
Membranas biologicas composicion
13
MEMBRANAS BIOLOGICAS COMPOSICION M. EN C. RAÚL HERRERA FRAGOSO Fisiología animal
-
Upload
raul-herrera -
Category
Education
-
view
49 -
download
0
Transcript of Membranas biologicas composicion

MEMBRANAS BIOLOGICAS COMPOSICIONM. EN C. RAÚL HERRERA FRAGOSO
Fisiología animal

LIPIDOS
La fosfatidilserina tiene una carga negativa, a diferencia de los demás que tienen carga neutra a pH fisiológico.
También están los fosfatidiletanolamina pero se hallan en pequeña cantidad.
Los fosfolípidos son como un disolvente tridimensional para las proteínas de la membrana.
En las membranas de las células bacterianas hay un solo tipo de fosfolípidos, a diferencia de las eucarióticas que tienen más de un tipo de fosfolípido.

MOVILIDAD
FLIP-FLOP: pueden saltar de una monocapa a la otra, pero se produce poco porque requiere gran gasto de energía
DIFUSIÓN LATERAL: cambian de lugar con fosfolípidos vecinos, dentro de la misma monocapa unas 107 veces por segundo. Esto da lugar a la difusión lateral de los fosfolípidos, a unos 10-8 cm2/seg.
ROTACIÓN: giran sobre su eje longitudinal con rapidez.
FLEXIÓN: Separación y aproximación de los extremos de las colas, por flexión de las cadenas carbonadas de los ácidos grasos.

SINTESIS DE FOSFOLIPIDOS
Se sintetizan principalmente en la monocapa citosólica de la membrana del Retículo Endoplásmico. Pasan hacia la otra monocapa, para que no se acumulen fosfolípidos en una monocapa, por medio de una enzima unida al Retículo Endoplásmico, que cataliza el flip-flop de los fosfolípidos formados específicos de la monocapa. Dicha enzima se llama TRANSLOCADORA DE FOSFOLÍPIDOS
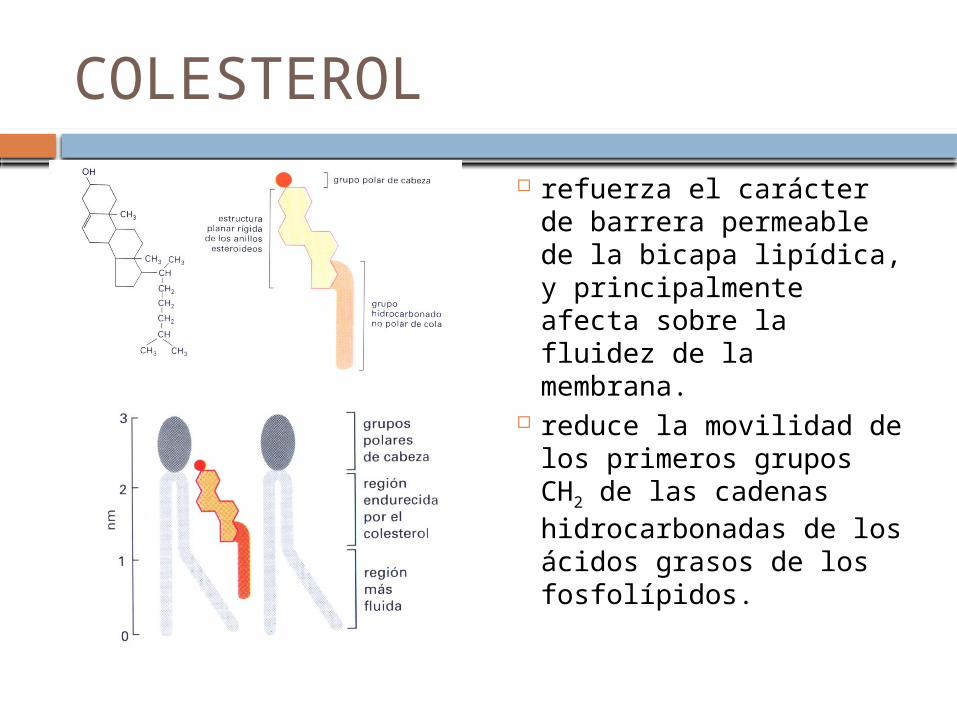
COLESTEROL
refuerza el carácter de barrera permeable de la bicapa lipídica, y principalmente afecta sobre la fluidez de la membrana.
reduce la movilidad de los primeros grupos CH2 de las cadenas hidrocarbonadas de los ácidos grasos de los fosfolípidos.

AZÚCARES Las moléculas de azúcares de la
membrana plasmática son los GLUCOLÍPIDOS.
Se encuentran exclusivamente en la mitad no citoplasmática de la bicapa lipídica, donde se autoasocian conformando microagregados mediante la formación de enlaces de hidrógenos entre ellas.
En la membrana plasmática los grupos de azúcar quedan al descubierto en la superficie de la célula donde desempeña un tipo de función en las interacciones de la célula con su entorno.
La distribución asimétrica de glucolípidos en la bicapa resulta de la adición de grupos de azúcar en las moléculas lipídicas en el lumen del complejo de Golgi que topográficamente equivale a exterior celular.

FUNCION
Gangliósidos: tiene carga negativa. Son abundantes en las células nerviosas y tiene importancia por su carga. Su presencia alteraría el campo eléctrico a través de la membrana y de iones en su superficie externa. Pueden tener papel en aislamiento eléctrico ya que se hallan en la cara no citoplasmática de la bicapa de la membrana mielínica, que aísla eléctricamente los axones de las células nerviosas.
En las células epiteliales intestinal se halla en la superficie apical de la membrana de la célula, donde protege la membrana de condiciones adversas como pH bajo y enzimas digestivas

GLUCOCÁLIZ las proteínas de membrana no sobresalen
desnudas al exterior celular, sino que están cubiertas por carbohidratos presentes en la superficie de todas las células eucarióticas.
Estos carbohidratos son cadenas de oligosacáridos unidas covalentemente a las proteínas de membrana (glucoproteína)y a los lípidos (glucolípidos)y, como cadena de polisacáridos de moléculas de proteoglucanos integrales de membrana.
Los proteoglucanos consisten en largas cadenas de polisacáridos unidas covalentemente a un núcleo proteico; se encuentran principalmente en el exterior celular como parte de la matriz extracelular. Pero en el caso de los proteoglucanos integrales de membrana, el núcleo proteico se extiende a través de la bicapa o está anclado a la bicapa mediante un glucosilfosfatidilinositol (GPI).

glucocáliz
El término glucocáliz se usa para referirse a la zona de superficie celular rica en carbohidratos. El Glucocáliz contiene (además de carbohidratos unidas a las proteínas intrínsecas de la membrana) glucoproteínas y proteoglucanos que han sido secretados al espacio exterior y que luego son adsorbidos en la superficie celular. Muchas de estas macromoléculas adsorbidas son componentes de la matriz extracelular.
Las cadenas laterales de oligosacáridos de las glucoproteínas y de los glucolípidos son diversas en cuanto a los azúcares. Generalmente contienen menos de 15 residuos glucídicos, a menudo están ramificados y los azúcares pueden estar unidos entre sí por enlaces covalentes

PROTEÍNAS
Las proteínas si bien se hallan, en la membrana plasmática, en menor cantidad que los lípidos, tienen funciones específicas. En la superficie celular pueden estar relacionadas a cadenas con carbohidratos (glucoptroteínas) formando parte del glucocáliz.
Las proteínas de membrana pueden estar relacionadas a la membrana de diferentes maneras

Proteínas transmembrana (1 y 2): Es una proteína anfipática. Ya que las regiones de la proteínas son hidrofóbicas interacciona con las colas de los fosfolípidos vecinos en el interior de la bicapa; y las regiones hidrofílicas quedan expuestas a ambos lados de la membrana hacia el medio acuoso.
Algunas de estas proteínas pueden tener una o más uniones covalentes a cadenas de ácidos grasos insertadas en la mitad citoplasmática de la bicapa. Estas uniones aumentan el carácter hidrofóbico de las proteínas transmembrana (1).
Las parte hidrofóbica de las proteínas transmembrana tiene aminoácidos de cadena lateral no polar. Y como las uniones peptídicas son en sí polares, todas las uniones peptídicas de las parte hidrofóbica, tienden a formar enlaces hidrógeno entre sí; éstas se maximizan si la cadena polipeptídica adopta forma de hélice regular al cruzar la bicapa.
Proteínas transmembrana de paso único, cuando atraviesan una sola vez la bicapa y, de proteínas de paso múltiple si la cadena atraviesa bicapa varias veces, y tienen varios segmentos hidrofóbicos intercalados con segmentos hidrofílicos. Ejemplo: bacteriorrodopsina, atraviesa la bicapa en forma de 7 hélices en la membrana de la bacteria Halobacterium halobium y es una bomba de protones.

Hay proteínas de membrana, no transmembrana, se localizan en el citosol y se asocian a la membrana sólo a través de una o más cadenas de ácidos grasos a las que se unen covalentemente o por otro tipo de cadenas lipídicas llamadas “grupos prenil” (3). Este tipo de proteínas se generan como proteínas solubles en el citosol y luego se trasladan directo a la membrana uniéndose al grupo lipídico.
Otras proteínas (4) de membrana quedan expuestas en la superficie celular externa, ancladas a la bicapa por medio de unión covalente, por medio de un oligosacárido específico, al fosfatidilinositol de la monocapa lipídica externa de la membrana plasmática. Estas proteínas se sintetizan en el RE como proteínas transmembrana de paso único, luego el segmento transmembranoso se escinde y se le añade un glucosilfosfatidilinositol (GPI), así la proteína queda unida a la superficie no citoplasmática de la membrana sólo por medio del anclaje. (el GPI se reconoce por uso de fosfolipasa C específico para GPI)
Algunas proteínas de membrana están unidas a una o a otra cara de la membrana mediante interacciones no covalentes con otras proteínas de membrana (5 y 6).

Proteínas periféricas: de la membrana, las que no son transmembrana, pueden ser liberadas mediante procedimientos de extracción relativamente suaves (de alta o baja fuerza iónica o de pH extremo) que interfieren en las interacciones proteicas pero mantienen intacta la bicapa lipídica.
Proteínas integrales: proteínas transmembrana, como varias proteínas unidas a la bicapa por cadenas de ácidos grasos y otras íntimamente ligadas a la membrana, que no pueden ser liberadas por éstos métodos.
Difusión de la proteínas: las proteínas no hacen flip-flop a través de la bicapa, sino que giran alrededor de un eje algo perpendicular al plano de la bicapa: difusión rotacional. También son capaces de desplazarse lateralmente por la membrana: difusión lateral. La velocidad de difusión varía según la proteína pero generalmente es de una décima o centésima parte de la velocidad que alcanzan las moléculas de fosfolípidos de la misma membrana.
Agrupamientos o patching: cuando los ligandos, como los anticuerpos, que tienen más de un sitio de unión (ligandos multivalentes) se unen a proteínas específicas de la superficie celular, las proteínas tienden a agregarse, mediante enlaces cruzados, formando grandes grupos o patches, lo cual indica que las proteínas son capaces de desplazarse lateralmente por la bicapa lipídica.
Caperuzas: Una vez formados los agregados en la superficie de una célula capaz de moverse, como un leucocito, son trasladados activamente a uno de los polos celulares, formando una caperuza.


















