
MUSCULAR ESQUELETICO - mmegias.webs.uvigo.es · genera el movimiento y de una parte lamentosa que...
10
Atlas de Histolog´ ıa Vegetal y Animal Tipos celulares MUSCULAR ESQUEL ´ ETICO Manuel Meg´ ıas, Pilar Molist, Manuel A. Pombal Departamento de Biolog´ ıa Funcional y Ciencias de la Salud. Fcacultad de Biolog´ ıa. Universidad de Vigo (Versi´ on: Enero 2018)
Transcript of MUSCULAR ESQUELETICO - mmegias.webs.uvigo.es · genera el movimiento y de una parte lamentosa que...

Atlas de Histologıa Vegetal y Animal
Tipos celularesMUSCULARESQUELETICO
Manuel Megıas, Pilar Molist, Manuel A. Pombal
Departamento de Biologıa Funcional y Ciencias de la Salud.Fcacultad de Biologıa. Universidad de Vigo
(Version: Enero 2018)

Este documento es una edicion en pdf del sitio
http://mmegias.webs2.uvigo.es/inicio.html.
Todo el contenido de este documento se distribuye bajo
la licencia Creative Commons del tipo BY-NC-SA
(Esta licencia permite modificar, ampliar, distribuir y usar
sin restriccion siempre que no se use para fines comerciales,
que el resultado tenga la misma licencia y que se nombre
a los autores)
La edicion de este documento se ha realizado con el software LATEX
(http://www.latex-project.org/), usando Texstudio
(www.texstudio.org/) como editor.

Contenidos
1 Muscular esqueletico 1

Tipos celulares. Muscular esqueletico. 1
1 Muscular esqueletico
Las celulas musculares esqueleticas estriadas formanlos musculos de contraccion voluntaria, los cuales nor-malmente estan anclados a los huesos mediante lostendones, aunque tambien hay musculos voluntar-ios no asociados a huesos. Las celulas muscularesesqueleticas son en realidad sincitios, es decir, uncitoplasma rodeado por membrana que incluye a nu-merosos nucleos. Son celulas muy alargadas y se car-acterizan por poseer un citoesqueleto muy desarrol-lado que permite el acortamiento de la longitud celu-lar, lo que provoca la contraccion muscular, y por elloel movimiento.
Morfologıa
Son celulas muy largas, de ahı el nombre de fibramuscular con el que tambien se les conoce, pudiendoir desde varios milımetros hasta mas de un metro. Enun corte transversal pueden tener entre 10 y 100 µmde diametro. Los nucleos se situan perifericamente enel citoplasma, justo debajo de la membrana, denomi-nada sarcolema. Esto deja la mayor parte del espaciointracelular para la maquinaria de contraccion.
El contenido interno de la celulas musculares es-queleticas esta dominado por el citoesqueleto, prin-cipalmente formado por filamentos de actina y demiosina II, una proteına motora. Ambas se asocian enhaces denominados miofibrillas que se pueden obser-var como puntos negros cuando las celulas muscularesse cortan transversalmente. Cada miofibrilla estarodeada por membranas del retıculo endoplasmaticoliso, denominado especıficamente como retıculo sar-coplasmico. Entre las miofibrillas tambien hay mito-condrias y cumulos de glucogeno.
El nombre de celula muscular esqueletica estriadase debe a que cuando se observa longitudinalmentecon el microscopio optico aparece un patron de bandasclaras y oscuras, a modo de estrıas, dispuestas perpen-dicularmente al eje mayor de la celula. Este bandeadoes consecuencia de la superposicion de los filamentosdel citoesqueleto en las miofibrillas. Las bandas os-curas se denominan bandas A (por anisotropıa, la cualse produce en la desviacion de la luz cuando se mirancon el microscopio de luz polarizada) y las claras ban-
das I (por isotropıa, la cual se produce en la desviacionde la luz). En el interior de ambas bandas se puedenobservar unas lıneas divisorias denominadas discos Z ydiscos H, respectivamente. Los discos Z tienen formatıpica de ziz-zag, cuando se observan con el micro-scopio electronico. Incluso el disco H tiene otra lıneainterna mas oscura denominada lınea M. La porcionde miofibrilla comprendida entre dos discos Z se de-nomina sarcomero.
Composicion molecular
Los sarcomeros de las celulas musculares estriadasson el resultado de la organizacion de una gran can-tidad de proteınas con diferentes funciones. Vamos aver las mas destacadas y algunas de sus funciones.
Filamentos delgados
Actina. Las moleculas de activa son el princi-pal componente del filamento delgado del sarcomero.Cada filamento de actina esta formado por dos helicesalfa enrolladas que estan asociadas con las proteınastropomiosinas y troponinas. Los filamentos madurosde actina miden aproximadamente 1 µm en longitud,la cual es extraordinariamente constante en las celulasmusculares estriadas y se consigue por la participacionde numerosas moleculas asociadas (ver mas adelante).Existen diferentes isoformas de actina que, aunquemuy parecidas estructuralmente, se expresan en tiposcelulares y momentos del desarrollo diferentes. Dosde ellas se expresan especıficamente en el musculo es-triado, tanto cardiaco como esqueletico, y otras dospresentes en el musculo estriado se expresan tambienen otros tejidos.
Tropomiosina. Esta formada por dos cadenas alfahelicoidales ensambladas como hileras y asociadas alos filamentos de actina. Hay muchas isoformas detropomiosina derivadas por maduracion selectiva de4 genes. Las hileras de tropomiosina se extienden a lolargo de todo el filamento de actina y se localizan enlos dos surcos que dejan dichos filamentos. Su princi-pal papel, junto con la troponina, es regular la inter-accion entre los filamentos de actina y las cabezas demiosina. Su accion en el contraccion muscular se debea un cambio conformacional que permite o impide elcontacto de las cabezas de miosina con los filamentosde actina. En el estado de relajacion, la tropomiosina
Atlas de la Universidad de Vigo

Tipos celulares. Muscular esqueletico. 2
Figura 1: Celulas musculares estriadas esqueleticas de la lengua de rata. En la imagen de la derecha se muestra el puntode union al hueso hioides, no se forma tendon en este caso.
bloquea el contacto actina-miosina, pero el aumentode la concentracion de calcio lo permite, y por tanto seproduce la contraccion muscular. Es interesante quela posicion de la tropomiosina varıa ligeramente de-pendiendo de las isoformas de actina y tropomiosinaque se expresen en el musculo.
Troponina En realidad hay tres troponinas asoci-adas a los filamentos de actina, C, I y T, que formanun complejo que coopera con la tropomiosina parapermitir o no la interaccion entre filamentos de actinay miosina. Hay una expresion diferencial de isoformasde troponina, de manera que cada una modula la con-traccion muscular de forma distinta en diferentes teji-dos. El papel de estas moleculas es menos conocidopero el 25 % de las cardiomiopatıas hipertroficas seatribuyen a mutaciones en las troponinas.
CapZ y Tropomodulina. Son moleculas que se unena los extremos de los filamentos de actina controlandosu elongacion y acortamiento. CapZ se une al ex-tremo + e influye en la nucleacion y estabilizacion delos filamentos de actina. CapZ es un heterodımeroformado por unidades alfa y beta, que en el musculoestriado estan embebidas en las lıneas Z, donde seunen a la actina. La tropomodulina se une al ex-tremo menos del filamento de actina, el extremo li-bre. Su capacidad de unirse y estabilizar el extremo
menos de los filamentos de actina depende de su unionsimultanea a la tropomiosina. El papel de la tropo-modulina es fundamental para establecer la longitudy estabilidad de los filamentos de actina, lo cual deter-mina las propiedades contractiles de la celula muscu-lar. Por ejemplo, las alteraciones de la tropomodulinaprovocan filamentos de actina mas largos que llevana ritmos de contraccion mucho mas lentos.
Filamentos gruesos
Miosina. El filamento grueso esta formado pormiosina y otras proteınas asociadas. Hay cientos demoleculas de miosina de la clase II. La miosina poseeuna parte globular donde se encuentra la cabeza quegenera el movimiento y de una parte filamentosa quees la encargada de anclarse y formar, junto con el restode moleculas de miosina, el filamento grueso. Existenvarias isoformas de miosina para la zona globular yfilamentosa que resultan en diferentes propiedades decontraccion, y se expresan de diferente forma segun eltipo de musculo. Los extremos del filamento grueso seinterdigitan con los filamentos de actina para formarla banda A. Las cabezas globulares de la miosina for-man los puentes con los filamentos de actina que per-mitiran el movimiento. Las cabezas de miosina estanen una porcion del filamento grueso denominado lıneaC, mientras que la lınea M es la que carece de dichas
Atlas de la Universidad de Vigo
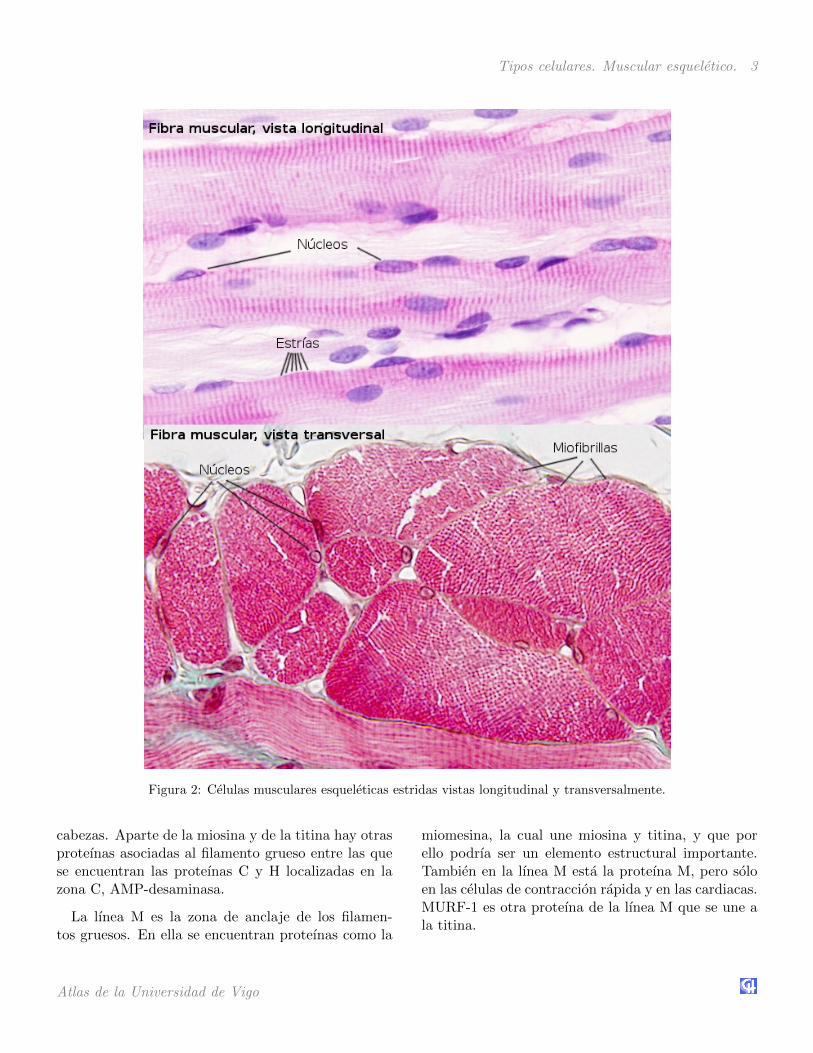
Tipos celulares. Muscular esqueletico. 3
Figura 2: Celulas musculares esqueleticas estridas vistas longitudinal y transversalmente.
cabezas. Aparte de la miosina y de la titina hay otrasproteınas asociadas al filamento grueso entre las quese encuentran las proteınas C y H localizadas en lazona C, AMP-desaminasa.
La lınea M es la zona de anclaje de los filamen-tos gruesos. En ella se encuentran proteınas como la
miomesina, la cual une miosina y titina, y que porello podrıa ser un elemento estructural importante.Tambien en la lınea M esta la proteına M, pero soloen las celulas de contraccion rapida y en las cardiacas.MURF-1 es otra proteına de la lınea M que se une ala titina.
Atlas de la Universidad de Vigo

Tipos celulares. Muscular esqueletico. 4
Figura 3: Esquema de un sarcomero.
Titina
La titina se puede considerar como el tercer fila-mento de los sarcomeros. Tras la actina y la miosina,es la tercera proteına mas abundante en el musculoy es la proteına mas larga identificada hasta la fecha(38138 aminoacidos). Ancla su extremo N-terminalen la lınea Z y el extremo C-terminal en la lınea M.Tanto uno como otro extremos se solapan con los
de otras titinas de sarcomeros adyacentes formandoası un sistema continuo. Parte de la proteına, lo-calizada en la banda I, tiene propiedades elasticas yparece contribuir a la resistencia al estiramiento delos sarcomeros, ademas de establecer una longitudmaxima para el sarcomero.
Nebulina
Es otra proteına gigante, que va desde la lınea Z
Atlas de la Universidad de Vigo
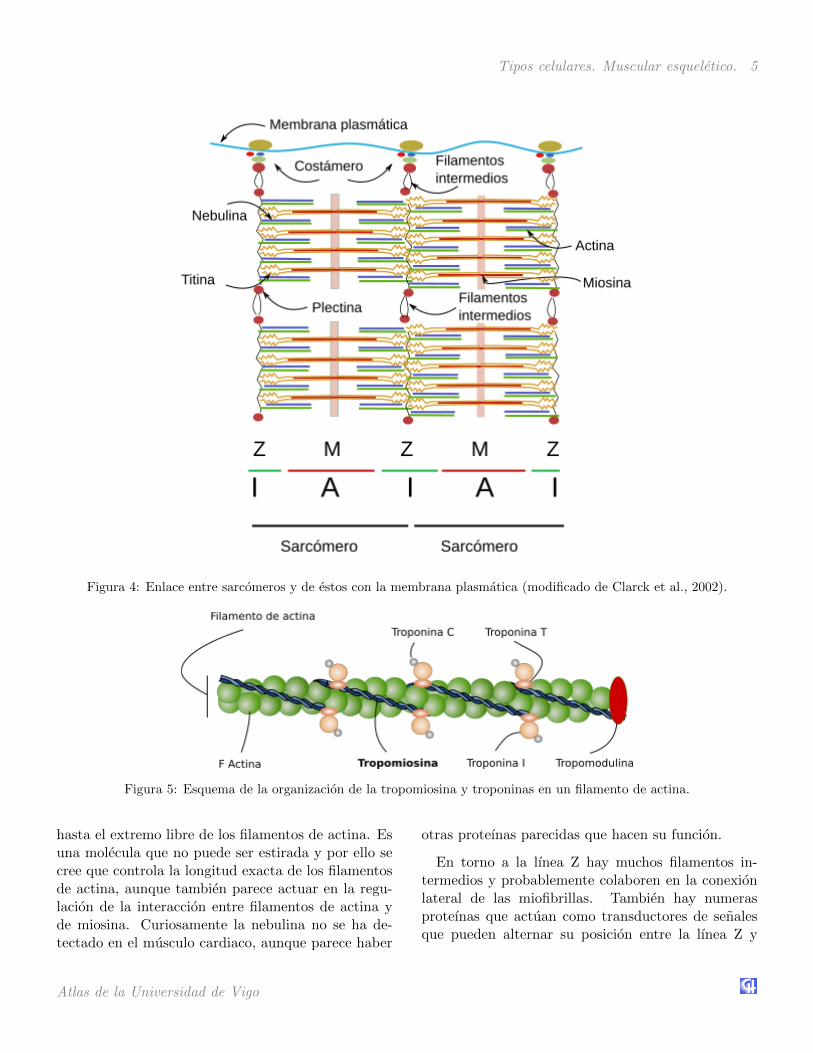
Tipos celulares. Muscular esqueletico. 5
Figura 4: Enlace entre sarcomeros y de estos con la membrana plasmatica (modificado de Clarck et al., 2002).
Figura 5: Esquema de la organizacion de la tropomiosina y troponinas en un filamento de actina.
hasta el extremo libre de los filamentos de actina. Esuna molecula que no puede ser estirada y por ello secree que controla la longitud exacta de los filamentosde actina, aunque tambien parece actuar en la regu-lacion de la interaccion entre filamentos de actina yde miosina. Curiosamente la nebulina no se ha de-tectado en el musculo cardiaco, aunque parece haber
otras proteınas parecidas que hacen su funcion.
En torno a la lınea Z hay muchos filamentos in-termedios y probablemente colaboren en la conexionlateral de las miofibrillas. Tambien hay numerasproteınas que actuan como transductores de senalesque pueden alternar su posicion entre la lınea Z y
Atlas de la Universidad de Vigo

Tipos celulares. Muscular esqueletico. 6
Figura 6: Esquema de la organizacion del filamento grueso, compuesto por moleculas de miosia enlazadas por sus cabezas.
otras partes de la celula. Aquı se encuentran proteınascomo la alfa-actinina que establece enlaces entre losfilamentos de actina, y la MLP que ayuda a la integri-dad del sarcomero y comunicacion con los nucleos dela celula. Otras son las proteınas FATZ, las miopal-adinas, filamina, telotonina, etcetera. En la lınea Ztambien se encuentra la desmina, un filamento inter-medio que tambien aparece asociada a los costameros,cuya mision es trabar los discos Z y establecer conex-iones de estos discos con la membrana plasmatica,nucleos, mitocondrias, y probablemente con los mi-crotubulos.
Costameros
Las miofibrillas de las celulas estriadas deben an-clarse a la membrana plasmatica. Los costameros
son agregados proteicos localizados en la membranaplasmatica que sirven de punto de anclaje a laslıneas Z y M. La celula muscular esta rodeada poruna matriz extracelular especial denominada laminabasal que aporta integridad estructural a la celula.Los costameros son los intermediarios que anclan lasmiofibrillas a la lamina basal. Entre las proteınasque median la adhesion con la matriz extracelularestan las integrinas, que ademas de adhesion actuancomo sensores mecanicos. Una proteına central de loscostameros es la distrofina que se une a las lamini-nas de la lamina basal. Mutaciones en la distrofinaproducen distrofia muscular. La espectrina es otraproteına presente en los costameros que ayuda en laconexion con el citoesqueleto.
Atlas de la Universidad de Vigo

Tipos celulares. Muscular esqueletico. 7
Los filamentos intermedios son elementos delcitoesqueleto que tambien ayudan a mantener la inte-gridad de la celulas muscular y colaboran en la unionde las miofibrillas con la membrana plasmatica. Laplectina, por otra parte, es una moleculas importanteintermediaria entre las miofibrillas y los filamentosintermedios. Las anquirinas favorecen la asociacionde los costameros y la membrana plasmatica, perotambien ayudan a organizar las proteınas en la mem-brana como canales y bombas en dominios especıficosde la superficie celular.
Hay proteınas asociadas a los sarcomeros quetambien se han encontrado en el interior del nucleo.Estas proteınas pueden cruzar la envuelta nuclear enlos dos sentidos y se cree que son mensajeras quellevan a los nucleos informacion del estado de lossarcomeros.
Funcionamiento
Las celulas musculares se contraen por la accionde su citoesqueleto. La unidad de contraccion se de-nomina sarcomero que es la parte de la miofibrillacomprendida entre dos discos Z consecutivos. Estoes aproximadamente unas 4 µm en un musculo rela-jado. Durante la contraccion los discos Z se puedenacercar hasta 1 µm de distancia. Esto ocurre por unacortamiento de la banda I, mientras que la A es deanchura fija. Ocurre porque los filamentos de actinason arrastrados sobre los de miosina II.
El acortamiento de la banda I se explica de la sigu-iente manera. Los filamentos de miosina II estan for-mados de manera que las cabezas de la miosina II, lascuales ejercen una fuerza tractora se encuentran ori-entadas de forma antiparalela, de manera que en losdos extremos la fuerza tractora es hacia la zona me-dia del filamento. Lo que arrastran estas cabezas demiosina II son los filamentos de actina de las bandasI.
Inervacion
Los miocitos estan inervados por neuronas denomi-nadas neuronas motoras, o motoneuronas, localizadasen la medula espinal y en el tronco encefalico. Cada
neurona es capaz de inervar a varios miocitos (unaneurona mas los miocitos inervados por ella consti-tuyen lo que se llama unidad motora), pero cada mioc-ito esta inervado por una solo motoneruona. Los ax-ones de las motoneuronas terminan sobre los miocitosformando una estructura denominada placa motorao union neuromuscular. Es una sinapsis especial es-tablecida por el terminal axonico y la membrana delmiocito, todo ello cubierto por expansiones gliales delas celulas de Schwann. El neurotransmisor implicadoen las placas motoras es la acetilcolina. Tambien hayuna inervacion sensitiva que se compone de los re-ceptores encapsulados y de los receptores tendinosos,los cuales llevan informacion sobre el estado de con-traccion y posicion de las celulas musculares.
Funcion
La funcion de los miocitos es la contraccion y comoresultado la produccion de movimiento. Mediantetecnicas histoquımicas se pueden detectar tres tiposde celulas musculares: rojas, claras e intermedias.Los tres tipos estan presentes en todos los musculospero su proporcion varıa segun la funcion particu-lar de dicho musculo. La diferencia entre ellos esla presencia de mas o menos mioglobina, que es unproteına presente en el interior de la celula y con ca-pacidad para transportar oxıgeno. Ası, las fibras rojasson celulas de pequeno tamano con una gran concen-tracion de mioglobina en su interior y con muchas mi-tocondrias, y esto les permite una contraccion lenta ysostenida, por ejemplo en aquellas que permiten man-tener la postura. Las fibras blancas poseen menosmioglobina y menos mitocondrias, y tienen un tamanocelular mayor. Son celulas con capacidad para unacontraccion rapida, pero se fatigan facilmente. Es-tos musculos participan en los movimientos precisosy rapidos. Las fibras intermedias tienen propiedadesmixtas.
Bibliografıa
Clark KA, McElhinny AS, Beckerle MC, GregorioCC. 2002. Striated muscle cytoarchitecture: an intri-cate web of form and function. Annual review of celland development biology. 18: 637-706.
Atlas de la Universidad de Vigo


















