Herencia Citoplásmica o Extranuclear · se debe a diferencias genética entre los individuos y...
82
• Rama de la genética que se ocupa de la herencia de los caracteres que varían de forma continua, llamados caracteres cuantitativos o caracteres métricos, porque su estudio depende de la medición en lugar de la enumeración. • Estudio de los caracteres poligénicos y sus propiedades genéticas estadísticas cuando son sujetos a la selección artificial y a las fuerzas microevolutivas. Genética Cuantitativa
Transcript of Herencia Citoplásmica o Extranuclear · se debe a diferencias genética entre los individuos y...

• Rama de la genética que se ocupa de la herencia de los caracteresque varían de forma continua, llamados caracteres cuantitativos ocaracteres métricos, porque su estudio depende de la medición enlugar de la enumeración.
• Estudio de los caracteres poligénicos y sus propiedades genéticasestadísticas cuando son sujetos a la selección artificial y a lasfuerzas microevolutivas.
Genética Cuantitativa
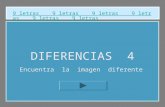
Variación cuantitativa vs mendeliana (o cualitativa)
Caracteres cualitativos mendelianosDistribución de la altura de varones

Preguntas qué trata de responder la genética cuantitativa
•¿Qué parte de la variación fenotípica de un carácter cuantitativo se debe a diferencias genética entre los individuos y qué parte a diferencias en el ambiente?
•¿Qué parte de la variación fenotípica puede ser seleccionada por un mejorador o por la selección natural?
•¿Cuántos genes o loci influyen sobre el carácter?
•¿Cómo se distribuyen los loci por el genoma?
•¿Qué efecto tienen los loci y como interactúan entre sí?

• Darwin (1859) y Mendel (1865 1900)
• Se creyó durante mucho tiempo que los caracteres cuantitativos y los
mendelianos obedecen a leyes distintas. La variación es muy importante para la
evolución: Biométricos (caracteres cuantitativos) vs. mendelianos (caracteres
cualitativos).
BASE MENDELIANA DE LA VARIACIÓN CONTINUA
• 1918: Sir Ronald Fisher integra el mendelismo con la biometría.
Demuestra que la variación cuantitativa es una consecuencia natural de la
herencia mendeliana.

• Los principios genéticos sobre los que se fundamenta la herencia de los
caracteres métricos o cuantitativos son los mismos que los que hasta ahora
hemos visto que rigen la transmisión de los caracteres cualitativos.
BASE MENDELIANA DE LA VARIACIÓN CONTINUA
• La naturaleza de los genes con efecto sobre los caracteres cuantitativos no es
diferente de la de los demás genes.
• Tras el redescubrimiento de las leyes de Mendel, a principios del siglo XX,
surgió el problema de explicar la herencia de los caracteres cuantitativos,
algunos opinaban que estos caracteres no se heredaban, sino que la variación
era debida al ambiente y no tenían nada que ver con la genética mendeliana.
• Los estudios de Wilhelm Johansen y Herman Nilsson-EhIe, que se plasmaron
respectivamente en la teoría de las líneas puras y la teoría de los factores
polímeros (poligénicos), aclararon esta dificultad inicial, y constituyen lo que se
denomina base mendeliana de la variación continua.

Teoría de las líneas puras
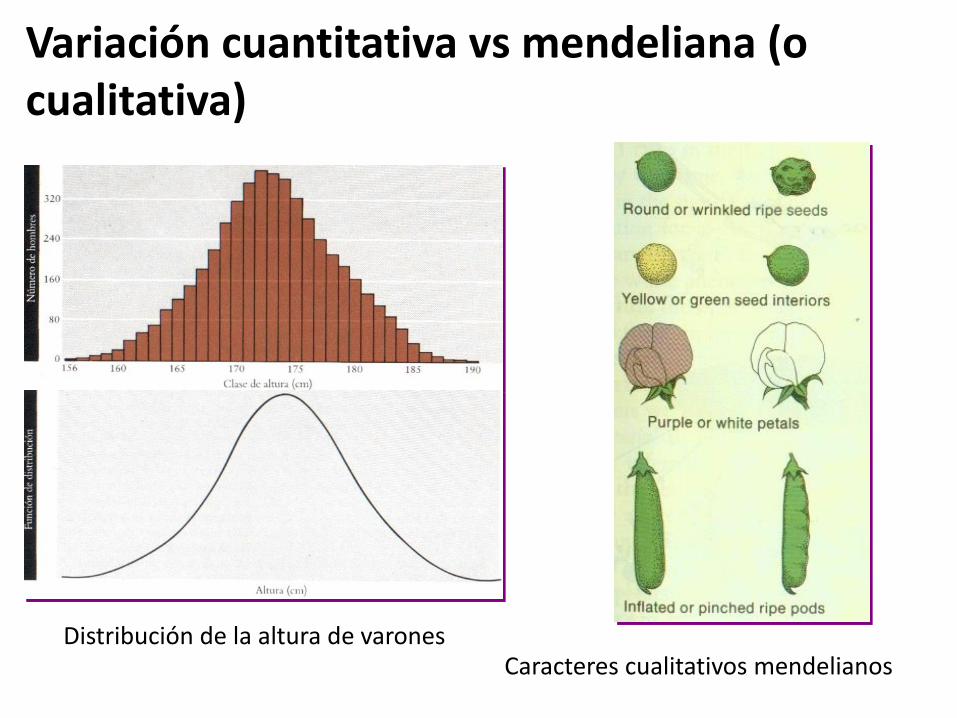
Variación ambiental
aa Plantas enanas AA Plantas altas
Altura planta
aa AA
Alguna variación ambiental
aa AA
Altura planta
Mucha variación ambiental
Altura planta
aa AA
Sin variación ambiental
Frec
uen
cia

P
F1
F2
Altura plantaPlantas enanas x plantas altas
F1 x F1
Altas x altas
Fre
cuenc
iaF
recu
enc
ia
Altas 3/4
Enanas 1/4
Fre
cuenc
ia
Pisum sativum
6045 75 80 110
95
80 110
95

P
F1
F2
Longitud de la espiga
Cortas x largas
F1 x F1
Fre
cuenc
iaF
recu
enc
iaF
recu
enc
ia
Zea mays (maíz) Longitud de espiga
1510 20 25 35
30
15 30
22,5
6 3922,5

Pisum sativum Zea mays
P
F1
F2
Carácter Mendeliano ---- Carácter cuantitativo
Fre
cuenc
iaF
recu
enc
iaF
recu
enc
ia

CARACTERES CUANTITATIVOS Y CUALITATIVOS
CUANTITATIVOS CUALITATIVOS
No pueden clasificarse en clases discretas.
Son rasgos fácilmente clasificables en distintas
categorías fenotípicas.
Gobernados por muchos genes, cada uno
contribuyendo con una pequeña cantidad al
fenotipo (poligenes).
Bajo control genético de sólo uno o unos cuantos
genes.
Tienen componentes ambientales además de
genéticos.
Tienen poca o ninguna modificación ambiental
que oculte sus efectos.
Variabilidad continua. Variabilidad discontinua.
Se estudian poblaciones y todos los tipos de
cruzamientos.
Se estudian apareamientos individuales y su
progenie.
El análisis es de tipo estadístico, proporcionado
cálculos aproximados de los parámetros de
las poblaciones.
El análisis es por medio de cálculos de
proporciones y relaciones.

EJEMPLOS DE CARACTERES CUANTITATIVOS
• La altura de las plantas maduras
• La tasa de producción de huevo o leche
• La producción de granos por hectárea
• Resistencia a algunas enfermedades en las plantas
• Tamaño de camada
• Crecimiento de los niños
• Peso de los adultos
• Cantidad de colesterol en el suero
• Longevidad

La importancia de la Genética Cuantitativa
• Rasgos de importancia economica
son cuantitativos
• Teórias de Genética Cuantitativa son
la base de los programas de
mejoramiento.
• La varación ambiental reduce la
eficiencia de la selección.

EL AMBIENTE INFLUYE EN LA EXPRESIÓN
• El ambiente influye en su expresión fenotípica de tal modo que las clasesfenotípicas observables no se pueden relacionar con genotipos concretos.
• La norma de reacción es el conjunto de vías que puede seguir ungenotipo, en todos los ambientes favorables y desfavorables, hasta darlugar a los diversos fenotipos posibles.

• Indica la relación existente entre el fenotipo (manifestación)correspondiente a una constitución genética determinada y lascondiciones ambientales.
Fenotipo= Factores genéticos + Factores ambientales
• Ejemplo: un genotipo AABbccDd. Que afecta el carácter “peso”. Silos alelos en mayúsculas aportan 5 gramos, y los en minúsculas 1gramo, el peso “genético” será de 24 gramos. Pero este peso estáinfluido por el ambiente, y tal individuo puede pesar mucho menossi come muy poco, o mucho más si come demasiado.

DISTRIBUCIÓN NORMAL
• El estudio de un rasgo cuantitativo en una población granderevela comúnmente, que muy pocos individuos poseenfenotipos extremos y que progresivamente más individuos seencuentran cerca del valor promedio de esa población.
• Este tipo de distribución simétrica tiene, por característica,forma de campana y se llama distribución normal.

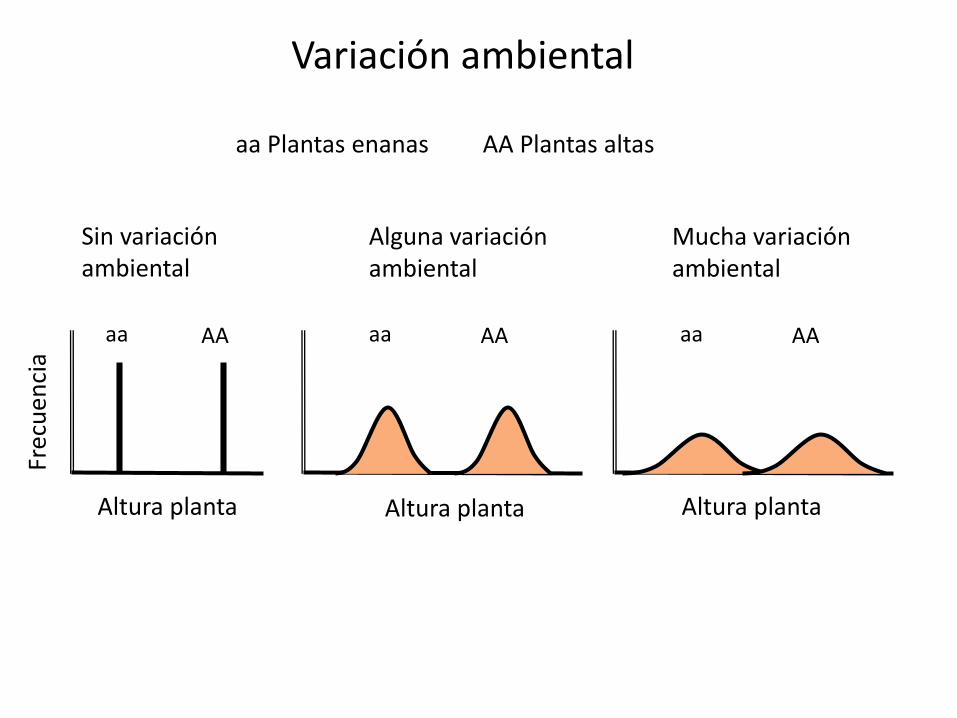
•Experimento Nilsson-Ehle (1909) cruzó dos variedades de trigo puras que diferían en el color de los granos de trigo, rojo y blanco. La F1 era intermedia en color y al cruzarla entre sí obtuvo al menos 7 clases de color en la F2. ¿Cómo explicarlo?
Supongamos control del carácter por un gen con dos alelos sin dominancia
Rojo X BlancoAA aa
Color intermedioAaRojo : Intermedio : BlancoAA Aa aa
1: 2: 1
P
F1
F2

Supongamos control del carácter por dos genes idénticos con dos alelos cada uno, sin dominancia, y donde la intensidad del color rojo depende del número de alelos mayúsculas (que son los que producen el pigmento rojo)
P
F1
F2
Rojo X BlancoAABB aabb
Color intermedio (Rojo medio) AaBb
Rojo oscuro: Rojo medio oscuro : Rojo medio : Rojo claro : Blanco
AABB AaBB AaBb Aabb aabbAABb AAbb aaBb
aaBB
1: 4 : 6: 4 : 1

Mismo supuesto anterior pero con tres genes
Rojo X BlancoAABBCC aabbcc
Color intermedio X Color intermedio (autofecundación)
P
F1
F2
•Fenotipo Rojo -------------> Blanco
•Número alelos que dan color
6 : 5 : 4 : 3 : 2 : 1 : 0•Proporción
1 : 6 : 15 : 20 : 15 : 6 : 1
AaBbCc AaBbCc
Supuestos: los genes segregan independientemente y sus efectos son aditivos
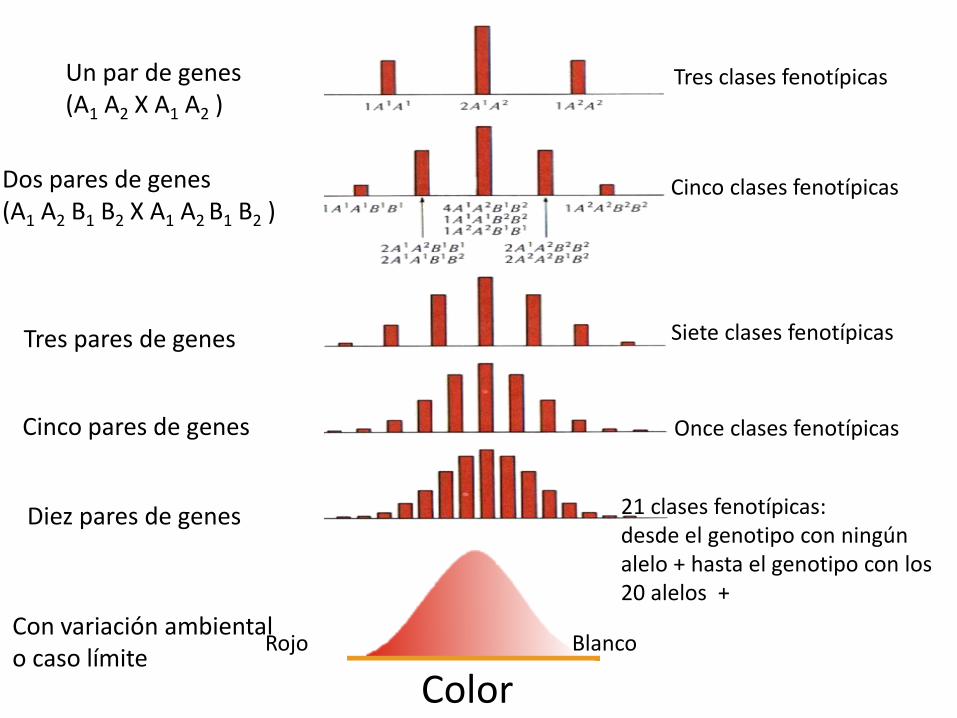
ColorRojo Blanco
Un par de genes(A1 A2 X A1 A2 )
Dos pares de genes(A1 A2 B1 B2 X A1 A2 B1 B2 )
Tres pares de genes
Cinco pares de genes
Diez pares de genes
Tres clases fenotípicas
Cinco clases fenotípicas
Siete clases fenotípicas
Once clases fenotípicas
21 clases fenotípicas: desde el genotipo con ningún alelo + hasta el genotipo con los 20 alelos +
Con variación ambientalo caso límite

ESTADÍSTICA DE LOS CARACTERES CUANTITATIVOS
• Como los caracteres cuantitativos exhiben una distribucióncontinua de fenotipos, no pueden ser analizados de la mismamanera que los caracteres controlados por genes mayores.
• Estos caracteres se describen entonces en términos deparámetros estadísticos.
• Los dos utilizados principalmente son: la media y la varianza.
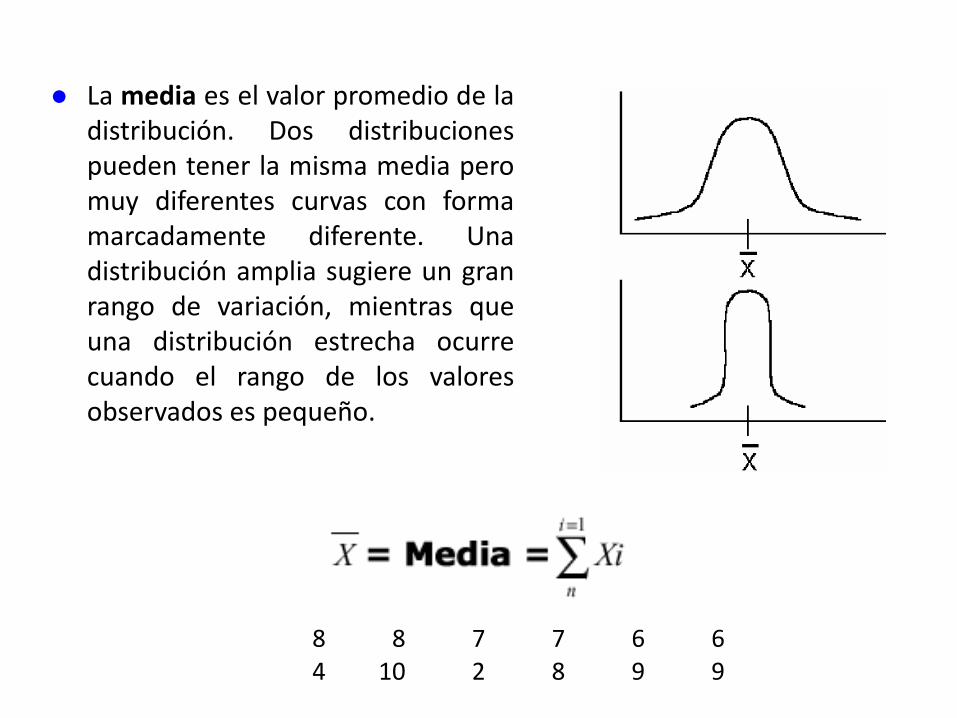
La media es el valor promedio de ladistribución. Dos distribucionespueden tener la misma media peromuy diferentes curvas con formamarcadamente diferente. Unadistribución amplia sugiere un granrango de variación, mientras queuna distribución estrecha ocurrecuando el rango de los valoresobservados es pequeño.
8 8 7 7 6 64 10 2 8 9 9

• La varianza es una medida de la variabilidad o dispersión de la distribución.
La desviación estándar es un parámetro estadístico tambiénrelevante porque se encuentra expresado en las mismas unidades
que la media. Es una medida de dispersión usada enestadística que nos dice cuánto tienden a alejarse los valorespuntuales del promedio en una distribución.


La distribución normal
2)(2
1
2
1)(
x
exf
Dos parámetros determinan la forma de la distribución:
•la media, y
•la desviación estándar (raíz cuadrada de
la varianza 2 = [(x-)2]/n)
•A y B difieren en sus medias (4 y 8)•B y C difieren en sus desviaciones típicas (1 y 0,5)
Función de densidad
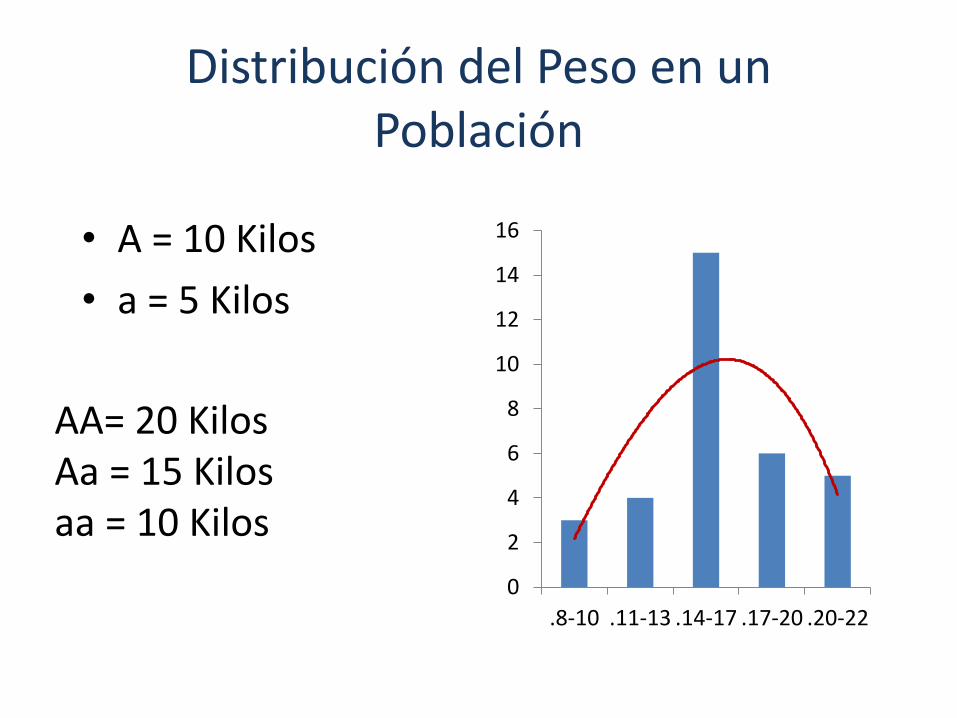
Distribución del Peso en un Población
• A = 10 Kilos
• a = 5 Kilos
0
0.5
1
1.5
2
2.5
20 15 100
2
4
6
8
10
12
14
16
.8-10 .11-13 .14-17 .17-20 .20-22
AA= 20 KilosAa = 15 Kilosaa = 10 Kilos
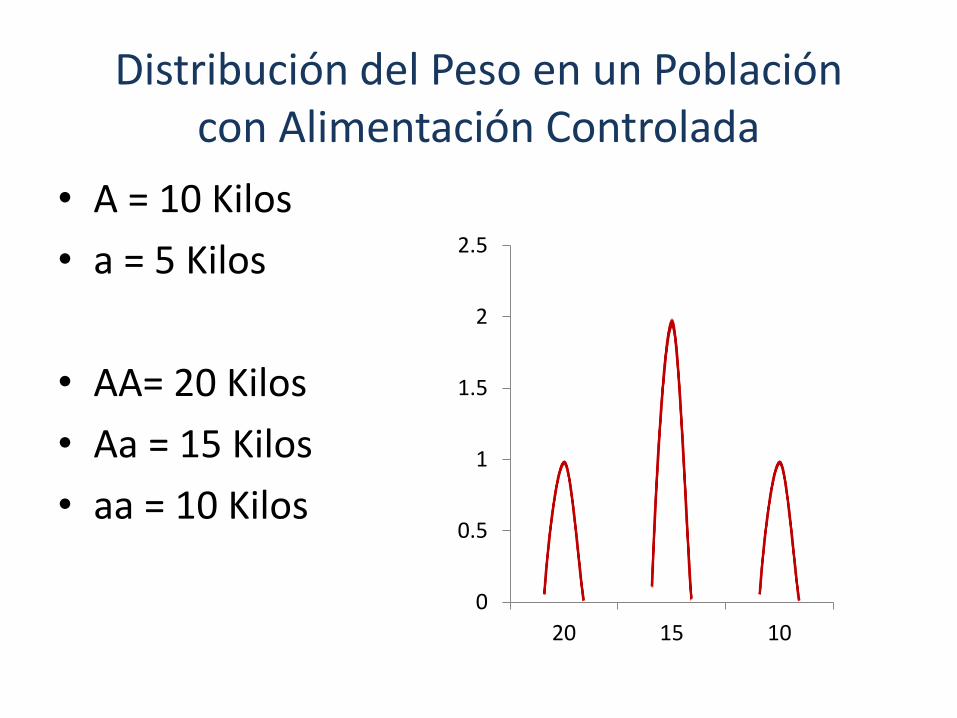
• A = 10 Kilos
• a = 5 Kilos
• AA= 20 Kilos
• Aa = 15 Kilos
• aa = 10 Kilos
0
0.5
1
1.5
2
2.5
20 15 10
Distribución del Peso en un Población con Alimentación Controlada

Heredabilidad de un carácter
Maa MAAMAa
aa Aa AA
•Si distintos genotipos de una población presentan distintas distribuciones para un carácter, decimos que el carácter es heredable
Distribución fenotípica de la población en su conjunto
Valor fenotípico

PARTICIÓN de la VARIANZA y HEREDABILIDAD
• Como sólo se puede medir directamente los valores fenotípicos, elcomponente genético del carácter medido se puede estimar haciendo usode las varianzas. Se puede expresar:
Vp = VG + VE
• Es decir, la variación que se observa en una población para un carácterfenotípico tiene dos componentes, uno genético y otro ambiental.
• Se define como heredabilidad el cociente entre la varianza genotípica y lavarianza fenotípica:
H2 = VG /VP
• Esta fórmula define la heredabilidad como la proporción de la variabilidadde la población que es debida a diferencias genéticas; el resto de lavariación sería debida a la influencia ambiental.

• Puntos muy importantes para recordar sobre laheredabilidad:
1. La heredabilidad de un carácter es una estimación específicade la población y del ambiente que uno está analizando.
2. Esta estimación es un parámetro de una población y no deun individuo.
3. La heredabilidad no indica en qué grado un carácter esgenético, mide solamente la proporción de la VarianzaFenotípica que es el resultado de factores genéticos.

MEDICIÓN DE LA HEREDABILIDAD
• Pueden usarse varios métodos para estimarlas heredabilidades de las característicasmétricas:
1. Componentes de la varianza
2. Semejanza genética de parientes
3. Respuesta a la selección

• El modo más directo es estimar la varianza ambientalextrayendo un conjunto de líneas homocigóticas de lapoblación, cruzándolas y midiendo la varianza fenotípica.
MEDICIÓN DE LA HEREDABILIDAD
• A= 10 Kilos
• a= 5 kilos
0
1
2
3
20 15 10
AA x AA
aa x aa
0
0.2
0.4
0.6
0.8
1
1.2
20 10

COMPONENTES DE LA VARIANZA
• El modo más directo es estimar la varianza ambientalextrayendo un conjunto de líneas homocigóticas de lapoblación, cruzándolas y midiendo la varianza fenotípica.
• Como no hay varianza genética en cada clase genotípica, estasvarianzas proporcionarán (cuando se promedien) unaestimación de la varianza ambiental.
• Se puede restar este valor al de la varianza fenotípica de lapoblación original para obtener la varianza genética.

EJEMPLOEstatura
1.51.81.92.11.51.71.81.91.51.21.52
1.71.81.51.61.71.81.61.5
Estatura1
1.21.31.20.91
1.41.21.21.21
1.21.21.31.11
1.11.21.11.3

EJEMPLO
Población Variación fenotípica
Apareamientos aleatorios VG + VE = 0.045895
Línea pura derivada de la primera VE = 0.016259
• Se calcula VG:VG = Vp– VE = 0.045895-0.016259
VG = 0.029636
• También se calcula H2:
H2 = 0.029636/ 0.045895H2 = 0.6457
H2 = VG /VP

• La mayoría de las características métricas no son altamenteheredables. No se ha definido rígidamente que se consideraalta o baja heredabilidad, pero los siguientes valores seaceptan por lo general.
• La heredabilidad puede tomar valores de 0 a 1. En general seasume que:
VALORES DE LA HEREDABILIDAD

EJEMPLO
1 0.15 8 0.22
2 0.38 9 0.31
3 0.26 10 0.4
4 0.45 11 0.36
5 0.17 12 0.45
6 0.36 13 0.25
7 0.29 14 0.58
0.3310.119
Campo Invernadero
0.3490.078
1 0.34 8 0.29
2 0.38 9 0.32
3 0.27 10 0.58
4 0.33 11 0.33
5 0.28 12 0.4
6 0.31 13 0.38
7 0.3 14 0.37
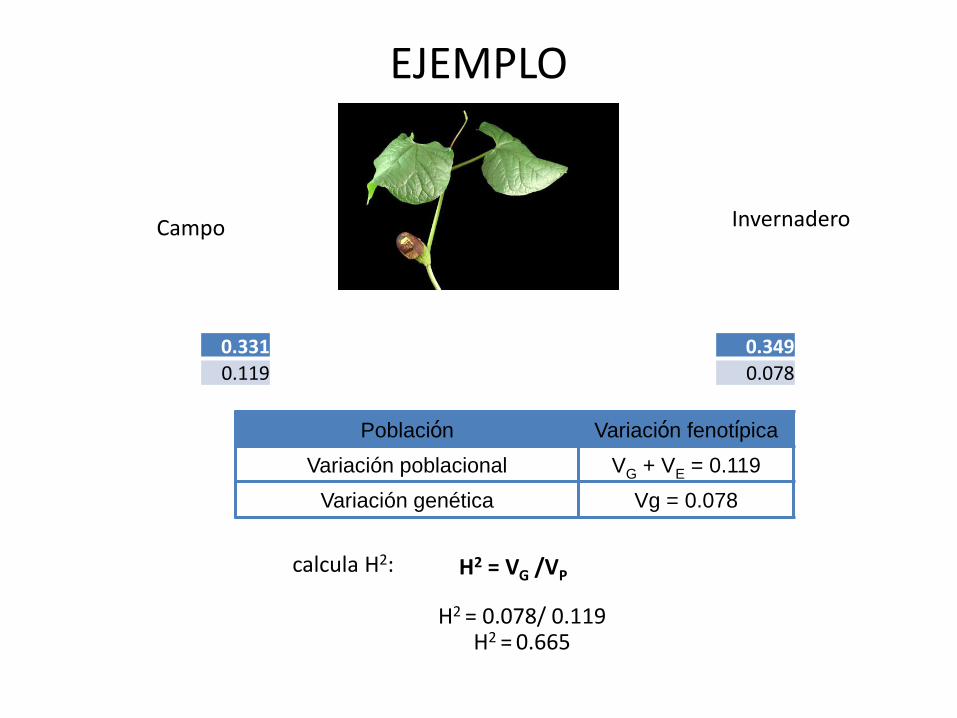
EJEMPLO
0.3310.119
Campo Invernadero
0.3490.078
Población Variación fenotípica
Variación poblacional VG + VE = 0.119
Variación genética Vg = 0.078
calcula H2:
H2 = 0.078/ 0.119H2 = 0.665
H2 = VG /VP

SEMEJANZA GENÉTICA DE PARIENTES• Es necesario construir un número de familias de un grado conocido de
relación.
• El tipo más simple de estructura familiar es la de hermanos. Los parentales se eligen de la población al azar y son cruzados por parejas para producir n número de familias .
• La medición del grado de parecido entre parientes se basa en la partición de la varianza fenotípica en componentes observables.
• Considerando el caso de individuos agrupados en familias de hermanos, la varianza fenotípica se descompone en:
VP = VB + VWdonde:
VW = la varianza de los individuos respecto a las medias dentro de los grupos (W: within families).
VB = la varianza de las medias de los grupos con respecto a la media de la población (B: between families).

1.5 1.8 1.7 1.6 1.6 1.5 1.6
SEMEJANZA GENÉTICA DE PARIENTES
1.8 2.0 1.9 1.8 1.9 2.2 1.9 1.7 1.5 1.9

SEMEJANZA GENÉTICA DE PARIENTES
VP = VB + VW
VP = 0.0388 + 0.0114 VP = 0.0388 + 0.0338
VP = 0.0502 VP = 0.0726
H2 = VG / VP
H2 = 0.0338 / 0.0726H2 = 0.0114 / 0.0502
H2 = 0.2271 H2 = 0.4656

Experimentos de selección artificial para la determinación de la heredabilidad de un carácter cuantitativo
Cruce entre individuos con valores fenotípicos extremos (mismo ambiente)
No heredable
Heredable
Generaciónparental
Media

RESPUESTA A LA SELECCIÓN
• La heredabilidad puede estimarse por selección artificial enanimales y plantas.
• La selección artificial opera cuando el hombre determina acuáles organismos debe permitirse que generendescendencia.
• Se conoce que cuando en una población heterogénea seseleccionan como reproductores individuos de un extremo dela población, sean los de mayor o menor valor fenotípico, losdescendientes muestran una media del carácter que tiende aaproximarse a la media de la generación parental.

• Por ejemplo, en las variedades de plantascultivadas obtenidas a partir de plantas silvestres medianteselección para diversas características.

• La heredabilidad mide la relación entre la media del carácter en lapoblación parental y en la descendiente mediante el cociente:
h2 = X1 – X0 / Xs – X0
• Llamamos X0 a la media del carácter en la generación parental, Xs a lamedia del carácter en los individuos seleccionados, y X1 a la media delcarácter de los descendientes.
• A la diferencia X1 – X0 entre la media de la población parental y la mediade la población seleccionada se le llama respuesta a la selección.
• La diferencia Xs – X0 entre la media de los individuos seleccionados y lamedia de la población parental se llama diferencial de selección.
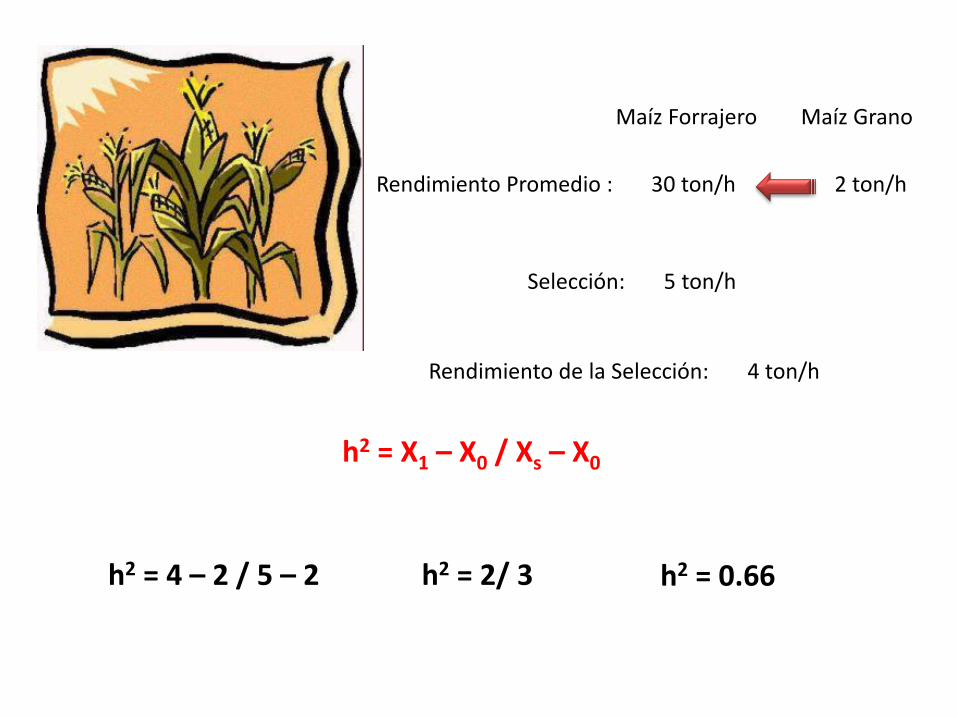
Rendimiento Promedio : 30 ton/h 2 ton/h
Maíz Forrajero Maíz Grano
Selección: 5 ton/h
Rendimiento de la Selección: 4 ton/h
h2 = X1 – X0 / Xs – X0
h2 = 4 – 2 / 5 – 2 h2 = 2/ 3 h2 = 0.66

92 +17 39 50 +13 48 45 +18 +6 *12
88 52 55 67 79 56 48 *11 50 77
*0 23 +15 59 *2 64 72 71 82 92
*4 28 40 82 46 51 85 +16 36 41
+21 94 80 87 +18 54 48 100 *8 *8
•Usted realiza su tesis en recuperación de ecosistemas, en la cual se reforestara una zona
mermada por una plaga, de acuerdo a los datos de invernadero se evaluaron 50 árboles
jóvenes para determinar la susceptibilidad a la plaga, de estos se eligieron dos grupos (+ y *)
para la obtención de simillas, de acuerdo a los datos de resistencia a la plaga de los
descendientes de estos dos grupos, cuáles recomendaría para su cultivo en el campo.
Población original
p 47.+ 16.857.* 6.429

12 9 8 14 12 12 15 8
13 10 13 12 10 11 9 14
14 14 11 13 12 13 14 11
Descendientes del grupo +
Descendientes del grupo *
5 5 8 1 0 6 1 7
8 6 5 5 0 5 6 7
3 9 6 2 5 1 7 5
.+ 11.683
.* 4.708

h2 = X1 – X0 / Xs – X0
p 47.+ 16.857.* 6.429
.+ 11.683
.* 4.708
-30.143 / -35.317 = 0.853 h2 + =
-40.571 / -42.2927 = .959 h2 * =
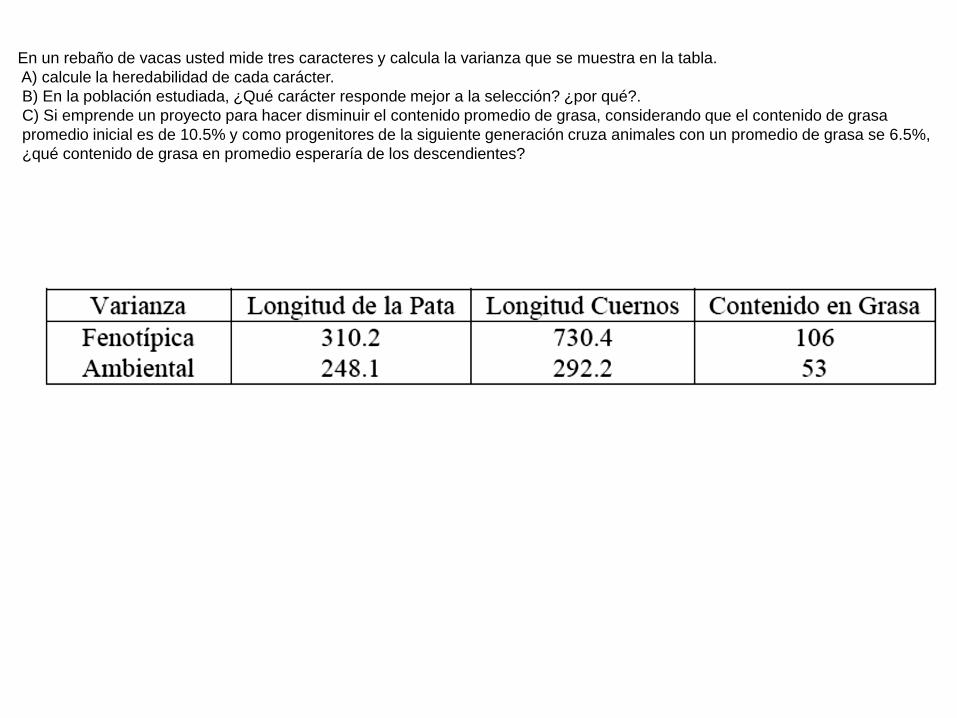
En un rebaño de vacas usted mide tres caracteres y calcula la varianza que se muestra en la tabla.
A) calcule la heredabilidad de cada carácter.
B) En la población estudiada, ¿Qué carácter responde mejor a la selección? ¿por qué?.
C) Si emprende un proyecto para hacer disminuir el contenido promedio de grasa, considerando que el contenido de grasa
promedio inicial es de 10.5% y como progenitores de la siguiente generación cruza animales con un promedio de grasa se 6.5%,
¿qué contenido de grasa en promedio esperaría de los descendientes?

Genética De Poblaciones
"Where Do We Come From? What Are We? Where Are We Going?" [1897] by Paul Gauguin

¿Cómo se manifiestan los alelos en una población?

Genética De Poblaciones

• El estudio de las diferencias genéticas que se dan de forma natural entre los organismos.
• Las diferencias genéticas entre los organismos DENTRO de una especie se denominan polimorfismos genéticos.
• Las diferencias genéticas que se acumulan ENTRE especies constituyen la divergencia genética.
• La genética de poblaciones es el estudio del polimorfismo y la divergencia.
Genética De Poblaciones

Objetivos
• Entender las leyes de Hardy-Weinberg y su aplicación en el estudio de genética de poblaciones.
• Conocer los diferentes mecanismos en la herencia de carácteres en una población.
• Emplear las leyes de Hardy-Weinberg para hallar frecuencias alélicas, genotípicas y fenotípicas.

Definición:
• Estudio de la herencia colectiva y la variación en los organismos que habitan un área o región.

Genética de poblaciones
• La segregación y variabilidad en la población está
gobernada por las Leyes Mendelianas. (Ley de
dominancia, Ley de segregación y Ley de segregación independiente).
– Se asume que los individuos contribuyen igualmente al “pool genético” y tienen la misma oportunidad de reproducirse.
– La frecuencia de los genes y sus alelos tienden a mantenerse constante por generaciones.• Se deduce que los cruces son al azar, no por selección.

La problemática de la genética de poblaciones es la descripción y explicación de la variación genética dentro y entre poblaciones
Theodosious Dobzhansky
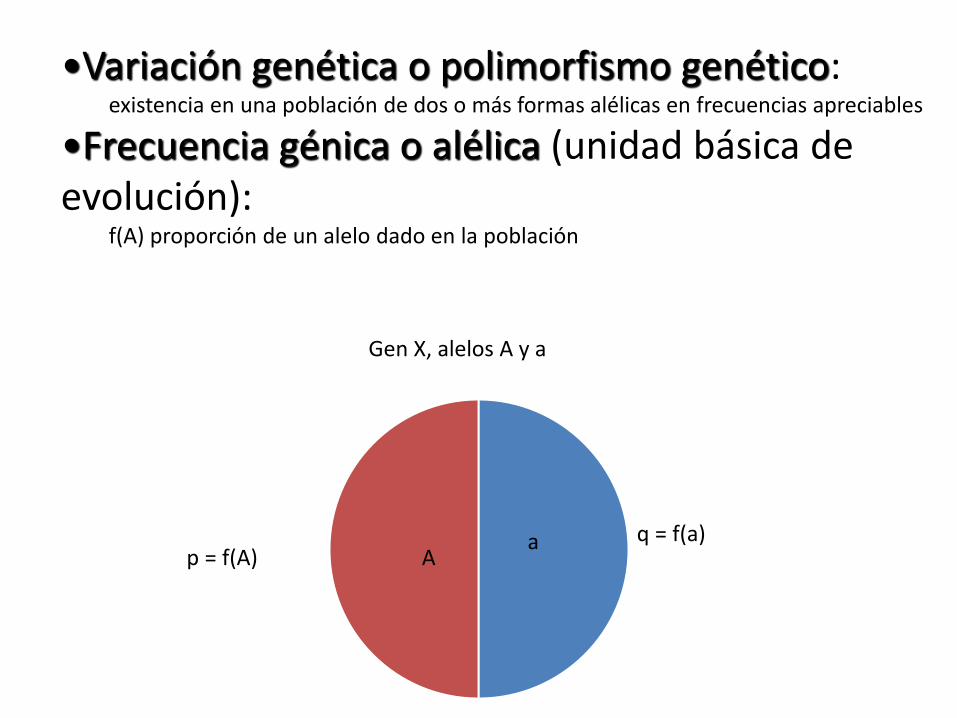
•Variación genética o polimorfismo genético: existencia en una población de dos o más formas alélicas en frecuencias apreciables
•Frecuencia génica o alélica (unidad básica de evolución):
f(A) proporción de un alelo dado en la población
q = f(a)
Gen X, alelos A y a
Aa
p = f(A)

La Genética de Poblaciones es una Teoría de Fuerzas
p = f(A)
Deriva genética
Selección natural
Mutación
Migración
Factores que cambian las frecuencias génicas en las poblaciones

Teorías de la variación en los 60
•Ausencia de variación
•Selección purificadora
•Genotipo silvestre es óptimo
•Muller (laboratorio)
•Eugenesia
•Variación ubicua
•Selección equilibradora
•No existe un genotipo silvestre
•Dobzhansky (naturalista)
•¡Viva la diversidad!, no interferencia
Teoría clásica Teoría equilibradora

Ley de Hardy - Weinberg• Establece que la frecuencia de un alelo y las
frecuencias genotípica de una población tienden apermanecer igual por generaciones.
• Si ocurre algún cambio en la frecuencia indica que ha ocurrido evolución.
[p+q]2 = p2 + 2pq + q2 = 1

Esperma
Ovulos
AAp2
Aapq
Aapq
aaq2
A
pa
qa q
A p
Frecuencias alélicas
Los supuestos implican una unión aleatoria de los alelos para formar genotipos

Ley de Hardy-Weinberg (cont.)
• Para sacar la frecuencia del alelo A
– p(A)= [p2+½(2pq)] / (p2 + 2pq + q2)
• Para sacar la frecuencia del alelo a
– q(a)= [q2+½(2pq)] / (p2 + 2pq + q2)

Predicción de frecuencias
• Para poder predecir las frecuencias genotípicas, frecuencia de un gen o frecuencia fenotípica de una población se puede hacer solo si se conoce cómo se hereda la característica a estudiarse.

0 0,1 0,2 0,3 0,4 0,5 0,6 0,7 0,8 0,9 1,0
0 0
,1 0
,2 0
,3 0
,4 0
,5 0
,6 0
,7 0
,8 0
,9 1
,0
p = f(A)
Frec
uen
cia
2pq (Aa)
p2 (AA)q2 (aa)
Gráfico de p2, 2pq y q2.

Genotipo
MM MN NN TotalN. individuos 1787 3037 1305 6129
N. alelos M 3574 3037 0 6611
N. alelos N 0 3037 2610 5647
N. alelos M + N 3574 6074 2610 12258
Prueba de ajuste a Hardy-Weinberg
Frecuencia alélica M = 6611/12258 = 0,53932 = pFrecuencia alélica N = 5647/12258 = 0,46068 = q
Frecuencia esperada p2 = 0,2908 2pq = 0,4969 q2 = 0,2122 1,000
Número esperada 1782,7 3045,6 1300,7 6129(Frecuencia X 6129)
84,3
04887,0)(
2
..1;05,0
22
lg
esperadonúmero
esperadonúmeroobservadonúmeroX

Tabla 2

Codominancia o dominancia incompleta
• Codominancia - los alelos producen efectos independientes en forma heterocigota
• Ej. Tipo de sangre AB
• Dominancia incompleta – hay expresión de dos alelos en un heterocigoto que lo hace diferente (de fenotipo intermedio)a los parentales homocigotos.

Codominancia (ejemplo):• Antígenos de la serie M-N en los eritrocitos humanos:
Población total: 200 personas
58 tipo M
101 tipo MN
41 tipo N
p(LM)= 58 + (½) 101 / 200 = 0.543
q(LN)= 41 + (½) 101 / 200 = 0.458
Al expandir el binomio:
[p(M)+q(N)]2 = 0.294LMLM+ 0.496LMLN + 0.209LNLN
Si se multiplica cada una de las frecuencias x200, vemos que se acercan a
los valores observados: .294x200=58.8 , .496x200=99.2 , .209x200=41.8

Dominancia Completa• En este caso los individuos heterocigotos no se pueden
diferenciar de los homocigotos dominantes.
• Ejemplo:
Asumiendo que la presencia del antígeno Rh (Rh+) se debe a un alelo dominante ‘‘R’’ y que la ausencia del antígeno (Rh-) se debe al alelo recesivo ‘‘r’’.
Un genotipo Rr y RR producen Rh+, mientras que rrproduce Rh-.

Dominancia Completa (ejemplo):• Se tomaron 100 personas al azar de una población y se obtuvieron:
25 Rh- (ausencia del antígeno Rh)
75 Rh+ (presencia del antígeno Rh)
La frecuencia de r se estima: p2 + 2pq + q2
q2 (rr)= 25/100= .25 q (r) = √.25 = 0.5
Si:
p+q = 1 1- q = p 1 – 0.5 = 0.5
La frecuencia estimada de los genotipos RR y Rr son:
p2 (RR)= (0.5) 2= 0.25, 2pq(Rr)= 2(0.5)(0.5)= 0.50
Y el porcentaje de cada uno de ellos: 25 RR y 50 Rr.

Alelos Múltiples
• En el caso en que un gen en particular se encuentra en tres o más formas alélicas en una población.
• Para los genes con múltiples alelos las proporciones de la Ley H-W se expanden:
(p+q+r)2= p2+q2+r2+2pq+2qr+2pr

Serie ABO en tipos de sangre
Tipo de sangre
GenotipoAntígenos presentes
Frecuencia fenotípica observada
Frecuencia fenotípica esperada
A IAIA, IAi A A p2 + 2pr
B IBIB, IBi B B q2 + 2qr
AB IAIB AB AB 2pq
O ii ninguno O r2

Alelos Múltiples (ejemplo):
Fenotipo Observados Frecuencia fenotípica
A 207
B 71
AB 21
O 301
total 600
Se encuestaron 600 estudiantes para saber su tipo de sangre, y obtuvieron los siguientes resultados:
Fenotipo Observados Frecuencia fenotípica
A 207 A =207/600=0.345
B 71 B =71/600=0.118
AB 21 AB =21/600=0.035
O 301 O =301/600=0.502
total 600 1.00

Frecuencias alélicas:
r(i)=√ o =√0.502 =0.708
p(IA)=1-√B+O = 1- √0.118+0.502 = 0.213
q(IB)=1-√A+O = 1- √0.345+0.502 = 0.080
p2+2pr = 0.045 + 0.301=0.346 x 600 personas = 207.8 (TipoA)
q2+2qr = 0.0064+0.113=0.119 x600 personas = 71.6 (TipoB)
2pq = 2[(0.213)(0.080)]=0.0340 x 600 personas = 20.4 (TipoAB)
r2 = 0.502 x600 personas = 301.2 (TipoO)

Genes ligados a X
• Se refiere a genes que se encuentran en el cromosoma X.
• Un ejemplo lo es la condición de hemofilia, la cual se transmite por un gen recesivo (Xh).
• La frecuencia del alelo se estima utilizando la frecuencia del fenotipo en hombres (hemicigotos) en la población.
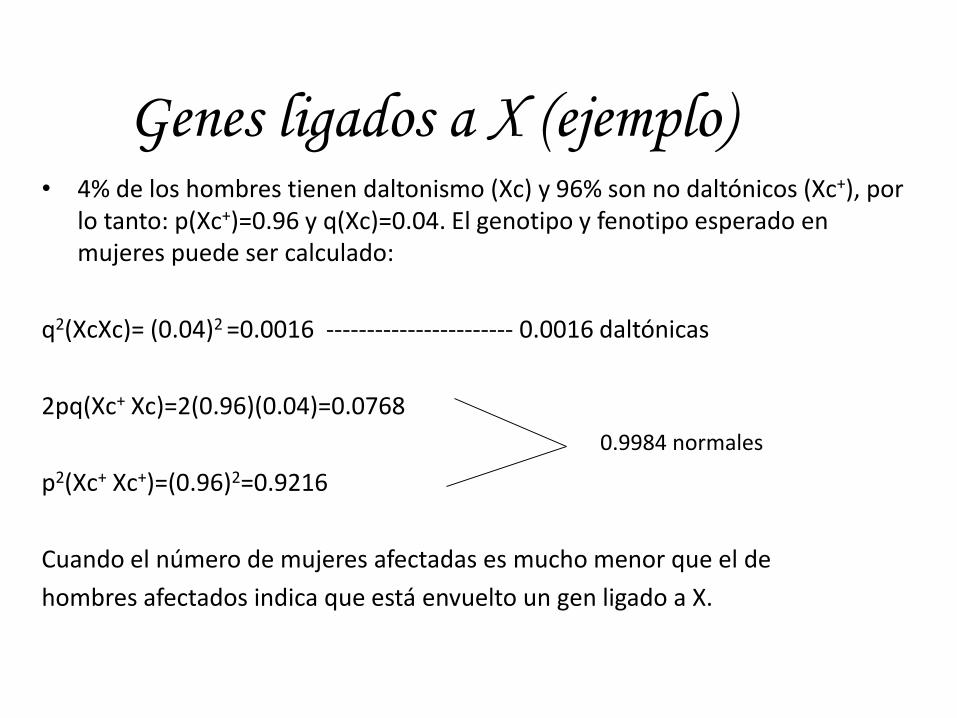
Genes ligados a X (ejemplo)• 4% de los hombres tienen daltonismo (Xc) y 96% son no daltónicos (Xc+), por
lo tanto: p(Xc+)=0.96 y q(Xc)=0.04. El genotipo y fenotipo esperado en mujeres puede ser calculado:
q2(XcXc)= (0.04)2 =0.0016 ----------------------- 0.0016 daltónicas
2pq(Xc+ Xc)=2(0.96)(0.04)=0.0768
p2(Xc+ Xc+)=(0.96)2=0.9216
Cuando el número de mujeres afectadas es mucho menor que el de
hombres afectados indica que está envuelto un gen ligado a X.
0.9984 normales

Condiciones necesarias para mantener el
equilibrio de H-W• Mutaciones
– Asumen que no hay mutaciones
– No es muy significativo ya que normalmente estas ocurren en el orden de 1x10-5 o 1x10-6.
• Migración
– Asumen que no hay migración.
– Si ocurre migración se pueden introducir nuevos genes a la población, puede ocurrir variabilidad.

Condiciones necesarias para mantener el
equilibrio de H-W• Selección
– Asume que no hay selección.
– Pero en la “vida real” algunos genotipos tienen mayor “preferencia” para reproducirse que otros.
• Deriva genética
– Asume que no hay cambios en la frecuencia alélica debido a fluctuación al azar.
– Asume que las poblaciones son grandes.

Condiciones necesarias para mantener el
equilibrio de H-W
• Todos los individuos se cruzan.
• Todos producen la misma cantidad de hijos.
– Si una o todas estas condiciones ocurren en una población no hay evolución.
– Este no es el caso de las poblaciones en la naturaleza.

Condiciones para mantener una población en equilibrio:
• No mutaciones
• No emigración o inmigración
• No preferencias genotípicas o fenotípicas
• Sorteo independiente de alelos
P 18 P 6 P 12 - P 12 +
R %
r %