Los efectos de la desnutrición sobre los procesos de … · DOS POR LA DESNUTRICION. En tØrminos...
21
Los efectos de la desnutricin sobre los procesos de aprendi- zaje y memoria en modelos ani- males. Solicitud de sobretiros: Sofa Daz-Cintra, Centro de Neurobiologa, Universidad Nacional Autnoma de MØxico. Ciudad Universitaria. MØxico. Apdo. Post. 70-228, MØxico, 04510, D.F., MØxico. Recibido el 16/Dic./1993. Aceptado para publicacin 26/Abril/1994. Mayra Garca-Ruiz, Leticia Parra, Sofa Daz-Cintra. Centro de Neurobiologa, Universidad Nacional Autnoma de MØxico. Ciudad Universitaria. MØxico, D.F., MØxico. RESUMEN. El sistema nervioso central (SNC) es extre- madamente vulnerable a diversos factores del me- dio ambiente. Uno de los factores que provoca efectos drÆsticos en el desarrollo de este sistema es la desnutricin. En los œltimos aæos se han tratado de relacionar los daæos provocados por la desnutricin sobre el SNC con los procesos cognitivos. Sin embargo y a pesar de las eviden- cias que muestran que los seres humanos desnutri- dos presentan cambios conductuales y sobretodo una actividad mental subnormal, es sorprendente el hecho de que los resultados obtenidos hasta la fecha sean controversiales. Algunos autores des- criben que la habilidad para el aprendizaje de los organismos desnutridos es deficiente, otros que esta habilidad no cambia y otros mÆs que es mejor que aquella mostrada en los organismos bien nu- tridos. Es por esto que el objetivo del presente escrito es hacer una revisin del trabajo experi- mental realizado acerca de los efectos de la desnutricin en el comportamiento y especialmen- te en los procesos de aprendizaje y memoria en modelos animales. Palabras clave: Desnutricin, aprendizaje y me- moria. SUMMARY. EFFECTS OF MALNUTRITION ON 88 Rev Biomed 1994; 5:88-108. Vol. 5/No. 2/Abril-Junio, 1994.
Transcript of Los efectos de la desnutrición sobre los procesos de … · DOS POR LA DESNUTRICION. En tØrminos...

Los efectos de la desnutriciónsobre los procesos de aprendi-zaje y memoria en modelos ani-males.
Solicitud de sobretiros: Sofía Díaz-Cintra, Centro de Neurobiología, Universidad Nacional Autónoma de México. Ciudad Universitaria. México. Apdo.Post. 70-228, México, 04510, D.F., México.Recibido el 16/Dic./1993. Aceptado para publicación 26/Abril/1994.
Mayra García-Ruiz, Leticia Parra, Sofía Díaz-Cintra.
Centro de Neurobiología, Universidad Nacional Autónoma de México. Ciudad Universitaria. México,D.F., México.
RESUMEN.
El sistema nervioso central (SNC) es extre-madamente vulnerable a diversos factores del me-dio ambiente. Uno de los factores que provocaefectos drásticos en el desarrollo de este sistemaes la desnutrición. En los últimos años se hantratado de relacionar los daños provocados por ladesnutrición sobre el SNC con los procesoscognitivos. Sin embargo y a pesar de las eviden-cias que muestran que los seres humanos desnutri-dos presentan cambios conductuales y sobretodouna actividad mental subnormal, es sorprendenteel hecho de que los resultados obtenidos hasta lafecha sean controversiales. Algunos autores des-criben que la habilidad para el aprendizaje de losorganismos desnutridos es deficiente, otros que
esta habilidad no cambia y otros más que es mejorque aquella mostrada en los organismos bien nu-tridos. Es por esto que el objetivo del presenteescrito es hacer una revisión del trabajo experi-mental realizado acerca de los efectos de ladesnutrición en el comportamiento y especialmen-te en los procesos de aprendizaje y memoria enmodelos animales.
Palabras clave: Desnutrición, aprendizaje y me-moria.
SUMMARY.
EFFECTS OF MALNUTRITION ON
88
Rev Biomed 1994; 5:88-108.
Vol. 5/No. 2/Abril-Junio, 1994.

LEARNING AND MEMORY PROCESSES INANIMAL MODELS. The central nervous system(CNS) is ext remely vulnerable to var iousenvironmental factors. Malnutrition is a factorwhich causes the most drastic effects on thedevelopment of the CNS. In recent years attemptshave been made to relate the alterations producedby malnutrition on the CNS with the cognocitiveprocesses of the malnourished organisms. Evidenceshows that malnourished human beings presentbehavioural changes and above all subnormal men-tal activity. However, it is surprising to note thatresults obtained hitherto are controversial. Severaldifferent groups have reported that malnourishedanimals can perform worse, equal to or even betterthan well nourished organisms. In this review weanalyzed those experimental studies with referenceto the effects of malnutrition on behaviour andespecially on learning and memory processes inanimal models.
Key words: Malnutrition, learning and memory.
INTRODUCCION.
En años recientes se ha enfatizado la impor-tancia que tiene la nutrición adecuada para el de-sarrollo funcional óptimo de los organismos(1).Esto es debido a la alta incidencia de la desnutricióninfantil en el mundo y a las evidencias que mues-tran que esta alteración provoca daños cerebralesque traen como consecuencia un deterioro en suscapacidades de aprendizaje y memoria y por tanto,una actividad mental subnormal (2-6).
Los estudios referentes a los efectos de ladesnutrición pre-, postnatal o ambas, han descritoseveras alteraciones neurales, a saber: a) modifi-caciones en el ritmo de división celular (7,8), b)reducciones del número de neuronas, de sus rami-ficaciones dentríticas y de sus espinas así como delnúmero de sinapsis en diversas regiones cerebrales
tales como, el núcleo del raphe dorsal, el locuscoeruleos, el cerebelo, el área visual 17 de lacorteza cerebral y el hipocampo (7,9-12), c) re-ducciones en la densidad de las células gliales delcuerpo cal loso (13) y d) inhibic ión de lamielinización cerebral (14).
En vista de estas alteraciones, ha surgido ungran interés por investigar los efectos de ladesnutrición sobre los procesos de aprendizaje ymemoria. Desafortunadamente, los estudios de ladesnutrición en los seres humanos son difíciles deevaluar, ya que en el desarrollo del niño influyendiversos factores, tales como, los genéticos, losmedio ambientales, los culturales y los nutricionalesque hacen difícil especificar la causa exacta deldeterioro mental y más aún, se dificulta el controlde aquellas condiciones antecedentes provocadaspor dichos factores. El uso de modelos animalesparece ser adecuado debido a que en éstos esposible controlar de manera más precisa los diver-sos factores que pueden intervenir sobre su desa-rrollo (15,16).
El objetivo de la presente revisión es haceruna evaluación de los trabajos experimentales másrelevantes -a nuestro juicio- que a la fecha se hanllevado a cabo sobre el efecto de la desnutriciónen el comportamiento y especialmente en los pro-cesos de aprendizaje y memoria en algunos mode-los animales.
En la primera parte, es nuestro propósitorevisar de manera general los cambios conductualesproducidos por la desnutrición y en la segundadiscutiremos los efectos de este factor en los pro-cesos de aprendizaje y memoria a través de dife-rentes paradigmas experimentales.
CAMBIOS CONDUCTUALES PRODUCI-DOS POR LA DESNUTRICION.
En términos generales se ha descrito que ladesnutrición provoca un retraso en la aparición delos reflejos y de las primeras conductas de las ratasdesnutridas en desarrollo (17-19). Diversos estu-
89
M García-Ruiz, L Parra, S Díaz-Cintra.
Revista Biomédica

dios han mostrado que las crías desnutridas com-piten menos con sus hermanos por el alimentomaterno (16,20), tienen alteraciones en el com-portamiento de orientación hacia el nido (21-26),son menos activas, disminuyen su frecuencia deacercamiento hacia sus madres (27) y emiten unmenor número de vocalizaciones que estimulan enla hembra las respuestas de lamido, de acarreo ode alimentación de las crías, por lo que se dice quelas crías desnutridas demandan menos atención dela hembra lactante (28).
Las crías desnutridas tardan algunos díasmás que las controles en que les aparezca el pela-je, se les abran los conductos auditivos externos ylos párpados y les inicie el ciclo estral (29,30). Lasconductas motoras tales como, el desplazamientopara salir del nido, la exploración y el nado ocu-rren más tardíamente (20,31,32); además presen-tan deterioro en la coordinación motora (32-35).Con respecto a la conducta de aseo, también se haobservado un retraso inicial en su desarrollo, queposteriormente se intensifica persistiendo así hastala edad adulta (36).
Se han observado también incrementos en laresponsividad en situaciones en las que el objetivoes el acceso a satisfactores, tales como agua ocomida (37-39), confusión en la discriminaciónentre un olor conocido y el olor materno (40) ymayor reactividad a est ímulos dolorosos oaversivos (41-44).
A largo plazo se observan deficienciasconductuales entre las que son de llamar la aten-ción, por parte de las hembras, descuido en elmantenimiento del nido (45,46) y una disminuciónen la atención maternal hacia las crías (27). En unoy otro sexo, se han encontrado alteraciones en elcomportamiento social (47), respuestas emocio-nales exageradas y reducción en los comporta-mientos explora tor ios (48) . Al terac ionesconductuales similares se han observado tambiénen cerdos desnutridos (49).
Por otra parte, con la prueba de campoabierto se ha observado que ratas desnutridaspostnatalmente permanecen más tiempo en el cua-
dro de salida antes de empezar la exploración(49). El desplazamiento dentro del campo abiertoparece ser sin una finalidad definida y sin husmeo(15,50-52). En esta situación de actividad libre lasratas desnutridas muestran un estado de altonerviosismo de tal manera que cualquier eventoextraño, como un ruido, puede provocar en ellasrespuestas de congelamiento de mayor duración ychillidos (42,50,53). En ambientes novedosos losanimales desnutridos (esto se ha observado enratas y monos) muestran apatía y temor a losobjetos o estímulos del medio ambiente, poca ha-bilidad de los animales para adaptarse a cambiosque se les presenten (44) y disminución de laatención (16,54). Este tipo de respuestas ha lleva-do a la conclusión de que el estado emocionalexagerado de los animales desnutridos interfierecon su capacidad de actuar y explorar en situacio-nes novedosas.
Se ha sugerido que alteraciones en la con-ducta podrían deberse a la alta emotividad quemuestran los organismos desnutridos, ya que ladesnutrición provoca que los animales sean extre-madamente activos, temerosos y agresivos cuan-do se encuentran ante situaciones novedosas(15,48,52,55-58). Un ejemplo de ello es cuandose somete a los animales a pruebas conductuales,en las cuales se ha observado que la reacciónemocional (e.g.: en el caso de las ratas desnutri-das) impide la expresión de una respuesta adecua-da o eficiente (59). Massaro y col. (20) y Salas ycol. (36) describieron un exceso en la conducta deaseo de ratas desnutridas, lo que podría interferircon la expresión de una conducta adecuada o bienpodría ser la expresión de una respuesta emocio-nal exagerada de estos animales. De la mismamanera podría suponerse que la conducta maternainadecuada de hembras sea una consecuencia de laalta emocionalidad que presentan los animales des-nutridos (27,59).
Por tanto, parece ser que las alteracionesconductuales observadas son principalmente debi-das a que la desnutrición modifica el estado emo-cional de los organismos.
90
Vol. 5/No. 2/Abril-Junio, 1994.
Desnutrición, aprendizaje y memoria.

EL EFECTO DE LA DESNUTRICION SO-BRE EL APRENDIZAJE Y LA MEMORIA.
En el estudio de los efectos de la desnutriciónen los procesos de aprendizaje y memoria, conti-nuamente se encuentran inconsistencias en los re-sultados. Se reportan ejecuciones inferiores, igua-les o inclusive mejores en las tareas desempeñadaspor los animales desnutridos. En esta sección ana-lizaremos diversos estudios acerca de este tema ydiscutiremos las posibles causas que han provoca-do estas inconsistencias.
APRENDIZAJE INSTRUMENTAL.
Se sabe que la desnutrición temprana puedeproducir secuelas conductuales tardías, que nologran compensarse a pesar de la rehabilitaciónnutricional posterior. Para indagar acerca de lascausas de estas secuelas, se ha empleado elcondicionamiento operante o instrumental y deéste el que involucra el reforzamiento a diferentesintervalos de tiempo y razón.
El condicionamiento operante (también lla-mado condicionamiento instrumental o aprendiza-je de ensayo y error), descubierto por E. Thorndikey estudiado de manera sistemática por B.F. Skinner,consiste en colocar una rata hambrienta en unacámara de prueba sonoamortiguada (a la que se leconoce como �caja de Skinner�) que tiene unapalanca emergiendo de una de las paredes (Figura1). La rata eventualmente tenderá a presionar lapalanca y obtendrá una recompensa apetitiva. Larata entonces aprenderá que una cierta respuestaes recompensada con comida (60). Posteriormen-te, a este dispositivo experimental se le hicierondiversas modificaciones, entre ellas tenemos a losreforzamientos intermitentes, los cuales son dedos clases, los de intervalo y los de razón. Encuanto a los reforzamientos de intervalo, éstospueden ser de tipo fijo o variable. El reforzamientode intervalo fijo se refiere que la recompensa oreforzador se proporciona a la primera respuestaque ocurra después de un intervalo designado
(fijo); por ejemplo, un minuto después de que larata apretó la palanca se le proporciona la recom-pensa. En el reforzamiento de intervalo variable elreforzador se le proporc iona a la ra taaleatoriamente en intervalos, desde uno muy bre-ve hasta uno muy largo, en un orden variable. Aligual que los intervalos, el reforzamiento de razónpuede ser fijo o variable. En el reforzamiento derazón fija el reforzador se proporciona después deun número predeterminado de respuestas, por ejemplodespués de que la rata presione 10 veces lapalanca se le proporciona la recompensa. En el derazón variable, la razón varía aleatoriamente depequeña a grande (61).
Halas y col. (37) mostraron que ratas des-nut r idas pos tna ta lmente probadas en uncondicionamiento operante apretaban un mayornúmero de veces la palanca que las controles yestos resultados fueron confirmados años más tar-
Figura 1.- Esquema que muestra una caja de Skinnerutilizada en los experimentos de aprendizaje instrumental.
91
M García-Ruiz, L Parra, S Díaz-Cintra.
Revista Biomédica

de por Tonkiss y col. (62). Este experimento mos-tró que las ratas desnutridas estaban más motiva-das para obtener la comida que las controles.Smart y Dobbing (38), explicaron que las ratasdesnutridas mostraban una alta motivación a lacomida debido a que la desnutrición modifica sumetabolismo basal provocando que se requieramás alimento. Por tanto, parece ser que la ejecu-ción de esta tarea es afectada por dos variablescríticas: la habilidad de aprendizaje y la motiva-ción.
Por otra parte, Stephens y Tonkiss (63)compararon la habilidad de ratas controles y des-nutridas postnatalmente (durante las primeras tressemanas de v ida) , para asociar es t ímulosredundantes con un castigo. Esto es, las ratastenían que aprender a suprimir la presión de unapalanca que les proporcionaba una recompensaapetitiva cuando estímulos medioambientales, ta-les como un tono y/o una luz, señalaban que lapresión de la palanca sería castigada con un cho-que eléctrico en las patas. No hubo diferenciaentre los grupos en el ritmo al cual aprendieron asuprimir la respuesta cuando una luz fue usadacomo señal de castigo, ni cuando un tono fueprobado al mismo tiempo con la luz. Pero cuandosubsecuentemente fue probado el tono solo, losdesnutridos fueron incapaces de asociarlo con elcastigo, por lo que los autores sugirieron que eltono podría ser un inhibidor condicionado -ya queel tono no estuvo pareado con el castigo- en losdesnutridos, pero no así en los controles, portanto parece ser que hay diferencias intrínsecas(e .g . : mot ivación, cur iosidad o fuerza delcondicionamiento) entre los dos grupos de anima-les en su habilidad para formar asociaciones entreestímulos. Resultados similares fueron reportadospor Stephens y col. (64).
Por esa misma época, Roberts y col. (65)propusieron que una estrategia que podría evitarlos problemas asociados con las diferenciasconductua les en té rminos de var iablesmotivacionales, era usar las técnicas estándar deanálisis experimental del comportamiento, por
ejemplo paradigmas de reforzamiento (65). En susexperimentos compararon ratas controles con ra-tas desnutridas postnatalente (desde el período delactancia hasta el día 43 posterior al destete), porreducción al 50% del alimento de las hembraslactantes y posteriormente a las crías destetadas.Una vez que los animales llegaron a la etapa adul-ta, fueron entrenados y probados en un paradigmaoperacional de reforzamiento de intervalos fijo yvariable. No hubo diferencias significativas entrecontroles y desnutridos durante el paradigma deintervalo fijo y en el caso del reforzamiento deintervalo variable, el grupo desnutrido respondiósignificativamente más rápido que los controles.Lo que permitiría pensar que la ocurrencia dediferencias entre controles y desnutridos duranteun condicionamiento operante depende del tipo deparadigma de recompensa que se emplee. Porejemplo, si cada vez que los animales presionan lapalanca reciben la recompensa será difícil encon-trar diferencias entre los grupos, pero si varía elintervalo entre la presión de la palanca y la apari-ción de la recompensa, de tal manera que el orzadorse haga más impredecible, las diferencias entrecontroles y desnutridos se hacen más grandes yevidentes. Estos autores concluyeron que ladesnutrición temprana no necesariamente produceincremento en las respuestas de los animales pro-bados en paradigmas que incluyen recompensasapetitivas.
Otro paradigma utilizado para analizar losefectos de la desnutrición temprana en el compor-tamiento de animales adultos, es el reforzamientodiferencial de tasas bajas de respuesta (RDB).Este es un paradigma operante en el cual la rata,debe esperar un intervalo específico entre res-puestas para presionar una palanca y en conse-cuencia adquirir el reforzamiento (comida). Unejemplo de ello es el trabajo de Tonkiss y col.(66). Estos investigadores probaron, en esteparadigma, ratas cuyas madres fueron desnutridas(con una dieta baja en caseína) desde 5 semanasantes del apareamiento hasta el término de la ges-tión y encontraron que el efecto de la desnutrición
92
Vol. 5/No. 2/Abril-Junio, 1994.
Desnutrición, aprendizaje y memoria.

93
se manifestó como una adquisición más lenta de latarea -estos resultados corroboran los experimen-tos de Laughlin y col. (30)-, por lo que sugirieronvarias hipótesis para explicar sus resultados: a) losanimales desnutridos prenatalmente podrían sermenos capaces de cronometrar sus respuestas ade-cuadamente, b) el desarrollo de la estrategia decronometraje podría haber sido alterado por ladesnutrición prenatal, c) los animales desnutridosse encontraban en un estado motivacional diferen-te que los controles, ya que se ha reportado que elnivel de privación tiene efecto en el RDB (67) y d)las ratas desnutridas estaban inicialmente más sen-sibles a la frustración provocada por la no-recom-pensa, ya que se ha mostrado (44) que las ratasdesnutridas muestran una resistencia mayor a laextinción. Esta resistencia a la extinción se obser-vó en una prueba de alternación espacial en unlaberinto en T con recompensa apetitiva, lo cualsugiere un decremento en la sensibilidad a la no-recompensa. Cualquiera que sea la explicaciónmás adecuada, lo que parece ser claro es que larehabilitación nutricional postnatal no es suficien-te para recompensar el daño prenatal.
Posteriormente, Tonkiss y col. (68) evalua-ron el efecto de la desnutrición prenatal en laextinción en un paradigma de reforzamiento par-cial, bajo la hipótesis de que el efecto de unreforzamiento parcial en la extinción (RPE) seríaatenuado o abolido en animales desnutridos. ElRPE se refiere a que las ratas entrenadas en unparadigma de reforzamiento parcial (con una re-compensa apetitiva) en una proporción de las prue-bas elegida al azar (típicamente 50%), son másresistentes a la extinción de la conducta, cuandola recompensa es presentada de manera completa-mente discontinua, que esas ratas entrenadas enun paradigma de reforzamiento continuo. Los re-sultados mostraron que el RPE no fue afectadopor la desnutrición prenatal, lo cual no es sorpren-dente ya que previamente se había observado (44)en animales desnutridos un incremento de resis-tencia a la extinción de la conducta en un paradig-ma de alternación espacial en un laberinto en T. Con
base en estos resultados estos investigadores pos-tulan que la desnutrición prenatal probablementeno interfiere con los procesos de extinción de laconducta per se en ratas maduras, sino más bienpuede causar pérdida de la flexibilidad en las es-trategias de respuesta cuando cambian las condi-ciones de prueba.
APRENDIZAJE DE DISCRIMINACION VI-SUAL.
Otra estrategia para determinar la capacidadde aprendizaje en animales desnutridos, la consti-tuyen las pruebas que exploran los procesosinvolucrados en resolver los problemas de la dis-criminación visual.
Rogers y col. (69) probaron, en ratas desnu-tridas postnatalmente (desde el nacimiento hastalos 45 días de edad), la habilidad para discriminarestímulos visuales (i.e., círculos y triángulos), alos que habían estado expuestos previamente, usan-do la versión no apetitiva de la técnica de Lashley.En esta prueba se coloca al animal en una platafor-ma frente a un pizarrón que contiene dos com-puertas, detrás de las cuales se encuentran losestímulos visuales y el sujeto experimental debeaprender a discriminar entre los estímulos a travésde un salto hacia la compuerta abierta que contie-ne el estímulo correcto. Los resultados mostraronque el aprendizaje de discriminación no fue dete-riorado por la desnutrición postnatal -estosresultados confirman experimentos anteriores(70,71)-. Sin embargo, sólo los controles mejora-ron la ejecución de la tarea de discriminación conla exposición previa, aparentemente esta exposi-ción no influyó en el aprendizaje de discriminaciónde los animales desnutridos. Estos resultados apo-yan la hipótesis de que los animales desnutridosparecen sufrir deficiencias en lo que Levitsky (72)llamó �aprendizaje incidental�, el cual se refiere aque los animales bien nutridos muestran �ham-bre� por aprender todo acerca de su medio am-biente, mientras que en los desnutridos el �ham-
M García-Ruiz, L Parra, S Díaz-Cintra.
Revista Biomédica

94
bre� o interés por aprender se ve inhibido. Losdesnutridos aparentemente responden sólo a esascaracterísticas medio ambientales que son de con-secuencia biológica inmediata para ellos.
Por otra parte, debido a que los efectos de laRPG pueden ser a largo plazo se ha sugerido quela desnutrición no disminuye el aprendizaje duran-te la exposición temprana, sino que afecta la ex-presión de este aprendizaje durante el entrena-miento de discriminación en la etapa adulta.
Por el contrario, Castro y Rudy (73) cuandocompararon animales normales y desnutridos (des-de el 2o. al 16o. postnatal) de diferentes edades(20-40 días y 70-77 días) en la ejecución de unatarea de discriminación de estímulos visuales deacuerdo a su posición, patrón y brillantez a travésde una versión acuática de la técnica de Lashley,encontraron deficiencias en la discriminación vi-sual. Estos investigadores observaron que la RPGdeterioró la ejecución en la tarea de discrimina-ción de brillantez (en crías de 20 y 40 días deedad) y en la discriminación de patrones (en ani-males de 40-70 días), mientras que en la discrimi-nación de la posición (i.e., izquierda vs. derecha),no encontraron diferencias significativas, por loque concluyeron que RPG no sólo tiene influenciaen la ejecución de la discriminación visual, sinoque puede dañar especialmente a los organismosque todavía están en desarrollo y permanecer eldaño hasta la etapa adulta.
Tonkiss y col. (74) encontraron resultadossimilares, empleando un paradigma de desnutricióndiferente (ver 66). Estos investigadores compara-ron animales normales y prenatalmente desnutri-dos en la ejecución de una tarea de discriminaciónde estímulos visuales en cuanto a la brillantez (i.e.,negro vs. blanco), a los patrones de orientación(i.e., barras horizontales vs. verticales) y de forma(i.e., círculos vs. cuadrados). Los animalesprenatalmente desnutridos de 91 días de edad,mostraron un número mayor de errores y latenciasmás largas en los tres problemas de discriminaciónvisual, aunque estos resultados no fueron signifi-cativos. Sin embargo, sólo en el caso de la discri-
minación de patrones de orientación los animalesdesnutridos mostraron deficiencias que sí alcanza-ron significación estadística. Los resultados dees te t rabajo indican que la desnut r ic iónhipoproteínica prenatal, deteriora el aprendizajede discriminación visual en animales adultos, loque estaría indicando efectos a largo plazo. Resul-tados similares fueron previamente reportados porCeledón y col. (75).
De los resultados de Castro y Rudy (73) ylos de Tonkiss y col. (74) emanan evidencias quemuestran que el efecto de la desnutrición tempra-na es selectivo, ya que influye sobre ciertas habili-dades, mientras que otras parecen no modificarse.
Lashley (76) propuso que el aprendizaje dediscriminación involucra 2 procesos: a) atención ala dimensión relevante del estímulo y b) asociacio-nes de los valores específicos de la dimensión conlos resultados de la elección correcta e incorrecta.Por tanto, los animales podrían fallar en los pro-blemas de discriminación por cualquiera de estosdos procesos. La propuesta de Lashley ofrece unaexplicación en cuanto a las deficiencias encontra-das en el aprendizaje de discriminación visual.Empero, a la fecha no hay un consenso con res-pecto a los efectos de la desnutrición en este tipode aprendizaje ya que, como hicimos notar previa-mente, existen estudios que no confirman que ladesnutrición pueda alterar el aprendizaje de discri-minación visual. Es necesario por tanto, enfatizarque los efectos de la desnutrición, tienen que serevaluados e interpretados según las condicionesbajo las cuales se lleven a cabo los experimentos.Esto es, deben de tomarse en cuenta diversosfactores que podrían intervenir en la ejecución delas tareas de aprendizaje, tales como la edad de losanimales, el método de desnutrición, el grado dedificultad de la tarea, etc..
APRENDIZAJE Y MEMORIA ESPACIAL.
Los intentos para identificar cambios fun-cionales permanentes en el aprendizaje y memoria
Vol. 5/No. 2/Abril-Junio, 1994.
Desnutrición, aprendizaje y memoria.

95
espacial que pueden reflejar cambios en la estruc-tura y función de determinadas regiones cerebra-les, han producido resultados disímbolos. Por ejem-plo, deficiencias en el aprendizaje de discrimina-ción espacial -en el cual el animal debe aprender adiscriminar entre un lugar en donde se encuentrala recompensa y otro vacío (e.g.: laberintos en T,Y o radiales, figura 2)-, que es un efecto consis-tente con la disfunción hipocampal, fueron encon-tradas por Barnes y col. (49), Simonson y Chow(77) y Hsueh y col. (78). Sin embargo, estosresultados no fueron confirmados en otros estu-dios (79,80).
Posteriormente, Jordan y col. (81) reporta-ron que ratas adultas desnutridas (a través de larestricción de alimento) desde la última semana degestación y durante el período de lactancia mos-traron deficiencias en la ejecución en el laberintoradial, (el cual ha sido vinculado a la función delhipocampal (82)) y fallaban en visitar de maneraalternada los brazos del laberinto (i.e., alternaciónespontánea) que es la respuesta típica de los bien
nutridos (83).Hall (84) por su parte, no encontró deficien-
cias en la ejecución del laberinto radial o laalternación espontánea en ratas cuyas madres fue-ron desnutridas por una dieta restringida en pro-teína, desde cinco semanas antes del apareamientohasta el período de lactancia. Parece ser que ladiscrepancia entre Hall y Jordan y col., se debe aque Hall sobre-entrenó a sus ratas y por esta razónHall podría haber reducido aspectos emocionales.Esto sugeriría que el sobre-entrenamiento podríareducir las deficiencias conductuales producidaspor la desnutrición. Sin embargo, existen otrasposibilidades, la primera de ellas se refiere a losdiferentes protocolos de dieta empleados en am-bos trabajos, ya que Jordan y col., utilizaron unmétodo de desnutrición pluricarencial y Hall sola-mente restringió la cantidad de proteína. La se-gunda, es que la desnutrición fue impuesta enperíodos diferentes, como fue mencionado. Jordany col. privaron a sus animales desde la últimasemana de gestación hasta la lactancia, mientrasque Hall impuso la dieta hipoproteínica a las hem-bras desde cinco semanas antes del apareamientohasta la lactancia.
Por otra parte y con el objetivo de evaluar elefecto de la experiencia previa en el aprendizaje deanimales desnutridos, Levitsky (85) comparó ungrupo control contra un grupo de animales desnu-tridos durante la lactancia y posdestete temprano(con una dieta baja en proteína), en el campoabierto una vez y, semanas después de la rehabili-tación nutricional, expuso a ambos grupos a unlaberinto con recompensa de comida. Encontróque el grupo desnutrido cometía más errores en ellaberinto que el grupo control, por lo que conclu-yó que el efecto de la experiencia -adquirida en elcampo abierto- en el aprendizaje tardío es blo-queado por la desnutrición.
En 1981 Morr is (86) in t rodujo unametodología (el ahora llamado laberinto de Morris,Figura 3) para estudiar los procesos de aprendiza-je y la memoria espacial a través de dos formas denavegación espacial. En ambas formas la rata es
Figura 2.- Ejemplo de un laberinto radial de 8 brazos. Larata es colocada en el centro para que se desplace a travésde los brazos del laberinto en busca de una recompensaapetitiva.
M García-Ruiz, L Parra, S Díaz-Cintra.
Revista Biomédica

96
colocada en un tanque circular con agua, el cualdebe nadar hacia una plataforma de escape. En laprimera de ellas, llamada de �señal-proximal�, laplataforma esta visible y la rata debe nadar direc-tamente hacia ella, la señal proximal. En la segun-da, llamada de �señal-distal�, la plataforma estáoculta bajo la superficie del agua, pero siempreestá colocada en una posición fija relativa a seña-les externas al laberinto, por lo que la rata debeaprender a utilizar las señales distales del medioambiente para navegar hacia la plataforma de es-cape.
Goodlett y col. (87) analizaron los efectosde la desnutrición hipoproteínica en dos tareas,una tarea de alternación espacial recompensada(i.e., los animales tenían que visitar de maneraalternada cada brazo del laberinto en T para obte-ner la recompensa) y otra de navegación en ellaberinto de Morris de señal-distal. Sus ratascrónicamente malnutridas cometieron más erroresque las controles en la tarea de alternación espa-cial. Tonkiss y Galler (44) observaron resultados
similares , aunque en animales desnutr idosprenatalmente. Por el contrario, en el laberinto deMorris, observaron que los sujetos desnutridosaprendieron a usar las señales distales tan biencomo los controles. Por lo que concluyeron que elprocesamiento complejo de información espaciales deteriorado por la desnutrición, mientras que lalocalización y el mapeo espacial per se no sonafectados por tal alteración. Además, la falta dedeterioro en la localización espacial en el laberin-to de Morris podría deberse a que -como se ha re-portado (72,88,89)- las ratas con antecedentes dedesnutrición a menudo no tienen dificultades conproblemas simples de aprendizaje, sino que losefectos de esta alteración son fácilmente detectablescuando las tareas de evaluación son más comple-jas.
Resultados similares, en cuanto al laberintode Morris, fueron encontrados por Campbell yBedi (90) y Bedi(91) los cuales sugieren que elhecho de no haber encontrado diferencias entre lasratas controles y las desnutridas podría indicar queel cerebro de la rata tiene un grado considerablede �capacidad disponible�. Esto es, que puedafuncionar de manera aparentemente normal aúncon un daño substancial y alteraciones morfológicasde varias regiones cerebrales, o bien que ladesnutrición temprana no afecta permanentemen-te el comportamiento de aprendizaje espacial de laratas.
Por su parte, Castro y Rudy (92) trataron deexplotar otra alternativa que pudiese ser la causade las incons is tencias en los resul tadosconductuales de animales desnutridos. Estos in-vestigadores notaron que los efectos conductualesde la desnutrición temprana se evaluaban en ani-males adultos, por lo que decidieron probar laposibilidad de que la desnutrición temprana pudie-se tener efectos mayores en el comportamiento delos animales que todavía estaban en desarrollo. Elmétodo de desnutrición que emplearon fue el deseparación de la camada durante los primeros 16días de vida y, a través del laberinto de Morris, ensus dos modalidades, probaron a sus animales a
Figura 3.- Representación esquemática del laberinto acuá-tico de Morris. La línea ondulada muestra un ejemplo dela trayectoria que puede seguir una rata para encontrar laplataforma de escape.
Vol. 5/No. 2/Abril-Junio, 1994.
Desnutrición, aprendizaje y memoria.

97
los 19, 22, 24, y 30 días de edad. Estos autoresencontraron que las ratas desnutridas, desde los19 días de edad eran capaces de localizar la plata-forma de escape en el laberinto de señal-proximal.Con respecto a los animales de 27 días probadosen el laberinto de señal-distal, observaron que lasratas desnutridas no aprendieron a utilizar las se-ñales distales para localizar la plataforma oculta.A los 30 días de edad, no hubo diferencias entrelos controles y desnutridos con respecto al uso deseñales distales para localizar la plataforma. Sinembargo, las latencias de escape fueron más largasen los animales desnutridos de esta edad que enlos controles. Estos experimentos mostraron quela desnutrición temprana no produce deficienciasen el aprendizaje con señal proximal, pero sí dete-riora la ejecución de la navegación en el paradigmade señal distal. Los resultados de estos autoressugieren que los efectos de la desnutrición tem-prana son dependientes de la edad y de la tarea deaprendizaje.
El efecto de la desnutrición en el aprendiza-je y la memoria espacial no solamente ha suscitadocontroversias en cuanto a si se producen deficien-cias o no se producen, sino también, en cuanto aun mejoramiento en la ejecución de tareas espa-ciales. Un ejemplo de ello es el trabajo de Idrobo ycol. (93) en el cual fue mostrado que ratonesdesnutridos postnatalmente desde los tres hastalos quince meses, tuvieron un mayor número derespuestas correctas en el paradigma del laberintoradial que los controles, por tanto parecería quelos animales desnutridos aprenden mejor la tareaque los controles, lo cual ya había sido reportadoaños antes por Laughlin y col. (30).
Sin embargo, es necesario hacer notar quelos paradigmas conductuales que involucran unarecompensa apetitiva (i.e., los laberintos en Y, T,radiales y algunos condicionamientos operantes),presentan problemas de interpretación, ya quecomo mostraron Halas y col. (37) y Tonkiss y col.(94) los animales desnutridos presentan una ma-yor motivación a la recompensa apetitiva que loscontroles, por tanto no es claro si los cambios en
el aprendizaje que se observan en este tipo dedispositivos son debidos a la desnutrición o a quelos animales están en un estado motivacional dis-tinto.
APRENDIZAJE DE EVITACION A ESTIMULOSNOCIVOS.
Como una alternativa a los paradigmas queinvolucran recompensas apetitivas se ha recurridoa paradigmas de aprendizaje con consecuenciasaversivas (e.g.: castigos). Por ejemplo, que el su-jeto aprenda a evitar un evento desagradable (e.g.:un choque eléctrico) inhibiendo (e.g.: la preven-ción pasiva, figura 4) o alternando su movimiento(e.g.: prevención activa), o bien que el animalasocie cierto tipo de estímulo (e.g.: un tono o unaluz) con la presentación de un castigo.
En una situación no apetitiva Barnes y col.(49) notaron que a cerdos con desnutrición tem-prana (con dietas bajas en proteína) les tomaba
Figura 4.- Esquema que muestra el dispositivo experi-mental más común, utilizado para la prevención pasiva.En este tipo de dispositivo la rata debe aprender a evitarun estímulo nocivo a través de la inhibición de su pasohacia el comportamiento oscuro (lado derecho del esquema).
M García-Ruiz, L Parra, S Díaz-Cintra.
Revista Biomédica

98
más tiempo extinguir una respuesta de evitacióncondicionada a un estímulo nocivo que a los orga-nismos controles, aunque no encontraron diferen-cias en el ritmo de adquisición de la respuesta (i.e.,en el ritmo de evitación del estímulo nocivo).Resultados similares encontraron Franková yBarnes (41) en ratas. Sin embargo, estos autoresenfatizaron que aunque no hubo diferencia entrecontroles y desnutridos en el ritmo de aprendizaje(expresado en términos de latencias), las ratasdesnutridas mostraron grandes al teracionesconductuales, tales como ansiedad y excitaciónexageradas. Por esta misma época Guthrie (15)reportó que las ratas desnutridas durante la lactanciay posdestete temprano, eran tan capaces como lascontroles de asociar un estímulo auditivo paraevitar un choque eléctrico.
Posteriormente, Levitsky y Barnes (42) com-pararon las respuestas de ratas adultas desnutridascon una dieta baja en caseína (impuesta durante lalactancia y 4 semanas después del destete), con lasrespuestas de ratas bien alimentadas, en losparadigmas de prevención pasiva y activa y obser-varon que las ratas desnutridas mostraban unarespuesta emocional más intensa en la evitación alchoque, i.e., permanecían más tiempo en la plata-forma después de haber recibido el choque eléctri-co que las ratas controles. En el caso de la preven-ción activa observaron que las ratas desnutridasmostraban una respuesta más rápida y más prolon-gada que las ratas normales. Estos autores conclu-yeron que la desnutrición temprana produce efec-tos a largo plazo disminuyendo el umbral de res-puesta. Franková (16), observó resultados simila-res aunque en sus experimentos las ratas desnutri-das mostraron aprender mejor que las controles.El mejoramiento en el aprendizaje, previamentefue reportado por Smart y Dobbing (95), por loque sugerimos que, además de la desnutrición,podría haber otros factores de tipo metodológico,como el grado y duración del per íodo dedesnutrición, la motivación, la edad de los anima-les, la manipulación temprana, etc. que estuvieseninterfiriendo para producir las diferentes respues-
tas de los animales.Por su parte, Smart y Dobbing (96) no en-
contraron alteraciones en la ejecución de una tareade prevención pasiva en ratas desnutridas (porreducción al 50% de la cantidad de alimento)durante la gestación y la lactancia. Sin embargo,años más tarde fue reportado (39,53,58) que lasratas previamente desnutridas mostraban umbra-les de alerta más bajos que las controles, lo cualpodría explicar el incremento en las respuestas delas ratas desnutridas ante un estímulo aversivo.
Con un tipo de desnutrición diferente, en elcual ratones fueron desnutridos a partir del sépti-mo día de la gestación hasta el destete, desde elnacimiento hasta el destete o ambos, Bush yLeathwood (29) observaron que todos los ratonesdesnutridos probados en un paradigma de evitación(i.e., que incluía un tono y una luz que anunciabanel choque eléctrico), lograron un número menorde evitaciones exitosas que los controles y, másaún que la manipulación temprana no tuvo ningúnefecto en el comportamiento de evitación.Adicionalmente, notaron que la desnutrición pre-natal tenía un mayor efecto que la postnatal. Estosresultados fueron confirmados por Tonjes y col.(98).
Posteriormente, fue sugerido que las dife-rencias en el comportamiento de ratas desnutridasy bien nutridas podrían deberse a las alteracionesen las interacciones maternales y sociales más quea la desnutrición per se (99), por lo que se llevarona cabo experimentos para probar los efectos de ladesnutrición y la privación maternal como facto-res que interfiriesen en la ejecución de una tareade prevención pasiva (43). En cuanto a las altera-ciones en el comportamiento se encontraron re-sultados similares a los previamente reportados(39,42,58). En cuanto a las interacciones materna-les se llegó a la conclusión de que la desnutrición ylos efectos de ésta en el crecimiento corporal ycerebral, son los factores primordiales que afectanel comportamiento tardío de las ratas previamentedesnutridas, más que las alteraciones madre-infan-te asociadas con un régimen nutricional (43).
Vol. 5/No. 2/Abril-Junio, 1994.
Desnutrición, aprendizaje y memoria.
99
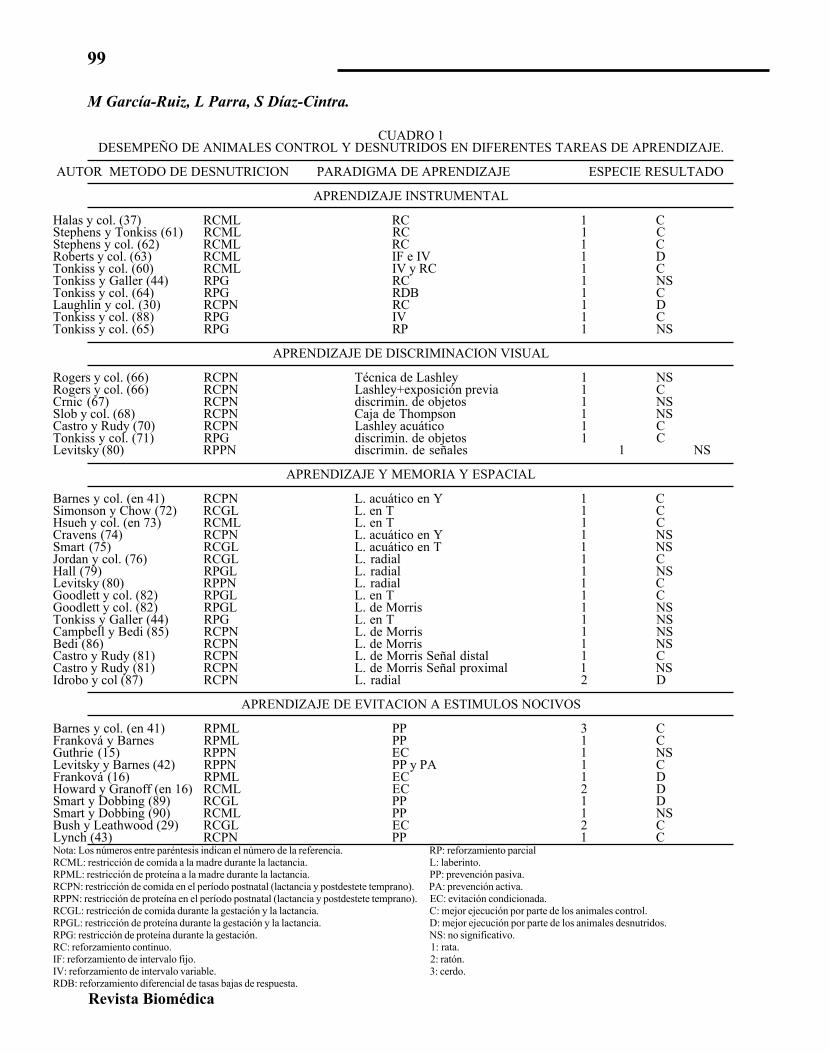
CUADRO 1DESEMPEÑO DE ANIMALES CONTROL Y DESNUTRIDOS EN DIFERENTES TAREAS DE APRENDIZAJE.
AUTOR METODO DE DESNUTRICION PARADIGMA DE APRENDIZAJE ESPECIE RESULTADO
APRENDIZAJE INSTRUMENTAL
Halas y col. (37) RCML RC 1 CStephens y Tonkiss (61) RCML RC 1 CStephens y col. (62) RCML RC 1 CRoberts y col. (63) RCML IF e IV 1 DTonkiss y col. (60) RCML IV y RC 1 CTonkiss y Galler (44) RPG RC 1 NSTonkiss y col. (64) RPG RDB 1 CLaughlin y col. (30) RCPN RC 1 DTonkiss y col. (88) RPG IV 1 CTonkiss y col. (65) RPG RP 1 NS
APRENDIZAJE DE DISCRIMINACION VISUAL
Rogers y col. (66) RCPN Técnica de Lashley 1 NSRogers y col. (66) RCPN Lashley+exposición previa 1 CCrnic (67) RCPN discrimin. de objetos 1 NSSlob y col. (68) RCPN Caja de Thompson 1 NSCastro y Rudy (70) RCPN Lashley acuático 1 CTonkiss y col. (71) RPG discrimin. de objetos 1 CLevitsky (80) RPPN discrimin. de señales 1 NS
APRENDIZAJE Y MEMORIA Y ESPACIAL
Barnes y col. (en 41) RCPN L. acuático en Y 1 CSimonson y Chow (72) RCGL L. en T 1 CHsueh y col. (en 73) RCML L. en T 1 CCravens (74) RCPN L. acuático en Y 1 NSSmart (75) RCGL L. acuático en T 1 NSJordan y col. (76) RCGL L. radial 1 CHall (79) RPGL L. radial 1 NSLevitsky (80) RPPN L. radial 1 CGoodlett y col. (82) RPGL L. en T 1 CGoodlett y col. (82) RPGL L. de Morris 1 NSTonkiss y Galler (44) RPG L. en T 1 NSCampbell y Bedi (85) RCPN L. de Morris 1 NSBedi (86) RCPN L. de Morris 1 NSCastro y Rudy (81) RCPN L. de Morris Señal distal 1 CCastro y Rudy (81) RCPN L. de Morris Señal proximal 1 NSIdrobo y col (87) RCPN L. radial 2 D
APRENDIZAJE DE EVITACION A ESTIMULOS NOCIVOS
Barnes y col. (en 41) RPML PP 3 CFranková y Barnes RPML PP 1 CGuthrie (15) RPPN EC 1 NSLevitsky y Barnes (42) RPPN PP y PA 1 CFranková (16) RPML EC 1 DHoward y Granoff (en 16) RCML EC 2 DSmart y Dobbing (89) RCGL PP 1 DSmart y Dobbing (90) RCML PP 1 NSBush y Leathwood (29) RCGL EC 2 CLynch (43) RCPN PP 1 CNota: Los números entre paréntesis indican el número de la referencia. RP: reforzamiento parcialRCML: restricción de comida a la madre durante la lactancia. L: laberinto.RPML: restricción de proteína a la madre durante la lactancia. PP: prevención pasiva.RCPN: restricción de comida en el período postnatal (lactancia y postdestete temprano). PA: prevención activa.RPPN: restricción de proteína en el período postnatal (lactancia y postdestete temprano). EC: evitación condicionada.RCGL: restricción de comida durante la gestación y la lactancia. C: mejor ejecución por parte de los animales control.RPGL: restricción de proteína durante la gestación y la lactancia. D: mejor ejecución por parte de los animales desnutridos.RPG: restricción de proteína durante la gestación. NS: no significativo.RC: reforzamiento continuo. 1: rata.IF: reforzamiento de intervalo fijo. 2: ratón.IV: reforzamiento de intervalo variable. 3: cerdo.RDB: reforzamiento diferencial de tasas bajas de respuesta.
M García-Ruiz, L Parra, S Díaz-Cintra.
Revista Biomédica

100
paradigmas que involucran una recompensaapetitiva, como es el caso de algunos laberintos ydel condicionamiento operante, es el hecho de quepara que el animal lleve a cabo la conducta se ledebe de privar de alimento horas antes de la prue-ba, por tanto de los resultados emanados de estaspruebas se pueden originar conclusiones inade-cuadas, por lo que no queda claro si el cambioconductual observado se debe a la desnutrición, ala motivación o a la privación de alimento. Detodos estos factores, la motivación parece ser laque más influye, ya que se ha reportado que ladesnutrición produce diversas alteraciones fisioló-gicas que persisten aún después del período en elque fue impuesta. Estas alteraciones incluyen cam-bios en la eficiencia de la asimilación del alimento(78) y cambios en los niveles de glucosa en sangre(100). Parece ser que estos cambios fisiológicosson la base de la gran motivación a la comida quemuestran los animales desnutridos.
Por otra parte, si no son los nutrientes losque se usan como reforzadores son los estímulosnocivos. Con este tipo de reforzadores, también sehan encontrado inconsistencias en los resultados.Algunos autores reportan deficiencias en laevitación a los estímulos nocivos, mientras queotros o no encuentran diferencias o reportan unnúmero mayor de evitaciones exitosas por partede los animales desnutridos. Parece ser, que labase de estas inconsistencias es la diferencia en elgrado de emocionalidad entre animales normalesy desnutridos, ya que el común denominador detodos estos trabajos, como mencionamos en pá-rrafos anteriores, es que los animales desnutridospresentan una alta emocionalidad y exhiben unarespuesta exagerada, probablemente debido a quelos umbrales para provocar un estado de alertason más bajos que en los animales bien nutridos(39,58,72).
Otro elemento que influye en la ocurrenciao no de diferencias entre controles y desnutridoses el grado de dificultad de la tarea del aprendizajeque se utilice. Por ejemplo, hay datos que mues-tran que el sobre-entrenamiento reduce la alta
Aunque, no queda claro si la desnutriciónproduce deficiencias en la habilidad de aprendizajede evitación a estímulos nocivos, es posible con-cluir que la desnutrición por una parte, disminuyeel umbral de respuesta de los animales y por otraproduce anormalidades conductuales tales comola ocurrencia de comportamientos estereotipados(e.g.: un acicalamiento exagerado) que en los ani-males bien nutridos no se presentan cuando sonexpuestos a pruebas de aprendizaje de este tipo.
En el cuadro 1, se resumen los resultadosobtenidos por los grupos de investigación -anali-zados en el presente trabajo-, sobre el efecto delos diferentes procedimientos de desnutrición enla ejecución de diversas tareas de aprendizaje.
Vol. 5/No. 2/Abril-Junio, 1994.
DISCUSION.
A lo largo de esta revisión hemos tratado deanalizar los efectos que produce la mala nutrición,no sólo en ciertos patrones conductuales, sinotambién en los procesos involucrados en el apren-dizaje. Con respecto al primer punto parece haberun consenso en cuanto a las alteraciones que ladesnutrición provoca, ya que los trabajos reporta-dos por diversos grupos de investigación conver-gen en los mismos puntos. Por ejemplo en que ladesnutrición produce un retraso en la aparición delas primeras conductas, alteraciones en el compor-tamiento social, menor exploración y sobretodoalgo que consistentemente se ha observado es unarespuesta emocional exagerada. Empero, en cuan-to a las alteraciones que produce la desnutriciónen los procesos de aprendizaje y memoria, losresul tados obtenidos has ta ahora soncontroversiales.
Sin embargo, hay muchas variables que po-drían afectar la ejecución que no tienen nada quever con el aprendizaje. Este aspecto es particular-mente serio, ya que en muchos trabajos se usannutrientes como reforzadores para la ejecución dela tarea de aprendizaje. El problema de los
Desnutrición, aprendizaje y memoria.

101
emocionalidad que caracteriza a los desnutridos(84), por tanto se podría esperar que las deficien-cias conductuales producidas por la mala nutri-ción se redujeran si se sobreentrena a los animales.Más aún, se tienen datos que sugieren que losorganismos desnutridos no tienen dificultades conproblemas simples de aprendizaje (e.g.: asociacio-nes entre estímulos), sino los efectos de ladesnutrición son fácilmente detectables como lastareas de evaluación son más complejas (72,88,89).Es posible que los parámetros de evaluación de lacapacidad para resolver problemas simples deaprendizaje de algunos paradigmas, no sean lasmedidas adecuadas de una deficiencia en el desa-rrollo cognitivo (85).
Hasta ahora, hemos mencionado algunascondiciones bajo las cuales se llevan a cabo losexperimentos para analizar los posibles efectos dela desnutrición en los procesos de aprendizaje ymemoria. Sin embargo, existen otros factores quepueden enmascarar estos efectos, entre los másimportantes tenemos a la edad, el sexo, el períodoen el que se instala la desnutrición y el grado deseveridad de la desnutrición.
Es importante que se tome en cuenta la edadde los animales, ya que se ha observado que ladesnutrición temprana tiene efectos más drásticosen los organismos en desarrollo que en los adul-tos, aunque en algunos casos el daño puede per-manecer hasta la etapa adulta (73,89). A esterespecto se ha sugerido que un efecto de ladesnutrición durante la vida temprana es deterio-rar la habilidad para aprender aspectos del medioambiente que no están inmediatamente relaciona-dos al desempeño de la tarea de ese momento(72). Con respecto al sexo, la mayoría de losexperimentos se han llevado a cabo en machos,pero los pocos experimentos que se han realizadoen hembras muestran que los efectos de ladesnutrición parecen ser menos severos o quizásea más difícil evaluar los resultados en las hem-bras, ya que en ellas el ciclo estral puede provocarvariaciones conductuales (101).
En cuanto al período de instalación de la
desnutrición y al grado de severidad de ésta, hayevidencias que muestran que la intensidad del dañodepende, de la etapa de la vida del animal en quese presente la desnutrición. Se sabe que existe unperíodo crítico en el desarrollo del SNC (i.e.,período de crecimiento rápido) que para el hom-bre sería el último trimestre de la gestación y losdos primeros años de vida y para la rata la últimasemana de la gestación y todo el período delactancia. Si la desnutrición se instala durante esteperíodo crítico, los daños podrían ser irreversibles(102,103) y desde luego, las alteraciones en losprocesos de aprendizaje serían más evidentes. In-clusive, se ha postulado que el mejoramiento en elaprendizaje por parte de los animales desnutridos,se debe a que la desnutrición fue instalada despuésdel período crítico. Un ejemplo de ello es el traba-jo de Idrobo y col. (93), en el cual los animalesfueron desnutridos postnatalmente desde el 3o. al15o. mes, cuando el período crítico ya había pasa-do.
Por otra parte, Katz (104) y Smart (101) sedieron a la tarea de hacer una evaluación de lostrabajos reportados hasta esas fechas sobre la in-fluencia de la desnutrición en el aprendizaje. Estosautores encontraron que en aproximadamente enel 60% de los experimentos no se reportaban dife-rencias significativas entre controles y desnutri-dos, en el 30% de los casos animales bien nutridosdesempeñaban mejor la tarea que los desnutridosy en el 10% restante los desnutridos tenían mejo-res ejecuciones que los controles. En la actualidadestos porcentajes han variado un poco. Con res-pecto a los trabajos que se mencionan en estarevisión, notamos que aproximadamente el 37%de los reportes no muestran diferencias significati-vas entre controles y desnutridos, el 49% favorecea los controles y el 14% muestran una mejorejecución por parte de los desnutridos. Como esposible notar, a la fecha se han incrementado losestudios que apoyan la idea de que los animalesbien nutridos se desempeñan mejor que los desnu-tridos, ha disminuido el número de estudios queno muestran diferencias significativas y han au-
M García-Ruiz, L Parra, S Díaz-Cintra.
Revista Biomédica

102
mentado muy ligeramente los trabajos en los quese favorece a los desnutridos. Probablemente, es-tos cambios desde 1984 a la fecha se deban a quese han empleado mejores diseños experimentales ydesde luego a que se ha tratado de controlar elmayor número de variables posibles que pudiesenconfundir los resultados.
Los estudios más recientes parecen apoyarla idea de que la desnutrición puede alterar lahabilidad de aprendizaje, sin embargo se debe te-ner cuidado en el momento de interpretar los re-sultados, por lo cual queremos finalizar enfatizandodos puntos que consideramos de gran importan-cia; en primer lugar que el agua o el alimento sonreforzadores inapropiados en los estudios de apren-dizaje en animales privados nutricionalmente y ensegundo, que se deben de tomar en cuenta losdiversos factores que pueden influir en el desem-peño de las tareas de aprendizaje de los animalesdesnutridos (e.g.: la motivación, el período de ladesnutrición, la edad de los sujetos, el grado dedificultad de la tarea, etc.) antes de llegar a unaconclusión definitiva sobre los efectos de ladesnutrición en los procesos de aprendizaje y me-moria.
AGRADECIMIENTOS.
Los autores expresan su agradecimiento alas siguientes personas: Dr. Manuel Salas Alvaradoy Dra. Carolina Escobar Briones, por sus valiosose inapreciables comentarios a este trabajo y, alapoyo proporcionado por la DGAPA, No. de Pro-yecto IN204892.
REFERENCIAS.
1.-Morgane PJ, Austin-LaFrance R, Bronzino J, ycol. Prenatal malnutrition and development of thebrain. Neurosci Biobehav Rev 1993; 17:91-128.
2.-Barnes RH. Reflections on the study ofmalnutrition and mental development. En: LevitskyDa Ed. Malnutrition, environment, and behavior.Cornell University Press Ltd, 1979:19-27.
3.-Galler JR, Ramsey F, Solimano G. The influenceof early malnutrition on subsequent behavioraldevelopment III. Learning disabilities as a sequelto malnutrition. Pediatric Res 1984; 18:309-313.
4.-Galler JR, Ramsey F, Forde V, Salt P, ArcherE. Long-term effects of early kwashiorkorcompared wi th marasmus. I I . In te lec tua lperformance. J Pediatric Gastroenterol Nutr 1987;6:847-854.
5.-Galler JR, Ramsey F, Salt P, Archer E. Long-term effects of early kwashiorkor compared withmarasmus. III. Fine motor skills. J PediatricGastroenterol Nutr 1987; 6:855-859.
6.-Winick M. Nutrition and brain development.En: Serban G ed. Nutrition and mental functions.New York: Plenum Press, 1975:65-73.
7 . -Dobbing J , Sands J . Vulnerabi l i ty ofdevelopment brain IX. The effect of nutritionalgrowth retardation on the timing of the braingrowth-spurt. Biol Neonate 1971; 19:363-378.
8.-West CD, Kemper T. The effect of low proteindiet on the anatomical development of the ratbrain. Brain Res 1976; 107:221-237.
9.-Cintra L, Díaz-Cintra S, Galván A, Kemper T,Norgane PJ. Effects of protein undernutrition onthe dentate gyrus in rats of three age groups. BrainRes 1990; 532:271-277.
10.-Díaz-Cintra S, Cintra L, Galván A, Kemper T,Norgane PJ. Effects of prenatal protein deprivationon the postnatal development of granule cells inthe fascia dentata. J Comp Neurol 1991; 310:356-364.
Vol. 5/No. 2/Abril-Junio, 1994.
Desnutrición, aprendizaje y memoria.

103
11.-García-Ruiz M, Díaz-Cintra S, Cintra L,Corkidi G. Effect of protein malnutrition on CA3hippocampal pyramidal cells in rats of three ages.Brain Res 1993; 625:203-212.
12.-Lewis P, Patel A, Balázs R. Effect ofundernutrition on cell generation in the adult ratbrain. Brain Res 1979; 168:186-189.
13.-De Oliveira LM, Almeida SS, Lima JG,Rocinholi LF. Environmental stimulation andanxiolytic drugs: Differential effects in earlymalnour ished animals . En: Cint ra L. ed .International workshop on malnutrition and thedeveloping central nervous system: Sociedad Mexi-cana de Ciencias Fisiológicas A.C. y UniversidadAmericana de Acapulco, 1993:15.
14.-Davison AN, Dobbing J. The developing brain.En: Davison AN, Dobbing J eds . Appliedneurochemistry. Philadelphia: FA Davis Co,1968:253-286.
15.-Guthie HA. Severe undernutrition in earlyinfancy and behavior in rahabilitated albino rats.Physiol Behav 1968; 3:619-623.
16.-Franková S. Influence of the familiarity whatthe environment and early malnutrition on theavoidance learning and behavior in rats. Act NervSup 1973; 15:207-216.
17.-Cowley JJ, Griesel RD. The effect on growthand behaviour of rahabilitating first and secondgeneration low protein rats. Anim Behav 1966;14:506-517.
18.-Simonson M, Sherwin RW, Anilane JK, YuWY, Chow BF. Neuromotor development inprogeny of inderfed mother rats. J Nutr 1969;98:18-24.
19.-Samrt JL, Dobbing J . Vulnerabil i ty ofdeveloping brain: II. Effects of early nutritional
deprivation on reflex ontogeny and developmentof behaviour in the rat. Brain Res 1971; 28:85-95.
20.-Massaro TF, Levitsky D, Barnes RH. Earlyprotein malnutrition in the rat: Behavioral changesduring rehabilitation. Dev Psychobiol 1977;10:105-111.
21.-Fleischer SF, Turkewitz G. Effect of neonatalstunting on development of rats: Large litterrearing. Dev Psychobiol 1979; 12:137-149.
22.-Fleischer SF, Turkewitz G. Behavioral effectsof rotation between lactating and nonlactatingfemales. Dev Psychobiol 1979; 12:245-254.
23.-Fleischer SF, Turkewitz G, Finklestein H.Sensory influences on homing of stunted rap pups.Dev Psychobiol 1981; 14:29-39.
24.-Galler JR. Home orientation in nursling rats:The ef fec ts of rahabi l i ta t ion fo l lowingintergenerational malnutrition. Dev Psychobiol1979; 12:499-508.
25.-Galler JR. Home-orienting behavior in rat pupssurviv ing pos tna ta l of in tergenera t ionalmalnutrition. Dev Psychobiol 1980; 13:563-572.
26.Galler JR, Seelig C. Home-orienting behaviorin rat pups: The effect of 2 and 3 generations ofrahabi l i ta t ion fo l lowing in tergenera t ionalmalnutrition. Dev Psychobiol 1981; 14:541-548.
27. -Franková S . Inf luence of ear ly socia lenvironment on behavioural development and onlater maternal behaviour of protein deprived rats.Activ Ner Sup 1981; 23:81-91.
28.-Hunt L, Smotherman W, Wiener S y Levine S.Nutritional variables and their effects on thedevelopment of ultrasonic vocalizations in rat pups.Physiol Behav 1976; 17:1037-1039.
M García-Ruiz, L Parra, S Díaz-Cintra.
Revista Biomédica

104
29.-Bush M, Leathwood PD. Effects of differentregimens of early malnutrition on behavioraldevelopment and avoidance learning in Swiss whitmice. Br J Nutr 1975; 33:373-385.
30.-Laughlin NK, Sranley F, Bell J . Earlyundernutrition and later hippocampal damage:effect of spontaneous behaviors and reversallearning. Physiol Psychol 1984; 11:268-277.
31.-Altman J, Sudarshan K, Das G, McCormickN, Barnes D. The influence of nutrition on neuraland behavioral development: II. Development ofsome motor, particularly locomotor patterns duringinfancy. Dev Psychobiol 1971; 4:97-114.
32.-Salas M, Cintra L. Behavioral effects ofundernutrition on the rat. Bol Estud Med BiolMex 1973; 28:23-27.
33.-Chase HP, Lindsley WFB, O�Brien D.Undernutrition and cerebellar development. Nature1969; 221:554-555.
34. -Cul ley WF, Lineberger RD. Effec t ofundernutrition on the size and composition of therat brain. J Nutr 1968; 96:375-381.
35.-Wallingford JC, Shrader RE, Zeman FJ. Effectof maternal protein-calorie malnutrition on fetalcerebellar neurogenesis. J Nutr 1980; 110:543-551.
36.-Salas M, Pulido S, Torrero C, Escobar C.Neonatal undernutr i t ion and self-groomingdevelopment in the rat: long-term effects. PhysiolBehav 1991; 50:567-572.
37.-Halas ES, Burger PA, Sandstead HH. Foodmotivation of rehabilitated malnourished rats:Implications for learning studies. Anim LearnBehav 1980; 8:152-158.
38.-Smart JL, Dobbing J. Increased thirst and
hunger in adult rats undernourished as infants: Analternative explanation. Br J Nutr 1977; 37:421-429.
39.-Smart JL, Dobbing J, Adlard BPF, Lynch A,Sanda J. Vulnerability of developing brain: Relativeeffects of growth restriction during the fetaland suckling periods on behavior and braincomposition of adult rats. J Nutr 1973; 103:1327-1338.
40.-Escobar C, Pulido S, Caldelas I. Earlyolfactory response in neonatally undernourishedrats. En: Cintra L, ed International workshop onmalnutrition and the developing central nervoussystem. Sociedad Mexicana de Ciencias Fisiológi-cas A.C. y Universidad Americana de Acapulco,1993:13-14.
41.-Franková S, Barnes RH. Effect of malnutritionin early life on avoidance conditioning and behaviorof adult rats. J Nutr 1968; 96:485-493.
42.-Levitsky DA, Barnes RH. Effect of earlymalnutrition on the reaction of adult rats toaversive stimuli. Nature 1970; 225:468-469.
43.-Lynch A. Passive avoidance behavior andresponse thresholds in adult male rats after earlypostnatal undernutrition. Physiol Behav 1976;16:27-32.
44.-Tonkiss J , Gal ler JR. Prenatal proteinmalnutrition and working memory performance inadult rats. Behav Brain Res 1990; 40:95-107.
45.-Salas M, Torrero C. Maternal behavior of ratsundernourished in the early postnatal period. BolEstud Med Biol Mex 1979; 30:237-244.
46.-Wiener SG, Fi tzpatr ick KM, Levin R,Smotherman Wp, Levin S. Alterations in the ma-ternal behavior of rats rearing malnourishedoffspring. Dev Psychobiol 1977; 10:243-254.
Vol. 5/No. 2/Abril-Junio, 1994.
Desnutrición, aprendizaje y memoria.

105
47.-Whatson TS, Smart JL. Social behaviour ofra ts fo l lowing pre and ear ly pos tna ta lundernutrition. Physiol Behav 1978; 20:749-753.
48.-Escobar C, Salas M. Ameliorating effects ofearly sensory stimulation on the behavior of adultrats underfed during the lactating period. Bol EstudMed Biol Mex 1987; 35:195-202.
49.-Barnes RH, Neely CS, Kwong E, LabadanBA, Franková S. Postnatal nutritional deprivationsas determinants of adult rat behavior toward food,its consumption and utilization. J Nutr 1968;96:467-476.
50.-Salas M, Cintra L. Undernutrition and noveltyresponses influence of early food restriction on theresponsiveness to novel stimuli in rats. Bol EstudMed Biol Mex 1979; 30:201-204.
51.-File SE, Dale S. Effects of time of day andfood deprivation on exploratory activity in therat. Anim Behav 1972; 20:758-762.
52.-Wiener SG, Robinson L, Levine S. Influenceof perinatal malnutrition on adult physiologicaland behavioral reactivity in rats. Physiol Behav1983; 30:41-50.
53.-Levitsky DA. Malnutrition and the hunger tolearn. En: Levi tsky DA, ed. Malnutr i t ion,environment and behavior . I thaca: CornellUniversity Press 1979:161-179.
54.-Zimermman RR, Strobel DA, Maguire D.Neophobic reactions in protein malnourished infantmonkeys. Proc 78th An Conv APA 1970; 6:197.
55.-Barnett SA, Smart JL, Winddowson EM. Earlynutrition and the activity and feeding of rats in anartificial environment. Dev Psychobiol 1971; 4:1-15.
56.-Levistsky DA, Barnes RH. Nutritional and
environmental interactions in the development ofthe rat: Long-term effects. Science 1972; 176:67-71.57. -Whatson TS, Smar t JL, Dobbin J .Undernutrition in early life: Lasting effects onactivity and social behavior of male and femalerats. Dev Psychobiol 1976; 9:529-538.
58.-Stern W, Morgane PJ, Miller M, Resnick O.Protein malnutrition in rats: Response of brainamines and behavior to food shock stress. ExpNeurol 1975; 47:56-67.
59.-Escobar BC. Alteraciones morfológicas en laamígdala y el claustrum asociadas a la desnutri-ción neonatal en la rata. Tesis Doctorado. UNAM1992.
60.-Kupfermann I. Learning and memory. En:Kandel ER, Schwartz JH, Jessel TM. ed. Principlesof Neural Science. 3a ed. USA: Elsevier 1991:997-1008.
61.-Bower G, Hilgard E. El condicionamiento ope-rante de Skiner. En: Bower GH. ed. Teorías delAprendizaje. 2a ed. México: Trillas, 1992:16-267.
62.-Tonkiss J, Smart JL, Massey RF. Effects ofearly life undernutrition in artificially-reared rats2. Subsequent behaviour. Physiol Behav 1987;41:555-562.
63.-Stephens DN, Tonkiss J. Rats undernourishedas infants differ from controls in learning about acompound but not a simple discriminative stimulus.Physiol Behav 1981; 26:803-808.
64.-Stephens DN, Tonkiss J , Wearden JH.Dif ferences be tween ra ts undernour ishedpreweaning and controls in learning aboutredundant st imulus during acquisit ion of aconditioned emotional response. Physiol Behav1982; 28:95-101.
M García-Ruiz, L Parra, S Díaz-Cintra.
Revista Biomédica

106
1991; 19:247-250.
75.-Celedón JM, Smart JL, Dobbing J. Effects oflevel of motivation on visual discrimination,transfer of learning, and long-term memory inpreviously undernourished and control rats. NutrBehav 1982; 1:89-97.
76.-Lashley KS. Brain mechanisms and intelligence,a quantitative study of injuries to the brain. NewYork: Dover Publications, Inc., 1929:1-185.
77.-Simonson M, Chow BF. Maze studies onprogeny of underfed mother rats. J Nutr 1970;100:685-690.
78.-Hsueh AM, Simonson M, Chow BF, HansonAM. The importance of period of dietary restrictionof the dam on behavior and growth in the rat. JNutr 1974; 104:37-46.
79 . -Cravens RW. Effec ts of materna lundernutrition on offspring behavior: Incentivevalue of food reward and escape from water. DevPsychobiol 1974; 7:61-69.
80.-Smart JL. Reversal of spatial discriminationlearn in in a water maze by previous lyundernourished rats. Anim Learn Behav 1976; 4:313-316.
81.-Jordan TC, Cane SE, Howells KF. deficits inspatial memory performance induced by earlyundernutrition. Dev Psychobiol 1981; 14:317-325.
82.-Olton DS, Becker JT, Handelmann GE.Hippocampus, space and memory. Behav BrainSci 1979; 2:313-365.
83.-Jordan TC, Howells KF, McNaugthon N,Heatlie P. Effects of early undernutrition onhippocampal development and function. Res ExpMed 1982; 180:201-207.
Vol. 5/No. 2/Abril-Junio, 1994.
Desnutrición, aprendizaje y memoria.
65.-Roberts HJ, Smart JL, Wearden JH. Early lifeundernutrition and operant responding in the rat:The effect of the reinforcement schedule employed.Physiol Behav 1982; 28:777-785.
66.-Tonkiss J, Galler JR, Formica N, Shukitt H,Timm RR. Fetal protein malnutrition impairsacquisition of a DRL task in adult rats. PhysiolBehav 1990; 48:73-77.
67.-Kramer TJ, Rilling M. Diferential reinformentof low rates: a selective critique. Psychol Bull1970;74:225-254.
68.-Tonkiss J, Foster GA, Galler JR. Prenatalprotein malnutrition and hippocampal function:partial reinforcement extinction effect. Brain ResBull 1991; 27:809-813.
69.-Rogers PJ, Tonkiss J, Smart JL. Incidentallearn ing i s impai red dur ing ear ly- l i feundernutrition. Dev Psychobiol 1986; 19:113-124.
70.-Crnic L. Effects of nutrition on brain chemistryand behavior. Dev Psychobiol 1983; 16:129-145.
71.-Slob AK, Snow CE, Natris-Mathot E. Absenceof behaviora l def ic i t s fo l lowing neonata lundernutrition in the rat. Dev Psychobiol 1973;6:177-186.
72.-Levitsky DA. Malnutrition and the hunger tolearn. En: Levitsky DA. Malnutrition, Environmentand Behavior. Ithaca: Cornell University Press,1979:161-179.
73.-Castro CA, Rudy JW. Early-life malnutritionimpairs the performance of both young and adultrats on visual discrimination learning tasks. DevPsychobiol 1989; 22:15-28.
74.-Tonkiss J, Galler JR, Shukitt-Hale B, RoccoFJ. Prenatal protein malnutrition impairs visualdiscrimination learning in adult rats. Psychobiol

107
M García-Ruiz, L Parra, S Díaz-Cintra.
Revista Biomédica
84.-Hall RD. Is hippocampal function in the adultrat impaired by early protein or protein-caloriedeficiencies? Dev Psychobiol 1983; 16:395-411.
85.-Levitsky DA. Malnutrition and animal modelsof cognitive development. En: Serban G., ed.Nutrition and Mental Function. New York: PlenumPress; 1975: 75-89.
86.-Morris RGM. Spatial localization does notdepend on the presence of local cues. LearningMotiv 1981; 12:239-260.
87.-Goodlett CR, Valentino ML, Morgane PJ,Resnick O. Spatial cue utilization in chronicallymalnourished rats: Task-specific learning deficits.Dev Psychobiol 1986; 19:1-15.
88.-Galler JR, Mane M. Gender differences in vi-sual discr iminat ion by rats in response tomalnutrition of varying durations. Dev Psychobiol1980; 13:409-416.
89.-Rudy JW, Castro CA. Undernutrition duringthe brain growth period of the rat significantlydelays development of processes mediatingPavlovian trace conditioning. Behav Neural Biol1990; 53:307-320.
90.-Campbell LF, Bedi KS. The effects ofundernutrition in early life on spatial learning.Physiol Behav 1989; 45:883-890.
91.-Bedi KS. Spatial learning ability of ratsundernourished during early postnatal life. PhysiolBehav 1992; 51:1001-1007.
92.-Castro CA, Rudy JW. Early-life malnutritionselectively retards the development of distal-butnot proximal-cue navigation. Dev Psychobiol 1987;20:521-537.
93.-Idrobo F, Nandy K, Mostofsky DI, Nandy L.Dietary restriction: effects on radial maze learning
and l ipofusc in p igment depos i t ion in thehippocampus and frontal cortex. Arch GerontolGeriatr 1987; 6:355-362.
94.-Tonkiss J, Shukitt-Hale B, Formica RN, RoccoFJ, Galler JR. Prenatal protein alters response toreward in adult rats. Physiol Behav 1990; 48:675-680.
95.-Smart JL, Dobbing J . Vulnerabil i ty ofdeveloping brain. VI. Relative effects of fetal andearly postnatal undernutrition on reflex ontogenyand development of behaviour in the rat. BrainRes 1971; 33:303-314.
96.-Smart JL, Dobbing J . Vulnerabil i ty ofdeveloping brain. IV. Passive avoidance behaviorin young rats following maternal undernutrition.Dev Psychobiol 1972; 5:129-136.
97.-Morgane PJ, Austin-LaFrance RJ, BronzinoJD, Tonkiss J, Galler JR. Malnutrition and thedeveloping central nervous system. En: Robert L,LK Jensen , ed . The vulnerable bra in andenvironmental risk (volumen 1): Malnutrition andhazard assessment. New York: Plenum Press,1992:3-44.
98.-Tonges R, Hecht K, Hinz G, Docke F, DonerG. Effects of neonatal pyridostigmine on sexualmaturation and adult behavior in female ratssubjected to maternal deprivation. Endocrinol1981; 78:1-11.
99.-Plaut SM. Studies of undernutrition in theyoung rat: methodological considerations. DevPsychobiol 1970; 3:157-167.
100.-Wapnir RA, Lifshitz F. Fasting-inducedhypoglycemia in experimentally malnourished rats.J Nutr 1977; 107:383-390.
101.-Smart JL. Undernutri t ion,learning andmemory: a critical analysis. Clin Neuropharma

108
Vol. 5/No. 2/Abril-Junio, 1994.
Desnutrición, aprendizaje y memoria.
1984; 7(suppl 1):358-359.
102.-Winick M, Noble A. Cellular response in therats during maturation at various ages. J Nutr1966; 89:300-306.
103.-Cordero ME, Trejo M, García E, Barros T,Rojas AM, Colombo M. Dentritic development inthe neocortex of adult rats following a maintainedprenatal and/or early postnatal life undernutrition.Early Hum Dev 1986; 14:245-258.
104.-Katz HB. The influence of undernutrition onlearning performance in rodents. Common BureauNut 1980; 50:767-784.