

Membrana plásmática - iib.unsam.edu.ar · Inserción de proteínas en la membrana plasmática: PL...
40
Regula los límites externos de las células y regula el tráfico a través de los mismos. En eucariotas también divide el espacio interno en compartimientos discretos. En ella tienen lugar diversas actividades biológicas. Es flexible, autosellante y selectivamente permeable a solutos polares. Exocitosis , endocitosis , división celular. Membrana plásmática
Transcript of Membrana plásmática - iib.unsam.edu.ar · Inserción de proteínas en la membrana plasmática: PL...

Regula los límites externos de las células y
regula el tráfico a través de los mismos.
En eucariotas también divide el espacio
interno en compartimientos discretos.
En ella tienen lugar diversas actividades
biológicas.
Es flexible, autosellante y selectivamente
permeable a solutos polares.
Exocitosis, endocitosis, división celular.
Membrana plásmática

Rol de los lípidos en la función
de las membranas celulares
No constituyen un “mar” en el que se
encuentran las proteínas.
Se asocian a funciones específicas al igual
que las proteínas.
Sus dominios hidrofóbicos se asocian
minimizando la superficie total en contacto
con el agua.
Sus porciones hidrofílicas interactúan a
través de puentes de hidrógeno o
interacciones iónicas, con el agua o con
otras cabezas polares de lípidos.

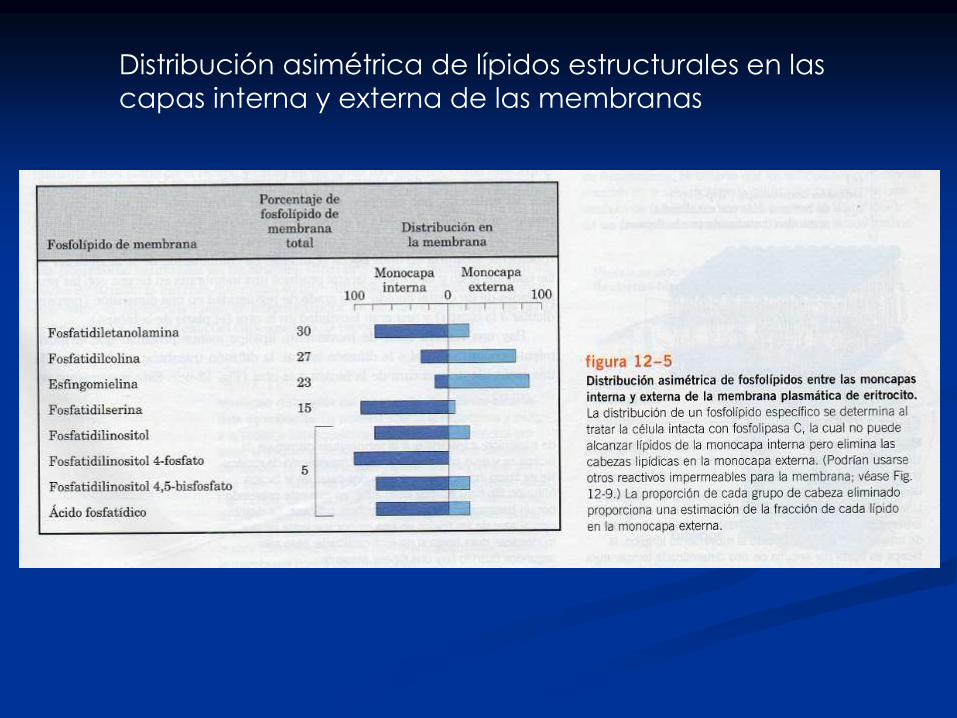
Distribución asimétrica de lípidos estructurales en las
capas interna y externa de las membranas

Modelo del mosaico fluido
Las membranas están compuestas por
lípidos y proteínas, glucolípidos y
glucoproteínas.
Al microscopio electrónico tienen
apariencia trilaminar:
cabezas polares de los fosfolípidos y esteroles:
láminas interior y exterior.
restos no polares: espacio intermedio.
Proteínas y lípidos pueden difundir en el
plano bidimensional de la membrana.
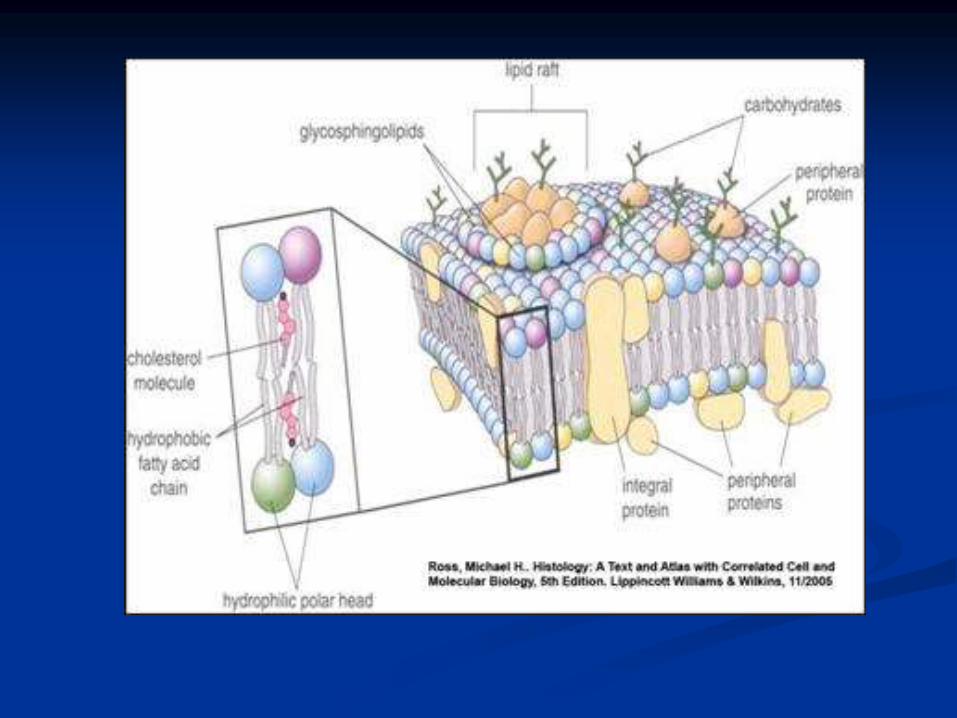


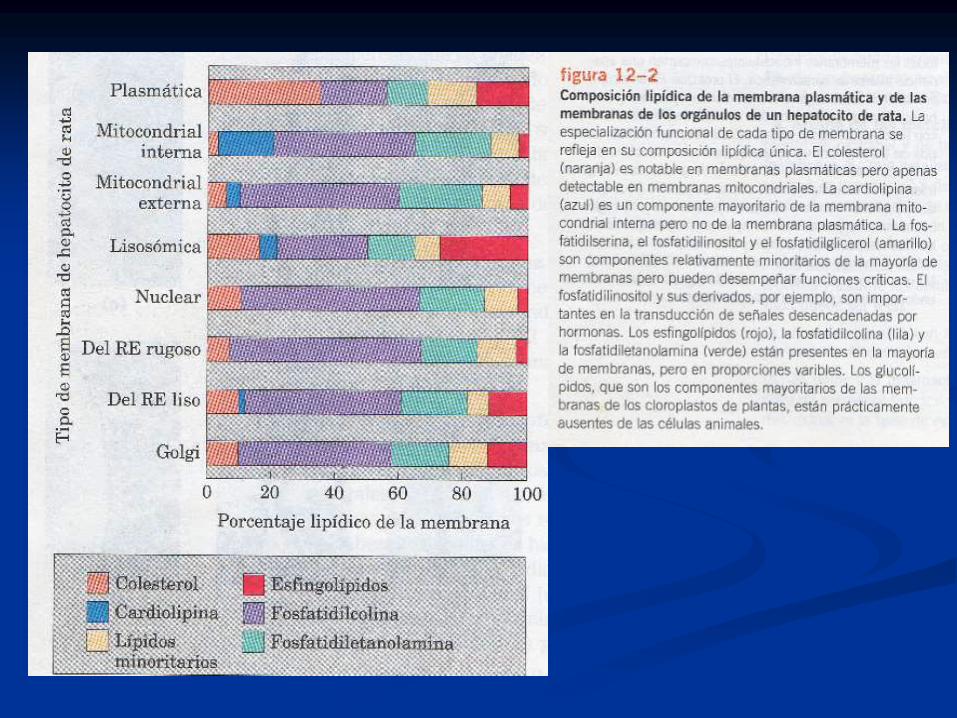
Esfingolípidos: presentes en todas las membranas
de células eucarióticas.
Colesterol: presente en todas las membranas de
células eucarióticas y algunas de bacterias.
Glicerofosfolípidos: fosfato en sn-3-glicerol y ácidos
grasos de cadena larga en sn 1 y 2: presentes en
bacterias y cél eucariotas en general.
Diacilglicerolglicanos neutros: componentes
mayoritarios de membranas de bacterias gram + y
de plantas.
LÍPIDOS PRESENTES EN LAS MEMBRANAS

Diacilglicerolglicanos (ej: diacil galactosil sn1 glicerol)
con mono o disacáridos unidos a C3.
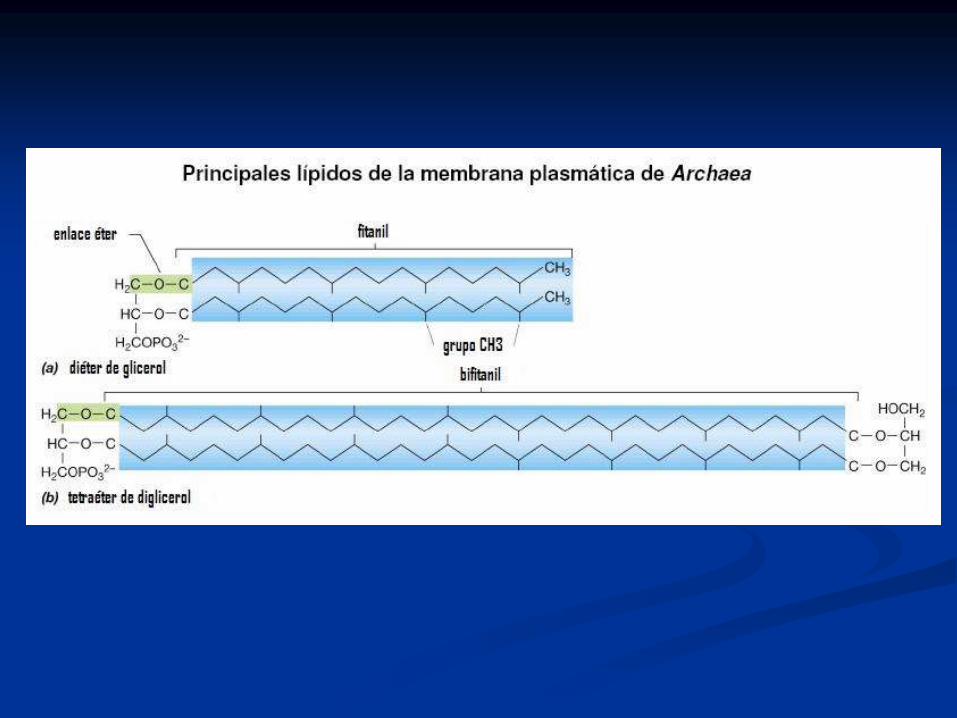
Otros lípidos, presentes en Arquea
(archaebacteria):
Diglicéridos y fosfoglicéridos + isoprenos (unión éter)
Difitanilglicerol (C20-C20 diéter): sn-1-glicerol y dos
isoprenilos saturados (fitanilos) en unión éter en sn 2 y
3.
Bifitanilglicerol (C40) diéter cíclico.
Bifitanilo (C40) diglicerol diéter.
Gliceroglicano con mono o disacárido en posición 1
de sn-1-glicerol.
Sacarolípido (lípido A): componente mayoritario en
membrana externa de bacterias gram –

Sacarolípidos en bacterias gram –
membrana externa: Lípido A
membrana interna: glicerofosfolípidos
E. coli: Lípido A: fosfolípido que contiene dos grupos
glucosamina en unión b 1-6, ácido R3-
hidroximirístico (C14) en posiciones 2, 3, 2´ y 3´ y
fosfato en posiciones 1 y 4´. Modificado en 6´ con
un disacárido KDO (dos 3-desoxi-D-mano ácidos
octulosónicos en unión 1-3) Lípido A - KDO2
modificado con Ag 0
Responsable del síndrome tóxico
Otras bacterias: contienen diferentes ácidos grasos
en posiciones 2, 3, 2´ y 3´

Pírez y Mota. Morfología y estructura bacteriana. Temas
de Bacteriología y virología médica.

Lipopolisacárido: Porción hidrofílica: Ag O (polisacárido O):
especificidad serológica, Núcleo (polisacárido): une al
lípido A.
Presente en la membrana externa de la
envoltura bacteriana
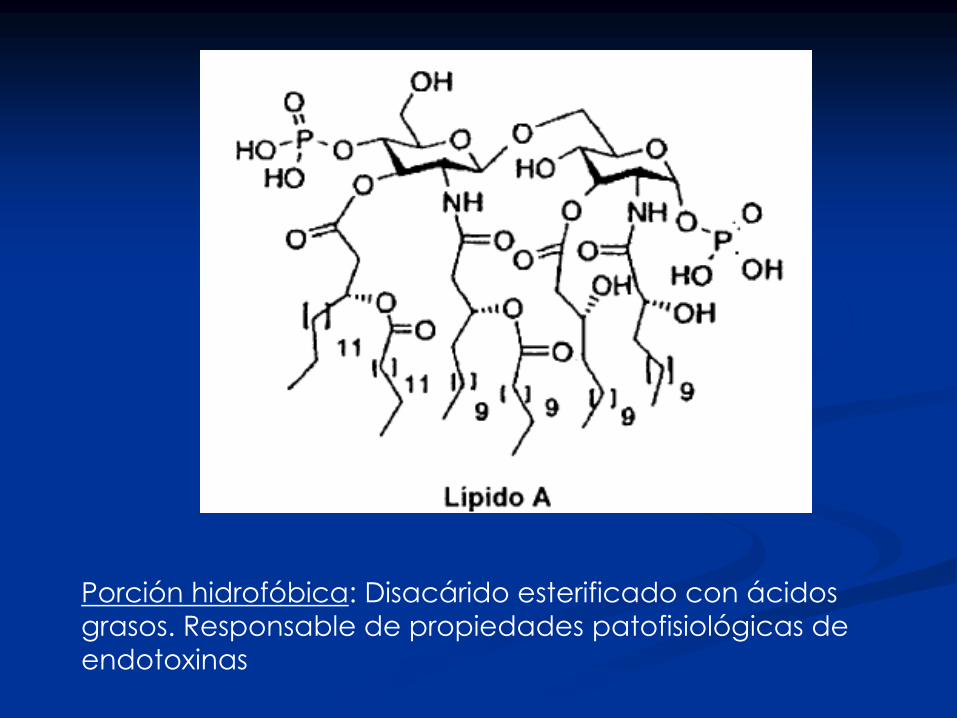
Porción hidrofóbica: Disacárido esterificado con ácidos
grasos. Responsable de propiedades patofisiológicas de
endotoxinas

- Lípido A es liberado cuando la célula se lisa como consecuencia
de la fagocitosis o de la acción antibióticos.
- Cuadro clínico: depende de la cantidad de endotoxina
circulante, desde un simple cuadro infeccioso con fiebre hasta
sepsis, falla multiorgánica y muerte.
- Pequeñas cantidades de endotoxina provocan: fiebre, activación
del complemento por la vía alternativa, activación de los
macrófagos y estimulación de linfocitos B.
- Cuatro blancos primarios de la endotoxina: fagocitos
mononucleares (macrófagos del bazo, de la médula ósea, de los
alvéolos pulmonares y de la cavidad peritoneal, monocitos de la
sangre periférica y células de Kupffer), neutrófilos, plaquetas y
linfocitos B.
- Grandes cantidades de endotoxina producen un shock
endotóxico, (con frecuencia letal): caída severa de la presión
arterial y coagulación intravascular diseminada (CID), entre otros
síntomas.
- CID: depósito de trombos en los vasos de pequeño calibre, con
daño en las áreas privadas de irrigación sanguínea; el consumo de
plaquetas, así como de factores de la coagulación (II, V y VII)
exceden la velocidad de producción conduciendo a hemorragias
internas y falla orgánica (fundamentalmente en pulmón, riñón e
hígado).

Polimorfismo lipídico
https://books.google.com.ar/books?hl=es&lr=&id=LZprT
sjcvIMC&oi=fnd&pg=PP1&dq=lipids+biochemistry&ots=
R2YEGsFPsq&sig=3GtnSa8yHUg7Wv435J_gEr6ejlE#v=on
epage&q=lipids%20biochemistry&f=false
Biochemistry of lipids, lipoproteins and membranes.
Vance y Vance 2008

Bajas cc de lípidos anfipáticos
monómeros en Sc
Mayores cc
asociación de dominios hidrofóbicos
Equilibrio entre monómeros en Sc y micelas
(concentración micelar crítica, CMC)
Para sust anfípáticas con carga neta:
fuerza iónica (NaCl 0,5 M) CMC
Urea cte dieléctrica del agua CMC
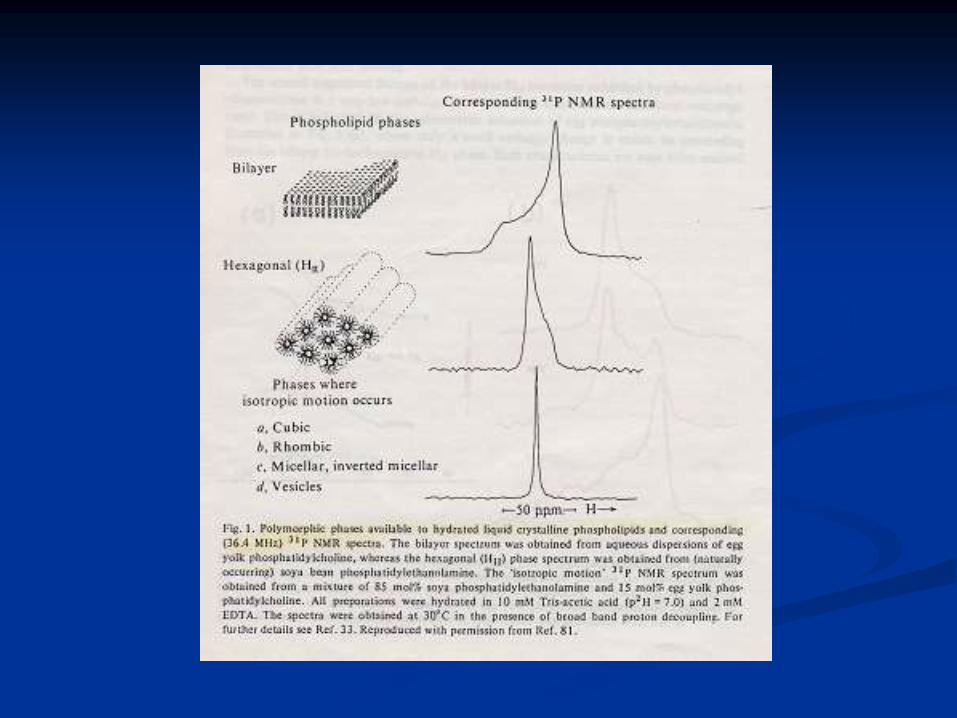

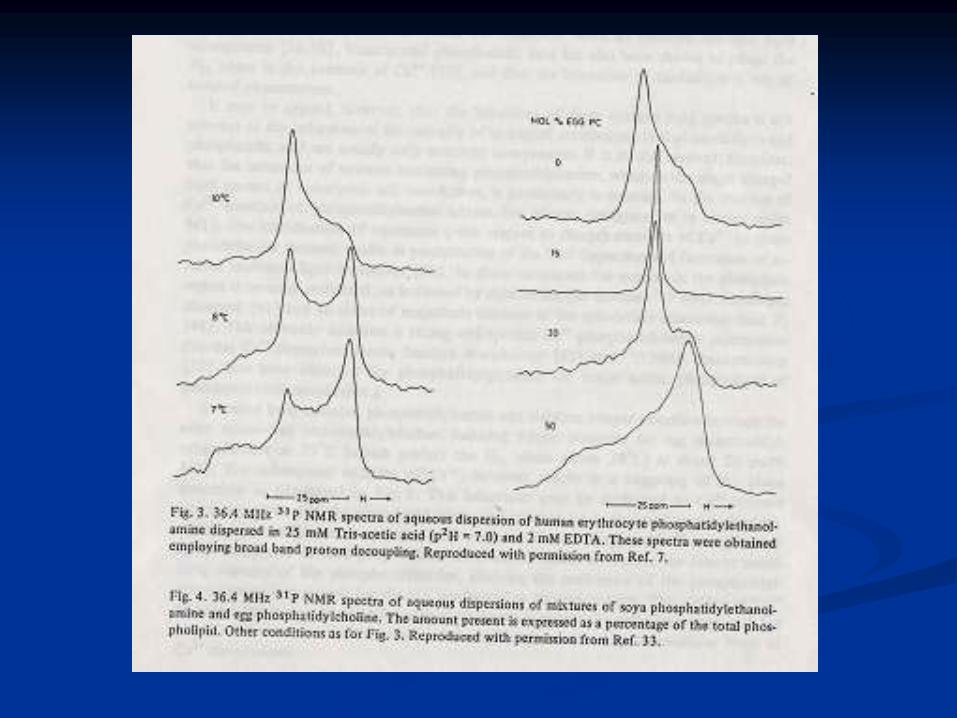

Lípidos puros en solución
Fases “ordered gel” Lb, o “liquid cristalline” La
Viscosidad o fluidez de dominios hidrofóbicos se
modifican con:
Temperatura
Largo de cadena carbonada de ácidos grasos
Insaturaciones
PC en solución: Lb o La
PL no bicapa como PE, CL y PA + Ca2+ (dependiendo de
T):
Lb La HII (arreglos hexagonales)
El agregado de PL no bicapa, a mezcla de PL bicapa
produce cambios de forma en la bicapa, con
discontinuidades y porciones hidrofóbicas de PL no
bicapa expuestas a la fase acuosa, que se atenúan con
la inserción de proteínas.

Adición de colesterol no permite la transición normal:
Inhibe Lb produciendo un estado menos fluido y más
ordenado que La.
Efectos del solvente:
Agregado de Ca2+ u otros cationes divalentes:
Reduce la carga efectiva de grupos de cabeza de CL y
PA favoreciendo HII.
pH ácido, Idem para PS.
Ca2+ papel en procesos de liberación de
contenidos en vesículas.
CL: PL bicapa o no bicapa, dependiendo de presencia
de Ca2+
PL único, actúa como conductor de H+ en las membranas
Deficiencia de CL en mitocondria de mamíferos
liberación de cyt C
inicio de apoptosis

PL afectan la función celular a través de efectos sobre la
estructura y función de las proteínas, mediante dos
mecanismos:
Interacción directa.
Modificaciones de las membranas (fluidez, espesor, forma y
propiedades de empaquetamiento).
Membranas de mamíferos: no modifican su composición
lipídica de acuerdo a su entorno, sí debido a la presencia
de Colesterol (las estabiliza en fase similar a La).
Membranas de microorganismos, frente a diferentes condiciones de crecimiento:
E. coli regula el contenido en ag insaturados en PL no bicapa.
Acholeplasma laidlawii regula el contenido en ácidos grasos
saturados e insaturados (T)
A bajas T: el contenido en ácidos grasos insaturados en
MGlcDG (Monoglucosil DG) o
MGlcDG/DGlcDGR (DiglucosilDG)
Membrana en La con potencial para pasar a HII

Importancia biológica de lo PL no bicapa
Procesos que requieren discontinuidad de la
bicapa: fusión y fisión de vesículas, inserción y
movimiento de proteínas.
Inserción de proteínas en la membrana plasmática: PL
anulares como interfaz entre estructura irregular de la
proteína y estructura regular de la bicapa.
PKC: unión reversible a la capa interna de la membrana plasmática:
activada por: 6 moléc de PS, 1 moléc DAG y Ca2+
En presencia de Ca2+ el isómero natural de PS favorece La HII
DAG siempre favorece HII
PLC: actividad regulada por la geometría y composición
de la bicapa
SPH estabiliza La e inhibe PLC.
No directamente influenciada por PL no bicapa, pero en presencia
de PE, se encuentra activada.

Los PL se asocian a las proteínas de las
membranas (solo a modo informativo
pág 28 a 33):
Con a hélices:
CL en Rhodobacter sphaeroides: cabeza
polar en centro de reacción de proteína
fotosintética y unida con puentes de
hidrógeno a la porción en contacto con el
citoplasma; cadenas hidrocarbonadas en
ranuras de la hélice.
PE en Thermochromatioum tepidum: unión en
centro de reacción de proteína fotosintética,
a Arg y Lys por interacción electrónica y a Tyr
y Gly por puentes de hidrógeno; cadenas
hidrocarbonadas en ranuras de la hélice.

Con proteínas b-Barrel (cadenas bantiparalelas):
Lipopolisacárido unido a proteína formadora
de poro en E. coli: cadenas acílicas orientadas
en paralelo a las hojas b (F de Van der Waals),
cabeza polar interactúa con 8 residuos
cargados en la superficie de la membrana.
Organización de complejos proteínicos:
PL calzan en el interior de subunidades de
complejos oligoméricos:
CL mantiene activ óptima de complejos de
membrana mitcondrial interna: NADH
deshidrogenasa, cyt bc1, ATP sintasa, cyt C oxidasa,
ATP/ADP translocasa.
CL es parte integral de la estructura de succinato
deshidrogenasa de E. coli.

Ubiquinol cyt c reductasa en levaduras
(complejo III de memb mit int): se identificaron
14 moléculas de PL en la estructura molecular
del dímero. 4 CL, 2 PI, 6 PE y 2 PC.
2 PE: contacto entre monómeros, 2 CL cerca de 2
PE, 2 PI intercaladas entre las 3 subunid catalíticas de
c/monómero.
El resto: lípidos anulares inmovilizados en la sup del
complejo (CL y PE: interfase entre complejos III y IV).
Complejos supermoleculares:
Respirasoma: Complejos I a IV de cad resp en
mamíferos en equilibrio con “respirasomas”
(unión de los 4 complejos). CL: rol importante
en la unión. CL en memb int de la mitocondria
atrapa protones, haciendo más eficiente la
síntesis de ATP

Sitios de unión de proteínas periféricas:
Interacción coulómbica entre dominios de la
sup proteica cargada + y dominios aniónicos
de PL de membrana: PA, PG, PI, CL ó PS.
Anillos aromáticos de proteínas expuestos,
interactúan a través de fuerzas “pi”
(cuadrupolo) con cationes lipídicos (cabezas
polares con carga + de PC y PE.
Interacción de sitios específicos de proteínas
con segundos mensajeros lipídicos: PI
fosforilados o DAG.
Inserción parcial de dominios hidrofóbicos de
las proteínas en la membrana: inserción de
una a hélice anfipática.

Translocación de proteínas a través de
la membrana plasmática:
PL aniónicos no bicapa son necesarios:
PG, CL, PE y Dgal DG. Demostrado en
estudios de reconstitución con cepas
mutantes de E coli y Bacillus subtilis,
deficientes en la síntesis de esos PL.
Plegado de proteínas de membrana,
asistido por lípidos: se requiere PE durante
el ensamblado y para la función de 3
proteínas transportadoras en E coli:
lactosa permeasa, fenilalanina permeasa
y g butirato permeasa.

Ensayos de reconstitución:
con PE transporte activo y facilitado
Con CL, PG o PC solo transporte facilitado
Determinantes moleculares de la
topología proteica:
La topología final de las proteínas de
membrana es el resultado de una interacción
finamente sintonizada entre las señales
topogénicas de las proteínas y los
determinantes topológicos dentro de la
membrana, influenciados por la carga neta
de los dominios extramembrana de las
proteínas, los grupos de cabeza de PL y la
interacción PL – proteínas en cada lado de la
membrana.

Dominios lipídicos Visión actual: Las membranas contienen microdominios,
de diferente composición lipídica y proteica, que
favorecen la compartimentalización de las funciones
celulares.
Mezclas definidas de lípidos experimentan separación
de fases debidas a polimorfismo lipídico, diferencias en
largos de cadenas acílicas, etc.
Lípidos no-bicapa experimentan transiciones de fase a
diferentes temperaturas segregación de dominios
en la bicapa.
Lípidos análogos anfipáticos se asocian en los dominios
a pesar de poseer = carga neta, debido a interacciones
hidrofóbicas de sus cadenas carbonadas.

SPH contiene acidos grasos con mayor grado de
saturación y de cadenas más largas (C20-C24) que
los glicerofosfolípidos (C16-C18).
Transición de Lb a La:
Glicerofosfolípidos 0°C
Esfingolípidos 37°C
Dominios compactos hidrofóbicos de SPH
Se separan de los desordenados dominios de
ácidos grasos insaturados de PL
Interacciones tipo puente de H entre lípidos y
entre lípidos y proteínas, contribuyen a la
segregación.

Melkonian et al, (1999) Role of Lipid Modifications in Targeting Proteins to
Detergent-resistant Membrane Rafts
MANY RAFT PROTEINS ARE ACYLATED, WHILE FEW ARE PRENYLATEDhttp://www.jbc.org/content/274/6/3910.long



Science 327, 46
DOI: 10.1126/science.1174621
Lipid Rafts As a Membrane-
Organizing Principle
Daniel Lingwood, et
al. (2009)