

PAPEL DE LA FAUNA EDÁFICA EN EL FUNCIONAMIENTO DE LOS ...
205
PAPEL DE LA FAUNA EDÁFICA EN EL FUNCIONAMIENTO DE LOS ECOSISTEMAS AGRÍCOLAS TESE DE DOUTORAMENTO JESÚS CASTRO LÓPEZ 2017 PAPEL DE LA FAUNA EDÁFICA EN EL FUNCIONAMIENTO DE LOS ECOSISTEMAS AGRÍCOLAS Jesús Castro López 2017 Tese de Doutoramento
Transcript of PAPEL DE LA FAUNA EDÁFICA EN EL FUNCIONAMIENTO DE LOS ...

PA
PE
L D
E L
A F
AU
NA
ED
ÁF
ICA
EN
EL
FU
NC
ION
AM
IEN
TO
DE
LO
S
EC
OSIS
TE
MA
S A
GR
ÍCO
LA
S
T
ESE
DE
DO
UT
OR
AM
EN
TO
JE
SÚS C
AST
RO
LÓ
PE
Z
2017
PAPEL DE LA FAUNA EDÁFICA EN EL FUNCIONAMIENTO
DE LOS ECOSISTEMAS AGRÍCOLAS
Jesús Castro López 2017 Tese de Doutoramento

Jesús Castro López
TESE DE DOUTORAMENTO
PAPEL DE LA FAUNA EDÁFICA EN EL FUNCIONAMIENTO DE LOS ECOSISTEMAS AGRÍCOLAS
Dirigida por los doctores:
María Jesús Iglesias Briones y Pedro Pablo Gallego Veigas
Año: 2017


DRA. MARÍA JESÚS IGLESIAS BRIONES y DR. PEDRO PABLO GALLEGO VEIGAS
HACEN CONSTAR que el presente trabajo, titulado Papel de la fauna edáfica en el funcionamiento de los ecosistemas agrícolas ue p ese ta Jesús Castro López para la obtención del título de Doctor fue realizado bajo su dirección en el Programa de Doctorado Biodiversidad y Ecosistemas .
Vigo, 20 de abril de 2017
Directores de la tesis doctoral
Fdo. Dra. María Jesús Iglesias Briones Fdo. Dr. Pedro Pablo Gallego Veigas


Este trabajo fue financiado por la Red de Investigación titulada REDUSO: Red de Uso Sostenible de los Recursos Naturales y
Agroalimentarios , subvencionada por la Xunta de Galicia
(CN 2012/108)


“Sin importar que tan urbana sea nuestra vida, nuestros cuerpos viven de la agricultura; nosotros venimos de la Tierra y
retornaremos a ella, y es así que existimos en la agricultura tanto como existimos en nuestra propia carne.”
Wendell Berry
Comunicaciones en Congresos Internacionales:
Autores: Castro J, Barciela A, Barreal ME, Gallego PP y Briones MJI Título: Earthworm ecological groupings as soil quality indicators of agricultural management Congreso: 10th International Symposium on Earthworm Ecology (I.S.E.E. 10) Lugar celebración: Athens, Georgia (USA) Fecha: 22-27 junio 2014 Autores: Castro J, Lago MCF, Barreal ME, Gallego PP y Briones MJI Título: Effects of agricultural practices on soil fauna communities in kiwifruit plantations Congreso: 8thInternational Symposium on Kiwifruit Lugar celebración: Dujiangyan city, Chengdu (China) Fecha: 18-22 septiembre 2014 Autores: Castro J, Barreal ME, Gallego PP y Briones MJI. Título: Structure and diversity of soil fauna communities under contrastingbiogeographical regions and agricultural managements Congreso: XVII International Colloquium on Soil Zoology and XIV International Colloquium on Apterygota Lugar celebración: Nara (Japón) Fecha: 22-26 de agosto de 2016
Publicaciones derivadas de Congresos:
Castro J, Lago MCF, Briones MJI, Gallego PP y Barreal ME. 2015. Effects of agricultural practices
on soil fauna communities in kiwifruit plantations. Acta Horticulturae, 1096, 267-273.
DOI: 10.17660/ActaHortic.2015.1096.29


AGRADECIMIENTOS
Dicen que a veces acertamos y otras muchas nos equivocamos, pero que siempre aprendemos. Este documento es un buen ejemplo de este refrán pues durante estos últimos cuatro años y medio no sólo he aprendido de fauna edáfica, ecología, agricultura, estadística multivariante y análisis isotópicos, también he aprendido sobre la confianza, paciencia, perseverancia y relatividad pues las cosas nunca son blancas o negras. Todo esto lo he aprendido a base de más de un tropiezo, pero a pesar de ello mis directores, Pedro Pablo y María Jesús, han continuado confiando en mí y en este proyecto, mejorándolo y mejorándome a cada paso con sus diferentes pero complementarios, puntos de vista.
Esther se merece una mención especial. Ha estado desde antes del primer muestreo hasta el fin poniendo las bases de la gestión de datos y muchas más cosas que no vienen al caso. Parece que al final conseguiste mejorar incluso al Jesús 2.0 y convertirlo en Jesús 3.0. Sólo puedo desearte que sigas teniendo Largos días y placenteras noches, sai.
Me gustaría agradecer a mis primeras guías personales en este complejo mundo de la fauna edáfica, Noela y Carmen, cuya ayuda con la identificación de las primeras muestras fue fundamental para el resto del camino.
Recientemente se ha publicado un artículo que demuestra que los estudiantes de doctorado tenemos una mayor tendencia hacia los desórdenes psiquiátricos comunes. Un buen ejemplo de ello es la fauna del CUVI, tanto los que pasaron por allí (Paula, Dani, Vero, Bego, Valeria, Luisa, Sara, Andrea y Pelayo) como los que aún buscan su camisa blanca de mangas largas (Pascual, Lolo, Gusu, Radhia, María, Nuria, Miguel) y los nuevos fichajes (Pascual gigante, Raquel y Hamza). A todos ellos me gustaría agradecerles su empatía y todos los buenos ratos compartidos en el comedor, en la cafeta y en los laboratorios.
Hay que agradecer a los propietarios y propietarias de las fincas que nos permitieran robarles las lombrices y llenarles los cultivos de agujeros.
La ciencia y el conocimiento deberían de estar al alcance de todo el mundo, pero tristemente no es así. Me gustaría dar las gracias a cierta web sin la cual no podría haber tenido acceso al 97% de los trabajos aquí referenciados (otros muchos leídos pero no referenciados).
Esta Tesis, la Tesina y el Máster no se podrían haber realizado sin el hueco para poder dormir que Marlene, Teresa y su familia me han regalado. Muchas Gracias.
Quisiera dar las gracias a Carpio puesto que gracias a los trabajos paralelos he podido complementar y mejorar este trabajo. ¿Quién nos diría que llegaríamos hasta aquí cuando empezamos con aquel trabajo con tus gallinas para Francis?
El apoyo y ánimos de la familia Garnil-García, en forma de tuppers llenos de delicatesen gallegas de Clara, han sido fundamentales para mí. Me hacéis sentir que tengo una familia aun estando a 1000 kilómetros de la mía y me habéis enseñado que la familia no siempre tiene lazos de sangre. Muchas gracias por todo.
Esta Tesis, y mi vida entera, se la tengo que agradecer a mis padres; Jesús y Mª del Carmen, y a mi hermana Helena. Siempre habéis creído en mí y en mis capacidades, me habéis apoyado hasta lo i posi le e ha éis dado el e pujó Dos huevos y al toro pa a la Tesi a, Para adelante, como los de Alicante pa a la Tesis) cuando más negro veía todo, sabiendo, como sólo saben los padres, hasta dónde es capaz de llegar uno. Me habéis apoyado y ayudado incluso cuando ello ha supuesto separarme de vosotros. No se le puede pedir más a una familia de lo que me habéis dado sin pedirlo.

Con la Tesina tampoco supe cómo empezar este párrafo dedicado a ti, Sandra. Quisiera saber agradecerte de alguna manera mejor que un párrafo todo lo que has hecho y haces por mí, pero no sé cómo hacerlo. Me haces mejor persona y estas siempre a mi lado. A pesar de aguantar mis cambios de humor y mi mal genio, siempre tienes una sonrisa y una recetilla para mejorar el día. Hoy es un mínimo ejemplo; llevo casi 10 horas sentado delante del PC en tu casa y me has traído, sonriendo como siempre, un yogurt helado de fresa y plátano y me has preguntado cuanto pan quiero para cenar. Ahora mismo no tenemos mucho, pero sinceramente, creo que lo tenemos todo. Me da la i p esió de ue se ueda o to, pe o… g a ias. G a ias “a d a, g a ias po todo.

ÍNDICE
RESUMEN ..................................................................................................................................... 1
SUMMARY..................................................................................................................................... 7
INTRODUCCIÓN GENERAL
1. Cambios en los sistemas de manejo agrícola .......................................................... 13
2. Los cultivos frutícolas en las regiones biogeográficas peninsulares ........................ 14
3. La fauna edáfica ....................................................................................................... 16
4. Areas de estudio ...................................................................................................... 19
5. Bibliografía ............................................................................................................... 25
OBJETIVOS................................................................................................................................... 31
CAPÍTULO I. Propiedades edafoquímicas y microbiológicas de los agrosistemas atlánticos y mediterráneos
1. Introducción ............................................................................................................. 35
2. Materiales y métodos .............................................................................................. 37
3. Resultados ................................................................................................................ 40
4. Discusión .................................................................................................................. 57
5. Conclusiones ............................................................................................................ 61
6. Bibliografía ............................................................................................................... 62
CAPÍTULO II. Composición y estructura de las comunidades edáficas en los agrosistemas atlánticos y mediterráneos
1. Introducción ............................................................................................................. 69
2. Materiales y métodos .............................................................................................. 72
3. Resultados ................................................................................................................ 80
4. Discusión ................................................................................................................ 110
5. Conclusiones .......................................................................................................... 117
6. Bibliografía ............................................................................................................. 117
CAPÍTULO III. Relaciones ecológicas entre comunidades edáficas y factores ambientales en los agrosistemas atlánticos y mediterráneos
1. Introducción ........................................................................................................... 125
2. Materiales y métodos ............................................................................................ 129
3. Resultados .............................................................................................................. 130
4. Discusión ................................................................................................................ 139
5. Conclusiones .......................................................................................................... 145
6. Bibliografía ............................................................................................................. 146
CAPÍTULO IV. Variación en la composición isotópica y estabilidad de las comunidades edáficas en los agrosistemas atlánticos y mediterráneos
1. Introducción ........................................................................................................... 153
2. Materiales y métodos ............................................................................................ 157
3. Resultados .............................................................................................................. 159
4. Discusión ................................................................................................................ 170
5. Conclusiones .......................................................................................................... 174
6. Bibliografía ............................................................................................................. 175
CONCLUSIONES GENERALES ..................................................................................................... 181
ANEXOS ................................................................................................................................... 183


RESUMEN


Resumen
1
Resumen Los sistemas agrícolas han aportado a la sociedad bienes fundamentales, principalmente
proporcionando alimentos pero también fibras textiles, combustibles y medicamentos. Para
poder satisfacer las necesidades de una población humana en continuo crecimiento se han
desarrollado técnicas agrícolas que tienen casi como único objetivo el aumentar la
p odu ti idad, lo ue se e glo a de t o del té i o ag i ultu a o e io al . E t e los
desarrollos tecnológicos para incrementar la producción agrícola están el uso de pesticidas,
fitorreguladores, fertilizantes inorgánicos y el uso de maquinaria pesada para las labores de
siembra, poda y cosecha. Sin embargo, en las últimas décadas se ha constatado un gran número
de efectos negativos que este tipo de agricultura intensiva tiene sobre el medio ambiente a largo
plazo, como son la disminución de la fertilidad del suelo, contaminación de las aguas
subterráneas, pérdida de biodiversidad, etc. Para mitigar estos efectos negativos sobre el
ecosistema, durante el siglo XX comenzaron a desarrollarse métodos alternativos más
respetuosos con el medio ambiente. A esta nueva metodología agrícola se la denomina como
ag i ultu a e ológi a u o gá i a se a a te iza fundamentalmente por no emplear pesticidas
y permitir únicamente fertilizantes orgánicos. La agricultura orgánica se ha implementado con
éxito en Europa de tal manera que se han creado diversos organismos internacionales con el fin
de establecer una regulación internacional de estas prácticas.
Las regiones bioclimáticas se definen como áreas con características climáticas, geológicas y
vegetacionales similares. De las tres regiones biogeográficas presentes en la Península Ibérica,
atlántica, mediterránea y alpina, las dos primeras ocupan la mayor extensión y contienen la
mayor superficie agrícola útil. La región atlántica se ubica en la zona norte y noroeste de la
Península y se caracteriza por presentar temperaturas frescas y lluvia moderada a lo largo del
año, mientras que la región mediterránea presenta una marcada estacionalidad con veranos
cálidos y secos e inviernos húmedos y frescos. Las diferencias geológicas de las dos regiones se
reflejan en las distintas propiedades físico-químicas de los suelos, sobre rocas graníticas en el
caso de los ubicados en la región atlántica y sobre materiales calcáreos en los suelos
mediterráneos.
Estas diferencias climáticas y edafoquímicas entre ambas regiones condicionan el éxito
productivo de distintas especies agrícolas, de tal manera que en Galicia (región atlántica)
destacan los cultivos de kiwi (Actinidia deliciosa) y vid (Vitis vinifera), las cuales representan una
importante actividad económica para la región. Por otro lado, el cultivo de olivo (Olea europea)
y de mandarino (Citrus clementina) son especies claves en el sector agrícola de Cataluña (región
mediterránea).

Resumen
2
Las diferencias climáticas existentes entre las dos regiones determinan no sólo la selección
del tipo de planta que se cultiva sino también las prácticas agrícolas que se aplican, con el fin de
asegurar su buen crecimiento. Por ello, el primer objetivo de esta memoria fue describir las
propiedades edafoclimáticas, edafoquímicas y microbiológicas de cada sistema de cultivo
frutícola en cada región. En el capítulo I se describen los valores de pH, humedad, temperatura,
carbono (C), nitrógeno (N), relación C/N y carbono de la biomasa microbiana de las 8 fincas
agrícolas investigadas. Los resultados indicaron la existencia de importantes diferencias entre
las regiones atlántica y mediterránea. La humedad del suelo destacó como la variable que mejor
diferencia a los suelos de cultivo de las dos regiones seguida del pH, de tal forma que los suelos
atlánticos son más húmedos y ácidos que los mediterráneos. Estas condiciones inhiben las
actividades microbianas en los cultivos atlánticos y como resultado se favorece la retención de
nutrientes (C y N) y que las relaciones C/N sean más altas que las de la región mediterránea.
Sin embargo, el ambiente edafoquímico de los suelos de cultivo pueden verse alterado por
el tipo de cultivo y manejo; así los cultivos con mayor requerimiento hídrico (kiwi y mandarino)
presentaron suelos más húmedos que los plantados con vid y olivo y los sometidos a manejos
convencionales, sometidos a una mayor frecuencia de riego, más húmedos que los orgánicos.
Esto unido a un pH más favorable y una material vegetal menos recalcitrante puede acelerar los
procesos de descomposición de la materia orgánica. Confirmando resultados previos, los
sistemas de manejo orgánico favorecen una mayor retención de C en los suelos que los
convencionales y por tanto, se prevén a largo plazo pérdidas de C y disminuciones de las
relaciones C/N en los manejos intensivos, lo cuales requerirán mayores aportes orgánicos para
compensar las pérdidas de fertilidad. Sin embargo, la magnitud de esta respuesta dependerá en
gran medida de las reservas iniciales y cuánto tiempo lleva sometido el suelo a un manejo
convencional.
Además de las propiedades físico-químicas de los suelos, las propiedades biológicas también
determinan el funcionamiento de los agrosistemas. Las comunidades de invertebrados edáficos
tienen un papel clave en el mantenimiento de la estructura y fertilidad del suelo, ya que aceleran
los procesos de descomposición y reciclado de nutrientes beneficiando así a las especies
cultivadas. La reducida movilidad y la gran sensibilidad de los organismos edáficos a las prácticas
agrícolas hacen que sean considerados indicadores del estado de salud de los agrosistemas .
Dentro de la fauna edáfica, la mesofauna (entre 200 µm y 2 mm) y la macrofauna (>2 mm) son
los que dominan en los sistemas agrícolas, en términos de abundancia y biomasa,
respectivamente. Los taxones de mesofauna de mayor relevancia son los ácaros, colémbolos y
enquitreidos que participan en la descomposición de los restos vegetales y mantienen la
fertilidad del suelo y, en el caso de los enquitreidos, además, aumentan la porosidad del mismo.

Resumen
3
Por otro lado, las lombrices de tierra (macrofauna) están consideradas como ingenieras del
ecosistema y, gracias de las galerías que construyen, aumentan porosidad, drenaje y aireación
del suelo. Además, facilitan incorporación de materia orgánica hacia las capas más profundas
del perfil e incrementan las actividades microbiológicas estimulando así la humificación y
mineralización de la materia orgánica. En función del tipo de alimentación y de sus actividades
excavadoras las lombrices de tierra se clasifican en distintas categorías ecológicas, con efectos
diferentes sobre los procesos edáficos.
Por ello, en el segundo capítulo de la tesis, se describe la composición y estructura de las
comunidades edáficas en los ocho cultivos frutícolas. Se pudo comprobar que
independientemente del tipo de manejo agrícola y de la región bioclimática, los taxones de
mesofauna constituyen los grupos más numerosos en los suelos agrícolas, confirmando
observaciones previas. Sin embargo, las temperaturas suaves y los suelos más húmedos y más
ácidos de la región atlántica incrementan la abundancia de enquitreidos frente a los
microartrópodos, mientras que los suelos más cálidos y secos y con un pH más alto de la región
mediterránea favorecen a los ácaros.
La macrofauna (lombrices de tierra) también resultó ser muy sensible a las variaciones
climáticas y como consecuencia, las especies que viven en la superficie (epigeas) desaparecen
de los suelos de la región mediterránea. Además, sus comunidades también se vieron muy
afectadas por las prácticas agrícolas, haciendo que fuesen muy disimilares. Los sistemas
convencionales reducen la diversidad y aumentan la equitabilidad entre especies de lombrices
en ambas regiones, pero el efecto del manejo sobre estructura funcional varió según la región y
el tipo de cultivo: en la región atlántica, los manejos convencionales redujeron drásticamente el
número de lombrices endogeas y anécicas, mientras que en la región mediterránea dio como
resultado que las lombrices anécicas fueran la categoría ecológica dominante. Estas
observaciones fueron corroboradas por las comparaciones de curvas de abundancia y biomasa
de la macrofauna, las cuales indicaron una menor perturbación en las fincas con manejos
orgánicos con respecto a los convencionales, lo que provocó que en los últimos sus poblaciones
desviasen más energía a la supervivencia que al crecimiento individual.
En el tercer capítulo se aborda el estudio de las relaciones ecológicas entre las variables
ambientales que caracterizan a los cultivos frutícolas (capítulo I) y las comunidades edáficas de
estos suelos, que fueron descritas en el capítulo II. El objetivo fue determinar qué propiedades
edafoquímicas de los suelos de cultivo tienen una mayor influencia sobre la estructura de las
comunidades edáficas que viven en los suelos de cultivo frutícolas de las regiones atlántica y
mediterránea investigadas. Los resultados señalaron que la humedad del suelo, el pH y la
materia orgánica son las variables que regulan el tamaño de las poblaciones de organismos en

Resumen
4
los suelos de ambas regiones. Entre ellas, destaca en importancia la humedad para los
oligoquetos (enquitreidos y lombrices de tierra), los cuales pueden poder sufrir serias
reducciones en sus poblaciones en los periodos de sequía. Por otro lado, los ácaros, y en especial
los oribátidos, presentaron una mayor resistencia a la desecación facilitando su supervivencia
en la región mediterránea, mientras que el pH del suelo fue la variable más influyente para las
poblaciones de colémbolos.
Unos suelos más secos y menos ricos en materia orgánica, pero con valores altos de pH y
biomasa microbiana, explican la ausencia de lombrices epigeas y la abundancia de anécicas en
los cultivos mediterráneos y por qué en los suelos de la región atlántica dominan las especies
endogeas. Estas relaciones ecológicas observadas tienen una importancia crucial para los
procesos edáficos puesto que una menor densidad de especies excavadoras puede provocar
reducciones en la porosidad y la capacidad de drenaje del suelo con consecuencias negativas
para las especies vegetales cultivadas.
Dado que las variaciones climáticas y en el ambiente edafoquímico causadas por las prácticas
agrícolas modifican tanto las abundancias como la estructura de las comunidades, es necesario
conocer si las relaciones tróficas entre los distintos taxones también se ven afectadas. Por ello,
en el capítulo IV se investigaron las variaciones en las composiciones isotópicas (δ13C y δ15N) de
los taxones dominantes en los cultivos frutícolas de ambas regiones. Los resultados de estos
análisis permitieron constatar que las condiciones más cálidas de la región mediterránea
favorecen la dominancia de los organismos excavadores y euedáficos que consumen materia
orgánica más humificada. En cambio, los manejos orgánicos favorecen cadenas tróficas más
complejas que incluyen consumidores primarios y secundarios, lo que garantiza una mayor
estabilidad del agrosistema y por tanto, representa un indicador fiable del grado de perturbación
de un suelo cultivado que podría ser utilizado en el desarrollo de estrategias de manejo más
sostenibles.
En vista de todo ello, se puede concluir que las comunidades edáficas son altamente
sensibles a los cambios ambientales, tanto a las condiciones climáticas como los resultantes
de las prácticas agrícolas, viéndose alteradas tanto sus abundancias como la estructura y
composición de sus comunidades. Si bien los manejos orgánicos son más beneficiosos para los
organismos, la magnitud de los efectos negativos de los manejos convencionales pueden verse
contrarrestados mediante el mantenimiento de unas condiciones ambientales favorables para
la supervivencia de las especies que viven en la superficie del suelo (riegos regulares y una
cubierta vegetal que evite el calentamiento y la evaporación excesiva en las capas más
superficiales y además, aporte alimento a las comunidades descomponedoras).

SUMMARY


Summary
7
Summary Agricultural systems have provided essential goods and services to society, mainly food
provision but also textile fibers, fuels and medicines. In order to meet the needs of a constantly
growing human population, agricultural techniques have been developed that aim to increase
productivity, usually known as "conventional agriculture". Among the technological
developments to increase agricultural production is the use of pesticides, phytorregulators,
mineral fertilizers and heavy machinery for sowing, pruning and harvesting. However, in the last
decades, a great number of long-term negative environmental effects by these intensive
approaches have been detected, such as the reduction in soil fertility, contamination of
groundwaters, biodiversity losses, etc. To mitigate these negative effects on the ecosystem,
more environmentally friendly alternative methods began to be developed during the 20th
century. This new agricultural methodology is known as "organic farming" which does not use
pesticides and only organic fertilizers are allowed. Organic farming has been successfully
implemented in Europe and consequently, several international organizations have been
created in order to establish international regulations to ensure good practices.
Bioclimatic regions are areas with similar climatic, geological and vegetation properties. Of
the three biogeographical regions present in the Iberian Peninsula, Atlantic, Mediterranean and
Alpine, the former two occupy the greatest area and contain the largest arable surface. The
Atlantic region is located in the north and northwest of the Peninsula and is characterized by
cool temperatures and moderate rainfall throughout the year, while the Mediterranean region
has a marked seasonality with warm and dry summers and cool and wet winters. The geological
differences of the two regions are reflected in the different physicochemical properties of their
soils, granitic rocks in the case of those located in the Atlantic region and on calcareous materials
for the Mediterranean ones.
These climatic and edaphochemical differences between the two regions determine the
success of different cultivated plant species and thus, kiwifruit (Actinidia deliciosa) and
grapevine (Vitis vinifera) orchards represent an important economic activity in Galicia (Atlantic
region). On the other hand, olive trees (Olea europea) and clementines (Citrus clementina) are
important crops for the agricultural sector of Catalonia (Mediterranean region).
The climatic differences between the two regions determine not only the selection of the
plant species to be cultivated but also the agricultural practices to be applied in order to ensure
their healthy growth. For this reason, the first objective of this thesis was to describe the
edaphoclimatic, edaphochemical and microbiological properties of each orchard at each region.
In Chapter I, the values of soil temperature and moisture, pH, carbon (C) and nitrogen (N)

Summary
8
contents, C/N ratio and microbial biomass carbon of the eight investigated farms are described.
The results indicated important differences between the edaphochemical properties of the
Atlantic and Mediterranean regions. Soil moisture was the variable that best differentiates the
cultivated soils from the two regions followed by pH; accordingly, the Atlantic soils are wetter
and more acidic than the Mediterranean ones. These conditions limit microbial activities in
Atlantic orchards and as a result, nutrient retention (C and N) is favoured and C/N ratios are
higher than those measured in the Mediterranean region.
However, the type of cultivated plant and agricultural management can also modify the soil
chemical environment of the cultured soils. Thus, those orchards with greater water
requirements (kiwifruit and clementine) had wetter soils than those planted with vines and olive
trees, and those under conventional management, with more frequent irrigation, were moister
than those under organic treatments. This together with a more favorable pH and a less
recalcitrant plant material can accelerate decomposition processes of soil organic matter.
Confirming previous results, organic managements promote a higher soil C retention than
conventional ones and therefore, in the long-term, C losses and decreases in the C/N ratios are
expected in the most intensive systems. Consequently, conventional soils will require higher
organic inputs to compensate for fertility losses. However, the magnitude of this response will
largely depend on the initial reserves present in the soil and for how long the agricultural system
has been subjected to conventional management.
In addition to the physical-chemical properties of soils, the biological properties also
determine the functioning of the agrosystems. The soil invertebrate communities play a key role
in maintaining soil structure and fertility, since they accelerate the decomposition processes and
nutrient cycling, thus benefiting cultivated species. The reduced mobility of the soil organisms
and their strong sensitivity to agricultural practices make them good indicators of the health
status of agricultural systems. Within the soil fauna, mesofauna (between 200 μm and 2 mm)
and macrofauna (>2 mm) dominate in the agricultural soils, in terms of abundance and biomass,
respectively. The most important mesofauna taxa are mites, collembolans and enchytraeids that
participate in the decomposition of the plant remains and maintain the fertility of the soil and,
in the case of the enchytraeids, also increase soil porosity. On the other hand, earthworms
(macrofauna) are considered to be ecosystem engineers since their burrowing activities
increase soil porosity and improve drainage and aeration. In addition, they facilitate the deep
incorporation of organic matter and stimulate microbial activities, which accelerates the
humification and mineralization of the organic matter. Depending on the food consumed and
their burrowing abilities, earthworms are classified into different ecological categories, with
different effects on soil processes.

Summary
9
Therefore, the second chapter of this thesis describes the composition and structure of the
soil communities in the eight fruit orchards. It was possible to state that, regardless of the type
of agricultural management and of the bioclimatic region, mesofauna taxa represent the most
abundant groups in the agricultural soils, confirming previous observations. However, the mild
temperatures and the wetter and acidic soils in the Atlantic region promote enchytraeids against
microarthropods, whereas the hotter and drier soils with a higher pH of the Mediterranean
region favour mite populations.
Macrofauna (earthworms) also proved to be very sensitive to climatic variations and
therefore, species living on the surface (epigeic) became totally absent from the soils of the
Mediterranean region. Earthworms were also greatly affected by agricultural practices, making
their communities very dissimilar among treatments. Conventional systems reduced their
diversity and increased equitability among species at both regions, but the effect of
management on their functional structure varied according to region and plant type: in the
Atlantic region, conventional management drastically reduced the number of endogeic and
anecic worms, whereas in the Mediterranean region anecic worms became the dominant
ecological grouping. These observations were corroborated by the results derived from
comparing the abundance and biomass curves, which indicated less environmental disturbance
in the organic farms than in the conventional ones, which led to the populations in the latter
treatments to divert more energy to survival than to individual growth.
The third chapter aims to study the ecological relationships between the environmental
variables that characterise the fruit orchards (Chapter I) and their soil communities, which were
described in Chapter II. The objective of this study was to determine which edaphochemical
properties had the greatest influence on the structure of the soil communities in the cultivated
soils of the Atlantic and Mediterranean regions. The results indicated that soil moisture, pH and
organic matter were the variables that control the size of the soil faunal populations at both
regions. Among them, soil moisture was crucial for oligochaetes (earthworms and enchytraeids),
which are expected to suffer serious reductions in their population numbers with frequent
periods of drought. On the other hand, mites, but more specifically oribatids, exhibited a strong
tolerance to desiccation in the Mediterranean region, whereas soil pH was the most influential
variable for collembolans.
Dryer soils that are less rich in organic matter, but show high values of pH and microbial
biomass, are the main explanation for the absence of epigeic worms and the abundance of
anecics in the Mediterranean orchards and also for endogeic worms dominating in the soils of
the Atlantic region. These ecological relationships have a strong influence on soil processes since

Summary
10
a lower density of deep dwelling species will result in decreasing soil porosity and water drainage
with negative consequences for the cultivated plant species.
Since climatic variations and the soil chemical environment caused by agricultural practices
modify both the abundances and the structure of the soil communities, it was desirable to know
whether the trophic relationships between the different taxa could be also affected. For this
reason, the variations in the isotopic compositions (δ13C and δ15N) of the dominant faunal groups
from the different regions, agricultural managements and orchards were investigated in Chapter
IV. The results of these analyses allowed to verify that warmer conditions in the Mediterranean
region promote the dominance of euedaphic species that consume more humified organic
matter. In addition, organic managements favour more complex soil food webs that include
primary and secondary consumers and consequently, ensuring a greater stability of the
agroecosystem. Therefore, they represent a reliable indicator of the degree of disturbance of
the cultivated soil that could be used in the development of more sustainable agricultural
strategies.
In view of all this, it can be concluded that soil communities are highly sensitive to
environmental changes, either in climatic conditions or those resulting from agricultural
practices, with both their abundance and the structure and composition of their communities
becoming altered. Although organic managements seem to be the most beneficial for soil
organisms, the magnitude of the negative effects of conventional managements can be
counteracted by ensuring favourable environmental conditions for the survival of those
species living on the soil surface (i.e. by regular watering of the soil and by keeping a plant
cover that prevents heating and excessive evaporation from the surface layers and that also
provides food to the decomposing communities).

INTRODUCCIÓN GENERAL


Introducción General
13
1. CAMBIOS EN LOS SISTEMAS DE MANEJO AGRÍCOLA
Desde hace más de 10.000 años la agricultura proporciona una de las principales vías de
producción de alimentos para la sociedad, pero además genera otros bienes económicos: fibras
textiles, combustibles y medicamentos (Lindsey y Jones 1991). Los avances en las tecnologías
agrícolas han permitido que la superficie dedicada a la agricultura haya aumentado
considerablemente, de tal modo que actualmente el 38% de la superficie terrestre se encuentra
cultivada (FAOSTAT 2015). Esta intensificación de la producción agrícola conlleva acciones como
la eliminación de la cubierta vegetal, el volteo del suelo, el empleo de maquinaria agrícola
pesada y el uso excesivo de productos químicos que conducen a un empobrecimiento de los
suelos y a pérdidas de diversidad (Chan 2001; Guzmán y Alonso 2004; Biaggini et al. 2007;
Bouvier et al. 2011).
El continuo incremento de la población humana ha propiciado el desarrollo de metodologías
de mejora vegetal y de técnicas agrícolas a fin de incrementar la productividad. Un ejemplo de
ello es la de o i ada agricultura convencional , la cual ha logrado mejorar los rendimientos
de las cosechas gracias al uso de sistemas de riego, fitorreguladores, pesticidas, fertilizantes
inorgánicos y la mecanización de las labores de siembra y cosecha (Porta et al. 2003). El objetivo
de este tipo de agricultura ha sido maximizar la producción y los beneficios económicos sin tener
en cuenta los efectos negativos sobre los ecosistemas a largo plazo, como la disminución de la
calidad química y biológica de los suelos. Como consecuencia, a lo largo de las últimas décadas
se han producido importantes pérdidas de fertilidad, un consumo excesivo de agua y abuso de
agroquímicos que, en último término, afectan a los procesos edáficos (Reidsma et al. 2006;
Hendrickx et al. 2007; Menta 2012) y a los servicios ecosistémicos que prestan estos
agrosistemas (Hole et al. 2005; Postma-Blaauw et al. 2010).
Con el fin de mitigar los efectos medioambientales negativos de la agricultura convencional,
a mediados del siglo XX comienzan a desarrollarse métodos alternativos de producción agrícola
que tratan de conseguir un equilibrio entre productividad y medio ambiente, asegurando un uso
sostenible de los recursos naturales (van Eekeren et al. 2008; Palm et al. 2014). Entre ellos, el
cultivo orgánico o ecológico se ha implantado con bastante éxito en Europa. De hecho, según
datos de Eurostat, la UE-27 tenía en 2011 una superficie total de 9,6 millones de hectáreas
cultivadas como orgánico, frente a los 5,7 millones que había en 2002 (EU 2013). Como
consecuencia, se han creado diversos organismos internacionales encargados de regular las
actividades agrícolas enmarcadas en la denominación de agricultura orgánica a través de
diversos documentos legislativos (Reglamento CEE 2092/1991; Reglamento CE 1935/1995;
Reglamento CE 1804/1999; Reglamento CE 392/2004). En el caso concreto de España, este tipo

Introducción General
14
de agricultura está regulada desde 1989 por la Orden de 4 de Octubre (Orden 1989) por la que
se aprueba el Reglamento de la Denominación Genérica "Agricultura Ecológica" y su Consejo
Regulador. Esta normativa fue de aplicación hasta la entrada en vigor del Reglamento CEE
2092/91 sobre la producción agrícola ecológica y su indicación en los productos agrarios y
alimenticios. Actualmente, esta norma también está derogada y la producción,
almacenamiento, transformación, transporte, comercialización y etiquetado de los productos
ecológicos se regula por los Reglamentos CE 834/2007 y CE 889/2008.
Además de obtener productos agrícolas de mejor calidad, aprovechar mejor los recursos
naturales y respetar el medio ambiente, los suelos de cultivos orgánicos poseen una mayor
diversidad edáfica y actividad microbiológica que los convencionales, lo que favorece además
una mayor disponibilidad de nutrientes para las plantas cultivadas (Marinari et al. 2006).
2. LOS CULTIVOS FRUTÍCOLAS EN LAS REGIONES BIOGEOGRÁFICAS PENINSULARES
Las regiones biogeográficas son áreas con características climáticas, geológicas y de
comunidades vegetales similares. La Directiva Hábitats (92/43/EEC) ha favorecido que los
estados miembros realicen una mejor caracterización biogeográfica de sus regiones con el fin
de facilitar la conservación de hábitats y especies sometidos a condiciones similares,
independientemente de las divisiones políticas y administrativas. En la última revisión realizada
en el 2011 para la Europa de los 27 (EAA 2012), de las nueve regiones biogeográficas definidas,
la España Peninsular contiene territorios de tres de ellas (Fig. 1):
i) Región Atlántica: Se extiende desde el norte del Reino Unido e Irlanda hasta las costas
septentrionales de España y Portugal, pasando por los Países Bajos y parte de Alemania,
Dinamarca, Bélgica y Francia, en el centro. Debido a la proximidad del mar y a que la
mayor parte del territorio no alcanza altitudes elevadas, el clima oceánico penetra hasta
el interior, lo que conlleva inviernos suaves, veranos frescos y lluvia moderada a lo largo
de todo el año debido a la predominancia de los vientos del oeste. En esta región los tipos
de hábitats mayoritarios son, en primer lugar, los agrícolas/jardines, representando el
38% de su superficie y en segundo, las praderas con el 30% (recopilado de varias fuentes
por EEA/ETC BD & ETC Water en EEA 2002).
ii) Región Alpina: Comprende siete de las cordilleras de mayor altitud y extensión como son
la de los Alpes (en el centro de la Unión Europea), los Apeninos (eje norte-sur de Italia),
los Pirineos (la frontera entre España y Francia), los Alpes escandinavos (recorren Suecia,
Finlandia y Noruega) y los Cárpatos (forman un arco de Eslovaquia a Rumanía). Con la
adhesión de Bulgaria en 2007, se han añadido dos nuevas estructuras montañosas a la

Introducción General
15
región alpina: la cordillera de los Balcanes y los montes de Ródope. Debido a las grandes
altitudes, el clima de esta región es relativamente frío y árido. Los bosques cubren las
vertientes más bajas (representando 41% de toda la superficie); pero, según va
incrementándose la altitud y descienden las temperaturas, los árboles escasean y dejan
paso a los pastos alpinos, páramos y matorrales de brezo (23% de la superficie; estimas
basadas en el 80% de la región según ETC/NPB en EEA 2002).
iii) Región Mediterránea: Abarca siete Estados miembros, bien de manera parcial (España,
Francia, Italia y Portugal) o total (Chipre, Grecia y Malta). El clima se caracteriza por
veranos secos y cálidos e inviernos húmedos y frescos, pero con grandes contrastes,
siendo frecuentes los repentinos episodios de aguaceros torrenciales o de vientos fuertes
(por ejemplo, el siroco, el mistral) en distintos momentos del año. Los principales hábitats
que caracterizan esta región son los agrícolas (36% de su superficie) y los bosques,
brezales y matorrales (54% del área total; recopilado de varias fuentes por EEA/ETC BD en
EEA 2002).
Figura 1: Mapa de las regiones biogeográficas en Europa (EEA 2012).
Por tanto, las condiciones orográficas y climáticas ejercen un efecto profundo en la
vegetación y la vida silvestre y condicionan los usos del suelo. En el caso de los suelos agrícolas,
los factores climatológicos tienen una fuerte influencia en la productividad (Iglesias y Minguez

Introducción General
16
1997), de tal forma que condiciones climáticas extremas como sequías, heladas o lluvias
torrenciales pueden provocar enormes pérdidas económicas (Parry et al. 2004). Esto conlleva a
que las especies que se cultivan en la Península Ibérica sean muy diferentes de una región a otra
(MAGRAMA 2015).
Así, en el extremo oeste de la región atlántica peninsular (Galicia), destacan los cultivos
frutícolas del kiwi y de la vid. El cultivo de kiwi representa una importante actividad económica,
con el 69% de la superficie agrícola dedicada a este cultivo y alcanzando el 80% de la producción
nacional (MAGRAMA 2015). La provincia más productiva es Pontevedra, con el 60% de la
producción total, siendo la que dedica más hectáreas a este cultivo (560 ha; MAGRAMA 2015).
En el caso de la vid, existen actualmente unas 26.022 ha dedicadas a este cultivo en Galicia, de
las cuales el 58% se localizan en la provincia de Pontevedra (MAGRAMA 2015). En concreto, en
Gali ia se ela o a i os o de o i a io es de o ige dife e tes: Mo te ei , ‘ias Bai as ,
‘i ei a “a a , ‘i ei o Valdeo as . Gali ia ue ta ade ás o a iedades autó to as de
u a o o so las u as Al a iño T ei adu a . El é ito del i o ela o ado o u a Al a iño
ha promovido un incremento considerable de su producción en la región vinícola Rias Baixas en
los últimos años y de hecho, desde el año 2008 las exportaciones de vino con esta denominación
de origen no han parado de aumentar (Vinos de España 2015).
A diferencia de la región atlántica, los cultivos de cítricos y el olivo son claves en el sector
productivo agrícola de la región mediterránea. En concreto, en el extremo noreste (Cataluña) la
superficie dedicada al cultivo de mandarinas es de 121.869 ha, de las cuales más del 99% se
localizan en la provincia de Tarragona (IDESCAT 2015). De hecho, España es el segundo país del
mundo en producción de mandarinas sólo superado por China (FAOSTAT 2015); pero, mientras
España ejerce un papel hegemónico en la exportación mundial de mandarina, la producción de
China se destina al consumo interno. Por otro lado, la región catalana ocupa el segundo lugar a
nivel nacional en extensión dedicada al cultivo del olivar (115.843 ha), siendo la provincia de
Tarragona donde se localiza el 59% de la superficie dedicada a este cultivo (IDESCAT 2015).
3. LA FAUNA EDÁFICA
Las comunidades edáficas son muy sensibles al tipo de manejo agrícola que se aplica a los
cultivos debido a su reducida capacidad para desplazarse a zonas con condiciones ambientales
más favorables. Esta sensibilidad a las perturbaciones ambientales ha llevado a varios
investigadores a considerar a la fauna edáfica como un indicador fiable del estado de salud de
los ecosistemas (Parisi et al. 2005; Parisi y Menta 2008; Yan et al. 2012; Paoletti et al. 2013).

Introducción General
17
De hecho, los organismos edáficos cumplen un papel en el mantenimiento de la estructura y
la fertilidad del suelo (Wolters 2000; Osler y Sommerkorn 2007), al acelerar los procesos de
descomposición de la materia orgánica y el reciclado de nutrientes.
Existen diversas clasificaciones para describir la diversidad edáfica y una muy utilizada es la
propuesta por Swift et al. (1979) que se basa en clasificar los organismos del suelo en tres
grandes grupos según su diámetro corporal:
i) Microfauna: organismos con un diámetro comprendido entre las 20 y las 200 µm (por
ejemplo, nematodos y protozoos)
ii) Mesofauna: organismos con un diámetro entre 200 µm y 2 mm (por ejemplo, ácaros,
colémbolos, enquitreidos, proturos, sínfilos, pseudoescorpiones, etc.).
iii) Macrofauna: organismos con un diámetro superior a los 2 mm (por ejemplo,
gasterópodos, la mayoría de insectos, miriápodos, lombrices de tierra, etc.).
En términos de biomasa, la macrofauna y mesofauna son los organismos que dominan en
suelos agrícolas frutícolas (Lago 2015). En concreto, la mesofauna participa activamente en la
acondicionamiento de la hojarasca facilitando el ataque por parte de microorganismos y por
tanto, acelerando la descomposición de los restos vegetales y contribuyendo al mantenimiento
de la fertilidad del suelo (Coleman et al. 2004). Entre los grupos taxonómicos que integran la
mesofauna, los ácaros (Phylum Chelicerata, Clase Arachnida, Subclase Acari) y los colémbolos
(Phylum Hexapoda, Subphylum Hexapoda, Clase Entognatha, Subclase Collembola) son los que
dominan numéricamente, mientras que los enquitreidos (Phylum Annelida, Clase Clitellata,
Subclase Oligochaeta, Familia Enchytraeidae) suelen representar la mayor biomasa.
A pesar de su pequeño tamaño, los ácaros tienen un papel relevante tanto en la aireación
del suelo como en los ciclos de nutrientes (Hoy 2011), aunque algunas especies son plagas de
ciertos cultivos. Los oribátidos constituyen el suborden de ácaros numéricamente dominante en
los sistemas edáficos y en general, se caracterizan por ser organismos beneficiosos para los
suelos agrícolas debido a su importante papel en la descomposición de la materia orgánica (Hoy
2011). A pesar de ser considerados organismos resistentes a las perturbaciones ambientales
suelen responder a las actividades agrícolas (como pueden ser por el uso de sustancias tóxicas
como pesticidas o fertilizantes), por lo que podrían ser utilizados como bioindicadores de la
calidad del suelo (Behan-Pelletier 1999). Estudios como el de Lebrun y van Straalen (1995) y
Siepel (1996) han demostrado que las poblaciones de oribátidos se reducen cuando su hábitat
sufre perturbaciones.
Los colémbolos están presentes en una gran variedad de suelos y destacan por su papel clave
en la descomposición de la materia orgánica y por su capacidad para favorecer el crecimiento
de las micorrizas, incrementando la disponibilidad de nutrientes para las plantas (Hopkin 1997).

Introducción General
18
Además, actúan en el control biológico de plagas reduciendo las poblaciones de ciertas especies
de hongos perjudiciales para la agricultura. Son organismos sensibles a los pesticidas utilizados
comúnmente en los manejos convencionales (Frampton 1999).
Los enquitreidos, gracias a sus actividades excavadoras, incrementan la porosidad del suelo,
mejoran la filtración del agua y aceleran los procesos de transformación de la materia orgánica
(Didden 1990; van Vliet et al. 1993; Carrera et al. 2009). Otro beneficio que aporta este grupo a
la agricultura es su potencial capacidad para controlar plagas de nematodos fitoparásitos (Jegen
1920; O´Connor 1967).
Entre los taxones incluidos en la macrofauna, las lombrices de tierra son el grupo con mayor
relevancia en los sistemas agrícolas (Lee 1985; Wardle 1995; Lavelle et al. 1997; Paoletti et al.
2013) al mejorar las propiedades físicas del suelo a una mayor escala debido a su mayor tamaño:
además de la construcción de túneles y galerías que facilitan el movimiento de aire y agua a lo
largo del perfil del suelo, incorporan materia orgánica hacia capas más profundas. También
mejoran las propiedades químicas de los suelos al incrementar la disponibilidad de nutrientes
fácilmente asimilables para las plantas y contribuyen a la humificación y mineralización de la
materia orgánica gracias a su interacción con las poblaciones microbianas (sus he es o asts
depositadas e la supe fi ie o e sus gale ías so o side adas hostpots pa a los
microorganismos) (Edwards y Bohlen 1996; Lavelle y Spain 2001).
En base al tipo de alimentación y sus actividades excavadoras, las lombrices de tierra se
clasifican en tres categorías ecológicas (Bouché 1972):
(i) Lombrices epigeas: lombrices de pequeño tamaño (entre 0,5 y 5 cm) que presentan
colores rojizos y rosados, habitan en las capas más superficiales del suelo y se alimentan
de materia orgánica fresca. Suelen tener tasas metabólicas y reproductivas altas, por lo
que responden muy rápido a cambios en las condiciones ambientales en la superficie
del suelo.
(ii) Lombrices anécicas: lombrices que suelen superar los 5 cm de longitud, están muy
pigmentadas y construyen galerías verticales que descienden hasta los horizontes
minerales, aunque se alimentan deposita sus he es asts en superficie. Tienen
buenas capacidades excavadoras gracias al gran desarrollo de la musculatura en la
región anterior del cuerpo. Su tasa reproductiva es relativamente baja.
(iii) Lombrices endogeas: el tamaño de estas lombrices oscila entre 1 y 8 cm, presentan
escasa pigmentación y viven en los horizontes minerales dónde construyen galerías
horizontales y se alimentan de materia orgánica mineralizada. Sus tasas de reproducción
son bajas y sus ciclos de vida largos.

Introducción General
19
A pesar de esta clasificación, no todas las especies de lombrices encajan perfectamente
en estas definiciones. Por ello, Bouché (1977) amplió el número de categorías para incluir
especies con características intermedias, como por ejemplo, las categorías epianécica y
epiendogea, para englobar a aquellas especies que muestran características intermedias
entre dos categorías ecológicas.
4. AREAS DE ESTUDIO: FINCAS DE CULTIVOS FRUTÍCOLAS EN DOS REGIONES BIOGEOGRÁFICAS PENINSULARES
Las áreas de estudio se localizaron en las comarcas del Bajo Miño y Vigo (Pontevedra, Galicia)
en la vertiente noroeste de la región atlántica y en las comarcas catalanas del Bajo Campo y
Tarragonés (Tarragona, Cataluña) en la vertiente noreste mediterránea (Fig. 2). En cada una de
estas comarcas se seleccionaron un cultivo frutícola que estuvieran sometidos a dos manejos
agrícolas distintos, uno con manejo convencional y otro orgánico (en total ocho fincas).
Figura 2: Localización de las áreas de estudio en las dos regiones investigadas.
4.1. Caracterización climática de las fincas
La provincia de Pontevedra se caracteriza por tener un clima subtipo Atlántico y variedad
Gallego (de Castro et al. 2005). Los datos de temperatura del aire y precipitaciones durante el
periodo de estudio fueron obtenidos a partir de las estaciones meteorológicas más próximas
(estaciones de As Eiras en el ayuntamiento de Tui y la de Entenza situada en el municipio de
Salceda de Caselas; MeteoGalicia 2013). El climograma elaborado a partir de estos datos (Fig. 3)
muestra que las mayores precipitaciones se recogieron en los meses invernales (enero-marzo)
con valores superiores a los 300 mm mensuales, que además fueron los más fríos (8-10 oC); en
cambio, julio y agosto del 2013 fueron extremadamente secos (6 mm) y cálidos (21,9 oC).

Introducción General
20
Figura 3: Variación mensual en la temperatura del aire y la precipitación en las fincas atlánticas a lo largo del periodo de estudio.
La región mediterránea se caracteriza por presentar un clima subtipo de fachada oriental y
variedad Catalán (de Castro et al. 2005). Los datos de precipitación y temperatura de la estación
meteorológica más próxima a las fincas de estudio (estación del municipio de Constantí; Servei
Metereológic de Catalunya, 2013) indican una reducida precipitación a lo largo del año de
estudio (Fig. 4), siendo marzo el mes que registró los mayores valores de precipitación mensual
(92,4 mm) y julio los más bajos (15,2 mm). En cuanto a la temperatura del aire medida a lo largo
del periodo investigado (Fig. 4), agosto fue el mes más caluroso fue (23,9 oC) y el más frío
diciembre (9,5 oC).
Figura 4: Variación mensual en la temperatura del aire y la precipitación en las fincas mediterráneas a largo del periodo de estudio.
4.2. Cultivos frutícolas
4.2.1. Cultivo de kiwi (Actinidia deliciosa cv. Hayward)
Las fincas dedicadas al cultivo de kiwi y objeto de este estudio se localizan en los
ayuntamientos de Figueiró (manejo convencional) y Goián (manejo orgánico) de la provincia de
Pontevedra.
0
50
100
150
200
250
300
350
400
0
5
10
15
20
25
30
Mar-13 Abr-13 May-13 Jun-13 Jul-13 Ago-13 Sep-13 Oct-13 Nov-13 Dic-13 Ene-14 Feb-14
Pre
cipitació
n (m
m)Te
mp
era
tura
(oC
)
Precipitación Temperatura del aire
0
50
100
150
200
250
300
350
400
0
5
10
15
20
25
30
Mar-13 Abr-13 May-13 Jun-13 Jul-13 Ago-13 Sep-13 Oct-13 Nov-13 Dic-13 Ene-14 Feb-14
Pre
cipitació
n (m
m)Te
mp
era
tura
(oC
)
Precipitación Temperatura del aire

Introducción General
21
La finca dedicada al cultivo convencional de kiwi (41o 58´ 20´´ N, 8o 46´ 34´´ W) tiene una
extensión de 21,03 ha (Fig. 5A). El cultivo de este fruto comenzó en 1987 en sustitución al uso
forestal al cual estaba dedicada anteriormente.
El riego de esta finca es por microaspersión. En primavera se realiza un abonado nitrogenado
químico de fondo y también durante los meses de mayo a julio mediante fertirrigación, con el
fin de que la planta obtenga los nutrientes necesarios durante los periodos más críticos para el
crecimiento del fruto.
Además, se aplican dos productos herbicidas a mediados de mayo (glufosinato amónico 15%,
3-5 L ha-1 y terbutilazina al 50%, 2 L ha-1) y se realizan enmiendas calcáreas periódicas con el fin
de mantener un pH adecuado para el desarrollo de la planta. Sin embargo, no se realizaron
enmiendas durante el año de estudio.
La finca de kiwi orgánico (41o 57´ N, 8° 46´ W) ocupa 0,79 ha (Fig. 5B) y anteriormente
el uso del terreno del cultivo estuvo destinado a invernadero de horto-fruticultura y
previamente a la producción de forraje para el ganado. Estas prácticas cesaron en 1998, año en
el que comenzó la producción orgánica de kiwi.
El periodo de riego se inicia con el aumento de las temperaturas y se realiza mediante
microaspersión. Periódicamente se realizan labores de encalado para mantener un pH óptimo
para el desarrollo del cultivo, pero no se aplicaron enmiendas calizas durante el periodo de
estudio. Para la fertilización se emplean fosfatos naturales blandos, sulfato de potasio y un
compost elaborado en la propia finca a partir de los restos de las podas y excrementos de aves
de corral. El abonado realizado en el año 2013 consistió en la adición al suelo de 70,7 Kg ha-1 de
P, 28,3 kg ha-1 de Ca; 6,54 kg ha-1 de Mg, 237,25 kg ha-1 de K y 56,04 kg ha-1 de S.
Figura 5: Fincas de cultivo de kiwi. (A) Convencional y (B) Orgánico.
4.2.2. Cultivo de vid (Vitis vinífera cv. Albariño)
Las fincas destinadas al cultivo de la vid se localizan en los ayuntamientos de Figueiró (manejo
convencional) y Salceda de Caselas (manejo orgánico) de la provincia de Pontevedra.
A B

Introducción General
22
La finca dedicada al cultivo de vid con manejo convencional (41o 58´ 23´´ N, 8o 47´ 5´´ W) tiene
una superficie de 17 ha, de las cuales sólo 7,7 ha están dedicadas al cultivar albariño (Fig. 6A). El
uso agrícola comenzó en esta finca en 1998, cambiando el uso forestal al que estaba dedicada
anteriormente. En esta finca se emplea la espaldera como sistema de soporte para la planta,
que es el sistema más utilizado en las plantaciones de uva de vinificación en todo el mundo y en
el que las dos ramas principales del tronco de la cepa se alinean en el sentido del surco, atándose
al alambre más cercano al suelo. Con este sistema, junto con una orientación adecuada de la
plantación, se aumenta la insolación en la planta mejorando la capacidad de maduración de la
uva.
La técnica empleada para el riego de esta finca es la microaspersión, aunque en general los
requerimientos de riego son muy bajos para este cultivo. Se realizaron labores de encalado a
finales del mes de marzo de 2013 con el objetivo de mantener un pH adecuado. Además, se
aplicaron dos tratamientos con herbicidas (glifosato 48%) en dosis de 5,4 L de producto por
hectárea y desde abril hasta agosto se realizaron de 2 a 4 tratamientos mensuales con fungicidas
y dos tratamientos con insecticidas. Al igual que en los cultivos convencionales anteriores, se
realizó un abonado químico de fondo (960 kg) que aportaron al suelo 30-40 kg ha-1 de N 20-30
kg ha-1 de P2O5 y 15-30 kg ha-1 de MgO.
La finca con vid sometida a manejo orgánico (42o 4´ 20´´ N, 8o 33´ 42´´ W) tiene una superficie
de 2,02 ha dedicadas al cultivo de Vitis vinífera cv. Albariño (Fig. 6B). El cultivo ecológico
certificado comenzó en el año 2009, aunque previamente se llevaba cultivando vid de forma
convencional desde 1978.
Como sistema de soporte para la vid en esta finca es de tipo pérgola que consiste en una
serie de postes verticales de apoyo de un andamio en posición horizontal (techo). Es un sistema
adecuado para tierras fértil, con buena disponibilidad de agua ya que busca lograr un buen
equilibrio entre producción de la planta y una buena exposición al sol.
Como ocurre en el cultivo convencional, los requerimientos de riego del viñedo son muy
bajos. El fertilizante aplicado consiste en una enmienda orgánica obtenida a partir de algas y
estiércoles de equino aportando 30 kg ha-1 de N, 36 kg ha-1 de P, 43,2 kg ha-1 de K y 630 kg de
ha-1 de C al suelo.

Introducción General
23
Figura 6: Fincas de cultivo de vid. (A) Convencional y (B) Orgánico.
4.2.3. Cultivo de mandarino (Citrus clementina var. Clemenules)
Las fincas de este cultivo se sitúan en los municipios de El Morrel (manejo convencional) y de
La Selva del Camp (manejo orgánico) de la provincia de Tarragona.
El cultivo de mandarino convencional (41o 12´ 6´´ N, 1o 11´ 18´´E) tiene una superficie de 1,8
ha y dedicado a esta labor productiva desde el año 1998 (Fig. 7A). Anteriormente el terreno
estaba dedicado a la producción de árboles frutales de melocotón (Prunus persica).
El riego se realiza a través de microaspersores y entre las adiciones de agroquímicos se
incluyen insecticidas, acaricidas, fungicidas y herbicidas. El abonado de fondo, aplicado en
invierno, es a base de estiércol de gallina (7.000 kg ha-1), mientras que el de primavera se basa
en abonos nitrogenados, fósforo y potasio a través de fertilizantes sólidos químicos (32% N).
Además, también se aplica un abonado de cobertera mediante fertirrigación (NPK).
La finca dedicada al cultivo de mandarino orgánico (41o 13´ 5´´N, 1o 9´ 7´´E) ocupa 0,76 ha
(Fig. 7B) y cuenta con aproximadamente 320 árboles de Citrus clementina var. Clemenules
injertados sobre un patrones híbridos de carrizo [Citris sinensis (L.) Osb. x Poncirus trifoliata (L.)
Raf.].
Tras su uso anterior de producción de mandarina mediante técnicas convencionales, en el
año 2002 se inicia el manejo orgánico de esta finca, cumpliendo desde entonces con los
estándares de la agricultura orgánica. De acuerdo con esta normativa, no se utilizan pesticidas,
herbicidas, ni fungicidas y sólo se emplea abono orgánico como fertilizante una vez al año (150
kg N ha-1, 21,8 kg P ha-1 y 24 kg K ha-1). Además, en el abonado de fondo realizado en enero del
año 2013 se aplicó abono mineral (199 kg K ha-1 y 37 kg P ha-1) para compensar los posibles
desequilibrios en potasio y fosforo. El tipo de riego se realiza mediante el sistema de
microaspersión.
A B

Introducción General
24
Figura 7: Fincas de cultivo de mandarino. (A) Convencional y (B) Orgánico.
4.2.4. Cultivo de olivo (Olea europea L. cv. Arbequina)
Los cultivos de olivo están ubicados en los municipios de El Morrel (manejo convencional) y
de La Selva del Camp (manejo orgánico) de la provincia de Tarragona.
La finca dedicada al cultivo de olivo con manejo convencional (41o 12´ 9´´ N, 1o 11´ 20´´ E)
abarca unas 0,31 ha (Fig. 8A). Las prácticas agrícolas utilizadas en esta finca incluyen la aplicación
de insecticidas, acaricidas, fungicidas y herbicidas. En los meses de primavera y otoño se añade
al suelo un humectante para incrementar la persistencia de las aplicaciones fitosanitarias que
contienen como principio activo alquil poliglicol (0,661 L ha-1).
A diferencia de los cultivos de mandarino, no se abona por fertirrigación y la frecuencia de
riego es más baja. La fertilización en invierno consiste en purín de gallinaza (7.000 kg ha-1) y en
primavera abonos químicos nitrogenados (NPK). De forma habitual se añaden 50-150 kg ha-1 de
N y con 0,5 kg/árbol de P2O5 cuando se observan carencias de fósforo.
La finca de olivo con manejo orgánico (41o 13´ 4´´N, 1o 8´ 34´´E) tiene una extensión de 0,56
ha (Fig. 8B).
Durante el año de estudio no se realizaron labores de fertilización y se suspendió el riego;
pero en años anteriores se añadió abono verde de veza que aporta 250 kg ha-1 de K2SO4 y 25 kg
ha-1 de P.
Figura 8: Fincas de cultivo de olivo. (A) Convencional y (B) Orgánico.
A B
A B

Introducción General
25
5. BIBLIOGRAFÍA
Behan-Pelletier V M. 1999. Oribatid mites in agroecosystems: role for bioindication. Agriculture, Ecosystems & Environment, 74, 411–423.
Biaggini M, Consortu R, Dapporto L, Dellacasa M, Paggetti E y Corti E. 2007. The taxonomic level order as a possible tool for rapid assessment of arthropod diversity in agricultural landscapes. Agriculture, Ecosystems & Environment, 122(2), 183-191.
Bouché M B. 1972. Lombriciens de France, Ecologie et Systematique. Institut National de la Recherche Agronomique, Paris.
Bouché M B. 1977. Strategies Lombriciences. En: Lohm U y Persson T (Eds.) Soil Organisms as Components of Ecosystems, pp. 122-132. Ecological Bulletins. Stockholm.
Bouvier J, Ricci B, Agerberg J y Lavigne C. 2011. Apple orchard pest control strategies affect bird communities in southeastern France. Environmental Toxicology & Chemistry, 30(1), 212-219.
Carrera N, Barreal M E, Gallego P P y Briones M J I. 2009. Soil invertebrates control peatland c fluxes in response to warming. Functional Ecology, 23, 637-648.
Chan K Y. 2001. An overview of some tillage impacts on earthworm population abundance and diversity - implications for functioning in soils. Soil & Tillage Research, 57, 179-191.
Coleman D C, Crossley D A y Hendrix P F. 2004. Fundamentals of Soil Ecology. Elsevier Academic Press. Burlington.
de Castro M, Martín-Vide J y Alonso S. 2005. 1. El clima de España: pasado, presente y escenarios de clima para el siglo XXI. En: Moreno Rodríguez J M (Ed.) Evaluación preliminar de los impactos en España por efecto del Cambio Climático, pp. 1-64. Ministerio de Medio Ambiente. Madrid.
Didden W A M. 1990. Involvement of enchytraeidae (Oligochaeta) in soil structure evolution in agricultural fields. Biology & Fertility of Soils, 9, 152-158.
Directiva 92/43/CEE del Consejo de 21 de mayo de 1992 relativa a la conservación de los hábitats naturales y de la fauna y flora silvestre. Diario Oficial de las Comunidades Europeas de 22 de julio de 1992, Nº 250, 7-50.
Edwards C A y Bohlen P J. 1996. Biology and Ecology of Earthworms. Chapman & Hall, London. EU 2013. Facts and figures on organic agriculture in the European Union. Disponible en
http://ec.europa.eu/agriculture/markets-and-prices/more-reports/pdf/organic-2013_en.pdf.[Acceso en septiembre 2015].
European Environment Agency (EEA) 2012. Biogeographical Regions. Revisión 1 (http://www.eea.europa.eu/data-and-maps/figures/biogeographical-regions-in-europe-1).
FAOSTAT. 2015. Food and Agriculture Organization of the United Nations. Statistics division. Disponible en: www.faostat3.fao.org [Acceso en marzo 2015].
Frampton G K. 1999. Spatial variation in non-target effects of the insecticides chlorpyrifos, cypermethrin and pirimicarb on collembola in winter wheat. Pesticide Science, 55(9), 875-886.
Guzmán G C y Alonso M. 2004. Manual De Olivicultura Ecológica. Instituto de sociología y estudios campesinos. Universidad de Córdoba. Córdoba.
Hendrickx F, Maelfait J-P, van Wingerden W, Schweigers O, Speelmans M, Aviron S, Augenstein I, Billeter R, Bailey D, Bukacek R, Burel F, Diekötter T, Dirksen J, Herzog F, Liira J, Roubalova M, Vandomme V y Bugter R. 2007. How landscape structure, land-use intensity and habitat diversity affect components of total arthropod diversity in agricultural landscapes. Journal of Applied Ecology, 44(2), 340-351.
Hole D G, Perkins A J, Wilson J D, Alexander I H, Grice P V y Evans A D. 2005. Does organic farming benefit biodiversity? Biological Conservation, 122(1), 113-130.
Hopkin S P. 1997. Biology of the Springtails (lnsecta: Collembola). Oxford University Press, Oxford. Hoy M A. 2011. Agricultural Acarology - Introduction to Integrated Mite Management. CRC Press, Taylor
& Francis Group. Boca Raton. Iglesias A y Minguez M I. 1997. Modelling crop-climate interactions in Spain: vulnerability and adaptation
of different agricultural systems to climate change. Mitigation & Adaptation Strategies for Global Change, 1(3), 273-288.
Institut d´Estadística de Catalunya (IDESCAT). Disponible en: www.idescat.cat. [Acceso en marzo 2015]. Jegen G. 1920. Die Bedeutung der Enchytraeiden für die Humusbildung. Landwirtschaftliches Jahrbuch
der Schweiz, 34, 55-71. Lago C F. 2015. Efecto del manejo agrícola sobre las propiedades biológicas y químicas del suelo y los
procesos de transformación de la materia orgánica en cultivos de kiwi (Actinidia deliciosa). (Tesis Doctoral). Universidad de Vigo, Vigo.

Introducción General
26
Lavelle P y Spain A V. 2001. Soil Ecology. Kluwer Academic Publishers. Boston. Lavelle P, Bignell D, Lepage M, Wolters V, Roger P, Ineson P, Heal O W, Dhillion S. 1997. Soil function in a
changing world: the role of invertebrate ecosystem engineers. European Journal of Soil Biology, 33(4), 159-193.
Lebrun P y Straalen N M. 1995. Oribatid mites: prospects for their use in ecotoxicology. Experimental and Applied Acarology, 19(7), 361-379.
Lee K E, 1985. Earthworms: Their Ecology and Relationships with Soils and Land Use. Academic Press, New York.
Lindsey K y Jones M G K. 1991. Biotecnología Vegetal Agrícola. Ed. Acribia. Madrid. MAGRAMA. 2015. Avance del anuario de estadística agroalimentaria del Ministerio de Agricultura,
Alimentación y Medio ambiente. Disponible en: www.magrama.gob.es. [Acceso en marzo 2015]. Marinari S, Mancinelli R, Campigla E y Grego S. 2006. Chemical and biological indicators of soil quality in
organic and conventional farming systems in central Italy. Ecological Indicators, 6, 701-711. Menta C. 2012. Soil fauna diversity-function, soil degradation, biological indices, soil restoration. En:
Lameed G A (Eds.) Biodiversity Conservation and Utilization in a Diverse World, pp. 59-94. INTECH Open Access Publisher.
MeteoGalicia 2013. Xunta de Galicia. Disponible en: www.meteogalicia.es. [Acceso en marzo 2015]. O Co o F B. 1967. The Enchytraeidae. En: Burges A y Raw F (Eds.) Soil Biology, pp 213-256. Academic
Press, New York. Orden de 4 De octubre de 1989 por la que se aprueba el reglamento de la denominación genérica
«agricultura ecológica» y su consejo regulador. Boletín Oficial del Estado, Nº 239, de 5 de octubre de 1989, pp 31348-31352.
Osler G H R y Sommerkorn M. 2007. Toward a complete soil C and N cycle: incorporating the soil fauna. Ecology, 88(7), 1611-1621.
Palm C, Blanco-Canqui H, DeClerck F, Gatere L y Grace P. 2014. Conservation agriculture and ecosystem services: an overview. Agriculture, Ecosystems & Environment, 187, 87-105.
Paoletti M G, Sommaggio D y Fusaro S. 2013. Proposta Di Indice Di Qualità Biologica Del Suolo (QBS-e) Basato Sui Lombrichi E Applicato Agli Agroecosistemi. Biología Ambientale, 27(2), 25-43.
Parisi V y Menta C. 2008. Microarthropods of the soil: convergence phenomena and evaluation of soil quality using QBS-ar and QBS-c. Fresenius Environmental Bulletin, 17(8), 1170-1174.
Parisi V, Menta C, Gardi C, Jacomini C y Mozzanica E. 2005. Microarthropod communities as a tool to assess soil quality and biodiversity: a new approach in Italy. Agriculture, Ecosystems & Environment, 105(1-2), 323-333.
Parry M L, Rosenzweig C, Iglesias A, Livermore M y Fischer G. 2004. Effects of climate change on global food production under sres emissions and socio-economic scenarios. Global Environmental Change, 14(1), 53-67.
Porta J, López-Acevedo M y Roquero C. 2003. Edafología para la Agricultura y el Medio Ambiente. Mundi-Prensa, Madrid.
Postma-Blaauw M B, de Goede R G M, Bloem J, Faber J H y Brussaard L. 2010. Soil biota community structure and abundance under agricultural intensification and extensification. Ecology, 91(2), 460-473.
Reglamento (CE) nº 834/2007 del Consejo de 28 de junio de 2007 sobre producción y etiquetado de los productos ecológicos y por el que se deroga el Reglamento (CEE) No 2092/91. Diario Oficial de la Unión Europea de 20 De Julio De 2007, Nº 189, 1-23.
Reglamento (CE) Nº 889/2008 de la Comisión, de 5 de septiembre de 2008, por el que se establecen disposiciones de aplicación del Reglamento (CE) Nº 834/2007 del Consejo sobre producción y etiquetado de los productos ecológicos, con respecto a la producción ecológica, su etiquetado y su control. Diario Oficial de la Unión Europea de 18 de septiembre de 2008, Nº 250, 1-84.
Reidsma P, Tekelenburg T, van den Berg M y Alkemade R. 2006. Impacts of land-use change on biodiversity: an assessment of agricultural biodiversity in the European Union. Agriculture, Ecosystems & Environment, 114(1), 86-102.
Servei Metereológic de Catalunya. 2013. Disponible en: www.meteo.cat. [Acceso en marzo 2015]. Siepel H. 1996. Biodiversity of soil microarthropods: the filtering of species. Biodiversity & Conservation,
5, 251–260. Swift M J, Heal O W y Anderson J M. 1979. Decomposition in Terrestrial Ecosystems. Blackwell, Oxford.

Introducción General
27
van Eekeren N, Bommelé L, Bloem J, Schouten T, Rutgers M, de Goede R, Reheul D y Brussaard L. 2008. Soil biological quality after 36 years of ley-arable cropping, permanent grassland and permanent arable cropping. Applied Soil Ecology, 40(3), 432-446.
van Vliet P C J, West L T, Hendrix P F y Coleman D C. 1993. The influence of Enchytraeidae (Oligochaeta) on the soil porosity of small microcosms. Geoderma, 56, 287-299.
Vinos de España. Disponible en: www.winesfromspain.com. [Acceso en marzo 2015]. Wardle D A. 1995. Impacts of disturbance on detritus food webs in agro-ecosystems of contrasting tillage
and weed management practices. Advances in Ecological Research, 26, 105-185. Yan S, Singh A N, Fu S, Liao C, Wang S, Li Y, Cui Y y Hu L. 2012. A soil fauna index for assessing soil quality.
Soil Biology & Biochemistry, 47, 158-165.


OBJETIVOS


Objetivos
31
OBJETIVOS
El objetivo general de este estudio es conocer el efecto de los dos manejos agrícolas más
comúnmente empleados (convencional y orgánico) sobre las comunidades edáficas en cultivos
frutales localizados en dos regiones biogeográficas peninsulares diferentes: la vertiente
noroeste de la región atlántica y la vertiente noreste mediterránea (Fig. 1). Se ha puesto especial
énfasis en conocer los efectos del manejo sobre las comunidades de lombrices de tierra debido
a su papel beneficioso para la agricultura y a su rápida respuesta a perturbaciones en los
ecosistemas edáficos.
Po ta to, la hipótesis i i ial pa a este t a ajo se fo ula de la siguie te a e a: Las
condiciones climáticas históricas (regímenes de temperatura y precipitaciones) determinan la
composición específica de las comunidades edáficas que serán las mejor adaptadas a esos
suelos, por lo que aquellas intervenciones agrícolas que impliquen alteraciones en el ambiente
edafoquímico de los suelos provocarán alteraciones en sus abundancias y en la estructura de
sus o u idades .
Para evaluar la validez de esta hipótesis se plantearon los siguientes objetivos específicos:
1. Describir las propiedades edafoclimáticas, edafoquímicas y microbiológicas en cada
sistema de cultivo frutícola de cada región con el fin de identificar cuáles son las
variables que mejor caracterizan estos agrosistemas. [CAPÍTULO I]
2. Describir la composición y estructura de las comunidades edáficas (macrofauna y
mesofauna) en términos de abundancia y diversidad en función de las condiciones
climáticas, el tipo de cultivo y de manejo, así como determinar cómo el grado de
perturbación de los ecosistemas afecta el balance de crecimiento
somático/reproductivo de la macrofauna. [CAPÍTULO II]
3. Establecer una relación directa entre las propiedades edafoquímicas de los suelos de
cultivo y las propiedades biológicas con el fin de poder determinar qué factores edáficos
tienen una mayor influencia sobre la estructura de las comunidades de organismos que
viven en los suelos de cultivo frutícolas de las regiones atlántica y mediterránea
investigadas. [CAPÍTULO III]
4. Utilizar técnicas de isotopos estables para identificar qué factores (región, tipo de cultivo
y/o manejo) determina la estructura trófica de las comunidades edáficas así como el
grado de sensibilidad de la cadena alimentaria edáfica a las perturbaciones originadas
por los manejos intensivos. [CAPÍTULO IV]


CAPÍTULO I:
Propiedades edafoquímicas y microbiológicas de los agrosistemas atlánticos y mediterráneos


Capítulo I
35
1. INTRODUCCIÓN
Los agroecosistemas desempeñan servicios ecosistémicos importantes como son las
transformaciones del carbono (C), el reciclado de nutrientes, el mantenimiento de la estructura
del suelo y la regulación de plagas y enfermedades (Kibblewhite et al. 2008). Este tipo de
procesos pueden verse estimulados o inhibidos por las prácticas agrícolas (Power 2010), de tal
forma que cualquier alteración en las propiedades abióticas (características físico-químicas y
ambientales) y bióticas de los suelos de cultivo repercuten sobre el buen funcionamiento de
estos sistemas (Wardle 1995).
Entre los factores abióticos, la humedad del suelo es un factor con una relación directa con
el crecimiento de las plantas (Letey 1985) y por tanto, con la productividad de los cultivos. A
pesar de la importancia fundamental de este parámetro para el crecimiento del cultivo, no es
fácil obtener un valor fiable, puesto que la humedad del suelo depende de varios factores que
normalmente son muy difíciles de estimar como son la evapotranspiración y la cantidad de agua
que llega al suelo por precipitación y el riego (Letey 1985). Además, en la mayoría de los cultivos
la frecuencia de riego se programa con el fin de mantener el crecimiento de la planta a lo largo
del ciclo vital y por tanto, modificando los valores edáficos de humedad.
La temperatura es otro factor que interacciona con la humedad del suelo (Letey 1985) y que
también tiene un efecto directo sobre el crecimiento y desarrollo de los cultivos (Letey 1985;
Ritchie y NeSmith 1991) así como sobre la estructura y composición de las comunidades de los
organismos edáficos (Gkisakis et al. 2016). Varios autores han observado que la temperatura del
suelo puede verse modificada por el tipo de prácticas agrícolas. Así, los suelos de cultivo con
manejos convencionales en los que hay volteo del suelo, retirada de la cubierta vegetal y/o de
poda, experimentan variaciones bruscas de temperatura (Franzluebbers et al. 1995). En cambio,
en los sistemas donde se aplica agricultura orgánica y en los que se retienen los restos orgánicos
resultantes de las podas y cosecha del cultivo anterior ayudan a reducir la evapotranspiración y
las variaciones en temperatura y humedad del suelo (Franzluebbers et al. 1995; Palm 2014). Las
variaciones en estos dos factores abióticos también tienen un efecto directo sobre los
invertebrados edáficos y aquellas especies capaces de tolerar periódicamente valores extremos
de temperatura y humedad son capaces de incrementar sus abundancias (Kautz et al. 2006).
El pH del suelo es otro factor que limita la producción agrícola, de tal manera que los suelos
muy ácidos pueden alterar o inhibir el crecimiento y desarrollo de algunas especies vegetales.
Los pH bajos incrementan la movilidad de ciertos metales como el aluminio y manganeso,
llegando a alcanzar niveles tóxicos (von Uexküll y Mutert 1995; Kochian et al. 2004). El uso
excesivo de fertilizantes nitrogenados también contribuye a acidificar el suelo (Belay et al. 2002).

Capítulo I
36
Finalmente, solamente algunos organismos edáficos son capaces de tolerar valores bajos de pH
del suelo (Kibblewhite et al. 2008). Por todo esto, es importante regular el pH de los suelos de
cultivo para, en su caso, poder aplicar enmiendas correctoras y mantener comunidades edáficas
activas.
Entre los factores bióticos, la vegetación tiene un papel fundamental en la regulación de la
cantidad de carbono (C) que entra al suelo. Las plantas fijan C gracias a la fotosíntesis pero la
capacidad de retención de ese C depende del tipo de vegetación dominante y de las condiciones
ambientales de precipitación y de la temperatura (Gupta y Rao 1994). El uso anterior de un suelo
también tiene una gran importancia en la retención de C y por ejemplo, las transformaciones de
pastizales y bosques en cultivos produce pérdidas de este elemento que puede alcanzar niveles
del 80% (Guo y Gifford 2002).
Para el crecimiento de las plantas, además de C, son necesarios aportes de nitrógeno (N). El
N puede llegar al suelo del cultivo por fijación de bacterias simbiontes en las raíces (micorrizas),
por la adición de fertilizantes y/o a través de la mineralización de la materia orgánica (Urbano
Terrón et al. 1998). Para que el cultivo pueda seguir siendo productivo es necesario el aporte
regular de enmiendas nitrogenadas con el fin de reponer el N retirado cuando se cosecha la
cubierta vegetal (Robertson y Vitousek 2009). Para ello, el agricultor puede: i) incluir rotaciones
con plantas fijadoras de N2 (por ejemplo, leguminosas) o ii) aportar abonos nitrogenados
(orgánico o mineral). Desde los años 50, los fertilizantes minerales han favorecido la expansión
agrícola potenciando la productividad (LeBauer y Treseder 2008; Robertson y Vitousek 2009).
Las necesidades de N de cada cultivo dependen de diversos factores como son la variedad de
planta empleada, el rendimiento potencial y la calidad del producto cultivado (García-Serrano y
Ruano 2010). Además, varía a lo largo del ciclo fenológico del cultivo y por ejemplo, en los
cultivos arbóreos, las mayores necesidades de N se requieren durante la floración y fructificación
(García-Serrano y Ruano 2010).
Otro factor biológico importante en el mantenimiento de la fertilidad de los suelos de cultivo
son los microorganismos (Sparling 1985). La comunidad microbiana está constituida
principalmente por bacterias y hongos que tienen un efecto beneficioso sobre el desarrollo de
las plantas, gracias a su labor en la mineralización de la materia orgánica, la solubilidad de los
nutrientes y la fijación del N (Bender et al. 2016). La comunidad microbiana de los sistemas
agrícolas se ve afectada por diversos factores ambientales, las propiedades edafoquímicas y el
tipo de manejo agrícola (Bossio et al. 1998; Curiel Yuste et al. 2007). Por ejemplo, Fierer y
Jackson (2006) observaron una correlación positiva del pH del suelo con la diversidad y riqueza
de las comunidades microbianas; en cambio, Allison et al. (2013) detectaron una relación
negativa entre abundancia de bacterias y humedad del suelo (con reducciones del 86%).

Capítulo I
37
También se ha observado una buena correlación positiva entre poblaciones microbianas y el
contenido en C y N del suelo (Wardle 1992) así como una gran sensibilidad de los
microorganismos al uso de agroquímicos (Pampulha y Oliveira 2006; García-Orenes et al. 2010).
De ahí que manejos agrícolas que incluyan la retirada de los restos vegetales de la superficie del
suelo y/o herbicidas y pesticidas, reducirán la calidad y cantidad de nutrientes disponibles para
las poblaciones microbianas (Christensen 1996), modificarán las condiciones de humedad y
temperatura (Souza Andrade et al. 2003) y alterarán la composición y estructura de sus
comunidades de invertebrados edáficos (Pampulha y Oliveira 2006; García-Orenes et al. 2010).
Debido a que la mayoría de estos factores interactúan es muy difícil cuantificar la magnitud
de cada uno de ellos, por lo que no hay consenso en la comunidad científica sobre si las
condiciones climáticas o las agrícolas tienen mayor influencia sobre el funcionamiento de los
agrosistemas (Lobel y Field 2007; Licker et al. 2010).
2. MATERIALES Y METODOS
La toma de muestras se realizó mensualmente desde marzo de 2013 hasta febrero de 2014
en las fincas agrícolas situadas en la región atlántica. En cambio, en el caso de las fincas
localizadas en la región mediterránea, se concentraron en los meses en los que las condiciones
de temperatura y humedad (Fig. 4) no fuesen limitantes para las actividades microbiológicas
(Allison et al. 2013; Curiel Yuste el al. 2007) como ocurre en los meses estivales (Fig. 4) y por
tanto, se realizaron en primavera (de marzo a mayo de 2013) y otoño (de septiembre a
diciembre de 2013).
En cada una de las salidas al campo se tomaron tres muestras de suelo al azar en tres sectores
distintos en cada una de las fincas (elegidos también al azar), con el fin de incluir la variabilidad
espacial en el diseño experimental. Se emplearon cilindros de acero de 10 cm de diámetro x 20
cm de altura que fueron excavados cuidadosamente. Del suelo extraído se obtuvieron cuatro
muestras homogéneas de los 10 primeros cm de este cilindro de suelo que fueron utilizados
para las siguientes determinaciones: 1) la determinación de la humedad del suelo, 2) pH, 3)
contenido de C, N y 4) contenido de C de la biomasa microbiana.

Capítulo I
38
Figura 9: Cilindros de acero de 10 cm de diámetro x 20 cm de altura para la determinación de las propiedades edafo-químicas y del contenido de C la biomasa microbiana.
2.1. Medidas de temperatura, humedad, pH y contenidos totales de C y N del suelo
La temperatura del suelo fue registrada de manera continua mediante registradores de datos
(data-logger HOBO® H08) enterrados a una profundidad de 10 cm en cada una de las fincas. En
cada muestreo mensual, los datos de temperatura de los 30 días anteriores a la fecha del
muestreo fueron descargados en el laboratorio.
La humedad del suelo fue calculada gravimétricamente tras secar muestras de suelo (2 - 4 g
peso fresco) a 105 oC en estufa hasta obtener peso constante.
El pH del suelo fue determinado en agua (en una relación 1:2,5; suelo:agua) a partir de
muestras de suelo secas y homogeneizadas (fracción < 2 mm). Para ello se empleó un pHmetro
(Crison micropH 2000).
El contenido total de C y N (%) del suelo fue determinado mediante un analizador elemental
LECO (CN-2000, LECO Corp., St Joseph, MI) en los Servicios Centrales de la Universidad de Vigo
(Centro de Apoio Científico-Tecnolóxico á Investigación, CACTI). A partir de estos datos se
calculó la relación C/N para cada muestra.
2.2. Determinación del carbono de la biomasa microbiana
Se tomaron dos muestras de 10 g de suelo (peso fresco) de cada réplica y una de ellas fue
fumigada con cloroformo libre de etanol. Posteriormente, se extrajo el C orgánico, tanto de las
muestras fumigadas como de las no fumigadas, utilizando 100 mL de sulfato potásico (K2SO4)
0,5 M (Vance et al. 1987). Para la cuantificación del C orgánico se empleó un autoanalizador de
flujo continuo (GE Sievers Innovox TOC Analyzer). El C de la biomasa microbiana (Cmic) se
expresó en mg C g-1 suelo seco según la siguiente ecuación: � �� = EcKc

Capítulo I
39
dónde Ec es la cantidad de C orgánico extraído del suelo fumigado menos C orgánico extraído
del suelo sin fumigar y Kc es una constante (Brake et al. 1999), que para los suelos minerales
toma el valor de 45% (KC=0,45) (Wu et al. 1990).
2.3. Análisis estadísticos
Los datos fueron analizados, en conjunto y por regiones biogeográficas, mediante análisis de
la varianza (ANOVA) con el fin de conocer si la región geográfica, el tipo de cultivo, el manejo
agrícola, el tiempo y/o la interacción de estos factores tuvo un efecto significativo sobre las
propiedades edafo-químicas (temperatura, humedad del suelo, pH, C, N, relación C/N) y
microbiológicas (C de la biomasa microbiana) de los suelos de cultivo frutícola investigados.
Debido a la diferente intensidad de muestreo en las dos regiones, se empleó una ANOVA para
diseños no balanceados para analizar el efecto de dos factores principales: región y manejo. La
separación de las medias se realizó con el test HSD (Honestly-significant-difference) de Tukey
α= , .
Previamente a estos análisis, se emplearon los test de Shapiro Wilk y de Levene para
comprobar la normalidad y homogeneidad de varianzas de los datos, respectivamente. Los
resultados de estos análisis revelaron que algunas variables requerían ser transformadas para
cumplir los criterios de la ANOVA y así, el C, el N, la relación C/N y el C de la biomasa fueron
transformados logarítmicamente.
Todos los análisis univariantes fueron realizados con el paquete estadístico STATISTICA v7
(StatSoft, 2005).
Además, para identificar las variables edáficas que mejor explican la variabilidad observada
en los suelos de los distintos cultivos sometidos a distintos manejos de las dos regiones
biogeográficas se empleó la técnica de ordenación de componentes principales (PCA). Los PCA
se realizaron en base a matrices de correlaciones de Pearson y la significación de los
componentes fue calculada siguiendo el étodo scree plot des ito po Ja kso . Este
método consiste en representar gráficamente los autovalores de cada uno de los componentes
en el orden en que fueron obtenidos junto con los intervalos de confianza del 95%; la forma que
adopta la curva resultante (asintótica negativa) se emplea para evaluar qué componentes
explican mejor los resultados (es decir, el punto en el que la curva se vuelve lineal y se aproxima
a cero indica el número máximo de componentes que se deben utilizar y se consideran
estadísticamente significativos).
Estos análisis multivariantes fueron realizados con el software PAST (PAleontological
STatistica ver 3.11; Hammer et al. 2001).
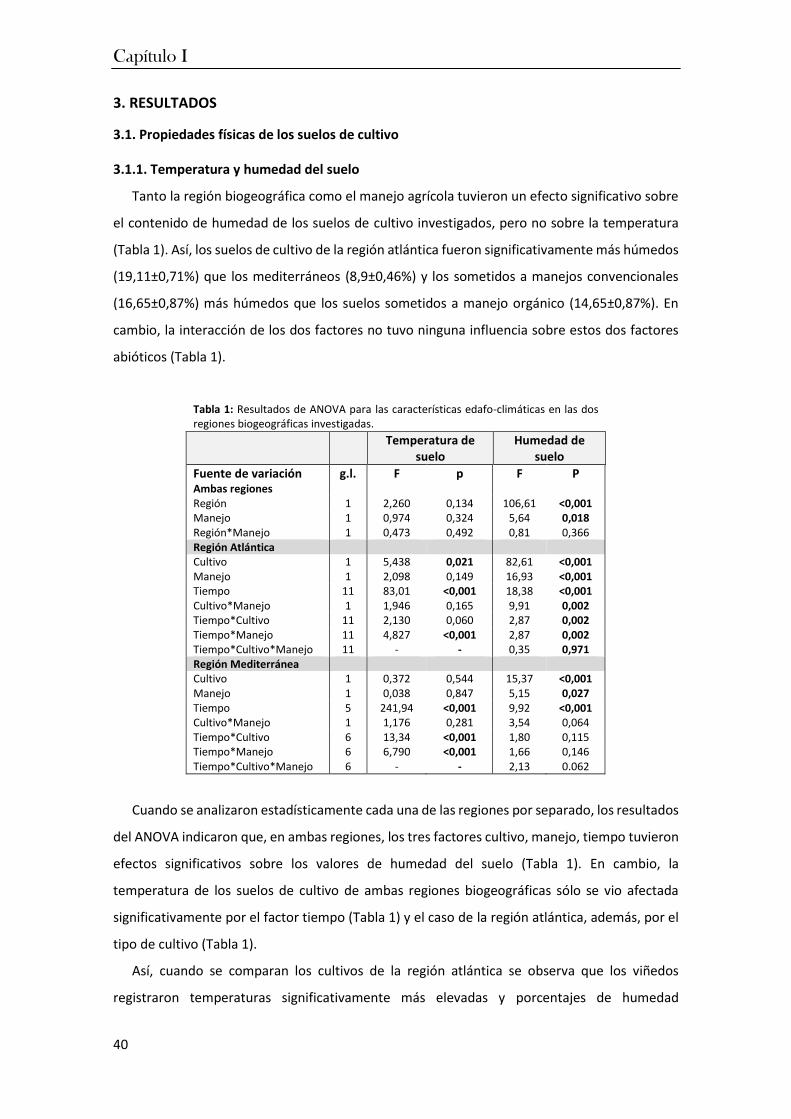
Capítulo I
40
3. RESULTADOS
3.1. Propiedades físicas de los suelos de cultivo
3.1.1. Temperatura y humedad del suelo
Tanto la región biogeográfica como el manejo agrícola tuvieron un efecto significativo sobre
el contenido de humedad de los suelos de cultivo investigados, pero no sobre la temperatura
(Tabla 1). Así, los suelos de cultivo de la región atlántica fueron significativamente más húmedos
(19,11±0,71%) que los mediterráneos (8,9±0,46%) y los sometidos a manejos convencionales
(16,65±0,87%) más húmedos que los suelos sometidos a manejo orgánico (14,65±0,87%). En
cambio, la interacción de los dos factores no tuvo ninguna influencia sobre estos dos factores
abióticos (Tabla 1).
Tabla 1: Resultados de ANOVA para las características edafo-climáticas en las dos regiones biogeográficas investigadas.
Temperatura de
suelo Humedad de
suelo
Fuente de variación g.l. F p F P Ambas regiones Región 1 2,260 0,134 106,61 <0,001 Manejo 1 0,974 0,324 5,64 0,018 Región*Manejo 1 0,473 0,492 0,81 0,366 Región Atlántica Cultivo 1 5,438 0,021 82,61 <0,001 Manejo 1 2,098 0,149 16,93 <0,001 Tiempo 11 83,01 <0,001 18,38 <0,001 Cultivo*Manejo 1 1,946 0,165 9,91 0,002 Tiempo*Cultivo 11 2,130 0,060 2,87 0,002 Tiempo*Manejo 11 4,827 <0,001 2,87 0,002 Tiempo*Cultivo*Manejo 11 - - 0,35 0,971 Región Mediterránea Cultivo 1 0,372 0,544 15,37 <0,001 Manejo 1 0,038 0,847 5,15 0,027 Tiempo 5 241,94 <0,001 9,92 <0,001 Cultivo*Manejo 1 1,176 0,281 3,54 0,064 Tiempo*Cultivo 6 13,34 <0,001 1,80 0,115 Tiempo*Manejo 6 6,790 <0,001 1,66 0,146 Tiempo*Cultivo*Manejo 6 - - 2,13 0.062
Cuando se analizaron estadísticamente cada una de las regiones por separado, los resultados
del ANOVA indicaron que, en ambas regiones, los tres factores cultivo, manejo, tiempo tuvieron
efectos significativos sobre los valores de humedad del suelo (Tabla 1). En cambio, la
temperatura de los suelos de cultivo de ambas regiones biogeográficas sólo se vio afectada
significativamente por el factor tiempo (Tabla 1) y el caso de la región atlántica, además, por el
tipo de cultivo (Tabla 1).
Así, cuando se comparan los cultivos de la región atlántica se observa que los viñedos
registraron temperaturas significativamente más elevadas y porcentajes de humedad

Capítulo I
41
significativamente más bajos que los de kiwi (Fig. 10A). En la región mediterránea los suelos de
cultivo sólo estuvieron afectados por la humedad y las fincas cultivadas con olivos las que
presentaron humedades significativamente más bajas que las de los suelos de las fincas de
mandarino (Fig. 10B).
Figura 10: Valores medios (±E.E.) de la humedad de suelo en los suelos de cultivo A) atlánticos y B) mediterráneos, Letras diferentes indican diferencias significativas (p<0,05) entre tratamientos.
En cuanto al efecto del manejo agrícola sobre la humedad del suelo (Tabla 1), en ambas
regiones los menores porcentajes de humedad fueron registrados en los cultivos orgánicos
frente a los convencionales (Figs. 11A,B).
Figura 11: Valores medios (±E.E.) de la humedad de suelo en los suelos de cultivo A) atlánticos y B) mediterráneos, en función del manejo agrícola. Letras diferentes indican diferencias significativas (p<0,05) entre tratamientos.
La interacción entre los factores cultivo y el manejo solo fue significativa para el caso del
porcentaje de humedad en los suelos de cultivos atlánticosl (Tabla 1). De acuerdo con esto, los
suelos de las fincas dedicadas al cultivo orgánico de la vid fueron los más secos, seguido del suelo
de cultivo de vid en sistema convencional (Fig. 12). En cambio los suelos de los cultivos de kiwi
producidos con ambos tipos de manejo mostraron valores de porcentaje de humedad muy
similares.
a
b
0
5
10
15
20
25
Mandarino Olivo
Hu
me
dad
del
Su
elo
(%
)
Ba
b
0
5
10
15
20
25
Kiwi Vid
Hu
med
ad d
el S
uel
o (
%)
A
b
a
0
5
10
15
20
25
OrgánicoConvencional
Hu
me
dad
del
Su
elo
(%
)
A
ba
0
5
10
15
20
25
OrgánicoConvencional
Hu
me
dad
del
Su
elo
(%
)
B

Capítulo I
42
Figura 12: Valores medios (±E.E.) de la humedad de suelo en los suelos de los cultivos atlánticos sometidos a diferentes manejos. Letras diferentes indican diferencias significativas (p<0,05) entre tratamientos.
En cuanto al factor tiempo, aunque se observó una clara estacionalidad en ambas regiones
(Fig. 13), destacaron el mes de septiembre por ser el más cálido (20,3±0,05 oC en la región
atlántica y 22,0±0,44 oC en la mediterránea) y el de diciembre por ser el más frio (8,6±0,12 y
8,9±0,38 oC, respectivamente). Además, en el caso de la región atlántica la menor humedad se
midió en los meses de verano (julio-septiembre), siendo septiembre el mes significativamente
más seco (9,07±2,22%) respecto a los meses de invierno (diciembre-febrero) y a los de primavera
(marzo-mayo), con porcentajes de humedad entre 19 y 26%. En la región mediterránea
septiembre también fue el más seco de los registrados, con valores del 5%.
En cuanto a las interacciones del factor tiempo con el resto de factores investigados (cultivo
y manejo), solamente la interacción tiempo*manejo tuvo un efecto significativo sobre la
temperatura del suelo de ambas regiones, mientras que la interacción tiempo*cultivo fue
significativa solamente para los suelos de la región mediterránea (Tabla 1). En el caso de la
humedad del suelo, todas las posibles interacciones entre estos tres factores resultaron ser
significativas en el caso de los suelos de la región atlántica pero no para los mediterráneos (Tabla
1).
Para simplificar la descripción de las variaciones temporales de la temperatura y la humedad
del suelo en relación a sus interacciones con el tipo de cultivo y de manejo, en la Fig. 13 se
muestran los climogramas de las 8 fincas estudiadas. Se puede observar que, en todos los suelos
investigados, los incrementos de temperatura ocurridos en los meses estivales coincidieron con
importantes descensos del contenido de agua del suelo (Fig. 13). En el caso de la región atlántica,
las oscilaciones en los valores de humedad del suelo a lo largo del tiempo fueron más acusadas
en los cultivos de viñedo que en los de kiwi (Figs. 13A-D) y en concreto, en los meses estivales la
humedad del suelo del cultivo de vid no superó el 12% (Figs. 13C,D). En la región mediterránea
(Figs. 13E-H), las mayores variaciones en los porcentajes de humedad fueron registradas en los
aa
b
c
0
5
10
15
20
25
30
Kiwi Con Kiwi Org Vid Con Vid Org
Hu
me
dad
del
Su
elo
(%
)

Capítulo I
43
olivares, siendo los meses de abril, septiembre y octubre los que registraron los menores
porcentajes de humedad (Figs. 13G,H); en cambio, los cultivos de mandarino presentaron suelos
más húmedos (Figs. 13E,F).
Figura 13: Valores medios (±E.E.) de la temperatura del suelo (o C) y la humedad (%) durante el periodo de estudio en A) kiwi convencional, B) kiwi orgánico, C) vid convencional, D) vid orgánico, E) mandarino convencional, F) mandarino orgánico, G) olivo convencional y H) olivo orgánico. Letras diferentes indican diferencias significativas entre fechas de muestreo para cada finca (p <0,05).
a a a
a a a a
0
5
10
15
20
25
30
0
10
20
30
40
50
60
Mar-13 Abr-13 May-13 Sep-13 Oct-13 Nov-13 Dic-13
Temp
eratura d
el suelo
( ºC)
Hu
med
ad (
%)
Humedad (%)F
Temperatura del suelo
ab ab abb
abab
a
0
5
10
15
20
25
30
0
10
20
30
40
50
60
Mar-13 Abr-13 May-13 Sep-13 Oct-13 Nov-13 Dic-13
Temp
eratura d
el suelo
( ºC)
Hu
med
ad (
%)
Humedad (%)E
Temperatura del suelo
ab
bc
a
c bc c
ab
0
5
10
15
20
25
30
0
10
20
30
40
50
60
Mar-13 Abr-13 May-13 Sep-13 Oct-13 Nov-13 Dic-13
Temp
eratura d
el suelo
( ºC)
Hu
med
ad (
%)
Humedad (%)H
Temperatura del suelo
ab
b
a
ab ab
abab
0
5
10
15
20
25
30
0
10
20
30
40
50
60
Mar-13 Abr-13 May-13 Sep-13 Oct-13 Nov-13 Dic-13
Temp
eratura d
el suelo
( ºC)
Hu
med
ad (
%)
Humedad (%)
GTemperatura del suelo
a
aa
a a
aa
a a
a
aa
0
5
10
15
20
25
30
0
10
20
30
40
50
60
Temp
eratura d
el suelo
( ºC)
Hu
med
ad (
%)
Humedad (%)B Temperatura del suelo
ab
a
ab
ab
ab bb
ab
ab
ab ab
ab
0
5
10
15
20
25
30
0
10
20
30
40
50
60
Temp
eratu
ra de
l sue
lo ( ºC
)
Hu
med
ad (
%)
Humedad (%)A Temperatura del suelo
abc
a
abc
c
dd d
bc
bcbc
ab
a
0
5
10
15
20
25
30
0
10
20
30
40
50
60
Temp
eratura d
el suelo
( ºC)
Hu
med
ad (
%)
Humedad (%)
CTemperatura del suelo
a
ab ab
c
d d d
abbc
ab ab
ab
0
5
10
15
20
25
30
0
10
20
30
40
50
60
Temp
eratura d
el suelo
( ºC)
Hu
med
ad (
%)
Humedad (%)D
Temperatura del suelo
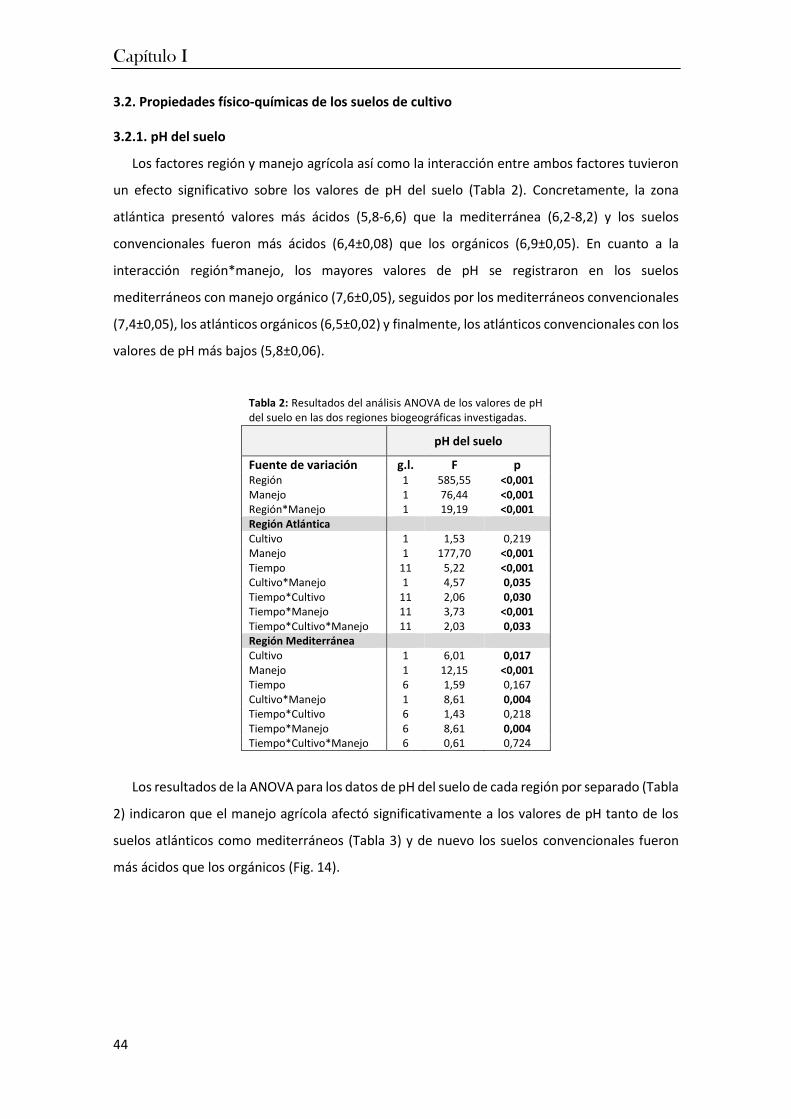
Capítulo I
44
3.2. Propiedades físico-químicas de los suelos de cultivo
3.2.1. pH del suelo
Los factores región y manejo agrícola así como la interacción entre ambos factores tuvieron
un efecto significativo sobre los valores de pH del suelo (Tabla 2). Concretamente, la zona
atlántica presentó valores más ácidos (5,8-6,6) que la mediterránea (6,2-8,2) y los suelos
convencionales fueron más ácidos (6,4±0,08) que los orgánicos (6,9±0,05). En cuanto a la
interacción región*manejo, los mayores valores de pH se registraron en los suelos
mediterráneos con manejo orgánico (7,6±0,05), seguidos por los mediterráneos convencionales
(7,4±0,05), los atlánticos orgánicos (6,5±0,02) y finalmente, los atlánticos convencionales con los
valores de pH más bajos (5,8±0,06).
Tabla 2: Resultados del análisis ANOVA de los valores de pH del suelo en las dos regiones biogeográficas investigadas.
pH del suelo
Fuente de variación g.l. F p Región 1 585,55 <0,001 Manejo 1 76,44 <0,001 Región*Manejo 1 19,19 <0,001 Región Atlántica Cultivo 1 1,53 0,219 Manejo 1 177,70 <0,001 Tiempo 11 5,22 <0,001 Cultivo*Manejo 1 4,57 0,035 Tiempo*Cultivo 11 2,06 0,030 Tiempo*Manejo 11 3,73 <0,001 Tiempo*Cultivo*Manejo 11 2,03 0,033 Región Mediterránea Cultivo 1 6,01 0,017 Manejo 1 12,15 <0,001 Tiempo 6 1,59 0,167 Cultivo*Manejo 1 8,61 0,004 Tiempo*Cultivo 6 1,43 0,218 Tiempo*Manejo 6 8,61 0,004 Tiempo*Cultivo*Manejo 6 0,61 0,724
Los resultados de la ANOVA para los datos de pH del suelo de cada región por separado (Tabla
2) indicaron que el manejo agrícola afectó significativamente a los valores de pH tanto de los
suelos atlánticos como mediterráneos (Tabla 3) y de nuevo los suelos convencionales fueron
más ácidos que los orgánicos (Fig. 14).

Capítulo I
45
Figura 14: Valores medios (±E.E.) del pH del suelo en los suelos de cultivo A) atlánticos y B) mediterráneos, en función del manejo agrícola. Letras diferentes indican diferencias significativas (p<0,05) entre tratamientos.
El tipo de cultivo sólo tuvo un efecto significativo en el caso de los suelos mediterráneos
(Tabla 2) y las fincas destinadas al cultivo de mandarino presentaron un pH inferior (7,46±0,32)
que las de olivos (7,63±0,39). En cambio, el factor tiempo solamente tuvo un efecto significativo
sobre el pH de los suelos atlánticos (Tabla 2) y los valores registrados fueron más bajos entre los
meses de octubre 2013 y febrero 2014 (valores en torno a 6) que en el resto de los meses
investigados (con valores comprendidos entre 6 y 6,2).
Los suelos de cultivo de ambas regiones estuvieron afectados significativamente por la
interacción cultivo*manejo y, en ambas regiones, se observó un gradiente decreciente de pH
desde las fincas orgánicas a las convencionales: en la región atlántica el orden fue viñedo
orgánico > kiwi orgánico > kiwi convencional > viñedo convencional (Fig. 15A) y en la
mediterránea, olivo orgánico > mandarino orgánico > mandarino convencional > olivo
convencional (Fig. 15B).
Figura 15: Valores medios (±E.E.) del pH del suelo en los suelos de cultivo A) atlánticos y B) mediterráneos, en función del tipo de cultivo y del manejo agrícola. Letras diferentes indican diferencias significativas (p<0,05) entre tratamientos.
En cuanto a las interacciones del factor tiempo con los otros dos factores (cultivo y manejo)
solamente la interacción entre tiempo y el manejo agrícola tuvo un efecto significativo sobre el
b
a
4
6
8
10
OrgánicoConvencional
pH
A
ba
4
6
8
10
OrgánicoConvencional
pH
B
a a ab
4
6
8
10
Mand Con Mand Org Olivo Con Olivo Org
pH
BB
a
b
a
b
4
6
8
10
Kiwi Con Kiwi Org Vid Con Vid Org
pH
A

Capítulo I
46
pH de los suelos de cultivo de ambas regiones (Tabla 2), destacando los valores más elevados de
los suelos con manejo orgánico durante el mes de agosto (6,9±0,1) frente al valor medio
registrado en febrero en los cultivos convencionales (5,3±0,16).
En cambio, solamente en las fincas atlánticas la interacción entre el factor tiempo y cultivo
así como la interacción tiempo*cultivo*manejo tuvieron una influencia significativa sobre los
valores de pH, de tal forma que los valores de pH más altos y más bajos fueron registrados en el
cultivo de kiwi (6,7±0,14 y 5,7±0,36 en el mes de agosto y en el de diciembre, respectivamente).
De forma similar, la interacción entre los factores tiempo, cultivo y manejo, también reflejó los
valores extremos de pH que se registraron en cultivos de kiwi, siendo más elevados en el mes
de marzo en la finca con manejo orgánico (6,8±0,11) y más bajos en el sistema convencional
durante el mes de diciembre (4,9±0,17).
3.2.2. Contenido de carbono del suelo
Los resultados de la ANOVA para el porcentaje de carbono del suelo indican un efecto
significativo de los factores región, manejo agrícola así como de la interacción entre ambos
(Tabla 3). Así, los valores más altos de C fueron registrados en la región atlántica (5,70±0,20%)
que la región mediterránea (2,39±0,09%) y en las fincas convencionales (4,97±0,29%) mayores
que las orgánicas (4,05±0,15%). Por último, los suelos de cultivo con manejo convencional en la
región atlántica presentaron los contenidos de carbono más elevados (7,02±0,39%), seguidos
por los suelos atlánticos orgánicos (4,73±0,17%) y finalmente, los valores más bajos fueron para
los suelos de cultivo sometidos a cualquiera de los dos manejos en la región mediterránea
(3,0±0,15% para manejo orgánico y 2,11±0,13% para el manejo convencional).

Capítulo I
47
Tabla 3: Resultados del análisis ANOVA de los valores de C del suelo en las dos regiones biogeográficas investigadas.
Carbono
Fuente de variación g.l. F p Región 1 162,53 <0,001 Manejo 1 8,24 0,001 Región*Manejo 1 23,02 <0,001 Región Atlántica Cultivo 1 2,99 0,087 Manejo 1 60,15 <0,001 Tiempo 11 13,50 <0,001 Cultivo*Manejo 1 0,05 0,824 Tiempo*Cultivo 11 1,28 0,242 Tiempo*Manejo 11 1,46 0,159 Tiempo*Cultivo*Manejo 11 1,39 0,188 Región Mediterránea Cultivo 1 9,69 0,003 Manejo 1 17,40 <0,001 Tiempo 6 3,26 0,009 Cultivo*Manejo 1 12,51 <0,001 Tiempo*Cultivo 6 2,07 0,074 Tiempo*Manejo 6 3,42 0,006 Tiempo*Cultivo*Manejo 6 1,15 0,346
Cuando se analizan las dos regiones por separado se observa que el contenido de C de los
suelos de cultivo mediterráneos se vieron afectados significativamente por los tres factores,
mientras que los de la región atlántica por el tipo de manejo agrícola y el factor tiempo (Tabla
3). Así, para el manejo, se observó que en la región atlántica los valores de C en los suelos de las
fincas orgánicas fueron significativamente inferiores a los registrados en las fincas
convencionales (Fig. 16A), mientras que las fincas convencionales de la región mediterránea
presentaron un contenido de C de suelo significativamente inferior al medido en las fincas
orgánicas (Fig. 16B).
Figura 16: Valores medios (±E.E.) del contenido de carbono del suelo en los suelos de cultivo A) atlánticos y B) mediterráneos, en función del manejo agrícola. Letras diferentes indican diferencias significativas (p<0,05) entre tratamientos.
a
b
0
2
4
6
8
10
Convencional Orgánico
C (
%)
A
ab
0
2
4
6
8
10
Convencional Orgánico
C (
%)
B

Capítulo I
48
En relación a las variaciones temporales del contenido de C en la región atlántica, los valores
significativamente más altos fueron registrados en los meses de primavera (8-9,2%) comparado
con el resto del año (4,3-6%). En cambio, en las fincas mediterráneas, destaca el elevado valor
de C registrado en el mes de mayo (2,9±0,16%), significativamente superior a los valores
registrados en septiembre, octubre y noviembre (2,04±0,25%, 2,11±0,30% y 1,89±0,27%,
respectivamente).
En cuanto al efecto del tipo de cultivo sobre el contenido de C, y para el caso de la región
mediterránea, destacan los valores significativamente mayores del cultivo de mandarino
(2,87±0,17%) comparados con los registrados en las fincas dedicadas al cultivo de olivo
(2,24±0,13%). Además, la interacción significativa entre tipo de cultivo y tipo de manejo
observada en los suelos de esta región (Tabla 3) reveló que el cultivo de mandarino mediante
manejo orgánico presentó contenidos de carbono significativamente más altos que el resto de
tratamientos (Fig. 17).
Figura 17: Valores medios (±E.E.) del carbono del suelo en los cultivos mediterráneos sometidos a diferentes manejos. Letras diferentes indican diferencias significativas (p<0,05) entre tratamientos.
En cuanto a las interacciones del factor tiempo con los otros dos factores investigados (Tabla
3) solamente la interacción tiempo*manejo resultó ser significativa y sólo en el caso de los
suelos de cultivo mediterráneos. De acuerdo con esto, destacan de nuevo los valores de
contenido de carbono significativamente más altos observados en el mes de mayo en los cultivos
orgánicos (2,9±0,7%), comparado con los valores tan bajos registrados en los cultivos
convencionales en los meses de octubre y noviembre (1,5±0,9 y 1,3±0,4%, respectivamente).
3.2.3. Contenido de nitrógeno del suelo
Los resultados de la ANOVA indicaron la existencia de diferencias significativas entre regiones
en cuanto al contenido de nitrógeno del suelo (Tabla 4), destacando los mayores valores
a
b
aa
0
1
2
3
4
Mandarino Con Mandarino Org Olivo Con Olivo Org
C(%
)
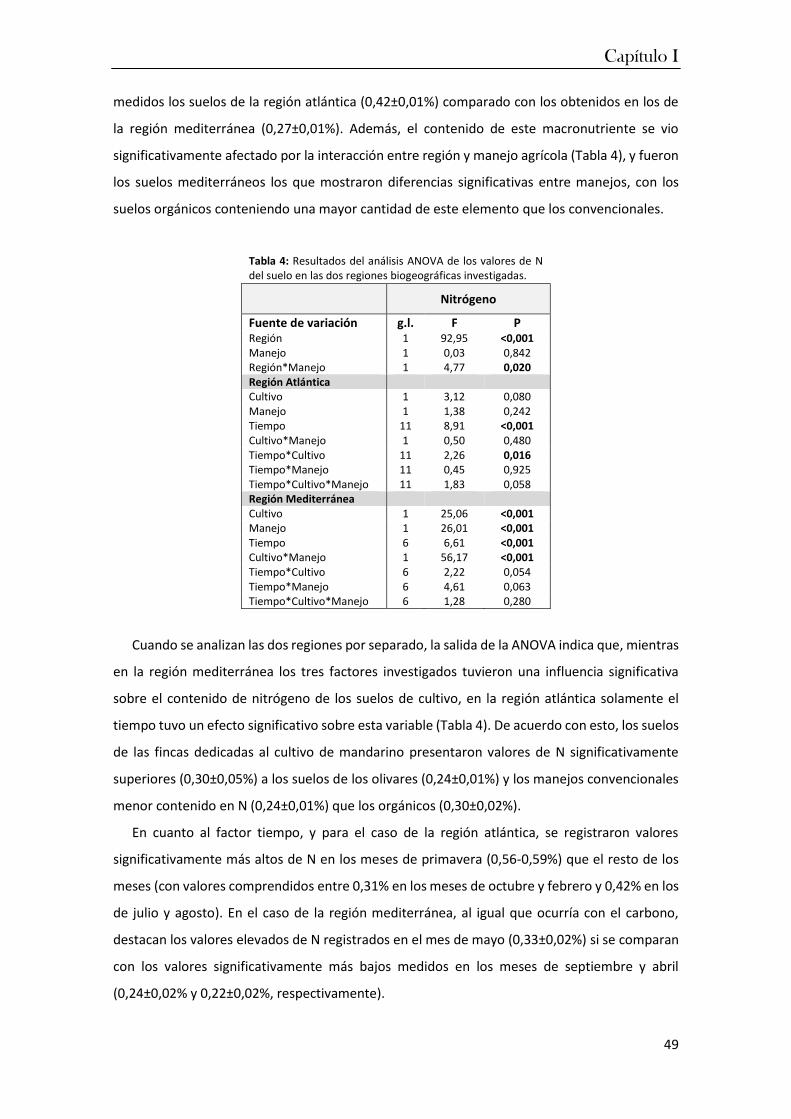
Capítulo I
49
medidos los suelos de la región atlántica (0,42±0,01%) comparado con los obtenidos en los de
la región mediterránea (0,27±0,01%). Además, el contenido de este macronutriente se vio
significativamente afectado por la interacción entre región y manejo agrícola (Tabla 4), y fueron
los suelos mediterráneos los que mostraron diferencias significativas entre manejos, con los
suelos orgánicos conteniendo una mayor cantidad de este elemento que los convencionales.
Tabla 4: Resultados del análisis ANOVA de los valores de N del suelo en las dos regiones biogeográficas investigadas.
Nitrógeno
Fuente de variación g.l. F P Región 1 92,95 <0,001 Manejo 1 0,03 0,842 Región*Manejo 1 4,77 0,020 Región Atlántica Cultivo 1 3,12 0,080 Manejo 1 1,38 0,242 Tiempo 11 8,91 <0,001 Cultivo*Manejo 1 0,50 0,480 Tiempo*Cultivo 11 2,26 0,016 Tiempo*Manejo 11 0,45 0,925 Tiempo*Cultivo*Manejo 11 1,83 0,058 Región Mediterránea Cultivo 1 25,06 <0,001 Manejo 1 26,01 <0,001 Tiempo 6 6,61 <0,001 Cultivo*Manejo 1 56,17 <0,001 Tiempo*Cultivo 6 2,22 0,054 Tiempo*Manejo 6 4,61 0,063 Tiempo*Cultivo*Manejo 6 1,28 0,280
Cuando se analizan las dos regiones por separado, la salida de la ANOVA indica que, mientras
en la región mediterránea los tres factores investigados tuvieron una influencia significativa
sobre el contenido de nitrógeno de los suelos de cultivo, en la región atlántica solamente el
tiempo tuvo un efecto significativo sobre esta variable (Tabla 4). De acuerdo con esto, los suelos
de las fincas dedicadas al cultivo de mandarino presentaron valores de N significativamente
superiores (0,30±0,05%) a los suelos de los olivares (0,24±0,01%) y los manejos convencionales
menor contenido en N (0,24±0,01%) que los orgánicos (0,30±0,02%).
En cuanto al factor tiempo, y para el caso de la región atlántica, se registraron valores
significativamente más altos de N en los meses de primavera (0,56-0,59%) que el resto de los
meses (con valores comprendidos entre 0,31% en los meses de octubre y febrero y 0,42% en los
de julio y agosto). En el caso de la región mediterránea, al igual que ocurría con el carbono,
destacan los valores elevados de N registrados en el mes de mayo (0,33±0,02%) si se comparan
con los valores significativamente más bajos medidos en los meses de septiembre y abril
(0,24±0,02% y 0,22±0,02%, respectivamente).
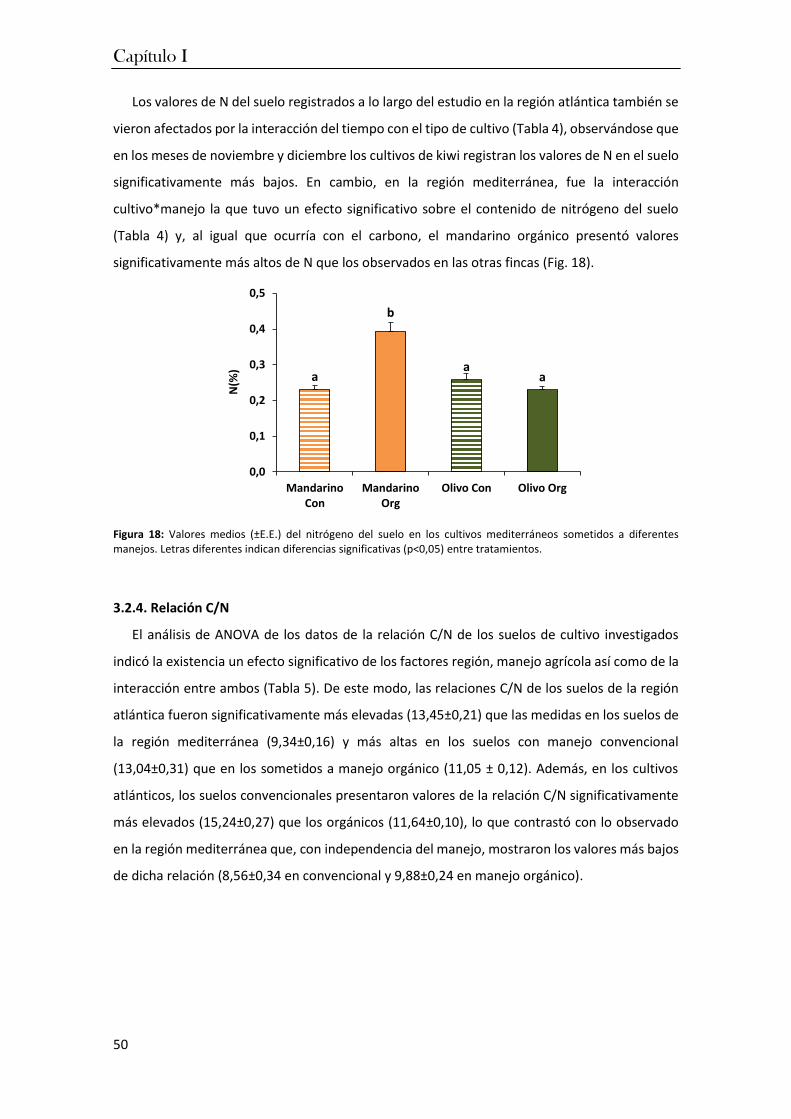
Capítulo I
50
Los valores de N del suelo registrados a lo largo del estudio en la región atlántica también se
vieron afectados por la interacción del tiempo con el tipo de cultivo (Tabla 4), observándose que
en los meses de noviembre y diciembre los cultivos de kiwi registran los valores de N en el suelo
significativamente más bajos. En cambio, en la región mediterránea, fue la interacción
cultivo*manejo la que tuvo un efecto significativo sobre el contenido de nitrógeno del suelo
(Tabla 4) y, al igual que ocurría con el carbono, el mandarino orgánico presentó valores
significativamente más altos de N que los observados en las otras fincas (Fig. 18).
Figura 18: Valores medios (±E.E.) del nitrógeno del suelo en los cultivos mediterráneos sometidos a diferentes manejos. Letras diferentes indican diferencias significativas (p<0,05) entre tratamientos.
3.2.4. Relación C/N
El análisis de ANOVA de los datos de la relación C/N de los suelos de cultivo investigados
indicó la existencia un efecto significativo de los factores región, manejo agrícola así como de la
interacción entre ambos (Tabla 5). De este modo, las relaciones C/N de los suelos de la región
atlántica fueron significativamente más elevadas (13,45±0,21) que las medidas en los suelos de
la región mediterránea (9,34±0,16) y más altas en los suelos con manejo convencional
(13,04±0,31) que en los sometidos a manejo orgánico (11,05 ± 0,12). Además, en los cultivos
atlánticos, los suelos convencionales presentaron valores de la relación C/N significativamente
más elevados (15,24±0,27) que los orgánicos (11,64±0,10), lo que contrastó con lo observado
en la región mediterránea que, con independencia del manejo, mostraron los valores más bajos
de dicha relación (8,56±0,34 en convencional y 9,88±0,24 en manejo orgánico).
a
b
aa
0,0
0,1
0,2
0,3
0,4
0,5
MandarinoCon
MandarinoOrg
Olivo Con Olivo Org
N(%
)

Capítulo I
51
Tabla 5: Resultados del análisis ANOVA de los valores de la relación C/N del suelo en las dos regiones biogeográficas investigadas.
C/N
Fuente de variación g.l. F P
Región 1 320,77 <0,001 Manejo 1 38,96 <0,001 Región*Manejo 1 88,750 <0,001 Región Atlántica Cultivo 1 0,19 0,613 Manejo 1 200,58 <0,001 Tiempo 11 2,34 0,022 Cultivo*Manejo 1 5,74 0,053 Tiempo*Cultivo 11 2,36 0,026 Tiempo*Manejo 11 0,79 0,745 Tiempo*Cultivo*Manejo 11 2,04 0,074 Región Mediterránea Cultivo 1 0,61 0,437 Manejo 1 8,16 0,001 Tiempo 6 3,53 0,006 Cultivo*Manejo 1 19,29 <0,001 Tiempo*Cultivo 6 1,32 0,263 Tiempo*Manejo 6 2,11 0,068 Tiempo*Cultivo*Manejo 6 1,70 0,139
Al analizar estadísticamente cada región por separado, se observa que el manejo y el tiempo
tuvieron un efecto significativo sobre los valores de la relación C/N de los suelos investigados
(Tabla 5). Así, los suelos atlánticos con manejo convencional presentaron valores de C/N
significativamente más altos (15,24±0,27) que los suelos orgánicos (11,65±0,11) y los suelos
mediterráneos con manejo orgánico más altos (9,69±0,16) que los registrados en los suelos
convencionales (8,96±0,27).
En cuanto a los cambios temporales de la relación C/N, en el caso de la región atlántica, los
valores se mantuvieron muy estables a lo largo de todo el periodo de estudio, pero en la región
mediterránea se observaron mayores variaciones (Tabla 5), destacando el mes de abril con los
mayores valores de esta relación (10,56±0,33) frente a los meses de mayo, octubre y noviembre,
con los valores más bajos (8,91±0,24; 8,71±0,43 y 9,02±0,69, respectivamente).
En el caso de la región atlántica la interacción tiempo*cultivo resultó ser significativa (Tabla
5), lo que fue debido a que en el mes de diciembre se pudo observar un descenso significativo
de la relación C/N en el cultivo de vid (10,10±0,10) con respecto al kiwi (13,60±1,30).
Al igual que ocurrió con los contenidos de C y N, la interacción cultivo*manejo también afectó
significativamente a los valores de la relación C/N en los suelos de cultivo mediterráneos (Tabla
5) y fueron los cultivos de olivo sometidos a manejo orgánico los que mostraron los valores
significativamente más altos que los de este cultivo en sistema convencional (Fig. 19).
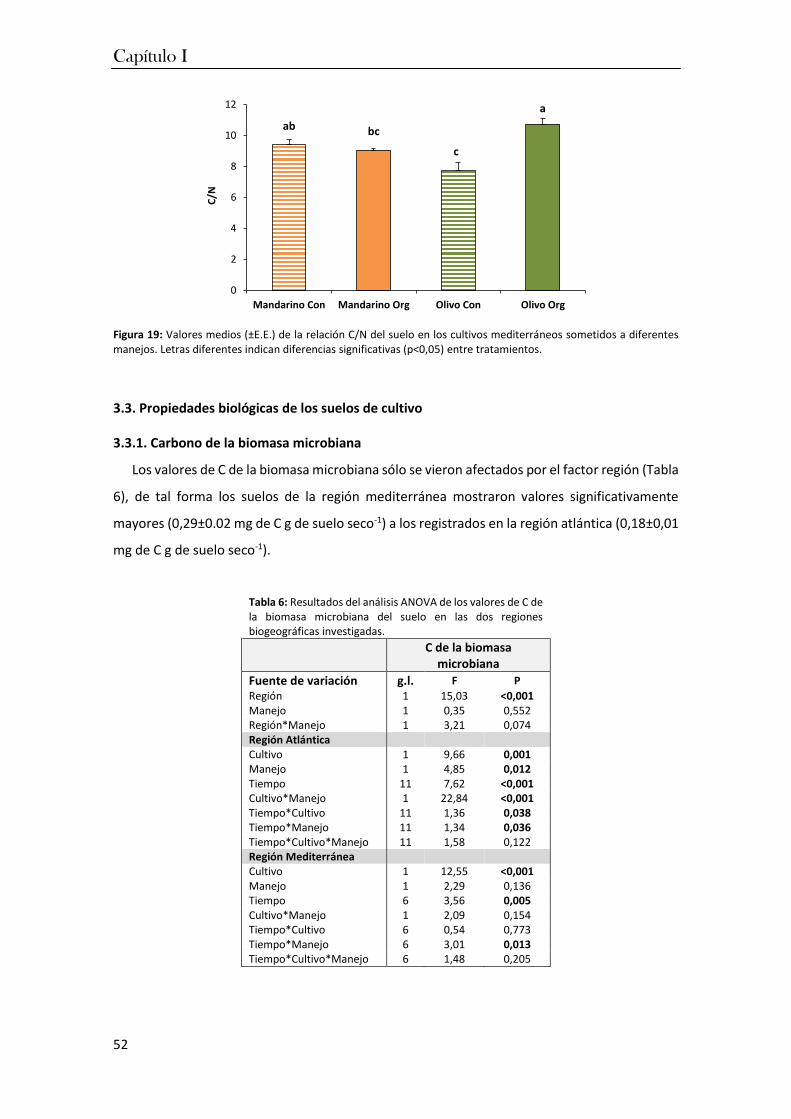
Capítulo I
52
Figura 19: Valores medios (±E.E.) de la relación C/N del suelo en los cultivos mediterráneos sometidos a diferentes manejos. Letras diferentes indican diferencias significativas (p<0,05) entre tratamientos.
3.3. Propiedades biológicas de los suelos de cultivo
3.3.1. Carbono de la biomasa microbiana
Los valores de C de la biomasa microbiana sólo se vieron afectados por el factor región (Tabla
6), de tal forma los suelos de la región mediterránea mostraron valores significativamente
mayores (0,29±0.02 mg de C g de suelo seco-1) a los registrados en la región atlántica (0,18±0,01
mg de C g de suelo seco-1).
Tabla 6: Resultados del análisis ANOVA de los valores de C de la biomasa microbiana del suelo en las dos regiones biogeográficas investigadas.
C de la biomasa
microbiana
Fuente de variación g.l. F P
Región 1 15,03 <0,001 Manejo 1 0,35 0,552 Región*Manejo 1 3,21 0,074 Región Atlántica Cultivo 1 9,66 0,001 Manejo 1 4,85 0,012 Tiempo 11 7,62 <0,001 Cultivo*Manejo 1 22,84 <0,001 Tiempo*Cultivo 11 1,36 0,038 Tiempo*Manejo 11 1,34 0,036 Tiempo*Cultivo*Manejo 11 1,58 0,122 Región Mediterránea Cultivo 1 12,55 <0,001 Manejo 1 2,29 0,136 Tiempo 6 3,56 0,005 Cultivo*Manejo 1 2,09 0,154 Tiempo*Cultivo 6 0,54 0,773 Tiempo*Manejo 6 3,01 0,013 Tiempo*Cultivo*Manejo 6 1,48 0,205
ab bc
c
a
0
2
4
6
8
10
12
Mandarino Con Mandarino Org Olivo Con Olivo Org
C/N

Capítulo I
53
Cuando se analizan las dos regiones por separado, la salida de la ANOVA indica que mientras
en la región atlántica los tres factores investigados tuvieron una influencia significativa sobre el
contenido de C de la biomasa microbiana, en la región mediterránea fueron el tipo de cultivo y
tiempo (Tabla 6). Así, en cuanto al factor cultivo en la región mediterránea, las fincas de
mandarino presentaron un mayor contenido en C de la biomasa microbiana en sus suelos
(0,36±0,03 mg de C g de suelo seco-1) que las de olivo (0,21±0,02 mg de C g de suelo seco-1). El
efecto del manejo en la región atlántica dio como resultado que los manejos convencionales
presentaran una mayor biomasa microbiana (0,20±0,01 mg de C g de suelo seco-1) que los suelos
con manejo orgánico (0,16±0,01 mg de C g de suelo seco-1).
En cuanto a las variaciones temporales observadas en la región atlántica, destacan los
elevados valores de C de la biomasa microbiana registrados en los meses de verano (junio, julio
y agosto con 0,30, 0,29 y 0,24 mg de C g de suelo seco-1, respectivamente) cuando se comparan
con los meses de enero y diciembre (0,05 mg de C g de suelo seco-1 en ambos casos). En los
suelos mediterráneos las principales diferencias se observaron entre los meses de marzo y
septiembre, con los valores más altos (0,39±0,27 y 0,36±0,18 mg de C g de suelo seco-1,
respectivamente), y el mes de diciembre, con los más bajos (0,12±0,09 mg de C g de suelo seco-
1).
La interacción tiempo*manejo resultó ser significativa para el contenido de C de la biomasa
microbiana de los suelos de cultivo de ambas regiones (Tabla 6). En la región atlántica, los
valores más elevados se registraron en los suelos con manejo convencional durante el periodo
de junio-septiembre (entre 0,32±0,03 en julio y 0,35±0,05 en junio) en comparación con los
medidos en diciembre de 2013 (0,04±0,02 mg de C g de suelo seco-1) y enero 2014 (0,05±0,03
mg de C g de suelo seco-1). En la región mediterránea sólo se observaron diferencias significativas
entre los valores más altos observados en los suelos orgánicos en el mes de marzo (0,50±0,03
mg de C g de suelo seco-1) y los valores más bajos obtenidos en los suelos convencionales
durante los meses de diciembre y enero (0,12±0,02 y 0,10±0,03 mg de C g de suelo seco-1,
respectivamente).
Al contrario de lo que ocurría con las propiedades químicas, la interacción cultivo*manejo
tuvo un efecto significativo sobre el contenido en C de la biomasa microbiana de los suelos en
la región atlántica (Tabla 6), destacando el cultivo de vid con manejo orgánico con los valores
significativamente más bajos de todos los tratamientos investigados (Fig. 20).

Capítulo I
54
Figura 20: Valores medios (±E.E.) de contenido de C de la biomasa microbiana en los cultivos atlánticos sometidos a diferentes manejos. Letras diferentes indican diferencias significativas (p<0,05) entre tratamientos.
3.4. Relaciones entre las propiedades físicas, químicas y biológicas de los suelos de cultivo
Los resultados del análisis de componentes principales (PCA) del conjunto de datos indican
que los tres primeros componentes explican más de un 85% de la varianza total (Tabla 7). Sin
embargo, el primer componente con un 54% de la varianza total explicada, fue el que resultó
ser significativo (Tabla 7).
Tabla 7: Resultados del PCA de los datos totales de las dos regiones. Se indican en negrita los autovalores de los componentes así como los coeficientes de correlación de Pearson de cada componente con cada variable que resultaron significativos; *(p<0,05), ** (p<0,01) y *** (p<0,001).
VARIABLES PC1 PC2 PC3 PC4
Humedad 0,446*** 0,092 -0,147 -0,172
pH -0,408*** 0,070 -0,440* 0,425
N 0,439*** -0,164 -0,359 0,382
C 0,459*** -0,193 -0,214 0,370
C/N 0,422*** -0,161 0,334 -0,255
Temp. suelo -0,163 -0,682*** 0,490** 0,396
C de la biomasa microbiana 0,160 -0,656*** -0,506** -0,532
Autovalores 3,810 1,202 0,957 0,486
Proporción de la varianza explicada (%) 54,438 17,183 13,684 6,950
Proporción de la varianza acumulada (%) 54,438 71,624 85,308 92,255
La representación gráfica de los datos multivariantes para los dos primeros componentes
(Fig. 21) permite constatar que las diferencias edafoclimáticas que caracterizan a las dos
regiones biogeográficas investigadas son las que mejor explican la distribución de los datos. De
tal forma, que hacia el lado positivo del PC1 se localizan las muestras de los suelos de cultivo de
la región atlántica caracterizadas por presentar los valores más altos de humedad y materia
orgánica (Fig. 21 y Tabla 7). En contraposición las muestras de suelo recogidas en los cultivos
mediterráneos aparecen fuertemente correlacionadas con el pH (Fig. 21 y Tabla 7).
aa
a
b
0
0,1
0,2
0,3
Kiwi Conv. Kiwi Org. Vid Conv. Vid Org.
Bio
mas
a m
ico
rbia
na
(mg
C g
su
elo
sec
o -1
)

Capítulo I
55
Tanto la temperatura del suelo como el C de la biomasa microbiana estuvieron fuertemente
correlacionadas con el segundo componente, que explica tan sólo un 17% de la varianza total, y
por tanto, no permite una buena ordenación de los datos (Fig. 21 y Tabla 7).
Figura 21: Gráfico bidimensional del análisis de componentes principales para las variables edáficas de las muestras de suelo de la región atlántica y mediterránea en función del tipo de manejo.
3.2.2. Análisis de los datos de la región atlántica.
Los resultados del PCA para los datos de los suelos atlánticos indican que los tres primeros
componentes explican entre un 21 y un 34% de la varianza total, siendo los tres significativos
(Tabla 8).
Tabla 8: Resultados del PCA de los datos de la región atlántica. Se indican en negrita los autovalores de los componentes así como los coeficientes de correlación de Pearson de cada componente con cada variable que resultaron significativos; *(p<0,05), ** (p<0,01) y *** (p<0,001).
VARIABLES PC1 PC2 PC3 PC4
Humedad -0,289 0,496*** -0,003 0,583**
pH 0,208 -0,133 -0,715*** 0,062
N -0,497*** 0,072 -0,465*** -0,169
C -0,602*** -0,023 -0,230 -0,208
C/N -0,470*** -0,128 0,466*** -0,181
Temp. suelo -0,054 -0,688*** 0,011 -0,090
C de la biomasa microbiana -0,195 -0,489*** 0,002 0,736***
Autovalores 2,418 1,839 1,498 0,785
Proporción de la varianza explicada (%) 34,546 26,273 21,404 11,215
Proporción de la varianza acumulada (%) 34,546 60,824 82,224 93,440
El gráfico bidimensional de los dos primeros componentes muestra de nuevo que la materia
orgánica y el pH son los factores que mejor explican la variabilidad de los datos (Fig. 22 y Tabla
8) separando el PC1 los suelos en función del manejo agrícola. Así, se puede constatar que los
valores más altos de %C, %N y de la relación C/N y los más bajos de pH correspondieron a los
manejos convencionales y los valores más bajos de estas variables a las muestras recogidas en
PC1
PC2
Mediterráneo con.
Mediterráneo org.
Atlántico con.
Atlántico org.

Capítulo I
56
los cultivos orgánicos (Fig. 22 y Tabla 8). Por otro lado, ni la humedad, ni la temperatura del suelo
o el C de la biomasa microbiana permiten discernir entre cultivos y/o manejos (Fig. 22).
Figura 22: Gráfico bidimensional del análisis de componentes principales para las variables edáficas de las muestras de suelo de la región atlántica en función de tipo de cultivo y manejo.
3.2.3. Análisis de los datos de la región mediterránea
Cuando se analizaron los datos de los suelos de la región mediterránea por separado
mediante PCA, se observó que los dos primeros componentes resultaron ser significativos y
explicaron un 58% de la varianza acumulada (Tabla 9).
Tabla 9: Resultados del PCA de los datos de la región mediterránea. Se indican en negrita los autovalores de los componentes así como los coeficientes de correlación de Pearson de cada componente con cada variable que resultaron significativos; *(p<0,05), ** (p<0,01) y *** (p<0,001).
VARIABLES PC1 PC2 PC3 PC4
Humedad -0,233 0,636*** 0,089 0,356
pH 0,346 0,199 -0,475 -0,310
N -0,563*** 0,062 -0,206 -0,323
C -0,546*** -0,021 -0,179 -0,455
C/N 0,131 -0,271 -0,744*** 0,232
Temp. suelo -0,070 -0,656*** 0,305 -0,068
C de la biomasa microbiana -0,432** -0,214 -0,209 0,637
Autovalores 2,561 1,556 1,230 0,657
Proporción de la varianza explicada (%) 36,595 22,236 17,573 9,387
Proporción de la varianza acumulada (%) 36,595 58,831 76,404 85,791
La visualización gráfica de los resultados para los dos primeros componentes, que explican
un 36 y un 22% de la varianza total (Tabla 9 y Fig. 23), permite observar la clara separación de
los cultivos orgánicos a lo largo del PC1, siendo los de mandarino los que presentan los valores
más altos de C de la biomasa microbiana, %C y %N, frente a las de olivo con valores bajos de
estas variables y altos de pH (Fig. 23). Las propiedades físicas (humedad y la temperatura del
PC1
PC2
Kiwi con.
Kiwi org.
Vid con.
Vid org.

Capítulo I
57
suelo) son las que mejor explican el PC2 (Tabla 9) pero no permitieron una buena ordenación de
los datos (Fig. 23).
Figura 23: Gráfico bidimensional del análisis de componentes principales para las variables edáficas de las muestras de suelo de la región mediterránea en función de tipo de cultivo y manejo.
4. DISCUSIÓN
Los resultados obtenidos tanto mediantes análisis univariantes como multivariantes indican
que la temperatura del suelo ha resultado ser un factor con poca relevancia a la hora de
caracterizar estos suelos de cultivo frutícolas. Este resultado resulta sorprendente ya que la
temperatura tiene un efecto directo sobre la actividad de las comunidades heterotróficas y los
procesos de descomposición de la materia orgánica (e.g. Hood 2001; Sierra et al. 2015).
A diferencia de la temperatura, los análisis de ANOVA y PCA indicaron que la humedad del
suelo es una variable que permite una buena diferenciación entre suelos atlánticos y
mediterráneos, ya que en los primeros el contenido de humedad es dos veces mayor (19 vs. 9%),
lo cual es un reflejo directo de las diferencias climáticas que existen entre estas dos regiones
biogeográficas (Rivas-Martínez et al. 2002). La región atlántica se caracteriza por presentar
inviernos muy húmedos y veranos frescos, con lluvia moderada a lo largo de todo el año,
mientras que la región mediterránea presenta veranos secos y cálidos e inviernos húmedos y
frescos, pero con episodios de aguaceros torrenciales en momentos muy puntuales (EEA 2002).
Sin embargo, el año de estudio no se ajustó demasiado al patrón climático característico de la
región atlántica (Carballeira et al. 1983), ya que si bien en el invierno la precipitación mensual
fue superior a los 300 mm, los meses de verano fueron extremadamente secos (<6 mm en julio);
aún así, en la mayoría de los meses (octubre-abril), la pluviosidad superó los 100 mm mensuales
(MeteoGalicia 2013). En cambio, en la región mediterránea los valores de lluvia fueron inferiores
a los 100 mm mensuales y tan sólo se registraron unos 15 mm en verano (Servei Metereológic
PC1
PC2
Mandarino con.
Mandarino org.
Olivo con.
Olivo org.

Capítulo I
58
de Catalunya 2013). Estas restricciones de agua en la región mediterránea pueden ser mitigadas
mediante una buena gestión del riego en las prácticas agrícolas (Lazzara y Rana 2010). De hecho,
el déficit hídrico en los cítricos mediterráneos supone un factor limitante del crecimiento del
fruto y de la producción (Vélez et al. 2012) y se hace necesario un riego regular, de ahí que los
suelos con manejo convencional presentaran una humedad significativamente mayor que los
orgánicos.
De este modo, en el caso de los suelos atlánticos, las diferentes necesidades hídricas de los
dos cultivos investigados explican las diferencias significativas observadas en el contenido de
agua entre los cultivos de kiwi (más húmedos) y los de vid (más secos). El kiwi es una especie
vegetal con una baja tolerancia a la sequía, ya que presenta un sistema radicular superficial y
por tanto, requiere de un régimen de riego continuado en los meses más cálidos (Otero 2008).
En el caso de la vid, sobre todo cuando su cultivo está destinado a la producción de vino, se
requiere un control riguroso del riego, ya que un riego excesivo o en una época no favorable
podría ocasionar cosechas de baja calidad (Acevedo et al. 2005). La mayor humedad observada
en el cultivo de vid convencional respecto al orgánico puede ser debido al mayor control del
riego en el sistema intensivo.
De forma similar, en la región mediterránea los suelos de mandarino presentaron valores de
humedad significativamente superiores a los de olivo, también debido a las diferentes
necesidades hídricas de estos dos cultivos. El olivo es un cultivo resistente a la sequía (Samish y
Spiegel 1961; Lavee et al. 1990) y por tanto, tiene requerimientos de riego mucho más bajos que
el mandarino. Sin embargo, las condiciones climáticas de la región mediterránea, sobre todo en
los meses estivales, pueden provocar condiciones estresantes en las etapas fenológicas más
sensibles del cultivo, reduciendo el rendimiento de los mismos (Rainey y Griffiths 2005), por lo
que se hace necesario un control minucioso del riego.
Los resultados de los análisis estadísticos también revelaron que el pH y la materia orgánica
explican una gran proporción de la varianza asociada a los datos obtenidos en este estudio. Los
valores de pH del suelo medidos en las 8 fincas son los esperados en base a la litología de las
regiones investigadas. El 88% de la superficie de la región gallega está ocupada por rocas ácidas
(Macías y Calvo de Anta 2001) y la acidez de los suelos atlánticos obliga a los agricultores a aplicar
periódicamente enmiendas calcáreas con el fin de mantener un pH adecuado para el desarrollo
del cultivo. Pero, además, en ambas regiones existen causas antrópicas asociadas al manejo
agrícola que producen cambios en el pH de los suelos, como son el uso de fertilizantes minerales
en cultivos que tienden a incrementar la acidez de los suelos (Kibblewhite et al. 2008). En los
cultivos convencionales de ambas regiones se emplearon fertilizantes nitrogenados y
presentaron valores inferiores de pH a los medidos en los suelos abonados con fertilizantes

Capítulo I
59
orgánicos. Un pH ácido en el suelo puede llegar a producir una movilización de metales pesados
en el suelo y alcanzar concentraciones tóxicas para las especies vegetales (Sacristán et al. 2015)
y en consecuencia, alterar la capacidad de las comunidades microbiológicas del suelo para
transformar la materia orgánica y el reciclado de nutrientes (Kemmitt et al. 2006). Además, en
el caso concreto de los suelos de Galicia, valores bajos de pH del suelo favorecen la movilización
del aluminio que causa toxicidad a los cultivos (Novoa Muñoz et al. 2012).
El contenido de materia orgánica (%C, %N y relación C/N) también resultaron ser variables
que permitieron una buena diferenciación de los suelos de cultivo de ambas regiones. Dado que
los procesos de descomposición y reciclaje de nutrientes de un suelo dependen de la naturaleza
de la vegetación y de los patrones de precipitación y de temperatura (Gupta y Rao 1994), es
previsible que existan diferencias en cuanto a la capacidad de retención de C y tasas de
mineralización entre las dos regiones biogeográficas. De acuerdo con esto, los mayores
contenidos de C se midieron en los suelos atlánticos comparado con los mediterráneos. Estos
resultados coinciden con las observaciones de Burke et al. (1989) que constataron que el C del
suelo aumenta con la precipitación y desciende con la temperatura. De este modo, las
condiciones climáticas que caracterizan a la región atlántica son un reflejo del diferente
porcentaje de materia orgánica presente en los suelos. Además, el manejo agrícola también
influye en la capacidad de retención de C de un suelo. En general, se sabe que las prácticas
agrícolas orgánicas tienden a incrementar el contenido de C de los suelos de cultivo (Azeez
2009). Sin embargo, en la región atlántica, el contenido de C de los suelos convencionales fue
muy superior al medido en los suelos orgánicos, lo que podría atribuirse al uso anterior de las
fincas convencionales (forestal), que favorecen el secuestro de C (Lal 2005), frente a la
producción de flor (uso extractivo) en el caso de las fincas orgánicas y que tiende a reducir las
reservas de C; pese a ello, estas fincas mantienen unos niveles adecuados de C en el suelo. En el
caso de las fincas mediterráneas, se observa una interacción importante entre tipo de cultivo y
manejo y los mayores valores de contenido de C fueron registrados en los cultivos de mandarino
orgánico, posiblemente por un efecto sinérgico de la humedad y el uso de fertilizantes orgánicos
(Blanco-Canqui et al. 2011; Palm et al. 2014).
Los suelos de cultivo de la región atlántica también presentaron mayores contenidos de N
que los de la región mediterránea, lo que puede atribuirse al hecho de que los procesos de
nitrificación se ven favorecidos en suelos húmedos (Stark y Firestone 1995; Paz-Ferreiro et al.
2010). En cambio, en la región mediterránea, tanto el tipo de cultivo como el manejo agrícola
tuvieron un efecto significativo sobre el contenido en N del suelo, siendo de nuevo los suelos de
mandarino sometidos a manejo orgánico los que presentaron la mayor cantidad de N total. Al
igual que en el caso anterior, esto podría estar relacionado con la humedad del suelo

Capítulo I
60
(significativamente superior en el mandarino) y al uso de de fertilizantes orgánicos que
aumentan el contenido de N del suelo (Birkhofer et al. 2008). Sin embargo, en el caso del olivar,
los resultados del PCA indican que los suelos de este cultivo sometidos a manejo orgánico se
caracterizan por los menores contenidos de N en el suelo, lo cual puede estar relacionado con
el pobre contenido de N de la hojarasca de olivo (Kherbouche et al. 2012) junto con los valores
bajos de humedad que caracterizan a estos cultivos y que hayan podido comprometer los
procesos de nitrificación en estos suelos (Paz-Ferreiro et al. 2010).
Por último, la relación C/N se emplea como un indicador de la calidad de la materia orgánica
en los ecosistemas aerobios y determina la tasa de disponibilidad de N para las plantas (Porta et
al. 2003). El valor de esta relación desciende a medida que la materia orgánica fresca se va
mineralizando y por tanto, los valores más elevados registrados en la región atlántica (por
encima de 13) que los de la región mediterránea (por debajo de 10) indican que los primeros
presentan un grado menor de humificación de la materia orgánica que los de la región
mediterránea y también que ambos valores están fuera del rango óptimo recomendado para
suelos agrícolas (10-12; Porta et al. 2003). Al igual que en el caso del C y N, el tipo de cultivo y el
manejo agrícola también tuvieron un efecto significativo sobre la relación C/N de los suelos de
cultivo de ambas regiones. En el caso de la región atlántica, los cultivos orgánicos presentan
valores de C/N menores que los manejos convencionales, debido al bajo contenido en C de los
primeros y relacionado (como se indicó anteriormente) con el uso anterior del suelo. En cambio,
en la región mediterránea, la relación C/N más alta observada en los suelos de cultivo de olivo
producidos con ambos tipos de manejo es probablemente el resultado de la suma de una baja
calidad de la hojarasca de este árbol (Kherbouche et al. 2012) y con unas condiciones edáficas
más secas que hacen que los procesos de mineralización se vean ralentizados (Paz-Ferreiro et
al. 2010; Hu et al. 2014; Bastida et al. 2017).
Dado que el C de la biomasa microbiana está muy influenciado por las condiciones climáticas
(Insam 1990; Insam et al. 1989; Guicharnaud et al. 2010) y por los contenidos de C y N del suelo
(Wardle 1992), esta variable también mostró diferencias importantes entre las dos regiones
geográficas estudiadas. La mayor actividad microbiológica fue observada en los suelos
mediterráneos, lo que no se explica por un mayor contenido orgánico de los suelos y en cambio,
podría estar relacionado con temperaturas más cálidas que tienden a estimular la actividad
microbiana (Insam et al. 1989; Insam 1990). Sin embargo, las diferencias entre las temperaturas
edáficas entre las dos regiones no fueron significativas, por lo que son otros factores los que
pueden haber tenido una mayor influencia sobre la actividad biológica de estos suelos de cultivo.
Así, un pH menos ácido en los suelos mediterráneos comparado con los atlánticos puede haber
favorecido la actividad microbiana, lo que coincide con estudios previos (Vance et al. 1987;

Capítulo I
61
Sparling y West 1988; Anderson y Joergensen 1997; Aciego Pietri y Brooks 2005). Además, en el
caso de esta región, el tipo de cultivo también tuvo un efecto significativo sobre este parámetro,
siendo menor en los olivares, lo que podría ser debido, como se comentó anteriormente, a la
baja humedad registrada en estos suelos (Orchard y Cook 1983; Stark y Firestone 1995; Averill
et al. 2016) junto con el elevado contenido en compuestos fenólicos, especialmente
oleuropeína, con propiedades antimicrobianas y antifúngicas de la hojarasca de olivo (Juven et
al. 1972; Aziz et al. 1998; Pereira et al. 2007), los cuales podrían haber resultado perjudiciales
para las actividades de las poblaciones microbianas en estos suelos. En cambio en la región
atlántica, el tipo de manejo más que el tipo de cultivo parece tener un efecto más importante
sobre el C de la biomasa microbiana, destacando los manejos convencionales por los valores
más altos de biomasa microbiana. Es posible que los contenidos significativamente mayores de
C y temperaturas de suelo más cálidas hayan proporcionado condiciones más favorables para la
actividad microbiológica en estos suelos con manejo convencional (Diacono y Montemurro
2010).
5. CONCLUSIONES
La humedad edáfica resultó ser la principal variable diferenciadora entre los suelos atlánticos
y mediterráneos, destacando los valores más elevados en los primeros y por tanto,
condicionando la frecuencia del riego en función de las necesidades hídricas de los cultivos con
mayor requerimiento de agua de cada región (kiwi en atlántico y mandarino en mediterráneo).
Por otro lado, la litología del terreno condiciona la movilidad de los cationes y por tanto, el pH
se convierte en el segundo factor edáfico que mejor diferencia las dos regiones, de modo que
los pH de los suelos de la región mediterránea son más altos que los atlánticos desarrollados
sobre rocas graníticas. Sin embargo, el ambiente edafoquímico de los suelos de cultivo se ve
alterado por el tipo de cultivo y de manejo agrícola implementado para asegurar el buen
crecimiento de la planta cultivada. Por tanto, además del riego, el uso de fertilizantes minerales
y la retirada o no de los restos de poda o de la cosecha pueden modificar la capacidad de un
suelo de cultivo para retener humedad y cationes. Así, el viñedo y sobre todo el olivar no reciben
riegos frecuentes y se suele eliminar la vegetación superficial entre árboles por lo que el estrés
hídrico es mayor.
Unos suelos más húmedos y más ácidos limitan las actividades de las poblaciones
microbianas lo que ralentiza los procesos de descomposición de la materia orgánica y favorece
la retención de nutrientes en el suelo (C y N). Como consecuencia de esto, los manejos orgánicos
tienden a favorecer la retencion de C en el suelo frente a los convencionales, con lo que estos

Capítulo I
62
últimos tienden a perder materia orgánica y obliga a realizar más aportes para compensar las
pérdidas. Sin embargo, la magnitud de esta respuesta dependerá en gran medida de las reservas
iniciales del suelo y cuánto tiempo lleva sometido el suelo a un manejo convencional.
En vista de todo ello, se puede concluir que aunque las condiciones climáticas de una
determinada región condicionan las propiedades edafoquímicas de los suelos y las actividades
de las comunidades microbianas, éstas pueden verse modificadas por el manejo agrícola y junto
con la calidad de los aportes de material vegetal determinarán finalmente las tasas de
descomposición de los mismos.
6. BIBLIOGRAFÍA
Acevedo C, Ortega-Farías S, Hidalgo C, Moreno Y y Córdova F. 2005. Efecto de diferentes niveles de agua aplicada en postcuaja y en postpinta sobre la calidad del vino cv. Cabernet Sauvignon. Agricultura Técnica, 65(4), 397-410.
Aciego Pietri J C y Brookes P C. 2005. Relationships between soil pH and microbial properties in a UK arable soil. Soil Biology & Biochemistry, 40, 1856-1861.
Allison S D, Lu Y, Weihe C, Goulden M L, Martiny A C, Treseder K K y Martiny J B. 2013. Microbial abundance and composition influence litter decomposition response to environmental change. Ecology, 94(3), 714-725.
Anderson T H y Joergensen R G, 1997. Relationship between SIR and FE estimates of microbial biomass C in deciduous forest soils at different pH. Soil Biology & Biochemistry, 29, 1033–1042.
Averill C, Waring B G y Hawkes C V. 2016. Historical precipitation predictably alters the shape and magnitude of microbial functional response to soil moisture. Global Change Biology, 22(5), 1957-1964.
Azeez G. 2009. Results of the comparative studies on the soil carbon levels of organic farming. En: Azeez G (Eds.) Soil Carbon and Organic Farming, pp. 44-68. Soil Association, London.
Aziz N H, Farag S E, Mousa L A y Abo-Zaid M A. 1998. Comparative antibacterial and antifungal effects of some phenolic compounds. Microbios, 93(374), 43-54.
Bastida F, Torres I F, Romero-T igue os C, Bald ia P, Vět o ský T, Ba o a J M, Alarcón J J, Hernández T, García C y Nicolás E. 2017. Combined effects of reduced irrigation and water quality on the soil microbial community of a citrus orchard under semi-arid conditions. Soil Biology & Biochemistry, 104, 226-237.
Beare M H, Parmelee W, Hendrix P F y Cheng W. 1992. Microbial and faunal interactions and effects on litter nitrogen and decomposition in agroecosystems. Ecological Monographs, 62(4), 569-591.
Belay A, Claassens A S y Wehner F C. 2002. Effect of direct nitrogen and potassium and residual phosphorus fertilizers on soil chemical properties, microbial components and maize yield under long-term crop rotation. Biology & Fertility of Soils, 35, 420-427.
Bender S F, Wagg C y van der Heijden M G. 2016. An underground revolution: biodiversity and soil ecological engineering for agricultural sustainability. Trends in Ecology & Evolution, 31(6), 440-452.
Birkhofer K, Bezemer T M, Bloem J, Bonkowski M, Christensen S, Dubois D, Ekelund F, Fliebach A, Gunst L, Hedlund K, Mäder P, Mikola J, Robin C, Setälä H, Tatin-Froux F, van der Putten W y Scheu S. 2008. Long-term organic farming fosters below and aboveground biota: implications for soil quality, biological control and productivity. Soil Biology & Biochemistry, 40(9), 2297-2308.
Blanco-Canqui H, Schlegel A J y Heer W F. 2011. Soil-profile distribution of carbon and associated properties in no-till along a precipitation gradient in the central great plains. Agriculture, Ecosystem & Environment, 144, 107-116.
Bossio D A, Scow K M, Gunapala N y Graham K J. 1998. Determinants of soil microbial communities: effects of agricultural management, season, and soil type on phospholipid fatty acid profiles. Microbial Ecology, 36(1), 1-12.
Brake M, Höper H y Joergensen R G. 1999. Land use-induced changes in activity and biomass of microorganisms in raised bog peats at different depths. Soil Biology & Biochemistry, 31, 1489-1497.

Capítulo I
63
Burke I C, Yonker C M, Parton W J, Cole C V, Schimel D S y Flach K. 1989. Texture, climate, and cultivation effects on soil organic matter content in US grassland soils. Soil Science Society of America Journal, 53(3), 800-805.
Carballeira A, Devesa C, Retuerto R, Santillán E y Uceda F. 1983. Bioclimatología de Galicia. Fundación Pedro Barrié de la Maza, Conde de Fenosa, A Coruña.
Christensen B T. 1996. Matching measurable soil organic matter fractions with conceptual pools in simulation models of carbon turnover: revision of model structure. En: Powlson D S, Smith P, Smith y J U (Eds.) Evaluation of Soil Organic Matter Models Using Existing Long-Term Datasets, pp. 143-159. Global Environmental Change. Springer, Berlin.
Curiel Yuste J, Baldocchi D D, Gershenson A, Goldstein A, Misson L y Wong S. 2007. Microbial soil respiration and its dependency on carbon inputs, soil temperature and moisture. Global Change Biology, 13(9), 2018-2035.
Diacono M y Montemurro F. 2010. Long-term effects of organic amendments on soil fertility. A review. Agronomy for Sustainable Development, 30, 401-422.
EEA (European Environment Agency) 2012. Biogeographical regions. Revisión 1 (http://www.eea.europa.eu/data-and-maps/figures/biogeographical-regions-in-europe-1).
Fierer N y Jackson R B. 2006. The diversity and biogeography of soil bacterial communities. Proceedings of the National Academy of Sciences of the United States of America, 103(3), 626-631.
Franzluebbers A J, Hons F M y Zuberer D A. 1995. Tillage and crop effects on seasonal dynamics of soil co 2 evolution, water content, temperature, and bulk density. Applied Soil Ecology, 2(2), 95-109.
García-Serrano P y Ruano S. 2010. Guía Práctica de la Fertilización Racional de los Cultivos en España. MARM.
Gkisakis V, Volakakis N, Kollaros D, Bàrberi P y Kabourakis E M. 2016. Soil arthropod community in the olive agroecosystem: determined by environment and farming practices in different management systems and agroecological zones. Agriculture, Ecosystems & Environment, 218, 178-189.
Guicharnaud R A, Arnalds O y Paton G I. 2010. The effect of season and management practices on soil microbial activities undergoing nitrogen treatments - interpretation from microcosm to field scale. Iceland Agricultural Sciences, 23, 123-134.
Guo L B y Gifford R M. 2002. Soil carbon stocks and land use change: a meta analysis. Global Change Biology, 8, 345-360.
Gupta R K y Rao D L N. 1994. Potential of wasteland for sequestring carbon by deforestation. Current Science, 66, 378-3801.
Guzmán G C y Alonso M. 2004. Manual De Olivicultura Ecológica. Instituto de Sociología y Estudios Campesinos. Universidad de Córdoba. Córdoba.
Hammer ∅, Harper D A T y Ryan P D. 2001. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Paleontologia Electronica, 4, 1-9.
Hood R C. 2001. The effect of soil temperature and moisture on organic matter decomposition and plant growth. Isotopes in Environmental & Health Studies, 37(1), 25-41.
Hu R, Wang X P, Pan Y X, Zhang Y F y Zhang H. 2014. The response mechanisms of soil n mineralization under biological soil crusts to temperature and moisture in temperate desert regions. European Journal of Soil Biology, 62, 66-73.
Insam H. 1990. Are the soil microbial biomass and basal respiration governed by the climatic regime? Soil Biology & Biochemistry, 22(4), 525-532.
Insam H, Parkinson D y Domsch K H. 1989. Influence of macroclimate on soil microbial biomass. Soil Biology & Biochemistry, 21, 211-221.
Insam H, Mitchel C C y Dormaar J F. 1991. Relationship of soil microbial biomass and activity with fertilization practice and crop yield of the three ultisols. Soil Biology & Biochemistry, 23(5), 459-464.
Jackson D A. 1993. Stopping rules in principal components analysis: a comparison of heuristical and statistical approaches. Ecology, 74, 2204-2214.
Juven B, Henis Y y Jacoby B. 1972. Studies on the mechanism of the anti-microbial action of oleuropein. Journal of Applied Bacteriology, 35, 559-567.
Kautz T, López-Fando C y Ellmer F. 2006. Abundance and biodiversity of soil microarthropods as influenced by different types of organic manure in a long-term field experiment in central Spain. Applied Soil Ecology, 33(3), 278-285.
Kemmitt S J, Wright D, Goulding K W T y Jones D L. 2006. pH regulation of carbon and nitrogen dynamics in two agricultural soils. Soil Biology & Biochemistry, 38, 898–911.

Capítulo I
64
Kherbouche D, Bernhard-Reversat F, Moali A y Lavelle P. 2012. The effect of crops and farming practices on earthworm communities in Soummam valley, Algeria. European Journal of Soil Biology, 48, 17-23.
Kibblewhite M G, Ritz K y Swift M J. 2008. Soil health in agricultural systems. Philosophical Transactions of the Royal Society of London Biological Sciences, 363(1492), 685-701.
Kirschbaum M U. 1995. The temperature dependence of soil organic matter decomposition, and the effect of global warming on soil organic c storage. Soil Biology & Biochemistry, 27(6), 753-760.
Kochian L V, Hoekenga O A y Piñeros M A. 2004. How do crop plants tolerate acid soils? Mechanisms of aluminium tolerance and phosphorous efficiency. Annual Review of Plan Biology, 55, 549-493.
Lal R. 2005. Forests soils and carbon sequestration. Forest Ecology & Management 220: 242-258. Lavee S, Nashef M, Wodner M y Harshemesh H. 1990. The effect ofcomplementary irrigation added to old
olive trees (Olea europaea L.) cv. Souri on fruit characteristics, yield and oil production. Advances in Horticultural Science, 4, 135–138.
Lazzara P y Rana G. 2010. The crop coefficient (Kc) values of the major crops grown under mediterranean climate. Italian Journal of Agrometeorology, 15(2), 25- 40.
LeBauer D S y Treseder K K. 2008. Nitrogen limitation of net primary productivity in terrestrial ecosystems is globally distributed. Ecology, 89, 371-379.
Letey J. 1985. Relationship between soil physical properties and crop production. En: Stewart B A (Eds.) Advances in Soil Science, pp. 277-294. Springer-Verlag New York Inc, New York.
Licker R, Johnston M, Foley JA, Barford C, Kucharik CJ, Monfreda C y Ramankutty N. 2010. Mind the gap: ho do li ate a d ag i ultu al a age e t e plai the ield gap of oplands around the world? Global Ecology & Biogeography, 19(6), 769-782.
Lindsey K y Jones M G K. 1991. Biotecnología Vegetal Agrícola. Ed. Acribia. Madrid. Lobell D B y Field C B. 2007. Global scale climate–crop yield relationships and the impacts of recent
warming. Environmental Research Letters, 2(1), 014002. Macías F y Calvo de Anta R. 2001. Los suelos. En: Atlas de Galicia. Sociedade para o Desenvolvemento
Comarcal de Galicia, pp 173-218. Xunta de Galicia. MeteoGalicia 2013. Xunta de Galicia. Disponible en: www.meteogalicia.es. [Acceso en marzo 2015]. Nóvoa-Muñoz J C, Taboada M T y García-Rodeja E. 2002. Influencia del contenido y formas de al extraíble
en los mecanismos de neutralización de ácidos de horizontes úmbricos. Edafología, 9(2), 115-128. Orchard V A y Cook F J. 1983. Relationship between soil respiration and soil moisture. Soil Biology &
Biochemistry, 15(4), 447-453. Otero V. 2008. Cultivo del kiwi: técnicas aplicadas a la mejora de la producción y conservación. (Tesis
Doctoral). Universidad de Vigo, Vigo. Palm C, Blanco-Canqui H, DeClerck F, Gatere L y Grace P. 2014. Conservation agriculture and ecosystem
services: an overview. Agriculture, Ecosystems & Environment, 187, 87-105. Pampulha M E y Oliveira A. 2006. Impact of an herbicide combination of bromoxynil and prosulfuron on
soil microorganisms. Current Microbiology, 53, 238–243. Paz-Ferreiro J, Trasar-Cepeda C, Leirós M C, Seoane S y Gil-Sotres F. 2010. Effect of management and
climate on biochemical properties of grassland soils from Galicia (NW Spain). European Journal of Soil Biology, 46, 136-143.
Pereira A P, Ferreira I C, Marcelino F, Valentão P, Andrade P B, Seabra R, Estevinho L, Bento A y Pereira J A. 2007. Phenolic compounds and antimicrobial activity of olive (Olea europaea L. cv. Cobrançosa) leaves. Molecules, 12(5), 1153-1162.
Porta J, López-Acevedo M y Roquero C. 2003. Edafología para la Agricultura y el Medio Ambiente. Mundi-Prensa, Madrid.
Power A G. 2010. Ecosystem Services and agriculture: tradeoffs and synergies. Philosophical Transactions of the Royal Society Biological Sciences, 365, 2959-2971.
Rainey K M y Griffiths P D. 2005. Differential response of common bean genotypes to high temperature. Journal of the American Society for Horticultural Science, 130 (1), 18-23.
Ritchie J T y NeSmith D S. 1991. Temperature and crop development. En: Hanks J y Ritchie JT (Eds.) Modeling Plant and Soil Systems, pp. 5-29. Agronomy Monograph, Madison, WI.
Rivas-Martínez S, Díaz T E, Fernández-González F, Izco J, Loidi J, Lousa M y Penas A. 2002. Vascular plant communities of Spain and Portugal. Addenda to the syntaxonomical checklist of 2001. Part I. Itinera Geobotanica, 15, 5-432.
Robertson G P y Vitousek P M. 2009. Nitrogen in agriculture: balancing the cost of an essential resource. Annual Review of Environment & Resources, 34, 97-125.

Capítulo I
65
Sacristán D, Peñarroya B y Recatalá L. 2015. Increasing the knowledge on the management of cu-contaminated agricultural soils by cropping tomato (Solanum lycopersicum L.). Land Degradation & Development, 26, 587-595.
Samish R y Spiegel P. 1961. The use of irrigation in growing olives for oil production. Israel Journal of Agricultural Research, 11, 87-95.
Servei Metereológic de Catalunya. 2013. Disponible en: www.meteo.cat. [Acceso en marzo 2015]. Sierra C A, Trumbore S E, Davidson E A, Vicca S y Janssens I. 2015. Sensitivity of decomposition rates of
soil organic matter with respect to simultaneous changes in temperature and moisture. Journal of Advances in Modeling Earth Systems, 7(1), 335-356.
Souza Andrade D, Colozzi-Filho A y Giller K E. 2003. The soil microbial community and soil tillage. En: El Titi A (Ed.) Soil Tillage in Agroecosystems. CRC Press, Boca Raton FL.
Sparling G P. 1985. The soil biomass. En: Vaughn D y Malcolm RE (Eds.) Soil Organic Matter and Biological Activity, pp 223-262. The Macaulay Institute for Soil Research, Aberdeen.
Sparling G P y West A W, 1988. A direct extraction method to estimate soil microbial C: calibration in situ using microbial respiration and 14C labelled cells. Soil Biology & Biochemistry, 20, 337–343.
Stark J M y Firestone M K. 1995. Mechanisms for soil moisture effects on activity of nitrifying bacteria. Applied & Environmental Microbiology, 61(1), 218-221.
StatSoft, Inc. 2005. STATISTICA, Version 7.1. StatSoft, Inc. Tulsa. Urbano Terrón P, Cornejo J y Cerdá A. 1998. Fertilidad y ciclos de los nutrientes en el suelo. En: Jiménez
Díaz R M y Lamo de Espinosa J (Eds.) Agricultura Sostenible, pp. 145-172. Ediciones Mundi-Prensa. Madrid.
Vance E D, Brookes P C y Jenkinson D S. 1987. An extraction method for measuring soil microbial C. Soil Biology & Biochemistry, 19, 703-708.
Vélez J E, Álvarez-Herrera J G y Alvarado-Sanabria O H. 2012. El estrés hídrico en cítricos (Citrus spp.): una revisión. Orinoquia, 16(2), 32-39.
von Uexküll H R y Mutert E. 1995. Global extent, development and economic impact of acid soils. Plant & Soil, 171, 1-15.
Wardle D A. 1992. A comparative assessment of factors which influence microbial biomass carbon and nitrogen levels in soil. Biological Reviews, 67, 321-358.
Wardle D A. 1995. Impacts of disturbance on detritus food webs in agro-ecosystems of contrasting tillage and weed management practices. Advances in Ecological Research, 26, 105-185.
Wu J, Joergensen R G, Pommerening B. 1990. Measurement of soil microbial biomass c by fumigation-extraction an automated procedure. Soil Biology & Biochemistry, 22, 1167-1169.


CAPÍTULO II:
Composición y estructura de las comunidades edáficas en los agrosistemas atlánticos y
mediterráneos


Capítulo II
69
1. INTRODUCCIÓN
Las comunidades edáficas son muy sensibles al tipo de manejo agrícola que se aplica en los
cultivos debido a su reducida movilidad, lo que les incapacita para desplazarse a zonas dónde
las condiciones ambientales sean más favorables (Eggleton et al. 2009; Blasi et al. 2013; Bardgett
y van der Putten 2014). Esta sensibilidad a las perturbaciones ambientales ha llevado a varios
investigadores a considerar a distintos taxones de la fauna edáfica como uno de los indicadores
más fiables del estado de salud de los ecosistemas (Parisi et al. 2005; Parisi y Menta 2008; Yan
et al. 2012; Paoletti et al. 2013).
Los invertebrados edáficos de mayor importancia, tanto en sistemas naturales como en
sistemas agrícolas, en términos de abundancia y/o biomasa, son los ácaros, colémbolos, los
enquitreidos y dentro de la macrofauna, las lombrices de tierra (Gardi et al. 2009).
Entre los ácaros, los representantes del suborden Oribatida suelen ser el grupo
numéricamente dominante en los sistemas edáficos (Norton 1990). Están considerados como
resistentes a las perturbaciones ambientales debido a que presentan bajas tasas metabólicas,
un desarrollo lento y reducida fecundidad (Behan-Pelletier 1999). Por ejemplo, Khalil et al.
(1999) encontraron un efecto positivo en la abundancia al convertir el uso del suelo de cultivos
anuales a cultivos de árboles frutales. En el caso concreto de olivares, los oribátidos pueden
llegar a ser el segundo taxón más abundante (después de los colémbolos) incluso cuando no se
emplean las técnicas más apropiadas para extraer artrópodos edáficos (Gonçalves y Pereira
2012). A pesar de su gran resiliencia, en algunos casos el manejo agrícola puede tener una gran
influencia sobre el tamaño de sus poblaciones y así Gagnarli et al. (2015) observaron que las
abundancias de estos ácaros eran dos veces mayor en viñedos orgánicos que en aquellos
cultivados intensivamente, donde el empleo de diversos pesticidas (hidróxido de cobre, sulfato
de cobre, piretroides y clorpirifós entre otros) podría haber incrementado las tasas de
mortalidad de estos organismos. Por el contrario, en estudios previos realizados en los mismos
cultivos de kiwi que el presente trabajo, Lago (2015) describió que las poblaciones de oribátidos
en los suelos con manejo convencional son mucho más numerosas que las habitan los cultivos
con manejo orgánico.
La diversidad de los hábitos alimenticios de este grupo (Behan-Pelletier 1999), su mayor
grado de esclerotización y tolerancia a valores de pH bajos podrían haberles otorgado ventajas,
frente a otros grupos de ácaros, para sobrevivir en los sistemas convencionales (Lago 2015). De
hecho, Salminen et al. (1997) demostraron en experimentos en mesocosmos que la cutícula más
esclerotizada de los oribátidos les confiere una mayor resistencia frente a la aplicación de
herbicidas que a los Mesostigmata (ácaros depredadores) con un exoesqueleto menos

Capítulo II
70
endurecido. En otros casos, las respuestas están relacionadas con cambios en la disponibilidad
de alimento. Así, Cao et al. (2011) estudiaron el efecto de fertilizantes orgánicos y minerales
sobre comunidades de ácaros a largo plazo y observaron una reducción en la abundancia y
diversidad de oribátidos, lo cual fue atribuido al elevado contenido en P de ambos fertilizantes,
ya que provocó una reducción en la abundancia de hongos del suelo de los cuales se alimentan
varias especies de estos ácaros.
Los colémbolos son también muy sensibles a las prácticas agrícolas y Sousa et al. (2004)
comprobaron que tanto la abundancia como la diversidad se reducen de forma importante en
los suelos de cultivo, comparado con otros usos del suelo menos intensivos como el forestal. De
forma similar, Filser et al. (1995) observaron una menor diversidad de colémbolos en suelos con
manejos agrícolas intensivos comparados con cultivos de rotación, reflejando el carácter
bioindicador de este grupo taxonómico. Una reducción en la disponibilidad de alimento parece
ser uno de los factores principales que mejor explican estos cambios en la estructura de sus
poblaciones cuando se cambia el manejo (Wardle et al. 2001). En este mismo sentido, Doles et
al. (2001) indican que la menor abundancia de colémbolos en cultivos de frutales con manejos
convencionales comparado con manejos agrícolas orgánicos se debe al mayor número de
recursos alimenticios que proporciona el abonado orgánico. Bedano et al. (2006) también
observaron una influencia negativa de manejos agrícolas convencionales en cultivos de
herbáceas sobre las poblaciones de colémbolos, destacando sus efectos perjudiciales sobre la
fertilidad del suelo. A pesar de estas evidencias, en otros estudios no se han observado apenas
diferencias entre las poblaciones de colémbolos en suelos de cultivo sometidos a distintos
sistemas de manejo (Czarnecki y Paprocki 1997; Yeates et al. 1997; Alvarez et al. 2001; Hole et
al. 2005). Esto puede ser debido a condiciones ambientales adversas para los colémbolos (por
ejemplo, condiciones de sequía en el suelo) que perjudicaron por igual a las poblaciones de
ambos manejos.
En general, los enquitreidos son el grupo de fauna edáfica menos estudiado en los suelos
agrícolas, debido a la dificultad que entraña su identificación taxonómica (Schmelz y Collado
2010). Además, ciertos factores como el clima, la región geográfica o el hábitat tienen una gran
influencia a la hora de regular el tamaño de sus poblaciones (revisado por Briones et al. 2007).
Sin embargo, son también muy sensibles a las prácticas agrícolas (Pelosi y Römbke 2016) y así,
por ejemplo, el uso de pesticidas en manejos convencionales han demostrado ser especialmente
dañino para sus poblaciones, agravado en los casos en los que se usan varios agroquímicos de
forma combinada (Karnatak et al. 2007). En cuanto al uso de fertilizantes, estudios como el de
Mitchell et al. (1978) y Andren y Lagerlöf (1983) destacan el efecto beneficioso de los

Capítulo II
71
fertilizantes orgánicos para sus poblaciones. En cambio, los estudios realizados sobre el efecto
de los fertilizantes minerales sobre los enquitreidos muestran resultados muy variados, tanto
positivos (De Goede et al. 2003) como negativos (Birkhofer et al. 2008) o incluso la ausencia de
efectos medibles (Lagerlöf et al. 1989; Vavoulidou et al. 2009; Payne et al. 2012).
Dentro de la macrofauna, las lombrices representan la mayor biomasa de invertebrados en
los suelos agrícolas (Lee 1985; Wardle 1995; Lavelle et al. 1997; Paoletti et al. 2013). Además,
los estudios previos indican que, de las tres categorías ecológicas en la que tradicionalmente se
clasifican (Bouché 1972), las lombrices epigeas son las que juegan el papel menos importante
en los ecosistemas agrícolas (Dinter et al. 2013), ya que suelen ser menos abundantes y aportan
la menor biomasa debido a su pequeño tamaño. Esto puede ser debido a que habitan las capas
más superficiales del suelo donde están más expuestas a las alteraciones físicas y químicas
provocadas por las retiradas de la cubierta vegetal, aplicaciones de productos agroquímicos, etc.
(Fragoso et al. 1999; Bertrand et al. 2015). De hecho, Paoletti (1999) concluyó que los pesticidas
tienen un efecto más perjudicial sobre las lombrices epigeas que sobre otras categorías
ecológicas. Sin embargo Pelosi et al. (2009) no encontraron diferencias significativas en cuanto
a riqueza de especies, abundancia o biomasa de lombrices epigeas entre manejos
convencionales y orgánicos. Una situación interesante es la descrita por Lago (2015) en cultivos
de kiwi, donde las abundancias de lombrices epigeas fueron menores a las del resto de
categorías ecológicas, pero mayores en el sistema convencional que en el orgánico, lo que fue
explicado por diferencias en el contenido de humedad del suelo, la relación C/N y la capacidad
de intercambio catiónico efectiva entre ambos manejos. En otros estudios, las mayores
abundancias de este grupo de lombrices se contabilizaron en sistemas orgánicos (Henneron et
al. 2015) atribuido al uso de fertilizantes ecológicos que constituyen una fuente de alimento
para estas especies.
La actividad excavadora vertical de las lombrices anécicas les permite poder refugiarse en
capas más profundas del perfil del suelo; sin embargo, se alimentan en superficie (Bouché 1972)
por lo que también son muy sensibles al manejo agrícola y se han registrado mortalidades
elevadas cuando aumenta la intensificación del uso del suelo (Lavelle et al. 1997; Briones et al.
2011). Sin embargo, en otros estudios apenas se vieron afectadas por las actividades agrícolas
(Pelosi et al. 2009; Schreck et al. 2012) y no se encontraron diferencias entre las abundancias de
este grupo de lombrices entre manejos convencionales y orgánicos. Sí se ha observado que el
uso de estiércol de ganado como fertilizante incrementa la abundancia de las poblaciones de
lombrices anécicas (Zaller y Köpke 2004), mientras que el uso de herbicidas perjudica a sus
poblaciones (Bertrand et al. 2015).

Capítulo II
72
Las lombrices endogeas construyen sus galerías horizontales en los horizontes minerales, por
lo que al realizar todas sus actividades en capas más profundas del perfil se encuentran más
protegidas frente a los efectos perjudiciales de las prácticas agrícolas (Wardle 1995; Lagerlöf et
al. 2002; van Eekeren et al. 2008; Pelosi et al. 2009). De hecho, se ha constatado que son las que
mejor soportan las agresiones debidas al manejo agrícola, incluso el uso de pesticidas y altas
concentraciones de metales pesados en el suelo (Paoletti 1999), lo que llevó a Calvin et al. (1987)
a considerar a las lombrices endogeas como dominantes en los suelos de los cultivos. Sin
embargo, al igual que las otras dos categorías ecológicas, las lombrices endogeas también se ven
favorecidas por las prácticas orgánicas (Henneron et al. 2015; Lago 2015). Como en casos
anteriores, el uso de fertilizantes orgánicos como fuente de alimento para sus poblaciones es
uno de los factores que explican los incrementos observados (Pérès et al. 1998; Zaller y Köpke
2004; Simonsen et al. 2010; Palm et al. 2013).
2. MATERIALES Y METODOS
2.1. Toma de muestras en el campo
La toma de muestras se realizó mensualmente desde marzo de 2013 hasta febrero de 2014
en las fincas agrícolas situadas en la fegión atlántica. Al igual que en el capítulo I, en el caso de
las fincas localizadas en la región mediterránea, las elevadas temperaturas registradas durante
los meses estivales, pero sobre todo las bajas precipitaciones en los meses con mayor contraste
térmico (verano e invierno; ver también Introducción General), hicieron que las abundancias de
las poblaciones de invertebrados edáficos disminuyeran drásticamente hasta su casi
desaparición, por lo que los muestreos en esta región se concentraron en los meses más
favorables para las actividades biológicas, de marzo a mayo y de septiembre a diciembre de
2014.
En cada uno de los muestreos se tomaron tres réplicas al azar en tres sectores (elegidos
también al azar) de cada una de las fincas, con el fin de incluir la variabilidad espacial en el diseño
experimental. Debido al diferente tamaño de la fauna edáfica los muestreos cuantitativos exigen
examinar un área mínima distinta:
A) Macrofauna (lombrices): se emplearon cuadrículas de 50 cm de lado y se excavó el suelo
cuidadosamente hasta unos 20 cm de profundidad (Fig. 24A). Las lombrices se
recolectaron manualmente y se almacenaron vivas en recipientes de plástico con suelo
húmedo original de la finca. Estas cajas perfectamente etiquetadas se transportaron en
nevera portátil y se procesaron inmediatamente al llegar al laboratorio.

Capítulo II
73
B) Mesofauna (microartrópodos y enquitreidos): se utilizaron cilindros de acero de 10 cm
de diámetro x 20 cm de altura (Fig. 24B). Las muestras de suelo con su vegetación
asociada intacta se introdujeron en bolsas de plástico etiquetadas y se transportaron en
nevera portátil al laboratorio. Una vez en el laboratorio se obtuvieron dos submuestras:
1/2 del área se empleó para la extracción de los microartrópodos (ácaros, colémbolos y
otros artrópodos de pequeño tamaño) y 1/4 para la extracción de enquitreidos. Estas
muestras perfectamente etiquetadas fueron almacenadas en cámara fría (4 oC) hasta
posteriores análisis.
Figura 24: Muestreo cuantitativo de los distintos grupos faunísticos: (A) cuadrícula de 50x50 cm para recoger manualmente lombrices hasta 20 cm de altura y (B) cilindros de acero de 10 cm de diámetro y 20 cm de altura para la extracción mesofauna.
2.2. Extracción e identificación de la fauna del suelo
2.2.1. Macrofauna: lombrices de tierra (Phylum Annelida: Clase Clitellata, Subclase
Oligochaeta)
La identificación de los ejemplares de lombrices recolectados se llevó a cabo hasta el nivel de
especie (Bouché 1972) con la ayuda de un microscopio estereoscópico (Nikon SMZ-U) provisto
de luz fría. A continuación, los especímenes fueron pesados utilizando una balanza digital
(Sartorius BL 1500 S, 0,01 g) y finalmente, se les asignó la categoría ecológica correspondiente
(Fig. 25) siguiendo las clasificaciones propuestas por Bouché (1972, 1977).
A B

Capítulo II
74
Figura 25: Ejemplares de lombrices pertenecientes a tres categorías ecológicas propuestas por Bouché (1972, 1977): A) epigeas (Dendrodrilus rubidus), B) anécica (Scheroteca gigas) y C) endogeas (Octolasion lacteum).
2.2.2. Mesofauna: Microartrópodos (Phylum Arthropoda: Subphyla Chelicerata, Myriapoda,
Crustacea y Hexapoda)
Para la extracción de microartrópodos se empleó una extracción seca (Tullgren 1918) y los
ejemplares obtenidos se fijaron en etanol al 70%. Para la clasificación taxonómica se adoptaron
las propuestas filogenéticas de Wheeler et al. (2001), Maddison et al. (2007) y Regier et al. (2010)
(Fig. 26).

Capítulo II
75
Figura 26: Esquema taxonómico adoptado para clasificar los microartrópodos investigados.
La identificación de los microartrópodos recogidos se llevó a cabo mediante una lupa
binocular (Nikon SMZ-U) y un microscopio (Nikon Eclipse 80i) hasta el nivel de orden, excepto
en el caso de los miriápodos que fueron identificados hasta nivel de clase. Se emplearon las
claves dicotómicas de Hopkin (2000) para colémbolos (Fig. 27A), Pérez-Iñigo (1993) para ácaros
(Fig. 27B) y las de Barrientos (1988) para los restantes grupos de microartrópodos (arácnidos,
miriápodos y órdenes de insectos; Fig. 27C).
Phylum Arthropoda Subphylum Chelicerata Clase Arachnida Subclase Micrura Orden Araneida Subclase Drompoda Orden Pseudoescorpionida Subclase Acari Superorden Parasitiformes Orden Mesostigmata (=Gamasida) Superorden Acariformes Orden Trombidiformes Suborden Prostigmata Orden Sarcoptiformes Suborden Oribatida Subphylum Myriapoda Clase Chilopoda Clase Diplopoda Clase Symphyla Subphylum Crustacea Clase Malacostraca Orden Isopoda Subphylum Hexapoda Clase Entognatha Orden Collembola Suborden Poduromorpha Suborden Entomobryomorpha Suborden Symphypleona Orden Diplura Orden Protura Clase Ectognatha (=Insecta) Subclase Pterygota Orden Hymenoptera Orden Hemiptera
Orden Coleoptera Orden Diptera Orden Thysanoptera Orden Lepidoptera Orden Psocoptera

Capítulo II
76
Figura 27: Ejemplares de diversos grupos de microartrópodos: A) colémbolo, B) ácaros, C) algunos ordenes de insectos (Hymenoptera, Coleoptera, Thysanoptera y Hemiptera), arañas y miriápodos (sínfilos).
2.2.3. Mesofauna: Enquitreidos (Phylum Annelida: Clase Clitellata, Subclase Oligochaeta)
Para los enquitreidos (Fig. 28) se empleó una extracción húmeda siguiendo el método de
O´Connor (1955). Los ejemplares vivos extraídos se fijaron en etanol al 70% y posteriormente,
se contaron con la ayuda de una lupa binocular (Nikon SMZ-U) y un microscopio (Nikon Eclipse
80i).
Figura 28: Ejemplares de enquitreidos (Familia Enchytraeidae).
2.3. Análisis estadísticos
2.3.1. Análisis factorial de la Varianza (ANOVA) de la composición y abundancia de los taxones
de invertebrados edáficos
Con el fin comprobar si la región geográfica, el tipo de cultivo y el manejo agrícola tuvieron
un efecto significativo sobre la composición y estructura de las comunidades edáficas se realizó
una ANOVA para diseños no balanceados (debido al diferente número de muestras recogidas en
cada región). Previamente se comprobó que los datos de abundancia cumpliesen los criterios
A Collembola B Acari
Hymenoptera
Araneida
Coleoptera Hemiptera
Thysanoptera
Symphyla
C

Capítulo II
77
de normalidad (Test de Shapiro Wilk) y la homogeneidad de varianzas (Test de Levene). Para la
separación de las medias se empleó, a posteriori, el test Tukey (α= , .
Para el estudio detallado del efecto del cultivo y el manejo en cada una de las regiones
geográficas investigadas también se empleó el análisis paramétrico ANOVA, pero solamente
incluyendo aquellos taxones que estuvieron presentes en al menos el 20% del número total de
muestras recogidas. Al igual que en el caso anterior, previamente se comprobó que los datos
cumpliesen los criterios de normalidad (Test de Shapiro Wilk) y homogeneidad de varianzas (Test
de Levene) y para la separación de las medias se empleó el test Tukey (α= , .
Todos los análisis fueron realizados con el programa estadístico STATISTICA v7 (StatSoft).
2.3.2. Análisis de la diversidad, similitud y dinámica temporal de las comunidades de lombrices
de tierra
Dado que las lombrices de tierra responden rápidamente a los cambios producidos por los
sistemas de laboreo (Pérès et al. 2008; van Eekeren et al. 2008; Postma-Blaauw et al. 2010), son
consideradas buenos indicadores de la calidad del suelo (Paoletti 1999; Pérès et al. 2008;
Pulleman et al. 2012) y por ello, se calcularon varios índices de diversidad y similitud y se
compararon cambios temporales de sus abundancias y biomasas, con el fin de describir la
estructura de sus comunidades en los diferentes cultivos frutícolas.
2.3.2.1. Análisis de diversidad
2.3.2.1.1. Índice de riqueza de especies (S)
Este índice proporciona una forma muy sencilla para medir la diversidad y se basa
únicamente en el número de especies presentes en una muestra. Se calcularon dos subtipos de
índices de riqueza de especies S:
SMensual: número de especies obtenidas en cada una de las fincas en cada muestreo mensual.
STotal: número de especies obtenidas en cada una de las fincas durante todo el periodo de
estudio.
2.3.2.1.2. Índice de diversidad de Shannon-Wiener (H´)
Este índice indica cómo se distribuyen las abundancias entre el número de especies presentes
en una muestra. Normalmente se ep ese ta o o H y se calcula según la siguiente fórmula
(Shannon y Wiener, 1949):
�´ = − ∑ �� � ���=

Capítulo II
78
dónde
S – número de especies (la riqueza de especies)
pi – proporción de individuos de la especie i respecto al total de individuos (ni/N)
ni – número de individuos de la especie i
N – número de todos los individuos de todas las especies
Po ta to, H =0 cuando sólo hay una especie en la muestra y será máxima cuando todas las
espe ies esté ep ese tadas po el is o ú e o de i di iduos H max).
En este estudio se calcularon dos subtipos de este índice en función de los datos de
abundancia obtenidos:
H´Mensual: valor de diversidad obtenido en cada una de las fincas en cada muestreo mensual.
H´Total: diversidad total en cada una de las fincas durante todo el periodo de estudio.
2.3.2.1.3. Índice de equitabilidad de Pielou (J´)
Este índice, propuesto por Pielou (1966), mide la proporción entre la diversidad observada y
la diversidad máxima teórica. Se calcula mediante la fórmula: �´ = H´ ���� H´Max
dónde H Total es el índice de Shannon-Wiener y H Max es la diversidad máxima que se obtendría si
la distribución de las abundancias de las especies en la comunidad fuesen perfectamente
equitativas.
H Max se calcula de la siguiente forma:
H´Max = Ln S Total
El valor del índice J´ oscila entre 0 y 1, alcanzándose este último cuando la diversidad
observada es igual a la máxima teórica.
2.3.2.2. Análisis de similitud/disimilitud en la estructura de las poblaciones de lombrices entre
distintas regiones y manejos agrícolas
Los datos de abundancia de lombrices fueron empleados para analizar el grado de
similitud/disimilitud en la estructura de sus comunidades en los distintos cultivos frutícolas
mediante el índice de similitud de Bray-Curtis (ajustada a cero), seguido de un análisis de Multi-
escalamiento Dimensional no Métrico (Multi-Dimensional Scaling, MDS) y el cálculo de los
porcentajes de similitud (Similarity Percentage Analysis, SIMPER; Clarke 1993).
La matriz de similitud ajustada a cero de Bray-Curtis se obtuvo al añadir un individuo ficticio
(denominado dummy) de cada especie a todas las muestras, de tal modo que las muestras vacías

Capítulo II
79
se convierten en similares entre ellas gracias a este individuo ficticio y así no es necesario
eliminarlas para realizar una correcta representación MDS posterior (Clarke et al. 2006).
Previamente al análisis se transformaron los datos mediante raíz cuadrada.
La significación de los resultados obtenidos en la matriz de similitud de Bray-Curtis se calculó
mediante el análisis de similitud (ANOSIM). El estadístico de este análisis (R) alcanza valores
comprendidos entre -1 y 1, pero normalmente sólo toma valores positivos. Cuando su valor está
cercano a 0 indica que los tratamientos no son diferentes y valores > 0 significa que sí hay
diferencias entre ellos. Para determinar la probabilidad de que los grupos sean diferentes,
ANOSIM recalcula el estadístico R utilizando un número fijo de permutaciones según el nivel sea
0,01 o 0,05 y calcula la probabilidad asociada al nivel de significación establecido (p) (Clarke et
al. 2006).
El análisis MDS permite visualizar el grado de similitud/disimilitud en la composición de las
comunidades obtenidas a partir del cálculo del índice de disimilitud de Bray-Curtis. Este índice
oscila entre 0 y 1, donde 0 indica que las dos muestras analizadas tienen la misma composición
(es decir, presentan las mismas especies), mientras que el valor de 1 significa que las dos
muestras no comparten ninguna especie. En el análisis MDS cuando la composición de una
comunidad de una zona determinada apenas varían entre muestras, los puntos aparecen
representados muy próximos entre sí y se mostrarán más separados cuando la composición de
dicha comunidad varía entre muestras.
Finalmente, el cálculo de los porcentajes de similitud (SIMPER) permite identificar qué
taxones contribuyen en mayor medida al valor de porcentaje de similitud. Este análisis compara
pares de especies o de muestras y calcula la contribución de cada uno de ellos a la
similitud/disimilitud total según su presencia/ausencia y/o su abundancia. Para este análisis
solamente se incluyeron las especies que contribuyeron en un 90% a la similitud/disimilitud total
acumulada.
2.3.2.3. Comparación de curvas de Abundancia y Biomasa (ABC)
La comparación de curvas de abundancia y biomasa (ABC) se basa en la superposición y
comparación de las curvas k-dominancia de Lambshead et al. (1983) y está considerada como
una técnica de análisis distribucional (Clarke y Warwick 2001). En general, se asume que en su
estado normal la comunidad está dominada por especies de crecimiento lento y tamaños
grandes y por tanto, la curva de biomasa está por encima de la curva de abundancia. En el eje
de abscisas se ordenan, en escala logarítmica, las especies en orden de contribución a la biomasa

Capítulo II
80
y abundancia respectivamente, y en el eje de ordenadas se representan el porcentaje de
dominancia (en escala acumulativa).
El estadístico W (Clarke 1990) representa el área de diferencia entre las dos curvas y permite
comparar los perfiles ABC para diferentes usos del suelo. Para el cálculo de W se emplea la
siguiente fórmula:
W = ∑ B� − A� /[5 S − ] �=
dónde S describe el cambio en el número total de especies, Ai es la abundancia total en cada
muestra i y Bi es la biomasa total en cada muestra i.
Los valores del estadístico W oscilan entre 1 y -1, aunque nunca alcanza los valores límite. El
estadístico W tiende a 1 cuando la curva de biomasa se localiza sobre la curva de abundancia,
es decir, en situaciones de ausencia de perturbación en las que las especies grandes y de
crecimiento lento suelen ser las dominantes; en cambio, W tiende a -1 en el caso de fuertes
perturbaciones, cuando la curva de abundancia se sitúa sobre la curva de biomasa y como
consecuencia, empiezan a dominar las especies oportunistas de pequeño tamaño y crecimiento
rápido. En los casos que las curvas están aproximadamente superpuestas (casos de
perturbaciones moderadas), el valor de W es próximo a 0, debido a que desaparecen las especies
de gran tamaño.
Los cálculos de los índices de diversidad, los análisis de similitud/disimilitud así como la
comparación de las curvas abundancia y biomasa fueron realizados con el estadístico
multivariante PRIMER (Plymouth Routines in Multivariate Ecological Research v6; Clarke y
Gorley 2006).
3. RESULTADOS
3.1. Composicion y estructura de las comunidades edáficas en los distintos sistemas de cultivo
3.1.1. Abundancias totales de fauna edáfica
Durante el estudio se capturaron un total de 11.883 invertebrados, de los cuales 6.036
pertenecieron al Phylum Annelida, con taxones incluidos tanto en la mesofauna (enquitreidos)
como en la macrofauna (lombrices de tierra). El resto de los especímenes capturados (5.847
ejemplares) fueron invertebrados pertenecientes al Phylum Arthropoda. Así, en los cultivos
estudiados se encontraron 3.356 ejemplares totales de la clase Arachnida (ordenes Araneida,
Pseudoescorpionida y subclase Acari), 118 individuos totales del subphylum Myriapoda (clases
Diplopoda, Chilopoda y Symphyla), 34 individuos totales del subphylum Crustacea (clase
Malacostraca, orden Isopoda). Finalmente, también se recolectaron 2.339 especímenes de
insectos, de los cuales 1.773 pertenecieron a la clase Entognatha (ordenes Collembola, Protura

Capítulo II
81
y Diplura) y 566 a la clase Ectognatha (ordenes Hymenoptera, Hemiptera, Coleoptera, Diptera,
Thysanoptera, Lepidoptera y Psocoptera).
Ni la región biogeográfica, ni el manejo tuvieron un efecto significativo sobre las abundancias
totales de fauna edáfica (Tabla 10). En cambio, el tipo de cultivo sí tuvo un efecto significativo
sobres sus densidades (Tabla 10) y concretamente, el cultivo de mandarino concentró las
mayores densidades de invertebrados edáficos, seguido del viñedo, kiwi y olivo (Fig. 29).
Tabla 10: Resultados de ANOVA para las abundancias totales de la fauna recolectada (ind. m-2).
Abundancia total Abundancia total en la Región Atlántica
Abundancia total en la Región
Mediterránea
Fuente de variación g.l. F p g.l. F p g.l. F p Región 1 1,01 0,317 - - - - - - Cultivo 2 4,35 0,014 1 0,04 0,831 1 6,75 0,011 Manejo 1 0,00 0,980 1 2,12 0,148 1 1,60 0,210 Tiempo - - - 11 2,94 0,002 6 2,95 0,014 Región*Manejo 1 3,41 0,066 - - - - - - Cultivo*Manejo 2 1,38 0,253 1 0,11 0,478 1 3,28 0,075 Tiempo*Cultivo - - - 11 1,72 0,079 6 0,86 0,526 Tiempo*Manejo - - - 11 0,97 0,478 6 0,79 0,574 Tiempo*Cultivo*Manejo - - - 11 0,91 0,526 6 0,72 0,632
Figura 29: Valores medios (±E.E.) de abundancia (ind. m-2) de fauna edáfica en cada uno de los cultivos investigados. Letras diferentes indican diferencias significativas entre cultivos (p<0,05).
Cuando se analizan los resultados de cada región por separado, los resultados de la ANOVA
(Tabla 10) indican que el tipo de cultivo sólo tuvo un efecto significativo sobre las abundancias
de las comunidades edáficas encontradas en los suelos mediterráneos, lo que puede estar
relacionado con el contraste de abundancias observado entre los cultivos de mandarino y olivo
(Fig. 29).
aa
b
a
0
5000
10000
15000
20000
25000
Kiwi Vid Mandarino Olivo
Ab
un
dan
cia
me
dia
(in
d. m
-2)

Capítulo II
82
En cambio, el factor tiempo tuvo un efecto significativo sobre las abundancias totales
registradas en ambas regiones (Tabla 1). Y así, en el mes de abril en la región atlántica se observó
una elevada densidad de organismos (28.470±5.907 ind. m-2), lo que contrastó con las
registradas en los meses de mayo, julio, agosto, septiembre, diciembre de 2013 y enero de 2014,
meses en los que no se superaron los 9.570 ind. m-2 de media (Fig. 30A). En el caso de la región
mediterránea, fueron los meses de marzo y diciembre los que presentaron significativamente
menores densidades (6.448±1.127 y 4.330±988 ind. m-2, respectivamente) que el mes de
septiembre (23.556±5.965 ind. m-2). En el resto de los meses las densidades de organismos
edáficos alcanzaron valores intermedios (Fig. 30B).
Figura 30: Valores medios (±E.E.) de abundancia mensual (ind. m-2) de fauna edáfica en la región atlántica (A) y Mediterránea (B). Letras diferentes indican diferencias significativas entre meses (p<0,05).
Del total de 11.883 individuos capturados en los suelos de cultivo investigados, el 71%
correspondió a la mesofauna, mientras que la macrofauna (lombrices de tierra) sólo representó
el 29%. Entre la mesofauna, los enquitreidos constituyeron el grupo mayoritario con un 47% del
total, seguidos de los arácnidos (30%) y los entognatos (16%), mientras que los insectos
ectognatos y el resto de artrópodos solamente contribuyeron con un 6% a la mesofauna total
(Fig. 31).
Figura 31: Abundancia relativa (%) de los principales grupos de mesofauna.
0%
20%
40%
60%
80%
100%
Mesofauna
Ab
un
dan
cia
rela
tiva
(%)
Ectognatha y otrosArthropoda
Entognatha
Arachnida
Enchytreidae

Capítulo II
83
Para el caso de la macrofauna, se identificaron un total 3.428 de individuos pertenecientes a
tres familias diferentes de lombrices: Lumbricidae (14 especies), Megascolecidae (1 especie;
Amyntas corticis) y Acanthodrildiae (1 especie; Microscolex phosphoreus).
En cuanto a la estructura ecológica de las comunidades de lombrices de tierra, estuvo
dominada por las lombrices anécicas y endogeas, representando el 47% y un 39% del total de
lombrices capturadas, respectivamente (Fig. 32). Esto fue debido al mayor número de
ejemplares capturados de Aporrectodea trapezoides (anécica) y Octolasion lacteum (endogea)
en estos suelos de cultivo (Fig. 32).
Figura 32: Abundancia relativa (%) de las especies de lombrices y categorías ecológicas.
El análisis de la ANOVA para las abundancias totales de estos dos grupos de fauna edáfica
(mesofauna y macrofauna) en cada una de las regiones investigadas (Tabla 11) indica que los
suelos de cultivo de ambas regiones estuvieron dominados por invertebrados de pequeño
tamaño (12.950±1.457 ind. m-2 de mesofauna frente a 56±10 ind. m-2 de macrofauna en la región
atlántica y 15.429±2071 ind. m-2 de mesofauna frente a 66±18 ind. m-2 de macrofauna en la
región mediterránea).
0%
20%
40%
60%
80%
100%
Macrofauna
Ab
un
dan
cia
rela
tiva
(%
)
Dendrobaena octaedra
Dendrodrilus rubidus
Lumbricus rubellus
Aporrectodea trapezoides
Lumbricus friendi
Lumbricus terrestris
Octodrilus complanatus
Scherotheca gigas
Allolobophora chlorotica
Aporrectodea caliginosa
Aporrectodea rosea
Octolasion cyaneum
Octolasion lacteum
Dendrobaena madeirensis
Microscolex phosphoreus
Amynthas corticis
Epiegas (8%)
Anécicas (47%)
Endoegas (39%)
Epiendoegas (4%)
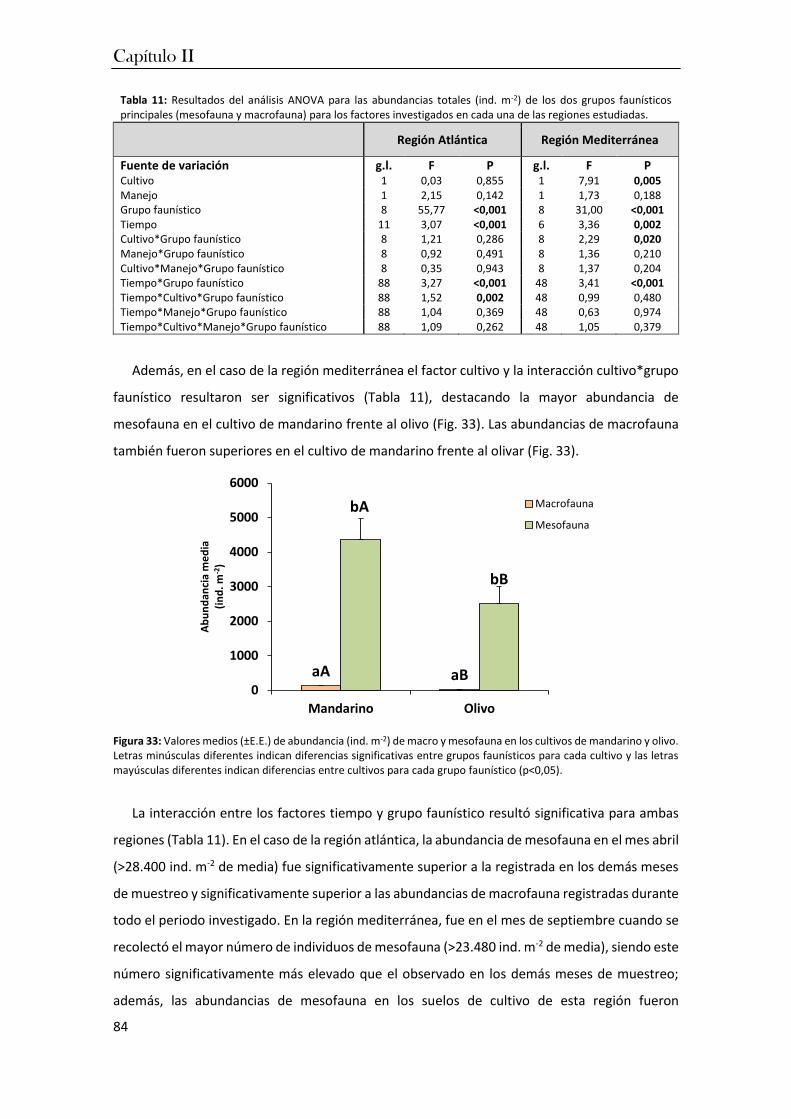
Capítulo II
84
Tabla 11: Resultados del análisis ANOVA para las abundancias totales (ind. m-2) de los dos grupos faunísticos principales (mesofauna y macrofauna) para los factores investigados en cada una de las regiones estudiadas.
Región Atlántica Región Mediterránea
Fuente de variación g.l. F P g.l. F P Cultivo 1 0,03 0,855 1 7,91 0,005 Manejo 1 2,15 0,142 1 1,73 0,188 Grupo faunístico 8 55,77 <0,001 8 31,00 <0,001 Tiempo 11 3,07 <0,001 6 3,36 0,002 Cultivo*Grupo faunístico 8 1,21 0,286 8 2,29 0,020 Manejo*Grupo faunístico 8 0,92 0,491 8 1,36 0,210 Cultivo*Manejo*Grupo faunístico 8 0,35 0,943 8 1,37 0,204 Tiempo*Grupo faunístico 88 3,27 <0,001 48 3,41 <0,001 Tiempo*Cultivo*Grupo faunístico 88 1,52 0,002 48 0,99 0,480 Tiempo*Manejo*Grupo faunístico 88 1,04 0,369 48 0,63 0,974 Tiempo*Cultivo*Manejo*Grupo faunístico 88 1,09 0,262 48 1,05 0,379
Además, en el caso de la región mediterránea el factor cultivo y la interacción cultivo*grupo
faunístico resultaron ser significativos (Tabla 11), destacando la mayor abundancia de
mesofauna en el cultivo de mandarino frente al olivo (Fig. 33). Las abundancias de macrofauna
también fueron superiores en el cultivo de mandarino frente al olivar (Fig. 33).
Figura 33: Valores medios (±E.E.) de abundancia (ind. m-2) de macro y mesofauna en los cultivos de mandarino y olivo. Letras minúsculas diferentes indican diferencias significativas entre grupos faunísticos para cada cultivo y las letras mayúsculas diferentes indican diferencias entre cultivos para cada grupo faunístico (p<0,05).
La interacción entre los factores tiempo y grupo faunístico resultó significativa para ambas
regiones (Tabla 11). En el caso de la región atlántica, la abundancia de mesofauna en el mes abril
(>28.400 ind. m-2 de media) fue significativamente superior a la registrada en los demás meses
de muestreo y significativamente superior a las abundancias de macrofauna registradas durante
todo el periodo investigado. En la región mediterránea, fue en el mes de septiembre cuando se
recolectó el mayor número de individuos de mesofauna (>23.480 ind. m-2 de media), siendo este
número significativamente más elevado que el observado en los demás meses de muestreo;
además, las abundancias de mesofauna en los suelos de cultivo de esta región fueron
aA aB
bA
bB
0
1000
2000
3000
4000
5000
6000
Mandarino Olivo
Ab
un
dan
cia
me
dia
(in
d. m
-2)
Macrofauna
Mesofauna

Capítulo II
85
significativamente diferentes a las de macrofauna en cada uno de los meses investigados.
Finalmente, la interacción tiempo*cultivo*grupo faunístico resultó ser significativa solamente
para la región atlántica (Tabla 11), lo que se asocia a la mayor abundancia de mesofauna
encontrada en el viñedo en el mes de abril (>35.150 ind. m-2 de media) con respecto a los valores
registrados en los demás meses de los restantes cultivos.
3.1.2. Abundancias de los distintos taxones de mesofauna y macrofauna
Al analizar el efecto de los distintos factores incluidos en el diseño experimental sobre las
abundancias de los diferentes taxones de meso y macrofauna en cada una de las regiones
biogeográficas (Tabla 12) se observaron diferencias significativas en las abundancias de los
distintos taxones identificados en cuanto a cultivo y manejo, así como importantes variaciones
temporales (Tabla 12).
Tabla 12: Resultados del análisis ANOVA para las abundancias (ind. m-2) de los principales taxones de mesofauna (Enchytraeidae, Arachnida, Entognatha, Ectognatha y otros Arthropoda) y las categorías ecológicas de la macrofauna (lombrices de tierra epigeas, anécicas, endogeas y epiendogeas).
Mesofauna Región Atlántica Región Mediterránea
Fuente de variación g.l. F P g.l. F P
Cultivo 1 0,06 0,799 1 7,47 0,006 Manejo 1 2,30 0,129 1 1,93 0,165 Taxón 3 42,29 <0,001 3 18,08 <0,001 Tiempo 11 3,06 <0,001 6 3,37 0,003 Cultivo*Taxón 3 1,42 0,236 3 1,39 0,244 Manejo*Taxón 3 0,64 0,583 3 1,22 0,301 Cultivo*Manejo*Taxón 3 0,40 0,748 3 0,87 0,454 Tiempo*Taxón 33 3,32 <0,001 18 3,42 <0,001 Tiempo *Cultivo*Taxón 33 1,46 0,049 18 1,00 0,458 Tiempo*Manejo*Taxón 33 1,05 0,390 18 0,58 0,910 Tiempo*Cultivo*Manejo*Taxón 33 1,12 0,297 18 1,09 0,356
Macrofauna Región Atlántica Región Mediterránea
Fuente de variación g.l. F P g.l. F P
Cultivo 1 172,06 <0,001 1 65,28 <0,001 Manejo 3 84,42 <0,001 1 58,93 <0,001 Categoría ecológica 3 74,62 <0,001 2 66,83 <0,001 Tiempo 11 5,66 <0,001 6 1,00 0,421 Cultivo*Categoría ecológica 3 84,21 <0,001 2 63,38 <0,001 Manejo*Categoría ecológica 3 91,63 <0,001 2 45,18 <0,001 Cultivo*Manejo*Categoría ecológica 3 90,73 <0,001 2 46,77 <0,001 Tiempo*Categoría ecológica 33 2,27 <0,001 12 2,48 0,005 Tiempo*Cultivo*Categoría ecológica 33 3,14 <0,001 12 2,23 0,012 Tiempo*Manejo*Categoría ecológica 33 1,89 0,002 12 2,40 0,006 Tiempo*Cultivo*Manejo*Categoría ecológica 33 1,67 0,012 12 2,33 0,008
3.1.2.1. Diferencias entre regiones biogeográficas
En el caso de la mesofauna, se observó que si bien en la región atlántica los enquitreidos
fueron el grupo mayoritario numéricamente (p<0,05; Fig. 34), en la región mediterránea el
grupo dominante fueron los arácnidos (p<0,05; Fig. 34), de los cuales más del 99% pertenecieron

Capítulo II
86
a la subclase Acari. En cambio, los insectos ectognatos y otros artrópodos (como arañas,
miriápodos, isópodos, etc.) tuvieron representaciones similares en ambas regiones, con menos
del 8% de representación total (Fig. 34) y, en concreto, 9 taxones (Pseudoescorpionida, Araneae,
Heteroptera, Thysanoptera, Lepidoptera, Psocoptera, Diplopoda, Chilopoda e Isopoda)
estuvieron muy poco representados (<0,2% de los individuos totales recogidos en todo el
estudio).
Figura 34: Abundancia relativa (%) de los distintos grupos integrantes de la mesofauna en cada una de las dos regiones biogeográficas estudiadas.
En el caso de la macrofauna, cinco especies de lombrices fueron encontradas solamente en
la región atlántica (Dendrobaena octaedra, D. madeirensis, Dendrodrilus rubidus, Lumbricus
friendi y L. rubellus), mientras que 6 especies (L. terrestris, Allolobophora chlorotica,
Aporrectodea rosea, Octolasion cyaneum, Octodrilus complanatus y Scherotheca gigas) fueron
recolectadas únicamente en la región mediterránea (Fig. 35). Por otro lado, las abundancias de
Octolasion lacteum, Microscolex phosphoreus y Amynthas corticis fueron significativamente más
abundantes en los suelos atlánticos que en los mediterráneos mientras que Aporrectodea
trapezoides estuvo mejor representada en los suelos mediterráneos (Fig. 35).
6 7
1320
13
55
68
18
0%
20%
40%
60%
80%
100%
Atlántica Mediterránea
Ab
un
dan
cia
rela
tiva
(%
)
Enchytreidae
Arachnida
Entognatha
Ectognatha y otrosArthropoda

Capítulo II
87
Figura 35: Abundancia relativa (%) de las lombrices encontradas en cada una de las regiones estudiadas. Para cada especie las diferencias significativas entre regiones se indican mediante un asterisco (p < 0,05).
La estructura ecológica de las comunidades de lombrices también fue significativamente
diferente en las dos regiones biogeográficas investigadas (Tabla 12 y Fig. 36). Así, en la región
atlántica las lombrices endogeas fueron el grupo dominante (Fig. 36), gracias a las elevadas
abundancias de la especie O. lacteum (Fig. 35), mientras que en la región mediterránea destaca
la ausencia de lombrices epigeas y la dominancia de las lombrices pertenecientes a la categoría
ecológica de las anécicas (Fig. 36), como resultado del elevado número de ejemplares de la
especie A. trapezoides (Fig. 35). En ambas regiones, las lombrices epiendogeas estuvieron muy
poco representadas (Fig. 36).
Figura 36: Valores medios (±E.E.) de abundancia (ind. m-2) de las lombrices de tierra pertenecientes a diferentes categorías ecológicas en cada una de las dos regiones estudiadas. Letras diferentes indican diferencias significativas entre categorías para cada región (p< 0,05).
0%
20%
40%
60%
80%
100%
De
nd
rob
ae
na
oct
ae
dra
De
nd
rod
rilu
s ru
bid
us
Lum
bri
cus
rub
ell
us
Ap
orr
ect
od
ea
tra
pe
zoid
es
Lum
bri
cus
frie
nd
i
Lum
bri
cus
terr
est
ris
Oct
od
rilu
s co
mp
lan
atu
s
Sch
ero
the
ca g
iga
s
All
olo
bo
ph
ora
ch
loro
tica
Ap
orr
ect
od
ea
ca
lig
ino
sa
Ap
orr
ect
od
ea
ro
sea
Oct
ola
sio
n c
ya
ne
um
Oct
ola
sio
n l
act
eu
m
De
nd
rob
ae
na
ma
de
ire
nsi
s
Mic
rosc
ole
x p
ho
sph
ore
us
Am
yn
tha
s co
rtic
is
Ab
un
dan
cia
rela
tiva
(%
)
Mediterránea
Atlántica
EPIGEAS ANÉCICAS ENDOGEAS EPIENDOGEAS
* ** *
EPIGEAS ANÉCICAS ENDOGEAS EPIENDOGEAS
a a
a
b
b
ab
0
10
20
30
40
50
60
70
80
Atlántica Mediterránea
Ab
un
dan
cia
med
ia (
ind
. m-2
)
Epigeas
Anécicas
Endogeas
Epiendogeas
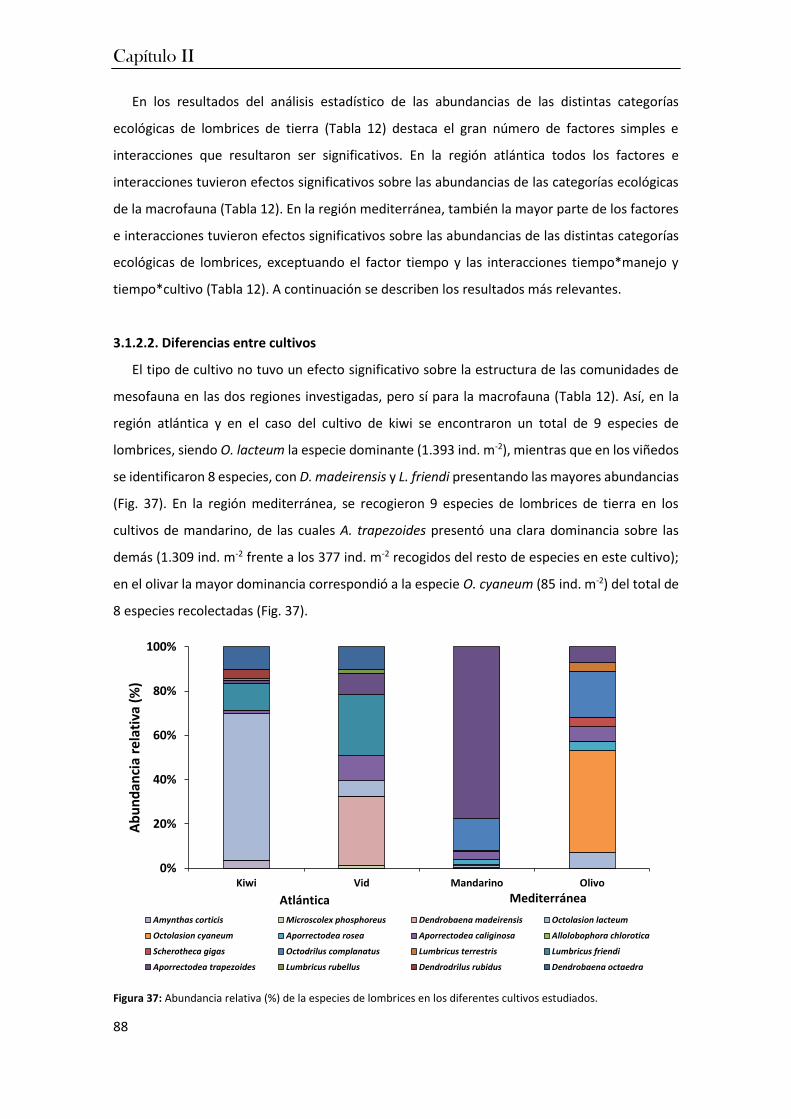
Capítulo II
88
En los resultados del análisis estadístico de las abundancias de las distintas categorías
ecológicas de lombrices de tierra (Tabla 12) destaca el gran número de factores simples e
interacciones que resultaron ser significativos. En la región atlántica todos los factores e
interacciones tuvieron efectos significativos sobre las abundancias de las categorías ecológicas
de la macrofauna (Tabla 12). En la región mediterránea, también la mayor parte de los factores
e interacciones tuvieron efectos significativos sobre las abundancias de las distintas categorías
ecológicas de lombrices, exceptuando el factor tiempo y las interacciones tiempo*manejo y
tiempo*cultivo (Tabla 12). A continuación se describen los resultados más relevantes.
3.1.2.2. Diferencias entre cultivos
El tipo de cultivo no tuvo un efecto significativo sobre la estructura de las comunidades de
mesofauna en las dos regiones investigadas, pero sí para la macrofauna (Tabla 12). Así, en la
región atlántica y en el caso del cultivo de kiwi se encontraron un total de 9 especies de
lombrices, siendo O. lacteum la especie dominante (1.393 ind. m-2), mientras que en los viñedos
se identificaron 8 especies, con D. madeirensis y L. friendi presentando las mayores abundancias
(Fig. 37). En la región mediterránea, se recogieron 9 especies de lombrices de tierra en los
cultivos de mandarino, de las cuales A. trapezoides presentó una clara dominancia sobre las
demás (1.309 ind. m-2 frente a los 377 ind. m-2 recogidos del resto de especies en este cultivo);
en el olivar la mayor dominancia correspondió a la especie O. cyaneum (85 ind. m-2) del total de
8 especies recolectadas (Fig. 37).
Figura 37: Abundancia relativa (%) de la especies de lombrices en los diferentes cultivos estudiados.
0%
20%
40%
60%
80%
100%
Kiwi Vid Mandarino Olivo
Ab
un
dan
cia
rela
tiva
(%
)
Amynthas corticis Microscolex phosphoreus Dendrobaena madeirensis Octolasion lacteum
Octolasion cyaneum Aporrectodea rosea Aporrectodea caliginosa Allolobophora chlorotica
Scherotheca gigas Octodrilus complanatus Lumbricus terrestris Lumbricus friendi
Aporrectodea trapezoides Lumbricus rubellus Dendrodrilus rubidus Dendrobaena octaedra
Atlántica Mediterránea

Capítulo II
89
La estructura ecológica de sus poblaciones de lombrices también fue significativamente
distinta en los cuatro cultivos estudiados (Tabla 12). Así, a diferencia de los cultivos atlánticos,
en la región mediterránea destaca la ausencia de lombrices que viven en los horizontes más
superficiales del suelo (epigeas), contabilizándose únicamente unos pocos ejemplares de
lombrices epiendogeas en el cultivo de mandarino (Fig. 38). Por otro lado, tanto el viñedo como
el olivo no mostraron diferencias significativas en las abundancias de las categorías ecológicas
presentes (Fig. 15). En cambio, en los cultivos de kiwi las lombrices endogeas dominaron
numéricamente frente a las demás categorías y en el mandarino lo hicieron las anécicas (Fig.
38).
Figura 38: Valores medios (±E.E.) de abundancia (ind. m-2) de las diferentes categorías ecológicas de lombrices encontradas en los cuatro cultivos estudiados. Letras minúsculas diferentes indican diferencias significativas entre categorías ecológicas para cada cultivo y las mayúsculas indican diferencias significativas entre cultivos para cada categoría ecológica (p < 0,05).
3.1.2.3. Diferencias entre manejos agrícolas
El análisis ANOVA de las abundancias de los 4 principales taxones de mesofauna identificados
indicó la ausencia de efectos significativos del manejo sobre sus abundancias en las dos regiones
investigadas (Tabla 12).
En el caso de la macrofauna, sí se observaron efectos significativos del manejo agrícola sobre
la estructura ecológica de las comunidades de lombrices encontradas en ambas regiones (Tabla
12). En la región atlántica y en el manejo convencional, todas las categorías ecológicas tuvieron
representaciones similares, mientras que en los suelos con manejo orgánico estuvieron
claramente dominadas por lombrices endogeas (Fig. 39A). En cambio, en la región
mediterránea, las lombrices anécicas fueron la categoría ecológica dominante en los suelos con
aA
aA
aAaA
aC
aA
bA
aB
bB aB
aA aA bA0
30
60
90
120
150
180
Kiwi Vid Mandarino Olivo
Ab
un
dan
cia
me
dia
(in
d. m
-2)
Epigeas
Anécicas
Endogeas
Epiendogeas
Atlántica Mediterránea

Capítulo II
90
manejo convencional, mientras que en los suelos con manejo orgánico las abundancias
estuvieron homogéneamente repartidas entre las distintas categorías ecológicas (Fig. 39B).
Figura 39: Valores medios (±E.E.) de abundancia (ind. m-2) de las diferentes categorías ecológicas de lombrices encontradas en los diferentes manejos agrícolas estudiados en la región atlántica (A) y mediterránea (B). Letras diferentes indican diferencias significativas entre categorías ecológica para cada manejo agrícola (p<0,05).
3.1.2.4. Diferencias entre cultivos sometidos a diferentes manejos agrícolas
El análisis ANOVA de las abundancias de los 4 principales taxones de mesofauna identificados
indicó la ausencia de efectos significativos de la interacción cultivo*manejo sobre sus
abundancias en ambas regiones (Tabla 12), siendo los enquitreidos el taxón dominante en todos
los suelos de cultivo atlánticos sometidos a los dos tipos de manejo agrícola y los arácnidos en
el caso de los cultivos mediterráneos en ambos manejos.
En el caso de la macrofauna, sí se observaron efectos significativos del manejo agrícola
aplicado a los distintos cultivos investigados sobre las categorías ecológicas de lombrices (Tabla
12), por lo que son estos resultados los que se describirán más detalladamente.
a
aa
a
a
b
a a
0
10
20
30
40
50
60
70
80
Convencional Orgánico
Ab
un
dan
cia
me
dia
(In
d. m
-2)
Epigeas Anécicas Endogeas Epiendogeas
A
a
bb
b b0
20
40
60
80
100
120
140
160
Convencional Orgánico
Ab
un
dan
cia
me
dia
(In
d. m
-2) B

Capítulo II
91
4.2.2.4.1. Cultivos de kiwi
En este cultivo atlántico, destacan las mayores abundancias de D. octaedra, D. rubidus y A.
trapezoides en la finca con manejo convencional (Fig. 40). En cambio, en la finca con manejo
orgánico O. lacteum dominó claramente al resto de las especies presentes en estos cultivos (Fig.
40). Además, L. friendi y A. corticis estuvieron ausentes en el suelo convencional y L. rubellus y
D. madeirensis del orgánico (Fig. 40)
Figura 40: Abundancia relativa (%) de las diferentes especies de lombrices encontradas en los cultivos de kiwi sometidos a diferentes manejos agrícolas. Para cada especie las diferencias significativas entre regiones se indican mediante un asterisco (p < 0,05).
En cuanto a la estructura ecológica de las comunidades de lombrices de tierra en los cultivos
de kiwi, hay que señalar que las lombrices epigeas dominaron en el manejo convencional,
mientras en el manejo orgánico fueron las endogeas la categoría ecológica dominante (Fig. 41).
Figura 41: Valores medios (±E.E.) de abundancia (ind. m-2) de las distintas categorías ecológicas de lombrices en los dos cultivos de kiwi sometidos a distinto manejo agrícola. Letras minúsculas diferentes indican diferencias significativas entre categorías ecológicas para cada manejo y las mayúsculas indican diferencias significativas entre manejos para cada categoría ecológica (p < 0,05).
0%
20%
40%
60%
80%
100%
De
nd
rob
ae
na
oct
ae
dra
De
nd
rod
rilu
s ru
bid
us
Lum
bri
cus
rub
ell
us
Ap
orr
ect
od
ea
tra
pe
zoid
es
Lum
bri
cus
frie
nd
i
Ap
orr
ect
od
ea
ca
lig
ino
sa
Oct
ola
sio
n l
act
eu
m
De
nd
rob
ae
na
ma
de
ire
nsi
s
Am
yn
tha
s co
rtic
is
Ab
un
dan
cia
rela
tiva
(%
)
Orgánico
Convencional
EPIGEAS ANÉCICAS ENDOGEAS EPIENDOGEAS
** ** *
aA
aBbA
bB
abA
cB
bAaA
0
20
40
60
80
100
120
Convencional Orgánico
Ab
un
dan
cia
me
dia
(in
d. m
-2)
Epigeas
Anécicas
Endogeas
Epiendogeas

Capítulo II
92
4.1.2.4.2. Cultivos de vid
En el cultivo convencional de vid se encontraron un total de 7 especies de lombrices, de las
cuales 6 de ellas pertenecen a la familia Lumbricidae y siendo D. madeirensis la especie
dominante. En el manejo orgánico se encontraron 5 especies, estando ausentes M. phosphoreus
y D. octaedra y con L. friendi dominando las poblaciones en estos suelos, una especie que no fue
recolectada en los suelos con manejo convencional (Fig. 42). Además, destacan las mayores
abundancias de L. rubellus, O. lacteum y las menores de A. trapezoides y D. madeirensis en el
sistema orgánico comparado con el manejo convencional (Fig. 42), aunque las diferencias no
fueron significativas.
Figura 42: Abundancia relativa (%) de las diferentes especies de lombrices encontradas en los cultivos de vid sometidos a diferentes manejos agrícolas.
Tampoco se detectaron diferencias significativas en la estructura ecológica de los suelos de
viñedo sometidos a diferente manejo, a pesar de que se apreció una cierta tendencia a ser las
lombrices anécicas el grupo ecológico dominante en el cultivo orgánico y una mayor
representatividad de las lombrices epiendogeas en el sistema convencional (Fig. 43).
0%
20%
40%
60%
80%
100%
De
nd
rob
ae
na
oct
ae
dra
Lum
bri
cus
rub
ell
us
Ap
orr
ect
od
ea
tra
pe
zoid
es
Lum
bri
cus
frie
nd
i
Ap
orr
ect
od
ea
ca
lig
ino
sa
Oct
ola
sio
n l
act
eu
m
De
nd
rob
ae
na
ma
de
ire
nsi
s
Mic
rosc
ole
x p
ho
sph
ore
us
Ab
un
dan
cia
rela
tiva
(%
)
Orgánico
Convencional
EPIGEAS ANÉCICAS ENDOGEAS EPIENDOGEAS

Capítulo II
93
Figura 43: Valores medios (±E.E.) de abundancia (ind. m-2) de las distintas categorías ecológicas de lombrices en los dos cultivos de vid sometidos a distinto manejo agrícola. Letras minúsculas diferentes indican diferencias significativas entre categorías ecológicas para cada manejo y las mayúsculas indican diferencias significativas entre manejos para cada categoría ecológica (p < 0,05).
4.1.2.4.3. Cultivos de mandarino
Las comunidades de lombrices en el cultivo de mandarino orgánico incluyeron 8 especies
pertenecientes a las tres familias identificadas (Lumbricidae, Acanthodrilidae y
Megascolecidae), aunque la especie O. complanatus presentó las mayores abundancias (Fig. 44).
Las cinco especies recolectadas en el suelo convencional pertenecen todas ellas a la familia
Lumbricidae y A. trapezoides fue la especie dominante (Fig. 44). Destacan también las mayores
abundancias de A. caliginosa y A. rosea cuando se compara el sistema convencional frente al
orgánico (Fig. 44).
Figura 44: Abundancia relativa (%) de las diferentes especies de lombrices encontradas en los cultivos de mandarino sometidos a diferentes manejos agrícolas. Para cada especie las diferencias significativas entre regiones se indican mediante un asterisco (p < 0,05).
aA
aA
aA
aA
aA
aA
aA
aA
0
2
4
6
8
10
12
14
Convencional Orgánico
Ab
un
dan
cia
me
dia
(in
d. m
-2)
Epigeas
Anécicas
Endogeas
Epiendogeas
0%
20%
40%
60%
80%
100%
Ap
orr
ect
od
ea
tra
pe
zoid
es
Oct
od
rilu
s co
mp
lan
atu
s
All
olo
bo
ph
ora
ch
loro
tica
Ap
orr
ect
od
ea
ca
lig
ino
sa
Ap
orr
ect
od
ea
ro
sea
Oct
ola
sio
n c
ya
ne
um
Oct
ola
sio
n l
act
eu
m
Mic
rosc
ole
x p
ho
sph
ore
us
Am
yn
tha
s co
rtic
is
Ab
un
dan
cia
rela
tiva
(%
)
Orgánico
Convencional
ANÉCICAS ENDOGEAS EPIENDOGEAS
*

Capítulo II
94
En cuanto a la estructura ecológica de las poblaciones de lombrices en los cultivos de
mandarino, hay que señalar que las lombrices anécicas dominaron en ambos manejos,
constituyendo más del 90% de los ejemplares capturados en el cultivo convencional y del 75%
en el orgánico (Fig. 45).
Figura 45: Valores medios (±E.E.) de abundancia (ind. m-2) de las distintas categorías ecológicas de lombrices en los dos cultivos de mandarino sometidos a distinto manejo agrícola. Letras minúsculas diferentes indican diferencias significativas entre categorías ecológicas para cada manejo y las mayúsculas indican diferencias significativas entre manejos para cada categoría ecológica (p < 0,05).
3.1.2.4.4. Cultivos de olivo
Se identificaron un total de 5 especies en cada una de las dos fincas, de las cuales 4 fueron
exclusivas de cada manejo agrícola y O. cyaneum estuvo presente en ambos (Fig. 46). Aunque
no se detectaron diferencias significativas entre manejos para las abundancias de cada especie,
se puede señalar que O. cyaneum fue la especie dominante en el sistema convencional,
representando más del 50% de las abundancias totales, mientras que en el sistema orgánico no
hubo una dominancia de una única especie sino que las cuatro especies exclusivas de este
sistema agrícola (A. trapezoides, A. caliginosa, A. rosea y S. gigas) alcanzaron representaciones
similares (ente un 20% y 30%).
aA
aBbA
aA a0
50
100
150
200
250
Convencional Orgánico
Ab
un
dan
cia
me
dia
(in
d. m
-2)
Anécicas
Endogeas
Epiendogeas

Capítulo II
95
Figura 46: Abundancia relativa (%) de las diferentes especies de lombrices encontradas en los cultivos de olivo sometidos a diferentes manejos agrícolas.
La estructura ecológica de las poblaciones de lombrices en estos cultivos estuvo únicamente
constituida por lombrices anécicas y endogeas. Aunque las diferencias en las abundancias de
estas dos categorías ecológicas entre manejos no fueron significativas, las lombrices endogeas
fueron más abundantes en el olivo con manejo convencional, mientras que en la finca de olivo
con manejo orgánico estas dos categorías tuvieron representaciones similares (Fig. 47).
Figura 47: Valores medios (±E.E.) de abundancia (ind. m-2) de las distintas categorías ecológicas de lombrices en los dos cultivos de olivo sometidos a distinto manejo agrícola. Letras minúsculas diferentes indican diferencias significativas entre categorías ecológicas para cada manejo y las mayúsculas indican diferencias significativas entre manejos para cada categoría ecológica (p < 0,05).
0%
20%
40%
60%
80%
100%
Ap
orr
ect
od
ea
tra
pe
zoid
es
Lum
bri
cus
terr
est
ris
Oct
od
rilu
s co
mp
lan
atu
s
Sch
ero
the
ca g
iga
s
Ap
orr
ect
od
ea
ca
lig
ino
sa
Ap
orr
ect
od
ea
ro
sea
Oct
ola
sio
n c
ya
ne
um
Oct
ola
sio
n l
act
eu
m
Ab
un
dan
cia
rela
tiva
(%
)
Orgánico
Convencional
ANÉCICAS ENDOGEAS
aA
aA
aA
aA
0
5
10
15
20
25
30
Convencional Orgánico
Ab
un
dan
cia
me
dia
(in
d. m
-2)
Anécicas
Endogeas

Capítulo II
96
3.2. Efectos de las prácticas agricolas sobre las abundancias de los grupos edáficos de meso y macrofauna dominantes en los cultivos atlánticos y mediterráneos
Como se indicó en la sección de materiales y métodos, con el fin de incrementar la robustez
de los análisis estadísticos y poder analizar con más detalle el efecto de los factores investigados
sobre la estructura de las comunidades edáficas en cada una de las dos regiones, se repitieron
los análisis estadísticos eliminando aquellos taxones con baja representatividad (<20%), de tal
forma que solamente se incluyeron las abundancias de siete taxones de mesofauna (ácaros
totales y los órdenes Oribatida y Mesostigmata, colémbolos totales y los subordenes
Poduromorpha y Symphypleona así como la familia Enchytraeidae) y cinco de macrofauna
(familia Formicidae, lombrices totales así como tres categorías ecológicas de lombrices de
tierra). Como se observa en la Tabla 13, de los siete taxones de mesofauna solamente cinco
estuvieron presentes en los cultivos de las dos regiones investigadas, mientras que los ácaros
Mesostigmata y los colémbolos Symphypleona solo estuvieron bien representados en una de
las dos regiones. De manera similar, en el caso de la macrofauna, la familia Formicidae y las
lombrices epigeas tuvieron escasa representación en las regiones atlántica y mediterránea,
respectivamente.
Tabla 13: Taxones con un porcentaje de presencia superior al 20% en el total de muestras recogidas en cada una de las regiones estudiadas.
Mesofauna Región
Atlántica (%) Región
Mediterránea (%)
Acari 45,17 83,32
Oribatida 40,35 80,95
Mesostigmata (=Gamasida) 20,23
Collembola 44,73 78,57
Poduromorpha 37,71 76,19
Symphypleona 20,17
Enchytraeidae 50,87 58,33
Macrofauna Región
Atlántica (%) Región
Mediterránea (%)
Formicidae 30,95
Lombrices Totales 50 76,19
Lombrices Epigeas 21,05
Lombrices Anécicas 23,24 63,09
Lombrices Endogeas 29,82 35,71
Los resultados del análisis ANOVA para aquellos grupos bien representados en ambas
regiones (Tabla 14) indicaron que, salvo las lombrices totales, todos los grupos faunísticos de
mesofauna resultaron significativamente afectados por la región biogeográfica y en el caso de
los colémbolos y lombrices totales, también se vieron afectados por la interacción
región*manejo (Tabla 14).
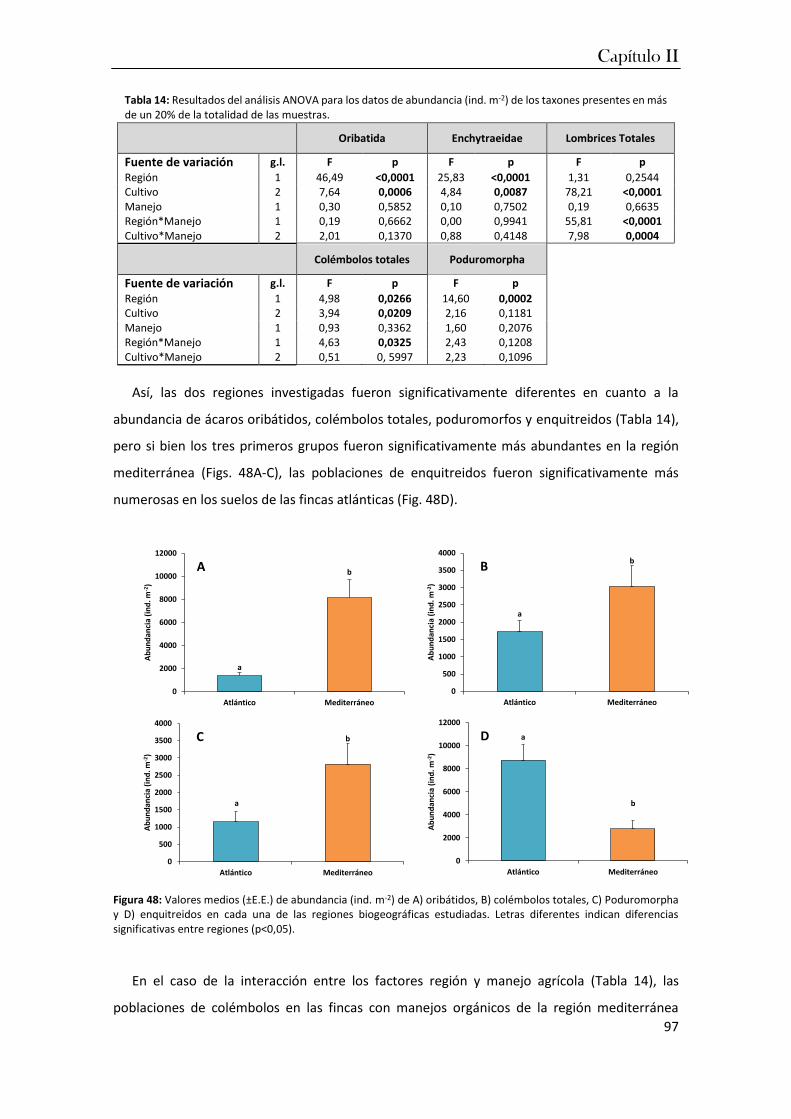
Capítulo II
97
Tabla 14: Resultados del análisis ANOVA para los datos de abundancia (ind. m-2) de los taxones presentes en más de un 20% de la totalidad de las muestras.
Oribatida Enchytraeidae Lombrices Totales
Fuente de variación g.l. F p F p F p
Región 1 46,49 <0,0001 25,83 <0,0001 1,31 0,2544 Cultivo 2 7,64 0,0006 4,84 0,0087 78,21 <0,0001 Manejo 1 0,30 0,5852 0,10 0,7502 0,19 0,6635 Región*Manejo 1 0,19 0,6662 0,00 0,9941 55,81 <0,0001 Cultivo*Manejo 2 2,01 0,1370 0,88 0,4148 7,98 0,0004
Colémbolos totales Poduromorpha
Fuente de variación g.l. F p F p
Región 1 4,98 0,0266 14,60 0,0002 Cultivo 2 3,94 0,0209 2,16 0,1181 Manejo 1 0,93 0,3362 1,60 0,2076 Región*Manejo 1 4,63 0,0325 2,43 0,1208 Cultivo*Manejo 2 0,51 0, 5997 2,23 0,1096
Así, las dos regiones investigadas fueron significativamente diferentes en cuanto a la
abundancia de ácaros oribátidos, colémbolos totales, poduromorfos y enquitreidos (Tabla 14),
pero si bien los tres primeros grupos fueron significativamente más abundantes en la región
mediterránea (Figs. 48A-C), las poblaciones de enquitreidos fueron significativamente más
numerosas en los suelos de las fincas atlánticas (Fig. 48D).
Figura 48: Valores medios (±E.E.) de abundancia (ind. m-2) de A) oribátidos, B) colémbolos totales, C) Poduromorpha y D) enquitreidos en cada una de las regiones biogeográficas estudiadas. Letras diferentes indican diferencias significativas entre regiones (p<0,05).
En el caso de la interacción entre los factores región y manejo agrícola (Tabla 14), las
poblaciones de colémbolos en las fincas con manejos orgánicos de la región mediterránea
a
b
0
2000
4000
6000
8000
10000
12000
Atlántico Mediterráneo
Ab
un
dan
cia
(in
d. m
-2)
A
a
b
0
500
1000
1500
2000
2500
3000
3500
4000
Atlántico Mediterráneo
Ab
un
dan
cia
(in
d. m
-2)
B
a
b
0
500
1000
1500
2000
2500
3000
3500
4000
Atlántico Mediterráneo
Ab
un
dan
cia
(in
d. m
-2)
C a
b
0
2000
4000
6000
8000
10000
12000
Atlántico Mediterráneo
Ab
un
dan
cia
(in
d. m
-2)
D
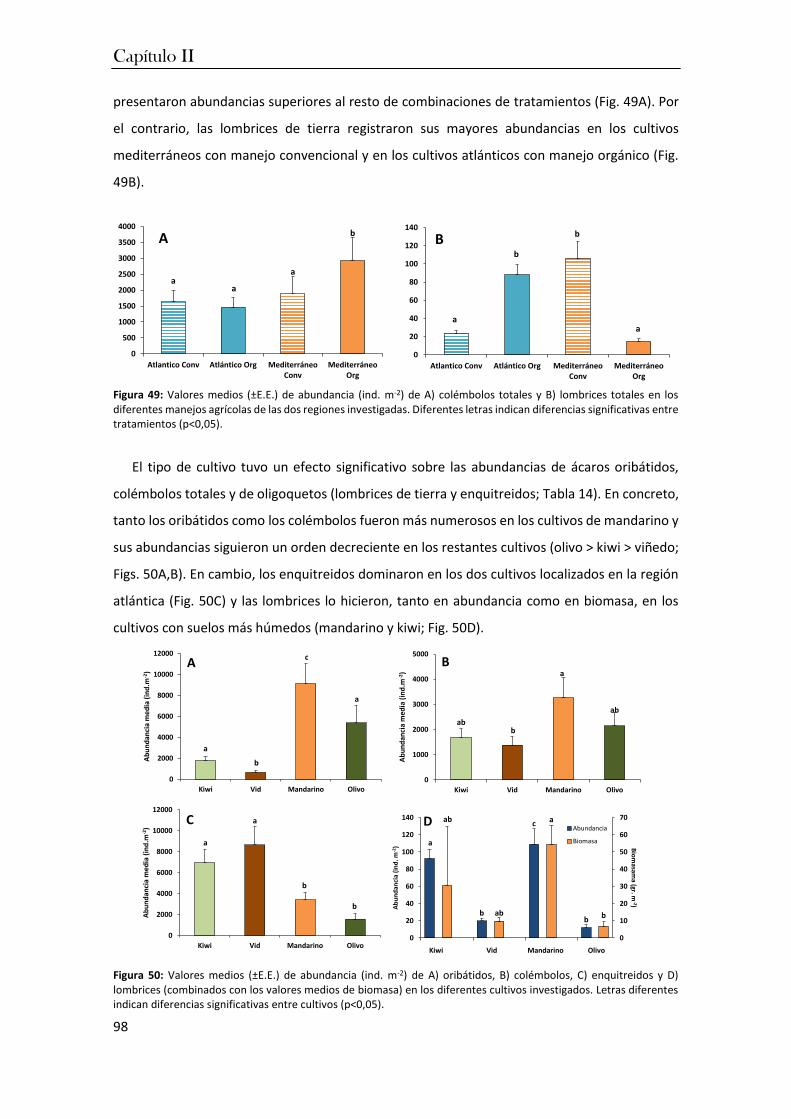
Capítulo II
98
presentaron abundancias superiores al resto de combinaciones de tratamientos (Fig. 49A). Por
el contrario, las lombrices de tierra registraron sus mayores abundancias en los cultivos
mediterráneos con manejo convencional y en los cultivos atlánticos con manejo orgánico (Fig.
49B).
Figura 49: Valores medios (±E.E.) de abundancia (ind. m-2) de A) colémbolos totales y B) lombrices totales en los diferentes manejos agrícolas de las dos regiones investigadas. Diferentes letras indican diferencias significativas entre tratamientos (p<0,05).
El tipo de cultivo tuvo un efecto significativo sobre las abundancias de ácaros oribátidos,
colémbolos totales y de oligoquetos (lombrices de tierra y enquitreidos; Tabla 14). En concreto,
tanto los oribátidos como los colémbolos fueron más numerosos en los cultivos de mandarino y
sus abundancias siguieron un orden decreciente en los restantes cultivos (olivo > kiwi > viñedo;
Figs. 50A,B). En cambio, los enquitreidos dominaron en los dos cultivos localizados en la región
atlántica (Fig. 50C) y las lombrices lo hicieron, tanto en abundancia como en biomasa, en los
cultivos con suelos más húmedos (mandarino y kiwi; Fig. 50D).
Figura 50: Valores medios (±E.E.) de abundancia (ind. m-2) de A) oribátidos, B) colémbolos, C) enquitreidos y D) lombrices (combinados con los valores medios de biomasa) en los diferentes cultivos investigados. Letras diferentes indican diferencias significativas entre cultivos (p<0,05).
aa
a
b
0
500
1000
1500
2000
2500
3000
3500
4000
Atlantico Conv Atlántico Org MediterráneoConv
MediterráneoOrg
A
a
b
b
a
0
20
40
60
80
100
120
140
Atlantico Conv Atlántico Org MediterráneoConv
MediterráneoOrg
B
a
b
c
a
0
2000
4000
6000
8000
10000
12000
Kiwi Vid Mandarino Olivo
Ab
un
dan
cia
med
ia (
ind
.m-2
)
A
abb
a
ab
0
1000
2000
3000
4000
5000
Kiwi Vid Mandarino Olivo
Ab
un
dan
cia
med
ia (
ind
.m-2
)
B
a
a
b
b
0
2000
4000
6000
8000
10000
12000
Kiwi Vid Mandarino Olivo
Ab
un
dan
cia
med
ia (
ind
.m-2
)
C
a
b
c
b
ab
ab
a
b
0
10
20
30
40
50
60
70
0
20
40
60
80
100
120
140
Bio
masam
a (gr. m-2)
Ab
un
dan
cia
(in
d. m
-2)
Abundancia
Biomasa
D
Kiwi Vid Mandarino Olivo
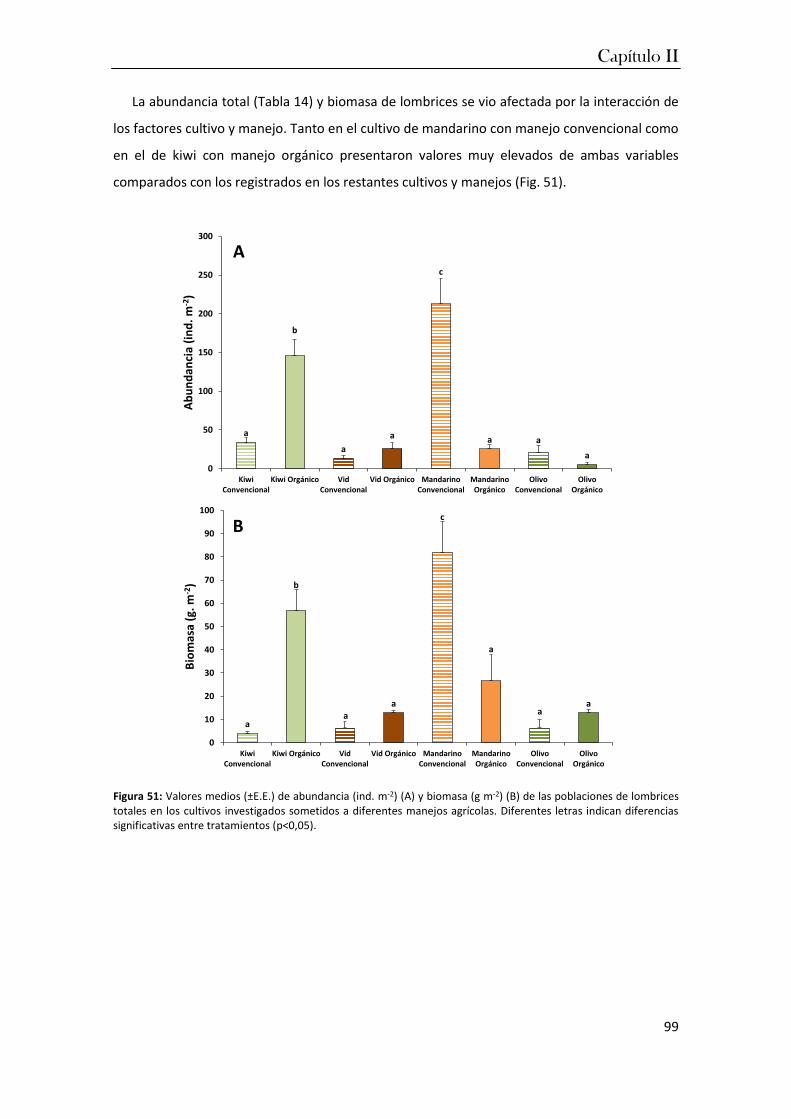
Capítulo II
99
La abundancia total (Tabla 14) y biomasa de lombrices se vio afectada por la interacción de
los factores cultivo y manejo. Tanto en el cultivo de mandarino con manejo convencional como
en el de kiwi con manejo orgánico presentaron valores muy elevados de ambas variables
comparados con los registrados en los restantes cultivos y manejos (Fig. 51).
Figura 51: Valores medios (±E.E.) de abundancia (ind. m-2) (A) y biomasa (g m-2) (B) de las poblaciones de lombrices totales en los cultivos investigados sometidos a diferentes manejos agrícolas. Diferentes letras indican diferencias significativas entre tratamientos (p<0,05).
a
b
a
a
c
a a
a
0
50
100
150
200
250
300
KiwiConvencional
Kiwi Orgánico VidConvencional
Vid Orgánico MandarinoConvencional
MandarinoOrgánico
OlivoConvencional
OlivoOrgánico
Ab
un
dan
cia
(in
d.
m-2
)
A
a
b
a
a
c
a
aa
0
10
20
30
40
50
60
70
80
90
100
KiwiConvencional
Kiwi Orgánico VidConvencional
Vid Orgánico MandarinoConvencional
MandarinoOrgánico
OlivoConvencional
OlivoOrgánico
Bio
mas
a (g
. m-2
)
B
Capítulo II
100
3.3. Efectos de las prácticas agricolas sobre la diversidad y estructura de las comunidades de lombrices de tierra en los cultivos atlánticos y mediterráneos
3.3.1. Índices de diversidad
3.3.1.1. Riqueza de especies
El análisis ANOVA aplicado al conjunto de datos obtenidos en este estudio indicó la ausencia
de efectos significativos de la región, tipo de cultivo, manejo y de la interacción entre estos
factores sobre el número de especies de lombrices identificadas (Tabla 6). De forma similar,
tampoco se detectaron efectos significativos de estos tres factores, ni de sus interacciones,
sobre la riqueza específica de lombrices en cada una de las regiones biogeográficas (Tabla 15).
Sin embargo, el tiempo resultó ser el único factor que afectó significativamente el valor de
SMensual de las comunidades de lombrices en el caso de la región mediterránea (Tabla 15).
Tabla 15: Resultados de ANOVA para SMensual
Riqueza mensual Riqueza mensual en la
Región Atlántica Riqueza mensual en la Región Mediterránea
Fuente de variación g.l. F p g.l. F p g.l. F p
Región 1 0,03 0,852 - - - - - - Cultivo - - - 1 2,72 0,103 1 1,82 0,186 Manejo 1 1,56 0,214 1 1,04 0,310 1 0,00 0,980 Tiempo - - - 11 0,72 0,541 6 3,89 0,002 Región*Manejo 1 2,20 0,150 - - - - - - Cultivo*Manejo - - - 1 2,21 0,141 1 0,00 0,980 Tiempo*Cultivo - - - 11 0,96 0,418 6 1,61 0,160 Tiempo*Manejo - - - 11 0,56 0,644 6 0,31 0,929 Tiempo*Cultivo*Manejo - - - 11 0,41 0,746 6 0,91 0,491
Es destacable que la finca con mayor riqueza de especies (SMensual y STotal) fue el mandarino
orgánico (Tabla 16). Además, tres de las fincas atlánticas (los dos cultivos de kiwi sometidos a
ambos manejos y el de vid convencional) presentaron el mismo valor de STotal (7). En cambio, el
cultivo orgánico de vid y las fincas de olivo orgánico y mandarino convencional registraron los
valores más bajos (Tabla 16).
Tabla 16: Índices de riqueza de especies en los distintos cultivos sometidos a los dos manejos agrícolas (CONV y ORG).
Índices de diversidad KIWI VID MANDARINO OLIVO CONV ORG CONV ORG CONV ORG CONV ORG
Riqueza de Especies Total (STotal) 7 7 7 5 5 8 4 5
Riqueza de Especies Media Mensual (SMensual) 3 3,5 1,75 1,92 2,71 3,43 1,29 1,29
Riqueza de Especies por cultivo 9 8 9 8

Capítulo II
101
3.3.1.2. Índice de diversidad de Shannon
Al igual que para los valores de riqueza específica, la región no tuvo un efecto significativo
sobre los valores de este índice de diversidad (Tabla 17). Sin embargo, el tipo de manejo agrícola
de los cultivos y la interacción de este factor con el de región biogeográfica tuvieron un efecto
significativo sobre los valores del índice de diversidad de Shannon (Tabla 17).
Tabla 17: Resultados de ANOVA para H´Mensual.
Diversidad mensual Diversidad mensual en
la Región Atlántica Diversidad mensual en la
Región Mediterránea
Fuente de variación g.l. F p g.l. F p g.l. F p
Región 1 0,18 0,665 - - - - - - Cultivo - - - 1 0,54 0,462 1 1,15 0,290 Manejo 1 5,94 0,016 1 0,69 0,407 1 7,62 0,009 Tiempo - - - 11 0,28 0,837 6 2,57 0,028 Región*Manejo 1 13,58 <0,001 - - - - - - Cultivo*Manejo - - - 1 0,00 0,989 1 0,37 0,543 Tiempo*Cultivo - - - 11 0,60 0,614 6 2,04 0,075 Tiempo*Manejo - - - 11 0,11 0,951 6 0,23 0,963 Tiempo*Cultivo*Manejo - - - 11 1,75 0,171 6 0,47 0,821
Analizando por separado los valores de H´ obtenidos en cada región, se observó que en el
caso de la región atlántica los valores de H´Total estuvieron comprendidos entre 0,69 en el kiwi
orgánico y 1,51 del kiwi convencional (Tabla 18), sin que ninguno de los factores investigados ni
sus interacciones tuviesen un efecto significativo sobre los valores de este índice de diversidad
(Tabla 17).
En el caso de los suelos mediterráneos, el rango de valores de H´Total fue de entre 0,59 del
mandarino convencional y 1,45 del olivo orgánico (Tabla 18).
Tabla 18: Índices de diversidad de Shannon en los distintos cultivos sometidos a los dos manejos agrícolas (CONV y ORG).
Índice KIWI VID MANDARINO OLIVO
CONV ORG CONV ORG CONV ORG CONV ORG
Índice Shannon Medio (H´Mensual) 0,83 0,62 0,39 0,41 0,51 0,84 0,07 0,30
Índice Shannon Total (H´Total) 1,51 0,69 1,44 1,17 0,59 1,32 1,06 1,45
El análisis de la ANOVA aplicado a los valores de diversidad de las muestras mediterráneas
indicó que tanto el manejo agrícola como tiempo tuvieron efectos significativos sobre los valores
de H´ (Tabla 17). Así, los cultivos con manejo orgánico presentaron una mayor diversidad de
especies que los cultivos convencionales (Fig. 52).

Capítulo II
102
Figura 52: Valores medios (±E.E.) del índice de diversidad de Shannon (H´Mensual) registrados en cultivos de la región mediterránea sometidos a distinto manejo agrícola. Letras diferentes indican diferencias significativas entre manejos (p < 0,05).
3.3.1.3. Índice de equitabilidad de Pielou (J´)
Los análisis estadísticos indicaron que el tipo de manejo tuvo un efecto significativo sobre los
valores del índice J´, así como también lo tuvo la interacción entre el factor región y el tipo de
manejo agrícola (Tabla 19). Así, el manejo agrícola convencional fue más equitativo que el
orgánico (Fig. 53A).
En cuanto a la interacción entre región y manejo agrícola, los valores del índice J´ en los suelos
de cultivo con el manejo convencional en la región atlántica y con manejo orgánico en la región
mediterránea fueron superiores a los valores medidos en los cultivos orgánicos de la región
atlántica y los convencionales de la región mediterránea (Fig. 53B).
En las fincas atlánticas se observó la existencia de interacciones significativas entre cultivo y
manejo y entre fecha, cultivo y manejo sobre los valores del índice J´ (Tabla 19). La finca con
mayor valor de índice J´ fue el kiwi convencional seguida del viñedo orgánico, el viñedo
convencional y finalmente, el kiwi orgánico con un valor muy bajo de J´ (Fig. 53C).
En el caso de los valores obtenidos para las fincas mediterráneas, no fue posible analizar
todas las interacciones entre factores debido al elevado número de casos en los que este índice
obtuvo el valor 0. Sin embargo, se pudo constatar que el manejo agrícola tuvo una influencia
significativa sobre los valores medios del índice J´ (Tabla 19) y en el sistema orgánico el valor de
este índice fue significativamente mayor que el obtenido en el manejo convencional (Fig. 53D).
b
a
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
OrgánicoConvencional
Índ
ice
de
div
ers
idad
de
Sh
ann
on
(H´)
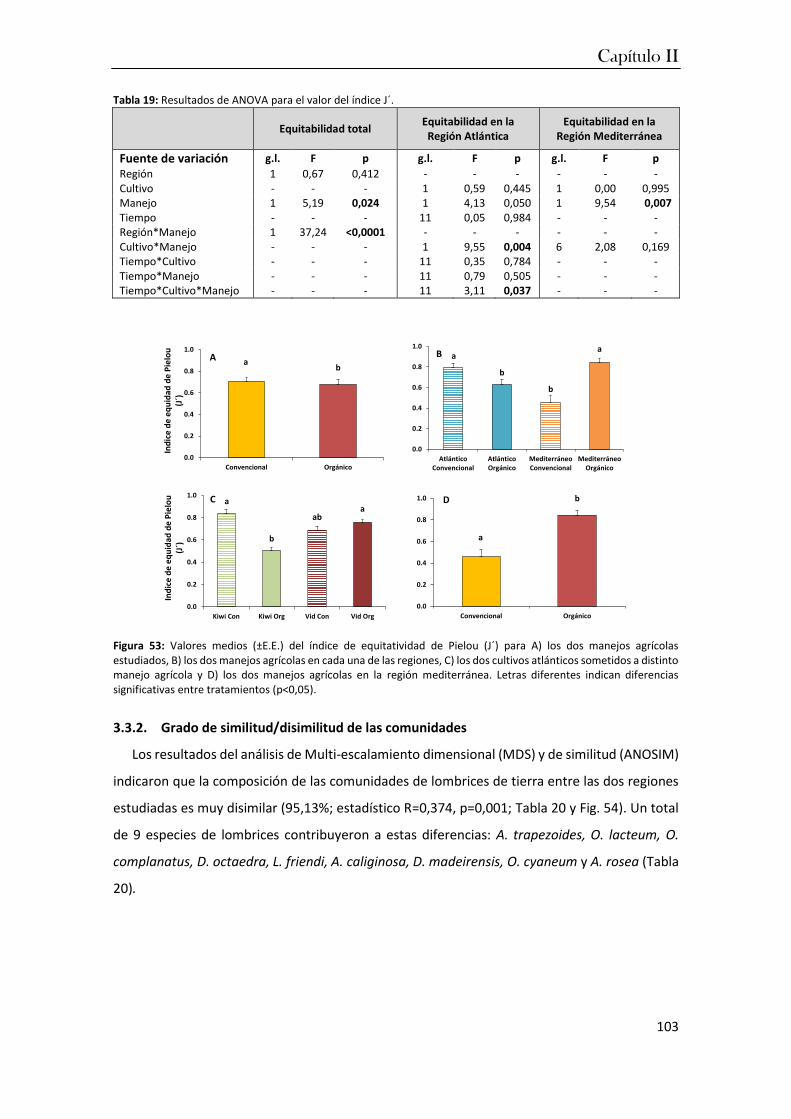
Capítulo II
103
Tabla 19: Resultados de ANOVA para el valor del índice J´.
Equitabilidad total Equitabilidad en la
Región Atlántica Equitabilidad en la
Región Mediterránea
Fuente de variación g.l. F p g.l. F p g.l. F p
Región 1 0,67 0,412 - - - - - - Cultivo - - - 1 0,59 0,445 1 0,00 0,995 Manejo 1 5,19 0,024 1 4,13 0,050 1 9,54 0,007 Tiempo - - - 11 0,05 0,984 - - - Región*Manejo 1 37,24 <0,0001 - - - - - - Cultivo*Manejo - - - 1 9,55 0,004 6 2,08 0,169 Tiempo*Cultivo - - - 11 0,35 0,784 - - - Tiempo*Manejo - - - 11 0,79 0,505 - - - Tiempo*Cultivo*Manejo - - - 11 3,11 0,037 - - -
Figura 53: Valores medios (±E.E.) del índice de equitatividad de Pielou (J´) para A) los dos manejos agrícolas estudiados, B) los dos manejos agrícolas en cada una de las regiones, C) los dos cultivos atlánticos sometidos a distinto manejo agrícola y D) los dos manejos agrícolas en la región mediterránea. Letras diferentes indican diferencias significativas entre tratamientos (p<0,05).
3.3.2. Grado de similitud/disimilitud de las comunidades
Los resultados del análisis de Multi-escalamiento dimensional (MDS) y de similitud (ANOSIM)
indicaron que la composición de las comunidades de lombrices de tierra entre las dos regiones
estudiadas es muy disimilar (95,13%; estadístico R=0,374, p=0,001; Tabla 20 y Fig. 54). Un total
de 9 especies de lombrices contribuyeron a estas diferencias: A. trapezoides, O. lacteum, O.
complanatus, D. octaedra, L. friendi, A. caliginosa, D. madeirensis, O. cyaneum y A. rosea (Tabla
20).
b
a
0.0
0.2
0.4
0.6
0.8
1.0
OrgánicoConvencional
Da
b
aba
0.0
0.2
0.4
0.6
0.8
1.0
Kiwi Con Kiwi Org Vid Con Vid Org
Ind
ice
de
eq
uid
ad d
e P
ielo
u(J
´)
C
ba
0.0
0.2
0.4
0.6
0.8
1.0
OrgánicoConvencional
Ind
ice
de
eq
uid
ad d
e P
ielo
u(J
´)
A a
b
b
a
0.0
0.2
0.4
0.6
0.8
1.0
AtlánticoConvencional
AtlánticoOrgánico
MediterráneoConvencional
MediterráneoOrgánico
B
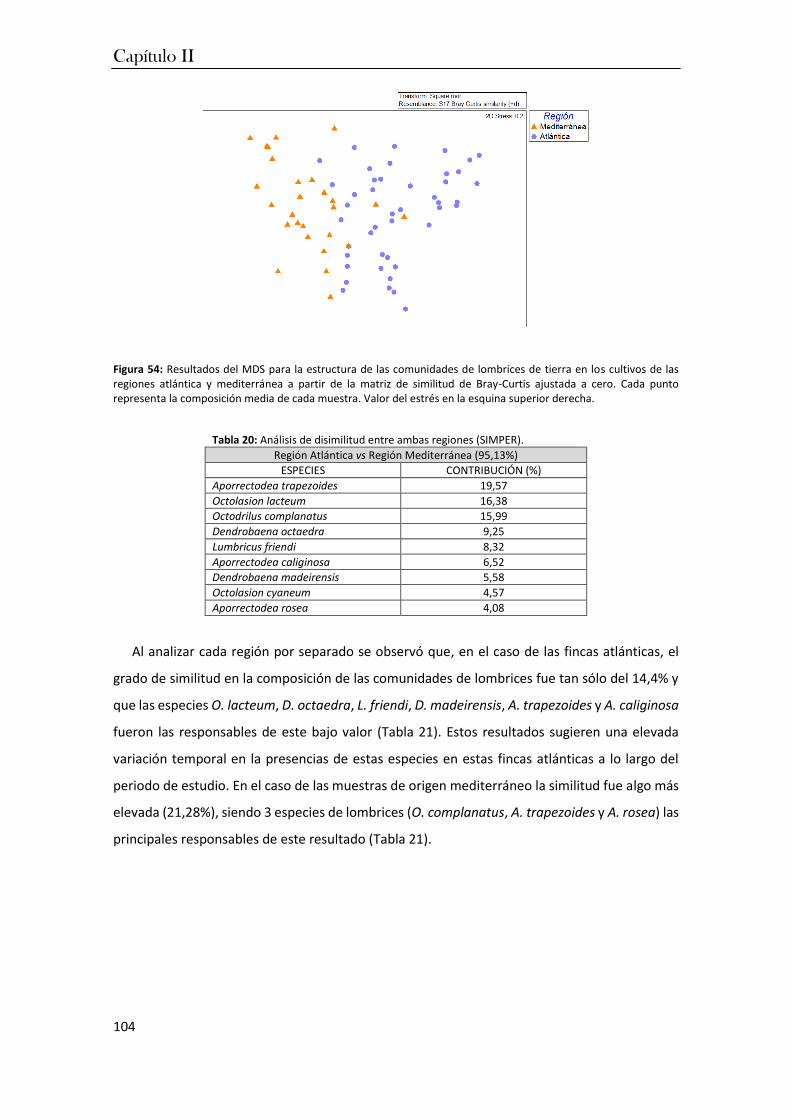
Capítulo II
104
Figura 54: Resultados del MDS para la estructura de las comunidades de lombrices de tierra en los cultivos de las regiones atlántica y mediterránea a partir de la matriz de similitud de Bray-Curtis ajustada a cero. Cada punto representa la composición media de cada muestra. Valor del estrés en la esquina superior derecha.
Tabla 20: Análisis de disimilitud entre ambas regiones (SIMPER).
Región Atlántica vs Región Mediterránea (95,13%)
ESPECIES CONTRIBUCIÓN (%)
Aporrectodea trapezoides 19,57
Octolasion lacteum 16,38
Octodrilus complanatus 15,99
Dendrobaena octaedra 9,25
Lumbricus friendi 8,32
Aporrectodea caliginosa 6,52
Dendrobaena madeirensis 5,58
Octolasion cyaneum 4,57
Aporrectodea rosea 4,08
Al analizar cada región por separado se observó que, en el caso de las fincas atlánticas, el
grado de similitud en la composición de las comunidades de lombrices fue tan sólo del 14,4% y
que las especies O. lacteum, D. octaedra, L. friendi, D. madeirensis, A. trapezoides y A. caliginosa
fueron las responsables de este bajo valor (Tabla 21). Estos resultados sugieren una elevada
variación temporal en la presencias de estas especies en estas fincas atlánticas a lo largo del
periodo de estudio. En el caso de las muestras de origen mediterráneo la similitud fue algo más
elevada (21,28%), siendo 3 especies de lombrices (O. complanatus, A. trapezoides y A. rosea) las
principales responsables de este resultado (Tabla 21).

Capítulo II
105
Tabla 21: Resultados del análisis de similitud y contribuciones de las distintas especies de lombrices (SIMPER) en cada una de las regiones biogeográficas. Se muestran las especies que contribuyen al 90% de la similitud acumulada.
Región Atlántica (14,40%) Región Mediterránea (21,28%)
ESPECIES CONTRIBUCIÓN (%) ESPECIES CONTRIBUCIÓN (%)
Octolasion lacteum 33,35 Octodrilus complanatus 55,55
Dendrobaena octaedra 24,69 Aporrectodea trapezoides 34,23
Lumbricus friendi 14,78 Aporrectodea rosea 5,01
Dendrobaena madeirensis 9,79
Aporrectodea trapezoides 6,80
Aporrectodea caliginosa 5,63
Los resultados del análisis de Multi-escalamiento dimensional y de similitud también
indicaron que las comunidades de lombrices en cada una de las regiones fueron
significativamente diferentes entre cultivos y manejos, corroborado por los resultados del
estadístico R del análisis ANOSIM (Tabla 22) y del análisis SIMPER (Tabla 23). En el caso de las
fincas atlánticas, las diferencias en la estructura de las comunidades de lombrices fueron
especialmente evidentes tanto en el caso de las dos fincas de kiwi sometidas a distinto manejo
(84%, Tabla 23 y Fig. 55, clústeres A y B) como de los dos cultivos de viñedo (> 90%, Tabla 23 y
Fig. 55, clústeres C y D).
Figura 55: Resultados del MDS para la estructura de las comunidades de lombrices de tierra en los cultivos atlánticos sometidos a distintos manejos agrícolas a partir de la matriz de similitud de Bray-Curtis ajustada a cero. Cada punto representa la composición media de cada muestra. Las elipses indican agrupamientos con similitud superior al 40% (p < 0,05). Valor del estrés en la esquina superior derecha.
En el caso de los cultivos de kiwi la elevada disimilitud entre los manejos convencional y
orgánico de kiwi fue debida a la mayor abundancia de O. lacteum en kiwi orgánico, la ausencia
de L. friendi en kiwi convencional y la mayor abundancia de D. octaedra en kiwi convencional
(Tabla 23). Sin embargo, las comunidades de lombrices encontradas en estos cultivos de kiwi,
A
B
C
D
E
Capítulo II
106
en ambos manejos, apenas sufrieron cambios a lo largo del tiempo y en el caso de sistema
convencional el porcentaje de similitud alcanzó el valor de 46,49% (Tabla 23) y las especies que
contribuyeron a esta similitud fueron D. octaedra y D. rubidus. En el caso de la finca orgánica la
similitud fue todavía más elevada, del 72,89% (Tabla 23) y las especies que más contribuyeron
fueron L. friendi y O. lacteum, siendo esta última la especie con mayor contribución (Tabla 23).
Tabla 22: Estadístico R del análisis de similitud (ANOSIM) y su nivel de significación para las dos regiones y para las fincas atlánticas y mediterráneas sometidas a
distintos manejos agrícolas. Estadístico R p
Región Atlántica 0,70 0,001
Kiwi Convencional vs. Kiwi Orgánico 0,91 0,001
Kiwi Orgánico vs. Vid Orgánico 0,77 0,001
Vid Convencional vs. Kiwi Orgánico 0,94 0,001
Kiwi Convencional vs. Vid Orgánico 0,83 0,001
Kiwi Convencional vs. Vid Convencional 0,44 0,001
Vid Convencional vs. Vid Orgánico 0,22 0,015
Región Mediterránea 0,60 0,001
Mandarino Convencional vs. Mandarino Orgánico 0,80 0,001
Mandarino Orgánico vs. Olivo Orgánico 0,66 0,001
Olivo Convencional vs. Mandarino Orgánico 0,25 0,006
Mandarino Convencional vs. Olivo Orgánico 0,87 0,003
Mandarino Convencional vs. Olivo Convencional 0,76 0,003
Olivo Convencional vs. Olivo Orgánico 0,34 0,008
En el caso de los dos viñedos, la disimilitud en las comunidades de lombrices entre los dos
manejos fue mucho mayor que la de los cultivos de kiwi, si bien la composición específica fue
muy similar en estas dos fincas (Tabla 23). Esto es debido a que la especie L. friendi no está
presente en el viñedo convencional, convirtiéndose en una de las especies que más contribuyó
a las diferencias observadas (Tabla 23).
Por otro lado, el porcentaje de similitud entre las comunidades de lombrices de estos cultivos
es mucho menor que la registrada los cultivos de kiwi (17,03%, Tabla 23), indicando una gran
variación en la composición de esta comunidad a lo largo del periodo de estudio. Dos especies,
D. madeirensis y A. caliginosa fueron las que más contribuyeron al porcentaje de similitud. En el
caso del viñedo orgánico, la similitud fue algo más elevada (21,39%, Tabla 23) atribuido a las
especies O. lacteum y L. friendi.

Capítulo II
107
Tabla 23: Resultados del análisis de similitud y disimilitud así como las contribuciones de las distintas especies de lombrices (SIMPER). Se muestran las especies que contribuyen al 70% de la similitud/disimilitud acumulada.
SIMPER Similitud Kiwi Convencional
(46,49%, Contribución >13%) Vid Convencional
(17,03, Contribución >14%) ESPECIES CONTRIBUCIÓN (%) ESPECIES CONTRIBUCIÓN (%) Dendrobaena octaedra 66,61 Dendrobaena madeirensis 61,46
Dendrodrilus rubidus 13,02 Aporrectodea caliginosa 14,59
Kiwi Orgánico (72,89%, Contribución >23%)
Vid Orgánico (21,39%, Contribución > 32%)
ESPECIES CONTRIBUCIÓN (%) ESPECIES CONTRIBUCIÓN (%) Octolasion lacteum 66,85 Octolasion lacteum 46,14
Lumbricus friendi 23,22 Lumbricus friendi 32,85
Mandarino Convencional (75,81%, Contribución >20%)
Olivo Convencional (16,35%, Contribución=100%)
Aporrectodea trapezoides 75,22 Octodrilus complanatus 100
Mandarino Orgánico (49,03%, Contribución >10%)
Olivo Orgánico (13,13%, Contribución > 20%)
ESPECIES CONTRIBUCIÓN (%) ESPECIES CONTRIBUCIÓN (%)
Octodrilus complanatus 78,50 Aporrectodea trapezoides 71,27
SIMPER Disimilitud
Kiwi Convencional vs Kiwi Orgánico (84,46%, Contribución >12%)
Vid Convencional vs Vid Orgánico (90,84%, Contribución >12%)
ESPECIES CONTRIBUCIÓN (%) ESPECIES CONTRIBUCIÓN (%)
Octolasion lacteum 41,52 Dendrobaena madeirensis 23,22
Lumbricus friendi 18,22 Lumbricus friendi 18,7
Dendrobaena octaedra 12,25 Octolasion lacteum 17,69
Aporrectodea trapezoides 12,7
Mandarino Convencional vs Mandarino Orgánico (64,68%, Contribución >8%)
Olivo Convencional vs Olivo Orgánico (99,73%, Contribución >5%)
ESPECIES CONTRIBUCIÓN (%) Octodrilus complanatus 30,23
Aporrectodea trapezoides 63,79 Aporrectodea trapezoides 13,63
Octodrilus complanatus 8,43 Octolasion cyaneum 11,48
Octolasion lacteum 9,43
Dendrobaena madeirensis 9
En el caso de las fincas mediterráneas, los resultados del MDS también reflejaron una cierta
diferenciación entre cultivos y manejos (Fig. 56), corroborado por los resultados del estadístico
R (Tabla 22) y siendo las comunidades de lombrices de las fincas de mandarino más similares
entre sí (Fig. 56, clústeres A y B) que las encontradas en las fincas dedicadas al cultivo de olivo
(Fig. 56, clústeres C y D). De hecho, las comunidades de lombrices de dos muestreos en las fincas
de olivo (4 de manejo convencional y 1 de orgánico) no pudieron ser incluidas en ningún clúster
por no llegar a alcanzar el valor mínimo del índice de similitud necesario (Fig. 56), a pesar de que
el estadístico R indicó una separación significativa entre estas fincas (Tabla 22).

Capítulo II
108
Figura 56: Resultados del MDS para la estructura de las comunidades de lombrices de tierra en los cultivos mediterráneos sometidos a distintos manejos agrícolas a partir de la matriz de similitud de Bray-Curtis ajustada a cero. Cada punto representa la composición media de cada muestra. Las elipses indican agrupamientos con similitud superior al 40% (p<0,05). Valor del estrés en la esquina superior derecha.
En cuanto al manejo agrícola, y para el caso del mandarino, la diferencia entre los dos
sistemas agrícolas alcanzó el 64%, siendo A. trapezoides la especie que más contribuyó a las
diferencias observadas (Tabla 23). En cambio, el grado de similitud en la comunidad de lombrices
presentes en el cultivo de mandarino convencional fue del 75,81% (Tabla 23), lo cual indica que
este cultivo experimentó escasas variaciones a lo largo del periodo de estudio y fue de nuevo la
especie A. trapezoides la que más contribuyó a estas similitudes (Tabla 23). En el caso del
mandarino orgánico se observó un porcentaje de similitud algo menor (49,03%; Tabla 23), lo
que sugiere que también hubo pocas variaciones temporales en esta comunidad de lombrices y
en este caso, la especie O. complanatus presentó la mayor contribución al porcentaje de
similitud medido en esta finca (Tabla 23).
En el caso del cultivo de olivo, el grado de disimilitud entre manejos agrícolas fue todavía
mayor (>99%), posiblemente debido a que sólo una especie (O. cyaneum) estuvo presente en
ambas fincas (Tabla 23). Debido a esto, los porcentajes de similitud fueron bastante bajos en
ambos manejos (Tabla 23), indicando una gran variación de la comunidad entre muestreos. Es
destacable que si bien A. trapezoides fue la especie que más contribuyó a la similitud de la finca
de olivo orgánico, O. complanatus fue la única especie responsable del total de la similitud
observada en la finca convencional (100%; Tabla 23).
A
B
C
D

Capítulo II
109
3.3.3. Comparación curvas abundancia y biomasa (ABC)
Los resultados derivados de comparar las curvas de biomasa y abundancia indican que las
fincas con manejos orgánicos presentaron una dominancia de la curva de biomasa sobre la curva
acumulada de abundancia. En cambio, las fincas dedicadas a la producción agrícola convencional
presentaron resultados más variados.
Así, en el caso de las fincas atlánticas y para el cultivo de kiwi con manejo convencional, la
curva de abundancia domina claramente sobre la curva de biomasa (Fig. 57A) haciendo que el
estadístico W alcance un valor negativo (WCONV=-0,248) lo que indica cierto grado de
perturbación en los suelos de esta finca. Sin embargo, en el cultivo orgánico de kiwi, las dos
curvas se superponen (Fig. 57B), lo que sugiere que la perturbación es moderada (WORG=-0,041).
En el caso de los cultivos de vid y en los dos manejos, la biomasa domina sobre la abundancia
(Figs. 57C,D), indicando una ausencia de perturbación ecológica (WCONV=0,168 y WORG=0,188).
Figura 57: Curvas de abundancia y biomasa de las lombrices de tierra presentes en los cultivos atlánticos de A) kiwi con manejo convencional, B) kiwi con manejo orgánico, C) vid con manejo convencional y D) vid con manejo orgánico.
En el caso de las fincas mediterráneas y en el cultivo de mandarino con manejo convencional,
las curvas de biomasa y abundancia de especies se cruzan y superponen (Fig. 58A), lo que parece
sugerir que este tipo de manejo está algo perturbado (WCONV=-0,046). En cambio, en el caso del
A B
C D
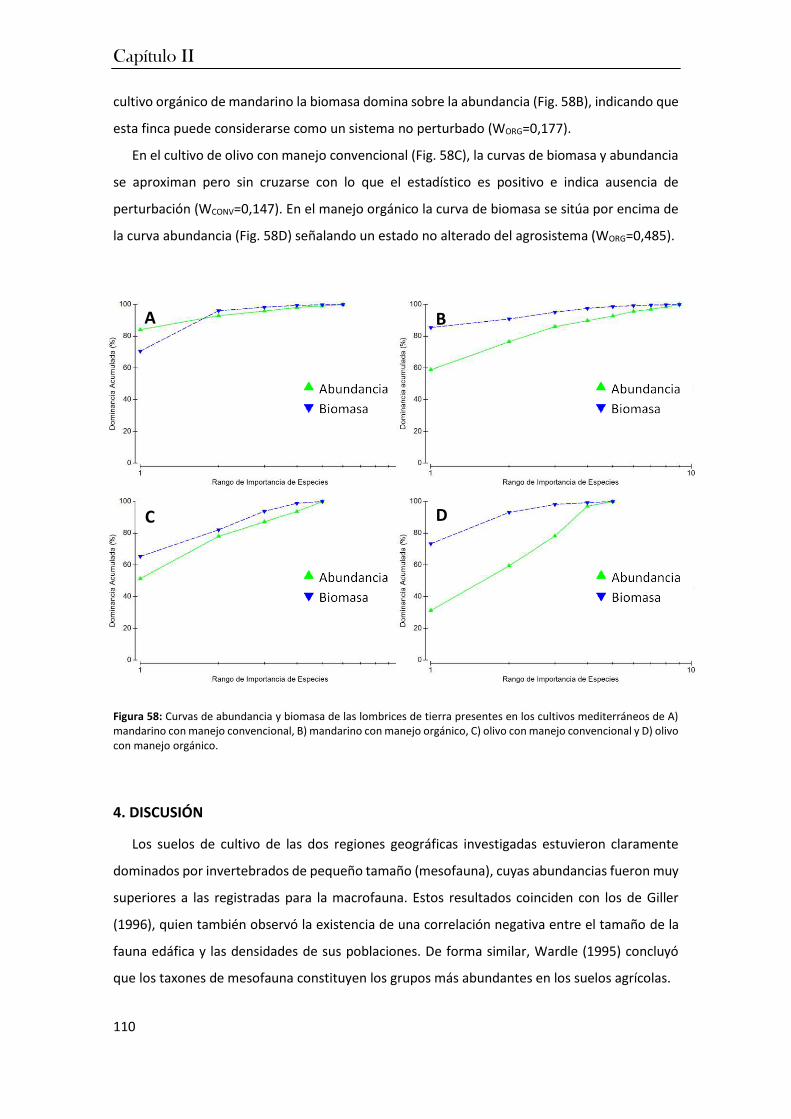
Capítulo II
110
cultivo orgánico de mandarino la biomasa domina sobre la abundancia (Fig. 58B), indicando que
esta finca puede considerarse como un sistema no perturbado (WORG=0,177).
En el cultivo de olivo con manejo convencional (Fig. 58C), la curvas de biomasa y abundancia
se aproximan pero sin cruzarse con lo que el estadístico es positivo e indica ausencia de
perturbación (WCONV=0,147). En el manejo orgánico la curva de biomasa se sitúa por encima de
la curva abundancia (Fig. 58D) señalando un estado no alterado del agrosistema (WORG=0,485).
Figura 58: Curvas de abundancia y biomasa de las lombrices de tierra presentes en los cultivos mediterráneos de A) mandarino con manejo convencional, B) mandarino con manejo orgánico, C) olivo con manejo convencional y D) olivo con manejo orgánico.
4. DISCUSIÓN
Los suelos de cultivo de las dos regiones geográficas investigadas estuvieron claramente
dominados por invertebrados de pequeño tamaño (mesofauna), cuyas abundancias fueron muy
superiores a las registradas para la macrofauna. Estos resultados coinciden con los de Giller
(1996), quien también observó la existencia de una correlación negativa entre el tamaño de la
fauna edáfica y las densidades de sus poblaciones. De forma similar, Wardle (1995) concluyó
que los taxones de mesofauna constituyen los grupos más abundantes en los suelos agrícolas.
A B
C D

Capítulo II
111
Ninguno de los factores investigados (región, tipo de cultivo y el de manejo agrícola) alteró
este contraste entre las abundancias de mesofauna y macrofauna y, aunque sí se observaron
variaciones temporales en las abundancias de estos dos grupos de fauna edáfica, la mesofauna
siempre fue mayoritaria. Estos cambios en las densidades poblacionales a lo largo del tiempo se
asocian a cambios en la disponibilidad de alimento, la presencia de depredadores y a las
variaciones estacionales de las condiciones de temperatura y humedad (Menta et al. 2013). En
el caso de los cultivos atlánticos investigados, las mayores densidades se observaron en el mes
de abril, fecha en la que la precipitación se redujo a casi la mitad del mes anterior y que,
combinado con la aplicación de fertilizantes en la primavera, podría haber propiciado el
incremento de poblaciones de ciertos taxones como los colémbolos, muy sensibles a los aportes
nitrogenados (Bardgett y Cook 1998; Ochoa-Hueso et al. 2014). En la región mediterránea, las
mayores densidades de fauna edáfica fueron registradas tras el periodo estival, en el mes de
septiembre, cuando las temperaturas comenzaron a descender permitiendo la colonización de
las capas más superficiales del suelo (Nannelli y Simoni 2002), donde se concentran los mayores
aportes de materia orgánica fresca.
4.1. Diferencias biogeográficas en la estructura de las comunidades edáficas
4.1.1. Mesofauna
Los resultados de este estudio indican que, si bien los suelos de cultivo frutícola están
dominados por mesofauna, los organismos más numerosos en la región atlántica fueron los
enquitreidos (68% del total de la mesofauna presente en esta región), mientras que los ácaros
constituyeron el grupo mayoritario de la mesofauna en la región mediterránea (55%).
La mayor abundancia relativa de enquitreidos en los cultivos atlánticos podría ser debida a
que estos son organismos especialmente sensibles a los valores de humedad y temperatura del
suelo (O´Connor 1957; Kasprzak 1982, Briones et al. 2007), aunque algunas especies son capaces
de producir formas de resistencia como capullos (Lagerlöf et al. 1989; Dash 1990) y desplazarse
a capas más profundas del perfil (Carrera 2012). Sin embargo, estas son estrategias temporales
y no permiten sobrevivir largos periodos de tiempo si las condiciones adversas persisten durante
largos periodos de tiempo (Briones et al. 1997). Por ello, es bastante probable que las
condiciones edafoclimáticas más favorables registradas en los suelos atlánticos pueden haber
favorecido una mayor proliferación de enquitreidos frente a los demás taxones de mesofauna.
En cambio, la marcada estacionalidad de la región mediterránea es la causa más probable de
que sus abundancias en estos suelos hayan sido muy inferiores a las de los artrópodos. En
concreto, la dominancia de oribátidos sobre el resto de grupos de mesofauna en los cultivos

Capítulo II
112
mediterráneos puede ser atribuida a su mayor capacidad para resistir la desecación (Block 1996;
Lindberg y Bengtsson 2006). Al igual que los oribátidos, la abundancia de colémbolos totales (y
en particular del suborden Poduromorpha) en los cultivos de la región mediterránea fue también
superior a la registrada en la región atlántica. Varios estudios (e.g. Verhoef y Selm 1983; Block
1996; Lindberg y Bengtsson 2006) señalan la capacidad de estos organismos edáficos de tolerar
condiciones de sequía y de recuperarse rápidamente gracias a sus altas tasas de reproducción.
4.1.2. Macrofauna
En el caso de la macrofauna, los análisis de simillaridad (MDS) evidenciaron las diferencias en
la estructura de las poblaciones de lombrices de tierra entre ambas regiones biogeográficas. Así,
de la totalidad de especies recolectadas, cinco de ellas solo fueron encontradas en la región
atlántica: D. octaedra, D. madeirensis, D. rubidus, L. rubellus (epigeas) y L. friendi (anécica) y seis
estuvieron presentes únicamente en la región mediterránea: L. terrestris, O. complanatus, S.
gigas, A. chlorotica, A. rosea y O. cyaneum (las tres primeras pertenecen a la categoría ecológica
de las anécicas y las tres restantes son endogeas). Sin embargo, a pesar estas diferencias entre
las dos comunidades de macrofauna, los análisis univariantes de abundancia y riqueza específica
no mostraron la existencia de diferencias significativas entre las dos regiones, lo que subraya la
necesidad de utilizar varias aproximaciones estadísticas para poder comparar en detalle la
estructura y composición de varias comunidades ecológicas (Clarke 1993).
Las especies epigeas más abundantes en la región atlántica, D. octaedra y D. rubidus, son
especies ampliamente distribuidas en Europa (Boag et al. 1997; Berman et al. 2001) y abundan
en los terrenos agrícolas (Baker et al. 1997). Su elevado polimorfismo genético y su capacidad
para reproducirse partenogenéticamente (Perel 1979; Uvarov 1998; Tiunov et al. 2006) les
facilita la rápida colonización de nuevos hábitats así como la rápida recuperación de sus
poblaciones cuando las condiciones ambientales vuelven a ser favorables.
A diferencia de la región atlántica, en los suelos mediterráneos las lombrices epigeas
estuvieron ausentes, lo que puede atribuirse a que las especies de esta categoría ecológica son
muy sensibles a las temperaturas elevadas y a la desecación, principalmente debido a que
tienden a ocupar los horizontes más superficiales del suelo (Gerard 1967; Lee 1985; Briones et
al. 2009). Además, a diferencia de otros grupos ecológicos, sus limitadas capacidades
excavadoras no les permite refugiarse en las capas más profundas del perfil del suelo durante
las épocas con condiciones ambientales más desfavorables.

Capítulo II
113
4.2. Efecto del tipo de cultivo y manejo agrícola sobre las comunidades de meso y macrofauna
No se observó un efecto significativo del tipo de cultivo ni del manejo agrícola sobre los
grupos mayoritarios de la mesofauna recolectada en los suelos de cultivo investigados
(enquitreidos, oribátidos y colémbolos). Esto puede ser debido a que los invertebrados de
menor tamaño son menos sensibles a las perturbaciones ambientales que los de gran tamaño
(Briones et al. 2009; Postma-Blaauw et al. 2010). Autores como Mazzoncini et al. (2010) tampoco
encontraron diferencias significativas entre las densidades de oribátidos y colémbolos en suelos
sometidos a diferentes manejos agrícolas (tanto orgánico como convencional), señalando que
las características de los suelos y condiciones climáticas típicas de las zonas de estudio son los
principales factores que controlan el tamaño de las poblaciones de estos microartrópodos.
4.2.1. Región atlántica
Las comunidades de lombrices de la región atlántica fueron claramente diferentes según el
tipo de cultivo y manejo (>80% de disimilitud). En el caso de los cultivos de kiwi se identificaron
9 especies, lo que representa un valor muy elevado comparado con los valores de riqueza
específica registrados en cultivos de kiwi de Nueva Zelanda (3 especies; Springett et al. 1994),
con un clima similar. En estos cultivos, O. lacteum fue la especie dominante, una lombriz
endogea que se caracteriza por estar muy bien adaptada a ambientes húmedos, estando
considerada cómo típica de la región atlántica de la Península Ibérica (Trigo et al. 1988;
Rodríguez et al. 1997).
Por el contrario, el reducido número de individuos encontrados en el cultivo de vid coinciden
con los resultados previos obtenidos por Paoletti et al. (1998), dónde el viñedo fue también el
cultivo con menor número de ejemplares y especies identificadas en comparación con otros
cultivos investigados como los de manzano, melocotón y kiwi. El requerimiento de agua por
parte del viñedo es muy bajo, de ahí que el riego está muy restringido en el manejo de estos
cultivos, por lo que es previsible que el contenido de humedad del suelo haya constituido un
factor limitante en la composición de la comunidad de lombrices. Y, por tanto, es posible que
pueda haber favorecido la presencia de especies de pequeño tamaño como D. octaedra y D.
rubidus, que son más tolerantes a las perturbaciones ambientales (Arnold et al. 2008).
El efecto del manejo agrícola sobre las comunidades de lombrices en estos suelos atlánticos
también se vio reflejado en los cambios observados en la estructura ecológica de sus
poblaciones. Así, las lombrices epigeas fueron más abundantes en el manejo convencional que
en el manejo orgánico. Se trata de organismos estrategas de la r (Bouché 1977), lo cual implica
una mayor tasa reproductora y ciclos de vida cortos, características que les permiten una mayor

Capítulo II
114
tasa de supervivencia frente a las perturbaciones ambientales, como son la aplicación de
agroquímicos en los cultivos convencionales (Feldmanstern 2013). De forma similar, estudios
previos realizados en fincas de kiwi (Lago 2015) indican una mayor abundancia de lombrices
epigeas en manejo convencional frente al orgánico, atribuido a su alta tasa reproductiva y a la
mayor cantidad de hojarasca depositada en el suelo sometido a manejo convencional.
Entre las lombrices anécicas, L. friendi estuvo únicamente presente en los cultivos orgánicos.
Su ausencia en los cultivos convencionales coincide con las observaciones realizadas por Schreck
et al. (2012), quienes señalaron una elevada sensibilidad de esta especie a los agroquímicos
empleados en los manejos convencionales. Una característica común de los suelos de los
cultivos convencionales atlánticos fue la escasa profundidad del perfil, lo que también podría
haber limitado las actividades excavadoras de estas lombrices. La única especie anécica
recolectada en las fincas con manejo convencional fue A. trapezoides, especie de menor tamaño
que L. friendi y con elevada capacidad de adaptación a condiciones ambientales extremas
(Fernández 2012).
Finalmente, los valores del estadístico W de las comparaciones de curvas de acumulación de
abundancia/biomasa indican que, aunque los valores para las cuatro fincas estudiadas fueron
próximos a 0 (indicando que estos agrosistemas sufren perturbaciones moderadas), estos
valores fueron negativos en los dos cultivos de kiwi, sobre todo con manejo convencional. Así,
en comparación con el sistema orgánico, el suelo convencional contó con la presencia de
especies de menor biomasa (49 y 718 g en el kiwi convencional y orgánico, respectivamente) y
una mayor dominancia de individuos juveniles frente a los adultos (56% en kiwi convencional
frente al 12% en kiwi orgánico), como resultado de una desviación de la energía hacia la
supervivencia en lugar de hacia el crecimiento individual.
4.2.2. Región mediterránea
En esta región destaca el elevado número de especies encontradas en los cultivos de cítricos
(9 especies en total), lo que contrasta con resultados previos (Dlamini y Haynes 2004;
Kherbouche et al. 2012) en los que se registraron entre 4 y 6 especies. En el caso de los cultivos
de olivo se identificaron 6 especies, lo que coincide con el número de especies identificadas por
Solomou et al. (2012), pero algo superior a las 4 especies identificadas por Kherbouche et al.
(2012).
Además, aunque está establecido que las especies de lombrices se suelen distribuir de forma
equitativa en los cultivos de cítricos (Dlamini y Haynes 2004; Kherbouche et al. 2012), los valores
del índice de uniformidad o equidad de Pielou (J´) en los cultivos mediterráneos de mandarino

Capítulo II
115
son más bajos (0,36) que los estimados para los olivos (que se aproximan a 1), lo que indica un
desequilibrio del reparto de individuos entre especies. En el caso del cultivo de mandarino
investigado se observó una dominancia de la especie A. trapezoides y junto con la presencia de
O. complanatus contribuyeron en gran medida a las diferencias observadas entre este cultivo y
el de olivo (resultados del procedimiento SIMPER). Esta divergencia podría asociarse a la mayor
frecuencia de riego en los cultivos de mandarino, lo que podría haber favorecido el buen
desarrollo de comunidades de lombrices a lo largo de todo el periodo de estudio, tal como ya
ha sido constatado en estudios previos (e.g. Evans y Guild 1948; Gerard 1967; Boag et al. 1997;
Jones et al. 2001; Crumsey et al. 2014). En cambio, en los cultivos de olivo, la mayor dominancia
correspondió a la especie O. cyaneum, cuando otros estudios destacan como especies
dominantes en olivares O. complantaus (Solomou et al. 2012) y A. rosea (Kherbouche et al.
2012).
No se observó una influencia significativa del manejo sobre la riqueza específica de las
poblaciones de lombrices de tierra en los suelos de cultivo mediterráneos. Sin embargo, los
valores del índice de Shannon obtenidos para las fincas investigadas indican que los suelos
sometidos a manejo orgánico contuvieron una comunidad de lombrices significativamente más
diversa que los convencionales. Además, también se observaron diferencias significativas para
los valores del índice de Pielou en función del manejo, presentando los sistemas orgánicos
valores de este índice significativamente mayores que los observados en el manejo
convencional, lo que puede atribuirse a la ausencia del uso de fertilizantes inorgánicos y
pesticidas en estos manejos y a las adiciones de enmiendas orgánicas que proporcionan un
aporte de nutrientes para las poblaciones de lombrices (Paoletti 1999; Hole et al. 2005; Palm et
al. 2013; Bertrand et al. 2015).
El manejo agrícola también tuvo un efecto muy importante sobre la estructura ecológica de
las comunidades de lombrices en los suelos mediterráneos, destacando la elevada abundancia
de individuos de lombrices anécicas en el mandarino convencional, con una clara dominancia
de la especie A. trapezoides (90% de la abundancia total de anécicas). Esto puede ser debido la
adición de abono de ``gallinaza´´, una enmienda muy rica en carbono, nitrógeno, fósforo y
potasio (Estrada Pareja 2005), que proporciona mayor cantidad de nutrientes disponibles para
estas lombrices que se alimentan en superficie (Leroy et al. 2008). La importancia de los abonos
orgánicos como fuente de alimento para las lombrices (Rodríguez et al. 1997; Bilalis et al. 2009;
Kherbouche et al. 2012) puede ser también la razón por la que la especie O. complanatus
(perteneciente también a la categoría ecológica de las anécicas) estuviese ausente en los suelos
de cultivo de olivo con manejo orgánico. Sin embargo, estos resultados contrastan con los

Capítulo II
116
obtenidos para otra especie anécica, S. gigas, capturada únicamente en el cultivo de olivo con
manejo orgánico. Esta especie de lombriz alcanza tamaños grandes (250-300 mm; Bouché 1972)
y tienen una representación muy escasa en el territorio europeo debido a su sensibilidad a las
prácticas agrícolas, por lo que su distribución está considerada como relicta en Europa (Paoletti
1999). Finalmente, L. terrestris, aunque es una especie anécica ampliamente distribuida en
Europa (Dinter et al. 2013), tuvo escasa representación en las fincas mediterráneas,
recogiéndose algunos ejemplares en el cultivo de mandarino orgánico y unos pocos en el olivo
convencional.
Por último, los valores del estadístico W fueron negativos en las fincas con manejos
convencionales y positivos en las fincas con manejo orgánico de la región mediterránea. En el
mandarino convencional, la especie dominante en abundancia y biomasa A. trapezoides empleó
gran parte de su energía en la reproducción, corroborado por la elevada proporción de
individuos juveniles en sus poblaciones (70%). Las restantes especies (O. complanatus, A. rosea
y A. caliginosa) obtuvieron entre un 20 y un 40% menos de peso medio por individuo que las
recogidas en el manejo orgánico. Estos resultados sugieren que, aunque este cultivo mantuvo
unas prácticas agrícolas con un nivel aceptable para el mantenimiento de sus comunidades de
lombrices (Paoletti 1999), cualquier perturbación que afecte a la especie dominante podría
tener repercusiones importantes en la calidad de estos suelos (Ehrlich y Ehrlich 1981; Dunne et
al. 2002). En cambio, en los cultivos de mandarino orgánico, las especies presentes no necesitan
invertir la mayor parte de la energía en la reproducción (supervivencia de la especie) y pueden
emplearla en crecimiento (reflejado en una mayor biomasa de los individuos). Así en esta finca
los individuos alcanzaron mayores tamaños (por ejemplo, el peso medio O. complanatus fue 1,4
g ind.-1 en el orgánico frente a 1 g ind.-1 observado en el convencional). Además, la ausencia de
perturbaciones favorece que haya mayor número de especies que pueden convivir gracias al
aprovechamiento de los diferentes nichos que les proporciona el cultivo (Abrams 1983;
Ekschmitt y Griffiths 1998). En el caso de los cultivos de olivo, la mayor contribución a la biomasa
fue de las lombrices anécicas, con un elevado porcentaje de estas lombrices siendo ejemplares
juveniles (78%). Estas fincas presentaron un elevado valor del estadístico W y por tanto, una
perturbación baja de sus suelos; sin embargo, registraron el menor número de especies e
individuos, lo cual puede ser debido a que la hojarasca de olivo tiene una menor calidad
nutricional para las lombrices, como resultado de su bajo contenido en nitrógeno y a la mayor
cantidad de compuestos fenólicos (Kherbouche et al. 2012).

Capítulo II
117
5. CONCLUSIONES
En ambas regiones biogeográficas la mesofauna fue mayoritaria comparada con la
macrofauna, lo que confirma resultados previos que indican que los taxones de pequeño
tamaño dominan las comunidades edáficas de los suelos agrícolas. La región atlántica con suelos
más húmedos favorece la mayor abundancia de enquitreidos, mientras que en la región
mediterránea los componentes mayoritarios de la mesofauna fueron los ácaros, estando
además ausentes las lombrices epigeas.
Ni la región biogeográfica, ni el manejo tuvieron un efecto significativo sobre las abundancias
totales de fauna edáfica; en cambio, el tipo de cultivo fue un factor muy influyente y los cultivos
de mandarino contuvo las mayores densidades totales de organismos edáficos.
El manejo agrícola no tuvo efecto sobre la mesofauna pero sí alteró significativamente la
diversidad y estructura funcional de la macrofauna, haciendo que las comunidades de los
distintos cultivos frutícolas fueran muy disimilares. En la región atlántica, los manejos
convencionales redujeron drásticamente el número de lombrices endogeas y anécicas, mientras
que en la región mediterránea dio como resultado que las lombrices anécicas fueran la categoría
ecológica dominante. Los cultivos de cítricos destacan presentar la mayor riqueza específica de
lombrices que tierra; sin embargo, son los cultivos que presentan mayores perturbaciones
debidas al manejo convencional, haciendo que la macrofauna dedique más energía a la
reproducción que al crecimiento individual.
6. BIBLIOGRAFÍA
Abrams P. 1983. The theory of limiting similarity. Annual Review of Ecology & Systematics, 14, 359-376. Alvarez T, Frampton G K y Goulson D. 2001. Epigeic collembola in winter wheat under organic, integrated
and conventional farm management regimes. Agriculture, Ecosystems & Environment, 83, 95-110. Andren O y Lagerlöf J. 1983. Soil fauna (Microarthropods, Enchytraeids, Nematodes) in Swedish
agricultural cropping systems. Acta Agriculturae Scandinavica, 33, 33-52. Arnold B E, Hodson M E, Charnock J y Peijnenburg W J G M. 2008. Comparison of subcellular partitioning,
distribution, and internal speciation of cu between cu-tolerant and naive populations of Dendrodrilus rubidus Savigny. Environmental Science & Technology, 42, 3900–3905.
Baker G H, Thumlert T A, Meisel L S, Carter P J y Kilpin G P. 1997. "Earthworms downunder": a survey of the earthworm fauna of urban and agricultural soils in Australia. Soil Biology & Biochemistry, 29(3-4), 589-597.
Bardgett R D y Cook R. 1998. Functional aspects of soil animal diversity in agricultural grasslands. Applied Soil Ecology, 10, 263-276.
Bardgett R D y van der Putten W H. 2014. Belowground biodiversity and ecosystem functioning. Nature, 515(7528), 505-511.
Barrientos J A. 1988. Bases para un Curso Práctico de Entomología. Asociación Española de Entomología. Barcelona.
Bedano J C, Cantú M P y Doucet M E. 2006. Soil springtails (Hexapoda: Collembola), Symphylans and Pauropods (Arthropoda: Myriapoda) under different management systems in agroecosystems of the subhumid Pampa (Argentina). European Journal of Soil Biology, 42(2), 107-119.

Capítulo II
118
Behan-Pelletier V M. 1999. Oribatid mite biodiversity in agrosystems: role for bioindication. Agriculture, Ecosystems & Environment, 74, 411-423.
Berman D I, Meshcheryakova E N, Alfimov A V y Leirikh A N. 2001. Spread of the earthworm Dendrobaena octaedra (Lumbricidae: Oligochaeta) from Europe to northern Asia is restricted by its insufficient frost resistance. Doklady Biological Sciences, 377, 145-148.
Bertrand M, Barot S, Blouin M, Whalen J, de Oliveira T y Roger-Estrade J. 2015. Earthworm services for cropping systems. A Review. Agronomy for Sustainable Development, 35, 553-567.
Bilalis D, Sidiras N, Vavoulidou E y Konstantas A. 2009. Earthworm populations as affected by crop practices on clay loam soil in a mediterranean climate. Acta Agriculturae Scandinavica Section B - Soil & Plant Science, 59, 440-446.
Birkhofer K, Bezemer T M, Bloem J, Bonkowski M, Christensen S, Dubois D, Ekelund F, Fliessbach A, Gunst L, Hedlund K, Mader P, Mikola J, Robin C, Setälä H, Tatin-Froux F, van der Putten W H y Scheu S. 2008. Long-term organic farming fosters below and aboveground biota: implications for soil quality, biological control and productivity. Soil Biology & Biochemistry, 40, 2297-2308.
Blasi S, Menta C, Balducci L, Conti F D, Petrini E y Piovesan G. 2013. Soil microarthropod communities from mediterranean forest ecosystems in central Italy under different disturbances. Environmental Monitoring and Assessment, 185(2), 1637-1655.
Block W. 1996. Cold or drought - the lesser of two evils for terrestrial arthropods? European Journal of Entomology, 93, 325-339.
Boag B, Palmer L F, Neilson R, Legg R y Chambers S J. 1997. Distribution, prevalence and intensity of earthworm populations in arable land and grassland in Scotland. Annals of Applied Biology, 130(1), 153-165.
Bouché M B. 1972. Lombriciens De France, Ecologie et Systematique. Institut National de la Recherche Agronomique, Paris.
Bouché M B. 1977. Strategies Lombriciences. En: Lohm U y Persson T (Eds.) Soil Organisms as Components of Ecosystems, pp. 122-132. Ecological Bulletins, Stockholm.
Briones M J I, Ineson P y Piearce TG. 1997. Effects of climate change on soil fauna: responses of enchytraeids, diptera larvae and tardigrades in a transplant experiment. Applied Soil Ecology, 6, 117–134.
Briones M J I, Ineson P y Heinemeyer A. 2007. Predicting potential impacts of climate change on the geographical distribution of enchytraeids: a meta-analysis approach. Global Change Biology, 13, 2252–2269.
Briones M J I, Ostle N J, McNamara N P y Poskitt J. 2009. Functional shifts of grassland soil communities in response to soil warming. Soil Biology & Biochemistry, 41(2), 315-322.
Briones M J I, Barreal M E, Harrison A C y Gallego P P. 2011. Earthworms and nitrogen applications to improve soil health in an intensively cultivated kiwifruit orchard. Applied Soil Ecology, 49, 158-166.
Calvin E B, Mato S, García R M y Díaz Cosín D J. 1987. Fauna de lombrices de tierra del valle del Tambre (La Coruña). ii. Cultivos, prados y estudio conjunto. Munibe (Ciencias Naturales), 39, 95-102.
Carrera N. 2012. Efecto do incremento de temperatura sobre a estrutura das comunidades e os fluxos de carbono en solos de turbeira. (Tesis Doctoral). Universidad de Vigo, Vigo.
Cao Z, Han X, Hu C, Chen J, Zhang D y Steinberger Y. 2011. Changes in the abundance and structure of a soil mite (Acari) community under long-term organic and chemical fertilizer treatments. Applied Soil Ecology, 49, 131-138.
Clarke K R y Gorley R N. 2006. PRIMER v6: User Manual/Tutorial. PRIMER-E, Plymouth. Clarke K R y Warwick R M. 2001. Change In Marine Communities: an Approach to Statistical Analysis and
Interpretation. National Environment Research Council, Plymouth. Clarke K R, Somerfield P J y Chapman M G. 2006. On resemblance measures for ecological studies,
including taxonomic dissimilarities and a zero-adjusted Bray–Curtis coefficient for denuded assemblages. Journal of Experimental Marine Biology & Ecology, 330(1), 55-80.
Clarke K R. 1990. Comparisons of dominance curves. Journal of Experimental Marine Biology & Ecology, 138, 143-157.
Clarke K R. 1993. Non-parametric multivariate analyses of changes in community structure. Australian Journal of Ecology, 18, 117- 143.
Crumsey J M, Le Moine J M, Vogel C S y Nadelhoffer K J. 2014. Historical patterns of exotic earthworm distributions inform contemporary associations with soil physical and chemical factors across a northern temperate forest. Soil Biology & Biochemistry, 68, 503-514.

Capítulo II
119
Czarnecki A J y Paprocki R. 1997. An attempt to characterize complex properties of agroecosystems based on soil fauna, soil properties and farming system in the north of Poland. Biological Agriculture & Horticulture, 15, 11-23.
Dash M C. 1990. Oligochaeta: Enchytraeidae. En: Dindal D L (Ed.) Soil Biology Guide, pp 311-340. Wiley & Sons, Inc., New York.
De Goede R G M, Brussaard L y Akkermans A D L. 2003. On-farm impact of cattle slurry manure management on biological soil quality. NJAS - Wageningen Journal of Life Sciences, 51, 103-133.
Dinter A, Oberwalder C, Kabouw P, Coulson M, Ernst G, Leicher T, Miles M, Weyman G y Klein O. 2013. Occurrence and distribution of earthworms in agricultural landscapes across Europe with regard to testing for responses to plant protection products. Journal of Soils & Sediments, 13 (2), 278-293.
Dlamini T C y Haynes R J. 2004. Influence of agricultural land use on the size and composition of earthworm communities in northern kwazulu-natal, South Africa. Applied Soil Ecology, 27, 77-88.
Doles J L, Zimmerman R J y Moore J C. 2001. Soil microarthropod community structure and dynamics in organic and conventionally managed apple orchards in western Colorado, USA. Applied Soil Ecology, 18(1), 83-96.
Dunne J A, Williams R J y Martínez N D. 2002. Network structure and biodiversity loss in food webs: robustness increases with connectance. Ecology Letters, 5, 558-567.
Eggleton P, Inward K, Smith J, Jones D T y Sherlock E. 2009. A six year study of earthworm (Lumbricidae) populations in pasture woodland in southern england shows their responses to soil temperature and soil moisture. Soil Biology & Biochemistry, 41(9), 1857-1865.
Ehrlich P R y Ehrlich A H. 1981. Extinction: The Causes and Consequences of the Disappearance of Species. Random. New York.
Ekschmitt K y Griffiths B S. 1998. Soil biodiversity and its implications for ecosystem functioning in a heterogeneous and variable environment. Applied Soil Ecology, 10, 201-215.
Estrada Pareja M M. 2005. Manejo y procesamiento de la gallinaza. Revista Lasallista de Investigación, 2(1), 43-48.
Evans A C y Guild W J M. 1948. Studies on the relationships between earthworms and soil fertilty. V. Field populations. Annals of Applied Biology, 35(4), 485-493.
Feldmanstern C. 2013. Sustaining biodiversity in peach orchards: baseline differences between conventional, low input and organic production systems. Trabajo Fin de Máster. Norwegian University of Life Sciences.
Fernández R M. 2012. Aporrectodea trapezoides (Dugès, 1828) (Oligochaeta, Lumbricidae): Filogeografía, Filogenia Y Biología Reproductiva (Tesis Doctoral). Universidad Complutense de Madrid. Madrid.
Filser J, Fromm H, Nagel R F y Winter K. 1995. Effects of previous intensive agricultural management on microorganisms and the biodiversity of soil fauna. Plant & Soil, 170(1), 123-129.
Fragoso C, Lavelle P, Blanchart E, Senapati B K, Jimenez J J, Martínez M A, Decaëns T y Tondoh J. 1999. Earthworm communities of tropical agroecosystems: origin, structure and influence of management practices. En: Lavelle P, Brussard L y Hendrix P (Eds.) Earthworm Management in Tropical Agroecosystems, pp 27-55. CABI, Wallingford.
Gagnarli E, Goggioli D, Tarchi F, Guidi S, Nannelli R, Vignozzi N, Valboa G, Lottero M R, Corino L y Simoni S. 2015. Case study of microarthropod communities to assess soil quality in different managed vineyards. Soil, 1(2), 527-536.
Gardi C, Montanarella L, Arrouays D, Bispo A, Lemanceau P, Jolivet C, Mulder C, Ranjard L, Römbke J, Rutgers M., Menta, C., 2009. Soil biodiversity monitoring in Europe: ongoing activities and challenges. European Journal of Soil Science, 60 (5), 807-819.
Gerard B M. 1967. Factors affecting earthworms in pastures. Journal of Animal Ecology, 36(1), 235-252. Giller P S. 1996. The di e sit of soil o u ities, the `poo a s t opi al ai fo est´. Biodiversity &
Conservation, 5(2), 135-168. Gonçalves M F y Pereira J A. 2012. Abundance and diversity of soil arthropods in the olive grove
ecosystem. Journal of Insect Science, 12, 20. Henneron L, Bernard L, Hedde M, Pelosi C, Villenave C, Chenu C, Bertrand M, Girardin C y Blanchart E.
2015. Fourteen years of evidence for positive effects of conservation agriculture and organic farming on soil life. Agronomy for Sustainable Development, 35, 169-181.
Hole D G, Perkins A J, Wilson J D, Alexander I H, Grice P V y Evans A D. 2005. Does organic farming benefit biodiversity? Biological Conservation, 122(1), 113-130.

Capítulo II
120
Hopkin S P. 2000. A Key to the Springtails of Britain and Ireland, AIDGAP (Aids to the Identification in Difficult Groups of Animals and Plants): Test Version. Shrewsbury: Field Studies Council.
Jones H D, Santoro G, Boag B y Neilson R. 2001. The diversity of earthworms in 200 Scottish fields and the posible effect of New Zealand land flatworms (Arthurdendyus Triangulates) on earthworms populations. Annals of Applied Biology, 139, 75-92.
Karnatak A K, Thakur S S y Shukla A. 2007. Impact of commonly used pesticides on the population of beneficial soil inhabitants (Enchytraeids) and yield in rice crop ecosystem. Pantnagar Journal of Research, 5(1), 27-29.
Kasprzak K. 1982. Review of enchytraeid (Oligochaeta, Enchytraeidae) community structure and function in agricultural ecosystems. Pedobiologia, 23, 217-232.
Khalil M A, Abdel-Lateif HM y Al-Assiuty AI. 1999. Changes in oribatid faunal structure associated with land conversion from annual crop into orchard. Pedobiologia, 43, 85-96.
Kherbouche D, Bernhard-Reversat F, Moali A y Lavelle P. 2012. The effect of crops and farming practices on earthworm communities in soummam valley, Algeria. European Journal of Soil Biology, 48, 17-23.
Lagerlöf J, Andrén O y Paustian K. 1989. Dynamics and contribution to carbon flows of enchytraeidae (oligochaeta) under four cropping systems. Journal of Applied Ecology, 26, 183–199.
Lagerlöf J, GoffreB y Vincent C. 2002. The importance of field boundaries for earthworms (lumbricidae) in the Swedish agricultural landscape. Agriculture, Ecosystems & Environment, 89(1), 91-103.
Lago C F, 2015. Efecto del manejo agrícola sobre las propiedades biológicas y químicas del suelo y los procesos de transformación de la materia orgánica en cultivos de kiwi (Actinidia deliciosa) (Tesis Doctoral). Universidad de Vigo, Vigo.
Lambshead P J D, Platt H M y Shaw K M. 1983. The detection of differences among assemblages of marine benthic species based on an assessment of dominance and diversity. Journal of Natural History, 17, 859-874.
Lavelle P, Bignell D, Lepage M, Wolters V, Roger P, Ineson P, Heal O W, Dhillion S. 1997. Soil function in a changing world: the role of invertebrate ecosystem engineers. European Journal of Soil Biology, 33(4), 159-193.
Lee K E, 1985. Earthworms: their Ecology and Relationships with Soils and Land Use. Academic Press, New York.
Leroy B L M, Schmidt O, van den Bossche A, Reheul D y Moens M. 2008. Earthworm population dynamics as influenced by the quality of exogenous organic matter. Pedobiologia, 52, 139-150.
Lindberg N y Bengtsson J. 2006. Recovery of forest soil fauna diversity and composition after repeated summer droughts. Oikos, 3, 494-506.
Maddison D R, Schulz K S y Maddison W P. 2007. The Tree of Life Web Project. http://tolweb.org. Mazzoncini M, Canalis S, Giovannetti M, Castagnoli M, Tittarelli F, Antichi D, Nannelli R, Cristani C y Bàrberi
P. 2010. Comparison of organic and conventional stockless arable systems: a multidisciplinary approach to soil quality evaluation. Applied Soil Ecology, 44, 124-132.
Menta C, García-Montero L G, Conti F D y Pinto S. 2013. The role and diversity of soil fauna in different woodlands. En: Manzanares L. (Ed.) Woodlands Structure, Species Diversity and Sustainable Management, pp. 43-74. Nova Publishers, New York.
Mitchell M J, Hartenstein R, Swift B L, Neuhauser E F, Abrams B I, Mulligan R M, Brown B A, Craig D y Kaplan D. 1978. Effects of different sewage sludges on some chemical and biological characteristics of soil. Journal of Environmental Quality, 7, 551-559.
Nannelli R y Simoni S. 2002. The influence of different types of grass mulching on the communities of oribatid mites in the vineyards. En: Bernini F, Nannelli R, Nuzzaci G y de Lillo E (Eds.) Acarid Phylogeny and Evolution: Adaptation in Mites and Ticks. Proceedings of the IV Symposium of the European Association of Acarologists, pp 363-371. Kluwer Academic Publishers.
Norton R A. 1990. Acarina: Oribatida. En: Dindal D L (Ed.) Soil Biology Guide, pp 779-803. Wiley, New York. Ochoa-Hueso R, Rocha I, Stevens C J, Manrique E y Luciañez M J. 2014. Simulated nitrogen deposition
affects soil fauna from a semiarid mediterranean ecosystem in central spain. Biology & Fertility of Soils, 50, 191-196.
O'Connor F B. 1955. Extraction of enchytraeid worms from coniferous forest soil. Nature, 175, 815-816. O'Connor F B. 1957. An ecological study of the enchytraeids worms from a coniferous forest soil. Oikos,
8, 161 -169. Palm J, van Schaik L M B y Schröder B. 2013. Modelling distribution patterns of anecic, epigeic and
endogeic earthworms at catchment-scale in agro-ecosystems. Pedobiologia, 56(1), 23-31.

Capítulo II
121
Paoletti M G, Sommaggio D y Fusaro S. 2013. Proposta di indice di qualità biologica del suolo (QBS-e) basato sui lombrichi e applicato agli agroecosistemi. Biología Ambientale, 27(2), 25-43.
Paoletti M G, Sommaggio D, Favretto M R, Petruzzelli G, Pezzarossa B y Barbafieri M. 1998. Earthworms as useful bioindicators of agroecosystem sustainability in orchards and vineyards with different inputs. Applied Soil Ecology, 10, 137- 150.
Paoletti M G. 1999. The role of earthworms for assessment of sustainability and as bioindicators. Agriculture, Ecosystems & Environment, 74, 137-155.
Parisi V y Menta C. 2008. Microarthropods of the soil: convergence phenomena and evaluation of soil quality using QBS-ar and QBS-C. Fresenius Environmental Bulletin, 17(8), 1170-1174.
Parisi V, Menta C, Gardi C, Jacomini C y Mozzanica E. 2005. Microarthropod communities as a tool to assess soil quality and biodiversity: a new approach in Italy. Agriculture, Ecosystems & Environment, 105(1-2), 323-333.
Payne R J, Thompson A M, Standen V, Field C D y Caporn S J M. 2012. Impact of simulated nitrogen pollution on heathland microfauna, mesofauna and plants. European Journal of Soil Biology, 49, 73-79.
Pelosi C, Bertrand M y Roger-Estrade J. 2009. Earthworm community in conventional, organic and direct seeding with living mulch cropping systems. Agronomy for Sustainable Development, 29(2), 287-295.
Pelosi C y Römbke J. 2016. Are enchytraeidae (Oligochaeta, Annelida) good indicators of agricultural management practices? Soil Biology & Biochemistry, 100, 255-263.
Perel T S. 1979. Range and Distribution of Earthworms of the USSR Fauna. Nauka Publishing House. Moscow.
Pérès G, Piron D, Bellido A, Goater C, Cluzeau D. 2008. Earthworms used as indicators of agricultural managements. Fresenius Environmental Bulletin, 17, 1181-1189.
Pérez-Iñigo C. 1993. Acari. Oribatei, Poronota. En: Ramos M A (Eds.) Fauna Ibérica, vol. 3. Museo Nacional de Ciencias Naturales, CSIC. Madrid.
Pielou E C. 1966. The measurement of diversity in different types of biological collections. Journal of Theoretical Biology, 13, 131-144.
Postma-Blaauw M B, de Goede R G M, Bloem J, Faber J H y Brussaard L. 2010. Soil biota community structure and abundance under agricultural intensification and extensification. Ecology, 91(2), 460-473.
Pulleman M, Creamer R, Hmaer U, Herlder J, Pelosi C, Peres G y Rutgers M. 2012. Soil biodiversity, biological indicators and soil ecosystems services – an overview of European approaches. Current Opinion in Environmental Sustainability, 4, 529-538.
Regier J C, Shultz J W, Zwick A, Hussey A, Ball B, Wetzer R, Martin J W y Cunningham C W. 2010. Arthropod relationships revealed by phylogenomic analysis of nuclear protein-coding sequences. Nature, 463, 1079-1083.
Rodríguez T, Trigo D y Cosín D J D. 1997. Biogeographical zonation of the western Iberian Peninsula on the basis of the distribution of earthworm species. Journal of Biogeography, 24(6), 893-901.
Salminen J, Setälä H y Haimi J. 1997. Regulation of decomposer community structure and decomposition processes in herbicide stressed humus soil. Applied Soil Ecology, 6(3), 265-274.
Schmelz R M y Collado R. 2010. A guide to European terrestrial and freshwater species of Enchytraeidae (Oligochaeta). Senckenberg Museum für Naturkunde Görlitz.
Schreck E, Gontier L, Dumat C y Geret F. 2012. Ecological and physiological effects of soil management practices on earthworm communities in french vineyards. European Journal of Soil Biology, 52, 8-15.
Shannon C E y Wiener W. 1949. The Mathematical Theory of Communication. University of Illinois Press, Urbana.
Solomou A D, Sfougaris A I, Vavoulidou E M y Csuzdi C. 2012. Advances of the 5th International Oligochaeta Taxonomy Meeting. Zoology in the Middle East, Supplementum, 4, 119-126.
Sousa J P, da Gama M M, Pinto C, Keating A, Calhôa F, Lemos¡ M, Castro C, Luz T, Leitão P y Dias, S. 2004. Effects of land-use on collembola diversity patterns in a mediterranean landscape. Pedobiologia, 48(5), 609-622.
Springett J A, Gray R A J, Reid J B y Petrie R. 1994. Deterioration in soil biological and physical properties associated with kiwifruit (Actinidia deliciosa). Applied Soil Ecology, 1, 231-241.
StatSoft, Inc. 2005. STATISTICA, Version 7.1. StatSoft, Inc. Tulsa.

Capítulo II
122
Tiunov A V, Hale C M, Holdsworth R y Vsevolodova-Perel T S. 2006. Invasion patterns of lumbricidae into the previously earthworm-free areas of northeastern Europe and the western great lakes region of North America. Biological Invasions, 8, 1223-1234.
Trigo D, Mascato R, Mato S y Cosín D J D. 1988. Biogeographical divisions of continental Portugal as regards earthworm fauna. Bolletino di Zoologia, 55(1-4), 85-92.
Tullgren A. 1918. Ein Sehr Einfacher Ausleseapparat Für Tierricole Tier-Formen. Zeitschrift für Angewandte Entomologie, 4, 149-150.
Uvarov A V. 1998. Respiration activity of Dendrobaena Octaedra (Lumbricidae) under constant and diurnally fluctuating temperature regimes in laboratory microcosms. European Journal of Soil Biology, 34, 1-10.
van Eekeren N, Bommelé L, Bloem J, Schouten T, Rutgers M, de Goede R, Reheul D y Brussaard L. 2008. Soil biological quality after 36 years of ley-arable cropping, permanent grassland and permanent arable cropping. Applied Soil Ecology, 40(3), 432-446.
Vavoulidou E, Coors A, Dózsa-Farkas K y Römbke J. 2009. Influence of farming practice, crop type and soil properties on the abundance of enchytraeidae (oligochaeta) in Greek agricultural soils. Soil Organisms, 81(2), 197-212.
Verhoef H A y van Selm A J. 1983. Distribution and population dynamics of collembola in relation to soil moisure. Holartic Ecology, 6, 387-394.
Wardle D A. 1995. Impacts of disturbance on detritus food webs in agro-ecosystems of contrasting tillage and weed management practices. Advances in Ecological Research, 26, 105-185.
Wardle D A, Yeates G W, Bonner K I, Nicholson K S y Watson R N. 2001. Impacts of ground vegetation management strategies in a kiwifruit orchard on the composition and functioning of the soil biota. Soil Biology & Biochemistry, 33(7), 893-905.
Wheeler W C, Whiting M, Wheeler Q D y Carpenter J M. 2001. The phylogeny of the extant hexapod orders. Cladistics, 17, 113-169.
Yan S, Singh A N, Fu S, Liao C, Wang S, Li Y, Cui Y y Hu L. 2012. A soil fauna index for assessing soil quality. Soil Biology & Biochemistry, 47, 158-165.
Yeates G W, Bardgett R D, Cook R, Hobbs P J, Bowling P J y Potter J F. 1997. Faunal and microbial diversity in three welsh grassland soils under conventional and organic management regimes. Journal of Applied Ecology, 34, 453-470.
Zaller J G y Köpke U. 2004. Effects of traditional and biodynamic farmyard manure amendment on yields, soil chemical, biochemical and biological properties in a long-term field experiment. Biology & Fertility of Soils, 40(4), 222-229.

CAPÍTULO III:
Relaciones ecológicas entre comunidades edáficas y factores ambientales en los agrosistemas atlánticos y
mediterráneos


Capítulo III
125
1. INTRODUCCIÓN
De la i teg a ió de los o eptos de e ología ag i ultu a a e el té i o agroecosistema
(Harper 1974), el cual se basa en considerar la producción agrícola entendida como un
ecosistema (Gliessman 1998). Este concepto proporciona un marco de trabajo para estudiar los
sistemas de producción agrícola teniendo en cuenta los flujos de nutrientes y energía así como
las interrelaciones de las comunidades bióticas y el medio. El origen antrópico de los
agroecosistemas hace que sean diferentes a los ecosistemas naturales (Tabla 24), sin embargo,
ambos están regulados por los mismos procesos (Gliessman 1998; Magdoff 2007).
Tabla 24: Principales características de los ecosistemas naturales y agroecosistemas (Gliessman 1998; Magdoff 2007).
Ecosistemas naturales Agroecosistemas
Flujo de energía Eficientes y autosuficientes Alterado antrópicamente, pocas
veces autosuficientes
Ciclo de nutrientes Eficiente
Reducido en comparación con los ecosistemas naturales y con
pérdidas de nutrientes debido a la recolección de las cosechas
Diversidad y estructura de comunidades
Diversidad elevada y comunidades complejas
Diversidad reducida y comunidades simplificadas
Resilencia Mayor resistencia a alteraciones y
capacidad de recuperación
Mayor sensibilidad a alteraciones debido a una menor diversidad
biológica
Por tanto, al igual que los ecosistemas naturales, los agroecosistemas se componen de
diversos componentes funcionales interconectados: componentes físicos (la energía solar, el
suelo, la atmósfera y la lluvia), componentes biológicos (cultivo de especies vegetales,
organismos descomponedores, animales y sus desechos) y factores humanos y sociales como
las entradas de enmiendas orgánicas (fertilizantes), el consumo de productos y el mercado
(Gliessman 2006). Entre todos ellos, destaca el nivel edáfico por sus repercusiones en el sistema
productivo, ya que el suelo proporciona el soporte físico y los nutrientes para las especies
cultivadas. A este nivel, el tipo de vegetación, la estructura del suelo, el ambiente físico
(especialmente la humedad y la temperatura) y químico (cantidad de materia orgánica, pH) del
suelo y las poblaciones microbianas han sido identificados como los factores con mayor
influencia sobre los organismos edáficos (El Titi y Ulber 1991; Beare et al. 1992; Wood y James
1993; Lavelle 1996; Lindberg et al. 2002; Wardle et al. 2004; Osler y Somerkon 2007).
La teo ía de iodi e sidad a idada p opuesta po Lavelle (1996) se basa en que la
diversidad de las comunidades de plantas determina la diversidad de las comunidades de
organismos edáficos que viven debajo. En el caso de los sistemas agrícolas, son numerosos los
trabajos que establecen una relación entre plantas cultivadas y fauna edáfica y así por ejemplo,

Capítulo III
126
Paoletti (1998) observó importantes diferencias en la abundancia y biomasa de lombrices al
comparar cultivos de manzana, vid, kiwi y melocotón, destacando los reducidos valores en los
viñedos frente a los demás cultivos. Los resultados de esta memoria (capítulo II) también
parecen apoyar la idea de que existen diferencias significativas entre las comunidades edáficas
de los suelos con diferentes especies cultivadas. Así, al igual que en el caso del estudio anterior,
el mayor contenido de compuestos recalcitrantes de la hojarasca de olivares y viñedos (Tian et
al. 1993) hace que sean menos palatables para los organismos edáficos (Kherbocuhe et al. 2012).
Por otro lado, se ha observado que alteraciones en la estructura del suelo también pueden
repercutir sobre el tamaño de las comunidades edáficas (Wardle 1995). En el caso de los suelos
agrícolas, prácticas como el arado con maquinaria pesada incrementa la compactación y reduce
la porosidad del suelo. En este sentido, Whalley et al. (1995) señalaron que la compactación del
suelo limita considerablemente las actividades de las lombrices y Lee y Foster (1991) que la
reducción en la porosidad del suelo dificulta los movimientos de la mesofauna, organismos que
sólo pueden mover pequeñas partículas de suelo.
Respecto al ambiente físico y a escala global, la importancia del clima como modulador de la
fauna edáfica varía considerablemente entre diferentes biomas y además, sus efectos pueden
ser directos o indirectos (García-Palacios et al. 2013). Por ejemplo, en el caso de las lombrices
de tierra (macrofauna), el efecto sobre los procesos edáficos viene condicionado por su
estructura funcional, la cual varía a lo largo del gradiente latitudinal y entre tipos de ecosistema
presentes en un mismo rango de latitud. Así, en climas fríos la mayor parte de la biomasa
corresponde a lombrices epigeas, mientras que en climas tropicales dominan las endogeas
(Lavelle 1983). Esto es debido a que las distintas especies han desarrollado adaptaciones morfo-
funcionales (como la secreción de moco para mantener la superficie corporal húmeda y la
retención de agua corporal; Bouché 1984) y etológicas (como la quiescencia y la diapausa) que
les permiten sobrevivir en los periodos con condiciones climáticas más adversas (Lavelle 1983;
Lee 1985). Sin embargo, otro grupo de oligoquetos, los enquitreidos (mesofauna), al carecer de
estructuras de protección contra la sequedad, sus comunidades son especialmente sensibles a
los cambios en la humedad y temperatura del suelo (Briones et al. 2007).
Entre la mesofauna, los colémbolos presentan una amplia distribución estando presentes en
ambientes húmedos y secos, en climas fríos como los de la región ártica y sistemas alpinos pero
también en zonas tropicales (Rusek 1998). Sin embargo, su biomasa en estos ecosistemas varía
enormente, desde 10-20 mg de peso seco por m-2 en pastos y bosques tropicales hasta 150 mg
de peso seco por m-2 en ecosistemas de tundra (Petersen 1982), lo que es un claro reflejo de su
gran sensibilidad a las condiciones ambientales. Al igual que los colémbolos, los ácaros también
están presentes en un gran número de biomas y también siguen un gradiente latitudinal, de tal

Capítulo III
127
manera que su diversidad disminuye al alejarnos del ecuador (Maraun et al. 2007). Parte de su
éxito se debe a que presentan distintas estrategias alimenticias (herbívoros, fungívoros,
detritívoros, parásitos, depredadores, omnívoros), lo que les permite explotar diferentes nichos
ecológicos (Illig et al. 2005; Maraun et al. 2007).
En los suelos agrícolas la mesofauna y la macrofauna dominan en términos de biomasa y
juegan un papel importante en el funcionamiento de estos ecosistemas (Lago 2015), por lo que
cualquier alteración en sus abundancias, biomasa y en la estructura de sus comunidades tiene
importantes efectos sobre los procesos edáficos. La abundancia de los enquitreidos en suelos
agrícolas puede llegar a alcanzar valores muy elevados (de unos pocos centenares a más de
30.000 ind. m-2; Coleman et al. 2004) y en suelos con acidez elevada son los principales
responsables de la descomposición de la materia orgánica (Briones et al. 2007). Los enquitreidos
son organismos principalmente saprófagos y su actividad excavadora aumenta la porosidad del
suelo, mejorando la aireación y la infiltración de agua (Didden 1990; van Vliet et al. 1993; Jänsch
et al. 2005; Carrera et al. 2009). Además, se ha observado en ciertos cultivos pueden llegar a
ejercer un control sobre las poblaciones de nematodos que infestan las raíces de las plantas
(Jegen 1920; O´Connor 1967), lo cual puede resultar un aspecto muy beneficioso en la salud de
los cultivos.
Otros grupos de mesofauna que son también relevantes para el funcionamiento de los suelos
agrícolas son los ácaros y los colémbolos (Menta 2012), pero debido a su elevada diversidad,
tanto morfológica como trófica, hace muy difícil determinar qué factores abióticos son los que
determinan sus respuestas a los diferentes tipos de gestión agrícola. Por ejemplo, el uso de
fertilizantes minerales parecen tener efectos negativos sobre los microartrópodos, reduciendo
sus abundancias (Tabaglio et al. 2009). En el caso de los colémbolos, su dieta ha sido descrita
como generalista (Johnson et al. 2005), aunque otros autores (Sechi et al. 2014) indican que las
distintas especies pueden presentar diferentes estrategias alimenticias en función de la
palatabilidad y disponibilidad de los recursos alimenticios. Debido a su reducida biomasa, la
contribución directa de los colémbolos a los ciclos del C y al N es reducida (Petersen y Luxton
1982), sin embargo su importancia para la movilización de nutrientes aumenta con la relación
C/N y cuando desciende la disponibilidad de agua (Verhoef y Brussaard 1990). Por ejemplo, Song
et al. (2016) observaron una rápida reducción en la abundancia y diversidad de las comunidades
de colémbolos en humedales al incrementar las entradas de N en el suelo (causado por el uso
de fertilizantes en los cultivos circundantes a la zona de estudio). Por otro lado, en la revisión
realizada por Filser (2002) se destaca, además de la disponibilidad de agua, el pH del suelo como
un factor limitante para las actividades de estos microartrópodos. Así, Ochoa-Hueso et al. (2014)
observaron que aportes de N del orden de 50 kg de N ha-1 año-1 provocan importantes

Capítulo III
128
reducciones de sus poblaciones, lo que fue atribuido a un incremento de la acidificación por
adiciones excesivas de fertilizantes nitrogenados.
La reducida capacidad de dispersión de los oribátidos en comparación con los colémbolos
compromete de forma importante su recuperación ante alteraciones del ecosistema (Lindberg
y Bengtsson 2005). Por otro lado, las propiedades químicas de los suelos también tienen un
efecto directo sobre el tamaño de sus poblaciones y así, Ochoa-Hueso et al. (2014) observaron
incrementos en las densidades de ácaros con el pH. De forma similar, Vreeken-Bujis et al. (1998)
constataron que, para el caso de los ácaros oribátidos, la cantidad y calidad de la materia
orgánica presente en los suelos juega un papel fundamental en el tamaño de sus poblaciones.
En este sentido, se ha observado efectos negativos a largo plazo de la fertilización, tanto
orgánica como mineral, sobre las poblaciones de oribátidos, principalmente debido al alto
contenido de fósforo de estas formulaciones (Cao et al. 2011). Sin embargo, Osler y Murphy
(2005) señalan que la abundancia de oribátidos en suelos agrícolas suele estar más condicionada
por alteraciones en la estructura física del suelo debidas al manejo que por el contenido de C y
N de la materia orgánica.
Dentro de la macrofauna, las lombrices de tierra tienen un papel destacado en los sistemas
agrícolas por su mayor biomasa y su papel fundamental en el mantenimiento de la estructura
del suelo (Zanella et al. 2001). Además, gracias a sus interacciones con las poblaciones
microbianas, favorecen la humificación de la materia orgánica (Edwards y Bohlen 1996; Lavelle
y Spain 2001). Entre los factores abióticos que afectan a sus poblaciones, se ha observado que
el contenido de C (Edwards 1983), N (Satchell 1967; Fragoso y Lavelle 1992; Curry 2004) y la
humedad (Lowe y Butt 2005) tienden a favorecer aumentos en las densidades y biomasa de
lombrices. En este sentido, Burtelow et al. (1998) concluyeron que una mayor cantidad de la
fracción mineralizable de materia orgánica (C y N de la biomasa microbiana) incrementa las
abundancias de lombrices. Por tanto, la calidad de la materia orgánica que entra en los
ecosistemas también es de vital importancia para sus comunidades y así, Kherbouche et al.
(2012) observaron que aportes orgánicos de baja calidad (pobres en N y con concentraciones
elevadas de compuestos fenólicos) reducen sus abundancias. El pH del suelo es otro factor
limitante para sus comunidades (Syers y Springett 1984; Edwards y Bohlen 1996) y algunas
especies disminuyen significativamente sus abundancias cuando el pH del suelo está fuera del
rango de 5,0-6,0 (Edwards y Lofty 1975; Chan y Mead 2003).
No existen muchos estudios que hayan abordado los efectos directos de los microorganismos
sobre las comunidades edáficas; sin embargo, se sabe que pueden condicionar la presencia y la
abundancia de estos invertebrados. Esta relación es especialmente evidente en el caso de
aquellos taxones cuya dieta es microbívora, puesto que sus abundancias pueden verse reducidas

Capítulo III
129
al disminuir la actividad microbiana (Zvereva y Kozlov 2010). La mayoría de los organismos que
viven en el suelo son detritívoros y dependen en cierta medida de las comunidades
microbiológicas, puesto que se alimentan de materia orgánica muerta una vez que ésta ha sido
acondicionada previamente por los microorganismos (Moore et al. 1988).
El objetivo principal de este estudio es identificar qué factores ambientales (climáticos y/o
edáficos) tienen una mayor influencia sobre la estructura de las comunidades edáficas que viven
en los suelos de cultivo frutícolas de las regiones atlántica y mediterránea investigados. Las
regiones biogeográficas peninsulares difieren en sus patrones climáticos y tipos de suelo y
determinan los patrones de distribución de los organismos a gran escala. Sin embargo, a nivel
local, las características edafoclimáticas de los hábitats condicionan la adaptación y la resiliencia
de las poblaciones a las perturbaciones ambientales y por tanto, la composición y estructura de
las comunidades edáficas será el resultado de la interacción entre los factores climáticos,
geológicos e históricos y sus capacidades de respuesta ante los cambios continuos en los
factores abióticos a los que se ven sometidas debido al manejo del cultivo.
2. MATERIALES Y MÉTODOS
Para este estudio se emplearon las matrices de datos de los parámetros físico-quimicos de
los distintos suelos de cultivo investigados (capítulo I) y las matrices de composición de la fauna
edáfica recolectada en cada uno de ellos (capítulo II).
Previamente a los análisis multivariantes se realizaron análisis previos de correlacion para
detectar si existía autocorrelacion entre las variables ambientales. Dado que relación C/N se
obtiene tras un cálculo directo a partir del C y el N, se eliminó de los análisis.
Con el fin de conseguir la normalidad de los datos se utilizaron distintas transformaciones: el
N fue normalizado logarítmicamente mediante el cálculo del log10(x) y el carbono de la biomasa
microbiana mediante log10(X+1).
Se emplearon análisis de correspondencias canónicas (ACC; Ter Braak 1986) que permiten
relacionar directamente una matriz de presencia/abundancia de especies recogidas en varias
localizaciones y una matriz de datos ambientales, parámetros físico-quimicos, etc. de las
localizaciones donde se recogieron las especies. Los CCA son, por tanto, análisis de gradiente
di e to , dónde el gradiente de las variables ambientales permite ordenar la respuesta de las
especies a dichos patrones ambientales. Esto o t asta o los a álisis de g adie te i di e to
donde primero se ordenan las especies en función de las muestras y la ordenación resultante se
intenta correlacionar con los factores ambientales (ordenación en dos pasos). El análisis CCA
extrae los ejes, que son combinaciones lineales de las variables ambientales, en orden

Capítulo III
130
decreciente del porcentaje de varianza absorbida. La significación de los ejes fue obtenida
mediante un test de Montecarlo con 9999 permutaciones y la traza de Pillai (Pillai 1955).
La representación gráfica de los ACC muestra los taxones faunísticos como puntos y las
variables ambientales como vectores que representan gradientes. La longitud de los vectores es
un reflejo de su importancia en la ordenación resultante (a mayor longitud, mayor importancia).
Los vectores pueden extenderse en ambos extremos y la mayor o menor proximidad de los
grupos de fauna a un determinado vector ambiental permite establecer la mayor o menor
influencia de esa variable sobre en las abundancias de un determinado taxón. Como
consecuencia, la gran media coincide con el origen de coordenadas del gráfico bidimensional.
Para estos análisis se emplearon las matrices de datos de abundancias totales de todos los
organismos recolectados (macrofauna y mesofauna) en las dos regiones investigadas y como
variables ambientales el contenido de C y N, el pH del suelo, los valores de temperatura y
humedad del suelo y el C de la biomasa microbiana. En primer lugar, se analizó el conjunto de
todos los datos incluyendo todos los grandes grupos de mesofauna (ácaros, colémbolos y
enquitreidos) y la abundancia total de lombrices (macrofauna) recolectados en las dos regiones.
Posteriormente se analizaron la mesofauna (a nivel de orden y con una presencia de al menos
el 20% en el total de muestras analizadas) y la macrofauna (a nivel de categorías ecológicas) por
separado en ambas regiones y en cada una de ellas.
Todos estos análisis se llevaron a cabo con el software PAST (PAleontological STatistica ver
3.11; Hammer et al. 2001), un software libre para análisis de datos con funciones para
estadísticos univariantes y multivariantes. La implementación en PAST para realizar un CCA sigue
el algoritmo de autoanálisis propuesto por Legendre y Legendre (1998) y se empleó el
escalamiento tipo 2 centrado en las especies en lugar de en las muestras (escalamiento tipo 1).
3. RESULTADOS
3.1. Efectos de los parámetros edafoquímicos sobre los principales grupos de mesofauna y macrofauna en las dos regiones investigadas
El análisis de correspondencias canónicas (ACC) realizado para las abundancias totales de
lombrices (macrofauna) y de enquitreidos, ácaros y colémbolos (mesofauna) resultó ser
significativo (traza de Pillai=0,04; p=0,0001). Los dos primeros ejes de ordenación explicaron un
98% de la varianza total, siendo ambos significativos (Tabla 25).
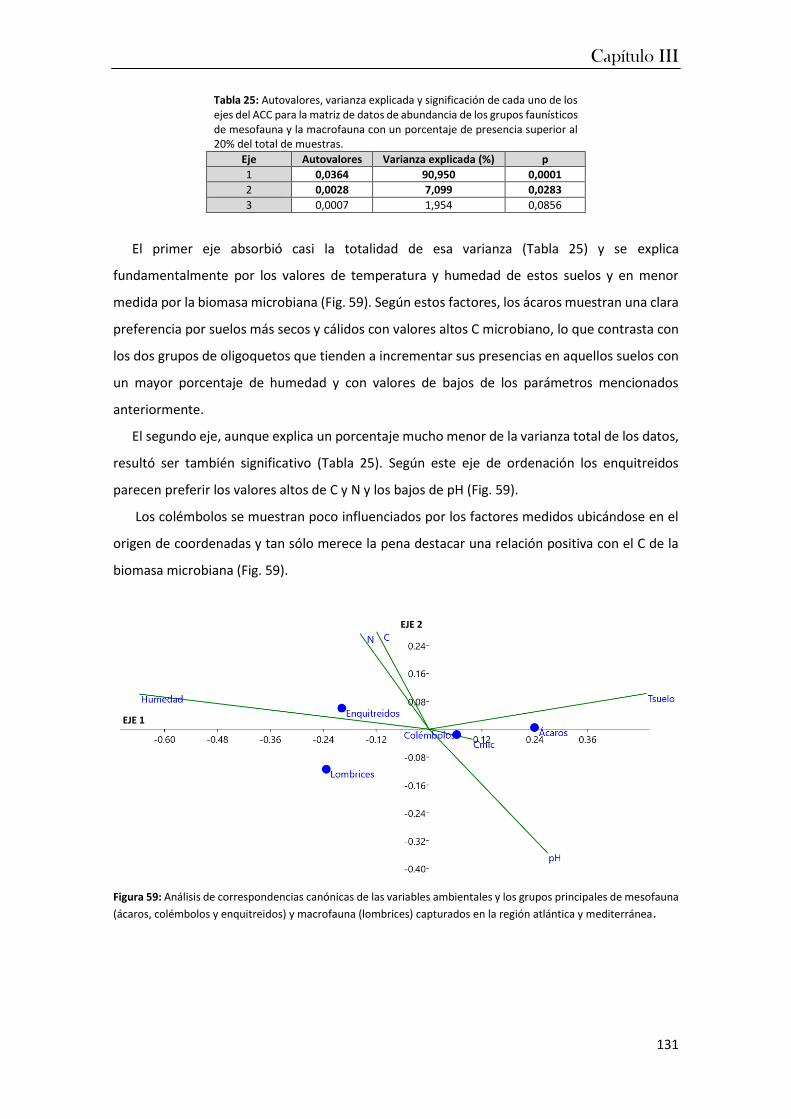
Capítulo III
131
Tabla 25: Autovalores, varianza explicada y significación de cada uno de los ejes del ACC para la matriz de datos de abundancia de los grupos faunísticos de mesofauna y la macrofauna con un porcentaje de presencia superior al 20% del total de muestras.
Eje Autovalores Varianza explicada (%) p
1 0,0364 90,950 0,0001
2 0,0028 7,099 0,0283
3 0,0007 1,954 0,0856
El primer eje absorbió casi la totalidad de esa varianza (Tabla 25) y se explica
fundamentalmente por los valores de temperatura y humedad de estos suelos y en menor
medida por la biomasa microbiana (Fig. 59). Según estos factores, los ácaros muestran una clara
preferencia por suelos más secos y cálidos con valores altos C microbiano, lo que contrasta con
los dos grupos de oligoquetos que tienden a incrementar sus presencias en aquellos suelos con
un mayor porcentaje de humedad y con valores de bajos de los parámetros mencionados
anteriormente.
El segundo eje, aunque explica un porcentaje mucho menor de la varianza total de los datos,
resultó ser también significativo (Tabla 25). Según este eje de ordenación los enquitreidos
parecen preferir los valores altos de C y N y los bajos de pH (Fig. 59).
Los colémbolos se muestran poco influenciados por los factores medidos ubicándose en el
origen de coordenadas y tan sólo merece la pena destacar una relación positiva con el C de la
biomasa microbiana (Fig. 59).
Figura 59: Análisis de correspondencias canónicas de las variables ambientales y los grupos principales de mesofauna
(ácaros, colémbolos y enquitreidos) y macrofauna (lombrices) capturados en la región atlántica y mediterránea.
EJE 1
EJE 2

Capítulo III
132
3.2. Efectos de los parámetros edafoquímicos sobre la estructura de las comunidades de mesofauna en las dos regiones investigadas
El análisis de correspondencias canónicas para las abundancias de los principales grupos de
mesofauna capturados en ambas regiones dio como resultado una ordenación multivariante
significativa (traza de Pillai=0,067; p=0,047). Aunque los dos primeros ejes explicaron casi un
86% de la varianza total de los datos (Tabla 26), solamente el primero de ellos resultó ser
significativo (Tabla 26).
Tabla 26: Autovalores, varianza explicada y significación de cada uno de los ejes del ACC para la matriz de datos de abundancia de los grupos faunísticos de mesofauna (con un porcentaje de presencia superior al 20% del total de muestras): ácaros (órdenes Oribatida, Mesostigmata y Prostigmata), colémbolos (subórdenes Poduromorpha, Entomobryomorpha y Symplypleona) y enquitreidos recogidos en ambas regiones.
Eje Autovalores Varianza explicada (%) p
1 0,0440 65,410 0,0408
2 0,0138 20,570 0,2594
3 0,0067 10,050 0,2072
Al igual que en el caso anterior, el primer eje separa hacia su lado positivo a los
microartrópodos con claras preferencias por los valores altos de temperatura y pH del suelo,
que en este caso fueron los oribátidos y los colémbolos poduromorfos (Fig. 60). En cambio, en
el lado negativo del eje se sitúan por un lado, los enquitreidos y los colémbolos del suborden
Symphypleona estrechamente relacionados con los valores altos de humedad y bajos de pH,
junto con los ácaros del suborden Prostigmata con claras preferencias por los suelos más ácidos
con altos contenidos de materia orgánica y por otro, los ácaros del orden Mesostigmata con
menores abundancias en suelos más cálidos (Fig. 60).
Finalmente, en el caso de los colémbolos entomobriomorfos la única variable edáfica
investigada que parece tener una influencia positiva sobre sus poblaciones fue el C de la biomasa
microbiana (Fig. 60).

Capítulo III
133
Figura 60: Análisis de correspondencias canónicas de las variables ambientales y los grupos principales de mesofauna (presentes en al menos un 20% de las muestras): ácaros (órdenes Oribatida, Mesostigmata y Prostigmata), colémbolos (subórdenes Poduromorpha, Entomobryomorpha y Symplypleona) y enquitreidos capturados en la región
atlántica y mediterránea.
Cuando se analizan las dos regiones por separado, el ACC realizado con la matriz de datos de
los suelos atlánticos dio como resultado una ordenación significativa de las variables (traza de
Pillai=0,062; p=0,009). En este caso se observa que la varianza está más repartida entre los ejes,
siendo los tres primeros significativos y explicando más del 99% de la varianza total (Tabla 27).
Tabla 27: Autovalores, varianza explicada y significación de cada uno de los ejes del ACC para la matriz de datos de abundancia de los grupos faunísticos de mesofauna con un porcentaje de presencia superior al 20% del total de muestras: ácaros (órden Oribatida), colémbolos (subórdenes Poduromorpha, Entomobryomorpha y Symplypleona) y enquitreidos capturados en la región atlántica.
Eje Autovalores Varianza explicada (%) p
1 0,0395 62,820 0,029
2 0,0150 23,910 0,066
3 0,0083 13,210 0,003
La representación gráfica de los dos primeros ejes indicó que el primero de ellos se explica
fundamentalmente por los dos factores abióticos, temperatura y humedad del suelo, y en menor
medida por el contenido de C del suelo, mientras que el segundo muestra una mayor relación
con el pH y en menor medida con los contenidos de N del suelo y de C de la biomasa microbiana
(Fig. 61).
De acuerdo con esto, la región atlántica se caracteriza por la mayor abundancia de ácaros
oribátidos en aquellos suelos de cultivo con una mayor temperatura de suelo y menor humedad,
mientras que los colémbolos entomobriomorfos abundan en los suelos más frios y húmedos de
esta región (Fig. 61). Los enquitreidos, además de preferir suelos fríos y húmedos, también
resultan beneficiados por valores bajos de pH, N y C de la biomasa microbiana (Fig. 61).
EJE 1
EJE 2
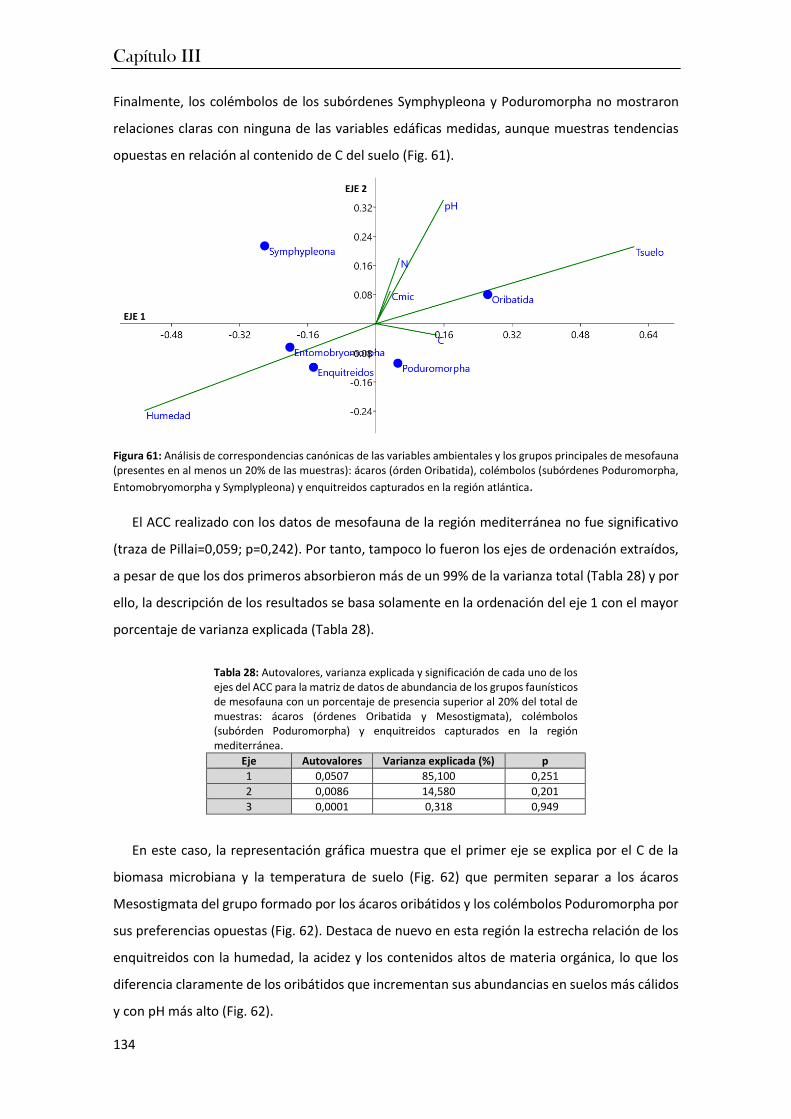
Capítulo III
134
Finalmente, los colémbolos de los subórdenes Symphypleona y Poduromorpha no mostraron
relaciones claras con ninguna de las variables edáficas medidas, aunque muestras tendencias
opuestas en relación al contenido de C del suelo (Fig. 61).
Figura 61: Análisis de correspondencias canónicas de las variables ambientales y los grupos principales de mesofauna (presentes en al menos un 20% de las muestras): ácaros (órden Oribatida), colémbolos (subórdenes Poduromorpha,
Entomobryomorpha y Symplypleona) y enquitreidos capturados en la región atlántica.
El ACC realizado con los datos de mesofauna de la región mediterránea no fue significativo
(traza de Pillai=0,059; p=0,242). Por tanto, tampoco lo fueron los ejes de ordenación extraídos,
a pesar de que los dos primeros absorbieron más de un 99% de la varianza total (Tabla 28) y por
ello, la descripción de los resultados se basa solamente en la ordenación del eje 1 con el mayor
porcentaje de varianza explicada (Tabla 28).
Tabla 28: Autovalores, varianza explicada y significación de cada uno de los ejes del ACC para la matriz de datos de abundancia de los grupos faunísticos de mesofauna con un porcentaje de presencia superior al 20% del total de muestras: ácaros (órdenes Oribatida y Mesostigmata), colémbolos (subórden Poduromorpha) y enquitreidos capturados en la región mediterránea.
Eje Autovalores Varianza explicada (%) p
1 0,0507 85,100 0,251
2 0,0086 14,580 0,201
3 0,0001 0,318 0,949
En este caso, la representación gráfica muestra que el primer eje se explica por el C de la
biomasa microbiana y la temperatura de suelo (Fig. 62) que permiten separar a los ácaros
Mesostigmata del grupo formado por los ácaros oribátidos y los colémbolos Poduromorpha por
sus preferencias opuestas (Fig. 62). Destaca de nuevo en esta región la estrecha relación de los
enquitreidos con la humedad, la acidez y los contenidos altos de materia orgánica, lo que los
diferencia claramente de los oribátidos que incrementan sus abundancias en suelos más cálidos
y con pH más alto (Fig. 62).
EJE 1
EJE 2

Capítulo III
135
Figura 62: Análisis de correspondencias canónicas de las variables ambientales y los grupos principales de mesofauna (presentes en al menos un 20% de las muestras): ácaros (órdenes Oribatida y Mesostigmata), colémbolos (subórden
Poduromorpha) y enquitreidos capturados en la región mediterránea. 3.3. Efectos de los parámetros edafoquímicos sobre la estructura de las comunidades de macrofauna en las dos regiones investigadas
El análisis de correspondencias realizado con los datos de abundancia de lombrices
identificadas a nivel de especie en ambas regiones resultó ser significativo (traza de Pillai=1,541;
p=0,0001), explicando los tres primeros ejes más de un 89% de la varianza total extraída (Tabla
29).
Tabla 29: Autovalores, varianza explicada y significación de cada uno de los ejes del ACC para la matriz de datos de abundancia de las distintas especies de lombrices recogidas en ambas regiones.
Eje Autovalores Varianza explicada (%) p
1 0,743 48,25 0,0001
2 0,492 31,94 0,0001
3 0,149 9,72 0,0025
En la representación gráfica de los dos primeros ejes se puede comprobar que el primer eje
aparece explicado fundamentalmente por el C de la biomasa microbiana, pero también por los
valores de humedad, materia orgánica (%C y %N) y pH (Fig. 63). La primera variable permite
identificar a Scherotheca gigas y Aporrectodea trapezoides como las especies más
correlacionadas con los valores altos de biomasa microbiana (Fig. 63).
Por otro lado, en la parte positiva de los dos ejes se sitúan las lombrices de pequeño tamaño
que viven en los horizontes superficiales del suelo (Lumbricus rubellus, Dendrodrilus rubidus,
EJE 1
EJE 2

Capítulo III
136
Dendrobaena octaedra, D. madeirensis y Microscolex phosphoreus) y con preferencia por suelos
húmedos, orgánicos y ácidos (Fig. 63). El grupo de especies formado por Lumbricus friendi,
Octolasion lacteum y Amynthas corticis también parece estar favorecido, aunque en menor
medida, por estas variables, siendo la humedad la variable más correlacionada con estas
especies (Figura 63).
Enn la parte negativa de ambos ejes se sitúan las especies de mayor tamaño como Octolasion
cyaneum, Octodrilus complanatus y Lumbricus terrestris, junto con Aporroctodea rosea y
Allolobophora chlorotica, que parecen preferir suelos con menos carga orgánica, más básicos y
menos húmedos (Fig. 63).
Fuera de este gradiente de pH, humedad y materia orgánica se sitúa Aporrectodea caliginosa
asociada con valores medios de todas las variables analizadas.
Figura 63: Análisis de correspondencias canónicas de las variables ambientales y las especies de lombrices de tierra capturadas en la región atlántica y mediterránea. Abreviaturas de las especies: ACh, Allolobophora chlorotica; ACo, Amynthas corticis; AC, Aporrectodea caliginosa; AR, Aporrectodea rosea; AT, Aporrectodea trapezoides; DO, Dendrobaena octaedra; DM, Dendrobaena madeirensis; DR, Dendrodrilus rubidus; LF, Lumbricus friendi; LR, Lumbricus rubellus; LT; Lumbricus terrestris; OC, Octodrilus complanatus; OCy, Octolasion cyaneum; OL, Octolasion lacteum; MP, Microscolex phosphoreus; SG, Scherotheca gigas.
El análisis de correspondencias realizado con la matriz de datos de abundancias de lombrices
capturadas en ambas regiones pero agrupadas por categorías ecológicas dio como resultado un
ordenación multivariante significativa (traza de Pillai=0,356; p=0,0001), en la que los dos
primeros ejes fueron significativos y explicaron un 97% de la varianza total extraída (Tabla 30).
EJE 1
EJE 2

Capítulo III
137
Tabla 30: Autovalores, varianza explicada y significación de cada uno de los ejes del ACC para la matriz de datos de abundancia de las distintas categorías ecológicas de lombrices (epigeas, anécicas, endogeas y epiendogeas) recogidas en ambas regiones.
Eje Autovalores Varianza explicada (%) p
1 0,2877 80,820 0,0001
2 0,0584 16,410 0,0006
3 0,0098 2,776 0,1083
La salida del análisis (Fig. 64) muestra que el eje 1 se explica fundamentalmente por el
contenido de C del suelo y menor medida por el pH. En cambio, el segundo eje sólo muestra una
estrecha relación con el C de la biomasa microbiana y con mucha menor relevancia la
temperatura del suelo. De las cuatro categorías ecológicas investigadas, la de las epigeas es la
que está más positivamente influenciada por los altos contenidos de C del suelo. En cambio, las
lombrices anécicas y epiendogeas se sitúan a lo largo de un gradiente de pH, materia orgánica y
humedad con tendencias opuestas (Fig. 64). La ordenación del eje 2 permite separar a las
lombrices anécicas de las endogeas y epiendogeas, por sus relaciones con los valores altos o
bajos del C de la biomasa microbiana, respectivamente (Fig. 64).
Figura 64: Análisis de correspondencias canónicas de las variables ambientales y las categorías ecológicas de lombrices de tierra (epigeas, anécicas, endogeas y epiendogeas) identificadas en las regiones atlántica y
mediterránea.
Cuando se analizan las matrices de datos de cada una de las regiones por separado mediante
ACC, se observa que, para la región atlántica, la ordenación fue significativa (traza de
Pillai=0,252; p=0,0001) y que los dos primeros ejes, también significativos, explicaron más de un
97% de la varianza total (Tabla 31).

Capítulo III
138
Tabla 31: Autovalores, varianza explicada y significación de cada uno de los ejes del ACC para la matriz de datos de abundancia de las cuatro categorías ecológicas de lombrices de tierra (epigeas, anécicas, endogeas y epiendogeas) capturadas en la región atlántica.
Eje Autovalores Varianza explicada (%) p
1 0,2001 79,450 0,0003
2 0,0442 17,560 0,0137
3 0,0075 2,995 0,1724
En el gráfico resultante (Fig. 65) se observa que el primer eje está correlacionado
principalmente con el contenido de C y N y la humedad del suelo y en menor medida por el pH.
Los valores altos de humedad, %C y %N caracterizan a los suelos de cultivo atlánticos dominados
por lombrices epigeas (Fig. 65). En cambio, los suelos más secos y menos orgánicos contienen
una mayor abundancia de lombrices anécicas, endogeas y epiendogeas y muy pocas epigeas
(Fig. 65).
Aunque el segundo eje no muestra correlación alta con ninguna variable edáfica, sí permite
separar las lombrices epiendogeas y en menor medida, las anécicas, con preferencias por los
valores más bajos de biomasa microbiana, de las endogeas con una mayor predilección por los
suelos menos ácidos con abundante biomasa microbiana (Fig. 65).
Figura 65: Análisis de correspondencias canónicas de las variables ambientales y las categorías ecológicas de
lombrices de tierra (epigeas, anécicas, endogeas y epiendogeas) identificadas en la región atlántica.
En el caso de la región mediterránea, no se encontraron lombrices epigeas por lo que los
análisis de ACC se realizaron para los datos de abundancia de las otras tres categorías ecológicas.
La ordenación multivariante de estos datos fue significativa (traza de Pillai=0,298; p=0,0004),
siendo además los dos ejes extraídos significativos (Tabla 32).
EJE 1
EJE 2

Capítulo III
139
Tabla 32: Autovalores, varianza explicada y significación de cada uno de los ejes del ACC para la matriz de datos de abundancia de las cuatro categorías ecológicas de lombrices de tierra (anécicas, endogeas y epiendogeas) capturadas en la región mediterránea.
Eje Autovalores Varianza explicada (%) p
1 0,2093 70,180 0,0030
2 0,0889 29,820 0,0033
El primer eje, que absorbe el mayor porcentaje de varianza, aparece correlacionado sobre
todo con el pH y el C microbiano y en menor medida con la humedad, el pH y el contenido de C
del suelo (Fig. 66). Estas variables permiten separar a los suelos de cultivo con valores más bajos
del pH del suelo, más húmedos y con mayor contenido de C y de biomasa microbiana por su
mayor abundancia de lombrices anécicas y la menor de endogeas, que muestran preferencias
contrarias (Fig. 66).
Debido a la escasa abundancia de lombrices epiendogeas en los suelos de cultivo de esta
región, éstas tienden a ubicarse en la periferia del diagrama de ordenación. A pesar de esto, esta
categoría ecológica de lombrices parece mostrar preferencias ecológicas similares a las de las
lombrices anécicas y además, con una clara preferencia por los valores bajos de temperatura
del suelo (Fig. 66).
Figura 66: Análisis de correspondencias canónicas de las variables ambientales y las categorías ecológicas de
lombrices de tierra (anécicas, endogeas y epiendogeas) identificadas en la región mediterránea.
4. DISCUSIÓN
Los resultados de los análisis de correspondencias canónicas de las comunidades edáficas y
de los parámetros ambientales medidos evidenciaron, en primer lugar, la importancia de la
humedad del suelo para la supervivencia de los dos grupos de oligoquetos (enquitreidos y
lombrices) en comparación con los microartrópodos. A diferencia de los ácaros y colémbolos
EJE 1
EJE 2

Capítulo III
140
con exoesqueletos quitinosos, estos anélidos necesitan que su tegumento esté siempre húmedo
para poder realizar el intercambio gaseoso (respiración cutánea). La dependencia de los
enquitreidos de la humedad ha sido descrita por varios autores como Kasprzak (1982) y Briones
et al. (1997) y es un reflejo de su gran vulnerabilidad frente a condiciones de desecación (Jänsch
et al. 2005). En el caso de las lombrices de tierra, si bien muchas especies han desarrollado
adaptaciones fisiológicas y de comportamiento (como la migración vertical hacia horizontes más
profundos y la adopción de estrategias de resistencia como la quiescencia y la diapausa) para
poder sobrevivir durante periodos secos (Lavelle 1983; Bouché 1984; Lee 1985), el contenido de
agua del suelo sigue siendo un factor limitante para la reproducción. Al igual que los
enquitreidos, la elevada permeabilidad de la piel hace que puedan llegar a deshidratarse cuando
se exponen al aire (Adolph 1927).
Los resultados de la ordenación del conjunto de los datos también permiten concluir que los
dos grupos de microartrópodos (ácaros y colémbolos) muestras preferencias ecológicas
distintas, destacando la estrecha relación entre la temperatura del suelo y las abundancias de
ácaros. En este sentido, se ha podido comprobar que ciertos grupos de ácaros edáficos,
concretamente los Prostigmata y los oribátidos, incrementan sus poblaciones durante la
primavera y el verano, cuando las temperaturas son más altas y hay menos humedad en los
suelos (Perdue y Crossley 1989). Sin embargo, temperaturas demasiado elevadas pueden
resultar limitantes para su supervivencia (Badejo 1990; Adejuyigbe et al. 1999).
Ninguna de las variables analizadas parece explicar de forma clara las densidades de
colémbolos en los suelos investigados, aunque sí se observó una cierta relación con el pH y con
el C de la biomasa microbiana. La respuesta de estos organismos a los diferentes valores de pH
en el suelo varía mucho según las especies; de hecho Rusek (1998) estableció una clasificación
de colémbolos en función a sus respuetas a acidificación/encalado en el suelo. Sin embargo,
otros autores (Thibaud y Christian 1989; Pozo 1986; Cutz‑Pool et al. 2003) han demostrado que
el pH del suelo es un factor clave en la abundancia de los colémbolos y por ejemplo, Hutson
(1978) observó que la longevidad de estos organismos se incrementa en el rango de pH=4-6,
mientras que para la reproducción el rango aumenta hasta 5-7, aunque no describe los
mecanismos específicos de cómo el pH interactúa con la biología de los colémbolos. Por otro
lado, se ha observado que los efectos positivos de la biomasa microbiana sobre sus poblaciones
es mayor en el caso de las especies euedáficas (suborden Poduromorpha) que sobre aquellos
que colonizan las capas más superficiales del suelo (subórdenes Entomobryomorpha y
Symphypleona) debido a la mayor estabilidad en las condiciones ambientales que caracateriza
a los horizontes más profundos y que favorece a estas especies con menor capacidad de
dispersión (Chagnon et al. 2000).

Capítulo III
141
Debido a que las posibles relaciones entre las variables edáficas y la macrofauna podrían
estar enmascaradas por las elevadas abundancias de los taxones de mesofauna registradas en
estos suelos, resulta adecuado un análisis detallado de estos dos grupos faunísticos por
separado. Así, para el caso de la mesofauna, en el análisis conjunto de los datos atlánticos y
mediterráneos, se volvió a confirmar la estrecha relación entre los ácaros oribátidos y la
temperatura del suelo y la de los enquitreidos y la humedad. La relación positiva entre las
abundancias de oribátidos y la temperatura del suelo ha sido descrito previamente (Webb et al.
1998; Briones et al. 2009) y por ejemplo, han permitido constatar un incremento de entre un 8-
10% en la abundancia de oribátidos al calentar experimentalmente suelos con alto contenido de
materia orgánica (Briones et al. 2009). En cambio, los ácaros Mesostigmata respondieron
negativamente a la temperatura del suelo y en este sentido, Manu et al. (2016) también
observaron importantes reducciones en sus poblaciones con los incrementos de temperatura.
Por último, los ácaros Prostigmata recolectados en ambas regiones aparecen asociados a los
valores altos de C y N y bajos de pH. Estos resultados coinciden con observaciones previas y así,
Hasegawa et al. (2013) encontraron una relación positiva entre abundancias de Prostigmata y
mayores contenido de N en el suelo y Mitchell et al. (2016) con los pH ácidos. Sin embargo, otros
estudios muestran preferencias ecológicas muy distintas y por ejemplo, Hågvar y Amundsen
(1981) detectaron un descenso de la abundancia y dominancia de Prostigmata al reducir el pH
del suelo, por lo que hacen falta más estudios para determinar la importancia relativa de estos
factores sobre sus tamaños poblacionales. La relación entre los valores altos de C y los ácaros
Prostigmata también ha sido descrita previamente (Noble et al. 1996; Hasegawa et al. 2013) y
se explica por una preferencia por los microorganismos como fuente de alimento y que
proliferan en los depósitos orgánicos (Hasegawa et al. 2013).
En el caso de los distintos subórdenes de colémbolos destacan las distintas preferencias
ecológicas en cuanto a pH y humedad del suelo de los Entomobryomorpha y Symphypleona. Así,
mientras los primeros abundan en aquellos suelos con valores altos de pH y bajos de humedad,
los segundos incrementan sus abundancias en suelos con pH ácidos y humedades altas. Aunque
ambos grupos son muy vulnerables a la desecación, se sabe que algunas especies son capaces
de vivir en condiciones ambientales inestables (Sousa y Gama 1994; Sousa et al. 1997) y
sobrevivir periodos de sequía durante periodos cortos (Álvarez et al. 1999). Por otro lado, el
efecto de la humedad y el pH sobre la distribución de las especies de colémbolos es indirecto,
ya que son los microorganismos los que responden de forma directa a variaciones de estos dos
factores abióticos y al ser una fuente de alimento para los colémbolos, éstos también se ven
afectados (Salmon et al. 2014). Además, a pesar de que la mayoría de las especies de colémbolos
de estos dos subórdenes son epiedáficas, es muy probable que especies diferentes ocupen

Capítulo III
142
distintos nichos para reducir la competencia entre ellos y, por tanto, que las respuestas a los
factores abióticos también varíen a nivel específico.
En el estudio de la mesofauna de cada región por separado, se confirma que el grupo
mayoritario de ácaros (Oribatida) muestra un comportamiento más homogéneo en las dos
regiones investigadas que los restantes integrantes de la mesofauna. Aunque en ambas regiones
prefieren suelos cálidos y secos, fueron especialmente abundantes en la región mediterránea,
donde se vieron también favorecidos por el pH del suelo. La relación observada entre oribátidos
y temperatura se ve corroborada por las observaciones de Stamou y Sgardelis (1989), quienes
concluyeron que la temperatura del suelo es el principal factor que controlan los cambios
poblacionales de los oribátidos en los climas mediterráneos. La epicutícula de estos organismos
es muy impermeable y evita la evapotranspiración (Madge 1965), lo cual les aporta una ventaja
frente al resto de taxones de mesofauna en condiciones de desecación. Block (1996) destaca la
resistencia de los oribátidos a la desecación al alifi a los o o o ga is os d -ai fo s . “i
embargo, Madge (1965) estudió las temperaturas letales (thermal death temperature) de estos
organismos y observó una menor mortalidad de oribátidos a temperaturas elevadas si las
condiciones humedad eran favorables. Esto es debido a que en ambientes muy secos los
oribátidos son incapaces de retener agua (Madge 1965). En cambio, el efecto positivo del pH
sobre las poblaciones de oribátidos contrasta con lo obtenido por otros autores en suelos
forestales donde generalmente la acidez del suelo favorece el aumento de sus densidades
(Hågvar y Amundsen 1981). En este sentido, se ha podido constatar que en la respuesta de los
oribátidos frente al pH también intervienen otros factores como la disponibilidad de alimento,
la humedad (van Straalen y Verhoef 1997) y la competencia inter específica (Hågvar 1990).
Otro orden de ácaros especialmente abundantes en la región mediterránea fueron los
Mesostigmata, pero en este caso la variable que más influyó en el tamaño de sus poblaciones
fue el carbono de la biomasa microbiana. Mitchell et al. (2016) observaron que ciertas
características del hábitat (por ejemplo, acumulación de materia orgánica) regulan la estructura
de las comunidades de estos ácaros explicando un 6% de su variación. La presencia de estos
ácaros depredadores podría haber reducido la abundancia de organismos microbívoros en el
suelo (puesto que la mayoría de las especies de Mesostigamata son predadoras) y haber
beneficiado, de manera indirecta, la biomasa microbiana (Hendrix et al. 1986; Dirilgen et al.
2016).
A diferencia de los ácaros, los distintos subórdenes de colémbolos muestran respuestas más
variables en las dos regiones. Por ejemplo, destaca el hecho de que el pH fue especialmente
importante para los colémbolos del suborden Poduromorpha, pero solamente en la región
mediterránea. De hecho, el número de individuos de este taxón en los suelos de esta región fue

Capítulo III
143
dos veces superior al registrado en los suelos atlánticos, lo que sugiere que estos colémbolos
son más sensibles a la acidez del suelo que los representantes de los otros dos subórdenes
(Entomobryomorpha y Symphypleona), lo cual se vio confirmado por la mayor abundancia de
especímenes de estos dos subórdenes en la región atlántica. Sin embargo, Fernandes et al.
(2009) en suelos de Brasil observaron que los colémbolos poduromorfos exhiben diferentes
rangos de tolerancia al pH edáfico. Por otro lado, los colémbolos entomobriomofos parecen
estar muy influenciados por la temperatura del suelo y la humedad, un hecho que se ve
confirmado por las escasas presencias de este grupo de colémbolos en la región mediterránea,
donde no llegaron a alcanzar el 20% de presencias en las muestras analizadas. Según Potapov et
al. (2016) la mayor parte de las especies de este suborden habitan las capas más superficiales
del suelo, en dónde están más expuestas a las condiciones climáticas adversas.
Los análisis realizados para el conjunto de datos de abundancia de lombrices (macrofauna)
muestran la existencia de una relación positiva entre las lombrices epigeas (en particular,
Lumbricus rubellus, Dendrobaena octaedra y Dendrodrilus rubidus) con altos contenidos de C y
N y de agua del suelo y bajos de pH. La presencia de una capa de hojarasca en el suelo de manera
permanente que conserve la humedad y proporcione alimento es un factor clave para la
supervivencia de las lombrices epigeas (Dunger 1989; Curry 2004). A diferencia del resto de
categorías ecológicas, estas lombrices toleran bien la la acidez del suelo (Curry 2004) y por tanto,
son especialmente abundantes en suelos ácidos y ricos en materia orgánica (Trigo et al. 1988).
Debido a que ocupan las capas superficiales del suelo (Gerard 1967; Lee 1985; Briones et al.
2009) son muy sensibles a condiciones de desecación y al aumento de temperatura. Además,
las lombrices epigeas tienen una menor capacidad excavadora, lo que les impide escapar
migrando hacia capas inferiores del suelo cuando la capas superficiales se secan (Keudel y
Schrader 1999). Están consideradas como estrategas de la r (Bouché 1977), es decir, ciclos de
vida muy cortos que responden muy rápidamente a los cambios en las condiciones ambientales.
Sin embargo, no parece que sus poblaciones hayan sido capaces de soportar las condiciones
extremas (elevadas temperaturas y sequías prolongadas) que caracterizan a los suelos de cultivo
mediterráneos investigados, ya que no se encontraron ejemplares de esta categoría ecológica
durante todo el periodo de estudio. La desaparición de esta categoría ecológica tiene
importantes implicaciones para el funcionamiento del ecosistema, ya que estas especies
favorecen la retención de C en el suelo hasta u % del a o o e o t ado e sus casts
proviene de la hojarasca del suelo; Jégou et al. 1998). Desde un punto de vista funcional están
i luidas e el g upo de t asfo ado es de la hoja as a La elle al t a sfo a físi a
químicamente los restos vegetales, reduciendo la relación C/N y favoreciendo el ataque
microbiano (Fragoso et al. 1997).

Capítulo III
144
Los análisis multivariantes para el conjunto de datos también indican la existencia de una
cierta relación positiva entre la abundancia de las lombrices anécicas (en particular, Scherotheca
gigas, Lumbricus terrestris, Octodrilus complanatus y Aporrectodea trapezoides) y valores altos
de pH y de biomasa microbiana y bajos de humedad y materia orgánica (%C y %N). Estas
relaciones explican su mayor dominancia en la región mediterránea. La mayoría de los estudios
previos coinciden en que los valores altos de pH favorecen a las lombrices anécicas (Muys et al.
1992; Muys y Granval 1997; Reich et al. 2005), lo que ha llevado a concluir que estas lombrices
son intolerantes a la acidez en el suelo, especialmente los juveniles (Hlava y Kopecký 2013).
Además, gracias a sus galerías verticales que pueden alcanzar los horizontes más profundos del
suelo (Bertrand et al. 2015) pueden buscar refugio y sobrevivir a periodos de sequía. Dado que
aceleran la descomposición de la materia orgánica depositada (Wardle 2000), el C y el N
contenidos en la hojarasca son asimilados rápidamente por las comunidades microbiológicas,
reduciéndose así la presencia de estos elementos en el suelo (Fahey et al. 2013). Estas lombrices
deposita sus he es asts e la supe fi ie que son muy ricas en compuestos de carbono que
sirven de fuente de alimento para los microorganismos (Shipitalo y Protz 1988; Marinissen 1994;
Guggenberger et al. 1996). Estas acumulaciones de heces junto con restos vegetales en las
proximidades de las aberturas de las galerías (los deno i ados idde s ) sirven también de
refugio para las poblaciones microbianas (Bohlen et al. 1997; Subler y Kirsch 1998; Stroud et al.
2016) por lo que explicaría la estrecha relación entre esta categoría ecológica de lombrices y los
microorganismos.
En cambio, las lombrices endogeas mostraron correlaciones con valores altos de humedad y
bajos de pH para el conjunto de datos obtenidos de ambas regiones. Sin embargo, cuando se
analizaron las dos regiones por separado, estas preferencias ecológicas solamente fueron
observadas en los suelos de la región mediterránea, mientras que en la región atlántica las
poblaciones de estas lombrices mostraron preferencias opuestas. Esto puede ser debido a que
las especies de lombrices endogeas recogidas exclusivamente en la región mediterránea
(Allolobophora chlorotica, Aporrectodea rosea y Octolasion cyaneum) son más tolerantes a las
condiciones de sequía que la especie endogea típicamente atlántica Octolasion lacteum,
caracterizada por estar bien adaptada a ambientes húmedos (Trigo et al. 1988; Rodríguez et al.
1997). Además, Ma et al. (1990) concluyeron que las lombrices endogeas son más sensibles que
las epigeas a la acidificación del suelo causada por la adición prolongada de fertilizantes
minerales. De hecho, han sido descritas por Bouché (1972) como lombrices neutrófilas, cuyo
rango óptimo de pH es de entre 6.0 y 7.0. Estos resultados demuestran que la acidificación del
suelo es uno de los factores que afecta más negativamente a la abundancia de lombrices (Ivask

Capítulo III
145
et al. 2006), hasta el punto de que en ocasiones se ha llegado a observar la desaparición total
de lombrices anécicas y endogeas en suelos acidificados (Potthoff et al. 2008).
Las lombrices epiendogeas recolectadas en los suelos de cultivo de ambas regiones
mostraron relaciones positivas con la humedad y negativas con la temperatura de suelo y el pH,
lo cual explica por qué las tres especies incluídas en esta categoría (Dendrobaena madeirensis,
Amynthas corticis y Microscolex phosphoreus) fueron más abundantes en la región atlántica que
en la mediterránea. En la región atlántica, además, se vieron favorecidas por los valores bajos
de biomasa microbiana, mientras que en la región mediterránea ocurrió lo contrario. Desde un
punto de vista ecológico presentan características intermedias entre epigeas y endogeas
(Bouché 1977) y Lavelle (1979) las considera como un subgrupo de lombrices endogeas que
habitan en las capas más superficiales del suelo. De hecho, en la región atlántica dos de ellas,
Dendrobaena madeirensis y Microscolex phosphoreus, parecen tener preferencias ecológicas
muy similares a las de las especies epigeas (es decir, suelos ácidos con valores altos de C, N y
humedad). Sin embargo, Amynthas corticis, una especie invasora originaria de Asia (Burtelow et
al. 1998), no presentó ninguna relación con las variables edáficas estudiadas.
5. CONCLUSIONES
Los resultados de este estudio reflejan que la humedad, el pH del suelo y la materia orgánica
fueron los factores con una mayor influencia sobre las abundancias y la estructura de las
poblaciones de organismos edáficos en los suelos de cultivo de ambas regiones. Destaca la
humedad como factor clave para las poblaciones de los dos grupos de oligoquetos investigados,
de ahí que periodos de sequía mas frecuentes y prolongados pueden llegar a comprometer
seriamente su supervivencia. En cambio, los ácaros, y en especial los oribátidos, con sus
exoesqueletos quitinosos son capaces de sobrevir condiciones de temperatura y humedad
extremas como las que caracterizan los suelos de la región mediterránea. El pH del suelo fue la
variable más influyente para las poblaciones de colémbolos, pero la respuesta fue diferente
según el grupo.
Por otro lado, las condiciones de humedad, el pH, la materia orgánica y la biomasa
microbiana explican la diferente estructura funcional de las comunidades de lombrices de tierra
en las dos regiones geográficas. La ausencia de lombrices epigeas en los cultivos mediterráneos
se debe a que estas especies prefieren suelos ácidos con altos contenidos de C y N. Los suelos
ácidos y húmedos son también los preferidos por las lombrices endogeas, a diferencia de las
anécicas que incrementan sus densidades en suelos con valores de pH y biomasa microbiana
más altos. Por tanto, si bien en la región atlántica la materia orgánica fresca puede ser procesada

Capítulo III
146
rápidamente gracias a la abundancia de especies que viven en la superficie, el efecto positivo
sobre la porosidad y la incorporación de la materia orgánica hacia los horizontes más profundos
del suelo se ve comprometido por las menores densidades de lombrices anécicas.
6. BIBLIOGRAFÍA
Adejuyigbe C O, Tian G y Adeoye G O. 1999. Soil microarthropod populations under natural and planted fallows in southwestern Nigeria. Agroforestry Systems, 47, 263-272.
Adolph E F. 1927. The regulation of volume and concentration in the body fluids of earthworms. Journal of Experimental Zoology Part A: Ecological Genetics & Physiology, 47(1), 31-62.
Alvarez T, Frampton G K y Goulson D. 1999. The effects of drought upon epigeal collembola from arable soils. Agricultural & Forest Entomology, 1, 243-24.
Badejo M A. 1990. Seasonal abundance of soil mites (Acarina) in two contrasting environments. Biotropica, 22, 382-390.
Bardgett R D y Cook C R. 1998. Functional aspects of soil animal diversity in agricultural grasslands. Applied Soil Ecology, 10, 263-276.
Beare M H, Parmelee R W, Hendrix P F, Cheng W, Coleman D C y Crossley D A. 1992. Microbial and faunal interactions and effects on litter nitrogen and decomposition in agroecosystems. Ecological Monographs, 62(4), 569-591.
Bertrand M, Barot S, Blouin M, Whalen J, De Oliveira T y Roger-Estrade J. 2015. Earthworm services for cropping systems. A review. Agronomy for Sustainable Development, 35(2), 553-567.
Block W. 1996. Cold or drought-the lesser of two evils for terrestrial arthropods? European Journal of Entomology, 93, 325-340.
Bohlen P J, Parmelee R W, McCartney D A, Edwards C A. 1997. Earthworm effects on carbon and nitrogen dynamics of surface litter in corn agroecosystems. Ecological Applications, 7, 1341-1349.
Bouché M B. 1972. Lombriciens de France. Ecologie et Systématique. Annales de Zoologie et Ecologie Animale, Numéro Hors-Série. Institut National de la Recherche Agronomique, Paris.
Bouché M B. 1977. Strategies lombriciences. En: Lohm U y Persson T (Eds.) Soil Organisms as Components of Ecosystems, pp. 122-132. Ecological Bulletins, Stockholm.
Bouché M B. 1984. Les modalités d'adaptation des lombriciens à la sécheresse. Bulletin de la Société Botanique de France. Actualités Botaniques, 131(2-4), 319-327.
Briones M J I, Ineson P y Heinemeyer A. 2007. Predicting potential impacts of climate change on the geog aphi al dist i utio of e h t aeids: a eta‐a al sis app oa h. Global Change Biology, 13(11), 2252-2269.
Briones M J I, Ineson P y Piearce T G. 1997. Effects of climate change on soil fauna: responses of enchytraeids, diptera larvae and tardigrades in a transplant experiment. Applied Soil Ecology, 6, 117–134.
Briones M J I, Ostle N J, McNamara N P y Poskitt J. 2009. Functional shifts of grassland soil communities in response to soil warming. Soil Biology & Biochemistry, 41(2), 315-322.
Burtelow A E, Bohlen P J y Groffman P M. 1998. Influence of exotic earthworm invasion on soil organic matter, microbial biomass and denitrification potential in forest soils of the northeastern United States. Applied Soil Ecology, 9(1), 197-202.
Cao Z, Han X, Hu C, Chen J, Zhang D y Steinberger Y. 2011. Changes in the abundance and structure of a soil mite (Acari) community under long-term organic and chemical fertilizer treatments. Applied Soil Ecology, 49, 131-138.
Carrera N, Barreal M E, Gallego P P y Briones M J I. 2009. Soil invertebrates control peatland c fluxes in response to warming. Functional Ecology, 23, 637-648.
Chagnon M, Hebert C y Pare D. 2000. Community structures of collembola in sugar maple forests: relations to humus type and seasonal trends. Pedobiologia, 44, 148–174.
Chan K Y y Mead J A. 2003. Soil acidity limits colonisation by Aporrectodea trapezoides, an exotic earthworm. Pedobiologia, 47(3), 225-229.
Coleman D C, Crossley D A y Hendrix PF. 2004. Fundamentals of Soil Ecology. London: Academic Press. Curry J P. 2004. Factors affecting the abundance of earthworms in soils. En: Edwards C A (Ed.) Earthworm
Ecology. pp 91-114. CRC Press LLC, Boca Raton, FL.

Capítulo III
147
Cutz-Pool L Q, Palacios-Vargas J y Vázquez M M. 2003. Comparación de algunos aspectos ecológicos de collembola en cuatro asociaciones vegetales de Noh-Bec, Quintana Roo, México. Folia Entomológica Mexicana, 42(1), 91-101.
Dirilgen T, Arroyo J, Dimmers W J, Faber J, Stone D, da Silva PM, Carvalho F, Schmelz R, Griffiths B S, Francisco R, Creamer R E, Sousa J P y Bolger T. 2016. Mite community composition across a European transect and its relationships to variation in other components of soil biodiversity. Applied Soil Ecology, 97, 86-97.
Didden W A M. 1990. Involvement of Enchytraeidae (Oligochaeta) in soil structure evolution in agricultural fields. Biology & Fertility of Soils, 9, 152-158.
Dunger W. 1989. The return of soil fauna to coal mined areas in the german democratic republic. En: Majer J D (Ed.) Animals in Primary Succession: The Role of Fauna in Reclaimed Lands, pp 307-337. Cambridge University Press, Cambridge.
Edwards C A. 1983. Earthworm ecology in cultivated soils. En: Satchell J E (Ed.) Earthworm Ecology: From Darwin to Vermiculture, pp 123-137. Chapman & Hall, London.
Edwards C A y Bohlen P J. 1996. Biology and Ecology of Earthworms. Chapman & Hall, London. Edwards C A y Lofty J R. 1975. The influence of cultivations on soil animal populations. En: Vanek J (Ed.)
Progress in Soil Zoology, pp 399-407. Academia Publishing House, Prague. El Titi A y Ulber B. 1991. Significance of biotic interactions between soil fauna and microflora in integrated
arable farming. Developments in Agricultural & Managed Forest Ecology, 23, 1-19. Fahey T J, Yavitt J B, Sherman R E, Maerz J C, Groffman P M, Fisk M C y Bohlen P J. 2013. Earthworm effects
on the incorporation of litter C and N into soil organic matter in a sugar maple forest. Ecological Applications, 23(5), 1185-1201.
Fernandes L H, Nessimian J L y Mendonça M C D. 2009. Structure of Poduromorpha (Collembola) communities in "restinga" environments in Brazil. Pesquisa Agropecuaria Brasileira, 44(8), 1033-1039.
Filser J. 2002. The role of collembola in carbon and nitrogen cycling in soil: Proceedings of the X International Colloquium on Apte gota, České Budějo i e : Apte gota at the Begi i g of the Third Millennium. Pedobiologia, 46(3-4), 234-245.
Fragoso C y Lavelle P. 1992. Earthworm communities of tropical rain forests. Soil Biology and Biochemistry, 24, 1397-408.
Fragoso C, Brown G G, Patron J C, Blanchart E, Lavelle P, Pashanasi B, Senepati B y Kumar T. 1997. Agricultural intensification, soil biodiversity and agroecosystem function in the tropics: the role of earthworms. Applied Soil Ecology, 6(1), 17-35.
Ga ía‐Pala ios P, Maest e F T, Kattge J Wall D H. . Climate and litter quality differently modulate the effects of soil fauna on litter decomposition across biomes. Ecology Letters, 16(8), 1045-1053.
Gerard B M. 1967. Factors affecting earthworms in pastures. Journal of Animal Ecology, 36(1), 235-252. Gliessman S R. 1998. Agroecology. Ecological Processes in Sustainable Agriculture. Ann Arbor Press,
Chelsea, MI, ISBN 1-57504-043-3. Gliessman S R. 2006. The agroecosystem concept. En: Gliessman S R (Ed.) Agroecology: The Ecology of
Sustainable Food Systems, pp. 23-32. CRC Press. Guggenberger G, Thomas R J y Zech W. 1996. Soil organic matter within earthworm casts of an anecic-
endogeic tropical pasture community, Colombia. Applied Soil Ecology, 3(3), 263-274.
Hammer ∅, Harper D A T y Ryan P D. 2001. PAST: Paleontological Statistics software package for education and data analysis. Paleontologia Electronica, 4(1), 9.
Harper J L. 1974. Agricultural Ecosystems. Agroecosystems, 1, 1-6. Hasegawa M, Okabe K, Fukuyama K, Makino S I, Okochi I, Tanaka H, Goto H, Mizoguchi T y Sakata T. 2013.
Community structures of Mesostigmata, prostigmata and oribatida in broad-leaved regeneration forests and conifer plantations of various ages. Experimental & Applied Acarology, 59(4), 391-408.
Hågvar S. 1990. Reactions To Soil Acidification In Microarthropods: Is Competition A Key Factor? Biology & Fertility of Soils, 9(2), 178-181.
Hågvar S y Amundsen T. 1981. Effects of liming and artificial acid rain on the mite (Acari) fauna in coniferous forest. Oikos, 37, 7-20.
Hendrix P F, Parmelee R W, Crossley D A, Coleman D C, Odum E P y Groffman P M. 1986. Detritus food webs in conventional and no-tillage agroecosystems. BioScience, 36, 374-380.
Hlava J y Kopecký O. 2013. Earthworm population responses to deciduous forest soil acidity and vegetation cover. Scientia Agriculturae Bohemica, 44(3), 133-137.
Hutson B R. 1978. Influence of pH, temperature and salinity on the fecundity and longevity of four species of Collembola. Pedobiologia, 18, 163-179.

Capítulo III
148
Illig J, Langel R, Norton R A, Scheu S y Maraun M. 2005. Where are the decomposers? Uncovering the soil food web of a tropical montane rain forest in southern Ecuador using stable isotopes (15N). Journal of Tropical Ecology, 21(5), 589-593.
Ivask M, Kuu A, Truu M y Truu J. 2006. The effect of soil type and soil moisture on earthworm communities. Journal of Agricultural Science, 17, 7-11.
Jänsch S, Römbke J y Didden W. 2005. The use of enchytraeids in ecological soil classification and assessment concepts. Ecotoxicology & Environmental Safety, 62(2), 266-277.
Jegen G. 1920. Die Bedeutung Der Enchytraeiden Für Die Humusbildung. Landwirtschaftliches Jahrbuch der Schweiz, 34, 55-71.
Jégou D, Cluzeau D, Balesdent J y Tréhen P. 1998. Effects of four ecological categories of earthworms on carbon transfer in soil. Applied Soil Ecology, 9(1), 249-255.
Johnson D, Krsek M, Wellington E M, Stott A W, Cole L, Bardgett R D, Read D J y Leake J R. 2005. Soil invertebrates disrupt carbon flow through fungal networks. Science, 309(5737), 1047-1047.
Kasprzak K. 1982. Review of Enchytraeid (Oligochaeta, Enychtraeidae) community structure and function in agricultural ecosystems. Pedobiologia, 23, 217-232.
Keudel M y Schrader S. 1999. Axial and radial pressure exerted by earthworms of different ecological groups. Biology & Fertility of Soils, 29(3), 262-269.
Kherbouche D, Bernhard-Reversat F, Moali A y Lavelle P. 2012. The effect of crops and farming practices on earthworm communities in Soummam valley, Algeria. European Journal of Soil Biology, 48, 17-23.
Lago C F, 2015. Efecto del manejo agrícola sobre las propiedades biológicas y químicas del suelo y los procesos de transformación de la materia orgánica en cultivos de kiwi (Actinidia deliciosa). (Tesis Doctoral). Universidad de Vigo, Vigo.
Lavelle P. 1979. Relations entre typesecologiques et profiles demographiques chez les vers de terre de la savanne De La to Cote d I o e . Revue d'Écologie et de Biologie du Sol, 16, 85-101.
Lavelle P. 1983. The structure of earthworm communities. En: Stachell J E (Ed.) Earthworm Ecology: From Darwin to Vermiculture, pp 449-466. Chapman & Hall, London.
Lavelle P. 1996. Diversity of soil fauna and ecosystem function. Biology International, 33, 3-16. Lavelle P. 1997. Faunal activities and soil processes: adaptive strategies that determine ecosystem
function. Advances in Ecological Research, 27, 93-132. Lavelle P y Spain A V. 2001. Soil Ecology. Kluwer Academic Publishers, Boston. Lee K E y Foster R C. 1991. Soil fauna and soil structure. Soil Research, 29(6), 745-775. Lee K E. 1985. Earthworms: their Ecology and Relationships with Soils and Land Use. Academic Press, New
York. Legendre P y Legendre L. 1998. Numerical Ecology: Second English Edition. Developments in
Environmental Modelling, 20. Lindberg N y Bengtsson J. 2005. Population responses of oribatid mites and collembolans after drought.
Applied Soil Ecology, 28(2), 163-174. Lindberg N, Bengtsson J B y Persson T. 2002. Effects of experimental irrigation and drought on the
composition and diversity of soil fauna in a coniferous stand. Journal of Applied Ecology, 39(6), 924-936.
Lowe C N y Butt K R. 2005. Culture techniques for soil dwelling earthworms: a review. Pedobiologia, 49(5), 401-413.
Ma W C, Brussaard L y De Ridder J A. 1990. Long-term effects of nitrogenous fertilizers on grassland earthworms (Oligochaeta: Lumbricidae): their relation to soil acidification. Agriculture, Ecosystems & Environment, 30(1-2), 71-80.
Madge D S. 1965. The effects of lethal temperatures on oribatid mites. Acarologia, 7(1), 121-130. Magdoff F. 2007. Ecological agriculture: principles, practices, and constraints. Renewable Agriculture &
Food Systems, 22(02), 109-117. Ma u M, Io da he V, Bă ilă ‘ I, Bodescu F y Onete M. 2016. The influence of environmental variables on
soil mite communities (Acari: Mesostigmata) from overgrazed grassland ecosystems–Romania. Italian Journal of Zoology, 83(1), 89-97
Maraun M, Schatz H y Scheu S. 2007. Awesome or ordinary? Global diversity patterns of oribatid mites. Ecography, 30(2), 209-216.
Marinissen J C Y. 1994. Earthworms and stability of soil structure: a study in a silt loam soil in a young Dutch polder. Agriculture, Ecosystems & Environment, 51, 75–87.

Capítulo III
149
Menta C. 2012. Soil fauna diversity-function, soil degradation, biological indices, soil restoration. En: Lameed G A (Eds.) Biodiversity Conservation and Utilization in a Diverse World, pp. 59-94. INTECH Open Access Publisher.
Mitchell R J, Urpeth H M, Britton A J, Black H y Taylor A R. 2016. Relative importance of local-and large-scale drivers of alpine soil microarthropod communities. Oecologia, 182(3), 913-924.
Moore J C, Walter D E y Hunt H W. 1988. Arthropod regulation of micro-and mesobiota in below-ground detrital food webs. Annual Review of Entomology, 33(1), 419-435.
Muys B y Granval P H. 1997. Earthworms as bio-indicators of forest site quality. Soil Biology & Biochemistry, 29(3-4), 323-328.
Muys B, Lust N y Granval P H. 1992. Effects of grassland afforestation with different tree species on earthworm communities, litter decomposition and nutrient status. Soil Biology & Biochemistry, 24(12), 1459-1466.
Noble J C, Whitford W G y Kaliszweski M. 1996. Soil and litter microarthropod populations from two contrasting ecosystems in semi-arid eastern Australia. Journal of Arid Environments, 32(3), 329-346.
O Co o F B. . The E h t aeidae. En: Burges A y Raw F (Eds.) Soil Biology, pp 213-256. Academic Press, New York.
Ochoa-Hueso R, Rocha I, Stevens C J, Manrique E y Luciañez M J. 2014. Simulated nitrogen deposition affects soil fauna from a semiarid mediterranean ecosystem in central Spain. Biology & Fertility of Soils, 50(1), 191-196.
Osler G H R y Murphy D V. 2005. Oribatid mite species richness and soil organic matter fractions in agricultural and native vegetation soils in Western Australia. Applied Soil Ecology, 29(1), 93-98.
Osler G H R y Sommerkorn M. 2007. Toward a complete soil C and N Cycle: incorporating the soil fauna. Ecology, 88(7), 1611-1621.
Paoletti M G, Sommaggio D, Favretto M R, Petruzzelli G, Pezzarossa B y Barbafieri M. 1998. Earthworms as useful bioindicators of agroecosystem sustainability in orchards and vineyards with different inputs. Applied Soil Ecology, 10, 137-150.
Perdue J C y Crossley D A. 1989. Seasonal abundance of soil mites (Acari) in experimental agroecosystems: effects of drought in no-tillage and conventional tillage. Soil & Tillage Research, 15(1-2), 117-124.
Petersen H y Luxton M. 1982. Acomparative analysis of soil fauna populations and their rolein decomposition processes. Oikos, 39, 287-388.
Petersen H. 1982. Basis for evaluation of the density and biomass data. Oikos, 39, 295-305. Petersen H. 1994. A review of collembolan ecology in ecosystem context. Acta Zoologica Fennica, 195,
111-118. Pillai K C S. 1955. Some new test criteria in multivariate analysis. The Annals of Mathematical Statistics,
26, 117-121. Potapov A A, Semenina E E, Korotkevich A Y, Kuznetsova N A y Tiunov A V. 2016. Connecting taxonomy
and ecology: trophic niches of collembolans as related to taxonomic identity and life forms. Soil Biology & Biochemistry, 101, 20-31.
Potthoff M, Asche N, Stein B, Muhs A y Beese F. 2008. Earthworm communities in temperate beech wood forest soils affected by liming. European Journal of Soil Biology, 44(2), 247-254.
Pozo J. 1986. Ecological factors affecting collembola populations. Ordination of communities. Revue d'Écologie et de Biologie du Sol, 23(3), 299-311.
Reich P B, Oleksyn J, Modrzynski J, Mrozinski P, Hobbie S E, Eissenstat D M, Chorover J, Chadwick O A, Hale C M y Tjoelker M G. 2005. Linking litter calcium, earthworms and soil properties: a common garden test with 14 tree species. Ecological Letters, 8, 811-818.
Rodríguez T, Trigo D y Cosín D J D. 1997. Biogeographical zonation of the western Iberian Peninsula on the basis of the distribution of earthworm species. Journal of Biogeography, 24(6), 893-901.
Rusek J. 1998. Biodiversity of Collembola and their functional role in the ecosystem. Biodiversity & Conservation, 7(9), 1207-1219.
Salmon S, Ponge J F, Gachet S, Deharveng L, Lefebvre N y Delabrosse F. 2014. Linking species, traits and habitat characteristics of collembola at European scale. Soil Biology & Biochemistry, 75, 73-85.
Satchell J E. 1967. Lumbricidae. En: Burges A y Raw F (Eds.) Soil Biology, pp 259-322. Academic Press, London.
Sechi V, D'Annibale A, Ambus P, Sárossy Z, Krogh P H, Eriksen J y Holmstrup M. 2014. Collembola feeding habits and niche specialization in agricultural grasslands of different composition. Soil Biology & Biochemistry, 74, 31-38.

Capítulo III
150
Shipitalo M J y Protz R. 1988. Factors influencing the dispersibility of clay in worm casts. Soil Science Society of America Journal, 52, 764-769.
Simonsen J, Posner J, Rosemeyer M Y, Baldock J. 2010. Endogeic and anecic earthworm abundance in six midwestern cropping systems. Applied Soil Ecology, 44(2), 147-155.
Song L, Liu J, Yan X, Chang L y Wu D. 2016. Euedaphic and hemiedaphic collembola suffer larger damages than epedaphic species to nitrogen input. Environmental Pollution, 208, 413-415.
Sousa J P y Gama M M. 1994. Rupture in a collembola community structure from a quercus rotundifolia lam. Forest due to the reafforestation with Eucalyptus globulus Labill. European Journal of Soil Biolology, 30, 71–78.
Sousa J P, Vingada J V, Barrocas H y Gama M M. 1997. Effects of introduced exotic tree species on collembola communities: the importance of management techniques. Pedobiologia, 41, 145–153.
Stamou G P y Sgardelis S P. 1989. Seasonal distribution patterns of oribatid mites (Acari: Cryptostigmata) in a forest ecosystem. The Journal of Animal Ecology, 58(3), 893-904.
Stroud J L, Irons D E, Carter J E, Watts C W, Murray P J, Norris S L y Whitmore A P. 2016. Lumbricus terrestris middens are biological and chemical hotspots in a minimum tillage arable ecosystem. Applied Soil Ecology, 105, 31-35.
Subler S y Kirsch A S. 1998. Spring dynamics of soil carbon, nitrogen, and microbial activity in earthworm middens in a no-till cornfield. Biology and Fertility of Soils, 26, 243-249.
Syers J K y Springett J A. 1984. Earthworms and soil fertility. Plant & Soil, 76, 93-104. Tabaglio V, Gavazzi C y Menta C. 2009. Physico-chemical indicators and microarthropod communities as
influenced by no-till, conventional tillage and nitrogen fertilisation after four years of continuous maize. Soil & Tillage research, 105(1), 135-142.
Ter Braak C F J. 1986. Canonical correspondence analysis: a new eigenvector technique for multivariate direct gradient analysis. Ecology, 67, 1167-1179.
Thibaud J M y Christian E. 1989. Collemboles interstitiels aériens des sables littoraux méditerranéens. Annales de la Société Entomologique de France, 25(1), 71-81.
Tian G, Brussaard L y Kang B T. 1993. Biological effects of plant residues with contrasting chemical compositions under humid tropical conditions: effects on soil fauna. Soil Biology & Biochemistry, 25(6), 731-737.
Trigo D, Mascato R, Mato S y Cosín D J D. 1988. Biogeographical divisions of continental Portugal as regards earthworm fauna. Bolletino di Zoologia, 55(1-4), 85-92.
van Straalen N M y Verhoef H A. 1997. The development of a bioindicator system for soil acidity based on arthropod pH preferences. Journal of Applied Ecology, 34, 217-232.
van Vliet P C J, West L T, Hendrix P F y Coleman D C. 1993. The influence of Enchytraeidae (Oligochaeta) on the soil porosity of small microcosms. Geoderma, 56, 287-299.
Verhoef H A y Brussaard L. 1990. Decomposition and nitrogen mineralization in natural and agro-ecosystems: the contribution of soil animals. Biogeochemistry, 11, 175–211.
Vreeken-Buijs M J, Hassink J y Brussaard L. 1998. Relationships of soil microarthropod biomass with organic matter and pore size distribution in soils under different land use. Soil Biology & Biochemistry, 30(1), 97-106.
Wardle D A, Bardgett R D, Klironomos J N, Setälä H, van Der Putten W H y Wall D H. 2004. Ecological linkages between aboveground and belowground biota. Science, 304(5677), 1629-1633.
Wardle D A. 1995. Impacts of disturbance on detritus food webs in agro-ecosystems of contrasting tillage and weed management practices. Advances in Ecological Research, 26, 105-185.
Wardle D A. 2000. Communities and Ecosystems. Princeton University Press, Princeton, New Jersey. Webb N R,Coulson S J, Hodkinsonz I D, Block W, Bale J S y Strathdee A T. 1998. The effects of experimental
temperature elevation on populations of cryptostigmatic mites in high arctic soils. Pedobiologia, 42, 298-308.
Whalley W R, Dumitru E y Dexter A R. 1995. Biological effects of soil compaction. Soil & Tillage Research, 35(1-2), 53-68.
Wood H B y James S W. 1993. Native and introduced earthworms from selected chaparral, woodland, and riparian zones in southern California. General Technical Report PSW, 142, 1-19.
Zanella A, Tomasi M, De Siena C, Frizzera L, Jabiol B, Nicolini G. 2001. Humus forestali: manuale di ecologia pe il i o os i e to e l i te p etazio e: appli azio e alle faggete. Viote di Monte Bondone (TN), Centro di Ecologia Alpina.
Zvereva E L y Kozlov M V. 2010. Responses of terrestrial arthropods to air pollution: a meta-analysis. Environmental Science and Pollution Research, 17, 297–311.

CAPÍTULO IV:
Variación en la composición isotópica y estabilidad de las comunidades edáficas en los agrosistemas
atlánticos y mediterráneos


Capítulo IV
153
1. INTRODUCCIÓN
1.1. El uso de isótopos estables en ecología terrestre
El uso de los isótopos estables está muy extendido en diversos campos de la ecología para
investigar la dieta y la posición que ocupan los organismos dentro de la cadena alimenticia
(Vander Zanden et al. 1999; Post et al. 2000; Post 2002). Estas técnicas presentan la ventaja de
que permiten identificar la fuente de alimento asimilada por los organismos a largo plazo en
contraposición con otros métodos basados en la descripción del contenido intestinal y que
meramente reflejan el alimento ingerido (DeNiro y Epstein 1981). Otra de las ventajas de
emplear técnicas de isótopos estables es que proporcionan información cuantitativa acerca de
la transmisión de energía o masa a través de los distintos niveles de las cadenas tróficas (Post
2002). Los principales elementos de interés para estudios isotópicos son el carbono (13C y 12C),
azufre (34S y 32S), nitrógeno (15N y 14N), oxígeno (18O y 16O) e hidrógeno (1H y 2H), pero son los de
carbono y de nitrógeno los que más se emplean en los estudios ecológicos centrados en los
ecosistemas terrestres.
Sin embargo, uno de los problemas en el uso de los isótopos estables es que reaccionan
químicamente y se metabolizan de forma diferente, lo que conduce a que sean incorporados en
los tejidos del consumidor en una proporción distinta a la que está presente en la fuente original.
Esto da luga a lo ue se o o e o o f a io a ie to isotópi o Ehleringer y Rundel 1988) y
varía de unos organismos a otros, en función de su tamaño, edad y estado fisiológico, con los
cambios estacionales de la dieta, así como a lo largo de la cadena trófica (Briones y Schmidt
2004). De forma general, los organismos discriminan en contra del isótopo más pesado y los
diferentes procesos metabólicos (respiración, digestión, formación de distintos tejidos,
excreción, etc.) dan lugar a un un ligero incremento en el valor isotópico del el animal con
respecto a su dieta (1-4‰; Ehleringer y Rundel 1988), que es lo que se conoce como
enriquecimiento isotópi o (Mestre et al. 2013). Por ello, se suele emplear la relación entre el
isótopo pesado y el ligero del mismo elemento (13C/12C o 15N/14N) como una estima de la
cantidad de nutriente asimilado por los distintos eslabones de la cadena trófica (Rounick y
Winterbourn 1986; Peterson y Fry 1987; France y Peters 1997; Post 2002).
Cuando la diferencia isotópica entre las posibles fuentes de alimento es amplia, es posible
identificar la dieta de un organismo. Por ejemplo, en el caso del carbono, la diferente
composicion isotópica que presentan las plantas con diferentes rutas fotosintéticas (C3 vs. C4)
permite determinar el origen del carbono asimilado por un organismo (Rounick y Winterbourn
1986; Peterson y Fry 1987; O Lea et al. . De fo a si ila , la relacion isotópica entre los
dos isótopos de nitrogeno se puede emplear también para identificar dietas, pero más

Capítulo IV
154
habitualmente para estimar el nivel que ocupa un organismo en la cadena trófica (Gannes et al.
1997; Post 2002). Dado que en cada nivel trófico se incrementa la proporcion de isótopo pesado
asimilado en torno al 3-4‰ DeNi o Epstei ; Mi aga a Wada 1984; Peterson y Fry
1987) es posible establecer si el individuo en cuestión es un herbívoro, carnívoro o consumidor
secundario (Mestre et al. 2013).
1.2. Composición isotópica de los organismos edáficos
El empleo de análisis isotópicos en agroecosistemas todavía no está muy extendido y si bien
existen algunos estudios que han abordado la estructura trófica en sistemas de producción
agrícola mediante estas técnicas (McNabb et al. 2001; Schmidt et al. 2004; Wise et al. 2006;
Mestre et al. 2013), muy pocas investigaciones se han centrado en cultivos frutícolas.
Sin embargo, los resultados de estos estudios han permitido profundizar en el conocimiento
de las preferencias alimenticias de los invertebrados que viven en los suelos cultivados. Así por
ejemplo, Schmidt et al. (2004) midieron la composición isotópica de C y N en varios grupos de
organismos edáficos (lombrices, enquitreidos y babosas) en cultivos de cebada y concluyeron
que cuando se consideran los isótopos de C y N por separado estos invertebrados forman un
continuo, pero si se combina la información de ambos se pueden diferenciar claramente los
herbívoros de los detritívoros.
Una gran parte de la bibliografía disponible se ha centrado en utilizar técnicas isotópicas para
conocer los hábitos alimenticios de las lombrices (Schmidt et al. 1997, 2004; Schmidt y Curry
1999; Briones et al. 1999a, 2001; Briones y Bol 2003; Dungait et al. 2008; Eck et al. 2015; Larsen
et al. 2016). Estos estudios han permitido concluir que las variaciones en la composición
isotópica de C de estos organismos son pequeñas cuando los cultivos contienen únicamente
plantas C3 (Schmidt et al. 1997); en cambio, los valores de δ15N permiten diferenciar varios
niveles tróficos en sus comunidades (Schmidt et al. 1997, 2004; Briones et al. 1999a, 2001 y
2003; Briones y Bol 2003; Dungait et al. 2008). Por ejemplo, se ha podido constatar que algunas
especies asimilan preferentemente materia orgánica lábil procedente de exudados radicales de
plantas en el caso de cultivos de guisante (Eck et al. 2015) y aminoácidos de origen vegetal y
bacteriano en praderas con manejo orgánico (Larsen et al. 2016) y que sus actividades
alimenticias pueden alterar la distribución del nitrógeno en los tejidos de las plantas en aquellos
sistemas agrícolas en los que se combina el cultivo de cereales y leguminosas (Schmidt y Curry
1999).
En el caso de los enquitreidos, apenas existe información sobre la posición trófica de las
diferentes especies y por tanto, tampoco se ha podido establecer una relación entre estrategias

Capítulo IV
155
alimenticias y la posición que ocupan a lo largo del perfil del suelo (Briones e Ineson 2002). Sin
embargo, analizando la composición isotópica de C de aminoácidos esenciales, Larsen et al.
(2016) encontraron que las poblaciones de enquitreidos que viven en pastos orgánicos obtienen
sus fuentes de proteínas tanto de las plantas como de bacterias. Además, los análisis de isotópos
de C y N de tres especies de enquitreidos (Fridericia galva, Fridericia christeri y Enchytraeus
buchholzi) indican que para el caso de las especies del género Fridericia son muy similares entre
sí (Schmidt et al. 2004), lo que podría sugerir que se alimentan de una dieta diferente a otros
géneros.
Entre la gran diversidad de taxones que integran el grupo de ácaros del suelo, los oribátidos
han recibido especial atención, pero la mayoría de los estudios isotópicos se han llevado a cabo
en sistemas forestales (Scheu y Falca 2000; Schneider et al. 2004; Erdmann et al. 2007; Heethoff
y Scheu 2016). Una de las principales conclusiones que se pueden extraer de estos trabajos es
que, contrariamente a la idea tradicional que asumía que los oribátidos presentan una
alimentación generalista con muy poca diferenciación trófica (Maraun et al. 1998; Scheu y Setälä
2002), el tipo de dieta tiene una gran influencia sobre los valores de δ13C de los oribátidos
(Heethoff y Scheu 2016). De hecho, en investigaciones realizadas en campos de cultivo de maíz,
Scheunemann et al. (2015) encontraron que la mayor parte de las especies de oribátidos
obtienen hasta un 26-32% del C de las raíces frente a un porcentaje menor de especies que lo
obtienen de los tallos (12%).
Otro grupo de organismos edáficos investigados mediante técnicas isotópicas son los
colémbolos, comúnmente considerados como descomponedores generalistas (Addison et al.
2003; Scheu y Simmerling 2004). Así, Potapov et al. (2016) constataron la existencia de un
gradiente isotópico condicionado por un gradiente vertical de humificación de la materia
orgánica, de tal forma que los valores de δ13C y δ15N son más bajos en los colémbolos
atmobiontes (asociados a macrófitas, troncos, arbustos y pastizales) que en los que habitan las
capas minerales del suelo. De forma similar, Chahartaghi et al. (2005) encontraron una gran
amplitud en la relación isotópica 15N/14N de 20 especies de colémbolos que van desde las
especies ficófagas/herbívoras, con los menores contenidos de N-15, hasta las especies
descomponedoras primarias y secundarias, con los valores más altos y, por tanto, podrían
incluirse en un rango de tres niveles tróficos. Sin embargo, las diferentes estrategias alimenticias
que exhiben los colémbolos también vienen condicionados por la disponibilidad y palatabilidad
de sus fuentes de alimento (Briones et al. 1999b; Sechi et al. 2014).
Debido a la importancia que tiene la abundancia y calidad de las dietas potenciales que
pueden ser utilizadas por los organismos edáficos, las técnicas isotópicas también pemiten
detectar si cambios en el uso del suelo pueden alterar el tipo y cantidad de un determinado

Capítulo IV
156
sustrato. Por ejemplo, se ha podido constatar que conversiones desde un pastizal a un
monocultivo de maíz provocan cambios en los hábitos alimenticios de varias especies de
lombrices de tierra (Briones et al. 1999a, 2001; Briones y Bol 2003). En este sentido, Crotty et al.
(2014) concluyeron que el tipo de vegetación funciona como el principal conductor del
funcionamiento trófico de la comunidad a todos los niveles y que los mismos taxones pueden
utilizar diferentes recursos en diferentes ecosistemas.
1.3. La cadena trófica edáfica como indicadora del grado de perturbación de los
agrosistemas
Desde hace tiempo se reconoce la importancia de las cadenas alimentarias para comprender
el funcionamiento de los ecosistemas (Wardle et al. 2004). En concreto, la cadena trófica
encargada de descomponer los aportes orgánicos es fundamental en los ecosistemas terrestres
debido a su papel clave en el ciclo de nutrientes y la producción primaria (De Angelis 1992).
La estructura y naturaleza de estas asociaciones son muy sensibles a las perturbaciones
ambientales, de tal forma que la intensificación del uso del suelo reduce la biomasa de todos los
componentes de la cadena trófica (de Vries et al. 2012). Esto es debido a que los diferentes
manejos en los sistemas agrícolas ejercen fuertes presiones selectivas sobre las comunidades
edáficas (Paustian et al. 1990). El uso de herbicidas, fertilizantes y la presencia o no de una
cubierta vegetal condicionan la supervivencia de muchos organismos del suelo (Menta 2012).
Por otro lado, la mayor o menor diversidad de las plantas cultivadas y su abundancia afectan al
número de hábitats y a la cantidad y calidad de los nutrientes disponibles (Maharning et al.
2008). El número de microhábitats depende de las propiedades físicas y químicas del sistema
(Bardgett y Cook 1998) y la disponibilidad de recursos de la palatabilidad del material vegetal
susceptible de ser descompuesto (Maharning et al. 2008).
Wardle (1995) observó una relación negativa entre intensificación agrícola y diversidad
edáfica en base a clases de tamaño corporal, lo cual puede tener importantes consecuencias
para la estabilidad del agrosistema. Esto es debido a que los organismos de mayor tamaño
suelen ocupar las posiciones más altas de la cadena trófica y tienen un mayor efecto
estabilizador; por tanto, si el manejo agrícola afecta con mayor intensidad a los organismos
edáficos con un determinado tamaño, la estabilidad del agrosistema se verá negativamente
afectada y consecuentemente, la estructura de la comunidad más que la diversidad per se es la
que determina la estabilidad de un ecosistema (Brussaard et al. 2004).
En vista de todo lo expuesto, parece claro que el uso de técnicas isotópicas permite obtener
información más detallada del uso de los diferentes sustratos disponibles por los diferentes
grupos de organismos así como su posición a lo largo de la cadena trófica. Dado que el tipo de

Capítulo IV
157
vegetación y el uso del suelo afectan al modo en que los organismos edáficos se alimentan e
interaccionan, se trata de técnicas muy sensibles que permiten conocer mejor los flujos de masa
y energía entre los distintos eslabones y por tanto, el funcionamiento de los ecosistemas.
Por todo ello, el objetivo de este estudio es utilizar técnicas isotópicas para conocer la
estructura trófica de los principales grupos faunísticos que dominan en los cultivos frutícolas de
dos regiones geográficas distintas con el fin de determinar si el clima, el tipo de cultivo y/o el de
manejo (convencional y orgánico) pueden alterar sus hábitos alimentarios. Además, dado que
los distintos grupos muestran distinta sensibilidad al grado de intensificación agrícola en función
de su tamaño es importante determinar si, además de alteraciones en las abundancias, existe
una relación entre perturbaciones debidas al manejo y la estabilidad del agrosistema.
2. MATERIALES Y MÉTODOS
2.1. Toma de muestras y procesado
Para los análisis isotópicos se emplearon ejemplares de macrofauna y los taxones más
abundantes de mesofauna (oribátidos, colémbolos y enquitreidos) recogidos desde mayo a junio
del año 2013, abarcando así la estación de la primavera con mayor actividad biológica. Los
métodos de recogida de los distintos grupos fauísticos son los mismos que los indicados en el
capítulo II, es decir, recolección manual de lombrices de tierra a partir de cuadriculas (50 x 50
cm) y extracciones de mesofauna a partir de muestras de suelo obtenidas mediante cilindros de
acero (10 cm de diámetro x 20 cm de altura). Pero, en este caso, los ejemplares recogidos no
fueron fijados, sino que tras ser identificados en vivo fueron congelados hasta su procesamiento
posterior. En el caso de lombrices previamente se seleccionaron ejemplares maduros de cada
especie (siempre que fuese posible) que fueron sacrificados y diseccionados para eliminar el
contenido del tubo digestivo (Briones et al. 1999b).
Posteriormente todos los ejemplares fueron liofilizados (TELSTAR CRYODOS-45) y el tejido
seco pulverizado manualmente en mortero. Aproximadamente 0,6 y 1,2 mg de muestra seca y
molida de cada taxon fue introducido en cápsulas de aluminio de 4 x 6 mm para ser analizadas
isótopicamente (δ13C y δ15N).
Los ejemplares de cada taxón recogidos en cada uno de los tres puntos aleatorios de cada
finca y muestreo tuvieron que ser agrupados para poder obtener una masa suficiente para
realizar los análisis isotópicos.

Capítulo IV
158
2.2. Análisis isotópicos
Las muestras secas de macrofauna y mesofauna fueron enviadas al James Hutton Institute
(Aberdeen, Escocia) para ser analizadas isotópicamente mediante un analizador elemental
(Flash EA 1112; Thermo Finnigan, Bremen, Alemania) conectado a un espectrofómetro de masas
(DeltaPlus XP; Thermo Finnigan, Bremen, Alemania).
Las o posi io es isotópi as se e p esa ‰, utiliza do la o e latu a δ, para indicar las
desviaciones de las abundancias naturales del material estándar (Peterson y Fry 1987):
δ13C ‰ =(RMuestra/REstándar) x 1000
δ15N ‰ =(RMuestra/REstándar) x 1000
dónde R=13C ⁄12C o 15N/14N
Como referencias convencionales de material estándar del δ13C y δ15N se emplearon Vienna
PeeDee Belemnite y el N2 atmosférico, respectivamente.
2.3. Índice V
El índice V (Wardle 1995) permite describir para evaluar la magnitud de respuesta de de un
organismo (o de un recurso basal) frente a un manejo agrícola intensivo (convencional). Se
calcula mediante la siguiente fórmula:
Índice V = ������+��� -1
dónde MCT es la abundancia media del taxón faunístico (ind. m-2) o el valor medio del sustrato
base (C, N) en el manejo convencional y MNT es la abundancia media de individuos o valor medio
del recurso basal en ausencia de perturbación agrícola (en este estudio el manejo orgánico). Los
valores el índice V varían entre -1 (inhibición extrema del manejo convencional) y 1
(estimulación máxima del manejo intensivo), mientras que un valor de 0 indica la ausencia de
efecto del manejo. Se han establecido hasta seis categorías para expresar el grado de respuesta
al manejo en función de los rangos de valores del índice V (Tabla 33; Wardle 1995):
Tabla 33: Categorías establecidas según los rangos de valores adoptados por el índice V (Wardle 1995).
Respuesta Rango de valores del índice
V Inhibición extrema V < -0,67
Inhibición moderada -0,33 > V > -0,67
Inhibición leve 0 > V > -0,33
Estimulación leve 0 < V < -0,33
Estimulación moderada 0,33 < V < 0,67
Estimulación extrema V > 0,67

Capítulo IV
159
2.4. Análisis estadísticos
Con el fin de comprobar la existencia de diferencias significativas en la composición isotópica
(δ15N y δ13C) de los grupos de organismos edáficos dominantes en los suelos de cultivo
investigados (oribátidos, colémbolos, enquitreidos y lombrices) se empleó un análisis de
PERMANOVA (ANOVA multivariante permutacional). Se emplearon matrices de similitud
basadas en las distancias Euclídeas sin transformación previa de los datos. Al igual que ANOVA,
PERMANOVA calcula un valor de la F (pseudo-F) de la suma de las distancias cuadradas entre los
grupos (Anderson et al. 2008). Como test a posteriori se empleó la distribución de la t, calculando
una pseudo-t a partir de la raíz cuadrada de la pseudo-F, pero la probabilidad fue calculada con
999 permutaciones.
En este análisis factorial se incluyeron como factores fijos región geográfica (atlántica y
mediterránea), tipo de cultivo (kiwi, vid, mandarino y olivo) y manejo agrícola (convencional y
orgánico), así como las interacciones entre cultivo y manejo.
Los análisis PERMANOVA fueron realizados con el programa estadístico PRIMER v.6
(Anderson et al. 2008).
3. RESULTADOS
3.1. Efecto de la región geográfica, tipo de cultivo y manejo sobre la composición isotópica de la mesofauna
3.1.1. Ácaros oribátidos
Los resultados del PERMANOVA de los contenidos en isótopos de C y N de los oribátidos
indican que la región geográfica no tuvo un efecto significativo sobre los valores de δ13C y δ15N,
lo que sugiere que este grupo de ácaros muestran preferencias alimenticias similares en ambas
regiones (Fig. 67A).
Tampoco el tipo de cultivo agrícola tuvo una influencia significativa sobre el contenido
isotópico de los oribátidos, observándose un gran solapamiento en los valores δ13C y δ15N (Fig.
67B). Sin embargo, merece la pena destacar ciertas diferencias en la posición trófica de los
oribátidos recolectados en los cultivos de kiwi y mandarino frente a los de vid y olivo
(δ15N=9,3± , ‰ , ± , ‰, espe ti a e te; Fig. B . Ade ás, fue posi le o stata ie tas
diferencias en cuanto al contenido isotópico de carbono entre los oribátidos de los suelos de
mandarino y los de kiwi, con una mayor preferencia de los primeros por los sustratos
enriquecidos en el isótopo más pesado (Fig. 67B).
En el caso del manejo agrícola, si bien las diferencias tampoco alcanzaron la significación
estadística, se puede apreciar que las poblaciones de oribátidos que viven en suelos orgánicos

Capítulo IV
160
tienden a utilizar sustratos enriquecidos en N-15 respecto a los de los suelos convencionales
(con una diferencia de 3 unidades delta; Fig. 67C).
Figura 67: Valores medios (±E.E.) de δ15N y δ13C en los oribátidos capturados en (A) las dos regiones geográficas investigadas, (B) en los cuatro cultivos y (C) en los dos manejos agrícolas.
En cambio, la interacción entre el tipo de cultivo y el manejo sí tuvo un efecto significativo
sobre la composición isotópica de C y N de los oribátidos (Pseudo-F=3,54; p=0,01). A pesar de la
gran variabilidad en los valores de δ13C, se observaron amplias diferencias entre las poblaciones
que habitan los cultivos sometidos a manejo orgánico, de tal modo que las presentes en los
suelos de mandarino y vid prefieren fuentes de carbono enriquecidas con C-13 en comparación
a las que viven en los suelos plantados con kiwi y olivo. La posición trófica de estos ácaros
también muestra importantes variaciones con el tipo de cultivo y manejo, destacando aquellos
ejemplares recolectados en los cultivos de kiwi y mandarino orgánico frente a los de olivo
orgánico (con una dife e ia de ≈ ‰; Fig. . E el aso de los ulti os so etidos a a ejo
convencional, aunque se observa una tendencia a incorporar el isótopo más pesado de N desde
los cultivos atlánticos (vid y kiwi) frente a los mediterraneos (mandarino y olivo), las diferencias
son muy pequeñas (Fig. 68).
0
2
4
6
8
10
12
-26,0 -25,5 -25,0 -24,5 -24,0
d15N
(0/ 0
0)
d13C (0/00)
A
AtlánticoMediterráneo
0
2
4
6
8
10
12
-26,5 -26,0 -25,5 -25,0 -24,5 -24,0 -23,5 -23,0
d15N
(0/ 0
0)
d13C (0/00)
KiwiVidMandarinoOlivo
B
0
2
4
6
8
10
12
14
-26,0 -25,5 -25,0 -24,5 -24,0
d15N
(0/ 0
0)
d13C (0/00)
ConvencionalOrgánico
C
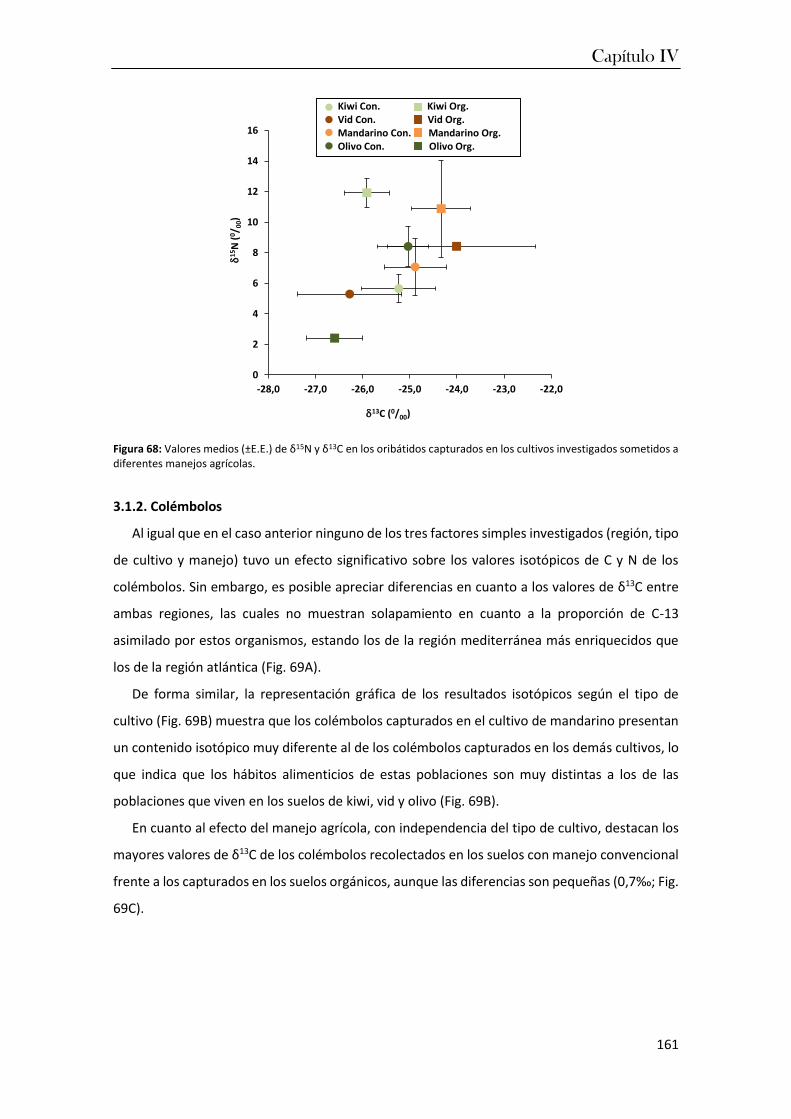
Capítulo IV
161
Figura 68: Valores medios (±E.E.) de δ15N y δ13C en los oribátidos capturados en los cultivos investigados sometidos a diferentes manejos agrícolas.
3.1.2. Colémbolos
Al igual que en el caso anterior ninguno de los tres factores simples investigados (región, tipo
de cultivo y manejo) tuvo un efecto significativo sobre los valores isotópicos de C y N de los
colémbolos. Sin embargo, es posible apreciar diferencias en cuanto a los valores de δ13C entre
ambas regiones, las cuales no muestran solapamiento en cuanto a la proporción de C-13
asimilado por estos organismos, estando los de la región mediterránea más enriquecidos que
los de la región atlántica (Fig. 69A).
De forma similar, la representación gráfica de los resultados isotópicos según el tipo de
cultivo (Fig. 69B) muestra que los colémbolos capturados en el cultivo de mandarino presentan
un contenido isotópico muy diferente al de los colémbolos capturados en los demás cultivos, lo
que indica que los hábitos alimenticios de estas poblaciones son muy distintas a los de las
poblaciones que viven en los suelos de kiwi, vid y olivo (Fig. 69B).
En cuanto al efecto del manejo agrícola, con independencia del tipo de cultivo, destacan los
mayores valores de δ13C de los colémbolos recolectados en los suelos con manejo convencional
f e te a los aptu ados e los suelos o gá i os, au ue las dife e ias so pe ueñas , ‰; Fig.
69C).
0
2
4
6
8
10
12
14
16
-28,0 -27,0 -26,0 -25,0 -24,0 -23,0 -22,0
d15N
(0/ 0
0)
d13C (0/00)
Kiwi Con. Kiwi Org.Vid Con. Vid Org.Mandarino Con. Mandarino Org.Olivo Con. Olivo Org.

Capítulo IV
162
Figura 69: Valores medios (±E.E.) de δ15N y δ13C en los colémbolos capturados en (A) las dos regiones geográficas investigadas, (B) en los cuatro cultivos y (C) en los dos manejos agrícolas.
En cuanto a la interacción tipo de cultivo*manejo, los resultados isotópicos muestran
diferencias significativas (Pseudo-F=4,99; p=0,002), destacando el mandarino convencional por
presentar los valores más altos de ambos isótopos (Fig. 70). Así, para el caso del carbono, a
excepción del olivar, en el resto de cultivos el manejo convencional induce a que los colémbolos
se alimenten preferentemente de sustratos con una mayor proporción de C-13 (Fig. 70). En
cuanto a los valores de δ15N, si bien no se obtuvo suficiente muestra de tejido para poder
analizar el contenido isotópico de los colémbolos capturados en el mandarino orgánico, destaca
la diferencia en la posición trófica que ocupan estos organismos en el kiwi orgánico comparado
al observado en el kiwi convencional, con una mayor enriquecimiento en N-15 de los primeros
(Fig. 70). Aunque en menor medida, esta misma situación se observa en el caso del viñedo, pero
se invierte en el caso del olivar, mostrando las poblaciones de los cultivos convencionales una
mayor cantidad de isótopo pesado asimilado en sus tejidos (Fig. 70).
0
1
2
3
4
5
6
7
8
9
10
-27,0 -26,5 -26,0 -25,5 -25,0
d15N
(0/ 0
0)
d13C (0/00)
AtlánticoMediterráneo
A
0
1
2
3
4
5
6
7
8
9
-27,0 -26,5 -26,0 -25,5 -25,0
d15N
(0/ 0
0)
d13C (0/00)
ConvencionalOrgánico
C
0
2
4
6
8
10
12
14
-27,5 -27,0 -26,5 -26,0 -25,5 -25,0 -24,5 -24,0
d15N
(0/ 0
0)
d13C (0/00)
B
KiwiVidMandarinoOlivo

Capítulo IV
163
Figura 70: Valores medios (±E.E.) de δ15N y δ13C en los colémbolos capturados en los cultivos investigados sometidos a diferentes manejos agrícolas.
3.1.3. Enquitreidos
A diferencia de los microartrópodos, la región geográfica tuvo un efecto significativo sobre
la composición isotópica de los enquitreidos (Pseudo-F=4,5; p=0,01). Se pudo constatar que si
bien los enquitreidos capturados en las dos regiones estudiadas utilizan fuentes de carbono
similares (Fig. 71A), ocupan posiciones tróficas diferentes, siendo los valores de δ15N de los
e uit eidos de la egió edite á ea as altos ue los de los e uit eidos atlá ti os , ‰
de diferencia; Fig. 71A).
En cambio, no se observaron diferencias significativas en el contenido isotópico de los
enquitreidos en relación al tipo de cultivo. Sin embargo, al igual que ocurría con los colémbolos,
las poblaciones de enquitreidos en los suelos de cultivo de mandarino presentan los valores más
altos de δ13C y δ15N que en el resto de cultivos (Fig. 71B). Los contenidos isotópicos de C y N mas
bajos fueron para los enquitreidos en los suelos del viñedo (-25,1± ‰ pa a δ13C y 6,8± , ‰
para δ15N; Fig. 71B).
Tampoco se detectaron diferencias significativas en la composición isotópica de los
enquitreidos que viven en los suelos de cultivo sometidos a distinto manejo, aunque los
enquitreidos capturados en los cultivos orgánicos presentaron valores de δ13C y δ15N más altos
que los presentes en los convencionales (Fig. 71C).
0
2
4
6
8
10
12
14
-28,0 -27,0 -26,0 -25,0 -24,0 -23,0 -22,0
d15N
(0/ 0
0)
d13C (0/00)
Kiwi Con. Kiwi Org.Vid Con. Vid Org.Mandarino Con. Olivo Con. Olivo Org.
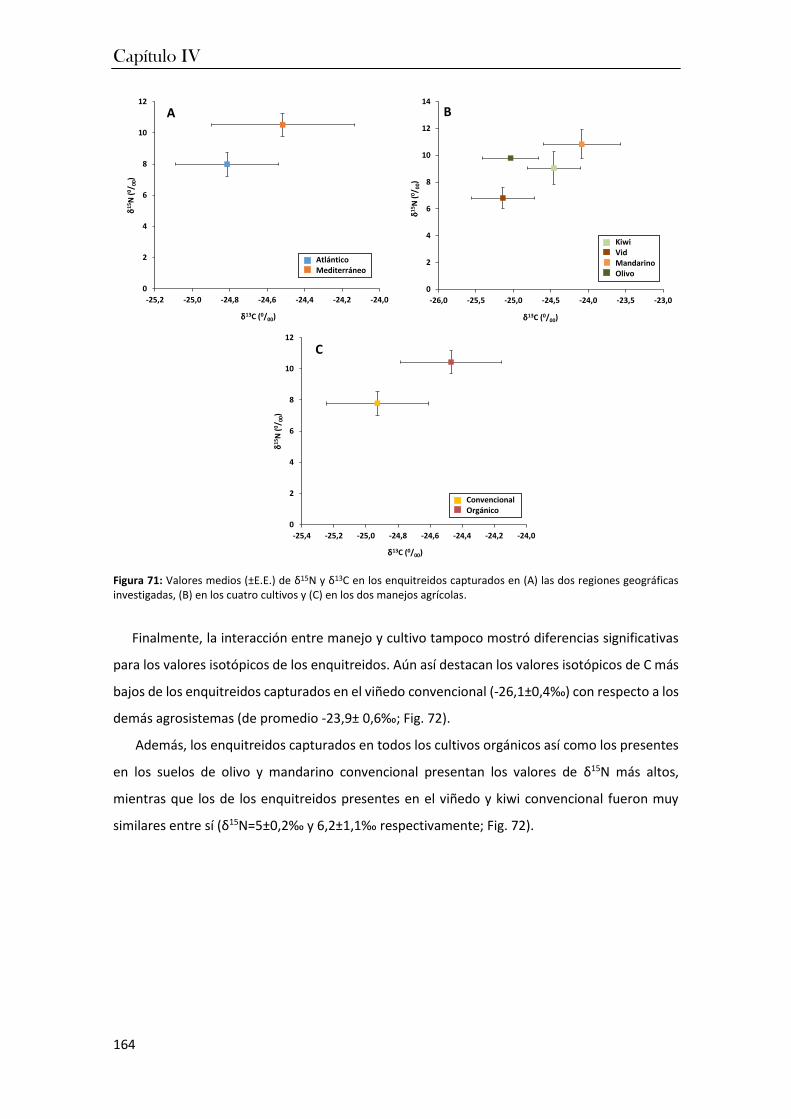
Capítulo IV
164
Figura 71: Valores medios (±E.E.) de δ15N y δ13C en los enquitreidos capturados en (A) las dos regiones geográficas investigadas, (B) en los cuatro cultivos y (C) en los dos manejos agrícolas.
Finalmente, la interacción entre manejo y cultivo tampoco mostró diferencias significativas
para los valores isotópicos de los enquitreidos. Aún así destacan los valores isotópicos de C más
bajos de los enquitreidos capturados en el viñedo convencional (-26,1± , ‰ con respecto a los
demás agrosistemas (de promedio -23,9± , ‰; Fig. 72).
Además, los enquitreidos capturados en todos los cultivos orgánicos así como los presentes
en los suelos de olivo y mandarino convencional presentan los valores de δ15N más altos,
mientras que los de los enquitreidos presentes en el viñedo y kiwi convencional fueron muy
similares entre sí (δ15N=5± , ‰ y 6,2± , ‰ respectivamente; Fig. 72).
0
2
4
6
8
10
12
-25,2 -25,0 -24,8 -24,6 -24,4 -24,2 -24,0
d15N
(0/ 0
0)
d13C (0/00)
AtlánticoMediterráneo
A
0
2
4
6
8
10
12
-25,4 -25,2 -25,0 -24,8 -24,6 -24,4 -24,2 -24,0
d15N
(0/ 0
0)
d13C (0/00)
ConvencionalOrgánico
C
0
2
4
6
8
10
12
14
-26,0 -25,5 -25,0 -24,5 -24,0 -23,5 -23,0
d15N
(0/ 0
0)
d13C (0/00)
B
KiwiVidMandarinoOlivo

Capítulo IV
165
Figura 72: Valores medios (±E.E.) de δ15N y δ13C en los enquitreidos capturados en los cultivos investigados sometidos a diferentes manejos agrícolas.
3.2. Efecto de la región geográfica, tipo de cultivo y manejo sobre la composición isotópica de la macrofauna
No se detectaron diferencias significativas en cuanto al contenido isotópico de las lombrices
para el factor región. Aunque los valores de δ13C de los individuos de ambas regiones muestran
una gran variabilidad (Fig. 73A), las lombrices de la región mediterránea presentaron un mayor
enriquecimiento de N-15 (δ15N=10,2± , ‰ ue las atlá ti as δ15N=7,9± , ‰; Fig. A .
Tampoco se observaron diferencias significativas en la composición isotópica de las
lombrices en función del tipo de cultivo agrícola, lo cual fue debido a la gran variabilidad
encontrada en los valores de δ13C y δ15N (Fig. 73B). Sin embargo, se puede apreciar un cierto
gradiente desde las lombrices recogidas en el viñedo, con los valores más bajos de ambos
isótopos, hasta los más altos medidos en las lombrices capturadas en los suelos de cultivo de
mandarino (con una diferencia de 4 unidades delta en el caso del 15N; Fig. 73B). En cambio, las
poblaciones de lombrices presentes en los cultivos de olivo y kiwi presentaron valores
intermedios de ambos isótopos (Fig. 73B).
No obstante sí se detectaron diferencias significativas en los contenidos isotópicos según el
manejo agrícola (Pseudo-F=3,7; p=0,03). Al igual que ocurría con los enquitreidos, los valores de
δ13C más altos fueron registrados en las lombrices presentes en los suelos con manejo orgánico
(-22,9± , ‰) frente a los medidos en las lombrices de los suelos convencionales (-23,4±0,1; Fig.
73C). Además, las lombrices de los suelos orgánicos presentaron también un mayor
0
2
4
6
8
10
12
14
-27,0 -26,5 -26,0 -25,5 -25,0 -24,5 -24,0 -23,5 -23,0 -22,5
d15N
(0/ 0
0)
d13C (0/00)
Kiwi Con. Kiwi Org.Vid Con. Vid Org.Mandarino Con. Mandarino Org.Olivo Con. Olivo Org.

Capítulo IV
166
enriquecimiento en el isótopo pesado de N en comparación con los convencionales
(δ15N=10,1±0,5 y 7,7±0,6‰, respectivamente; Fig. 73C).
Figura 73: Valores medios (±E.E.) de δ15N y δ13C en las lombrices de tierra capturados en (A) las dos regiones geográficas investigadas, (B) en los cuatro cultivos y (C) en los dos manejos agrícolas.
También se observaron diferencias significativas en la composición isotópica de las lombrices
de tierra en relación a la interacción entre tipo de cultivo y manejo (Pseudo-F=3,5; p=0,004). En
cuanto a δ13C, se aprecia la diferencia entre las lombrices del olivo orgánico y las de kiwi y viñedo
convencionales que presentaron los valores más bajos (<- , ‰ las lo i es de los esta tes
cultivos (Fig. 74). Por otro lado, los alo es de δ15N permiten separar por un lado a las lombrices
presentes en los cultivos de mandarino sometidos a los dos tipos de manejo, junto con las del
olivo convencional y kiwi orgánico por presentar los mayores contenidos de N-15 (δ15N=10-
‰ f e te a los edidos e las lombrices capturadas en los viñedos con ambos tipos de
manejo agrícola, el kiwi convencional y el olivo orgánico por otro, con los menores valores de
este isótopo δ15N e to o a ‰; Fig. 74).
0
2
4
6
8
10
12
-23,6 -23,4 -23,2 -23,0 -22,8
d15N
(0/ 0
0)
d13C (0/00)
AtlánticoMediterráneo
A
0
2
4
6
8
10
12
-23,8 -23,6 -23,4 -23,2 -23,0 -22,8 -22,6
d15N
(0/ 0
0)
d13C (0/00)
ConvencionalOrgánico
C
0
2
4
6
8
10
12
14
-23,9 -23,7 -23,5 -23,3 -23,1 -22,9 -22,7 -22,5
d15N
(0/ 0
0)
d13C (0/00)
B
KiwiVidMandarinoOlivo

Capítulo IV
167
Figura 74: Valores medios (±E.E.) de δ15N y δ13C en las lombrices capturados en los cultivos investigados sometidos a diferentes manejos agrícolas.
3.3. Efecto del tipo de cultivo y manejo sobre la estructura trófica de la comunidad en base a clases de tamaño corporal
3.3.1. Cultivos de kiwi
El cálculo del índice V para los datos obtenidos en los cultivos de kiwi muestra una
estimulación leve del manejo convencional sobre la mesofauna, así como un ligero incremento
del contenido de C y N del suelo (Fig. 75). La abundancia de oribátidos fue la que mayor
estimulación experimentó, seguidos de los colémbolos, mientras que los enquitreidos fueron los
menos afectados por el manejo convencional en estos cultivos (Fig. 75).
Sin embargo, la macrofauna sufrió una inhibición moderada-alta de sus abundancias en el
manejo convencional de kiwi (Fig. 75).
0
2
4
6
8
10
12
14
-25,0 -24,5 -24,0 -23,5 -23,0 -22,5 -22,0 -21,5
d15N
(0/ 0
0)
d13C (0/00)
Kiwi Con. Kiwi Org.Vid Con. Vid Org.Mandarino Con. Mandarino Org.Olivo Con. OlivoOrg.
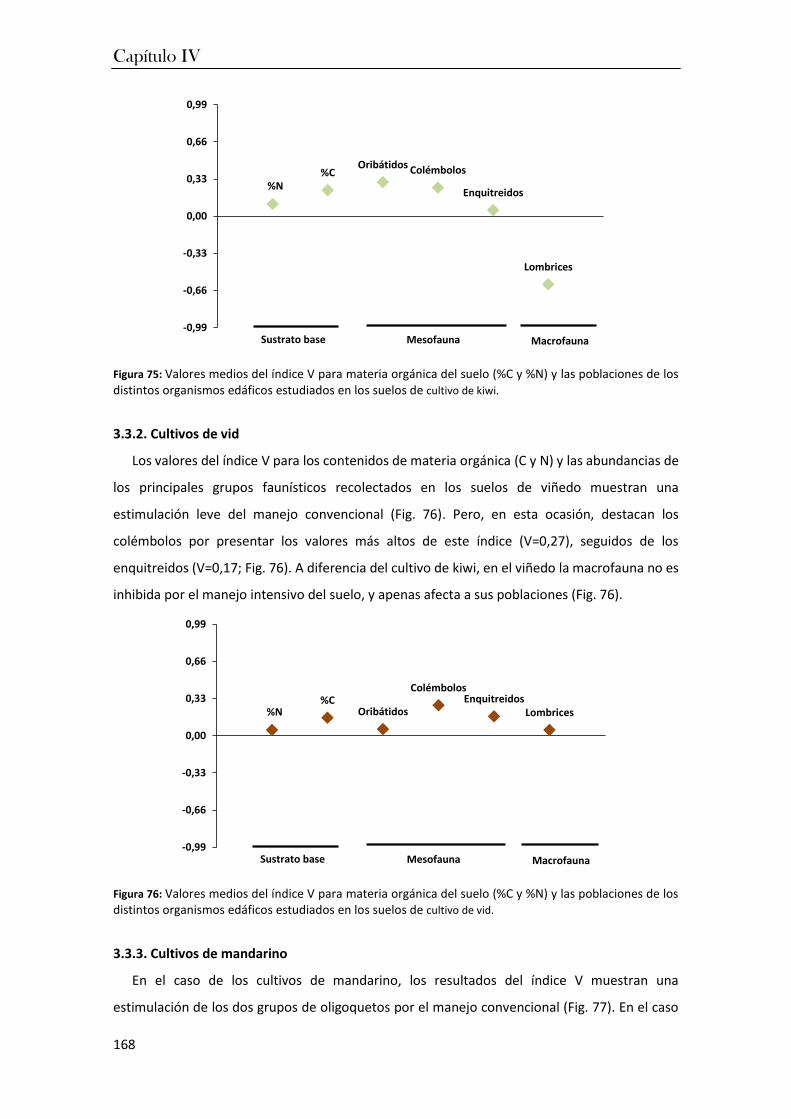
Capítulo IV
168
Figura 75: Valores medios del índice V para materia orgánica del suelo (%C y %N) y las poblaciones de los distintos organismos edáficos estudiados en los suelos de cultivo de kiwi.
3.3.2. Cultivos de vid
Los valores del índice V para los contenidos de materia orgánica (C y N) y las abundancias de
los principales grupos faunísticos recolectados en los suelos de viñedo muestran una
estimulación leve del manejo convencional (Fig. 76). Pero, en esta ocasión, destacan los
colémbolos por presentar los valores más altos de este índice (V=0,27), seguidos de los
enquitreidos (V=0,17; Fig. 76). A diferencia del cultivo de kiwi, en el viñedo la macrofauna no es
inhibida por el manejo intensivo del suelo, y apenas afecta a sus poblaciones (Fig. 76).
Figura 76: Valores medios del índice V para materia orgánica del suelo (%C y %N) y las poblaciones de los distintos organismos edáficos estudiados en los suelos de cultivo de vid.
3.3.3. Cultivos de mandarino
En el caso de los cultivos de mandarino, los resultados del índice V muestran una
estimulación de los dos grupos de oligoquetos por el manejo convencional (Fig. 77). En el caso
%N%C
Oribátidos Colémbolos
Enquitreidos
Lombrices
-0,99
-0,66
-0,33
0,00
0,33
0,66
0,99
Sustrato base Mesofauna Macrofauna
%N%C
Oribátidos
ColémbolosEnquitreidos
Lombrices
-0,99
-0,66
-0,33
0,00
0,33
0,66
0,99
Sustrato base Mesofauna Macrofauna
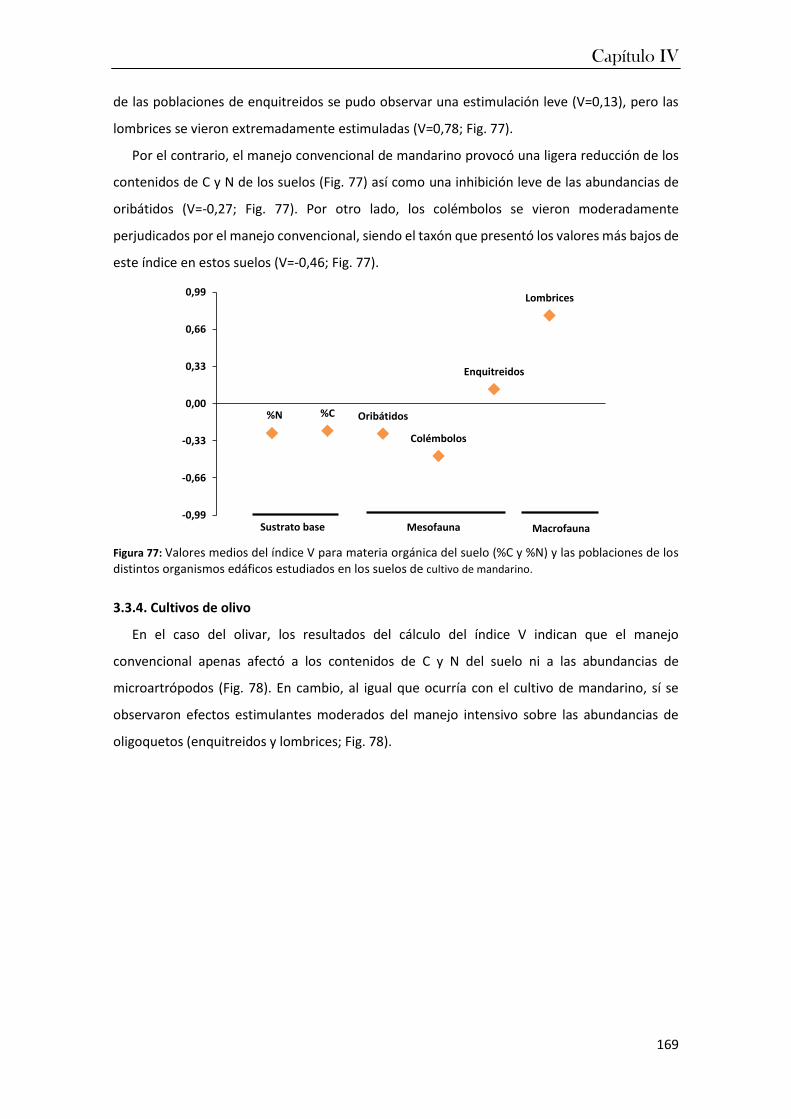
Capítulo IV
169
de las poblaciones de enquitreidos se pudo observar una estimulación leve (V=0,13), pero las
lombrices se vieron extremadamente estimuladas (V=0,78; Fig. 77).
Por el contrario, el manejo convencional de mandarino provocó una ligera reducción de los
contenidos de C y N de los suelos (Fig. 77) así como una inhibición leve de las abundancias de
oribátidos (V=-0,27; Fig. 77). Por otro lado, los colémbolos se vieron moderadamente
perjudicados por el manejo convencional, siendo el taxón que presentó los valores más bajos de
este índice en estos suelos (V=-0,46; Fig. 77).
Figura 77: Valores medios del índice V para materia orgánica del suelo (%C y %N) y las poblaciones de los distintos organismos edáficos estudiados en los suelos de cultivo de mandarino.
3.3.4. Cultivos de olivo
En el caso del olivar, los resultados del cálculo del índice V indican que el manejo
convencional apenas afectó a los contenidos de C y N del suelo ni a las abundancias de
microartrópodos (Fig. 78). En cambio, al igual que ocurría con el cultivo de mandarino, sí se
observaron efectos estimulantes moderados del manejo intensivo sobre las abundancias de
oligoquetos (enquitreidos y lombrices; Fig. 78).
%N %C Oribátidos
Colémbolos
Enquitreidos
Lombrices
-0,99
-0,66
-0,33
0,00
0,33
0,66
0,99
Sustrato base Mesofauna Macrofauna

Capítulo IV
170
Figura 78: Valores medios del índice V para materia orgánica del suelo (%C y %N) y las poblaciones de los distintos organismos edáficos estudiados en los suelos de cultivo de olivo.
4. DISCUSIÓN
4.1. Composición isotópica de las comunidades edáficas en los agrosistemas atlánticos y
mediterráneos
El manejo de los cultivos frutícolas con plantas leñosas, al no ser una actividad extractiva
completa (sólo se extraen los frutos), no implica arado del suelo ni la eliminación de toda la
planta cultivada en el periodo de cosecha; sin embargo, otras actividades comunes en este tipo
de sistemas como son el uso de fertilizantes, herbicidas y el destino de los residuos de la poda y
cosecha tienen una influencia directa sobre la supervivencia y la disponibilidad de alimento para
los organismos edáficos. Por otro lado, las características climáticas debidas a condicionantes
geográficos, también determina el tipo de planta a cultivar así como la implementación de
diversas prácticas para garantizar su crecimiento (e.g. sistemas de riego), las cuales también
tienen un efecto directo sobre las actividades de la fauna edáfica. Por todo ello, el manejo
agrícola condiciona no sólo el tamaño de sus poblaciones sino también la competencia por los
recursos alimenticios, y en definitiva la estabilidad del agrosistema y su funcionamiento.
La variación composición isotópica de los organismos detritívoros investigados indica que las
condiciones climáticas de la región mediterránea dieron como resultado comunidades edáficas
con un contenido más alto de 15N que los observados en aquellas que habitan los suelos de
cultivo de la región atlántica. Las mayores temperaturas que reinan en esta región favorecen,
por un lado, a aquellos organismos eudáficos y hemiedáficos (Potapov et al. 2016) que
desarrollan sus ciclos vitales dentro del suelo (dónde las condiciones de humedad y temperatura
son más favorables) y no en la superficie (como son los colémbolos Symphypleona; Anexo I), así
como aquellos que pueden buscar refugio en los horizontes mas profundos del suelo donde la
%N
%C
Oribátidos
Colémbolos
EnquitreidosLombrices
-0,99
-0,66
-0,33
0,00
0,33
0,66
0,99
Sustrato base Mesofauna Macrofauna

Capítulo IV
171
materia orgánica está más humificada (enquitreidos y lombrices). Además, se sabe que
temperaturas cálidas aceleran los procesos de descomposición lo que suele llevar asociado un
incremento en el contenido de 15N de la materia orgánica (Tiunov 2007). La única excepción a
este patrón fueron los oribátidos, lo cual puede ser debido a la enorme variedad de estrategias
alimenticias que exhibe este grupo, ocupando diferentes nichos ecológicos (Schneider et al.
2004).
El tipo de cultivo también tuvo un efecto significativo sobre la utilización de los recursos
alimentarios disponibles por parte de los organismos del suelo, lo que corrobora resultados
previos que indican que el tipo de vegetación funciona como el principal conductor del
funcionamiento trófico de la comunidad edáfica (Schmidt et al. 1997; Briones et al. 2001; Crotty
et al. 2014). De forma general, la comunidad edáfica presente en el cultivo de mandarino es la
que presenta los valores isotópicos, tanto en δ13C como en δ15N, más altos y fue especialmente
evidente en el caso de los colémbolos, enquitreidos y lombrices. Esto parece indicar que estos
tres taxones en este cultivo se comportan como consumidores secundarios, posiblemente
predando microorganismos saprófagos (Potapov et al. 2013). Temperaturas más cálidas y un pH
del suelo menos ácido junto con el uso de fertilizantes orgánicos y un riego frecuente
constituyen condiciones que favorecen el aumento de la actividad microbiana (Fig. 23; capítulo
I) y de las abundancias de colémbolos (Fig. 50B; capítulo II) y oligoquetos, sobre todo lombrices
(Fig. 50D; capítulo II), en los cultivos de mandarino. De hecho, las lombrices constituyeron el
taxón con los mayores enriquecimientos en 13C y 15N en en este cultivo, posiblemente debido a
la dominancia de lombrices anécicas (Fig. 38; capítulo II). Estudios previos (Briones y Schmidt
2004) indican que dado que las lombrices anécicas construyen galerías verticales pueden
alimentarse de la materia orgánica más humificada presente en horizontes más profundos del
suelo, lo que explicaría los mayores valores isotópicos medidos en los tejidos de estos
organismos.
Los resultados isotópicos obtenidos para los restantes cultivos indican que la comunidad
edáfica estaría constituida fundamentalmente por consumidores primarios en que el material
vegetal forma una parte importante de su dieta (Potapov et al. 2013). De todos ellos, los cultivos
de vid destacan por presentar los valores isotópicos más bajos. Al ser un cultivo de la región
atlántica, las temperaturas más frescas y los pH de suelo más ácidos hacen que la actividad
microbiológica está disminuida (Fig. 22; capítulo I) así como la abundancia de meso y
macrofauna (Fig. 50; capítulo II), con lo que los procesos de descomposición se ralentizan.
Por último, merece la pena destacar que, en el caso de los oribátidos, los valores de δ15N más
altos en los cultivos de mandarino y el kiwi podrían indicar la presencia de especies carnívoras o
carroñeras. “i e a go, los alo es egist ados > ‰ po té i o edio dista u ho de los

Capítulo IV
172
descritos para estos grupos tróficos en otros estudios, dó de el alo de δ15N no suele superar
las 6 unidades delta (Scheu y Falca 2000; Schneider et al. 2004; Maraun et al. 2011). En este
sentido, Mestre et al. (2013) registraron valores de δ15N similares en arañas (en torno a los 10-
‰ ue fue at i uido a dep eda ió i t ag e ial, po lo ual es posible que las comunidades
de ácaros de los suelos de estos dos cultivos incluyesen ácaros que se alimenten de otros
oribátidos carnívoros o incluso de enquitreidos. Sin embargo, es difícil conocer las causas
específicas del enriquecimiento en isótopos de N en los oribátidos debido a las dificultades
existentes a la hora de determinar la firma isotópica de los recursos basales (Maraun et al. 2011).
En función del manejo agrícola también se pudo observar que tanto los oribátidos como los
enquitreidos se alimentan de fuentes de C similares en ambos tipos de sistemas, convencional
y orgánico, mientras que los colémbolos y las lombrices utilizan fuentes diferentes y asimilaron
una mayor cantidad de 13C en el sistema convencional y en el orgánico, respectivamente. Estos
resultados indican un cambio en los hábitos alimenticios de estos dos taxones como
consecuencia de un manejo agrícola distinto. El mayor enriquecimiento isotópico de los
colémbolos del sistema convencional sugiere quese trata de especies euedáficas que se
alimentan de microorganismos (Chahartaghi et al. 2005; Ferlian et al. 2014a), mientras que en
el manejo orgánico, donde no se emplean herbicidas, su dieta consistiría sobre todo en material
vegetal. En el caso de las lombrices de tierra, la diferente composición isotópica podría ser
debida al mayor número de lombrices endogeas recolectadas en el sistema orgánico frente el
convencional (36,4±5,82 y 8,04±2,03 ind. m-2, respectivamente), las cuales se caracterizan por
consumir sustratos con un mayor contenido de 13C que el resto de categorías ecológicas
(Schmidt et al. 1997; Briones y Schmidt 2004).
En cambio, para el caso del N, la mayoría de los taxones edáficos presentaron los valores más
altos de δ15N en los suelos orgánicos en comparación con los convencionales. La única excepción
fueron los colémbolos, que ocuparon la misma posición trófica en ambos manejos. La ausencia
de uso de productos herbicidas en el manejo orgánico permite el crecimiento de comunidades
vegetales y una mayor acumulación de materia orgánica que proporciona condiciones de
temperatura, humedad y cantidad de alimento más favorables para las comunidades edáficas
(Edwards y Pimentel 1989; Moreby et al. 1994) que fueron más diversas (Fig. 52; capítulo III)
permitiendo el establecimiento de cadenas tróficas más complejas.
En cuanto a la interacción cultivo y manejo, se observó una gran variabilidad en los valores
isotópicos. En general, oribátidos, enquitreidos y lombrices muestran un gran solapamiento en
la asimilación de fuentes de C; en cambio, los colémbolos que habitan los cultivos orgánicos de
kiwi, olivo y vid así como el convencional de vid parecen basar sus dietas en recursos poco
enriquecidas en 13C por lo que éstas consistirían fundamentalmente de material vegetal,

Capítulo IV
173
mientras que en los sistemas convencionales de kiwi, mandarino y olivo sus comunidades
estarían constituidas por especies descomponedoras secundarias, basando su alimentación en
microorganismos que se alimentan de exudados radicales, experimentando así un
enriquecimiento de C en sus tejidos (Ferlian et al. 2014b).
En el caso del d15N, destacan los mayores valores registrados en todos los taxones edáficos
presentes en la finca de kiwi orgánico comparado con los observados en la finca convencional
de este cultivo y que sugiere de nuevo una mayor complejidad de las relaciones tróficas entre
los diferentes taxones de estos organismos en los suelos con manejo orgánico. De hecho, las
lombrices fueron más abundantes en la finca orgánica, en especial las endogeas y anécicas (Fig.
41; capítulo II), que presentan el mayor enriquecimiento de 15N (Briones y Schmidt 2004). Los
enquitreidos también fueron más abundantes en los suelos con el manejo menos intensivo lo
que coincide con resultados previos (Lago 2015). En cambio, los ácaros y los colémbolos fueron
más abundantes en el sistema convencional lo que indica que, como se indicó anteriormente,
en este sistema sobreviven las especies más edáficas que basan su alimentación en sustratos
más humificados (Ferlian et al. 2014a)
4.2. Estabilidad de los agrosistemas atlánticos y mediterráeos frente al manejo agrícola
Los resultados del índice V indican la existencia de importantes diferencias en la resiliencia
de los distintos taxones edáficos frente a la intensificación del sistema productivo. En el caso de
los cultivos atlánticos (kiwi y vid), las temperaturas más bajas y los suelos más húmedos y ácidos
favorecen la acumulación de materia orgánica en el suelo. Pero, es en los cultivos
convencionales de esta región dónde se registran los mayores valores de humedad (Fig. 11A;
capítulo I), %C (Fig. 16A; capítulo I) y biomasa microbiana (0,20±0,01 frente a 0,16±0,01 mg de
C g de suelo seco-1 en los manejos convencional y orgánico, respectivamente). Los colémbolos
constituyen el taxón que se ve más estimulado por el manejo convencional de los dos cultivos
atlánticos debido el incremento en los recursos basales (Wardle et al. 2001) y por una mayor
dominancia de especies eudáficas (tal como se indicó anteriormente en base a su mayor
enriquecimiento en 13C) a expensas de las epiedáficas, ya que son más resistentes a la pérdida
de la cubierta vegetal protectora causada por las aplicaciones de herbicidas (Alvarez et al. 2001;
Kautz et al. 2006) y por tanto, se beneficiarían de esa abundante biomasa microbiana.
En cambio, los otros dos grupos de mesofauna, aunque también muestran respuestas
positivas al manejo convencional, los oribátidos se ven especialmente favorecidos por un
manejo más intensivo en los cultivos de kiwi y los enquitreidos en los de vid. Dado que estos dos
cultivos fueron muy similares en cuanto relación C/N y contenido de biomasa microbiana, las
diferentes respuestas pueden atribuirse al tiempo que llevan las fincas dedicadas al cultivo

Capítulo IV
174
convencional (30 años en la kiwi frente a 20 del de vid) y a las prácticas de encalado más
frecuentes en las fincas de kiwi que perjudican a los enquitreidos (Abrahamsen 1983). En
cambio, la mayor tolerancia de los oribátidos a la desecación (Block 1996; Lindberg y Bengtsson
2006) les habría aportado una ventaja competitiva en los cultivos con mayor retirada de la
cubierta vegetal, como ocurre en estos suelos de kiwi convencional. De hecho estudios previos
en las mismas fincas de kiwi (Lago 2015) dieron como resultado una fuerte inhibición de los
enquitreidos frente a una estimulación de los oribátidos en estos suelos convencionales.
Coincidiendo también con resultados previos (Lago 2015) las lombrices se vieron claramente
desfavorecidas por la intensificación del cultivo de kiwi, lo que confirma la mayor sensibilidad
de los organismos de mayor tamaño (macrofauna) a las prácticas agrícolas más agresivas
(Wardle 1995) y resalta su valor como indicadoras de perturbaciones ambientales (Römbke et
al. 2005; Paoletti et al. 2013; Falco et al. 2015).
Todos estos resultados indican que la producción intensiva de kiwi en la región atlántica tiene
importantes consecuencias negativas para la estabilidad de estos agrosistemas al verse
reducidas drásticamente las abundancias de oligoquetos, en especial de lombrices, cuya mayor
biomasa se traduce en efectos a mayor escala de los procesos edáficos, como son la reducción
de la capacidad de aireación e infiltración de agua en el suelo, alteraciones en las comunidades
microbiológicas y en consecuencia, reducción de la productividad agrícola (Lee 1985; Wardle
1995; Buckerfield et al. 1997; Lavelle et al. 1997; Paoletti et al. 2013).
Una situación muy diferente se observa en los cultivos de la región mediterránea, donde la
producción de mandarino y olivo empleando un manejo convencional tiende a promover las
poblaciones de oligoquetos frente a las de microartrópodos, lo que se traduce en una mayor
estabilidad de estos agrosistemas. La reducción en los recursos basales posiblemente afectó
negativamente a la mesofauna (Wardle et al. 2001). En cambio, las poblaciones de lombrices
resultan claramente estimuladas en ambos cultivos, pero sobre todo en el de mandarino
convencional. La mayor actividad de estas especies aceleran los procesos de descomposición de
tal forma que la materia orgánica depositada es rápidamente mineralizada y explicaría la mayor
reducción del contenido de C en este agrosistema de producción intensiva en esta región
(Lavelle et al. 1997; Nielsen et al. 2011; Brito-Vega et al. 2013).
5. CONCLUSIONES
En contraposición con la idea generalizada de que las comunidades edáficas están
constituidas fundamentalmente por organismos saprófagos y generalistas, la composición
isotópica de los taxones edáficos recolectados en los cultivos de ambas regiones y sometidos a
distintos manejos indican que existe una fuerte diferenciación trófica. Así, las condiciones

Capítulo IV
175
climáticas más cálidas de la región mediterránea favorecen a los organismos excavadores y
euedáficos que consumen materia orgánica más humificada. En esta región, las condiciones
edafoquímicas de los suelos de cultivo de mandarino permiten el establecimiento comunidades
tróficas más diversas, lo que resalta la importancia de las interacciones planta-suelo para
comprender mejor el funcionamiento de los ecosistemas edáficos.
Aunque de forma general, los manejos orgánicos favorecen cadenas tróficas más complejas
que incluyen consumidores primarios y secundarios, el efecto de un manejo agrícola
convencional sobre la estructura trófica de las comunidades edáficas puede ser positivo o
negativo dependiendo del tipo de planta cultivada que determina la calidad de los aportes
orgánicos y de las prácticas aplicadas que modifican las condiciones de temperatura y humedad
del suelo (e.g. sistemas de riego, existencia de una cubierta vegetal protectora entre árboles
frutales, etc.). Aquellas prácticas que mejoren las condiciones edafo-climáticas permitirán que
incluso los organismos más sensibles a las prácticas convencionales, que suelen ser de mayor
tamaño y cuyas actividades tienen efectos sobre los procesos edáficos de mayor magnitud,
puedan sobrevivir e incluso incrementar sus tamaños poblacionales. Una comunidad edáfica
más compleja, con un mayor número y diversidad de eslabones tróficos, garantiza una mayor
estabilidad del agrosistema y por tanto, representa un indicador fiable del grado de perturbación
de un suelo cultivado que podría ser utilizado en el desarrollo de estrategias de manejo más
sostenibles.
6. BIBLIOGRAFÍA
Anderson M J, Gorley R N y Clarke K R, 2008. PERMANOVA+ for PRIMER. Guide to Software and Statistical Methods. PRIMER–E, Plymouth.
Alvarez T, Frampton G K y Goulson D. 2001. Epigeic Collembola in winter wheat under organic, integrated and conventional farm management regimes. Agriculture, Ecosystems & Environment, 83(1), 95-110.
Abrahamsen G. 1983. Effects of lime and artificial acid rain on the enchytraeid (Oligochaeta) fauna in coniferous forest. Ecography, 6(3), 247–254.
Addison J A, Trofymow J A y Marshall V G. 2003. Functional role of Collembola in successional coastal temperate forests on Vancouver Island. Canadian Journal of Applied Soil Ecology, 24, 247-261.
Bardgett R D y Cook R (1998) Functional aspects of soil animal diversity in agricultural grasslands. Applied Soil Ecology, 10, 263-276.
Block W. 1996. Cold or drought - the lesser of two evils for terrestrial arthropods? European Journal of Entomology, 93, 325-339.
Briones M J I y Ineson P. 2002. Use of 14C carbon dating to determine feeding behaviour of enchytraeids. Soil Biology & Biochemistry, 34(6), 881-884.
Briones M J I y Bol R. 2003. Natural abundance of C-13 and N-15 in earthworms from different cropping treatments. Pedobiologia, 47, 560–567.
Briones M J I y Schmidt O. 2004. Stable isotope techniques in studies of the ecological diversity and functions of earthworms. Recent Research Developments in Crop Science, 1, 11-26.
Briones M J I, Bol R, Sleep D y Sampedro L. 1999a. Isotopic ecology of earthworms under grassland and arable cropping systems. Pedobiologia, 43, 675– 683.
Briones M J I, Ineson P y Sleep D. 1999b. Use of δ13C to determine food selection in collembolan species. Soil Biology & Biochemistry, 31(6), 937-940.

Capítulo IV
176
Briones M J I, Bol R, Sleep D, Allen D y Sampedro L. 2001. Spatio-temporal variation of stable isotope ratios in earthworms under grassland and maize cropping systems. Soil Biology & Biochemistry, 33(12), 1673-1682.
Brito-Vega H, Espinoza-Victoria D, Salaya-Domínguez J M y Gómez-Méndez E. 2013. The soil biota: importance in agroforestry and agricultural systems. Tropical & Subtropical Agroecosystems, 16(3), 445-453.
Buckerfield J C, Lee K E, Davoren C W y Hannay J N. 1997. Earthworms as indicators of sustainable production in dryland cropping in southern Australia. Soil Biology & Biochemistry, 29(3-4), 547-554.
Brussaard L, Kuyper T W, Didden W A M, Goede R G M, de Bloem J. 2004. Biological soil quality from biomass to biodiversity - importance and resilience to management stress and disturbance. En: Schjønning P, Elmholt S, Christensen B T (Eds.) Managing Soil Quality: Challenges in Modern Agriculture, pp 139-162. CABI Publishing, Wallingford.
Chahartaghi M, Langel R, Scheu S y Ruess L. 2005. Feeding guilds in Collembola based on nitrogen stable isotope ratios. Soil Biology & Biochemistry, 37(9), 1718-1725.
Crotty F V, Blackshaw R P, Adl S M, Inger R y Murray P J. 2014. Divergence of feeding channels within the soil food web determined by ecosystem type. Ecology & Evolution, 4(1), 1-13.
Curry J P y Schmidt O. 2007. The feeding ecology of earthworms - a review. Pedobiologia, 50, 463-477. DeNiro M J y Epstein S. 1981. Influence of diet on the distribution of nitrogen isotopes in animals.
Geochimica et Cosmochimica Acta, 45, 341-351. De Angelis D L. 1992. Dynamics of Nutrient Cycling and Food Webs. Chapman & Hall, London. de Vries F T, Liiri M E, Bjørnlund L, Bowker M A, Christensen S, Setälä H M y Bardgett R D. 2012. Land use
alters the resistance and resilience of soil food webs to drought. Nature Climate Change, 2(4), 276-280.
Dungait J A, Briones M J I, Bol R y Evershed R. 2008. Enhancing the understanding of earthworm feeding behaviour via the use of fatty acid delta C-13 values determined by gas chromatography-combustion-isotope ratio mass spectrometry. Rapid Communications in Mass Spectrometry, 22(11), 1643-1652.
Eck T, Potthoff M, Dyckmans J, Wichern F y Joergensen R G. 2015. Priming effects of Aporrectodea caliginosa on young rhizodeposits and old soil organic matter following wheat straw addition. European Journal of Soil Biology, 70, 38-45.
Edwards C A y Pimentel D. 1989. Impact of herbicides on soil ecosystems. Critical Reviews in Plant Sciences, 8(3), 221-257.
Ehleringer J R y Rundel P W. 1988. Stable isotopes: history, units and instrumentation. En: Rundel P W, Ehleringer J R y Nagy K A (Eds.) Stable Isotopes in Ecological Research, pp 2-15. Springer-Verlag, New York.
Erdmann G, Otte V, Langel R, Scheu S y Maraun M. 2007. The trophic structure of bark-living oribatid mite communities analysed with stable isotopes (15N, 13C) indicates strong niche differentiation. Experimental & Applied Acarology, 41(1-2), 1-10.
Falco L B, Sandler R, Momo F, Di Ciocco C, Saravia L y Coviella C. 2015. Earthworm assemblages in different intensity of agricultural uses and their relation to edaphic variables. PeerJ, 3, e979.
Ferlian O, Klarner B, Langeneckert A E y Scheu S. 2014a. Trophic niche differentiation and utilisation of food resources in collembolans based on complementary analyses of fatty acids and stable isotopes. Soil Biology & Biochemistry, 82, 28-35.
Ferlian O, Cesarz S, Marhan S y Scheu S. 2014b. Carbon food resources of earthworms of different ecological groups as indicated by 13C compound-specific stable isotope analysis. Soil Biology & Biochemistry, 77, 22-30.
France R L y Peters R H. 1997. Ecosystem differences in the trophic enrichment of 13C in aquatic food webs. Canadian Journal of Fisheries & Aquatic Sciences, 54, 1255-1258.
Gannes L Z, Obrien D M y del Rio C M. 1997. Stable isotopes in animal ecology: Assumptions, caveats, and a call for more laboratory experiments. Ecology, 78, 1271-1276.
Heethoff M y “ heu “. . ‘elia ilit of isotopi f a tio atio Δ15N, Δ13C) for the delimitation of trophic le els of o i atid ites: Diet st o gl affe ts Δ13C ut ot Δ15N. Soil Biology & Biochemistry, 101, 124-129.
Kautz T, López-Fando C y Ellmer F. 2006. Abundance and biodiversity of soil microarthropods as influenced by different types of organic manure in a long-term field experiment in Central Spain. Applied Soil Ecology, 33(3), 278-285.

Capítulo IV
177
Lago C F. 2015. Efecto del manejo agrícola sobre las propiedades biológicas y químicas del suelo y los procesos de transformación de la materia orgánica en cultivos de kiwi (Actinidia deliciosa). (Tesis Doctoral). Universidad de Vigo, Vigo.
Larsen T, Pollierer M M, Holmstrup M, D'Annibale A, Maraldo K, Andersen N y Eriksen J. 2016. Substantial nutritional contribution of bacterial amino acids to earthworms and enchytraeids: A case study from organic grasslands. Soil Biology & Biochemistry, 99, 21-27.
Lavelle P, Bignell D, Lepage M, Wolters V, Roger P, Ineson P, Heal O W, Dhillion S. 1997. Soil function in a changing world: the role of invertebrate ecosystem engineers. European Journal of Soil Biology, 33(4), 159-193.
Lee K E. 1985. Earthworms: their ecology and relationships with soils and land use. Academic Press, New York.
Lindberg N y Bengtsson J. 2006. Recovery of forest soil fauna diversity and composition after repeated summer droughts. Oikos, 3, 494-506.
Maharning A R, Mills A A y Adl S M. 2008. Soil community changes during secondary succession to naturalized grasslands. Applied Soil Ecology, 41, 137-147.
Maraun M, Migge S, Schaefer M, Scheu S. 1998. Selection of microfungal food by six oribatid mite species (Oribatida, Acari) from two different beech forests. Pedobiologia, 42, 232-240.
Maraun M, Erdmann G, Fischer B M, Pollierer M M, Norton R A, Schneider K y Scheu S. 2011. Stable isotopes revisited: their use and limits for oribatid mite trophic ecology. Soil Biology & Biochemistry, 43(5), 877-882.
McNabb D M, Halaj J y Wise D H. 2001. Inferring trophic positions of generalist predators and their linkage to the detrital food web in agroecosystems: a stable isotope analysis. Pedobiologia, 45(4), 289-297.
Menta C. 2012. Soil fauna diversity - function, soil degradation, biological indices, soil restoration. En: Gbolagade Akeem Lameed (Ed.) Biodiversity Conservation & Utilization in a Diverse World, pp. 59-94. InTech, DOI: 10.5772/51091.
Mestre L, Piñol J, Barrientos J A, Espadaler X, Brewitt K, Werner C y Platner C. 2013. Trophic structure of the spider community of a Mediterranean citrus grove: a stable isotope analysis. Basic & Applied Ecology, 14(5), 413-422.
Minagawa M y Wada E. 1984. Stepwise enrichment of 15N along food chains: further evidence and the relation between 15N and animal age. Geochimica et Cosmochimica Acta, 48, 1135–1140.
Moreby S J, Aebischer N J, Southway S E y Sotherton N W. 1994. A comparison of the flora and arthropod fauna oforganically grown winter wheat in Southern England. Annals of Applied Biology, 125, 13-27.
Nielsen U N, Ayres E, Wall D H y Bardgett R D. 2011. Soil biodiversity and carbon cycling: a review and synthesis of studies examining diversity–function relationships. European Journal of Soil Science, 62(1), 105-116.
O Lea M H, Madha a “ Pa eth P. . Physical and chemical basis of carbon isotope fractionation in plants. Plant, Cell & Environment, 15, 1099-1104.
Paoletti M G, Sommaggio D y Fusaro S. 2013. Proposta di indice di qualità biologica del suolo (QBS-e) basato sui lombrichi e applicato agli agroecosistemi. Biologia Ambientale, 27(2), 25-43.
Paustian K, Andren O, Clarholm M, Hansson A-C, Johansson G, Lagerlof J, Lindberg T, Pettersson R y Sohlenius B. 1990. Carbon and nitrogen budgets of four agro-ecosystems with annual and perennial crops, with and without N fertilization. Journal of Applied Ecology, 27(1), 60-84.
Peterson B J y Fry B. 1987. Stable isotopes in ecosystem studies. Annual Review of Ecology & Systematics, 18, 293–320.
Post D M, Pace M L y Hairston N G. 2000. Ecosystem size determines food-chain length in lakes. Nature, 405, 1047-1049.
Post D M. 2002. Using stable isotopes to estimate trophic position: models, methods, and assumptions. Ecology, 83(3), 703-718.
Potapov A M y Tiunov A V. 2016. Stable isotope composition of mycophagous collembolans versus mycotrophic plants: do soil invertebrates feed on mycorrhizal fungi? Soil Biology & Biochemistry, 93, 115-118.
Potapov A M, Semenina E E, Kurakov A V y Tiunov A V. 2013. Large 13C/12C and small 15N/14N isotope fractionation in an experimental detrital foodweb (litter–fungi–collembolans). Ecological Research, 28(6), 1069-1079.
Potapov A M, Semenina E E, Korotkevich A Y, Kuznetsova N A y Tiunov A V. 2016. Connecting taxonomy and ecology: Trophic niches of collembolans as related to taxonomic identity and life forms. Soil Biology & Biochemistry, 101, 20-31.

Capítulo IV
178
Römbke J, Jänsch S y Didden W. 2005. The use of earthworms in ecological soil classification and assessment concepts. Ecotoxicology & Environmental Safety, 62, 249-265.
Rounick J S y Winterbourn M J. 1986. Stable carbon isotopes and carbon flow in ecosystems. BioScience, 36, 171-177.
Scheu S y Falca M. 2000. The soil food web of two beech forests (Fagus sylvatica) of contrasting humus type: stable isotope analysis of a macro-and a mesofauna-dominated community. Oecologia, 123(2), 285-296.
Scheu S y Setälä H. 2002. Multitrophic interactions in decomposer foodwebs. En: Tscharntke T y Hawkins B A (Eds.) Multitrophic Level Interactions, pp 233-264. Cambridge University Press, Cambridge.
Scheu S y Simmerling F. 2004. Growth and reproduction of fungal feeding Collembola as affected by fungal species, melanin and mixed diets. Oecologia, 139, 347-353.
Scheunemann N, Digel C, Scheu S, Butenschoen O. 2015. Roots rather than shoot residues drive soil arthropod communities of arable fields. Oecologia, 179(4), 1135-1145.
Schmidt O y Curry J P. 1999. Effects of earthworms on biomass production, nitrogen allocation and nitrogen transfer in wheat-clover intercropping model systems. Plant & Soil, 214 (1-2), 187-198.
Schmidt O, Scrimgeour C M y Handley L L. 1997. Natural abundance of N-15 and C-13 in earthworms from a wheat and a wheat-clover field. Soil Biology & Biochemistry, 29(9-10), 1301-1308.
Schmidt O, Curry J P, Dyckmans J, Rota E y “ i geou C M. . Dual sta le isotope a al sis δ13C and δ15N) of soil invertebrates and their food sources. Pedobiologia, 48(2), 171-180.
Schneider K, Migge S, Norton R A, Scheu S, Langel R, Reineking A y Maraun M. 2004. Trophic niche differentiation in soil microarthropods (Oribatida, Acari): evidence from stable isotope ratios (15N/14N). Soil Biology & Biochemistry, 36(11), 1769-1774.
Sechi V, D'Annibale A, Ambus P, Sárossy Z, Krogh P H, Eriksen J y Holmstrup M. 2014. Collembola feeding habits and niche specialization in agricultural grasslands of different composition. Soil Biology & Biochemistry, 74, 31-38.
Tiunov A. 2007. Stable isotopes of C and N in soil ecological studies. Biology Bulletin, 34(4), 395-407. Wardle D A. 1995. Impacts of disturbance on detritus food webs in agro-ecosystems of contrasting tillage
and weed management practices. Advances in Ecological Research, 26, 105-185. Wardle D A, Yeates G W, Bonnera K I, Nicholson K S, Watson R N. 2001. Impacts of ground vegetation
management strategies in a kiwifruit orchard on the composition and functioning of the soil biota. Soil Biology & Biochemistry, 33(7-8), 893-905.
Wardle D A, Bardgett R D, Klironomos J N, Setälä H, van Der Putten W H y Wall D H. 2004. Ecological linkages between aboveground and belowground biota. Science, 304(5677), 1629-1633.
Wise D H, Moldenhauer D M y Halaj J. 2006. Using stable isotopes to reveal shifts in prey consumption by generalist predators. Ecological Applications, 16(3), 865-876.
Vander Zanden M J, Shuter B J, Lester N y J. B. Rasmussen J B. 1999. Patterns of food chain length in lakes: a stable isotope study. American Naturalist, 154, 406-416.

CONCLUSIONES GENERALES


Conclusiones
181
CONCLUSIONES
1) Las prácticas agrícolas modifican significativamente el ambiente físico, de tal forma que los
suelos donde se cultivan frutos con mayor requerimiento hídrico (kiwi y mandarino) son más
húmedos y los sometidos a manejos convencionales más húmedos que los orgánicos.
2) De las siete variables edáficas investigadas, la humedad y el pH son las que mejor caracterizan
a los suelos cultivados: unos suelos más húmedos y más ácidos limitan las actividades de las
poblaciones microbianas, ralentizan los procesos de descomposición y favorecen la retención
de nutrientes en el suelo (C y N).
3) De los tres factores estudiados (región, tipo de cultivo y manejo) sólo el cultivo tuvo un efecto
significativo sobre las densidades de organismos edáficos: el cultivo de mandarino fue el que
concentró las mayores abundancias y presentó las comunidades tróficas más diversas.
4) Los taxones de mesofauna fueron más abundantes que la macrofauna en los suelos agrícolas.
Mientras en la región atlántica dominan los enquitreidos, en los suelos mediterráneos fueron
los ácaros, con mayor tolerancia a la desecación.
5) La macrofauna fue muy sensible a las prácticas agrícolas convencionales, viéndose reducida
su diversidad y alterada su estructura funcional. Además, en los cultivos de cítricos más
perturbados sus poblaciones dedican más energía hacia la supervivencia que hacia el
crecimiento individual.
6) Los sistemas de manejo orgánico favorecen una mayor retención de C en el suelo que los
convencionales, por lo que estos últimos requerirán mayores aportes orgánicos para compensar
las pérdidas de fertilidad. Sin embargo, la magnitud de esta respuesta dependerá de las reservas
iniciales y cuánto tiempo lleva sometido el suelo a un manejo convencional.
7) Los manejos orgánicos favorecen el establecimiento de cadenas tróficas más complejas. Sin
embargo, un manejo convencional podría favorecer la supervivencia de la macrofauna si las
prácticas agrícolas aplicadas mantienen unas condiciones de temperatura y humedad de suelo
favorables.

Conclusiones
182
8) Una comunidad edáfica con mayor complejidad trófica y abundancia de macrofauna garantiza
una mayor homeostasis del agrosistema y podría ser un indicador fiable para el desarrollo de
estrategias de manejo más sostenibles.
En vista de todo ello, se puede concluir que las comunidades edáficas son altamente sensibles
a los cambios ambientales, tanto a las condiciones climáticas como los resultantes de las
prácticas agrícolas, viéndose alteradas tanto sus abundancias como la estructura y
composición de sus comunidades. Si bien los manejos orgánicos son más beneficiosos para los
organismos, la magnitud de los efectos negativos de los manejos convencionales pueden verse
contrarrestados mediante el mantenimiento de unas condiciones ambientales favorables para
la supervivencia de las especies que viven en la superficie del suelo (riegos regulares y una
cubierta vegetal que evite el calentamiento y la evaporación excesiva en las capas más
superficiales y además, aporte alimento a las comunidades descomponedoras).

ANEXOS

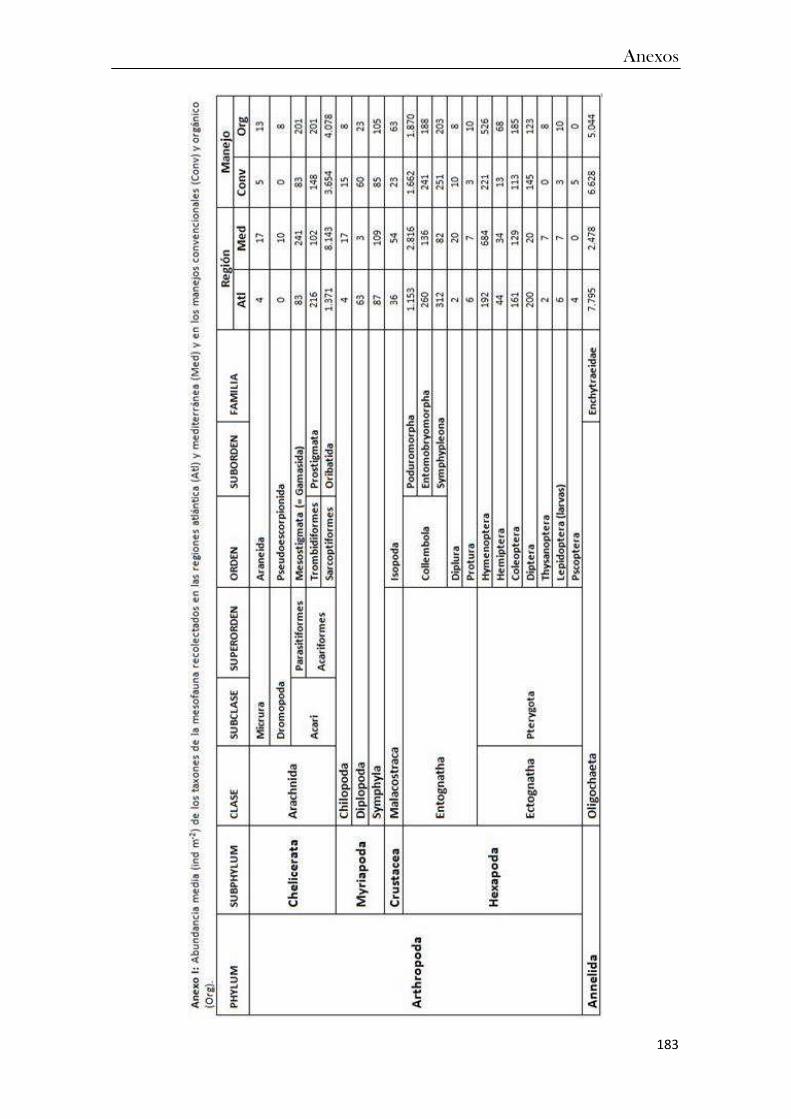
Anexos
183

Anexos
184

Anexos
185

Anexos
186

Anexos
187


















