
Estudio de la función de péptidos tipo snakin
11
Estudio de la función de péptidos tipo snakin Jiménez-Manzano Camila 1 , López-Martínez Alma Lucía 2 , Philippe Monnier Aguilar 3 , Marithé Carrera Lobato 4 & Torres-Huerta Ana Laura 5 . Instituto Tecnológico y de Estudios Superiores de Monterrey, Departamento de Bioingeniería 1 [email protected], 2 [email protected], 3 [email protected], 4 [email protected], 5 [email protected] Palabras clave: péptidos antimicrobianos, snakin/GASA, Allium sativum, Mangifera indica, Eucalyptus globulus RESUMEN Los péptidos antimicrobianos representan una parte fundamental del sistema inmune de diversos organismos multicelulares, al presentar actividad de defensa frente a bacterias, hongos, levaduras, virus y parásitos. Debido a sus distintos mecanismos de acción antibacterial, se han posicionado como una alternativa prometedora a los antibióticos y fungicidas. Diversas especies de gran interés comercial y por sus propiedades medicinales se han seleccionado para identificar sus péptidos antimicrobianos. Las especies de estudio corresponden al ajo (Allium sativum), mango (Mangifera indica) y el eucalipto (Eucalyptus globulus), ya que en diversas publicaciones se ha demostrado su efecto antimicrobiano, aunque no se han analizado los metabolitos y péptidos involucrados en este efecto. Se realizó la identificación in silico de péptidos con homología alta a snakins previamente reportadas en otras especies como referencia, se realizó un alineamiento de la secuencia de aminoácidos de los péptidos de ajo, mango y eucalipto respecto a la Snakin-1 de Solanum tuberosum (como referencia, que se encuentra bien caracterizada). Se confirmó la presencia de las 12 cisteínas características de la familia Snakin/GASA y con la predicción de su estructura se observa que se mantienen conservadas las estructuras características en proteínas de esta familia. El ajo es un gran candidato para la obtención de Snakin-1 (SN-1), un péptido que posee propiedades antimicrobianas y antifúngicas, lo que significa que pueden ser de alto interés en el área de agricultura y farmacéutica. A partir de tejido de ajo se realizó la extracción de ARN total y se amplificó la secuencia codificante madura del péptido snakin de ajo (AsSN-1). Adicionalmente se realizó una amplificación mediante qRT-PCR de AsSN-1 para comparar el nivel de expresión de dicho péptido en los diferentes tejidos de ajo. Con estos resultados se propone la posterior expresión recombinante de este péptido para su caracterización y posterior uso como agente de control de enfermedades. Adicionalmente se tiene como perspectiva avanzar en la caracterización de los péptidos de mango y eucalipto. INTRODUCCIÓN Las proteínas de la familia Snakin/GASA están ampliamente distribuidas entre diferentes especies de plantas. Su expresión es tejido específico y solo es temporal, además su localización subcelular es variable. Todas las proteínas de esta familia mantienen conservados en su secuencia 12 cisteínas que son esenciales para su actividad bioquímica y para mantener una estructura definida por la formación de puentes disulfuro (Yeung H. y cols., 2016). Aún no se han caracterizado por completo las funciones de estas proteínas, aunque se les ha relacionado con la respuesta de las plantas frente a estrés biótico y abiótico, así como una posible participación en procesos hormonales y la homeostasis redox (Nahirñak, V. y cols., 2012). Adicionalmente se ha visto que tienen propiedades antimicrobianas, causa la rápida agregación de bacterias Gram positivas y negativas, aunque su actividad antimicrobiana no está correlacionada con su capacidad de agregación.
Transcript of Estudio de la función de péptidos tipo snakin

Estudio de la función de péptidos tipo snakin Jiménez-Manzano Camila1, López-Martínez Alma Lucía2, Philippe Monnier Aguilar3, Marithé Carrera Lobato4 & Torres-Huerta Ana Laura5.
Instituto Tecnológico y de Estudios Superiores de Monterrey, Departamento de Bioingeniería
[email protected], [email protected], [email protected], [email protected], [email protected]
Palabras clave: péptidos antimicrobianos, snakin/GASA, Allium sativum, Mangifera indica, Eucalyptus globulus
RESUMEN
Los péptidos antimicrobianos representan una parte fundamental del sistema inmune de diversos organismos multicelulares,
al presentar actividad de defensa frente a bacterias, hongos, levaduras, virus y parásitos. Debido a sus distintos mecanismos
de acción antibacterial, se han posicionado como una alternativa prometedora a los antibióticos y fungicidas. Diversas
especies de gran interés comercial y por sus propiedades medicinales se han seleccionado para identificar sus péptidos
antimicrobianos. Las especies de estudio corresponden al ajo (Allium sativum), mango (Mangifera indica) y el eucalipto
(Eucalyptus globulus), ya que en diversas publicaciones se ha demostrado su efecto antimicrobiano, aunque no se han
analizado los metabolitos y péptidos involucrados en este efecto. Se realizó la identificación in silico de péptidos con homología
alta a snakins previamente reportadas en otras especies como referencia, se realizó un alineamiento de la secuencia de
aminoácidos de los péptidos de ajo, mango y eucalipto respecto a la Snakin-1 de Solanum tuberosum (como referencia, que
se encuentra bien caracterizada). Se confirmó la presencia de las 12 cisteínas características de la familia Snakin/GASA y
con la predicción de su estructura se observa que se mantienen conservadas las estructuras características en proteínas de
esta familia. El ajo es un gran candidato para la obtención de Snakin-1 (SN-1), un péptido que posee propiedades
antimicrobianas y antifúngicas, lo que significa que pueden ser de alto interés en el área de agricultura y farmacéutica. A partir
de tejido de ajo se realizó la extracción de ARN total y se amplificó la secuencia codificante madura del péptido snakin de ajo
(AsSN-1). Adicionalmente se realizó una amplificación mediante qRT-PCR de AsSN-1 para comparar el nivel de expresión de
dicho péptido en los diferentes tejidos de ajo. Con estos resultados se propone la posterior expresión recombinante de este
péptido para su caracterización y posterior uso como agente de control de enfermedades. Adicionalmente se tiene como
perspectiva avanzar en la caracterización de los péptidos de mango y eucalipto.
INTRODUCCIÓN
Las proteínas de la familia Snakin/GASA están ampliamente distribuidas entre diferentes especies de plantas. Su expresión
es tejido específico y solo es temporal, además su localización subcelular es variable. Todas las proteínas de esta familia
mantienen conservados en su secuencia 12 cisteínas que son esenciales para su actividad bioquímica y para mantener una
estructura definida por la formación de puentes disulfuro (Yeung H. y cols., 2016). Aún no se han caracterizado por completo
las funciones de estas proteínas, aunque se les ha relacionado con la respuesta de las plantas frente a estrés biótico y abiótico,
así como una posible participación en procesos hormonales y la homeostasis redox (Nahirñak, V. y cols., 2012).
Adicionalmente se ha visto que tienen propiedades antimicrobianas, causa la rápida agregación de bacterias Gram positivas
y negativas, aunque su actividad antimicrobiana no está correlacionada con su capacidad de agregación.

Los genes de las proteínas de la familia Snakin/GASA codifican para
proteínas pequeñas que contienen 3 dominios, un péptido señal de 18-
29 aminoácidos, una región variable que es altamente divergente entre
especies y la región carboxilo terminal de aproximadamente 60
aminoácidos que contiene 12 residuos de cisteínas (dominio GASA) con
una posición conservada cada uno (ver figura 1). Ninguna de las
proteínas de esta familia contiene motifs de sitio activo.
Figura 1. Alineamiento y estructura de proteínas de la familia snakin/GASA. A) Alineamiento de proteínas donde se resaltan los residuos de cisteína y su interacción para la formación de puentes disulfuro. B) Modelo de estructura de la proteína SN-1 de Solanum tuberosum (papa). C) Esquema de la estructura de una proteína tipo snakin donde se muestra los puentes disulfuro y las alfa hélices. D) Alineamiento de diferentes clases de proteínas de la familia snakin. Todas las figuras fueron tomadas de Porto W. F. and Franco O. L., 2013; y de Yeung, H. y cols., 2016.
Diversos estudios (PlantPepDB, Manually curated Plant Peptide Database) han determinado la existencia de más de 3848
péptidos identificados en plantas (de 443 especies analizadas). Los principales tipos de péptidos antimicrobianos son:
defensinas, tioninas, LTPs (Non-specific lipid transfer proteins), Péptidos tipo Heveina, Knottins, α-Harpinas, Ciclotidos y
Snakins (Das D. y cols., 2020). En estudios de transcriptoma de 1267 plantas se identificaron 4849 secuencias pertenecientes
a la familia Snakin. Este tipo de péptidos resultan sumamente interesantes porque mantienen su conformación en condiciones
de pH bajo (vacuola por ejemplo) o ambientes proteolíticos (sistema de digestión de herbívoros). Algunos ejemplos de péptidos
de la familia Snakin/GASA identificados en algunas especies se observan en la siguiente tabla.
Tabla 1. Algunos péptidos pertenecientes a la familia Snakin/GASA que se han identificado en algunas plantas. Especie Nombre de las proteínas
jitomate (Solanum lycopersicum) GAST1, RSI-1
Petunia (Petunia hybrida) GIP1-5
Arabidopsis (Arabidopsis thaliana) GASA 1-15
Papa (Solanum tuberosum) SN1-2
Frijol (Phaseolus vulgaris) FBCBP
Arroz (Oryza sativa) OsGASR1-2 OsGSR1
Gerbera (Gerbera hybrida) GEG
Hayuco, fruto del haya (Fagus sylvatica) FsGASA4
Fresa (Fragaria ananassa) FaGAST
Maíz (Zea mays) ZmGSL1-10
Soya (Glicine soja) GsGASA1
Pimiento (Capsicum annuum) CaSn
Los péptidos tipo snakin causan ruptura celular mediante la ruptura de membranas, formación de poros, modificación de canales ióinicos y eventualmente la muerte celular, aunque aún no se han caracterizado estos mecanismos. Sin embargo, se cree que es mediante la disrupción o agrupamiento de lípidos aniónicos de membrana bacteriana. En hongos y levaduras interactúa con componentes de carga negativa como: Glucofosfolípidos, Esfingolípidos y Manoproteínas.

Figura 2. Efecto antimicrobiano de los péptidos tipo Snakin. A) Aglomeración observada de microorganismos en presencia de Snakin de papa (figura tomada de Berrocal-Lobo M. y cols., 2002), B) Posible mecanismo de descripción de membranas (figura modificada de Mookherjee, N. y cols-, 2020).
Expresar proteínas en plantas resulta costoso y requiere de mucho tiempo, por ello algunas de las proteínas de tipo snakin se han producido de forma recombinante en bacterias, levaduras y células de insecto, lo que ha resultado en poca eficiencia en la expresión y purificación, aunque las proteínas mantienen su funcionalidad (Kovalskaya N. and Hammond R. W., 2009), para ver detalles y comparar diferentes sistemas ver siguiente esquema. En el caso de bacterias, se ha optado por utilizar la fusión de tags como pelB y GST para poder disminuir el efecto tóxico contra la bacteria de expresión, sin embargo, el gran inconveniente es la remoción de los tags para poder obtener un péptido funcional (Kikukawa, T. y cols., 2017). Esto reduce el rendimiento. En levaduras se ha reportado un mayor rendimiento, así como en células de insecto aunque aún se tienen algunas desventajas con los costos y un proceso más complicado. En el caso de la síntesis química se tiene la desventaja de la poca actividad funcional porque se requiere un tratamiento especial para la formación de puentes disulfuro.
En los últimos años, la disponibilidad de información obtenida mediante herramientas de diversas omicas ha permitido tener datos de especies de plantas tanto modelo como de especies nativas. Éstas últimas se destacan por su importancia en la medicina popular y permanecen poco exploradas. Allium sativum, es una planta con muchas propiedades, como por ejemplo antibacterial, antiviral, antifúngico, anti-parasitante, con efecto farmacológico en sistema inmune, cardiovascular, entre otros (Mikaili P., y cols., 2013). En ajo se ha identificado una secuencia con alto parecido al péptido snakin de Solanum tuberosum mediante la herramienta de blast. Con el modelado in silico del péptido, se observó que conserva la estructura de la familia de los péptidos tipo snakin. Por otro lado, se ha reportado actividad antibacteriana y fungicida en Mangifera Indica, frente a cepas de Escherichia Coli, Klebsiella pneumoniae, Candida krusei y Candida albican (Mahalik, G., y cols., 2017). Mientras que, en el caso de eucalipto, esta actividad antimicrobiana se ha visto con el aceite esencial y se debe a la presencia de compuestos como a los componentes tales como 1,8-cineol, citronelal, citronelol, acetato de citronelilo, p-cimeno, eucamalol,
limoneno, linalol, β-pineno, γ-terpineno, α-terpinol, aloocimeno y aromadendreno (Nezhad, F. M., 2009). Sin embargo, no se

ha estudiado el potencial de péptidos de la familia Snakin, cuya caracterización resultaría de gran utilidad en el reto de la resistencia antimicrobiana, especialmente por el mecanismo de acción de estas proteínas a través del sistema inmune de organismos.
Justificación
Las proteínas tipo snakin de algunas especies han mostrado tener propiedades antimicrobianas y por ende tienen potencial aplicación en
el desarrollo de productos para aumentar la vida de anaquel de frutas y verduras, así como su uso para desinfección de superficies entre
otras aplicaciones. Debido a que solo algunas proteínas tipo snakin han sido caracterizadas y han demostrado eficiencia en la eliminación
de gran diversidad de bacterias y hongas, es importante estudiar las pertenecientes a mano, ajo y eucalipto, que son especies de
importancia comercial en nuestro país, así como su relevancia en la medicina tradicional.
Hipótesis
Si los péptidos identificados en mango, ajo y eucalipto contienen conservados los dominios característicos de la familia snakin entonces al
ser producidas de forma recombinante mostrarán actividad antimicrobiana y fungicida.
Objetivo general
Producir los péptidos de tipo snakin de ajo, mango y eucalipto de forma recombinante y mediante síntesis química (versión normal y
conteniendo aminoácidos no canónicos) para evaluar su actividad antimicrobiana y fungicida contra especies que representan problemas
graves en la agricultura mexicana.
Objetivos particulares
1. Estudio del perfil de expresión de los péptidos tipo snakin de ajo, mango y eucalipto
a. Aislamiento de ARN de las plantas
b. Diseño de primers para PCR en tiempo real (amplificación de los péptidos y gen control de carga)
c. PCR en tiempo real para verificar el perfil de expresión en diversos tejidos de las plantas
2. Identificación de las secuencias de los péptidos tipo snakin de ajo, mango y eucalipto
a. Identificar los genes y las secuencias codificantes de las proteínas tipo snakin.
b. Realizar alineamientos para identificar los dominios característicos de las proteínas tipo snakin.
c. Analizar las secuencias para determinar cuál es la región codificante que corresponda la proteína madura y al péptido señal.
3. Producción y purificación de los péptidos recombinantes tipo snakin de ajo, mango y eucalipto.
a. Diseño y optimización de la secuencia codificante para mandar a síntesis química de la secuencia
b. Clonar las secuencias codificantes en el vector de expresión
c. Transformar/transfectar el plásmido en el sistema de expresión.
d. Verificar la expresión de las proteínas tipo snakin mediante SDS-PAGE y western blot.
4. Obtención de péptidos mediante síntesis química (versión con aminoácidos normales y con aminoácidos no convencionales)
a. Selección de la plataforma de síntesis más conveniente para la obtención de los péptidos
b. Selección del aminoácido no convencional a sustituir y simulación de su plegamiento.
c. Síntesis de los péptidos
5. Estudiar la función de las proteínas tipo snakin
a. Analizar propiedades anti-microbianas
b. Estudiar propiedades fungicidas
c. Estudiar estabilidad

METODOLOGÍA (PRIMERA PARTE)
Identificación de secuencias con homología a proteínas de la familia snakin Mediante la herramienta de BLAST se usó la secuencia del péptido de papa (Uniprot B6E1W5) como referencia para buscar en todas las secuencias codificantes de los genomas de mango y eucalipto. Las secuencias con el porcentaje más alto de identidad fueron analizadas para determinar el número de cisteínas y la presencia de un péptido líder característico de las proteínas de este tipo, se utilizó el programa PrediSi (http://www.predisi.de/predisi/). Adicionalmente se realizaron modelados de estructura mediante iTASSER y SWISS model. Extracción de ARNm Para obtener el perfil de expresión de la proteína SN-1 en ajo, se deben de separar los tejidos a estudiar en un ajo vivo. Primeramente, estos se deben desinfectar adecuadamente para obtener condiciones asépticas siguiendo los siguientes pasos [10]. Lavar con una solución de SDS al 10%, posteriormente lavar con etanol 70% durante 30 segundos y durante 4 minutos con hipoclorito de sodio al 20%. Después enjuagar con agua de tal manera que no haya material genético externo que pueda alterar los resultados del procedimiento. Una vez que ha sido desinfectado en su totalidad, se procede a separar los tejidos con un bisturí. Los tejidos a separar son las raíces, las hojas, el tallo, el sistema radical, y el bulbo debe de ser separado en sus componentes, como lo son los dientes y las catáfilas. Una vez que se han obtenido los tejidos, se debe de seguir un protocolo de extracción de ARNm. Primero, se deben homogeneizar aproximadamente 100mg de las muestras de tejido en nitrógeno líquido, se resuspende en 400uL de Buffer de extracción (Tris-HCl pH 8 100 mM, EDTA pH 8 20mM, NaCl 1.4M, CTAB 2% p/v) y se incuba en un baño a 65°C (el buffer se debe precalentar previamente). La incubación debe durar 10 minutos y durante este tiempo procurar invertir 3 veces para mezclar. Después, añadir 400uL fenol-cloroformo (25:24 v/v) y agitar. Incubar 5 minutos a temperatura ambiente, para después centrifugar a 10,000xg 5 minutos a temperatura ambiente. Trasladar el sobrenadante a un tubo nuevo y añadir un volumen cloroformo-alcohol isoamílico (24:1 v/v), agitar. Incubar 5 minutos a temperatura ambiente. Posteriormente, centrifugar a 10,000xg 5 minutos a temperatura ambiente. Transferir la fase superior a un tubo nuevo y agregar un volumen de acetato de amonio 5M. Incubar en hielo 20 minutos (mínimo) y centrifugar a 14,000xg 20 minutos a 4 grados Celsius. Descartar el sobrenadante y lavar con Etanol 70% dos veces con (700uL). Centrifugar a 14,000xg 5 minutos y finalmente, secar y resuspender el pellet en 50uL de agua o Buffer TE (30-50uL). Tras seguir el protocolo anteriormente descrito, se realizó una electroforesis en gel de agarosa para visualizar la presencia y calidad del ARN obtenido. Síntesis de cDNA Antes de realizar la qRT-PCR, es necesario realizar la cuantificación y síntesis de cDNA de las muestras extraídas. Para la síntesis de cDNA se preparó una dilución y posteriormente un mix 1 (500ng de ARN, 1µl de Oligonucleótido dT, y agua grado biología molecular para ajustar a 12 µl). Después se procede a una incubación a 65°C durante 5 min, posteriormente se coloca en hielo. A continuación, se prepara una segunda mezcla (mix2). Se juntan ambas mezclas. Las mezclas se incuban en el termociclador a 42°C durante 60 minutos y posteriormente 70°C por 5 minutos. Finalmente, se almacenaron a -20°C.
Tabla 2. Ejemplo de cantidades de reactivo de Mix 1 para cada reactivo
Tabla 3. Ejempo de cantidades de reactivos de Mix 2 adicionadas a cada reactivo
qRT-PCR Con cDNA se realiza una reacción de qRT-PCR con los siguientes primers: Forward (5’-GAATTCGCATCGCCTTCTTCAATCTC-3’) y Reverse (5’-TCTAGAGTAGGACACTTGCGAGCTCCAC3’). Para tener una referencia de las especificaciones de los primers, se obtuvieron con TM Calculator de Thermo Fisher Scientific, tomando en cuenta la polimerasa a utilizar, así como la concentración de los primers (0.3μM). Se muestra a continuación la información correspondiente:

Figura 2. Datos de los primers a utilizar, por ejemplo, peso molecular, Tm de cada primer y temperatura de alineamiento
sugerida. Para medir el perfil de expresión, se utiliza el kit de qRT-PCR fabricado por Thermo Scientific Maxima SYBR Green/ROXq PCR MasterMix (2X) (código de catálogo: K0221). Se prepara un master mix de reacción adicionando los siguientes componentes (excepto cDNA templado) para cada 25μl de reacción en un tubo a temperatura ambiente.
Tabla 4. Reactivos y volúmenes para preparar master mix.
Primero se mezcla el master mix y se agregan volúmenes adecuados a tubos de PCR. Después se agrega el ADN templado (500 ng por reacción). Se mezcla gentilmente sin crear burbujas (centrifugar si es necesario). Finalmente se programa el termociclador.
Tabla 5. Programa de tiempos y temperatura por ciclos de amplificación.
Se realizaron reacciones por triplicado para cada muestra. El control de cada muestra se realizó con la amplificación del gen de actina, gen de tipo housekeeping que permite la comparación de la expresión. Por lo tanto, se depositó cada reacción con los contenidos descritos previamente en una placa de 96 pozos, con un total de 30 reacciones. Por último, se realizó una electroforesis en gel de agarosa con los triplicados de la muestra, es decir, 30 muestras en total.

RESULTADOS Y DISCUSIÓN
La secuencia de un péptido similar a SN-1 fue identificado mediante un análisis de Blast utilizando como secuencia de referencia la SN-1 de Solanum tuberosum. Se observó un porcentaje de identidad del 47% en el caso del ajo y 83% de identidad con un péptido de mango y de 77% con un péptido de eucalipto; en ambos casos resulto ser el porcentaje más alto de identidad. En todos los casos se observó que se mantienen conservados todos los puentes disulfuro presencia de 12 cisteínas (figura 3). Para corroborar que la estructura de los péptidos candidatos se mantiene conservada, se realizó la predicción de estructura de sus secuencias considerando que la forma madura de este péptido no incluye la secuencia líder. Para determinar la secuencia líder se utilizó el programa SignalP-5.0 Server, sin embargo, no fue posible identificar tal secuencia líder para los péptidos identificados de mango y eucalipto.
Figura 3. Identificación de los péptidos tipo Snakin en Ajo, mango y eucalipto. En la parte superior se aprecia el modelado de la estructura de los péptidos, en rojo se resalta la región amino terminal que resulta ser la región con mayor diferencia. En la parte inferior se aprecia el resultado del alineamiento del péptido identificado de SN1 de Mangifera Indica y Eucalyptus globulos contra la secuencia del péptido maduro de Allium sativum y Solanum tuberosum. Las flechas rojas señalan los 12 residuos de cisteínas que se mantienen conservados.
Para comenzar con el estudio de estos péptidos se decidió comenzar con el péptido candidato de ajo, ya que presentaba una señal péptido líder así como la conservación de las cisteínas. Como podemos observar en la figura 4, se identificó la secuencia tanto de aminoácidos como de nucleótidos que codifican para la versión madura de ajo. A partir de la secuencia identificada se realizó el diseño de primers para amplificar la región codificante de la versión madura de la snakin de ajo para poder estudiar su perfil de expresión así como clonar la secuencia codificante para su expresión recombinante.
Figura 4. Identificación de la secuencia del péptido tipo SN-1 de Allium sativum. A) Resultado de la predicción del péptido señal contenido en la secuencia de SN-1, B) y C) Representación del péptido señal identificado en color rojo, D) Secuencia codificante del péptido de ajo, mostrando en verde los codones que codifican para cisteínas, en letas rojas la secuencia codificante del péptido señal y en amarillo el codón de paro.

Para tener mejor detalle de la estructura del péptido de ajo y la formación de puentes disulfuro, que son característicos de este tipo de proteínas se realizó una predicción de su estructura (figura 5). Se identificaron dos alfa hélices y la formación de puentes disulfuro como ha sido reportado para otras proteínas de esta familia. Esto nos corrobora y nos sugiere que también podrá tener las funciones de los péptidos tipo Snakin. Con base en el análisis por cristalografía realizado por Kuddus, M. et al (2017) SN-1 presenta una estructura comprendida por 2 hélices cortas (α1 y α2) que forman una hélice-vuelta-hélice, una sección adicional que consiste de una hélice-310 corta, dos loops rígidos y seis enlaces disulfuro entre Cys5 -Cys30, Cys9 -Cys26, Cys13-Cys22, Cys29-Cys62, Cys33-Cys49 y d Cys35-Cys47. Estos últimos confieren estabilidad térmica, química y proteolíticas al péptido. La formación de los puentes disulfuro sucede de forma similar en ajo y las otras especies estudiadas.
Figura 5. Predicción de la estructura de SN-1 de ajo, mango y eucalipto mediante iTASSER, en amarillo se pueden apreciar los posibles puentes disulfuro que se forman entre los residuos de cisteína.
Tomando en cuenta la información previamente descrita, ya que SN-1 se encuentra expresada de forma constitutiva para algunos tejidos, es posible inferir que en el caso del ajo se encontrará expresado este péptido también de forma diferencial, dependiendo del tejido, por ello quisimos corroborar en hojas, raíces y diente de ajo. Para encontrar el perfil de expresión de SN-1 en el ajo, se realizó la extracción de ARN de diferentes tejidos con el fin de extraer su ARNm. En la figura 6 podemos apreciar que la extracción de ARN resultó con ARN de buena calidad, como se puede inferir al observar las bandas correspondientes a las bandas de ARN ribosomal.
Figura 6. Electroforesis de ARN aislado de diferentes tejidos de ajo. [A] Ejemplos de extracción de ARN de diente, hoja y raíz. En los carriles se tienen las siguientes muestras: 1) Marcador 100 pb Invitrogen (3 µl). 2) ARN obtenido a partir del tallo naciente del bulbo de ajo (5 µl). 3) ARN aislado del diente de la planta de ajo (5 µl). 4) ARN una hoja de ajo (5 µl). 5) ARN obtenido a partir de raiz de ajo (5 µl). [B] ejemplos de extracción de ARN de tallo de ajo (5 µl).
Al visualizar el gel, es claro que se cuenta con ARN para síntesis de cDNA previo a la RT-PCR. Adicionalmente se realizó una prueba de amplificación de SN1 para verificar la funcionabilidad de los primers diseñados y tener un primer acercamiento de la expresión de este gen en diversos tejidos antes de realizar la técnica de PCR en tiempo real. Se puede notar en la figura 7 que pudimos detectar un amplificado del tamaño esperado (288 pb) correspondiente a la secuencia codificante de la SN1 de ajo.
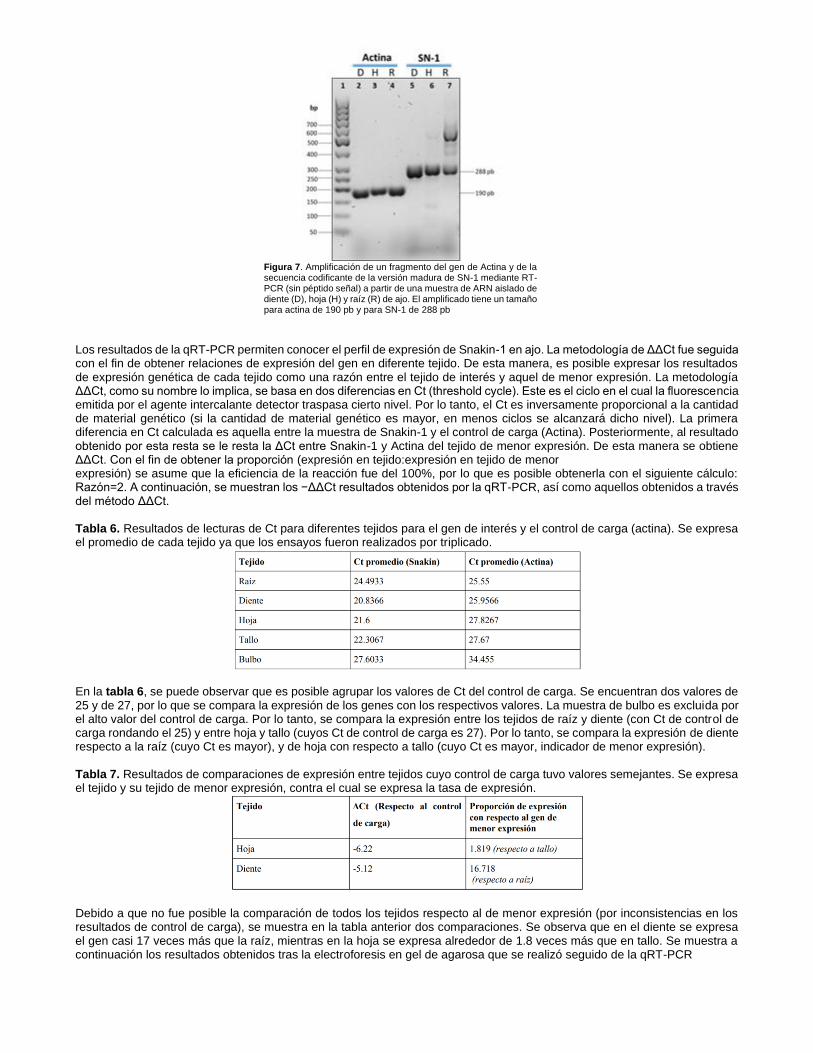
Figura 7. Amplificación de un fragmento del gen de Actina y de la secuencia codificante de la versión madura de SN-1 mediante RT-PCR (sin péptido señal) a partir de una muestra de ARN aislado de diente (D), hoja (H) y raíz (R) de ajo. El amplificado tiene un tamaño para actina de 190 pb y para SN-1 de 288 pb
Los resultados de la qRT-PCR permiten conocer el perfil de expresión de Snakin-1 en ajo. La metodología de ΔΔCt fue seguida con el fin de obtener relaciones de expresión del gen en diferente tejido. De esta manera, es posible expresar los resultados de expresión genética de cada tejido como una razón entre el tejido de interés y aquel de menor expresión. La metodología ΔΔCt, como su nombre lo implica, se basa en dos diferencias en Ct (threshold cycle). Este es el ciclo en el cual la fluorescencia emitida por el agente intercalante detector traspasa cierto nivel. Por lo tanto, el Ct es inversamente proporcional a la cantidad de material genético (si la cantidad de material genético es mayor, en menos ciclos se alcanzará dicho nivel). La primera diferencia en Ct calculada es aquella entre la muestra de Snakin-1 y el control de carga (Actina). Posteriormente, al resultado obtenido por esta resta se le resta la ΔCt entre Snakin-1 y Actina del tejido de menor expresión. De esta manera se obtiene ΔΔCt. Con el fin de obtener la proporción (expresión en tejido:expresión en tejido de menor expresión) se asume que la eficiencia de la reacción fue del 100%, por lo que es posible obtenerla con el siguiente cálculo: Razón=2. A continuación, se muestran los −ΔΔCt resultados obtenidos por la qRT-PCR, así como aquellos obtenidos a través del método ΔΔCt. Tabla 6. Resultados de lecturas de Ct para diferentes tejidos para el gen de interés y el control de carga (actina). Se expresa el promedio de cada tejido ya que los ensayos fueron realizados por triplicado.
En la tabla 6, se puede observar que es posible agrupar los valores de Ct del control de carga. Se encuentran dos valores de 25 y de 27, por lo que se compara la expresión de los genes con los respectivos valores. La muestra de bulbo es excluida por el alto valor del control de carga. Por lo tanto, se compara la expresión entre los tejidos de raíz y diente (con Ct de control de carga rondando el 25) y entre hoja y tallo (cuyos Ct de control de carga es 27). Por lo tanto, se compara la expresión de diente respecto a la raíz (cuyo Ct es mayor), y de hoja con respecto a tallo (cuyo Ct es mayor, indicador de menor expresión). Tabla 7. Resultados de comparaciones de expresión entre tejidos cuyo control de carga tuvo valores semejantes. Se expresa el tejido y su tejido de menor expresión, contra el cual se expresa la tasa de expresión.
Debido a que no fue posible la comparación de todos los tejidos respecto al de menor expresión (por inconsistencias en los resultados de control de carga), se muestra en la tabla anterior dos comparaciones. Se observa que en el diente se expresa el gen casi 17 veces más que la raíz, mientras en la hoja se expresa alrededor de 1.8 veces más que en tallo. Se muestra a continuación los resultados obtenidos tras la electroforesis en gel de agarosa que se realizó seguido de la qRT-PCR
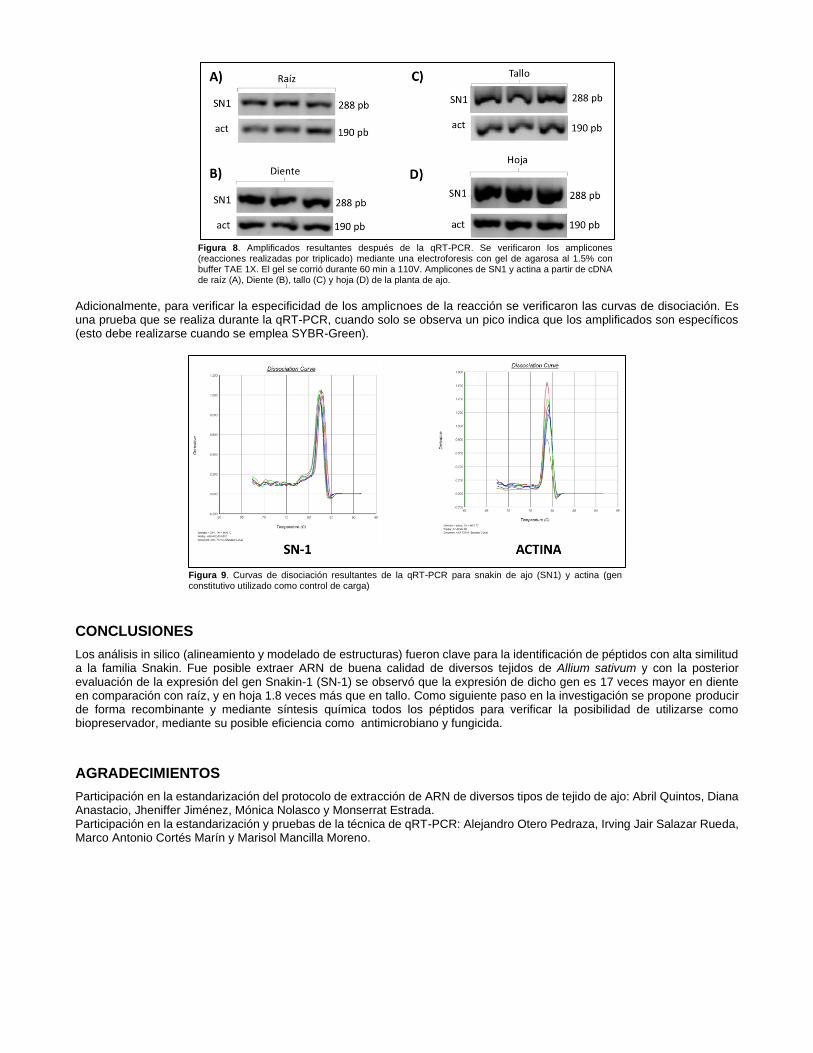
Figura 8. Amplificados resultantes después de la qRT-PCR. Se verificaron los amplicones (reacciones realizadas por triplicado) mediante una electroforesis con gel de agarosa al 1.5% con buffer TAE 1X. El gel se corrió durante 60 min a 110V. Amplicones de SN1 y actina a partir de cDNA de raíz (A), Diente (B), tallo (C) y hoja (D) de la planta de ajo.
Adicionalmente, para verificar la especificidad de los amplicnoes de la reacción se verificaron las curvas de disociación. Es una prueba que se realiza durante la qRT-PCR, cuando solo se observa un pico indica que los amplificados son específicos (esto debe realizarse cuando se emplea SYBR-Green).
Figura 9. Curvas de disociación resultantes de la qRT-PCR para snakin de ajo (SN1) y actina (gen constitutivo utilizado como control de carga)
CONCLUSIONES
Los análisis in silico (alineamiento y modelado de estructuras) fueron clave para la identificación de péptidos con alta similitud a la familia Snakin. Fue posible extraer ARN de buena calidad de diversos tejidos de Allium sativum y con la posterior evaluación de la expresión del gen Snakin-1 (SN-1) se observó que la expresión de dicho gen es 17 veces mayor en diente en comparación con raíz, y en hoja 1.8 veces más que en tallo. Como siguiente paso en la investigación se propone producir de forma recombinante y mediante síntesis química todos los péptidos para verificar la posibilidad de utilizarse como biopreservador, mediante su posible eficiencia como antimicrobiano y fungicida.
AGRADECIMIENTOS
Participación en la estandarización del protocolo de extracción de ARN de diversos tipos de tejido de ajo: Abril Quintos, Diana Anastacio, Jheniffer Jiménez, Mónica Nolasco y Monserrat Estrada. Participación en la estandarización y pruebas de la técnica de qRT-PCR: Alejandro Otero Pedraza, Irving Jair Salazar Rueda, Marco Antonio Cortés Marín y Marisol Mancilla Moreno.

REFERENCIAS
1. Berrocal-Lobo M., Segura A., Moreno M., López G., Garcı́a-Olmedo F., Molina M. (2002). Snakin-2, an Antimicrobial
Peptide from Potato Whose Gene Is Locally Induced by Wounding and Responds to Pathogen Infection. Plant Physiology Mar 2002, 128 (3) 951-961.
2. Das D, Jaiswal M, Khan F N, Ahamad S and Kumar S. (2020). PlantPepDB: A manually curated plant peptide
database.Scientific reports 10:2194
3. Kovalskaya N. and Hammond R. W. (2009). Expression and functional characterization of the plant antimicrobial snakin-1 and defensin recombinant proteins, Protein Expr. Purif., vol. 63, no. 1, pp. 12–17, Jan. 2009.
4. Kikukawa, T., Demura, M., & Aizawa, T. (2017). Enhanced expression of cysteine-rich antimicrobial peptide snakin-1 in Escherichia coli using an aggregation-prone protein coexpression system. Biotechnology progress, 33(6), 1520–1528.
5. Mahalik, G., Sahoo, S. & Satapathy, K. (2017). Evaluation of phytochemical constituents and antimicrobial properties of Mangifera Indica L. Leaves against urinary tract infection-causing pathogens. Asian Journal of Pharmaceutical and Clinical Research. 10. 169.
6. Mikaili P., Maadirad S., Moloudizargari M., Aghajanshakeri S., and Sarahroodi S. (2013). Therapeutic uses and pharmacological properties of garlic, shallot, and their biologically active compounds. Iranian Journal of Basic Medical Sciences, vol. 16, no. 10. pp. 1031–1048.
7. Mookherjee, N., Anderson, M.A., Haagsman, H.P. et al. Antimicrobial host defence peptides: functions and clinical potential. Nat Rev Drug Discov 19, 311–332 (2020).
8. Nahirñak, V., Almasia N. I., Hopp H. E., and Vazquez-Rovere C. (2012). “Snakin/GASA proteins,” Plant Signal. Behav., vol. 7, no. 8, pp. 1004–1008.
9. Nezhad, F. M., Zeigham, H., Mota, A., Sattari, M., & Yadegar, A. (2009). Antibacterial activity of Eucalyptus extracts on methicillin resistance Staphylococcus aureus. Research Journal of Biological Sciences, 4(8), 905-908.
10. Porto W. F. and Franco O. L. (2013). “Theoretical structural insights into the snakin/GASA family”. Peptides, Volume
44, Pages 163-167.
11. Yeung, H., Squire, C., Yosaatmadja, Y., Panjikar, S., López, G., Molina, A., Baker, E., Harris, P., & Brimble, M. (2016). Radiation Damage and Racemic Protein Crystallography Reveal the Unique Structure of the GASA/Snakin Protein Superfamily. Angewandte Chemie, 55 28, 7930-3 .











![Péptidos y proteínas[1]](https://static.fdocuments.ec/doc/165x107/557b2192d8b42a726a8b4a97/peptidos-y-proteinas1.jpg)

![Péptidos de colágeno – Soluciones versátiles en salud y ... · ¿Qué son los péptidos de colágeno? [2] Los péptidos de colágeno, son cadenas pequeñas de proteínas, que](https://static.fdocuments.ec/doc/165x107/5ac079cc7f8b9ac6688c3499/pptidos-de-colgeno-soluciones-verstiles-en-salud-y-son-los-pptidos-de-colgeno.jpg)




