lipidos de mebrana
9
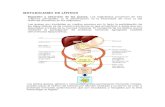
1 CONTENIDO: 1. Introducción: Lípidos estructurales de las membranas. 1.1 Glicerofosfolípidos o fosfoglicéridos. 1.2 Esfingolípidos. 1.3 Esteroles. 2. Biosíntesis de los Glicerofosfolípidos o Fosfoacilgliceroles. 3. Biosíntesis de los Esfingolípidos. 4. Biosíntesis del colesterol. 1. INTRODUCCIÓN: LÍPIDOS ESTRUCTURALES DE LAS MEMBRANAS. La membrana celular o plasmática es una estructura laminar que engloba a las células, define sus límites y contribuye a mantener el equilibrio entre el interior y el exterior de éstas. Además, se asemeja a las membranas que delimitan los orgánulos de células eucariotas (Figura 1). Está constituida por diversas biomoléculas (lípidos, proteínas carbohidratos) y mezcla de estos. En general, predominan los lípidos (Tabla 1). La característica arquitectónica central de las membranas biológicas es una doble capa lipídica que constituye una barrera al paso de moléculas polares y de iones. Los lípidos de las membranas son anfipáticos; un extremo de la molécula es hidrofóbico y el otro hidrofílico (Figura 2). Sus interacciones hidrofóbicas entre ellos y las hidrofílicas con el agua dirigen su empaquetamiento hacia la formación de láminas bicapas membranosas. Figura 1: Estructura general de las membranas biológicas. Figura 2: Estructura de los Fosfolípidos. En este tema se describen tres tipos de membrana: Los Glicerofosfolípidos, comúnmente llamados Fosfolípidos o fosfoacilglicéridos; los esfingolípidos y los esteroles. 1.1. Glicerofosfolípidos: Los Glicerofosfolípidos; también llamados Fosfoglicéridos, Fosfoacilgliceroles o Fosfolípidos; son lípidos de membrana en los que dos ácidos grasos están unidos por enlace éster al primer y segundo carbonos del glicerol y un grupo de cabeza muy polar o cargado está unido por enlace fosfodiéster al tercer carbono. El glicerol es una proquiral; no tiene carbonos asimétricos pero la unión del fosfato a cualquiera de los dos extremos lo convierte en un compuesto quiral que se denomina correctamente L-glicerol-3-fosfato, D-glicerol-1-fosfato o sn-glicerol (Figura 3). Estos compuestos, como derivados del ácido fosfatídico, se nombran según el alcohol polar en el grupo de cabeza (figura 4). Figura 3: D-glicerol-3-fosfato, el armazón de los Fosfolípidos
-
Upload
isai-millan -
Category
Documents
-
view
1.280 -
download
3
Transcript of lipidos de mebrana

1
CONTENIDO:
1. Introducción: Lípidos estructurales de las membranas. 1.1 Glicerofosfolípidos o fosfoglicéridos. 1.2 Esfingolípidos. 1.3 Esteroles.
2. Biosíntesis de los Glicerofosfolípidos o Fosfoacilgliceroles. 3. Biosíntesis de los Esfingolípidos. 4. Biosíntesis del colesterol. 1. INTRODUCCIÓN: LÍPIDOS ESTRUCTURALES DE LAS MEMBRANAS.
La membrana celular o plasmática es una estructura
laminar que engloba a las células, define sus límites y contribuye a mantener el equilibrio entre el interior y el exterior de éstas. Además, se asemeja a las membranas que delimitan los orgánulos de células eucariotas (Figura 1). Está constituida por diversas biomoléculas (lípidos, proteínas carbohidratos) y mezcla de estos. En general, predominan los lípidos (Tabla 1).
La característica arquitectónica central de las membranas biológicas es una doble capa lipídica que constituye una barrera al paso de moléculas polares y de iones. Los lípidos de las membranas son anfipáticos; un extremo de la molécula es hidrofóbico y el otro hidrofílico (Figura 2). Sus interacciones hidrofóbicas entre ellos y las hidrofílicas con el agua dirigen su empaquetamiento hacia la formación de láminas bicapas membranosas.
Figura 1: Estructura general de las membranas biológicas.
Figura 2: Estructura de los Fosfolípidos.
En este tema se describen tres tipos de membrana: Los Glicerofosfolípidos, comúnmente llamados Fosfolípidos o fosfoacilglicéridos; los esfingolípidos y los esteroles.
1.1. Glicerofosfolípidos:
Los Glicerofosfolípidos; también llamados Fosfoglicéridos, Fosfoacilgliceroles o Fosfolípidos; son lípidos de membrana en los que dos ácidos grasos están unidos por enlace éster al primer y segundo carbonos del glicerol y un grupo de cabeza muy polar o cargado está unido por enlace fosfodiéster al tercer carbono. El glicerol es una proquiral; no tiene carbonos asimétricos pero la unión del fosfato a cualquiera de los dos extremos lo convierte en un compuesto quiral que se denomina correctamente L-glicerol-3-fosfato, D-glicerol-1-fosfato o sn-glicerol (Figura 3). Estos compuestos, como derivados del ácido fosfatídico, se nombran según el alcohol polar en el grupo de cabeza (figura 4).
Figura 3: D-glicerol-3-fosfato, el armazón de los Fosfolípidos

2
Figura 3: Glicerofosfolípidos. Los Glicerofosfolípidos comunes son Diacilgliceroles unidos a
Alcoholes del grupo de cabeza mediante un enlace fosfodiéster.
1.2. Esfingolípidos:
Estos lípidos también tienen una cabeza polar y dos
colas apolares pero, a diferencia de los Glicerofosfo-lípidos, no contiene glicerol. Están compuestos por una molécula de amino-alcohol de cadena larga, la esfingo-sina (También llamada 4-esfingenina) o uno de sus derivados, una molécula de un ácido graso de cadena larga y un grupo de cabeza polar unido por enlace glucosídico en algunos casos y por enlace fosfodiéster en otros (Figura 4).
Figura 4: Los tres primeros carbonos del extremo polar de la
esfingosina son análogos a los tres carbonos del glicerol.
3
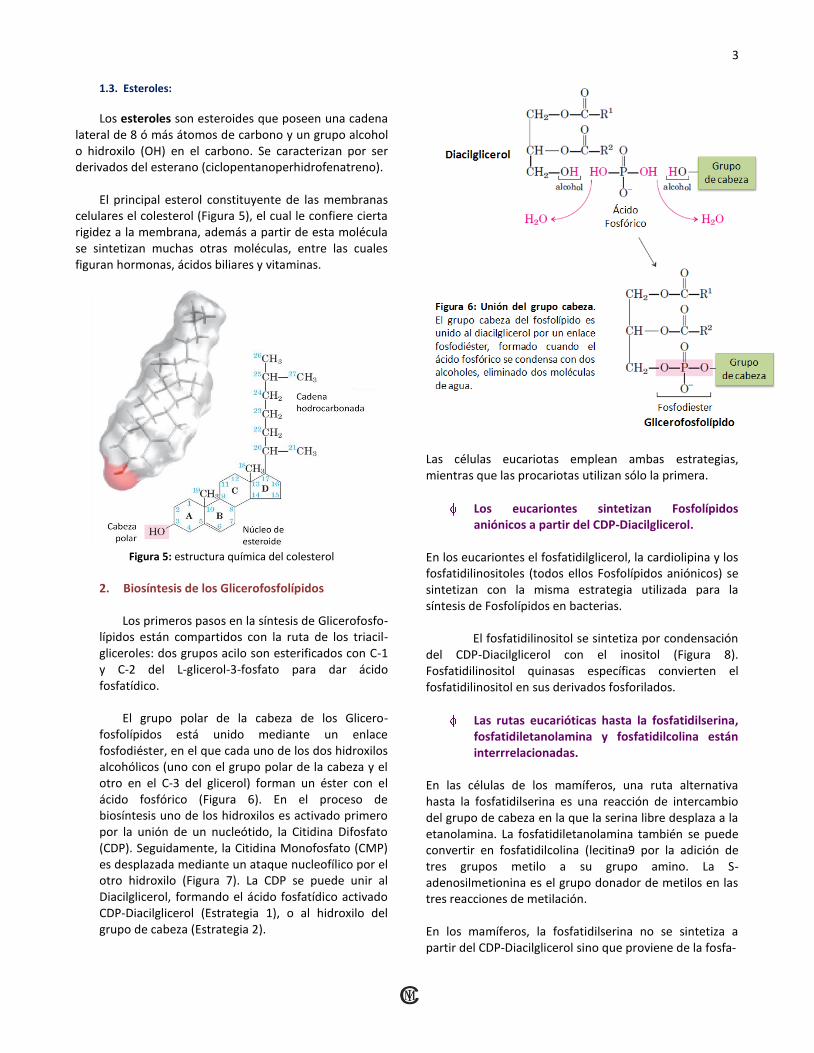
1.3. Esteroles:
Los esteroles son esteroides que poseen una cadena lateral de 8 ó más átomos de carbono y un grupo alcohol o hidroxilo (OH) en el carbono. Se caracterizan por ser derivados del esterano (ciclopentanoperhidrofenatreno).
El principal esterol constituyente de las membranas celulares el colesterol (Figura 5), el cual le confiere cierta rigidez a la membrana, además a partir de esta molécula se sintetizan muchas otras moléculas, entre las cuales figuran hormonas, ácidos biliares y vitaminas.
Figura 5: estructura química del colesterol
2. Biosíntesis de los Glicerofosfolípidos
Los primeros pasos en la síntesis de Glicerofosfo-
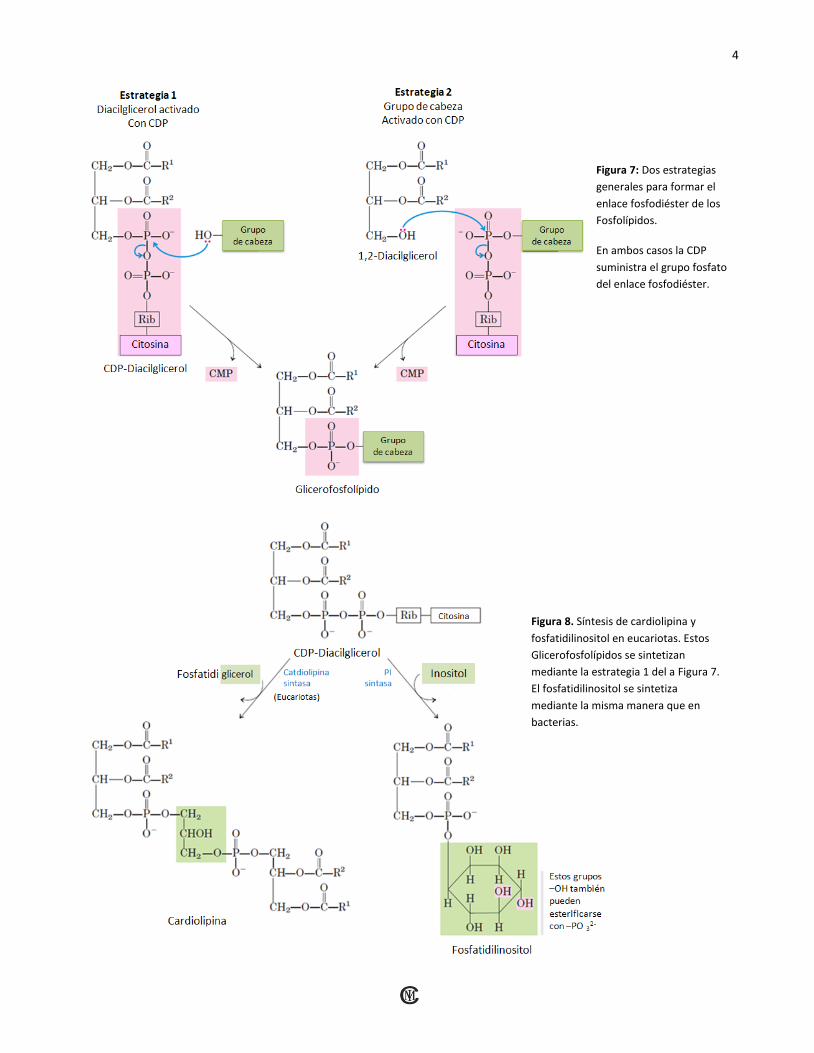
lípidos están compartidos con la ruta de los triacil-gliceroles: dos grupos acilo son esterificados con C-1 y C-2 del L-glicerol-3-fosfato para dar ácido fosfatídico. El grupo polar de la cabeza de los Glicero-fosfolípidos está unido mediante un enlace fosfodiéster, en el que cada uno de los dos hidroxilos alcohólicos (uno con el grupo polar de la cabeza y el otro en el C-3 del glicerol) forman un éster con el ácido fosfórico (Figura 6). En el proceso de biosíntesis uno de los hidroxilos es activado primero por la unión de un nucleótido, la Citidina Difosfato (CDP). Seguidamente, la Citidina Monofosfato (CMP) es desplazada mediante un ataque nucleofílico por el otro hidroxilo (Figura 7). La CDP se puede unir al Diacilglicerol, formando el ácido fosfatídico activado CDP-Diacilglicerol (Estrategia 1), o al hidroxilo del grupo de cabeza (Estrategia 2).
Las células eucariotas emplean ambas estrategias, mientras que las procariotas utilizan sólo la primera.
Los eucariontes sintetizan Fosfolípidos aniónicos a partir del CDP-Diacilglicerol.
En los eucariontes el fosfatidilglicerol, la cardiolipina y los fosfatidilinositoles (todos ellos Fosfolípidos aniónicos) se sintetizan con la misma estrategia utilizada para la síntesis de Fosfolípidos en bacterias. El fosfatidilinositol se sintetiza por condensación del CDP-Diacilglicerol con el inositol (Figura 8). Fosfatidilinositol quinasas específicas convierten el fosfatidilinositol en sus derivados fosforilados.
Las rutas eucarióticas hasta la fosfatidilserina, fosfatidiletanolamina y fosfatidilcolina están interrrelacionadas.
En las células de los mamíferos, una ruta alternativa hasta la fosfatidilserina es una reacción de intercambio del grupo de cabeza en la que la serina libre desplaza a la etanolamina. La fosfatidiletanolamina también se puede convertir en fosfatidilcolina (lecitina9 por la adición de tres grupos metilo a su grupo amino. La S-adenosilmetionina es el grupo donador de metilos en las tres reacciones de metilación. En los mamíferos, la fosfatidilserina no se sintetiza a partir del CDP-Diacilglicerol sino que proviene de la fosfa-
4
Figura 7: Dos estrategias
generales para formar el
enlace fosfodiéster de los
Fosfolípidos.
En ambos casos la CDP
suministra el grupo fosfato
del enlace fosfodiéster.
Figura 8. Síntesis de cardiolipina y
fosfatidilinositol en eucariotas. Estos
Glicerofosfolípidos se sintetizan
mediante la estrategia 1 del a Figura 7.
El fosfatidilinositol se sintetiza
mediante la misma manera que en
bacterias.

5
tidiletanolamina mediante una reacción de intercambio del grupo cabeza (figura 9).La síntesis de la fosfatidiletanolamina y la fosfatidilcolina en los mamíferos tiene lugar según la estrategia de la figura 7: fosforilación y activación del grupo de cabeza seguidas por la condensación con Diacilglicerol. Por ejemplo, la colina se reutiliza al ser fosforilada para luego convertirse en CDP-colina por condensación con CTP. Un Diacilglicerol desplaza CMP de la CDP-colina produciendo fosfatidilcolina (Figura 10).
Figura9: Ruta de recuperación de la fosfatidilserina a la
fosfatidiletanolamina. Una ruta de recuperación análoga convierte la etanolamina obtenida de la dieta fosfatidiletanolamina. En el hígado la fosfatidilcolina se produce también por metilación de la fosfatidiletanolamina (utilizando S-adenosilmetionina, como se ha descrito anteriormente), pero en el resto de los tejidos la fosfatidilcolina se produce por condensación del Diacilglicerol con CDP-colina. Las rutas para la formación del fosfatidilcolina y fosfatidiletanolamina en diversos organismos se resume en la figura 11.
Figura 10: Ruta de la síntesis de fosfatidilcolina a partir de
la colina en mamíferos.
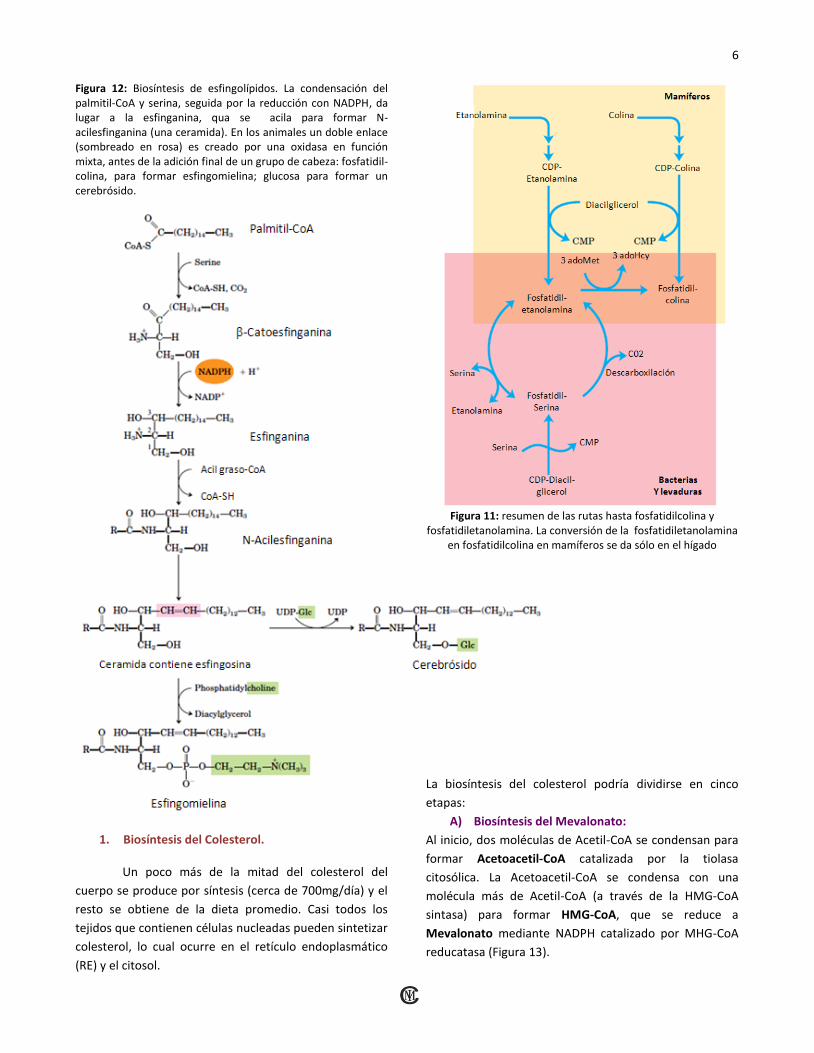
3. Biosíntesis de los Esfingolípidos. La biosíntesis de esfingolípidos tiene lugar en cuatro fases:
A. Síntesis de la mina de 18 carbonos esfinganina a partir de palmitil-CoA y serina.
B. Unión de un ácido graso en enlace amida para formar n-acilesfinganina.
C. Desaturación de la porción de la esfinganina para formar N-acilesfingosina (ceramida).
D. Unión de un grupo de cabeza para producir un esfingolípido tal cono un cerebrósido o esfingomielina (Figura 12).
6
Figura 12: Biosíntesis de esfingolípidos. La condensación del palmitil-CoA y serina, seguida por la reducción con NADPH, da lugar a la esfinganina, qua se acila para formar N-acilesfinganina (una ceramida). En los animales un doble enlace (sombreado en rosa) es creado por una oxidasa en función mixta, antes de la adición final de un grupo de cabeza: fosfatidil-colina, para formar esfingomielina; glucosa para formar un cerebrósido.
1. Biosíntesis del Colesterol.
Un poco más de la mitad del colesterol del
cuerpo se produce por síntesis (cerca de 700mg/día) y el
resto se obtiene de la dieta promedio. Casi todos los
tejidos que contienen células nucleadas pueden sintetizar
colesterol, lo cual ocurre en el retículo endoplasmático
(RE) y el citosol.
Figura 11: resumen de las rutas hasta fosfatidilcolina y
fosfatidiletanolamina. La conversión de la fosfatidiletanolamina en fosfatidilcolina en mamíferos se da sólo en el hígado
La biosíntesis del colesterol podría dividirse en cinco
etapas:
A) Biosíntesis del Mevalonato:
Al inicio, dos moléculas de Acetil-CoA se condensan para
formar Acetoacetil-CoA catalizada por la tiolasa
citosólica. La Acetoacetil-CoA se condensa con una
molécula más de Acetil-CoA (a través de la HMG-CoA
sintasa) para formar HMG-CoA, que se reduce a
Mevalonato mediante NADPH catalizado por MHG-CoA
reducatasa (Figura 13).

7
Figura 13: Biosíntesis del Mevalonato.
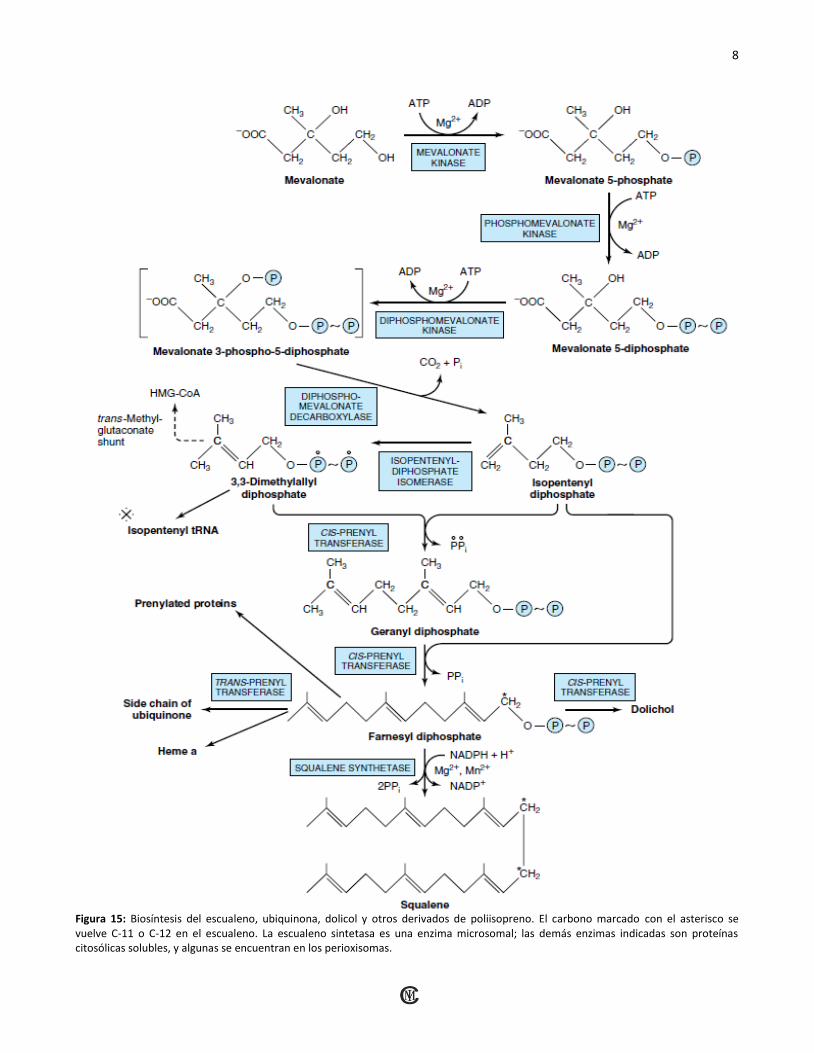
B) Formación de unidades Isoprenoides:
El ATP fosforila en forma sucesiva al Mevalonato
mediante tres cinasas, y después de la descarboxilación
(Figura 14), se forma la unidad isoprenoide activa
difosfato de isopentilo.
C) Seis unidades Isoprenoides forman escualeno:
El difosfato de isopentilo se isomeriza mediante un
desplazamiento del doble enlace para formar el
intermediario de 10 carbonos difosfato de geranilo
(figura 15). Una condensación más con difosfato de
isopentilo forma difosfato de farnesilo. Dos moléculas de
difosfato de farnesilo se condensan en el extremo de
difosfato para formar escualeno.
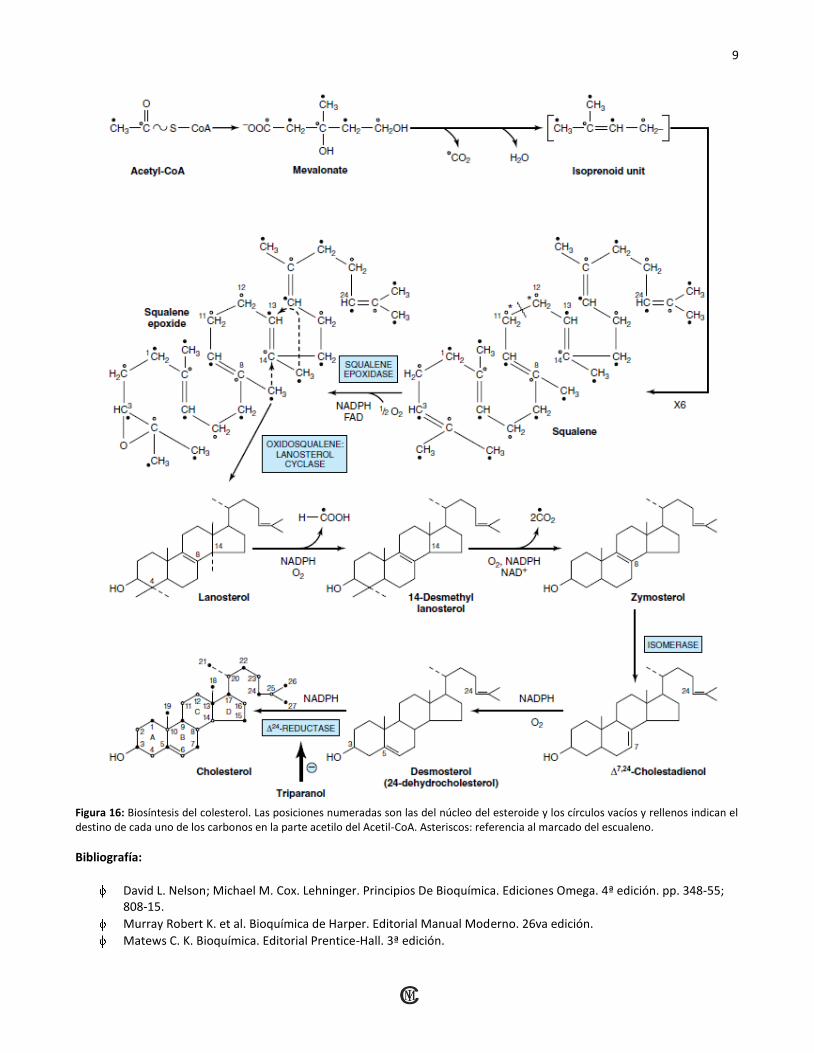
D) Formación del Lanosterol:
El escualeno se pliega en una estructura que se asemeja
bastante al núcleo de esteroide (Figura 16). Antes de que
ocurra el cierre del anillo, el escualeno se convierte en
2,3-epíxido de escualeno por medio de una oxidasa de
función mixta en el RE, la escualeno epoxidasa. El grupo
metilo en C-14 se transfiere al C-13 y el de C-8 a C-14
cuando ocurre la ciclización cuyo catalizador es la
oxidoescualeno: Lanosterol ciclasa.
E) Formación del Colesterol:
La formación del Colesterol a partir de Lanosterol se lleva
a cabo en las membranas del RE y se realiza en una serie
de pasos de los cuales no se sabe con certeza el orden en
que tiene lugar las reacciones (Figura 16).
Figura 14: Formación de unidades Isoprenoides.
8
Figura 15: Biosíntesis del escualeno, ubiquinona, dolicol y otros derivados de poliisopreno. El carbono marcado con el asterisco se vuelve C-11 o C-12 en el escualeno. La escualeno sintetasa es una enzima microsomal; las demás enzimas indicadas son proteínas citosólicas solubles, y algunas se encuentran en los perioxisomas.
9
Figura 16: Biosíntesis del colesterol. Las posiciones numeradas son las del núcleo del esteroide y los círculos vacíos y rellenos indican el destino de cada uno de los carbonos en la parte acetilo del Acetil-CoA. Asteriscos: referencia al marcado del escualeno.
Bibliografía:
David L. Nelson; Michael M. Cox. Lehninger. Principios De Bioquímica. Ediciones Omega. 4ª edición. pp. 348-55; 808-15.
Murray Robert K. et al. Bioquímica de Harper. Editorial Manual Moderno. 26va edición.
Matews C. K. Bioquímica. Editorial Prentice-Hall. 3ª edición.












![Lipidos Y Lipidos Simples [Modo De Compatibilidad]](https://static.fdocuments.ec/doc/165x107/557d1d50d8b42a6e4f8b4d75/lipidos-y-lipidos-simples-modo-de-compatibilidad.jpg)