
Evaluacion de la resistencia a costra negra en materiales germoplasmicos de papa en invernadero
104
i DEPARTAMENTO DE CIENCIAS DE LA VIDA Y DE LA AGRICULTURA CARRERA DE INGENIERÍA EN BIOTECNOLOGÍA PROYECTO DE TITULACIÓN PREVIO A LA OBTENCIÓN DEL TÍTULO DE INGENIERO EN BIOTECNOLOGÍA AUTOR: EDISON ANDRÉS GALARRAGA GALARZA TEMA: EVALUACIÓN DE LA RESISTENCIA A COSTRA NEGRA (Rhizoctonia solani Kühn) AG3 AISLADA DE LA PROVINCIA DE CARCHI EN MATERIAL GERMOPLÁSMICO DE PAPA A NIVEL DE INVERNADERO. DIRECTORA: ALMA KOCH, MSc. CODIRECTOR: ANDRÉS IZQUIERDO, PhD. SANGOLQUÍ, ABRIL 2015
-
Upload
andres-galarraga -
Category
Documents
-
view
24 -
download
1
description
Evalucion de resistencia a costra negra. Rhizoctonia solani Kuhn AG3 en germoplasmas de papa
Transcript of Evaluacion de la resistencia a costra negra en materiales germoplasmicos de papa en invernadero

i
DEPARTAMENTO DE CIENCIAS DE LA VIDA Y DE LAAGRICULTURA
CARRERA DE INGENIERÍA EN BIOTECNOLOGÍA
PROYECTO DE TITULACIÓN PREVIO A LA OBTENCIÓN DELTÍTULO DE INGENIERO EN BIOTECNOLOGÍA
AUTOR: EDISON ANDRÉS GALARRAGA GALARZA
TEMA: EVALUACIÓN DE LA RESISTENCIA A COSTRA NEGRA(Rhizoctonia solani Kühn) AG3 AISLADA DE LA PROVINCIA DE
CARCHI EN MATERIAL GERMOPLÁSMICO DE PAPA A NIVELDE INVERNADERO.
DIRECTORA: ALMA KOCH, MSc.
CODIRECTOR: ANDRÉS IZQUIERDO, PhD.
SANGOLQUÍ, ABRIL 2015

ii
HOJA DE LEGALIZACIÓN DEL PROYECTO
ELABORADO POR
________________________Sr. Edison Andrés Galárraga Galarza
DIRECTORA DE LA CARRERA
________________________M.Sc. María Augusta Chávez
Sangolquí, Abril de 2015

iii
CERTIFICACIÓN
Alma Koch, MSc. Andrés Izquierdo, Ph.D.
Certifican que el trabajo titulado “Evaluación de la resistencia a costra negra(Rhizoctonia solani Kühn) AG3 aislada de la provincia de Carchi en materialgermoplásmico de papa a nivel de invernadero” fue realizado en su totalidad por elSr. Edison Andrés Galarraga Galarza, como requerimiento parcial a la obtención deltítulo de INGENIERO EN BIOTECNOLOGÍA, ha sido guiado y revisadoperiódicamente y cumple con las normas estatutarias establecidas por la Universidad delas Fuerzas Armadas-ESPE, en el Reglamento de Estudiantes.
El mencionado trabajo consta de un documento empastado y de un disco compacto, quecontiene los archivos en formato portátil de Acrobat (pdf). Autorizan a Edison AndrésGalarraga Galarza que lo entregue a la señora María Augusta Chávez, Ms.C, en sucalidad de Coordinadora de la Carrera.
Sangolquí, Abril 2015.
______________________ ______________________
ALMA KOCH, MSc. ANDRÉS IZQUIERDO, Ph.D.
DIRECTOR CODIRECTOR

iv
DECLARACIÓN DE RESPONSABILIDAD
Edison Andrés Galarraga Galarza
Declaro que:
El proyecto de grado denominado “Evaluación de la resistencia a costra negra
(Rhizoctonia solani Kühn) AG3 aislada de la provincia de Carchi en material
germoplásmico de papa a nivel de invernadero” ha sido desarrollado en base a una
investigación exhaustiva, respetando todos los derechos intelectuales de terceros,
conforme las citas que constan al pie de las páginas correspondientes, cuyas fuentes se
incorporan en la bibliografía. Consecuentemente este trabajo es de mi autoría.
En virtud de esta declaración, me responsabilizo del contenido, veracidad y alcance
científico del proyecto de grado en mención.
Sangolquí, Abril 2015.
_____________________________
Edison Andrés Galárraga Galarza

v
AUTORIZACIÓN
Yo, Edison Andrés Galarraga Galarza
Autorizo a la Universidad de las Fuerzas Armadas-ESPE, la publicación en la biblioteca
virtual de la Institución del trabajo “Evaluación de la resistencia a costra negra
(Rhizoctonia solani Kühn) AG3 aislada de la provincia de Carchi en material
germoplásmico de papa a nivel de invernadero” cuyo contenido y criterios son de mi
exclusiva responsabilidad y autoría.
Sangolquí, Abril 2015.
_____________________________
Edison Andrés Galárraga Galarza

vi
DEDICATORIA
A Dios por ser muy bueno conmigo, por su infinito amor y misericordia.
A mis padres, que con mucha paciencia y amor me han sabido guiar y tener paciencia a
lo largo de mi carrera universitaria.
A mi abuelo, mi papi “Coke”, que me enseñado como a un hijo, me ha acompañado en
todos mis logros y los ha vuelto suyos; esta tesis va para ti mi viejo.
Edison Andrés Galárraga Galarza

vii
AGRADECIMIENTOS
A Dios por darme la fuerza y oír todas mis oraciones, a mi familia por haber estado
conmigo siempre en las buenas y en las malas, los amo.
A mis profesores que de buena o mala manera han sabido guiarme a lo largo de mi vida
universitaria.
A Almita Koch, MSc. por haberme hecho parte de su familia microbiología y por
apoyarme durante el desarrollo de mi tesis. Siempre admiraré su paciencia y sus
conocimientos.
Al Doctor Andrés Izquierdo, PhD. por haber aceptado la codirección y brindarme sus
conocimientos.
A Jessy Maisincho por la ayuda y la paciencia brindada durante todo este tiempo,
muchas gracias.
Al Instituto Nacional Autónomo de Investigaciones Agropecuarias (INIAP) por haberme
acogido durante la realización de mi tesis, una gran institución. Al Ing. Eloy Mora por
su, paciencia, confianza y apertura en el Departamento de Protección Vegetal de INIAP.
Al Ing. Pedrito Romero por su apertura, respaldo, confianza y conocimientos.
Al Ing. Geovanni Rodríguez mi colega y amigo, gracias por el apoyo y la confianza mi
amigo.
Al Ing. Javier Molina por haberme acompañado a lo largo de mi carrera, por brindarme
sus conocimientos y ser un amigo. Gracias a ti aprendí que “vale más un burro bien
dormido que un genio amanecido” (Molina, 2010).
A todos mis amigos: Pablo Espinosa, Alexandra Rueda, Alexandra reyes, Andrés
Almeida, Gabriela Fuentes, Wilson Castro, Isaac Armendáriz a todos en general, faltan
muchos pero a tod@s los llevaré siempre en el corazón.
DIOS, PATRIA Y LIBERTAD

viii
ÍNDICE DE CONTENIDOS
CERTIFICACIÓN………………………………………………………………………iii
DECLARACIÓN DE RESPONSABILIDAD…………………………………………..iv
AUTORIZACIÓN………………………………………………………………………..v
DEDICATORIA………………………………………………………………………...vi
AGRADECIMIENTOS………………………………………………………………...vii
CAPÍTULO 1 – INTRODUCCIÓN……………………………………………………...1
1.1. Justificación del problema…………………………………………………………1
1.2. Objetivos de la investigación……………………………………………………....3
1.3. Marco Teórico…………………………………………………………………….4
1.3.1. Importancia socio-económica……………………………………………..4
1.3.2. Consumo…………………………………………………………………...4
1.3.3. La papa (Solanum tuberosum L.)………………………………………….5
1.3.4. Genética de la papa………………………………………………………...5
1.3.5. Relevancia de la especie…………………………………………………...6
1.3.6 Distribución de la especie………………………………………………….7
1.3.7. Producción a nivel Nacional………………………………………………7
1.3.8. Particularidades del Cultivo………………………………………………9

ix
1.3.9. Morfología de la papa…………………………………………………….12
1.3.10. Fenología del Cultivo…………………………………………………...15
1.3.11. Enfermedades del cultivo de papa………………………………………15
1.3.12. Características de enfermedades por fitopatógenos……………………..16
1.3.13. Enfermedades fúngicas que afectan al cultivo de papa………………....18
1.3.14. Costra negra…………………………………………………………..…18
1.3.15. Morfología y citología de Rhizoctonia solani Kühn….……………...…20
1.3.16. Grupos de Anastomosis (AG)…………………………………………..22
1.3.17. Epidemiología…………………………………………………………..23
1.3.18. Ciclo de Vida de R. solani Kühn………………………………………..24
1.3.19. Síntomas de costra negra………………………………………………..25
1.3.20. Control de la enfermedad……………………………………………….28
CAPÍTULO 2 - MATERIALES Y MÉTODOS……………………………………….29
2.1. Participantes………………………………………………….……..……………29
2.2. Zona de estudio……………………………………………………………….….29
2.3. Revisión bibliográfica……………………………………………………………29
2.4. Fase de campo-toma de muestras………………………………………………..30
2.5. Fase de laboratorio………………………………………………………………30
2.6. Aislamiento de Rhizoctonia solani Kühn……………………………………..…31

x
2.6.1. Preparación de medio de cultivo………………………………………….31
2.6.2. Desinfección de los explantes tubérculos de Solanum tuberosum……….31
2.6.3. Aislamiento………………………………………………………………32
2.6.4. Observación del cultivo al microscopio………………………………….32
2.6.5. Extracción de ADN de R. solani Kühn…………………………………..33
2.6.6 PCR anidada para la determinación del Grupo de Anastomosis 3 (AG3)...34
2.6.7. Amplificación de las regiones ITS1-5.8S-ITS2………………………….34
2.6.8. Amplificación de regiones específicas para AG-3……………………….35
2.6.9. Electroforesis en gel de agarosa………………………………………….36
2.6.10. Multiplicación de Rhizoctonia solani Kühn…………………………….36
2.6.11. Preparación del inóculo de R. solani Kühn……………………………..37
2.6.12. Inoculación de Tierra con germoplasmas……………………………….38
2.6.13. Control de Inóculo………………………………………………………39
2.6.14. Manejo del experimento………………………………………………...40
2.7. Método de evaluación………………………………………………………40
CAPÍTULO 3 – RESULTADOS………………………………………………………42
3.1. Aislamiento de Rhizoctonia solani Kühn………………………………………..42
3.2. Extracción de ADN de Rhizoctonia solani Kühn………………………………..43
3.3. Nested PCR para la confirmación de Rhizoctonia solani AG3……………….…44

xi
3.4. Preparación del Inóculo…………………………………………………………..46
3.5. Sintomatología aérea……………………………………………………………..47
3.6. Análisis estadístico……………………………………………………………….47
3.7. Porcentaje de emergencia………………………………………………………..48
3.8. Altura de las variedades inoculadas……………………………………………...48
3.9. Evaluación a la cosecha de los tubérculos……………………………………….51
3.10. Severidad de la infección……………………………………………………….52
3.11. Número de tubérculos…………………………………………………………..57
3.12. Rendimiento total en gramos……………………………………………………61
CAPÍTULO 4 – DISCUSIÓN……………………………………………………….…64
4.1. Aislamiento e identificación de Rhizoctonia solani Kühn……………………….65
4.2. Preparación del inóculo de Rhizoctonia solani Kühn……………………………66
4.3. Síntomas de R. solani Kühn……………………………………………………...67
4.4. Control de altura de la planta…………………………………………………….69
4.5. Evaluación de los tubérculos…………………………………………………….69
4.6. Severidad de la Infección………………………………………………………...70
4.7. Rendimiento……………………………………………………………………...71
4.8. Plagas y control del ensayo………………………………………………………71
CAPÍTULO 5 – CONCLUSIONES……………………………………………………74

xii
CAPÍTULO 6 –
RECOMENDACIONES………………………………………………………………..75
BIBLIOGRAFÍA……………………………………………………………………….76
ANEXO I………………………………………………………………………………..81
ANEXO II………………………………………………………………………………84
ANEXO III……………………………………………………………………………...85

xiii
LISTADO DE FIGURAS
Figura 1.1 Aporte de la producción de papas por provincias……………………………8
Figura 1.2 Flor de la planta de papa en invernadero…………………………………...15
Figura 1.3 Sintomatología de R. solani ………………………………………………..27
Figura 2.1 Dispensación de medio PDA en cajas petri dentro de una cámara de flujo
laminar…………………………………………………………………………………..31
Figura 2.2 Armada de PCR dentro de una cámara de flujo laminar…………………...36
Figura 2.3 Preparación de inóculo de R. solani………………………………………..37
Figura 2.4 Inoculación de tierra para la siembra de germoplasmas……………...…….39
Figura 2.5 Invernadero de la estación científica Santa catalina………………………..40
Figura 3.1 Cultivo aislado de Rhizoctonia solani Kühn en medio PDA en caja petri…42
Figura 3.2 Tinción de hifas de Rhizoctonia solani Kühn vista en el objetivo…………43
Figura 3.3 Electroforesis de la región ITS1-5,8-ITS2 con su control negativo………..45
Figura 3.4 Amplificación de la región específica para AG3 con su control negativo…46
Figura 3.5. A: Preparación del inóculo de Rhizoctonia solani Kühn en semillas de
avena…………………………………………………………………………………….47
Figura 3.6 Diagrama de barras para las alturas de las plantas con su error estándar….50
Figura 3.7 Necrosis en tallo de la variedad I-Rubí del cuarto bloque…………………52
Figura 3.8 Tubérculos con costra negra de la variedad mejorada I-Rubí………………54
Figura 3.9 Tubérculos del clon promisorio 12-4-170………………………………….55

xiv
Figura 3.10 Diagrama de cajas con sus media de la severidad de las variedades de papa
inoculadas……………………………………………………………………………….56
Figura 3.11. Diagrama de la variable número de tubérculos por variedad con sus
respectivas medias………………………………………………………………………58
Figura 3.12. Diagrama de bloques de la variable rendimiento en variedades inoculadas y
no inoculadas………………………………………………………………………..….63

xv
LISTADO DE TABLAS
Tabla 1.1 Composición química óptima del suelo para el cultivo de papa.……………10
Tabla 1.2 Aporte nutricional de papa fresca para consumo humano…………………..12
Tabla 2.1 Materiales germoplásmicos evaluados……….……………………………..38
Tabla 2.2 Criterio de Clive para la evaluación de tubérculos………………………….41
Tabla 3.1. Valores de concentración y absorbancias de ADN total extraído de
Rhizoctonia solani………………………………………………………………………44
Tabla 3.2 Análisis de varianza de la altura de las plantas……………………………...49
Tabla 3.3 Alturas promedio y pruebas de Tukey para la variable altura………………51
Tabla 3.4 Análisis de varianza del porcentaje de severidad de la infección de papas…53
Tabla 3.5 Resultados del análisis de tukey, rangos de los valores promedios del
porcentaje de infección de costra negra…………………………………………………57
Tabla 3.6 Análisis de varianza para la variable número de tubérculos de tratamientos
inoculados y testigo……………………………………………………………………..59
Tabla 3.7 Prueba de Tukey para la variable número de tubérculos…………………….60
Tabla 3.8 Análisis de varianza para la variable gramos por maceta de tratamientos
inoculados y testigo…………………………………………………………………….61
Tabla 3.9 Prueba de Tukey para la variable “rendimiento por maceta”……………….62

xvi
RESUMEN
La papa (Solanum tuberosum) es uno de los cultivos tradicionales de mayor importancia
económica en el Ecuador y el cuarto a nivel mundial. En el Ecuador se ha visto afectado
por Rhizoctonia solani Kühn, agente causal de la Rizoctoniosis o costra negra,
enfermedad fúngica que ataca a brotes, tallos, estolones, tubérculos, causa necrosis,
marchitez foliar y en ocasiones anula la germinación de la plántula. Puede acabar con
hasta el 25% de un cultivo en campos de agricultores. La búsqueda y selección de
variedades resistentes es de importancia crucial para hacer frente a problemas que
afectan directamente a los cultivos, sean estos de origen fitopatológico, físico-químico o
de carácter ambiental. Se buscaron variedades de papa con resistencia a costra negra
AG3.
R. solani AG3 fue aislada a partir de esclerocios de papa provenientes de la provincia de
Carchi. Se confirmó mediante biología molecular el grupo de anastomosis 3 (AG3) para
el género Rhizoctonia con primers específicos. Se evaluaron frente a R. solani veinte
materiales germoplásmicos proporcionados por INIAP, en los que se incluían variedades
mejoradas y clones promisorios. El material vegetal fue estudiado durante seis meses a
nivel de invernadero. Se evaluaron y seleccionaron los tubérculos a la cosecha mediante
el criterio de Clive (1971) en base al porcentaje de infección. Se encontró que la
variedad I-Puca Shungo y el clon promisorio 12-6-1 presentan Alta tolerancia ante R.
solani.
Palabras clave: costra negra, anastomosis, fitopatógenos, germoplásmico, resistencia.

xvii
ABSTRACT
The potato (Solanum tuberosum) is one of the traditional crops of major economic
importance in Ecuador and the fourth worldwide. In Ecuador, it has been affected by
Rhizoctonia solani Kühn, causal agent Rizoctoniosis or black scab, fungal disease that
attacks buds, stems, stolons, tubers, cause necrosis, leaf wilting and sometimes nullifies
seedling germination. R. solani can kill up to 25% of a crop in farmers' fields. The
search and selection of resistant varieties is crucial to address problems directly affecting
crops, these problems can be: phytopathological origin, physical-chemical or
environmental nature. Potato varieties were sought with black scab resistance AG3.
R. solani AG3 was isolated from sclerotia of potatoes from the province of Carchi.
Anastomosis group 3 (AG3) for the genus Rhizoctonia was confirmed by molecular
biology with specifics primers. Twenty potato seeds provided by INIAP, which includes
improved varieties and promising clones, were evaluated.
The plant material was studied for six months at the level of greenhouse. They were
evaluated and selected the harvested tubers by Clive criteria (1971) by the percentage of
infection. It was found that the variety I-Puca Shungo promising clone 12-6-1 present
high tolerance for R. solani.
Keywords: black crust, anastomosis, plant pathogens, potato-seed, resistance.

1
CAPÍTULO 1 – INTRODUCCIÓN
La papa (Solanum tuberosum) es uno de los cultivos de mayor importancia
económica en el Ecuador. Según la FAO, la producción se vincula a 82000 productores
en alrededor de 90 cantones. El cultivo del tubérculo abarca la mayor extensión de
terreno con un 5,5% del área total de cultivos de la serranía ecuatoriana (Paca, 2009).
La presencia de la enfermedad de la costra negra se ha incrementado en el país
durante los últimos años, llegando a causar hasta la muerte del 70% de las plántulas de
papa. Se han documentado pérdidas en campo que llegan hasta el 20% y que se han
reflejado en importantes pérdidas económicas (INIAP, 2005).
La búsqueda de variedades con tolerancia genética ha sido el objetivo del sector
agrícola durante los últimos años. El uso de productos químicos para el control de
fitopatógenos ha dado lugar a plagas secundarias, por lo que, el uso de semilla con
tolerancia a Rhizoctonia solani Kühn representaría menor costo en la producción del
tubérculo, disminuiría el uso de fungicidas y generaría menor impacto ambiental (Prado
et al., 2001).
1.1. Justificación del problema
Se estima que la producción global de papa genera 16.5 billones de dólares anuales.
El cultivo de papa es económicamente importante a nivel mundial. En el Ecuador, la
papa constituye el cuarto cultivo más importante por su producción, alto consumo,
aporte nutricional y por ser una fuente de ingresos para numerosos agricultores,
principalmente del sector rural. Representa el 7.4% del Producto Interno Bruto Agrícola
(PIBA) (Devaux et al., 2010).

2
En los últimos años, debido al incremento de tierras cultivadas, se han desarrollado
problemas fitosanitarios que han afectado directamente a los agricultores, situación
debida al mal manejo del cultivo, al desconocimiento de las causas de enfermedad y a la
falta de tecnología adecuada que permita controlar estos problemas oportunamente y así
evitar que se propague a otras zonas (INIAP, 2005).
Torres (1989), en su estudio acerca de las principales enfermedades fúngicas de la
papa relacionadas con la producción de tubérculos-semillas, indica que todas estas
enfermedades son de gran importancia, ya que pueden llegar a destruir cultivos enteros o
afectar en el rendimiento de la producción, restándole calidad al tubérculo, este es el
caso de la costra negra (Rhizoctonia solani Kühn). Se conocen en Ecuador más de 30
enfermedades fúngicas que afectan al cultivo de Solanum tuberosum siendo la
Rizoctoniosis o costra negra la más común en suelos paperos. R. solani se perpetúa en
cultivos por acciones mecánicas como la labranza, la utilización de implementos
agrícolas infectados, restos de tallos y hojas infectadas, y semillas con esclerocios.
Rhizoctonia solani Kühn ataca brotes, estolones y tubérculos, la mayor sintomatología
ocurre en el estado de plántula; causando efectos irreversibles para la planta, llegando a
causar una pérdida de casi el 70%. En el año 2005 (INIAP, 2005) en la Estación
Experimental de Santa Catalina (EESC), se obtuvieron porcentajes de incidencia y de
severidad del 81%; al realizar controles de calidad internos de semilla de papa categoría
registrada INIAP Fripapa. En consecuencia de las altas infecciones, la extracción de
semilla para la variedad I-Fripapa disminuyó considerablemente. Se obtuvieron valores
de 50% de extracción frente a 60-70% que se considera rentable.

3
Con estos antecedentes, enfocados en el impacto de pérdidas económicas para los
agricultores, es necesaria la generación permanente de nuevas variedades de papa que
posean una tolerancia alta o parcial a R. solani; conociendo que la semilla se considera
como el factor principal para una producción agrícola saludable.
1.2. Objetivos de la investigación
1.2.1. Objetivo general
Encontrar materiales germoplásmicos de papa tolerantes a costra negra, mediante la
evaluación de la severidad en sintomatología en parte aérea, en tallos, raíces y
tubérculos, para disponer de tubérculo semilla con una tolerancia moderada y alta a
Rhizoctonia solani Kühn.
1.2.2. Objetivos específicos
a) Aislar Rhizoctonia solani Kühn AG3 de esclerocios de papa provenientes de la
provincia de Carchi.
b) Identificar el comportamiento de diferentes materiales germoplásmicos de papa,
proporcionados por INIAP, frente a la enfermedad costra negra (Rhizoctonia solani
Kühn).
c) Seleccionar las variedades tolerantes a costra negra.

4
1.3. Marco Teórico
1.3.1. Importancia socio-económica
Se estima que 225000 personas subsisten directamente del cultivo de la papa,
115000 personas están involucradas indirectamente en actividades de comercialización,
transporte y procesamiento del tubérculo. En general, el 7% de la población
económicamente activa del país tienen a la papa como su principal fuente de ingreso. El
cultivo de la papa se caracteriza por su alto uso de mano de obra. Se emplean alrededor
de 150 jornales en cada hectárea por ciclo, es decir, en 49000 hectáreas cultivadas por
año se usan 7.5 millones de jornales anuales, un equivalente a 36.7 millones de dólares
de ingresos directos. Anualmente por cada hectárea cultivada se requiere una inversión
de 2500 dólares (Reinoso, 2011).
En los Andes, los países con mayor producción por hectárea sembrada son:
Colombia y Venezuela con 16 t/ha, mientras que los países con menor producción son
Bolivia y Ecuador con un promedio de 6 a 7 t/ha. En Ecuador se siembran comúnmente
30 cultivares, de los cuales, INIAP Gabriela y Superchola representan el 50% del área
sembrada (INIAP, 2002).
1.3.2. Consumo
La papa es uno de los alimentos de mayor consumo en la dieta de los habitantes de la
serranía ecuatoriana. Según el Instituto Autónomo de Investigaciones Agropecuarias
(INIAP) el consumo anual de una persona por ciudad es: 122 kg en Quito, 80 kg en

5
Cuenca y 50 kg en Guayaquil. Actualmente las industrias alimenticias utilizan 50000
toneladas al año, lo cual representa el 10% de la producción nacional (Andrade, 2002).
1.3.3. La papa (Solanum tuberosum L.)
Se conoce como papa o patata al tubérculo formado por engrosamiento de raíz de la
planta Solanum tuberosum L. Según Dimitri (1972), a la papa (Solanum tuberosum), le
corresponde la siguiente clasificación taxonómica: Reino Plantae; Subreino Antophyla;
División Angiosperma; Clase Dicotiledónea; Subclase Simpetalae; Orden Tubuflorae;
Familia Solanaceae; Género Solanum; Especie tuberosum.
Es una planta de altura por lo que sus cultivos se realizan desde los 2700 a 3400
msnm, y se desarrolla adecuadamente a una temperatura que oscila entre los 9 y 11°C.
Aunque el cultivo se encuentra en varios tipos de suelos, le van mejor aquellos de
textura arenosa, con buen drenaje y estructura suelta, es sensible a suelos compactos por
lo que se recomienda tener una profundidad de al menos 30 centímetros. Se desarrolla en
un rango de pH de 5.0 a 7.0 (Paca, 2009).
1.3.4. Genética de la papa
Existen alrededor de 4000 variedades de papa, variedad génica distribuida en más de
100 países alrededor del mundo. Esta gran diversidad genética ha sido preservada
mediante prácticas tradicionales de los agricultores. El cuidado de las semillas por parte
de los pequeños productores paperos de la región andina han permitido el
mantenimiento de la gran variedad de papas; que presentan adaptaciones a altura,
temperaturas y suelos (Borba, 2008).

6
El Ecuador posee una inmensa diversidad genética de papa. El Instituto Autónomo
de Investigaciones Agropecuarias (INIAP) ha identificado 393 variedades de papa, 232
pertenecen a variedades silvestres y 161 a variedades nativas. La gran diversidad de
materiales junto con germoplasmas modificados, han sido la respuesta a las limitaciones
del cultivo, dando resistencia a factores adversos como: heladas, condiciones
ambientales y fitopatógenos (Reinoso, 2011).
El flujo genético de la especie es alto entre variedades cultivadas y silvestres, con la
subespecie andigena y con especies hexaploides (EBN=4). El fenómeno genético en la
solanácea ha sido atribuido a la dispersión del polen, a la coexistencia geográfica y a la
relación taxonómica. Aparentemente la evolución de las especies de papa se originó a
partir del nivel diploide. El gametofito de la papa es 2n=12. El esporofito encontrado en
diferentes variedades puede ser: 2n=24 y 48; 2n=24, 36, 48 y 60. Solanum tuberosum
presenta alta heterocigosidad (he) que oscila entre 0.46 y 0.52 (Ortiz & Peloquin, 1994).
1.3.5. Relevancia de la especie
El cultivo de papa ocupa el cuarto lugar dentro de las plantaciones de mayor
importancia a nivel mundial; debido a su sabor y consistencia, a su alto aporte
nutricional y sus múltiples usos. El almidón es utilizado por industrias farmacéuticas,
textiles, de la madera y del papel, en la elaboración de pegamentos, piensos y etanol. En
Europa, a nivel industrial, es utilizada en la producción de vodka, whisky, almidón y
otras industrias la emplean como comidas rápidas (CONABIO, 2008).

7
A diferencia de otros cultivos, la papa se puede multiplicar de manera vegetativa a
través de tubérculos semilla. Este cultivo tiene un alto rendimiento y no requiere de
cuidados especiales (Estrada, 2000).
1.3.6 Distribución de la especie
La papa es una planta originaria de los Andes sudamericanos y se extiende desde
Colombia, pasando por Ecuador, Perú, Bolivia, Venezuela, Chile hasta Argentina. En el
Ecuador se distribuye en tres zonas geográficas: norte, centro y sur. El cultivo se
desarrolla en laderas irregulares en terrenos con una inclinación de hasta el 45%, en un
intervalo de 2400 a 3800 msnm. El cultivo se desarrolla en condiciones de subpáramo
húmedo (Hurtado & Román, 2002).
1.3.7. Producción a nivel Nacional
Las provincias de Carchi, Pichincha, Cotopaxi, Tungurahua y Chimborazo,
aportaron el 79.5% de la producción, las mayores extensiones de cultivo de papa
corresponden (Figura 1.1) en orden a: Carchi (26%), Chimborazo (15%), Cotopaxi
(15%), Tungurahua (14%) y Pichincha (11%) (Reinoso, 2011).

8
Figura 1.1: Aporte de la producción de papas por provincias (Reinoso, 2011).
La producción papera en el país se divide en tres zonas:
Zona Norte: Carchi e Imbabura
Carchi e Imbabura presentan el mayor índice de producción a nivel nacional, poseen
un rendimiento de 21.7 t/ha y produce el 40% de la cosecha anual en el país. El área
papera zonal comprende alturas entre los 2800 msnm a 3200 msnm y presenta un clima
frío característico de alta montaña, con temperaturas promedio de 11-12°C. En esta zona
las probabilidades de heladas son relativamente bajas (Pumisacho & Sherwood, 2002).
Zona centro: Pichincha, Cotopaxi, Tungurahua, Chimborazo y Bolívar
En la zona centro, la provincia de Chimborazo, a una altura comprendida entre los
2200-3600 msnm, tiene la mayor superficie dedicada al cultivo de la solanácea, sin
embargo, el rendimiento de la provincia es bajo (11 t/ha). La temperatura varía entre 6°
y 15°C. Existen tres zonas de producción papera: occidente, nororiente y cordillera

9
central. En la zona de occidente se encuentran los cantones Colta y Riobamba. El cantón
Chambo corresponde a la zona nororiental y el cantón Guano a la cordillera central. En
este último la siembra se puede dar todo el año. En la provincia de Chimborazo la
producción del tubérculo se da en diferentes tipos de suelo. Los más importantes:
Inceptisoles, Mollisoles, Entisoles y Afisoles (Pumisacho & Sherwood, 2002).
Zona Sur: Cañar, Azuay y Loja
La zona productora sur es la de menor importancia a nivel nacional. Tiene un
promedio de temperatura entre los 13°C y 15°C. La mayor producción se encuentra en la
provincia de Cañar, a una altura de 2000 msnm, presenta un rendimiento de 8-10 t/ha.
En las provincias de Azuay y Loja, la producción de papa es casi nula debido a las bajas
precipitaciones de la zona (Pumisacho & Sherwood, 2002).
1.3.8. Particularidades del Cultivo
1.3.8.1 Requerimientos de Suelo
En el cultivo de papa, su forma, apariencia y rendimiento depende de la textura y
naturaleza del suelo. La papa se produce en suelos bien drenados, que posean materia
orgánica y nutrientes en su composición. La planta de papa necesita de un suelo suelto,
inclinado, y con buen drenaje para evitar que el agua se empoce; preferentemente el
suelo debe ser profundo y fértil con un pH en un intervalo de 5.0 a 6.0 (Paca, 2009).
Dentro de los mejores suelos para el cultivo de la solanácea se encuentran: los
francos, franco-arenosos, franco-limosos y franco-arcillosos. Estos suelos se caracterizan
por ser de color oscuro, poseer una textura liviana y permitir el libre crecimiento de

10
raíces y estolones. La cantidad de materia orgánica adecuada en el suelo debe ser mayor
al 3.5% en la composición del terreno (Hurtado & Román, 2002).
En la sierra ecuatoriana el suelo predominante es de origen volcánico, con altas
concentraciones de aluminio. La ceniza volcánica forma un complejo entre materia
orgánica y minerales. Estas tierras tienen gran capacidad de retención de agua y alto
poder de fijación de fósforo (INIAP, 2002).
1.3.8.2. Composición química óptima
Según Hurtado & Román (2002), la composición química óptima del suelo apta para
el cultivo de papa se muestra en la Tabla 1.1.
Tabla 1.1 Composición química óptima del suelo para el cultivo de papa (Hurtado & Román, 2002).
Propiedades Químicas Rango óptimopH 5.5 - 6N Variable
P > 28 mg/kgK > 5 %
Ca++ 65%Mg++ 18%
Acidez total < 10 %Conductividad eléctrica < 4 dsm- 1
1.3.8.3. Intensidad lumínica
La cantidad de luz en un cultivo es de importancia para la floración y fructificación.
Durante el inicio del crecimiento del tubérculo tendrá un efecto marcado sobre su futuro
desarrollo, es fundamental en la síntesis de carbohidratos a través del proceso de
fotosíntesis (CORPOICA, 2000).

11
La intensidad lumínica es de gran importancia en los fotoperíodos requeridos por las
plantas; así, uno corto disminuirá el crecimiento vegetativo, pero a su vez acumulará
mayor cantidad de carbohidratos que aumentará la producción de tubérculos, y los
fotoperiodos largos favorecen el crecimiento. La cantidad óptima de horas luz para el
cultivo de papa son 12 horas al día (INTA, 2004).
1.3.8.4. Disponibilidad de agua
El efecto de la falta de humedad en el suelo afecta en el rendimiento final de los
cultivos, depende del estado fenológico de la planta al momento del déficit hídrico. La
disponibilidad de agua en el suelo es uno de los factores que influirán directamente en el
crecimiento del cultivo, en la fotosíntesis y en la absorción de nutrientes; si existe poca
disponibilidad de agua se producirá clorosis, marchitamiento foliar y por consiguiente la
disminución en el rendimiento. Un exceso de humedad ayuda al desarrollo de
enfermedades. La precipitación o cantidad mínima de agua requerida es de 600 mm,
distribuida en todo su ciclo vegetativo; las mayores demandas se dan en las etapas de
germinación y crecimiento de los tubérculos, por lo cual es necesario efectuar riegos
suplementarios en los períodos críticos o cuando no se presenta lluvia. Un rango óptimo
de precipitación para el cultivo de papa es de 800 a 1200 mm durante el ciclo (INIAP,
1984).
1.3.8.5. Temperatura
La papa es una planta termoperiódica, es necesario que exista una variación de por lo
menos 10°C entre la temperatura diurna y la nocturna. Si la diferencia es menor, el
crecimiento y tuberización se ven afectados. Cuando esta situación se da a menudo, a lo

12
largo del ciclo vegetativo, el rendimiento y la calidad son afectados, pues las
temperaturas altas son ideales para el crecimiento de tallos y hojas, pero no para los
tubérculos (Hurtado & Román, 2002).
La temperatura interviene directamente en la producción de brotes en los
germoplasmas, controla la utilización de nutrientes, la pérdida de agua e influye en las
etapas fenológicas del cultivo (INTA, 2004).
La papa requiere de condiciones que permitan su desarrollo como los climas
templados fríos y las altitudes que van desde los 2 500 a 3 500 metros, la temperatura
óptima para el cultivo es de 9 a 11ºC. El cultivo soporta hasta una temperatura máxima
de 18ºC y debe encontrarse libre de heladas y granizadas (INIAP, 2005).
Las plantas producen más follaje a temperaturas bajo los 29ºC. A estas temperaturas
se induce el desarrollo de tallos, el crecimiento de las hojas es atrofiado debido a la
reducción de la expansión del área foliar, los niveles de almidón son bajos en las hojas y
los estolones son más largos. Se retrasa el comienzo del crecimiento del tubérculo y se
demora el inicio del llenado del mismo. Las temperaturas elevadas favorecen el
desarrollo de plagas y enfermedades. La humedad ambiental excesiva empeora el ataque
de lancha negra y la aparición de otros fitopatógenos (Estrada, 2000; CORPOICA,
2000).
1.3.9. Morfología de la papa
El desarrollo de la planta puede ser producida a partir de un tubérculo semilla y de
semilla de papa. En el caso de ser producido por medio de un tubérculo, las raíces

13
tendrán formación en los brotes del mismo, mientras que el tallo se constituye de nudos
que se encuentran en los brotes del tubérculo (Piñeros, 2009).
El tamaño de la planta va desde los 0.4 a 1.4 m, poseen tallos leñosos y gruesos de
color verde o rojo púrpura. El follaje normalmente alcanza una altura entre 0.60 a 1 m.
Las hojas son compuestas y pignadas. Las hojas primarias de plántulas pueden ser
simples, pero una planta madura contiene hojas compuestas en par y alternadas, y
produce un tubérculo, la papa misma, con abundante contenido de almidón. Las hojas se
ordenan en forma alterna a lo largo del tallo, dando un aspecto frondoso al follaje,
especialmente en las variedades mejoradas (Solano, 2007).
El tubérculo de la papa es un tallo subterráneo en su totalidad, el cual funciona como
almacén. A medida que madura, la cantidad de elementos conductores reducen y no
existe una zona de cambium continuo. Sobre la superficie de la exodermis, se confinan
depresiones de la superficie, dentro de las cuales se ubican las yemas vegetativas, y
donde se originan tallos. La formación de tubérculos es consecuencia de la proliferación
del tejido de parenquimatoso que estimula el aumento de células. El tejido vascular de
los tallos, estolones y tubérculos toma inicialmente la forma de haces bicolaterales.
Mientras el tubérculo crece, se forman tubos cribosos, células acompañantes y elementos
del parénquima conductor. Los hidratos de carbono se almacenan dentro de las células
del parénquima de reserva; de la médula y la corteza en forma de gránulos de almidón
(Aucancela et al., 2012; EDIFARM & CIA, 2010).
El aporte nutricional de los tubérculos está dado por el contenido de macro y
micronutrientes y por la biodisponibilidad. En promedio 100 gramos de papa, porción

14
que consume un individuo adulto, contiene gran cantidad de micro y macro elementos
los cuales se muestran en la Tabla 1.2 El tubérculo contiene gran cantidad de Ca, P y
vitamina C. La vitamina C contenida en una papa es equivalente a la cantidad diaria que
necesita una persona (FAO, 2008).
Tabla 1.2 Aporte nutricional de papa fresca para consumo humano (FAO, 2008)
Componente ValorAgua 77 gFibra 1.80 g
Valor calórico 87 KcalProteína 1.87 g
Carbohidratos 20.13 gLípidos 0.10 g
Vitamina C 13 mgHierro 0.31 mgCalcio 5 mg
Fósforo 44 mg
Dependiendo de la variedad, las flores se localizan en racimos terminales al tallo.
Existen factores como el fotoperiodo, la temperatura y el clima que estimulan la
floración de la planta. El inicio del periodo de floración indica la tuberización, uno de
los estados fenológicos más importantes para la planta (Figura 1.2). Las flores llegan a
tener 4 cm de diámetro, poseen cinco pétalos (Pentámeras) unidos por los bordes
formando una corola (Borja et al., 2010).

15
Figura 1.2 Flor de la planta de papa en invernadero (Galárraga, 2014).
1.3.10. Fenología del Cultivo
La descripción fenológica del cultivo de papa se da en respuesta al comportamiento
fisiológico varietal frente a condiciones climatológicas, insumos agrícolas y programas
agronómicos. Las etapas fenológicas de la planta de papa se inicia con la germinación
del tubérculo semilla, pasando por varias fases fisiológicas de tipo vegetativo hasta
alcanzar la madurez fisiológica y termina con la tuberización, dependiendo la variedad.
(Yzarra & Martín, 2011; Borja et al., 2010).
1.3.11. Enfermedades del cultivo de papa
El suelo es el hábitat de muchos microorganismos, en su mayoría beneficiosos para
las plantas, una pequeña parte causan patologías. Los microorganismos causantes de
enfermedad en las plantas se los conoce como fitopatógenos y pueden ser: bacterias,
virus, hongos, insectos, maleza y nematodos. Los patógenos del suelo han sido
considerados como secundarios dentro de los problemas de los cultivares, pero, en las
últimas décadas por el mal manejo del suelo y de la semilla, su presencia ha ido
incrementando preocupantemente. Los fitopatógenos se encuentran en todos los suelos
15
Figura 1.2 Flor de la planta de papa en invernadero (Galárraga, 2014).
1.3.10. Fenología del Cultivo
La descripción fenológica del cultivo de papa se da en respuesta al comportamiento
fisiológico varietal frente a condiciones climatológicas, insumos agrícolas y programas
agronómicos. Las etapas fenológicas de la planta de papa se inicia con la germinación
del tubérculo semilla, pasando por varias fases fisiológicas de tipo vegetativo hasta
alcanzar la madurez fisiológica y termina con la tuberización, dependiendo la variedad.
(Yzarra & Martín, 2011; Borja et al., 2010).
1.3.11. Enfermedades del cultivo de papa
El suelo es el hábitat de muchos microorganismos, en su mayoría beneficiosos para
las plantas, una pequeña parte causan patologías. Los microorganismos causantes de
enfermedad en las plantas se los conoce como fitopatógenos y pueden ser: bacterias,
virus, hongos, insectos, maleza y nematodos. Los patógenos del suelo han sido
considerados como secundarios dentro de los problemas de los cultivares, pero, en las
últimas décadas por el mal manejo del suelo y de la semilla, su presencia ha ido
incrementando preocupantemente. Los fitopatógenos se encuentran en todos los suelos
15
Figura 1.2 Flor de la planta de papa en invernadero (Galárraga, 2014).
1.3.10. Fenología del Cultivo
La descripción fenológica del cultivo de papa se da en respuesta al comportamiento
fisiológico varietal frente a condiciones climatológicas, insumos agrícolas y programas
agronómicos. Las etapas fenológicas de la planta de papa se inicia con la germinación
del tubérculo semilla, pasando por varias fases fisiológicas de tipo vegetativo hasta
alcanzar la madurez fisiológica y termina con la tuberización, dependiendo la variedad.
(Yzarra & Martín, 2011; Borja et al., 2010).
1.3.11. Enfermedades del cultivo de papa
El suelo es el hábitat de muchos microorganismos, en su mayoría beneficiosos para
las plantas, una pequeña parte causan patologías. Los microorganismos causantes de
enfermedad en las plantas se los conoce como fitopatógenos y pueden ser: bacterias,
virus, hongos, insectos, maleza y nematodos. Los patógenos del suelo han sido
considerados como secundarios dentro de los problemas de los cultivares, pero, en las
últimas décadas por el mal manejo del suelo y de la semilla, su presencia ha ido
incrementando preocupantemente. Los fitopatógenos se encuentran en todos los suelos

16
de todas las zonas paperas del país, con mayor incidencia en zonas frías, húmedas y en
suelos donde es predominante el monocultivo (Jager et al., 1982).
La diseminación de los fitopatógenos entre cultivos es provocada por el uso de
semilla contaminada, por restos de material vegetal contaminado, por suelo infectado e
implementos agrícolas contaminados. Dentro de las principales enfermedades causadas
por hongos se encuentra el tizón temprano ocasionado por Alternaria solana, el tizón
tardío que es causado por Phytophthora infestans y el cancro de la papa provocado por
Rhizoctonia solani Kühn. Estas plagas en la actualidad son controladas mediante el uso
de fungicidas. No existe ningún control totalmente eficaz por lo que se ha optado por
combinar el control por medios químicos, la resistencia genética y el manejo cultural.
Hay que recordar que el tizón tardío fue el causante de la hambruna producida en Irlanda
durante los años 1845 y 1848, en donde hubo una epidemia ocasionada por P. infestans
en los cultivos de papa, y como consecuencia produjo la muerte de más de un millón de
personas, además de un alto porcentaje de emigración de los pobladores (Borba, 2008).
Entre los principales fitopatógenos de suelos paperos se encuentran: la costra negra,
el nematodo del quiste de la papa; que afectan directamente al rendimiento, la sarna y
roña que afectan la apariencia del tubérculo, así como la pérdida de valor comercial
(Aucancela et al., 2012).
1.3.12. Características de enfermedades por fitopatógenos
La gran cantidad de pérdidas económicas es consecuencia de las enfermedades que
contraen los cultivos de papas, independientemente de la manera de contagio. La
infección puede ser de diferentes tipos como: bacterial, viral, fúngico, por nematodos,

17
insectos, maleza u oomicetos, los mismos que pueden ser clasificados por la especie de
plaga o enfermedad (Pérez & Forbes, 2011).
En el caso de enfermedades bacterianas existen de tres clases que son las más
comunes en la zona andina, marchitez bacteriana, pudrición blanda, pierna negra o pie
negro y la sarna común. Las condiciones favorables para la plaga que se encuentran
comúnmente en las enfermedades bacterianas son campos infestados, semillas
infestadas; dependiendo de la enfermedad se pueden encontrar una mayor propagación
en climas cálidos y en campos anegados (Pérez & Forbes, 2011). En el caso de pie negro
y pudrición blanda, pueden ser infectados en el momento de almacenamiento del
tubérculo o en el suelo antes de ser cultivado. Los tubérculos son más propensos a
adquirir esta enfermedad en el proceso de transporte y cosecha, ya que pueden sufrir
cortes (Castro & Contreras, 2011).
La sintomatología de la sarna común no puede ser observada en la parte aérea de la
planta, debido a que no afecta la parte área sino que la enfermedad se ubica en el
tubérculo. En fases tempranas de la enfermedad se observa rajaduras en forma de
estrellada, las mismas que aumentan de tamaño según la edad del tubérculo, pueden
llegar alcanzar hasta 8 mm de diámetro, en los tubérculos se observan de diferentes
formas geométricas las cuales pueden cubrir la mayor parte de la epidermis (Mora et al.,
2011).
La virosis es uno de los tipos de enfermedades virales que se encuentran en la zona
andina. Los síntomas que se reflejan en el cultivo de papa son enanismo en la planta,
amarillento, deformación de hojas, rigidez, necrosis en nervaduras, entre otros. En

18
algunos casos no presentan síntomas. Las condiciones favorables en la que la plaga se
propaga son por medio de las semillas infectadas, uso de las semillas varias veces, entre
otras (Pérez & Forbes, 2011)
1.3.13. Enfermedades fúngicas que afectan al cultivo de papa
En climas de alta humedad y con alta precipitación, las plantas son propensas a
enfermedades causadas por hongos; una enfermedad fúngica es reconocida por la
producción de manchas o necrosis de consistencia seca en las hojas, tallos y tubérculos.
1.3.14. Costra negra
Rhizoctonia solani fue descrito por Julios Kühn en el año de 1858. Este hongo
pertenece a la clase Basidiomycete y se caracteriza por no producir esporas asexuales. R.
solani únicamente en condiciones especiales puede producir esporas sexuales
(basidiosporas). En la naturaleza R. solani se reproduce asexualmente y permanece
como micelio vegetativo, el cual forma estructuras de resistencia llamadas esclerocios,
masas de hifas entretejidas con superficies duras y resistentes que actúan como
propágulos. En la superficie del tubérculo R. solani asocia sus hifas con masas de tierra
dando el aspecto de una costra razón por la cual ha tomado vulgarmente el nombre de
costra negra. Su estado sexual es conocido como Thanatephorus cucumeris, se
desarrolla en ciertas condiciones de humedad y de alta temperatura. La forma sexual de
R. solani ha sido encontrado en cultivares de la provincia de Carchi, Tungurahua y
Chimborazo (Alba, 2006).
Rhizoctonia solani Kühn, se mantiene en el suelo por restos de plantas enfermas y
principalmente por implementos agrícolas utilizados en la labranza de la tierra de

19
cultivo. La costra negra se caracteriza por atacar a brotes, tallos, estolones y tubérculos,
causando necrosis, marchitez foliar, retardando su emergencia y en ocasiones anulando
la germinación de la plántula. En tallos, la necrosis estrangula el tejido frenando el
desarrollo y dando paso a la formación de tubérculos aéreos. En la epidermis de los
tubérculos maduros se forman los esclerocios, estructuras que ayudarán a la
conservación del hongo en el campo (Hooker, 1980; Agrios, 2002).
Rhizoctonia. Solani Kühn es un habitante natural del suelo y está presente en todas
las tierras paperas del mundo. Costra negra es una enfermedad persistente, muy
adaptable y versátil, es considerada de alta importancia ya que disminuye la calidad de
los tubérculos-semillas debido a la presencia de los esclerocios en la piel de los
tubérculos. Cuando la enfermedad afecta la base de la planta ocasiona cierto
enrollamiento de las hojas semejante a los síntomas de PLRV e interfiere en la
evaluación, puede ocasionar el descarte de plantas supuestamente infectadas con el virus
del enrollamiento (PLRV), cuando en realidad se trata de plantas afectadas por R. solani
(Agrios, 2002).
La presencia de la costra negra se ha incrementado en los últimos años en el país.
Según Martín & Torres (1989), R. solani puede llegar a causar la muerte de hasta el 70%
en plántulas. Fankhauser (1997) manifiesta que las pérdidas en campos de agricultores
están alrededor del 25% (INIAP, 2005).
Rhizoctonia solani Kühn puede atacar 250 especies de plantas, está presente en
campo y en las bodegas de almacenamiento de tubérculos donde se encuentra el material

20
vegetal que será utilizado como material de propagación. De las 250 especies
identificadas, 52 corresponden a plantas asociadas al cultivo de papa (Jager et al., 1982).
Costra negra se caracteriza por presentar tolerancia a la acidez, esta propiedad le
permite mayor supervivencia en condiciones de estrés. Se han documentado que ataques
moderados de este hongo pueden causar pérdidas de hasta 20% en los suelos negro
andinos del país. En el Ecuador R. solani ataca una gran diversidad de plantas,
incluyendo arvejas, habas, cebada y trigo. Se ha demostrado que existe una clara
especialización dentro de la población por determinados huéspedes. Existen formas no
patógenas del hongo que eventualmente actúan como antagonistas de las formas
patógenas (Torres, 1997).
1.3.15. Morfología y citología de Rhizoctonia solani Kühn
En el medio de cultivo, las hifas maduras de Rhizoctonia solani Kühn presenta
ramificaciones en ángulos de 90° y 45° a partir de la hifa principal, las cuales se forman
por la constricción de los septos. En la mayoría de las hifas principales se desarrolla una
nueva ramificación al final. Las hifas jóvenes tienen un aspecto transparente y se forman
a manera de “malla” cubriendo la superficie del medio de cultivo. En el campo existe
una variación de color entre hifas aéreas y subterráneas, las subterráneas tienden a
permanecer hialinas, mientras, las aéreas presentan colores amarillentos y marrones
debido a la acumulación de melanina dentro de las paredes celulares. R. solani posee
hifas binucleadas y se caracterizan por ser más finas que las multinucleadas. Su diámetro
oscila entre los 3 a 17 µm y la longitud celular varía entre 50 y 25 µm. Es importante

21
conocer que R. solani no presenta conexiones clamp, conidios, esclerocios diferenciados
dentro de la corteza y médula, tampoco se ha conocido de pigmentos diferentes al
marrón en el color en hifas (Marshall, 1980).
Los esclerocios de R. solani están formados por masas compactas de células
monilioides llamadas clamidiosporas o células escleróticas, se forman en la superficie de
los tejidos vegetales. Durante la formación de esclerocios no hay un patrón claro de
organización hifal. El color de los esclerocios va desde tonalidades marrones a rosáceas.
La forma y el tamaño del micelio varía según el tipo de Rhizoctonia. Los esclerocios de
los grupos de anastomosis 1-1c son pequeños y con formas redondas, mientras que los
esclerocios de otros grupos presentan formas irregulares que varían de 1 a 8 mm de
diámetro (Taheri, 2010).
Los aislados de Rhizoctonia solani Kühn pueden esporular en condiciones
ambientales especiales. Las células de las hifas cortas se bifurcan frecuentemente y
producen densas acumulaciones donde se forman los basidios, que pueden ser
cilíndricos, ovales o esféricos (Whitney, 1964).
En el citoplasma de R. solani se encuentran vacuolas alargadas ubicadas en los
extremos hifales. Se han encontrado grandes vacuolas en células maduras. En las células
monilioides, los esclerocios presentan depósitos con aceite. Dentro del citoplasma de
Rhizoctonia solani se pueden encontrar otros orgánulos celulares como: núcleo,
mitocondria, vacuolas, retículo endoplasmático, ribosomas, micro túbulos, lomasomas,
paredes de hifas, poros y septos (Marshall, 1980).

22
La pared celular, la constituye una simple capa de 80 Å de espesor, con la madurez
del tejido se forman capas adicionales, algunas llegan al espesor de 1 µm. La pared del
septo consta de dos zonas ricas en electrones. El septo puede contener de una a varias
lamelas de acuerdo a la edad celular (Whitney, 1964).
El retículo endoplasmático se encuentra en células jóvenes, se compone de dos capas
paralelas que forman un conjunto cerrado de cisternas porosas o de láminas apiladas.
Suele ser continuo con el parentosoma alrededor del poro del septo y luego se extiende
hacia el citoplasma (Marshall, 1980).
El número de núcleos depende de la Rhizoctonia, por ejemplo, en R. repens las
células son binucleadas. Las células de R. solani, R. zeae y R. oryzae son
multinucleadas. El número de núcleos en células viejas es bajo, lo cual ocurre debido a
la formación de septos en el citoplasma. Los núcleos son de forma oval y se pueden
observar los nucléolos claramente bajo el microscopio de contraste, la membrana
nuclear suele tener conexiones con el retículo endoplasmático (Taheri, 2010).
1.3.16. Grupos de Anastomosis (AG)
De acuerdo con observaciones realizadas in vitro por Carling et al. (2002) se
describe la habilidad de fusionar las hifas del fitopatógeno, a este fenómeno se lo conoce
como anastomosis. Se ha logrado clasificar a R. solani Kühn en 13 grupos de
anastomosis (AG), que se diferencian tanto morfológicamente y en su fisiología por
hospedero, además de características moleculares y secuencias de ADN (Anexo I).
El grupo de anastomosis 3 (AG3) son los más frecuentes en plantas de papa y son
asociados a lesiones que producen necrosis en el tejido, además, de la formación de

23
costras negras en los tubérculos, sus esclerocios pueden llegar a soportar variaciones de
temperaturas. El grupo de anastomosis 4 (AG4) es considerado el más patogénico; a
diferencia del grupo de anastomosis 3, no forma esclerocios, soporta altas temperaturas
y afecta otros cultivos asociados a la papa, también produce la pudrición de raíces; sin
embargo, de este mismo grupo se han encontrado aislamientos no patogénicos en papa
(Acosta et al., 2011).
Se han encontrado varios géneros de Rhizoctonia spp. que forman un grupo endófilo,
estos se asocian a raíces de orquídeas que viven en simbiosis, y son esenciales para la
germinación de semillas y el desarrollo de la planta (Mesa, 2011).
1.3.17. Epidemiología
Rhizoctonia solani Kühn está presente en el suelo de cultivo así como también en la
superficie del tubérculo en forma de esclerocios. El grupo de anastomosis 3 se mantiene
en el suelo en forma de esclerocios asociados a masas de tierra. El grupo de anastomosis
4 permanece como micelio en rastrojos (restos de tejidos de hospedantes). La
inoculación del suelo ocurre cuando un tubérculo-semilla enfermo con costra negra tiene
contacto con la tierra de cultivo; el inóculo se incrementa cuando no se hacen rotaciones
de cultivos como maíz, trigo o cebada; principalmente con el grupo de anastomosis 3
(AG3) en papa. Las condiciones favorables para el desarrollo del hongo se dan con
suficiente humedad del suelo y temperatura de alrededor de 18°C. Es solo en estas
condiciones en donde los esclerocios y los micelios del hongo proliferan y afectan
tejidos jóvenes, especialmente en el estado de plántula del cultivo (Acosta et al., 2011).

24
1.3.18. Ciclo de Vida de R. solani Kühn
Los esclerocios presentes en la tierra de cultivo son estimulados por exudados
producidos durante el crecimiento celular de las plantas y también por la
descomposición de restos de materia orgánica. Estas estructuras producen micelio que al
tener contacto con la planta ataca las primeras barreras externas del tejido vegetal; el
proceso de infección es promovido por la producción de diferentes enzimas
extracelulares (celulasas) que degradan la pared celular de la planta. El hongo continúa
su desarrollo en la superficie externa de la planta causando enfermedad por la formación
de laceraciones en el tejido vegetal que penetran a las células tomando nutrientes para
acelerar su metabolismo (Cotes, 2007).
Como el hongo destruye las células de la planta, las hifas continúan creciendo y
colonizando el tejido necrosado, muchas veces formando más esclerocios. El inóculo es
producido en el interior o exterior del tejido hospedero, ocurriendo sucesivamente
nuevos ciclos cuando los tubérculos están formados y el material vegetal está disponible
(Cotes, 2007).
Hofman & Jongebloed (1988) tras un estudio sobre la infección de plantas de papa,
señalaron la relación entre el desarrollo del micelio de R. solani Kühn en la superficie de
los brotes con la agresividad de la enfermedad. Observaron que bajo el micelio durante
la infección, las hifas pueden penetrar en las células de la epidermis del tejido, proceso
causado por la actividad enzimática del patógeno. La colonización del tejido de la planta
es inicialmente restringida a una o dos capas celulares bajo el micelio, la degradación de
la pared celular y la muerte de sus células es más común en la tercera y cuarta capa

25
celular; finalmente las lesiones pueden llegar hasta cerca de doce capas de células,
alcanzando los haces vasculares del tallo, llegando a diseminarse la infección al floema
y en casos agresivos también el xilema. Las lesiones que circundan un brote causan
necrosis, ocasionando la aparición de rebrotes jóvenes. Concluyeron que el tamaño de la
laceración causada por la infección de R. solani Kühn es proporcional al tamaño del
colchón de infección en la superficie de los brotes, indicando que el daño es producido
después de la penetración del hongo desde el colchón de infección; el tamaño de la
lesión constituye en el tallo un limitante para su crecimiento. Excepto durante la
colonización inicial de células externas, el micelio nunca se observa en tejido sano; por
lo tanto, el tejido colonizado de la planta se restringe a la parte directamente bajo el
colchón de infección. R. solani Kühn coloniza el tejido muerto después de pocos días,
donde puede utilizar los nutrientes liberados de zonas necrosadas para su crecimiento.
Brotes emergidos saludables pueden infectarse severamente y caerse (Hofman &
Jongebloed, 1988).
1.3.19. Síntomas de costra negra
El género Rhizoctonia se caracteriza por la formación de estructuras de resistencia
con redes hifales que unen el micelio con la planta. Esto ha dado origen a la clasificación
de muchos hongos en el género. Los síntomas típicos de la enfermedad de costra negra
en la papa incluyen: la muerte de brotes emergentes, necrosis de tallos subterráneos y
estolones, disminución del sistema radical, cambio de coloración a marrón en las raíces
y la formación de esclerocios en tubérculos maduros y en desarrollo (Tsor et al., 2001).

26
Rhizoctonia solani Kühn ataca a brotes y tallos a partir de esclerocios presentes en el
suelo o en la semilla. Es fácil identificar campos paperos en el país con pobre
emergencia debido a costra negra. En las raíces, los estolones y la parte baja del tallo, el
hongo forma lesiones alargadas, hundidas de color café rojizo. La infección trae consigo
la “poda” de estolones y raíces por necrosis, afectando directamente al volumen de raíz.
En cuadros más avanzados, la infección de raíces y tallos se expresa en la parte aérea
con un enrollamiento hacia la cara superior de las hojas. Se puede presentar clorosis
foliar y formación de tubérculos aéreos como producto de la acumulación de azúcares
que no pueden ser transportados a los tubérculos debido a la estrangulación del tallo. A
veces se observa una mancha de color blanquecina al pie del tallo (INIAP, 2005).
Los síntomas en los tubérculos se pueden parecer a los de la sarna de pradera, pero
difieren de estos últimos por tener esclerocios y rajaduras. Un ataque severo a las yemas
deforma los tubérculos y causa un fenómeno conocido como tubérculos en “formas de
muñecas”. Cuando el follaje madura y muere, el hongo forma esclerocios delgados y
negros en la superficie del tubérculo. Los síntomas son especialmente visibles después
de lavar el tubérculo (INIAP-CIP, 2002; Torres, 1997).
Esta enfermedad no siempre está asociada a reducciones de rendimientos, pero la
formación de esclerocios sobre los tubérculos reduce su calidad comercial, lo que
asociado con el desarrollo de tubérculos mal formados, alteraciones en el tamaño y en el
número de los mismos, resulta en pérdidas económicas considerables en el cultivo (Jager
et al., 1996).

27
Es importante destacar que las plántulas y tubérculos enfermos (Figura 1.3)
presentan:
Estrangulamiento del cuello de la planta
Una mancha algodonosa en el cuello de la planta
Retardo de desarrollo
Hojas superiores marchitas y enrolladas
Clorosis foliar
Formación de tubérculos aéreos
Tubérculos deformes
En la superficie de los tubérculos maduros se forman costras negras llamadas
esclerocios que son estructuras de reproducción de R. solani Kühn.
Figura 1.3. A: Esclerocios de R. solani Kühn en tubérculo maduro. B: Necrosis de raíz causada por R.solani Kühn (Galárraga, 2014).

28
1.3.20. Control de la enfermedad
La incidencia de costra negra puede disminuir significativamente si se mantiene
cuidadosamente el cultivo de la solanácea. Torres (1997) aconseja los siguientes
cuidados para los cultivos:
Usar material germoplásmico libre de esclerocios.
Evitar el monocultivo haciendo rotaciones con cultivos no hospedantes del
hongo, como los cereales.
Realizar la desinfección del material vegetal con fungicidas sistémicos, para
proteger a las plantas durante la emergencia.
Aplicar fungicidas al suelo en el momento de la siembra, disminuye los daños
pero no incrementa los rendimientos.
Sembrar el tubérculo de forma superficial para que la planta tenga una mayor
emergencia. La susceptibilidad de la planta a la infección disminuye con el
desarrollo del follaje.
La rotación con papa debe ser amplia, se aconseja realizarlo a 1:5 ciclos.
Acortar el tiempo en que los tubérculos permanecen en el suelo una vez cortado
el follaje.
No todos los esclerocios son viables. No es necesaria una desinfección de
semillas de papa para un cultivo si la semilla tiene menos de un 25%.

29
CAPÍTULO 2 - MATERIALES Y MÉTODOS
2.1. Participantes
El trabajo de tesis fue realizado por Edison Galárraga, con la dirección de Alma
Koch, MSc. y codirección de Andrés Izquierdo, PhD.
2.2. Zona de estudio
La investigación fue ejecutada en conjunto con el Instituto Nacional Autónomo de
Investigaciones Agropecuarias (INIAP). Se buscó evaluar el comportamiento de
diferentes materiales germoplásmicos de papa frente al fitopatógeno R. solani Kühn a
nivel de invernadero. Se utilizaron ocho variedades de papa de INIAP y doce clones
promisorios distribuidos en cuatro repeticiones y dos testigos aleatorizados en seis
bloques del invernadero.
El estudio molecular se realizó en los laboratorios de Microbiología de la
Universidad de las fuerzas Armadas-ESPE, campus Sangolquí, y en los invernaderos de
la Estación científica de Santa Catalina de INIAP, ubicada en Cutuglagua.
Duración de la Investigación
El proyecto de tesis fue desarrollado en el periodo de abril-diciembre del 2014.
2.3. Revisión bibliográfica
El proceso investigativo se inició con una revisión bibliográfica sobre el manejo de
Rhizoctonia solani Kühn en el laboratorio, su biología y manejo. También se

30
consultaron métodos de aislamiento del hongo a partir de esclerocios, métodos de
inoculación en tierra y la caracterización de la enfermedad tanto en la plántula como en
el tubérculo.
2.4. Fase de campo-toma de muestras
Se tomaron muestras de papa con Rizoctoniosis o costra negra de la localidad de
San Gabriel, Cantón Montúfar, provincia de Carchi, coordenadas 0.6, -77.81667, altura:
2900 msnm, el aislamiento del fitopatógeno se realizó en los laboratorios de
Microbiología de las Fuerzas Armadas (ESPE).
Las muestras se envolvieron en toallas de papel, se colocaron en un sobre de papel y
se rotularon respectivamente. Las muestras de los tubérculos con sintomatología de
costra negra se transportaron en un termo.
2.5. Fase de laboratorio
Esta fase se realizó en los Laboratorios de Microbiología de la Carrera de Ingeniería
en Biotecnología de la Universidad de las fuerzas Armadas (ESPE), que se encuentran
ubicados en Sangolquí, Av. El Progreso, s/n, Cantón Rumiñahui, provincia de Pichincha,
0º 18,81 S; 78º 26, 64 O; altitud: 2516 msnm.

31
2.6. Aislamiento de Rhizoctonia solani Kühn
2.6.1. Preparación de medio de cultivo
Se preparó medio PDA, usando 39 g de PDA (Difco®) por cada litro de agua
destilada, se esterilizó a una temperatura de 121°C durante 15 min, una vez que la
temperatura bajó a los 37°, se colocó 2 µL de cloranfenicol por cada litro de PDA, con el
fin de inhibir el crecimiento bacteriano, y se dispensó 25 mL en cada caja petri estéril
dentro de una cámara de flujo laminar (Figura 2.1) para evitar la contaminación
ambiental (Castellanos et al., 2007).
Figura 2.1 Dispensación de medio PDA en cajas petri dentro de una cámara de flujo laminar (Galárraga,2014).
2.6.2. Desinfección de los explantes tubérculos de Solanum tuberosum
Los tubérculos con esclerocios fueron lavados bajo agua corriente para eliminar el
exceso de tierra, se colocaron las muestras en un vaso de precipitación de 1000 mL
cubierto con una gasa, y se dejó caer el agua sobre el vaso durante una hora. Dentro de
31
2.6. Aislamiento de Rhizoctonia solani Kühn
2.6.1. Preparación de medio de cultivo
Se preparó medio PDA, usando 39 g de PDA (Difco®) por cada litro de agua
destilada, se esterilizó a una temperatura de 121°C durante 15 min, una vez que la
temperatura bajó a los 37°, se colocó 2 µL de cloranfenicol por cada litro de PDA, con el
fin de inhibir el crecimiento bacteriano, y se dispensó 25 mL en cada caja petri estéril
dentro de una cámara de flujo laminar (Figura 2.1) para evitar la contaminación
ambiental (Castellanos et al., 2007).
Figura 2.1 Dispensación de medio PDA en cajas petri dentro de una cámara de flujo laminar (Galárraga,2014).
2.6.2. Desinfección de los explantes tubérculos de Solanum tuberosum
Los tubérculos con esclerocios fueron lavados bajo agua corriente para eliminar el
exceso de tierra, se colocaron las muestras en un vaso de precipitación de 1000 mL
cubierto con una gasa, y se dejó caer el agua sobre el vaso durante una hora. Dentro de
31
2.6. Aislamiento de Rhizoctonia solani Kühn
2.6.1. Preparación de medio de cultivo
Se preparó medio PDA, usando 39 g de PDA (Difco®) por cada litro de agua
destilada, se esterilizó a una temperatura de 121°C durante 15 min, una vez que la
temperatura bajó a los 37°, se colocó 2 µL de cloranfenicol por cada litro de PDA, con el
fin de inhibir el crecimiento bacteriano, y se dispensó 25 mL en cada caja petri estéril
dentro de una cámara de flujo laminar (Figura 2.1) para evitar la contaminación
ambiental (Castellanos et al., 2007).
Figura 2.1 Dispensación de medio PDA en cajas petri dentro de una cámara de flujo laminar (Galárraga,2014).
2.6.2. Desinfección de los explantes tubérculos de Solanum tuberosum
Los tubérculos con esclerocios fueron lavados bajo agua corriente para eliminar el
exceso de tierra, se colocaron las muestras en un vaso de precipitación de 1000 mL
cubierto con una gasa, y se dejó caer el agua sobre el vaso durante una hora. Dentro de

32
la cámara de flujo laminar se cortaron las secciones que contenían esclerocios y se
realizó una desinfección con Hipoclorito de Sodio (NaClO) 0.5% durante 3 min. Se
lavaron tres veces en agua destilada estéril para eliminar los residuos de cloro.
Posteriormente se procedió con la ayuda de un bisturí y unas pinzas estériles a separar
los esclerocios del tubérculo para sembrarlos en medio PDA con cloranfenicol, este
último inhibe el crecimiento de bacterias presentes en el medio ambiente y en el
tubérculo (CIAT, 2008).
2.6.3. Aislamiento
Se incubaron los medios con los esclerocios a 22°C durante cinco días y según las
características morfológicas de cada colonia, se procedió a separar las cepas de
Rhizoctonia solani Kühn haciendo un corte con bisturí en el extremo más alejado de las
demás colonias existentes para garantizar la pureza del cultivo. Se realizó un pase de
medio en PDA con cloranfenicol y se dejó incubar. Se observaron las cajas todos los
días para diferenciar las colonias presentes (CIAT, 2008).
2.6.4. Observación del cultivo al microscopio
Se multiplicaron los medios con Rhizoctonia solani Kühn para luego observar al
microscopio y garantizar la pureza del cultivo. Se cortó un pedazo de cinta adhesiva
transparente y se pegó al micelio del hongo dentro de la caja petri. En un portaobjetos
se colocó una gota de azul de metileno y sobre esta un pedazo de cinta adhesiva con el
micelio del fitopatógeno. R. solani Kühn al microscopio muestra una estructura peculiar,
entre sus ramificaciones forma ángulos de 90°. No se observaron esporas asexuales, R.

33
solani forma esporas sexuales únicamente en condiciones especiales (Acosta et al.,
2011).
2.6.5. Extracción de ADN de R. solani Kühn
Para obtener el material genético se utilizó el protocolo de extracción de ADN en
hongos a partir de medio sólido de Goodwin & Lee (1993), modificado por Galárraga
(2014).
Se tomó aproximadamente 0.5 g de medio sólido, raspando el micelio con un bisturí
estéril y se colocó en vial de 1,5 mL. Se adicionó 200 µL de buffer de lisis (50mM Tris
HCL pH 7.2, 50 mM EDTA, 3% SDS y 1% de Mercaptoetanol) y se calentó al
microondas durante 10 s al máximo nivel.
Se adicionó 200 µL del buffer de lisis y se incubó a 80°C durante 10 min. Se enfrió
a 50°C, se adicionó también 12 µL de proteínasa K y se incubó a 50°C durante 30 min.
Se agregó v/v de etanol absoluto a la master mix y se centrifugó a 10000 g durante
10 min. Una vez centrifugado se tomó el sobrenadante y se transfirió a un tubo nuevo
colocando v/v de cloroformo alcohol isoamílico 24:1 y se centrifugó a 10000 g durante
10 min.
Se tomó el sobrenadante y se colocó en un nuevo tubo eppendorf adicionando 700
µL de isopropanol con 70 µL de acetato de sodio. La mezcla se dejó precipitar 20°C
durante toda la noche. Se centrifugó a 10000 g durante 10 min. Se adicionó 10 µL de
RNAsa (1mg/mL) dejando incubar a 37°C durante 30 min.

34
Se descartó el pellet y se realizaron dos lavados con etanol al 70%. Se eliminó el
etanol y se secó durante 10 min a 50°C en el Termo bloque, se resuspendió con 50 uL de
agua ultra pura.
2.6.6 PCR anidada para la determinación del Grupo de Anastomosis 3 (AG3)
Se realizó una nested PCR para la confirmación del grupo de anastomosis 3 para las
muestras aisladas desde esclerocios (Figura 2.2). En primera instancia se amplificó
regiones ITS (Internal Transcripts Spaces). Estos transcritos tienen un sin número de
ventajas como: un gran número de copias lo que facilita su amplificación (Van Tuinen et
al., 1999), representación de una huella informativa para cada hongo (Gardes & Bruns,
1993) y regiones conservadas (5.8S, 18S y 28S) lo que permite un fácil diseño de
primers (Henrion et al., 1994).
En la primera etapa se utilizaron los primers generalistas ITS1 e ITS4 para
amplificar un fragmento de ADN ribosomal de aproximadamente de 700 pb, con el fin
de lograr mayor especificidad. Con el producto de PCR obtenido al amplificar la región
ITS1-5.8S-ITS2 se utilizaron los primers Rs1F2 y Rs2R1 específicos para AG 3 que
amplifican un fragmento de alrededor de 500 pb (Lees, K. et al., 2002).
2.6.7. Amplificación de las regiones ITS1-5.8S-ITS2
En la primera PCR para la amplificación de las regiones ITS1-5.8S-ITS2 del ADN
ribosomal de R. solani Kühn se utilizó: el primer forward ITS1 (5´-
TCCGTAGGTGAACCTGCGG-3´) y el primer reverse ITS4 (5´-
TCCTCCGCTTATTGATATGC-3´). Amplifican un fragmento de un tamaño

35
aproximado de 700 pb. La reacción de PCR con un volumen final de 25 µL contenía: 1X
buffer de PCR, 1.5 mM de MgCl2, 0.2 mM de dNTP´s, 10 mM de cada primer, una
unidad de Taq DNA polimerasa y 1,5 µL de ADN. Los ciclos de amplificación
utilizados fueron modificados de Fernández (2011) como se describe a continuación: un
ciclo inicial de desnaturalización a 95 °C por 2 min; seguido de 35 ciclos de 95 °C por
45 s, 63 °C por 60 s y 72 °C por 90 s; con un ciclo final de extensión de 72 °C por 5 min
(Fernández, 2011; White et al., 1990).
2.6.8. Amplificación de regiones específicas para AG-3
La segunda PCR para la confirmación del grupo de anastomosis 3 (AG-3) utilizó los
iniciadores forward Rs1F2 (5′-TTGGTTGTAGCTGGTCTATTT-3′) y reverse Rs2R1
(5′TATCACGCTGAGTGGAACCA-3′). La reacción de PCR con un volumen final de
25 µL contenía: 1X buffer de PCR, 2 mM de MgCl2, 0.2 mM de dNTP´s, 0.3 mM de
cada primer, una unidad de Taq DNA polimerasa y 2 µL del producto de PCR de la
primera fase. Los ciclos de amplificación a utilizarse fueron: un ciclo inicial de
desnaturalización a 95 °C por 2 min; seguido de 35 ciclos de 95 °C por 45 s, 63 °C por
60 s y 72 °C por 90 s; con un ciclo final de extensión de 72 °C por 5 min (Fernández,
2011).

36
Figura 2.2 Armada de PCR dentro de una cámara de flujo laminar (Galárraga, 2014).
2.6.9. Electroforesis en gel de agarosa
Los productos de PCR se corrieron en un gel de agarosa al 2%, para lo cual se pesó
1.6 g de agarosa (DifcoTM) por cada 80 mL de TBE 1X, se calentó en estufa con
agitación y se llevó al microondas donde fue calentada a máxima potencia durante
cortos intervalos de tiempo para evitar que hierva y se derrame. Diluida la agarosa, se
dejó enfriar y se colocó el SYBR® Green (InvitrogenTM) 1.5X como fluoróforo para la
visualización de ADN una vez corrida la muestra.
Las muestras se corrieron a 300mA y 100V durante una hora en un ambiente libre de
luz.
2.6.10. Multiplicación de Rhizoctonia solani Kühn
Se realizaron cortes de aproximadamente 0.5 cm en el cultivo madre aislado y se
transfirieron a cajas petri con medio PDA fresco, se sellaron con parafilm y se incubaron
a 24°C.
36
Figura 2.2 Armada de PCR dentro de una cámara de flujo laminar (Galárraga, 2014).
2.6.9. Electroforesis en gel de agarosa
Los productos de PCR se corrieron en un gel de agarosa al 2%, para lo cual se pesó
1.6 g de agarosa (DifcoTM) por cada 80 mL de TBE 1X, se calentó en estufa con
agitación y se llevó al microondas donde fue calentada a máxima potencia durante
cortos intervalos de tiempo para evitar que hierva y se derrame. Diluida la agarosa, se
dejó enfriar y se colocó el SYBR® Green (InvitrogenTM) 1.5X como fluoróforo para la
visualización de ADN una vez corrida la muestra.
Las muestras se corrieron a 300mA y 100V durante una hora en un ambiente libre de
luz.
2.6.10. Multiplicación de Rhizoctonia solani Kühn
Se realizaron cortes de aproximadamente 0.5 cm en el cultivo madre aislado y se
transfirieron a cajas petri con medio PDA fresco, se sellaron con parafilm y se incubaron
a 24°C.
36
Figura 2.2 Armada de PCR dentro de una cámara de flujo laminar (Galárraga, 2014).
2.6.9. Electroforesis en gel de agarosa
Los productos de PCR se corrieron en un gel de agarosa al 2%, para lo cual se pesó
1.6 g de agarosa (DifcoTM) por cada 80 mL de TBE 1X, se calentó en estufa con
agitación y se llevó al microondas donde fue calentada a máxima potencia durante
cortos intervalos de tiempo para evitar que hierva y se derrame. Diluida la agarosa, se
dejó enfriar y se colocó el SYBR® Green (InvitrogenTM) 1.5X como fluoróforo para la
visualización de ADN una vez corrida la muestra.
Las muestras se corrieron a 300mA y 100V durante una hora en un ambiente libre de
luz.
2.6.10. Multiplicación de Rhizoctonia solani Kühn
Se realizaron cortes de aproximadamente 0.5 cm en el cultivo madre aislado y se
transfirieron a cajas petri con medio PDA fresco, se sellaron con parafilm y se incubaron
a 24°C.

37
La multiplicación de los cultivos se realizó con el fin de tener suficiente material
biológico para la siembra en semillas de avena, que sirven como medio de soporte en la
inoculación de macetas con tierra, en donde se sembró el material germoplásmico de
variedades INIAP y clones promisorios, para determinar su resistencia ante costra negra.
2.6.11. Preparación del inóculo de R. solani Kühn
El inóculo de R. solani Kühn se preparó en semilla de avena previamente
autoclavada. El método utilizado fue tomado de Paulitz & Schroeder, 2005 y
modificado. Se utilizó una proporción de 2:1 entre avena y agua. En Erlenmeyers de 1 L
se distribuyó la mezcla y se autoclavó dos veces a 121°C durante 15 minutos. Una vez
enfriada la mezcla ya estéril, en una cámara de flujo laminar, se procedió a colocar 5 g
de cultivo de R. solani Kühn de siete días en cada Erlenmeyer (Figura 2.3). El inóculo
permaneció en los Erlenmeyers durante 18 días en un ambiente de 24°C sin luz.
Se realizaron dos hidrataciones al inóculo, dentro de una cámara de flujo, cada una
de 100 mL (Paulitz & Schroeder, 2005).
Figura 2.3 A: cortes en el medio PDA con R. solani. B: Preparacion del inóculo dentro de una cámara deflujo laminar (Galárraga, 2014).

38
2.6.12. Inoculación de Tierra con germoplasmas
Las germoplasmas (20 germoplasmas distintos entre variedades INIAP y clones
promisorios) de papa fueron lavados con agua corriente y secados a la sombra, para
después ser sembrados y tapados con una parte de tierra estéril. Se realizaron cuatro
repeticiones con dos tratamientos testigos (sin inóculo) (Tabla 2.1).
Tabla 2.1 Materiales germoplásmicos evaluados (Galárraga, 2014)
N° Variedad Tipo1 INIAP-ESTELA Variedad mejorada2 INIAP-FRIPAPA Variedad mejorada3 INIAP-PUCA
SHUNGOVariedad mejorada
4 INIAP-NATIVIDAD
Variedad mejorada
5 INIAP-SUPERCHOLA
Variedad mejorada
6 INIAP-VICTORIA Variedad mejorada7 INIAP-YANA
SHUNGOVariedad mejorada
8 RUBI Variedad mejorada9 07-32-15 Clon promisorio
10 12-2-6 Clon promisorio
11 12-2-18 Clon promisorio12 12-3-3 Clon promisorio13 12-4-71 Clon promisorio14 12-4-46 Clon promisorio15 12-4-170 Clon promisorio16 12-6-1 Clon promisorio17 12-6-24 Clon promisorio18 97-25-3 Clon promisorio19 98-2-6 Clon promisorio20 98-38-12 Clon promisorio

39
Se utilizaron 20 g de avena con R. solani para inocular 3 Kg de tierra autoclavada
(una parte de arena de río y tres partes de arena de zanja). Se inoculó un total de 120
macetas con una capacidad de 3 Kg cada una (Figura 2.4).
Figura 2.4 A: Inoculación de tierra para la siembra de germoplasmas. B: Materiales inoculados eninvernadero (Galárraga, 2014).
2.6.13. Control de Inóculo
Se realizó un control de inóculo a los 90 días de haber empezado el experimento en
el invernadero. Se tomó 100 g de muestra de suelo a una profundidad de 10 cm dentro
de la maceta. Se mezclaron con 2.5 L de agua, se dejó reposar la suspensión durante 30
s y se filtró. Se colocaron entre dos capas de papel estéril y se secó. Se colocó la tierra en
platos petri con PDA más cloranfenicol y se incubó 48 h a 24°C. Se traspasó el hongo a
un nuevo medio con PDA se incubaron 48 h a 24°C. Se observó el micelio al
microscopio.

40
2.6.14. Manejo del experimento
Luego de la siembra, se hizo un riego y posteriormente se mantuvo el sustrato en
capacidad de campo. A los 47, 62 y 77 días se realizaron fertilizaciones con 20-19-19-
0.5 MgO (Nitrofosca foliar). Los controles de plagas y enfermedades se realizaron de
acuerdo a la exigencia del cultivo.
El ensayo se mantuvo durante seis meses en condiciones de invernadero en la
estación experimental “Santa Catalina” de INIAP en la parroquia Cutuglagua del cantón
Quito (Figura 2.5).
Figura 2.5 A: Invernadero de la estación científica Santa catalina donde se realizó el ensayo. B:Disposición de los bloques en el invernadero (Galárraga, 2014).
2.7. Método de evaluación
El diseño estadístico que se utilizó fue en bloques al azar con un arreglo factorial
con cuatro repeticiones y dos tratamientos testigo. Las evaluaciones se realizaron

41
semanalmente tomando datos de la altura de las plantas, y de la sintomatología aérea de
costra negra (hojas deformes y amarillentas, defoliación, plantas marchitas y muertas).
Se midió el porcentaje de emergencia a los 25, 32 y 49 días después de la siembra. A
la cosecha se evaluó el porcentaje de daño de raíz y base de tallo (Clive, 1971), el
porcentaje de esclerocios en el tubérculo y el rendimiento expresado en kg/maceta (ver
Tabla 2.2).
= 0 ∗ + 1 ∗ + 2 ∗ + 3 ∗ + 4 ∗4 ∗ é ∗ 100Tabla 2.2 Criterio de Clive para la evaluación de tubérculos (Clive, 1971).
Escala Infección
(%)
Reacción
0 0 Sana
1 1 a10 Muy ligera
2 11 a 25 ligera
3 26 a 50 Moderada
4 > 51 Severa
Se realizó un ANOVA y pruebas de medias con diferencia mínima de significancia
además de un análisis de Tukey para cada variable para poder seleccionar los materiales
germoplásmicos que presentan tolerancia a R. solani.

42
CAPÍTULO 3 – RESULTADOS
3.1. Aislamiento de Rhizoctonia solani Kühn
El aislamiento de R. solani a partir de esclerocios de tubérculos infectados fue
conseguido en medio PDA con cloranfenicol (Figura 3.1). Para obtener un cultivo puro
se hicieron constantes traspasos de medio realizando cortes en los extremos del micelio
(Castellanos et al., 2007).
Figura 3.1 Cultivo aislado de Rhizoctonia solani Kühn en medio PDA en caja petri (Galárraga, 2014).
Se observó al microscopio la morfología del micelio del hongo con la técnica de
tinción de azul de lactofenol (Figura 3.2) para la comprobación morfológica del género
Rhizoctonia (French, 1982).
El cultivo puro de Rhizoctonia solani Kühn se caracterizó por tener un color blanco
en el micelio joven. A medida que maduraba, se tornó de un color marrón. R. solani se
42
CAPÍTULO 3 – RESULTADOS
3.1. Aislamiento de Rhizoctonia solani Kühn
El aislamiento de R. solani a partir de esclerocios de tubérculos infectados fue
conseguido en medio PDA con cloranfenicol (Figura 3.1). Para obtener un cultivo puro
se hicieron constantes traspasos de medio realizando cortes en los extremos del micelio
(Castellanos et al., 2007).
Figura 3.1 Cultivo aislado de Rhizoctonia solani Kühn en medio PDA en caja petri (Galárraga, 2014).
Se observó al microscopio la morfología del micelio del hongo con la técnica de
tinción de azul de lactofenol (Figura 3.2) para la comprobación morfológica del género
Rhizoctonia (French, 1982).
El cultivo puro de Rhizoctonia solani Kühn se caracterizó por tener un color blanco
en el micelio joven. A medida que maduraba, se tornó de un color marrón. R. solani se
42
CAPÍTULO 3 – RESULTADOS
3.1. Aislamiento de Rhizoctonia solani Kühn
El aislamiento de R. solani a partir de esclerocios de tubérculos infectados fue
conseguido en medio PDA con cloranfenicol (Figura 3.1). Para obtener un cultivo puro
se hicieron constantes traspasos de medio realizando cortes en los extremos del micelio
(Castellanos et al., 2007).
Figura 3.1 Cultivo aislado de Rhizoctonia solani Kühn en medio PDA en caja petri (Galárraga, 2014).
Se observó al microscopio la morfología del micelio del hongo con la técnica de
tinción de azul de lactofenol (Figura 3.2) para la comprobación morfológica del género
Rhizoctonia (French, 1982).
El cultivo puro de Rhizoctonia solani Kühn se caracterizó por tener un color blanco
en el micelio joven. A medida que maduraba, se tornó de un color marrón. R. solani se

43
caracteriza por formar esclerocios a través de la fusión de hifas, en este proceso se pudo
observar la segregación de una sustancia de color ambar previo a la formación de los
esclerocios. Estas estructuras son capaces de sobrevivir a la deshidratación y a
temperaturas bajas en condiciones de campo (Torres, 1997).
Figura 3.2 Tinción de hifas de Rhizoctonia solani Kühn vista en el objetivo de 40X. A: Se observaramificación de la hifa principal en angulo de 90°. B: Se observa constricción de septos y ramificaciones
de hifas jóvenes (Galárraga, 2014).
3.2. Extracción de ADN de Rhizoctonia solani Kühn
La metodología de Goodwin & Lee (1993) modificada por Galárraga (2014)
permitió la extracción de ADN en concentraciones elevadas y sin contaminación de
compuestos fenólicos que interfieran con los demás procedimientos.
La concentración promedio de las muestras procesadas presentó un valor de 1676
ng/µL y una pureza promedio de 1.87; lo que sugiere que el protocolo empleado fue
aceptable para este tipo de muestras (Tabla 3.1).

44
Las concentraciones óptimas necesarias para una PCR rodean los valores de 15
ng/µL, por lo que se realizó alícuotas con agua destilada grado PCR y se conservaron las
muestras a -20°C para su posterior uso. Las muestras de material genético pueden
conservarse a -20°C durante periodos intermedios de tiempo, luego se debe conservar a -
80°C, temperatura en la cual puede mantenerse durante años (Goodwin & Lee, 1993).
Tabla 3.1. Valores de concentración y absorbancias de ADN total extraído de Rhizoctonia solani
Kühn con el método de extracción a partir de medio sólido de Goodwin & Lee (1993).
Muestra Concentración(ng/µL)
A(260/230)
A(260/280)
M1-1 357,8 1,9 1,897
M1-2 1853 2,26 1,875M1-3 3339 1,46 1,678
M2-1 950,8 2,05 1,875M2-2 2994 1,775 1,75
M2-3 563 2,03 2,16
La cuantificación fue realizada en el equipo Nanodrop® y los resultados dados por
el sistema se muestran en el ANEXO II.
3.3. Nested PCR para la confirmación de Rhizoctonia solani AG3
3.3.1. Amplificación de Regiones ITS1-5,8S-ITS2
La amplificación de la región ITS1-5,8S-ITS2 del ADN de Rhizoctonia solani Kühn
obtenido de esclerocios de Solanum tuberosum mostró buena integridad de bandas, sin
productos inespecíficos y sin la amplificación del control negativo (Figura 3.3). La
región ITS1-5,8S-ITS2 mostró un tamaño aproximado de 700 pb (Hidalgo et al., 1997).

45
Figura 3.3 Electroforesis de la región ITS1-5,8-ITS2 con su control negativo C (-).Gel de agarosa al2% teñido con 0,05 µg/mL (v/v) de SYBR® Safe gel stain. Corrida a 100V por 1 h. MM: Marcador de
peso molecular 100 bp (Invitrogen ®) (Galárraga, 2014).
3.3.2. PCR específica para Grupo de Anastomosis 3 (AG3) de Rhizoctonia solani
Kühn
Para la confirmación del Grupo de Anastomosis 3 (AG3) se utilizaron los productos
de PCR de la amplificación de las regiones ITS1-5,8S-ITS2. Los primers utilizados
Rs1R2 y Rs2R1, forward y reverse respectivamente, fueron diseñados por Lees et al.
(2002). Las muestras de Rhizoctonia solani Kühn fueron positivas al grupo de
anastomosis 3 (AG3), presentando bandas de aproximadamente 500 pb, lo cual reporta
Lees et. al (2002) en sus ensayos. No se presentaron amplificaciones de productos
inespecíficos ni del control negativo C (-) (Figura 3.4).

46
Figura 3.4 Amplificación de la región específica para AG3 con su control negativo C (-).Gel deagarosa al 2% teñido con 0,05 µg/mL (v/v) de SYBR® Safe gel stain. Corrida a 100V por 1 h. MM:
Marcador de peso molecular 100 bp (Invitrogen ®) (Galárraga, 2014)..
3.4. Preparación del Inóculo
La semilla de avena fue humedecida dentro de una cámara de flujo laminar con
agua estéril, conforme requerían las condiciones del medio (Figura 3.5-A). Al principio
se cubrieron los Erlenmeyers con parafilm pero a la segunda semana se colocaron
tapones de algodón con gasa estériles para mantener la aireación del inóculo. El inóculo
estuvo listo a los 29 días (Figura 3.5-B) y en ese tiempo se realizaron alrededor de cinco
humidificaciones de 100 mL cada una.

47
Figura 3.5. A: Preparación del inóculo de Rhizoctonia solani Kühn en semillas de avena. B: InoculoR. solani a los 29 días (Galárraga, 2014).
3.5. Sintomatología aérea
Dentro de invernadero el comportamiento de Rhizoctonia solani Kühn fue distinto al
comportamiento de campo descrito por Acosta et al. (2011). Los cuatro bloques del
invernadero presentaron marchitez foliar a partir de los 45 días de empezado el ensayo.
La sintomatología de costra negra fue enmascarada por otras enfermedades originadas
del tubérculo semilla, además, de plagas secundarias y no pudo ser evaluada. Se observó
marchitez foliar en tratamientos testigo, este es el caso de los materiales
germoplásmicos: I-Victoria, 98-2-6, 12-4-170 y I-Rubí. Como control de plagas y con el
fin de fortalecer las plantas hasta la tuberización se utilizó nitrofosca foliar (20-19-19-
0,5 MgO). Durante el ensayo no se observó la formación de tubérculos aéreos en
ninguno de los bloques.
3.6. Análisis estadístico
Con los datos obtenidos de las mediciones y de la cosecha de los tubérculos se
realizó un análisis de normalidades para ver el comportamiento de los datos. Una vez

48
identificado el tipo de datos (paramétricos o no paramétricos) se procedió a realizar un
análisis comparativo de Tukey para encontrar la significancia de los datos y así poder
clasificar las variedades según su sintomatología. Se agruparon las variedades en:
altamente tolerantes (AT), tolerantes (T), parcialmente tolerantes (PT) o susceptibles
(S). Se utilizó el software estadístico INFOSTAT.
3.7. Porcentaje de emergencia
Todas las variedades de todos los tratamientos mostraron una emergencia del 100%
a partir de los ocho días de cultivo a excepción de la variedad mejorada I-Superchola la
cual mostró un 83.3% de emergencia, la primera repetición de este tratamiento mostró
pudrición del tubérculo por Erwinia carotovora.
3.8. Altura de las variedades inoculadas
Se realizaron controles periódicos de la altura de las plantas a partir de los tres
meses de iniciado el ensayo. El análisis de varianza para la altura de la planta presentó
una baja significación estadística de la diferencia de alturas entre los tratamientos
inoculados respecto a los tratamientos testigo. La media de la altura fue 41.83 cm y el
coeficiente de variación para la altura de las plantas fue de 30.05% como se muestra en
la Tabla 3.2.

49
Tabla 3.2 Análisis de varianza de la altura de las plantas de papa del ensayo de evaluación de laresistencia ante Rhizoctonia solani Kühn.
F.V. GL CM FcTOTAL 117TRATAMIENTOS 38 284.90 *ERROR 79 151.54X (%) 41.83CV (%) 30.05
La media aritmética del promedio de altura para los 40 tratamientos fue analizada
estadísticamente mediante un análisis de Tukey. La variedad I-Rubí resultó ser la de
menor tamaño en ambos tratamientos (Figura 3.6).
En los tratamientos inoculados, la menor altura de planta, se presentó en la variedad
mejorada I-Rubí con un valor de 20.25 cm. En rangos de alturas compartidas (AB) se
encontraron el resto de tratamientos con un rango de 28.75 cm (clon promisorio 12-6-1)
a 55.50 cm (variedad mejorada I-Puca) (Tabla 3.6).
En los tratamientos testigos con la menor altura de planta, se presentó la variedad
mejorada I-Rubí con 21 cm de altura de planta mientras que el resto de variedades
mostraron rangos compartidos (AB) con un promedio de 27.50 cm (Variedad mejorada
I-Estela) hasta 61.50 cm (Clon promisorio 07-32-15), la mayor altura para este
tratamiento la tiene el clon promisorio 12-3-3 con un valor promedio de 65.50 cm (Tabla
3.3).

50
Figura 3.6 Diagrama de barras para las alturas de las plantas con su error estándar (Galárraga,
2014).
Las alturas de todos los tratamientos incluyendo inoculados y testigos se mostraron
similares. Los tratamientos testigo mostraron una mínima diferencia estadística en su
altura a comparación con los bloques inoculados.

51
Tabla 3.3 Alturas promedio y pruebas de Tukey para la variable altura de planta en el ensayo deevaluación de la resistencia ante Rhizoctonia solani Kühn.
TRATAMIENTOS INOCULADOS TRATAMIENTOS TESTIGOSPROMEDIOS RANGOS PROMEDIOS RANGOS
I-RUBI 20.25 A* I-RUBI 21.00 A*
12-6-1 28.75 AB I-ESTELA 27.50 AB
I-VICTORIA 31.50 AB I-VICTORIA 33.00 AB
12-4-46 33.50 AB I-FRIPAPA 33.00 AB
98-2-6 34.25 AB 12-2-18 33.00 AB
I-FRIPAPA 35.25 AB 98-38-12 35.00 AB
I-NATIVIDAD 35.75 AB 12-6-1 36.50 AB
I-SUPERCHOLA 35.75 AB I-YANASHUNGO
38.50 AB
12-2-6 36.50 AB SUPERCHOLA 40.00 AB
I-YANA SHUNGO 38.50 AB 98-2-6 40.50 AB
I-ESTELA 40.00 AB I-NATIVIDAD 45.00 AB
98-38-12 44.50 AB 12-2-6 45.50 AB
12-4-170 45.25 AB 12-6-24 47.50 AB
12-2-18 45.75 AB 12-4-46 47.50 AB
12-3-3 46.00 AB 12-4-170 55.50 AB
12-6-24 46.25 AB 97-25-3 55.50 AB
12-4-71 47.00 AB 12-4-71 56.00 AB
07-32-15 48.50 AB I-PUCA SHUNGO 56.00 AB
97-25-3 51.25 AB 07-32-15 61.50 AB
I-PUCA SHUNGO 55.50 AB 12-3-3 65.50 B
3.9. Evaluación a la cosecha de los tubérculos
La evaluación de los tubérculos se realizó a los seis meses de comenzado el ensayo
en invernadero. Se cortó el follaje una semana antes de la cosecha con el fin de acelerar
la madurez de los tubérculos, garantizando la acumulación de materia seca, factor
importante en la calidad del tubérculo.
A la cosecha de los tubérculos se observó que los veinte materiales inoculados,
mostraron similar sintomatología en todos los bloques, la necrosis de raíz, fue el factor

52
común en todas las variedades. El 80% de las raíces y tallos de los tratamientos
inoculados se encontraban necrosadas por R. solani. Este síntoma se observó con mayor
intensidad en la variedad I-Rubí del cuarto bloque (Figura 3.7-A) y en el clon promisorio
97-25-3 del segundo bloque (Figura 3.7-B).
Figura 3.7 A: Necrosis en tallo de la variedad I-Rubí del cuarto bloque a los cinco meses deiniciado el ensayo. B: Necrosis de tallo en el clon promisorio 97-25-3 del segundo bloque a los 135 días
del cultivo (Galárraga, 2014).
Se encontró la formación de tubérculos en forma de muñecos en las variedades: I-
Estela, I-Victoria y en el clon promisorio 12-2-6. En las variedades: I-Fripapa, I-
Superchola, I-Yana Shungo, I-Rubí, y en los clones 98-2-6 y 98-38-12 se encontró
síntomas de sarna causados por la bacteria Streptomyces scabies. Los tratamientos
testigo no presentaron sintomatología en los tubérculos y mostraron buena apariencia.
En el ANEXO III se describe la sintomatología presentada por maceta de cada
variedad por cada bloque.
3.10. Severidad de la infección
Se utilizó el criterio de Clive para la evaluación y clasificación de los tubérculos
según la sintomatología de costra negra. Se realizó un análisis de varianza y se encontró

53
un coeficiente de variación (C.V.) de 39.16% y una media de infección de esclerocios de
22.50% en variedades inoculadas, una suma de cuadrados de 351.74 y un valor de
contraste (FC) de baja significancia (≤ 0.05) (Tabla 3.4).
Tabla 3.4 Análisis de varianza del porcentaje de severidad de la infección de papas del ensayo de
evaluación de la resistencia ante Rhizoctonia solani Kühn.
F.V. GL CM FcTOTAL 79TRATAMIENTOS 19 351.74 *REPETICIONES 3 60.64ERROR 57 77.66X (%) 22.52C.V. (%) 39.16
Para el análisis de Tukey del porcentaje de infección del tubérculo (Tabla 3.8) se
encontraron cuatro rangos de significancia los cuales relacionaron y clasificaron los
materiales germoplásmicos evaluados según la tolerancia presentada. Los resultados
muestran que al menos uno de los tratamientos inoculados fue diferente a los demás. En
el primer rango compartido (AB), se encuentran 17 de los materiales germoplásmicos
evaluados. La variedad mejorada I-Rubí presentó el valor más alto de infección con
34.78%, seguido por el clon 97-25-3 con 33.10%, I-Fripapa con 30.13% y el clon 98-38-
12 con 29.30% de severidad (Figura 3.8). El clon promisorio 12-2-6 obtuvo el menor
valor dentro de este rango con un promedio de infección de 14.38%, mostrando una
tolerancia parcial (TP) a Rhizcotonia solani Kühn.

54
Figura 3.8 Tubérculos con costra negra de la variedad mejorada I-Rubí. A: I-Rubí primera repetición. B:I-Rubí segunda repetición. C: I-Rubí tercera repetición. D: I-Rubí cuarta repetición (Galárraga, 2014).
Los más bajos niveles de infección por costra negra se encontraron dentro del rango
compartido CD. El clon promisorio 12-4-170 obtuvo un valor de 6,93% de infección en
tubérculos (Figura 3.9-A). El clon promisorio 12-6-1 obtuvo un valor de 5.62% de
severidad en el tubérculo, mostrando una tolerancia moderada (TM) a costra negra
(Figura 3.9-B). La variedad mejorada I-Puca es considerada como altamente tolerante
(AT) a costra negra con un valor de 0.00% de severidad (Figura 3.9- C).

55
Figura 3.9 A: Tubérculos del clon promisorio 12-4-170, primer bloque. B: Tubérculos del clonpromisorio 12-6-1, primer bloque. C: planta y tubérculos de la variedad 3 correspondiente a I-Puca, cuarto
bloque (Galárraga, 2014).
De los 20 materiales germoplásmicos en estudio: la variedad (I-Puca) presentó alta
tolerancia; dos se mostraron tolerantes (clones promisorios: 12-4-170 y 12-6-1); nueve
se presentaron parcialmente tolerantes y ocho fueron encontrados susceptibles (Tabla
3.5). Las variedades más susceptibles a Rhizoctonia solani Kühn fueron: I-Rubí, seguido
del clon promisorio 97-25-3 e I-Fripapa. Ninguno de los testigos presentó
sintomatología (Figura 3.10).

56
Figura 3.10 Diagrama de cajas con sus media de la severidad de las variedades de papa inoculadas(Galárraga, 2014).
Las variedad y clones promisorios: I-Estela, 98-2-6, I-Yana Shungo, 12-3-3, mostraron
Suceptibilidad (S) a Rhizoctonia solani Kühn con una media de 28.67% de severidad.
Estas variedades mostraron gran número de esclerocios en el tubérculo y altos niveles de
necrosis en raíz.

57
Tabla 3.5 Resultados del análisis de tukey, rangos de los valores promedios del porcentaje deinfección de costra negra en tubérculos de papa. Tipo de resistencia: alta resistencia (AR), resistencia ®,
resistencia parcial (RP) o susceptibilidad (S).
TRATAMIENTOS INOCULADOS TRATAMIENTOS TESTIGO
VARIEDAD % DEINFECCIÓN
RANGO TIPO DERESISTENCIA
VARIEDAD %INF
RUBI 34.78 A S RUBI 0.00
97-25-3 33.10 A S 97-25-3 0.00
I-FRIPAPA 30.13 A S I-FRIPAPA 0.00
98-38-12 29.30 AB S 98-38-12 0.00
12-3-3 29.15 AB S 12-3-3 0.00
I-YANASHUNGO
28.63 ABC S I-YANA SHUNGO 0.00
98-2-6 28.60 ABC S 98-2-6 0.00I-ESTELA 28.33 ABC S I-ESTELA 0.00
I-NATIVIDAD 25.00 ABC PT I-NATIVIDAD 0.0012-6-24 24.75 ABC PT 12-6-24 0.0012-2-18 24.53 ABC PT 12-2-18 0.0012-4-71 24.38 ABC PT 12-4-71 0.00
I-SUPERCHOLA 23.03 ABCD PT I-SUPERCHOLA 0.0007-32-15 22.33 ABCD PT 07-32-15 0.00
I-VICTORIA 19.10 ABCD PT I-VICTORIA 0.0012-4-46 18.03 ABCD PT 12-4-46 0.0012-2-6 14.38 ABCD PT 12-2-6 0.00
12-4-170 6.93 BCD T 12-4-170 0.0012-6-1 5.62 CD T 12-6-1 0.00
I-PUCA SHUNGO 0.00 D AT I-PUCA SHUNGO 0.00
3.11. Número de tubérculos
En el análisis de varianza del número de tubérculos por tratamiento, no se
encontraron diferencias estadísticas significativas entre los tratamientos. Se encontró un
coeficiente de variación (C.V.) de 50.08% y una media de producción de 7.91 tubérculos
por maceta (Tabla 3.6). Para los tratamientos testigo se observó coeficientes de variación
de 38.60%, una media de producción de 9.90 tubérculos por maceta, una suma de
cuadrados de 5.97 para los tratamientos inoculados y de 3.18 para los tratamientos no

58
inoculados. Se encontró que en ambos tratamientos la variable número de tubérculos con
respecto al estado de la planta (inoculada y no inoculada) no tienen relación (Figura
3.11).
Figura 3.11. Diagrama de la variable número de tubérculos por variedad con sus respectivas medias(Galárraga, 2014)
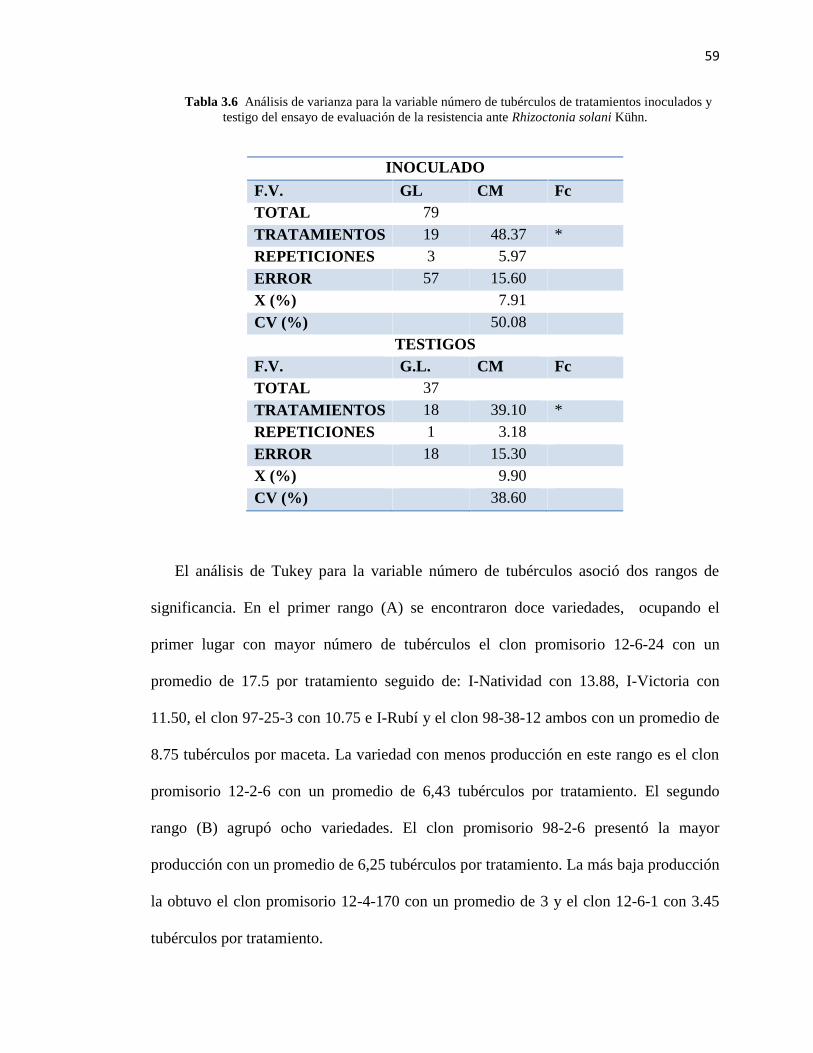
59
Tabla 3.6 Análisis de varianza para la variable número de tubérculos de tratamientos inoculados ytestigo del ensayo de evaluación de la resistencia ante Rhizoctonia solani Kühn.
INOCULADO
F.V. GL CM FcTOTAL 79
TRATAMIENTOS 19 48.37 *REPETICIONES 3 5.97
ERROR 57 15.60X (%) 7.91
CV (%) 50.08TESTIGOS
F.V. G.L. CM FcTOTAL 37
TRATAMIENTOS 18 39.10 *REPETICIONES 1 3.18
ERROR 18 15.30X (%) 9.90
CV (%) 38.60
El análisis de Tukey para la variable número de tubérculos asoció dos rangos de
significancia. En el primer rango (A) se encontraron doce variedades, ocupando el
primer lugar con mayor número de tubérculos el clon promisorio 12-6-24 con un
promedio de 17.5 por tratamiento seguido de: I-Natividad con 13.88, I-Victoria con
11.50, el clon 97-25-3 con 10.75 e I-Rubí y el clon 98-38-12 ambos con un promedio de
8.75 tubérculos por maceta. La variedad con menos producción en este rango es el clon
promisorio 12-2-6 con un promedio de 6,43 tubérculos por tratamiento. El segundo
rango (B) agrupó ocho variedades. El clon promisorio 98-2-6 presentó la mayor
producción con un promedio de 6,25 tubérculos por tratamiento. La más baja producción
la obtuvo el clon promisorio 12-4-170 con un promedio de 3 y el clon 12-6-1 con 3.45
tubérculos por tratamiento.

60
En los tratamientos testigo se observó tres rangos de significancia. En el primer
rango (A) se destacan: el clon promisorio 12-6-24 con un promedio de 20.50 tubérculos
por tratamiento y la variedad mejorada I-Natividad con un promedio de 15.50
tubérculos. El segundo rango compartido (BC), agrupó a 17 germoplasmas en los que
incluyen las variedades: I-Victoria, I-Superchola, I-Friapa, I-Yana Shungo, I-Estela e I-
Puca Shungo. En el último rango (C), la variedad I-Yana Shungo con un promedio de
4.50 tubérculos por tratamiento (Tabla 3.7).
Tabla 3.7 Prueba de Tukey para la variable número de tubérculos del ensayo de evaluación de laresistencia ante Rhizoctonia solani Kühn.
TRATAMIENTOSINOCULADOS
TRATAMIENTOSTESTIGOS
VARIEDAD PROMEDIO RANGO VARIEDAD PROMEDIO RANGO12-6-24 17.50 A 12-6-24 20.50 AI-NATIVIDAD 13.88 AB I-NATIVIDAD 15.50 ABI-VICTORIA 11.50 ABC 12-4-46 15.00 BC97-25-3 10.75 ABC 97-25-3 14.50 BCI-RUBI 8.75 ABC I-SUPERCHOLA 14.00 BC98-38-12 8.75 ABC 98-2-6 13.00 BC12-4-71 8.50 ABC 12-2-18 13.00 BC07-32-15 8.25 ABC I-VICTORIA 11.00 BCI-SUPERCHOLA 7.75 ABC 12-4-71 9.50 BC12-2-18 7.50 ABC 12-2-6 8.50 BCI-FRIPAPA 7.25 ABC 12-4-170 8.00 BCI-ESTELA 7.25 ABC 98-38-12 8.00 BC12-2-6 6.25 BC I-ESTELA 8.00 BC98-2-6 6.25 BC 12-3-3 7.00 BC12-3-3 6.00 BC RUBI 6.50 BCI-YANASHUNGO
6.00 BC I-FRIPAPA 5.50 BC
12-4-46 4.50 BC 07-32-15 5.50 BCI-PUCASHUNGO
4.75 BC I-PUCASHUNGO
5.50 BC
12-6-1 3.45 BC 12-6-1 5.00 BC12-4-170 3.00 C I-YANA
SHUNGO4.50 C

61
3.12. Rendimiento total en gramos
En la Tabla 3.8, se muestra el análisis de varianza para los tratamientos inoculados y
testigos. El coeficiente de variación fue de 54,88% para los tratamientos inoculados y
de 36,93% para los tratamientos testigo. Los promedios de rendimiento en gramos por
maceta fueron de 69,55 g en los tratamientos inoculados y de 95,55 g en los tratamientos
testigo. Existe una relación con significación estadística entre la variable rendimiento
respecto al estado de la planta (inoculada y no inoculada).
Tabla 3.8 Análisis de varianza para la variable gramos por maceta de tratamientos inoculados ytestigo del ensayo de evaluación de la resistencia ante Rhizoctonia solani Kühn.
TRATAMIENTOS INOCULADOSF.V. GL CM FcTOTAL 79TRATAMIENTOS 19 57.20 *REPETICIONES 3 545.16ERROR 57 1404.16X (%) 69.55CV (%) 54.88
En la prueba de Tukey para la variable de producción (g/maceta) se encontraron dos
rangos de significancia distinguidos (Tabla 3.9). El primer rango (A), presentó 19
variedades, el clon promisorio 98-38-12 obtuvo el mayor rendimiento promedio con
120,18 gramos por maceta. La variedad mejorada I-Puca tuvo el menor rendimiento con
13,51 gramos por maceta. El segundo rango (AB), abarcó 19 variedades, el rendimiento
más alto se lo tuvo la variedad mejorada I-Fripapa con 118,58 gramos por maceta. El
menor rendimiento presentó el clon 12-6-1 con 11,80 gramos por maceta (Figura 3.12).

62
Los tratamientos testigo se distribuyeron en cuatro rangos de significancia. El
primer rango (A) presentó 15 variedades en el clon promisorio 98-2-6 obtuvo un
rendimiento de 192 gramos por maceta. En el segundo (AB) se agruparon 17
tratamientos. El clon promisorio 98-38-12 y la variedad mejorada I-Natividad tuvieron
los mejores rendimientos, con 163,75 y 141,95 gramos por maceta respectivamente.
El tercer rango (BCD) abarcó 13 tratamientos. La variedad mejorada I-Rubí obtuvo
el mayor rendimiento con un promedio de 140 gramos, el más bajo rendimiento lo
obtuvo el clon promisorio12-3-3 con 60.50 gramos. El último rango (D) presentó cuatro
materiales germoplásmicos, la de menor rendimiento fue el clon promisorio 12-6-1 con
un promedio de 10.25 gramos por maceta seguido de los materiales vegetales: I-Puca
Shungo con 15.50 g, el clon 12-2-6 con 22.50 g y el clon 12-4-170 con 30.50 g por
maceta (Tabla 3.9).
Los materiales inoculados tuvieron un rendimiento menor en comparación a los no
inoculados. Existió una reducción de rendimiento de casi un 30% en los tratamientos
inoculados.
Tabla 3.9 Prueba de Tukey para la variable “rendimiento por maceta” del ensayo de evaluación de la
resistencia ante Rhizoctonia solani Kühn.
TRATAMIENTO INOCULADO TRATAMIENTO TESTIGOVARIEDAD PROMEDIO RANGO VARIEDAD PROMEDIO RANGO98-38-12 120.18 A 98-2-6 192.00 AI-FRIPAPA 118.58 A 98-38-12 163.75 BI-NATIVIDAD 115.70 AB I-NATIVIDAD 141.95 BCRUBI 112.95 ABC I-RUBI 140.00 BCD98-2-6 111.63 ABC I-VICTORIA 138.25 BCDI-ESTELA 96.85 ABCD I-ESTELA 128.75 BCD12-6-24 85.13 ABCD I-FRIPAPA 120.20 BCD
CONTINÚA

63
I-YANASHUNGO
78.50 ABCD I-YANASHUNGO
119.85 BCD
12-4-71 76.75 ABCD 07-32-15 111.25 BCD07-32-15 74.75 ABCD 12-2-18 106.25 BCDI-VICTORIA 68.63 ABCD I-SUPERCHOLA 90.50 BCD97-25-3 65.38 ABCD 12-6-24 86.50 BCD12-2-18 63.13 ABCD 12-4-46 85.25 BCDI-SUPERCHOLA
51.33 ABCD 12-4-71 80.00 BCD
12-2-6 45.63 ABCD 97-25-3 67.25 BCD12-3-3 27.63 ABCD 12-3-3 60.50 BCD12-4-170 27.50 BCD 12-4-170 30.50 CD12-4-170 22.93 AB 12-2-6 22.50 CDI-PUCASHUNGO
13.51 AB I-PUCASHUNGO
15.50 CD
12-6-1 11.80 B 12-6-1 10.25 D
Figura 3.12. Diagrama de bloques de la variable rendimiento en variedades inoculadas y no inoculadas(Galárraga, 2014).

64
CAPÍTULO 4 – DISCUSIÓN
Prado et al. (2001) manifiesta que la utilización de variedades resistentes a
fitopatógenos se considera primordial en el manejo integrado de costra negra en el
campo, debido a que representa menor costo de producción, además de que su
utilización genera bajo impacto ambiental por la disminución de fungicidas para el
control de Rhizoctonia solani Kühn.
El cancro de la papa o costra negra ocasiona lesiones, principalmente a nivel de
tallos, raíces y tubérculos. Aucancela et al. (2012) corrobora que R. solani afecta la
apariencia del tubérculo y consecuentemente causa la pérdida de valor comercial. Torres
(1997) reporta pérdidas de hasta el 20% de cultivos por R. solani. Estos son algunos de
los argumentos por los que se ha buscado en esta investigación, materiales
germoplásmicos que presenten mayor resistencia al hongo causante de la costra negra.
Según INIAP-DNPV (2009) el uso de semilla sana se considera el componente más
importante para reducir las enfermedades causadas por fitopatógenos de suelo y afirma
que es posible reducir su incidencia hasta un 20%. INIAP-DNPV (2010) explica que
mediante el uso de semilla sana sumado a la rotación de cultivos se pueden disminuir las
infecciones hasta un 28%. La utilización de una semilla de calidad y con tolerancia a
costra negra, reducirá en gran porcentaje las pérdidas económicas del sector papero del
país.
En este ensayo se evaluaron frente a costra negra veinte germoplasmas de papa de
diferentes procedencias genéticas. Dentro del grupo de germoplasmas a evaluarse se
incluían variedades mejoradas que han sido el resultado de cruces con líneas nativas que

65
han permanecido hasta la actualidad (I-Puca Shungo e I-Yana Shungo). Siempre los
agricultores han sido un factor importante en la domesticación, la selección y
mejoramiento de las variedades de cultivos importantes para el hombre, un buen ejemplo
es la papa (Estrada, 2000).
4.1. Aislamiento e identificación de Rhizoctonia solani Kühn
El cultivo utilizado en este ensayo fue aislado a partir de tubérculos que presentaban
esclerocios del fitopatógeno. Según Chavarro (2011) los cultivos de R. solani jóvenes
en medio PDA se caracterizan por tener el micelio de un color blanco, y que a medida
que envejece se torna de un color café claro y empieza a formar esclerocios. El cultivo
aislado siempre contó con esas características. Es importante durante el aislamiento
utilizar antibióticos que controlen el crecimiento de bacterias en el medio, en nuestro
caso se utilizó cloranfenicol. Agrios (2002) recomienda el uso de cloranfenicol al
0,005% en medio PDA para aislar fitopatógenos de suelo.
Giovannini et al. (2005) afirma que Rhizoctonia solani Kühn visto al microscopio
presenta ramificaciones en ángulo recto, constricción de ramificaciones y la formación
de un septo cerca del punto de origen. De acuerdo con estos criterios la cepa aislada
concordó con las características y se la identificó como R. solani Kühn. De acuerdo a la
capacidad de fusionar hifas, R. solani se clasifica en grupos de anastomosis, los grupos
más recurrentes según Giovanini et al. (2005) son los: AG3, AG4, AG5 y AG8, siendo
el grupo de anastomosis 3 (AG3) el más patogénico. Este grupo se caracteriza por
formar estructuras de resistencia fusionando sus hifas con masas de tierra o materia

66
orgánica. En medio de cultivo PDA de la cepa aislada se pudo constatar la formación de
esclerocios a partir de la segunda semana de la siembra (Cotes, 2007).
Sumado a la confirmación morfológica de las colonias se realizó una PCR de
confirmación de grupo de anastomosis, según Lees et al. (2002) la región genética que
confirma el grupo de anastomosis 3 (AG3) se encuentra alrededor de los 500 pb. En el
ensayo se obtuvieron bandas de alrededor de 500 pb que confirmó que el fitopatógeno
aislado se trató de R. solani AG3.
Al realizar la amplificación de las regiones ITS1-5,8S-ITS2 las temperaturas de
ciclos propuestas por White et al. (1990) de un ciclo a 95°C por 3 min, seguido de 35
ciclos a 55°C durante 30 s, 72°C por 2 min, 95°C por 30 s y un ciclo final de extensión
por 10 min a 72°C no amplificaron de buena manera, por lo que se procedió a realizar
pruebas de variación de temperaturas, y se tuvo que la mayor amplificación para la
región ITS1-5,8S-ITS2 se obtiene con un ciclo inicial de desnaturalización a 95°C por 2
min; seguido de 35 ciclos de 95°C por 45 s, 63°C por 60 s y 72°C por 90 s; con un ciclo
final de extensión de 72°C por 5 min, temperaturas que coinciden con la amplificación
de la región AG3 propuesta por Fernández (2011).
4.2. Preparación del inóculo de Rhizoctonia solani Kühn
Carling et al. (2002) menciona que el grupo AG3 de Rhizoctonia es uno de los
grupos de anastomosis más dañinos para la planta, debido a la formación acelerada de
esclerocios y la colonización agresiva de tallos y raíces, esto se corroboró durante la
etapa de multiplicación de los cultivos. Se observaron condiciones de crecimiento
aceleradas y con formación temprana de esclerocios. Como en el ensayo se utilizarían

67
germoplasmas experimentales y se tendrían que evaluar hasta su cosecha, se optó por
utilizar 20 g de inóculo en avena por cada 3 Kg de tierra y no 20 g por cada 1 Kg como
lo propone Farah et al. (2008). Con la modificación de la cantidad de inóculo por Kg de
tierra se obtuvieron buenos resultados, las plantas enfermaron y se pudo seleccionar los
materiales germoplásmicos con tolerancia a costra negra. Wharton et al. (2007) afirma
que el inóculo en semilla de avena es tan dañino como el proveniente de suelo en el
campo, si los órganos de la planta se desarrolla en proximidad al inóculo.
4.3. Síntomas de R. solani Kühn
Durante las primeras instancias del trabajo de invernadero se presentaron pocas
características aéreas de costra negra. Se pudo observar algunas hojas de la base que
presentaban clorosis en varios tratamientos. Tsor et al. (2001) describe algunas de las
características de la colonización de Rhizoctonia solani Kühn e incluye la clorosis foliar
como un síntoma visible en etapas tempranas de la colonización del tejido del
hospedero. I-Estela e I-Rubí fueron las variedades que mostraron una clorosis foliar
temprana durante los primeros 45 días a partir de la siembra, se observó también un
cambio en la coloración de la base del tallo hasta presentar una necrosis de color marrón
con severos daños a nivel de epidermis. En el resto de materiales los síntomas de
clorosis foliar fueron enmascarados por plagas secundarias que presentaron similar
sintomatología. La necrosis en tallo es característica en una planta de papa con la
enfermedad de costra negra. La zonas necrosadas tuvieron un color amarillento en sus
primeras instancias, después de colonizado todo el tejido del área afectada se volvió de
color marrón. En condiciones avanzadas de la enfermedad puede presentar superficies
blanquecinas con el micelio de Rhizoctonia solani Kühn (INIAP, 2005).

68
La sintomatología de costra negra, a nivel de campo, incluye síntomas como: la
formación de tubérculos aéreos, la presencia de hojas enrolladas hacia el envés en las
partes superiores, estrangulamiento del cuello de la planta, tubérculos deformes y
esclerocios en la superficie del tubérculo (Aucancela et al., 2012). En el ensayo a nivel
de invernadero no se manifestó toda la sintomatología descrita por Aucancela et
al.(2012). No se observó la formación de tubérculos aéreos, enrollamiento de hojas
característico de la Rizoctoniosis, ni una clara estrangulación de tallo, pero a la cosecha
se encontró que R. solani había colonizado tejidos radicales y de tallo en todos los
tratamientos evaluados como los describe el ANEXO III. La presencia o ausencia de
algunos de los síntomas de costra negra en los materiales evaluados dependen de la
tolerancia y del comportamiento del cultivar (Mesa, 2011).
Las raíces y estolones pueden ser infectados por R. solani en cualquier etapa
fenológica de la planta, pero la mayor incidencia ocurre en etapas tempranas del
desarrollo Wharton et. al. (2007) menciona que la tolerancia de la variedad incrementa
después de la emergencia del brote, limitando la colonización de los tejidos. La
severidad de la infección por R. solani depende principalmente de la tolerancia genética
de la variedad infectada.
A la cosecha se observó tallos y raíces necrosadas en los 20 tratamientos con
inóculo y solo en la variedad I-Rubi fue evidente la presencia de manchas algodonosas
sobre la base del tallo, evidencia de la fase sexual de R. solani, Tsor (2001) menciona
que las basidiosporas y el micelio formado en esta fase sexual no causa daño a la planta.

69
4.4. Control de altura de la planta
Durante el ensayo se realizaron controles periódicos de altura a partir de los tres
meses de la siembra. Los datos obtenidos fueron sometidos a un análisis estadístico en
INFOSTAT en el cual se identificó que la variable altura no presentó una diferencia
significativa entre tratamientos inoculados y tratamientos testigo. Se observaron
variedades que en ambos tratamientos fueron las más pequeñas, este es el caso de I-Rubi
con una altura promedio de 20,25 cm y 21 cm en los tratamientos inoculados y testigo
correspondientemente. INIAP (2005) corrobora que la variedad I-Rubi se caracteriza por
ser una variedad semirastrera y de baja altura. A pesar del fenotipo de la variedad, las
plagas secundarias pudieron afectar el crecimiento de las plantas ya que los áfidos y
Oidium afectan directamente a las hojas, por ende, a la fotosíntesis; afectando el
metabolismo. I-Puca Shungo se destacó por su gran altura en todos los bloques. Se
tuvieron valores promedio de 55,50 cm y 65,50 cm en los tratamientos inoculados y
testigos correspondientemente. Andrade et al. (1995) manifiesta que el factor altura
puede deberse a aspectos genéticos de los materiales y no a la presencia de costra negra.
4.5. Evaluación de los tubérculos
I-Estela, I-Yana Shungo, I-Rubí y el clon promisorio 12-2-6, presentaron tubérculos
en forma de muñecos. Giovanini (2005) describe que esta deformidad es característica
de un estado avanzado de la enfermedad de costra negra y que estos se producen por
inhibición del crecimiento de las aéreas de tejido colonizado por costra negra. En las
variedades: I-Fripapa, I-Superchola, I-Yana Shungo, I-Rubí, y en los clones promisorios
98-2-6 y 98-38-12 se encontró síntomas de sarna causados por la bacteria Streptomyces
scabies. Pérez & Forbes (2011) afirma que la sarna que afecta a la papa se transmite en

70
el tubérculo semilla y que es asociada a la enfermedad de costra negra, en este ensayo se
pudo observar que nueve de las veinte variedades presentaron síntomas de sarna a la
cosecha (ANEXO III).
4.6. Severidad de la Infección
En ocho de los materiales evaluados se observaron altos porcentajes de severidad.
Existen materiales germoplásmicos que se encuentran en una escala de 3 según el
criterio de clasificación de Clive (1971) con un porcentaje de infección de hasta 35%, el
análisis estadístico de este parámetro junto con los criterios de clasificación utilizados en
el ensayo (Tabla 2.2) hacen es factible la clasificación de materiales por la tolerancia
presentada.
Mediante los análisis de varianza y las prueba de Tukey realizadas se encontró que
existió alta significancia estadística que relacionan la variedad inoculada y la severidad.
Mediante los parámetros de severidad ante costra negra dados por Clive (1971) se logró
separar las variedades que presentaron tolerancia de las susceptibles, esto basándose en
una clasificación de los tubérculos dependiendo del porcentaje de infección y la reacción
presentada, se agrupó los materiales germoplásmicos en cuatro categorías: altamente
tolerantes (AT), tolerantes (T), parcialmente tolerantes (PT) y susceptibles (S). Los
materiales tolerantes encontrados en esta investigación deben ser probados en campo
para continuar con la selección de un tubérculo semilla con resistencia a costra negra.
Los datos analizados en INFOSTAT y la sintomatología observada en invernadero
declaran a la variedad I- Puca Shungo como altamente tolerante. La variedad I-Rubí se
la encontró susceptible ante Rhizoctonia solani Kühn, el análisis realizado junto a las

71
características de sintomatología aérea descritas por Tsor et al. (2001) se confirma la
relación entre sintomatología aérea y severidad de la colonización del tubérculo.
4.7. Rendimiento
Después de realizar el análisis estadístico de la variable rendimiento por maceta se
encontró que el coeficiente de variación (C.V.) fue de 50.08% y una media de
producción de 7.91 tubérculos por tratamiento. Se observó que para los tratamientos
testigo, el coeficiente de variación dió un valor de 38.60% y una media de producción de
9.90. El análisis de Tukey reporta una diferencia estadística significativa en la variable
número de tubérculos con respecto al inóculo. En el ensayo se encontraron reducciones
de rendimiento de hasta un 28%. INIAP (2005) reporta perdidas de rendimiento de hasta
un 30% en cultivares del país, indicando que R. solani AG3 es causante de grandes
pérdidas económicas del sector papero, además, Jager et al. (1996) corrobora que las
enfermedades del suelo afectan a la pérdida de valor comercial de los tubérculos como
se observa en los datos de severidad obtenidos en este estudio.
4.8. Plagas y control del ensayo
Se observó en primeras instancias la muerte de una plántula del segundo bloque de
la variedad Superchola y se encontró que el tubérculo estaba en putrefacción por
Erwinia carotovora spp. carotovora.
Existieron plagas no esperadas que afectaron a la sintomatología aérea. Una de estas
fue el Oidium, que ingresó al invernadero en plantas contaminadas provenientes de otro
ensayo. En cuestión de 30 días, a pesar de haber eliminado el foco de contaminación, ya

72
había contaminado algunas plantas del tercer bloque del invernadero. En cuestión de
cinco meses todos los bloques se encontraban con sintomatología de oídium. Castillo
(2008) describe la sintomatología como un micelio enteramente superficial que atacan
órganos jóvenes que se manifiesta en forma de manchas blanquecinas pulvurulentas, las
manchas causan lesión en las hojas semejantes a los de una quemazón, conforme ha
avanzado la enfermedad, produce el secamiento y marchitez de la planta. Se realizaron
tres fumigaciones con NIMROD, un fungicida sistémico preventivo y curativo para el
control de Oidium.
Un factor crítico durante la aparición de síntomas aéreos fue los síntomas de virosis
en las hojas de la mayoría de variedades. Esto posiblemente fue transmitido por vectores
que se encontraban en el tubérculo antes de la siembra. INIAP-DNVP (2009) aconseja el
uso de semilla sana para evitar enfermedades durante el crecimiento de la planta de
papa. En los bloques del invernadero se hizo evidente una gran presencia de áfidos que
se extendió a partir del segundo mes desde la siembra. Según Hooker (1980) los áfidos
son vectores para la transmisión de virus y describe que la sintomatología se presenta
fundamentalmente en las hojas jóvenes de la parte apical, y que toman una coloración
pálida, amarillenta; pudiendo llegar a enrollarse hacia arriba. Las características de
infección de virus estuvieron presentes en el ensayo, enmascarando la sintomatología
aérea de costra negra, debido al similar marchitamiento y enrollamiento de hojas. A
pesar de estos factores imprevistos, se realizaron tres fertilizaciones foliares para ver si
los síntomas desaparecían o cuales persistían, las plantas mejoraron el aspecto pero
volvieron a decaer enseguida.
French (1982) aconseja que para una buena investigación fitopatológica se necesita
tener conocimiento del origen del sujeto de estudio (en este caso los materiales

73
germoplásmicos), también menciona que si se va a trabajar con un fitopatógeno en
especial se debe esterilizar la tierra del experimento, y tener cuidado en la preparación
del inóculo, enfatiza que el control de enfermedades y plagas comunes se puede llevar a
cabo durante todo el ciclo experimental. Todas esas medidas fueron tomadas a lo largo
del ensayo en el invernadero. Se cree también que la sintomatología aérea como
tubérculos aéreos y el estrangulamiento del cuello de la planta, no fueron observados
con claridad debido a las condiciones de estrés de las plantas, Castellanos (2007) indica
que el comportamiento de Rhizoctonia solani Kühn es diferente en campo, laboratorio e
invernadero; esto debido a que en campo puede existir mayor humedad y una
temperatura favorable para el desarrollo del micelio. Las plantas en el invernadero
estuvieron libres de estrés climático; las variaciones de temperatura y de humedad de los
materiales evaluados en el ensayo estuvieron controlados.
La presencia de plagas secundarias en el ensayo no intervino con la selección de los
materiales germoplásmicos. Castillo (2008) afirma que Oidium es una enfermedad que
afecta solo a partes aéreas de la planta, no existe influencia sobre la tuberización. A
pesar de que la sintomatología aérea fue enmascarada, los bloques del invernadero se
mantuvieron hasta la cosecha con la ayuda de fungicidas foliares.
Hooker (1980) menciona que la virosis puede afectar a los rendimientos solo cuando
es una infección secundaria, en el caso del ensayo los áfidos fueron los vectores de la
enfermedad y esta ocurrió durante el desarrollo de las plantas, por ende se trató de una
infección primaria y no influenció en la producción de los bloques. La clasificación de
los materiales germoplásmicos se basó en la severidad de la infección como lo describe
Clive en sus criterios de selección.

74
CAPÍTULO 5 – CONCLUSIONES
Rhizoctonia solani Kühn AG3 fue aislada a partir de esclerocios de papa infectada
con costra negra.
Se encontró que la variedad I-Puca shungo presenta tolerancia a Rhizoctonia solani
Kühn con un porcentaje de infección del 0%. Las variedades 12-6-1 y 12-4-170
presentan una tolerancia moderada a costra negra con porcentajes de infección de
5.62% y 6.93% respectivamente.
La variable altura no mostró diferencias en el comportamiento de tratamientos
inoculados respecto al grupo control.
Los materiales inoculados con Rhizoctonia solani Kühn presentaron una
disminución en el rendimiento de hasta un 28% en comparación con sus tratamientos
testigo.

75
CAPÍTULO 6 – RECOMENDACIONES
Se recomienda realizar pruebas de campo para poner a prueba la resistencia de las
variedades seleccionadas. El comportamiento de Rhizoctonia solani Kühn en las
variedades evaluadas puede ser diferente en condiciones de campo. Las condiciones
ambientales en este ensayo fueron controladas y la inoculación fue homogénea.
Existen muchas variedades nativas que pueden presentar resistencia a costra negra.
Deberían ser puestas a prueba como se lo ha hecho con variedades de INIAP en este
ensayo. Variedades nativas han permanecido hasta hoy en día por presentar resistencia
fitopatógenos de suelo y a factores de estrés climático. El estudio de variedades con
resistencia genética a fitopatógenos es de gran importancia. Se pueden disminuir
impactos ambientales del uso de fungicidas mediante su uso.
Se debería ampliar el estudio molecular de la interacción patógeno-huésped para R.
solani Kühn, así se lograría conocer los factores moleculares y genes que le dan
resistencia a especies nativas de papas. Los genes de resistencia a costra negra podrían
ser utilizados en cultivos asociados a Rhizoctonia solani para el control de la
enfermedad.

76
BIBLIOGRAFÍA
Agrios, H. 2002. Fitopatología. Editorial Limusa, Grupo Noriega Editores,
Segunda edición. México, D.F. 838p.
Andrade, H. et al 2002. El cultivo de la papa en el Ecuador. Pumisacho y
Sherwood, Editores. INIAP, CIP, Quito-Ecuador. pp 21-28.
Aucancela,R, et al. 2012. Conozca y maneje las enfermedades del suelo en el
cultivo de papa. Boletín divulgativo No 142. INIAP. Quito-Ecuador.
Borba, N. 2008, La papa un alimento básico. Posibles impactos frente a la
introducción de la papa transgénica. RAP-AL. Uruguay.
Borja, C. et. al, 2010. Manual de cultivo de papa. EDIFARM & CÍA. Quito-
Ecuador.
Carling, D. et al. 2002. Hyphal anastomosis reactions, rDNA-Internal
Transcribed Spacer Sequences, and virulence levels among subsets of
Rhizoctonia solani Anastomosis Group-2 (AG-2) and AG-BI. Phytopathology.
92(1):43-50.
Castellanos, G. et al. 2007. Rhizoctonia solani. Manejo del hongo en laboratorio.
Centro Internacional de Agricultura Tropical (CIAT). Departamento de
fitopatología del frijol. Cali-Colombia.
CESA. 1996. Las papas, Cultivo de ciclo corto. 2da ed. Quito, Ecuador. Edit.
CESA.
CORPOICA (Corporación Colombiana de Investigación Agropecuaria). 2000.
Manejo Integrado del cultivo de papa. Colombia.

77
Cotes. 2007. Biología y manejo de Rhizoctonia solani. Laboratorio de Control
Biológico Centro de Biotecnología y Bioindustria (CBB). Bogotá-Colombia.
Devaux, A. et al. 2010. El sector papa en la región andina: Diagnóstico y
elementos para una visión estratégica (Bolívar, Ecuador y Perú). Centro
Internacional de la Papa. Lima, Perú. 385 p.
Dimitri, M. 1972. Enciclopedia de agricultura y jardinería. 2da. ed. Buenos,
Aires, Argentina, Acne S. A. V. 1 p. 197- 198, 273-278.
Dimitri, M. 1972. Enciclopedia de agricultura y jardinería. 2da. ed. Buenos,
Aires, Argentina, Acne S. A. V. 1 p. 197- 198, 273-278.
Escobar, W. 1997. Caracterización morfológica, agronómica y bioquímica de la
colección ecuatoriana de papa subgrupo tardías. Ingeniero Agrónomo, Facultad
de Ciencias Agropecuarias, Escuela de Ingeniería Agronómica, Universidad
Estatal de Bolívar, Guaranda, Ecuador.
Estrada, N. 2000. La biodiversidad en el mejoramiento genético de la papa.
Plural Editores. La Paz, Bolivia. 372 p.
Farah, N. et al. (2008). Influence of inoculum levels of Rhizoctonia solani and
susceptibility on new potato germplasm. Pak. J. Bot., 2199-2209.
French, E. & T. Hebert. 1982. Métodos de investigación fitopatológica. Instituto
interamericano de cooperación para la agricultura. San José-Costa Rica.
Giovannini, C. et al. 2005. Caracterización morfológica y patogénica de
Rhizoctonia solani Kühn a partir de tubérculos de papa (solanum tuberosum L)
provenientes de diferentes predios de la x región de Chile. IDESIA 23(1): pp. 13-
18.

78
Hidalgo, E. et al. 1997. PCR: Una nueva herramienta para el estudio de hongos
ectomicorrícicos. Revista Catalana Micol., vol. 20: 187-198. Barcelona-España
Hofman & Jongebloed. 1998. Infection process of Rhizoctonia solani on
Solanum tuberosum and effects of granular nematicidas. Neth. J. Pl. Path.
94:243-252.
Hooker, W. 1980. Compendio de enfermedades de la papa. Centro
Internacional de la papa, Lima, Perú 111p.
Huaman, Z. 1980. Botánica sistemática y morfológica de la papa. Boletín de
Información Técnica N° 6. Centro Internacional de la Papa, Lima, Perú. 20 p.
Hurtado, G. & M. Román, 2002. Guía técnica del cultivo de papa. Centro
nacional de tecnología agropecuaria y forestal. Arce-El Salvador.
INIAP (Instituto Nacional Autónomo de Investigaciones Agropecuarias).
Ecuador. 1984. Manual del cultivo de papa. Manual No. 5 p 1 – 2.
Instituto Nacional Autónomo de Investigaciones Agropecuarias. 2008. Informe
anual técnico. Estación experimental Sta. Catalina. Departamento de protección
vegetal. Quito, Ecuador. 26 pp.
Instituto Nacional Autónomo de Investigaciones Agropecuarias. 2009. Informe
anual técnico. Estación experimental Sta. Catalina. Departamento de protección
vegetal. Quito, Ecuador. 26 pp.
Instituto Nacional Autónomo de Investigaciones Agropecuarias. 2010. Informe
anual técnico. Estación experimental Sta. Catalina. Departamento de protección
vegetal. Quito, Ecuador.

79
INTA (Instituto Nicaragüense de Tecnología Agropecuaria). 2004. Guía Técnica
sobre Manejo Integrado de Plagas (MIP) en el cultivo de la papa (Solanum
tuberosum L.). Nicaragua.
Jager, G. & H. Velvi. 1988. Inactivation of sclerotia of Rhizoctonia solani on
potato tubers by Verticillium biguttatum, a soil-borne mycoparasite. Neth. J. Pl.
Path. 94(5):225-231.
Kerh, A. et-al. 1967. Producción Comercial de la papa.. México, México.Edit.
Centro Regional de Ayuda Técnica para el desarrollo Internacional (I. A. D.) P
22-24.
Lees, D. et al. 2002. Development of conventional and quantitative real-time
PCR assays for the detection and identification of Rhizoctonia solani AG-3 in
potato and soil. Plant Pathology (2002) 51, 293–302.
Marshall, D. 1980. Infection cushion on rice sheaths by Rhiuzoctonia solani.
The American Phytopathological Society, 1980, 70(10): 947-950.
Mesa, E. 2011. Variabilidad genética y detección molecular de poblaciones del
hongo Rhizoctonia solani en regiones colombianas productoras de papa.
Universidad Nacional de Colombia. Bogotá-Colombia.
Ortiz, R. & S.J. Peloquin, 1994. Effect of sporophytic heterozygosity on the male
gametophyte of the tetraploid potato (Solanum tuberosum). Ann. Bot. 73: 61-64.
Paca, J. 2009. Respuesta del cultivo de la papa (Solanum tuberosum L.) variedad
chaucha a la aplicación de cuatro tipos de abonos en tres dosis. Proyecto de tesis.
ESPOCH. Riobamba-Ecuador.

80
Paulitz, T. & K. Schroeder. 2005. A new method for the quantification of
Rhizoctonia solani and R. oryzae from soil. Plant Disease, 767-772.
Pérez, W. & G. Forbes, 2011. Guía de identificación de plagas que afectan a la
papa en la zona andina. Centro Internacional de la papa (CIP). Lima-Perú.
Pérez, W. & G. Forbes. 2011. Manual de observaciones fenológicas. Servicio
Nacional de Meteorología e Hidrología-SENAMHI. Lima-Perú.
Piñeros, C. 2009. Recopilación de la investigación del Sistema Productivo Papa
Criolla FEDERACIÓN COLOMBIANA DE PRODUCTORES DE PAPA Y
GOBERNACIÓN DE CUNDINAMARCA. Bogotá-Colombia. 154 p.
Prado, G. et al. 2001. Caracterización preliminar de la resistencia de
germoplasma de arroz al añublo de la vaina (Rhizoctonia solani Kuhn). Foro
arrocero latinoamericano 7(1):8-11.
Reinoso, I. 2011. El cultivo de papa y su participación en la economía
ecuatoriana. INIAP-Estación experimental Santa catalina. Quito-Ecuador.
Taheri, P. 2010. Molecular and cytological aspects of tomato-Rhizoctonia solani
interaction. ISHS Acta Horticulturae 914.
Torres, H. 1997. Manual de las enfermedades más importantes de la papa en
el Perú. CIP-SENASA-PAPA ANDINA, Lima-Perú 19-22.
Tsor & Barak. 2001. Biological control of Black scurf on potato under organic
management. Crop Protection. 20, p. 145-150.
Whitney, H. 1964. Physiological and cytological studies of basidiospores
repetition in Rhizoctonia solani Kühn. Canadian Journal of Botany, 1964,
42(10): 1397-1404.

81
ANEXO I
Grupos de anastomosis para Rhizoctonia solani Kühn (Chavarro, 2011)
AG1-1 A (= AG tipo1)
Crece rápidamente, 30 mm/día a 28-30ºC. Su óptimo es a 30ºC
AG 1 IC (AG1 TIPO 3)
Patógeno virulento que se cría en climas fríos, causa camping off a zanahoria, alforfón,
lino soja y pino
AG 1 inespecífico
Patógeno de remolacha azucarera, col, repollo, judías y soja.
AG2-1
Este grupo tiene crecimiento lento (13mm/día a 25ºC). Son patógenos de cultivos de
invierno formando esclerocios rojos en forma de anillo. Causa camping off en
crucíferas, fresa, tabaco japonés, tulipán y en trébol subterráneo
AG2-2IIIB
Es un grupo que crece a altas temperaturas (óptimo aproximadamente 30ºC). Son
patogénicos de arroz, jengibre, gladiolos, césped, trigo, remolacha azucarera, batata y
provoca camping off en plántulas de árboles, crisantemos y batata china.
AG2-2IV
Patógeno de remolacha azucarera, césped

82
AG2-2-2 (inespecífico)
AG3
Crece unos 12mm/día a 25ºC. Patógeno de patata, tomate y tabaco.
AG-4
Se define como tipo “pratícola”. Según algunos autores representa a algunas especies (R.
pratícola.).
Se suele aislar del suelo con plantas infectadas en países de clima húmedo. Ataca a
tomate, guisante, espinaca, patata. Camping off en pinos y otros hospedantes en estado
de plántula, cebolla, guisante, patata y judía.
Hay aislados no patógenos que protegen a rabanitos, zanahoria, algodón, patata y trigo
de camping off, estimulando el crecimiento de la planta.
AG-5
Patógenos débiles de patata, césped, judía, soja y azuki. Algunos se micorrizan con
orquídeas
AG-6
No son patógenos, algunos micorrizan a orquídeas.
AG-7
No son patógenos.
AG-8
Patógeno de cereales.

83
AG-9
Patógeno de crucíferas, patata pero menos patogénicos que AG-3.
AG-10
Probablemente no son patógenos
AGBI
No son patógenos.
AGUNK
Patógeno de de arroz, no se fusionan con ningún grupo.

84
ANEXO II
Resultados de la cuantificación de ADN de Rhizoctonia solani Kühn

85
ANEXO III
Síntomas observados en veinte materiales de papa evaluados en invernadero
(Galárraga, 2014)
N° Variedad Bloque Sintomatología de Tubérculos
1 I-ESTELA 1 Tubérculos en forma de muñecos
2 Raíces necrosadas
3 Necrosis café oscura en raízprincipal
4 Necrosis raíz-Tubérculos en formade muñecos
2 I-FRIPAPA 1 Necrosis de raíz
2 Raíces necrosadas – síntomas desarna en tubérculos
3 Necrosis café oscura en raízprincipal – síntomas sarna
4 Necrosis raíz – síntomas de sarna
3 I- PUCA SHUNGO 1 Necrosis de raíz
2 Necrosis de raíz
3 Necrosis de raíz
4 Necrosis raíz principal
4 I-NATIVIDAD 1 Necrosis de raíces
2 Necrosis raíz principal
3 Necrosis raíz principal
4 Necrosis de raíces
5 I-SUPERCHOLA 1 No presentó necrosis raíz
2 Planta no brotó
3 Necrosis raíz – síntomas de sarna
4 Síntomas de sarna
6 I-VICTORIA 1 Necrosis en raíz
2 Necrosis en raíz - tubérculos enforma de muñecos
3 Necrosis en raíz - tubérculos enforma de muñecos
4 Necrosis en raíz - tubérculos enforma de muñecos
7 I-YANA SHUNGO 1 Necrosis en raíz
2 Necrosis en raíz
3 Tubérculos en forma de muñecos

86
4 Tubérculos forma de muñecos –Síntomas de sarna
8 RUBI 1 Necrosis en raíz – Síntomas desarna
2 Tubérculos en forma de muñecos
3 Síntomas de sarna
4 Necrosis de raíz
9 07-32-15 1 Necrosis en tallo y raíces
2 Necrosis en tallo y raíces
3 Necrosis en tallo y raíces
4 Necrosis en tallo y raíces
10 12-2-6 1 Tubérculos pequeños y en forma demuñecos
2 Tubérculos pequeños y en forma demuñecos
3 Tubérculos pequeños y en forma demuñecos
4 Necrosis de raíz
11 12-2-18 1 Necrosis en raíz
2 Necrosis en raíz
3 Necrosis en raíz
4 Necrosis en raíz
12 12-3-3 1 Raíz necrosada – Tubérculos enforma de muñecos y podridos
2 Raíz necrosada – Tubérculos enforma de muñecos y podridos
3 Síntomas de sarna
4 Raíz necrosada
13 12-4-71 1 Necrosis en raíz
2 Necrosis en raíz
3 Necrosis en raíz
4 Necrosis en raíz
14 12-4-46 1 Raíz y tallo necrosado
2 Raíz y tallo necrosado
3 Raíz y tallo necrosado
4 Necrosis en tallo – sin tubérculos
15 12-4-170 1 Necrosis en raíz
2 Raíz y tallo necrosado
3 Necrosis raíz – no hay producción
4 Necrosis de raíz
16 12-6-1 1 Necrosis en raíz
2 Necrosis de raíz
3 Necrosis de raíz
4 Necrosis de raíz

87
17 12-6-24 1 Necrosis raíz coloración caféoscuro
2 Necrosis raíz coloración caféoscuro
3 Necrosis raíz coloración caféoscuro
4 Necrosis raíz coloración caféoscuro
18 97-25-3 1 Necrosis raíz coloración caféoscuro
2 Necrosis raíz coloración caféoscuro
3 Necrosis raíz coloración caféoscuro
4 Necrosis raíz coloración caféoscuro – síntomas de sarna
19 98-2-6 1 Necrosis raíz
2 Necrosis raíz – síntomas de sarna
3 Necrosis raíz – síntomas de sarna
4 Necrosis raíz
20 98-38-12 1 Necrosis raíz – Síntomas de sarna
2 Necrosis raíz – Síntomas de sarna
3 Necrosis raíz – Síntomas de sarna
4 Necrosis raíz














