
MEMBRANA CELULAR - mmegias.webs.uvigo.esmmegias.webs.uvigo.es/...03-membrana-celular.pdf · Esquema...
31
Atlas de Histología Animal y Vegetal MEMBRANA CELULAR Manuel Megías, Pilar Molist, Manuel A. Pombal DEPARTAMENTO DE BIOLOGÍA FUNCIONAL Y CIENCIAS DE LA SALUD. FACULTAD DE BIOLOGÍA.UNIVERSIDAD DE VIGO. (VERSIÓN:JULIO 2017)
Transcript of MEMBRANA CELULAR - mmegias.webs.uvigo.esmmegias.webs.uvigo.es/...03-membrana-celular.pdf · Esquema...

Atlas de Histología Animal y Vegetal
MEMBRANA CELULAR
Manuel Megías, Pilar Molist, Manuel A. Pombal
DEPARTAMENTO DE BIOLOGÍA FUNCIONAL Y CIENCIAS DE LA SALUD.FACULTAD DE BIOLOGÍA. UNIVERSIDAD DE VIGO.
(VERSIÓN: JULIO 2017)

Este documento es una edición en pdf del sitiohttp://webs.uvigo.es/mmegias/inicio.html
y
ha sido creado con el programa Scribus
(http://www.scribus.net/)
Todo el contenido de este documento se distribuye bajo la licenciaCreative Commons del tipo BYNCSA (Esta licencia permite modificar,
ampliar, distribuir y usar sin restricción siempre que no se use para finescomerciales, que el resultado tenga la misma licencia y que se nombre a
los autores).

ÍNDICE
Membrana celular ........................................................ 4
Lípidos ........................................................................ 6
Proteínas ..................................................................... 10
Glúcidos ...................................................................... 12
Permeabilidad, fluidez, heterogeneidad ...................... 13
Asimetría, reparación, fusión ...................................... 18
Síntesis ........................................................................ 22
Transporte ................................................................... 23
Adhesión ..................................................................... 26
Complejos ................................................................... 28
Bibliografía ................................................................... 31
Atlas de histología vegetal y animal. Universidad de Vigo.

La célula. Membrana celular. 4
Atlas de histología vegetal y animal. Universidad de Vigo.
Tras dejar el espacio pericelular en nuestro viajehacia la célula tropezamos con la membrana plasmáticade la célula. Ésta es una estructura vital. La rotura de lamembrana plasmática durante más de unos pocossegundos lleva irremisiblemente a la muerte celular. Esuna barrera física que separa el medio celular internodel externo. En las células eucariotas, y en algunasprocariotas, también hay membranas intracelulares quedelimitan a los orgánulos, separando el medio internodel orgánulo del citosol.
Composición y estructura. Las membranas celularesestán formadas por lípidos, proteínas y, en menormedida, por glúcidos. La estructura y la organizaciónde las membranas celulares, así como sus propiedades,están condicionadas fundamentalmente por los lípidos.Éstos son moléculas anfipáticas, con una partehidrofílica y otra hidrofóbica, que se disponenformando una bicapa lipídica donde las parteshidrofóbicas se encuentran en el centro de la membranay las hidrofílicas en contacto con el agua. Entre loslípidos se anclan las proteínas denominadas integrales.Las proteínas transmembrana son proteínas integralesque poseen secuencias de aminoácidos hidrofóbicosentre las cadenas de los ácidos grasos de los lípidos, y
dominios hidrofílicos que están en contacto con lasolución acuosa intra y extracelula. También hayproteínas asociadas a una u otra superficie de la bicapalipídica y que pueden disociarse de ella. Los glúcidosno aparecen en todas las membranas, por ejemplo enalgunas intracelulares, pero son abundantes en la quedelimita la célula con el medio externo, la membranaplasmática. Se localizan en la superficie extracelular.Los glúcidos se encuentran unidos covalentemente alos lípidos o a las proteínas.
Por tanto las membranas son como láminas extensasque cuando se observan en secciones transversales,perpendiculares a sus superficies, con el microscopioelectrónico presentan un aspecto trilaminar: dos franjasoscuras que corresponden con las partes hifrofílicas delos lípidos y una franja clara más ancha entre ellas queson sus cadenas de ácidos grasos. A esto se denominaunidad de membrana y es así para todas las membranascelulares. El espesor de las membranas varía entre los 6y los 10 nm, lo cual indica que no todas las membranasson exactamente iguales.
Las propiedades fisiológicas y estructurales de lasmembranas dependen de la proporción y del tipo de
Esquema de la organización de una membrana plasmática según la visión actual del modelo de mosaico fluido de Singer y
Nicolson (1972). Es una bicapa fluida estructurada por los lípidos pero heterogénea en su organización. Determinados
lípidos se asocian entre sí para formar agrupaciones más densas denominadas balsas de lípidos, en la cuales se sitúan
ciertas proteínas por afinidad eléctrica. El colesterol se localiza entre las cadenas de ácidos grasos, cerca de la zona
hidrofílica ("cabezas" de los lípidos). Las proteínas transmembrana comunican el exterior (arriba) con el interior (abajo) de la
célula. Los glúcidos se localizan en la parte extracelular formando el glucocálix. En este esquema no se muestran las
interacciones con la matriz extracelular ni con las moléculas del citoesqueleto. (Modificado de Edidin, 2003 y Nicolson 2014)

Atlas de histología vegetal y animal. Universidad de Vigo.
moléculas que las componen: lípidos, proteínas yglúcidos. Así, la membrana de los eritrocitos de ratacontiene un 50 % de lípidos, un 40 % de proteínas y un10 % de glúcidos. Una proporción similar a ésta es lamás común entre las membranas plasmáticas de todaslas células animales, con algunas excepciones. Porejemplo, la mielina formada por las membranasplasmáticas de las células de Schwan, que rodean a losaxones situados fuera del sistema nervioso central,contienen un 80 % de lípidos y un 20 % de proteínas.Las membranas intracelulares suelen contener unamayor proporción de proteínas que la membranaplasmática. La mayor diferencia la encontramos en lasmitocondrias donde el porcentaje de proteínas de sumembrana interna llega hasta el 80 %. Por supuesto,lípidos, proteínas y glúcidos son grupos heterogéneosde moléculas y también las membranas celulares sediferencian en la composición y en la proporción dedistintos tipos de lípidos, de proteínas y de glúcidos.Además, como dijimos anteriormente, las membranasestán en una constante renovación que permite a lacélula cambiar su composición.
Propiedades. Parte de las funciones de lasmembranas son debidas a sus propiedades físicoquímicas: a) es una estructura fluida que hace que susmoléculas tengan movilidad lateral, como si de unalámina de líquido viscoso se tratase; b) essemipermeable, por lo que puede actuar como unabarrera selectiva frente a determinadas moléculas; c)posee la capacidad de romperse y repararse de nuevosin perder su organización, es una estructura flexible ymaleable que se adapta a las necesidades de la célula;d) está en permanente renovación, es decir, eliminacióny adición de moléculas que permiten su adaptación alas necesidades fisiológicas de la célula.
Funciones. Cada tipo de membrana estáespecializada en una o varias funciones dependiendo
del compartimento celular del que forme parte. Entrelas múltiples funciones necesarias para la célula querealizan las membranas están la creación ymantenimiento de gradientes iónicos, los cuales hacensensible a la célula frente a estímulos externos,permiten la transmisión de información y la producciónde ATP, son necesarios para la realización deltransporte selectivo de moléculas , etcétera. Lasmembranas también hacen posible la creación decompartimentos intracelulares donde se realizanfunciones imprescindibles o la envuelta nuclear queencierra al ADN. En las membranas se disponenmúltiples receptores que permiten a la célula "sentir" lainformación que viaja en forma de moléculas por elmedio extracelular. Por ejemplo, dan a las neuronas suspropiedades y capacidades, también a las musculares.Tambén poseen enzimas asociadas que realizannumerosas actividades metabólicas, como la síntesis decelulosa o de ácido hialurónico, fosforilaciones,producción de energía, síntesis de lípidos, etcétera.Como hemos visto en el apartado anterior, laadhesividad celular a la matriz extracelular o a otrascélulas en los tejidos animales se debe a las moléculaspresentes en la membrana plasmática.
En los siguientes apartados veremos loscomponentes moleculares, para después tratar laspropiedades de las membranas celulares y algunas desus funciones más importantes. En capítulosposteriores veremos que las membranas celulares delos orgánulos participan de forma determinante en susfunciones, en el trasiego de moléculas en el interior dela célula mediante el denominado tráfico vesicular, asícomo en la incorporación y liberación demacromoléculas entre el interior y el exterior celular enlos procesos de endocitosis y exocitosis,respectivamente.
La célula. Membrana celular. 5

Atlas de histología vegetal y animal. Universidad de Vigo.
La organización de las membranas celulares estádeterminada por las características de suscomponentes, fundamentalmente por los lípidos. Losotros componentes importantes de las membranascelulares son las proteínas, principales actores en lasfunciones celulares asociadas a las membranas, y losglúcidos. Sin embargo, la diversidad (hay más de miltipos de lípidos diferentes) y su organización espacial(formando un picapa) hacen a los lípidos esenciales.Así, ellos definen las propiedades físicas de lasmembranas. La longitud y el grado de saturación de susácidos grasos regulan la fluidez y el grosor de lamembrana, y su distribución desigual crea asimetría enlas membranas. En la membrana plasmática las cargasasociadas a sus partes hidrofílicas contribuyen a crearun gradiente eléctrico entre la cara externa y la interna,y por tanto a modular el potencial eléctrico. Medianteinteracciones electroquímicas son capaces de modularla actividad de las proteínas de membrana. Se hapostulado que las interacciones moleculares entreciertos lípidos producen la segregación de dominiosespaciales y funcionales en áreas restringidas de lamembrana que afectan también a la localización de lasproteínas y a sus funciones. Son las denominadasbalsas de lípidos o "lipid rafts". Pero ademáspueden actuar como segundos mensajeros queabandonan la membrana, viajan a compartimentosintracelulares y desencadenan respuestas celulares.
Los lípidos constituyen aproximadamente el 50% del peso de las membranas, con unos 5 millonesde moléculas por µm2. Las membranas celularesde una célula eucariota contienen más de 1000tipos de lípidos que aparecen en distintaproporción según el tipo de membrana queestemos considerando. Se estima queaproximadamente el 5 % de los genes de unacélula están dedicados a producir sus lípidos.
Los lípidos de membrana se caracterizan porposeer una parte apolar o hidrófoba que constituyela parte interna de la membrana y por una partehidrofílica que está en contacto con el medioacuoso. Por ello se dice que son moléculasafipáticas. Se clasifican en tres grupos según suestructura y composición molecular:
glicerofosfolípidos (también denominadosglicerolípidos, fosfoglicéridos o simplementefosfolípidos), los esfingolípidos y los estéroles.
Fosfoglicéridos o glicerofosfolípidos
Son los lípidos más abundantes (representan más del70 %) de las membranas celulares y estructuralmenteconstan de tres partes: dos cadenas de ácidos grasos,glicerol, un ácido fosfato al que se unen moléculas dediversa naturaleza y que aportan gran parte de lavariabilidad de estos lípidos. Las cadenas de ácidosgrasos contienen de 13 a 19 átomos de carbono delongitud. La mayoría de los enlaces entre estoscarbonos son simples y por tanto se dice que sonenlaces saturados. Sin embargo, más de la mitad deestos ácidos grasos tienen al menos un doble enlaceentre dos átomos de carbono, hablamos entonces deácidos grasos insaturados. Los dobles enlaces hacenque la cadena de ácido graso se doble y, aunquerestrinja las posibilidades de movimiento de la cadena,un aumento de la proporción de estos dobles enlacesaumenta la fluidez de la membrana puesto que provocamás separación entre moléculas. Los ácidos grasos
Estructura y tipos de glicerofosfolípidos más abundantes de las
membranas eucariotas.
La célula. Membrana celular. 6

constituyen la parte hidrofóbica (fobia por el agua) delos glicerofosfolípidos y son los que constituyen laparte interna de las membranas.
El glicerol hace de puente entre los ácidos grasos yla parte hidrofílica (apetencia por el agua). Estecomponente hidrofílico está formado por un grupofosfato al que se pueden unir una variedad demoléculas, tales como la etonalamina, colina, serina,glicerol, inositol, el inositol 4,5bifosfato, etcétera.Estos componentes son los que dan nombre a losdistintos tipos de glicerofosfolípidos. El tipofosfatidilcolina representa más del 50 % de losfosfoglicéridos en las membranas eucariotas.
Esfingolípidos
Deben su nombre a que poseen una molécula deesfingosina, un alcohol nitrogenado con una cadenacarbonada larga, a la cual se le une una cadena de ácidograso, formando la estructura básica denominadaceramida (ver figura ). A la ceramida se le une unaparte hidrofílica que puede de ser de diversanaturaleza. Por tanto queda una estructura similar a lade los glicerofosfolípidos, dos cadenas hidrofóbicasunidas a una estructura hidrofílica. Los esfingolípidosconstituyen la mayoría de los denominadosglicolípidos de las membranas, es decir, lípidos queposeen uno o más azúcares unidos formando parte desu zona hidrofílica. Otro tipo de esfingolípidos son lasesfingomielinas que poseen una etanolamina o unacolina fosforiladas en sus zonas hidrofílicas. Losesfingolípidos son más abundantes en las mebranasplasmáticas que en las de los orgánulos, y se lespropone como lo principales responsables, junto con elcolesterol, de la segregación lateral de la membrana endominios moleculares (balsas de lípidos).
Esteroles
El colesterol es el esterol másimportante de las células animales y eltercer tipo de lípido más abundante enla membrana plasmática (hasta el 25 %del total de lípidos), mientras queaparece en pequeñas proporciones enlas membranas de los orgánuloscelulares (1 % en el retículoendoplasmático). El colesterol selocaliza entre las cadenas de ácidosgrasos de los otros lípidos. Es importante para laestructura de la membrana puesto que junto con losesfingolípidos parece contribuir a formarheterogeneidades en la distribución molecular ytambién participa en ciertos procesos metabólicosvitales como la síntesis de hormonas esteroideas o desales biliares, entre otras. El colesterol no aparece enlas membranas de las plantas, en algunas célulaseucariotas, ni en las bacterias, pero estas células tienenotro tipo de esteroles. Los estéroles son esenciales parala integridad y funcionamiento de las membranaseucariotas. Sirven para modular la rigidez, la fluidez yla permeabilidad.
La distribución de lípidos depende de lamembrana
Aunque hay tres grandes grupos de lípidos demembrana (glicerolípidos, esfingolípidos y esteroles),hay miles de tipos moleculares diferentes de lípidosdistribuidos por las membranas celulares. Lacomposición de lípidos varía entre las membranas delos diferentes compartimentos membranosos de lacélula. Se propone que la propia identidad de losorgánulos viene determinada por la composición de sus
membranas, tanto proteínascomo lípidos. Por ejemplo, lamembrana plasmática tieneuna composición lipídicadiferente a la membrana delretículo endoplasmático o a ladel aparato de Golgi. Estasdiferencias se mantienen apesar del flujo constante delípidos desde suscompartimentos de síntesis,principalmemente el retículoendoplasmático, hasta otras
Atlas de histología vegetal y animal. Universidad de Vigo.
Esfingolípidos.
Colesterol
La célula. Membrana celular. 7
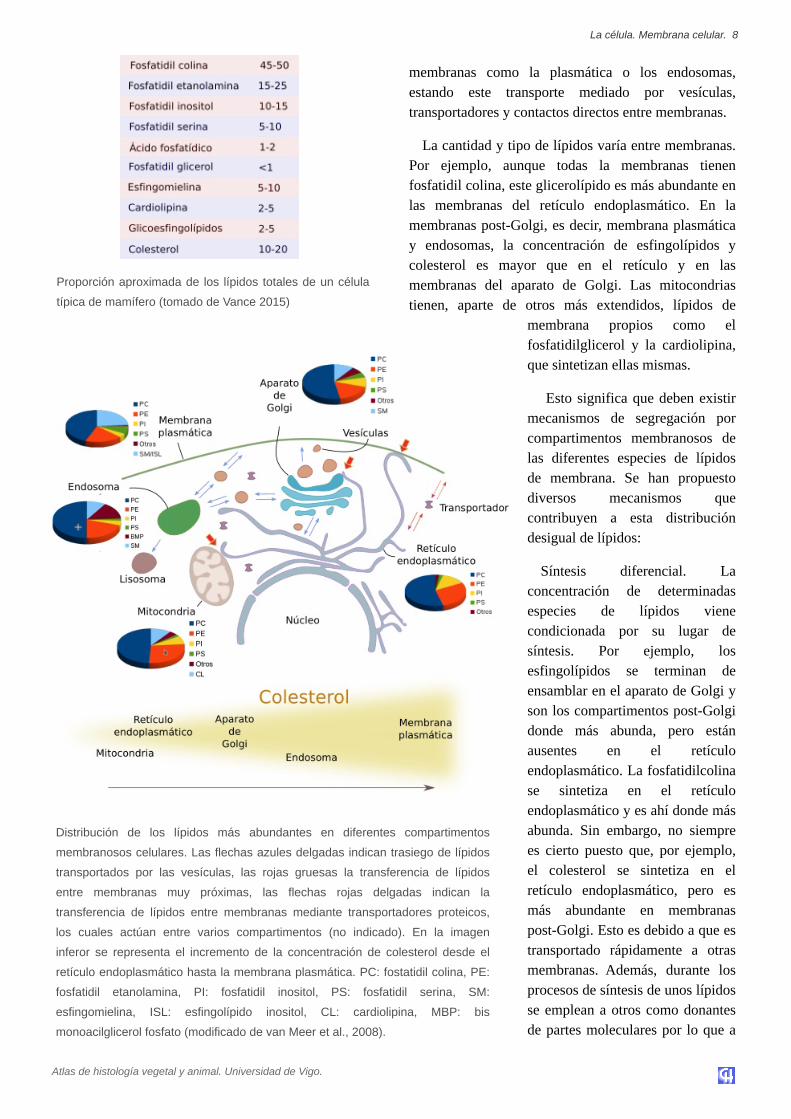
membranas como la plasmática o los endosomas,estando este transporte mediado por vesículas,transportadores y contactos directos entre membranas.
La cantidad y tipo de lípidos varía entre membranas.Por ejemplo, aunque todas la membranas tienenfosfatidil colina, este glicerolípido es más abundante enlas membranas del retículo endoplasmático. En lamembranas postGolgi, es decir, membrana plasmáticay endosomas, la concentración de esfingolípidos ycolesterol es mayor que en el retículo y en lasmembranas del aparato de Golgi. Las mitocondriastienen, aparte de otros más extendidos, lípidos de
membrana propios como elfosfatidilglicerol y la cardiolipina,que sintetizan ellas mismas.
Esto significa que deben existirmecanismos de segregación porcompartimentos membranosos delas diferentes especies de lípidosde membrana. Se han propuestodiversos mecanismos quecontribuyen a esta distribucióndesigual de lípidos:
Síntesis diferencial. Laconcentración de determinadasespecies de lípidos vienecondicionada por su lugar desíntesis. Por ejemplo, losesfingolípidos se terminan deensamblar en el aparato de Golgi yson los compartimentos postGolgidonde más abunda, pero estánausentes en el retículoendoplasmático. La fosfatidilcolinase sintetiza en el retículoendoplasmático y es ahí donde másabunda. Sin embargo, no siemprees cierto puesto que, por ejemplo,el colesterol se sintetiza en elretículo endoplasmático, pero esmás abundante en membranaspostGolgi. Esto es debido a que estransportado rápidamente a otrasmembranas. Además, durante losprocesos de síntesis de unos lípidosse emplean a otros como donantesde partes moleculares por lo que a
Distribución de los lípidos más abundantes en diferentes compartimentos
membranosos celulares. Las flechas azules delgadas indican trasiego de lípidos
transportados por las vesículas, las rojas gruesas la transferencia de lípidos
entre membranas muy próximas, las flechas rojas delgadas indican la
transferencia de lípidos entre membranas mediante transportadores proteicos,
los cuales actúan entre varios compartimentos (no indicado). En la imagen
inferor se representa el incremento de la concentración de colesterol desde el
retículo endoplasmático hasta la membrana plasmática. PC: fostatidil colina, PE:
fosfatidil etanolamina, PI: fosfatidil inositol, PS: fosfatidil serina, SM:
esfingomielina, ISL: esfingolípido inositol, CL: cardiolipina, MBP: bis
monoacilglicerol fosfato (modificado de van Meer et al., 2008).
Atlas de histología vegetal y animal. Universidad de Vigo.
Proporción aproximada de los lípidos totales de un célula
típica de mamífero (tomado de Vance 2015)
La célula. Membrana celular. 8

la vez que se sintetiza una nueva especie lipídicadesaparece otra, y todo contribuye a cambiar lacomposición lipídica de la membrana de esecompartimento.
Selección y transporte. Los lípidos deben sertransportados entre membranas puesto que no difundenlibremente por el citosol. Ello implica que eltransportador puede seleccionar qué lípidos acarrea deun lado a otro cambiando así las proporciones dedeterminadas especies de lípidos. Estos transportadoresson fundamentalmente las vesículas, las cuales formansus membranas con lípidos de las propias membranasdel compartimento de partida. Por ejemplo, lasvesículas que van desde el Golgi a la membrana y a losendosomas están enriquecidas en esfingolípidos ycolesterol, respecto la concentración de estas moléculasen las propias membranas del Golgi. También haytransportadores de lípidos individuales que sonproteínas que son capaces de extraer un lípido de unamembrana, transportarlo por el citosol y colocarlo enotra membrana.
Contactos entre membranas. Las membranas dedeterminados orgánulos pueden observarseextremadamente próximas. Hay proteínas que secolocan entre las membranas de dos orgánulos cuandoestán muy próximas espacialmente, haciendo de puentepara el trasiego de lípidos entre las membranas deambos orgánulos. Esto ocurre entre membranas decompartimentos que no están comunicados mediante
vesículas, por ejemplo, entre el retículo endoplasmáticoy las mitocondrias. Pero también entre compartimentoscomunicados por membranas como el retículoendoplasmático y el lado trans del aparato de Golgi,entre el retículo endoplasmático y los endosomas, oentre el retículo endoplasmático y la membranaplasmática. Por ejemplo, el complejo proteico CERT(ceramide transfer protein) transfiere ceramida desdelas membranas del retículo endoplasmático al ladotrans del aparato de Golgi.
Degradación y reciclado diferencial. Todas lasmembranas sufren un proceso de reciclado de lípidos,bien por degradación in situ de los lípidos o por susalida en vesículas de reciclado. En ambos procesos esposible un mecanismo de selección de unos tipos delípidos respecto a otros que condicionará la poblaciónde lípidos de la membrana.
Asimetría
La asimetría es la distribución diferencial de lípidosentre las dos monocapas de lípidos que componen lasmembranas. Mientras en el reticulo endoplasmáticoambas monocapas son muy similares,pero no idénticas,en cuanto a la composición lipídica, las membranas dellado trans del aparato de Golgi y los compartimentospostGolgi suelen tener una clara diferencia decomposición entre monocopas. Para saber más ver(asimetría de membrana).
La célula. Membrana celular. 9
Atlas de histología vegetal y animal. Universidad de Vigo.

Atlas de histología vegetal y animal. Universidad de Vigo.
Las proteínas son las responsables de muchas de lasfunciones que tienen su base en las membranascelulares. El contenido de proteínas de las membranasrespecto a los lípidos es varibale dependiendo del tipode membrana, pero puede ser de cerca de 1:40 ennúmero de moléculas, lo que indicaría que lasmembranas contienen bastantes proteínas. Inclusopuede ser más alta en aquellas membranas confunciones netamente metabólicas como la membranainterna de las mitocondrias. Hay dos grandes grupos deproteínas relacionadas con las membranas: lasintegrales y las asociadas.
Proteínas integrales
Las proteínas integrales son aquellas que formanparte de la mambrana de manera permanente. Sedividen en tres grupos: las transmembrana, laintegradas en una monocapa y las unidascovalentemente a moléculas que forman parte de lamembrana.
Las proteínas transmembrana poseen tres tipos dedominios en sus secuencias de aminoácidos: uno
extracelular, uno intracelular y otro en el interior en lapropia membrana. En el intracelular poseen secuenciasde aminoácidos con radicales hidrofóbicos que sesitúan entre las cadenas de ácidos grasos de los lípidosde la membrana, mientras que los dominios intra yextracelular poseen secuencias de aminoácidos conradicales hidrofílicos. Estas proteínas sonmayoritariamente producidas en el retículoendoplasmático y repartidas por otras membranas de lacélula principalmente mediante el tráfico vesicular,como veremos en capítulos posteriores.
Existen proteínas transmembrana cuya cadena deaminoácidos cruza una sola vez la membrana mientrasque otras pueden hacerlo hasta 7 veces. Muchasproteínas transmembrana realizan su función cuando seasocian con otros polipéptidos también integrales paraformar estructuras oligoméricas (más de un elemento).Las funciones son muy variadas, pero destacan laadhesión llevada a cabo por las integrinas, cadherinas,selectinas y otras; el intercambio de iones, calcio, sodioo potasio, entre ambos lados de la membrana comohacen las bombas de iones y los canales iónicos, loscuales permiten gradientes que hacen posible, porejemplo, la sínteis de ATP; permitir el cruce demoléculas como la glucosa por parte de lostransportadores; algunas participan en la comunicacióncelular y actúan como receptores de señales como losque reconocen los factores de crecimiento,neurotransmisores, hormonas y otros. En este últimocaso, la organización en dominios extracelular eintracelular permite una comunicación entre amboslados de la membrana, lo cual hace que unainformación extracelular, por ejemplo una hormonareconocida por el dominio extracelular, provoque uncambio conformacional en el dominio intracelular yesto desencadene una cascada de señales intracelularesque alteren la fisiología celular, incluso la expresióngénica.
Hay proteínas integrales que forman parte de lamembrana pero que no son transmembrana. Un tiposon aquellas que poseen parte de su secuencia deaminoácidos integrada entre los lípidos de una de lasmonocapas de la membrana, y por tanto tienen undominio extramembranoso que puede ser citosólico oextracelular, según en que monocapa de la membrana
En este esquema se muestran los principales tipos de
proteínas transmembrana. Las hay con un solo cruce
como la glicoforina o con varios como algunos receptores.
En ambos casos la secuencia o secuencias de
aminoácidos localizadas entre las cadenas de ácidos
grasos adoptan una conformación en alfa hélice. La
aquaporina, un canal que cruza numerosas veces la
membrana, posee secuencias de aminoácidos de la zona
hidrofóbica que se disponen en hebras beta. (Modificado
de Pollard et al., 2007)
La célula. Membrana celular. 10

estén integradas. Otro tipo de proteínas integrales, notransmembrana, son aquellas que se encuentranancladas por enlaces covalentes a moléculas de lamembrana, como puede ser un ácido graso. En estecaso toda la secuencia de aminoácidos esextramembranosa, pudiendo ser citosólica oextracelular. En este tipo podemos también incluir a lasproteínas que están unidas covalentemente a losglúcidos de los lípidos. A las proteínas que no sontransmembrana se les puede llamar también periféricaspuesto que sólo están relacionadas con una monocapa.Las proteínas periféricas Incluyen también a lasproteínas asociadas (ver más abajo).
Proteínas asociadas
Las proteínas asociadas a las membranas plasmáticasson aquellas que no forman parte permanente de lasmembranas, es decir, no son proteínas integrales, y suunión a la membrana se produce por interaccioneseléctricas con moléculas de la propia membrana(fuerzas de van der Waals). Estas asociaciones son máslábiles y las proteínas pueden abandonar la membranacon cierta facilidad. Son proteínas hidrosolubles.
Atlas de histología vegetal y animal. Universidad de Vigo.
Esquema de las principales formas de proteínas periféricas:
integrales y asociadas. De izquierda a derecha: proteínas
que tienen una parte de su secuencia de aminoácidos
inserta en una de las monocapas de la membrana, proteínas
que están unidas covalentemente a azúcares de los
glucolípidos, proteínas que tienen ácidos grasos unidos
covalente, lo que les permite insertarse en la zona hidrófoba
de la membrana. Proteínas que interactúan eléctricamente
con las cabezas de los lípidos; y proteínas que interactúan
con los dominios extramembrana de proteínas
transmembrana. (Modificado de Alberts et al., 2002).
La célula. Membrana celular. 11

Los glúcidos presentes en las membranas estánunidos covalentemente a los lípidos formando losglicolípidos y a las proteínas formando lasglicoproteínas de membrana. Algunos proteoglicanosinsertan sus cadenas de aminoácidos con radicaleshidrófobos en la membrana, quedando losglicosaminoglicanos hacia el exterior. Aunque existenglúcidos en las membranas intracelulares, tantoglucolípidos como glucoproteínas son mucho másabundantes en la membrana plasmática,preferentemente localizados en la monocapa externa.Los glúcidos de las membranas se ensamblanprincipalmente en el aparato de Golgi, aunque tambiénen el retículo endoplasmático.
Hay tres tipos de glicolípidos: losglicoesfingolípidos, que son los más abundantes, losgliceroglicolípidos y los glicosilfosfatidilinositoles.Los gliceroglicolípidos son típicos de las membranasplasmáticas de las plantas. Sin embargo, la mayoría delos glúcidos de membrana se encuentran asociados alas proteínas, denominadas glicoproteínas. Mientrasque prácticamente todas las proteínas contienensacáridos unidos, sólo un 5 % de los lípidos los poseen.Al conjunto de glúcidos localizados en la membranaplasmática se les denomina glicocálix. En algunos tiposcelulares la cantidad de glúcidos que se encuentran enlas superficie celular es tan grande que puedeobservarse con el microscopio electrónico. En algunascélulas como en los enterocitos el glicocálix se puedeextender más de 1 µm desde la membrana celular. Lacélula queda así recubierta por una envuelta deglúcidos que representa entre el 2 y el 10 % del peso dela membrana plasmática. El grado de desarrollo delglicocálix depende del tipo celular.
No se puede considerar a los glúcidos comomoléculas inertes sino que tienen papeles importantesen el funcionamiento celular, fundamentalmente actúancomo lugares de reconocimiento y unión. Por ejemplo,los grupos sanguíneos vienen determinados porglúcidos de la membrana, lo que implica que tienencapacidad de respuesta inmunitaria. Cuando se produceuna infección, las células endoteliales próximasexponen una serie de proteínas llamadas selectinas que
reconocen y unen sacáridos de los linfocitos circulantesen el torrente sanguíneo y permiten su adhesión y elcruce del propio endotelio para dirigirse hacia la zonainfectada. El reconocimiento celular mediado por losglúcidos es también muy importante durante eldesarrollo embrionario. Son también unos de losprincipales lugares de reconocimiento por parte de lospatógenos para unirse e infectar a las células. Los viruscomo el de la gripe, bacterias como las E. colipatógenas y protozoos patógenos deben adherirse a lasuperficie celular para infectar, de otra manera seránbarridos por los mecanismos de limpieza delorganismo. Estos organismos patógenos poseen unasproteínas de membrana denominadas lectinas quetienen afinidad por determinados azúcares o cadenas deazúcares y por tanto sólo reconocerán a las células quelos posean. La selectividad en la infección dedeterminados tipos celulares depende de lacomposición de azúcares de su glicocálix. Existendiferencias entre los glúcidos de la membranas devertebrados, invertebrados y protozoos.
Atlas de histología vegetal y animal. Universidad de Vigo.
Esquema de algunas moléculas glicosidadas de la
membrana plasmática. Los glicolípidos son principalmente
esfingolípidos con diferente composición de glúcidos.
Algunos proteoglicanos tienen su parte proteica insertada
entre las cadenas de ácidos grasos. Numerosos glúcidos
de la membrana forman parte de las glicoproteínas,
La célula. Membrana celular. 12

La composición química de las membranas hace queposean unas propiedades esenciales para las funcionesque desempeñan en la célula. Podemos agrupar dichaspropiedades en cinco: semipermeabilidad, asimetría,fluidez, reparación y renovación.
Semipermeabilidad
Esta propiedad es consecuencia del ambientehidrófobo interno de la membrana creado por lascadenas de ácidos grasos de los lípidos, difícil decruzar por las moléculas con carga eléctrica neta. Estopermite a las membranas crear compartimentosintracelulares o mantener separados el mediointracelular del extracelular y, por tanto, impedir lalibre difusión de diversos tipos de moléculas a sutravés. Sin embargo, la permeabilidad es selectiva. Lasvariables que más influyen en la difusión pasiva son lapolaridad y el tamaño de la molécula. Así, moléculaspequeñas sin carga, por ejemplo el CO2, N2, O2, omoléculas con alta solubilidad en grasas como el etanolcruzan las membranas prácticamente sin oposición, porun proceso de difusión pasiva. La permeabilidad de lamembrana es menor para aquellas moléculas concargas pero globalmente neutras (el número de cargasnegativas iguala al de cargas positivas) como el agua oel glicerol. Se podría pensar que el agua difundelibremente por las membranas pero no es así y por elloen determinadas membranas existen unas moléculas
denominadasacuaporinas quefacilitan el crucede la membranapor parte delagua. Es menoraún la capacidadde atravesar lamembrana paramoléculasgrandes neutraspero con cargas,como la glucosa.Sin embargo, esaltamenteimpermeable alos iones y a lasmoléculas que
también carga neta. Algunos valores del coeficiente depermeabilidad a través de membranas por difusiónpasiva son: O2: 2.3 cm/s, CO2: 0,35 cm/s, H2O: 0,0034cm/s, glicerol: 10 6 cm/s, sodio y potasio: 10 14 cm/s.
La desigual distribución de iones y moléculas entreambos lados de la membrana es la base para la creaciónde los gradientes químicos y eléctricos. La medida deesa diferencia de concentración de cargas es lo que sellama potencial de membrana, que se usa para muchasfunciones celulares, como por ejemplo la síntesis deATP o la transmisión del impulso nervioso. Lasemipermeabilidad de la membrana también permite elfenómeno de la ósmosis, es decir, el flujo de agua haciadonde más concentración de solutos haya. Las célulasvegetales deben su crecimiento a este proceso. Comoveremos más adelante, las moléculas que no cruzan lasmembranas libremente son interesantes para las célulaspuesto que la variación de sus concentraciones a unlado u otro de la membrana puede actuar como señaleso como herramientas. Por ello se han inventadoproteínas integrales de membrana que permitenselectivamente el paso de estas sustancias de un lado alotro. Por ejemplo, la contracción muscular se debe auna rotura de ese gradiente eléctrico.
Fluidez y heterogeneidad
Es la capacidad de una molécula que forma parte deuna membrana para desplazarse por ella. Lasmembranas son fluidas, prácticamente son láminas degrasa, donde las moléculas se encuentran en un estadode líquido viscoso. Esto implica que, en teoría, lasmoléculas podrían difundir y desplazarse por ella sinrestricciones. Consideremos un glicerofosfolípido queestá situado en la membrana plasmática en sumonocapa externa. Tendría dos posibilidades demovimiento: uno lateral donde se desplazaría entre lasmoléculas contiguas, y otro en el que saltaría a lamonocapa interna, movimiento denominado "flipflop".Los dos tipos de movimientos se han demostradoexperimentalmente en membranas artificiales pero elprimero es mucho más frecuente que el segundo. Unamolécula lipídica puede recorrer 30 micras en unos 20segundos por difusión pasiva lateral, es decir, podríadar la vuelta a una célula de tamaño medio enaproximadamente un minuto. Sin embargo, los saltos
Atlas de histología vegetal y animal. Universidad de Vigo.
El cruce de la membrana por parte
de las moléculas depende de su
tamaño y de sus características
eléctricas (Modificado de Alberts et
al., 2002).
La célula. Membrana celular. 13
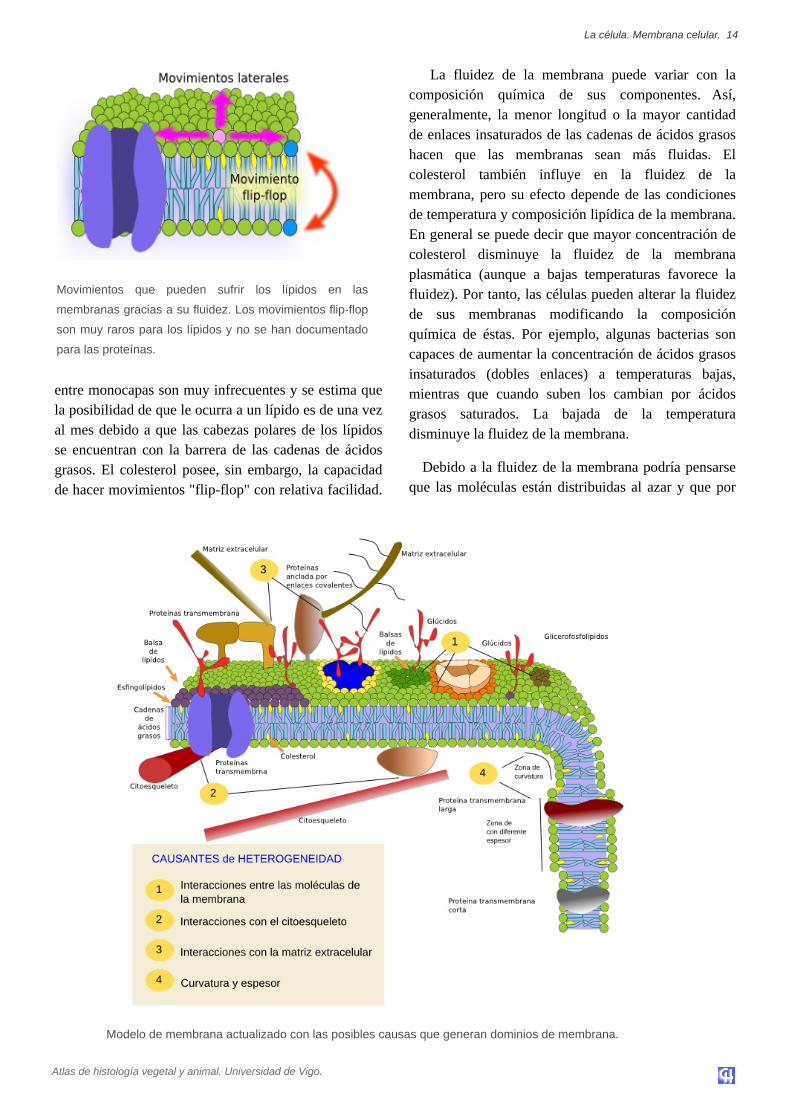
entre monocapas son muy infrecuentes y se estima quela posibilidad de que le ocurra a un lípido es de una vezal mes debido a que las cabezas polares de los lípidosse encuentran con la barrera de las cadenas de ácidosgrasos. El colesterol posee, sin embargo, la capacidadde hacer movimientos "flipflop" con relativa facilidad.
La fluidez de la membrana puede variar con lacomposición química de sus componentes. Así,generalmente, la menor longitud o la mayor cantidadde enlaces insaturados de las cadenas de ácidos grasoshacen que las membranas sean más fluidas. Elcolesterol también influye en la fluidez de lamembrana, pero su efecto depende de las condicionesde temperatura y composición lipídica de la membrana.En general se puede decir que mayor concentración decolesterol disminuye la fluidez de la membranaplasmática (aunque a bajas temperaturas favorece lafluidez). Por tanto, las células pueden alterar la fluidezde sus membranas modificando la composiciónquímica de éstas. Por ejemplo, algunas bacterias soncapaces de aumentar la concentración de ácidos grasosinsaturados (dobles enlaces) a temperaturas bajas,mientras que cuando suben los cambian por ácidosgrasos saturados. La bajada de la temperaturadisminuye la fluidez de la membrana.
Debido a la fluidez de la membrana podría pensarseque las moléculas están distribuidas al azar y que por
Atlas de histología vegetal y animal. Universidad de Vigo.
Movimientos que pueden sufrir los lípidos en las
membranas gracias a su fluidez. Los movimientos flipflop
son muy raros para los lípidos y no se han documentado
para las proteínas.
Modelo de membrana actualizado con las posibles causas que generan dominios de membrana.
La célula. Membrana celular. 14

tanto la membrana sería homogénea en cuando acomposición molecular en cualquier lugar de susuperficie. Esto no es así y hay diversas restricciones alas movilidad lateral de las moléculas que hacen que lamembrana tenga dominios con composicionesmoleculares diferentes. Las resticciones pueden secausadas por diferentes mecanismos como interaccióncon moléculas del citoesqueleto o la matrixextracelular, interacciones moleculares entre laspropias moléculas de la membrana, densidad (menorfluidez local), concentraciónn de cargas eléctricas,grado de curvatura de la membrana o espesor de lamisma.
Una restricción al movimiento de las moléculas enlas membranas de las células se debe a lasinteracciones y asociaciones moleculares entre laspropias moléculas de las membranas. Esto afecta tantoa proteínas como a los lípidos y crea dominios,distribución heterogénea de moléculas en lasmembranas.
Los esfingolípidos y el colesterol se pueden asociarentre sí espontáneamente haciendo que su movilidaddisminuya y por tanto se conviertan en una región
membranosa más densa que el resto, como si de unabalsa en un mar se tratara. Se cree que estasasociaciones, denominadas balsas de lípidos ("lipidrafts"), son muy abundantes y dinámicas y hacen quelas membranas celulares sean en realidad un mosaicode dominios más densos que viajan entre losglicerofosfolípidos, más fluidos. Hay experimentos queapoyan la idea de que ciertas proteínas tendrían mayorapetencia por estas balsas y por tanto viajarían en elinterior de ellas. Este confinamiento de proteínas endominios celulares es importante puesto que permitiríaagrupar o segregar conjuntos de proteínas quefavorecerían o no el inicio de cascadas de señalizaciónintracelulares. Además, se postula que la altaconcentración de ciertos tipos de lípidos en dichasbalsas crea un ambiente químico propicio paradeterminadas reacciones químicas o interaccionesmoleculares. Por ejemplo, se cree que la infección delos linfocitos por parte del virus del SIDA necesita laexistencia de dichas balsas de lípidos. En cualquiercaso tales dominios de esfingolípidos y colesterol sólose han postulado para la monocapa externa de lamembrana plasmática, aunque también se propone suexistencia en las membranas de los orgánulos celularesdonde algunas funciones del propio orgánulo estaríansegregadas en distintos dominios de sus membranas.
Las proteínas integrales o asociadas a la membranatambién pueden interaccionar entre sí y puedenensamblarse en estructuras macromoleculares quepueden favorecer la transmisión de señales,reconocimiento celular, activación enzimática,movimiento celular, etc. También hay proteínasmultiméricas que sólo son activas cuando tienen todassus subunidades asociadas entre sí. Por ejemplo, elreceptor de la insulina está compuesto por cuatrosubunidades. Por supuesto, también hay interaccionesentre lípidos y proteínas que crean dominios demembrana.
Las proteínas integrales de membrana tambiéntienen la posibilidad de una libre difusión lateral. Perose ha comprobado que las proteínas tienen numerosasrestricciones a la movilidad, principalmente por culpade las interacciones de sus dominios intra yextracelulares con moléculas del citoesqueleto y de lamatriz extracelular, respectivamente. Estasinteracciones anclan por tiempos más o menosprolongados las proteínas de membrana a lugares
Atlas de histología vegetal y animal. Universidad de Vigo.
Las interacciones entre lípidos, sobre todo esfingolípidos
y colesterol, pueden crear zonas de mayor densidad
lípidica. También la interacción entre proteínas y ciertos
lípidos pueden crear zonas o conchas de lípidos
característicos alrededor de las proteínas.
La célula. Membrana celular. 15

concretos de la superficie celular. Las células tienenotros mecanismos para confinar proteínas adeterminados dominios celulares. Por ejemplo, en las
células epiteliales del digestivo ciertos transportadoresy enzimas están localizados sólo en la zona apical yotros en la basal gracias al cierre a modo de cinturónque realizan las uniones estrechas, como vimos en elcapítulo dedicado a la matriz extracelular. Tal asimetríaes esencial para el funcionamiento de la célula epitelial.
Curvar una membrana es otra manera de creardominios, y esta curvatura puede ser el proceso inicialde la formación de una vesícula, la extensión de unaexpansión celular, el cambio o crecimiento de unorgánulo, o simplemente un pliegue que actúa comobarrera a las difusión lateral de moléculas.
La maquinaria necesaria para curvar una membranarequiere a su vez un dominio de membrana parallevarlo a cabo. Determinadas composiciones lipídicaso zonas con diferente carga eléctrica son sitios deatracción para la maquinaria. Los fosfoinosítidos sonlípidos que participan en este reclutamiento,particularmente PIP2 y PIP3. Son apropiados para estopuesto que su cabeza puede cambiar en carga y
estructura mediantemodificaciones químicaslocales. Estos lípidos son asu vez mantenidos en elsitio por afinidad con lasproteías reclutadas. Lacreación inicial de undominio para curvar unamembrana ocurre tambiéncon la fostatidil serina, lacual, cuando cambia dehemicapa por acción de lasflipasas, es capaz de ayudara generar curvatura, y esretenida por proteínas comola caveolina. Los filamentosde actina pueden tambiéncontrolar la difusión delípidos, además deproteínas, creando barreras amodo de vallas en superficiecitosólica de la membrana,creando así dominios delípidos sin la intervencióndel colesterol o losesfingolípidos.
Los microdominios de
Atlas de histología vegetal y animal. Universidad de Vigo.
Los movimientos de las moléculas pueden estar
restringidos por las interacciones directas con la matriz
extracelular, con el citoesqueleto, aunque también se
pueden limitar los movimientos por la inclusión en las
balsas de lípidos o por la disposición del citoesqueleto
(imagen de la derecha).
Algunos mecanismos moleculares que curvan las membranas de las células.
La célula. Membrana celular. 16

lípidos atraen a las proteínas que realmente curvan alas membranas de manera efectiva. Hay proteínasespecializadas en crear curvatura de membranas. Eldomimio proteico BAR (Bin/amphyphysin/Rsv161) esuno de ellos. Lo hace de dos maneras: mediante lacreación de un andamiaje curvado sobre el quedescansa la membrana o mediante inserciones desecuencias de aminoácidos en la membrana a modo decuñas. Otros ejemplos son las caveolinas que tambiéngeneran curvatura para formar caveolas, lastetraspaninas que generan túbulos en las membrana, losESCRT que son responsables en los endosomas decrear las vesículas de los cuerpos multivesiculares,etcétera. Por último, los filamentos de actina son unasauténticos curvadores de membranas mediantepolimerización, mediante la cual empuja a lamembrana plasmática hacia afuera creando
expansiones celulares. Muchas de las proteínas capacesde curvar la membrana son también unas activadorasde la polimerización de filamentos de actina.
Otros dominios físicos de las membranas, aunquecreados por moléculas presentes en la propiamembrana como las proteínas transmembrana, sonregiones de diferente espesor o altura. Son formadospor proteínas transmembrana que tienen del dominiohidrofóbico más largo de lo habitual y por tanto seacomodan mejor en la membrana cuando se rodean delípidos con cadenas de ácidos grasos largos. Estasagrupaciones de lípidos y proteínas crean áreas demayor grosor que excluyen a otras proteínas consecuencias de aminoácidos hidrofóbicos más cortas o alípidos con cadenas de ácidos grasos con menosátomos de carbono.
Dominio de membrana creado por diferencias de espesor.
La célula. Membrana celular. 17
Atlas de histología vegetal y animal. Universidad de Vigo.

Las membranas celulares están formadas por unabicapa lipídica con dos hemicapas. En las membranasde los orgánulos y en la plasmática existe unahemicapa orientada hacia el citosol y otra orientadahacia el interior del orgánulo o al exterior celular,respectivamente. La composición en lípidos, glúcidos yproteínas periféricas es distinta en ambas hemicapas.Además, las proteínas transmembrana tienen unaorientación precisa. Esta desigual distribución demoléculas entre ambas hemicapas se denominaasimetría de membrana, y se conocía incluso antes deformularse el modelo de mosaico fluido de lamembrana en 1972.
La generación y mantenimiento de esta asimetría esesencial para la célula. En la membrana plasmática, lahemicapa orientada hacia el exterior contiene unamayoría de los lípidos que poseen colina, como lafosfatidilcolina y la esfingomielina, mientras que lafosfatidiletanolamina, fosfatidilinositol y lafosfatidilserina se localizan en la hemicapa interna.Esto es interesante porque crean una distribucióndiferente de cargas entre ambas superficies de lamembrana, que contribuye al potencial de membrana.Se ha visto que en ausencia de iones la membranaplasmática es capaz de producir un potencial demembrana por sí misma debido a la mayorconcentración de cargas negativas de la monocapainterna. Además, esta asimetría facilita la asociaciónespecífica de proteínas que necesitan un ambienteeléctrico determinado y que es aportado por lanaturaleza química de las cabezas de los lípidos. Otroejemplo es el lípido fosfatidil inositol, localizado en lahemicapa interna, que al ser modificado por ciertasfosfolipasas se divide en dos moléculas, una de lascuales viaja por el citosol y actúa como segundomensajero. También son importantes las propiedadesfísicas que tal asimetría aporta a las membranas y, porejmplo, parece que una determinada composiciónlipídica de la hemicapa citosólica facilita la formaciónde vesículas hacia el citosol, es decir, se curva másfácilmente hacia el citosol.
La rotura de esta asimetría lipídica funciona a vecescomo una señal de que hay alteraciones en la célula.Por ejemplo, las células que sufren apoptosis, muertecelular programada, exponen rápidamente la
fosfatidilserina en la monocapa externa y esto es unaseñal para que los macrófagos eliminen esa célula. Dehecho, en la membrana plasmática de las células sanashay proteínas "patrullando" en la membrana plasmáticaque cuando detectan una fosfatidilserina en lamonocapa externa la devuelven rápidamente a lainterna. También es importante la rotura de la asimetríade los lípidos en los eritrocitos para el inicio de lacoagulación sanguínea. Incluso algunos virus conmembrana exponen en su monocapa externafosfatidilserina y fostatidiletanolamina para serincorporados a las células con más facilidad pormacropinocitosis o fagocitosis.
Aunque cuando se habla de asimetría estamosrefiriéndonos sobre todo a los lípidos, también hay unadistribución u organización desigual de los glúcidos yde las proteínas entre las dos monocapas de lasmembranas celulares. Los glúcidos se localizanpreferentemente en la hemicapa externa de lamembrana plasmática formando el glicocálix, y en lano citosólica de los lisosomas y endosomas, comoveremos más adelante. Esto hace que actúen comocentros de reconocimiento y protección para lascélulas. Las proteínas también tienen una orientaciónprecisa en la membrana, con un dominio citosólico yotro extracelular o en el interior de los orgánulos. Estoes claro cuando obervamos a los receptores de lamembrana plasmática, los cuales tienen que tener sucentro de reconocimiento orientado hacia el ladoextracelular.
¿Dónde y cómo se produce la asimetría?
Lípidos. La distribución asimétrica de los lípidos seproduce principalmente en el aparato de Golgi y enotros compartimentos celulares Curiosamente, en elretículo endoplasmático, donde mayoritariamente sesintetizan los lípidos, hay una distribución muyparecida entre las dos hemicapas. Para los lípidos concabeza polar grande es difícil cruzar de una hemicapa ala otra (movimiento del tipo “flipflop”) por la barreraque supone el ambiente hidrófobo que generan lascadenas de ácidos grasos. Sin embargo, para otroslípidos con zona polar poco voluminosa, como elcolesterol, diacilglicerol, ceramida o ácidos grasosprotonados, el cambio entre monocapas es muy
La célula. Membrana celular. 18
Atlas de histología vegetal y animal. Universidad de Vigo.

frecuente. Los glicerofosfolípidos de cabezas polaresgrandes pueden salvar la barrera hidrófoba de losácidos grasos mediante unos transportadoresespecíficos, o traslocasas, localizados en lasmembranas. Hay tres tipos: flipasas, flopasas ymezcladores ("scramblases"). Estas proteínas seencargan de transportar glicerofosfolípidos entre lasdos hemicapas y generar asimetría. Las flipasastransportan lípidos hacia la hemicapa interna, lasflopasas hacia la hemicapa externa y las mezcladorasen ambas direcciones. Las mezcladoras no necesitanATP para llevara cabo su función. Esta desigualdistribución de los lípidos se mantiene por la propiadificultad de los movimientos “flipflop”. Por ejemplo,lo esfingolípidos, que no son translocados, permanecenen la monocapa donde se sintetizan: la interna delaparato de Golgi, que se será posteriormente la externade la membrana plasmática. Más del 80 % de losesfingolípidos de la membrana plasmática se localizanen la monocapa externa.
Estas proteinas se distribuyen desigualmente por lasdiferentes membranas de la célula para aportar a cadamembrana sus asimetría lipídica característica. Inclusodentro de una misma familia puede haber subtipos condiferente regulación. Como se mencionó antes, laapoptosis provoca la rotura de la asimetría de lamembrana plasmatica que desencadena la fagocitosisde la célula. Esta rotura se debe a una activaciónpuntual de una proteína mezcladora que se encuentra
en dicha membrana. Sin embargo, otra proteínamezcladora que actúa en el retículoendoplasmático está funcionando siempre, y poreso las dos hemicapas de este orgánulo son muyparecidas. Otro ejemplo, en la membrana deleritrocito hay proteínas mezcladoras que seactivan con la entrada masiva de calcio yfavorecen la coagulación sanguínea.
Glúcidos. La distribución de los glúcidos,localizados sobre todo en la hemicapa externade la membrana plasmática, se produce durantesu síntesis, primero en el retículoendoplasmático y posteriormente en el aparatode Golgi.
Proteínas. La asimetría de las proteínas seproduce durante su síntesis en el retículoendoplasmático, aunque las proteínas asociadasa la cara citosólica se sintetizan en el citosol.
La distribución y organización de las moléculas en las dos
hemicapas de las membranas puede ser diferente. Esto es
claro para los lípidos en la membrana plasmática (arriba),
pero no tanto en la del retículo endoplasmático (abajo). Sin
embargo, las proteínas se orientan y disponen de manera
asimétrica tanto en las membranas del retículo como en la
plasmática. Se indica la carga negativa de la fosfatidil
serina para mostrar la diferente distribución de cargas entre
ambas monocas de la membrana plasmática, pero no corre
lo mismo en la membrana del retículo endoplasmático.
Proteínas encargadas de redistribuir los diferentes lípidos entre las
dos monocapas de las membranas. La localización de estas
proteínas varía entre orgánulos y entre tipos celulares. Flipasas,
flopasas son en realizadad dos familias de proteínas cuyos
miembros se distribuyen diferencialmente. (Modificado de Quazi y
Molday, 2011).
La célula. Membrana celular. 19
Atlas de histología vegetal y animal. Universidad de Vigo.

Rotura y fusión de membranas
Una de las propiedades de las membranas más útilespara la célula es la capacidad de romperse y volver aser fusionadas. Ello permite que los compartimentosintracelulares puedan ser tremendamente plásticos, esdecir, crecer, dividirse, fusionarse, liberar fragmentosen forma de vesículas membranosas en uncompartimento que viajan a otro con el que sefusionan, etcétera. Ésta es la base del transportevesicular que veremos en los apartados siguientes. Estacaracterística de las membranas es también necesariadurante la etapa de la mitosis denominada citocinesisdonde la membrana citoplasmática debe crecer ensuperficie, romperse y luego fusionarse para formardos células hijas independientes. Estos procesos derotura y de fusión de membranas están gobernadosprinciplamente por las proteínas, entre las que sedestacan las SNARE (soluble Nethylmaleimidesensitive factor attachment receptor), pero también porlas propiedades fisicoquímicas de los lípidos quecomponen las membranas.
Reparación de membranas
Numerosos procesos naturales o la manipulaciónexperimental de las células provocan la rotura de lasmembranas celulares. Por ejemplo, en los tejidos vivossometidos a tensiones hay un proceso de rotura de lamembrana plasmática, como ocurre frecuentemente enlas células musculares. Pero también en losexperimentos de clonación se necesita meter unapipeta, la captación de vectores o ADN supone a vecesla poración de las membranas celulares, la propiamanipulación supone roturas de membrana. La roturade la membrana plasmática es letal para la célula si seprolonga más de unos cuantos segundos. Las célulascuentan con mecanismos para reparar estos daños ymantener así las diferencias entre el medio interno yexterno. Los tejidos que no son capaces de reemplazara sus células como es el caso del sistema nervioso,estos mecanismos de reparación son especialmenteinteresantes.
Una rotura de la membrana plasmática rápidamentedispara la entrada masiva de calcio en la célula, lo queproduce alteraciones celulares que, si se mantienedurante mucho tiempo, desencadenan la apoptosis(muerte celular programada). Per este incremento decalcio dispara también los mecanismos de reparación.El calcio dispara numerosas vías enzimáticas en
paralelo que confluyen en la generación de vesículas yfusión de estas para sellar la membrana. Curiosamenteni el cloro, ni el sodio, ni el potasio paracen participaren los mecanimos de reparación de las membranas.
Hay dos maneras de sellar la membrana según eltipo de daño que se produzca. Cuando los daños sonpequeños (normalmente menores a 0.2 µm) laspropiedades de los lípidos de la membrana sonsuficientes para repararlos. Ello es debido a que loslípidos en el borde de la membrana adoptan unadisposición inestable que fuerza a dichos bordes aencontrarse y a sellarse. La rapidez con que esteproceso ocurre depende de la tensión de la membrana,que depende a su vez de los puntos de anclaje, bien alcitoesqueleto o a la matriz extracelular. Cuando seproduce una rotura entra calcio a favor de gradiente deconcentración, lo que hace que el citoesqueleto sedesorganice parcialmente en la zona dañada y su efectosobre la membrana disminuye, se rebaja así la tensión yaumenta la velocidad de resellado. Las proteínasESCRT, que participan en los procesos membranososde formación de vesículas internas en los cuerposmultivesiculares, también participan en el sellado depequeñas roturas de membrana (<100 nm de anchura).
Cuando los daños son grandes (más de 0.20.5 µm)los bordes rotos libres de la membrana están demasiadolejos para se que puedan autosellar y se pone enfuncionamiento un mecanismo de exocitosis masiva, esdecir, fusión de vesículas con la membrana plasmática,aunque en este caso también incluye grandes
Cuando las roturas de la membrana son pequeñas,
menores a unas 0.2 micras, las propiedades moleculares
de los lípidos son suficientes para cerrar el hueco
(Modificado de McNeil y Steinhardt, 2003)
La célula. Membrana celular. 20
Atlas de histología vegetal y animal. Universidad de Vigo.

compartimentos membranosos. El proceso sería elsiguiente: la amplia rotura produce una gran entrada decalcio, éste provoca la formación y fusión decompartimentos membranosos próximos al lugar de larotura, las cuales terminarán por fusionarse con losbordes de la membrana plasmática. Entre loscompartimentos implicados en la fusión estarían losendosomas, los lisosomas, vesículas próximas y otroscompartimentos especializados de distintos tiposcelulares. Los lisosomas parecen especialmenteimportantes en este proceso. La endocitosis en lapropia célula y la desorganización de las cisternas delreiculo ayudarían también a crear compartimentos quese fusionarían con la zona de rotura. El calcio tambiénactiva a enzimas protesas que favorecen el proceso desellado, probablemente porque eliminan elcitoesqueleto, lo que favorece el monvimiento de loscompartimentos membranosos en la zona de rotura. Seha propuesto que el mecanismo de fusión demembranas desarrollado por los eucariotas fue enrealidad inventado para reparar las roturas que seproducían en las membranas de las primeras células.Estos mecanismos se usaron después para el tráficointracelular.
El mecanismo propuesto según el cual se forma ungran compartimento por la fusión de vesículas internas,y es éste el que se fusiona con la zona rota, propuestopor McNeil, no tiene soporte experimental. No se haobservado en microscopía electrónica y las medicioneseléctricas sugieren que el proceso de sellado es unmecanismo progresivo. Además, el sellado es másrápido en aquellas células con muchoscompartimentos.
Las células y los tejidos tienen mecanismos paraadaptarse a las tensiones mecánicas repetitivas: lamatriz extracelular se especializa, aumentan loscomplejos de unión, aumentan los filamentosintermedios del citoesqueleto, aumenta las dimensionesy el número de compartimentos membranosos
celulares, etcétera. En los cultivos celulares se puedenestudiar las respuestas de las células a las tensionesmecánicas. Se ha comprobado que ante un estiramientodel 1015% las células aumentan su superficie demembrana por fusión de compartimentos internos. Estoocurre normalmente en las células de la vejigaurinaria,que sufren grandes variaciones de tensión.Cuando las células en cultivo son estiradas dos veces,en la segunda se produce una reparación más rápidaque en la primera. Se observa que la cantidad devesículas producidas por el aparato de Golgi es mayorde lo normal, por lo que la célula puede responder conmayor eficacia. La sujeción de la células a la matrizextracelular también se ve reforzada. Así, las proteínasdel citoesqueleto y de la matriz extracelular seincrementan en número para adaptarse a las tensionesmecánicas repetitivas.
Cuando las roturas de la membrana son grandes, más de
0.20.5 micras, ocurre una gran entrada de calcio que
dispara procesos similares a los de la exocitosis. Los
orgánulos próximos son conducidos a la zona del daño, se
fusionan con la membrana plasmática.
La célula. Membrana celular. 21
Atlas de histología vegetal y animal. Universidad de Vigo.

El origen de las moléculas que forman parte de lasmembranas lo trataremos con más detalle cuandohablemos de los orgánulos que las producen. Así, lasíntesis de lípidos se estudiará cuando hablemos delretículo endoplasmático y aparato de Golgi, la de lasproteínas cuando nos centremos en el retículoendoplasmático rugoso y los glúcidos cuandoabordemos el aparato de Golgi. Asimismo, en elapartado dedicado a cada orgánulo membranoso de lacélula estudiaremos cómo y de dónde proceden lasmoléculas que lo forman. Aquí haremos una síntesis.
Las membranas están en perpetua renovación, susmoléculas son degradadas y sintetizadas de nuevocontinuamente. Mediante marcaje de aminoácidos seha comprobado que las proteínas de alto pesomolecular de la membrana plasmática se renuevan cada25 días, mientras que las de bajo peso molecular lohacen cada 713 días. Los lípidos lo hacen cada 35días. La mayoría de los componentes de las membranascelulares se sintetizan en el retículo endoplasmático.Esto es cierto para las proteínas y para la mayoría delos lípidos, mientras que algunos lípidos y la mayorparte de los glúcidos son sintetizados en el aparato deGolgi. El transporte de estas moléculas desde elretículo endoplasmático o desde el aparato de Golgihasta su destino final sigue generalmente las rutas deltráfico vesicular, que son aquellas que comunican loscompartimentos celulares membranosos mediantevesículas. Sin embargo, los lípidos también se puedentransportar mediante proteínas transportadoras ocontactos directos entre membranas.
El retículo fabrica proteínas y lípidos para sí mismo,para la envuelta nuclear, que en realidad es unacontinuación de las membranas del propio retículo,para el aparato de Golgi, para los endosomas, para loslisosomas, para la membrana plasmática y para laspropias vesículas que sirven de transportadores. Elretículo también fabrica lípidos para otros orgánulosque quedan fuera de la ruta vesicular como son lasmitocondrias y los cloroplastos, los cuales reciben loslípidos gracias a proteínas transportadoras o porcontactos directos de sus membranas. Algunos de loslípidos de los compartimentos que participan en el
tráfico vesicular, además de en vesículas, también setransportan de esta forma. Las proteínas de lasmembranas de las mitocondrias y de los cloroplastos sesintetizan en ribosomas libres en el citosol o sonproducidas por el propio orgánulo, puesto que estos dosorgánulos contienen ADN, ribosomas y toda lamaquinaria para la síntesis proteica.
Merece mencionar como caso aparte a losperoxisomas. Recientemente se ha postulado que sonderivados de las membranas del retículoendoplasmático, de las que se originarían porevaginación. Sin embargo, es un orgánulo que norecibe ni envía vesículas a otros orgánulos. Además, seha demostrado que proteínas producidas por losribosomas citosólicos forman parte de las proteínasinternas de los peroxisomas. Luego las fuentes de susmoléculas de membrana son variadas.
En el caso de la membrana plasmática la principalfuente de moléculas son las vesículas que se fusionancon ella. Los compartimentos que producen dichasvesículas son principalmente los endosomas y elaparato de Golgi. Pero en última instancia las proteínasy los lípidos que conforman las vesículas sonsintetizados en el retículo endoplasmático y aparato deGolgi. Por ejemplo, el glucocálix desarrollado presenteen algunas células se ensambla principalmente en elaparato de Golgi. La renovación de las moléculas delas membrana plasmática se completa con la formaciónde vesículas en la propia membrana que viajarán enuna serie de pasos hasta los lisosomas donde serándegradadas. A veces las moléculas que se retiran de lamembrana por parte de las vesículas son devueltas aella, de nuevo en forma de vesículas, gracias a unproceso de reciclaje que tiene a los endosomas comocompartimento de relevo. Esto hace que la membranaplasmática pueda variar la cantidad y proporción de susmoléculas sin la necesidad de un costoso proceso dedegradación y síntesis.
Por último, las membranas de los endosomas y de loslisosomas, y de las vacuolas de las plantas provienenen gran parte de vesículas de endocitosis, pero tambiénde vesículas enviadas por el Golgi.
La célula. Membrana celular. 22
Atlas de histología vegetal y animal. Universidad de Vigo.

Las membranas suponen una barrera a la libredifusión de iones y moléculas cargadas eléctricamente.Sin embargo, muchas de esas moléculas son esencialespara la célula. Pensemos, por ejemplo, en la glucosa oen los iones que crean gradientes electroquímicos.Conocer el transporte de membrana no sólo esimportante para saber cómo funciona una célula sinopor ejemplo para sintetizar fármacos que lleguen a susdianas en el interior de las células. Como dijimos enapartados anteriores la creación de gradientes entreambos lados de la membrana es necesaria puesto quese usan en muchos aspectos de la fisiología celular.Aproximadamente el 10 % de los genes de una célulaestán relacionadas con transportadores de membrana,lo que nos da una idea de la importancia de estemecanismo para el célula. Pero para que estosgradientes sean útiles es necesario que la célula puedacrearlos, regularlos y romperlos cuando lo necesite. Enla membrana existen unas proteínas especializadastanto en el transporte de moléculas necesarias para elmetabolismo como en la creación y modificación delos gradientes electroquímicos. Son proteínastransmembrana que se agrupan en tres tipos: bombas,transportadores y canales.
Las bombas son proteínas transmembrana quetransportan iones o moléculas de un lado al otro de lamembrana en contra de sus gradientes deconcentración, con gasto de energía. La energía lapueden obtener de diferentes fuentes: de la luz, dereacciones de óxidoreducción o de la hidrólisis delATP, que es lo más frecuente. Las bombas soncreadoras de gradientes puesto que transforman energíaquímica o electromagnética (luz) en gradienteelectroquímico que luego será usado para otrasnecesidades celulares, como veremos más adelante. Noexiste una gran diversidad molecular de bombas y sepueden clasificar según la fuente de energía. a) Usan laluz como fuente energética. Por ejemplo, labacteriorodosina utiliza la luz para crear un gradientede protones en las membranas de algunos procariotas.b) Utilizan potenciales de óxidoreducción para creargradientes. Los complejos de las cadenas de transportede electrones de las mitocondrias aprovechan cambiosde óxidoreducción para mover protones desde la
matriz al espacio intermembranoso, entre las dosmembranas. c) Usan ATP como fuente de energía.Dentro de este grupo hay varios tipos. Las hay queintroducen protones en los orgánulos, como laspresentes en las membranas de los lisosomas queproducen pH ácidos para permitir la degradación demoléculas. A este grupo también pertenecen lasATPasas de las mitocondrias y de los cloroplastos querealizan el proceso contrario, utilizan un gradiente deprotones para sintetizar ATP. Aunque también puedenconsumir ATP para producir un gradiente de protones.Otro tipo de bombas que utilizan ATP transportan ionesde un lado a otro de la membrana. Una de las másimportantes es la ATPasa de Na+/K+, responsable de lacreación de los gradientes iónicos de las membranasplasmáticas y que permite la excitabilidad de lasneuronas, de las células musculares, la absorción de losalimentos por las células del aparato digestivo, etcétera.Nos podemos hacer una idea de la importancia de estasbombas Na+/K+ en las células animales sabiendo queconsumen hasta el 25 % del total del ATP celular. Aeste grupo pertenecen también las bombas quetransportan cationes como el calcio, por ejemplo la quesaca calcio del retículo endoplasmático de las célulasmusculares en cada contracción muscular. Por últimotenemos los bombas denominadas ABC que usan ATPpara mover una gran variedad de moléculas entre
Ejemplos de proteínas que permiten el transporte de iones
con gasto de energía, denominadas bombas. El primer
ejemplo (más a la izquierda) es un complejo de la cadena
respiratoria de las mitocondrias. A continuación una
bacteriorodopsina, que usa la luz visible para mover
protones a través de la membrana y por último , una bomba
que intercambia sodio y potasio, ayudando a establecer los
gradientes de estos iones en la membrana plasmática.
(Modificado de Alberts et al., 2002).
La célula. Membrana celular. 23
Atlas de histología vegetal y animal. Universidad de Vigo.

ambos lados de la membrana. Aparecen en todas lascélulas conocidas y son capaces de transportar desdeiones, hasta monosacáridos, aminoácidos, poliscáridosy polipéptidos.
Los transportadores son proteínas integrales queusan gradientes electroquímicos para mover moléculasentre ambos lados de la membrana. Este tipo demovimiento se denomina difusión facilitada: difusiónporque es un movimiento pasivo generado por elgradiente electroquímico existente y facilitada puestoque es el transportador el que la permite. Lostransportadores son muy numerosos, más de 100familias, y aparecen en todas las membranas de lacélula. El mecanismo de transporte supone unreconocimiento de la molécula o moléculas a las quevan a transportar y un cambio conformacional deltransportador que posibilita el trasiego de las moléculasentre ambos lados de la membrana. El transporte puedeser de distintos tipos. El transporte uniporte suponemover una molécula a favor de su gradiente deconcentración. El cotransporte permite la translaciónsimultánea de dos moléculas entre ambos lados de lamembrana. Si el sentido en el que viaja una moléculases contrario al de la otra se denomina antiporte y si lasdos moléculas viajan en el mismo sentido se denominasimporte. En los movimientos de cotransporte uno delas moléculas suele viajar a favor de gradiente deconcentración y utiliza esa fuerza para mover a la otramolécula que viaja en contra de su gradiente de
concentración. Por ejemplo, las células cardiacasutilizan el transportador Na+/Ca2+ para sacar calcio encontra de gradiente desde el citosol hacia el exteriorcelular aprovechando el gradiente de Na+, que siemprees mayor fuera de la célula que en el citosol. Lostransportadores que realizan antiporte suelenintercambiar elementos parecidos: catión por catión,anión por anión, azúcar por azúcar, etcétera. Sinembargo, en el simporte se pueden transportarmoléculas diferentes. Por ejemplo, en las célulasintestinales se emplea el gradiente de Na+ paraincorporar Dglucosa.
Los canales son proteínas integrales que crean poroso conductos hidrofílicos que comunican ambos ladosde la membrana. Tienen la propiedad de poder abrir ocerrar dicho conducto según ciertas condiciones. Suprincipal función es regular los gradientes iónicos entreambos lados de la membrana, por tanto alterar elpotencial eléctroquímico de ésta, hecho que setransformará en información para la célula. También
Esquema de los distintos tipos de transporte llevado a cabo
por los transportadores. Pueden ser uniporte si transportan
una sólas molécula a favor de gradiente. También hay
transportadores que cotransportan dos moléculas distintas
aprovechando el gradiente de concentración de una de
ellas. Si las dos viajan en un mismo sentido tenemos un
cotransporte simporte y si lo hacen en sentido contrario
antiporte (Modificado de Alberts et al., 2002).
Los canales iónicos son conductos hidrofílicos que permiten
el paso de iones a favor de gradiente de concentración.
Pueden ser canales dependientes de voltaje cuando su
apertura o cierre están regulados por el potencial de la
membrana o dependientes de ligando cuando es el
reconocimiento de una molécula, por ejemplo un
neurotransmisor, lo que provoca su apertura. (Modificado
de Alberts et al., 2002).
La célula. Membrana celular. 24
Atlas de histología vegetal y animal. Universidad de Vigo.

son necesarios para la secreción o absorción desustancias como ocurre en el riñón. En cualquier casoes siempre un transporte pasivo puesto que los ionessiempre viajan a favor de gradiente de concentración yla selección de los iones por los distintos tipos decanales depende del diámetro del canal hidrofílico. Hayuna gran diversidad de canales: canales de sodio, depotásio, de calcio, de cloro, incluso de agua(acuaporinas), etcétera. Como hemos dicho, la aperturao cierre del canal puede modularse. Si cambia con la
variación del potencial electroquímico de la membranase denominan canales dependientes de voltaje.También pueden modularse por la unión de ligandos opor modificaciones covalentes, por ejemplo porfosforilación. Realizan funciones trascendentales parael organismo como la excitabilidad neuronal,contracción muscular, prevención de la poliespermiatemprana (evitar que más de un espermatozoide se unaa un ovocito), etcétera.
La célula. Membrana celular. 25
Atlas de histología vegetal y animal. Universidad de Vigo.

Las células de los organismos pluricelulares seorganizan en tejidos y órganos. Esta disposicióndepende en gran medida de su capacidad para adherirsebien a la matriz extracelular o a otras células. Laadhesión no sólo sirve para anclar y situar a las célulaspara formar andamiajes tridimensionales, sino tambiénpara comunicarse entre sí. Es decir, el grado deadhesión y a quién se adhieren las células es un tipo deinformación útil para la célula. La adhesión se realizapor medio de las denominadas proteínas de adhesión,las cuales se encuentran ancladas a la membranaplasmática. Hay dos tipos, aquellas que anclan la célulaa la matriz extracelular y aquellas que stablecenuniones directas entre dos células contiguas.
Adhesión de la célula de la matriz extracelular
Las integrinas son las moléculas más importantes enla adhesión de la célula a la matriz extracelular. Sonuna gran familia de proteínas presentes enprácticamente todos los animales. Son proteínastransmembrana formadas por dos subunidades (alfa ybeta). En mamíferos hay 18 unidades alfa y 3 unidadesbeta que por combinación pueden formar hasta 24integrinas diferentes, las cuales se expresan según eltejido o estado fisiológico de la célula. Cada una tiene3 dominios moleculares: un dominio intracelular quecontacta con los filamentos de actina del citoesqueleto,otro extracelular globular que es capaz de unirse alcolágeno, fibronectinas y lamininas, y un dominiotransmembrana entre las cadenas de ácidos grasos de lamembrana. Su capacidad de unirse a moléculas de lamatriz extracelular y al citoesqueleto permite unacontinuidad estructural mecánica entre el interior yexterior de la célula. Pero además permite modificar elcomportamiento celular en función de las moléculaspresentes en la matriz extracelular. Esto es posibleporque el estado de adhesión de la integrina setransmite a su dominio citosólico, el cual interactúacon proteínas que son capaces de viajar al interior delcitoplasma para afectar a rutas moleculares o viajar alinterior del núcleo para alterar la expresión génica.También la célula puede modificar su capacidad deadhesión, y por tanto su movilidad, cambiando el juegode proteínas de adhesión en su superficie.
Las integrinas suelen aparecer asociadas en lamembrana plasmática formando las denominadasadhesiones focales y también formar agregadosmayores como son los hemidesmosomas. La fuerza deunión de una célula a la matriz extracelular dependepues de la cantidad, tipo y estado de las integrinas quepresenta en su membrana plasmática.
Adhesión entre células
Estas moléculas se encargan de adherir directamenteunas células a otras. Hay cuatro tipos: cadherinas,inmunoglobulinas, selectinas y algunos tipos deintegrinas. Las cadherinas se encuentran en lasuperficie de la mayoría de las células animales yforman uniones homotípicas, es decir, reconocen aotras cadherinas en la célula adyacente. Son una gransuperfamilia de proteínas cuyos miembros suelenaparecer característicamente en ciertos tejidos. Así, laNcadherina se expresa en el tejido nervioso, la Ecadherina en el tejido epitelial, etcétera. Es por ello quejuegan un papel importante en la segregación depoblaciones celulares de los distintos tejidos. Sonespecialmente importantes durante el desarrolloembrionario. Las cadherinas son parte estructural delos desmosomas (macula adherens). Las moléculas deadhesión del tipo inmunoglobulina, también llamadasCAM (cell adhesion molecule) forman unioneshomofílicas con inmunoglobulinas presentes en lacélula adyacente, aunque también pueden realizar
Integrinas en diferentes estado de activación (Modificado
de Luo et al., 2007).
La célula. Membrana celular. 26
Atlas de histología vegetal y animal. Universidad de Vigo.

uniones heterofílicas con otro tipo de moléculas. Estambién una gran familia de proteínas con distribuciónespecífica de sus miembros en los distintos tejidos. Porejemplo, NCAM aparece en el sistema nervioso. Susuniones no son tan fuertes como las de las cadherinas yparece que actúan ajustando de forma más fina laasociación entre células de un mismo tejido. Lasselectinas son también proteínas de adhesión entrecélulas, pero forman uniones heterofílicas, es decir, seunen a moléculas de diferente tipo de la otra célula, enconcreto a glúcidos presentes en la célula vecina. Estoes gracias a que poseen un dominio que tiene apetenciapor determinados azúcares (ácido siálico y fucosa). Sonimportantes en la unión de los glóbulos blancos a lasparedes del endotelio cuando abandonan el torrentesanguíneo para adentrarse en los tejidos. Lasintegrinas, que antes vimos como moléculas quemedian la adhesión de las células a la matrizextracelular, también pueden mediar adhesiones célulacélula. En concreto, algunas integrinas pueden formar
uniones con algunas moléculas transmembrana del tipode las inmunoglobulinas.
Integrinas en diferentes estado de activación (Modificado
de Luo et al., 2007).
La célula. Membrana celular. 27
Atlas de histología vegetal y animal. Universidad de Vigo.

Como hemos visto en el apartado anterior las célulasse anclan a la matriz extracelular y a otras célulasmediante unas proteínas especializadas. Las integrinas,cadherinas, selectinas e inmunoglobulinas son las másimportantes. A veces se producen uniones tanespecializadas y desarrolladas que forman estructurasmacromoleculares denominadas complejos de unión yuniones focales, las cuales son fundamentales paramantener la cohesión de muchos tejidos,principalmente los epitelios, el tejido muscular y elnervioso.
Los complejos de unión se clasifican según su forma,las moléculas de adhesión que los componen, loselementos a los que se unen y sus interacciones con elcitoesqueleto. La primera vez que se observaron fuecon el microscopio electrónico y se clasificaronmorfológicamente, pero fueron las técnicas de biologíamolecular las que permitieron desentrañar susestructuras moleculares.
Las uniones estrechas o zonula occludens seencuentran en diferentes tipos celulares, como en laspartes apicales de los epitelios, en los endotelios delsistema nervioso, o en el tejido muscular cardiaco.Establecen uniones tan fuertes y estrechas entre lascélulas contiguas que prácticamente no dejan espaciointercelular entre sus membranas plasmáticas,limitando la difusión de sustancia solublesextracelulares.
En el caso de las células epiteliales forman unaespecie de cinturón que rodea todo el perímetro celular.Además de mantener cohesionadas fuertemente a lascélulas realizan otras funciones. En los epitelios, porejemplo en el epitelio digestivo, impiden la difusiónintercelular evitando que las sustancias del interior deltubo digestivo penetren en el organismo por losespacios intercelulares. Esto obliga a las sustancias aser captadas selectivamente por parte de las célulasepiteliales, donde son transformadas y liberadas altorrente sanguíneo. Pero, además, las uniones estrechaspermiten la polaridad de las células epiteliales puestoque impiden la difusión lateral de moléculas insertas ensus membranas celulares. Es decir, actúan como unabarrera física a la difusión lateral de las moléculas de la
membrana plasmática. Con ello se consigue una zona odominio apical con un juego de moléculas distinto alque hay en el domino laterobasal de la célula epitelial.Esta separación es importante para establecer uncamino de captación y liberación de sustancias desde elexterior hacia el interior.
En los capilares del sistema nervioso central lascélulas endoteliales están unidas por uniones estrechasque contribuyen a establecer la barrerahematoencefálica, la cual es un filtro importante paralas moléculas que tienen intercambiarse entre la sangrey las neuronas y glía.
Las uniones estrechas están formadas por la ocludinay por una familia de moléculas denominadas claudinas,que son las proteínas transmembrana encargadas deestablecer los contactos célulacélula. Las claudinas
parecen ser las más importantes en el establecimientode la unión y en estas uniones forman unos poros quedejan pasar ciertos iones por el espacio extracelular, nomás de 1 nanometro de diámetro. Hay 20 tipos declaudinas, cada una de las cuales forma uno poroextracelular distinto y así los epitelios puedenmodificar la selectividad de su permeabilidadintercelular según el tipo de claudina que expresen. Eldominio intracelular de estas moléculas interactúa conotras moléculas denominadas ZO, las cuales forman un
Esquema de las uniones estrechas de las células epiteliales
del digestivo. La estructura molecular parece ser similar en
los distintos tipos de epitelios. (Modificado de Niessen
2007).
La célula. Membrana celular. 28
Atlas de histología vegetal y animal. Universidad de Vigo.
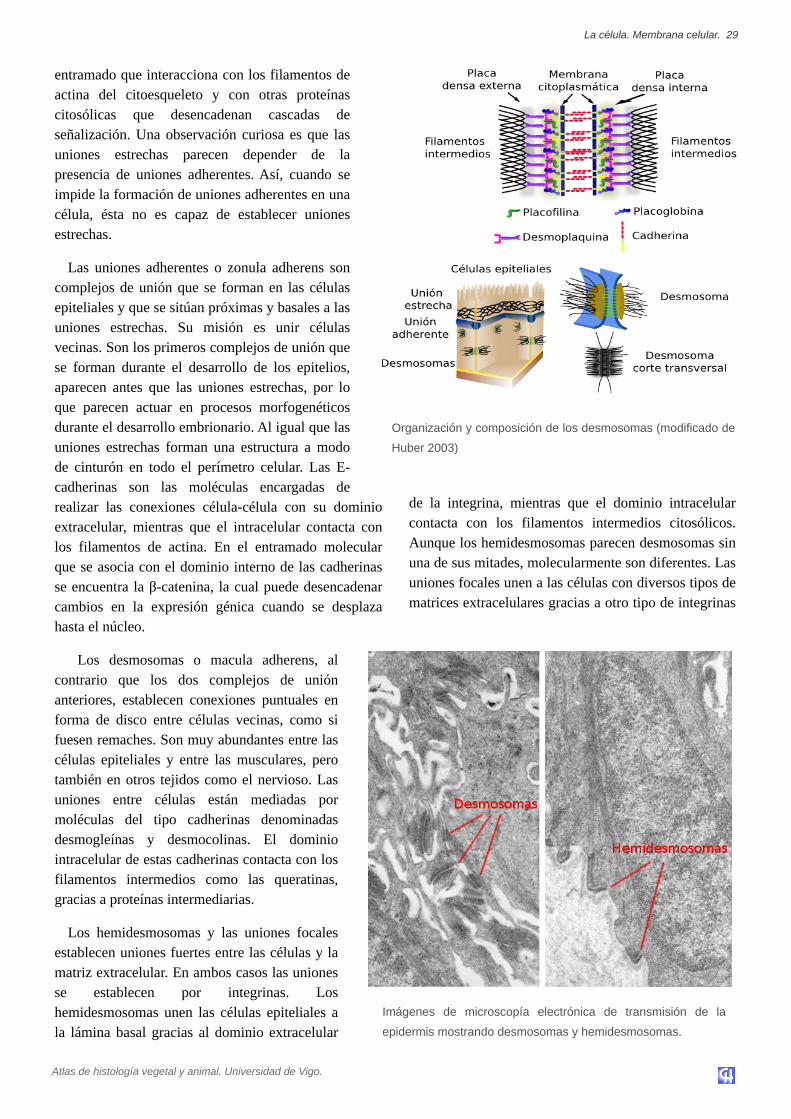
entramado que interacciona con los filamentos deactina del citoesqueleto y con otras proteínascitosólicas que desencadenan cascadas deseñalización. Una observación curiosa es que lasuniones estrechas parecen depender de lapresencia de uniones adherentes. Así, cuando seimpide la formación de uniones adherentes en unacélula, ésta no es capaz de establecer unionesestrechas.
Las uniones adherentes o zonula adherens soncomplejos de unión que se forman en las célulasepiteliales y que se sitúan próximas y basales a lasuniones estrechas. Su misión es unir célulasvecinas. Son los primeros complejos de unión quese forman durante el desarrollo de los epitelios,aparecen antes que las uniones estrechas, por loque parecen actuar en procesos morfogenéticosdurante el desarrollo embrionario. Al igual que lasuniones estrechas forman una estructura a modode cinturón en todo el perímetro celular. Las Ecadherinas son las moléculas encargadas derealizar las conexiones célulacélula con su dominioextracelular, mientras que el intracelular contacta conlos filamentos de actina. En el entramado molecularque se asocia con el dominio interno de las cadherinasse encuentra la βcatenina, la cual puede desencadenarcambios en la expresión génica cuando se desplazahasta el núcleo.
Los desmosomas o macula adherens, alcontrario que los dos complejos de uniónanteriores, establecen conexiones puntuales enforma de disco entre células vecinas, como sifuesen remaches. Son muy abundantes entre lascélulas epiteliales y entre las musculares, perotambién en otros tejidos como el nervioso. Lasuniones entre células están mediadas pormoléculas del tipo cadherinas denominadasdesmogleínas y desmocolinas. El dominiointracelular de estas cadherinas contacta con losfilamentos intermedios como las queratinas,gracias a proteínas intermediarias.
Los hemidesmosomas y las uniones focalesestablecen uniones fuertes entre las células y lamatriz extracelular. En ambos casos las unionesse establecen por integrinas. Loshemidesmosomas unen las células epiteliales ala lámina basal gracias al dominio extracelular
de la integrina, mientras que el dominio intracelularcontacta con los filamentos intermedios citosólicos.Aunque los hemidesmosomas parecen desmosomas sinuna de sus mitades, molecularmente son diferentes. Lasuniones focales unen a las células con diversos tipos dematrices extracelulares gracias a otro tipo de integrinas
Organización y composición de los desmosomas (modificado de
Huber 2003)
Imágenes de microscopía electrónica de transmisión de la
epidermis mostrando desmosomas y hemidesmosomas.
La célula. Membrana celular. 29
Atlas de histología vegetal y animal. Universidad de Vigo.

que en su dominio intracelular contacta con losfilamentos de actina.
Algunos autores suelen colocar en este apartado deestructuras cohesivas macromoleculares a las unionesen hendidura. Estas son uniones entre célulasestablecidas por unas moléculas denominadas
conexinas. Sin embargo, las uniones en hendidura notienen como principal misión cohesionar tejidos sinopermitir la comunicación directa entre citoplasmas decélulas vecinas, gracias a los canales que crean lasconexinas. Por tanto, veremos estas estructuras cuandohablemos de la comunicación celular.
Esquema un hemidesmosma localizado en la base de un epitelio de
mamífero. (Modificado de Hahn 2001)
La célula. Membrana celular. 30
Atlas de histología vegetal y animal. Universidad de Vigo.

Membrana celular
Edidin M. 2003. Lipids on the frontier: a century of cell
membrane bilayers . 2003. Nature reviews in molecular and
cell biology. 4:414418.
Nicolson GL. 2014. The fluidmosaic model of membrane
structure: still relevant to understanding the structure,
function and dynamics of biological membranes after more
than 40 years. Biochimica and biophysica acta. 1838:1451
1466.
Lípidos
Bissig C, Gruenberg J. 2013. Lipid sorting and multivesicular
endosome biogenesis. Cold Spring Harbour perspectives in
biology. 5:a016816.
Janmey PA, Kinnunen PKJ. 2006. Byophisical properties of
lipids and dynamic membranes. Trends in cell biology.
16:538546.
Vance JE. 2015. Phospholipid synthesis and transport in
mammalian cells. Traffic. 16:1.
van Meer G, Voelker DR, Feigenson GW. 2008. Membrane
lipids: where are they a how they behave. Nature reviews in
molecular cell biology. 9:112124.
Glúcidos
Fuster MM, Esko JD . The sweet and sour of cancer: glycans
as novel therapeutic targets. Nature reviews cancer. 2005.
5(7):526542.
Permeabilida, fluidez, heterogeneidad
Jarsch IK, Daste F, Gallop JL (2014). Membrane curvature in
cell biology: an integration of molecular mechanisms. Journal
of cell biology. 214: 275387.
Asimetría, fusión, reparación
Bittner GD, Spaeth CS, Poon AD, Burgess ZS, McGill CH.
2016. Repair of traumatic plasmalemmal damage to neurons
and other eukaryotic cells. Neuronal regeneration research
11:10331042.
Daleke DL. 2007. Phospholipid flippases. The journal of
biological chemistry 282:821825.
McNeil PL, Steinhardt RA. 2003. Plasma membrane
disruption: repair, prevention, adaptation. Annual review in
cell and development biology. 19:697731.
Nicolson GL. 2014. The Fluid—Mosaic Model of Membrane
Structure: Still relevant to understanding the structure,
function and dynamics of biological membranes after more
than 40years. Biochimica et Biophysica Acta (BBA)
Biomembranes. 1838(6): 14511466.
Quazi F, S. Molday RS 2011. Lipid transport by mammalian
ABC proteins. Essays in biochemistry. 50, 265–290.
Adhesión
Hynes RO. 1999. Cell adhesion: old and new questions.
Trends in neurosciences. 9(12): M33M37.
Luo BH, Carman CV, Springer TA. 2007. Structural basis of
integrin regulation and signaling. Annual review of
immunology. 24: 619647.
Complejos de unión
Hynes RO. 1999. Cell adhesion: old and new questions.
Trends in neurosciences. 9(12): M33M37.
Luo BH, Carman CV, Springer TA. 2007. Structural basis of
integrin regulation and signaling. Annual review of
immunology. 24: 619647.
Atlas de histología vegetal y animal. Universidad de Vigo.
La célula. Membrana celular. 31















![[PPT]LA MEMBRANA PLASMÁTICA - Biologia Blog | … · Web viewTEMA 8. LA MEMBRANA PLASMÁTICA Y LAS ENVOLTURAS EXTERNAS 1. LA MEMBRANA PLASMÁTICA 1.1. MEMBRANA UNITARIA Membrana](https://static.fdocuments.ec/doc/165x107/5bcd58cf09d3f206798d90a2/pptla-membrana-plasmatica-biologia-blog-web-viewtema-8-la-membrana-plasmatica.jpg)


