Fosforilacion oxidativa
35
DOCENTE Q.F. Lázaro Alarcón Galvez CURSO Bioquímica ALUMNO CONTRERAS MOGOLLON HANS J Lambayeque, Setiembre del 2012
Transcript of Fosforilacion oxidativa

DOCENTE
Q.F. Lázaro Alarcón Galvez
CURSO
Bioquímica
ALUMNO
CONTRERAS MOGOLLON HANS J
Lambayeque, Setiembre del 2012


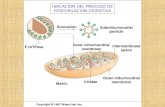
La fosforilación oxidativa es un proceso metabólico
que utiliza energía liberada por la oxidación de
nutrientes para producir adenosín trifosfato (ATP).
Se le llama así para distinguirla de otras rutas que
producen ATP con menor rendimiento, llamadas "a
nivel de sustrato". Se calcula que hasta el 90% de la
energía celular en forma de ATP es producida de
esta forma.

El descubrimiento de Albert
Lehninger en 1943 de que en
las mitocondrias se realiza la
fosforilación oxidativa en los
eucariotas marcó el inicio de
la fase moderna de los estudios
sobre las transducciones
biológicas de energía.

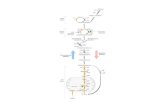
La fosforilación oxidativa empieza con la entrada de
electrones en la cadena respiratoria. La mayor parte
de dichos electrones provienen de la acción de
deshidrogenasas que captan electrones de vías
catabólicas y los canalizan hacia aceptores
universales de electrones: nucleótidos de
nicotinamida (NAD* o NADP*) o nucleótidos de
flavina (FMN o FAD).

Los transportadores de electrones de la cadena
respiratoria están organizados en complejos
supramoleculares incrustados en membranas que se
pueden separar físicamente. El tratamiento suave de
la membrana mitocondrial interna con detergentes
permite la resolución de cuatro complejos distintos de
transportadores electrónicos, siendo cada uno de ellos
capaz de catalizar la transferencia electrónica a
través de una porción de la cadena.

COMPLEJO I: NADH A UBIQUINONA:
también llamado NADH: ubiquinona
oxidorreductasa o NADH deshidrogenasa, es un
enzima enorme compuesto por 42 cadenas
polipeptídicas diferentes.
COMPLEJO II: SUCCINATO A
UBIQUINONA. Es la única enzima del ciclo
cítrico ligado a membrana. Contiene cinco
grupos prostéticos de dos tipos y cuatro
subunidades prostéticas diferentes.


COMPLEJO III: UBIQUINONA A CITOCROMO
C: acopla la transferencia de electrones desde el
ubiquinol al citocromo c con el transporte vectorial
de protones de la matriz al espacio intermembrana.
COMPLEJO IV: CITOCROMO C A O2. En el
último paso de la cadena respiratoria, el Complejo
IV, también llamado citocromo oxidasa, transporta
electrones desde el citocromo c al oxígeno
molecular, reduciéndolo a H2O. El Complejo IV es
un enzima muy grande de la membrana
mitocondrial interna.

El modelo quimiosmotico (Peter Mitchell) : según el modelo
la energía electroquímica inherente a la diferencia en la
concentración de protones y ala separación de cargas a
través de la membrana mitocondrial interna, la fuerza
protón – motriz, impulsa la síntesis de ATP a medida q los
trotones fluyen de manera pasiva de regreso a la matriz a
través de un poro que se encuentra asociado a la ATP
sintasa.
Quimiosintetico: son las reacciones enzimáticas en las que
intervienen simultáneamente, una reacción química y un
proceso de transporte. Cuando se suspenden mitocondrias
aisladas en un tampón que contiene ADP, Pi


Experimento de intercambio de18O. F1solubilisado a partir de
membranas mitocondriales se
incuba ATP en presencia de
agua marcada con 18O. A
intervalos de toma una muestra
de la solución y se analiza la
incorporación de 18O. lo que
indica que tanto la hidrólisis
como la síntesis de ATP han
tenido lugar varias veces
durante la incubación

Probable complejo del periodo de transición para la síntesis e hidrólisis
de ATP en la ATP sintasa. la subunidad se muestra en verde; en
gris.los residuos cargados positivamente -Arg182 y -Arg176
coordiProbable complejo del periodo de transición para la síntesis e
hidrólisis de ATP en la ATP sintasa. la subunidad se muestra en verde;
en gris.los residuos cargados positivamente -Arg182 y -Arg176
coordinan dos oxígenos del intermediario fosfato pentacovalente; -Lys135
interacciona con un tercer oxigeno y el ion Mg2+ (esferas verdes)
estabelcienda aun más el intermediario las esferas establecida aun así el
intermediario. Las esferas azules representas el grupo saliente (salientes)
aun mas en estas intersc tienen cono te resulta como resultado un rápido
equilibrado de ATP y ADP y A DEP +p1 en el sitio activa nan dos
oxígenos del intermediario fosfato pentacovalente; -Lys135 interacciona
con un tercer oxigeno y el ion Mg2+ (esferas verdes) estableciendo aun
más el intermediario las esferas establecida aun así el intermediario. Las
esferas azules representas el grupo saliente (salientes) aun mas en estas
intersc tienen cono te resulta como resultado un rápido equilibrado de
ATP y ADP y A DEP +p1 en el sitio activa


La energía conservada en un gradiente de protones puede
impulsar al transporte de solutos contra gradiente a través de
una membrana





El ATP sintasa lleva a
cabo catálisis
rotacional en la que el
flujo de protones a
través de F0 hace q
cada uno de los tres
sitios de unión de
nucleótidos de F1 cicle
desde la conformación
que une (ADP + Pi) a
la que une ATP y
finalmente a la vacía

La proporción del ATP
sintetizado por 1/2 O2
reducido a agua H2O
(la razón P/O) es unos
2,5 cuando los
electrones entran en la
cadena respiratoria en
el complejo I y de 1,5
cuando los electrones
entran en CoQ

La formación del
ATP por enzima
requiere poca
energía; el papel de
la fuerza protón-
motriz es empujar
el ATP fuera de su
sitio de unión

La membrana mitocondrial interna es impermeable
al NADH y al NAD-, pero se transportan
equivalentes de NADH desde el citosol a la matriz
por una de dos lanzaderas. Los equivalentes de
NADH que se transporten del malatoaspartato
entran en la cadena respiratoria por el complejo I
dando una razón P/O de 2,5; los que se transportan
por la lanzadera del glicerol 3-fosfato entran por el
CoQ y dan una razón P/O de 1,5


Fosforilación Oxidativa
Sintetiza ATPEn Cel.
Aeróbicas
1 mol Glucosa a CO2 = 30 o
32 ATP

La tasa respiratoria (consumo de O2) en las mitocondrias está
bajo una regulación precisa, está limitada generalmente por la
disponibilidad de ADP como sustrato de la fosforilación. La
dependencia de la velocidad de consumo de O2 con respecto a
la concentración de ADP del aceptor de Pi, denominada control
por aceptor de la respiración, puede ser espectacular. En
algunos tejidos animales la razón del control por aceptor, la
proporción de la velocidad máxima de consumo de O2 inducida
por el ADP a la velocidad basal en ausencia de ADP, es al
menos de 10.

Cuando una célula esta isquémica (privada de oxigeno), tal como
sucede en un ataque de corazón o una embolia, cesa la
transferencia de electrones al oxígeno y cesa también el bombeo
de protones. La fuerza protón – motriz desaparece al cabo de
poco. Esto no se produce gracias a un inhibidor proteico pequeño
(84 aminoácidos), IF1, que simultáneamente a dos moléculas de
ATP sintasa inhibiendo su actividad ATPasa. IF1 solo es
inhibidor de forma dimerica, que está favoreciendo a un pH
inferior a 6.5. En una célula sin oxígeno, la principal fuente de
ATP es la glucolisis, con lo que los ácidos pirúvico o láctico así
formados disminuyen en pH en el citosol y la matriz mitocondrial.


El resultado de este cortocircuito
de protones es que la energía de
oxidación no se conserva mediante
la formación de ATP sino que se
disipa en forma de calor, lo que
contribuye al mantenimiento de la
temperatura corporal del recién
nacido. Los animales hibernantes
también dependen de las
mitocondrias desacopladas del
tejido adiposo marrón para
generar calor durante largos
periodos de letargo.

Las mitocondrias contienen su propio genoma, una molécula circular de
ADN de doble cadena. Cada una de las cientos o miles de mitocondrias
de una célula tiene cinco copias de este genoma. Hay 37 genes (16 569
pares de bases) en el cromosoma mitocondrial humano, incluidos 13 que
codifican proteínas de la cadena respiratoria: los restantes genes
codifican moléculas de rRNA y tRNA esenciales para la maquinaria
sintetizadora de proteínas de la mitocondria.

La fotosíntesis es el proceso
metabólico primario de las plantas y
de hecho se considera el primario en
la biosfera, es así debido a que
permite utilizar la energía radiante
del sol para transformarla en energía
química de ATP, que es necesaria
para la de carbohidratos y de hecho
para el anabolismo en general. Esta
capacidad solo la desarrollan las
plantas ya que cuentan con células
parenquimatosas clorofilianas, las
cuales se encuentran en las
hojas, partes verdes de tallos, flores y
frutos verdes.

Fot
ofos
fori
laci
on
Fotofosforilacion
Fotosistema I
Fotosistema II
Reducción CO2

En 1954, Arnon demostró que la
incidencia de luz sobre
cloroplastos, poseyendo estas cantidades
suficientes de ADP y grupos fosfato, da
lugar a la síntesis de ATP, siendo
proporcional su cantidad al flujo
electrónico inducido por luz. Como se ha
mencionado este proceso depende de la
luz, además al igual que la fosforilación
oxidativa, requiere una membrana
tilacoide intacta y puede desacoplarse del
transporte electrónico, el cual es
impulsado por la luz por compuesto tales
como 2,4-dinitrofenol.

• Fotosistema I: En este fotosistema un electrón es captado por una
molécula aceptara de electrones, o aceptora primaria, el cual se
desplaza a través de una serie de reacciones de tipo redox hasta
llegar a la fijación del electrón a la coenzima NADP.
• Fotosistema II Este se encuentra en la membrana tilacoide de los
cloroplastos, es una proteína intrínseca al igual que la cadena
transportadora d electrones, además de ser el lugar donde el agua
sufre una oxidación.

Dentro de esta etapa
encontramos el ciclo de
Calvin-Benson, para el
cual la fuente de
energía para la ruta
son el NADPH y el ATP
producidos durante las
reacciones de la
fotosíntesis
dependientes de la luz.
e glucosa