismos comportamientos reproductivos
observados ya en el AMPNM. Por lo tanto, en Ecuador existen dos zonas de permanencia
temporal de ballenas jorobadas en las cuales se cumple la estación de reproducción de M.
novaeangliae: las costas de Esmeraldas y las del AMPNM.
A partir de estas observaciones, los procesos reproductivos de estos cetáceos fueron
investigados científicamente e incluso los pobladores locales comenzaron a interesarse por
las ballenas jorobadas para desarrollar en esta área la industria de la observación comercial
de ballenas (whale watching), la misma que actualmente se ha convertido en una de las
actividades económicas más importantes de la región.
Conocer que las costas del AMPNM fueron identificadas y descritas como la
primera área de reproducción de ballenas jorobadas en Ecuador y posteriormente la

117
comprobación de que las costas de Esmeraldas también son una zona de reproducción para
M. novaeangliae, nos coloca en la posición de plantear la posibilidad de que las dos zonas
de reproducción de ballenas jorobadas tengan la presencia de individuos que se desplazan
entre las dos zonas indistintamente.
La pregunta que surge de este hecho es: ¿Las ballenas que están en ambas zonas son
comunes entre éstas o son diferentes para cada sitio?, pues es muy poco lo que se sabe de
movimientos entre sitios específicos de reproducción durante el invierno austral (Flores-
Gonzáles, et al., 1998). Este cuestionamiento es el objetivo de la presente investigación.
3.3 LAS ÁREAS DE REPRODUCCIÓN DE Megaptera novaeangliae EN ECUADOR
Mackintosh (1942; 1965) y Carwardine (1995) reconocen la existencia de seis
stocks de ballenas jorobadas distribuidos alrededor del continente Antártico durante el
verano austral; por otro lado la IWC (2001) identifica siete stocks de ballenas jorobadas en
el hemisferio sur, debido a que subdividen al stock oriental de Australia en dos
componentes: oriental y occidental; conocemos que las ballenas presentes en las costas del
Ecuador y Colombia durante sus períodos reproductivos pertenecen al stock de ballenas
jorobadas del Pacífico Sudeste que se alimenta en la Antártida (Dawbin, 1966;
Leatherwood y Reeves, 1983; IWC, 1999); asumimos por lo tanto que las ballenas
jorobadas objeto del presente estudio pertenecen a diferentes agregaciones de los stocks que
se distribuyen alrededor del continente Antártico durante el verano austral y que forman
parte de la población del hemisferio sur (Figura 2a y 2b).
Sin embargo Castro et al. (2002) mencionan que la base de individuos identificados
hasta el momento en la zona del AMPNM servirá sólo para compararlos con catálogos de

118
otros países siendo que los individuos presentes en Ecuador necesariamente deben ser
comparados en primer lugar entre las dos zonas de reproducción del Ecuador (AMPNM y
costas de Esmeraldas) y en segundo lugar, con las ballenas jorobadas que se localizan
alrededor de la Isla Gorgona en Colombia, correspondientes al stock de la Península
Antártica e Isla Gorgona, Colombia y América del Sur (Baker et al. 1998).
3.4 ESTADO DE CONSERVACIÓN DE LOS CETÁCEOS
Durante siglos, las poblaciones de ballenas jorobadas se vieron amenazadas y
disminuidas hasta niveles cercanos a la extinción debido a la cacería indiscriminada
realizada por los barcos balleneros alrededor del mundo. En el año 1966, la Comisión
Ballenera Internacional (IWC), dispuso mediante la emisión del Acuerdo Mundial de
Protección y Conservación de las Ballenas y otros cetáceos, la prohibición de la cacería y
muerte de todas las especies de cetáceos en el mundo. Todos los países fueron signatarios
de este acuerdo, con excepción de Japón y Noruega, quienes hasta la actualidad continúan
cazando a las ballenas de los océanos del mundo.
La legislación ecuatoriana protege también a las ballenas con el Acuerdo Ministerial
No.196, mediante el Registro Oficial No. 458 del 14 de junio de 1990, el cual dispone que
todas las especies de ballenas presentes dentro de las 200 millas náuticas (aguas territoriales
ecuatorianas), sean protegidas y que Galápagos sea declarado como “Santuario de
Ballenas”. El 28 de enero de 2000, mediante el Registro Oficial No.5 se declara la
prohibición para cualquier actividad de cacería de ballenas en el Ecuador.
Actualmente, organizaciones mundiales de protección de la fauna silvestre han
determinado el estado de conservación de los cetáceos. Según el Libro Rojo de la Unión

119
Internacional para la Conservación de la Naturaleza (IUCN, por sus siglas en inglés) las
ballenas jorobadas (Megaptera novaeangliae) se ubican dentro de la categoría VU
(Vulnerable, IUCN, 2002) criterio que es compartido por Baillie y Groombridge (1996) y
por Tirira (2001). Esta categoría agrupa a un taxón que no está ni En Peligro ni en Peligro
Crítico, pero que en estado silvestre enfrenta un alto riesgo de extinción a mediano plazo.
Carwardine (1995) coloca a esta especie en una categoría con estado de conservación
RARO. Según la Convención sobre el Comercio Internacional de Especies Amenazadas
(CITES, por sus siglas en inglés) las ballenas jorobadas se ubican dentro del Apéndice I
(CITES, 1996), que incluye todas las especies en peligro de extinción cuyo comercio
internacional, tanto para especimenes muertos, vivos o alguna de sus partes, tiene
prohibición absoluta.
Si bien en la actualidad la cacería de ballenas aparentemente no constituye un riesgo
importante que afecte a las poblaciones de Megaptera novaeangliae en el mundo, sí lo son
las capturas incidentales en artes pesqueras artesanales e industriales que se producen
frecuentemente (Alava et al., 2001), mismas que por la magnitud de la actividad pesquera
mundial, se transforman en una de las mayores amenazas para esta y otras especies de
animales marinos.
Asimismo la contaminación del mar por evacuación de desechos urbanos e
industriales y la creciente actividad turística en las zonas de distribución de las ballenas
jorobadas son otras amenazas de importancia al momento de considerar el estado de
conservación y protección de Megaptera novaeangliae en todo el planeta.
3.5 USO DEL RECURSO BALLENA JOROBADA EN EL ÁREA DE ESTUDIO

120
La actividad turística de observación de ballenas (whale watching) en el AMPNM
se inició hace aproximadamente diez años; en las costas de Esmeraldas esta actividad tiene
un origen más reciente, aproximadamente dos años. En un principio, las personas, empresas
e infraestructura dedicadas y utilizadas para el turismo en el PNM y el AMPNM, eran
pocas. Con el pasar de los años estas actividades productivas se han incrementado hasta que
en la actualidad ocupan el segundo lugar como el rubro económico más importante de la
zona.
Hoy en día muchos de los pobladores de Esmeraldas y del AMPNM se dedican de
manera total o parcial a actividades de observación de ballenas, mismas que se
complementan con todo el aparato turístico que se ha desarrollado en la zona. Así,
restaurantes, hoteles, hosterías, tiendas de recuerdos y artesanías se han visto beneficiadas
por la gran cantidad de turistas, nacionales y extranjeros que anualmente acuden a la zona
para realizar la observación de ballenas.
Actualmente la gran vistosidad y belleza de las ballenas jorobadas las convierten sin
duda en un recurso natural de gran importancia para las poblaciones que las utilizan como
un recurso turístico y medio de subsistencia.
3.6 ASPECTOS BIOLÓGICOS Y REPRODUCTIVOS DE Megaptera novaeangliae
Las ballenas jorobadas son mamíferos marinos de gran tamaño que pertenecen al
orden de los cetáceos, nombre que proviene del latín cetus y del griego ketus y que significa
“monstruo grande del mar”, atribuido por los pueblos antiguos a estos animales que han
poblado los mares desde tiempos remotos.

121
La ballena jorobada está dentro del suborden de las “ballenas con barbas” o
Misticetos, grupo caracterizado por poseer placas córneas en su boca, estructuras que les
permite alimentarse por un mecanismo de filtración.
El nombre científico de este mamífero proviene de las palabras en latín mega,
grande; ptera, alas; nova, nueva; angliae, Inglaterra; “alas grandes de Nueva Inglaterra” es
el nombre que confirma una de sus características más llamativas, sus inmensas aletas
pectorales, estructuras que pueden llegar a medir hasta un tercio del largo total de su cuerpo
(Figura 3).
Son de color gris oscuro brillante, sus extremidades adaptadas para la natación han
formado aletas (dos pectorales y una caudal) y en la parte superior delantera de su cabeza
presentan protuberancias que sobresalen a manera de callosidades. Poseen pulmones y
respiran mediante la expulsión de aire, a presión, a través de dos orificios (espiráculos)
localizados en la parte superior de su cabeza. Como homeotermos, estos pulmones y otros
órganos han sufrido un extraordinario proceso de adaptación evolutiva a lo largo de miles
de años lo que actualmente les permite habitar los ambientes marinos.
Las ballenas jorobadas, nombre que se han ganado por su prominente aleta dorsal
que sobresale frecuentemente al momento de la inmersión, también son conocidas como
yubartas o humpback whales.
Su tamaño promedio en estado adulto es entre 15 a 16 m, siendo las hembras
frecuentemente más grandes que los machos; su peso puede alcanzar entre 15 y 30
toneladas. Las crías miden al nacer entre 3 y 4 m aproximadamente y alcanzan un peso
entre 2 a 3 toneladas. Puede llegar a los 40 o 50 años de edad.
Durante la etapa reproductiva, las ballenas jorobadas machos y hembras desarrollan
comportamientos reproductivos típicos especialmente utilizados en el cortejo (saltos,

122
coletazos, golpes de cola, golpes consecutivos de la aleta pectoral). Como sucede en otros
mamíferos que comparten un mismo origen filogenético con las ballenas (como los
delfines, cachalotes y cérvidos, Harvey-Pough, et al., 1999), los alces macho (Alce alce)
que son silenciosos durante casi todo el año, durante la época de celo, en septiembre, dejan
de comer y emiten vocalizaciones complicada consistentes en bramidos para atraer a las
hembras; macho y hembra se llaman recíprocamente con gemidos para atraerse y aparearse
(Runtz, 1991). Las ballenas por su lado, utilizan cantos que son verdaderos sistemas de
comunicación, calificados como los más complejos del reino animal (Kaufman y Forestell
1999).
Según Tyack (1981) los cantos de las ballenas jorobadas macho tienen doble
función en el comportamiento reproductivo de las ballenas jorobadas: atraer a las hembras
y alejar a otros machos que pueden representar competencia. Según Baker y Herman
(1984), los cantos de los machos sirven también para sincronizar la ovulación en las
hembras. Los cantos de las ballenas jorobadas cumplen un rol importante en la interacción
entre individuos, en medio de la compleja estructura social de las ballenas jorobadas.
Matilla et al. (1987) reportan que individuos de M. novaeangliae del hemisferio
norte inician cantos completos en sus zonas de alimentación en otoño, y continúan con los
cantos en su migración a aguas tropicales (Kibblewhite et al., 1967; Levenson y Leapley,
1978) y según Payne y McVay (1971) y Stone et al.(1987) los cantos de las ballenas
jorobadas del hemisferio norte duran por lo menos hasta la primera mitad del viaje de
retorno a aguas del Ártico.
Los cantos en M. novaeangliae son un elemento importante que pueden ayudar a
determinar las rutas migratorias de estos cetáceos (Clapman y Mattila, 1990), sin embargo,
no se han realizado estudios de cantos de ballenas jorobadas del hemisferio sur, y en

123
especial del stock de la Antártida que llega a aguas ecuatorianas y colombianas para
aparearse y reproducirse.
La estructura social en M. novaeangliae no está definida específicamente y no es
fácil caracterizarla, pues no existe ningún vínculo estrecho entre los miembros de los
grupos. La composición grupal está relacionada en gran medida con la capacidad auditiva
de la especie. La única relación estable es la formada por el “núcleo familiar”, constituido
por la hembra y la cría, misma que dura solamente la temporada de la lactancia, entre seis y
diez meses (Chittleborough, 1958). Este núcleo familiar puede tener como acompañante a
un macho que se denomina “escolta”, que no permanece indefinidamente con la madre y la
cría y su acompañamiento es inestable, pudiendo ser desplazado por otros machos (Brtnik,
2000). Las relaciones entre machos y hembras se caracterizan por el establecimiento
temporal de parejas para cumplir el apareamiento, siendo normal las relaciones polígamas
entre individuos.
Las estructuras y comportamientos sociales en las ballenas jorobadas han
evolucionado específicamente en relación con las áreas en donde se ubican en determinada
temporada. Los grupos en las áreas de alimentación son grandes y mantienen asociaciones a
largo plazo aunque pueden durar semanas y volver a establecerse en el año siguiente; su
comportamiento es básicamente no competitivo y parcialmente cooperativo, tienen estratos
sociales diferentes, dependiendo de la preferencia por el alimento (Brtnik, 2000); por
ejemplo aquellas que se alimentan de krill forman grupos pequeños e inestables y las que se
alimentan de peces pequeños forman grupos grandes y estables con asociaciones a largo
plazo (Baker y Herman, 1984).
En el área de reproducción se forman grupos pequeños, inestables y dinámicos
debido a la formación continua de nuevos grupos por parte de machos que se apartan de las

124
hembras (Tyack y Whitehead, 1983; Baker y Herman, 1984; Mobley y Herman, 1985); la
estructura social es diferente y las ballenas muestran un comportamiento competitivo de
apareamiento, con peleas entre los machos (Brtnik, 2000).
El apareamiento de la ballena jorobada sincroniza el ciclo sexual de las hembras,
que dura aproximadamente un año, con su permanencia en aguas cálidas y tropicales. Los
eventos de cópula en el mar son hechos no fácilmente observados con facilidad pues se
realiza debajo de la superficie del agua. Después del apareamiento las ballenas migran a su
área de la alimentación y en el siguiente año regresan al área de la reproducción para el
nacimiento de las crías, luego de un período de gestación de once meses y medio. Posterior
a un período de lactancia de aproximadamente seis a diez meses, tiempo en el que la
hembra permanece con la cría, las hembras vuelven a ser receptivas y fértiles, a pesar de
esto, generalmente la mayor parte de hembras esperan un año más antes de aparearse otra
vez. La época de fertilidad en las hembras se reduce a una vez por año y determina los
ciclos sexuales de los machos; por ello en las zonas de reproducción los machos presentan
una abundante producción de esperma y consecuentemente un crecimiento de los testículos.
Esto les permite tener mayor posibilidad de fecundar a varias hembras (Chittleborough,
1958).
Los comportamientos migratorios varían de una especie a otra, pero en general se
relacionan con las épocas de alimentación y épocas de reproducción en zonas específicas
para cada actividad. En los cetáceos, estos comportamientos dependen de dos factores
interdependientes: cambios estacionales y disponibilidad de alimentos. Clapman et al.
(1990) mencionan que las ballenas jorobadas del hemisferio norte (Golfo de Maine, Golfo
de San Lawrence, Provincia de Newfoundland, Península de Labrador, Isla Groenlandia e
Islandia), migran hacia aguas tropicales en las Antillas para cumplir sus actividades de

125
apareamiento, reproducción y crianza. Best et al. (1995) describen las características
migratorias de la ruta del sur de las ballenas jorobadas en las costas occidentales de
Sudáfrica. Estos autores concentraron sus esfuerzos en la ruta sur y no en la norte debido a
ventajas en el clima de la primera. Las preferencia de las ballenas por una zona debido a la
conformación de las costas, provocó que las ballenas fueran vistas frecuentemente pero sin
que aparentemente exista migración dentro del área.
Las zonas de alimentación de la población de ballenas que llega a Ecuador y
Colombia se encuentran en la Antártida, en donde la disponibilidad de plancton les permite
alimentarse (en el período entre octubre y marzo) y prepararse para el viaje anual a los
sitios de reproducción en aguas tropicales (entre junio y septiembre).
En el mes de marzo, la población de ballenas jorobadas inicia el viaje por la costa
Pacífica Este de Sudamérica hacia el norte, de esta manera, a finales de mayo (casi tres
meses después) los primeros individuos son observados en las aguas cálidas tropicales,
características de las zonas de reproducción, de las costas de Ecuador y Colombia. El
desplazamiento de las hembras preñadas hacia zonas de aguas tropicales indica la necesidad
de las madres de tener sus crías en sitios que faciliten un aumento en su tamaño y peso para
poder sobrevivir en las frías aguas de los polos a su regreso, luego de tres meses de
permanencia en la zona de reproducción.
La Antártida, continente que permanece congelado todo el año es el hábitat de las
ballenas jorobadas del hemisferio sur durante el verano austral. Este continente constituye
una zona con abundancia de alimento para estos cetáceos, quienes aprovechan la gran
cantidad de chumumo (Anchoa aregentivittata, A. exigua, A. ischana, peces pequeños de la
familia Engraulidae, Bearez, 1996) y de krill (Euphausia superba, invertebrados
microscópicos, Dieckmann, 2003) que constituyen parte importante de su dieta.

126
3.7 OBJETIVO GENERAL
La finalidad de este trabajo es registrar, identificar y reconocer individuos en las dos
áreas de reproducción de ballenas jorobadas en el Ecuador y, determinar si existen
individuos que están presentes en las dos áreas de reproducción durante los mismos
períodos, entre los meses de junio – septiembre de cada año mediante la generación de
información sobre los posibles desplazamientos que llevan a cabo las ballenas jorobadas
(M. novaeangliae) entre las zonas de reproducción ubicadas en el AMPNM y en las costas
de Esmeraldas.
3.8 OBJETIVOS ESPECÍFICOS
3.8.1 Identificar individuos comunes a las dos zonas de reproducción en las costas del
Ecuador durante la temporada 2001.
3.8.2 Establecer un patrón de distribución temporal y espacial de M. novaeangliae en las
dos zonas de reproducción en el Ecuador.

127
4. MATERIALES Y MÉTODOS
4.1 ÁREA DE ESTUDIO
El área donde se desarrolló esta investigación está localizada en la región marino-
costera del Ecuador, frente a las provincias de Esmeraldas y Manabí. En este documento
haré referencia a dos zonas de estudio (denominadas A y B) que constituyen las dos áreas
de trabajo.
La zona de estudio A, frente a la provincia de Esmeraldas, se extiende desde las
costas de Atacames al norte (0°52´N), hasta el Cabo San Francisco al sur (0°39´N). Hacia
el límite oriental encontramos la línea de costa de la provincia de Esmeraldas y, la línea
batimétrica (cota o curva de nivel, en profundidad) de 10 a 20 m al oeste. El relieve de la
línea de costa en este sector es irregular y presenta una conformación pronunciada en la
zona de Punta Galera (0°50´N y 80°30´W) y Cabo San Francisco (0°39´S y 80°50´W). Los
estratos submarinos del fondo en esta zona están compuestos por elementos que los
caracterizan como rocosos y con sustratos de barro (INOCAR, 1982; 1984; Figura 1).
Las costas de la provincia de Esmeraldas sufren la influencia de la corriente fría de
Humboldt, modificando el clima con vientos, oleajes, cielos nublados y presencia de garúa
en este sector, en la época comprendida entre junio y julio hasta septiembre y octubre. El
valor promedio de temperatura del mar en el período julio – septiembre de 2001 fue de
23,13 °C (Tabla 1). Los valores de temperatura del mar durante los meses de julio (Figura
4a), agosto (Figura 4b) y septiembre (Figura 4c) se mantienen relativamente constantes y

128
no existen diferencias en las condiciones del mar (oleajes y vientos) en la zona de estudio A
(IMARPE, 2001, Figura 5).
Las costas de la provincia de Esmeraldas tienen una ocupación humana histórica
muy antigua. En ellas se han desarrollado actividades pesqueras y de uso de los recursos
desde épocas remotas.
La zona de estudio B, correspondiente al Área Marina del Parque Nacional
Machalilla (AMPNM) se localiza en la zona costera del sur de la provincia de Manabí,
frente a la zona ocupada por el Parque Nacional Machalilla (Figura 1). El PNM es una de
las 31 áreas del Sistema Nacional de Áreas Protegidas del Ecuador (SNAP, Ministerio del
Ambiente, 2002). Fue establecido legalmente el 26 de julio de 1979 mediante el Acuerdo
Ministerial 0322, publicado en el Registro Oficial 069 del 20 de noviembre del mismo año.
El área total del PNM es de 56 184 ha, repartidas entre las localidades continentales
extremas de: Agua Blanca, Salaite, Ayampe y Punta Piqueros en la zona continental y las
Islas Salango y De la Plata en el área marina (INEFAN/GEF, 1998). Posee también una
reserva marina de dos millas marítimas a lo largo de su perfil costanero y alrededor de las
islas, en el Océano Pacífico.
El área de estudio B se extiende desde los 01°00´S hasta los 01°16´ S. El límite
occidental está marcado por la Isla de la Plata, en los 81°06´ W, zona con profundidades
aproximadas de 100 m. La línea costera del PNM, entre los 0 y 20 m de profundidad,
marca el límite de la zona de estudio hacia el oriente. Los fondos submarinos en esta zona
se presentan compuestos por sustratos de arena, roca, grava y zonas de arrecifes de coral
(Ayón, 1988). CDC-Ecuador/Fundación Natura (1998) califican a los ecosistemas de
arrecifes presentes en el AMPNM, en el sector de los bajos de Cantagallo, Copé y los
alrededores de la Isla de la Plata como estructuras de arrecife rocoso con parches o sectores

129
definidos de coral. INEFAN/GEF (1998) consideran que en el Ecuador y en la zona sur de
Manabí no se encuentran arrecifes coralinos verdaderos, pues éstos tienen distribución
restringida a regiones del Caribe y Australia; los que existen en este sector son arrecifes
rocosos con parches de coral, que aunque no son los mismos, tienen características
similares.
Las características climáticas del AMPNM están influenciadas por corrientes
marinas durante todo el año. En los meses de verano (temporada en la región costera del
Ecuador caracterizada por la presencia de garúa, temperaturas ambientales promedio de 16
– 18 grados, notable ausencia de sol, condiciones oceanográficas cambiantes y el mar
mostrando mayor agitación) entre junio y septiembre, la Contra-corriente Ecuatorial llega
desde el oeste y gira frente al continente dirigiéndose hacia al norte (Corriente Norte
Ecuatorial) y hacia el sur (Corriente Sur Ecuatorial). La corriente de Humboldt también
tiene influencia directa sobre la zona en esta época. Ésta llega proveniente del sur y luego
de encontrarse con la Corriente Sur Ecuatorial, forman el Frente Ecuatorial.
Los aspectos oceanográficos del mar son factores determinantes a la hora de la
distribución de los organismos marinos. La presencia de ballenas jorobadas, tortugas y
otros animales, en determinados lugares del mar están relacionados estrechamente con la
disponibilidad de alimento, la configuración geográfica de las costas, etc.
Según Hall, M. (comunicación personal) las corrientes marinas determinan la
distribución de especies de acuerdo a la estacionalidad de su permanencia en determinadas
localidades, facilitando o haciendo más difícil su supervivencia.
El área B, durante la temporada de verano, se caracteriza por la presencia de
abundante nubosidad; la temperatura promedio del área de estudio B durante el período
julio – septiembre de 2001 fue de 19,9 °C (IMARPE, 2001; Figuras 4a, 4b, 4c y 5).

130
La zona sur de Manabí, donde se ubica el territorio ocupado por el PNM y su área
marina tiene una referencia de presencia humana histórica ancestral con cerca de 5000 años
(Cultura Valdivia 3200 años A.C – Cultura Manteña 1530 años D.C., CDC-
Ecuador/Fundación Natura, 1998).
La pesca artesanal ha sido y sigue siendo la principal actividad productiva de la
zona del AMPNM y de las costas de Esmeraldas. En algunos casos, esta actividad ha sido
complementada o sustituida por otras que han tenido un crecimiento importante en los
últimos años (turismo, artesanía, agricultura).
Se sabe que las zonas de reproducción de M. novaeangliae están distribuidas en las
regiones de aguas tropicales alrededor del mundo. Sin embargo, no se conoce a detalle las
características de los individuos de la población de ballenas jorobadas que permanecen en
las dos zonas de reproducción en aguas ecuatorianas entre junio y septiembre.
4.2 REGISTRO DE DATOS EN EL CAMPO
La investigación de campo fue realizada durante la temporada de permanencia de
las ballenas jorobadas (Megaptera novaeangliae) en el AMPNM y en las costas frente a la
provincia de Esmeraldas, entre julio y septiembre de 2001.
La actividad fue desarrollada simultáneamente en las dos zonas de estudio (A y B)
por un equipo de trabajo con un investigador principal y cuatro asistentes de campo,
utilizando siempre la misma metodología.
4.2.1 FOTOGRAFÍAS

131
Los registros de datos en el campo se basaron en la recopilación de fotografías, en
formato de diapositivas, de aletas caudales de las ballenas jorobadas en las áreas de estudio.
Para la obtención de las fotografías se utilizaron, como plataformas de observación, los
botes que realizan la actividad turística de observación de ballenas en ambas zonas. En
Esmeraldas los recorridos se realizaron a bordo de embarcaciones de pescadores que
realizan actividad de observación de ballenas en la zona marina frente a las costas de Súa,
Same y Atacames. Los recorridos en el AMPNM fueron realizados a bordo de
embarcaciones turísticas para observación de ballenas, en la ruta bidireccional desde Puerto
López hacia la Isla de la Plata.
Los botes tienen una dimensión promedio de 8 a 10 m de eslora (largo) y 3 a 4 m de
manga (ancho). Las embarcaciones utilizan dos motores fuera de borda con capacidad
promedio de 100 a 115 caballos de fuerza.
Los registros fotográficos fueron realizados desde la proa o techo de los botes, a una
altura aproximada de 2,5 – 3,5 m de la superficie del mar (Figura 6).
4.2.2 EQUIPO
Las fotografías de las aletas caudales fueron obtenidas con cámaras de fotos Nikon
F3; Nikon FE3692797 y Cannon AV1, equipadas con lentes Cannon 200M 100––200 mm
1:5,6; MC P×ÁÈHAP 4,5/300 MAKPO 950234 y TAMRON AF 28––200 mm,
respectivamente, utilizando películas para diapositivas a color.
En la recolección de datos en el campo también se emplearon binoculares Nikon
Sprint II (10 x 21), cuaderno de notas, lápices, brújula y una Radio Banda Marina VHF,

132
marca ICOM de 69 canales para comunicación con guías y capitanes entre los botes de
observación.
4.2.3 TRANSECTOS
El uso de transectos en estudios con cetáceos facilita la obtención de datos en áreas
en donde los animales se pueden dispersar ampliamente. En general otro tipo de
metodología basado en marcaje o seguimiento dificultan el trabajo, principalmente por
incrementar los costos de la investigación y por la poca factibilidad de marcar ballenas
jorobadas.
La metodología utilizada en este estudio fue el registro fotográfico de individuos
desde un punto móvil llamado “plataforma de observación”, que en este caso fueron los
botes de observación de ballenas. El componente plataforma de observación, así como los
número de registro de individuos y la distancia entre el individuo y el observador son parte
de la metodología de transectos en banda según Chamberlain (1982). Los transectos en
banda son utilizados para estudios de poblaciones animales cuando no se puede realizar
otro tipo de transectos. En nuestro caso la plataforma de observación fue únicamente un
elemento logístico.
Los transectos realizados para este trabajo fueron coincidenciales y arbitrarios pues,
a pesar de que fue una ruta establecida la que se tomó como base para los registros, en cada
avistamiento de ballenas, el bote de observación se detuvo.
La distancia de observación entre la plataforma de avistamiento y las ballenas fue
arbitraria, tomando en cuenta que existen regulaciones sobre la distancia máxima de

133
acercamiento hacia las ballenas, pero que estas no siempre se cumplen. Generalmente, la
distancia no fue mayor a 80 - 100 m. Todos los animales observados durante el recorrido
son contabilizados para la estimación de abundancia.
La superficie recorrida durante los trayectos fue variable, pues la finalidad de los
viajes era encontrar grupos de ballenas, por lo que nunca la distancia recorrida y el área
abarcada fue la misma. Esta dependió siempre de la localización de las ballenas.
4.2.4 DATOS
La recolección de datos en el campo, para cada registro se clasificaron en las siguientes
categorías:
4.2.4.1 Temporales
Fecha, hora de salida y de regreso de la travesía, hora de inicio y finalización del
registro de observaciones de la ballena o grupo de ballenas.
4.2.4.2 Geográficos
Dirección del desplazamiento, ubicación aproximada.
4.2.4.3 Climáticos
Según Laevastu (1980), se considera que el estado del clima en el mar tiene influencia
directa en mayor o menor grado con el comportamiento de especies que normalmente se
encuentran en zonas superficiales del mar (pelágicas). Los datos de estado del mar son
utilizados generalmente para cálculos de densidades poblacionales, pues el mar en un
estado agitado influye directamente en la posibilidad de registro del individuos. A pesar de
que nuestro estudio no se relaciona con dinámica poblacional, hemos utilizado la misma
metodología que se usó desde el principio del monitoreo.

134
a. Cobertura del cielo, con un rango de 0 a 8, donde: 0= totalmente despejado y
8= totalmente cubierto. (Figura 7).
b. Estado del mar, utilizando el rango de 0 a 4, donde: 0= muy tranquilo, sin
oleaje (como un espejo) y 4= muy agitado con olas de tamaño entre 3 o 4 m.
4.2.4.4 Biológicos y etológicos
Identificación de la especie, número grupal (número de individuos registrados),
composición grupal (descripción de los individuos registrados con relación al tamaño y/o
edad) y comportamiento general más llamativo de acuerdo a las categorías utilizadas según
Brtnik (2000, Anexo 1).
4.2.4.5 Fotográficos
Aletas caudales y otros detalles importantes para la identificación (como cicatrices,
heridas o manchas).
4.2.5 INGRESO DE DATOS
Los datos tomados durante el trabajo de campo se registraron en un cuaderno de
notas y posteriormente se ingresaron en formularios diseñados en hojas de Excel. Los
formularios diseñados son cuatro y en cada uno se incluye información específica para cada
observación.
4.2.5.1 Formulario de Fotoidentificación
Incluye los datos del viaje. Fecha, observador, cobertura del cielo, estado del mar.
Anexo 2.
4.2.5.2 Formulario de Esfuerzo

135
Hora de inicio y de finalización de cada observación de ballenas. Anexo 3.
4.2.5.3 Formulario de Registros
Detalle de cada observación: tamaño del grupo (número de individuos),
composición del grupo (adultos, juveniles y/o crías observadas) y actividad general
desarrollada durante la observación categorizada en patrones definidos (saltos). Anexo 4.
4.2.5.4 Formulario de Fotografías
Detalle del número y código del rollo de diapositivas utilizado, el número de la
fotografía al inicio de la observación, el número y detalle del registro de cada diapositiva y
el número de la fotografía al finalizar el día de observación. Anexo 5.
4.2.6 FOTOIDENTIFICACIÓN
Se utilizó la fotoidentificación como la herramienta principal en esta investigación.
Ésta metodología se basa en la obtención de fotografías de individuos o partes de ellos para
su registro y posterior identificación. Esta herramienta generalmente se asocia con el
método de marcaje y recaptura para estimar poblaciones animales silvestres, método
estándar usado para este tipo de estudios (Lien y Katona, 1991).
Esta herramienta fue usada como metodología de registro de las aletas caudales y
dorsales de las ballenas, para su reconocimiento gracias a marcas permanentes naturales o
provocadas, pigmentaciones, heridas o muescas en sus aletas y otras partes del cuerpo
(Katona y Whitehead, 1981), que son únicas tanto para ballenas como delfines y marsopas
(Lien y Katona, 1991).
Existen numerosos ejemplos de estudios que han utilizado las marcas especiales del
cuerpo de animales como cebras (Klingel, 1965; Peterson, 1972), rinocerontes negros

136
(Goddard, 1966; Mukinya, 1973), jirafas (Foster, 1966), elefantes africanos (Douglas-
Hamilton, 1973) , leones (Schaller, 1972), chimpancés (Goodall, 1986) y tiburones martillo
(Myrberg y Gruber, 1974), para identificar individuos en las poblaciones silvestres
(Würsig, et al. 1990). Asimismo, los pelajes de focas marcados con variedad de puntos y
anillos que contrastan entre fondos claros y oscuros tienen patrones únicos de coloración
útiles para identificar a cada individuo (Crowley et al., 2002).
En general, las estimaciones de poblaciones, productividad y tasas de supervivencia
específica desarrollados durante años han utilizado diferentes métodos de marcaje de
animales; para ello se han utilizado diversas herramientas como pintura, marcas
permanentes, cortes, amputaciones, collares, anillos y otros insumos. En otros casos, la
metodología para estudios de mamíferos marinos y otras grandes especies del mar utiliza
elementos y materiales mucho más efectivos y precisos, pero asimismo mucho más
costosos (Klimley et al. 2001) como el uso de microchips insertados en la piel de ballenas,
cachalotes y delfines; dispositivos de radio-telemetría usando sistemas satelitales de rastreo,
cámaras de registro de navegación e inmersión atados a aletas dorsales o dorsos, etc.
La metodología de marcaje-captura y recaptura se ha usado en estudios de campo
para estimar la abundancia de poblaciones animales, la ocupación de hábitat, los cambios
en la abundancia relativa de éstas, composición y tamaño grupal, comportamiento, etc,
(Castro et al., 2002). Sin embargo esta metodología en la mayoría de los casos presenta
inconvenientes por ser una metodología cara que requiere gran esfuerzo y que muchas
veces no cubre las expectativas sobre los resultados esperados.
Generalmente el marcaje de animales involucra gran desgaste energético, de
recursos y el número general de individuos marcados es bajo. Además marcar grandes
números de animales puede ser logísticamente problemático e implica perturbación a los

137
mismos. La fotoidentificación, herramienta que fundamenta la metodología del marcaje-
captura y recaptura, ha sido frecuentemente utilizada para estos fines en estudios con
cetáceos. En comparación con otros métodos de marcaje-captura y recaptura, ésta presenta
ventajas significativas por ser un método de marcaje no-invasivo que puede marcar grandes
números de animales, con un gasto relativamente bajo de energía y recursos relativamente
menores que el de otros métodos de captura-recaptura. Además el tiempo requerido para
adquirir las imágenes, ingreso de datos y búsqueda de individuos previamente registrados
es pequeño en relación con la información conseguida (Crowley, et al., 2002).
4.2.7 ANÁLISIS
4.2.7.1 Registros fotográficos
Los registros fotográficos fueron analizados mediante la comparación de las
fotografías de aletas caudales, mediante la digitalización (scanning) de las diapositivas con
un Escáner Nikon MA-20 Side Mount Adapter y con el software Nikon Scan 2.5.1 y su
posterior ingreso a una base de datos digital.
Las imágenes fueron editadas en Microsoft Photo Editor de Office 2000 y en
Micrograph Picture Publisher 8, retocadas (en algunos casos) en Adobe Photo Shop 5.5 y
almacenadas en formato gráfico JPEG.
A las imágenes digitales se les asignó un número de trabajo para su clasificación;
esta metodología permite utilizar estos números de trabajo para identificar cada imagen
utilizando la sigla fl que significa fluke (aleta caudal) más un número de tres dígitos
asignado en orden ascendente (001 – 361), en consecuencia cada código consta de un par
de letras y un número (por ejemplo: fl 023).

138
El análisis de las fotografías fue realizado según parámetros diferentes: calidad de
imagen, patrones de aletas caudales según coloración, según marcas y según la forma de la
hendidura media de la aleta caudal.
4.2.7.2 Calidad de la imagen
Para analizar la calidad de imagen se tomó en cuenta la metodología propuesta por
Mizroch, et al.(1990) la cual califica y categoriza a las fotografías con puntajes que van del
1 al 3, donde se atribuye 1 a la mejor calidad y 3 a la de calidad menor.
La categoría 1 califica a la fotografía como de “muy buena calidad” lo que significa
que es muy útil para la identificación. La categoría 2 asigna a la fotografía una “calidad
media”, que la hace útil para la identificación. La categoría 3 califica a la fotografía de
“mala calidad”, lo que la hace no útil para la identificación.
La calidad de las fotografías puede ser dividida en dos variables:
a. Calidad de la fotografía propiamente dicha: claridad, resolución, cercanía,
plano de la imagen.
b. Calidad de la fotografía para identificación: claridad, enfoque, señas
particulares de la aleta, colores específicos que permitan identificar al
individuo (marcas, cicatrices).
Las diferentes categorías de “calidad de fotografía” se refieren a que si bien una
fotografía puede tener categoría 1 con relación a su enfoque y cercanía (es decir a la
“calidad de fotografía”), esta no necesariamente será una foto de categoría 1 para la
identificación del individuo (por lo que la categoría “calidad para identificación” podría ser
menor o igual).
4.2.7.3 Patrón de las aletas caudales según la coloración

139
Esta calificación forma parte de la metodología propuesta por Mizroch, et al. (1990)
para clasificar las aletas caudales según un patrón genérico de colas. Este patrón analiza los
componentes y distribución de colores básicos (blanco y negro) en las aletas caudales. Este
esquema toma en cuenta seis posibles patrones básicos, analizando el borde superior
(trailing) e inferior (leading) de la aleta. En cada caso se analiza si estos bordes son blancos
o negros y según este detalle se le asigna el nombre de la categoría como black trailing o
white trailing y correspondientemente el nombre para el borde inferior black leading o
white leading. En cada nivel existen sin embargo diferentes posibilidades para la
distribución de los colores en la aleta caudal.
Cada uno de estos patrones incluye a su vez cinco, seis o siete posibilidades de
distribución de los colores en las aletas (Figura 8). Cada aleta asignada con un patrón de
color dispone de un código que le agrupa dentro de un conjunto de aletas con esas
características; por ejemplo: btwl 34, en donde las dos primeras siglas corresponden al
patrón del borde superior (black trailing), las dos siglas siguientes corresponden al patrón
del borde inferior (white leading) y la cifra siguiente indica cual de las opciones de
combinación de colores de la aleta le corresponden específicamente Mizroch et al. (1990,
Figura 8, Tabla 2).
4.2.7.4 Patrón de las aletas caudales según marcas
Otro patrón analiza la presencia de marcas, cicatrices y huellas en algún lugar de la
aleta caudal. Mizroch et al. (1990, Tabla 3). En esta clasificación se toma en cuenta
aspectos de marcas visibles en la aleta caudal. Puede haber diferencias entre uno u otro
registro, pues hay marcas que se modifican por diferentes razones.
4.2.7.5 Patrón de las aletas caudales según la forma de la hendidura media

140
Mizroch et al. (1990) proponen otro nivel de caracterización de las aletas caudales,
basado en el tipo y forma que presenta la hendidura media de la aleta. Según estos autores
existen seis diferentes posibles tipos de hendidura media entre los lóbulos de las aletas
caudales (Figura 9, Tabla 4).
4.2.7.6 Análisis espacial y temporal de la distribución de ballenas en las zonas de
reproducción
Se compararon los datos espaciales y temporales obtenidos en las dos zonas de
reproducción de ballenas jorobadas en el Ecuador tomando los datos de frecuencia y
abundancia de ballenas jorobadas en las dos zonas de estudio en períodos mensuales y
semanales de la temporada 2001.
En ambos casos se hizo relación de los números de individuos registrados (número
grupal) con el día o semana de la observación. A partir de esta relación se obtuvieron datos
de abundancia de ballenas en las dos zonas por semana.
El esfuerzo aplicado en la observación también fue una variable utilizada para este
análisis. En este caso se hizo relación entre el esfuerzo invertido (número de horas de
observación) en las dos zonas y, el número efectivo de ballenas/hora registrados durante el
período del estudio.
4.2.7.7 Desplazamientos entre la zonas de reproducción en el Ecuador
El registro de individuos en las zonas de estudio A y B, nos dan información de las
preferencias de estos sitios por determinado individuo, según el número y fecha de su
registro en determinada zona.
La presencia de un individuo en uno de los dos sitios nos propone que este mismo
individuo podría potencialmente ser registrado en la otra zona de reproducción; de esta
manera, si existiera evidencia de su presencia en ambas zonas, podríamos comprobar su

141
desplazamiento entre las dos zonas de reproducción en Ecuador. A su vez, la evidencia de
estos desplazamientos puede ser diferenciada espacial y temporalmente.
4.2.7.8 Análisis estadístico
a. De los datos de temperaturas superficiales del mar en las zonas de estudio:
Se ingresaron los datos en hojas de cálculo agrupándolos en valores de temperatura
superficial del mar, por día, por semana y por mes.
Se obtuvo el promedio de las temperaturas superficiales del mar para cada semana
y para cada mes durante la temporada.
Se aplicó la prueba de t dependiente, misma que relaciona las medias de cada grupo
de datos que tienen cierto nivel de incertidumbre, mas no las medidas independientes. Para
la aplicación de esta prueba se utilizó el programa estadístico BioStat 1.0., con un valor de
significancia del 0.05.
Los valores de la prueba de t nos muestran si existen o no diferencias significativas
entre las temperaturas superficiales del mar en las dos zonas de estudio.
4.2.8. TÉRMINOS Y RELACIONES
4.2.8.1 Población
La oposición estacional de los hemisferios divide a la distribución mundial de
ballenas jorobadas en dos poblaciones: la de los hemisferios norte y sur (Flores-Gonzáles,
et al., 1998).
En este estudio se utiliza el término población para definir al conjunto de ballenas
jorobadas localizadas en las zonas de alimentación y reproducción del hemisferio sur
(Figura 2a y 2b).

142
4.2.8.2 Stock
El término stock fue inicialmente utilizado en relación con temas de pesquería; con
éste se denomina a los individuos de una especie que ocupan un rango espacial bien
definido. Posteriormente este término ha sido aplicado para recursos que no son
actualmente explotados por el ser humano, como es el caso de las ballenas jorobadas.
Para nuestro tema de investigación, se define a stock como el conjunto de
individuos temporal y/o espacialmente separados del resto de conjuntos de una misma
población, que ocupa sitios diferentes en regiones de alimentación y puede o no tener
intercambios de sus individuos con otros stock de la misma población (Caballero et al.,
2000a; 2000b).
Se puede decir que el grupo de individuos de una especie que ocupa un rango
espacial bien definido, independiente de otros de la misma especie es un stock. En estos
puede darse una distribución al azar y puede haber migraciones dirigidas por actividades
estacionales reproductivas o alimenticias.
Las ballenas jorobadas presentes en las costas del Ecuador y Colombia durante su
temporada de reproducción pertenecen a uno de los seis stocks de ballenas jorobadas de la
población del continente Antártico (Figura 2a y 2b).
4.2.8.3 Agregación
Se entiende por agregación al conjunto de individuos de ballenas jorobadas que
siendo parte de un mismo stock, al llegar a las zonas de reproducción, se separan y cada una
de las agregaciones permanecen separadas en los centros de reproducción; posteriormente,
al retornar a las zonas de alimentación vuelven a juntarse formando parte del mismo stock;
este término puede ser equivalente al de substocks que mencionan Volgenau et al. (1995).

143
Los términos de “agregaciones reproductivas” y “agregaciones alimentarias” son
utilizados por un investigador (Olavaria, comunicación personal) en un estudio que trata de
caracterizar genéticamente dichas agregaciones de los stocks de la población antártica de
ballenas jorobadas; éste se basa en la utilización de marcadores moleculares mitocondriales
y nucleares de muestras de ADN.
En el presente trabajo de investigación se utilizará el término agregación para
nombrar a las ballenas de cada zona de estudio (Figura 2a y 2b).
4.2.8.4 Grupo
La definición de grupo es un tanto complicada. En unos casos es definido como “los
animales que mantienen contacto acústico mediante los cantos que pueden tener un alcance
de hasta 10 km a la redonda” (Brtnik, 2000). Este criterio significaría que todos los
individuos de un área determinada son miembros de un mismo grupo.
Otra definición califica a un grupo como aquellos animales que se encuentran en un
área de 100 metros a la redonda del conjunto de ballenas observado y cuyas actividades y
direcciones están sincronizados (Whitehead, 1983; Mobley y Herman, 1985; Perry et
al.1990) o, como los individuos que nadan juntos y mantienen direcciones y respiraciones
sincronizados (Clapham, 1993), pero con una estructura inestable y dinámica, la cual dura
solamente pocas horas (Brtnik, 2000). Whitehead (1983), Weinrich et al. (1991) y Matilla
et al. (1994) mencionan que las ballenas jorobadas que en las áreas de reproducción se
encuentran solas, en parejas o en grupos con cría. Una ballena solitaria es considerada
como grupo por sí misma.
Para este estudio se toma la definición de Clapham (1993) de base para definir al
grupo, como el conjunto de individuos que se encuentran en un área de observación

144
controlada por el observador, mismos que mantienen algún tipo de interacción en cuanto a
actividad, dirección de navegación, comportamiento o respiraciones.
Las ballenas analizadas en Esmeraldas y Machalilla nos muestran que podrían ser
considerados como “subconjuntos” de la población de ballenas jorobadas de la Antártida,
sin embargo, el término que utilizaremos en este punto para identificar a las ballenas de
Esmeraldas y de Machalilla será el de “agregación”.
Por lo tanto, es poco probable que con la ausencia de relaciones permanentes entre
ellas, las ballenas formen grupos de una manera estable (Figura 2a y 2b).
4.2.9 INTERCAMBIO ENTRE INDIVIDUOS
Los desplazamientos que existen entre las zonas de reproducción de Colombia y
Ecuador están probados; así también estudios acerca del ADNm de ballenas jorobadas del
hemisferio Sur muestran que existe algún patrón de intercambio dentro y entre las cuencas
oceánicas, por la presencia de nucleotipos idénticos, compartidos entre las ballenas de los
hemisferios norte y sur (Baker et al ., 1998).
Estos estudios prueban que hay diferencias significativas entre los nucleotipos y
heterogeneidad en la diversidad de haplotipos de las ballenas de cuatro regiones analizadas
(grupo IV de Australia occidental; grupo V “componente occidental” de Australia oriental;
grupo V “componente oriental” de Nueva Zelanda y Tonga y grupo VI de la Península
Antártica, Isla Gorgona, Colombia y América del Sur) correspondientes a tres stocks de
ballenas jorobadas (Baker et al., 1998).
Baker et al. (1998) menciona que las diferencias significativas entre los nucleotipos
de los stocks de la Antártida y la heterogeneidad en la diversidad de los haplotipos de los

145
grupos analizados, prueban teóricamente que existe un movimiento e intercambio de
individuos en forma longitudinal y no latitudinal. Este intercambio estaría favoreciendo que
las ballenas del grupo VI del stock de Antártida, que se localizan en aguas sudamericanas
de las costas del Pacífico, incluidas las ecuatorianas, podrían mostrar la tendencia a
desplazarse longitudinalmente entre las zonas de reproducción en Ecuador y entre las zonas
de Ecuador - Colombia –Panamá.
Para la realización del presente estudio he tomado en cuenta los términos que mejor
describen a las ballenas jorobadas como miembros de una especie que se localizan en
determinada zona en un período definido.
4.2.10. INTERACCIONES CON OTROS CETÁCEOS
Debido a la existencia de pocas referencias que documenten aspectos etológicos de
los mamíferos marinos en Ecuador, creo importante hacer mención de eventos de
interacción interespecíficas que se registraron simultáneamente a la recopilación de datos
para este trabajo de investigación, pues estos hechos son un aporte a la ciencia y
contribuyen a conocer y aclarar aspectos de la historia natural de estos animales.
Se detallan dos eventos de interacción relacionados con delfines manchados
(Stenella attenuata) y con orcas (Orcinus orca); estos recogen los detalles más importantes
de las observaciones y son relatos descriptivos de las situaciones observadas.

146
5. RESULTADOS Y DISCUSIÓN
5.1 ASPECTOS CLIMÁTICOS Y FACTORES FÍSICOS EN LAS DOS ZONAS DE
ESTUDIO
Conociendo que la presencia de ballenas jorobadas en las costas de América del
Sur, frente a Colombia y Ecuador se debe a la necesidad de éstas de tener aguas cálidas
para su reproducción (nacimiento de crías y apareamiento), consideramos que la
temperatura del agua es un factor determinante sobre la presencia de esta especie en las
zonas de reproducción de Ecuador.
El análisis de las temperaturas superficiales diarias del mar nos ha dado información
sobre sus promedios. La temperatura promedio en la temporada de estudio (julio –
septiembre) de la zona A es de 23,1ºC y es ligeramente superior a la temperatura media de
la zona B (19,9ºC), con una diferencia de 3,2ºC (Tabla 1). Analizando los valores promedio
de temperatura superficial del mar en las dos zonas de estudio durante el período de
investigación vemos que la media se incrementa en un 16,2% (de 19,9 ºC a 23,1 ºC) en la
zona A con relación a la zona B (Tabla 1, Figura 5).
Los datos de las temperaturas superficiales del mar fueron tomadas de las imágenes
satelitales diarias de la temperatura superficial del mar en las dos zonas de estudio; la
interpretación de estas imágenes se basan en espectros fotocromáticos de las diferencias
térmicas del agua de mar (Figuras 4a, 4b y 4c).
Luego de su análisis podemos concluir que la zona de estudio A es más cálida que
la zona de estudio B, en esta temporada del año; la prueba de t pareada nos muestra que

147
estadísticamente existen diferencias altamente significativas entre las medias de las
temperaturas de las dos zonas de estudio t (9, 10 = 6,89**, p<0,05 ).
Estudios sobre preferencias de hábitat relacionadas con la temperatura superficial
del mar no han sido desarrollados aún, el estudio sobre uso de hábitat por parte de las
ballenas jorobadas en la zona de estudio B está siendo desarrollado en la actualidad
(Yumiseva, en prep.), sin embargo según los valores de temperaturas superficiales del mar
obtenidos y la diferencia entre ambas zonas podemos decir que la temperatura del agua del
mar sí puede ser un factor que determina la preferencia de una zona por parte de las
ballenas. En la zona de estudio A (que presentó temperatura más cálida) la frecuencia de
ballenas ha sido mayor (2.6 ballenas/hora) durante la temporada frente a la zona de estudio
B (2.4 ballenas/hora), hecho que podría explicarse por la mayor calidez de las aguas de esta
zona y la preferencia de las ballenas por aguas con estas características para que sus crías
permanezcan en la época de nacimiento y crianza.
5.2. DATOS OBTENIDOS
Se lograron recopilar 815 registros (fotografías) de aletas caudales de ballenas
jorobadas durante el período julio-septiembre de 2001, de las cuales 210 (correspondientes
a la calidad de identificación 1 y 2) fueron catalogadas, codificadas y asignadas con un
número de trabajo dentro de la base de datos de aletas caudales de cada una de las zonas.
Se tuvieron un total de 440 horas de muestreo, durante los viajes a bordo de
embarcaciones de observación de ballenas de operadoras turísticas (n = 256) en las costas
de Esmeraldas (107) y en el AMPNM (149, Tabla 8).

148
El número total (n) de aletas caudales identificadas como individuos diferentes fue
de 98. De ellas 18 % pertenecieron a registros hechos en Esmeraldas y 82 % corresponden
a registros del AMPNM. Los datos mencionados anteriormente no son indicadores
relacionados con la abundancia de ballenas en cada zona estudiada, es decir, aunque se
aplique un mayor esfuerzo de muestreo en el área B, los números de individuos
identificados podrían aumentar, mas no la frecuencia de individuos.
Una de las explicaciones para esta diferencia en las frecuencias de ballenas sería la
temperatura del agua (que es mayor en la zona A), otra sería la menor presión turística en la
zona A y por último la configuración de los fondos marinos de la zona A: zonas de bajas
profundidades (25 – 30 m.) están más cercanos a la costa que en el área B.
5.2.1 FRECUENCIA DE AVISTAMIENTO DE INDIVIDUOS EN LAS DOS ZONAS
DE ESTUDIO
Con un esfuerzo de muestreo promedio quincenal de 30,3 horas en la zona de estudio A
y de 27 horas en la zona de estudio B (Figura 10), se obtuvo que la frecuencia de ballenas
jorobadas en las zonas de estudio A y B entre la tercera semana de junio y la segunda de
julio es similar (2,7 y 2,8 ballenas/hora, respectivamente, Figura 11).
La frecuencia total de individuos registrados en las dos zonas (2,6 ballenas/hora en
Esmeraldas y 2,4 ballenas/hora en AMPNM) (Tabla 5) estuvo relacionada con el esfuerzo
de muestreo promedio, (201 horas en Esmeraldas y 240 horas en AMPNM (Tabla 8.
Figuras 11 y 12). Analizando las Figuras 11, 12, 13 y 14, vemos que existe una diferencia
temporal en relación con la frecuencia de ballenas en las dos zonas de estudio (Tabla 5).

149
En el área de estudio A durante la semana del 20 de agosto al 2 de septiembre, el
esfuerzo de muestreo fue de 24 horas. En el mismo período en el área de estudio B el
esfuerzo de muestreo fue de 41 horas. Sin embargo, a pesar de la disminución de esfuerzo
de muestreo en la zona A con respecto a la B (aproximadamente en un 58,5%), la
frecuencia de ballenas en la primera es del 3,3 ballenas/hora a diferencia del 1,5
ballenas/hora registradas en la segunda, lo que significa que la frecuencia de individuos en
la zona de estudio A es más alta que en la zona de estudio B.
Comparando los datos de ambas zonas de estudio del año 2000 (Brtnik, 2000;
Yumiseva, en prep.) y del 2001 (Figuras 11, 12, 13 y 14), se observa una diferencia en la
frecuencia de individuos de las dos zonas. Esta diferencia nos muestra un período de
incremento de la abundancia en la zona A y, simultáneamente una disminución en la
abundancia en la zona B entre el 20 de agosto y el 2 de septiembre del 2000. Este
incremento podría estar relacionado con el desplazamiento de individuos desde la zona B a
la zona A, mismos que fueron registrados en B en media temporada y que probablemente
estaban de paso hacia su zona de destino A; este decremento que se hace notar desde el
período del 13 de agosto en la zona B y se prolonga hasta el período del 2 de septiembre del
2000, fechas en las que probablemente empiezan a registrarse en la zona B individuos que
provienen de la zona A y que empiezan su viaje de regreso hacia la Antártida.
En el período del 28 de agosto al 2 de septiembre del 2000, la frecuencia de ballenas
en las zonas fue de 2,9 ballenas/hora en la zona A y de 1,7 ballenas / hora en la zona B
(Figura 12). Durante el período del 3 al 13 de septiembre de 2000, cuando el esfuerzo para
la zona A fue de 15 horas y para el área B fue de 43 horas, la frecuencia de individuos fue
de 1,6 ballenas / hora en la zona A y de 0,8 ballenas en la zona B (Figura 12).

150
En el período comprendido entre el 6 de agosto y el 17 de septiembre del 2001 se
vuelve a observar un incremento en la frecuencia de la zona A y una disminución
importante en la frecuencia de individuos de la zona B (Tabla 5, Figura 13 y 14) que
también podría estar relacionada con un registro previo en la zona B de los individuos que
se dirigían a la zona A. Posteriormente, en el período del 27 de agosto al 9 de septiembre
del 2001, el área B muestra otro decrecimiento en la frecuencia de individuos en B, lo cual
puede mostrar que se volvieron a registrar en la zona B individuos provenientes de la zona
A, que en este caso estaban de regreso hacia la Antártida.
Esta constante disminución en la abundancia de ballenas en la zona B con relación a
la zona A, comparativamente con el año 2000 y la temporada 2001puede darnos una idea
de que las ballenas que en un período determinado son registradas en la zona B pueden ser
individuos que se dirigen (en temporada media) hacia la zona A, o individuos que
provienen de la zona A y se dirigen de regreso a la Antártida y pasan por la zona B.
Pensamos también en la posibilidad de que algunos individuos de la zona B se desplacen a
la zona A y viceversa durante su permanencia en las dos zonas, evidencia de la que
carecemos ante la ausencia de fotografías de individuos en ambas zonas; esto podría
verificarse con mayor cantidad de datos y un seguimiento a las poblaciones durante las
temporadas siguientes.
5.2.2 COMPARACIÓN DE LOS REGISTROS DE M. novaeangliae DEL ÁREA A CON
LOS DEL ÁREA B
5.2.2.1 Patrón de clasificación por la presencia de marcas, cicatrices y huellas en algún
lugar de la aleta caudal

151
Las categorías de coloración en las aletas caudales permitieron que se identifiquen
grupos de aletas siguiendo los lineamientos de Mizroch, et al. (1990). Entre las más
abundantes tenemos aquellas que muestran líneas (209) y las que no tienen una marca
definida (200, Tabla 9). Otras categorías de aletas caudales son las de rasgaduras (166) y
puntos (156). En ningún caso existen reavistamientos de individuos en ambas zonas con los
patrones de marcas detallados en la metodología.
5.2.2.2 Patrón de clasificación por la muesca media entre los lóbulos de las aletas
caudales
Mizroch, et al. (1990) mencionan que la forma de la hendidura media de las aletas
caudales nos muestran una forma de agrupar a las colas en categorías. La mayor cantidad
de aletas (111) presentan una hendidura media redondeada, en el tipo de hendidura no
identificada se incluyen 90 aletas caudales y en el de forma de “u” tenemos 74 aletas
caudales (Tabla 10).
5.2.2.3 Categorías en la calidad de las fotografías analizadas
El análisis de la calidad de fotografía para fotoidentificación usando el rango 1- 3
nos muestra que las fotografías de calidad 1 son 95, las de calidad 2 son 115 y las de
calidad 3 son 43.
El mayor número de fotografías corresponde a la categoría 2, es decir la calidad de
la mayoría de fotografías es apta para la fotoidentificación. En la categoría 1 se incluyen 95
fotografías. Las de categoría 3 (43) no han sido consideradas para la fotoidentificación pues
no muestra características ni definición claramente diferenciables.
5.2.3 IDENTIFICACIÓN DE INDIVIDUOS EN LAS DOS ZONAS DE ESTUDIO

152
Entre el período julio – octubre de 2001, individuos de ballenas jorobadas en ambas
zonas de estudio fueron registradas, catalogadas e identificadas. La identificación de los
individuos se logró a partir de una combinación de métodos e instrumentos que se basaron
en la comparación de las aletas caudales.
En el área de estudio A fueron identificadas 17 ballenas como individuos diferentes,
en el área de estudio B fueron 82 las ballenas identificadas, a partir de 210 fotografías de
aletas caudales, dándonos un total de 99 ballenas identificadas en las dos zonas durante el
período de estudio (Tabla 11). De los 99 individuos registrados, uno (fl 0355) corresponde
a un reavistamiento (recaptura del individuo a través del registro fotográfico de su aleta
caudal) de una ballena registrada en el año 1999 (Castro, et al., 2002, Anexo 6), por lo que
el número total de individuos identificados es de 98.
5.2.4 PATRONES DE DISTRIBUCIÓN ESPACIAL Y TEMPORAL DE M. novaeangliae
EN LAS ZONAS DE REPRODUCCIÓN EN ECUADOR
Este estudio se hizo con el conocimiento previo de que el número de ballenas en la
zona B es de alrededor de 350 individuos (Scheidat, 2001), dato al que llegó con el método
de “captura – recaptura”, con una variación: “captura” por “marca” de los individuos
mediante el registro fotográfico de las aletas caudales (fotoidentificación). El principio de
este método se basa en que los individuos de una población marcados y liberados serán
recapturados en un número proporcional a su abundancia dentro de esa población.
El número de ballenas jorobadas determinado por Scheidat (2001) podría
modificarse en el caso de que se confirmen desplazamientos de individuos de cada
agregación entre las dos zonas, situación que esta investigación no ha probado.

153
Observando las figuras 13 y 14 vemos que la frecuencia de ballenas (número de
ballenas/hora) en cada una de las áreas de estudio varía y esta variación nos puede indicar
la preferencia que estos animales muestran por una u otra zona en determinada fecha. Esta
presencia de ballenas en determinada zona, en una fecha, nos da una idea de una
distribución espacial y temporal de las ballenas durante los procesos reproductivos en las
zonas A y B. Se ha evidenciado que las ballenas pueden permanecer como agregaciones
independientes en determinados períodos en ambas zonas, permanencia que varía según los
valores de frecuencia de ballenas registradas.
Se observa en estos períodos una disminución de la frecuencia en la zona B y
simultáneamente un incremento de la misma, en la zona A (Figuras 11 y 12, Tabla 5),
situación que podría estar relacionada con desplazamientos temporales de las ballenas entre
las zonas de estudio, pero que no ha sido probada aún, pues no han sido registradas ballenas
comunes a las dos zonas, hecho que evidenciaría que las zonas de reproducción en el
Ecuador comparten individuos que indistintamente se movilizan entre estas con finalidad
de reproducción.
Solamente con el registro de individuos en las dos zonas podríamos probar que las
agregaciones incluyen individuos que se movilizan entre ambas zonas, situación que
disminuiría la presión sobre las agregaciones de ballenas en cada zona por la industria de la
observación comercial de ballenas jorobadas.
En este caso el patrón de distribución temporal de las ballenas jorobadas de las
agregaciones A y B del Ecuador está determinado por las fechas (mes de agosto y
septiembre) de la temporada en la que se encuentran. El desplazamiento de los individuos
de la zona B hacia otro lugar puede darnos idea de que sí existe un patrón de distribución
espacial que no necesariamente corresponde a los movimientos entre las zonas de

154
reproducción del Ecuador cercanos a la costa sino que puede darse en zonas de aguas
profundas.
Los individuos que se estarían movilizando entre estas zonas tendrían otras rutas
fuera de la zona de estudio, por lo que no se han registrado a estos individuos en
desplazamiento.
5.2.4.1 Composición grupal en las zonas de reproducción del Ecuador
La composición grupal de las ballenas en ambas zonas de estudio difiere. Esta
variación radica en la frecuencia de ballenas en las diferentes fechas de la temporada en
cada zona. Durante el mes de julio se ve una mayor cantidad de ballenas registradas (120 y
103), durante las semanas del 9 al 22 de julio de 2001 y del 23 de julio al 5 de agosto del
2001 en el AMPNM. Las ballenas registradas durante este mismo período en Esmeraldas
es de 63 y 69 (Tabla 6).
Según los datos analizados, el promedio del tamaño grupal en la zona de estudio A
al inicio de temporada (mes de julio de 2001) fue de 2.1 ballenas/grupo. A mediados de
temporada (mes de agosto) se formaron grupos con un promedio de 2.6 animales/grupo y
los grupos más grandes fueron observados a finales de temporada (septiembre) con un
promedio de 2.8 animales/grupo.
La conformación de los grupos en la zona del AMPNM tuvo diferentes
proporciones con relación a los individuos registrados. El 49% de las ballenas observadas
formaron grupos de dos animales, 16% de las ballenas observadas estuvieron solas y 35%
de las ballenas observadas formaron grupos de 3 o mas animales.
Ballenas solitarias y grupos de dos animales fueron observados con mayor
frecuencia a inicios de temporada. Grupos grandes, de 3 o más animales fueron observados
durante toda la temporada.

155
En Esmeraldas, el 42.4% de las ballenas registradas formaban parte de grupos de 2
individuos, el 34.9% fueron ballenas solitarias y el 13.3% fueron grupos de 3 individuos.
Grupos de ballenas de un individuo fueron más frecuentes a inicios de temporada, grupos
de dos individuos a mediados y grupos de tres o más individuos fueron más comunes a
finales de temporada.
La composición grupal está determinada por la temporada en la que se registra
debido a que a inicios de temporada será más frecuente observar a ballenas que permanecen
en grupos de dos, lo que significan posibles parejas en busca de oportunidades de
apareamiento. En ambas zonas (A y B) el porcentaje de grupos de 2 individuos era más
frecuente (zona A 49% y zona B 42%). Mientras la temporada va avanzando, los grupos de
ballenas se hacen más numerosos pues los machos tratan de buscar hembras receptivas y
existe mayor posibilidad de grupos grandes a finales de temporada en donde se registran
machos competitivos disputándose las hembras, mismas que cada vez son menos
frecuentes.
5.2.5 DESPLAZAMIENTO DE LAS BALLENAS JOROBADAS ENTRE LAS ZONAS
DE REPRODUCCIÓN DEL ECUADOR
Durante el período 4 de julio - 2 de octubre de 2001 no se han registraron
reavistamientos de ballenas jorobadas en las zonas de estudio A y B. Conociendo que desde
inicios de la década de los noventa las costas ecuatorianas en el sur de Manabí y norte de
Guayas fueron identificadas como sitio de permanencia temporal de ballenas jorobadas
(Félix et al., 1994; Félix y Hasse, 1998; 2001a, 2001b), hecho que fue probado con la
presencia de ballenas jorobadas en las zonas A y B, es interesante mencionar como

156
resultado de la presente investigación que las zonas A y B no comparten individuos de
ballenas jorobadas entre sí.
La caracterización preliminar de las agregaciones de ballenas jorobadas de las áreas
de estudio A y B realizada en el año 2001 nos muestra que son grupos diferentes y
temporalmente independientes, es decir, que entre las ballenas que permanecen en el
AMPNM y en Esmeraldas durante la temporada de reproducción no existe intercambio de
individuos.
Según Garrigue et al. (2002) las agregaciones de ballenas que forman parte de uno
de los stocks de la Antártida, al llegar a las zonas de reproducción (Colombia y Ecuador en
las dos áreas de estudio) se separan y permanecen temporalmente alejadas en estos centros;
posteriormente, al retornar a las zonas de alimentación volverían a reunirse formando un
mismo stock, sin embargo algunos individuos registrados en Ecuador (alrededores de la Isla
de la Plata, 01º16´S, 81º06´W) han sido observados también en la zona circundante de la
Isla Gorgona en Colombia, 02º 47´N, 78º18´W en algunas ocasiones (Flores-Gonzáles y
Capella, 1998, Tabla 7), por lo que se puede afirmar que la agregación (población, según
Félix et al., 1998) de ballenas de Ecuador está en contacto con la de Colombia y que existe
un intercambio de individuos entre las dos agregaciones (poblaciones, según Félix et
al.,1998), aunque aún no tenemos evidencias de la presencia de los mismos individuos en
las dos zonas de reproducción de Ecuador (A y B) alternativamente.
Las fechas de reavistamientos de individuos mencionadas en la Tabla 7 nos hace
pensar que fueron registros de aquellos individuos que migraban hacia o desde la zona de
reproducción, provenientes o con destino de o hacia la Antártida, explicándose así su
presencia en ambas zonas; con estos datos podemos especular sobre las razones por las que
una ballena (0022COL, Flores-Gonzáles y Capella, 1998) fue registrada el 26 de agosto en

157
aguas colombianas y el 21 de septiembre, dos años más tarde, en aguas ecuatorianas. Si
tomamos en cuenta que la temporada de reproducción puede extenderse entre junio y
octubre (hasta noviembre en algunos casos), en agosto esta ballena pudo haber tenido su
apareamiento en aguas colombianas y el registro de la misma ballena en el mes de
septiembre en aguas ecuatorianas podría indicar que en estas fechas estaba retornando hacia
el sur, para lo cual utilizó el área costera ecuatoriana en su desplazamiento hacia la zona de
alimentación de la Antártida.
Otro ejemplo podemos observar en la ballena (0035COL, Flores-Gonzáles y
Capella, 1998), que fue registrada el 21 de agosto en aguas colombianas y el 21 de
septiembre, dos años después, en aguas ecuatorianas.
Estos registros de ballenas en aguas colombianas y en fechas posteriores en aguas
ecuatorianas nos pueden dar una idea de los patrones de desplazamiento que tiene esta
especie en las zonas de reproducción del Pacífico Sur. Las ballenas podrían retornar hacia
las zonas de alimentación navegando por aguas costeras, menos profundas y cercanas al
continente, como un posible mecanismo de protección contra depredadores.
La falta de reavistamientos de ballenas en las zonas de estudio A y B pueden darnos
una pauta para entender su preferencia por una de las dos zonas. Por lo tanto, si la
conformación geográfica de la costa de una zona de reproducción de ballenas jorobadas
presenta las condiciones adecuadas y hace que esta agregación de ballenas la prefiera, no
habría entonces necesidad de movimientos de ballenas entre áreas de reproducción
cercanas. El caso del AMPNM y de las costas de Esmeraldas serán entonces lo
suficientemente favorables para que las ballenas que se encuentran en cada una de ellas no
tenga la necesidad de movilizarse de una a otra, pues en ellas disponen de agua cálida apta
para que sus crías engorden lo suficiente antes de volver a sus zonas de alimentación en la

158
Antártida, de fondos poco profundos que hacen menos probable el ataque de depredadores
y que favorecen el aprendizaje de las crías sobre cómo respirar, saltar, escapar, etc.
Debido a la falta de registros fotográficos de individuos, que muestren
reavistamientos en las dos zonas de estudio, podemos concluir que no existe evidencia de
que los individuos se estén desplazando entre las dos zonas de reproducción del Ecuador
durante su permanencia temporal en estas, a pesar de que ambas se caracterizan por
factores físicos y ambientales que las hacen favorable para que las ballenas las busquen y
cumplan sus actividades reproductivas en ellas.
Los patrones de movilización de las ballenas en estas zonas son aún desconocidos,
aunque debido a referencias bibliográficas (Flores-Gonzáles et al., 1998), el stock de la
Península Antártica, Isla Gorgona, y Colombia es el mismo para América del Sur, en donde
se incluyen también a las ballenas que permanecen en aguas ecuatorianas, y si existe
evidencia de que las ballenas de Ecuador se movilizan entre esta zona y la Isla Gorgona
(Flores-González et al., 1998), las agregaciones de ballenas que permanecen en las dos
zonas de reproducción de Ecuador podrían estar desplazándose entre éstas durante su
permanencia en la zona.
Según los resultados de Baker et al. (1998) habría intercambio de individuos y
posibles cruzamientos entre individuos de los hemisferios norte y sur, más no así
intercambio y/o cruzamiento de individuos entre grupos y/o stocks de ballenas del
hemisferio sur.
Los datos de la presente investigación nos indican que aparentemente los individuos
registrados en el AMPNM y en Esmeraldas corresponden a dos agregaciones diferentes,
situación que, desde el punto de vista de manejo del recurso, perjudica su sustentabilidad a

159
largo plazo pues la presión que se aplica a las ballenas por parte de la actividad de
observación es acumulativa para cada una de ellas.
En este caso no sería factible que las ballenas que reciben cierta cantidad de presión
y estrés en Esmeraldas puedan bajar al AMPNM y disminuir o aumentar esta presión, sino
que la agregación de Esmeraldas estaría sometida a la misma presión de la actividad en
Esmeraldas y viceversa para la agregación del AMPNM.
Tomando en cuenta que la actividad en cada una de las áreas es diferente en
cantidad de embarcaciones, grupos de observación y presión por parte de la afluencia
turística, estaríamos enfrentados a una presión diferenciada para cada agregación.
En el caso de Esmeraldas, la presión, por el momento sería menor, pues hay menos
cantidad de embarcaciones y demanda turística que en el AMPNM, la que estaría
soportando la mayor presión.
El manejo del recurso en función de estos resultados son determinantes en la
conservación de las ballenas de ambas zonas de reproducción del Ecuador.
5.2.6 INTERACCIONES CON OTROS CETÁCEOS EN LAS ÁREAS DE
REPRODUCCIÓN
A pesar de que los objetivos de este estudio no se relacionan con las asociaciones
interespecíficas entre cetáceos es importante mencionar la presencia de este tipo de
relaciones entre ballenas jorobadas y otras especies de mamíferos marinos, que en el caso
particular de Orcinus orca puede ser determinante en la presencia o ausencia de ballenas
jorobadas en determinada zona. De hecho la mayor o menor cantidad de registros de O.
orca en zonas de reproducción de ballenas jorobadas se relaciona con una mayor o menor

160
posibilidad de encontrar grupos de ballenas jorobadas principalmente con crías, pues los
grupos de ballenas con cría generalmente tratarán de evitar encuentros con las orcas por ser
éstos los principales depredadores de esta especie.
5.2.6.1 Con delfines manchados (Stenella attenuata)
Grupos de ballenas jorobadas fueron observadas por tres veces, interactuando con
grupos de delfines en la localidad de Punta Escalera, Isla de la Plata, en el AMPNM. Dos
de ellas la interacción fue entre M. novaeangliae y Stenella attenuata (delfines manchados).
Los grupos de ballenas jorobadas, con una composición de 3 de animales (entre
adultos y cría) mostraron actividad de “integración”, consistente en navegación entre el
grupo de delfines, haciendo una serie de movimientos de navegación junto a los S.
attenuata, mientras que el grupo de éstos, de aproximadamente 300 individuos, número
calculado en base a lo propuesto por Carwardine (1995, Figura 14) que menciona como
método para este cálculo, multiplicar el número de delfines que se observan sobre la
superficie del mar por 1.75, cifra que nos dice el número de delfines bajo la superficie y
con este valor se puede calcular el número total aproximado de delfines de ese grupo.
Los delfines navegaban y se incluían mezclándose dentro del grupo de ballenas
jorobadas, haciendo para ello un sinnúmero de piruetas y atractivos movimientos. El
número aproximado de delfines fue calculado en base al principio, como muestra la figura,
que en cada conteo, bajo la superficie del agua puede haber dos veces el número de delfines
que se ve sobre la superficie (Carwardine, 1995).
Mientras los delfines acompañaban a las ballenas en su ruta, la que se mantuvo
invariable en las ballenas (dirección S-N), los delfines poco a poco fueron distanciando su
rumbo hasta que en un momento dado se desplazaron en un sentido opuesto al de las

161
ballenas. En este momento los dos grupos de cetáceos tomó rumbos diferentes y se alejaron
mutuamente.
5.2.7.2 Con orcas (Orcinus orca)
La presencia de O. orca en aguas ecuatorianas continentales ha sido reportada por
Tirira (1998) y sus registros constan como ocasionales. Sin embargo Eisemberg (1989) no
reporta la presencia de las orcas en territorio ecuatoriano. El registro de O. orca en la zona
de estudio “B” ha sido confirmada en numerosas ocasiones por pescadores artesanales
mediante comunicación personal, quienes identifican claramente a estos depredadores
como uno de los causantes de la muerte de ballenas jorobadas en este sector. En el año
1996, Denkinger (comunicación personal) reporta la presencia de orcas en las
inmediaciones de la zona de estudio B, con un claro comportamiento de búsqueda y
consecución de alimentación mediante el ataque a ballenas jorobadas. Scheidat (2001)
también reporta la presencia de orcas en la zona de estudio B.
Durante el mes de agosto de 2001 se registraron 4 individuos de O. orca (3 adultos
y una cría) en actitud de ataque a una ballena jorobada adulta (Moreno de los Ríos,
comunicación personal). Este registro de un grupo de orcas en actitud de ataque a una
ballena jorobada, muestra que es un evento que se produce de manera consecutiva en esta
zona de reproducción en el Ecuador y no es un hecho aislado, sino que podría considerarse
como un hecho constante en temporada de reproducción de ballenas jorobadas.
Según la observación de Moreno de los Ríos (comunicación personal), la ballena
jorobada se ubicó a estribor y el grupo de orcas se acercó al lugar. La ballena jorobada
detuvo su actividad y desapareció. Las orcas empezaron a alejarse y el bote se acercó a las
orcas y se inició la observación como a cualquier grupo de ballenas.

162
Se observó a las orcas viajando, con las aletas dorsales fuera de la superficie del
agua. Dos de ellas de aproximadamente 2 m de alto, las otras aletas dorsales eran de menor
tamaño. Uno de los individuos que viajaba con el grupo de orcas era una cría, de
aproximadamente un medio o tres cuartas partes del tamaño de los otros individuos.
La observación del grupo de orcas duró aproximadamente 10 min. La cría con un
adulto (posiblemente la madre) se acercó al bote (a 6-7 m), luego se aproximó aún más,
hacia la proa del bote; hicieron un par de inmersiones más, muy cerca del bote y luego se
alejaron. El resto del grupo permaneció como a 20 m del bote y cada vez se alejaban más.
Luego los adultos y la cría se unieron a este grupo y se alejaron.
Las orcas tomaron dirección hacia la izquierda y el bote lo hizo hacia la derecha. La
ballena que mostraba actividad a lo lejos mostró coletazos, medio saltos. A los 5 minutos
de iniciada la observación, se vio al grupo de orcas acercarse directamente a la ballena
(misma que se encontraba estaba entre el bote y las orcas) a estribor. Se observó entonces
que la ballena se empezó a alejar hacia la popa. Las orcas siguieron acercándose y tomaron
la misma dirección que la ballena y la siguieron. El bote giró su dirección con relación al
movimiento de la ballena. Se vio que la ballena emergió hacia la superficie, se quedó quieta
y las orcas se localizaron en algún lugar, probablemente rodeando a la ballena.
La cría de orca permaneció atrás y alejada de la ballena. Las orcas iniciaron el
ataque a la ballena, se sumergieron y emergieron por varias ocasione. La ballena en clara
actitud de defensa, daba coletazos y se movía con mucha agitación, sumergiéndose y
emergiendo constantemente. El ataque se hizo visible a intervalos de segundos en la
superficie. Esta actividad duró por aproximadamente 8 minutos.

163
En un momento dado las orcas emergieron y fueron observadas alejándose del sitio.
La ballena también se alejó en dirección contraria con una velocidad mayor a la que
normalmente se las observa en cualquier avistamiento.
El registro de este comportamiento agresivo y de ataque por parte de orcas a
ballenas jorobadas no fue efectivo para las orcas, pues no lograron su objetivo (matar y
comer a la ballena). Sin embargo, la presencia de una cría de orca nos puede llevar a pensar
en que no fue un ataque propiamente dicho sino más bien una actividad de enseñanza de la
madre y otros adultos de orca hacia la cría que todo el tiempo permaneció en actitud no
participativa pero pendiente (Figura 15).

164
6. RECOMENDACIONES
6.1 METODOLOGÍAS PARA CARACTERIZACIÓN DE AGREGACIONES DE
BALLENAS JOROBADAS
6.1.1 CANTOS
Si bien es cierto, la fotoidentificación es una herramienta altamente efectiva al
momento de caracterizar e identificar individuos de ballenas jorobadas; en la
caracterización de agregaciones o poblaciones, las marcas específicas de las aletas caudales
o dorsales de cada individuo se convierten en una herramienta con un menor rango de
efectividad, comparándola con otras que pueden darnos un espectro mayor de elementos
para caracterizar a una población, stock o agregación.
Los cantos en las ballenas jorobadas son elementos inherentes a su biología y
etología, de gran ayuda a la hora de determinar las características de las ballenas a nivel de
agregaciones, stocks o poblaciones. El conocimiento de las características de los cantos de
las agregaciones de ballenas jorobadas presentes en Ecuador (Esmeraldas y AMPNM) con
detalle, nos daría elementos para establecer posibles relaciones entre individuos de una y
otra área.
Se recomienda utilizar esta herramienta con este objetivo y en temporadas futuras
hacer grabaciones y registros de cantos para tener mayores elementos de juicio en el
proceso de caracterización de las agregaciones de ballenas jorobadas Ecuador.

165
6.1.2 ANÁLISIS GENÉTICO DE INDIVIDUOS
Numerosos son actualmente los estudios de poblaciones de fauna silvestre que
utiliza el componente genético para la caracterización de estas. Alrededor del mundo se han
implementado trabajos de investigación que utilizan muestras de piel, sangre y/o tejidos
para el análisis de estructuras celulares (ADN) para el establecimiento de relaciones entre
poblaciones o las líneas de parentesco entre especies.
Estudios del ADNm de los stocks de ballenas jorobadas del hemisferio Sur están
siendo desarrollados actualmente (Olavaria, comunicación personal), y tratan de establecer
las relaciones y parentescos entre ellos. Las agregaciones del Pacífico Sur Este (con
excepción de muestras obtenidas en algunos individuos de la zona de Isla Gorgona en
Colombia) no son objeto de este muestreo y por lo tanto se recomienda iniciar un estudio
al respecto, incluyendo muestras de individuos de las agregaciones de Ecuador, pues esta
información enriquecerá la información de la que se dispone para este sector.
Un análisis genético de ambas agregaciones nos dejará en claro las posibles
relaciones que existen entre individuos de Esmeraldas, AMPNM e Isla Gorgona.
6.2 TURISMO DE OBSERVACIÓN DE BALLENAS EN LAS ZONAS DE
REPRODUCCIÓN
Como sabemos, las áreas de estudio son actualmente centros de atracción turística
que ve en la observación de ballenas una fuente alternativa de distracción, utilizado tanto
por operadores turísticos como por turistas como uno de los principales destinos en el
Ecuador.

166
La utilización de recursos naturales como atractivos turísticos es una tendencia
mundial para dar alternativas de trabajo a comunidades y poblaciones. Las comunidades
que se asientan en las áreas de estudio y sus zonas de influencia no son la excepción.
El objetivo macro de esta investigación es principalmente brindar elementos
técnicos que permitan un mejor uso del recurso ballena jorobada. Los resultados del
presente estudio pueden ser utilizados para propender a un uso adecuado de este recurso
evitando o reduciendo el uso desordenado y la sobreexplotación así como el impacto y la
afectación de la actividad turística en la población de Megaptera novaeangliae que
anualmente se localizan en las costas de Esmeraldas y en el AMPNM.
El conocimiento de que los grupos de ballenas de ambas zonas son independientes
nos permite concluir que la presión que proviene de la actividad de observación de ballenas
y que es recibida por cada agregación será acumulativa, provocará mayores efectos en cada
agregación y el impacto humano a mediano y largo plazo será mayor de no encontrarse la
manera de garantizar que se cumplan las disposiciones que el Reglamento de Observación
de Ballenas y otros mamíferos marinos, propone.
Este menciona algunos aspectos, tres de los que merecen resaltarse: la distancia de
acercamiento y permanencia de los botes con relación a los grupos, el tiempo de
observación que cada bote debe cumplir y la actitud del bote con relación a la ballena o
grupo de ballenas, mismo que debe tratar respetar la actividad de los grupos en el momento
del acercamiento, especialmente si existen crías.
Es necesario también promover y apoyar todos los esfuerzos de investigación sobre
la interacción entre ballenas jorobadas y turismo, pues la investigación es la única
herramienta que tenemos para conocer y garantizar que las actividades que involucren a
este recurso estén siendo bien o mal desarrolladas y de esta manera hacer factible el manejo

167
sustentable y a largo plazo de este importante recurso natural del mundo, del que
numerosas comunidades se benefician actualmente.

168
7. BIBLIOGRAFÍA CITADA
Alava, J. J., Barragán, M. J., Castro, C., Carvajal, R. 2001. Breve nota acerca de los
varamientos de ballenas jorobadas (Megaptera novaeangliae) en la costa del Ecuador
durante la temporada 2001. En prep.
Ayón, H. 1988. Grandes rasgos geomorfológicos de la costa ecuatoriana. Serie Informes #
1. PMRC (Programa de Manejo de Recursos Costeros). Ecuador.
Baillie, J., Groombridge, B. 1996 IUCN Red list of threatened animals. International Union
for Conservation of Nature and Natural Resources. The IUCN special survival
commission. The World Conservation Union Gland. 378 pp.
Bajpai, S., Gingerich, P.D. 1998. A new Eocene archaeocete (mammalia, Cetacea) from
India and the time of origin of whales. Proc. Natl. Acad. Sci. USA. Vol. 95, pp.
15464 – 15468, December 1998. Evolution. [en línea] http://www-
personal.umich.edu/~gingeric/PDFfiles/PDG344.pdf [Consulta: 12 feb 2003].
Baker, C. S., Herman, L. M. 1984. Aggressive behavior between humpback whales
(Megaptera novaeangliae) wintering in Hawaiian waters. Can. J. Zool. 62:1932-1967.
Citado en: Abundance, Habitat Use, Behavior and Management of the Humpback
Whales (Megaptera novaeangliae) in the Machalilla National Park, Ecuador (M.
Scheidat). Dissertation. Zur Erlangung des Doktorgrades der Mathematisch-
Naturwissenschaftlichen Fakultät. Der Christian-Albrechts-Universität, Zu Kiel.
Baker, C. S. , Flores-González, L., Abernethy, B., Rosenbaum, H. C., Slade, R. W.,
Capella, J., Bannister, J. L. 1998. Mitochondrial DNA variation and maternal gene
flow among Humpback Whales of the Southern Hemisphere. Marine Mammal

169
Science: Vol. 14, No.4, pp.721-737. [en línea].
http://apt.allenpress.com/aptonline/?request=get-abstract&issn=0824-
0469&volume=014 [Consulta: 16 ene 2003].
Best, P. B., Sekiguchi, K., Findlay, K. P. 1995. A Suspended migration of humpback
whales Megaptera novaeangliae on the west coast of South Africa. Vol.118:1-12,
1995. Marine Ecology Progress Series Mar. Ecol. Prog. Ser. Published March 9.
Bearez, P. 1995. Lista de Peces Marinos del Ecuador Continental. Rev. Biol. Trop. 44(2):
731 – 741, 1996.
Brown, M. R., Corkeron, P. J., Hale, P. T., Schultz, K. W., Bryden, M. M. 1995. Evidence
for a sex-segregated migration in the humpback whale (Megaptera novaeangliae).
Proc. R. Soc. Lond. B (1995)259, 22-234. The Royal Society.
Brtnik, P. 2000. La influencia de la actividad de whale watching al comportamiento de la
ballena jorobada (Megaptera novaeangliae) en el Parque Nacional Machalilla. Tesis
de Masterado. Universidad Tuebingen.
Caballero, S., Hamilton, H., Flórez-González, L., Capella, J., Olavarria, C., Rosenbaum,
H.C., Baker, C.S. 2000a. Stock identity and diversity of humpback whale
mitochondrial DNA lineages on the colombian winter breeding grounds. SC/52/IA14.
En: Exploring the Greatest Iceberg in History. Antartica: The Ross Ice Shelf. [en
línea] http://www.iceisland.net/carlos.html [Consulta: 22 ene 2003].
Caballero, S., Hamilton, H., Jaramillo, C., Capella, J., Flórez-González, L., Olavarría, C.,
Rosembaum, H.C., Guhl F., Baker, C.S. 2000 b. Genetic characterization of the
colombian pacific coast humpback whale population using RAPDs and mitochondrial
DNA sequences Humpback Whale Conference. 29th august - 1st september 2000.
Brisbane, Australia. En: Exploring the Greatest Iceberg in History. Antartica: The

170
Ross Ice Shelf. [en línea] http://www.iceisland.net/carlos.html [Consulta: 22 ene
2003].
Castro, C., Denkinger, J., Gonzales, J., Scheidat, M. 1999. Historic presence of marine
mammals on the Ecuadorian coast. Abstracts of the 13th annual conference of the
European Cetacean Society in Valencia, Spain April 1999.
Carwardine, M. 1995. Whales, dolphins and porpoises. First American Edition. DK
Publishing Inc. New York, USA.
Carwardine, M., Hoyt, E., Ewan Fordyce, R. & Gill, P. s/a. Collins: Whales & Dolphins.
The Ultimate Guide to Marine Mammals.
Castro, C., González, J. 2002. Población de la Ballena Jorobada Megaptera novaeangliae,
Balaenopteridae, en el Parque Nacional Machalilla. Tesis presentada como requisito
para optar el título de Doctor en Biología. Universidad Central del Ecuador. Facultad
de Filosofía, Letras y Ciencias de la Educación. Escuela de Biología.
CDC-Ecuador, Fundación Natura. 1998. Machalilla: Guía Natural y Cultural. Editorial
Nuevo Arte. Quito.
Chittleborough, R. G. 1958. The breeding cycle of the female humpback whale Megaptera
nodosa (Bonna-terre). Australian Journal of Marine and Freshwater Research, 9:1-18.
CITES. 1996. Convención sobre el comercio internacional de especies amenazadas de
fauna y flora silvestres. Proyecto de conservación y manejo de fauna silvestre.
Ministerio de Agricultura. Santiago de Chile. 67 pp. En: Tirira, 1999. Mamíferos del
Ecuador. Publicación Especial 2. Museo de Zoología QCAZ/CBA/PUCE/SIMBIOE.
Quito.
Clapman, P. J., Mattila, D. K. 1990. Humpback Whale Songs as Indicators of Migration
Routes. Marine Mammal Science, 6(2): 155-160 (April 1990).

171
Clapham, P. J. 1993. Social organization of humpback whales on North Atlantic feeding
ground. Symmp. Zool. Soc. Lond. No. 66: 131-145.
Crowley, S. M., Kelly, B. P., Daniel, R. 2002. Individual identification of harbor seals for
application to population and behavioral studies. Manuscript. [en línea]
http://www.google.com/search?hl=es&ie=UTF-8&oe=UTF-
8&q=Crowley+Phoca+patterns&btnG=B%C3%BAsqueda+en+Google&lr=
[Consulta: 25 jun 2002].
Darling, et al. 1996. Movement of a Humpback Whale (Megaptera novaeangliae) from
Japan to British Columbia and Return. Marine Mammal Science, Vol. 12, No.2 1996.
Dawbin, W. H. 1966. The seasonal migratory cycle of humpback whales. Pp. 145-170. In:
(K. S. Norris, ed) Whales, Dolphins and Porpoises. University of California Press,
Berkley and Los Angeles. Xv+789 pp. En: A note on humpback whales off the coast
of Ecuador during the 1997 “El Niño” event (F. Félix y B. Haase) 3(1):000-000.
J.Cetacean Res. Manage. 2001.
Denkinger, J., Pairoa-Riofrío, C., Scheidat, M. 1997. Humpback Whales (Megaptera
novaeangliae) breeding off the coast of Manabí, Ecuador. Proceedings of the 11th
Annual Conference of the European Cetacean Society (ECS) in Stralsund, Germany.
Dieckmann, G. 2003. Stiftung Alfred-Wegener-Institut für Polar- und Meeresforschung in
der Helmholtz- Gemeinschaft. AWI. [en línea]. http://www.awi-
bremerhaven.de/Eistour/krill-e.html [Consulta: 12 dic 2002].
Douglas-Hamilton, I. 1973. On the ecology and behavior of the Lake Manyara elephants. E.
Afr. Wild. J. 11:401 – 3. En: Methods of Photo-identification for small cetaceans (B.
Würsig, T. A. Jefferson, 1990) Rep. Int. Whal. Comm. (Special Issue 12). Pp.45 - 52.
EDIDAC. 1994. Gran Diccionario Enciclopédico Visual.

172
Eisemberg, J. F. 1989. Mammals of the Neotropics. Volume I. Panamá, Colombia,
Venezuela, Guyana, Suriname, French Guyana. University of Chicago Press.
Chicago.
Estivill, A., Urbano, C. 1997. Cómo citar recursos electrónicos. Artículo encargado por la
revista Information World en Español. (publicado en el número de septiembre de
1997). [en línea]
http://www.puce.edu.ec/zoologia/S%20Burneo/Zoologia%20Lab/citas%20internet.do
c [Consulta: 17 abr 2003].
EVEREST Cúpula. Diccionario de la Lengua Española. Sexta Edición. España.
FAO.Guía FAO para la Identificación de Especies para los Fines de la Pesca.
Volumen III. Pacífico Centro Oriental. FAO/Comisión Europea/Instituto de
Investigación Senckenberg/Agencia Noruega para el Desarrollo Internacional.
Sección Mamíferos Marinos.
Findlay, K. P., Best, P. B. 1996. Estimates of the Numbers of Humpback Whales Observed
Migrating Past Cape Vidal, South Africa, 1988-1991. Marine Mammal Science,
12(3):354-370 (July 1996).
Félix, F., Samaniego, J. 1994. Incidental catches of small cetaceans in the artisanal fisheries
of Ecuador. Rep. Int. Whal. Comm. (special issue 15).Pp 475-480. En: La
Investigación de la Ballena Jorobada (Megaptera novaeangliae) alrededor de la Isla
de la Plata, Manabí, durante 1995 (F. Félix y B. Haase) Acta Oceanográfica del
Pacífico. INOCAR, Ecuador. 9(1), 1998.
Félix, F., Haase, B, Samaniego, J. 1994. Promotion of “whale watching” in Ecuador.
Project report to Whale & Dolphin Conservation Society, Bath, England. October
1994. 10 pp. (No publicado). En: La Investigación de la Ballena Jorobada (Megaptera

173
novaeangliae) alrededor de la Isla de la Plata, Manabí, durante 1995. Acta
Oceanográfica del Pacífico (F. Félix, F. y B. Haase) INOCAR, Ecuador; 9(1), 1998.
Félix, F., Haase, B., Chiluiza, D., Amador, P. 1996. Beaching and bycatch of sperm and
humpback whales in Ecuador: updating an alarming continuing story. Informe
No.SC/48/014 presentado a la 47ava Reunión de la Comisión Ballenera Internacional
en Aberdeen, Escocia. Junio 1996. Informe 5 pp. (No Publicado). En: La
Investigación de la Ballena Jorobada (Megaptera novaeangliae) alrededor de la Isla
de la Plata, Manabí, durante 1995. Acta Oceanográfica del Pacífico (F. Félix, F. y B.
Haase) INOCAR, Ecuador; 9(1), 1998.
Félix, F., Haase, B., Davis, J. W., Chiluiza, D., Amador, P. 1997. A Note on Recent
Strandings and Bycatches of Sperm Whales (Physeter macrocephalus) and
Humpback Whales (Megaptera novaeangliae) in Ecuador. Rep. Int. Whal. Comm. 47,
1997. pp. 917-919.
Félix, F., Haase, B. 1998. La Investigación de la Ballena Jorobada (Megaptera
novaeangliae) alrededor de la Isla de la Plata, Manabí, durante 1995. Acta
Oceanográfica del Pacífico. INOCAR, Ecuador; 9(1), 1998.
Félix, F., Haase, B. 2001a. A note on humpback whales off the coast of Ecuador during the
1997 “El Niño” event. J. Cetacean Res. Manage. 3(1):000-000, 2001.
Félix, F., Haase, B. 2001b. Towards an estimate of the Southeastern Pacific humpback
whale stock. J. Cetacean Res. Manage. 3(1):000-000, 2001.
Félix, F., Haase, B. 2001. The humpback whale off The Coast of Ecuador, population
parameters and behavior. Revista de Biología Marina y Oceanografía 36(1): 61 – 74,
julio de 2001.

174
Flores, L. B., Haase, B., Capella, J., Bravo, G., Félix, F., Lyrholm, T., Gerrote, T. 1994.
Movimiento de ballenas jorobadas Megaptera novaeangliae en la costa Oeste de
Sudamérica. Anais da 6ª Reuniao da Trábalo de Especialistas em Mamíferos
Aquáticos da América do Sul. Florianópolis, Brasil. 24-28 de Outubro de 1994, p.77
(resumen). En: La Investigación de la Ballena Jorobada (Megaptera novaeangliae)
alrededor de la Isla de la Plata, Manabí, durante 1995. Acta Oceanográfica del
Pacífico (F. Félix, F. y B. Haase) INOCAR, Ecuador; 9(1), 1998.
Flores-Gonzáles, L., Capella, J. 1998. Changes in Winter Destinations and the
Northernmost Records of Southeastern Pacific Humpback Whales. Marine Mammal
Science, 14(1):189-196 (January 1998).
Foster, J.B. 1966. The giraffe of Nairobi National Park: Home range, sex ratios, the herd,
and food. E. Afr. Wildl. J. 4:139–48. En: Methods of Photo-identification for small
cetaceans (B. Würsig, T. A. Jefferson, 1990) Rep. Int. Whal. Comm. (Special Issue
12). Pp.45 - 52.
Fundación Natura / CDC-Ecuador. 1998. Machalilla: guía natural y cultural. Editorial
Nuevo Arte. Quito.
Garrigue C., Aguayo, A., Amante-Helweg, V., Baker, C.S., Caballero, S., Clapham, P.,
Constantine, R., Denkinger, J., Donoghue, M., Florez-Gonzalez, L., Greaves, J.,
Hauser, N., Olavarria, O., Pairoa, C., Peckham, H., Poole, M. 2002. Movements of
humpback whales in Oceania, South Pacific. Journal of Cetacean Research and
Management 4 (3) : 255-260.
Goddard, J. 1966. Mating and courtship of the black rhinoceros (Diceros bicornis L.). E.
Afr. Wildl. J. 4:69 – 75. En: Methods of Photo-identification for small cetaceans (B.
Würsig, T. A. Jefferson, 1990) Rep. Int. Whal. Comm. (Special Issue 12). Pp.45 - 52.

175
Goodall, J. 1986. The Chimpanzees of Gombe: Patterns of Behavior. The Belknop Press of
Harvard University Press. 673 pp. En: Methods of Photo-identification for small
cetaceans (B. Würsig, T. A. Jefferson, 1990) Rep. Int. Whal. Comm. (Special Issue
12). Pp.45 - 52.
Haase, B. 1990. Resultados preliminares de observaciones sobre cetáceos en la costa
continental Suroeste del Ecuador. 4ta. Reunión de Trabajo de Especialistas en
Mamíferos Acuáticos de América del Sur. 12-15 de Noviembre. Valdivia, Chile. p.30.
(resumen). En: La Investigación de la Ballena Jorobada (Megaptera novaeangliae)
alrededor de la Isla de la Plata, Manabí, durante 1995. Acta Oceanográfica del
Pacífico (F. Félix, F. y B. Haase) INOCAR, Ecuador; 9(1), 1998.
Haase, B., Félix, F. 1991. La ballena jorobada (Megaptera novaeangliae) alrededor de la
Isla de la Plata, Parque Nacional Machalilla, Manabí. Informe presentado al Parque
Nacional Machalilla. Ecuador. Nov. 1991. 7 pp. (No Publicado). En: La Investigación
de la Ballena Jorobada (Megaptera novaeangliae) alrededor de la Isla de la Plata,
Manabí, durante 1995. Acta Oceanográfica del Pacífico (F. Félix, F. y B. Haase)
INOCAR, Ecuador; 9(1), 1998.
Haase B., Félix, F. 1993. Identification of the Humpback Whale population along the
Ecuadorian coast. Project report to Whale & Dolphin Conservation Society, Bath,
England. February 1993. 5 pp. (No Publicado). En: La Investigación de la Ballena
Jorobada (Megaptera novaeangliae) alrededor de la Isla de la Plata, Manabí, durante
1995. Acta Oceanográfica del Pacífico (F. Félix, F. y B. Haase) INOCAR, Ecuador;
9(1), 1998.
Haase B., Félix, F. 1994. A note on the incidental mortality of sperm whales (Physeter
macrocephalus) in Ecuador. Rep. Int. Whale Comm. (special issue 15). Pp. 481-483.

176
En: La Investigación de la Ballena Jorobada (Megaptera novaeangliae) alrededor de
la Isla de la Plata, Manabí, durante 1995. Acta Oceanográfica del Pacífico (F. Félix,
F. y B. Haase) INOCAR, Ecuador; 9(1), 1998.
Hammond, P. S., Mizroch, S. A., Donovan, G. P. 1990. Individual Recognition of
Cetaceans. Use of Photo-Identification and other techniques to estimate population
parameters. Report of the International Whaling Commission, Special Issue 12.
Cambridge.
Harvey Pough, F., Janis, C., Heiser, J. 1999. Vertebrate Life. Fifth Edition. Prentice Hall.
New Jersey.
Instituto del Mar del Perú (IMARPE). 2001. Temperaturas Superficiales del Mar Diaria.
NAVO/NOAA/UPRSIG. [en línea].
http://www.imarpe/gob.pe/argen/adc/web/2001/tsmdiaria2001.html [Consulta: 14
diciembre 2001].
INEFAN/GEF. 1997. Evaluación del Área Marina del Parque Nacional Machalilla. I Parte.
INEFAN/GEF. 1998. Plan de Manejo Parque Nacional Machalilla. Área Marina. Tomo II.
INEFAN/GEF. 1998. Plan de Manejo del Parque Nacional Machalilla. Resumen Ejecutivo.
INOCAR, 1982. Carta I.O.A. 101. Levantamiento Hidrológico. Punta Coquitos y Ensenada
de San Francisco. Primera Edición. Agosto 1982.
INOCAR, 1984. Carta I.O.A. 102. Levantamiento Hidrográfico. Cabo de San Francisco y
Punta Ballena. Primera Edición. Noviembre 1984.
International Whaling Commission. 1999. Report of the Scientific Committee. Anex E.
Report of the Sub-Committee on Other Great Whales. J. Cetacean Res. Manage I
(Suppl.): 117-155. En: A note on humpback whales off the coast of Ecuador during

177
the 1997 “El Niño” event (F. Félix y B. Haase) 3(1):000-000 2001a. J. Cetacean Res.
Manage.
International Whaling Commission. 2001. Report of the Scientific Committee. J. Cetacean
Res. Manage. (Suppl.)3:In press. En: Towards an estimate of the Southeastern Pacific
humpback whale stock (F. Félix y B. Haase) 3(1):000-000. J. Cetacean Res. Manage.
IUCN-Sur/GTNBD/EcoCiencia. 1997. Informe de los talleres para la identificación de las
prioridades de investigación y conservación para la biodiversidad de Ecuador.
Taller de Especialistas en Mamíferos del Ecuador, realizado el 5 y 6 de noviembre
de 1996. Informe Final preparado por EcoCiencia para la Embajada de los Países
Bajos y el Proyecto INEFAN/GEF. Quito.
IUCN. 2002. 2002 IUCN Red List of Threatened Species [en línea].
http://www.wcmc.org.uk/species/animals/categories.html
Katona, S. K., Whitehead, H.P. 1981. Identifying humpback whales using their natural
markings. Polar Rec. 20: 439-44.
Kaufman,G. y P. Forestell. 1999. Hawaii´s Humpback Whales. A complete whalewatchers
Guide. Pacific Whale Foundation. Hong Kong.
Kibblewhite, A. C., Denham, R. N., Barnes, D. J. 1967. Unusual low-frequency signals
observed in New Zealand waters. Journal of the Acoustical Society of America.
41:644-655, En: Humpback Whale Songs as Indicators of Migration Routes. (P. J.
Clapman y D. K. Mattila). 6(2): 155-160. Marine Mammal Science. April 1990.
Klimley, A. P., LeBoeuf, B. L., Cantara, K. M., Richert, J. E., Davis, S. F., Van
Sommeran, S. 2001. Radio-acoustic positioning as a tool for studying site-specific
behavior of the white shark and other large marine species. Marine Biology. Abstract
Volume 138 Issue 2 (2001) pp 429-446. ISSN: 1432-1793 [en línea]

178
http://www.Springer LINK:<font c.../b>Biology – Abstract Volume 138 Issue 2
[Consulta: 27 mar 2002].
Klingel, H. 1965. Notes on the biology of the plains zebra, Eqqus quagga boehmi,
Matschic. E. Afr. Wildl. J. 3:86 – 8. En: Methods of Photo-identification for small
cetaceans (B. Würsig, T. A. Jefferson, 1990) Rep. Int. Whal. Comm. (Special Issue
12). Pp.45 - 52.
Krebs, C. 1986. Ecología. Análisis experimental de la distribución y abundancia. Ediciones
Pirámide, S.A. México.
Laevastu, T. 1980. Manual de Métodos de Biología Pesquera. Organización de las
Naciones Unidas para la Agricultura y la Alimentación (FAO).
Leatherwood, S., Reeves, R. R. 1983. The Sierra Club Handbook of Whales and Dolphins.
Sierra Club Books, San Francisco xvii+302pp. En: A note on humpback whales off
the coast of Ecuador during the 1997 “El Niño” event (F. Félix y B. Haase) 3(1):000-
000 2001a. J. Cetacean Res. Manage.
Levenson, C., Leapley, W. T. 1978. Distribution of humpback whales (Megaptera
novaeangliae) in the Caribbean determined by a rapid acoustic method. Journal of the
Fisheries Research Board of Canada 35:1150-1152. En: En: Humpback Whale Songs
as Indicators of Migration Routes. (P. J. Clapman y D. K. Mattila). 6(2): 155-160.
Marine Mammal Science. April 1990.
Lien, J., Katona, S. 1991. A guide to the photographic identification of individual whales
based on their natural and acquired markings. The American Cetacean Society: 69 pp.
Mackintosh, N. A. 1942. The southern stocks of whalebone whales. Discovery Rep. XXII:
197-300. Citado en: Abundance, Habitat Use, Behavior and Management of the
Humpback Whales (Megaptera novaeangliae) in the Machalilla National Park,

179
Ecuador (M. Scheidat). Dissertation. Zur Erlangung des Doktorgrades der
Mathematisch-Naturwissenschaftlichen Fakultät. Der Christian-Albrechts-Universität,
Zu Kiel.
Mackintosh, N. A. 1965. The stocks of whales. The Buckland Foundation. Citado en:
Abundance, Habitat Use, Behavior and Management of the Humpback Whales
(Megaptera novaeangliae) in the Machalilla National Park, Ecuador (M. Scheidat).
Dissertation. Zur Erlangung des Doktorgrades der Mathematisch-
Naturwissenschaftlichen Fakultät. Der Christian-Albrechts-Universität, Zu Kiel.
Mattila, D. K., Guinee, L. N., Mayo, C. A. 1987. Humpback whale songs on a North
Atlantic feeding ground. Journal of Mammalogy 68:880-883. En: Humpback Whale
Songs as Indicators of Migration Routes. (P. J. Clapman y D. K. Mattila). 6(2): 155-
160. Marine Mammal Science. April 1990.
Matilla, D.K., Clapham, P.J., Vasquez, O., Bowman, R. S. 1994. Occurrence population,
composition and habitat use of the humpback whale in Samana Bay, Dominican
Republic. Can. J. Zool. 72: 1898-1907. En: Humpback Whale (Megaptera
novaeangliae) Research Literature Bibliography. [en línea].
www.oceania.org.au/soundnet/research/humpback.html [Consulta: 22 feb 2003].
Ministerio del Ambiente. 2002. Hacia un Nuevo Modelo de Gestión 2000-2002. Informe de
Actividades. Diciembre 2002.
Mizroch, S. A., Beard, J. A., Lynde, M. 1990. Computer Assisted Photo-Identification of
Humpback Whales. Rep. Int. Whal. Comm. (Special Issue 12). Pp. 63- 70.

180
Mobley, J. R. Jr., Herman, L. M. 1985. Transience of social affiliations among humpback
whales (Megaptera novaeangliae) on the Hawaiian wintering grounds. Can. J. Zool.
67: 762-772.
Mukinya, J.G. 1973. Density, distribution, population structure and social organization of
the black rhinoceros in Masai Mara Game Reserve. E. Afr. Wildl. J. 11:385 – 400.
En: Methods of Photo-identification for small cetaceans (B. Würsig, T. A. Jefferson,
1990) Rep. Int. Whal. Comm. (Special Issue 12). Pp.45 - 52.
Muñoz, E., Felix, F., Florez-Gonzalez, L., Haase, B., Katona, S., Lodi, L., McOsker, M.,
Robertson, K., Stevick, P., Siciliano, S. 1998 Migrations of individually identified
humpback whales (Megaptera novaeangliae) between the Antarctic Peninsula and
South America. Abstract. First Marine Mammal Science Conference. Mónaco, 20-24
January 1998. [VMML P 4028].
Myrberg, A.A. y Gruber, S.H. 1974. The behavior of the bonnethead shark, Sphyrna tiburo.
Copeia 1974:358 – 74. En: Methods of Photo-identification for small cetaceans (B.
Würsig, T. A. Jefferson, 1990) Rep. Int. Whal. Comm. (Special Issue 12). Pp.45 - 52.
NOAA. 2003. Humpback Whale (Megaptera novaeangliae): Western North Pacific Stock.
[en línea]
http://www.nmfs.noaa.gov/prot_res/PR2/Stock_Assessment_Program/Cetaceans/Hu
mpback_Whale_(Western_N._Pacific)/AK00humpbackwhale_WesternN.Pacific.pd
f [Consulta: 13 ene 2003].
Payne, R. S., Mc Vay, S. 1971. Songs of humpback whales. Science 173: 585-597. En:
Humpback Whale Songs as Indicators of Migration Routes. (P. J. Clapman y D. K.
Mattila). 6(2): 155-160. Marine Mammal Science. April 1990.

181
Perry, A., Baker, C.S., Herman, L.M. 1990. Population characteristics of individually
identified humpback whales in the central and eastern North pacific: a summary and
critique. Rep. Int. Whal. Comm. (Special Issue 12): 307-317. Citado en: Humpback
Whale (Megaptera novaeangliae): Western North Pacific Stock. [en línea]
http://www.nfms.noaa.gov/protres/PR2/StockAssessmentProgram/Cetaceans/Hump
back_Whale_(Western_N._Pacific)/AK00humpbackwhale_WesternN.Pacific.pdf
[Consulta: 13 ene 2003].
Peterson, J.C.B. 1972. An identification system for zebra (Eqqus burchelli Gray). E. Afr.
Wildl. J. 10:59 – 63. En: Methods of Photo-identification for small cetaceans (B.
Würsig, T. A. Jefferson, 1990) Rep. Int. Whal. Comm. (Special Issue 12). Pp.45 - 52.
Runtz, M.W.P. 1991. Moose behavior. [en línea].
http://www.mooseworld.com/behavior.htm [Consulta: 23 abr 2003].
SALVAT. 1975. El Mar. Gran Enciclopedia Salvat. Tomo 3 (bar-cha). Salvat S.A.
de Ediciones Pamplona, España.
Schaeff, C. 1997. Whale Blow Holes. Manuscript. [en línea].
http://whale.wheelock.edu/archives/ask97/0008.html [Consulta: 22 mar 2003].
Schaeff, C. 1998. Order Cetacea: whales, dolphins and porpoises [en línea].
http://animaldiversity.ummz.umich.edu/chordata/mammalia/cetacea.html
[Consulta: 22 abr 2003].
Schaller, G.B. 1972. The Serengeti Lion. University of Chicago Press. 480 pp. Citado en:
Methods of Photo-identification for small cetaceans (B. Würsig, T. A. Jefferson,
1990) Rep. Int. Whal. Comm. (Special Issue 12). Pp.45 - 52.

182
Scheidat, M., Castro, C., Denkinger, J., Gonzáles, J., Adelung, D. 2000. A breeding area for
humpback whales (Megaptera novaeangliae) off Ecuador. J. Cetacean Res. Manage.
2(3):165-171. 2000.
Scheidat, M. 2001. Abundance, Habitat Use, Behavior and Management of the Humpback
Whales (Megaptera novaeangliae) in the Machalilla National Park, Ecuador.
Dissertation. Zur Erlangung des Doktorgrades der Mathematisch-
Naturwissenschaftlichen Fakultät. Der Christian-Albrechts-Universität, Zu Kiel.
Stevick, P.T., Oien, N., Matilla, D.K. 1998. Migration of a humpback whale (Megaptera
novaeangliae) between Norway and the West Indies. Marine Mammal Science
14(1):162-166. (January 1998).
Stone, G. S., Katona, S. K., Tucker, E. B. 1987. History, migration and present status of
humpback whales, Megaptera novaeangliae, at Bermuda. Biological Conservation
42-133-145. En: Humpback Whale Songs as Indicators of Migration Routes. (P. J.
Clapman y D. K. Mattila). 6(2): 155-160. Marine Mammal Science. April 1990.
Tirira, D. 1999. Mamíferos del Ecuador. Publicación Especial 2. Museo de Zoología
QCAZ/CBA/PUCE/SIMBIOE. Quito.
Tirira, D. (Ed.). 2001. Libro Rojo de los Mamíferos del Ecuador Serie Rojos del Ecuador.
Tomo 1. Publicación Especial sobre los Mamíferos del Ecuador 4.
SIMBIOE/EcoCiencia/Ministerio del Ambiente/UICN. Quito.
Townsend, C. H. 1935. The distribution of certain whales as shown by logbood records of
american whaleships. Zoologica XIX (1):3-50, figs. 1-2. plates I-IV. Scientific
Contributions of the New York Zoological Society. The Society of the Zoological
Park: New York. Citado en: Abundance, Habitat Use, Behavior and Management of
the Humpback Whales (Megaptera novaeangliae) in the Machalilla National Park,

183
Ecuador (M. Scheidat). Dissertation. Zur Erlangung des Doktorgrades der
Mathematisch-Naturwissenschaftlichen Fakultät. Der Christian-Albrechts-Universität,
Zu Kiel.
Tyack, P. 1981. Interactions between singing Hawaiian whales and conspecifics nearby.
Behav. Ecol. Sociobiol. 8:106-116. Citado en: Abundance, Habitat Use, Behavior and
Management of the Humpback Whales (Megaptera novaeangliae) in the Machalilla
National Park, Ecuador (M. Scheidat). Dissertation. Zur Erlangung des Doktorgrades
der Mathematisch-Naturwissenschaftlichen Fakultät. Der Christian-Albrechts-
Universität, Zu Kiel.
Tyack, P., Whitehead, H. 1983. Male competition in large groups of wintering humpback
whales. Behavior. 83:132-154. Citado en: La influencia de la actividad de whale
watching al comportamiento de la ballena jorobada (Megaptera novaeangliae) en el
Parque Nacional Machalilla (P. Brtnik). Tesis de Masterado. Universidad
Tuebingen.
Utreras, V., Merlen, G., Tirira, D., Castro, C. 2001. Ballena Jorobada (Megaptera
novaeangliae). Pp. 77-79, En: Libro Rojo de los Mamíferos del Ecuador Serie
Rojos del Ecuador. Tomo 1. Publicación Especial sobre los Mamíferos del Ecuador
4. (D. Tirira, ed) SIMBIOE/EcoCiencia/Ministerio del Ambiente/UICN. Quito.
Valsecchi, E., Hale, P., Corkeron, P., Amos§, W. 2002. Social structure in migrating
humpback whales (Megaptera novaeangliae). Molecular Ecology (2002) 11: 507 –
518. Blackwell Sciece Ltd.
Volgenau, L., Kraus, S. D., Lien, J. 1995. The impact of entanglements on two substocks of
western North Atlantic humpback whale, Megaptera novaeangliae. Canadian
Journal of Zoology, 73, pp 1689-1698. Citado en: Humpback Whale (Megaptera

184
novaeangliae). Research Literature Bibliography: Oceanía Project. [en línea].
http://www.oceania.org.au/soundnet/research/humpback.html [Consulta: 15 mar
2003].
Weinrich, M.T.; Kuhlberg, A.K., 1991. Short-term association patterns of humpback whale
(Megaptera novaeangliae) groups on their southern Gulf of Maine feeding ground.
Can. J. Zool. 69:3005 – 3011. Citado en: Scientific Publications. Whale Center of
New England. WCNE Abstracts. [en línea]
http://www.whalecenter.org/abstract.htm#DNA [Consulta: 26 mar 2003].
Whitehead, H. 1983. Structure and stability of humpback whale groups off Newfoundland
Can. J. Zool. 61:1391-1397.
Whitehead, H., Moore, M. J. 1982. Distribution and movements of West Indian humpback
whales in winter. Can. J. Zoo. 60. 1982.
Würsig, B., Jefferson, T.A. 1990. Methods of Photo-identification for small cetaceans. Rep.
Int. Whal. Comm. (Special Issue 12). Pp.45-52.
Yumiseva, C. en prep. Uso de hábitat por parte de las ballenas jorobadas (Megaptera
novaeangliae) en el área marina del PNM. Tesis de Licenciatura. PUCE. En
elaboración.

185
8. FIGURAS
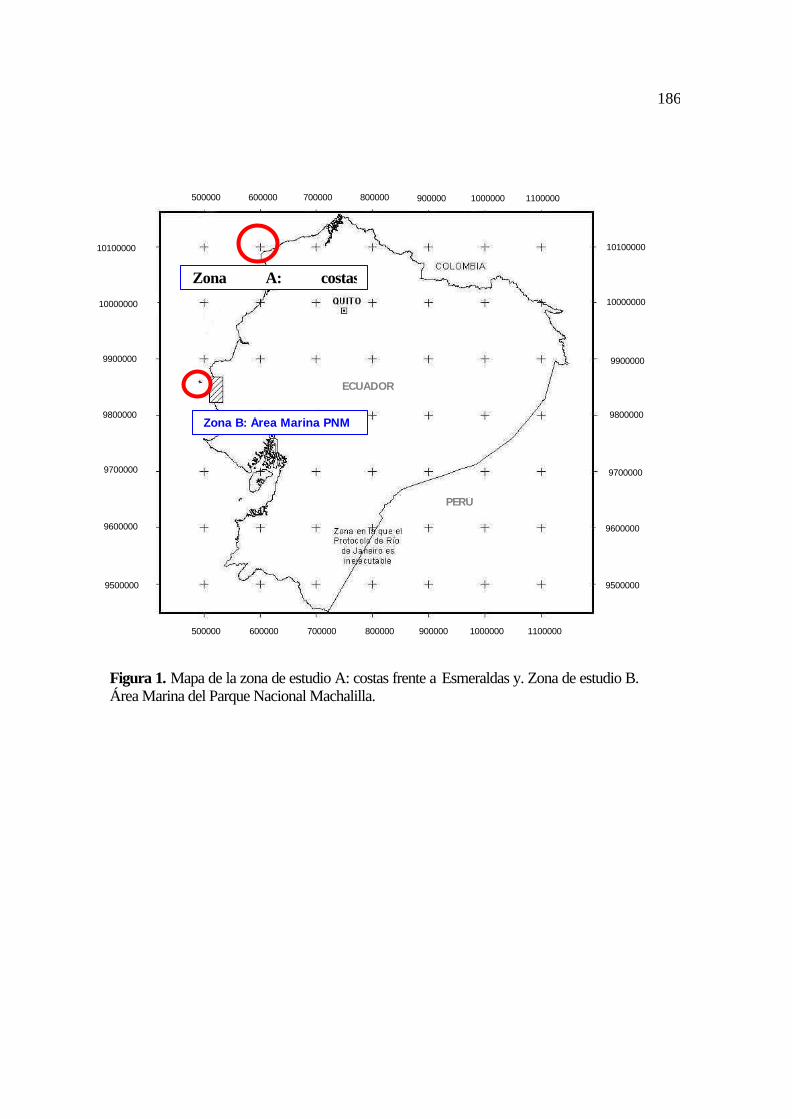
186
Figura 1. Mapa de la zona de estudio A: costas frente a Esmeraldas y. Zona de estudio B. Área Marina del Parque Nacional Machalilla.
Zona A: costas
Zona B: Área Marina PNM
500000 600000 700000 800000 900000 1000000 1100000
10100000
10000000
9900000
9800000
9700000
9600000
9500000
500000 600000 700000 800000 900000 1000000 1100000
10100000
10000000
9900000
9800000
9700000
9600000
9500000
ECUADOR
PERÚ
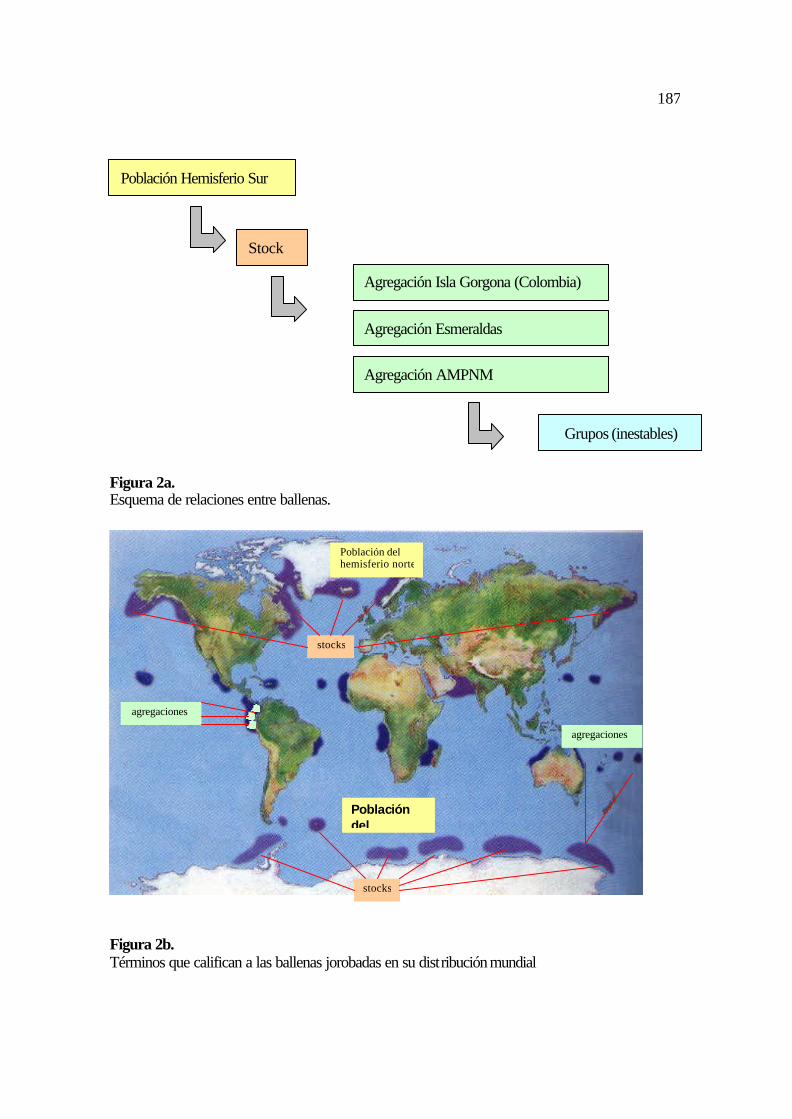
187
Figura 2a. Esquema de relaciones entre ballenas.
Figura 2b. Términos que califican a las ballenas jorobadas en su distribución mundial
Población del hemisferio norte
Población del
stocks
stocks
agregaciones
agregaciones
Grupos (inestables)
Agregación Isla Gorgona (Colombia)
Agregación Esmeraldas
Agregación AMPNM
Stock
Población Hemisferio Sur
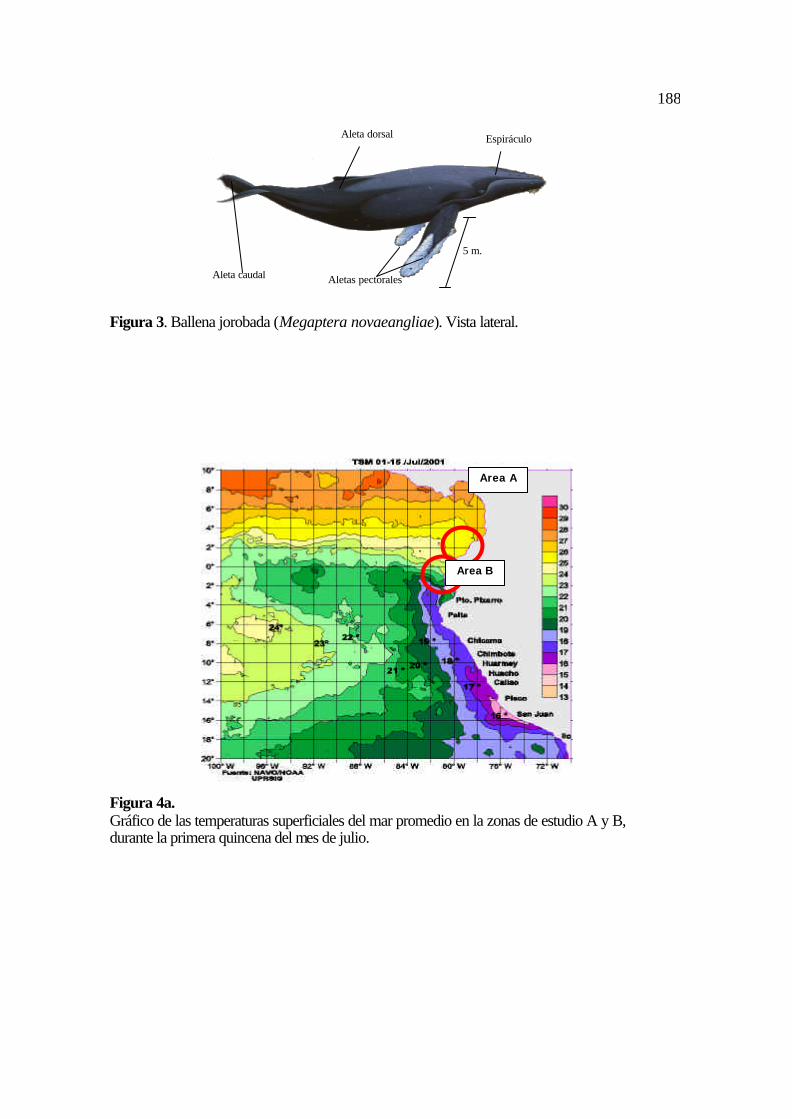
188
Figura 3. Ballena jorobada (Megaptera novaeangliae). Vista lateral.
Figura 4a. Gráfico de las temperaturas superficiales del mar promedio en la zonas de estudio A y B, durante la primera quincena del mes de julio.
Área A A
Área B
Aleta dorsal Espiráculo
Aletas pectoralesAleta caudal
5 m.
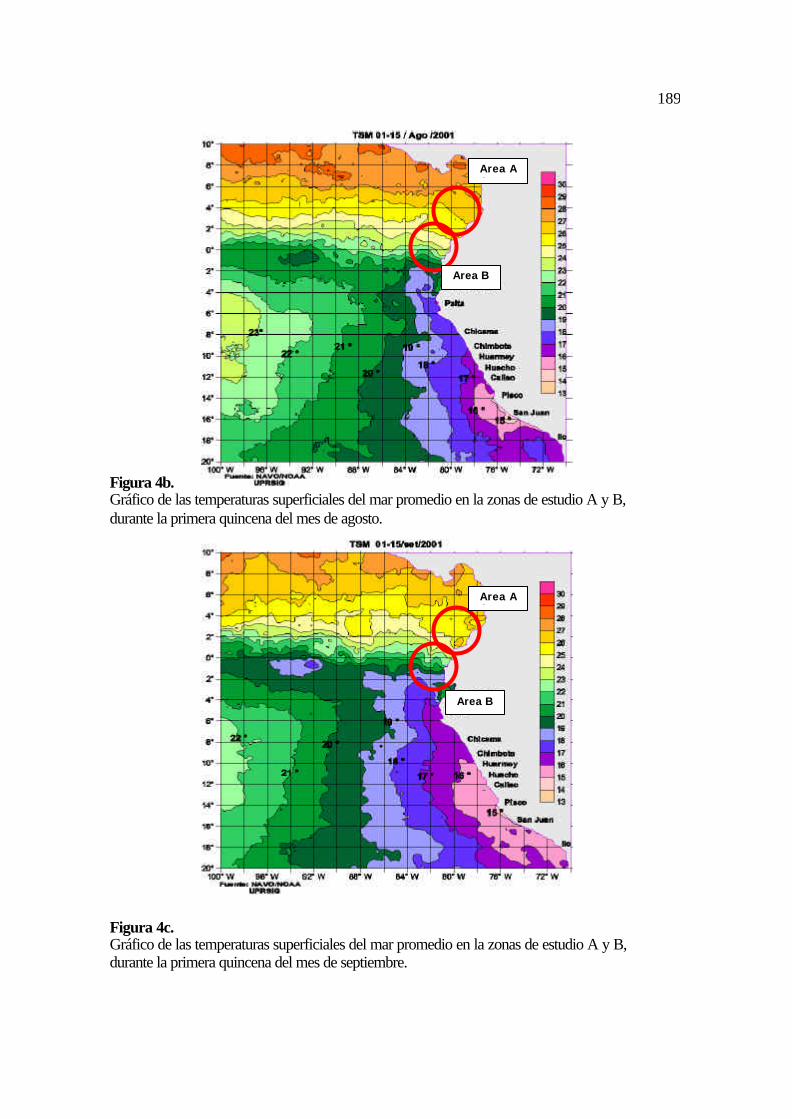
189
Figura 4b. Gráfico de las temperaturas superficiales del mar promedio en la zonas de estudio A y B, durante la primera quincena del mes de agosto.
Figura 4c. Gráfico de las temperaturas superficiales del mar promedio en la zonas de estudio A y B, durante la primera quincena del mes de septiembre.
Área A A
Área B
Área A A
Área B
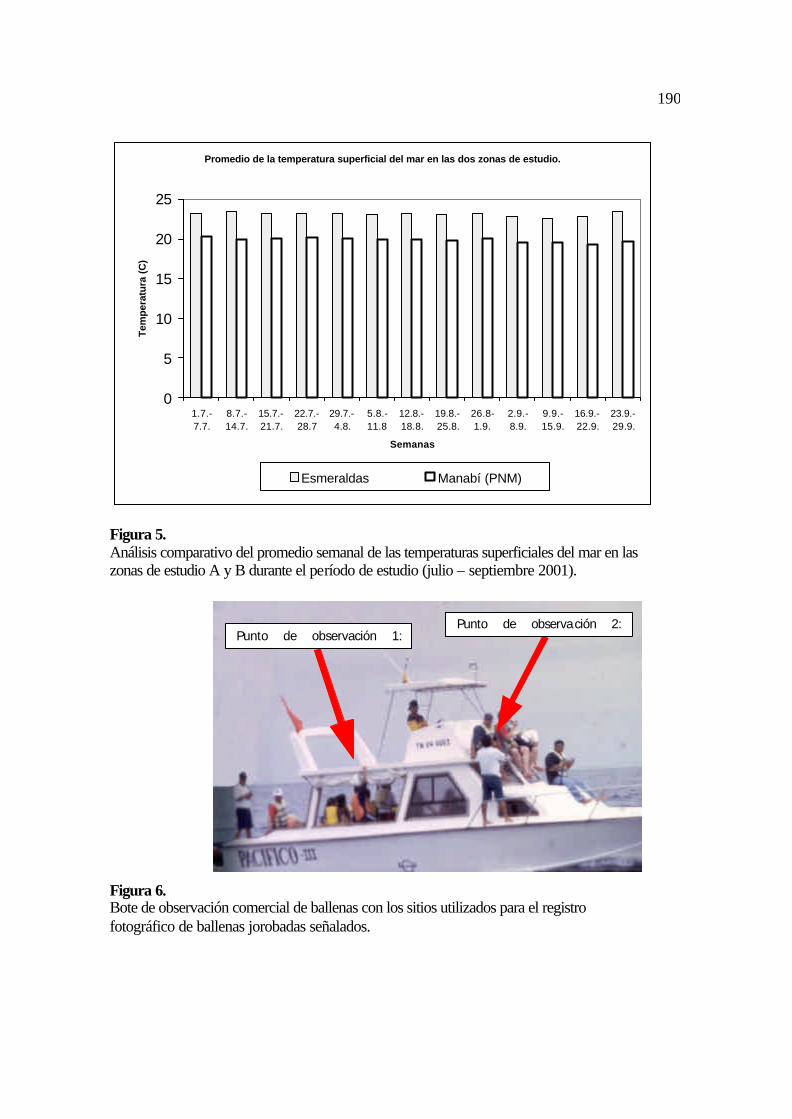
190
Promedio de la temperatura superficial del mar en las dos zonas de estudio.
0
5
10
15
20
25
1.7.-7.7.
8.7.-14.7.
15.7.-21.7.
22.7.-28.7
29.7.-4.8.
5.8.-11.8
12.8.-18.8.
19.8.-25.8.
26.8-1.9.
2.9.-8.9.
9.9.-15.9.
16.9.-22.9.
23.9.-29.9.
Semanas
Tem
per
atu
ra (
C)
Esmeraldas Manabí (PNM)
Figura 5. Análisis comparativo del promedio semanal de las temperaturas superficiales del mar en las zonas de estudio A y B durante el período de estudio (julio – septiembre 2001).
Figura 6. Bote de observación comercial de ballenas con los sitios utilizados para el registro fotográfico de ballenas jorobadas señalados.
Punto de observación 1: Punto de observación 2:
191
Figura 7. Esquema de registro para cobertura del cielo.
Figura 8 Patrones de coloración de aletas caudales y códigos utilizados (Mizroch et al., 1990).
192
Figura 9. Patrones de hendiduras medias de aletas caudales (Mizroch, et al., 1990).
0
0.51
1.5
2
2.5
3
3.5
4
25.6-8.7. 9.7-22.7. 23.7-5.8. 6.8-19.8 20.8-2.9. 3.9-17.9Períodos
Bal
len
as/h
ora
051015202530354045
esfu
erzo
en
ho
ras
po
r p
erío
do
Figura 10. Esfuerzo quincenal de muestreo en las dos zonas de estudio (A y B) durante la temporada 2001.
Ballenas/hora ESM esfuerzo ESM esfuerzo PNMBallenas/hora PNM
no identificada
1
2
3
4
5
6
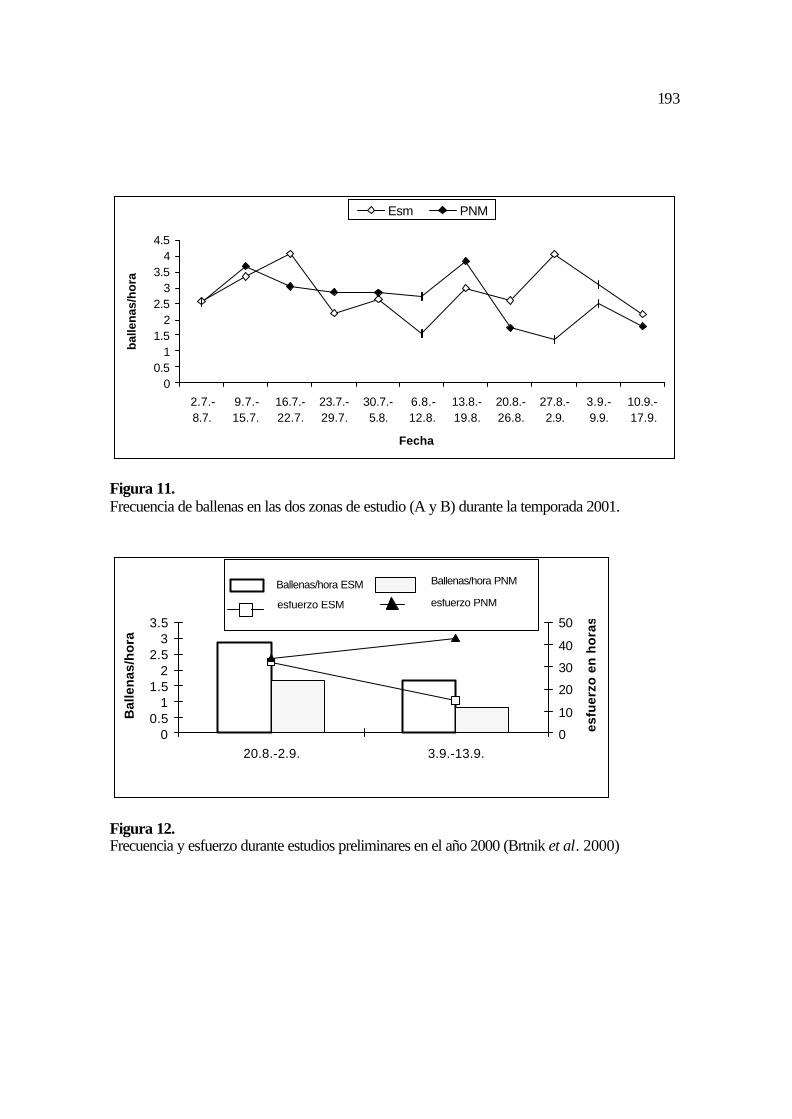
193
00.5
11.5
22.5
33.5
44.5
2.7.-8.7.
9.7.-15.7.
16.7.-22.7.
23.7.-29.7.
30.7.-5.8.
6.8.-12.8.
13.8.-19.8.
20.8.-26.8.
27.8.-2.9.
3.9.-9.9.
10.9.-17.9.
Fecha
bal
len
as/h
ora
Esm PNM
Figura 11. Frecuencia de ballenas en las dos zonas de estudio (A y B) durante la temporada 2001.
00.5
11.5
22.5
33.5
20.8.-2.9. 3.9.-13.9.
Bal
len
as/h
ora
0
10
20
30
40
50
esfu
erzo
en
ho
ras
Figura 12. Frecuencia y esfuerzo durante estudios preliminares en el año 2000 (Brtnik et al. 2000)
Ballenas/hora ESM
esfuerzo ESM esfuerzo PNM
Ballenas/hora PNM
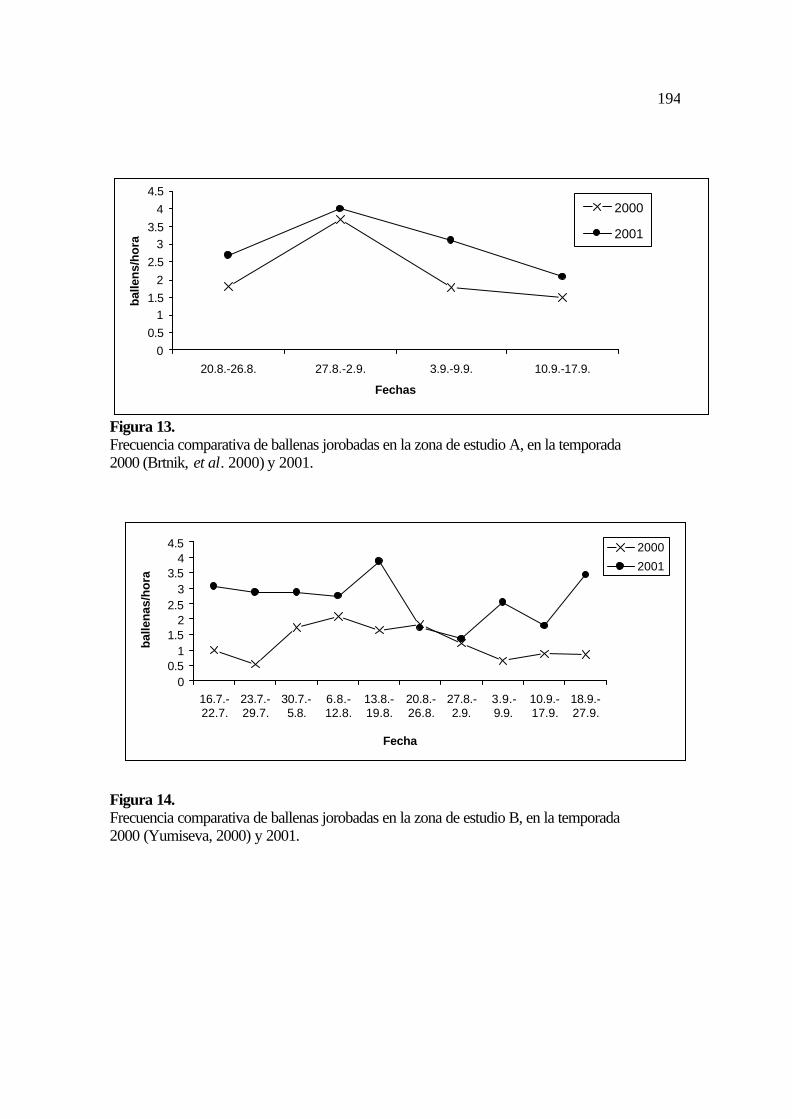
194
0
0.5
1
1.5
2
2.5
3
3.5
4
4.5
20.8.-26.8. 27.8.-2.9. 3.9.-9.9. 10.9.-17.9.
Fechas
bal
len
s/h
ora
2000
2001
Figura 13. Frecuencia comparativa de ballenas jorobadas en la zona de estudio A, en la temporada 2000 (Brtnik, et al. 2000) y 2001.
Figura 14. Frecuencia comparativa de ballenas jorobadas en la zona de estudio B, en la temporada 2000 (Yumiseva, 2000) y 2001.
00.5
11.5
22.5
33.5
44.5
16.7.-22.7.
23.7.-29.7.
30.7.-5.8.
6.8.-12.8.
13.8.-19.8.
20.8.-26.8.
27.8.-2.9.
3.9.-9.9.
10.9.-17.9.
18.9.-27.9.
Fecha
bal
len
as/h
ora
2000
2001
195
Figura 15. Esquema para contar individuos en grupos de delfines (Carwardine, 1995)
Figura 16. Esquema de observación del ataque de O. orca sobre una ballena jorobada.
Cría de orca
Ballena jorobada Orca adulta 1
Orca adulta 2
Orca adulta 3 bote

196
9. TABLAS
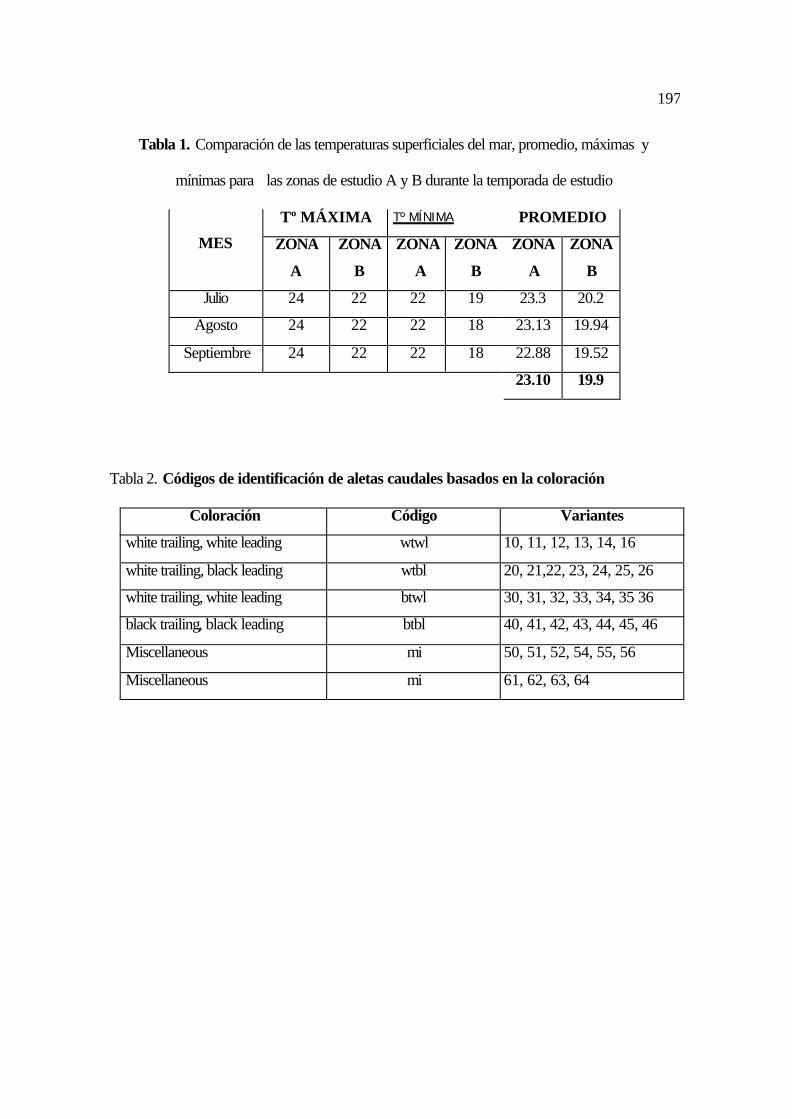
197
Tabla 1. Comparación de las temperaturas superficiales del mar, promedio, máximas y
mínimas para las zonas de estudio A y B durante la temporada de estudio
Tº MÁXIMA Tº MÍNIMA PROMEDIO
MES ZONA
A
ZONA
B
ZONA
A
ZONA
B
ZONA
A
ZONA
B
Julio 24 22 22 19 23.3 20.2
Agosto 24 22 22 18 23.13 19.94
Septiembre 24 22 22 18 22.88 19.52
23.10 19.9
Tabla 2. Códigos de identificación de aletas caudales basados en la coloración
Coloración Código Variantes
white trailing, white leading wtwl 10, 11, 12, 13, 14, 16
white trailing, black leading wtbl 20, 21,22, 23, 24, 25, 26
white trailing, white leading btwl 30, 31, 32, 33, 34, 35 36
black trailing, black leading btbl 40, 41, 42, 43, 44, 45, 46
Miscellaneous mi 50, 51, 52, 54, 55, 56
Miscellaneous mi 61, 62, 63, 64
198
Tabla 3. Códigos de marcas y señales de identificación para aletas caudales (Mizroch, et al . 1990)
Marca Código Descripción
Can´t tell * Sector de la aleta bajo el agua, fuera de marco o con un mal ángulo
Circles C Marcas circulares abiertas, blancas con centro negro o negras con centro blanco
Flecks F Pigmentos moteados, no parecen haber sido provocados por heridas o mordeduras
Hole H Hueco
Lines L Cualquier marca linear (rectas o curvas), cuyo largo sea al menos 4 veces su ancho
Missing M Un sector del animal ausente
Notch N Notch, nick o mordedura
Raches R Líneas paralelas, que parezcan causadas por mordeduras de depredadores
Spots S Marcas circulares, cerradas y no lineares
Distinctive X Cualquier marca distintiva que por si mismo pueda ayudar a la valoración del animal. Puede usarse una valoración con los otros códigos
Tabla 4. Código de clasificación de aletas caudales según la hendidura media, según Mizroch, et al ., (1990)
Tipo de hendidura media de las
aletas caudales
“v” aguda
“v” grave
redondeada
en cuadro
recta
no identificada
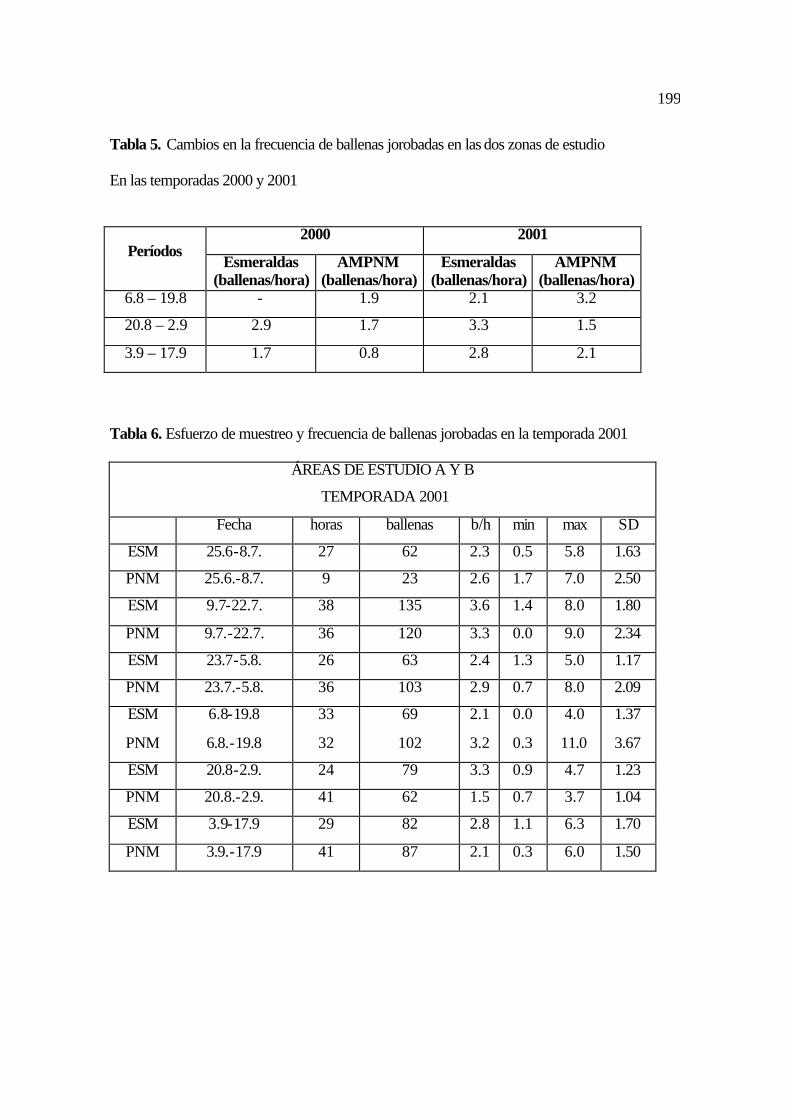
199
Tabla 5. Cambios en la frecuencia de ballenas jorobadas en las dos zonas de estudio
En las temporadas 2000 y 2001
2000 2001
Períodos Esmeraldas
(ballenas/hora) AMPNM
(ballenas/hora) Esmeraldas
(ballenas/hora) AMPNM
(ballenas/hora) 6.8 – 19.8 - 1.9 2.1 3.2
20.8 – 2.9 2.9 1.7 3.3 1.5
3.9 – 17.9 1.7 0.8 2.8 2.1
Tabla 6. Esfuerzo de muestreo y frecuencia de ballenas jorobadas en la temporada 2001
ÁREAS DE ESTUDIO A Y B
TEMPORADA 2001
Fecha horas ballenas b/h min max SD
ESM 25.6-8.7. 27 62 2.3 0.5 5.8 1.63
PNM 25.6.-8.7. 9 23 2.6 1.7 7.0 2.50
ESM 9.7-22.7. 38 135 3.6 1.4 8.0 1.80
PNM 9.7.-22.7. 36 120 3.3 0.0 9.0 2.34
ESM 23.7-5.8. 26 63 2.4 1.3 5.0 1.17
PNM 23.7.-5.8. 36 103 2.9 0.7 8.0 2.09
ESM 6.8-19.8 33 69 2.1 0.0 4.0 1.37
PNM 6.8.-19.8 32 102 3.2 0.3 11.0 3.67
ESM 20.8-2.9. 24 79 3.3 0.9 4.7 1.23
PNM 20.8.-2.9. 41 62 1.5 0.7 3.7 1.04
ESM 3.9-17.9 29 82 2.8 1.1 6.3 1.70
PNM 3.9.-17.9 41 87 2.1 0.3 6.0 1.50
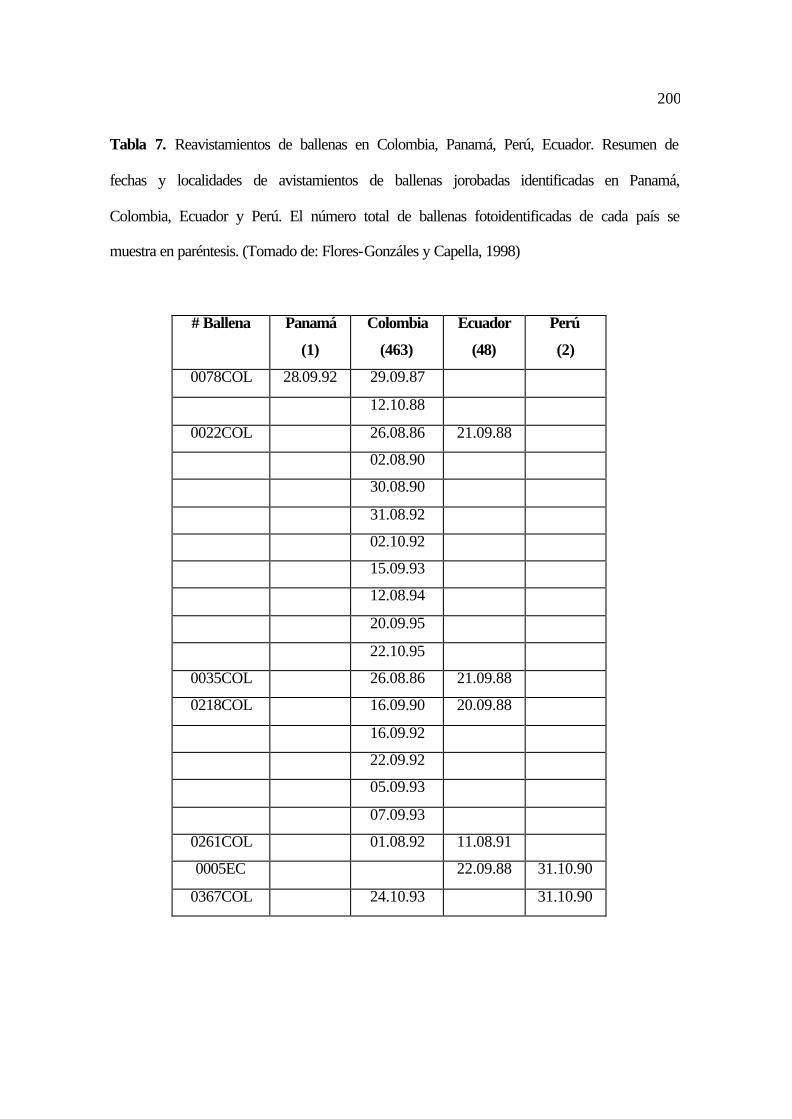
200
Tabla 7. Reavistamientos de ballenas en Colombia, Panamá, Perú, Ecuador. Resumen de
fechas y localidades de avistamientos de ballenas jorobadas identificadas en Panamá,
Colombia, Ecuador y Perú. El número total de ballenas fotoidentificadas de cada país se
muestra en paréntesis. (Tomado de: Flores-Gonzáles y Capella, 1998)
# Ballena Panamá
(1)
Colombia
(463)
Ecuador
(48)
Perú
(2)
0078COL 28.09.92 29.09.87
12.10.88
0022COL 26.08.86 21.09.88
02.08.90
30.08.90
31.08.92
02.10.92
15.09.93
12.08.94
20.09.95
22.10.95
0035COL 26.08.86 21.09.88
0218COL 16.09.90 20.09.88
16.09.92
22.09.92
05.09.93
07.09.93
0261COL 01.08.92 11.08.91
0005EC 22.09.88 31.10.90
0367COL 24.10.93 31.10.90
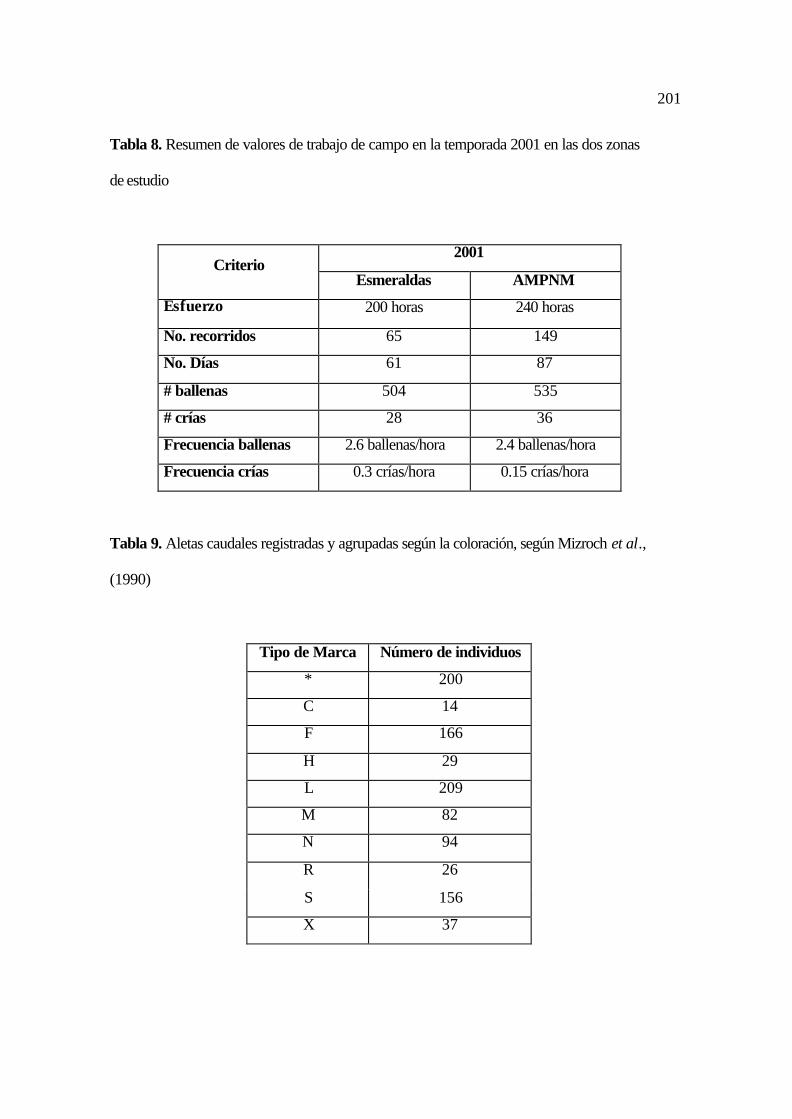
201
Tabla 8. Resumen de valores de trabajo de campo en la temporada 2001 en las dos zonas
de estudio
2001 Criterio
Esmeraldas AMPNM
Esfuerzo 200 horas 240 horas
No. recorridos 65 149
No. Días 61 87
# ballenas 504 535
# crías 28 36
Frecuencia ballenas 2.6 ballenas/hora 2.4 ballenas/hora
Frecuencia crías 0.3 crías/hora 0.15 crías/hora
Tabla 9. Aletas caudales registradas y agrupadas según la coloración, según Mizroch et al.,
(1990)
Tipo de Marca Número de individuos
* 200
C 14
F 166
H 29
L 209
M 82
N 94
R 26
S 156
X 37
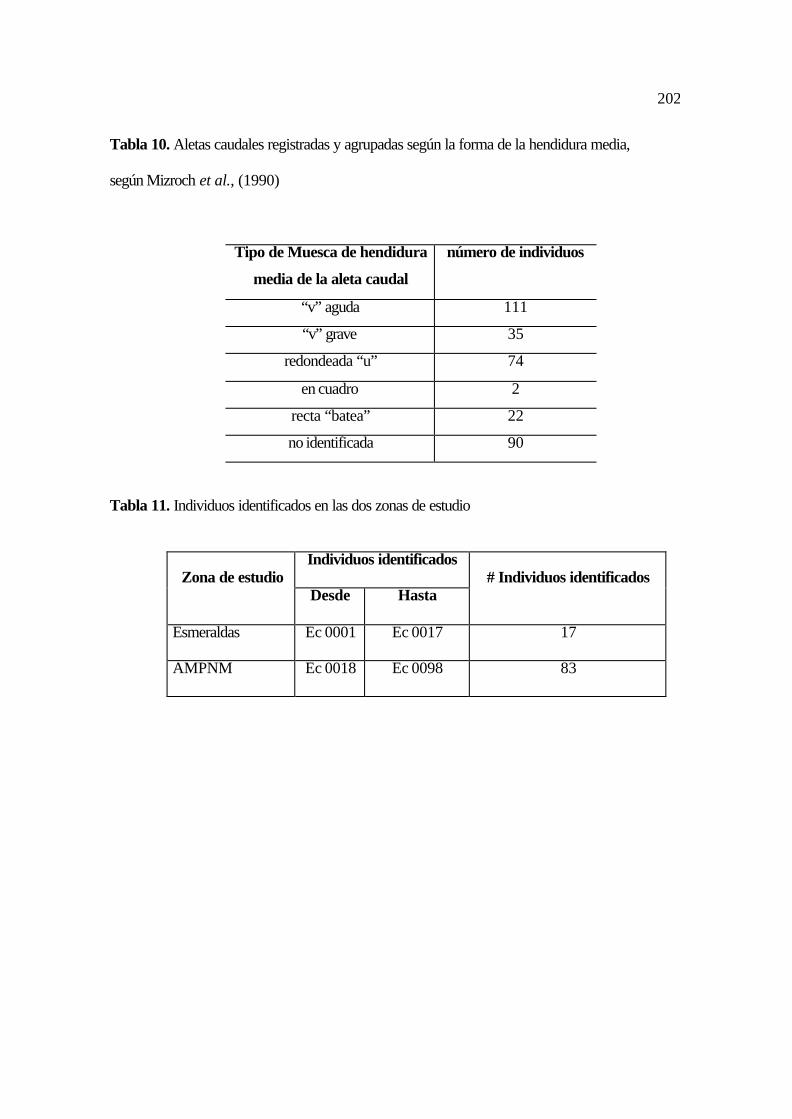
202
Tabla 10. Aletas caudales registradas y agrupadas según la forma de la hendidura media,
según Mizroch et al., (1990)
Tipo de Muesca de hendidura
media de la aleta caudal
número de individuos
“v” aguda 111
“v” grave 35
redondeada “u” 74
en cuadro 2
recta “batea” 22
no identificada 90
Tabla 11. Individuos identificados en las dos zonas de estudio
Individuos identificados Zona de estudio
Desde Hasta # Individuos identificados
Esmeraldas Ec 0001 Ec 0017 17
AMPNM Ec 0018 Ec 0098 83

203
10. ANEXOS
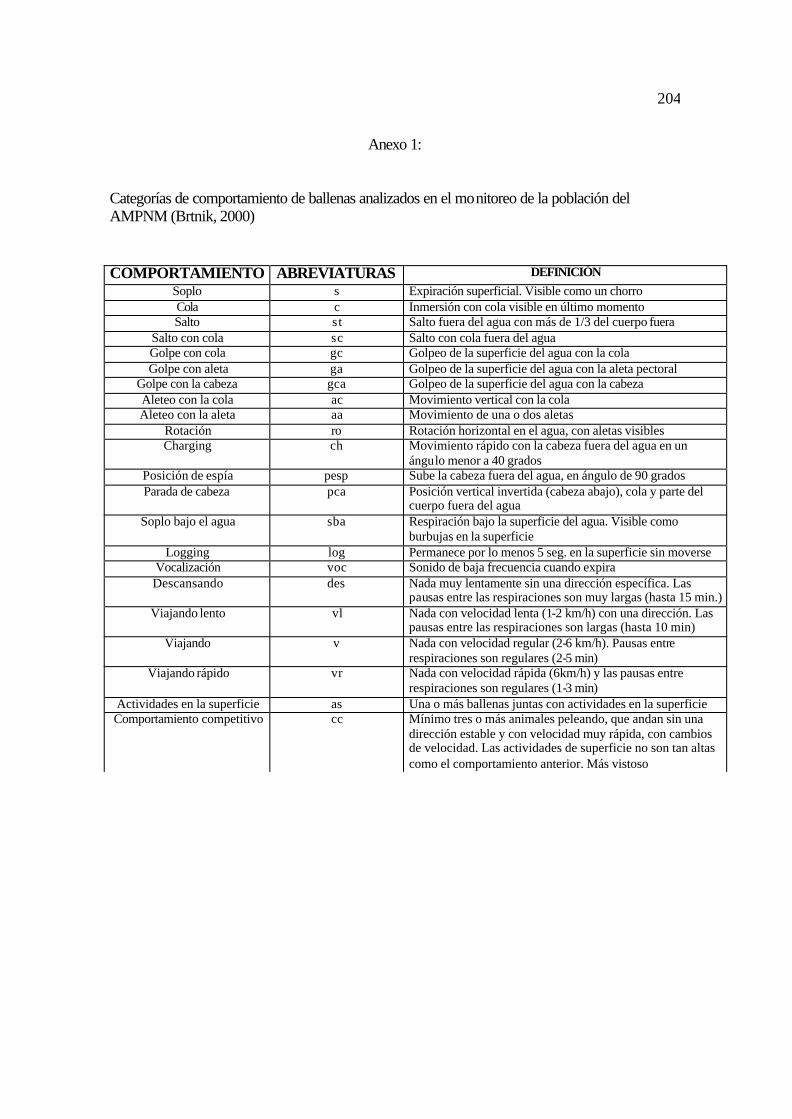
204
Anexo 1:
Categorías de comportamiento de ballenas analizados en el monitoreo de la población del AMPNM (Brtnik, 2000)
COMPORTAMIENTO ABREVIATURAS DEFINICIÓN Soplo s Expiración superficial. Visible como un chorro Cola c Inmersión con cola visible en último momento Salto st Salto fuera del agua con más de 1/3 del cuerpo fuera
Salto con cola sc Salto con cola fuera del agua Golpe con cola gc Golpeo de la superficie del agua con la cola Golpe con aleta ga Golpeo de la superficie del agua con la aleta pectoral
Golpe con la cabeza gca Golpeo de la superficie del agua con la cabeza Aleteo con la cola ac Movimiento vertical con la cola Aleteo con la aleta aa Movimiento de una o dos aletas
Rotación ro Rotación horizontal en el agua, con aletas visibles Charging ch Movimiento rápido con la cabeza fuera del agua en un
ángulo menor a 40 grados Posición de espía pesp Sube la cabeza fuera del agua, en ángulo de 90 grados Parada de cabeza pca Posición vertical invertida (cabeza abajo), cola y parte del
cuerpo fuera del agua Soplo bajo el agua sba Respiración bajo la superficie del agua. Visible como
burbujas en la superficie Logging log Permanece por lo menos 5 seg. en la superficie sin moverse
Vocalización voc Sonido de baja frecuencia cuando expira Descansando des Nada muy lentamente sin una dirección específica. Las
pausas entre las respiraciones son muy largas (hasta 15 min.) Viajando lento vl Nada con velocidad lenta (1-2 km/h) con una dirección. Las
pausas entre las respiraciones son largas (hasta 10 min) Viajando v Nada con velocidad regular (2-6 km/h). Pausas entre
respiraciones son regulares (2-5 min) Viajando rápido vr Nada con velocidad rápida (6km/h) y las pausas entre
respiraciones son regulares (1-3 min) Actividades en la superficie as Una o más ballenas juntas con actividades en la superficie Comportamiento competitivo cc Mínimo tres o más animales peleando, que andan sin una
dirección estable y con velocidad muy rápida, con cambios de velocidad. Las actividades de superficie no son tan altas como el comportamiento anterior. Más vistoso

205
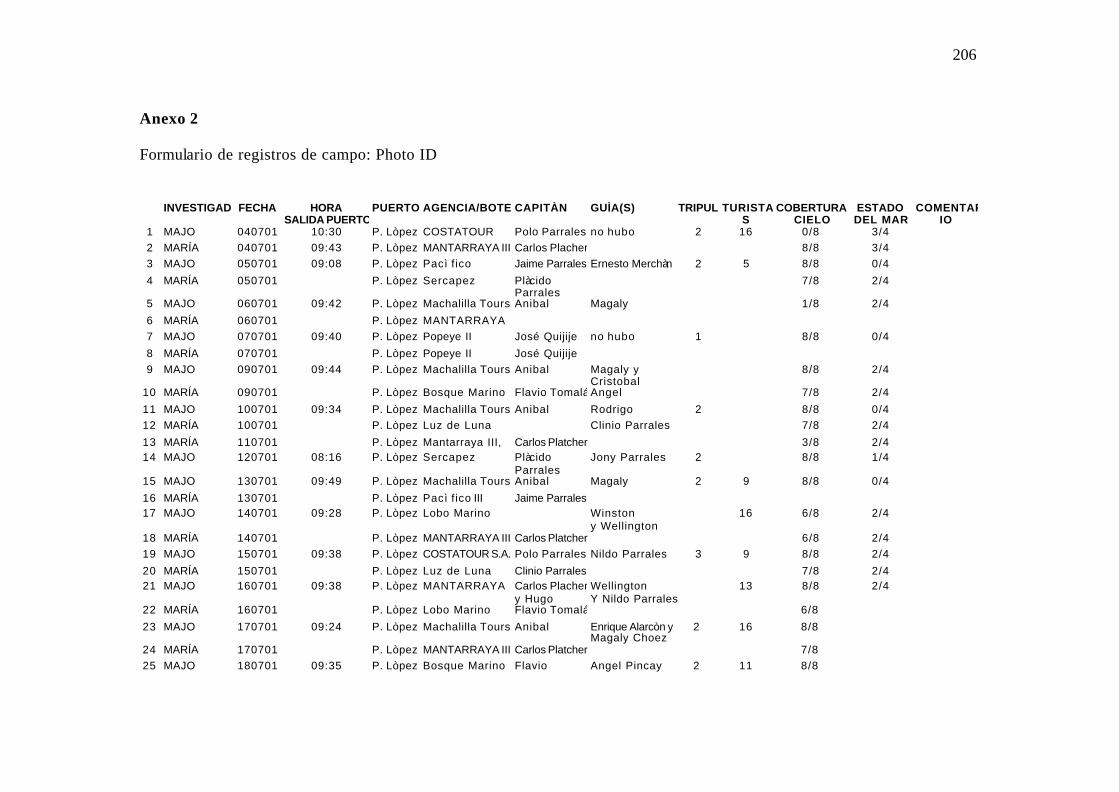
206
Anexo 2 Formulario de registros de campo: Photo ID
INVESTIGAD FECHA HORA SALIDA PUERTO
PUERTO AGENCIA/BOTE CAPITÀN GUÌA(S) TRIPUL TURISTAS
COBERTURA CIELO
ESTADO DEL MAR
COMENTARIO
1 MAJO 040701 10:30 P. Lòpez COSTATOUR Polo Parrales no hubo 2 16 0/8 3/4
2 MARÍA 040701 09:43 P. Lòpez MANTARRAYA III Carlos Placher 8/8 3/4
3 MAJO 050701 09:08 P. Lòpez Pacìfico Jaime Parrales Ernesto Merchàn 2 5 8/8 0/4
4 MARÍA 050701 P. Lòpez Sercapez Plàcido Parrales
7/8 2/4
5 MAJO 060701 09:42 P. Lòpez Machalilla Tours Anibal Magaly 1/8 2/4
6 MARÍA 060701 P. Lòpez MANTARRAYA
7 MAJO 070701 09:40 P. Lòpez Popeye II José Quijije no hubo 1 8/8 0/4
8 MARÍA 070701 P. Lòpez Popeye II José Quijije
9 MAJO 090701 09:44 P. Lòpez Machalilla Tours Anibal Magaly y Cristobal
8/8 2/4
10 MARÍA 090701 P. Lòpez Bosque Marino Flavio Tomalá Angel 7/8 2/4
11 MAJO 100701 09:34 P. Lòpez Machalilla Tours Anibal Rodrigo 2 8/8 0/4
12 MARÍA 100701 P. Lòpez Luz de Luna Clinio Parrales 7/8 2/4
13 MARÍA 110701 P. Lòpez Mantarraya III, Carlos Platcher 3/8 2/4 14 MAJO 120701 08:16 P. Lòpez Sercapez Plàcido
Parrales Jony Parrales 2 8/8 1/4
15 MAJO 130701 09:49 P. Lòpez Machalilla Tours Anibal Magaly 2 9 8/8 0/4
16 MARÍA 130701 P. Lòpez Pacìfico III Jaime Parrales 17 MAJO 140701 09:28 P. Lòpez Lobo Marino Winston
y Wellington 16 6/8 2/4
18 MARÍA 140701 P. Lòpez MANTARRAYA III Carlos Platcher 6/8 2/4
19 MAJO 150701 09:38 P. Lòpez COSTATOUR S.A. Polo Parrales Nildo Parrales 3 9 8/8 2/4
20 MARÍA 150701 P. Lòpez Luz de Luna Clinio Parrales 7/8 2/4 21 MAJO 160701 09:38 P. Lòpez MANTARRAYA Carlos Placher
y Hugo Wellington Y Nildo Parrales
13 8/8 2/4
22 MARÍA 160701 P. Lòpez Lobo Marino Flavio Tomalá 6/8
23 MAJO 170701 09:24 P. Lòpez Machalilla Tours Anibal Enrique Alarcòn y Magaly Choez
2 16 8/8
24 MARÍA 170701 P. Lòpez MANTARRAYA III Carlos Platcher 7/8
25 MAJO 180701 09:35 P. Lòpez Bosque Marino Flavio Angel Pincay 2 11 8/8
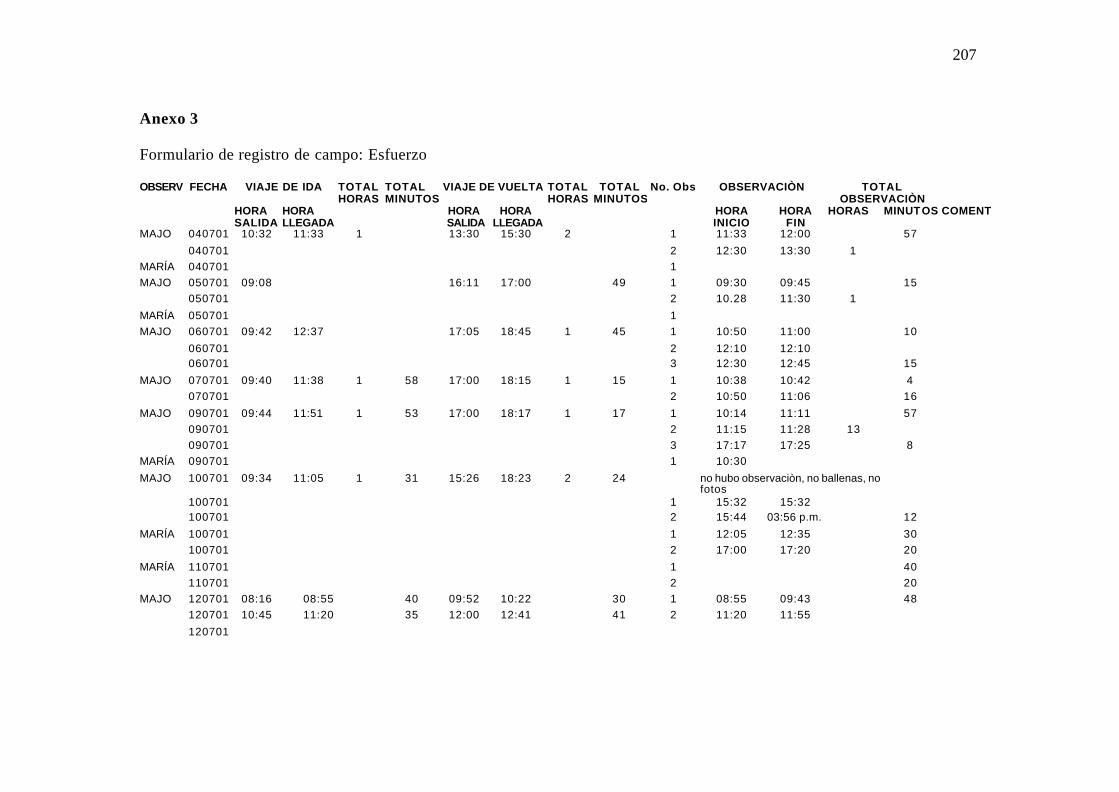
207
Anexo 3 Formulario de registro de campo: Esfuerzo OBSERV FECHA VIAJE DE IDA TOTAL
HORAS TOTAL MINUTOS
VIAJE DE VUELTA TOTAL HORAS
TOTAL MINUTOS
No. Obs OBSERVACIÒN TOTAL OBSERVACIÒN
HORA SALIDA
HORA LLEGADA
HORA SALIDA
HORA LLEGADA
HORA INICIO
HORA FIN
HORAS MINUTOS COMENT
MAJO 040701 10:32 11:33 1 13:30 15:30 2 1 11:33 12:00 57
040701 2 12:30 13:30 1
MARÍA 040701 1
MAJO 050701 09:08 16:11 17:00 49 1 09:30 09:45 15
050701 2 10.28 11:30 1
MARÍA 050701 1
MAJO 060701 09:42 12:37 17:05 18:45 1 45 1 10:50 11:00 10
060701 2 12:10 12:10 060701 3 12:30 12:45 15
MAJO 070701 09:40 11:38 1 58 17:00 18:15 1 15 1 10:38 10:42 4
070701 2 10:50 11:06 16
MAJO 090701 09:44 11:51 1 53 17:00 18:17 1 17 1 10:14 11:11 57
090701 2 11:15 11:28 13
090701 3 17:17 17:25 8
MARÍA 090701 1 10:30
MAJO 100701 09:34 11:05 1 31 15:26 18:23 2 24 no hubo observaciòn, no ballenas, no fotos
100701 1 15:32 15:32 100701 2 15:44 03:56 p.m. 12
MARÍA 100701 1 12:05 12:35 30
100701 2 17:00 17:20 20
MARÍA 110701 1 40
110701 2 20
MAJO 120701 08:16 08:55 40 09:52 10:22 30 1 08:55 09:43 48
120701 10:45 11:20 35 12:00 12:41 41 2 11:20 11:55
120701
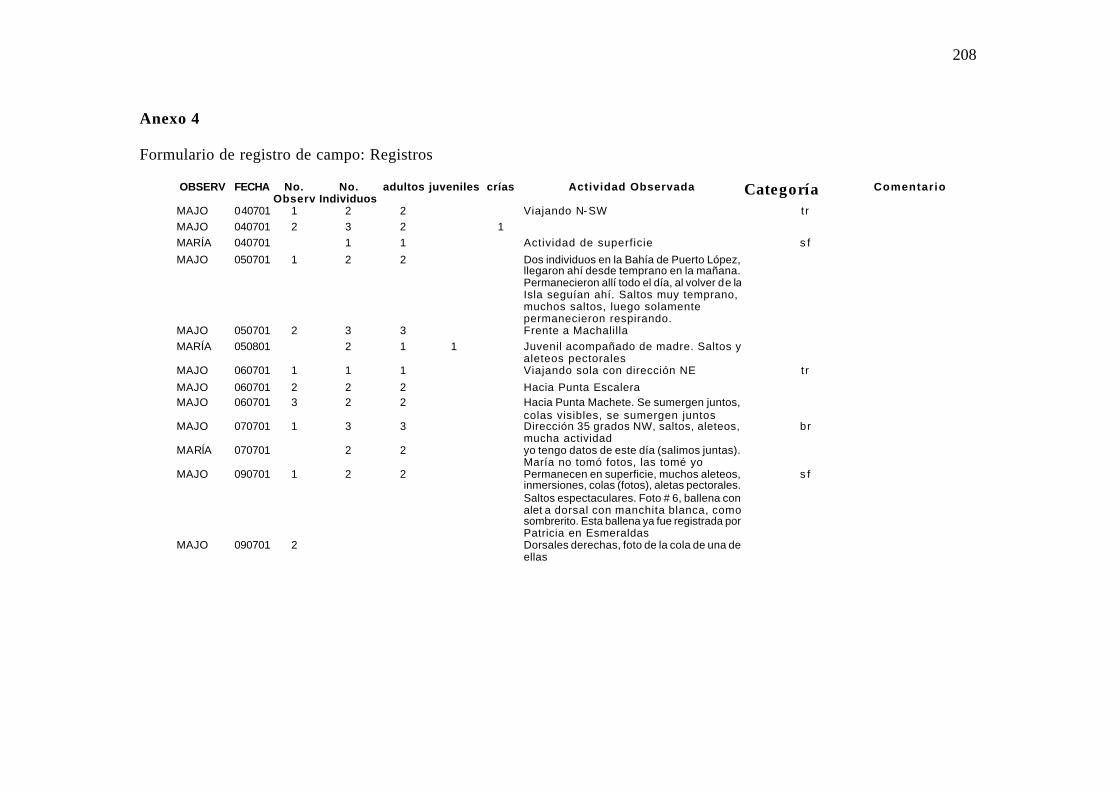
208
Anexo 4 Formulario de registro de campo: Registros
OBSERV FECHA No. Observ
No. Individuos
adultos juveniles crías Actividad Observada Categoría Comentario
MAJO 040701 1 2 2 Viajando N-SW tr
MAJO 040701 2 3 2 1
MARÍA 040701 1 1 Actividad de superficie s f
MAJO 050701 1 2 2 Dos individuos en la Bahía de Puerto López, llegaron ahí desde temprano en la mañana. Permanecieron allí todo el día, al volver de la Isla seguían ahí. Saltos muy temprano, muchos saltos, luego solamente permanecieron respirando.
MAJO 050701 2 3 3 Frente a Machalilla
MARÍA 050801 2 1 1 Juvenil acompañado de madre. Saltos y aleteos pectorales
MAJO 060701 1 1 1 Viajando sola con dirección NE tr
MAJO 060701 2 2 2 Hacia Punta Escalera MAJO 060701 3 2 2 Hacia Punta Machete. Se sumergen juntos,
colas visibles, se sumergen juntos
MAJO 070701 1 3 3 Dirección 35 grados NW, saltos, aleteos, mucha actividad
br
MARÍA 070701 2 2 yo tengo datos de este día (salimos juntas). María no tomó fotos, las tomé yo
MAJO 090701 1 2 2 Permanecen en superficie, muchos aleteos, inmersiones, colas (fotos), aletas pectorales. Saltos espectaculares. Foto # 6, ballena con alet a dorsal con manchita blanca, como sombrerito. Esta ballena ya fue registrada por Patricia en Esmeraldas
s f
MAJO 090701 2 Dorsales derechas, foto de la cola de una de ellas
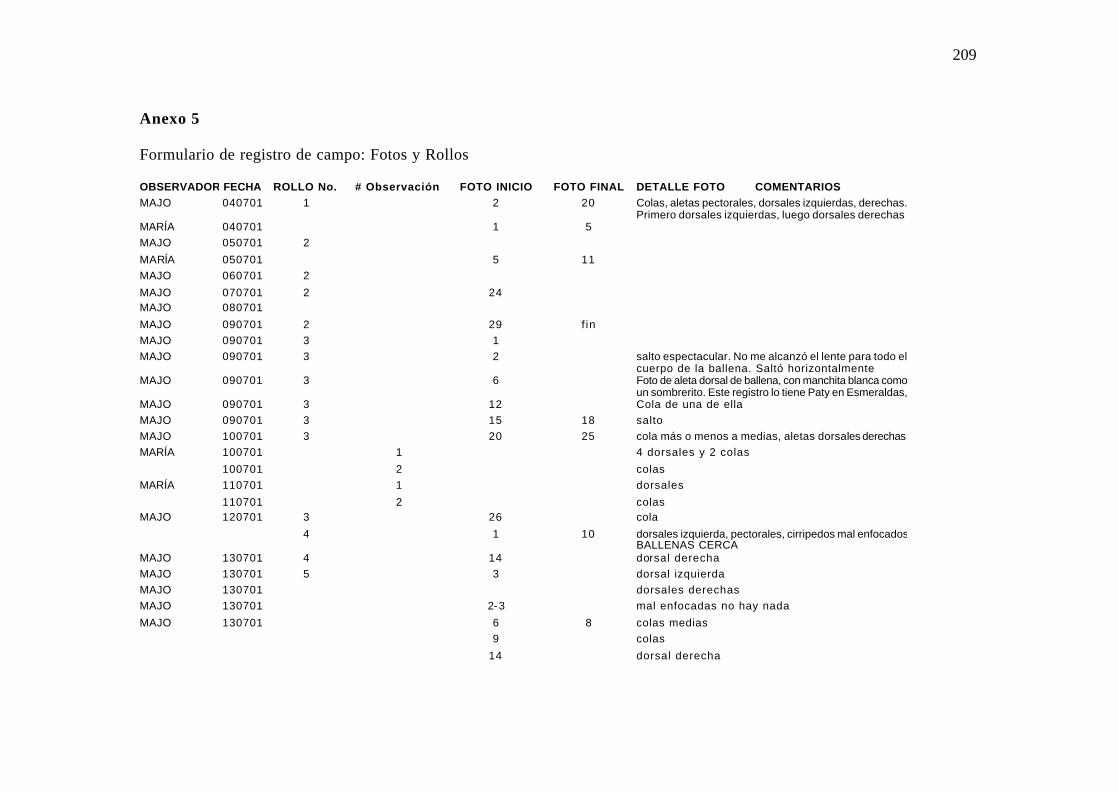
209
Anexo 5 Formulario de registro de campo: Fotos y Rollos OBSERVADOR FECHA ROLLO No. # Observación FOTO INICIO FOTO FINAL DETALLE FOTO COMENTARIOS MAJO 040701 1 2 20 Colas, aletas pectorales, dorsales izquierdas, derechas.
Primero dorsales izquierdas, luego dorsales derechas MARÍA 040701 1 5
MAJO 050701 2
MARÍA 050701 5 11
MAJO 060701 2
MAJO 070701 2 24 MAJO 080701
MAJO 090701 2 29 f in
MAJO 090701 3 1
MAJO 090701 3 2 salto espectacular. No me alcanzó el lente para todo el cuerpo de la ballena. Saltó horizontalmente
MAJO 090701 3 6 Foto de aleta dorsal de ballena, con manchita blanca como un sombrerito. Este registro lo tiene Paty en Esmeraldas,
MAJO 090701 3 12 Cola de una de ella
MAJO 090701 3 15 18 salto
MAJO 100701 3 20 25 cola más o menos a medias, aletas dorsales derechas
MARÍA 100701 1 4 dorsales y 2 colas
100701 2 colas
MARÍA 110701 1 dorsales
110701 2 colas MAJO 120701 3 26 cola
4 1 10 dorsales izquierda, pectorales, cirripedos mal enfocados BALLENAS CERCA
MAJO 130701 4 14 dorsal derecha
MAJO 130701 5 3 dorsal izquierda
MAJO 130701 dorsales derechas
MAJO 130701 2-3 mal enfocadas no hay nada
MAJO 130701 6 8 colas medias
9 colas
14 dorsal derecha

210
Anexo 6 Fotos de recaptura (2000 y 2001)
Foto por: D. Rosero Julio, 2000
Foto por: M.J. Barragán Agosto 2001

211
Anexo 7: Interacciones con otros cetáceos (Orcinus orca) Fotos: María José Barragán P.
Aleta dorsal de orca macho
Aleta dorsal y cabeza de orca macho
Cabeza y parte de aleta dorsal de orca macho
Grasa subdérmica de cría de ballena jorobada muerta por ataque de orcas


















