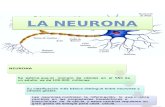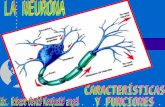
Organización Funcional de La Neurona
41
Organización funcional de la neurona 1. Cuerpo celular 2. Dendritas 3. Núcleo 4. Aparato de Golgi 5. Cono axónico 6. Cuerpos de Nissl 7. Mitocondria 8. Axón mielínico 9. Célula de Schwan 10. Nódulo de Ranvier 11. Colateral del axón 12. Telodendro 13. Botones terminales Las características estructurales de una neurona típica están dadas por las que presentan sus tres componentes básicos: el soma o pericarion, las dendritas y el axón. Sin embargo, existe una amplia variedad de formas y tamaños que dependen del soma y de los procesos neuronales mencionados. Así, el tamaño del soma varía entre los 6-8m (células granulosas del cerebelo) y los 60-80 m (células de Purkinje también en el cerebelo). En general, la morfología de las neuronas, igual que la de las células gliales, es extendida lo cual
-
Upload
lucia-lopez -
Category
Documents
-
view
70 -
download
3
description
como esta organizada la neurona
Transcript of Organización Funcional de La Neurona
Organizacin funcional de laneurona
1. Cuerpo celular2. Dendritas3. Ncleo4. Aparato de Golgi5. Cono axnico6. Cuerpos de Nissl7. Mitocondria8. Axn mielnico9. Clula de Schwan10. Ndulo de Ranvier11. Colateral del axn12. Telodendro13. Botones terminales
Las caractersticas estructurales de una neurona tpica estn dadas por las que presentan sus tres componentes bsicos: el soma opericarion,las dendritas y el axn. Sin embargo, existe una amplia variedad de formas y tamaos que dependen del soma y de los procesos neuronales mencionados. As, el tamao del soma vara entre los 6-8m (clulas granulosas del cerebelo) y los 60-80m (clulas de Purkinje tambin en el cerebelo). En general, la morfologa de las neuronas, igual que la de las clulas gliales, es extendida lo cual refleja una forma de adaptacin en clulas cuya funcin depende de las mltiples interacciones que puedan establecer.El cuerpo neuronal se encuentra rodeado de una membrana de alrededor de 7.5 nm de grosor, la membrana plasmtica. El citoplasma neuronal presenta una serie de sistemas membranosos (ncleo, retculo endoplasmtico, sistema de Golgi) que constituyen organelos y que, a pesar de estar conectados entre s, tienen caractersticas enzimticas especficas. En l se encuentran, adems, otros componentes como los lisosomas, grnulos de lipofucsina, mitocondrias, vesculas y complejos vesiculares, neurofilamentos, neurotbulos y ribosomas.Una de las caractersticas importantes de la neurona es organizacin membranosa.Las protenas neuronales adems de ser fundamentales para las funciones de estas clulas determinan la alta especificidad funcional de sus estructuras membranosas. As, ellas forman parte de sitios funcionales especficos en la membrana de organelos subcelulares, en receptores qumicos y en canales inicos.Los sitios especficos de las membranas son esenciales para el reconocimiento neuronal, tal como los receptores y los canales inicos lo son para la comunicacin y la excitabillidad neuronales. Muchas de las protenas que se ubican en las membranas de los organelos son esenciales para las interacciones que se dan entre ellos.Topogrficamente, las protenas se ubican en el citosol (protenas fibrilares y enzimas), en la membrana plasmtica y en la de organelos, como las mitocondrias y el ncleo.Protenas citoslicas. Representa uno de los grupos que tiene mayor abundancia de protenas. En l se distinguen: las protenas fibrilares: son las que constituyen el citoesqueleto (los neurofilamentos) y entre ellas se encuentran la tubulina, la actina y sus protenas asociadas. Representan alrededor de un 25% de las protenas totales de la neuronas. Enzimas: catalizan las reacciones metablicas de las neuronas.Lasprotenas citoslicasse forman en los poliribosomas libres o polisomas, ubicados en el citoplasma neuronal, cuando el mRNA para esas protenas se une a los ribosomas. En relacin a estas protenas hay que considerar a otra protena pequea, laubiquitina, que se une residuos de lisina de las protena para su posterior degradacin.Protenas nucleares y mitocondriales. Tambin se forman en los polirribosomas y luego son enviadas al ncleo o a las mitocondrias, donde existen receptores especficos a los que se unen para incorporarse al organelo, por el proceso detraslocacin. El mecanismo por el que se incorporan las protenas despus de su sntesis, es laimportacin post-transduccin.Hay dos categoras de protenas de membranas: lasprotenas integrales: se incluyen en este grupo los receptores qumicos de membrana (a neurotransmisores, a factores de crecimiento). Ellas estn incertadas o embebidas en la bicapa lipdica o estn unidas covalentemente a otras molculas que s atraviesan la membrana. Una protena que atraviesa la membrana y que ofrece un grupo N-terminal, hacia el espacio extraneuronal, es designada como deltipo I. Las hay tambin deltipo IIque son aquellas en que el grupo N-terminal se ubica en el citosol. Lasprotenas perifricas: se ubican en el lado citoslico de la membrana a la cual se unen por asociaciones que hacen con los lpidos de la membrana o con las colas citoslicas de protenas integrales o con otras protenas perifricas (protena bsica de la mielina o complejos de protenas).Las protenas de la membrana plasmtica y las de secrecin se forman en los polirribosomas que se unen al retculo endoplasmtico rugoso. Ellos constituyen un material de naturaleza basfila (se tien con colorantes bsicos como el azul de toluidina, el violeta de cresilo y el azul de metileno) que al microscopioi ptico se han identificado como la substancia de Nissl. Una vez que las protenas formadas en este sistema pasan al interior del retculo, ellas son modificadas por procesos que se inician el retculo y que continuan en el sistema de Golgi y an, posteriormente, en los organelos finales a donde son destinadas (vesculas de secrecin).Las protenas que son componentes de las membranas abandonan el retculo en una variedad de vesculas. Adems de las de secrecin, son muy importantes para las neuronas, las vesculas sinpticas. A travs de ambos tipos de vesculas las protenas son enviadas al espacio extraneural por la va constitutiva o la va regulada.1. Cuerpo celular2. Axn3. Ncleo4. Citoplasma5. Retculo endoplasmtico rugoso6. Retculo endoplasmtico liso7. Ribosomas8. Poro nuclear9. Lisosoma10. Vesculas secretoras11. mRNA12. Polisoma13. Protena14. Mitocondria15. Sistema de Golgi
Hay unas pocas proteinas que son sintetizadas en el genoma mitocondrial. La mayora de las proteinas de una neurona generan proteinas en el soma neuronal.Las seales que inducen la sntesis de nuevas molculas de un tipo dado de protena actan a nivel del ncleo donde se genera, como respuesta, un aumento del mRNA especfico para esa protena. Para las proteinas citoslicas este mensajero sale del ncleo por un poro para unirse en el citosol a ribosomas y constituir los llamados polisomas o polirribosomas. Es en esta estructura que el mensaje del mRNA se transforma en proteina. Por este mecanismo se forman las proteinas fibrilares, las enzimas y tambin proteinas que se incorporarn al ncleo y a mitocondrias. Las proteinas que van a incorporarse a molculas de secrecin se forman en polisomas que se unen a las membranas del retculo endoplsmico rugoso.El mRNA contiene la informacin que cuando es traducida y expresada en la secuencia polipeptdica determina el destino de la nueva protena formada: a la membrana plasmtica, a una vescula, etc..Desde el retculo endoplsmico, las protenas recin sintetizadas se mueven al Golgi donde son almacenadas en grnulos o vesculas secretoras que, posteriormente, se separan del Golgi y al llegar a la membrana pueden liberar su contenido por exocitosis.1. 13 protofilamentos forman un microtbulo2. Cada protofilamento est formado por unidadesyde tubulina que se alternan ordenadamente3. Microtbulo4. Neurofilamento formado por tres protofibrillas enrolladas una sobre la otra5. Cada protofibrilla est formada por dos protofilamentos6. Protofilamento. Es un filamento formado por 4 dmeros que forman un complejo tetramrico7. Complejo tetramrico8. Dmero formado por dos monmeros9. Monmeros formados de especies de citoqueratina, una variedad de protena10. Microfilamento formado por monmeros de actina globular. Dos hebras de monmeros se enrollan una sobre la otra. Cada microfilamento tiene alrededor de 7nm de dimetro
Esta formado por tres estructura de tipo fibrilar, de diferente dimetro: los microtbulos, los neurofilamentos y los microfilamentos. A ellos se asocian otras protenas. Cada tipo de estos filamentos corresponden a un polmero formado por un nmero variable de molculas de un monmero. En el caso de los microtbulos es latubulina, en el de los neurofilamentos es la citoqueratina y en el de los microfilamentos, la actina globular.El citoesqueleto cumple las siguientes funciones en la neurona: mediar el movimiento de organelos entre diferentes regiones de la neurona fijar la ubicacin de determinados componentes de la membrana, por ejemplo receptores qumicos, en los sitios adecuados. determinar la forma neuronal.Microtbulos: se presentan como las fibras de mayor dimetro del citoesqueleto (25 a 28 nm de dimetro externo) Cada fibra se presenta como un cilindro cuya pared est formada por 13 estructuras alargadas o protofilamentos de unos 5 nm de dimetro. Cada protofilamento est constituido por monmeros de subunidadesydetubulina, que se alternan en la estructura.Neurofilamentos: (o neurofibrillas o filamentos intermedios en otros tipos de clulas): tienen alrededor de 10 nm de dimetro, son los ms abundantes y representan el soporte del citoesqueleto. Cada neurofilamento est constituido por monmeros tambin organizados como estructuras filamentosas. Pero dos monmeros se unen, enrollados uno alrededor del otro para constituir un dmero. A su vez, dos dmeros enrollados uno en el otro, constituyen una fibra de mayor grosor, el complejo tetramrico. La unin de varios de estos complejos forman el protofilamento y dos protofilamentos forman la protofibrilla. Tres protofibrillas enrolladas constituyen el neurofilamento.Microfilamentos: Son polmeros en forma de filamentos de 3 a 5 nm de dimetro que estn formados por monmeros de actina globular, donde cada monmero tiene ATP o ADP. Cada microfilamento est formado por dos hebras de actina enrolladas en forma de hlice, es decir, una sobre la otra.1. Estructura filamentosa formada por la agregacin de monmeros que entran o salen en cada extremo de la fibra. Sistema en equilibrio porque el nmero de monmeros que se incorporan es igual al nmero de monmeros que se pierden2. Monmero3. Polmero4. Otra forma de equilibrio en que el nmero de monmeros que entran por un extremo es igual al nmero de los que se pierden por el otro5. Forma en que puede crecer un polmero6. Forma en que puede disminuir la longitud de un polmero
Las neuronas sintetizan los monmeros con los cuales construyen las estructuras filamentosas del citoesqueleto. Ello es posible por un proceso de automontaje que no requiere de energa inicial si no que es el resultado de un equilibrio entre la concentracin de monmeros y la de polmeros, configuracin esta ltima que es ms estable que la de los monmeros.El polmero, la hebra, se va formando por la agregacin de monmeros que ocurre en ambos extremos de ella. Pero tambin puede ocurrir en esos sitios, prdida de ellos. As, la mantencin de la estructura filamentosa de un largo dado, resulta de un verdadero equilibrio dinmico.Pero tambin puede haber unapolimerizacin polarizadaen la cual se aaden monmeros por uno de los extremos mientras se pierden por el otro (polimerizacin en molino de rueda o direccional). Este fenmeno ocurre cuando se introduce una perturbacin energtica en el sistema. Por ejemplo, cuando el ATP se disocia generando ADP.Pero, adems de estar en equilibrio dinmico o de polimerizacin direccional, filamentos como los microtbulos y los microfilamentos, pueden experimentar cambios bruscos de longitud. Esta situacin se designa deinestabilidad dinmica, es transiente y tiende a aumentar la formacin de monmeros, es decir, disminuye la longitud de los filamentos.La funcin fundamental de las neuronas es el manejo de informacin. La reciben, la procesan, la almacenan y la generan. Las neuronas producen molculas (neurotransmisores, neuromoduladores) para comunicarse y/o controlar y/o modular el funcionamiento de otras clulas. Tambin lo hace en relacin a su propio funcionamiento, a travs de segundos mensajeros, que genera en respuestas a las seales que le llegan desde otras clulas. La naturaleza de los procesos relacionados con la produccin de esa amplia gama de seales es bioqumica. Pero tambin puede ser bioelctrica cuando las neuronas se comunican con otras clulas a travs de iones (sinapsis elctricas).Desde el punto de vista bioqumico, los principales mensajeros entre las neuronas y otras clulas pueden ser: aminas (acetil-colina, serotonina, catecolaminas), amino-cidos (glutamato, aspartato, GABA, glicina), nucletidos (ATP, adenosina) y pptidos neuroactivos (opiceos, pptidos neurohipofisiarios, secretinas, insulinas, somatostatina, gastrinas). Estos mensajeros son almacenados en vesculas sinpticas y son liberados por mecanismos complejos generalmente iniciados por seales bioelctricas, los potenciales de accin.La informacin bsica ms importante relacionada con cada neurotransmisor se puede obtener a partir del recuadro correspondiente.
1. Modelo de mosaico fluido de la membrana plasmtica (bicapa de fosfolpidos)2. Lado externo de la membrana3. Lado interno de la membrana4. Protena intrnseca de la membrana5. Protena canal inico de la membrana6. Glicoprotena1. Molculas de fosfolpidos organizadas en bicapa2. Molculas de colesterol3. Cadenas de carbohidratos4. Glicolpidos5. Regin polar (hidroflica) de la molcula de fosfolpido6. Regin hidroflica de la molcula de fosfolpido
Esta estructura es una bicapa lipdica, formada por fosfolpidos, que actua como un esqueleto o soporte en el cual se insertan numerosas otras estructuras moleculares como canales inicos, receptores qumicos, transportadores, bombas inicas, enzimas que generan segundos mensajeros, protenas de reconocimiento y de conexin con otras clulas, protenas que sirven de soporte a elementos del citoesqueleto. La membrana plasmtica de la neurona puede, entonces, adems de limitar la estructura de esta clula cumplir un amplio rango de funciones. Adems de su naturaleza lipdica, la membrana se caracteriza por ser polarizada elctricamente ya que su lado interno esta cubierto por una nube de cargas negativas, mientras que su exterior lo est de cargas positivas.La membrana separa dos compartimientos: el intraneuronal y el extraneuronal. Por su composicin lipdica impide el paso a travs de ella de molculas hidroflicas (solubles en agua) y/o de aquellas que tengan cargas elctricas (iones) a travs de esa fase. Sin embargo, se comporta como una membrana semipermeable selectiva frente a este tipo de substancias. En efecto, en reposo es permeable al in potasio y al agua pero impermeable a otras especies inicas como el Na+o el Ca2+. Tambin es selectivamente permeable a ciertos metabolitos como la glucosa u a otras molculas, como los precursores de neurotransmisores.El paso de iones se hace a travs de protenas-canales, que son reguladas por seales qumicas (neurotransmisores, hormonas o drogas) o por cambios en la diferencia de voltaje que caracteriza a la membrana, la cual es mantenida dentro de rangos muy estrechos por el trabajo de las bombas inicas (bomba de Na+-K+, bomba de Ca2+).En base al funcionamiento coordinado de canales y bombas inicas existe en las membranas plasmticas celulares un sistema que regula la excitabilidad neuronal y que le permite responder en forma casi instantnea a una amplia variedad de estmulos, normales unos (neurotransmisores, hormonas) perturbaciones otros (drogas). La respuestas que generan las neuronas frente a estos estmulos son de naturaleza bioelctricas y estn representadas porpotenciales localesypropagados. Estos ltimos estn acoplados, en las neuronas, a la liberacin de neurotransmisores que son las seales a travs de las cuales ellas se comunican con otras clulas. Pero tambin las neuronas pueden responder generando segundos mensajeros, que pueden interactuar entre s e inducir cambios duraderos en la conducta neuronal. Este tipo de mecanismo le confiere a las neuronas una alta plasticidad funcional que es la base de procesos complejos como elaprendizajey lamemoria.1. Osciloscopio de rayos catdicos2. Pantalla del osciloscopio3. Barrido con circuito abierto4. Barrido con circuito cerrado5. Pila6. Interruptor7. Ctodo8. Chorro o rayo de electrones dirigidos hacia la pantalla9. Dispositivos de aceleracin10. Placas horizontales11. Placas verticales
Es un instrumento que permite medir con gran precisin diferencias de potencial, corrientes, resistencias y otros parmetros elctricos, en un ampio rango. Es una aplicacin prctica de los rayos catdicos o electrones (partculas cargadas con electricidad negativa). Esta partculas se desprenden desde el ctodo de un circuito elctrico cuando circula la corriente. Fsicamente, es un verdadero can de electrones que se ubica en el interior de un tubo de alto vaco en el cual, en la cara opuesta al ctodo, se instala un pantalla (vidrio) recubierta de material fluorescente (tungstato de cadmio) que emite luz al ser impactada por los electrones.Los electrones que salen del ctodo son acelerados en su trayectoria hacia el nodo terminal que se encuentra en la pantalla. En la trayectoria de los electrones, que van configurando un verdadero haz, se han instalado dos placas llamadas horizontales entre las cuales pasa el haz de electrones. Estas placas estn dispuestas verticalmente y ellas se pueden cargar elctricamente a velocidad variable y controlada. As, mientras una de ellas adquiere carga negativa la otra queda con carga positiva, establecindose entonces una diferencia de potencial entre ambas placas. Este cambio afecta al haz de electrones el cual se desva horizontalmente, yendo a dar al lado de la pantalla donde est la placa con carga positiva. As, el haz de electrones puede recorrer la pantalla de derecha a izquierda a la velocidad que es determinada a voluntad. Esto depende de la velocidad con que se cambien las cargas de una de las placas con respecto a la otra. Cuando esta velocidad es baja, se ver en la pantalla del osciloscopio un punto que se desplaza en sentido horizontal. Si la velocidad aumenta, se ver que el punto se desplaza con mayor rapidez y a velocidades ms altas, el punto en movimiento se transformar en una lnea.El osciloscopio dispone, adems, de otro juego de placas llamadas esta vez, placas verticales. Estas pueden conectarse con fuentes de poder elctrico (diferencias de voltaje), como por ejemplo, una pila elctrica, cuyo potencial se puede medir.Si el polo positivo de la pila (nodo) se conecta a la placa vertical inferior y el ctodo de la pila a la placa superior, esta ltima se cargar negativamente, lo cual provocar un salto vertical del haz de electrones en sentido descendente. Ello se traducira en un desplazamiento de la lnea de la pantalla a otra posicin, en la parte inferior de ella.La magnitud del salto depende de la magnitud del voltaje de la pila, es decir, de la diferencia de potencial entre el nodo y el ctodo. Si se desconecta la pila del osciloscopio, la lnea de la pantalla volver a su posicin inicial, que correspondera a una diferencia de voltaje igual a cero.1. Axn gigante (400 7002. Microelectrodo3. Electrodo de referencia4. Pantalla del osciloscopio5. Placa vertical superior6. Placa vertical inferior7. Medidor de voltajes8. Barrido9. Sistema generador de pulsos (estmulos elctricos) con dos electrodos: un ctodo (-) y un nodo (+)10. El microelectrodo penetra en el interior del axn11. El barrido da un salto y se ubica en esta nueva ubicacin. La diferencia entre las dos posiciones marca la diferecia de potencial que existe entre el lado extremo y el interno de la membrana del axn
Los microelectrodos son dispositivos de vidrio o de ciertos tipos de metal, o de sus aleaciones (oro, platino, platino-iridio), que permiten registrar en la inmediata vecindad de una neurona su actividad elctrica. Cuando los dispositivos son de vidrio y tienen una punta tan fina, que no es posible verla bajo el microscopio ptico (dimetro externo inferior a 0.01 nm), se les define como ultramicroelectrodos y con ellos se puede penetrar las clulas sin peligro de daarlas mecanicamente. Si estos microelectrosdos se ubican en un sistema de soporte adecuado y se les llena con un medio conductor elctrico (solucin salina de alta concentracin inica) y se les conecta al osciloscopio de rayos catdicos a travs de un medio adecuado de amplificacin, es posible conocer las caractersticas elctricas de las neuronas en reposo y durante su actividad.Si conectamos un ultramicroelectrodo a la placa vertical superior del osciloscopio y otro electrodo, llamado de referencia, para cerrar el circuito lo conectamos a la placa vertical inferior, podremos explorar la conducta elctrica de la neurona antes y despus de penetra con el ultramicroelectrodo o electrodo activo.Si ambos electrodos se encuentran fuera de la neurona, como se indica en el esquema, el barrido en la pantalla del osciloscopio (punto o lnea luminosa que atraviesa la pantalla del osciloscopio) no se altera ya que no hay diferencia de potencial entre las placas. Esa lnea y su ubicacin en la pantalla del osciloscopio nos sevirn de referencia y le daremos un valor igual a cero.Al penetrar con el microelectrodo activo al interior del soma neuronal, el barrido en la pantalla del osciloscopio da un salto hacia abajo y toma una nueva ubicacin donde queda estable. Al sacar el microelectrodo desde el interior de la neurona el barrido vuelve a la posicin cero. Qu significa este cambio en la posicin del barrido en la pantalla ? Cmo interpretamos que al estar ambos electrodos en el lado externo de la neurona, el barrido en la pantalla del osciloscopio permanece inalterable y en la misma posicin ?El cambio de posicin del barrido seala un cambio en el voltaje de la placa vertical superior, a la cual est conectado el microelectrodo, con respecto a la otra placa. El salto hacia abajo significa que el barrido ha sido rechazado por la nueva carga que adquiri la placa superior. Como los electrones tienen carga elctrica negativa, podemos inferir que dicha placa se ha cargado negativamente. En otras palabras, que el electrodo activo, al penetrar en el interior de la neurona ha penetrado en un campo elctrico ms negativo que el que exista en el lado exterior de la membrana y que, por lo tanto, existe una diferencia de potencial entre ambos lados de la membrana. Es el llamado potencial de membrana comunmente denominado tambin como potencial de reposo y que se caracteriza porque el interior de la neurona es ms negativo que el exterior.1. Lado externo de la membrana plasmtica2. Lado interno de la membrana plasmtica3. Membrana plasmtica4. Canal inico especfico para el in sodio5. Canal inico especfico para el in potasio6. Canal inico especfico para el in cloro
1. Compartimiento extracelular2. Iones en el compartimiento extracelular (Na+:in de sodio; K+:in potasio; Cl:in cloro)3. Membrana plasmtica4. Compartimiento citoplasmtico (intracelular) A:aniones5. Iones en el compartimiento intracelular6. Carga positiva (+) que predomina en el lado externo de la membrana7. Carga negativa (-) que predomina en el lado interno de la membrana
La membrana plasmtica presenta propiedades elctricas, ya que se presenta elctricamente polarizada (su lado interno es ms negativo que el externo), y determina efectos osmticos ya que es capaz de influir en la distribucin de los iones debido a que ejerce una permeabillidad selectiva sobre ellos. Estos se encuentran en diferente concentracin a uno u a otro lado de la membrana.Por su naturaleza lipdica la membrana es impermeable a los iones, pero ello no ha sido obstculo para que de la interaccin que hay entre ellos y la membrana se generen caractersticas de gran importancia funcional. Por un lado, un sistema que regula la excitabilidad de la neurona basado en la diferencia de potencial que existe entre el exterior y el interior (ms negativo) de la membrana plasmtica es el llamado potencial de reposo de la membrana. Por otra parte, un sistema de canales inicos regulados por el valor del potencial de reposo, cuya apertura en forma coordinada genera un sistema de seales que se transmite, normalmente desde el soma al terminal nervioso, es el potencial de accin o espiga. Su funcin en la neurona es inducir la liberacin de un mensaje qumico hacia una clula vecina, permitiendo as un flujo de informacin.Por qu existe el potencial de reposo? Los iones que existen en el interior o en el microambiente de la neurona tienden a distribuirse buscando igualar sus concentraciones en el compartimiento y entre el exterior y el interior de la neurona. Ello se debe a que para cada especie inica hay dos fuerzas que determinan su distribucin: las diferencias de su concentracin y la fuerza del campo elctrico en el que se encuentran. Cada in se comporta buscando entonces un equilibrio electroqumico. La gradiente de concentracin (fuerza osmtica) empuja en un sentido y la fuerza elctrica en el sentido opuesto.En condiciones de reposo la membrana es permeable solo al K+porque es el canal para este catin el nico que est abierto. Como en el interior de la neurona (o de cualquier clula) existen aniones (A), protenas con carga negativa, el K+se acumula en el interior tratando de neutralizar su carga. Hay mayor cantidad de K+en el interior de la neurona. Existe entonces una fuerza osmtica que induce un constante flujo de K+hacia el exterior, a travs de los canales de K+abiertos. Pero la nube de K+que tiende a salir de la neurona se acumula en el lado externo de la membrana dejando exceso de carga negativa, que acta como una fuerza que los tiende a retener. Se produce entonces un equilibrio en el cual la cantidad de K+que sale es igual a la que se recupera (bomba de Na+-K+), lo que explica la constancia del potencial de membrana.1. Neurona2. Terminales nerviosos excitadores3. Terminales nerviosos inhibidores4. Montculo axnico5. Axn mielnico6. Mielina7. Ndulo de Ranvier8. Electrodos de registro extracelular (plata, platino-iridio)9. Terminal nervioso excitador activado. Se est liberando neurotransmisor que excita localmente slo la regin que destella10. Placa vertical inferior11. Placa vertical superior12. Pantalla del osciloscopio13. Barrido14. Artefacto15. Medidor de voltaje
Al estimular a travs de terminales nerviosos excitadores la regin somato-dendrtica de una neurona, se originan desde la zona vecina al montculo axnico, en elsegmento inicial del axn, seales elctricas (potenciales de accin) que se propagan hasta el terminal nervioso.Si se ubican sobre el axn un par de electrodos metlicos (de plata, o de oro, o de iridio) es posible registrar en la pantalla del osciloscopio esa respuesta propagada. Observe las conexiones entre los electrodos (extracelulares) y las placas verticales del osciloscopio: el electrodo proximal al soma va conectadao a la placa vertical inferior y el distal, a la placa vertical superior.Si la neurona no est siendo estimulada, el barrido permanece en la pantalla del osciloscopio como una lnea horizontal o como un punto que se desplaza sobre la pantalla, siguiendo esa misma trayectoria horizontal.Al estimular los terminales nerviosos excitadores en forma individual, cada uno por separado, y a baja frecuencia (1 Hz) no se observarn cambios en el barrido. Sin embargo, al aumentar la frecuencia de estimulacin (E2 = 10 Hz), aparecer una perturbacin en el barrido, el cual se presentar como una onda bifsica que tendr la misma frecuencia que la del estmulo.Explique por qu la onda es bifsica.Observe que al estimular en estas condiciones se ha generado un potencial de accin (sumacin temporal).Observe la correlacin que existe entre el paso del potencial de accin por la regin donde se ubican los electrodos y las fases de la onda bifsica.Si se estimulan, simultneamente, varios terminales nerviosos excitadores (E3) a 10 Hz, tambin aparecer un potencial de accin bifsico (sumacin espacial).1. Axn gigante (400 700)2. Microelectrodo3. Electrodo de referencia4. Pantalla del osciloscopio5. Placa vertical superior6. Placa vertical inferior7. Medidor de voltajes8. Barrido9. Sistema generador de pulsos (estmulos elctricos) con dos electrodos: un ctodo (-) y un nodo (+)
Para entender las caractersticas del potencial de accin pensemos en un experimento ideal. Podemos disponer de un axn gigante. Podra ser el de una jibia, que puede alcanzar hasta 700 micrones de dimetro. Dicho axn se colocara en condiciones adecuadas de composicin inica, de pH y de temperatura. Si disponemos, adems, de un equipo estandar para estudios de registros intracelulares (osciloscopio, preamplificador) podremos ver el potencial de accin con registro intracelular. Pero, debemos disponer adems, de los llamadosultramicroelectrodos, que son microelectrodos de vidrio con una punta tan fina (< a 0.1de dimetro externo) que solo es posible verla con el microscopio electrnico. Por ello, estos electrodos estan llenos con un sistema conductor lquido, representado por una solucin salina de alta concentracin, por ejemplo, K+-Cl, 2 M. Adems del microelectrodo de registro, se utiliza otro electrodo, de referencia, que permite cerrar el circuito del sistema.Al estar ubicados los electrodos sobre la superficie del axn veremos que en la pantalla del osciloscopio el barrido se ubica en una cierta posicin, que se puede modificar a voluntad, y sin mostrar perturbacin alguna. (52-A).Al introducir el microelectrodo en el axn, el barrido cambiar bruscamente de posicin. Segn las conexiones que se muestran en el esquema, se ubicar en la parte baja de la pantalla y la distancia entre ambas posiciones representar el valor delpotencial de reposode la neurona, que corresponde a la diferencia de potencial que existe entre el lado externo y el interno de la membrana, alrededor de 70 mV (52-B).Si, en seguida, usamos los electrodos de estmulo elctrico, aplicados a la superficie del axn, podremos estudiar el efecto de esos estmulos sobre el axn. Se pueden usar estmulos de intensidad variable, entre 0.5 y 10 volts, de 1 mseg de duracin. Empezaremos a estimular con estmulos de baja intensidad (0.5 volts), la cual aumentaremos gradualmente. Veremos que con los estmulos de baja intensidad no hay perturbaciones en el barrido, con excepcin de una pequea deflexin vertical, elartefacto, que indica el momento en que llega el estmulo elctrico al axn (52-C). Al alcanzar unos 3.0 volts de intensidad (estmulo umbral), observamos que adems del artefacto, aparece en la pantalla del osciloscopio una gran deflexin, como una V invertida, que dura 3-5 mseg. Es elpotencial de accin(52-D). A partir de ese nivel de intensidad, cada vez que apliquemos un estmulo observaremos la aparicin de un potencial de accin. Pero tambin observaremos que todos los potenciales de accin tienen el mismo tamao (ley del todo o nada). Observaremos tambin que el potencial de accin consiste en una deflexin del barrido, hacia arriba, que alcanza el potencial cero (ubicacin que tena el barrido antes de la penetracin con el microelectrodo en el axn) y lo sobrepasa en alrededor de 30 mV. Se alcanza, entonces, en estafase ascendentedel potencial de accin un desplazamiento equivalente a 100 mV. Pero al alcanzar esa magnitud de cambio, el desplazamiento se detiene bruscamente (inactivacin) para volver a caer a la posicin que tena antes de la aplicacin del estmulo. Esta trayectoria es lafase descendentedel potencial de accin.Durante los 3-5 mseg que dura el evento si se trata de aplicar un segundo estmulo durante al fase ascendente del potencial de accin no se obtendr respuesta (perodo refractario absoluto). El segmento del potencial que queda sobre la lnea cero se llamaexcedente.1. Esquema que representa registro simultneo de un potencial de accin y de las conductancias al in sodio y al in potasio relacionadas con el potencial2. Potencial cero, es el potencial de referencia medido antes de la penetracin en la clula del microelectrodo3. Diferencia de potencial medida despus de la penetracin del microelectrodo4. Potencial de accin5. Conductancia al in sodio. Representa una corriente positiva queentrapor canales especficos para el in sodio. Corresponde a la fase ascendente del potencial de accin6. Conductancia al in potasio. Representa a una corriente positiva que sale de la clula. Corresponde a la fase descendente del potencial de accin.7. Escala que mide el potencial de membrana en mV8. Escala que representa el nmero de canales iones por unidad de superficie de membrana de la clula (m2)9. Artefacto
Si repitiendo el experimento descritoEl potencial de accin, medimos la conductancia a los iones durante las fases del potencial de accin, podremos entender el mecanismo inico de este fenmeno.Laconductanciaa los iones es una propiedad de la membrana del axn. Comunmente se la designa por la letraGy representa una medida de la facilidad con que los iones pasan o atraviesan un segmento de la membrana. Como los iones tienen carga elctrica, la conductancia se manifiesta en forma de corrientes elctricas que atraviesan a la membrana. La conductancia se mide en unidades llamadas siemens.Primero, gracias a los trabajos de K.S. Cole y H. T. Curtis, se encontr que durante el potencial de accin, cambiaba la conductancia de la regin de la membrana por donde pasaba el potencial. Posteriormente, A.L.Hodgkin y B. Katz, describieron, durante el potencial de accin, los cambios de conductancia para especies inicas especficas (Na+y K+)en las distintas fases de dicho fenmeno. Se encontr que durante la fase ascendente del potencial de accin est aumentada la conductancia al Na+y que durante la fase descendente, lo est la del K+.Qu sentido o direccin tendra la corriente relacionada con el aumento de la conductancia al Na+? Qu sentido tendra la corriente relacionada con el aumento de la conductancia al K+? De acuerdo a los cambios descritos en qu forma se relaciona el potencial de accin con los iones sodio y potasio.
Los NTs representan las molculas a travs de las cuales se comunican las clulas y especialmente las neuronas entre s. Son varios los criterios para definir a una molcula como NT: la molcula debe ubicarse en la clula presinptica la molcula debe liberarse cuando se hiperpolariza la parte presinptica en la clula post-sinptica se ubican receptores especficos para el NT debe existir un mecanismo que termine la accin del NTEntre las molculas que cumplen los requisitos mencionados se encuentran:1. molculas pequeas como la acetilcolina2. aminocidos3. purinas4. catecolaminas5. indolamina (serotonina o 5HT)6. histamina7. algunos pptidos cuyo tamao vara entre 3 y 30 aminocidos1. Sinapsis entre un axn terminal (2) y una dendrita (3)2. Axn terminal3. Dendrita4. Mitocondria5. Vesculas sinpticas pequeas, claras. Contienen neurotransmisor cuya molcula es de tamao pequeo6. Vesculas sinpticas grandes de centro denso, conteniendo neuropptidos o aminas biognicas
En la mayora de las neuronas las vesculas sinpticas son los organelos donde se almacenan los neurotransmisores gracias a lo cual, adems, estas molculas quedan protegidas contra la destruccin enzimtica. Tambin, juegan un papel fundamental en el proceso de liberacin del neurotransmisor por exocitosis. Se han descrito dos tipos de vesculas: las pequeas de un dimetro de alrrededor de 50 nm y las grandes que tienen entre 70 a 200 nm de dimetro.Las vesculas se forman en el soma neuronal desde donde son transportadas hasta los terminales nerviosos. Despus de participar en el proceso de liberacin del neurotransmisor las vesculas pueden serreusadasgracias al proceso de reciclaje de membranas que maneja la neurona.En vesculas sinpticas purificadas ha sido posible conocer su composicin qumica. Adems del neurotransmisor, que las define especficamente, ellas tambin almacenan otras molculas que parecen co-participar en el proceso de la neurotransmisin qumica, aunque no siempre este aclarado su papel funcional. As, en las vesculas noradrenrgicas se encuentran molculas como el ATP, o proteinas solubles como las cromograninas o enzimas como la dopamina-hidroxilasa, que cataliza la formacin de noradrenalina a partir de la dopamina.La concentracin del neurotransmisor en el interior de la vescula es muy alta. Ello se explica porque existe un sistema de almacenamiento para el neurotransmisor y porque, adems, en la membrana de la vescula existe un sistema de transporte (un transportador acoplado a una gradiente de H+que aporta energa) que permite la incorporacin del neurotransmisor contra gradiente de concentracin. En la pared de las vesculas existen, entonces, proteinas que son transportadoras y otras que son bombas inicas.Pero tambin hay en la pared de las vesculas otras proteinas que tienen que ver con su transporte hasta el terminal nervioso, con su ubicacin en esa regin, con su relacin con el citoesqueleto y con el proceso de su movilizacin en el terminal previa al de exocitosis.1. Parte presinptica (terminal nervioso o varicosidad)2. Parte post-sinptica3. Espacio sinptico4. Vesculas sinpticas con halo5. Vesculas sinpticas sin halo (estas vesculas no seran reusadas funcionalmente para la liberacin del neurotransmisor)6. Proyecciones densas7. Cisterna8. Vesculas sinpticas que han perdido el halo y son usadas para la liberacin de nuevo neurotransmisor
Esta claramente establecido que la participacin de las vesculas sinpticas en el proceso de liberacin del neurotransmisor involucra la fusin de su membrana con la del terminal axnico. Ello significa que despus de la expulsin del contenido vesicular (exocitosis) la membrana de la vescula queda incorporada en la del terminal. Posteriormente, vuelve a incorporarse al interior del terminal, en forma de vesculas que aparecen rodeadas de unhaloformado por una proteina, laclatrina. Hay entonces un proceso dereciclajede vesculas, pero an no esta claro el mecanismo involucrado. Algunas hiptesis han planteado que existiran dos vas para explicar el reciclaje de las vesculas:1. el exceso de membrana producido por la fusin de las vesculas con la membrana del terminal, genera en estas la formacin de pequeas fosas con halo de clatrina. Esta envoltura constituye una pequea red alrrededor de la cavidad de la fosa. Luego de formada la vescula, se desliga de su halo y queda la vescula lista para almacenar nuevas molculas del neurotransmisor, o recin sintetizadas o recin captadas. Esta hiptesis es la dela va de reciclaje.2. Una segunda va estara representada por segmentos de membrana del terminal (con la membrana vesicular incorporada) que en condiciones especiales, reentraran en el terminal como vesculas sin halo. Esta va se ha llamado detranscitosis.1. Terminal nervioso2. Vaina de mielina3. Citoesqueleto4. Vesculas sinpticas inmaduras5. Vesculas sinpticas maduras (aptas para la exocitosis)6. Vesculas sinptica en exocitosis7. Neurotransmisor8. Espacio sinptico9. Membrana presinptica10. Eudosoma11. Vescula sinptica en recuperacin con halo de clatrina12. Canales de calcio dependiente de voltaje13. Filamento de actina del citoesqueleto al cual se unen las vesculas cuando el terminal est en reposo14. Vesculas ubicadas en el sitio activo15. Sinaptobrevina16. Sinaptotagmina17. Sintaxina18. Complejo SNARES19. Activacin de sinaptotagmina por calcio20. Complejo calcio-sinaptotagmina cataliza la fusin de la membrana vesicular con la del terminal
El neurotransmisor (NT) es la seal qumica que libera una neurona para comunicarse con otras clulas. Como l se encuentra almacenado, en altas concentraciones, en vesculas sinpticas, el proceso de su liberacin involucra la activa participacin de estos organelos. La liberacin del NT ocurre desde el axn neuronal y slo en neuronas dopaminrgicas, ubicadas en la substancia nigra, se descrito liberacin del NT desde la dendritas y en clulas sensoriales de algunos rganos receptores (conos de la retina), que no presentan axn, se describe tambin liberacin de NT desde una regin denominada sinptica.Hay dos lugares en el axn desde los cuales se puede liberar el NT: desde la varicosidades o desde el terminal nervioso. Las varicosidades son ensanchamientos esfricos que se observan en los axones de algunas neuronas. Tanto en los terminales nerviosos como en las varicosidades se encuentran vesculas con alto contenido de NT. Desde esas ubicaciones el NT se libera constantemente en bajas cantidades (liberacin basal) que no representan una seal de comunicacin. Cuando el potencial de accin invade el terminal nervioso (o la varicosidad) se induce un aumento notable de la liberacin del NT, transformndose as en una seal de informacin. Tradicionalmente se acepta, entonces, que es el potencial de accin el que inicia la liberacin de un NT. El proceso por el cual sale el NT contenido en las vesculas es la exocitosis. La membrana de la vescula queda incorporada en la membrana del terminal, pero es selectivamente recuperada e incorporada en un proceso de regeneracin de nuevas vesculas (Ciclo exo-endocitsico) que permite el reuso de las vesculas en la funcin sinptica.Las vesculas que liberan el NT tienen que estar ubicadas en el llamado sitio activo del terminal en lugares muy cercanos al punto de liberacin (en las sinapsis rpidas) o en lugares ms alejados como ocurre en las sinapsis lentas en las cuales el NT es algn pptido o alguna amina biognica.El potencial de accin al invadir el terminal activa canales de calcio dependientes de voltaje los cuales se abren producindose una entrada de calcio al terminal con el consiguiente incremento de su concentracin en el terminal, en alrrededor de 10 nM, lo cual es suficiente para que actue como un seal. El blanco sobre el cual acta esta seal no slo se encuentra muy cercano al sitio de entrada sino que, adems, reacciona muy rpidamente con este calcio. El efecto de este catin es provocar una rpida fusin de la membrana de la vescula con la del terminal, sin embargo, aunque el mecanismo involucrado aun no est aclarado los eventos bioqumicos que ocurren durante este proceso han demostrado la participacin de importantes proteinas de las membranas de la vescula y del terminal nervioso.Cuando el terminal est en reposo, hay tres proteinas presentes en la membrana vesicular que han demostrado tener importancia funcional. La sinaptofisina, la sinaptobrevina, que estan unidas formando un complejo, y la sinapsina I, no fosforilada, a travs de la cual la vescula esta unida a filamentos de actina del citoesqueleto.Cuando aumenta la concentracin de calcio en el terminal, se une a otra proteina la calmodulina, activndola para que induzca la estimulacin de una enzima, la proteina quinasa II dependiente de calmodulina. Esta enzima activada provoca la fosforilacin (usando ATP) de la sinapsina I, lo cual provoca su separacin de la actina y de la vescula, la cual queda entonces liberada.Pero el calcio tambin provoca la separacin del complejo sinaptofisina-sinaptobrevina de modo que la sinaptobrevina de la pared vesicular se comporta como un complejo molecular (V-SNARE) que tiene afinidad por otro complejo anlogo (T-SNARE, formado por SNAP-25 y sintaxina) pero ubicado en la membrana del terminal. La unin entre ambos complejos permite que la vescula se ubique y se fije en un punto de la membrana del terminal.Otra proteina calcio-dependiente ubicada en la pared vesicular, la sinaptotagmina, provoca la fusin de ambas membranas. Luego se formar un poro y sobrevendr la exocitosis quedando la membrana de la vescula incorporada en la del terminal.
1. Terminal nervioso que representa el elemento presinptico2. Canales de Ca2+3. Vesiculas sinpticas que viajan por flujo axoplasmtico desde el soma4. Bomba de protones en la pared vesicular5. Vescula sinptica6. Vesculas sinpticas ancladas al citoesqueleto por sinapsina I7. Vesculas sinpticas ubicadas en el sitio activo8. Membrana post-sinptica9. Receptores post-sinpticos1. Vescula sinptica en proceso de exocitosis liberando su neurotransmisor2. Membrana de vescula fusionada con membrana del terminal a travs de la cual ocurre liberancin no-cuantica del neurotransmisor3. Vescula sinptica en recuperacin que tiene en su membrana clatrina (vesculas con halo)4. Vescula que se dirige al endosoma5. Formacin de vesculas desde los endosomas6. Sistema de enzimas hidrolticas de degradacin del neurotransmisor, mecanismo de trmino de la accin del NT7. Sistema de recaptacin del neurotransmisor (tambin es mecanismo de trmino de la accin del neurotransmisor)8. Receptores presinpticos al neurotransmisor liberado: autorreceptores
Las substancias puede efluir de los terminales nerviosos no slo por exocitosis. Tambin salen por difusin, como es el caso del cido araquidnico y de los derivados de eicosanoides como las prostaglandinas.Mas aun, hay otras substancias que se liberan desde los terminales nerviosos gracias a la participacin de molculas transportadoras. Estas molculas son proteinas que funcionan como bombas inicas y pueden transportar iones como el calcio, sacndoles contra gradiente de concentracin desde la clula.Algunos transportadores ubicados en los terminales nerviosos cumplen un papel central en el proceso del trmino de la accin del neurotransmisor. Este es un proceso fundamental en la neurotransmisin qumica ya que exceso de neurotransmisor en el espacio sinptico llevara al bloqueo de la sinapsis. Estos transportadores de los terminales nerviosos son capaces de secuestrar rpidamente al neurotransmisor liberado, reincorporndolo en el terminal nervioso. Algunos transportadores pueden funcionar, sin embargo, como transportadores reversos, es decir, sacan neurotransmisores desde el terminal. Tal tipo de liberacin se ha descrito para el glutamato y para el GABA en clulas de la retina.
1. Soma neuronal2. Dendrtas3. Montculo axnico4. Axn5. Terminal nervioso como botn terminal6. Varicosidad7. Vescula sinptica formada en el soma. Viaja hacia el terminal1. Autorreceptor ubicado en una varicosidad en el terminal2. Heterorreceptor ubicado en el terminal3. Receptor-canal ubicado en una espina dendrtica. Es excitador4. Receptor inhibidor ubicado en una espina dendrtica5. Receptor inhibidor ubicado en el soma6. Vesculas sinpticas ubicadas en una varicosidad7. Receptor metabotrpico ubicado en el terminal
Los receptores qumicos de las clulas han sido definidos como estructuras moleculares a las cuales se unen especficamente otras molculas consideradas comomensajeros qumicos(neurotransmisores, hormonas y otras molculas neuroactivas)Los receptores qumicos se pueden ubicar en la membrana plasmtica en la cual se insertan atravesndola. Se ha definido para ellos, dominios de membrana y tambin a ambos lados de la membrana. El del lado externo corresponde al sitio de unin para los mensajeros que vienen de otras clulas, situacin que define a los llamadosheterorreceptores. Si a ese sitio se unen mensajeros que vienen de la propia clula, se definen los llamadosautorreceptores.Pero tambin los receptores qumicos se pueden ubicar en el interior de las clula. Son losreceptores intracelularesy se les encuentra en el citoplasma o en el ncleo. Sobre ellos actuan mensajes, que por su naturaleza qumica pueden atravesar la membrana plasmtica como son las hormonas esteroidales, las tiroideas y los neurosteroides.Los receptores qumicos celulares se han ordenado en diferentes grupos:1. Receptores-canalesoreceptores ionotrpicosocanales inicos regulados por un ligando. Son proteinas que tienen un sitio de unin a un ligando especfico (el mensaje) y presentan, al mismo tiempo, un poro o canal en su estructura. En este grupo se ubican los receptores nicotnicos, los GABAA, los ionotrpicos a glutamato.2. Receptores unidos a enzimas. Son proteinas que presentan un sitio de unin a un ligando especfico en su dominio extracelular y hacia el citoplasma su estructuta presenta actividad enzimtica, generalmente comportndose como una proteina quinasa capaz de inducir la fosforilacin de otras proteinas intracelulares y cambiando, entonces, su funcionamiento. En este grupo se ubican receptores a factores de crecimiento.3. Receptores unidos a proteina G. En este grupo se encuentran una familia de proteinas cuya estructura cruza 7 veces el espesor de la membrana (receptores transmembrana 7). Cuando son activados por su ligando especfico (dominio extracelular) su respuesta se realiza a travs de su dominio intracelular y consiste en activar el complejo molecular de laproteina Gcon la cual est relacionado. Se han identificado cientos de receptores unidos a proteina G. Por intermedio de esta proteina se generan segundos mensajeros como el cAMP. En este grupo de receptores se ubican, los receptores-adrenrgicos, los receptores muscarnicos a la acetil-colina, los metabotrpicos a glutamato y receptores a hormonas peptidrgicas.4. Receptores intracelulares. Se ubican en el interior de la clula y su activacin lleva a un aumento de mRNA especficos y de sus correspondientes proteinas. En este grupo se ubican los receptores a hormonas esteroidales, a hormonas tiroideas y a neurosteroides.Loa receptores qumicos de la membrana plasmtica ubicados en el soma o en la regin dendrtica son los que reciben la informacin que les llegan desde los terminales nerviosos que inervan la neurona. Es la naturaleza inhibidora o excitadora de esos receptores la que determinar si esa neurona ser estimulada (aumento en ella de la generacin de potenciales de accin o de trenes de potenciales) o ser inhibida (disminucin del nmero de potenciales que genera en reposo o silenciada).Antes de penetrar a la neurona con el microelectrodo, cuando ambos electrodos estn en el lado externo de la membrana, no se percibe ninguna alteracin en el barrido presente en la pantalla del osciloscopio, ni en su ubicacin. Al penetrar con el microelectrodo, el barrido se ubicar de un salto en una nueva ubicacin. La diferencia entre la posicin inicial del barrido y su nueva ubicacin, representa la diferencia de potencial que existe en la membrana plasmtica de la neurona.Al estimular el terminala(E1) se observa en el barrido una breve deflexin vertical seguida de una onda de despolarizacin suave. Es un potencial excitatorio post-sinptico (PEPS). Si se estimulan dos terminales nerviosos,ayd, (E2), aparece un PEPS mayor. Finalmente, si se estimulan simultneamente los terminales nerviosos,a, d, e y b, (E3), se observar en la pantalla del osciloscopio, despus del artefacto un potencial de accin con prepotencial. Al repetir esta estimulacin (E3), pero estimulando tambin, al mismo tiempo, los terminales inhibidoresf y g (E4), slo se observar un PEPS leve.Se observa entonces que los NTs que actan sobre receptores excitadores provocan como respuestas locales, pequeas hipopolarizaciones en el dominio de esos receptores. Son los PEPS que se pueden sumar. Si su magnitud es mayor, generarn un potencial de accin que se ver deformado por la presencia de un prepotencial, que representa el PEPS. Al estimular simultneamente receptores activados por NTs inhibidores, se neutralizar el efecto de los NTs excitadores, al punto de bloquear la aparicin de potenciales de accin.
Cada receptor es una protena formada por la combinacin de 4 a 5 subunidades iguales o distintas. Esta caracterstica ofrece una enorme posibilidad de combinaciones lo que explica la existencia de un alto nmero y de una gran diversidad de receptores a pesar del relativamente bajo nmero seales-mensajes (neurotransmisores, hormonas u otro tipo de molculas neuroactivas).La protena-receptor va inmersa en la membrana plasmtica. La composicin subunitaria determina las caractersticas funcionales del receptor los cual explica que un mismo neurotransmisor pueda ejercer diversos efectos.En la tabla se muestran las distintas subunidades que en diferente proporcin pueden entrar en la constitucin de un subtipo dado de receptor. La composicin subunitaria parece depender de diversos factores, regionales algunos de ellos y otros de carcter ms general, como son los endocrinos.Para un mismo neurotransmisor pueden haber varios tipos de receptores. Por ejemplo, para el glutamato hay tres subtipos de receptores ionotrpicos, AMPA, NMDA Y KAINATO. Para cada subtipo, las subunidades son diferentes y se les identifica en forma especfica.En la tabla se indican las subunidades que forman los receptores a neurotransmisores como el GABA, la glicina, la ACh, la serotonina y las purinas.