mecanismos de defensa.pdf
17
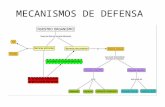
MECANISMOS DE DEFENSA Aquellos mecanismos que están presentes desde el nacimiento se denominan inespecíficos o innatos y actúan en forma inespecífica contra cualquier agente agresor. Por el contrario, los mecanismos específicos o adquiridos van apareciendo en el transcurso de la vida a medida que el individuo se enfrenta a diversos agentes microbiológicos, contra los cuales va desarrollando una respuesta específica que lo protege de futuros contactos. Ambos sistemas se complementan en sus funciones defensivas en forma muy armónica. Sin embargo, a veces la respuesta normal defensiva resulta de efectos deletéreos para el huésped. Por otro lado, los microorganismos han ido desarrollando estrategias para evadir ciertos mecanismos defensivos del huésped. MECANISMOS INESPECÍFICOS O INNATOS La capacidad de un virus de infectar a un huésped está condicionada por la presencia de receptores en la célula huésped, y determinada genéticamente. Como son propios de cada especie, los virus que afectan a animales rara vez encuentran receptores para invadir células humanas, aunque hay excepciones (rabia, ciertas cepas de influenza A, hantavirus). Es posible que los receptores varíen con la edad, sexo, estados fisiológicos (embarazo), enfermedades concomitantes, estrés, y sean un factor importante en la variación de la susceptibilidad que existe en los individuos de una población ante los virus. Los receptores no están diseñados en la célula específicamente para los virus, sino que cumplen otras funciones, y resultan estructuralmente adecuados para la adsorción de ciertos virus. Así se ha demostrado que los virus HIV usan como receptor al CD4 de los linfocitos T, el virus Epstein Barr utiliza el C3H, etcétera. La edad, sexo y estado nutritivo deben considerarse como una suma de factores. Por ejemplo, la desnutrición se asocia frecuentemente a carencias nutritivas específicas, hacinamiento, malos hábitos higiénicos, bajo nivel socioeconómico, falta de control de salud, siendo difícil determinar qué papel juega la desnutrición en sí en la susceptibilidad ante un agente específico. La integridad anatómica y funcional de las barreras mecánicas (piel, mucosas) es fundamental para evitar infecciones por diversos microorganismos (Figura 1). A nivel de mucosas respiratoria, digestiva, genital, conjuntival, etc., existe todo un sistema defensivo basado en la existencia de cilios, secreciones mucosas que contienen enzimas y otros elementos que impiden o dificultan la adsorción viral. En esta permanente lucha de las mucosas por eliminar los agentes agresores muchas veces se rompe el equilibrio por otras razones (tabaquismo, contaminación ambiental, alcoholismo, enfriamiento, infección masiva, etc), lo que facilita el progreso de la infección.
-
Upload
clint-benjamin -
Category
Documents
-
view
252 -
download
3
Transcript of mecanismos de defensa.pdf

MECANISMOS DE DEFENSA Aquellos mecanismos que están presentes desde el nacimiento se denominan inespecíficos o innatos y actúan en forma inespecífica contra cualquier agente agresor. Por el contrario, los mecanismos específicos o adquiridos van apareciendo en el transcurso de la vida a medida que el individuo se enfrenta a diversos agentes microbiológicos, contra los cuales va desarrollando una respuesta específica que lo protege de futuros contactos. Ambos sistemas se complementan en sus funciones defensivas en forma muy armónica. Sin embargo, a veces la respuesta normal defensiva resulta de efectos deletéreos para el huésped. Por otro lado, los microorganismos han ido desarrollando estrategias para evadir ciertos mecanismos defensivos del huésped. MECANISMOS INESPECÍFICOS O INNATOS La capacidad de un virus de infectar a un huésped está condicionada por la presencia de receptores en la célula huésped, y determinada genéticamente. Como son propios de cada especie, los virus que afectan a animales rara vez encuentran receptores para invadir células humanas, aunque hay excepciones (rabia, ciertas cepas de influenza A, hantavirus). Es posible que los receptores varíen con la edad, sexo, estados fisiológicos (embarazo), enfermedades concomitantes, estrés, y sean un factor importante en la variación de la susceptibilidad que existe en los individuos de una población ante los virus. Los receptores no están diseñados en la célula específicamente para los virus, sino que cumplen otras funciones, y resultan estructuralmente adecuados para la adsorción de ciertos virus. Así se ha demostrado que los virus HIV usan como receptor al CD4 de los linfocitos T, el virus Epstein Barr utiliza el C3H, etcétera. La edad, sexo y estado nutritivo deben considerarse como una suma de factores. Por ejemplo, la desnutrición se asocia frecuentemente a carencias nutritivas específicas, hacinamiento, malos hábitos higiénicos, bajo nivel socioeconómico, falta de control de salud, siendo difícil determinar qué papel juega la desnutrición en sí en la susceptibilidad ante un agente específico. La integridad anatómica y funcional de las barreras mecánicas (piel, mucosas) es fundamental para evitar infecciones por diversos microorganismos (Figura 1). A nivel de mucosas respiratoria, digestiva, genital, conjuntival, etc., existe todo un sistema defensivo basado en la existencia de cilios, secreciones mucosas que contienen enzimas y otros elementos que impiden o dificultan la adsorción viral. En esta permanente lucha de las mucosas por eliminar los agentes agresores muchas veces se rompe el equilibrio por otras razones (tabaquismo, contaminación ambiental, alcoholismo, enfriamiento, infección masiva, etc), lo que facilita el progreso de la infección.

La inflamación, con todos los mecanismos que involucra (fiebre, fagocitosis por macrófagos y neutrófilos, liberación de mediadores, etc), representa un mecanismo defensivo muy eficiente. Sin embargo, la alteración fisiológica que conlleva puede ser la causa de las manifestaciones clínicas, en ocasiones exagerada y deletérea. Por eso, debe considerarse que las drogas de acción antiinflamatoria pueden ser beneficiosas o dañinas en la evolución de la infección, dependiendo del mecanismo de lesión en juego. Este ha de tenerse especialmente presente al prescribir corticoides, que son potentes antiinflamatorios. La producción de interferones (alfa, beta, gama) es un mecanismo muy eficiente de defensa que actúa rápidamente y contribuye a la mejoría de la infección. Los interferones son proteínas producidas por leucocitos, fibroblastos y linfocitos T en respuesta a una infección viral u otros estímulos; tienen una triple acción como antiviral, inmunomodulador y citostático. Los interferones son específicos para la especie que los produce, debido a su interacción con el receptor de la membrana celular. pero tienen un amplio espectro de acción que les permite actuar sobre la mayoría de los virus, por lo que parecen ser elementos ideales para el control de las infecciones virales. Sin embargo, la rápida degradación que sufren en el organismo, los efectos secundarios que inducen y las dificultades en su biosíntesis, han limitado su uso terapéutico a ciertas situaciones, como se explica en el capítulo de antivirales. Naturaleza de los interferones. Hay 3 clases de interferón (IFN) humano: α, β y γ (Tabla 1). Los IFN-α y β son producidos con mayor frecuencia por leucocitos y fibroblastos respectivamente, en respuesta a la infección viral así como a una variedad de inductores. Estos se denominan interferones tipo I. El IFN-γ, conocido también como interferón inmune o interferón tipo II, es producido por células linfoides no sensibilizadas en respuesta a mitógenos y linfocitos T sensibilizados, estimulados por antígenos específicos. La separación de los interferones en estas 3 clases se basa principalmente en su antigenicidad; son inmunológicamente diferentes, y por ello, los antisueros contra una clase son incapaces de neutralizar a los interferones de las otras dos clases. Todos los interferones son derivados de precursores que tienen 166 aminoácidos (aa); poseen una secuencia señal en el extremo N-terminal de 20-23 aminoácidos de largo, siendo el tamaño final de estas proteínas de 143 aa para IFN-α, 145 aa para IFN-β y 146 aa para IFN-γ. Producción de los interferones. Las células en condiciones normales no contienen ni sintetizan interferón, lo que indica ausencia de transcripción de los genes que codifican para estas proteínas. Sin embargo, su formación puede inducirse por una variedad de factores, tales como una infección viral, moléculas de RNA de doble hebra, algunas bacterias y otros componentes que estimulan la respuesta inmune. El inductor más importante de la producción de interferón resulta ser la infección por virus; en general, los virus RNA son mejores inductores que los virus DNA. Durante la infección viral de una célula se produce la liberación del genoma viral (DNA o RNA). La presencia o replicación del genoma viral se asocia con una desrepresión de los genes que codifican para los interferones. Consecuentemente,

se produce la síntesis de mRNA y posterior producción de proteínas, en este caso, interferón. La producción de interferón es independiente del tipo viral infectivo; comienza alrededor de 4 horas postinfección, y, en forma paralela a la síntesis de proteínas virales, alcanza un máximo, para declinar posteriormente. Los interferones producidos son secretados al medio extracelular, uniéndose a receptores específicos expresados tanto en la misma célula como en las células vecinas no infectadas. Su vida media efectiva es relativamente corta (alrededor de 4-6 horas). Los interferones son capaces de disminuir la carga viral en células ya infectadas; sin embargo, son más efectivos en inhibir la replicación viral en células vecinas o ubicadas a distancia. Después de la unión interferón-receptor celular, se induce la síntesis de diversas moléculas que actúan a nivel de la transcripción del mRNA viral, en la traducción de las proteínas virales y en el ensamblaje y/o liberación de partículas virales. Mecanismo de acción. Una de las proteínas inducidas por el interferón es capaz de iniciar una serie de reacciones que llevan a la activación de una enzima presente en forma latente, la cual es capaz de degradar el mRNA viral. Este sistema de adeninas (2,5 Oligo-A), que actúa como activador de una ribonucleasa latente, implica la síntesis de una enzima, que a su vez sintetiza un activador de un pequeño oligonucleótido, latente en el citoplasma celular. Esta ribonucleasa degrada al mRNA viral, inhibiendo la traducción de las proteínas virales; aunque también es capaz de degradar mRNA celular, su efecto es mayor sobre los mRNA virales. Otro mecanismo que permite explicar la acción antiviral del interferón involucra la inducción de la síntesis de otra proteína y su subsecuente activación. Corresponde a una proteína quinasa de 67 kd que normalmente está inactiva; sin embargo, frente a una infección viral y a la producción de interferón, esta quinasa se autofosforila y es capaz, a su vez, de fosforilar el factor de iniciación 2 (elf-2) que participa en la traducción del mRNA en el ribosoma. Esto produce la inhibición de la síntesis de proteínas virales.
Además de inducir la síntesis de enzimas que inhiben la multiplicación viral, el interferón afecta una variedad de funciones celulares como: el crecimiento celular, tanto de células normales como tumorales; la diferenciación celular; el aumento de la actividad de células NK; la inhibición de la migración de los leucocitos; el aumento de la citotoxicidad mediada por anticuerpos, etc. Hoy en día se considera a los IFN como citoquinas capaces de transmitir señales regulatorias entre células, y por lo tanto, capaces de ejercer funciones inmunomoduladoras.
Actualmente, una de las áreas de mayor interés es la relacionada con el estudio de la actividad antiproliferativa e inmunomoduladora del interferón en el tratamiento del cáncer humano. El IFN-α ha sido empleado como terapia para una variedad de tumores con mejores resultados que el IFN-γ. MECANISMOS ESPECÍFICOS, INMUNES O ADQUIRIDOS El huésped inmunocompetente desarrolla mecanismos específicos de inmunidad adquirida, a nivel humoral (anticuerpos) y celular, dirigidos contra distintos

constituyentes del agente infectante que se expresan durante la infección. Ambos componentes de la respuesta inmune están interrelacionados, se pueden producir a nivel local, asociado a mucosas, y a nivel sistémico, y son esenciales para controlar la infección e inducir resistencia duradera frente a las reinfecciones. Los virus que destruyen las células para salir al medio extracelular son fundamentalmente controlados por la respuesta humoral, mientras que aquellos que lo hacen por yemación, sin lisis, están limitados básicamente por mecanismos de inmunidad mediados por células (Figuras 7 y 8). En este caso, los anticuerpos pueden participar en la neutralización viral si existe viremia, o en la lisis de células infectadas en conjunto con el complemento o con ciertas células citotóxicas asesinas (células killer o natural killer). Esta respuesta inmune adquirida está definida por su especificidad y por su capacidad de memoria, de tal forma que en un contacto posterior con el mismo antígeno, la respuesta secundaria será más intensa, precoz y prolongada, por existir un conocimiento previo almacenado como memoria. Cada individuo nace con una cantidad de clones de linfocitos, los cuales tienen un receptor específico capaz de reconocer un determinado epítopo. Durante la respuesta primaria ocurre una expansión clonal de poblaciones de linfocitos específicos para un antígeno, de tal forma que, posteriormente, perduran linfocitos B y T capaces de responder al antígeno en el futuro. Estas células de memoria poseen receptores más afines al antígeno, debido a la selección ocurrida durante la respuesta primaria. Inmunidad humoral. En la inmunidad humoral, los mecanismos efectores son los anticuerpos o inmunoglobulinas (Ig), glicoproteínas producidas por los linfocitos B y constituidas por 4 cadenas polipeptídicas, dos pesadas y dos livianas. Estos anticuerpos desempeñan diversas funciones: eliminar la infección viral primaria, limitar la viremia, limitar la enfermedad y prevenir las reinfecciones. El organismo produce anticuerpos contra todos los constituyentes inmunogénicos del virión con los que se contacta. Aquellos que se dirigen contra los constituyentes externos del virión serán capaces de neutralizar la acción de éste y se denominan anticuerpos neutralizantes, porque determinan la pérdida de la infectividad de los viriones. Esta acción se ejercería de muchas formas: inhibiendo la adsorción del virión a los receptores celulares, evitando la penetración viral, inhibiendo la decapsulación, provocando la lisis inmune de las partículas mediante anticuerpos y complemento, y favoreciendo la fagocitosis mediante opsonización y agregación de las partículas virales. En el primer contacto con un virus se producen los anticuerpos en la denominada respuesta primaria. Esta respuesta es bifásica, con una primera fase constituida por IgM (a los 5-8 días postinfección), y la segunda por IgG (15-20 días). La IgA aparece en el suero en menor cantidad y en forma posterior. La corta duración de la IgM, generalmente 2-3 meses, aunque puede variar según la infección viral, permite su utilización en el diagnóstico de una infección reciente. Por el contrario, la IgG dura meses, años y a veces, toda la vida.

En las respuestas secundarias (reinfecciones) puede haber una respuesta anamnésica de IgM de corta duración, pero lo que siempre está presente es una producción importante de IgG. Esta respuesta es más rápida e intensa que la primaria, cumpliendo un papel importante en el control temprano de la infección. La magnitud de la respuesta inmune dependerá de factores tales como la naturaleza del virus, su puerta de entrada, su forma de diseminación, etc. Es así como en el caso de existir viremia, la estimulación antigénica será intensa y afectará a todo el tejido linfoide del organismo y la respuesta de anticuerpos será de gran magnitud. En las infecciones locales como las respiratorias, el virus queda restringido a la mucosa, existiendo una producción local de IgA secretoria en los nódulos linfoides de la submucosa; en estos casos, la respuesta sérica es de escasa magnitud. Se ha detectado IgA secretoria específica antiviral en secreciones respiratorias, oculares, intestinales, genitales y en la leche materna. Muchos virus inducen la producción de esta inmunoglobulina y es de especial importancia en la prevención de las reinfecciones por virus respiratorios. Su corta duración, 1-4 años, contribuye a la repetición de episodios de afecciones virales respiratorias que se presentan durante nuestra vida. Inmunidad celular. La participación de la inmunidad celular requiere del procesamiento del material inmunogénico por las células monocíticas fijas (Kuppfer en hígado, microglia en sistema nervioso, macrófagos alveolares, etc.) y circulantes del huésped, quienes lo procesan y “presentan” a los linfocitos. Los elementos efectores son los linfocitos T que pueden agruparse en LTh CD4 (ayudadores), LTs CD8 (supresores) y LTc (citotóxicos). Existen otros linfocitos denominados asesinos naturales (NK) de funciones igualmente citotóxicas Estos linfocitos T expresan en su superficie una proteína fijadora de antígeno en forma específica, el receptor de las células T (TcR). La fijación de un antígeno a este receptor inicia una cascada de reacciones de activación que inducen la proliferación y la diferenciación de las células T. El reconocimiento de estos antígenos se realiza sólo cuando están presentes en la superficie de las células presentadoras de antígenos (APC), asociados a los antígenos del complejo mayor de histocompatibilidad (MHC) de clase I para LTs y c y de clase II para LTh. (Figuras 2 y 3) Tras la activación de las células T, éstas intervienen estimulando o suprimiendo las respuestas inmunes de otras células o destruyendo la célula blanco, mediante procesos de contacto entre membranas celulares (citotoxicidad) o por liberación de mediadores. Las células T citotóxicas CD8 reconocen el antígeno en el contexto de los antígenos de clase I del MHC, que se encuentran en prácticamente todas las células del organismo. La destrucción celular se llevaría a cabo por la liberación de enzimas (Figuras 3 y 4). Los LT CD4 o colaboradores regulan toda la respuesta inmune, incluyendo la regulación de las funciones de los LB y de los LT citotóxicos. El reconocimiento del

antígeno sólo se efectúa si está en asociación con los antígenos de clase II del MHC, presente sólo en algunas células presentadoras de antígenos del sistema inmune, como linfocitos B y macrófagos (Figura 6). Estos LTh se han clasificado según el tipo de respuesta inducida: Th1, si es predominantemente defensiva, por la producción de IL 2 e interferón gama; Th2, o inflamatoria, por producción de IL 4, IL5 e IL 13 que inducen síntesis de IgE, maduración, migración y degranulación de eosinófilos, fenómenos típicos de la respuesta alérgica.
En general, la inmunidad celular es importante en la resistencia a los patógenos que replican en el interior de las células, y dado el carácter de parásitos intracelulares que poseen los virus, es fácil comprender su importancia en el control de las infecciones virales (Figura 7). Como se mencionaba en un comienzo, las respuestas inmunes humoral y celular están interrelacionadas (Figura 2). Es así como la activación de los LB requiere del contacto con los LT, aunque también existen antígenos capaces de hacer una activación “T independiente” de los LB. Las células B ingieren y procesan el antígeno unido a la inmunoglobulina y presentan el inmunógeno junto con la proteína del MHC. Esta combinación es reconocida por receptores específicos para el antígeno, presentes en los LT CD4 o ayudadores, determinando la unión de ambas células (Figura 5). Este contacto directo permite el intercambio de señales de activación y la producción de proteínas inmunorreguladoras denominadas citoquinas que se unen a receptores específicos de los LB (Tabla 2). Los LT CD8 pueden inhibir la producción de anticuerpos actuando directamente sobre las células B, o a través de la supresión de una célula T que estimule la producción de anticuerpos. EVASIÓN DE LA RESPUESTA INMUNE. La respuesta inmune específica permite tanto la recuperación de una infección como la prevención de futuros episodios y tiende a controlar y eliminar el agente infectante. Ante esto se han desarrollado diversos mecanismos de evasión a esta respuesta que explican la persistencia de la infección y la sobrevida del agente (Tabla 3). A continuación se comentan algunos de los mecanismos mencionados en la Tabla: - Los herpesvirus se propagan por vecindad y pasan de la célula infectada a la vecina sin alcanzar el compartimento extracelular, evadiendo así los mecanismos defensivos extracelulares. - Algunos virus tienen la capacidad de experimentar variaciones estructurales que hacen ineficiente la inmunidad adquirida contra las cepas precedentes (influenza A). La gran variedad de serotipos existentes en la mayoría de los grupos de virus (3 poliovirus, más de 100 rinovirus, 46 adenovirus, 72 enterovirus, hepatitis A,B,C,D,E, etc.), significa que los virus se han diversificado evolutivamente a tal punto que no inducen una respuesta inmune común de grupo (heterotípica), sino que se requiere el montaje de una respuesta específica para cada virus (homotípica). La tecnología

actual permite estudiar más profundamente la estructura de los virus, y cada día surge la pregunta sobre cuántas variantes se han descrito de un virus y qué relación tiene con su patogenicidad, virulencia y con los otros aspectos de la virología. - La localización de algunos virus en sitios ocultos al sistema inmunocompetente facilita su persistencia (virus sarampión en sistema nervioso, virus herpes simplex en ganglios sensitivos). - La formación de complejos inmunes de los virus invasores con moléculas de anticuerpos dificulta su reconocimiento como partículas extrañas y su consiguiente eliminación. - La depresión inmunológica transitoria o permanente que inducen algunos virus dificulta la respuesta inmune. Se ha demostrado que muchos virus alteran la respuesta inmune (HIV, citomegalovirus, influenza, sarampión), ya sea al destruir las células inmunocompetentes o al producir un desequilibrio en su acción, como se ilustra en la Tabla 4.
AUTOINMUNIDAD. La respuesta inmune generalmente es beneficiosa y permite la eliminación del agente infectivo. Se ha mencionado que la respuesta inflamatoria puede ser responsable de los signos y síntomas de la infección. Una condición básica del sistema inmune es su capacidad de discriminar lo propio de lo ajeno, para evitar desarrollar una respuesta contra antígenos propios. Sin embargo, este reconocimiento puede fallar y el sistema inmunocompetente, en conjunto con la respuesta inflamatoria inespecífica, desarrolla una reacción contra el propio huésped que es deletérea. Existen mecanismos patogénicos de autoinmunidad y a los virus se les ha responsabilizado de muchos de ellos; algunos se mencionan en la Tabla 5. Los virus envueltos forman su manto con proteínas virales que se intercalan entre las proteínas de la membrana de la célula, de modo que el virus lleva en su manto algunos antígenos del huésped. Pueden darse dos situaciones: primero, que el virus se replique en un lugar de difícil acceso para el sistema inmunocompetente, como el sistema nervioso, y al salir los virus hacia la circulación general arrastren componentes propios (como mielina) que son considerados ajenos por el sistema inmune. Segundo, que las proteínas normales de la célula sufran pequeñas alteraciones al integrarse al manto viral. En ambos casos el sistema inmune va a desarrollar una respuesta contra antígenos celulares propios que daña a la misma célula. Otros ejemplos incluyen la desregulación del control de la respuesta inmune por alteración de linfocitos T ayudadores y supresores, con respuesta policlonal contra antígenos normales (HIV, virus Epstein- Barr).
CUESTIONARIO
1. Mencione mecanismos inespecíficos de defensa que sean importantes en la protección frente a una infección viral.
2. ¿Qué alteraciones de los mecanismos inespecíficos de defensa pueden ser

responsables de infecciones respiratorias repetitivas? 3. Mencione características de la respuesta inmune adquirida. 4. ¿En qué se diferencia una respuesta inmune primaria de una secundaria? 5. ¿En qué se diferencia la respuesta inmune frente a una infección local de una
infección sistémica? 6. ¿Qué papel cumple el MHC? 7. Frente a los virus extracelulares, ¿qué tipo de inmunidad específica es más
relevante y por qué? 8. Frente a los virus intracelulares, ¿qué tipo de inmunidad específica es más
relevante y por qué? 9. Explique 3 mecanismos de evasión de la respuesta inmune, dando ejemplos. 10. Mencione algunos virus capaces de determinar inmunosupresión en el ser
humano. 11. Describa 2 mecanismos de autoinmunidad, enfatizando las repercusiones
que pueden tener para el huésped. FIGURA 1.
MECANISMOS DE DEFENSA INESPECÍFICOS
Lisozima (lágrimas)Secreciones, remoción partículas Cilios y mucus
Ácidez gástrica
Flujo urinario
Piel: barrera mecánica
Secreciones genitales
TABLA 1. CARACTERÍSTICAS DE LOS INTERFERONES HUMANOS PARÁMETRO IFN-α IFN-β IFN-γ
CÉLULAS PRODUCTORAS Leucocitos periféricos Fibroblastos Linfocitos

AGENTES INDUCTORES
Infección viral, RNA doble hebra, bacteria
Infección viral, RNA doble hebra
Mitógenos, Antígenos
LOCALIZACIÓN CROMOSÓMICA
9 9 12
TAMAÑO (aa)
143 145 146
ESTABILIDAD A pH 2
sí sí no
ACTIVIDAD EN SDS
sí sí no
TABLA 2. ALGUNAS CITOQUINAS DEL SISTEMA INMUNE CITOQUINA
PRODUCIDA POR
CÉLULAS BLANCO
EFECTOS EN LA RESPUESTA INMUNE
IL-2
Células T Células T y NK Proliferación de células activadas por antígenos y de otras células inmaduras (ej: NK)
IL-3 (CSF)
Células T (CD4+)
Células progenitoras (Médula ósea)
Crecimiento/ diferenciación de todas las células
IL-4
Células T (CD4+)
Células B Células T
Estimulación del cambio de cadena pesada de la Ig a isotipo IgE Desarrollo y diferenciación de las células T
IL-5
Células T Células B Eosinófilos
Crecimiento y diferenciación de eosinófilos y activación de eosinófilos maduros
IL-7
Células estromales (médula ósea)
Linfocitos Factor de crecimiento (genera células pre- B y pre- T)
IL-10
Macrófagos Células T
Macrófagos Células T
Inhibición de la función de los macrófagos Control de la respuesta inmune
IL-15
Macrófagos Otros
Células NK y T Proliferación
IFN-α Tipo 1
Macrófagos Todas las células
Interfiere con la replicación viral
IFN-γ Tipo 1 Fibroblastos Células NK Activación

TNF
Macrófagos Células T
Leucocitos y Células endoteliales
Activación/proceso inflamatorio
TGF-β
Células T Macrófagos Otros
Células T Otras Macrófagos
Inhibe la activación y proliferación de linfocitos Inhibe la activación de macrófagos
IL = interleuquina CSF = factor estimulador de colonia IFN = interferón TNF = factor de necrosis tumoral TGF = factor transformador de
TABLA 3. MECANISMOS DE EVASIÓN A LA RESPUESTA INMUNE DE ALGUNOS VIRUS
------------------------------------------------------------------------------------------------------------------MECANISMOS EJEMPLOS ------------------------------------------------------------------------------------------------------------------ Respuesta humoral o celular pobre Virus respiratorio sincicial (RSV) Propagación a célula contigua Virus herpes simplex, virus rabia Variedad antigénica Virus influenza, rhinovirus, HIV Complejo virus-anticuerpo Virus hepatitis B Modulación presentación antígeno celular Virus sarampión, RSV Camuflaje de receptores Fc Herpesvirus Inmunosupresión HIV, virus sarampión Virus influenza A Latencia Virus herpes -----------------------------------------------------------------------------------------------------------------
TABLA 4. ALGUNOS VIRUS RESPONSABLES DE INMUNOSUPRESIÓN ------------------------------------------------------------------------------------------------------------------ VIRUS HUÉSPED FUNCIÓN AFECTADA OBSERVACIONES LT LB NK2 Macrófago ------------------------------------------------------------------------------------------------------------------ Sarampión humano + + +

Distemper perro + + + rofunda supresión HTLV-11 humano + + especialmente LT h3 HIV humano + + gran compromiso LT h3 Citomegalovirus humano + + + + ¿agente secundario? Epstein Barr humano + + activación policlonal LB Herpes simplex humano + sólo in vitro Influenza pollo + destruye linfocitos ------------------------------------------------------------------------------------------------------------------ 1. HTLV-1 : Virus humano linfotrópico de células T 2. NK: Células natural killer 3. LT h: Linfocito T helper
TABLA 5. POSIBLES MECANISMOS DE AUTOINMUNIDAD MEDIADOS POR VIRUS
------------------------------------------------------------------------------------------------------------------ 1. Cambios en el sistema inmune del huésped: - Activación directa policlonal de linfocitos B - Alteración directa o indirecta de células inmunoreguladoras, como linfocitos T supresores, natural killer, otros. - Alteración de liberación de linfoquinas (interferón, factor de necrosis tumoral). 2. Efecto dañino de anticuerpos antivirales: - Semejanza molecular. - Anticuerpos antiidiotipos. - Formación de complejos inmunes. 3. Cambios en los antígenos del huésped inducidos por virus: - Exposición de antígenos ocultos. - Modificación de antígenos de superficie. - Inducción de nuevos antígenos. - INcorporación de antígenos celulares por virus . ----------------------------------------------------------------------------------------------------------------

FIGURA 2. RESPUESTA INMUNE CELULAR
FIGURA 3.
ACTIVACIÓN DE LT CD8 ACTIVACIÓN DE LT CD8
Señal específica y Señal específica y coco--estimuladorestimulador
DestrucciónDestrucción celular celular
Célula infectadaCélula infectada
MHC-I
CD8
ACTIVACIÓN DE LT CD8ACTIVACIÓN DE LT CD8

FIGURA 4. DESTRUCCIÓN DE LA CÉLULA INFECTADA POR EL LT CD8
En la célula infectada se producen proteínas virales que se rompen en péptidos, los cuales son transportados al retículo endoplásmico, donde son rodeados por moléculas de MHC- I. Estos complejos viajan a la superficie celular, pudiendo ser detectados por un LT citotóxico que expresa una proteína CD8 (1). Posteriormente, el LT secreta compuestos que destruyen a la célula infectada.
ACCIÓN ACCIÓN DEL DEL LT LT
Estimulación del LT Estimulación del LT naive naive por APC por APC
LT reconoce antígenoLT reconoce antígenoen célula infectadaen célula infectada
LT activado mata aLT activado mata a célula infectada célula infectada
FIGURA 5. ACTIVACIÓN DEL LB
Señal específica y Señal específica y coco--estimuladorestimulador
ActivaciónActivación celular celular
LB LB
ACTIVACIÓN DE LBACTIVACIÓN DE LB

El anticuerpo presente en la superficie del LB actúa como su receptor, al cual se puede unir un antígeno, el que será depositado en una vesícula en el interior de la célula, rompiéndose en péptidos. Una molécula de MHC -II migra a la vesícula y se une al péptido, transportándose a la superficie celular. Un LT CD4 helper se une y activará la proliferación de los LB y la producción de anticuerpos. FIGURA 6.
ACTIVACIÓN DE LT CD4 ACTIVACIÓN DE LT CD4
Señal específica y Señal específica y coco--estimuladorestimulador
ActivaciónActivación celular celular
ACTIVACIÓN DE LT CD4ACTIVACIÓN DE LT CD4

RESPUESTA INMUNE ANTIRESPUESTA INMUNE ANTIVIRUS INTRACELULARVIRUS INTRACELULAR
VIRUS VIRUS INTRACELULARINTRACELULAR
ComplejoComplejo Ag Ag--AcAc
LISIS CELULARLISIS CELULAR
ADCC porADCC porK,K, Mf Mf, PMN,C´, PMN,C´
AcAc LTcLTc
LinfoquinasLinfoquinas
InterferónInterferónNKNK
PMNPMN Mf Mf
MIFMIFMAFMAF
LinfotoxinasLinfotoxinas
LEUCOLISINAS,LEUCOLISINAS,CITOTOXICIDAD DIRECTACITOTOXICIDAD DIRECTA
FIGURA 7.

FIGURA 8
RESPUESTA INMUNE ANTIRESPUESTA INMUNE ANTIVIRUS EXTRACELULARVIRUS EXTRACELULAR
VIRUS EXTRACELULARVIRUS EXTRACELULAR
ComplejoComplejoAgAg--AcAc
AcsAcs
NeutralizaciónNeutralización
LisisLisis
OpsonizaciónOpsonización
R.I R.Icelularcelular
LinfoquinasLinfoquinas
PMNPMN Mf Mf
MAFMAFMIFMIF
FagocitosisFagocitosis
BIBLIOGRAFíA
1.- Abbas A, Lichtman A, Pober J. Cellular and molecular immunology. Philadelphia: WB Saunders, 1991. 2.- Ahmed R, Stevens J. Viral persistence. En: Fields B. Virology, 2ª ed. New York: Raven Press, 1991; 241-265. 3. Collier L, Oxford J. How viruses cause disease. En: Collier L, Oxford J. Human Virology. Oxford: Oxford University Press, 1993; 39- 55. 4. Fields B. Biología de los virus. En: Schaechter M, Medoff G, Eisenstein B, Guerra H. Microbiología. 2ª ed. Buenos Aires: Editorial Médica Panamericana, 1994: 415- 436. 5. Hendley JO, Gwaltney J. Mechanisms of transmission of rhinovirus infections. Epidemiol. Rev. 1988; 10: 242- 58.

6. Jawetz E, Melnick J, Adelberg E. Patogenia y control de las enfermedades virales. En: Jawetz E, Melnick J, Adelberg E. Microbiología Médica, 14ª ed. México: El Manual Moderno, 1992; 421- 439. 7. Roitt I, Brostoff H, Male D. Immunology, 3ª ed. Londres: Mosby-Year Book Europe Limited, 1993. 8. Schattner A, Rager-Zisman R. Virus-induced autoimmunity. Rev Infect Dis 1990; 12 (2): 204-22. 9. Tramont E. Mecanismos de defensa generales o inespecíficos del huésped. En: Mandell G, Douglas R, Bennet J. Enfermedades infecciosas. Principios y prácticas, 3ª ed. Buenos Aires: Editorial Médica Panamericana, 1991; 35-42. 10. Tyler K, Fields B. Pathogenesis of viral infections. En: Fields BN, Knipe DM, Howley PM et al. Virology, 3ª ed. Philadelphia: Lippincott- Raven Publishers, 1996; 173- 218.