Clase 10 Y 11 Ciclo De Krebs Transporte Electronico Y Fosforilacion Oxidativa
Upload
geriatriauatCategory
view
5.536download
1

La bioenergética describe la transferencia
y la utilización de energía en los sistemas
biológicos.
Se utiliza unas cuantas ideas básicas del
campo de la termodinámica, en particular el
concepto de la energía libre.

La bioenergética tiene que ver con los
estados energéticos inicial y final,
componentes de una reacción, y no con el
mecanismo o el tiempo necesario para que
ocurra el cambio químico. La bioenergética
predice si es posible un proceso, en tanto
que la cinética mide la rapidez con que
ocurre la reacción.

Energía LibreLa dirección y la extensión a las que procede una reacción química
depende del grado al que cambian dos factores
durante esa reacción.Son: f) Entalpía: es una medición del cambio en el contenido de
calor de los reactivos y los productosb) Entropía es una medición del cambio de la reacción al azar
entre los reactivos y los productos.

Cambio de energía libre:
Produce en dos formas, delta G y delta G°.
Delta G, es la mas general porque
predice el cambio de la energía libre
y, por este motivo, la dirección de
una reacción a cualquier concentración
especifica de productos y reactivos.

La energía libre estándar, delta G°, es el
cambio de energía que ocurre cuando
reactivos y productos se encuentran en una
concentración de 1 mol/L.

El signo de G predice la dirección de una reacción
El cambio de la energía libre, delta G ( G),
se puede emplear para predecir la dirección
de una reacción a temperatura y presión
constantes. Considérese la reacción:
A B

1. Delta G negativo
Si delta G es un numero negativo habrá
perdida neta de energía y la reacción
no ira espontánea. A se convierte en B.
Esta es una reacción exergonica.

2. Delta G positiva
Si delta G es un numero positivo habrá
ganancia neta de energía y la reacción
no ira espontáneamente desde B hacia A.
Esta es una reacción endergonica.

3. Delta G tiene valor cero
Si delta G = 0 , los reactivos estarán en
equilibrio.

Delta G de las reacciones hacia adelante y hacia atrás
La energía libre de la reacción hacia
adelante (AB) es igual en magnitud, pero de signo opuesto a la reacción hacia (BA).
Delta G depende de la concentración de reactivos y productos
Delta G de la reacción A B depende de la concentración del reactivo y el producto.

Delta G° es el cambio estándar de energía
libre, R es la constante de los gases
(1.987 cal/mol-grado), T es la temperatura
absoluta (°K). [A] y [B] son la
concentraciones reales de reactivos y
producto. In representa al logaritmo natural.

La reacción en la que la concentración del reactivo, glucosa 6-fosfato, es mayor en comparación con la concentración del producto, factura 6-fosfato es grande y negativa, lo que hace que delta Gº sea negativa a pesar de que delta Gº es positiva.

Cambio de energía libre estándar, delta Gº Delta Gº se denomina
cambio de energía libre estándar porque es igual al cambio de energía libre, delta G, bajo condiciones estándar; esto es cuando los reactivos y los productos se conservan a la concentración de 1 mol/L.

Delta Gº tiene valor de predicción solo bajo condiciones estándar
Bajo condiciones estándar delta Gº puede emplearse para predecir la dirección en que procederá una reacción porque, en esta situación delta Gº es = delta G, delta Gº no puede predecir la dirección que seguirá la reacción bajo condiciones fisiológicas porque esta compuesta solo por constantes (R, T y Keq) no se altera con los cambios de la concentraciones del producto o sustrato.

Relaciones entre delta Gº y K eq
En una reacción A B, se llega a un punto de equilibrio en el que ocurre ningun cambio quimico, esto es cuando A se convierte B co igual rapidez como la que ocurrre cuando B se convierte en A. En este estado la razon [B] con respecto [A] es constante independientemente de las concetraciones reales de los dos compuestos

Los cambios de energía libre estándar( G°)
son adquiridos en cualquier secuencia de
reacciones consecutivas, como lo son los
cambios de energía libre ( G).
Por ejemplo:Glucosa+ATP glucosa6-fosfato+ATP G° =4000cal/mol
Glucosa6-fosfatofructosa6-fosfato G°=+400cal/mol
Glucosa + ATPfructosa6-fosfato+ADP G°=-3600cal/mol
3. Los valores delta G° de dos reacciones consecutivas son aditivos

4. Los delta G de una vía son aditivos:
Es muy importante en la vías bioquímicas
por las que los sustratos deben pasar en una
dirección particular (por ejemplo, ABCD..).
La suma de los deltas G de las reacciones
individuales sean negativos, la vía pude proceder
potencialmente como esta escrita incluso si
algunas reacciones componentes individuales de
esta tengan un delata G positivo.

ATP COMO TRANSPORTADOR DE ENERGIA
Un engrane que tiene
una pesa montada gira
en la dirección que
logra el estado mas
bajo de energía; en este
caso la pesa busca si
posición mas bajo.

El movimiento inverso
no se ve favorecido
desde el punto de vista
energético y no se produce
de manera favorecido
desde el punto de vista
energético y no se produce
de manera espontánea.

Puede emplearse el movimiento favorecido desde el punto de vista energético de un engrane para hacer girar a un segundo engrane en una dirección en la que no se movería de manera espontánea.

A. Las reacciones se acoplan mediante intermediarios en común
Dos reacciones químicas tiene un
intermediario en común cuando se producen
de manera sucesiva de modo que el
producto de la primera es un sustrato para
la segunda.

B. Energía transportada por el ATPEl ATP se constituye por
una molécula de
adenosina (adenina + ribosa) a la que se
unen tres grupos fosfatos. Se quita uno de
los fosfatos, el resultado será monofosfato
de adenosina (AMP).

CADENA DE TRANSPORTE DE ELECTRONESLas moléculas ricas en
energía, como glucosa, se
metabolizan por medio de una serie de
reacciones de oxidación que producen en ultima
instancia CO2 y agua.

Los intermediarios metabólicos de esta reacciones donde electrones a coenzimas especificas, como dinucleótido de nicotinamida y adenina (NAD+) y dinucleótido de adenina y flavina (FAD) para formar la coenzimas reducidas ricas en energía NADH y FADH2.

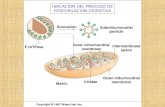
A. Mitocondria
La cadena de transporte de electrones se
encuentra en la membrana mitocondrial
interior, y es la vía final común por laque
fluyen los electrones derivados desde los
diferentes combustibles del cuerpo hacia el
oxigeno.

1. Estructura de la mitocondria
La membrana exterior contiene poros especiales que la vuelven libremente permeable ala mayor parte de los iones y las moléculas pequeñas, la membrana interior de la mitocondria es una estructura especializada impermeable a la mayor parte de los iones pequeños, como H+, Na+, y K+, moléculas pequeñas como ATP, ADP, piruvato y otros metabolitos importantes para la función de la mitocondria.

2. Complejas de sintasa de ATP
Estos complejos de proteínas se denominan
partículas membranosa interiores, y se
adhieren a la superficie interior de la
membrana mitocondrial interior.

3. Matriz de la mitocondria
La parte interior de la mitocondria esta constituida en 50% por proteínas. Estas moléculas son las enzimas encargadas de la oxidación del piruvato, aso como de aminoácidos, ácidos grasos y las que participan en el ciclo del acido tricarboxilico (TCA).Esta matriz contiene NAD+ y FAD y ADP y F¡, que emplean para producir ATP.

Organización de la cadenaLa membrana mitocondrial interior se puede desintegrar en cinco
complejos enzimáticos separados, designados con los números romano I a V. Cada complejo acepta o dona electrones a transportadores de estos relativamente móviles, como coenzima Q y citocromo c. Cada transportador de esta cadena puede recibir electrones de un donador de estos y donarlos de manera subsecuente al siguiente transportador en la cadena.

C. Reacciones de la cadena de transporte de electrones
Con excepción de la coenzima Q, todos los miembros de esta cadena son proteínas. Pueden funcionar como enzimas, como sucede en el caso de las deshidrogenasas, pueden contener hierro como parte de una centro de hierro y azufre, coordinarse por un anillo de porfirina, como sucede en el caso de los citocromos, o contener cobre, como el complejo de citocromo a + a3.

1. Formación de NADH
El NAD+ se reduce hasta NADH por acción
de las deshidrogenasas que quitan dos
átomos de hidrogeno a sus sustrato. Se
transfieren ambos electrones pero solo un
patrón al NAD+, como lo que se forman
NADH y un protón libre H9+.

2.Deshidrogenasa del NADH
Este complejo tiene una molécula firmemente unida de mononucleótido de flavina (FMN), coenzima relacionada desde el punto de vista estructura con el FAD que acepta los dos átomos de hidrogeno con lo que se convierte en FMNH2.

Coenzima Q
Es un derivado de la quinona con una cola isoprenoidea larga. Se llama también ubiquinona porque se ubica en los sistemas biológicos.

son citocromos los miembros restantes de la cadena de trasporte de electrones.cada uno tiene un grupo hem.
Citocromos

Citocromo a + a3
este complejo del citocromo es el único transportador de electrones en el que el hierro del hem tiene un ligando libre que puede reaccionar directamente con oxigeno molecular.

Inhibidores específicos de sitiose han identificado los inhibidores
específicos se sitio de trasporte de electrones.

Descarga de energía libre durante el transporte de electrones:Se descarga energía libre
conforme se transfieren electrones a lo largo de la cadena de trasporte de estos por un donador (agente reductor) a un aceptador (agente oxidante).

Pares redox
la oxidación (perdida de electrones) de un compuesto se acompaña siempre de reducción (ganancia de electrones) de una segunda sustancia.

Potencial de reducción estándar Eo
Los potenciales de reducción estándar de diversos pares redox se pueden colocar en lista ordenada desde el Eo mas negativo hasta el mas positivo.

FOSFORILACION OXIDATIVA
La transferencia de electrones por debajo de la cadena de trasportadores de estos se favorece desde el punto de vista energético porque el NADH es un donador potente de electrones y el oxigeno molecular es un receptor ávido de estos.

Hipótesis quimioosmótica
Explica la manera en que la energía libre que se genera por la actividad de la cadena de trasporte de electrones se emplea para producir ATP a partir de la reacción ADP+F.

Bomba de protones
El transporte de electrones esta acoplado con la fosforilación del ADP por el trasporte de protones (H+) a través de la membrana mitocondrial interior desde la matriz hacia el espacio intermembranoso.

Sintasa del ATP: sintetiza como su nombre lo indica ATP, valiéndose de la energía del grandiente de protones que se genera por la cadena de transporte de electrones.
Oligomicina: este fármaco se fija al tallo de la sintasa de ATP, con lo que cierra el canal de H+ y proviene del reingreso de protones en la matriz motocondrial.

Proteínas desacopladoras (UCP)
En la membrana mitocondrial interior de los mamíferos, con inclusión desde luego del ser humano.

Transporte de ATP y ADPLa membrana mitocondrial inferior
necesita trasportadores especializados de ADP y F desde el citosol.( en el que se emplea ATP y se convierte en ADP en muchas reacciones que necesitan energía)

Trasporte de equivalentes reductores
La membrana mitocondrial interior carece de proteína trasportadora de NADH y el NADH que se produce en el citosol no puede penetrar directamente en la mitocondria.

Defectos hereditarios de la fosforilación oxidativaUnos 13 de los cerca de
100 polipéptidos que se requieren para la fosforilación oxidativa son calificados por el DNA mitocondrial.las mutaciones del mtDNA son los causantes de diversas enfermedades entre ellas ciertos casos de miopatias mitocondriales.