
Heterocronía y variación morfológica en anuros - SciELO · males (¡el curso comprendía la...
20
29 Puntos de Vista Autor para correspondencia: [email protected] Heterocronía y variación morfológica en anuros Marissa Fabrezi Instituto de Bio y Geociencias. CONICET-Universidad Nacional de Salta. Mendoza 2. 4400-Salta. República Argentina Cuad. herpetol. 26 (1): 29-47 (2012) Recibido: 28 Julio 2011 Revisado: 28 Setiembre 2011 Aceptado: 4 Octubre 2011 Editor Asociado: F. Vera Candioti RESUMEN El concepto de heterocronía está ligado a la variación en la ontogenia y es frecuente en estu- dios de morfología comparada, involucra tanto a caracteres individuales como a todo el or- ganismo, al tamaño y a la forma y lleva implícito el tiempo en el cual ocurre una trayectoria ontogenética. El estudio de la heterocronía ha generado un conjunto de términos, definicio- nes y modelos en los que se confunde el origen de la variación (cambio heterocrónico) con sus consecuencias morfológicas en procesos evolutivos. En este trabajo se revisan algunos conceptos muy usados en el estudio de la heterocronía con ejemplos de la variación durante el desarrollo en anuros. Palabras clave: Pedomorfosis; Peramorfosis; Tamaño; Forma. ABSTRACT Heterochrony is a concept referred to ontogenetic variation and is frequently used in comparative morphology. It involves single traits, the whole organism, size, shape and has implicit the timing in which an ontogenetic trajectory occurs. The study of heterochrony has produced terms, definitions and models in which sometimes the heterochronic variation overlaps with its morphological results and evolutionary processes. Herein, the revision of some of these terms and definitions is discussed with data of developmental variation in anurans. Key words: Paedomorphosis; Peramorphosis; Size; Shape. Introducción El desarrollo animal en sí ofrece una infinidad de perspectivas para su estudio que convergen en lo que hoy llamamos Evo-Devo, un nuevo programa de investigación ubicado en la interfase de las interrelaciones causales–recíprocas entre el desarro- llo y la evolución de los organismos, a múltiples es- calas y niveles de análisis que une conceptos de la Biología del Desarrollo y de la Biología Evolutiva, desde genética a epigénesis; desde el nivel molecular al del organismo; desde un punto de vista histórico a uno funcional, con el intento de proporcionar una visión más comprensiva de la forma animal, enfatizando en el problema de la evolución de los fenotipos (Müller, 2007). En este contexto, la Mor- fología cobra un nuevo significado en estudios evo- lutivos, como lo han demostrado las investigacio- nes de los apéndices pares de los vertebrados en una serie de abordajes que combinan la expresión génica, la morfogénesis de los cartílagos primarios de las aletas pectorales en tiburones y la interpreta- ción del esqueleto de las aletas del fósil devónico Tiktaalik roseae (Daeschler et al., 2006; Shubin et al., 2006; Dahn et al, 2007; Shubin, 2009). El descu- brimiento de T. roseae y los estudios comparativos derivados, más amplios que aquellos centrados en organismos modelo, demuestran que el abordaje multidimensional que propone la Evolución del Desarrollo promete avanzar en nuestra compren- sión de los grandes temas de la Evolución. Aún cuando la Evolución del Desarrollo como disciplina en construcción, todavía es materia de debate desde que determinados grupos de investi-
Transcript of Heterocronía y variación morfológica en anuros - SciELO · males (¡el curso comprendía la...

29
Puntos de Vista
Autor para correspondencia: [email protected]
Heterocronía y variación morfológica en anuros
Marissa Fabrezi
Instituto de Bio y Geociencias. CONICET-Universidad Nacional de Salta. Mendoza 2. 4400-Salta. RepúblicaArgentina
Cuad. herpetol. 26 (1): 29-47 (2012)
Recibido: 28 Julio 2011Revisado: 28 Setiembre 2011Aceptado: 4 Octubre 2011Editor Asociado: F. Vera Candioti
RESUMENEl concepto de heterocronía está ligado a la variación en la ontogenia y es frecuente en estu-dios de morfología comparada, involucra tanto a caracteres individuales como a todo el or-ganismo, al tamaño y a la forma y lleva implícito el tiempo en el cual ocurre una trayectoriaontogenética. El estudio de la heterocronía ha generado un conjunto de términos, definicio-nes y modelos en los que se confunde el origen de la variación (cambio heterocrónico) consus consecuencias morfológicas en procesos evolutivos. En este trabajo se revisan algunosconceptos muy usados en el estudio de la heterocronía con ejemplos de la variación duranteel desarrollo en anuros.
Palabras clave: Pedomorfosis; Peramorfosis; Tamaño; Forma.
ABSTRACTHeterochrony is a concept referred to ontogenetic variation and is frequently used incomparative morphology. It involves single traits, the whole organism, size, shape and hasimplicit the timing in which an ontogenetic trajectory occurs. The study of heterochrony hasproduced terms, definitions and models in which sometimes the heterochronic variationoverlaps with its morphological results and evolutionary processes. Herein, the revision ofsome of these terms and definitions is discussed with data of developmental variation inanurans.
Key words: Paedomorphosis; Peramorphosis; Size; Shape.
Introducción
El desarrollo animal en sí ofrece una infinidad deperspectivas para su estudio que convergen en loque hoy llamamos Evo-Devo, un nuevo programade investigación ubicado en la interfase de lasinterrelaciones causales–recíprocas entre el desarro-llo y la evolución de los organismos, a múltiples es-calas y niveles de análisis que une conceptos de laBiología del Desarrollo y de la Biología Evolutiva,desde genética a epigénesis; desde el nivel molecularal del organismo; desde un punto de vista históricoa uno funcional, con el intento de proporcionar unavisión más comprensiva de la forma animal,enfatizando en el problema de la evolución de losfenotipos (Müller, 2007). En este contexto, la Mor-fología cobra un nuevo significado en estudios evo-lutivos, como lo han demostrado las investigacio-
nes de los apéndices pares de los vertebrados en unaserie de abordajes que combinan la expresióngénica, la morfogénesis de los cartílagos primariosde las aletas pectorales en tiburones y la interpreta-ción del esqueleto de las aletas del fósil devónicoTiktaalik roseae (Daeschler et al., 2006; Shubin etal., 2006; Dahn et al, 2007; Shubin, 2009). El descu-brimiento de T. roseae y los estudios comparativosderivados, más amplios que aquellos centrados enorganismos modelo, demuestran que el abordajemultidimensional que propone la Evolución delDesarrollo promete avanzar en nuestra compren-sión de los grandes temas de la Evolución.
Aún cuando la Evolución del Desarrollo comodisciplina en construcción, todavía es materia dedebate desde que determinados grupos de investi-

30
M. Fabrezi - Heterocronía en anuros
gación asumen que no existirían diferencias entreella y la Genética del Desarrollo, el hecho de queésta sea vista como un programa de investigaciónindependiente y conceptualmente diferente a laGenética del Desarrollo y a la Biología Evolutiva hagenerado un impulso en el diseño de nuevos pro-yectos en los que el re-descubrimiento de ideas yconceptos propuestos desde la Embriología clásicay la Anatomía se convierten en algo novedoso.
Como morfóloga, el concepto de heterocroníame ha resultado deslumbrante por su riqueza des-de el momento en que cursaba Embriología y Ana-tomía Comparadas, en la Facultad de Ciencias Na-turales de la Universidad Nacional de Tucumán.Utilizábamos como guía el Curso de Anatomía yFisiología Comparadas que el Dr. KonstatinGavrilov había elaborado para sus estudiantes deBiología y Odontología a los que llamábamos los«Cuadernos de Gavrilov». En los Cuadernos(Gavrilov, 1958–1981), había traducciones del In-glés, Alemán, Francés y Ruso de las grandes ideasde morfólogos del siglo XIX y principios del XX queeran ilustradas con ejemplos de la anatomía deembriones, larvas y adultos de distintos grupos ani-males (¡el curso comprendía la embriología y ana-tomía de los metazoos!) y más allá, cada clase con-sistía en observar y dibujar preparados anatómicosque llenaban el aula, discutir la variación de órga-nos y sistemas, aplicar los conceptos de analogía,homología, esbozo, vestigio, etc. y contrastarlo conliteratura, que incluía la variación en el registro fó-sil. Esta pequeña historia personal señala la orien-tación (o más bien el sesgo) del porqué voy a desa-rrollar este punto de vista considerando la varia-ción en la morfología y la heterocronía durante eldesarrollo tardío.
Para empezar, conviene repasar algunos con-ceptos e ideas generales que nos ayudarán a enten-der el punto de vista. En la Figura 1 se representaun esquema del desarrollo animal en el que se resu-men los procesos involucrados en una secuencia.Durante la gametogénesis, la meiosis es un tipo dedivisión celular exclusivo de las líneas germinales.En el desarrollo inicial, la determinación y la dife-renciación celular conducen a tipos celulares espe-cializados que indican que una célula o región re-presentará tal o cual grupo celular y en los que exis-te una dependencia de signos que rodean a las célu-las, como la inducción. Durante el desarrollo tar-dío, la cantidad procesos de división, interacción,organización e integración celular se expanden de
manera notable; generan crecimiento y originanestructuras tan diversas como los apéndices pares,el oído, o el sistema circulatorio que en conjuntoconducen al establecimiento de un plan corporaldistintivo. Entre el desarrollo inicial y el desarrollotardío, existe un estado (estado filotípico) en el cualel embrión, llamado faríngula, presenta todas lascaracterísticas fundamentales del plan corporal dellinaje al que pertenece (notocorda, sacos faríngeos,etc.). La faríngula es el estado del desarrollo quepresenta menor variación entre taxa y representaasí la forma animal que une la ontogenia de un taxóndado con la historia del linaje al que pertenece.
El concepto de heterocronía se usa para des-cribir alteraciones en el tiempo de los eventos dedesarrollo a través de los cuales los cambiosmorfológicos y las novedades se originan durantela evolución de un linaje. Por esta razón está ligadoa las influencias recíprocas entre desarrollo y evo-lución y entre ontogenia y filogenia. Desde su ori-gen, el concepto de heterocronía dio lugar a la pro-posición de una gran cantidad de términos e inter-pretaciones en los que los límites entre procesos (deldesarrollo y evolutivos) y patrones (morfológicos yfilogenéticos) se superponen entre sí y confunden.Sobre el tema, existe una abundante y variada lite-ratura pero la mayoría de los autores que teorizasobre la heterocronía insiste en los trabajos de Gould(1977) y Alberch et al. (1979) y en algunos ejem-plos clásicos como las curvas de edad y crecimien-to en primates. Quiero evitar proporcionar una grancantidad de citas o discutir interpretaciones pun-tuales que pueden desviar el objetivo inicial de estacontribución y me voy a limitar a presentar los tra-bajos en los que encuentro un marco conceptualpara interpretar los datos.
A partir del momento en que Haeckel formu-ló su Ley Biogenética, basada en secuencias de de-sarrollo en las que trataba a la heterocronía comoun cambio de secuencia, numerosos estudios handemostrado la recapitulación cuando se analizancaracteres simples (Richardson y Keuck, 2002). ParaHaeckel, la recapitulación, como adición de pasosterminales en una secuencia ontogenética, se opo-ne a la cenogénesis que se aplica a casos donde laalteración en una secuencia ocurre en estadios ini-ciales o intermedios, no al final de la ontogenia.
De Beer (1940) al comparar la variación entreestados «inmaduros» y «adultos» con respecto a unantecesor, definió ocho posibilidades de variaciónheterocrónica que:

31
Cuad. herpetol. 26 (1): 29-47 (2012)
Figura 1. Diagrama del reloj de arena (modificado de Carlson, 2003) donde se representa la secuencia en el desarrollo animal. Laexpansión de las bases indica un incremento en la diversidad en los procesos de desarrollo y también de la variación morfológicacon respecto al desarrollo inicial y el estado filotípico.
A.— aparecen en el estado inmaduro del an-tecesor y en la ontogenia descendiente se recono-cen como: 1) variación en estado inmaduro que
produce adaptación sin afectar la forma del adulto;cenogénesis; 2) variación en estado inmaduro y enel adulto, donde el nuevo estado adulto se desvía

32
M. Fabrezi - Heterocronía en anuros
del antecesor, desviación; 3) desarrollo somáticodemorado con respecto al desarrollo de los órga-nos reproductores; pedogénesis y neotenia, y 4) eldesarrollo de un carácter que conduce a un vesti-gio; reducción.
B.— se reconocen en el estado «adulto» delantecesor y en la ontogenia descendiente puedenaparecer como: 1) diferencias que distinguen indi-viduos en estados adultos, como variedades, razas;variación entre adultos (intraespecífica); 2) comocaracteres vestigiales en estados seniles; retarda-ción; 3) un exceso de ontogenia en los adultos;hipermorfosis, y 4) una precoz aparición de rasgosancestrales en estados juveniles, aceleración.
De Beer (1940) consideró que tanto lapedomorfosis (introducción de caracteres juveni-les del ancestro en estados adultos de la descenden-cia por desviación, pedogénesis o neotenia) y lagerontomorfosis (caracteres que estaban presentesen los adultos del ancestro y aparecen modificadosen estados adultos de la descendencia por variación,hipermorfosis o aceleración) tenían efectos en lafilogenia de manera opuesta ya que lagerontomorfosis produce recapitulación mientrasque las variaciones pedomórficas sonantirecapitulatorias o neogenéticas.
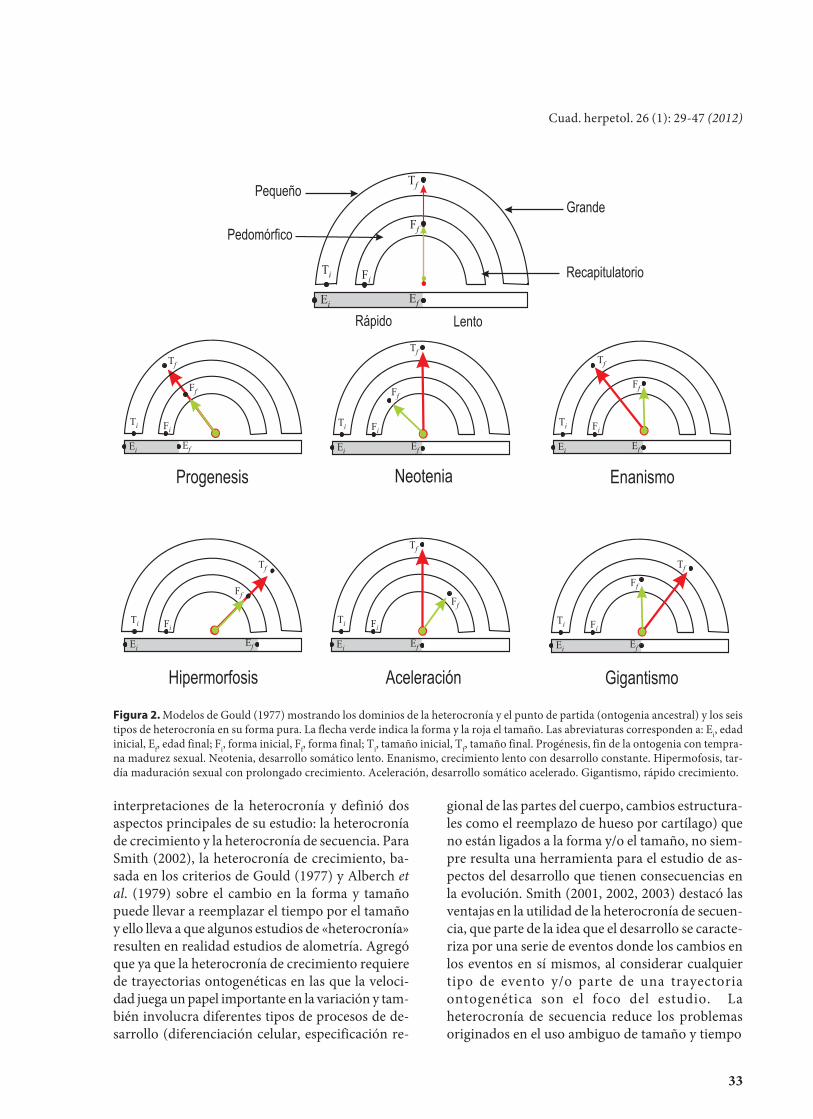
Después de varios años de latencia del con-cepto, Gould (1977) insistió en la idea del paralelis-mo entre ontogenia y filogenia y señaló a laheterocronía como proceso fundamental para lageneración de cambio con consecuencias en lafilogenia. En este contexto, caracterizó la variaciónde tres parámetros –tiempo, forma y tamaño– enlos linajes con respecto a una ontogenia ancestralpara definir los tipos de cambios y sus consecuen-cias en la evolución. Gould (1977) resaltó elparámetro tiempo en la interpretación de laheterocronía y la conceptualizó en seis tipos prin-cipales a los que representó gráficamente, utilizan-do un reloj, en el que la combinación de la posiciónde las agujas que representan la forma y el tamañodefinen cada tipo de heterocronía con respecto auna ontogenia ancestral en la que las agujas estánen posición 0 (Fig. 2). Cabe destacar que en losmodelos de Gould (1977) la distinta ubicación delas agujas que representan forma y tamaño con res-pecto al tiempo, ponen de manifiesto la indepen-dencia en el desarrollo de estas dos variables.
Posteriormente, Alberch et al. (1979) con laidea de proporcionar un marco conceptual más di-námico que el de Gould (1977) plantearon una se-
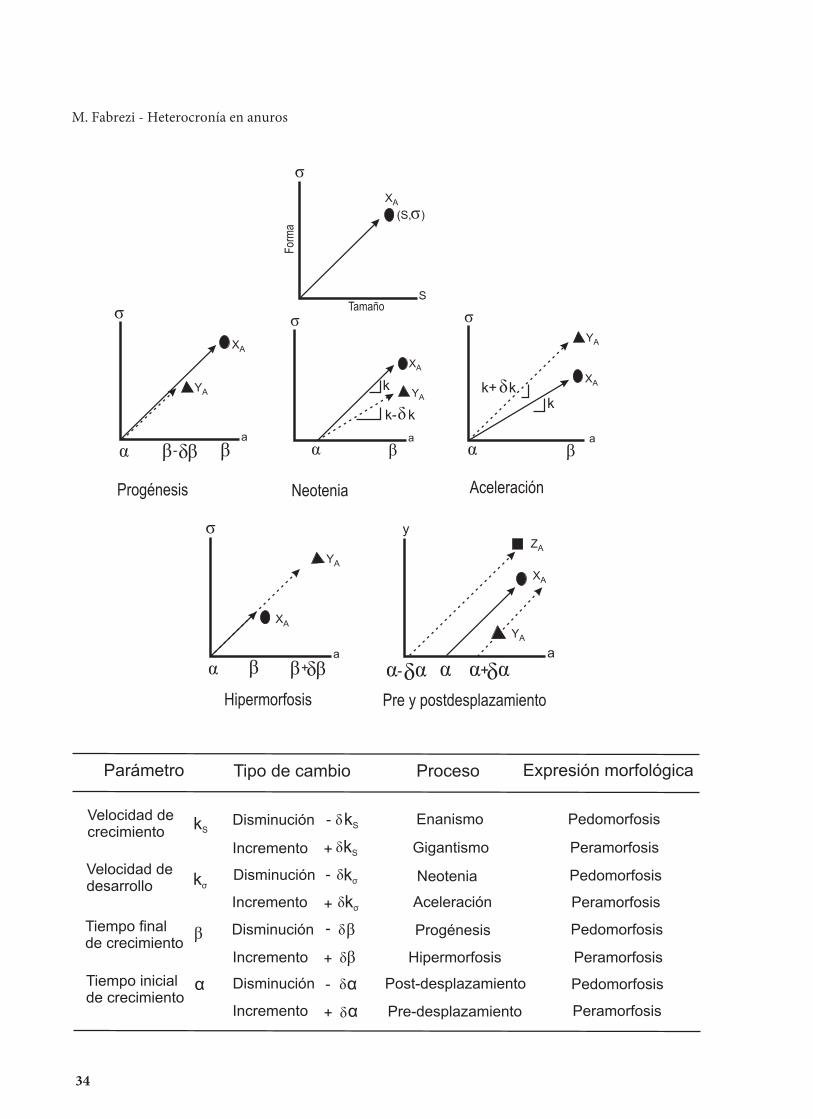
rie de modelos para ocho tipos de cambiosheterocrónicos con consecuencias en la filogeniarepresentados de manera sencilla en gráficos decoordenadas cartesianas. En estos modelos la for-ma y el tamaño fueron los elementos a partir de loscuales se representó la variación en los tiempos ini-cial y final y la velocidad de desarrollo (Fig. 3). Tantoen las interpretaciones de Gould (1977) y Alberchet al. (1979) se reconoce un marco conceptual paraentender el rol de las heterocronías sobre lasontogenias de todo el organismo y es en este punto,donde encontramos ciertas dificultades para deli-mitar con precisión las ideas de forma y tamaño yde desarrollo y crecimiento.
Reilly et al. (1997) revisaron la historia y evo-lución del concepto de heterocronía y centraron suestudio en los cambios simples que en conjuntoafectan a todo el organismo. Reconocieron que enla definición y el estudio de la heterocronía hayimplícitos seis aspectos importantes que no se de-ben dejar de lado: 1) la existencia de una hipótesisque proponga la trayectoria de la ontogenia ances-tral, 2) asumir que un cambio heterocrónico puedeo no tener consecuencias en la variaciónmorfológica, 3) delimitar con precisión el descriptordel rasgo a analizar; 4) contar con información realo inferencia explícita con respecto al tiempo (edad)en el que ocurren las trayectorias en la ontogeniaancestral y la descendiente, 5) contar con trayecto-rias ontogenéticas en las que las secuencias (varia-ción en el tiempo) sean completas, y 6) asumir laindependencia de los rasgos somáticos yreproductores. A partir de estas premisas, estos au-tores redefinieron la heterocronía con una nuevaterminología que distingue: 1) los patronesmorfológicos que ocurren al comparar ontogeniasindividuales de aquellos patrones que tienen con-secuencia en la variación intra e interespecífica, y2) los procesos de desarrollo en ontogenias indivi-duales de los procesos evolutivos intra einterespecíficos (Fig. 4). El trabajo de Reilly et al.(1997) contribuyó con una extensa argumentacióna la discusión de los términos neotenia y progénesispara identificar procesos heterocrónicos. Además,resaltaron la necesidad de definir el estado termi-nal de una trayectoria ontogenética como la por-ción de la curva donde la pendiente se acerca a 0para comparar curvas de un mismo rasgo sin tenerque referir el estado terminal a otro componentede la variación (madurez sexual, tamaño, etc.).
Smith (2001; 2002; 2003) revisó las distintas
33
Cuad. herpetol. 26 (1): 29-47 (2012)
interpretaciones de la heterocronía y definió dosaspectos principales de su estudio: la heterocroníade crecimiento y la heterocronía de secuencia. ParaSmith (2002), la heterocronía de crecimiento, ba-sada en los criterios de Gould (1977) y Alberch etal. (1979) sobre el cambio en la forma y tamañopuede llevar a reemplazar el tiempo por el tamañoy ello lleva a que algunos estudios de «heterocronía»resulten en realidad estudios de alometría. Agregóque ya que la heterocronía de crecimiento requierede trayectorias ontogenéticas en las que la veloci-dad juega un papel importante en la variación y tam-bién involucra diferentes tipos de procesos de de-sarrollo (diferenciación celular, especificación re-
gional de las partes del cuerpo, cambios estructura-les como el reemplazo de hueso por cartílago) queno están ligados a la forma y/o el tamaño, no siem-pre resulta una herramienta para el estudio de as-pectos del desarrollo que tienen consecuencias enla evolución. Smith (2001, 2002, 2003) destacó lasventajas en la utilidad de la heterocronía de secuen-cia, que parte de la idea que el desarrollo se caracte-riza por una serie de eventos donde los cambios enlos eventos en sí mismos, al considerar cualquiertipo de evento y/o parte de una trayectoriaontogenética son el foco del estudio. Laheterocronía de secuencia reduce los problemasoriginados en el uso ambiguo de tamaño y tiempo
EiEf
Tf
Ti Fi
Ff
EiEf
Tf
Ti Fi
Ff
EiEf
Tf
Ti Fi
Ff
EiEf
Tf
Ti Fi
Ff
EiEf
Tf
Ti Fi
Ff
EiEf
Tf
Ti Fi
Ff
EiEf
Tf
Ti Fi
Ff
Progenesis Neotenia
Hipermorfosis Aceleración
Enanismo
Gigantismo
PequeñoGrande
Recapitulatorio
Pedomórfico
Rápido Lento
Figura 2. Modelos de Gould (1977) mostrando los dominios de la heterocronía y el punto de partida (ontogenia ancestral) y los seistipos de heterocronía en su forma pura. La flecha verde indica la forma y la roja el tamaño. Las abreviaturas corresponden a: Ei, edadinicial, Ef, edad final; Fi, forma inicial, Ff, forma final; Ti, tamaño inicial, Tf, tamaño final. Progénesis, fin de la ontogenia con tempra-na madurez sexual. Neotenia, desarrollo somático lento. Enanismo, crecimiento lento con desarrollo constante. Hipermofosis, tar-día maduración sexual con prolongado crecimiento. Aceleración, desarrollo somático acelerado. Gigantismo, rápido crecimiento.
34
M. Fabrezi - Heterocronía en anuros
Parámetro Tipo de cambio Proceso Expresión morfológica
Disminución
Enanismo Pedomorfosis
Incremento Gigantismo Peramorfosis
Disminución
Disminución
Disminución
Incremento
Incremento
Incremento
Aceleración
Neotenia
Progénesis
Hipermorfosis
Post-desplazamiento
Pre-desplazamiento
Pedomorfosis
Pedomorfosis
Pedomorfosis
Peramorfosis
Peramorfosis
Peramorfosis
Velocidad decrecimiento
Velocidad dedesarrollo
Tiempo finalde crecimiento
Tiempo inicialde crecimiento
kS
kσ
α
-
-
-
+
+
+
-
+
kσ
kσ
α
α
kS
kS
Tamaño
For
ma
S
(S, )
XA
a
XA
YA
k- k
k
a
-
XA
YA
a
YA
k+ kk
XA
NeoteniaProgénesis Aceleración
a
+
YA
XA
Hipermorfosis
YA
XA
ZA
a
+-
y
Pre y postdesplazamiento

35
Cuad. herpetol. 26 (1): 29-47 (2012)
< Figura 3. La interpretación de Alberch et al. (1979) del tamaño S y la forma de un organismo (d), donde k es la tasa o velocidaddel crecimiento, a es la edad inicial del crecimiento y b la edad final del crecimiento que puede ser inferida por el limite del tamañoo la forma. XA representa la ontogenia ancestral. Progénesis y neotenia son perturbaciones negativas que producen descendientespedomórficos, la progénesis porque el crecimiento se detiene antes, y la neotenia por una tasa de crecimiento negativa de la forma.Aceleración e hipermorfosis son perturbaciones positivas que producen descendientes peramórficos. La aceleración por un incre-mento en la velocidad de crecimiento y la hipermorfosis por una prolongación del tiempo final del mismo. Pre y posdesplazamientose refieren a cambios en la edad a la cual una estructura comienza su ontogenia y resultan peramorficos o pedomórficos respectiva-mente a una misma velocidad de desarrollo. En la tabla, la distinción entre parámetros a analizar, tipos de cambio, procesos yconsecuencias morfológicas de estos autores.
y permite el análisis de los cambios temporales enel desarrollo cuando estos no pueden ser caracteri-zados por tamaño y forma. Smith (2001) propusoel uso de las secuencias como un método deestandarización en donde el desarrollo reconoce supropio tiempo para una evento dado cuando otroseventos han sido completados. La comparacióninterespecífica de trayectorias ontogenéticas permi-te establecer donde algunos caracteres de distintasunidades modulares del embrión se disocian y aque-llos que han co-evolucionado y para resaltarlo,Schlosser (2001) propuso un método gráfico muypráctico.
En el análisis de la variación morfológica,tanto la heterocronía de crecimiento como laheterocronía de secuencias, ofrecen un sinnúmerode herramientas para interpretar la variación comoconsecuencia del desarrollo. Sin embargo, la histo-ria y evolución de las ideas de heterocronía, las di-ficultades metodológicas para estudiar el desarro-llo y definir los parámetros objeto de estudio y lasparticularidades del desarrollo del grupo animal quese analiza convergen con cierta dificultad en la com-paración e interpretación de las ontogenias.
En la Figura 1, tres procesos importantes ca-racterizan el desarrollo tardío, organogénesis,morfogénesis y crecimiento, que se superponenpero son independientes. En la organogénesis y lamorfogénesis los procesos celulares implican laconstrucción de los componentes del plan corpo-ral mientras que el crecimiento involucra el arre-glo de esos componentes con un incremento en lasdimensiones espaciales a lo largo del tiempo. El cre-cimiento es un aspecto del desarrollo pero el desa-rrollo puede ocurrir sin crecimiento (Richtsmeier,2003). La morfogénesis conduce a la forma y el cre-cimiento actúa sobre la forma y el tamaño.
Tablas de desarrollo y tiempoEs importante destacar que para describir y com-parar trayectorias ontogenéticas necesitamos defi-nir puntos de referencia que sean un indicador de
tiempo para entender donde comienza y terminauna secuencia. Para ello son fundamentales las ta-blas de desarrollo que permiten fijar eventos deldesarrollo (estadios) en una secuencia estándar don-de ubicar las trayectorias. En anfibios hay tablas dedesarrollo estándar para especies con desarrollodirecto y con desarrollo larval. En los anuros condesarrollo directo, la eclosión es un evento en eltiempo de la ontogenia que indica que los procesosde organogénesis y morfogénesis han alcanzado unpunto que permiten la vida libre del individuo y co-mienza la etapa postembrionaria. En los anuros condesarrollo larval, el fin de la vida embrionaria se es-tablece en el momento en que las branquias exter-nas desaparecen y comienza la alimentación de lalarva, mientras que el fin de la vida larval es marca-do por la completa ausencia de la cola. Al compa-rar las ontogenias de dos anuros, la eclosión (en unaespecie con desarrollo directo) y la pérdida de lacola (en una especie con desarrollo larval) seríanpuntos de referencia equivalentes en la ontogenia(Schlosser, 2001).
A partir de la eclosión de un embrión y/o lapérdida completa de la cola (fin de la metamorfo-sis) existen varios estadios que se denominan concierta laxitud estadios postmetamórficos,metamorfoseados, juveniles, subadultos, adultos yque se organizan como series de tamaño sin preci-sar mayores detalles. En conjunto, deberían llamarseestadios «anuros propiamente dichos» ya queinvolucran diferentes etapas no madurassexualmente, maduras y también seniles que habríaque caracterizar. Estas etapas de la ontogeniainvolucran procesos morfogenéticos como diferen-ciación gonadal, osificación, desarrollo de caracte-res sexuales secundarios, o de crecimiento. Son muyimportantes en los estudios de la variación duranteel desarrollo y también exhiben una altísima varia-ción interespecífica e intraespecífica (especialmen-te diferencias entre los sexos).
El estado adulto se define por su capacidadreproductora, esto implica que en sus gónadas las

36
M. Fabrezi - Heterocronía en anuros
XA
YA
-k
Fo
rma XA
YA
Fo
rma
XA
YA
Fo
rma
Desaceleración Hipomorfosis Post-desplazamiento
tiempo tiempotiempo
PEDOMORFOSIS
PERAMORFOSIS
+k
XA XA XA
YA
YA YA
Fo
rma
Fo
rma
Fo
rma
tiempo tiempotiempo
Aceleración Hipermorfosis Pre-desplazamiento
Patrón ProcesoPatrón/Proceso
Desacelerado/Desaceleración
Hipomórfico/Hipomorfosis
Demorado/Post-dezplazamiento
Acelarado/Aceleración
Hipermórfico/Hipermorfosis
Adelantado/Pre-dezplazamiento
Combinación
Pedomorfosis
Pedogénesis
(Interespecífico)Pedomórfico
(Interespecífico)
Pedotípico(Intraespecífico) (Intraespecífico)
Peramorfosis(Interespecífico)
Peramórfico(Interespecífico)
Peratípico(Intraespecífico)
Peramorfosis(Intraespecífico)
Isotípico
Isomorfosis(Interespecífico)
Isomórfico(Interespecífico)
(Intraespecífico)Isogénesis
(Intraespecífico)

37
Cuad. herpetol. 26 (1): 29-47 (2012)
células sexuales (ovocitos vitelogénicos y esperma-tozoides) se encuentran listas para iniciar un nue-vo ciclo de vida. Algunos estudios de heterocronía(Alberch et al., 1979) en los que estados terminalesde desarrollo y crecimiento somático se ligan al es-tado adulto o la madurez sexual (ambosinvolucrados en las definiciones de neotenia yprogénesis) han generado confusión y por esta ra-zón, Reilly et al. (1997) propusieron el uso de tra-yectorias independientes para los caracteres de de-sarrollo somático y de desarrollo sexual, en dondeel estado terminal en cualquier trayectoria sea con-siderado como su asíntota.
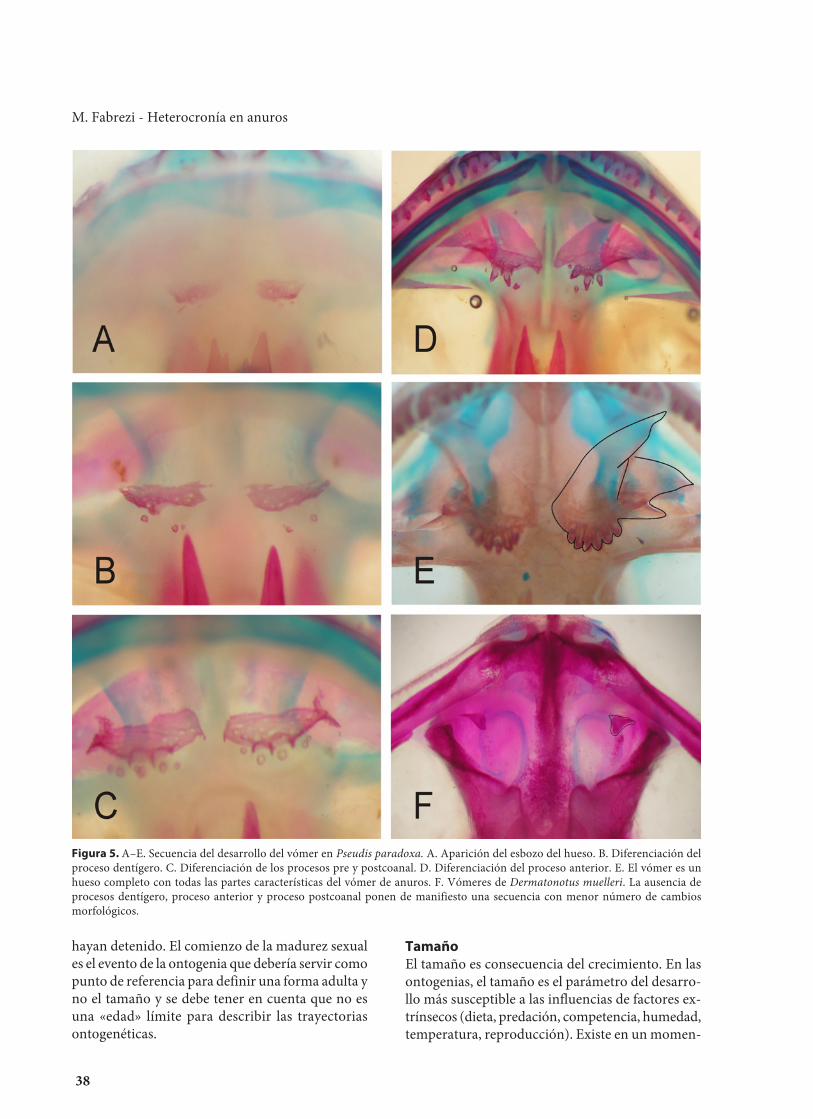
FormaEl parámetro forma se refiere tanto a la forma deun carácter individual, un complejo de caracteres oa la forma de un organismo en un momento dadoen el que se asume se adquieren una serie de carac-terísticas definitivas. La variación en el desarrollode la forma de un carácter se puede plantear inde-pendientemente del tamaño, considerando sola-mente aspectos cualitativos de su morfogénesis. Porejemplo, si la trayectoria a comparar entre dos es-pecies se refiere al fémur, la secuencia estaría dadapor los siguientes eventos: aparición del esbozocartilaginoso, diferenciación de las epífisis, desarro-llo de la cresta femoral, osificación de la diáfisis,osificación de epífisis. Si se toma como ejemplo elvómer de los anuros, reconocemos en una especieuna serie de cambios progresivos que culminan enun hueso complejo (Fig. 5 A–E); mientras que enotra especie la forma definitiva es más simple (Fig.5F). En ambos casos la forma es una característicaindependiente del tamaño y es posible sostener queen el segundo caso el vómer es incompleto. Hayformas complejas, definidas por la sumatoria de tra-yectorias ontogenéticas que implican morfogénesisy crecimiento. Por ejemplo la forma del autopodiode la extremidad posterior en los anuros que resul-ta de la combinación del crecimiento diferencial delos tejidos interdigitales y la morfogénesis y creci-miento diferencial de los metatarsales y falanges(Goldberg y Fabrezi, 2008).
Con respecto a la forma final del organismo,en los anfibios, la ontogenia implica dos entidadesmorfológicas y/o funcionales claramente diferen-
< Figura 4. Los seis tipos de cambios puros en los tiempos de desarrollo como fueron planteados por Reilly et al. (1997). Las flechasindican la hipótesis sobre la dirección del cambio si la ontogenia ancestral es XA. Estos autores sugirieron distinguir entre patronesy procesos según su nivel de ocurrencia como se indica en la tabla.
tes después del estado de faríngula (Fig. 1). En lasetapas postembrionarias, los procesos de desarro-llo y crecimiento continúan, ya sea directamentepara alcanzar la forma final de los adultos de algu-nas especies (desarrollo directo) o bien la formalarval de otras especies. Cabe destacar que la formafinal de cada gran etapa del ciclo de vida de un anu-ro, son sólo puntos de referencia para ubicar las tra-yectorias ontogenéticas y que no necesariamenterepresentan o están ligados a los extremos de tra-yectorias particulares.
Las larvas presentan un conjunto de caracte-res que desaparecen y otros que sufren transforma-ciones muy profundas durante la metamorfosis, apartir de ese momento se organizará una nueva for-ma que se desarrolla y crece en el adulto de la espe-cie. Aquí cabe plantearse cuál es la forma final de lalarva. Se podría decir que ésta es alcanzada durantelos estadios prometamórficos de Etkin (1936) ya queen ellos se encuentran completamente desarrolla-dos y son funcionales todos los caracteres larvalesque desaparecerán durante la metamorfosis (tuboanal, sistema de la línea lateral, disco oral y cola,por ejemplo) o bien, que la forma final de la larvase adquiere cuando la trayectoria ontogenética detodos los caracteres larvales se completa (fin de lametamorfosis) para continuar con procesos de de-sarrollo y crecimiento que conduzcan a la formadel adulto.
El criterio usado para definir el estadio adul-to se refiere al individuo sexualmente maduro. Enlas especies con dimorfismo sexual, algunos carac-teres sexuales secundarios como los sacos vocales yel canto, las callosidades nupciales, etc. correspon-den a un macho adulto; mientras que en especiessin dimorfismo sexual los machos adultos se iden-tifican por los testículos desarrollados y con signosde espermatogénesis. Los ovarios con ovocitosvitelogénicos confirman el estadio adulto de lashembras. Para definir la forma adulta en una espe-cie ¿lo hacemos siempre con la certeza de que es laforma de un individuo sexualmente maduro o in-ferimos que es una forma adulta por el tamaño? Sedebe tener en cuenta que en una serie ontogenéticapostembrionaria o postmetamórfica la forma adul-ta y/o madura sexualmente puede ser alcanzadaantes de que algunos procesos de crecimiento se
38
M. Fabrezi - Heterocronía en anuros
Figura 5. A–E. Secuencia del desarrollo del vómer en Pseudis paradoxa. A. Aparición del esbozo del hueso. B. Diferenciación delproceso dentígero. C. Diferenciación de los procesos pre y postcoanal. D. Diferenciación del proceso anterior. E. El vómer es unhueso completo con todas las partes características del vómer de anuros. F. Vómeres de Dermatonotus muelleri. La ausencia deprocesos dentígero, proceso anterior y proceso postcoanal ponen de manifiesto una secuencia con menor número de cambiosmorfológicos.
hayan detenido. El comienzo de la madurez sexuales el evento de la ontogenia que debería servir comopunto de referencia para definir una forma adulta yno el tamaño y se debe tener en cuenta que no esuna «edad» límite para describir las trayectoriasontogenéticas.
TamañoEl tamaño es consecuencia del crecimiento. En lasontogenias, el tamaño es el parámetro del desarro-llo más susceptible a las influencias de factores ex-trínsecos (dieta, predación, competencia, humedad,temperatura, reproducción). Existe en un momen-

39
Cuad. herpetol. 26 (1): 29-47 (2012)
to dado un tamaño final o al menos un tamaño enel cual el crecimiento parece detenerse (asíntota delcrecimiento corporal).
Como en la forma, el tamaño puede ser anali-zado considerando tanto a caracteres individualescomo a todo el organismo y para ello normalmenteusamos métodos cuantitativos de descripción.
Al analizar caracteres individuales es impor-tante tener en cuenta que la variación de tamañoen una estructura dada puede ser isométrica, es de-cir todas sus partes crecen de la misma manera oalométrica, donde una de las partes crece de mane-ra diferente al resto. Para algunos autores, esta va-riación con un cambio evolutivo en la cantidad dedesarrollo de una entidad se denomina heterometría(Arthur, 2004).
Para determinar el tamaño del organismo, enlos anuros, es importante considerar la etapa delciclo de vida. En algunos estudios, se consideró elpeso corporal como un indicador de tamaño peroéste no siempre resulta un buen parámetro. En elcaso de las larvas por ejemplo, la cantidad y el tipode contenido del intestino influyen notablementesobre el peso real del espécimen y por esta razón, esmás habitual el uso de longitudes. En especímenespostmetamórficos o posteclosión, se utiliza la lon-gitud entre el hocico y la cloaca para dar una aproxi-mación del tamaño del cuerpo. En el estudio de laslarvas, el tamaño de las mismas se refiere a la longi-tud entre el hocico y el extremo de la cola (siemprey cuando está sea completa). Sin embargo, para des-cribir la trayectoria ontogenética del tamaño en unaespecie dada, se debe considerar sólo el tamaño delcuerpo de la larva (longitud entre el hocico y el pun-to medio entre la bases de las extremidades poste-riores) ya que la cola no será un componente deltamaño definitivo. Sin embargo, la variación entrela longitud total de la larva y la longitud del cuerpode la larva de una especie en una trayectoria sonbuenos descriptores de crecimiento si la cola crecemás en relación al cuerpo o no.
TiempoUna trayectoria ontogenética tiene un comienzo yun final que ocurren dentro de límites de tiempopara cada especie y la finitud de la misma tiene comolímite máximo la duración de una generación. Tan-to el tiempo inicial como el tiempo final de una tra-yectoria ocurren en momentos dados que corres-ponden a determinadas «edades» y el tiempo entreambas edades indica la tasa o velocidad media en la
que esa trayectoria ocurre.A pesar de la importancia del parámetro
tiempo en los estudios de las ontogenias, la escasezde datos es un factor que muchas veces limita lacomprensión integral de la variación. La informa-ción sobre la duración en condiciones naturales delos ciclos de vida en anuros está limitada a muypocas especies y por lo tanto, algunos eventos seconsideran como «edades» de referencia sin contarcon valores absolutos de tiempo. Se ha menciona-do por ejemplo la madurez sexual como el eventoque denota el comienzo de la vida adulta, la apari-ción de las extremidades anteriores y la completadesaparición de la cola indican el comienzo y el finde la metamorfosis respectivamente. Estos eventospermiten de manera indirecta establecer edadespara describir una secuencia y formular compara-ciones.
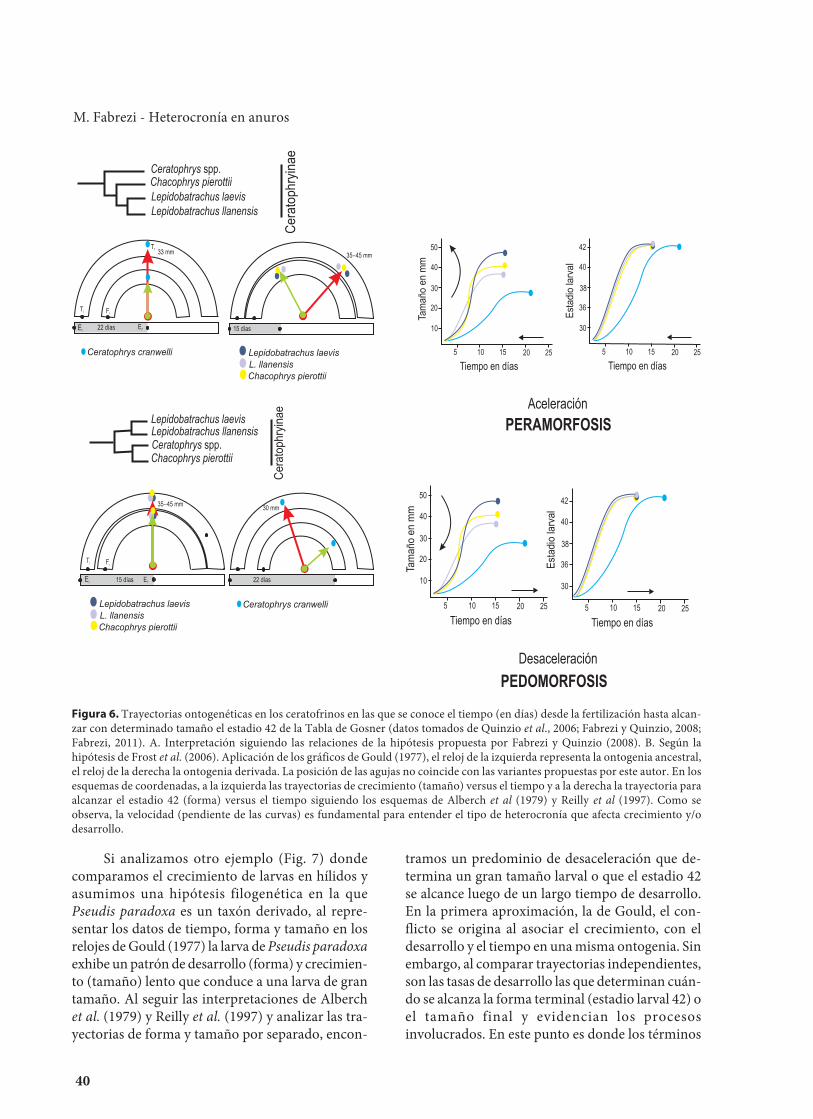
Trayectorias ontogenéticas y heterocronía decrecimientoEn la Figura 6 se grafican datos tomados de la lite-ratura (Fabrezi y Quinzio, 2008; Fabrezi, 2011) quesiguen las interpretaciones de Gould (1977),Alberch et al. (1979) y Reilly et al. (1997) y se con-sideran dos hipótesis filogenéticas para asignar laontogenia ancestral de los ceratofrinos. Si se asumeque el tamaño final de la larva es aquel en que lapendiente es 0 y que la forma final de la larva sealcanza cuando la piel del miembro anterior se fu-siona a la piel de la pared del cuerpo [que justa-mente coincide con la liberación de los miembrosanteriores del estadio 42 (Gosner, 1960) o al comen-zar la metamorfosis (Etkin, 1936)], las variacionesentre el tiempo del crecimiento y desarrollo que seobservan entre las distintas ontogenias no corres-ponden a las variaciones nombradas o definidas porGould (1977) al graficarlas en el esquema del reloj.Al aplicar las definiciones de cambios simples deAlberch et al. (1979) y las precisiones dadas porReilly et al. (1999) para comparar trayectorias, esposible determinar los tipos de variación en el cre-cimiento que están involucrados considerando lacurva de crecimiento en función del tiempo (sinnecesidad de considerar el estado de desarrollolarval) o la curva de desarrollo sin considerar el cre-cimiento. En cada caso y según la hipótesisfilogenética en la que se realice la interpretación elpatrón resultante será claramente acelerado odesacelerado, con incremento o disminución detamaño.
40
M. Fabrezi - Heterocronía en anuros
Aceleración
Desaceleración
PERAMORFOSIS
PEDOMORFOSIS
Est
ad
iola
rva
l
Tiempo en días
30
38
36
40
42
5 15 2510 20
Tam
añ
oe
nm
m
Tiempo en días
10
30
20
40
50
5 15 2510 20
Tam
año
en
mm
Tiempo en días
10
30
20
40
50
5 15 2510 20
Est
adio
larv
al
30
38
36
40
42
Tiempo en días
5 15 2510 20
Ce
rato
ph
ryin
ae
Lepidobatrachus laevis
Lepidobatrachus llanensis
Ceratophrys spp.Chacophrys pierottii
Ce
rato
ph
ryin
ae
Lepidobatrachus laevisLepidobatrachus llanensisCeratophrys spp.Chacophrys pierottii
EiEf
Tf
Ti Fi
22 días
33 mm
Ceratophrys cranwelli
22 días
30 mm
Ceratophrys cranwelli
15 días
35 45 mm–
Lepidobatrachus laevis
L. llanensis
Chacophrys pierottii
Ti Fi
15 días
35 45 mm–
Lepidobatrachus laevis
L. llanensis
Chacophrys pierottii
Ei Ef
Figura 6. Trayectorias ontogenéticas en los ceratofrinos en las que se conoce el tiempo (en días) desde la fertilización hasta alcan-zar con determinado tamaño el estadio 42 de la Tabla de Gosner (datos tomados de Quinzio et al., 2006; Fabrezi y Quinzio, 2008;Fabrezi, 2011). A. Interpretación siguiendo las relaciones de la hipótesis propuesta por Fabrezi y Quinzio (2008). B. Según lahipótesis de Frost et al. (2006). Aplicación de los gráficos de Gould (1977), el reloj de la izquierda representa la ontogenia ancestral,el reloj de la derecha la ontogenia derivada. La posición de las agujas no coincide con las variantes propuestas por este autor. En losesquemas de coordenadas, a la izquierda las trayectorias de crecimiento (tamaño) versus el tiempo y a la derecha la trayectoria paraalcanzar el estadio 42 (forma) versus el tiempo siguiendo los esquemas de Alberch et al (1979) y Reilly et al (1997). Como seobserva, la velocidad (pendiente de las curvas) es fundamental para entender el tipo de heterocronía que afecta crecimiento y/odesarrollo.
Si analizamos otro ejemplo (Fig. 7) dondecomparamos el crecimiento de larvas en hílidos yasumimos una hipótesis filogenética en la quePseudis paradoxa es un taxón derivado, al repre-sentar los datos de tiempo, forma y tamaño en losrelojes de Gould (1977) la larva de Pseudis paradoxaexhibe un patrón de desarrollo (forma) y crecimien-to (tamaño) lento que conduce a una larva de grantamaño. Al seguir las interpretaciones de Alberchet al. (1979) y Reilly et al. (1997) y analizar las tra-yectorias de forma y tamaño por separado, encon-
tramos un predominio de desaceleración que de-termina un gran tamaño larval o que el estadio 42se alcance luego de un largo tiempo de desarrollo.En la primera aproximación, la de Gould, el con-flicto se origina al asociar el crecimiento, con eldesarrollo y el tiempo en una misma ontogenia. Sinembargo, al comparar trayectorias independientes,son las tasas de desarrollo las que determinan cuán-do se alcanza la forma terminal (estadio larval 42) oel tamaño final y evidencian los procesosinvolucrados. En este punto es donde los términos

41
gigantismo/enanismo e hipermorfosis/hipo-morfosis, (Alberch et al., 1979; Reilly et al., 1997)no responden a un tipo de cambio en la ontogeniasino son su consecuencia.
La heterocronía de crecimiento es importan-te para interpretar la evolución del tamaño en unlinaje. Sin embargo, la combinación de distintasperturbaciones heterocrónicas tiene resultados queno necesariamente reflejan cuál de ellas ha predo-minado si se desconoce el tiempo en el que ocurrela trayectoria. Por esta razón conviene evitar el usode los términos pedomorfosis y peramorfosis cuan-do se aplican exclusivamente sobre la base del ta-maño final.
Trayectorias ontogenéticas y heterocronía desecuenciasEn esta segunda parte se hará referencia a la varia-ción en la morfogénesis, es decir aquella que con-diciona la forma y para ello también se tendrán encuenta las etapas larvales.
Para describir el desarrollo larval en anurosse utilizan tablas que caracterizan el cambio pro-gresivo en la forma de distintos rasgos exo-morfológicos. La tabla más usada es la Tabla deGosner (1960) que resume en 46 estadios la secuen-cia de desarrollo entre la fertilización y la ausenciacompleta de cola. Siguiendo esta tabla (Gosner,1960) se pueden describir las trayectoriasontogenéticas de algunos caracteres para formularcomparaciones que pueden ser usadas en análisisinterespecíficos y detectar heterocronías.
En la ontogenia de anuros con ciclos de vidabifásico, el desarrollo comprende tres conjuntos decaracteres, cuya diferenciación se inicia en etapasembrionarias y que tienen comportamientos dife-rentes en sus trayectorias (Fabrezi, 2011): 1) los ca-racteres exclusivos de las larvas, como el disco oral,sus estructuras queratinizadas y las papilas que lodelimitan, el espiráculo, el tubo anal y la cola; todosellos desaparecen durante la metamorfosis, es de-cir su trayectoria implica diferenciación, desarro-llo, forma final larval y desaparición; 2) caracteresque en las larvas tienen determinada configuracióny función y cambian durante la metamorfosis paraconstituir las configuraciones y funciones de la «for-ma adulta» típica. Por ejemplo el esqueleto de losarcos viscerales (mandíbulas, hioides y arcosbranquiales), el intestino, la piel; sus trayectoriascomprenden diferenciación, desarrollo, formalarval, transformación, desarrollo y forma definiti-
va; y 3) caracteres que comienzan a desarrollarseen etapas larvales y que adquieren funcionalidad enetapas postmetamórficas como las extremidades, lalengua, el complejo opercular-timpánico; con tra-yectorias que comprenden diferenciación, desarro-llo y forma definitiva. En los tres casos, las secuen-cias pueden mostrar variaciones heterocrónicas.
En la Figura 8 se representa la variación en lassecuencias de caracteres larvales, caracteres que setransforman y caracteres que pertenecen a los esta-dios «anuros propiamente dichos». Las áreassombreadas delimitan eventos de la ontogenia quepodrían ser considerados como puntos de referen-cia para inferir tiempos o «edades» relativas: el co-mienzo de la metamorfosis, identificado por la li-beración de las extremidades anteriores y el fin dela metamorfosis por la ausencia completa de cola.En varias especies de neobatracios (Leptodactylusbufonius, L. chaquensis, L. fuscus, L. latinasus,ceratofrinos, Physalaemus biligonigerus,Trachycephalus typhonius), el tubo anal desapareceantes del inicio de la metamorfosis. Inmediatamen-te, la cola comienza a acortarse y a sufrir una dis-minución general que implica la reducción de lasaletas y luego de la musculatura, hasta quedar re-ducida a un muñón de tejido atrofiado que puededesprenderse (Fabrezi, 2011). Al comparar la se-cuencia de desaparición de caracteres larvales en-tre hílidos (Phyllomedusa spp., Scinax spp; Pseudisparadoxa) tanto la desaparición del tubo anal comola reducción de la cola comienzan después de la li-beración de las extremidades anteriores con un re-traso notable en Pseudis paradoxa (Figura 8A).Fabrezi (2011) señaló que la desaparición de estoscaracteres larvales se encuentra demorada en espe-cies en las que el tubo anal está muy desarrollado(en forma de embudo) y la longitud de la cola esproporcionalmente mayor a la longitud del cuerpolarval.
En el segundo ejemplo (Fig. 8B) se presentanlas secuencias en el desplazamiento del ángulo dela boca que involucra el alargamiento y transfor-maciones de los cartílagos mandibulares(palatocuadrado, infrarostrales y cartílagos deMeckel). Cabe destacar que en las larvas de losneobatracios el ángulo de la boca siempre se en-cuentra por delante de la narina. A partir de estaposición inicial que se mantiene hasta el estadio 42(Gosner, 1960) se caracterizan cuatro morfologíassobre la base de la posición del ángulo de la bocaentre el comienzo y el fin de la metamorfosis. Esto
Cuad. herpetol. 26 (1): 29-47 (2012)
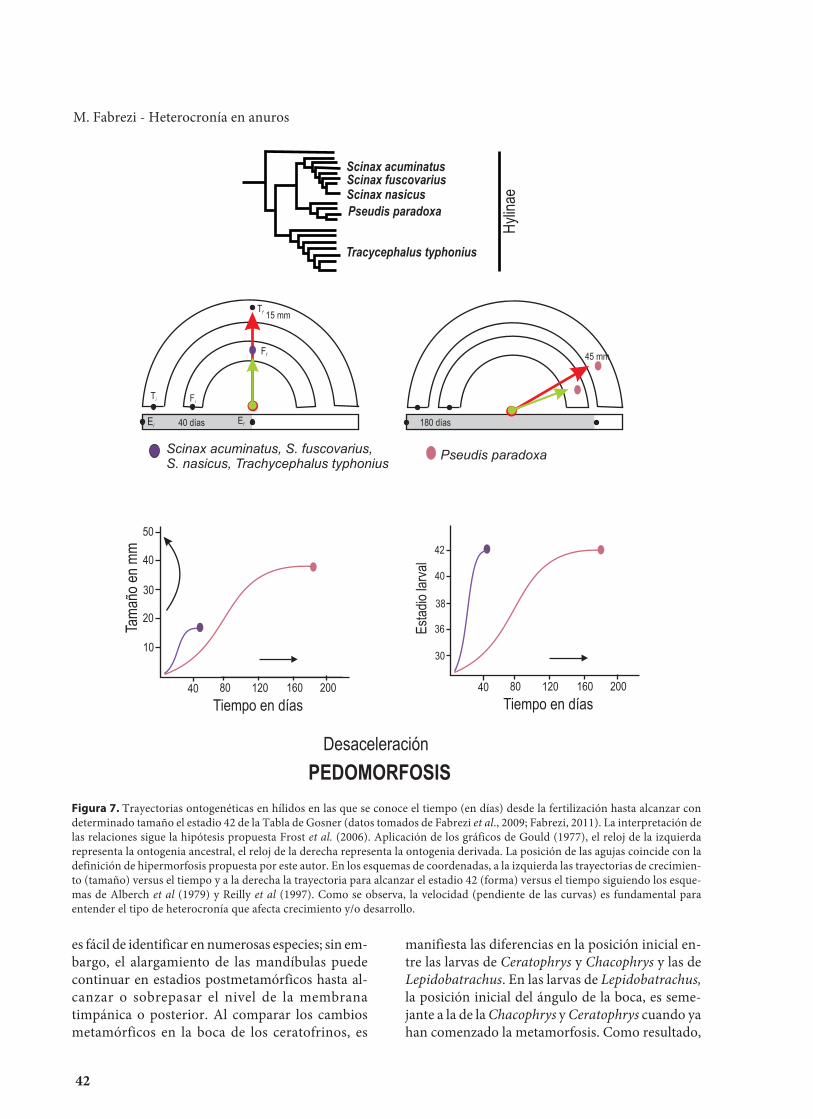
42
Hyl
inae
Scinax acuminatusScinax fuscovariusScinax nasicus
Tracycephalus typhonius
Pseudis paradoxa
EiEf
Tf
Ti Fi
Ff
15 mm
40 días
Scinax acuminatus, S. fuscovarius,S. nasicus, Trachycephalus typhonius
45 mm
180 días
Pseudis paradoxa
Desaceleración
PEDOMORFOSIS
Tam
año
enm
m
Tiempo en días Tiempo en días
10
30
20
40
50
40 40120 12080 80160 160200 200
Est
adio
larv
al
30
38
36
40
42
Figura 7. Trayectorias ontogenéticas en hílidos en las que se conoce el tiempo (en días) desde la fertilización hasta alcanzar condeterminado tamaño el estadio 42 de la Tabla de Gosner (datos tomados de Fabrezi et al., 2009; Fabrezi, 2011). La interpretación delas relaciones sigue la hipótesis propuesta Frost et al. (2006). Aplicación de los gráficos de Gould (1977), el reloj de la izquierdarepresenta la ontogenia ancestral, el reloj de la derecha representa la ontogenia derivada. La posición de las agujas coincide con ladefinición de hipermorfosis propuesta por este autor. En los esquemas de coordenadas, a la izquierda las trayectorias de crecimien-to (tamaño) versus el tiempo y a la derecha la trayectoria para alcanzar el estadio 42 (forma) versus el tiempo siguiendo los esque-mas de Alberch et al (1979) y Reilly et al (1997). Como se observa, la velocidad (pendiente de las curvas) es fundamental paraentender el tipo de heterocronía que afecta crecimiento y/o desarrollo.
es fácil de identificar en numerosas especies; sin em-bargo, el alargamiento de las mandíbulas puedecontinuar en estadios postmetamórficos hasta al-canzar o sobrepasar el nivel de la membranatimpánica o posterior. Al comparar los cambiosmetamórficos en la boca de los ceratofrinos, es
manifiesta las diferencias en la posición inicial en-tre las larvas de Ceratophrys y Chacophrys y las deLepidobatrachus. En las larvas de Lepidobatrachus,la posición inicial del ángulo de la boca, es seme-jante a la de la Chacophrys y Ceratophrys cuando yahan comenzado la metamorfosis. Como resultado,
M. Fabrezi - Heterocronía en anuros

43
osificación parsinterna plectrum
opérculopars media y externa
plectrum cartilaginosas
pars internaplectrum cartilaginosa
Amyetophrynus regularis (Sedra y Michael, 1959)
Prometamorfosis PostmetamorfosisMetamorfosis
osificación parsmedia plectrum
anillo timpánicodiferenciación
pars interna plectrumopérculo
pars mediaplectrum cartilaginosa
pars externaplectrum cartilaginosa
membrana timpánica
Pseudis paradoxa (Fabrezi y Goldberg, 2009)
Metamorfosis Postmetamorfosis
C
Pseudis paradoxa (Fabrezi et al., 2009)
cola completamente ausente
cola reducida a un muñón
reducción de la musculatura caudal
tubo analdesaparición
tubo anal
reducción de aletas dorsal y ventral
44 45 46
máxima longitudde la cola
43
cola completamenteausente
cola reducidaa un muñón
reducción de lamusculatura caudal
tubo anal
31–41
desaparicióntubo anal
reducción de aletasdorsal y ventral
43 44 45 46
máxima longitudde la cola
disco oralcompleto
desaparición deldisco oral
42
31–41
disco oralcompleto
desaparición deldisco oral
42
Scinax spp. (Fabrezi, 2011)
A
muy por detrás del ojo,posterior a la articulación cráneo-vertebralángulo de la boca
43 44 45 4628–41 42
alcanza elborde posterior del ojo
ángulo de la boca
por detrás del ojoángulo de la bocaángulo de la boca entre
el punto medio y bordeposterior del ojo
31–41 42
ángulo de la bocapor delante de la narina
ángulo de la bocaentre la narina y
borde anterior del ojo
43 44 45 46
Ceratophrys cranwelli, Chacophrys pierottii ( Fabrezi, 2006; 2011; )Fabrezi y Quinzio, 2008
alcanzael borde posterior del ojo
ángulo de la bocaentre
el punto medio y el bordeposterior del ojo
ángulo de la bocaángulo de la boca entrela narina y el borde
anterior del ojo
Lepidobatrachus spp. (Fabrezi, 2006; 2011; )Fabrezi y Quinzio, 2008
B
Figura 8. Heterocronías en las secuencias. A. Secuencias de desaparición de caracteres propios de las larvas. B. Secuencias decambios de caracteres larvales a adultos durante la metamorfosis. C. Secuencia de desarrollo de caracteres adultos.
Cuad. herpetol. 26 (1): 29-47 (2012)

44
en Lepidobatrachus las morfologías durante la me-tamorfosis también se encuentran adelantadas y tie-nen consecuencias en la forma definitiva, ya quelas especies de Lepidobatrachus son las únicas en-tre los anuros con la suspensión mandibular ubica-da tan atrás (Fabrezi, 2006; Fabrezi, 2011). Es im-portante detenerse aquí en un punto de la varia-ción durante el desarrollo en la que existe una va-riación temporal (heterocronía) pero también es-pacial (heterotopía). Al comenzar el desarrollolarval, en Lepidobatrachus los cartílagosmandibulares (palatocuadrado, cartílago de Meckele infrarostral) presentan una configuración clara-mente diferente a la del resto de las larvas que de-termina relaciones espaciales de la musculaturamandibular y las ramas del trigémino que se des-vían de la relación que exhiben todos los anurostanto en larvas como en adultos (Haas, 2001).
En el tercer caso (Fig. 8C) se comparan dossecuencias de desarrollo del sistema opercular-timpánico característico de los anuros. Aunque lainformación sobre este tema se conoce para muypocas especies (ver síntesis en Fabrezi y Goldberg,2009), la completa descripción en Amyetophrynusregularis (Sedra y Michael, 1959) podría ser exten-siva a otras especies o al menos coincide con lo ge-neralizado por Hetherington (1988) quien propu-so que la diferenciación del sistema opercular pre-cede a la del sistema timpánico que estaría asocia-do con las frecuencias relativas a las vocalizacionesen etapas reproductoras. En Pseudis paradoxa, lasecuencia difiere en varios aspectos: 1) hay un cam-bio en la secuencia del desarrollo del sistematimpánico, que en la mayoría de las especies co-mienza bien avanzada la metamorfosis o en esta-dios postmetamórficos; esta alteración podría te-ner alguna relación con el avanzado desarrollogonadal (ovarios con ovocitos previtelogénicos ytestículos con espermátidas) que presentan losrenacuajos de Pseudis al finalizar la metamorfosis yque indican que en Pseudis la edad reproductora oel estadio adulto es alcanzado casi inmediatamentefinalizada la metamorfosis (Downie, et al., 2009;Fabrezi et al., 2010); 2) hay un retraso en el tiempode diferenciación del sistema opercular hacia eta-pas postmetamórficas; como en otras ranas conadultos acuáticos [Xenopus laevis (Trueb y Hanken,1992), Pipa pipa (Trueb et al., 2000)]. Dado que elsistema opercular trasmite vibraciones de baja fre-cuencia del sustrato (Jaslow et al, 1988), se podríaespecular que completa su desarrollo antes de fina-
lizar la metamorfosis en el caso de las formas te-rrestres; 3) la alteración en la secuencia de desarro-llo de los distintos componentes del sistematimpánico no incide en su forma final ya que pre-senta la misma configuración que en todos aque-llos anuros en los que el sistema es completo; 4) laheterocronía de secuencias permite dilucidar cues-tiones de homología ya que la disociación de even-tos del desarrollo en este caso, pone de manifiestoque la pars externa del plectro y el anillo timpánicose originarían en relación con el palatocuadrado yno con la pars media-interna del plectro.
Heterocronías y terminologíaUno de los aspectos fundamentales en el estudio delas ontogenias y su variación, es la dificultad de ade-cuar el caso de estudio a modelos o enunciados pro-puestos y también a las diferentes terminologíasusadas para referirse a un mismo problema o a lainversa, el mismo término para distintos problemas.Por otro lado, la ausencia de variación no implicaque no exista heterocronía; la carencia de datos deldesarrollo deja de lado información muy importan-te para entender la diversificación y relaciones en-tre especies. A continuación se discutirán algunostérminos e interpretaciones que considero debería-mos usar con sumo cuidado en el estudio de la va-riación durante le desarrollo.
Los términos pedomorfosis y peramorfosis,se refieren a procesos y se aplican a heterocroníasque revelan menor o mayor cantidad de desarrollo,entendiendo que la cantidad de desarrollo está de-terminada por la combinación de la variación tem-poral de una trayectoria ontogenética (tiempo ini-cial, tiempo final y velocidad). Cuando la resultan-te es negativa, el resultado es pedomorfosis; si espositiva se trata de peramorfosis y si se compensan,seguramente el resultado es isomorfosis. El tiempoes un componente implícito en la heterocronía ypor lo tanto es necesario para entender los cambiosen el desarrollo.
Los términos hipomorfosis e hipermofosis,que se aplican a cualquier trayectoria en la que novarían la velocidad y el tiempo inicial, sino el tiem-po final (Alberch et al., 1979; Reilly et al, 1997), noson componentes de la variación en sí, sino su con-secuencia y no tienen una relación directa con pro-cesos de pedomorfosis o peramorfosis cuando nose tiene la certeza de que la velocidad y tiempo ini-cial sean los mismos en las trayectorias que se com-paran.
M. Fabrezi - Heterocronía en anuros

45
Un caso extremo de hipomorfosis (referido altamaño) se asocia al proceso evolutivo deminiaturización que parece ser característico de al-gunos linajes y que también se relaciona conpedomorfosis. Trueb y Alberch (1985) describie-ron la secuencia de desarrollo del los huesos del crá-neo en anuros y propusieron que algunos patronespodrían relacionar a las especies de tamaños peque-ños con la pérdida de algunos huesos (palatino,vómer, cuadradoyugal), huesos que no se pierdenen especies de tamaño grande. Yeh (2002) exploróentre los anuros la variación intraordinal en el ta-maño de los adultos, la forma del cráneo yosificaciones y describió algunos patrones que po-drían ser predictivos para algunos linajes que exhi-ben una tendencia a la reducción de tamaño. Enestos estudios, hay un establecimiento a priori querelaciona tamaño pequeño/desarrollo incompletocon un proceso de pedomorfosis.
Los términos progénesis y neotenia se refie-ren a casos muy estrictos en donde la comparacióninvolucra tamaño/ forma en una misma ontogenia.El primero se define por tamaño (pequeño) conmadurez sexual, se asocia a pedomorfosis pero nonecesariamente implica este proceso ya que quizásla forma final (definida por la madurez sexual) y eltamaño pueden ser también producto de acelera-ción. Los ejemplos de neotenia se refieren a larvascon madurez sexual, el término no se aplica a taxacon desarrollo directo, en anfibios, larvas madurassexualmente ocurren entre los urodelos y sólo hayun caso conocido entre los anuros(Spahenorhynchus bromelicola, Bokermann, 1974).Reilly et al. (1997) plantearon un análisis muy pre-ciso de la problemática de heterocronías enAmbystoma y explicaron los aspectos de la varia-ción heterocrónica que están involucrados en lo quese conoce como neotenia contrastando dos aspec-tos: la extensión en tiempo del desarrollo larval y laaceleración en la maduración gonadal. Tantoprogénesis como neotenia son términos que debe-rían ser usados en un contexto comparado en el quelos tiempos de desarrollo somático y gonadal y deltamaño sean conocidos.
Otros términos tales como cenogénesis, tie-nen muy poco uso pero quizás representen un tipode variación durante el desarrollo que no estaríacontemplada en los ejemplos. Haeckel la consideróopuesta a la recapitulación (adición de pasos en laparte final de una secuencia) y la refirió a caracte-res que se incorporan en etapas juveniles por
heterocronía, heterotopía o adaptación larval. Tantoel concepto propuesto por Haeckel como sus pos-teriores interpretaciones, incluida la de de Beer(1940) que lo restringe a «variación en estados ju-veniles (adaptación) sin afectar a los adultos» fue-ron ampliamente discutidas por Gould (1977). Eltérmino fue reintroducido por Blanco y Alberch(1992) en un análisis de la variabilidad en el desa-rrollo del carpo/tarso de los urodelos en el que seespecula que la heterocronía actúa sobre las extre-midades en desarrollo en respuesta a la existenciade demandas funcionales (cabe destacar que las ex-tremidades en desarrollo de los urodelos con ciclosde vida bifásico participan en la locomoción y ex-hiben alta variación intraespecífica en sus patronesde desarrollo). El término cenogénesis es compli-cado porque asocia la alteración en la trayectoriacon una explicación funcional (causal) sin afectarla forma final. Al analizar este concepto con infor-mación relacionada sobre la variación en laontogenia de los anuros surgen algunos posiblesejemplos: 1) variación intraespecífica en la presen-cia/ausencia del apéndice nasal durante el desarro-llo larval de Chacophrys pierottii (Quinzio et al.,2006) aunque no existe una explicación tentativasobre la adaptación o función que el mismo cum-ple; 2) la disociación en la secuencia de desarrollodel aparato opercular-timpánico en Pseudisparadoxa, que se especuló tendría alguna relacióncon la madurez sexual casi inmediatamente despuésde la metamorfosis.
Heterocronías, desarrollo larval y variaciónmorfológicaLos ejemplos presentados consideran la variacióndurante el desarrollo larval sobre la base de tama-ño, forma final y tiempos de desarrollo.
Al analizar heterocronías de crecimiento larvalel parámetro tiempo revela cuando hay aceleracióny en que casos se puede establecer peramorfosis (Fig.6) o cuando hay desaceleración y el resultado espedomorfosis (Fig. 7).
Al analizar las heterocronías en las secuencias,algunas de estas especies presentan también pertur-baciones. Una tardía desaparición de los caractereslarvales asociada a un temprano fin del desarrollode los caracteres funcionales de las etapaspostmetamórficas no tiene consecuencias en la for-ma terminal en el caso de Pseudis paradoxa (Fig.8A y C). Sin embargo, en el caso de las especies deLepidobatrachus en las que se observa una apari-
Cuad. herpetol. 26 (1): 29-47 (2012)

46
ción inicial en la larva de morfologías que general-mente ocurren durante la metamorfosis hay con-secuencias morfológicas en la forma final única quealcanzan los adultos (Fig. 8B). Sería este un ejem-plo en el que claramente la heterocronía, que seevidencia por procesos de desarrollo y crecimientoacelerados, sumada a la heterotopía, conduce a unainnovación morfológica.
Llama la atención que en la literatura queaborda tendencias evolutivas y diversificación delinajes, se ha enfatizado en la pedomorfosis comogeneradora de innovaciones (Clarke, 1996). Sinembargo, otros tipos de variaciones en el desarro-llo como las heterotopías juegan un papel clave enel origen de novedades morfológicas. Algunos ejem-plos frecuentes tienen que ver con el desarrollo deciertas estructuras esqueléticas como los elemen-tos intercalares en neobatracios (Manzano et al.,2007) y la osificación dorsal del hioides enLepidobatrachus (Fabrezi y Lobo, 2009). En estoscasos, en términos estrictos, la aparición de nuevasestructuras implica la adición de nuevas trayecto-rias a una ontogenia.
Desde mi punto de vista, el estudio de la va-riación durante el desarrollo debe profundizar lossiguientes aspectos principales: 1) distinguir entrelos parámetros forma y tamaño; 2) distinguir clara-mente la heterocronía de crecimiento yheterocronía de secuencias; 3) contar con muestrasque comprendan series ontogenéticas completas, enlas que las trayectorias puedan ser descriptas y com-paradas desde su comienzo hasta el final; 4) contarcon datos de los tiempos en los que ocurren loseventos; la heterocronía es básicamente un conceptoque se refiere a la variación temporal (tiempo deinicio, velocidad y tiempo final). Si bien obtenerestos datos en la ontogenia es difícil no es una tareaimposible; y 5) partir de una distinción clara entreel cambio heterocrónico y su consecuencia en lamorfología.
AgradecimientosEsta contribución surge de críticas, sugerencias ydiscusiones durante muchos años con VirginiaAbdala y Marcos Vaira, quienes además hicieronobservaciones sobre el manuscrito. Silvia Quinzioy Javier Goldberg aportaron información y discu-sión para profundizar el concepto de heterocronía.Demián Slodki sumó interesantes interrogantes aesta cuestión. David Buckley, Florencia VeraCandioti y un revisor anónimo hicieron comenta-
rios muy enriquecedores durante el proceso de re-visión. Este trabajo es parte del proyecto PIP-CONICET 0239.
Literatura citadaAlberch, P.; Gould, S.J.; Oster, G.F. & Wake, D.B. 1979. Size
and shape in ontogeny and phylogeny. Paleobiology 5:296–317.
Arthur, W. 2004. Biased embryos and evolution. CambridgeUniversity Press.
Blanco, M.J. & Alberch, P. 1992. Caenogenesis, developmentalvariability, and evolution in the carpus and the tarsus ofthe marbled newt Triturus marmoratus. Evolution 46:677–687.
Bokermann, W.C.A. 1974. Observações sobre desen-volvimento precoce em Sphaenorhynchus bromelicola Bok.1966 (Anura. Hylidae). Revista Brasileira de Biologia34:35–41.
Carlson, B.M. 2003. Developmental Mechanisms: Animal: 74–83. En B.K Hall& W. M. Olson (eds.), Keywords &Concepts in Evolutionary Developmental Biology.Harvard University Press.
Clarke, B.T. 1996. Small size in amphibians-its ecological andevolutionary implications. Symposium Zoological Societyof London 69:201–224.
Daeschler, E.B.; Shubin, N.H. & Jenkins, F.A.. 2006. ADevonian tetrapod-like fish and the evolution of thetetrapod body plan. Nature 440:757–763.
Dahn, R.D.; Davis, M.C.; Pappano, W.N. & Shubin, N.H.1.2007. Sonic hedgehog function in chondrichthyan fins andthe evolution of appendage patterning. Nature 445:311–314.
de Beer, G. 1940. Embryos and Ancestors. Oxford UniversityPress, Oxford.
Downie, J.R.; Sams, K. & Walsh, P.T. 2009. The paradoxicalfrog Pseudis paradoxa: Larval anatomical characteristics,including gonadal maturation. Herpetological Journal19:1–10.
Etkin, W. 1936. The phenomena of anuran metamorphosis.III. The development of the thyroid gland. Journal ofMorphology 59:68–89.
Fabrezi, M. 2006. Morphological evolution of Ceratophryinae(Anura, Neobatrachia). Journal of Zoological Systematicsand Evolutionary Research 44:153–166.
Fabrezi, M. 2011. Heterochrony in growth and developmentin anurans from the Chaco of South America. EvolutionaryBiology DOI:10.1007/s11692-011-9128-5.
Fabrezi, M. & Goldberg, J. 2009. Heterochrony during skeletaldevelopment of Pseudis platensis (Anura. Hylidae) and theearly offset of skeleton development and growth. Journalof Morphology 270: 205–220.
Fabrezi, M. & Lobo, F.J. 2009. Hyoid skeleton, related muscles,and morphological novelties in the frog Lepidobatrachus(Anura, Ceratophryidae). The Anatomical Record292:1700–1712.
Fabrezi, M. & Quinzio, S.I. 2008. Morphological evolution inCeratophyinae frogs (Anura. Neobatrachia): The effectsof heterochronic changes during larval development andmetamorphosis. Zoological Journal of the Linnean Society
M. Fabrezi - Heterocronía en anuros

47
© 2012 por el autor, licencia otorgada a la Asociación Herpetológica Argenti-na. Este artículo es de acceso abierto y distribuido bajo los términos y condi-ciones de una licencia Atribución-No Comercial 3.0 Unported de CreativeCommons. Para ver una copia de esta licencia, visite http://creativecommons.org/licenses/by-nc/3.0/deed.es
154:752–780.Fabrezi, M.; Quinzio, S.I. & Goldberg, J. 2009. The giant
tadpole and delayed metamorphosis of Pseudis platensisGallardo, 1961 (Anura. Hylidae). Journal of Herpetology43:228–243.
Fabrezi, M.; Quinzio, S.I. & Goldberg, J. 2010. The ontogenyof Pseudis platensis (Anura, Hylidae): Heterochrony andthe effects of larval development in the postmetamorphiclife. Journal of Morphology 271:496–510.
Frost, D.R.; Grant, T.; Faivovich, J.; Bain, R.H.; Haas, A.;Haddad, C.F.B.; de Sá, R.O.; Channing, A.; Wilkinson, M.;Donnellan, S.C.; Raxworthy, C.J.; Campbell, J.A.; Blotto,B.L.; Moler, P.; Drewes, R.C.; Nussbaum, R.A.; Lynch, J.D.;Green, D.M. & Wheeler, W.C. 2006. The amphibian treeof life. Bulletin of American Museum of Natural History297:1–370.
Gavrilov, K. 1958–1981. Curso de Anatomía y FisiologíaComparadas (La Evolución, Embriología, Tegumento,Esqueleto (2 fascículos). Órganos respiratorios (4fascículos). Escuela Universitaria de Ciencias Naturales yFacultad de Ciencias Naturales. Universidad Nacional deTucumán.
Goldberg, J. & Fabrezi, M. 2008. Development and variationof the anuran webbed feet (Amphibia. Anura). ZoologicalJournal of the Linnean Society 152:39–58.
Gosner, K.L. 1960. A simplified table for staging anuranembryos and larvae with notes on identification.Herpetologica 16:183–190.
Gould, S.J. 1977. Ontogeny and Phylogeny. Cambridge: TheBelknap Press of Harvard University Press.
Haas, A. 2001. Mandibular arch musculature of anurantadpoles, with comments on homologies of amphibian jawmuscles. Journal of Morphology 247:1–33.
Hetherington, T.E. 1988. Metamorphic changes in the middleear: 339–357. En: Fritzsch, B.; Ryan, M.J.; Wilczynski, W.;Hetherington, T.E. & Walkowiak, W. (eds.). The Evolutionof the Amphibian Auditory System. New York: WileyInterscience.
Jaslow, A.P.; Hetherington, T.E. & Lombard, R.E. 1988.Structure and function of the amphibian middle ear: 69–91. En: Fritzsch, B.; Ryan, M.J.; Wilczynski, W.;Hetherington, T.E. & Walkowiak, W. (Eds.). TheEvolution of the Amphibian Auditory System. New York:Wiley Interscience.
Manzano, A.S.; Fabrezi, M. & Vences, M. 2007. Intercalaryelements, treefrogs, and the early differentiation of acomplex system in the Neobatrachia. The AnatomicalRecord 290:1551–1567.
Müller, G.B. 2007. Six memos for Evo-Devo: 499–524. En:M.D. Laubichler & J Maienschein (eds.): From
Embryology to Evo-Devo. A History of DevelopmentalEvolution. The MIT Press, Cambridge.
Quinzio, S.I.; Fabrezi, M. & Faivovich, J. 2006. Redescriptionof the tadpole of Chacophrys pierottii (Vellard, 1948)(Anura: Ceratophryidae). South American Journal ofHerpetology 1:202–209.
Reilly, S.M.; Wiley, E.O. & Meinhardt, D.J. 1997. An integrativeapproach to heterochrony: The distinction betweeninterspecific and intraspecific phenomena. BiologicalJournal of the Linnean Society 60:119–143.
Richardson, M.K. & Keuck, G. 2002. Haeckel’s ABC ofevolution and development. Biological Reviews 77:495–528
Richtsmeier, J.T. 2003. 2003. Growth: Animal: 161–169. En:B.K Hall & W. M. Olson (eds.) Keywords & Concepts inEvolutionary Developmental Biology. Harvard UniversityPress.
Schlosser, G. 2001. Using heterochrony plots to detect thedissociated coevolution of characters. Journal ofExperimental Zoology 291:282–304.
Sedra, S.N. & Michael, M.L. 1959. The ontogenesis of the soundconducting apparatus of the Egyptian toad, Bufo regularisReuss, with a review of this apparatus in Salientia. Journalof Morphology 104:359–375.
Shubin, N. 2009. Your inner fish: a journey into the 3.5-billion-year history of the human body. New York: Vintage Books.
Shubin, N.H.; Daeschler, E.B. & Jenkins, F.A.. 2006. Thepectoral fin of Tiktaalik roseae and the origin of thetetrapod limb. Nature 440: 764–771.
Smith, K.K. 2001. Heterochrony revisited: The evolution ofdevelopmental sequences. Biological Journal of the LinneanSociety 73:169–186.
Smith, K.K. 2002. Sequence heterochrony and the evolutionof development. Journal of Morphology 252:82–97.
Smith, K.K. 2003. Time’s arrow: heterochrony and theevolution of development. Integrative Journal ofDevelopmental Biology 47:613–621.
Trueb, L. & Alberch P. 1985. Miniaturization and the anuranskull: A case study of heterochrony: 113–121. En: H-R.Duncker & G. Fleisher (eds.),Vertebrate Morphology.New York: Gustav Fischer Verlag.
Trueb, L. & Hanken, J. 1992. Skeletal development in Xenopuslaevis (Anura: Pipidae). Journal of Morphology 214:1–41.
Trueb, L.; Púgener, L.A. & Maglia, A.M. 2000. Ontogeny ofthe bizarre: An osteological description of Pipa pipa(Anura: Pipidae), with and account of skeletaldevelopment in the species. Journal of Morphology 243:75–104.
Yeh, J. 2002. The effect of miniaturized body size on skeletalmorphology in frogs. Evolution 56:2628–2641.
Cuad. herpetol. 26 (1): 29-47 (2012)

48


















