Origen y evolución temprana de las aves · dientes, y el dedo oponible en el pie, que le permite...
173
Origen y evolución temprana de las aves
Transcript of Origen y evolución temprana de las aves · dientes, y el dedo oponible en el pie, que le permite...

Origen y evolución temprana
de las aves

Las aves representan una de las
especializaciones mas notables
de vertebrados en el vuelo. Se
destacan por sus huesos
livianos, múltiples elementos
esqueléticos fusionados,
reducción de la fíbula, gran
desarrolllo del cinturón pectoral
en relación al aleteo, brazos
transformados en alas, la
presencia de plumas, reducción
de la cola, pérdida de los
dientes, y el dedo oponible en el
pie, que le permite aferrarse a
las ramas de los árboles.
El comprensión de cómo se
adquirieron todos estos rasgos
fue un procesos que comenzó
con el descubrimiento de
Archaeopteryx en el siglo XIX y
que ha aumentado notablemente
en las últimas décadas

Poco después de que el hecho de
la evolución ganara aceptación, los científicos se abalanzaron
sobre el registro fósil en busca de los representantes de los
ancestros de las formas actuales.
La evolución implicaba claramente que las aves habían
descendido de los reptiles, lo cual se delataba en sus pies
escamosos
Debido a esto el descubrimiento
del fósil de Archaeopteryx lithographica fue un
acontecimiento de gran
importancia, ya que esta criatura emplumada presentaba con
claridad dientes en sus mandíbulas, garras en sus manos
y una larga cola vertebral,
generando el vínculo esperado entre aves y reptiles.
Archaeopteryx
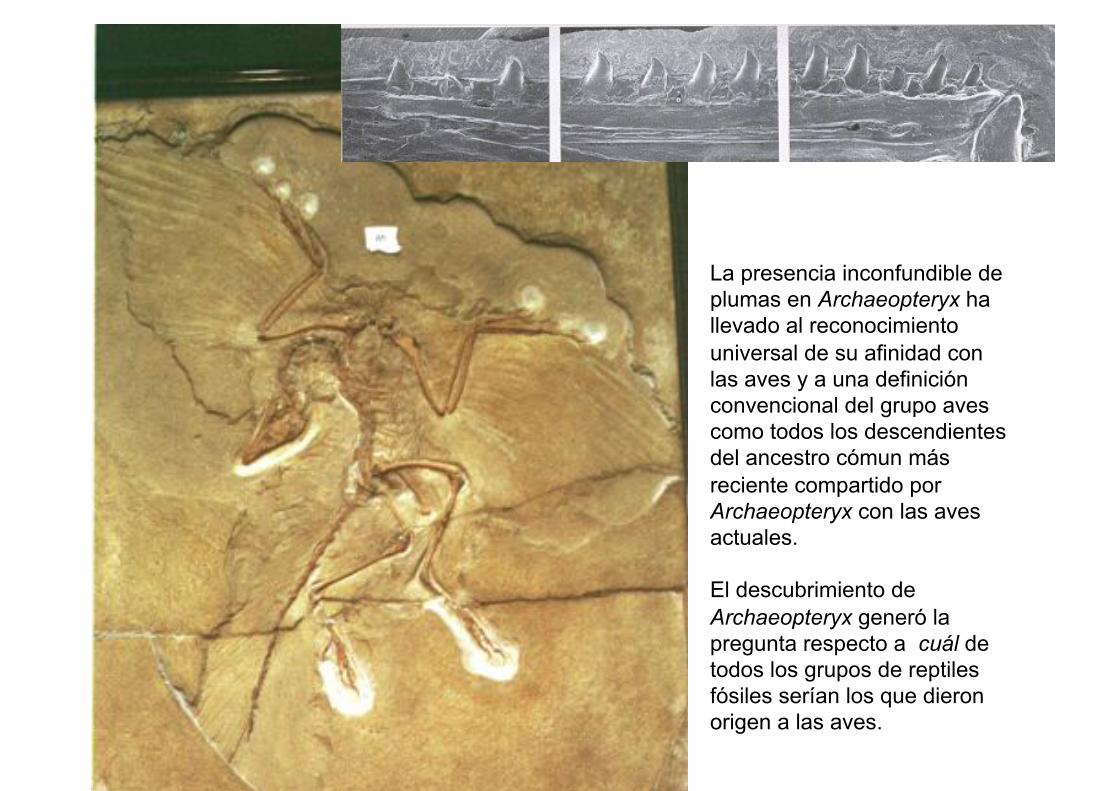
La presencia inconfundible de
plumas en Archaeopteryx ha
llevado al reconocimiento
universal de su afinidad con
las aves y a una definición
convencional del grupo aves
como todos los descendientes
del ancestro cómun más
reciente compartido por
Archaeopteryx con las aves
actuales.
El descubrimiento de
Archaeopteryx generó la
pregunta respecto a cuál de
todos los grupos de reptiles
fósiles serían los que dieron
origen a las aves.
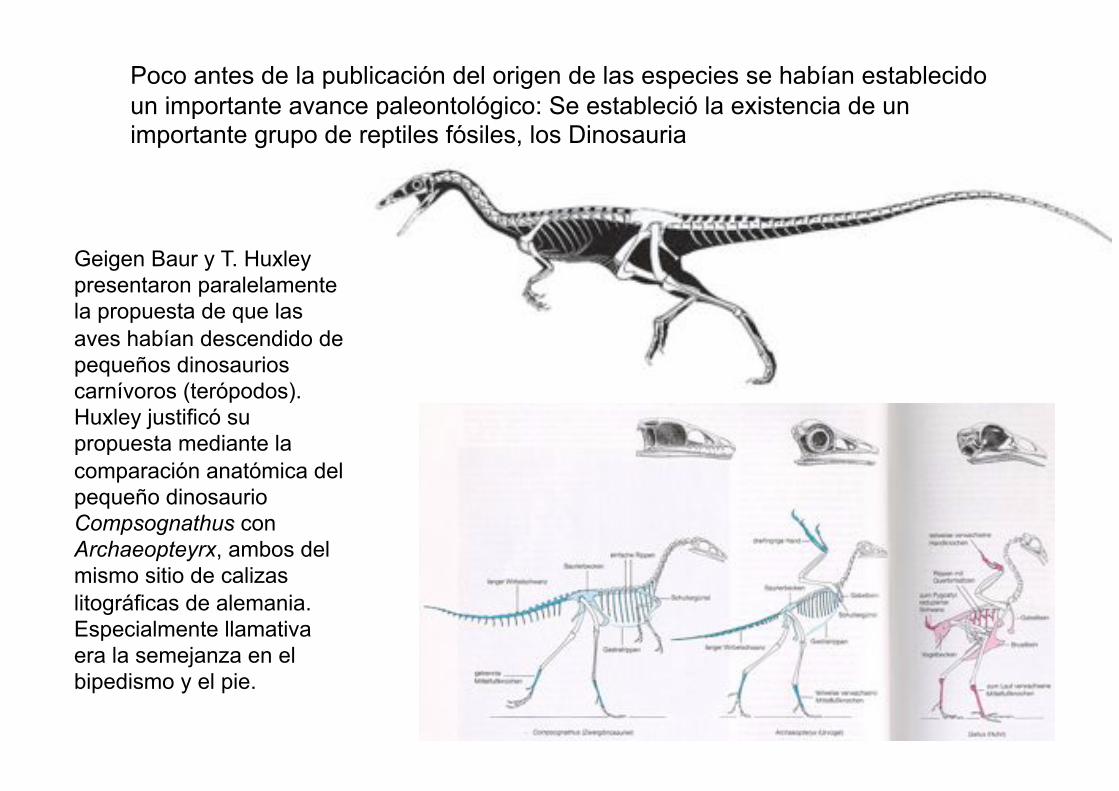
Geigen Baur y T. Huxley
presentaron paralelamente
la propuesta de que las
aves habían descendido de
pequeños dinosaurios
carnívoros (terópodos).
Huxley justificó su
propuesta mediante la
comparación anatómica del
pequeño dinosaurio
Compsognathus con
Archaeopteyrx, ambos del
mismo sitio de calizas
litográficas de alemania.
Especialmente llamativa
era la semejanza en el
bipedismo y el pie.
Poco antes de la publicación del origen de las especies se habían establecido
un importante avance paleontológico: Se estableció la existencia de un importante grupo de reptiles fósiles, los Dinosauria

Sin embargo, pronto se generaron hipótesis alternativas, que consideraban
que las semejanzas observadas entre dinosaurios terópodos y las aves habían sido adquiridas de manera independiente como el resultado de la
convergencia en torno al estilo de vida bípedo. Se señalaba además que los
terópodos evidenciaban la reducción del tamaño del brazo, lo cual se consideraba un impedimento para la evolución de las alas.

En 1929, un importante libro sobre el
origen de las aves por Heilmann reconocía las semejanzas existentes
entre aves y dinosaurios terópodos. Sin embargo, consideró que la ausencia de
clavícula en los terópodos implicaba que
estos no podían haber dado origen a las aves, en las cuales las clavículas son de
gran tamaño y componen la “fúrcula” o hueso de los deseos.
En aquel entonces era popular el concepto de la “ley de Dollo” que afirma
que un órgano o estrucutra perdido en la evolución tiene una probabilidad muy
baja de volver a aparecer bajo la misma
forma en los descendientes. Actualmente se sabe que esta
consideración no puede ser aplicada como ley y que las reversiones o
atavismos ocurren de manera natural y
pueden obtenerse experimentalmente (Ej. Dientes de pollo por Dr. David
Lemus, Chile)

Heilmann popularizó el concepto
de que ancestros cuadrúpedos y arborícolas habían dado origen a
las aves, evolucionando el vuelo en el contexto de los saltos entre
ramas, de manera similar a lo
ocurrido en las ardillas voladoras.
Los dinosaurios terópodos parecían impedidos de desarrollar
un estilo de vida arborícola por su
postura bípeda y brazos reducidos. Más aún, la ubicación
del centro de masa de los terópodos en la cadera, impediría
efectuar saltos entre rama y rama,
a diferencia de un centro de masa ubicado más hacia anterior

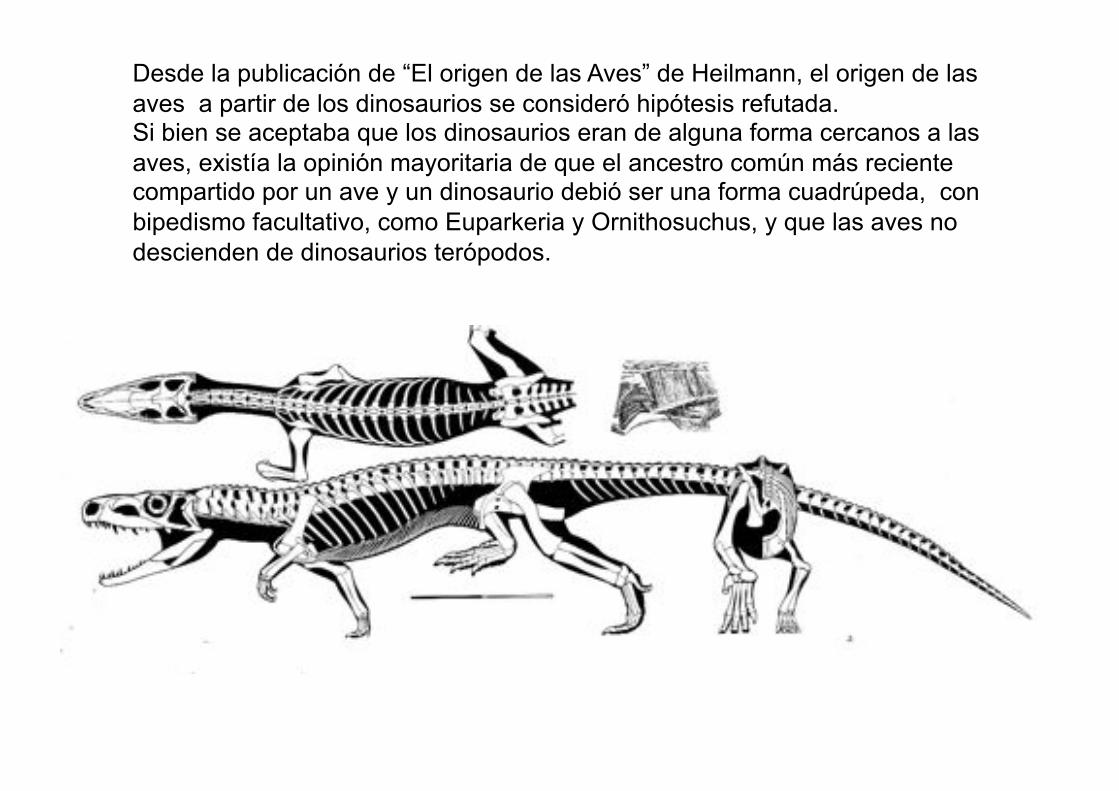
Desde la publicación de “El origen de las Aves” de Heilmann, el origen de las
aves a partir de los dinosaurios se consideró hipótesis refutada. Si bien se aceptaba que los dinosaurios eran de alguna forma cercanos a las
aves, existía la opinión mayoritaria de que el ancestro común más reciente compartido por un ave y un dinosaurio debió ser una forma cuadrúpeda, con
bipedismo facultativo, como Euparkeria y Ornithosuchus, y que las aves no
descienden de dinosaurios terópodos.

Megalancosaurus
Se ha propuesto al protorosuario Longisquama
como una posible alternativa para un ancestro no-dinosaurio las aves, dado el descubrimiento de
estructuras en su piel que han sido comparadas con plumas, pero que en realidad poseen diferencias
estructurales importantes

Desde los años 60’s hasta hoy, una serie de descubrimientos fósiles han
resucitado con fuerza la hipótesis de que las aves son dinosaurios terópodos
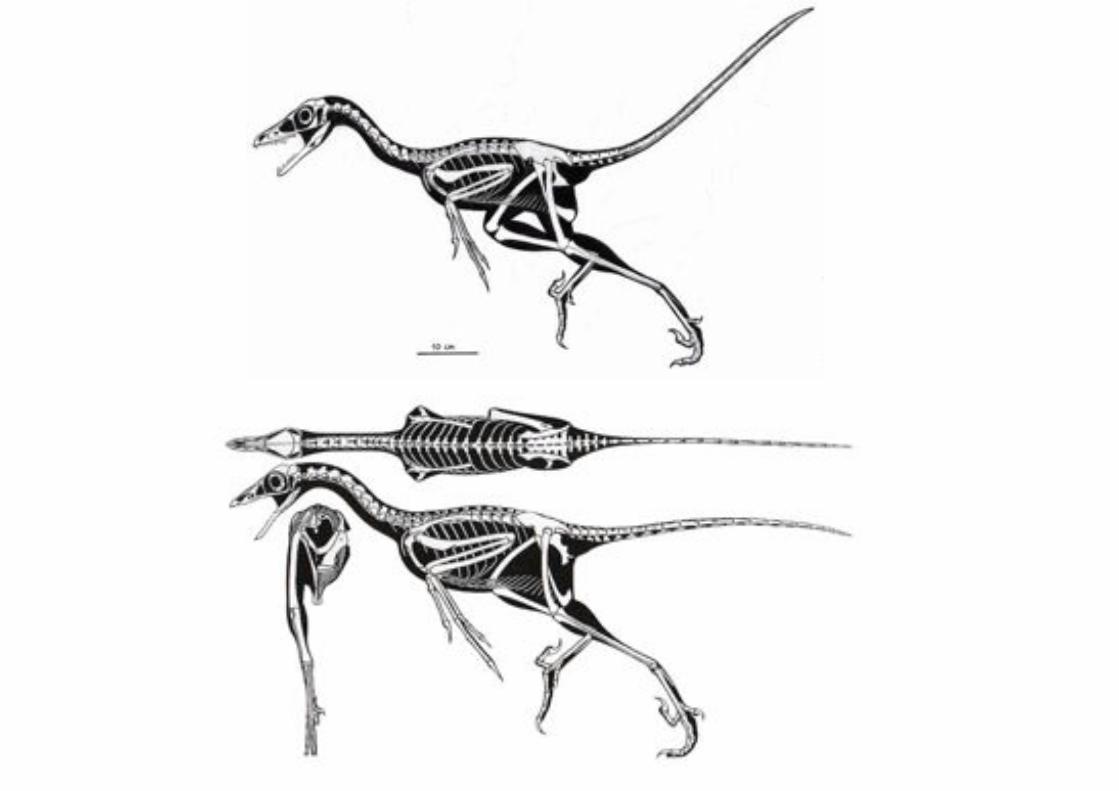
El descubrimiento del terópodo dromaeosaurido Deinonychus antirrhopus
reveló nuevas semejanzas entre terópodos y las aves. El Dr. John Ostrom de Yale resucitó la hipótesis de un origen terópodo de las aves a partir de
la comparación del esqueleto de Deinonychus con Archaeopteryx. Deinonychus revela que existieron dinosaurios terópdos muy semejantes a
alas aves que poseían largos brazos con manos raptoriales y una
impresionante garra en el pie que sugería un estilo de depredación activa. Resucitan las afirmaciones de que podía esperarse que algunos fósiles de
dinosaurios llegaran a ser descubiertos con plumas.

Deinonychus

Comienza la época del “renacimiento de los dinosaurios” Se argumenta
que los dinosaurios no eran animales de sangre fría e inactivos sino que eran más semejantes a las aves en la complejidad de su conducta y su
nivel de actividad y metabolismo. Se presentan varias evidencias en este sentido y se genera un importante debate repecto al metabolismo de los
dinosaurios.
John Ostrom Robert Bakker
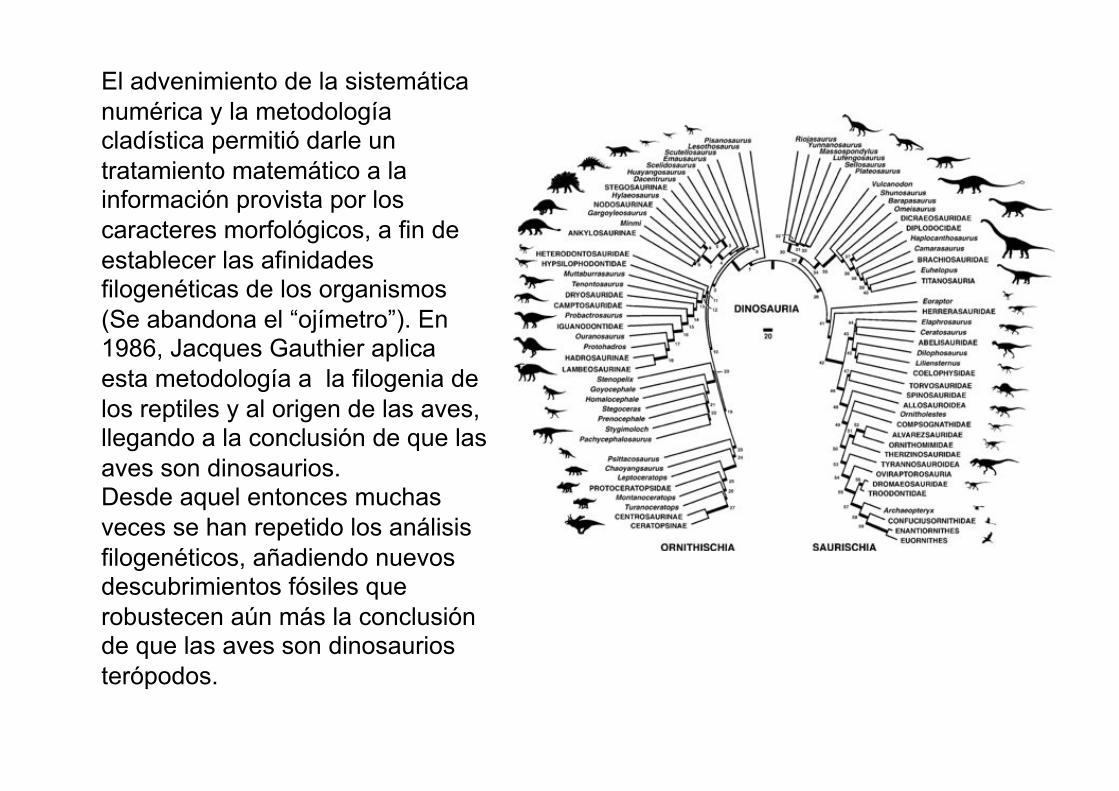
El advenimiento de la sistemática
numérica y la metodología cladística permitió darle un
tratamiento matemático a la información provista por los
caracteres morfológicos, a fin de
establecer las afinidades filogenéticas de los organismos
(Se abandona el “ojímetro”). En 1986, Jacques Gauthier aplica
esta metodología a la filogenia de
los reptiles y al origen de las aves, llegando a la conclusión de que las
aves son dinosaurios. Desde aquel entonces muchas
veces se han repetido los análisis
filogenéticos, añadiendo nuevos descubrimientos fósiles que
robustecen aún más la conclusión de que las aves son dinosaurios
terópodos.

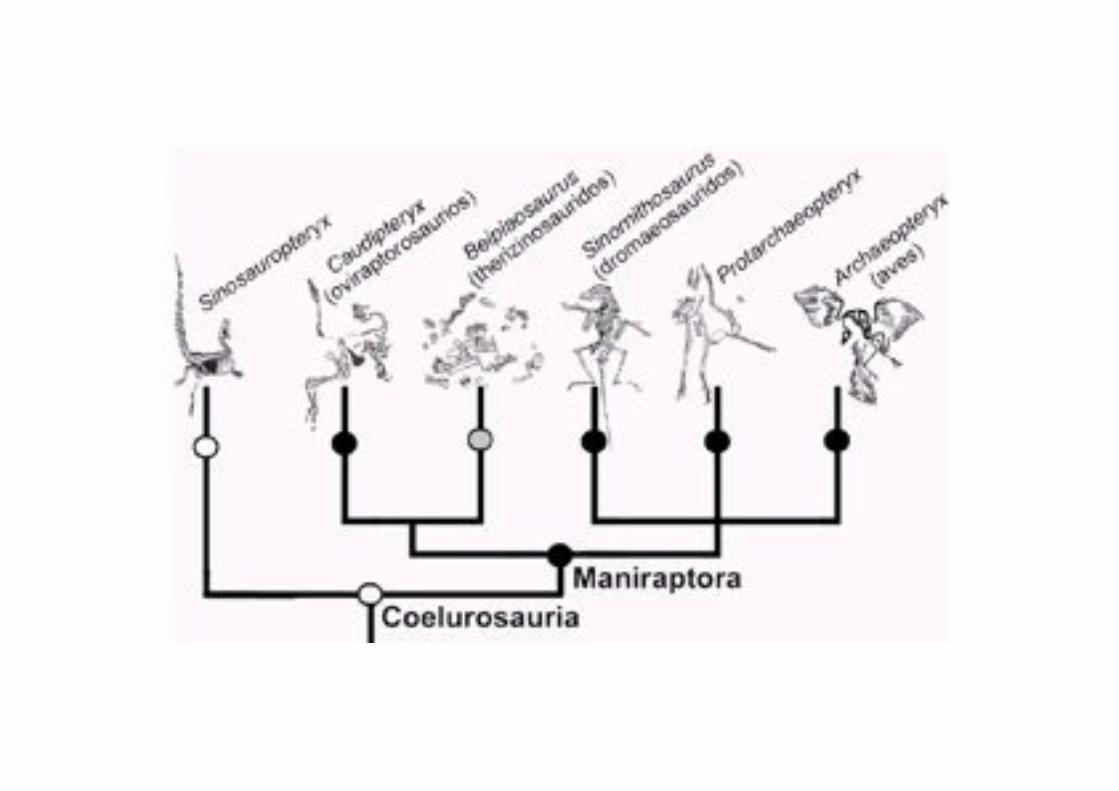
Sin embargo, han persistido un grado de polémica respecto a cuales
dinosaurios celurosaurios son los más cercanos a las aves
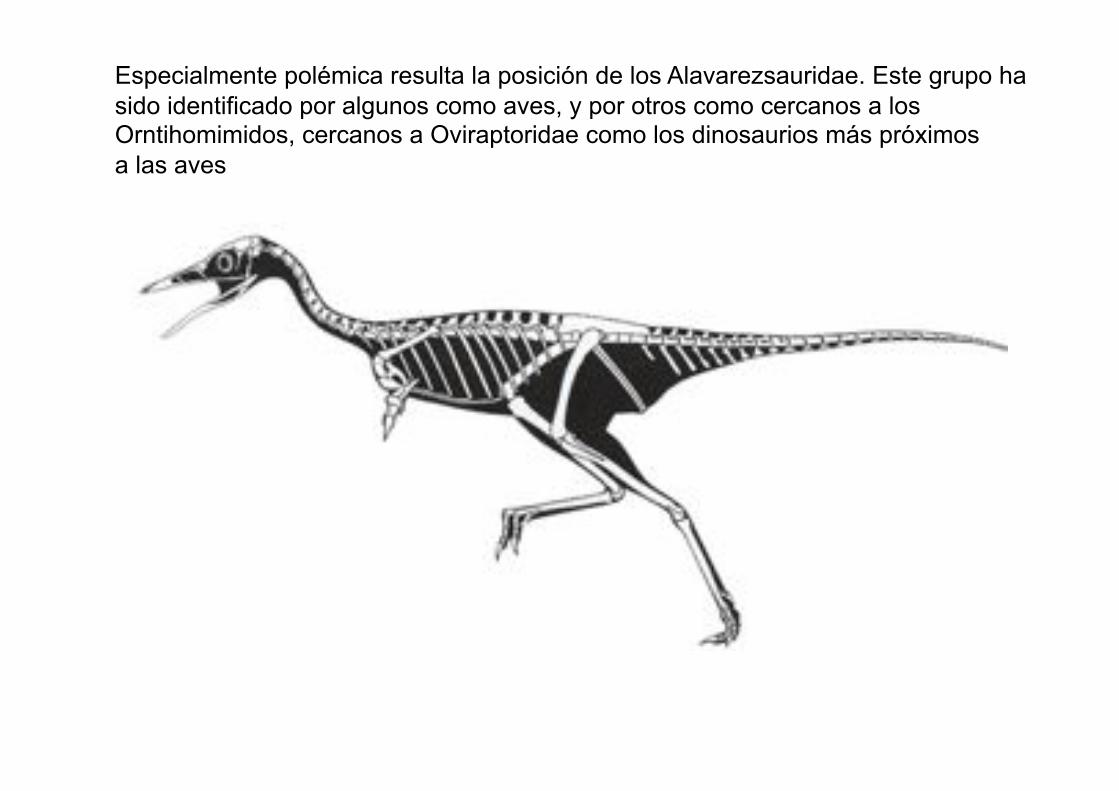
Especialmente polémica resulta la posición de los Alavarezsauridae. Este grupo ha
sido identificado por algunos como aves, y por otros como cercanos a los Orntihomimidos, cercanos a Oviraptoridae como los dinosaurios más próximos
a las aves

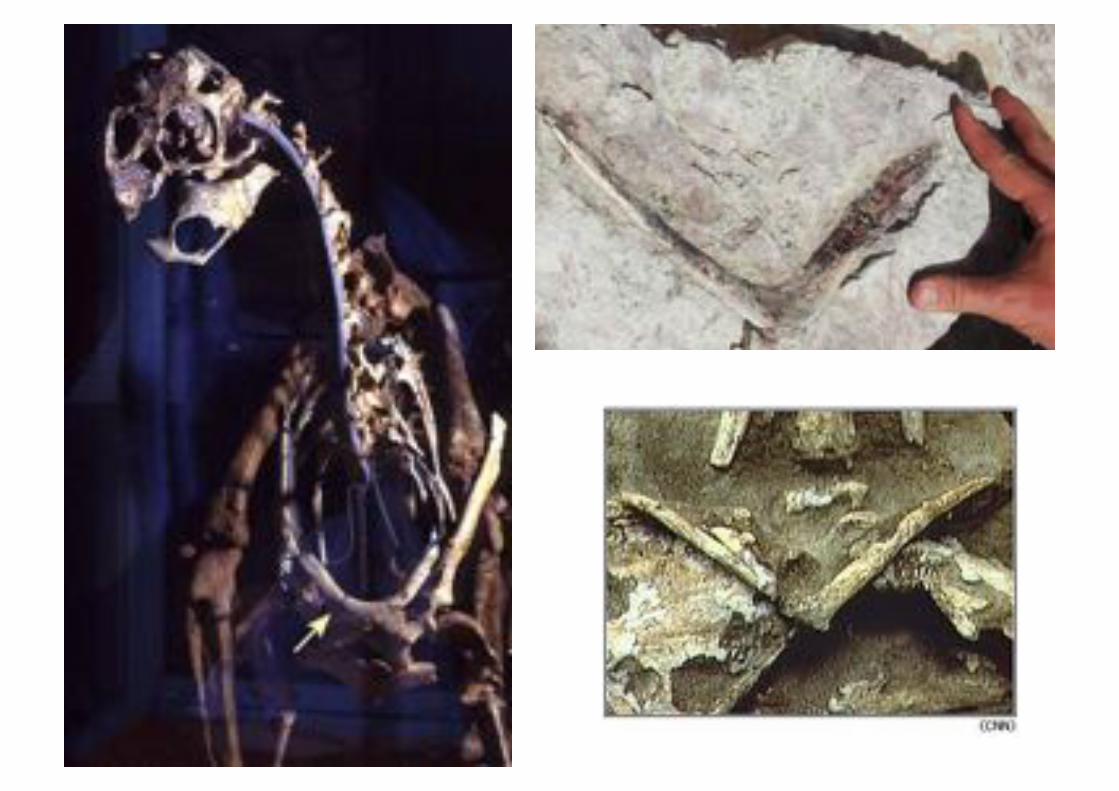
Comienzan a descubrirse dinosaurios terópodos con preservación de las
clavículas. Más impresionante aún, en muchos se aprecia que las clavículas se fusionan hacia la línea media, al igual que el hueso de los deseos. La fúrcula
se encuentra presente en dinosaurios terópodos de varios grupos, incluyendo formas grandes y pequeñas. Muchas veces la fúrcula fue confundida con una
costilla ventral (gastralia)
La objeción interpuesta por la ley de Dollo queda empíricamente descartada.
Con el advenimiento de la metodología cladística, las aves fueron aceptadas como un tipo de dinosaurio dentro de la comunidad dedicada a la sistemática
filogenética y la gran mayoría de la comunidad paleontológica


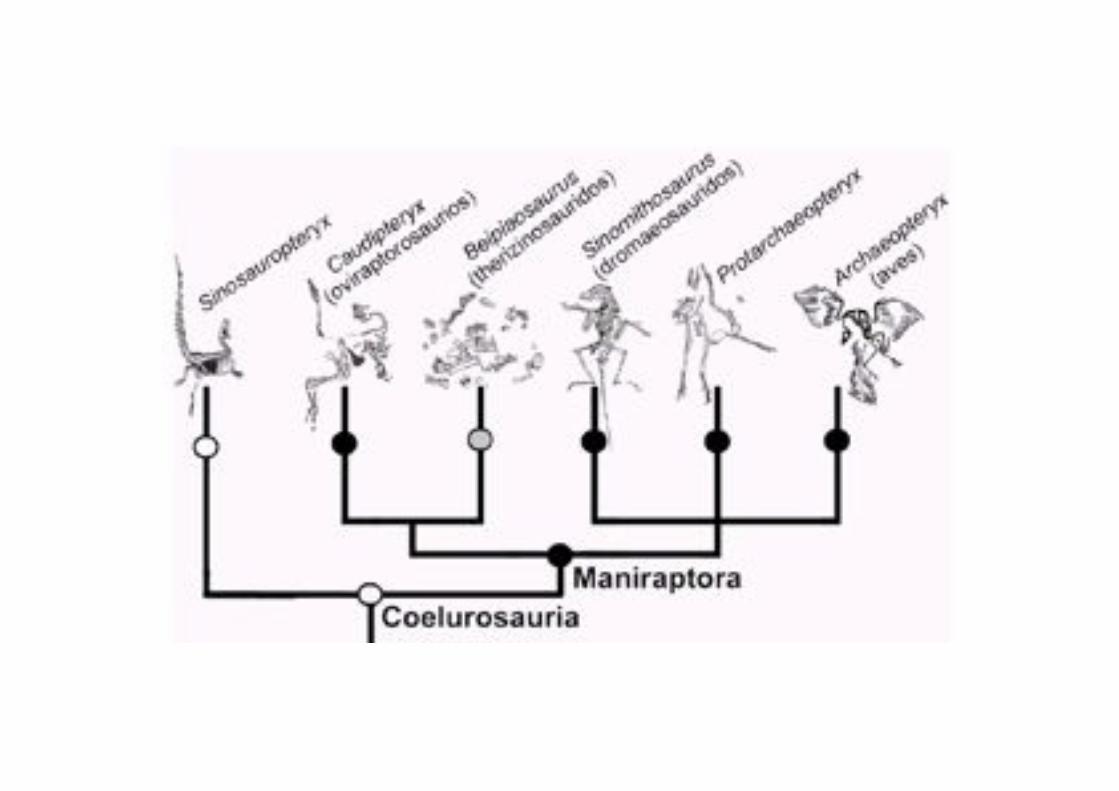
Hacia la década de los 90’s hubo un resurgimiento de objeciones teóricas en
contra de un origen de las aves a partir de dinosaurios, con argumentos construidos desde la perspectiva de la mecánica respiratoria. Estas objeciones
sin embargo se han demostrado infundadas, ya no corren dentro de la comunidad científica, y sus propios autores ahora confiesan que las aves son
dinosaurios.
El origen de las aves a partir de los dinosaurios ha recbido un fuerte
espaldarazo por el descubimiento a partir de 1997 de dinosaurios “peludos” (cubiertos con un integumento filiforme) y dinosaurios que presentan
plumas.
Sin embargo los detractores de siempre (especialmente Alan Feduccia, editor
de Science) han argumentado que estos no son dinosaurios sino aves que se parecen a dinosaurios, negando toda utilidad de los resultados del tratamiento
matemático de la información morfológica.

Sinosauropteryx
prima



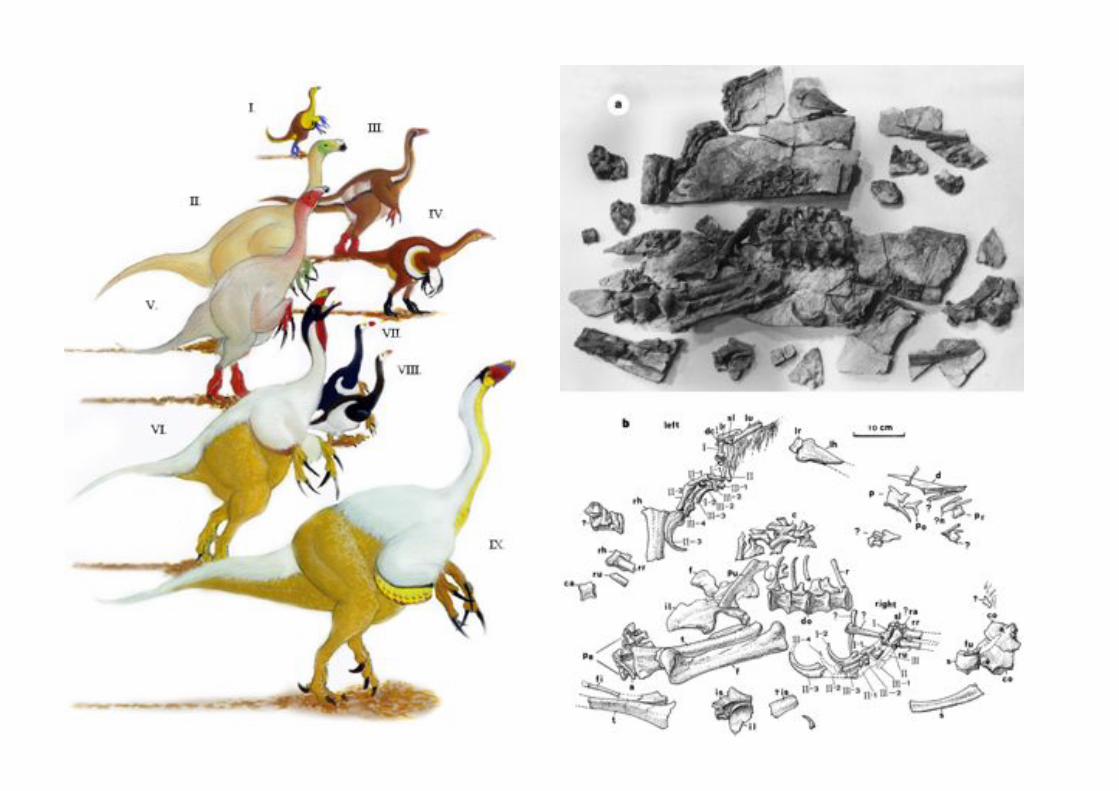




Dromaeosauridae








Confuciusornis


The problem of digit identity in the wing and the origin of birds
Alexander O. Vargas
Department of Anatomy, University of Wisconsin-Madison Department of Ecology and Evolutionary Biology
Yale University

Definition of birds (Avialae):
XVIIIth century. Linnaeus: Any animal
showing feathers
In the XIXth century, the discovery of
Archaeopteryx, was accordingly considered “the first bird”
Following the historical trail, a generally
accepted current group definition of Birds
has developed : All descendants of the most recent common ancestor shared by
Archaeopteryx with modern birds (Neornithes).
Introduction: The evolutionary origin of birds

Theropoda
Aves (Avialae)
DINOSAURIA Sereno 1999
With the advent of cladistic methods of phylogenetic analysis (based on
algorythms of parsimony) the hypothesis that birds descend from the bipedal predatory dinosaurs (Theropoda) has been reaffirmed

Birds belong in the Archosauria: the
group comprising all the descendants of the most recent common ancestor
shared by birds and crocodilia (The crocodilia are birds closest living
relatives).
Birds are at the same time a type of
archosaur, dinosaur and theropod in the same nested sense in which it is
possible to state that humans are at
the same time a type of mammal, primate and anthropoid (All bird is a
dinosaur, but not all dinosaurs are birds)
Dinosauria
Archosauria
Theropoda
Sumida & Brochu 2000 Aves
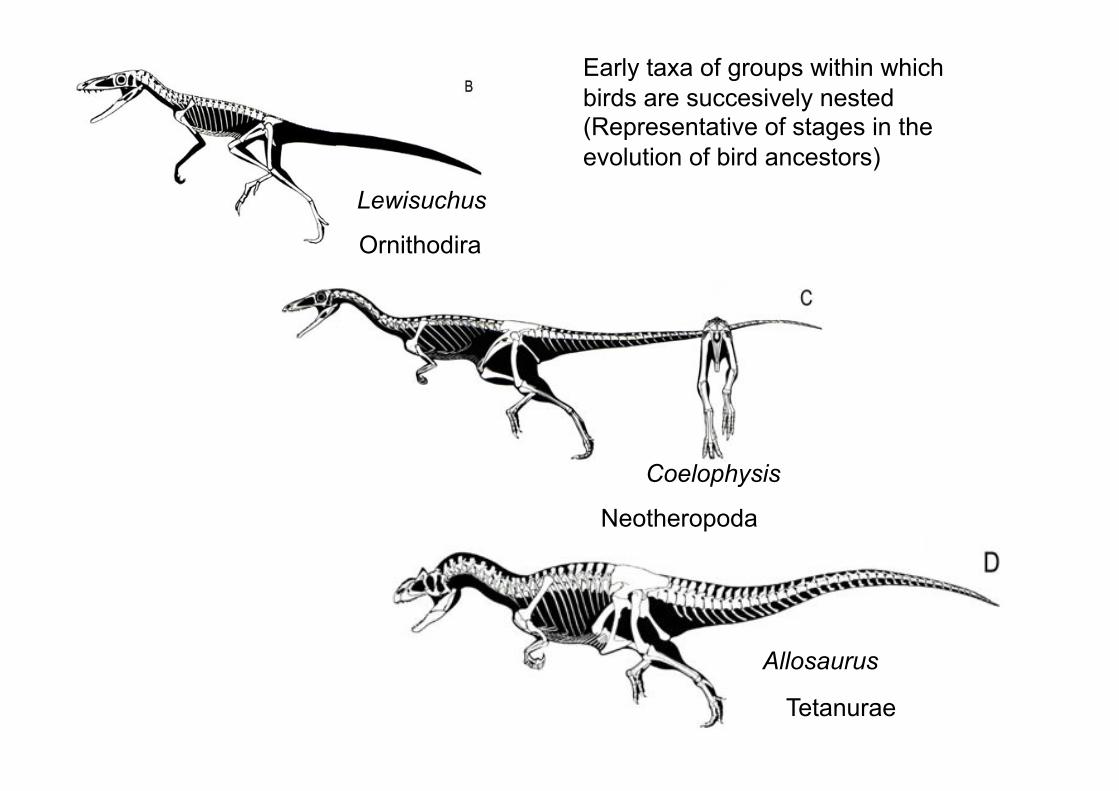
Ornithodira
Neotheropoda
Tetanurae
Allosaurus
Coelophysis
Lewisuchus
Early taxa of groups within which
birds are succesively nested (Representative of stages in the
evolution of bird ancestors)

Ornitholestes
Sinornithosaurus
Maniraptora
Eumaniraptora
(Paraves)
Avialae
Rahonavis

Because of the phylogenetic proximity between Archaeopteryx and some
theropod dinosaurs, it was long ago predicted that theropods bearing feathers should eventually be discovered.
There is now evidence that the origin of feathers preceded the origin of Archaeopteryx, and that feathers were present in the theropod ancestors of
birds. Therefore, the old definition of Linnaeus has been mostly abandoned.
Caudipteryx
Microraptor

Dromaeosauridae Sinornithosaurus

Microraptor

Protarchaeopteryx

However, a few researchers do not accept cladistic methods of phylogenetic analys and
maintain that birds do not descend from theropods. They argue that feathered theropods
are not such but in fact are birds that resemble theropods by convergence. Through
qualitative arguments rather than phylogenetic analysis, they propose that birds
descended from other fossil Archosaurs such as Longisquama, a possibly flying form that
presents large scales resembling feathers.
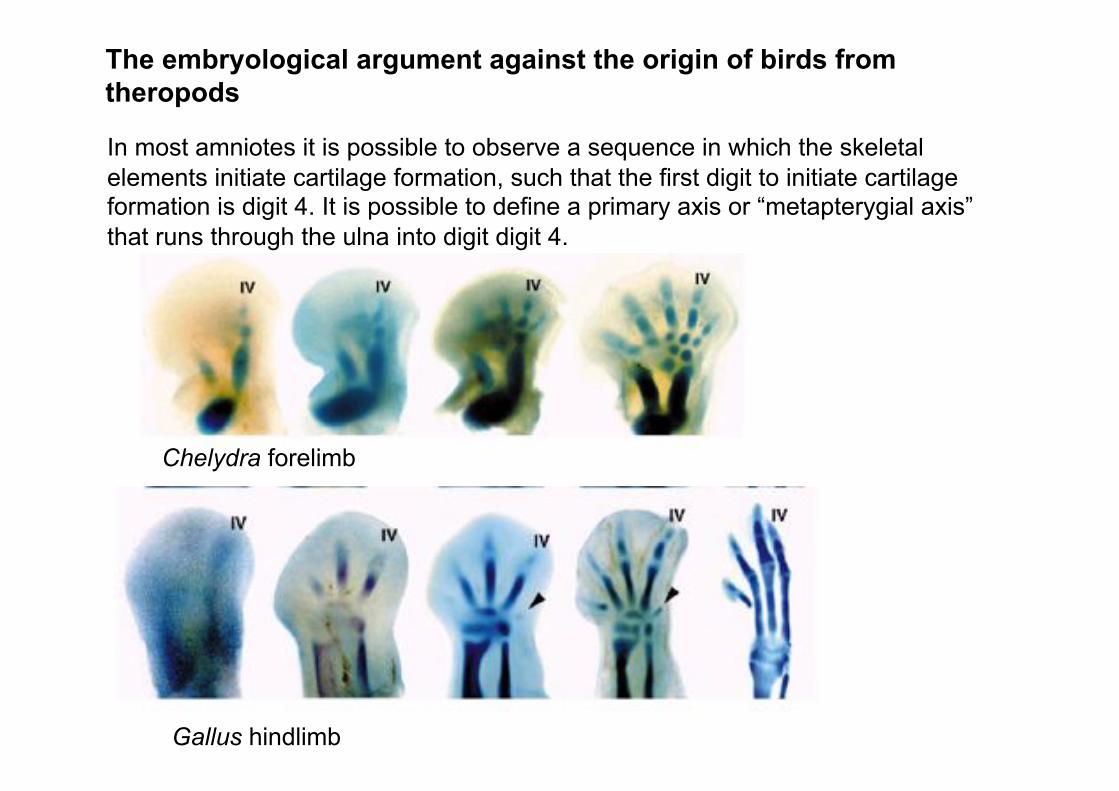
In most amniotes it is possible to observe a sequence in which the skeletal
elements initiate cartilage formation, such that the first digit to initiate cartilage formation is digit 4. It is possible to define a primary axis or “metapterygial axis”
that runs through the ulna into digit digit 4.
Chelydra forelimb
Gallus hindlimb
The embryological argument against the origin of birds from theropods

Within developmental biology, the digits of the wing are called 2, 3 and 4,
according to the fact that the primary axis of cartilage formation runs through the ulna into the posteriomost digit.
Alligator forelimb
Gallus forelimb
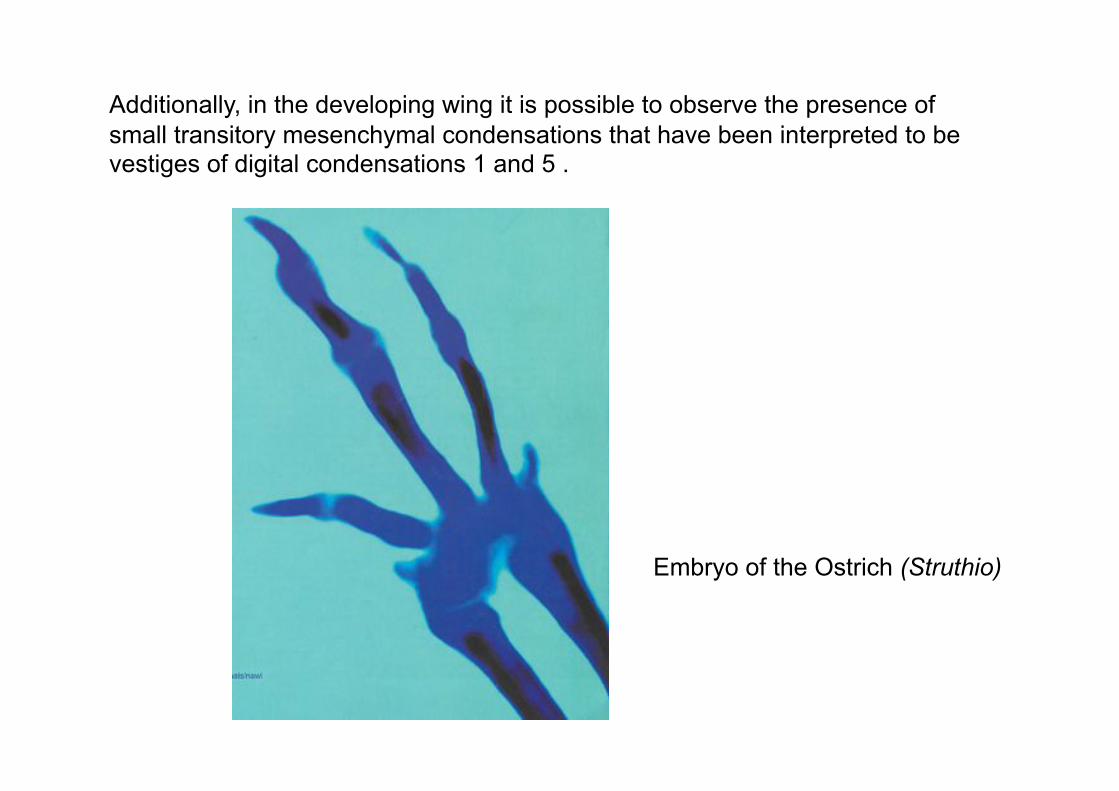
Additionally, in the developing wing it is possible to observe the presence of
small transitory mesenchymal condensations that have been interpreted to be vestiges of digital condensations 1 and 5 .
Embryo of the Ostrich (Struthio)
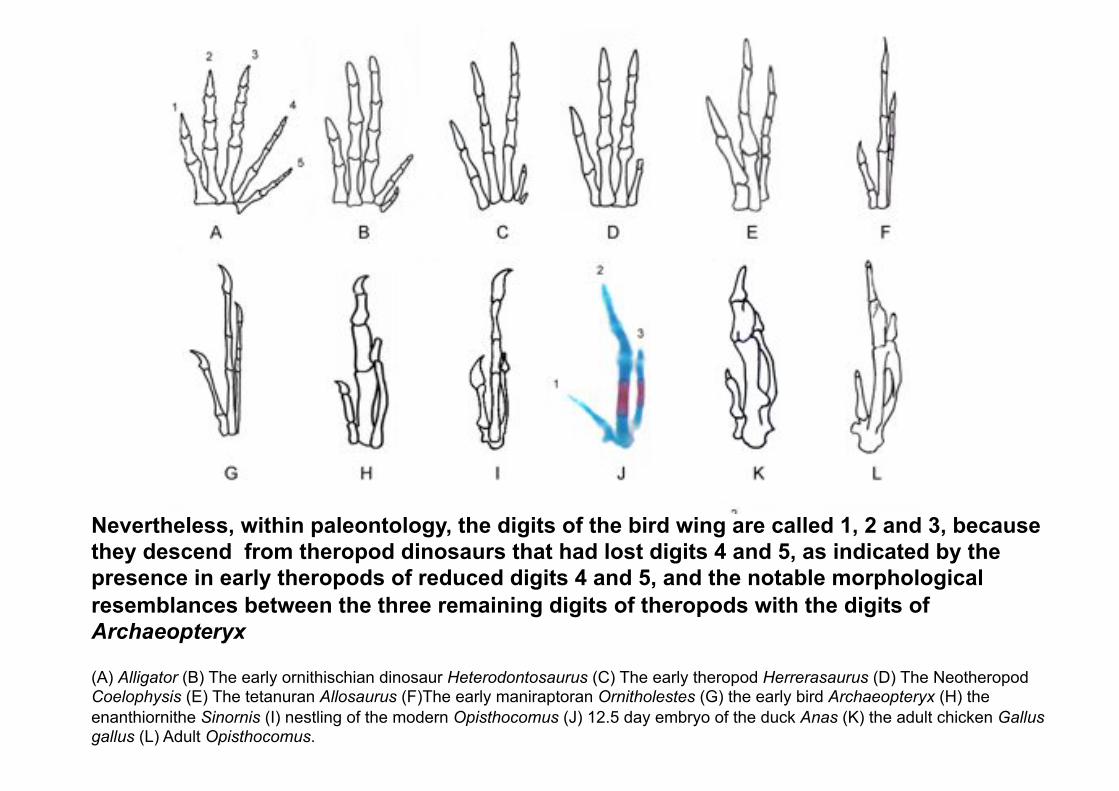
Nevertheless, within paleontology, the digits of the bird wing are called 1, 2 and 3, because they descend from theropod dinosaurs that had lost digits 4 and 5, as indicated by the presence in early theropods of reduced digits 4 and 5, and the notable morphological resemblances between the three remaining digits of theropods with the digits of Archaeopteryx
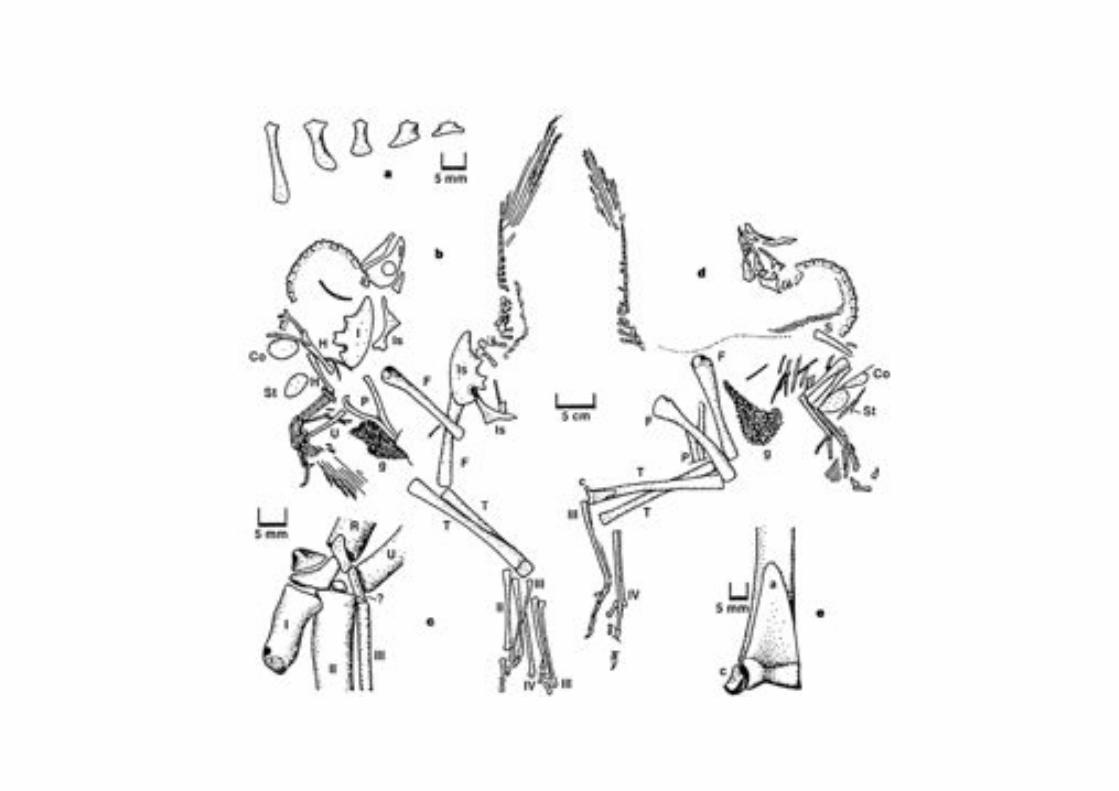
(A) Alligator (B) The early ornithischian dinosaur Heterodontosaurus (C) The early theropod Herrerasaurus (D) The Neotheropod Coelophysis (E) The tetanuran Allosaurus (F)The early maniraptoran Ornitholestes (G) the early bird Archaeopteryx (H) the
enanthiornithe Sinornis (I) nestling of the modern Opisthocomus (J) 12.5 day embryo of the duck Anas (K) the adult chicken Gallus gallus (L) Adult Opisthocomus.

It has been proposed that the embryological condition of the bird wing is
inconsistent with the origin of birds from theropod dinosaurs, and that birds alternatively must have descended from an ancestor that had lost digits 1
and 5

The apparent inconsistency with the embryology of the wing is considered the greatest obstacle to full acceptance of the hypothesis that birds descend from dinosaurs
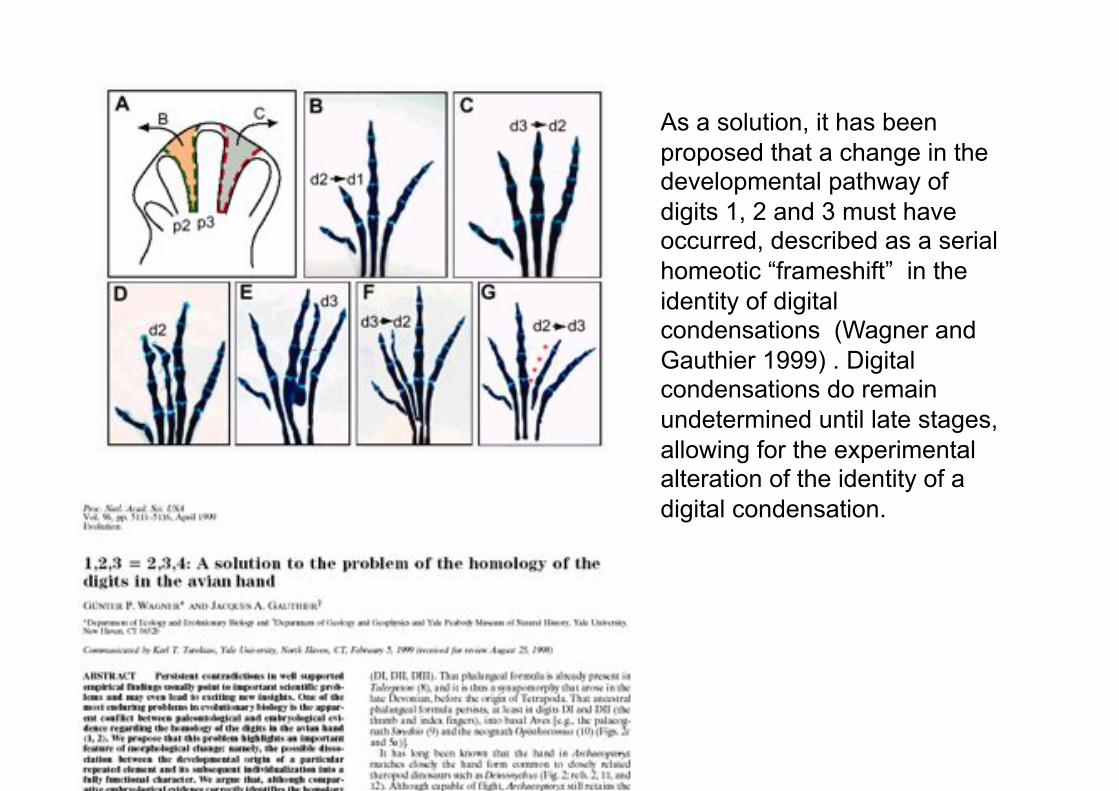
As a solution, it has been
proposed that a change in the developmental pathway of
digits 1, 2 and 3 must have occurred, described as a serial
homeotic “frameshift” in the
identity of digital condensations (Wagner and
Gauthier 1999) . Digital condensations do remain
undetermined until late stages,
allowing for the experimental alteration of the identity of a
digital condensation.

Fig 2.4.3 Homeotic transformation of the thumb to an index in humans.
Heiss (1957)
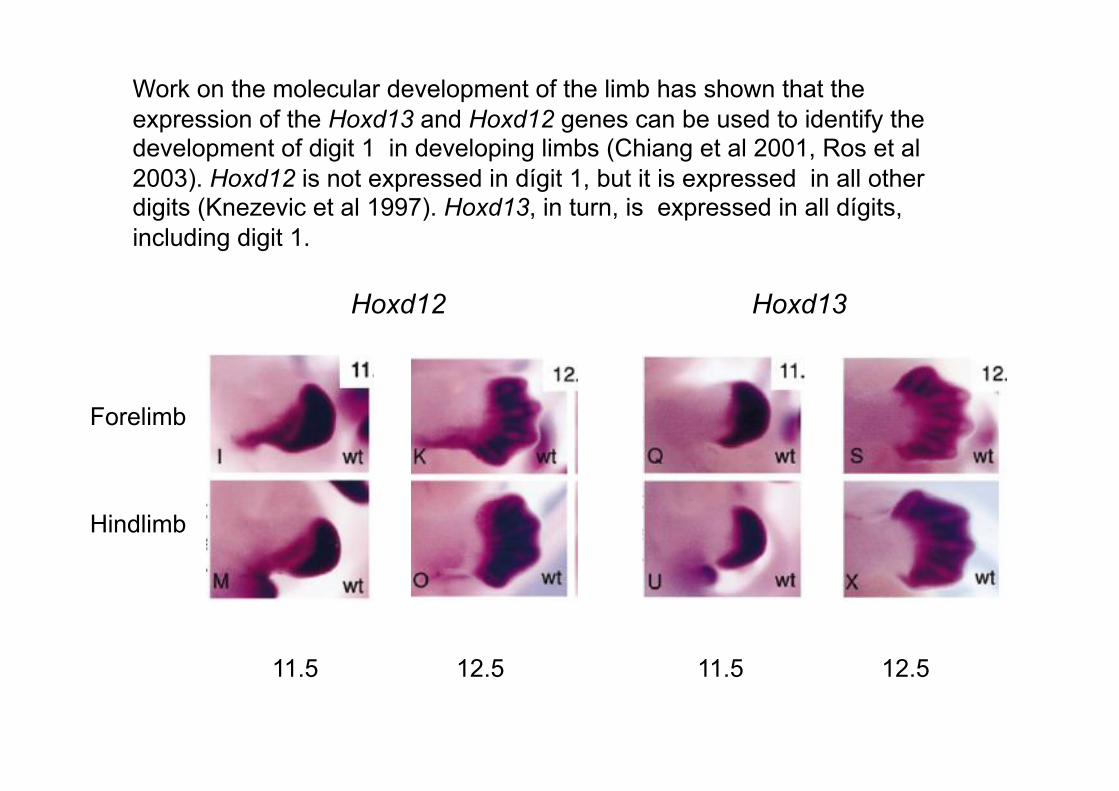
Work on the molecular development of the limb has shown that the
expression of the Hoxd13 and Hoxd12 genes can be used to identify the development of digit 1 in developing limbs (Chiang et al 2001, Ros et al
2003). Hoxd12 is not expressed in dígit 1, but it is expressed in all other digits (Knezevic et al 1997). Hoxd13, in turn, is expressed in all dígits,
including digit 1.
Hoxd12 Hoxd13
Forelimb
Hindlimb
11.5 12.5 11.5 12.5

Forced misexpression of Hoxd12 in all digits of the mouse can lead to the
homeotic transformation of dígit 1 to a longer, triphalangeal dígit 2 (Knezevic et al. 1997).
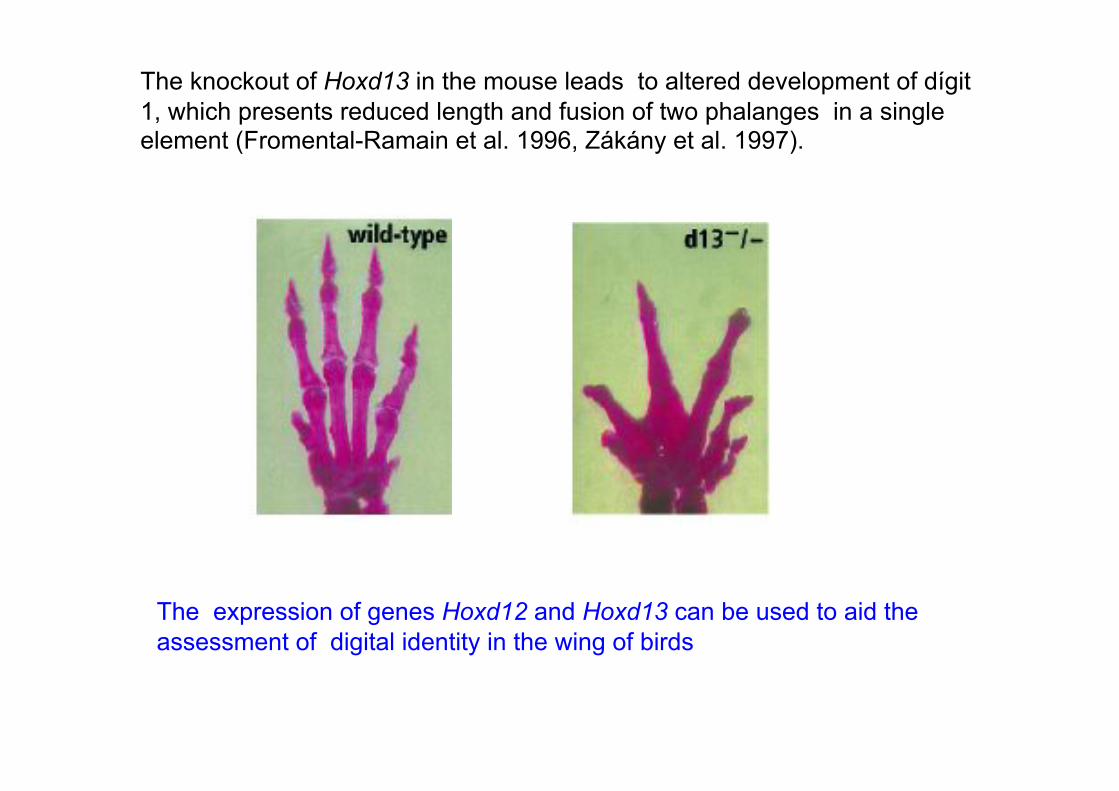
The knockout of Hoxd13 in the mouse leads to altered development of dígit
1, which presents reduced length and fusion of two phalanges in a single element (Fromental-Ramain et al. 1996, Zákány et al. 1997).
The expression of genes Hoxd12 and Hoxd13 can be used to aid the
assessment of digital identity in the wing of birds

Sin embargo, la morfología comparada (de taxa fósiles y vivientes) identifica a los dedos del ala de las aves como 1, 2 y 3

Wagner y Gauthier 1999 PNAS 96:5111
Solución
propuesta:
1,2,3 = 2,3,4.
Inferencia de un
evento de Desplazamiento
Homeótico en Serie (“DHS”)
(Homeotic Frame-
shift ) en la evolución del ala

Feduccia 1999 PNAS
A algunos puede tomarles un tiempo entusiasmarse con el DHS…
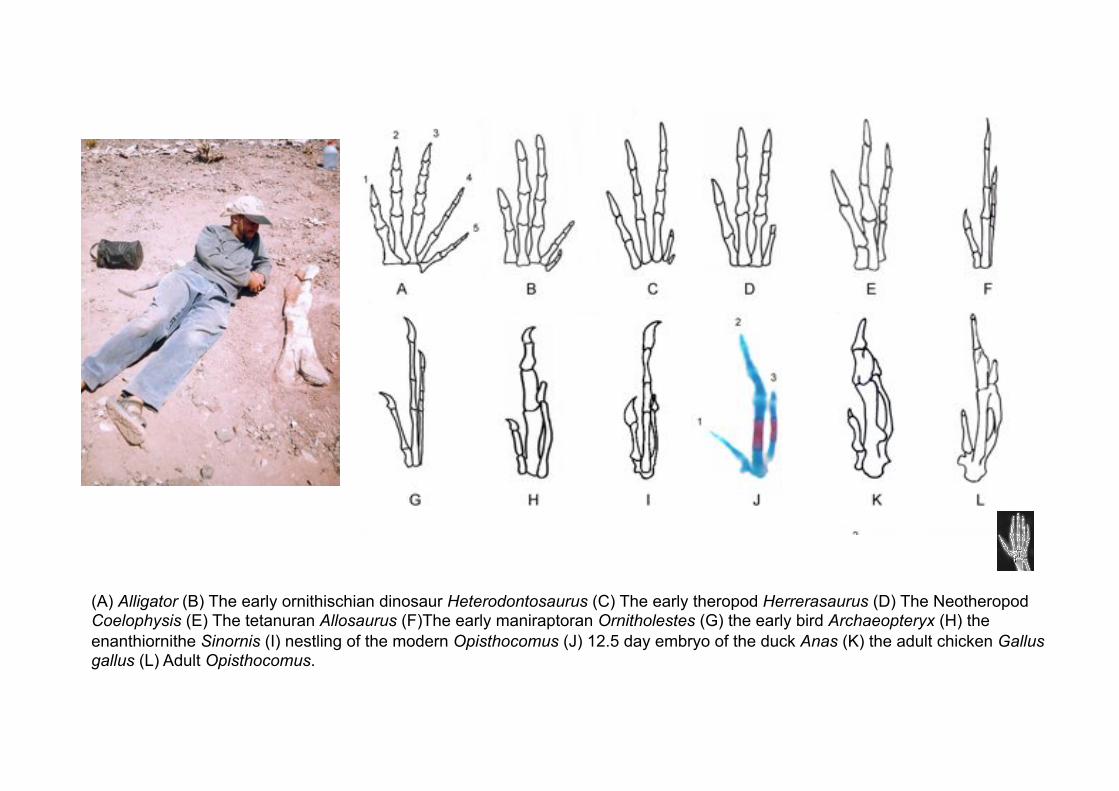
(A) Alligator (B) The early ornithischian dinosaur Heterodontosaurus (C) The early theropod Herrerasaurus (D) The Neotheropod Coelophysis (E) The tetanuran Allosaurus (F)The early maniraptoran Ornitholestes (G) the early bird Archaeopteryx (H) the
enanthiornithe Sinornis (I) nestling of the modern Opisthocomus (J) 12.5 day embryo of the duck Anas (K) the adult chicken Gallus gallus (L) Adult Opisthocomus.
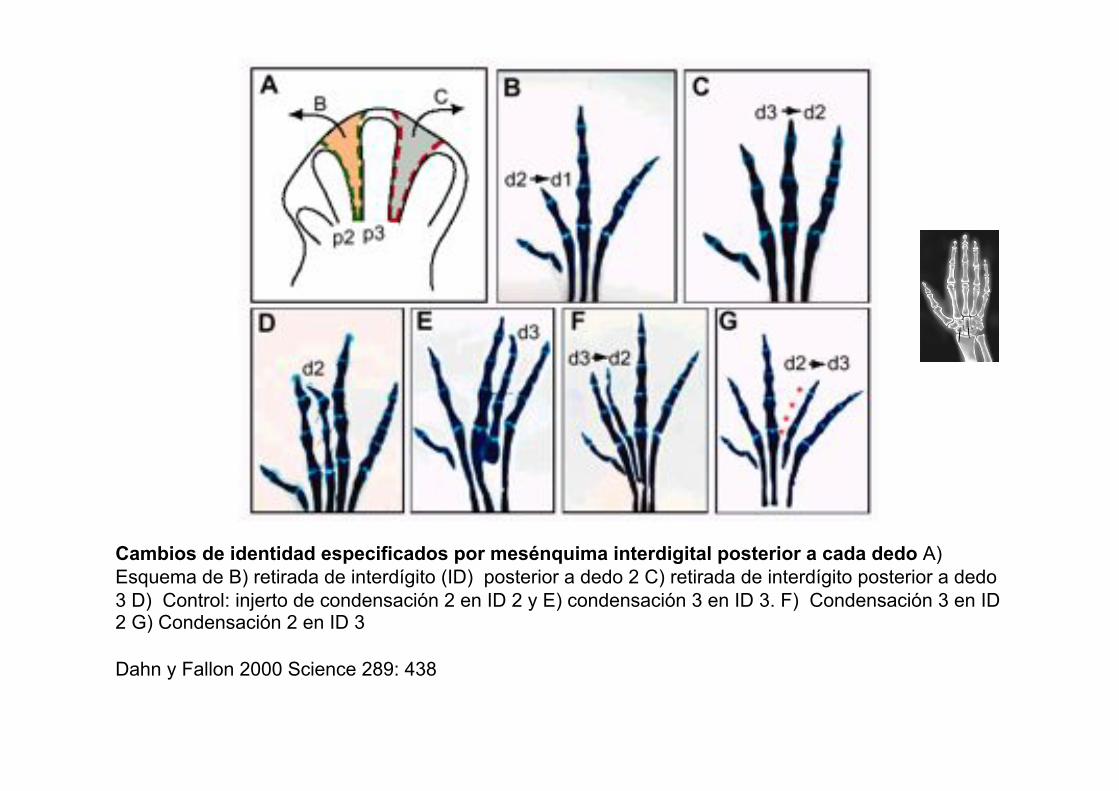
Cambios de identidad especificados por mesénquima interdigital posterior a cada dedo A)
Esquema de B) retirada de interdígito (ID) posterior a dedo 2 C) retirada de interdígito posterior a dedo
3 D) Control: injerto de condensación 2 en ID 2 y E) condensación 3 en ID 3. F) Condensación 3 en ID 2 G) Condensación 2 en ID 3
Dahn y Fallon 2000 Science 289: 438
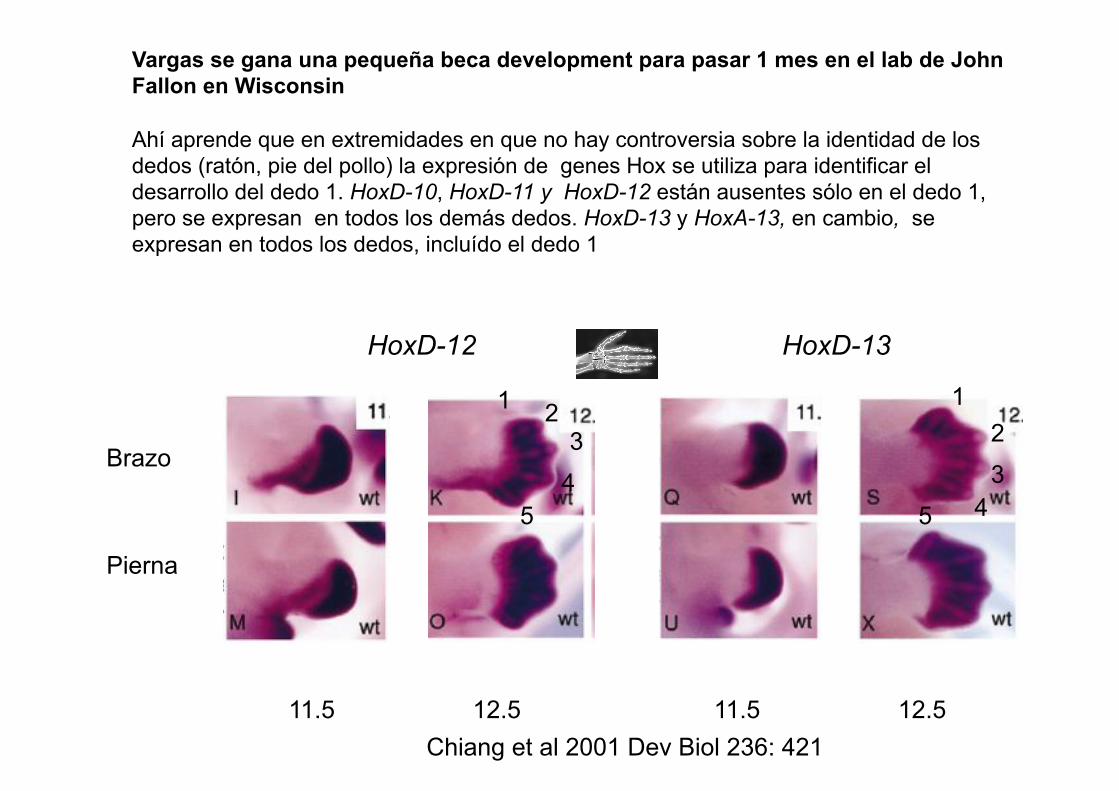
Vargas se gana una pequeña beca development para pasar 1 mes en el lab de John Fallon en Wisconsin
Ahí aprende que en extremidades en que no hay controversia sobre la identidad de los
dedos (ratón, pie del pollo) la expresión de genes Hox se utiliza para identificar el
desarrollo del dedo 1. HoxD-10, HoxD-11 y HoxD-12 están ausentes sólo en el dedo 1,
pero se expresan en todos los demás dedos. HoxD-13 y HoxA-13, en cambio, se
expresan en todos los dedos, incluído el dedo 1
HoxD-12 HoxD-13
Brazo
Pierna
11.5 12.5 11.5 12.5
Chiang et al 2001 Dev Biol 236: 421
1
3
2
4 5
1 2
3
4
5

The knockout of Hoxd13 in the mouse leads to altered development of dígit
1, which presents reduced length and fusion of two phalanges in a single element (Fromental-Ramain et al. 1996, Zákány et al. 1997).
The expression of genes Hoxd12 and Hoxd13 can be used to aid the
assessment of digital identity in the wing of birds

Expresión forzada de HoxD-12 en el dedo 1 de ratón:
transformación a un dedo 2
Expresión forzada de HoxD-12
en pie de pollo (colaboración con Fallon lab) sugiere
conservación funcional de HoxD-12 en ratón y pollo
(pese a 300 millones de años
de divergencia evolutiva)
Expresión forzada de
HoxD-11 en dedo 1 del pie
del pollo: transformación a
un dedo 2
EXPERIMENTOS FUNCIONALES

En el ala, HoxD-10, HoxD-11 y HoxD-12 están ausentes sólo en el dedo bifalangeal anterior del ala. La expresión genética sugiere que este es un dedo 1, pese a desarrollarse a partir de la posición embriológica del dedo 2.
Vargas & Fallon 2005 JEZB (MOL DEV EVOL) 304:86
Vargas and Fallon 2005 JEZ B (MDE) 304:86
Pie pollo Mano ratón
HoxD-13 HoxD-12 HoxD-13 HoxD-12
Mano de pollo (dias 7 y 8)

¿Qué tal si la ausencia de expresión de HoxD-12 es sólo cosa de ser el dedo más anterior? Los mutantes de pollo talpid2 y Silkie presentan un pie con un eje antero-posterior
alterado. talpid2 muestra un pie “posteriorizado” y polidáctilo, sin dedo 1. En Silkie, el
dedo más anterior no es el dedo 1 sino un dedo 2 ectópico
Silkie Talpid2
Vargas & Fallon 2005 JEZB 304:86

En ratón, HoxA-13 se expresa en todos los dedos, pero sólo el dedo 1 requiere HoxA-13
para su formación. Es el único dedo que desaparece en ausencia de función de HoxA-13
Image from Fromental-Ramain et al. 1996

Discussion Considering the fossil evidence, it is notable that in the development of the wing of the wildtype chicken, the expression of Hoxd12 is absent in the most
anterior digit, an the expression of Hoxd13 is present. Despite the fact this digit is described as developing from digital condensation 2, the expression of
Hoxd12 y Hoxd13 indicate it is a digit 1.


Para observar la “función” de HoxA-13 en el ala, un alelo dominante negativo (putativo)
para HoxA-13 fue contruido que comprende una fusion of dominio represor de la
transcripción del gen engrailed de drosphila y una región de HoxA-13 (EnHoxA-13) en
un vector retroviral que fue electroporado en el primordio del ala en estadio 17
Datos preliminares: Como resultado, el dedo anterior del ala dejó de formarse
sugiriendo que sólo este dedo requiere HoxA-13, tal como el dedo 1 del ratón.

Desplazamiento
Homeótico
en Serie:
Identidad y
expresión de
HoxD10-12 hacia
posterior
Ratón Pollo

Pero, el ratón es sólo un pariente lejano de las aves. Los crocodylia son los parientes vivos más cercanos a las aves.
Aproximación
comparativa:
Si el dedo más anterior del ala es un dedo 2, es posible que en los crocodilia el dedo 2 no exprese HoxD-10, HoxD-11 y HoxD-12
Si, de acuerdo al DHS, la ausencia de HoxD-10, HoxD-11 y HoxD-12 en el en el ala de las aves indica, como en el ratón, el desarrollo del dedo 1, la ausencia de expresión debiera observarse sólo en el dedo 1 de parientes más cercanos de las aves

Clonamos un fragmento del exon 1 de HoxD-11 de Alligator mississippiensis para observar su expresión en dedos embrionarios.
Vargas et al 2008 PLoS ONE 3(10): e3325

Como en el ratón, en Alligator, HoxD-11 está ausente sólo en el dedo 1 bifalangeal; el dedo anterior del ala se parece exclusivamente al dedo 1 de Alligator
Esto es consistente con el DHS, que mantiene que los dedos del ala son 1,2 y 3 pese a
desarrollarse a partir de las condensaciones 2,3,y 4
Vargas et al 2008 PLoS ONE 3(10): e3325

La transformación homeótica de dedos se ha documentado, pero sólo para dedos individuales
Dahn y Fallon 2000 Science 289:438
La evidencia experimental: ¿Es posible un DHS en el lab?

Sin embargo, nunca se ha demostrado el desplazamiento en serie, simultáneo de más de un dígito. ¿Podemos inducir un DHS de manera experimental?
Scherz et al. 2007 Dev Biol 308:343
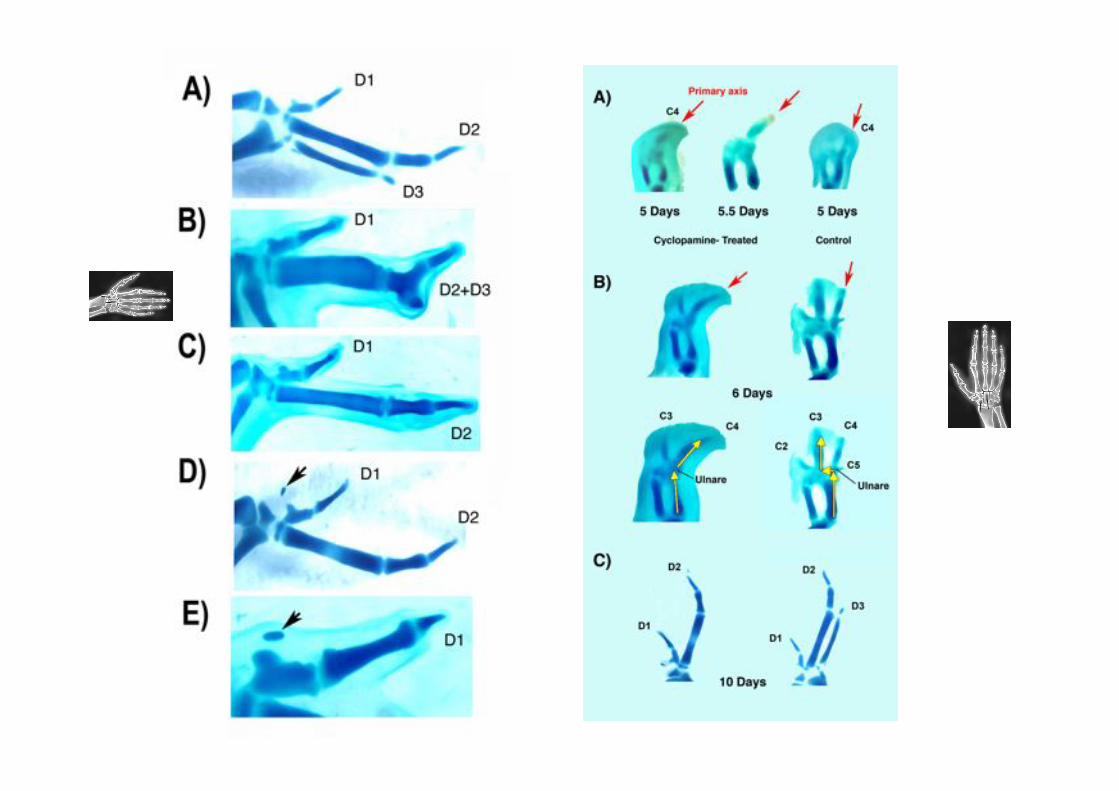
La aplicación de cyclopamina en estadios 18-20 produce alas bidáctilas con dedos 1 y 2.
¿Dónde está el eje primario de
formación de cartílago en estas
alas experimentales?
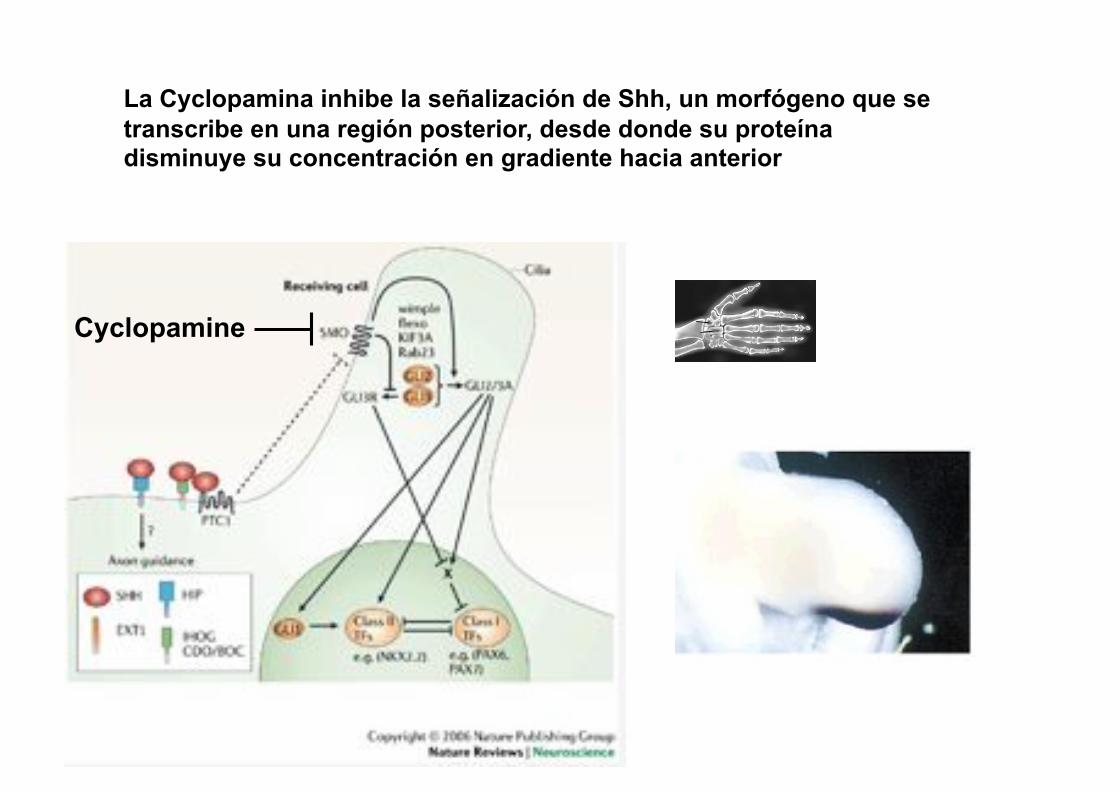
Cyclopamine
La Cyclopamina inhibe la señalización de Shh, un morfógeno que se transcribe en una región posterior, desde donde su proteína disminuye su concentración en gradiente hacia anterior

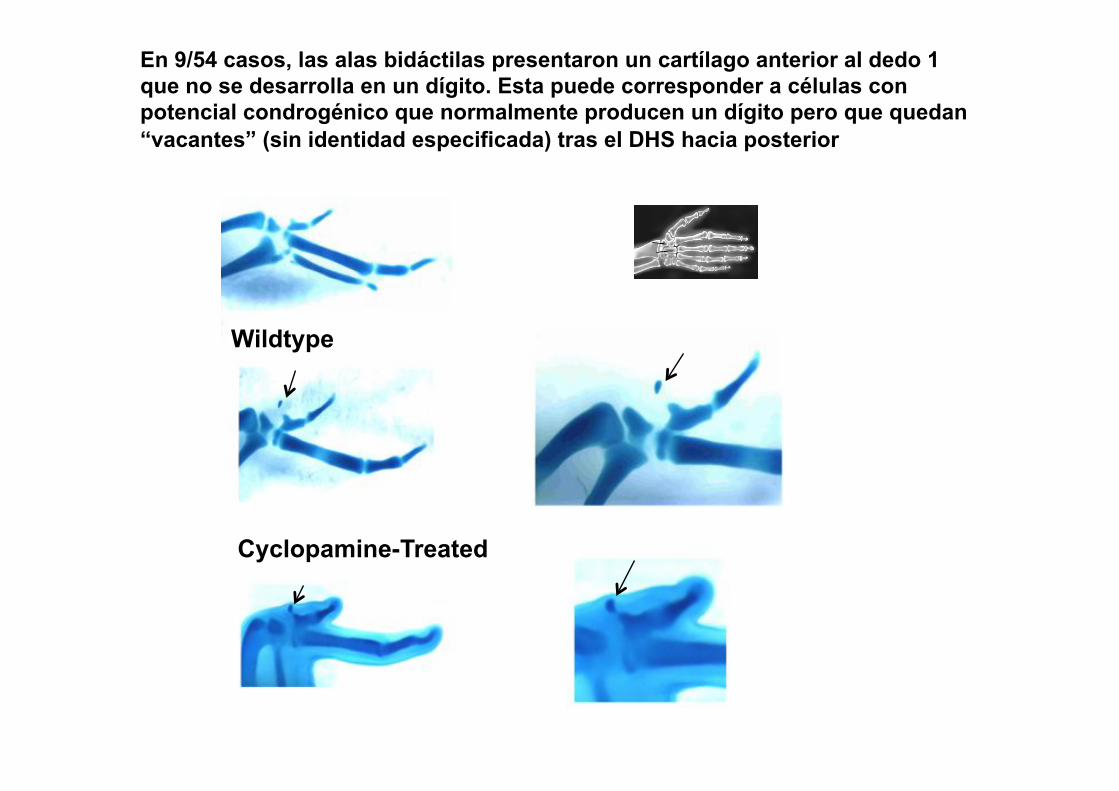
En 9/54 casos, las alas bidáctilas presentaron un cartílago anterior al dedo 1 que no se desarrolla en un dígito. Esta puede corresponder a células con potencial condrogénico que normalmente producen un dígito pero que quedan “vacantes” (sin identidad especificada) tras el DHS hacia posterior
Wildtype
Cyclopamine-Treated
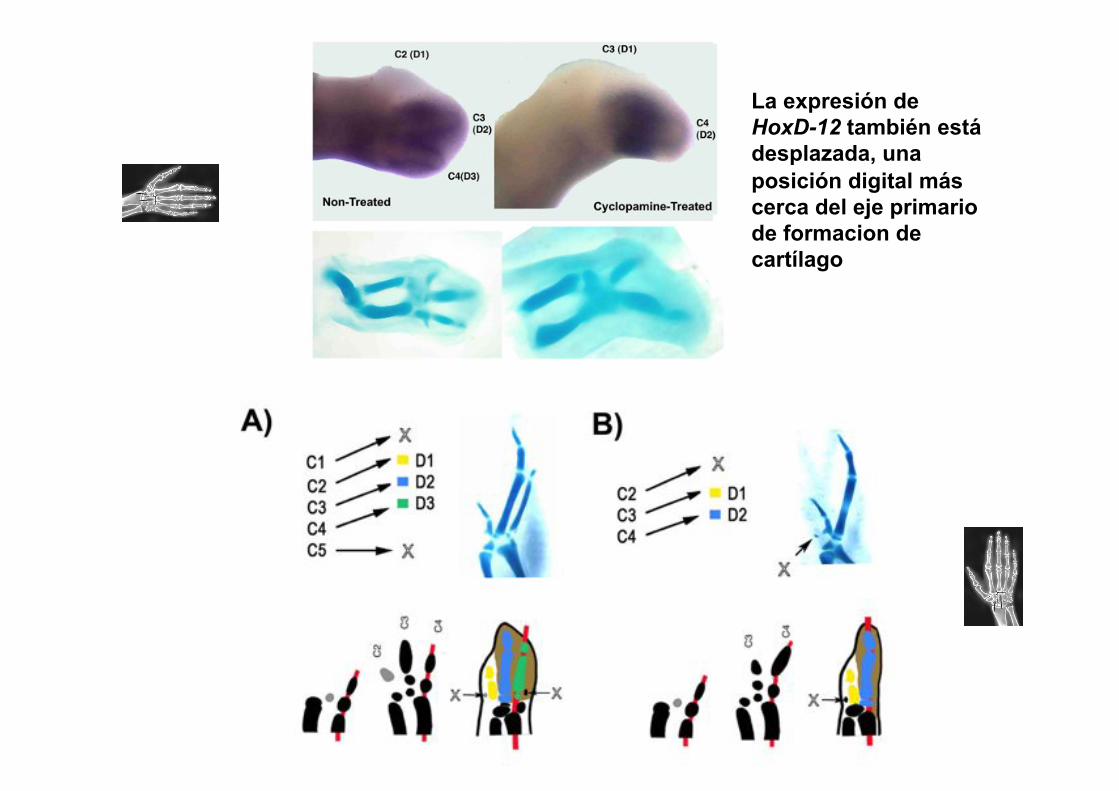
La expresión de HoxD-12 también está desplazada, una posición digital más cerca del eje primario de formacion de cartílago
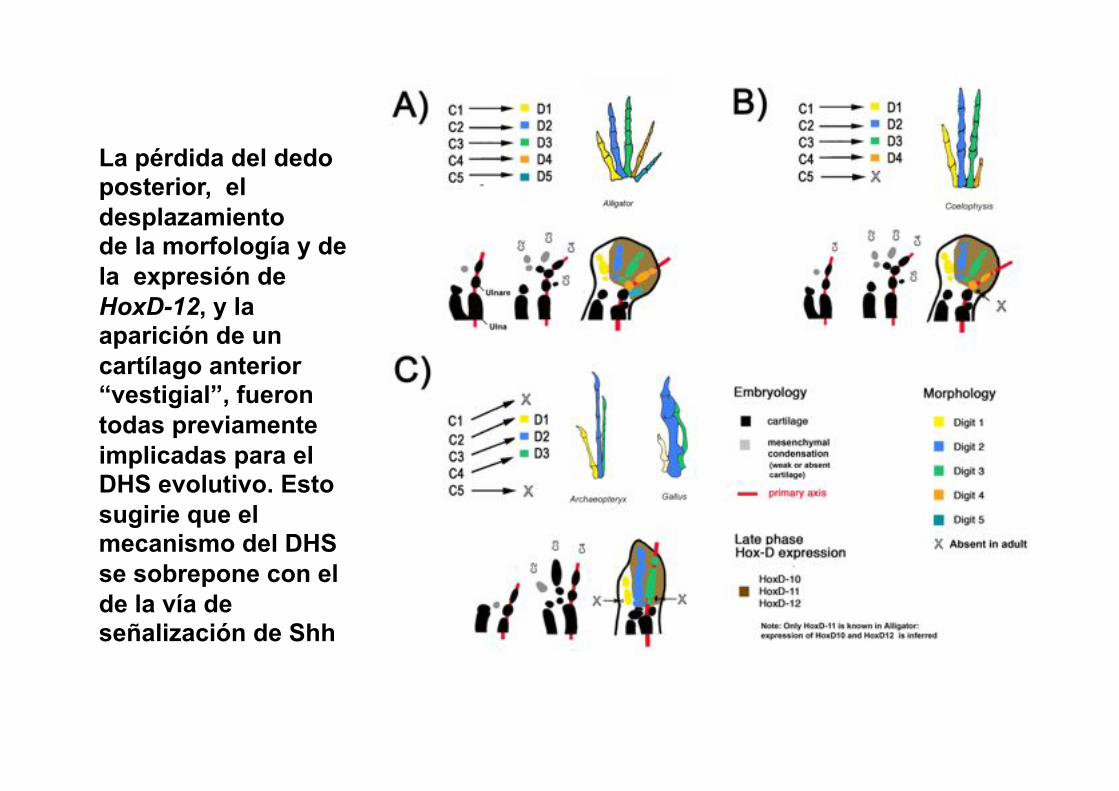
La pérdida del dedo posterior, el desplazamiento de la morfología y de la expresión de HoxD-12, y la aparición de un cartílago anterior “vestigial”, fueron todas previamente implicadas para el DHS evolutivo. Esto sugirie que el mecanismo del DHS se sobrepone con el de la vía de señalización de Shh

Conclusiones
La ausencia de HoxD-12, HoxD-11 y HoxD-10 en dedo 1 es ancestral para amniotos y conservada en ratón y pollo
El caso del ala de las aves ejemplifica cómo el origen embriológico puede no ser infalible como indicador de homología (pese a que generalmente funciona)
Como previamente no se habían descrito desplazamientos homeóticos en serie experimentales, la aplicación de cyclopamina confirma una posibilidad mecanística que previamente había sido inferida mediante comparaciones evolutivas.
(La naturaleza no hace distinción entre ciencias experimentales y comparativas; los mismos mecanismos se ven reflejados en ambas)



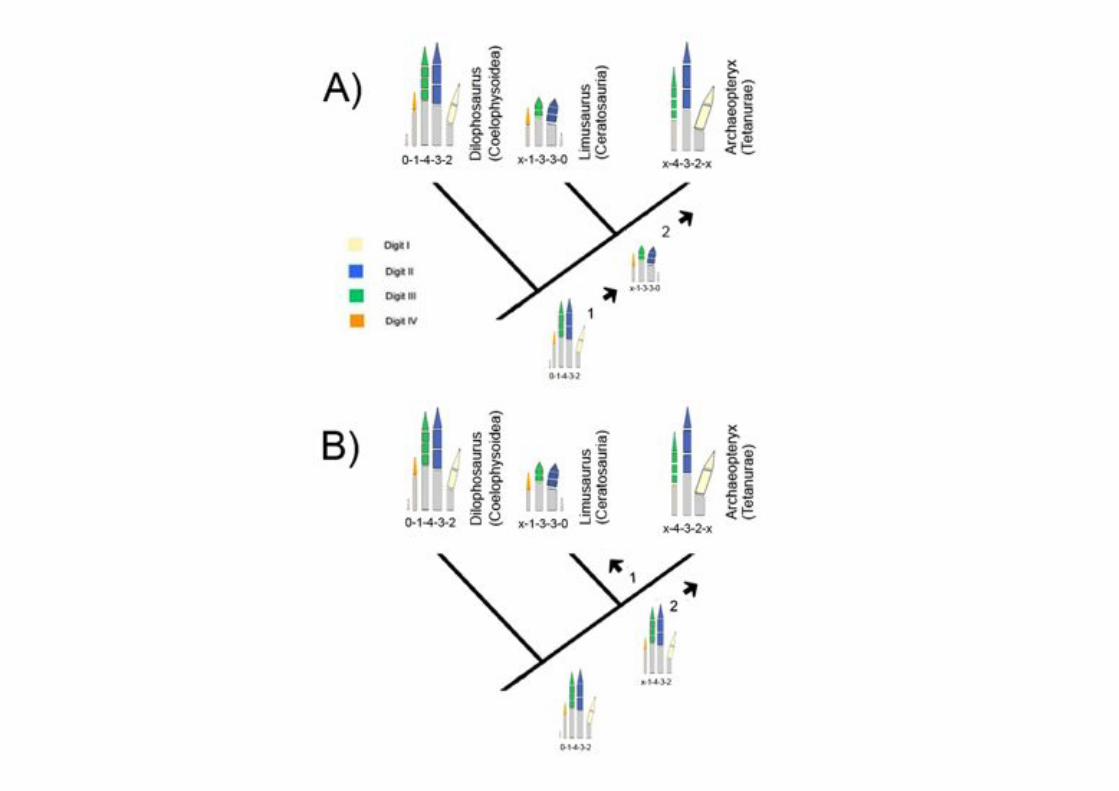

Actuales investigaciones de Vargaslab en el Departamento de Biología:
Deriva fenogenética: Cambio de la expresión de genes Tbx en un dedo 4 que
no cambia de morfología

Pese al descubrimiento de estos dinosaurios emplumados, persiste aún
una objeción importante, publicable en evistas académicas, y que guarda relación con la identidad de los dedos del ala de las aves.
De acuerdo a este argumento (La “objeción embriológica) las aves poseen
tres dedos en el ala que corresponden a los dedos índice , medio, y anular (2,3, y 4), mientras que los tres dedos de la mano de los dinosaurios son el
pulgar, índice y medio (1, 2y 3)
Este es el único actualmente argumento publicable en revistas científicas
para decir que las aves no son dinosaurios

Hinchliffe 2002
Gilbert 2002

Por lo tanto la paleontología y la biología del desarrollo parecen
ofrecer información conflictiva respecto a si los dedos de
la mano de las aves son 1,2 y 3 ó 2, 3 y 4.


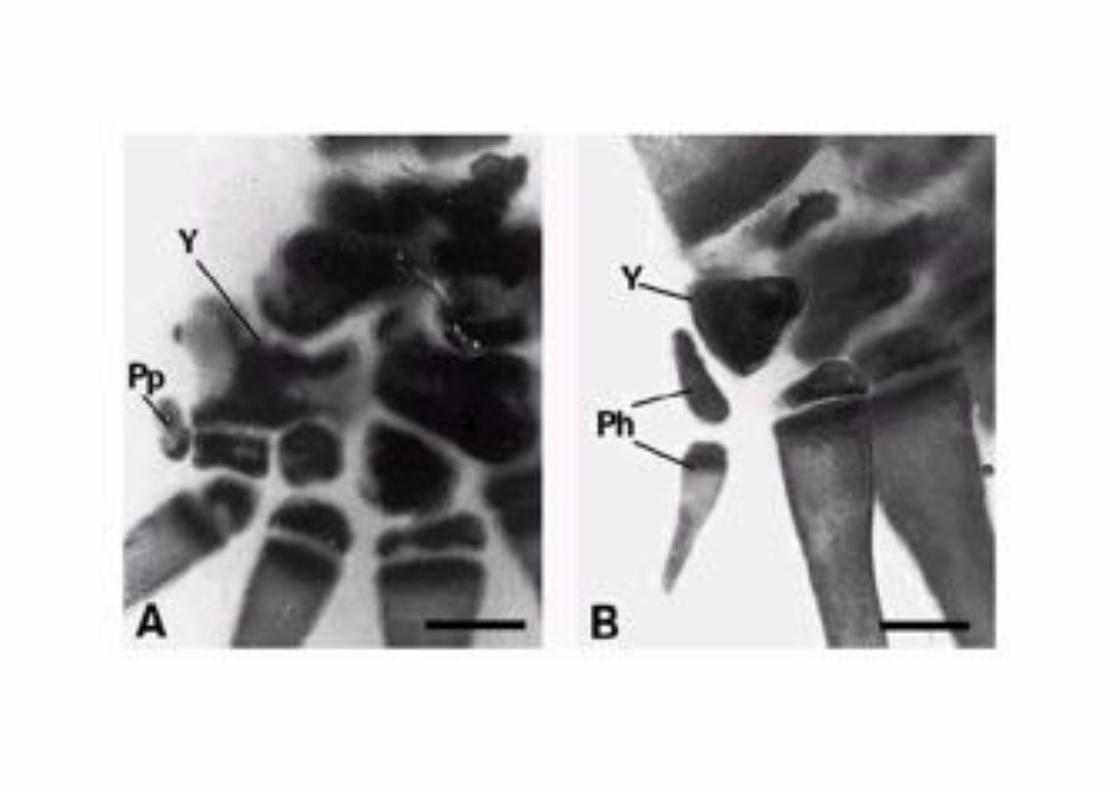

El argumento más perdurable y reconocido en contra del origen de las aves a partir de los
dinosaurios ha sido la identificación 2, 3, 4 de los dígitos del ala. La información aquí
presentada no favorece este argumento y consolida el hecho de que las aves son
representantes vivientes de los dinosaurios

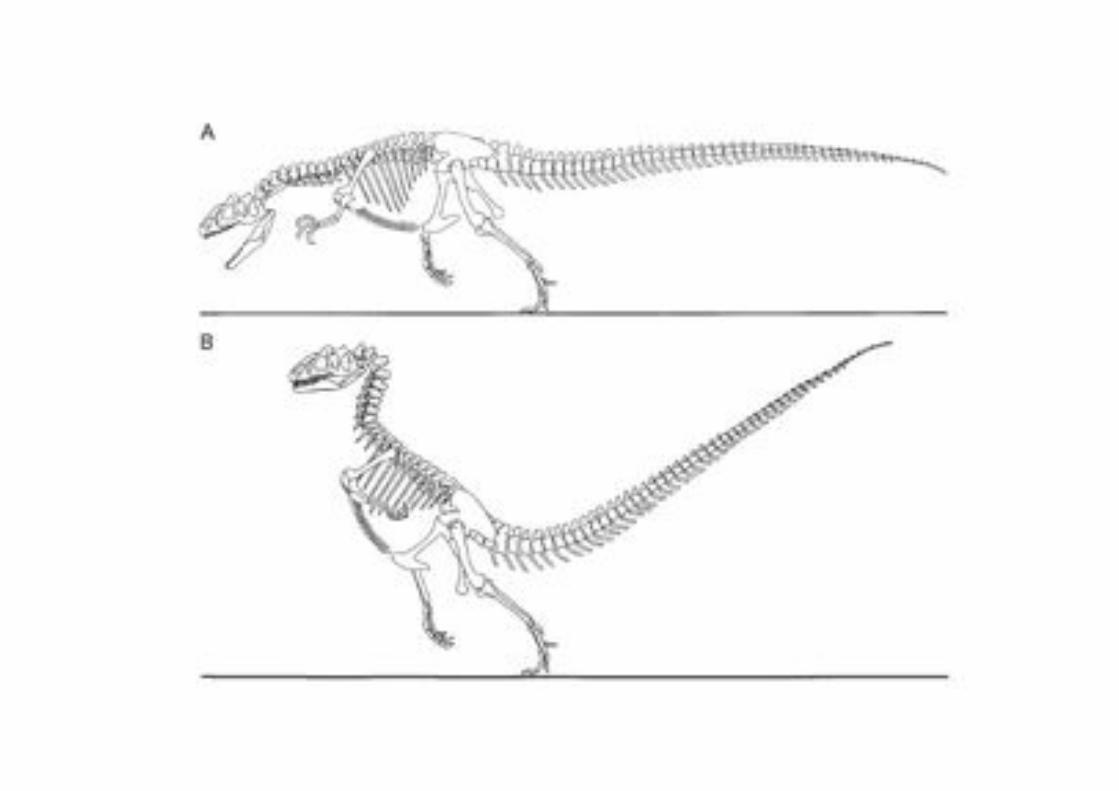
La transformación de dinosaurio en Ave Las aves presentan muchos rasgos que es posible describir cómo se adquirieron a
partir de los dinosaurios.
En las aves,la pantorrilla difiere de la de un dinosaurio por que el hueso de la fíbula se ve reducido en tamaño y su extremo distal no se contacta con el tarso (tobillo)
Importantemente, este cambio se ve
“recapitulado” en el desarrollo embrionario de las aves

Alargamiento de la pierna bajo la rodilla

Archer et al (1984) Afirman que la fíbula pierde su extremo distal (epifisis)
distal en el desarrollo, con la consecuente reducción del crecimiento
longitudinal hacia distal (“reducción” de la fíbula). Adicionalmente, plantean
que el trozo desprendido corresponde al hueso calcáneo (fibulare) del tobillo.

Archer describe un plano de células aplanadas (zona transparente)
cerca de la epifisis fibular y afirma que especifica el futuro plano de
fractura

Sin embargo, en la mayoría de los tetrápodos (con la excepción de
los urodelos) y en todos los amniotos, el calcáneo se presenta
como una condensación cartilaginosa independiente desde
estadios muy tempranos que se observa previo a las
condensaciones cartilaginosas del arco digital (Siguiendo la
secuencia de condrificación del “eje metapterigio”)

¿Qué ocurría en el desarrollo de un dinosaurio?
Si el fibulare se desprendía de la fíbula, ¿Porqué no disminuía
el crecimiento longitudinal de la fíbula?
Si se origina de forma diferente ¿Es homólogo el calcáneo de aves
al de otros tetrápodos?

Mediante diafanizados y cortes histológicos, observé que el calcáneoen el pollo y
en el pato se desarrolla como en cualquier tetrápodo. En el caso del pollo puede
ser observado como un elemento independiente entre los estadios 26 al 28, previo
al supuesto origen por desprendimiento propuesto por Archer
Pollo E26 Pato 6 días

E26
E27
E28

Resultados:
El fibulare (calcáneo) de las aves no se origina a partir de la epífisis
distal de la fíbula cartilaginosa, sino que inicia la condrogénesis
como un elemento independiente (E25) .Ulteriomente se aproxima
mucho a la fíbula (E28-29), momento en que se ha deescrito un
repentino alargamiento de la fíbula (Hall 1975, Archer 1984). La
zona de células aplanadas descrita por Archer et al. probablemente
corresponde a la interfase entre los pericondrios de ambos huesos.
Ulteriormente (E30) se pierde la proximidad física entre fíbula y
fibulare, interpretada erróneamente por Archer como el
desprendimiento de la epifisis distal de la fíbula. El fibulare despues
es incorporado por fusión al tibiotarso.

Alargamiento del coracoides y
evolución del músculo sterno-coracoideo

Fusión y expansión de las placas esternales

Unión, alargamiento y
cambio de ángulo entre
las clavículas


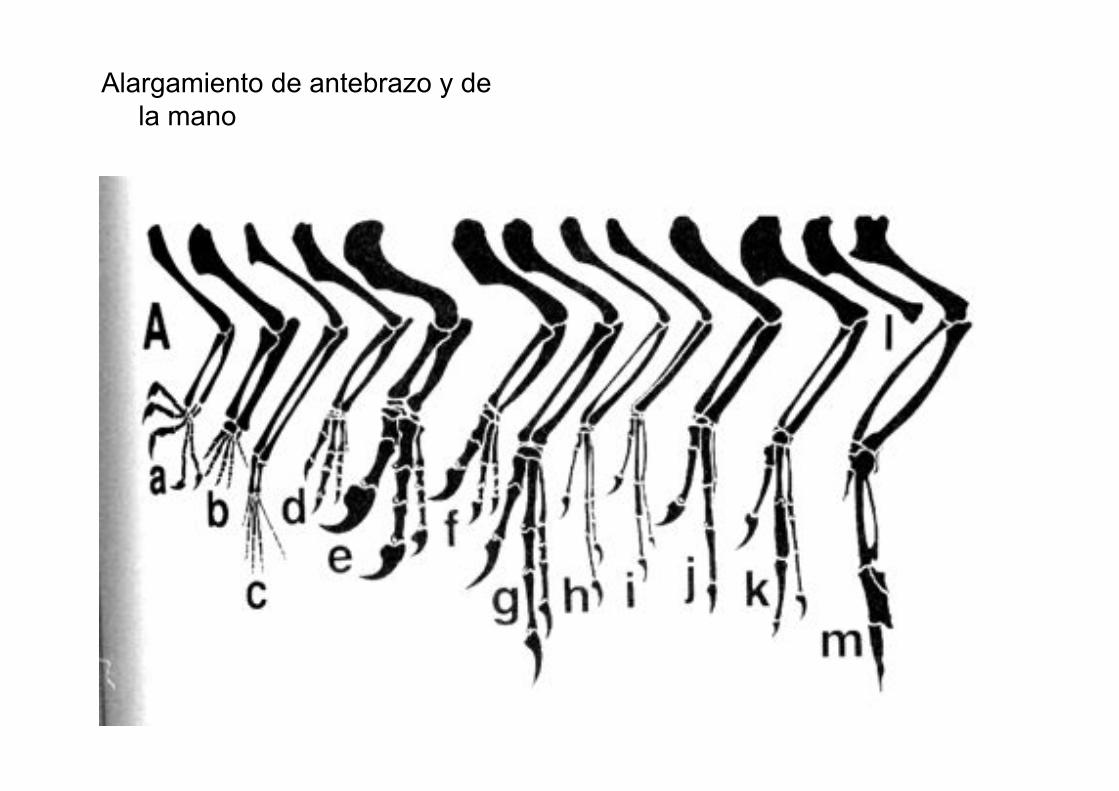
Alargamiento de antebrazo y de
la mano
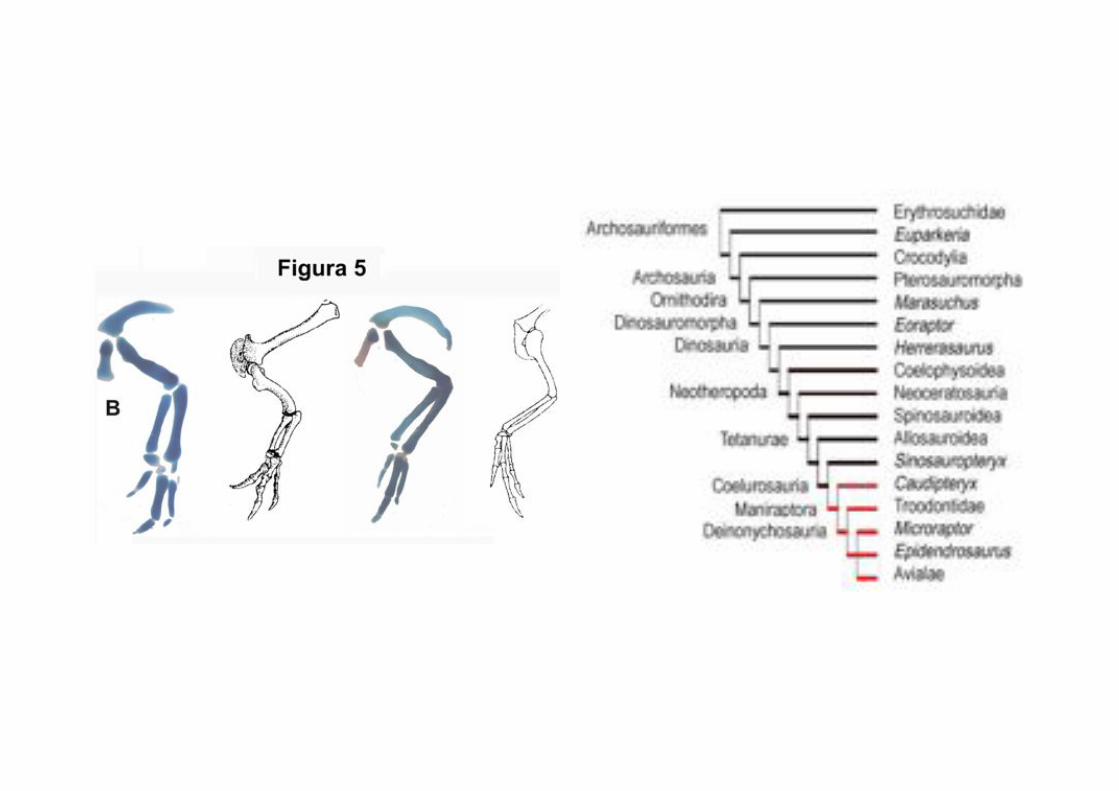


Metacarpo 1 reducido, no
fusionado al metacarpo2


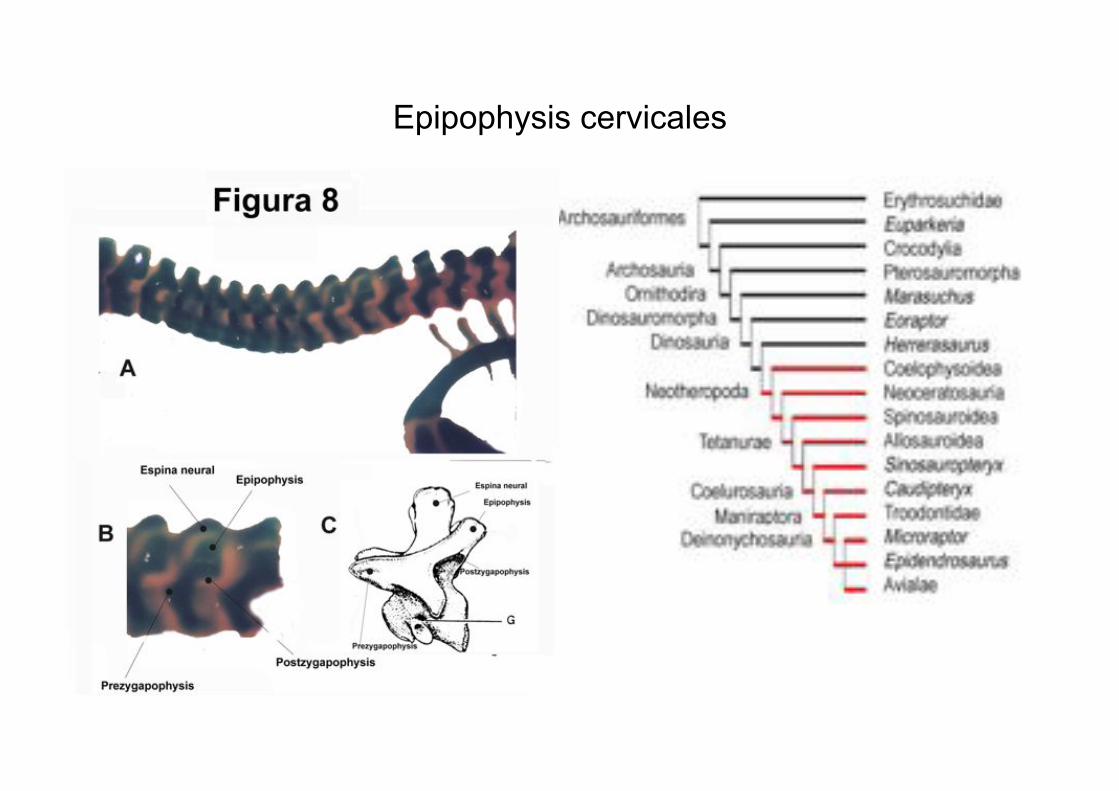
Epipophysis cervicales

Metatarso V reducido, no fusionado al tarso

Alargamiento de la pierna bajo la rodilla


Presencia de cresta cnémica

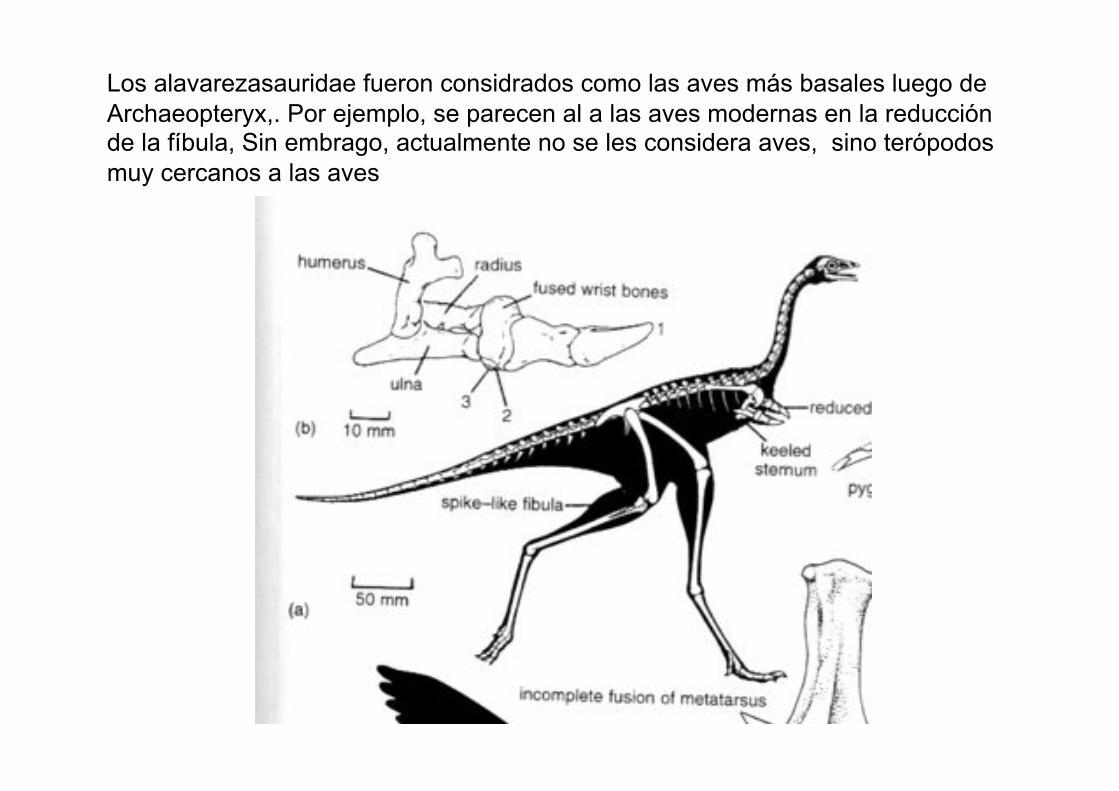
Los alavarezasauridae fueron considrados como las aves más basales luego de
Archaeopteryx,. Por ejemplo, se parecen al a las aves modernas en la reducción de la fíbula, Sin embrago, actualmente no se les considera aves, sino terópodos
muy cercanos a las aves


Rahonavis ostromi
Presenta la garra en forma de Hoz característica de los Deinonychosauria
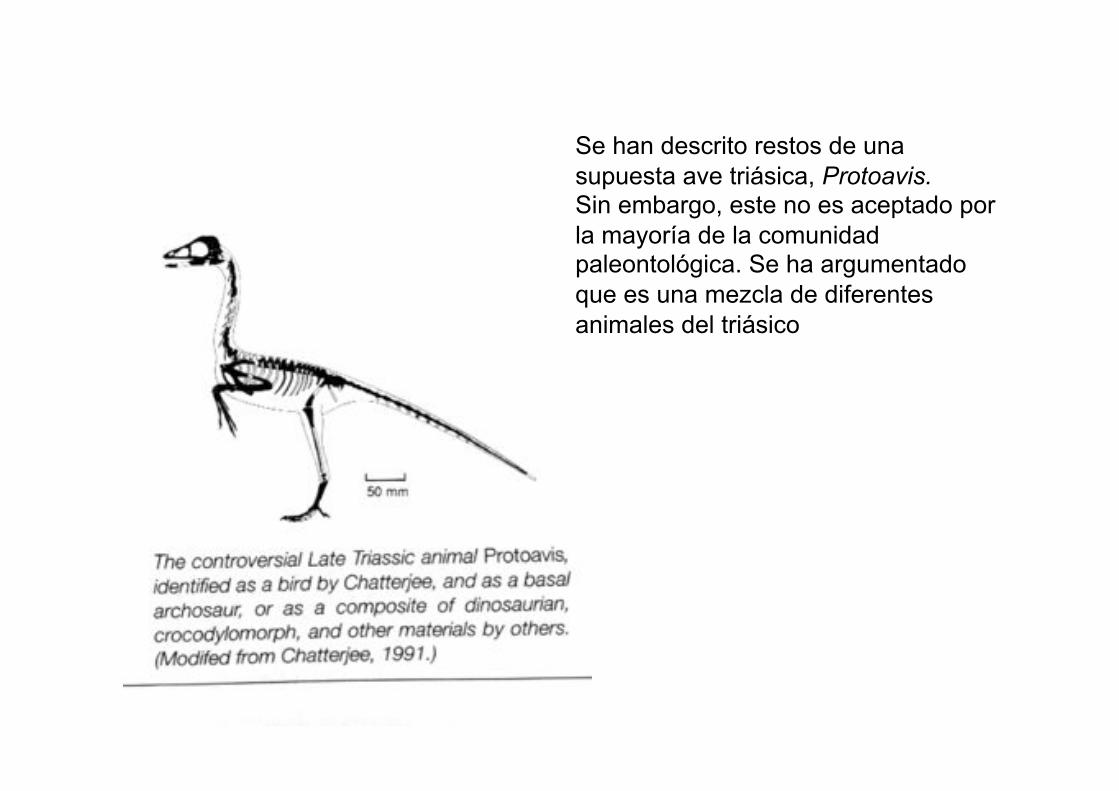
Se han descrito restos de una
supuesta ave triásica, Protoavis. Sin embargo, este no es aceptado por
la mayoría de la comunidad paleontológica. Se ha argumentado
que es una mezcla de diferentes
animales del triásico
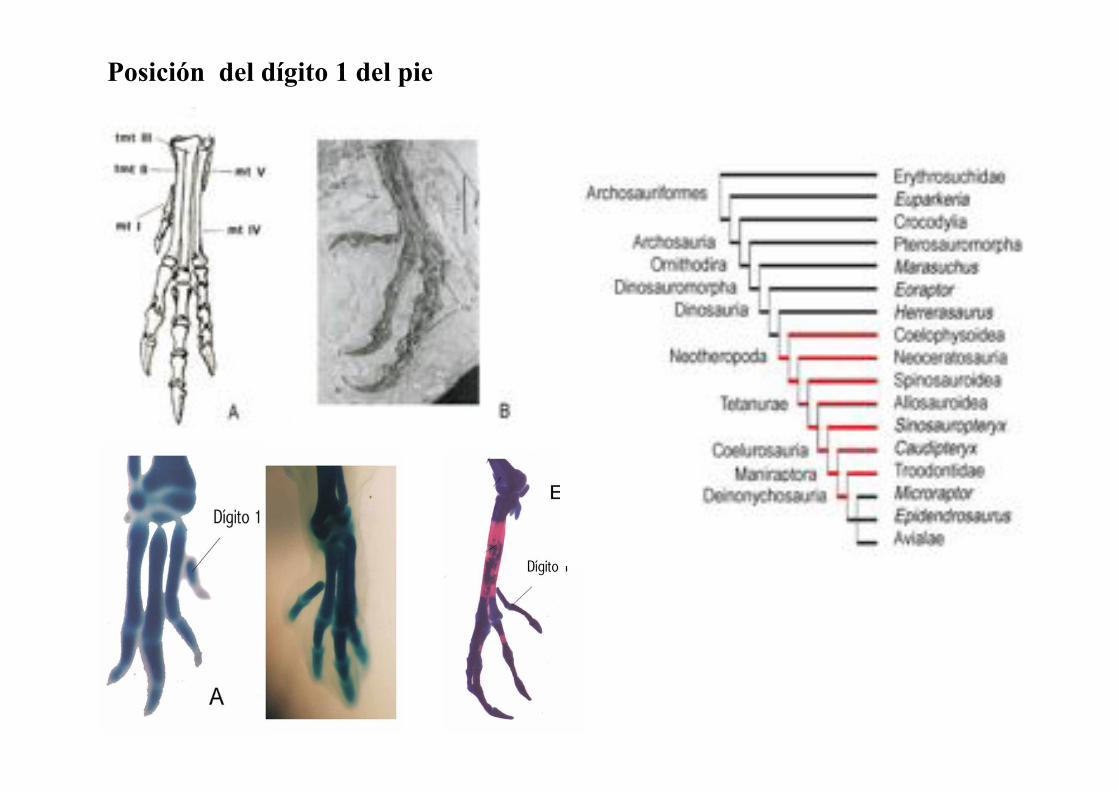
Posición del dígito 1 del pie

dígito 1 oponible

Para distinguir si este cambio de posición del dígito 1 es el resultado del
desplazamiento del dígito o más bien del mayor crecimiento hacia proximal de
los metatarsos, efectué inyecciones del colorante lipofílico fluorescente DiI en
la región proximal, media y distal de los metatarsos en desarrollo del pie de
pollos de 6 y 7 días de incubación. Cada sitio de inyección fue dibujado con una
camara lucida. Luego se fotografió el lugar en que se detectaron las celúlas
marcadas al día 9 del desarrollo, cuando el dígito 1 presenta una ubicación
distal


La orientación del pubis es cada vez más hacia
atrás, en la ontogenia y filogenia

Las aves poseen sacos
respiratorios, verdaderas extensiones de los pulmones
hacia distintas regiones corporales, incluynedo el interior
de huesos (no mostrado en la
figura) El aire ingresa hacia los sacos posteriores y luego hacia
los pulmones y sacos más anteriores, generando una
unidireccionalidad del flujo aéreo
completamente única a las aves.

El primer metacarpo, grande e independiente en Archaeopteryx, se hace pequeño
y se fusiona completamente al segundo metacarpo en aves actuales, presentando además un proceso de la musculatura extensora de la mano

En aves modernas frecuentemente se
describe que el calcáneo posee un proceso ascendente. Esto se considero
incompatible con el tobillo de dinosaurios terópodos, en los cuales el
astrágalo es el que proyecta un
proceso ascendente. Sin amebargo, una examinación de la emvrología
indica que en realidad este proceso ascendente no pertenence al
calacáneo, sino que al astrágalo, como
en dinosaurios
Deinonychus Archaeopteryx
Hesperornithiforme
Hoatzin


Pygostilia: Confuciusornithidae +
Ornithotoraces Poseen pygostilo, esternón osificado
con quilla Confuciusornithidae Poseen un pygostilo, pero el Tercer
dedo de la mano aún posee 4 falanges Destaca la pérdida de los dientes,
adquirida independientemente de las aves actuales (Neornithes)
Hallux oponible muy bien desarrollado

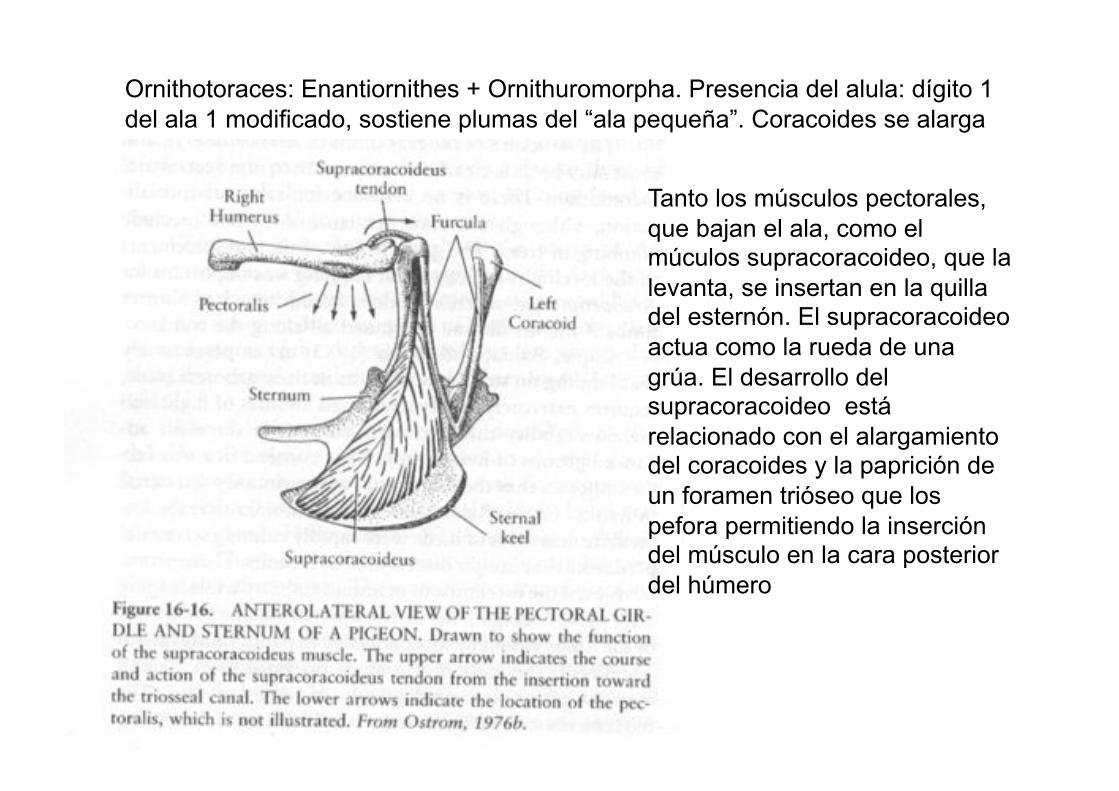
Tanto los músculos pectorales,
que bajan el ala, como el múculos supracoracoideo, que la
levanta, se insertan en la quilla del esternón. El supracoracoideo
actua como la rueda de una
grúa. El desarrollo del supracoracoideo está
relacionado con el alargamiento del coracoides y la paprición de
un foramen trióseo que los
pefora permitiendo la inserción del músculo en la cara posterior
del húmero
Ornithotoraces: Enantiornithes + Ornithuromorpha. Presencia del alula: dígito 1
del ala 1 modificado, sostiene plumas del “ala pequeña”. Coracoides se alarga

Las Enanthiornithes, a diferencia de Confuciusornithidae, eran dentadas, y habpainr educido el número de falanges en el dígito 3 de la amno, de 4
falanges, a una sola. Sin embargo, retienen los dígitos 1 y 2 con 2 y 3 falanges, las terminales siendo las falanges unguales (garras)
Poseen un metatarso IV muy delgado en el pie, los extremos más distales de
los metatarsos no se fusionan.

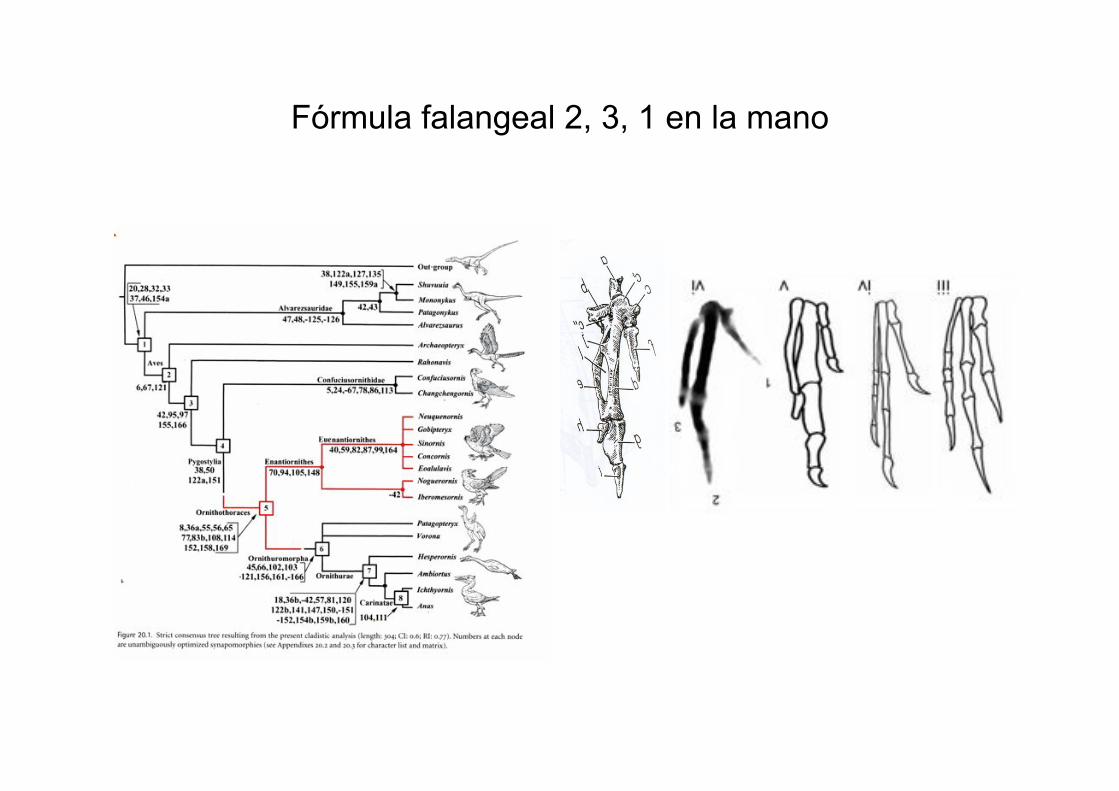
Fórmula falangeal 2, 3, 1 en la mano
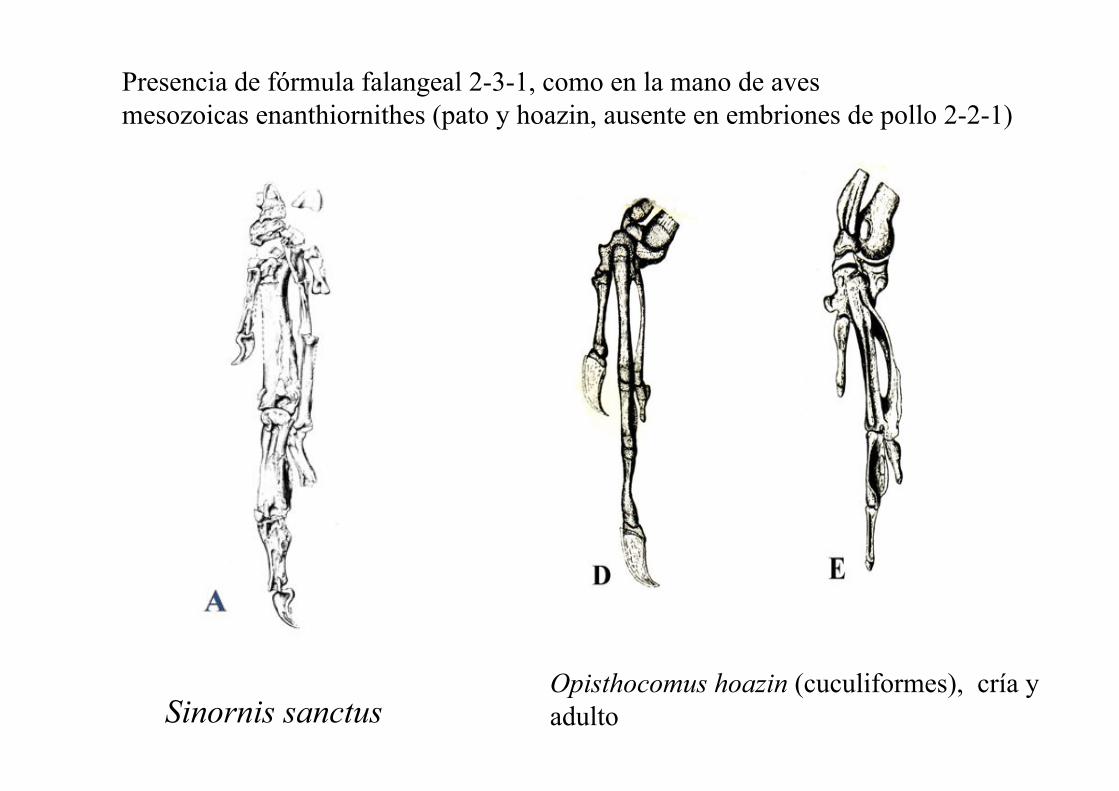
Presencia de fórmula falangeal 2-3-1, como en la mano de aves
mesozoicas enanthiornithes (pato y hoazin, ausente en embriones de pollo 2-2-1)
Sinornis sanctus Opisthocomus hoazin (cuculiformes), cría y
adulto

Anas platirrhynchos, 12.5 días de
incubación y adulto Gallus gallus

Gran tamaño del acetábulo en enantiornithes y el embrión del pollo, en
contratse con el pequeño tamaño del acetábulo en adultos


Ornithuromorpha:
Comparten una mano más similar a la de las aves actuales (neornithes) la
presencia de metacarpos más extensamente fusionados entre s{i
Patagopteryx deferrerasii
Es un ave terrestre del cretácico que ya había perdido la capacidad de volar

Ornithurae: Hesperornis + Carinatae
Poseen acetábulos reducidos
Hesperornothiformes: Formas marinas
dentadas con brazo vestigializado
Carinatae: Presentan mucho mayor desarrollo
de la quilla y una cabeza proxinal del húmero
de forma globular
Dea¿stacan los Ichthyornitidae, marinos,
voladores y dentados

Neornithes: Representan a todos los descendientes del ancestro en común más
rciente de todas las aves actuales. Son Carinatae que han perdido por completo los dientes. Se pierden defeinitivamente las costillas ventrales (gastralia). El
húmero es hueco y presenta una fosa para el ingreso de un saco pulmonar
Las Neornithes se pueden dividr en
dos grupos que se reconocen con facilidad por sus paladares.
Paleognathae: El paladar presenta un vomer grande firementente unido al
pterygoide, no hay articulación entre
pterygoideo y palatino, y hay una articulación movible entre el
pterygoideo y la base de la caja craneana.
Neognathae: paladar más suelto y
móvilLos vomners estan reducidos o perididos por completo, hay una
articulación movible entre el pterygoideo y el palatino, y se pierde
la articulación de pterygoideo con la
caja craneana.

Paleognatha:
Tinamiformes: Tinamúes Apterygiformes: Kiwis, Moas Struthioniformes: Avestruces Rheiformes: Ñandúes Casuariiformes: Casuarios y Emúes

Las Paleognathae son de hábitos terrestres y son pobres voladoras o no vuelan
en absoluto. Debido a esto poseen una quilla esternal ausente o muy pobremente desarrollada, lo cual las diferencia de Neognathae

Los avestruces han perdido por
completo el dígito 1 y 2 y corren sobre dos dedos, el 3 y el 4. El
metatarso 2 está drásticamente reducido.
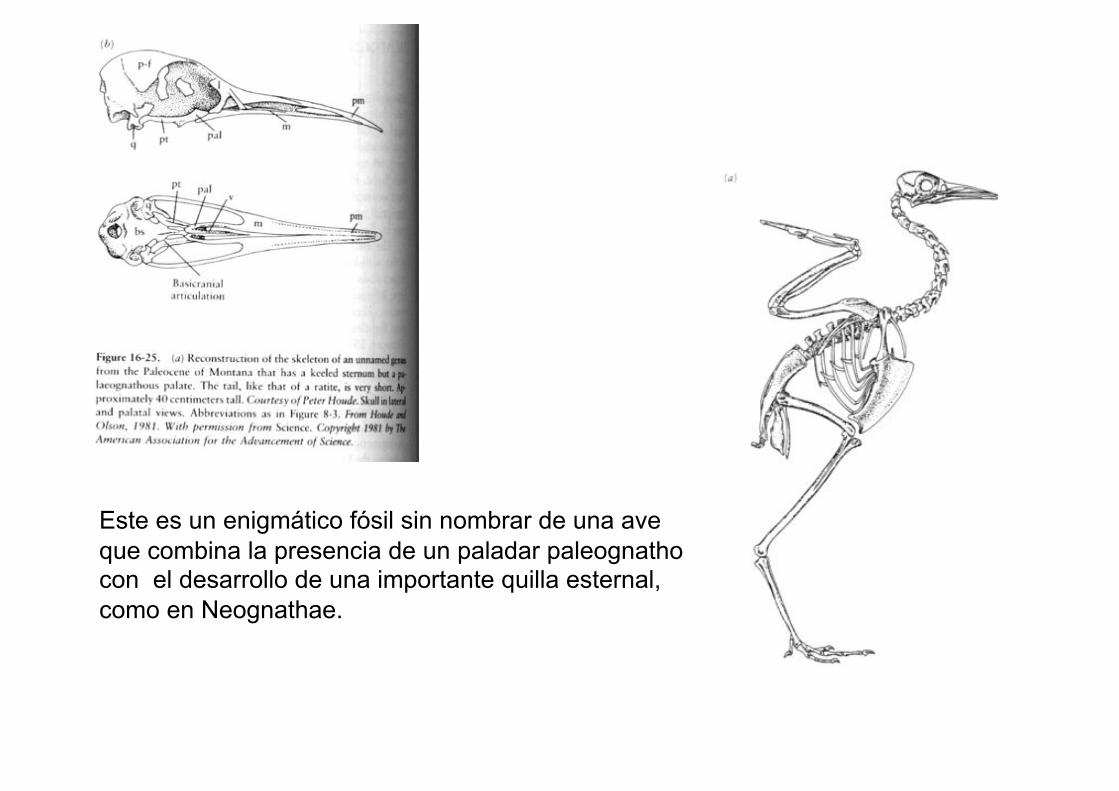
Este es un enigmático fósil sin nombrar de una ave
que combina la presencia de un paladar paleognatho con el desarrollo de una importante quilla esternal,
como en Neognathae.

Neognathae

Neognatha
Galliformes: pollo, pavo, faisan, Urogallos
Anseriformes: Patos y Cisnes Podicipedifomes: Somormujos
Gaviiformes: Colimbos
Spheniscifomes: Pingüinos Pelecaniformes: Rabijuncos, pelícanos, Cormoranes, fragatas,
Piqueros Procelariformes: Albatroses, Petreles
Gruiformes: Grullas, rascones, pollas de agua y fochas
Charadriformes: Gaviotas, Flamingos, Ostreros, Chorlitos, Zarapitos, Phorhusracidae, Alcas, Frailecillos, etc.
Ardeidae : Avetorillos, según otros, garzas también Columbiformes: palomas y tórtolas
Ciconiiformes: Garzas, Buitres del nuevo mundo
Falconiformes: Halcones, águilas, y buitres del viejo mundo Strigiformes: búhos
Caprimulgiformes: Nictibios, Egotelos Apodiformes: Picaflores y vencejos, otros incluyen chotacabras
Coracciformes: calaos, martín pescadores
Piciformes: carpinteros, tucanes Passeriformes: Aves cantoras

Galloanserae: Galliformes y Anseriformes

Columbiformes
Didus
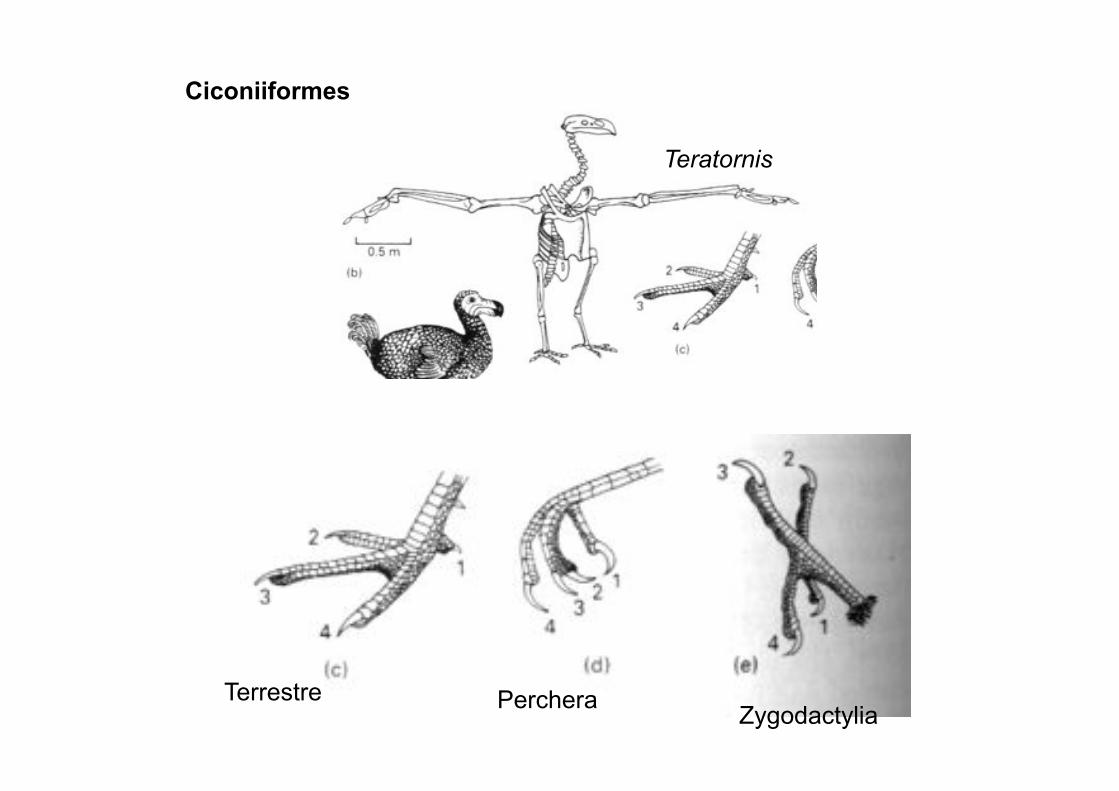
Ciconiiformes
Terrestre Perchera Zygodactylia
Teratornis

Charadriformes: Un grupo con una gran diversidad de estilos de Vida acuales: Flamingos, aves playeras, gaviotas,
Pingüinos falsos (Auk), y extintos grandes Depredadores terrestres como Phorusrhacidae

Sphenisciformes
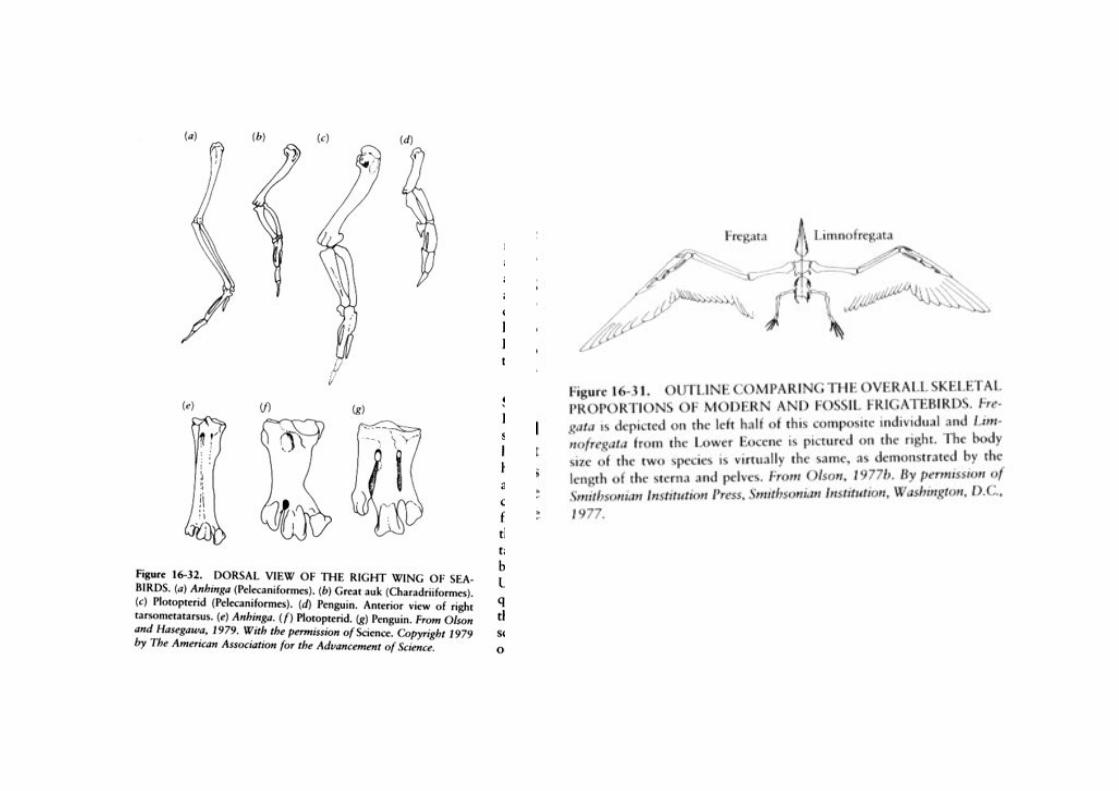
Pelecaniformes: Plotopteridae, Familia extinta
convergente con los pingüinos

Pelecaniformes: Pelagornithidae (Pseudodontornes)