
Métodos de muestreo binomial y secuencial para … · El objetivo de este trabajo es obtener un...
28
Bol. San. Veg. Plagas, 19: 559-586, 1993 Métodos de muestreo binomial y secuencial para Tetranychus urticae Koch (Acari: Tetranychidae) y Amblyseius californicus (McGregor) (Acari: Phytoseiidae) en fresón J. E. GONZÁLEZ ZAMORA, F. GARCÍA-MARÍ, A. RIBES, J. SAQUÉS, L. MASIELLO y S. ORENGA Desde agosto de 1988 hasta julio de 1991 se realizaron 484 muéstreos de la araña roja Tetranychus urticae Koch y del fitoseido Amblyseius californicus (MCGREGOR) en 10 parcelas de fresón en Valencia (España). Con estos datos se han desarrollado un mé- todo de muestreo enumerativo y un método de muestreo binomial (basado en el con- cepto de valor de corte) para estimar la densidad poblacional. También se ha desarro- llado un muestreo binomial de toma de decisión utilizando los valores de corte. Se ha comprobado que la población de T. urticae (hembras y formas móviles) y de A.califor- nicus (formas móviles) cumplen la ley potencial de Taylor con unos valores de b de 1,46 y de 1,58 para hembras y formas móviles de T. urticae respectivamente y 1,31 para formas móviles de A.californicus. Se ha encontrado que muestrear hembras de T. urticae es un índice aceptable de la población total de formas móviles, con un coefi- ciente de correlación r = 0,927. En el muestreo enumerativo se han obtenido las curvas que relacionan la densidad poblacional con el tamaño de muestra, con una precisión de E = 0,25, basándose en los índices de Taylor encontrados, necesitándose 25 y 35 folio- Ios para estimar adecuadamente la población de hembras y formas móviles de T. urti- cae cuando las poblaciones están alrededor de los umbrales de daño (7 hembras por fo- liólo y 20 formas móviles por foliólo); para formas móviles del fitoseido A. californi- cus son necesarios 75 foliólos para estimar su población a un nivel de 0,5 formas móvi- les por foliólo. El muestreo binomial para la estimación de la densidad poblacional se ha desarrollado en base a obtener la relación existente entre el número de ácaros por fo- liólo y el porcentaje de foliólos ocupados por To menos individuos, bien por medio de la distribución binomial negativa, bien a partir de una fórmula empírica. El criterio para la elección del valor de corte ha sido el de optimizar la robustez y minimizar el coste del método de muestreo, eligiéndose en el caso de T. urticae los valores de corte T = 4 para hembras y T = 9 para formas móviles, con un tamaño de muestra de 65 y 93 folio- Ios respectivamente cuando se muestrea con densidades poblacionales cercanas a los niveles de daño. Para fitoseidos, tomando como único valor de corte T = 0, se ha visto que utilizando la distribución binomial negativa se necesitan 125 foliólos para pobla- ciones alrededor de 0,5 formas móviles por foliólo. En el desarrollo del muestreo bino- mial secuencial de toma de decisión para T. urticae se han comparado dos de los proce- dimientos descritos en la literatura usando los valores de corte encontrados. Se han ob- tenido las curvas operativa característica (OC) y del número medio de muestras (ASN), necesarias para valorar el proceso de la toma de decisiones, en función de dichos valo- res de corte. J. E. GONZÁLEZ ZAMORA, F. GARCÍA-MARÍ, J. SAQUÉS, L. MASIELLO y S. ORENGA. De- partamento de Producción Vegetal. ETS Ingenieros Agrónomos. Universidad Politéc- nica, Camino de Vera, 14. 46022 Valencia. A. RIBES. Servei de Sanitat Vegetal (Conselleria de D'Agricultura i Pesca, Generalitat Valenciana). Ctra. Alacant-Valencia, km 249,5. Apartado 125. 46460 Silla (Valencia). Palabras clave: Muestreo enumerativo, muestreo binomial, muestreo secuencial, muestreo, Tetranychus urticae, Amblyseius californicus, fresón, índices de agregación de Taylor
Transcript of Métodos de muestreo binomial y secuencial para … · El objetivo de este trabajo es obtener un...

Bol. San. Veg. Plagas, 19: 559-586, 1993
Métodos de muestreo binomial y secuencial para Tetranychus urticae Koch (Acari: Tetranychidae) y Amblyseius californicus (McGregor) (Acari: Phytoseiidae) en fresón
J. E. GONZÁLEZ ZAMORA, F. GARCÍA-MARÍ, A. RIBES, J. SAQUÉS, L. MASIELLO y S. ORENGA
Desde agosto de 1988 hasta julio de 1991 se realizaron 484 muéstreos de la araña roja Tetranychus urticae Koch y del fitoseido Amblyseius californicus (MCGREGOR) en 10 parcelas de fresón en Valencia (España). Con estos datos se han desarrollado un método de muestreo enumerativo y un método de muestreo binomial (basado en el concepto de valor de corte) para estimar la densidad poblacional. También se ha desarrollado un muestreo binomial de toma de decisión utilizando los valores de corte. Se ha comprobado que la población de T. urticae (hembras y formas móviles) y de A.califor-nicus (formas móviles) cumplen la ley potencial de Taylor con unos valores de b de 1,46 y de 1,58 para hembras y formas móviles de T. urticae respectivamente y 1,31 para formas móviles de A.californicus. Se ha encontrado que muestrear hembras de T. urticae es un índice aceptable de la población total de formas móviles, con un coeficiente de correlación r = 0,927. En el muestreo enumerativo se han obtenido las curvas que relacionan la densidad poblacional con el tamaño de muestra, con una precisión de E = 0,25, basándose en los índices de Taylor encontrados, necesitándose 25 y 35 folio-Ios para estimar adecuadamente la población de hembras y formas móviles de T. urticae cuando las poblaciones están alrededor de los umbrales de daño (7 hembras por foliólo y 20 formas móviles por foliólo); para formas móviles del fitoseido A. californicus son necesarios 75 foliólos para estimar su población a un nivel de 0,5 formas móviles por foliólo. El muestreo binomial para la estimación de la densidad poblacional se ha desarrollado en base a obtener la relación existente entre el número de ácaros por foliólo y el porcentaje de foliólos ocupados por To menos individuos, bien por medio de la distribución binomial negativa, bien a partir de una fórmula empírica. El criterio para la elección del valor de corte ha sido el de optimizar la robustez y minimizar el coste del método de muestreo, eligiéndose en el caso de T. urticae los valores de corte T = 4 para hembras y T = 9 para formas móviles, con un tamaño de muestra de 65 y 93 folio-Ios respectivamente cuando se muestrea con densidades poblacionales cercanas a los niveles de daño. Para fitoseidos, tomando como único valor de corte T = 0, se ha visto que utilizando la distribución binomial negativa se necesitan 125 foliólos para poblaciones alrededor de 0,5 formas móviles por foliólo. En el desarrollo del muestreo binomial secuencial de toma de decisión para T. urticae se han comparado dos de los procedimientos descritos en la literatura usando los valores de corte encontrados. Se han obtenido las curvas operativa característica (OC) y del número medio de muestras (ASN), necesarias para valorar el proceso de la toma de decisiones, en función de dichos valores de corte.
J. E. GONZÁLEZ ZAMORA, F. GARCÍA-MARÍ, J. SAQUÉS, L. MASIELLO y S. ORENGA. Departamento de Producción Vegetal. ETS Ingenieros Agrónomos. Universidad Politécnica, Camino de Vera, 14. 46022 Valencia.
A. RIBES. Servei de Sanitat Vegetal (Conselleria de D'Agricultura i Pesca, Generalitat Valenciana). Ctra. Alacant-Valencia, km 249,5. Apartado 125. 46460 Silla (Valencia).
Palabras clave: Muestreo enumerativo, muestreo binomial, muestreo secuencial, muestreo, Tetranychus urticae, Amblyseius californicus, fresón, índices de agregación de Taylor

INTRODUCCIÓN
El acaro tetraníquido Tetranychus urticaeKoch es una de las principales plagas delfresón en todo el mundo. Se alimenta suc-cionando el contenido celular por el envésde las hojas, privándolas de su capacidad defotosintetizar. La acción prolongada de esteacaro lleva por tanto a una apreciable dismi-nución de la cosecha y un debilitamiento dela planta que puede terminar con la muertede ésta. El control químico ha sido la formatradicional de controlar sus poblaciones y seenfrenta a ciertos problemas como la apari-ción de resistencias debidas al uso frecuentey mala utilización de acaricidas, las prolife-raciones de ácaros inducidas por la elimina-ción de sus enemigos naturales (sobre todofitoseidos) y también la cada vez mayor im-portancia que se está dando a los residuosde plaguicidas en un fruto que se consumeen fresco.
Un medio para evitar o paliar estos pro-blemas es el de la aplicación del control in-tegrado sobre esta plaga, cosa que ya se estárealizando en algunos países como Italia(BARDUCCI, 1988; BATTAGLIA et al, 1990;
BENUZZI et al, 1990; BONOMO et al, 1991),Australia ( W A I T E , 1 9 8 8 ) O California(GLIESMAN et al, 1990). Uno de los prime-ros pasos es el de desarrollar un método demuestreo adecuado para cada una de las pla-gas más importantes, lo que nos permitirásaber el nivel poblacional que hay y obraren consecuencia. Además, el método demuestreo debe ser preciso, exacto y con lasimplificación suficiente para poder serpráctico en el campo.
Existen diversos procedimientos de sim-plificación del muestreo que manteniendola precisión deseada permiten su aplicaciónde forma razonablemente sencilla. El mues-treo por presencia-ausencia es un tipo demuestreo al que cada vez se le está dandomás importancia por las ventajas que puedemanifestar en cuanto ahorro de tiempo. Esespecialmente adecuado en poblaciones quemuestran una fuerte agregación, como losácaros, y consiste en contar el número de
unidades de muestreo ocupadas por los ar-trópodos y no el número de individuos encada unidad de muestreo. Su aplicación estátanto en la estimación de la media poblacio-nal como en la toma de decisiones cuandose sigue un programa de control integradode plagas. Autores como ANSCOMBE(1948), KONO y SUGINO (1958) o PIELOU(1960) empezaron a aplicar este conceptoen el control de plagas, relacionando lamedia poblacional (m) con la proporción dehojas no ocupadas (p0). Tanto KONO y Su-GINO (1958) como PIELOU (1960) ya indi-caban además en sus trabajos la posibilidadde elección de un valor de la poblaciónmayor de uno superado el cual se podíaconsiderar a la hoja como ocupada, aunqueesta posibilidad no había tenido hasta hacepoco una aplicación concreta, y en generalse ha aplicado considerando como hojaocupada aquella que contiene uno o más in-dividuos.
Sin embargo, recientemente algunos auto-res (BINNS y BOSTANIAN, 1990a, 1990b)han retomado esta idea y han puesto de ma-nifiesto que el error asociado a un muestreobinomial puede reducirse si se consideracomo hoja ocupada aquella en la que se en-cuentran más de T individuos (T > 0), consi-guiéndose de esta manera una mayor robus-tez en el muestreo. Además, se pueden esti-mar poblaciones a niveles de densidad máselevados que con el método tradicional deT = 0, que era una de las limitaciones quetenía, puesto que con niveles de poblacióncercanos al umbral de tratamiento se solíaencontrar que el porcentaje de hojas ocupa-das estaba cercano al 100 %. El término T,denominado por estos autores como cut-offo tally threshold, lo hemos traducido comovalor de corte. La aproximación a este tipode muestreo a través de una relación entre lamedia m y la proporción de hojas ocupadaspor T o menos individuos (pT) se puede hacerpor dos caminos. Uno es a través de unafunción de distribución conocida, como esel caso de la binomial negativa (BN); el otrocamino es a partir de expresiones empíricasformuladas por diversos investigadores.

El objetivo de este trabajo es obtener unmétodo de muestreo para la plaga de laaraña roja T. urticae en fresón basado en lapresencia-ausencia de ácaros que tenga pre-cisión en los valores estimados (es decir,que la varianza sea lo más pequeña posible),que sea robusto (es decir, que cambios enlos parámetros utilizados para desarrollar elmétodo de muestreo no influyan sensible-mente en las conclusiones), que sea exactoen los parámetros medidos (o sea, que hayaescasa diferencia entre el parámetro medidoy su valor real) y que además suponga unahorro de tiempo en los muestros efectua-dos en el campo. Los trabajos de BlNNS(1990), BINNS y BOSTANIAN (1990a, 1990b)y NYROP y BINNS (1992) hacen hincapié enestas necesidades y aportan soluciones queconjugan todas ellas. La elección de valorde corte adecuado es una de estas solucio-nes, que redundará principalmente en unamayor robustez en el método de muestreo, yque está unido al concepto de mayor exacti-tud dado que los métodos de muestreo bino-mial están basados en que se conoce la rela-ción que existe entre la media y pT, de formaque cuanto más robusto sea un método demuestreo binomial más exactitud habrá ensus predicciones (BlNNS, 1990).
En este trabajo se van a comparar por unaparte un muestreo de estimación poblacionalcuando se lleva de forma enumerativa conotro de forma binomial (tanto en el caso deseguir una función de distribución, la BN eneste caso, como si sigue un modelo empíri-co) en el que se calculará el valor de corteóptimo; por otro lado se va a proponer unmodelo secuencial binomial de toma de de-cisiones, también basado en premisas ante-riores (el valor de corte) junto con la com-paración de algunas de las fórmulas que seencuentran en la bibliografía. Simultánea-mente se desarrollará también un método demuestreo de ácaros fitoseidos de la especieAmblyseius californicus (MCGREGOR), unode los principales depredadores de la arañaroja en fresón, comparando la estimaciónpoblacional realizada según un método enu-merativo con otro de tipo binomial.
MATERIAL Y MÉTODOS
Los datos se han obtenido en una serie demuéstreos de la araña roja Tetranychus urti-cae Koch realizados sobre fresón en la pro-vincia de Valencia desde agosto de 1988hasta julio de 1991. Los muéstreos durante1988 hasta 1990 consistieron en la toma de40 foliólos centrales de la parte media-altade la planta, su traslado al laboratorio y elconteo con ayuda de una lupa binocular delas formas móviles (hembras, machos e in-maduros) y huevos presentes. Durante 1991y también en una de las parcelas de 1989 secogían de 25 a 50 foliólos y se contaban enla propia parcela las hembras, con ayuda deuna lupa de campo cuando era necesario. Entotal se realizaron 484 muéstreos correspon-dientes a 20.240 foliólos (11.640 en labora-torio y 8.600 en campo) contándose 30.480formas móviles y 42.151 hembras de T. urti-cae. De éstas últimas 31.579 fueron encampo y el resto, 10.572, en laboratorio. Elnúmero de fitoseidos contabilizados fue de3.990 formas móviles (1.849 en laboratorioy el resto en campo) de A. californicus, es-pecie prácticamente única aparecida en elfresón.
El número de parcelas muestreadas duran-te este período de tiempo fue de 10, con unaamplia variedad de situaciones. Las varieda-des que se muestrearon fueron las siguientes:Pájaro, Douglas, Chandler, Parker y Selva.
RESULTADOS Y DISCUSIÓN
Correlación entre estados de desarrollo
Una de las formas de simplificar el mues-tre o de un insecto plaga o de cualquier otroes elegir una forma de desarrollo que seafácil de detectar en el campo, incluso a sim-ple vista. Esto debe estar precedido por laconstatación de que el estado de desarrolloelegido es representativo de la poblacióntotal que nos interesa medir. En el caso dearaña roja la población que causa los dañosestá constituida por el total de las formas

Fig. 2.-En una de las parcelas muestreadas se detectó un fuerte foco de infestación producido por la araña roja en la zonaque llevaba el agricultor, llegando a la muerte de algunas plantas pese a los reiterados tratamientos que se efectuaron.
Fig. 1.-Vista general de una de las parcelas en la que se estuvo realizando los muéstreos de araña roja y fitoseidos.

Cuadro 1 .-Coeficientes de correlación encontrados entre los diferentes estados de desarrollode la araña roja Tetranychus urticae en foliólos de fresón, con las muestras llevadas al laboratorio
y aplicando la transformación Ln (m + 1)
Huevos Inmaduros Hembras Formas móviles
Formas móvilesTotal de formas
A los datos se les ha aplicado la transformación Ln (m + 1).El número de muestras utilizadas para obtener estos índices ha sido de 192.
móviles (hembras, machos, e inmaduros).Sin embargo el seguimiento de ésta en plenocampo puede ser algo engorroso, siendo im-prescindible el uso de una lupa de campo; elmuestreo de hembras en lugar de formasmóviles sería un ahorro de tiempo importan-te, primero porque habría que contar menosindividuos y segundo porque estos son demayor tamaño y de coloración distintiva .
Para comprobar si es posible el muestreode hembras se ha establecido una correla-ción en todos los muéstreos realizados entrelas diversas formas de desarrollo presentesen la población. En el Cuadro 1 se puedenver los coeficientes de correlación que exis-ten entre las diversas formas de araña roja
muestreadas. El valor del coeficiente de co-rrelación de 0,927 entre HEMBRAS y FORMASMÓVILES, una vez aplicada la transforma-ción Ln (m + 1), es elevado y puede consi-derarse suficiente para justificar el tomar alas hembras como índice de la poblacióntotal de formas móviles (Figura 3). Esto sig-nifica que puede ser factible el muestreosólo de hembras para evaluar la poblacióntotal de formas móviles, aunque es siempremás exacto el muestreo de formas móviles,ya que son éstas las que causan el daño a lasplantas. En los apartados siguientes desarro-llaremos los métodos de muestreo tanto parala población de hembras como para la po-blación de formas móviles de la araña roja.
Fig. 3.-Correlación entre hembras y formas móviles de Tetranychus urticae encontrada en foliólos de fresón con lasmuestras llevadas al laboratorio entre 1988 y 1990, aplicando la transformación Ln (m + 1) a los datos.

Fig. 4.-La araña roja Tetranychus urticae puedemultiplicarse velozmente llegando a alcanzar poblaciones
extremadamente elevadas, que acarrean la muertede la hoja y hasta de la planta si no hay algún factorlimitante como pueden ser los enemigos naturales.
Fig. 5.- Hembra adulta de Tetranychus urticae alimen-tándose en una hoja de fresón. La coloración roja lepermite destacar sobre el fondo verde del foliólo.
Fig. 6.- Hembra adulta del acaro fitoseido Amblyseiuscalifornicus cerca de un huevo de araña roja, de los quees un activo consumidor. La coloración rojiza del fitoseido
aparece después de haberse alimentado de araña roja.
Existen varios métodos para llevar a caboel muestreo de una población. Se puede in-tentar estimar la densidad poblacional, biende forma enumerativa, es decir, contando elnúmero de individuos presentes en la mues-tra, o bien de forma binomial, es decir, con-tando el número de unidades de muestreoocupadas o no por el insecto. Por otra parte,en vez de pretender estimar la densidad po-blacional se puede elaborar un método demuestreo para tomar una decisión, en estecaso la de tratar o no la araña roja, previa fi-jación de un umbral de daño para la plaga.
Estimación de la densidad poblacional.Método enumerativo
Para evaluar la abundancia de una poblaciónde insectos de forma enumerativa es necesariotomar una serie de unidades de muestreo ycontar en ellas el número de insectos presente.Para decidir el tamaño de muestra adecuadopodemos fijar la precisión del muestreo deforma que el error estándar (S/Vn) sea unaproporción fija (E) de la media (m):
El valor de n que nos interesa despejar seconsigue dando un valor a E (precisión relati-va del muestreo, p.e. 0,10 ó 0,25), siendo co-nocidas previamente las variables de la po-blación como son S y m. El problemainherente a esta expresión es que la media yla varianza muestral cambian y no son fijas,de ahí que GREEN (1970) propusiera una ex-presión que ponía a S en función de la media.Esta modificación se basa en la relación queexiste entre la media y la varianza propuestapor TAYLOR (1961) (S2= a • mb), donde aybson los denominados índices de Taylor. Laecuación anterior queda por tanto, una vezdespejada n, de la siguiente forma:

En un trabajo anterior (GARCÍA-MARÍet al, 1991) se demuestra que la agregaciónde las poblaciones de araña roja en nuestrasparcelas de fresón se ajusta de forma bastan-te aceptable a la ley potencial de Taylor,tanto en el caso de hembras como para for-mas móviles. En el Cuadro 2 se muestran losvalores de los índices de Taylor para arañaroja y ácaros fitoseidos, siendo estos valoresuna actualización con más muéstreos de lospresentados en la publicación citada. Aun-que los nuevos parámetros actualizados quefiguran en el cuadro se han calculado con unnúmero de muestras de más del doble en al-gunos casos, son casi idénticos a los queaparecen en la citada publicación.
En la Figura 7 aparecen las curvas que seobtienen para la anterior fórmula cuando seutiliza un valor de E = 0.25, tanto en formasmóviles como en hembras. Tomando un um-bral de daños para hembras de 7 por foliólo ypara formas móviles de 20 por foliólo, el nú-mero de foliólos muestrear para esos valoresde media sería de 25 y 35 foliólos para hem-bras y formas móviles respectivamente. En elcaso de fitoseidos, con las poblaciones mediasque se suelen encontrar en campo (0,5 a 2,0formas móviles por foliólo), harían falta unmáximo de hasta 75 foliólos para estimar ade-cuadamente su población a bajas densidades.
Estimación binomial
Un procedimiento de simplificación pro-puesto para evaluar poblaciones de insectos
que están fuertemente agrupadas en las uni-dades de muestreo es el de estimarlas con-tando, no el número de insectos por hoja,sino el número de hojas ocupadas o no. Aesto se denomina muestreo binomial o porpresencia-ausencia y puede aplicarse si seencuentra una relación entre la proporciónde órganos ocupados (o no ocupados) y elnúmero de insectos por órgano. Esta rela-ción puede deducirse si se conoce la funciónde distribución de la población.
Una de las distribuciones más usada es ladenominada Binomial Negativa (BN), queha demostrado en muchos casos ser aplica-ble al estudio de la distribución en poblacio-nes de insectos. Si una población se distri-buye según la función BN, entonces la rela-ción entre la proporción de hojas no ocupa-das [P(0)] y la media poblacional vienedada por la expresión (BLISS y FISHER,1953):
Previamente hay que conocer el pará-metro de la BN denominado k y que enprincipio no debería variar con el valorde la media. Sin embargo, en muchoscasos no es así y k suele variar al modifi-carse m. Uno de los procedimientos pro-puestos para evitar esto ( W I L S O N yROOM, 1983) es sustituir el valor de k enfunción de m a través de la expresión querelaciona k con m y S2 [k = m2/(S2 - m)] yponiendo el valor de la varianza S2 en
Cuadro 2.-Indices de agregación de Taylor para hembras y formas móviles de la araña rojaTetranychus urticae y formas móviles del fitoseido Amblyseius californicus encontrados
en los muéstreos llevados a cabo desde 1988 hasta 1991

Fig. 7.-Relación entre la media y el tamaño de muestra necesario para estimar la población con un error de E = 0,25 si-guiendo un método enumerativo para: a) hembras de Tetranychus urticae, b) formas móviles de Tetranychus urticae y
c) formas móviles de Amblyseius californicus.

función de m a través de la ley potencialde Taylor, suponiendo que los datos de losmuéstreos cumplan dicha ley. Con ello re-sulta:
Esta segunda opción parece algo más rea-lista que tratar de encontrar un valor de kconstante, aunque como han destacado algu-nos autores (BINNS y BOSTANIAN, 1990a;NYROP y BINNS, 1992) el ajuste existenteentre el valor de k calculado por esta expre-sión y el encontrado en los muéstreos nosuele ser muy bueno. Después de sustituir yreordenar la ecuación 4 en la 3 se obtiene laexpresión:
propuesta por WILSON y ROOM (1983).En aquellos casos en que la BN no sea
una buena aproximación a los muéstreos esposible utilizar una fórmula empírica querelacione m con p0. KONO y SUGINO (1958),y GERRARD y CHIANG (1970), de forma in-dependiente, propusieron la siguiente expre-sión:
mientras que NACHMAN (1984) obtuvo estaotra relación:
donde a, fi, a' y b' son constantes obtenidasal realizar la regresión. Como puede dedu-cirse fácilmente ambas expresiones sonequivalentes sin más que aplicar Ln a la pri-mera expresión. Esta última ecuación es laque aplicó RAWORTH (1986) en un métodode muestreo de araña roja en fresón tenien-do en cuenta el porcentaje de foliólos noocupados.
En la Figura 8 se pueden comparar ladistribución binomial y la expresión empí-
rica con los datos encontrados. El valordel coeficiente de regresión r2 que se ob-tiene al establecer una regresión linealentre los valores de p observados y los es-timados por ambas curvas puede permitir-nos establecer una comparación entreambos tipos de curvas para los distintostipos de ácaros. En T. urticae para hem-bras la BN tiene un r2 = 0,89 y la fórmulaempírica r2 = 0,88 y en el caso de formasmóviles la BN tiene un r2 = 0,88 y la fór-mula empírica r2= 0,84. Para formas mó-viles del fitoseido A. californicus en laBN resulta r2= 0,92 y en la expresión em-pírica r2= 0,92. Vemos que en ambas ex-presiones el ajuste obtenido resulta muysimilar.
En este trabajo, tal como se apuntó al co-mienzo en la introducción, la solucióndada para obtener robustez en el métodode muestreo se basa en la elección de loque denominamos como nivel o valor decorte (T) y que consiste en determinar unnivel de población por encima del cual seconsidera a la hoja ocupada, mientras quepara valores iguales o inferiores a él seconsidera no ocupada. El trabajo de BINNS(1990) ofrece una revisión del método aseguir para elegir el valor de T que nosasegure la máxima robustez; pero ademásde este criterio hay que tener en cuenta queelegir una T excesivamente elevada suponeun aumento del trabajo de muestreo altener que contar más ácaros a fin de deci-dir si una hoja se considera ocupada o no.Por ello es preferible centrarse en aquellosvalores de T lo más pequeños posibles,dentro de los que nos den una robustezaceptable.
El tamaño de muestra necesario parapoder estimar la media con fiabilidad partedel conocimiento de la varianza, tanto en elcaso de utilizar la BN como si seguimos elmodelo empírico. A continuación se va aproseguir con el cálculo del valor de T ópti-mo y el tamaño de muestra necesario paraese valor de corte, primero siguiendo el mo-delo empírico y posteriormente con la distri-bución BN.
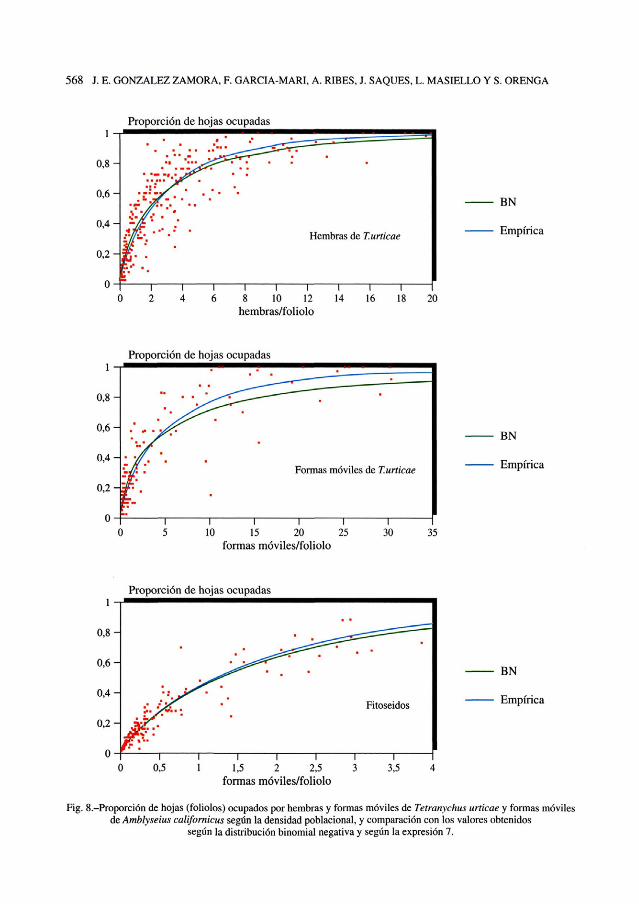
Fig. 8.-Proporción de hojas (foliólos) ocupados por hembras y formas móviles de Tetranychus urticae y formas móvilesde Amblyseius californicus según ía densidad poblacional, y comparación con los valores obtenidos
según la distribución binomial negativa y según la expresión 7.

MODELO EMPÍRICO
NACHMAN (1984) propuso una ecuaciónpara el cálculo de la varianza de Ln /Í:
donde /i es una estimación insesgada de my se obtiene mediante la ecuación 7. Estaestimación debe ser previamente corregidamediante la adición del término s2/2 a laconstante a' (NACHMAN, 1984). Los térmi-nos ay b provienen de la ecuación de la leypotencial de Taylor, S2 = a • m\ y que apa-recen en el Cuadro 2. El primer sumando dela ecuación nos indica lo bien que se ajus-tan los datos de partida a la ecuación y co-rresponde a la varianza de una regresión li-neal corriente; el segundo sumando corres-ponde a la varianza asociada a un muestreobinomial, mientras que el tercer sumandocorresponde a la varianza de los puntos demuestreo alrededor de la recta de regresión,s2, que NACHMAN expresa en función de losíndices de Taylor. Este término s2 nos indi-ca cuánto varía una parcela en concreto res-pecto a la estimación de la ecuación y espor tanto deseable que sea lo más pequeñaposible, siendo una característica del con-junto de datos. La substitución de este tér-mino s2 en función de los índices de Taylores controvertida, tal como lo reflejan lostrabajos de BINNS y BOSTANIAN (1990b) yde SCHAALJE et al., (1991), sin embargo nodeja de ser una aproximación que nos per-mite llegar a nuestro objetivo: el cálculo deun tamaño de muestra asociado a cada valorde corte.
En el caso de hembras y de formas móvi-les de araña roja los valores de los distintoscomponentes de la varianza (Fórmula 8) enfunción del valor de T aparecen en el Cuadro3. Los valores asociados a cada T se calcula-
ron mediante la regresión de la fórmula 7.De acuerdo con BINNS (1990) y BINNS yBOSTANIAN (1990b) la elección de T ha derealizarse siguiendo el criterio de asegurarla máxima robustez, fijándose, en éste caso,en que el valor de s2 (cuadrado medio resi-dual de la regresión) se el mínimo posible;otro de los parámetros a tener en cuenta esel coeficiente b', dado que cuanto más bajoes, mayor será la exactitud de las prediccio-nes de nuestro modelo de regresión. Tam-bién hay que tener en cuenta que valores ex-cesivos de T no son operativos en el campopor lo que hay que hacer una elección pon-derada. Con todas estas consideraciones nosha parecido adecuada la elección de unvalor de corte de T = 4 para hembras y deT = 9 para formas móviles.
Para formas móviles de A. californicus(Cuadro 3) el único valor de corte conside-rado ha sido T = 0 puesto que poner valoressuperiores no es muy realista, dada la difi-cultad de observar a estos ácaros y los nive-les poblacionales que suele alcanzar.
Para el cálculo del tamaño de muestra, yde acuerdo con GERRARD y CHIANG (1970)y NACHMAN (1984):
donde E es la precisión relativa del mues-treo y CV es el coeficiente de variación. Ese obtuvo mediante la expresión:
X y <|> son el intervalo de error admitido parala media m y no son iguales debido al pasode Ln a unidades métricas.

Cuadro 3.-Valores de corte considerados y parámetros asociados a la regresiónLn m = a1 + b'Ln(-Ln pT) para hembras y formas móviles de la araña roja Tetranychus urticae
y para formas móviles del fitoseido Amblyseius californicus en foliólos de fresón
Parámetros:s: = cuadrado medio residualN = número de pares de observaciones de m y pT utilizadas en la regresiónX = media de Ln(-Ln pT)SSDX = suma de las desviaciones cuadradas de Ln(—Ln pT)a'yb1 = coeficientes de la regresión
Dando pues un valor a X (p.e. 0,25) y co-nocidos los parámetros de la ecuación 8 esposible despejar de ella el término n basán-dose en las ecuaciones 10 y 11. Este métodoha sido aplicado por RAWORTH (1986) enfresón. NYROP et al, (1989) utilizan el
mismo método para calcular el tamaño demuestra en un sistema de muestreo binomialde Panonychus ulmi Koch donde comparanla utilización de la expresión 8 con otra pro-puesta por KUNO (1986) para el cáculo de n,también basada en la expresión empírica de

KONO y SUGINO (1958) (Fórmula 6), prefi-riendo la primera por comprobar estos auto-res que estima mejor la varianza asociada aun muestreo binomial.
El tamaño de muestra asociado a los valo-res de corte elegidos como adecuados se ob-tuvo substituyendo en la fórmula 8 en vezde pQ el valor de p4 para hembras, o sea, laproporción de foliólos en cada muestreo
conteniendo 4 o menos hembras y el valorp9 para formas móviles, es decir, la propor-ción de foliólos con 9 o menos formas mó-viles. Ambas curvas se pueden observar enla Figura 9. En el caso de HEMBRAS el nú-mero de foliólos a muestrear estaría alrede-dor de 65 mientras que para FORMAS MÓ-VILES serían necesarios unos 93 al tomarcomo medias las correspondientes a los res-
Fig. 9.-Tamaño de muestra en función de la media, para hembras y formas móviles de Tetranychus urücae, cuandose sigue un procedimiento de estimación binomial utilizando la expresión 7, para diversos valores de corte T,
siendo el error de estimación de E = 0,25.

Fig. lO.-Proporción de foliólos ocupados por 4 o menos hembras y por 9 o menos formas móviles de Tetranychus urticae(línea de trazo grueso) y proporción de foliólos ocupados por más de 4 hembras y más de 9 formas móviles
(línea de trazo fino) en función de la densidad poblacional, según la expresión 7.
pectivos umbrales de daños. En la misma fi-gura se puede ver el tamaño de muestra ne-cesario en caso de elegir T = 0: 80 foliólosen el caso de hembras y aproximadamente145 para formas móviles. En fitoseidos serí-an necesarios alrededor de 200 foliólos(T = 0) para poblaciones de 0,5 formas mó-viles por foliólo (Figura 12).
La relación existente de p4 y p9 (para hem-bras y formas móviles, respectivamente, de
T. urticae) con la media poblacional, segúnlas ecuaciones 6 y 7, pueden verse en la Fi-gura 10.
BINOMIAL NEGATIVA
El otro método que se ha venido utilizan-do para desarrollar el método de muestreobinomial se ha basado en la ecuación de la

BN, modelo teórico, a diferencia del ante-rior que es un modelo empírico.
Este tipo de modelo es más recomendablesiempre que pueda aplicarse debido a que sucarga teórica permite extraer más conclusio-nes y más razonadas que el modelo anterior.La aplicación de la binomial negativa nece-sita que previamente se haya comprobadoque nuestros datos se ajustan a esta distribu-ción. Para ello primero se calcula el coefi-ciente k de la binomial negativa; en nuestrocaso se aplicó el tercer método propuestopor SOUTHWOOD (1978, p. 29), que da lamáxima verosimilitud para dicho parámetro.El paso siguiente consiste en desarrollar lasfrecuencias esperadas para cada muestreo deacuerdo con el valor de k; esto se realizó si-guiendo la fórmula que proponen BLISS yFISHER(1953):
F(i) = ( k ^ Í - ' ) - , m • F(i - 1) (13)(k + m) • i
en donde N es el tamaño de muestra e / cadauna de las clases dentro de los muéstreos.
A continuación se compara la distribuciónreal de cada muestreo con la calculada pre-viamente mediante la aplicación del test X2.Con nuestros datos los resultados fueron queen el caso de HEMBRAS, de 393 muéstreosdisponibles 35 de ellos (el 9 %) no seguíanel modelo de la BN (p < 0,03); a estos últi-mos datos se les aplicó un segundo métodode comparación, el del tercer momento(SOUTHWOOD, 1978, p.31), dando como re-sultado que sólo tres de los 35 muéstreos nose ajustaban a una BN. En el caso de lasFORMAS MÓVILES se procedió de igualmanera: de 193 muéstreos 25 de ellos (13 %)no se ajustaban a la BN (p < 0,05); se aplicótambién el criterio del tercer momento resul-tando que sólo cuatro de estos muéstreos nose ajustaban tampoco a la BN. Con todosestos resultados parece posible la aplicaciónde la BN a nuestros datos.
Para fitoseidos no se han realizado loscálculos anteriores, pero como se puede de-ducir del valor de r = 0,92 obtenido al reali-zar la regresión entre la proporción de hojasocupadas según la BN y según los muéstreoses perfectamente admisible que la BN sepueda aplicar a las poblaciones de fitoseidos.
El cálculo de una k común no parece rea-lista, dado que al representar \lk frente alLn m se pudo apreciar una clara pendienteen la recta, lo que desaconseja su cálculo(SOUTHWOOD, 1978).
El procedimiento para realizar un método demuestreo binomial parte de la premisa de queconocemos el umbral de daño de la plaga encuestión, dado que a partir de éste vamos a re-alizar una serie de cálculos del parámetro k ba-sándonos en la relación que existe entre éste ylos coeficientes a y b de la ley potencial deTaylor (Fórmula 4). Esto permite dar un valorvariable para k en función de la media m loque parece ser mejor solución que la de darleun valor constante. Para hembras, con umbralde daños de 7 hembras por foliólo, el valor dek obtenido de la fórmula 4 es de 0,7 0 y paraformas móviles, con un umbral de daños de 20por foliólo, el valor de k calculado es de 0,48.
Una vez estimado este valor de k para elumbral de daños el paso siguiente es dar unintervalo donde k varía pero dentro de losmismos valores del umbral de daños. Deesta forma lo que se pretende es elegir unvalor de T donde las diferencias de probabi-lidades entre las distintas k sean lo más pe-queñas posibles y no afecten por tanto a lasdecisiones a tomar. El intervalo para elvalor de k se obtiene observando los valoresde los muéstreos, cuando la media poblacio-nal está alrededor del umbral de daños. Elproceso continúa con el cálculo de las pro-babilidades de que en un muestreo aparez-can hojas con más de 0,1, 2, .../ individuos

en donde las F(i) se han calculado según lasecuaciones 12 y 13 sustituyendo el valor dem por el que consideremos como umbral dedaños y k por el calculado en la ecuación 4para el umbral de daños y por los valoresextremos del intervalo.
El resultado de todos estos procesos sepuede ver en el Cuadro 4. En él se puedeobservar que en el caso de HEMBRAS, paraun nivel de daños estimado de 7 hembraspor foliólo la máxima semejanza de las pro-babilidades con k distintas se da en el casode P > 11, es decir, según esta cuadro habríaque tratar cuando el porcentaje de hojas conmás de 11 hembras superara el 20,5 %. ParaFORMAS MÓVILES, con un umbral dedaños de 20 formas móviles por foliólo,igualmente se puede ver que cuando P > 40las probabilidades para los diferentes valo-res de k son las más parecidas entre si; portanto cuando el porcentaje de foliólos ocu-pados con más de 40 formas móviles supereel 15,6 % habría que tratar.
Una vez elegido el valor de T que nos dala máxima robustez (T = 11 para hembras yT = 40 para formas móviles) el siguientepaso es el cálculo del tamaño de muestra
asociado a ese valor. En este caso se ha se-guido la expresión que aparece en KUNO(1986) para la BN, donde n es función de laprobabilidad de que aparezcan hojas no ocu-padas (p0) y del valor de k:
donde E es la precisión expresada en térmi-nos de error estándar. Tal como sugierenBiNNS (1990) y NYROP y BINNS (1992) elvalor de k se ha hecho variar en función dela media, de acuerdo con la ecuación 4.
El cálculo de las probabilidades de que enun muestreo aparezcan T o menos indivi-duos (pT) se ha realizado calculando las pro-babilidades de que en un muestreo aparez-can cuadrantes con í individuos, según lasfórmulas para las frecuencias esperadas dela BN (Fórmulas 12 y 13). La probabilidadde que aparezcan To menos individuos es:
T
pT = Sp(i)
Cuadro 4.-Porcentaje de foliólos en una muestra que supera la cantidad de i individuos (p > i)para el valor k calculado según la ecuación 4 y para los valores de k que se encuentran próximos,tanto en hembras como en formas móviles de Tretanychus urticae, cuando se toma como umbral
de daños 7 hembras por foliólo y 20 formas móviles por foliólo y utilizando la distribuciónbinomial negativa

y este valor es el que se va substituyendoen la ecuación 14. De esta forma todaslas ecuaciones están en función de lamedia m.
En la Figura 11 se puede ver el tamaño demuestra necesario para hembras y formasmóviles con los diferentes valores de T ópti-mos en cada caso, y comparándolos con elvalor de T = 0. Para HEMBRAS son nece-
sarios unos 105 foliólos mientras que paraFORMAS MÓVILES son necesarios unos 140foliólos cuando se muestrea en la zona delumbral de daños. En todos los casos se ne-cesitan menos foliólos cuando se utiliza elvalor de corte óptimo. En el caso de los fito-seidos serían necesarios alrededor de 125foliólos (con T = 0 ) para poblaciones de 0,5formas móviles por foliólo (Figura 12).
Fig. 11 .-Tamaño de muestra en función de la media, para hembras y formas móviles de Tetranvchus urticae, cuandose sigue un procedimiento de estimación binomial utilizando la distribución binomial negativa, para diversos valores
de corte T, siendo el error de estimación de E = 0,25.
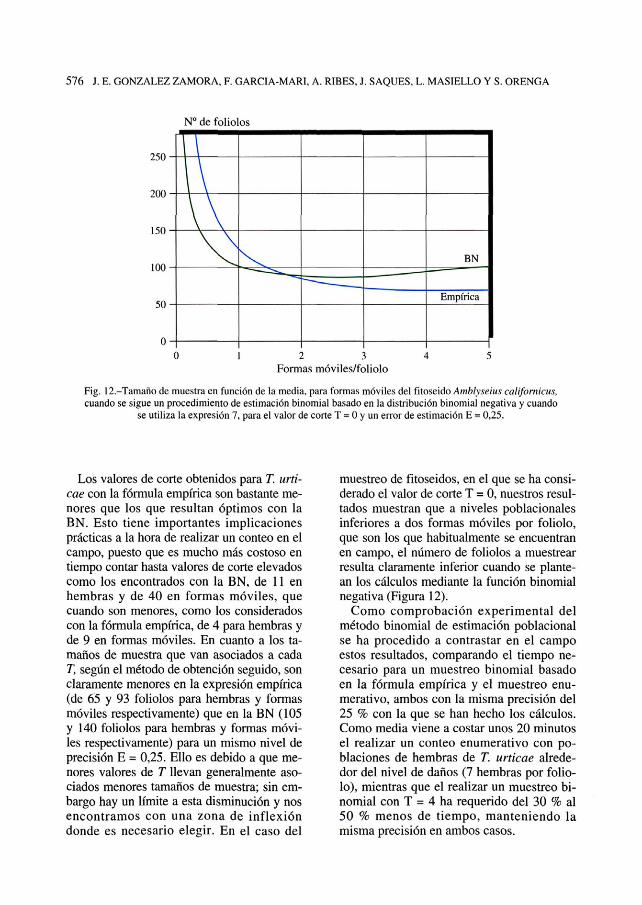
Fig. 12.-Tamaño de muestra en función de la media, para formas móviles del fítoseido Amblyseius californicus,cuando se sigue un procedimiento de estimación binomial basado en la distribución binomial negativa y cuando
se utiliza la expresión 7, para el valor de corte T = 0 y un error de estimación E = 0,25.
Los valores de corte obtenidos para T. urti-cae con la fórmula empírica son bastante me-nores que los que resultan óptimos con laBN. Esto tiene importantes implicacionesprácticas a la hora de realizar un conteo en elcampo, puesto que es mucho más costoso entiempo contar hasta valores de corte elevadoscomo los encontrados con la BN, de 11 enhembras y de 40 en formas móviles, quecuando son menores, como los consideradoscon la fórmula empírica, de 4 para hembras yde 9 en formas móviles. En cuanto a los ta-maños de muestra que van asociados a cada1] según el método de obtención seguido, sonclaramente menores en la expresión empírica(de 65 y 93 foliólos para hembras y formasmóviles respectivamente) que en la BN (105y 140 foliólos para hembras y formas móvi-les respectivamente) para un mismo nivel deprecisión E = 0,25. Ello es debido a que me-nores valores de T llevan generalmente aso-ciados menores tamaños de muestra; sin em-bargo hay un límite a esta disminución y nosencontramos con una zona de inflexióndonde es necesario elegir. En el caso del
muestreo de fitoseidos, en el que se ha consi-derado el valor de corte T = 0, nuestros resul-tados muestran que a niveles poblacionalesinferiores a dos formas móviles por foliólo,que son los que habitualmente se encuentranen campo, el número de foliólos a muestrearresulta claramente inferior cuando se plante-an los cálculos mediante la función binomialnegativa (Figura 12).
Como comprobación experimental delmétodo binomial de estimación poblacionalse ha procedido a contrastar en el campoestos resultados, comparando el tiempo ne-cesario para un muestreo binomial basadoen la fórmula empírica y el muestreo enu-merativo, ambos con la misma precisión del25 % con la que se han hecho los cálculos.Como media viene a costar unos 20 minutosel realizar un conteo enumerativo con po-blaciones de hembras de T. urticae alrede-dor del nivel de daños (7 hembras por folió-lo), mientras que el realizar un muestreo bi-nomial con T = 4 ha requerido del 30 % al50 % menos de tiempo, manteniendo lamisma precisión en ambos casos.

Muestreo para la toma de decisióno de clasificación
Un aspecto importante en el control inte-grado de plagas es el de tomar decisiones,generalmente la de tratar o no tratar, cuandose muestrea una plaga. Un ahorro importan-te de tiempo proviene de que si la poblaciónde la plaga está muy por debajo del umbralde daño unas pocas muestras (menos que lasdefinidas por los anteriores métodos) toma-das al azar por la parcela nos bastarán paratomar la decisión de no tratar; igualmente, sila población está muy por encima de dichoumbral también necesitaremos coger pocasmuestras para tomar la decisión. Además,en este tipo de muestreo siempre hay unazona de indecisión en la que hay que seguirtomando muestras hasta llegar a una deci-sión. También se puede desarrollar en térmi-nos de presencia-ausencia de la plaga, conlo que tendríamos un muestreo que combinael procedimiento binomial con la toma dedecisión.
Un primer paso es definir el umbral dedaños para la especie en cuestión. En hem-bras de araña roja ya hemos visto que se haadoptado el valor de 7 hembras por foliólo ypara formas móviles el valor de 20 formasmóviles por foliólo, umbrales que hemospodido comprobar que son aplicables ennuestras condiciones (GONZÁLEZ ZAMORAet. al, 1991), pudiendo considerarse comomoderados si se comparan con los encontra-dos por otros autores en California para cul-tivos de planta frigo de fresón (WYMANet. al, 1979; OATMAM et. al, 1981; SAN-CES et. al, 1981 ), que llegan hasta las 50formas móviles por foliólo sin una mermaapreciable de la cantidad ni la calidad de lacosecha; en el caso de planta fresca de fre-són estos autores aconsejan unos umbralesinferiores, del orden de 20 formas móvilespor foliólo. Un aspecto esencial es decidirtambién los errores asociados a las tomas dedecisiones, a saber: la probabilidad de tratarcuando el nivel de plaga está por debajo delnivel de daño (error tipo 1) representado pora y la probabilidad de no tratar cuando la
plaga ha superado el nivel de daño (errortipo 11) representado por /3. La elección deestos valores depende de varios factores,como el coste de la cosecha, coste de losproductos plaguicidas, efectos sobre otrasplagas y enemigos naturales, y relacionesentre la plaga y la planta; como ejemplo sepuede decir que si el precio de la cosechacayera el valor de a disminuiría mientrasque el de (3 aumentaría (WILSON, 1985). Ennuestro caso hemos tomado como valoresde a y ¡3 el de 0,1, que es un valor empleadofrecuentemente por diversos autores.
En la literatura se pueden encontrar diver-sas fórmulas que se han aplicado para obte-ner las rectas que nos indican las decisionesa tomar. Las más conocidas son las desarro-lladas por WALD para su test de la tasa deprobabilidad secuencial (SPRT, sequentialprobability ratio test) en función del tipo dedistribución (FOWLER y LYNCH, 1987;KREBS, 1989, pág. 246). En entomología lasmás utilizadas son las que se aplican a laBN y a la distribución binomial (Cuadro 5).En todos los casos se obtienen las ecuacio-nes de dos rectas, una superior, por encimade la cual hay que tratar, y otra inferior pordebajo de la cual no hay que tratar. El nivelsuperior de plaga está dado por el umbral dedaños mientras que el nivel inferior se esco-ge como un porcentaje más o menos arbitra-rio del nivel de daño.
Las ecuaciones para este tipo de muestreocon nuestros datos de araña roja en fresón,tanto para hembras como para formas mó-viles, se han dibujado en la Figura 13. Sólose tienen en cuenta las ecuaciones de la dis-tribución binomial por lo que se aplica eldesarrollo de la fórmula empírica y no la dela BN por ser este último más trabajoso ymenos realista, puesto que, como se havisto anteriormente, los valores de corteque se encuentran parecen demasiado ele-vados.
Las proporciones de foliólos asociadas alos umbrales de daño se han obtenido de laFigura 10. En este caso se han utilizado lasproporción de hojas ocupadas por más de 4hembras y por más de 9 formas móviles.

Cuadro 5.-Fórmulas para la obtención de las rectas de decisión de Wald cuando se realizaun muestreo secuencial de toma de decisión, según una distribución binomial
y según una distribución binomial negativa
* Recta superior* Recta inferior
En donde a y /3 vienen definidos, m es la media, k es el factor de la BN y los subíndices representan 0 el límite inferior de la plaga y 7 elnivel superior o de daño. En el caso de la distribución Binomial p es una proporción.
En las ecuaciones descritas por WALD, ade-más de las líneas de decisión, hay asociadasdos curvas, la curva operativa característica(operating characteristic, OC) y la curva delnúmero medio de muestras (average samplenumber, ASN) (FOWLER y LYNCH, 1987). Laprimera de ellas nos indica la probabilidad deaceptar la hipótesis nula o de no tratar, mien-tras que la segunda nos indica el númeromedio de observaciones necesarias para llegara una decisión terminante, en ambos casos enfunción de la densidad poblacional. Las cur-vas OC y ASN, tanto para hembras comopara formas móviles de T. urticae, aparecen
en la Figura 14. En la revisión que BlNNS yNYROP (1992) realizan de la aplicación de losmétodos de muestreo cuando el objetivo es latoma de decisiones en el control integrado deplagas, estos autores ponen de relieve la nece-sidad de obtener estas curvas como un mane-ra de valorar el método de muestreo propues-to. De esta forma, la curva ASN nos propor-ciona una forma de cuantificar el tiempo quehay que dedicar a tomar la decisión, mientrasque la curva OC nos informa de si el métodoes lo suficientemente discriminante paratomar la decisión (tratar o no tratar) en fun-ción de la media poblacional.

Fig. 13.-Rectas de decisión de Wadd para una distribución binomial, en hembras y en formas móviles de Tetranychusurticae, cuando se utilizan los valores de corte óptimos. Ecuaciones de las rectas: Hembras, recta superior
Y = 2,69 + 0,336n, recta inferior Y = -2,69 + 0,336n con pj = 0,43 y p0 = 0,25 y formas móviles, recta superiorY = 3,08 + 0,403n, recta inferior Y = -3,08 + 0,403n, con pj = 0,49 y p0 = 0,32.
Como se puede observar en la Figura 14,el número medio de foliólos necesarios paratomar una decisión oscila alrededor de 32para hembras, y de 40 para formas móviles,lo que parece ser un importante ahorro detiempo si lo comparamos con el número dehojas que habría que tomar en caso de sernecesario estimar también la densidad po-
blacional siguiendo un muestreo binomial.En las mismas figuras se comparan las cur-vas para los valores de corte elegidos y parael valor de T = 0; como se puede ver, hayuna apreciable disminución de alrededor del20 % del número de foliólos necesarios paratomar una decisión tanto en hembras comoen formas móviles.

Fig. 14.-Curvas operativas características (OC) y de! número medio de muestras (ASN), según Wald, para hembrasy formas móviles de Tetranychus urticae y para diferentes valores de T.

Una salvedad que hay que hacer es que elcálculo de estas curvas OC y ASN está some-tido a un error, tal como indican FOWLER yLYNCH (1987). Según estos autores, la conse-cuencia práctica de estos errores es que el nú-mero de foliólos indicados por la curva ASNes superior al que sería necesario realmentepara ajustamos a los errores predeterminadosa y /3, con lo cual estaríamos trabajando conunos errores inferiores a los elegidos.
WILSON et al, (1983) propusieron otrasfórmulas de muestreo binomial de toma dedecisión en donde sólo es necesario conocerel umbral de daños, no siendo necesariohacer ninguna suposición para el segundoumbral:
Fig. 15.-Rectas de decisión según las ecuaciones 15 de WILSON et al, (1983), tomando como nivel de daños T¡ = 0,43para hembras y T¡ = 0,49 para formas móviles de Tetranychus urticae, utilizando los valores de corte ótpimos.
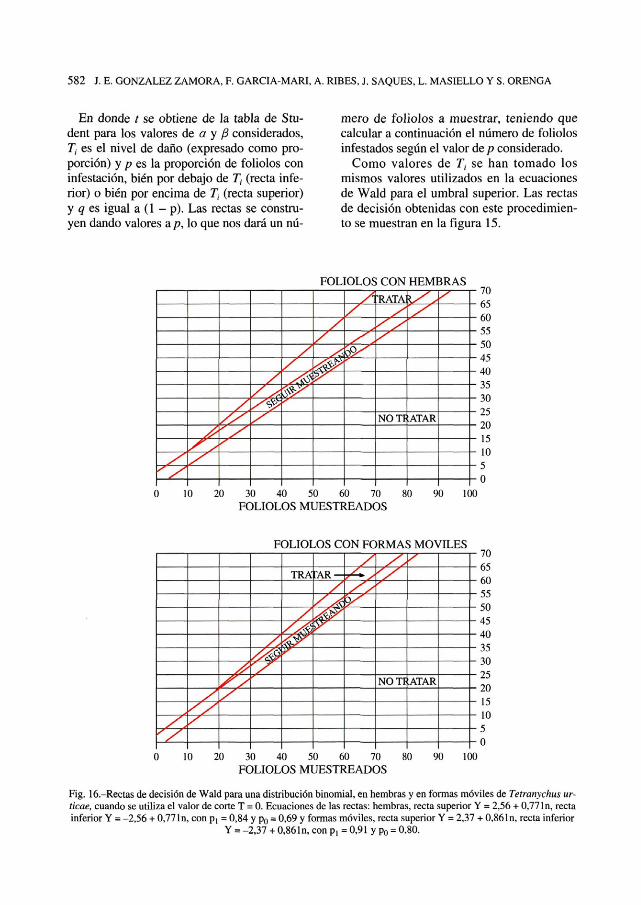
En donde t se obtiene de la tabla de Stu-dent para los valores de a y ft considerados,T¡ es el nivel de daño (expresado como pro-porción) y p es la proporción de foliólos coninfestación, bien por debajo de Tt (recta infe-rior) o bien por encima de T¡ (recta superior)y q es igual a (1 - p). Las rectas se constru-yen dando valores a p, lo que nos dará un nú-
mero de foliólos a muestrar, teniendo quecalcular a continuación el número de foliólosinfestados según el valor de p considerado.
Como valores de T¡ se han tomado losmismos valores utilizados en la ecuacionesde Wald para el umbral superior. Las rectasde decisión obtenidas con este procedimien-to se muestran en la figura 15.
Fig. 16.-Rectas de decisión de Wald para una distribución binomial, en hembras y en formas móviles de Tetranychus ur~ticae, cuando se utiliza el valor de corte T = 0. Ecuaciones de las rectas: hembras, recta superior Y = 2,56 + 0,771 n, rectainferior Y = -2,56 + 0,771n, con pj = 0,84 y p0 = 0,69 y formas móviles, recta superior Y = 2,37 + 0,861n, recta inferior
Y = -2,37 + 0,86 ln, con P! - 0,91 y p0 = 0,80.

Al comparar la Figura 13, donde se han re-presentado las líneas de decisión según lasecuaciones de Wald para hembras y formasmóviles, con la Figura 15, fórmulas de WlL-SON et ai, (1983), se observa que tanto enhembras como en formas móviles las fórmulaspropuestas por WILSON et ai, (1983) son másrestrictivas cuando la población se acerca al
umbral, necesitándose más foliólos ocupadospara llegar a tomar la decisión de tratar. Encuanto a tomar la decisión de no tratar no haytanta diferencia entre ambos modelos comoocurría en el caso anterior, pero la expresiónde WILSON et ai, (1983) sigue siendo másconservadora, necesitándose mayor número dehojas no ocupadas para tomar la decisión.
Fig. 17.-Rectas de decisión según las ecuaciones 15 de WILSON et al., (1983), tomando como nivel de daño T¡ = 0,84para hembras y T¡ = 0,91 para formas móviles de Tetranychus urticae, utilizando el valor de corte T = 0.

También se han obtenido las líneas de de-cisión cuando en vez de considerar el valorde corte óptimo se usa el valor de T = 0 (Fi-guras 16 y 17). Aunque en este caso el inter-valo de indecisión es algo más reducido quecuando se toma un valor de corte superior acero y puede parecer que se llega antes auna decisión, esto no es así ya que en elcaso de T = 0 la zona de tratar es tambiénmuy reducida. Ello es debido a que conside-rar como ocupado el foliólo a partir de unasola forma (hembra o forma móvil) noslleva a encontrar casi todos los foliólos ocu-pados cuando muestreamos una poblaciónde araña roja en fresón con densidad próxi-ma al umbral de daño, con lo que los puntosa representar en las figuras estarán siempreen la zona alta, próxima a las áreas de trataro de indecisión.
AGRADECIMIENTOS
Este proyecto se ha realizado gracias alapoyo económico de la Comisión Intermi-
nisterial de Ciencia y Tecnología (CICYT)en su proyecto AGR88-0175 y del Serviciode Sanidad de los Vegetales de la Conse-lleria de Agricultura y Pesca de la Genera-litat Valenciana. Asimismo agradecemos labeca concedida por la Consellería de Cul-tura, Educación y Ciencia de la GeneralitadValenciana al primer autor. Deseamos ex-presar nuestro agradecimiento a aquellaspersonas que han colaborado en los traba-jos de campo, Toñi Soto, Rafael Laborda,Elias Benages y Amparo Meseguer por suvaliosa ayuda. También agradecemos a losagricultores y cooperativas el que desinte-resadamente nos hayan cedido parte de susparcelas para poder realizar este trabajo: ala Cooperativa Agrícola de Alginet y a sutécnico D. Salvador García, a la Cooperati-va Agrícola de Bolbaite y especialmente aD. Juan Herrándiz y a los agricultores Sres.Espert, Zurriaga (padre e hijo), Quijal, yValero, todos ellos de Alginet, Nicasio yFrancisco Giner, de Bolbaite, SalvadorAses y Antonio Peiró de Anna y Josep Tu-dela de Alberic.
ABSTRACT
GONZÁLEZ ZAMORA, J. E.; GARCÍA-MARÍ, F.; RIBES, A.; SAQUES, J.; MASIELLO, L. yORENGA, S. (1993): Binomial and sequential sampling programs for Tetranychus urticaeKoch (Acari: Tetranychae) and Amblyseius californicus (MCGREGOR) (Acari: Phytoseii-dae) on strawberies. Bol. San. Veg. Plagas, 19(4): 559-586.
For August 1988 till July 1991, 484 samples of Tetranychus urticae Koch andAmblyseius californicus (MCGREGOR) on strawberry leaflets have been taken in 10commercial strawberry plots in Valencia (Spain). With these data a sequential estima-tion and a binomial estimation (based on the tally threslhold or cut-off concept) sam-pling program have been developed. Besides, a binomial decision sampling scheme hasbeen evolved using the tally threslhod. The distribution of the populations of the mitesagree with the Taylor power law, being b 1.46 and 1.58 for female and motile stage ofT. urticae and 1.31 for motile stage of A. californicus. Our data suggest that samplingonly females of T. urticae is a good aproximation to the sampling of all motile stagesof T. urticae, with a correlation coefícient r = 0.927. In the sequential estimation 25and 35 leaflets are needed to estimate the population density of females and motile sta-ges of T. urticae when they are around the threshold density of 7 females per leafletand 20 motile stages per leaflet, with a sampling error E = 0.25. In the case of motilestages of A. californicus, 75 leaflets are needed to estimate the population at low densi-ties (0.5 per leaflet) with the same sampling error. The binomial estimation samplingprogram has been developed by previously estimating the relationship between themite population density and the proportion of leaflets with T or less individuals, appl-ying either the negative binomial distribution and an empirical relationship. The maincriteria to select the cut-off number T have been maximize robustness and minimize thetime cost of the samplig method. For T. urticae T = 4 (females) and T = 9 (motile sta-ges) have been the cut-off selected, needing 65 and 93 leaflets to estimate the popula-tion near the threshold density. To sample A. californicus with a binomial method

based on the negative binomial distribution, 125 leaflets are necessary at low densities(0.5 mites per leaflet), using T = 0. Two methods have been compared in the process ofevolving the binomial decision sampling program for T. urticae when using the cut-offnumbers selected before. The operating characteristic (OC) and the average samplenumber (ASN) curves have been also obtained, based on the previous values of T as away of asses the proposed decision making plan.
Key words: Sequential sampling, binomial sampling, decision making sampling,sampling, Tetranychus urticae, Amblyseius californicus, strawberry, Taylor's powerlaw index.
REFERENCIAS
ANSCOMBE, F. J., 1948: On estimating the population ofaphids in a potato field. Ann. Appl. Bioi, 35: 567-571.
BARDUCCI, S., 1988: Lotta integratta e biológica per ladifesa della fragola in coltura protetta. L'lnformato-reAgrario, 21:89-91.
BATTAGLIA, D.; BORRIELLO, M. y SPICCIARELLI, R.,1990: Contrallo biológico del Tetranychus urticaeKoch con Phytoseiulus persimilis Athias-henriot sufragola in coltura protetta nel metapontino. Informa-tore Agrario, 40 (7/8): 44-46.
BENUZZI, M.; NICOLI, G. y BOTTEGHI, P., 1990: Lottabiológica e integrata sulla fragola. 4° SymposiumNacional sobre Agroquímicos, Sevilla: 181 -197.
BINNS, M. R., 1990: Robustness in binomial samplingfor decision-making in pest incidence. En: Monito-ring and Integrated Management of Arthropod Pestsof Small Fruits Crops. Editores N. J. Bostanian,L. T. Wilson, y T. J. Dennehy. Intercept Ltd., Ando-ver, Hampshire: 63-79.
BINNS, M. R., y BOSTANIAN, N. J., 1990a: Robust bino-mial decision rules for integrated pest managementbased on the negative binomial distribution. Am. En-tomoi, 36: 50-54.
BINNS, M. R.; y BOSTANIAN, N. J., 1990b: Robustnessin empirically based binomial decision rules for inte-grated pest management. J. Econ. Entomol., 83:420-427.
BINNS, M. R.; y NYROP, J. P., 1992: Samplig insect po-pulations for the purpose of IPM decision making.Annu. Rev. Entomol., 37: 427-453.
BLISS, C. I. y FISHER, R. A.; 1953: Fitting the negativebinomial distribution to biological data . Biometrics,9: 176-200.
BONOMO, G.; CATALANO, G.; MALTESE, V. y SPAR-TA, S., 1991: Esperienze di lotta biológica e integratanella fragolicultura marsalese. L'Informatore Agra-rio, 47:97-100.
FOWLER, G. W. y LYNCH, A. M, 1987: Sampling plansin insect pest management bassed on Wald' s sequen-tial probability ratio test. Environ. Entomol., 16:345-354.
GARCÍA-MARÍ, F.; GONZÁLEZ-ZAMORA, J. E.; ORENGAROYO, S.; SAQUES FERNÁNDEZ, J.; LABORDA CENJOR,R.; SOTO SÁNCHEZ, A. y RIBES KONINCKX, A., 1991:Distribución espacial y asociación entre especies deácaros fitófagos (Tetranychidae) y depredadores(Phytoseüdae) en hojas de fresón. Bol. San. Veg.Plagas, 17: 401 -415.
GERRARD, D.J. y CHIANG, H. C. 1970: Density estima-tion of corn rootworm egg populations based uponfrequency of ocurrence. Ecology, 51: 237-245.
GLIESMAN, S. R.; SWEZEY, S. L; ALLISON, J.; COCHRAN,J.; FARRELL, J.; KLUSON, R.; ROSADO-MAY, F. yWERNER, M., 1990: Strawberry production systemsduring conversion to organic management. Califor-nia Agriculture, 44(4): 4-7.
GONZÁLEZ-ZAMORA, J. E.; ORENGA, S.; GARCÍA-MARÍ,F. y LABORDA, R., 1991: Liberación de ácaros depre-dadores para el control de araña roja en fresón. Phy-toma-España, 32: 20-27.
GREEN, R. H., 1970: On fixed precision level sequentialsampling. Res. Popul. Ecoi, 12: 249-251.
KONO, T.; SUGINO, T., 1958: On the estimation of thedensity of rice stems infected by the rice stem borer.Jpn. J. Appl. Entomol. Zooi, 2: 184-188.
KREBS, C. J., 1989: Ecological methodology. HarperCollins Publishers, Inc., New York. 654 pp
KUNO, E., 1986: Evaluation of statistical precision anddesign of efficient sampling for the population esti-mation based on frequency of ocurrence. Res. Popul.Ecol, 28:305-319.
NACHMAN, G., 1984: Estimates of mean populationdensity and spatial distribution of Tetranychus urti-cae (Acariña: Tetranychidae) and Phytoseiulus per-similis (Acariña: Phytoseüdae) based upon the pro-portion of empty sampling units. J. Appl. Ecol, 21:903-913.
NYROP, J. P.; AGNELLO, A. M.; KOVACH J. y REISSIG,W. H., 1989: Binomial sequential classification sam-pling plans for european red mite (Acari: Tetrany-chidae) with special reference to performance crite-ria. J. Econ. Entomol., 82: 482-490.
NYROP, J. P. y BINNS, M. R., 1992: Algorithms forcomputing operating characteristic and averagesample number functions for sequential sam-pling plans based on binomial count modelsand revised plans for european red mite (Acari:Tetranychidae) on apple. J. Econ. Entomol., 85:1.253-1.273.
OATMAN, E. R.; WYMAN, J. A.; BROWNING, H. W. yVOTH, V., 1981: Effects of releases and varying in-festation levels of the twospotted spider mite onstrawberry yield in southern California. J. Econ. En-tomol., 74: 112-115.
PIELOU, D. P., 1960: Contagious distribution in the eu-ropean red mite, Panonychus ulmi (Koch), and a

method of grading population densities from a countof mite-free leaves. Can. J. Zool., 38: 645-653.
RAWORTH, D. A., 1986: Sampling statistics and a sam-pling scheme for the two-spotted spider mite, Te-tranychus urticae (Acari: Tetranychidae), on straw-berries. Can. Ent., 118: 807-814.
SANCES, F. V.; WYMAN, J. A.; TING, I. P.; VAN STE-ENWYK, R. A. y OATMAN, E. R., 1981: Spider miteinteractions with photosynthesis, transpiration andproductivity of strawberry. Environ. EntomoL, 10:442-448.
SCHAAUE, G. B.; BUTTS, R. A. y LYSKS, T. J., 1991: Si-mulation studies of binomial sampling: a new va-riance estimator and density predictor, with specialrefernce to the russian wheat aphid (Homoptera: Ap-hididae). J. Econ. EntomoL, 84: 140-147.
SOUTHWOOD, T. R. E., 1978: Ecological methods, withparticular reference to the studyof insect popula-tions. Chapman y Hall, London. 524 pp.
TAYLOR, L. R., 1961: Agregation, variance and themean. Nature, 189: 732-735.
WAITE, G. K., 1988: Integrated control of Tetranychusurticae in strawberry in south-east Queensland. Exp.Appl. Acaroi, 5: 23-32.
WILSON, L.T.; PICKEL, C ; MOUNT, R. C. y ZALOM,F. G., 1983: Presence-absence sequential samplingfor cabbage aphid and green peach aphid (Homopte-ra: Aphididae) on brussels sprouts. J. Econ. Ento-moL, 76: 476-479.
WILSON, L. T. y ROOM, P. M, 1983: Clumping patternsof fruit and arthropods in cotton, with implicationsfor binomial sampling. Environ. EntomoL, 12: 50-54.
WILSON, L. T., 1985: Estimating the abundance and im-pact of arthropod natural enemies in IPM systems.En Biological Control in Agricultural IPM Systems.Editores M. A. Hoy y D. C. Herzog. AcademicPress, Inc., Orlando, Florida.
WYMAN, J. A.; OATMAN, E. R. y VOTH, V., 1979: Effectsof varying twospotted spider mite infestations levelson strawberry yield. J. Econ. EntomoL, 72: 747-753.
(Aceptado para su publicación: 24 marzo 1993)


















