
Estructura y Diversidad - Universidade de Santiago de ... · distribución de las diferentes...
140
Estructura y Diversidad 141 Estructura y Diversidad La estructura y la diversidad son, junto con la densidad, las principales características de los rodales. La diversidad es un concepto que permite diferentes interpretaciones aunque, en general, se emplea este término como sinónimo de diversidad de especies. Por otro lado, la estructura de un bosque hace referencia a la distribución de las principales características arbóreas en el espacio, teniendo especial importancia la distribución de las diferentes especies y la distribución de las mismas por clases de dimensión. Por tanto, habitualmente son las distribuciones de frecuencia de los atributos de los árboles las herramientas empleadas para describir la estructura del bosque. Dicha estructura viene determinada no solo por la distribución más o menos regular de los árboles en el terreno, sino sobre todo, por la mezcla espacial de las distintas especies y el grado de mezcla de árboles con diferentes dimensiones.
Transcript of Estructura y Diversidad - Universidade de Santiago de ... · distribución de las diferentes...

Estructura y Diversidad 141
Estructura y Diversidad
La estructura y la diversidad son, junto con la densidad, las principales características
de los rodales. La diversidad es un concepto que permite diferentes interpretaciones
aunque, en general, se emplea este término como sinónimo de diversidad de especies.
Por otro lado, la estructura de un bosque hace referencia a la distribución de las
principales características arbóreas en el espacio, teniendo especial importancia la
distribución de las diferentes especies y la distribución de las mismas por clases de
dimensión. Por tanto, habitualmente son las distribuciones de frecuencia de los
atributos de los árboles las herramientas empleadas para describir la estructura del
bosque. Dicha estructura viene determinada no solo por la distribución más o menos
regular de los árboles en el terreno, sino sobre todo, por la mezcla espacial de las
distintas especies y el grado de mezcla de árboles con diferentes dimensiones.

Estructura y Diversidad 142
Diversidad de especies Una consecuencia directa de la extensa deforestación que viene sucediendo desde la
segunda mitad del sigo XX es la elevada desaparición de especies. Muchas especies de
animales y plantas se extinguieron o están amenazadas de extinción en amplias áreas
geográficas. Esta preocupante situación trajo como consecuencia una actividad mucho
más intensa en el campo de la investigación sobre la diversidad. Entre los resultados
obtenidos en los trabajos de investigación llevados a cabo en este campo destacan las
relaciones existentes entre el tamaño de la superficie analizada y el número de especies
y entre los diferentes estados de sucesión y el número de especies en el ecosistema
forestal.
El número de especies aumenta, como era de esperar, con la superficie analizada.
La figura 4-1 muestra la relación empírica entre el tamaño de la superficie muestreada
y el número de especies arbóreas y arbustivas en Panamá (izquierda según Hubbell,
2001). Como se observa en la figura, el número de especies aumenta
considerablemente al aumentar la superficie muestreada entre 100 m² y 50 ha. En
cambio, un aumento de la superficie de 50 a 1500 ha no tiene una influencia tan
marcada en el número de especies observadas. Al considerar superficies mucho más
extensas, como por ejemplo la región del Canal o la totalidad del territorio de Panamá
se observa de nuevo un claro incremento del número de especies arbóreas y arbustivas
presentes.
En la figura 4-1 derecha se muestra “la curva teórica de las tres fases” de la
diversidad (Hubbell, 2001). Cuando se considera una escala local, el número de
especies aumenta fuertemente con la superficie. Al considerar una escala regional, la
frecuencia acumulada de especies se ve menos afectada por la superficie analizada,
mientras que el empleo de una dimensión biogeográfica continental o intercontinental

Estructura y Diversidad 143
supone un fuerte incremento del número de especies a medida que aumenta la
superficie como consecuencia de un desarrollo separado del proceso evolutivo.
1,0
1,5
2,0
2,5
3,0
3,5
-3,0 -1,0 1,0 3,0 5,0 7,0
Log10 (Superficie ha)
Log
10 (
n°
de
esp
ecie
s)
Panamá (total)
Panamá (Canal)
50 ha Sup.
local regional continental
Log
(n
° d
e es
pec
ies)
Log (Superficie)
Figura 4-1. Relación empírica entre la superficie y el número de especies arbustivas y arbóreas en Panamá (izquierda) y el modelo general derivado (derecha. Según Hubbell, 2001, páginas 161 y 199).
A escala local la biodiversidad de los bosques se ve influida no solo por la superficie
considerada sino también por el manejo forestal. La figura 4-2 muestra la relación
entre los diferentes estados de sucesión natural y el número de especies en un
ecosistema forestal de haya según Jenssen y Hoffmann (2002).
Estado de árbol juvenil
Estado de árbol adulto
Estado de árbol deedad avanzada
Estado dedescomposición
Estado de crecimiento juvenil
2022
25 8
3
Figura 4-2. Número medio de especies de plantas en parcelas de 400 m² de superficie en cada uno de los diferentes estados de sucesión de un bosque de haya (según Jenssen y Hoffmann, 2002).

Estructura y Diversidad 144
El número medio de especies de plantas para una misma superficie aumenta
notablemente desde el estado de crecimiento juvenil hasta el estado de árbol adulto. El
mayor número de especies se observó en el estado de descomposición, debido a que al
producirse cambios a pequeña escala en las condiciones de luz y sombra se favorece la
existencia de diferentes micro-estaciones con características ligeramente diferentes
para el crecimiento de las especies.
En la figura 4-3 se muestra la relación entre las frecuencias relativas de
abundancia de especies y el rango o posición social que ocupa cada una de las especies
presentes en tres tipos de ecosistemas forestales (Hubbell, 2001).
Rango de la frecuencia de especies
0 50 100 150 200 250 10- 7
10 - 6
10- 5
10- 4
10 - 3
10 - 2
10 - 1
10 0
Fre
cuen
cia
rela
tiva
de
esp
ecie
s
Bosque boreal
Bosque tremplado
de forndosas
Bosque tropical
Figura 4-3. Representación simplificada de las relaciones de dominancia y diversidad para tres ecosistemas forestales según Hubbell (2001, página 116).
Una especie de rango elevado implica una mayor frecuencia relativa, es decir, una
mayor presencia o dominancia sobre el resto de las especies. La forma de las relaciones
diversidad-dominancia son similares. El primer ejemplo de la figura 4-3 muestra la
curva obtenida para un bosque boreal con menos de 10 especies. El segundo ejemplo
corresponde a un bosque de frondosas de zona templada en el que se encontraron 40
especies diferentes en una parcela de ensayo de 1 ha. En este caso, el intervalo de
variación de las frecuencias relativas de cada especie es menor que en el bosque boreal.

Estructura y Diversidad 145
La menor dispersión de la frecuencia relativa de cada especie se observa en un bosque
tropical en el que se encontraron más de 250 especies en una superficie de 4 ha.
Las especies más frecuentes se convierten en dominantes a medida que
disminuye la riqueza de especies. Como consecuencia, la dispersión de la frecuencia
relativa de especies aumenta en los ecosistemas con un menor número de especies.
Especialmente destacable es el hecho de que las relaciones representadas en la figura
anterior siguen una pauta en forma de S, similar en los tres casos. Esa relación en
forma de S es independiente de la superficie analizada y puede ser descrita mediante
un modelo matemático cuyos parámetros se determinan a partir del número de
especies presentes (Hubbell, 2001).
Distribución diamétrica unimodal La diversidad de un bosque se caracteriza no solo por el número de especies existentes
sino también por la distribución de las dimensiones de los árboles. Dos de las variables
de dimensión más relevantes para la práctica forestal son el diámetro a la altura de
pecho y la altura. La distribución de frecuencias de estas variables se muestra con un
ejemplo de una parcela de ensayo de pícea de 116 años en el Solling (Alemania). La
parcela de ensayo, cuya superficie es de 0,16 ha, tiene 41 árboles cuyos diámetros se
muestran en la Tabla 4-1.
Diámetro cm
41, 41, 38, 53, 44, 42, 50, 43, 40, 44, 40, 33, 39, 32, 49, 47, 38, 40, 37, 34, 47, 37, 41, 38, 38, 43, 40, 42, 34, 39, 41, 44, 41, 45, 43, 36, 36, 46, 46, 34, 50
Tabla 4-1. Lista de diámetros (cm) en una parcela de ensayo de pícea de 116 años en el Solling (Alemania).
Con los datos de diámetros del rodal se puede construir la distribución de frecuencias
de diámetros agrupando los árboles por clases diamétricas y asignando la frecuencia
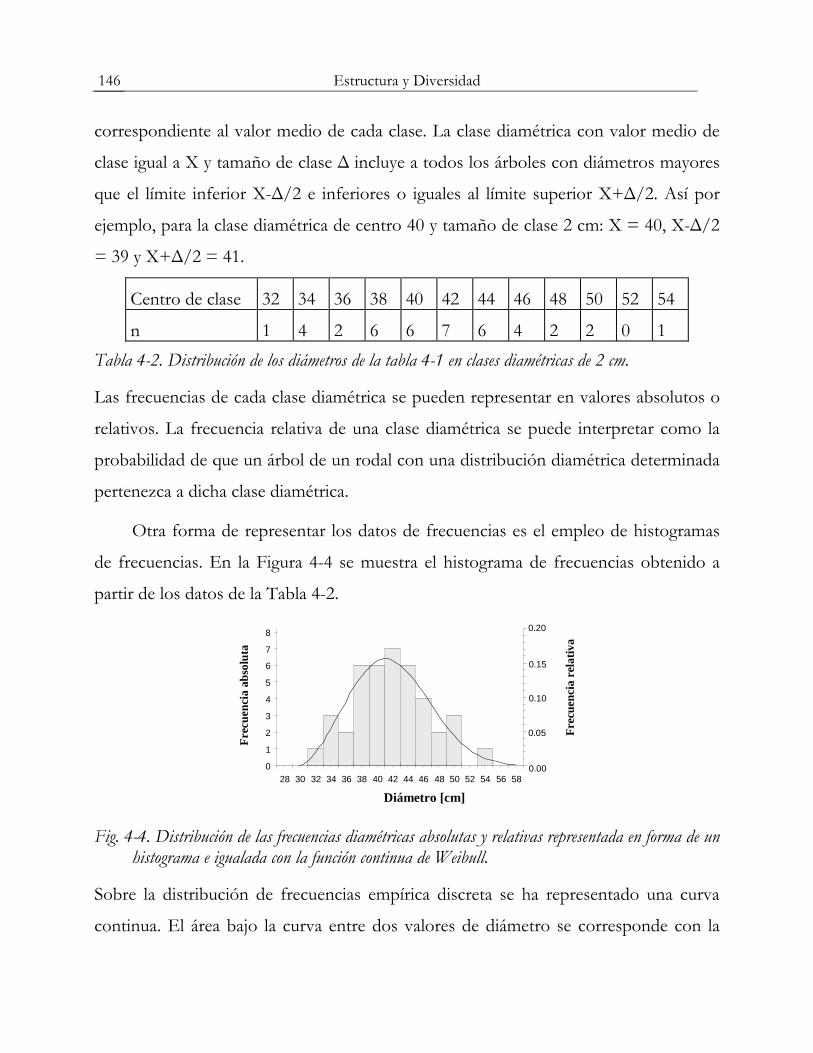
Estructura y Diversidad 146
correspondiente al valor medio de cada clase. La clase diamétrica con valor medio de
clase igual a X y tamaño de clase Δ incluye a todos los árboles con diámetros mayores
que el límite inferior X-Δ/2 e inferiores o iguales al límite superior X+Δ/2. Así por
ejemplo, para la clase diamétrica de centro 40 y tamaño de clase 2 cm: X = 40, X-Δ/2
= 39 y X+Δ/2 = 41.
Centro de clase 32 34 36 38 40 42 44 46 48 50 52 54
n 1 4 2 6 6 7 6 4 2 2 0 1 Tabla 4-2. Distribución de los diámetros de la tabla 4-1 en clases diamétricas de 2 cm.
Las frecuencias de cada clase diamétrica se pueden representar en valores absolutos o
relativos. La frecuencia relativa de una clase diamétrica se puede interpretar como la
probabilidad de que un árbol de un rodal con una distribución diamétrica determinada
pertenezca a dicha clase diamétrica.
Otra forma de representar los datos de frecuencias es el empleo de histogramas
de frecuencias. En la Figura 4-4 se muestra el histograma de frecuencias obtenido a
partir de los datos de la Tabla 4-2.
0 1 2 3 4 5 6 7 8
28 30 32 34 36 38 40 42 44 46 48 50 52 54 56 58
Diámetro [cm]
Frec
uenc
ia a
bsol
uta
0.00
0.20
0.15
0.10
0.05 Frec
uenc
ia r
elat
iva
Fig. 4-4. Distribución de las frecuencias diamétricas absolutas y relativas representada en forma de un histograma e igualada con la función continua de Weibull.
Sobre la distribución de frecuencias empírica discreta se ha representado una curva
continua. El área bajo la curva entre dos valores de diámetro se corresponde con la

Estructura y Diversidad 147
probabilidad de que un árbol cualquiera del rodal tenga un diámetro comprendido
entre esos dos valores. A este tipo de funciones continuas se las denomina funciones
de densidad. En el ejemplo de la figura 4-4 se ha empleado la función de densidad de
Weibull.
Estas funciones de densidad no se suelen emplear para estimar la probabilidad
con la que una variable continua, por ejemplo el diámetro, toma un valor determinado,
sino que más bien se usan para obtener la probabilidad con la que un valor se
encuentra en un intervalo determinado, por ejemplo una clase diamétrica.
Las curvas de las funciones de densidad para cada rodal se obtienen mediante la
estimación por métodos matemáticos de los parámetros que definen la expresión
matemática de la curva a partir de los datos reales. La mayoría de las funciones de
densidad tienen expresiones matemáticas complejas que dificultan la estimación de su
parámetros por lo que, a menudo es preferible emplear las funciones de distribución
de probabilidad que se obtienen empleando datos de frecuencias acumuladas. En el
caso de clases diámetricas, la frecuencia acumulada se obtiene sumando las frecuencias
de todas las clases diamétricas menores o iguales que la analizada. En el caso de
funciones continuas, esa suma es una integral y las funciones de distribución de
probabilidad se obtienen como integración de las funciones de densidad de
probabilidad. La función de distribución de Weibull se obtendría de la integración de
la función de densidad de Weibull (Ecuación. 4-1):
c
badd
dXXXdPd⎟⎠⎞
⎜⎝⎛
∞−
−−
−==≤= ∫ e1)D()()F( 4-1
Siendo X un diámetro escogido aleatoriamente, D el diámetro para el que se quiere estimar la probabilidad de que su valor sea menor o igual que X, D(X) la función de densidad de probabilidad de Weibull, F(d) es la función de distribución de probabilidad de Weibull, a es el parámetro de posición de la función Weibull, b es el parámetro de escala y c es el parámetro de forma de la función Weibull.

Estructura y Diversidad 148
En el ejemplo de la Tabla 4.2 los valores de los parámetros de posición, escala y forma
de la función de Weibull son a =30, b =13,4 y c =2,6. De este modo, se puede
emplear la ecuación 4-1 para determinar la frecuencia relativa de cualquier clase
diamétrica. La frecuencia absoluta de la clase diamétrica sería igual al producto de la
frecuencia relativa por el número total de árboles. Por ejemplo, la frecuencia relativa
del número de pies en la clase diamétrica de centro 44 y 2 centímetros de tamaño de
clase [P(43 ≤ d ≤ 45)], se obtiene mediante la resta P(d ≤ 45) - P(d ≤ 43) = 0,74 - 0,6 =
0,14, Finalmente, si se multiplica este valor por el número de pies del rodal se obtiene
la frecuencia absoluta correspondiente a esta clase diamétrica (41),, 0,14 = 5,74 - es
decir - aproximadamente 6 árboles.
La expresión matemática para estimar la frecuencia relativa de una clase
diamétrica empleando la función de distribución de Weibull es la siguiente:
( ) ( ) ( )c
b
adc
b
ad
eedFdFdXdP⎟⎠⎞
⎜⎝⎛
⎟⎠⎞
⎜⎝⎛ −
−−
−
−=−=≤<
infsup
infsupsupinf 4-2
Donde dinf y dsup son el límite inferior y el límite superior de la clase diamétrica,
respectivamente
Si se invierte la función de distribución de Weibull, es decir, se despeja el valor
del diámetro en función de su frecuencia relativa, se obtiene una expresión matemática
que permite simular distribuciones de diámetros teóricas:
( )dF = c
bad
⎟⎠⎞
⎜⎝⎛−
−
− e1
d = ( )( )[ ] cdnba1 F1−−⋅+ l = ( )( )[ ] c
dXPnba1
>−⋅+ l 4-3
Dónde P(X > d) = 1-F(d) es la probabilidad de que un diámetro escogido aleatoriamente X sea mayor que d (un número aleatorio de distribución homogénea en el intervalo [0, 1]) y a, b y c son los parámetros de la función de Weibull.

Estructura y Diversidad 149
Con la ecuación 4-3 se puede determinar, por ejemplo, cuál debe ser el diámetro de un
árbol para que haya un 50% de probabilidad de que sea menor que un árbol escogido
aleatoriamente, o lo que es lo mismo, cuál debe ser el diámetro de un árbol para que la
mitad de los árboles del rodal tengan un diámetro mayor que él. Para el rodal de la
figura 4-4 la respuesta sería ( )[ ] 64150ln41330 621
...x . =−⋅+= cm. De este modo, con la
ecuación 4.3 se pueden simular distribuciones diamétricas, para unos valores dados de
los parámetros de la función de Weibull, simplemente al generar números aleatorios
entre 0 y 1 y sustituirlos por el término P(X > d).
La distribución de Weibull puede ser similar a la distribución normal, pero
también puede presentar asimetría hacia la derecha o hacia la izquierda e incluso puede
representar la forma en J invertida de las distribuciones diamétricas de masas
irregulares. El parámetro de escala (b) esta relacionado con el recorrido o rango de los
diámetros de la distribución analizada mientras que el parámetro de forma (c) define la
forma de la curva obtenida de manera que: si c < 1 se obtienen curvas en forma de J
invertida típicas de distribuciones diamétricas de masas irregulares; si c = 1, coincide
con la distribución exponencial; si 1 < c < 3,6 la distribución presenta asimetría hacia
la derecha; si c = 3,6 la distribución de Weibull se aproxima a la normal y si c > 3,6 la
distribución presenta asimetría hacia la izquierda.
Distribuciones diamétricas multimodales Como se ha comentado anteriormente, las distribuciones diamétricas pueden presentar
diferentes formas dependiendo de cómo sea la estructura del rodal o del efecto de las
claras realizadas (Álvarez González et al., 2002). Además, en el caso de que se
consideren otras variables dendrométricas o dasométricas, la forma de las
distribuciones también puede variar. En la figura 4-5 se muestran algunos ejemplos de

Estructura y Diversidad 150
distribuciones diamétricas y de alturas típicas de rodales forestales con diferentes
estructuras.
0
2
4
6
8
10
12
28 32 36 40 44 48 52 56 60 Diámetro [cm]
N
0
2
4
6
8
10
12
14
26 28 30 32 34 36 38 40 42 Altura [m]
N
(a) (b)
0 5
10 15 20 25 30 35 40 45
5 10 15 20 25 30 35 40 45 50 55 60 65 70
Diámetro [cm]
N
0
2
4
6 8
10
12
14
8 12 16 20 24 28 32 36 40 Altura [m]
N
(c) (d)
0
10
20
30
40
50
60
70
80
0,5 1 1,5 2 2,5 3 Altura [m]
N
0
4
8
12
16
20
24
28
8 12 16 20 24 28 32 36 40 44 48 52 56 60 64
Diámetro [cm]
N
(e) (f)
Figura 4-5. Típicas distribuciones de frecuencia (a) el diámetro en rodales regulares puros (asimetría a la izquierda), (b) altura de los árboles en rodales regulares puros (asimetría a la derecha), (c) el diámetro en rodales de 2 estratos y (d) la altura de los árboles en rodales mixtos de dos estratos (dos máximos), (e) altura de las plantas en estadio juvenil, rodal regular puro (distribución normal) y (f) diámetro en monte irregular (exponencial negativa, forma de J invertida).
La función de Weibull se puede emplear para representar distribuciones bimodales, es
decir, con dos máximos (Puumalainen, 1996; Condés, 1997; Hessenmöller, 2001).
También se pueden representar mediante la función de Weibull distribuciones
diamétricas de rodales mixtos en las que se superponen las distribuciones
correspondientes a cada especie (ver, por ejemplo los trabajos de Chung, 1996 o Liu et
al., 2002). En la Figura 4-6 se muestra la distribución diamétrica de un rodal mixto de
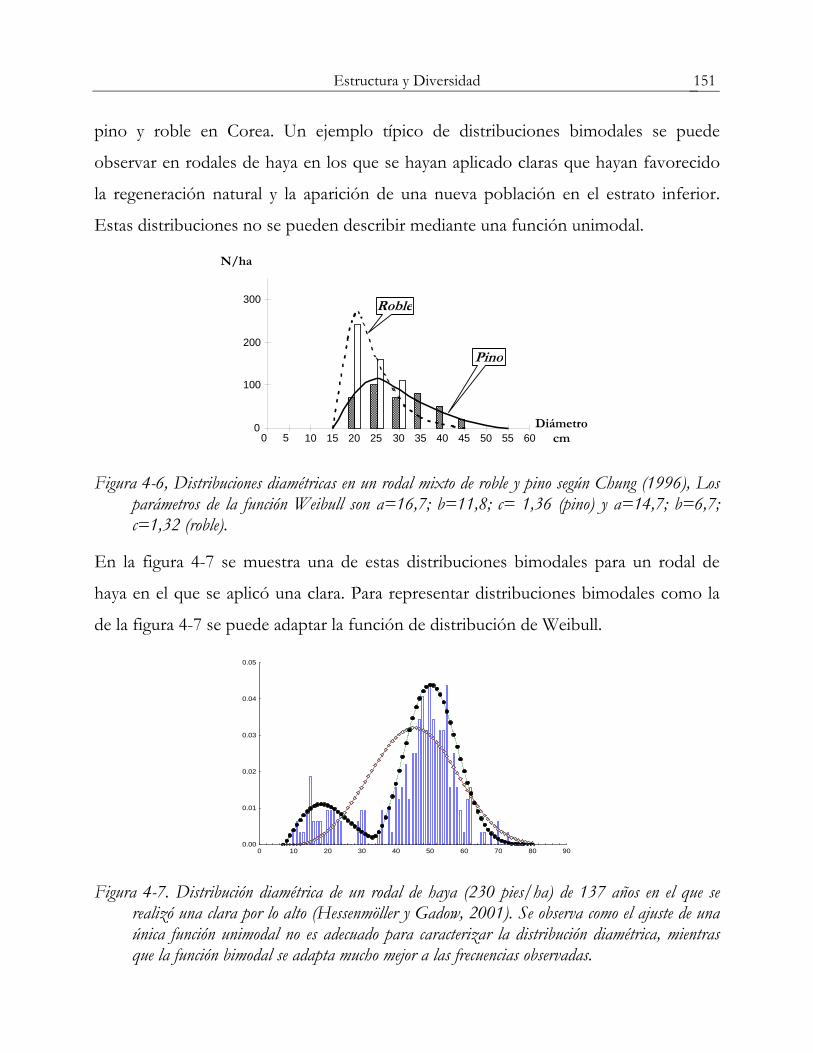
Estructura y Diversidad 151
pino y roble en Corea. Un ejemplo típico de distribuciones bimodales se puede
observar en rodales de haya en los que se hayan aplicado claras que hayan favorecido
la regeneración natural y la aparición de una nueva población en el estrato inferior.
Estas distribuciones no se pueden describir mediante una función unimodal.
0
100
200
300
0 5 10 15 20 25 30 35 40 45 50 55 60Diámetro
cm
N/ha
Roble
Pino
Figura 4-6, Distribuciones diamétricas en un rodal mixto de roble y pino según Chung (1996), Los parámetros de la función Weibull son a=16,7; b=11,8; c= 1,36 (pino) y a=14,7; b=6,7; c=1,32 (roble).
En la figura 4-7 se muestra una de estas distribuciones bimodales para un rodal de
haya en el que se aplicó una clara. Para representar distribuciones bimodales como la
de la figura 4-7 se puede adaptar la función de distribución de Weibull.
0.00
0.01
0.02
0.03
0.04
0.05
0 10 20 30 40 50 60 70 80 90
Figura 4-7. Distribución diamétrica de un rodal de haya (230 pies/ha) de 137 años en el que se realizó una clara por lo alto (Hessenmöller y Gadow, 2001). Se observa como el ajuste de una única función unimodal no es adecuado para caracterizar la distribución diamétrica, mientras que la función bimodal se adapta mucho mejor a las frecuencias observadas.

Estructura y Diversidad 152
En bosques de haya se pueden encontrar estructuras diferentes dependiendo del
estado de la sucesión en que se encuentre el rodal (ver, por ejemplo los trabajos de
Korpel, 1992; Košir, 1966; Emborg et al., 2000; Oheimb et al., 2005). Wenk (1996) y
Condés (1997) recomiendan emplear dos funciones de distribución, una para cada
estrato, calcular los parámetros de cada función separadamente y finalmente
establecer condiciones de continuidad entre ambas. La distribución bimodal típica de
rodales de estas características se obtendría empleando la siguiente expresión
matemática:
)()1()()( supinf dfgdfgdf ⋅−+⋅= 4-4
Donde y describen las funciones para el estrato inferior y superior,
respectivamente y g es el parámetro de conexión de las dos funciones. En el caso de
que se emplee la función de distribución de Weibull, su expresión para distribuciones
bimodales es la siguiente:
)(inf df )(sup df
⎪⎪⎪⎪⎪⎪⎪⎪
⎩
⎪⎪⎪⎪⎪⎪⎪⎪
⎨
⎧
<
⎥⎥⎥⎥
⎦
⎤
⎢⎢⎢⎢
⎣
⎡
⋅⎟⎟⎠
⎞⎜⎜⎝
⎛ −⋅⋅−+
⎥⎥⎥⎥
⎦
⎤
⎢⎢⎢⎢
⎣
⎡
⋅⎟⎟⎠
⎞⎜⎜⎝
⎛ −⋅⋅
≤<
⎥⎥⎥⎥
⎦
⎤
⎢⎢⎢⎢
⎣
⎡
⋅⎟⎟⎠
⎞⎜⎜⎝
⎛ −⋅⋅
≤
=
⎟⎟⎟
⎠
⎞
⎜⎜⎜
⎝
⎛ −−−
⎟⎟
⎠
⎞
⎜⎜
⎝
⎛ −−−
⎟⎟
⎠
⎞
⎜⎜
⎝
⎛ −−−
daeb
adbc
geb
adbcg
adaeb
adbcg
ad
df
c
b
adc
c
badc
c
badc
sup
sup
sup
sup1
sup
sup
sup
sup
inf
infinf1
inf
inf
inf
inf
supinf
inf
infinf1
inf
inf
inf
inf
inf
supinf
inf
)1(
0
)(
para
para
para
4-5
Esta expresión se utilizó para representar los datos observados en la figura 4-7 y,
posteriormente se evaluó la bondad del ajuste mediante el cálculo de un estadístico
denominado distancia genética (Gregorius, 1974; Pommerening, 1997) que, en este

Estructura y Diversidad 153
caso, evalúa la similitud entre las frecuencias observadas y las estimadas. La distancia
genética para esta comparación se calcula como (Niggemeyer, 1999):
1ˆ210
1≤−=≤ ∑
=
n
iii ddd 4-6
id y representan la frecuencia estimada y la observada para la clase diamétrica i
respectivamente y n es el número de clases diamétricas. Si id
1=d las distribuciones no
tienen nada en común, si las distribuciones son idénticas. 0=d
Esta función bimodal se aplicó a tres grupos de parcelas diferentes para analizar
su capacidad para caracterizar las distribuciones de rodales de haya en Alemania
(Hessenmöller y Gadow, 2001). El primer grupo de datos corresponde a una red de
parcelas de ensayo en las que no se habían realizado claras y con edades comprendidas
entre 51 y 150 años. El segundo grupo corresponde a una red de parcelas de ensayo
del Instituto de investigación forestal de la Baja Sajonia en las que se habían realizado
claras por lo alto de intensidad media y de edades comprendida entre 56 y 98 años. El
tercer grupo se corresponde con 30 rodales puros de haya del distrito forestal de
Paderborn de edad comprendida entre 59 y 137 años en los que se midieron todos los
diámetros. Los resultados obtenidos mostraban que, como era de esperar, las
funciones bimodales describen mejor los rodales adultos de haya que la función
unimodal.
Distribución de diámetros y alturas Otra característica estructural esencial de un rodal, frecuentemente resultado de los
tratamientos selvícolas realizados, es la distribución de las alturas. Esta distribución
describe la estructura vertical de un rodal y, al igual que la distribución diamétrica,
también se puede caracterizar mediante un histograma de frecuencias o mediante una
función de distribución continua. La figura 4-8 (izquierda) muestra la relación entre las
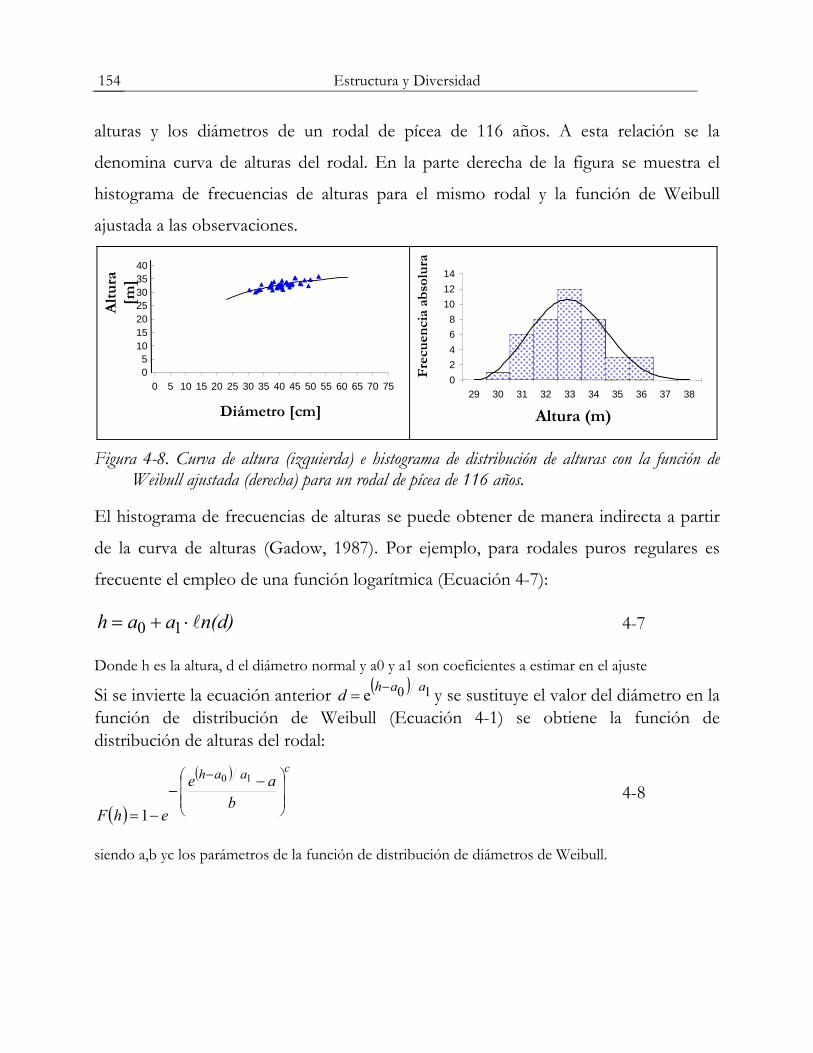
Estructura y Diversidad 154
alturas y los diámetros de un rodal de pícea de 116 años. A esta relación se la
denomina curva de alturas del rodal. En la parte derecha de la figura se muestra el
histograma de frecuencias de alturas para el mismo rodal y la función de Weibull
ajustada a las observaciones.
0 5
10 15 20 25 30 35 40
0 5 10 15 20 25 30 35 40 45 50 55 60 65 70 75
Diámetro [cm]
Alt
ura
[m
]
02468
101214
29 30 31 32 33 34 35 36 37 38
Altura (m) F
recu
enci
a ab
solu
ra
Figura 4-8. Curva de altura (izquierda) e histograma de distribución de alturas con la función de Weibull ajustada (derecha) para un rodal de pícea de 116 años.
El histograma de frecuencias de alturas se puede obtener de manera indirecta a partir
de la curva de alturas (Gadow, 1987). Por ejemplo, para rodales puros regulares es
frecuente el empleo de una función logarítmica (Ecuación 4-7):
n(d)aah l⋅+= 10 4-7
Donde h es la altura, d el diámetro normal y a0 y a1 son coeficientes a estimar en el ajuste
Si se invierte la ecuación anterior ( ) 10e aahd −= y se sustituye el valor del diámetro en la función de distribución de Weibull (Ecuación 4-1) se obtiene la función de distribución de alturas del rodal:
( )
( ) c
bae
ehF
aah
⎟⎟⎟
⎠
⎞
⎜⎜⎜
⎝
⎛ − −−
−=
10
1 4-8
siendo a,b yc los parámetros de la función de distribución de diámetros de Weibull.

Estructura y Diversidad 155
Curvas de altura de árbol individual
La medición de las alturas de los árboles en el terreno sigue siendo más costosa que la
medición de diámetros a pesar de las mejoras de los aparatos. Por lo tanto, para limitar
el número de mediciones de alturas se pueden emplear las curvas locales de altura de
rodal o las curvas generalizadas de altura (Kramer y Akça, 1995, página 138 y
siguientes). Las primeras describen la relación entre las alturas y los diámetros de un
rodal concreto mientras que las segundas relacionan la altura de los árboles con su
diámetro y también con variables dasométricas del rodal como puede ser su edad o su
densidad.
Ejemplos de ecuaciones de altura generalizada se pueden encontrar en los
trabajos de Hui y Gadow (1993a) para rodales regulares puros de la especie
Cunninghamia lanceolata en China; Soares y Tomé (2002) para plantaciones de eucalipto
en Portugal; López Sánchez et al. (2003) para Pinus radiata en Galicia; Bravo Oviedo y
Montero (2005) para Pinus pinea en España o Castedo et al. (2005) para Pinus pinaster en
Galicia incluyendo un término estocástico para tener en cuenta la variabilidad existente
en altura para un mismo diámetro.
Temesgen y Gadow (2003) desarrollaron curvas generalizadas de altura para
rodales mixtos en la región interior de British Columbia en Canadá. En este trabajo se
analizaron ocho especies diferentes: álamo (Populus tremuloides); thuja (Thuja plicata);
abedul (Betula papyrifera); pino de Oregón (Pseudotsuga menziesii); alerce (Larix occidentalis);
dos especies de pino (Pinus contorta y Pinus ponderosa) y pícea (Picea engelmanii). Se
analizaron cinco modelos diferentes con distintas combinaciones de variables
dasométricas. La inclusión en el modelo del índice de competencia BAL (que
considera al mismo tiempo la densidad del rodal y la posición relativa de un árbol
dentro de la distribución diamétrica) mejoró considerablemente la exactitud de las
estimaciones. Finalmente se recomendó el empleo del siguiente modelo:

Estructura y Diversidad 156
⎟⎠⎞
⎜⎝⎛ −⋅+= ⋅ cdbeah 13.1 4-9
Donde los parámetros a y c son función de variables dasométricas según las
relaciones: GaNaBALaaa ⋅+⋅+⋅+= 4321( y );65 BALaac ⋅+= BAL es el
área basimétrica de los árboles mayores que el analizado (m²/ha); G es el área
basimétrica del rodal (m²/ha); N es el número de pies/ha y b y ai (i=1,...,6) son
coeficientes propios de cada especie (ver tabla 4-3). A b C Especie a1 a2 a3 a4 b a5 a6
Álamo 20,655 0,08724 0,01509 1,399 Thuja 17,947 -0,0009 0,14087 0,03497 1,304 Abedul 20,446 -0,0007 0,13355 0,03576 1,262 Pino de Oregón 32,037 -0,3504 -0,0007 0,18308 0,01797 1,093 0,00802 Alerce 41,792 0,01709 1,118 0,00404 Pino contorta 20,852 0,3168 -0,0004 0,23962 0,03184 1,087 -0,0014 Pino ponderosa 32,208 0,01738 1,107 Pícea 17,080 0,0932 0,34276 0,01073 1,462
Tabla 4-3. Parámetros estimados para la ecuación 4-9 (Temesgen y Gadow, 2003).
Distribución de frecuencias bivariante
Las curvas de altura del rodal descritas hasta ahora permiten obtener distinguir por
especies e incluso por rodales, pero dentro de una misma especie y rodal, asignan la
misma altura a dos árboles del mismo diámetro. Por lo tanto estas curvas no describen
la dispersión real de alturas. En muchas ocasiones es necesario contar con curvas de
altura que tengan en cuenta esta dispersión, como por ejemplo en la estimación de la
calidad de estación, para la descripción de la estructura del rodal en bosques naturales
o para poder realizar simulaciones realistas en modelos de árbol individual. Una
posible solución es el ajuste de una función de distribución de frecuencias bivariante
que analice conjuntamente las alturas y los diámetros del rodal. Schmidt y Gadow
(1999) emplearon la función bivariante SBB (Johnson, 1949) para estimar la
distribución conjunta de alturas y diámetros.

Estructura y Diversidad 157
En el caso de rodales semi-regulares o irregulares no es suficiente con emplear
una única función bivariante puesto que existen dos o más estratos con diferentes
relaciones altura-diámetro. En estos casos se pueden combinar varias funciones
bivariantes para caracterizar mejor al rodal. Por ejemplo, Zucchini et al. (2001)
emplearon dos distribuciones bivariantes normales para caracterizar la distribución
combinada altura-diámetro en rodales irregulares de haya. La distribución final para el
rodal completo f(d,h) se obtiene empleando la siguiente expresión:
),()1(),(),( 21 hdnhdnhdf ⋅−+⋅= αα 4-10
Donde n1(d,h) y n2(d,h) son las distribuciones bivariantes ajustadas a las alturas y
diámetros de cada uno de los dos estratos que se observaron en la muestra y α es un
parámetro que varía en el intervalo [0,1] y que define la proporción de la población
total que pertenece a cada uno de esos dos estratos. El valor de α obtenido al ajustar
este modelo a los datos obtenidos por Schmidt y Gadow (1999) fue de 0,19, indicando
que un 19% de la muestra pertenecía al primer estrato y un 81% al segundo estrato
considerado. La representación gráfica de la función de densidad así obtenida (Figura
4-9) muestra claramente que existen dos subpoblaciones (figura derecha), y que en la
subpoblación más pequeña del estrato inferior la pendiente de la relación altura-
diámetro (figura derecha) es mayor que en la otra subpoblación. La bondad del ajuste
de este modelo con dos distribuciones de densidad bivariantes mejoró notablemente la
del obtenido solamente con la función bivariante SBB.
El modelo es fácil de interpretar y reproduce de forma realista las dos
subpoblaciones o estratos que constituyen el rodal y que se diferencian en la relación
altura-diámetro. El principal problema para la aplicación de este método es el elevado
número de mediciones que se deben realizar (como mínimo 50 pares de valores altura-
diámetro) para poder ajustar un modelo que estime con la suficiente exactitud. Una
forma de poder reducir los costes de las mediciones sería no medir las alturas

Estructura y Diversidad 158
exactamente sino utilizar clases de altura de intervalos entre 3 y 10 m, aunque esta
reducción de costes se consigue a costa de una pérdida de exactitud del modelo.
Figura 4-9. Representación en perspectiva y contorno de la función de densidad conjunta de dos distribuciones bivariantes normales ajustada a los pares de valores de diámetros y alturas del bosque natural Dreyberg (Solling).
Otra alternativa para representar la variabilidad de las alturas para cada diámetro es el
empleo de modelos estocásticos (Castedo et al., 2005). Esta metodología se basa en
añadir a la curva altura-diámetro local o generalizada ajustada un término estocástico
obtenido a partir de una función de distribución de probabilidad. De este modo, la
expresión utilizada para asignar las alturas a cada árbol es:
( ) iyUiestoci sFyy ˆ1ˆˆ −+= 4-11
donde es la estimación estocástica de la altura, es la estimación obtenida
mediante la curva altura-diámetro ajustada, es el valor de la inversa de la función
de distribución normal estándar para la variable aleatoria U generada uniformemente
en el intervalo (0,1), y es el error estándar de la predicción de altura que se obtiene
del ajuste de la relación altura-diámetro.
(estociy ) iy
1−UF
iys ˆ
Estructura y Diversidad 159
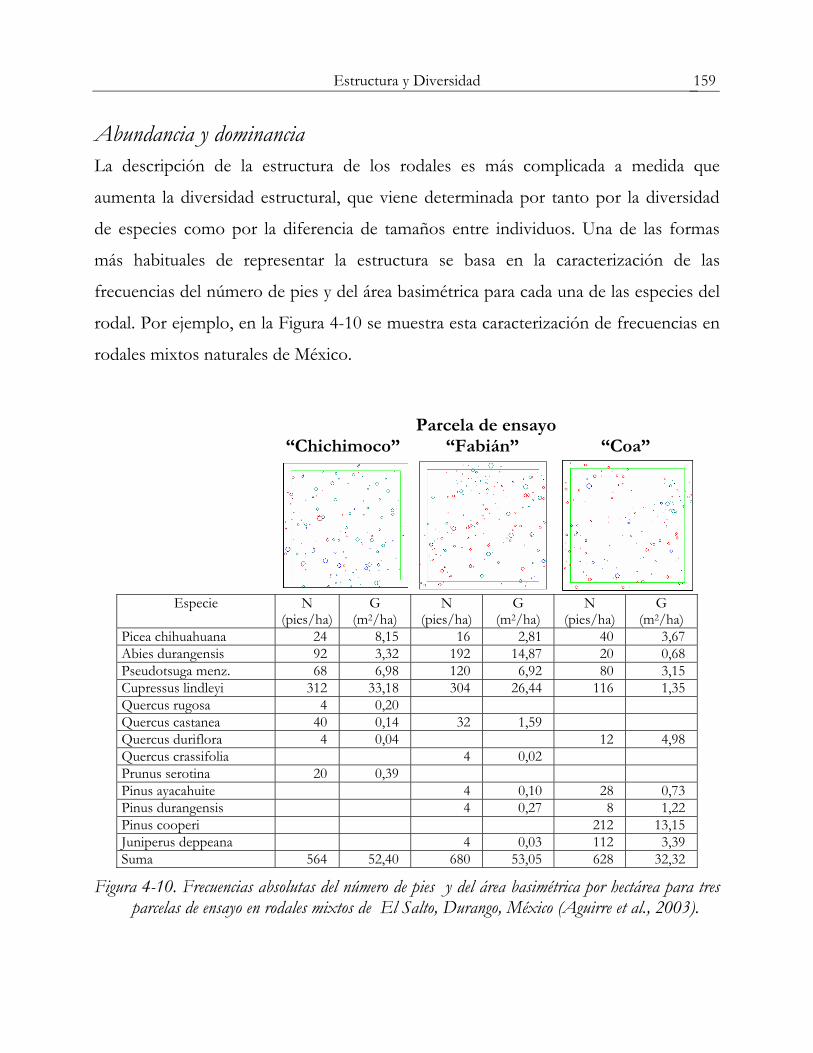
Abundancia y dominancia La descripción de la estructura de los rodales es más complicada a medida que
aumenta la diversidad estructural, que viene determinada por tanto por la diversidad
de especies como por la diferencia de tamaños entre individuos. Una de las formas
más habituales de representar la estructura se basa en la caracterización de las
frecuencias del número de pies y del área basimétrica para cada una de las especies del
rodal. Por ejemplo, en la Figura 4-10 se muestra esta caracterización de frecuencias en
rodales mixtos naturales de México.
Parcela de ensayo “Chichimoco” “Fabián” “Coa”
Especie N (pies/ha)
G (m2/ha)
N (pies/ha)
G (m2/ha)
N (pies/ha)
G (m2/ha)
Picea chihuahuana 24 8,15 16 2,81 40 3,67 Abies durangensis 92 3,32 192 14,87 20 0,68 Pseudotsuga menz. 68 6,98 120 6,92 80 3,15 Cupressus lindleyi 312 33,18 304 26,44 116 1,35 Quercus rugosa 4 0,20 Quercus castanea 40 0,14 32 1,59 Quercus duriflora 4 0,04 12 4,98 Quercus crassifolia 4 0,02 Prunus serotina 20 0,39 Pinus ayacahuite 4 0,10 28 0,73 Pinus durangensis 4 0,27 8 1,22 Pinus cooperi 212 13,15 Juniperus deppeana 4 0,03 112 3,39 Suma 564 52,40 680 53,05 628 32,32
Figura 4-10. Frecuencias absolutas del número de pies y del área basimétrica por hectárea para tres parcelas de ensayo en rodales mixtos de El Salto, Durango, México (Aguirre et al., 2003).

Estructura y Diversidad 160
A menudo se emplea el término abundancia como el valor de la frecuencia absoluta
del número de pies de una especie (Lamprecht, 1986). Por otro lado, el área
basimétrica es una variable dasométrica que aporta información no sólo de la
frecuencia de pies sino también de la dimensión de los mismos. Por esta razón,
también se suele emplear el término dominancia para referirse al valor del área
basimétrica absoluta de una especie en un rodal mixto. El producto de la abundancia y
dominancia relativa de una especie es lo que se conoce como importancia de la especie
dentro del rodal. En la Tabla 4-4 se muestran los valores de frecuencia relativa del
número de pies y del área basimétrica para la parcela de ensayo “Coa” de la figura 4-
10. Especie N% G% Importancia Picea chihuahuana 0,0637 0,1136 0,0072 Abies durangensis 0,0318 0,0210 0,0007 Pseudotsuga menziesii 0,1274 0,0975 0,0124 Cupressus lindleyi 0,1847 0,0418 0,0077 Quercus rugosa 0,0000 0,0000 0,0000 Quercus castanea 0,0000 0,0000 0,0000 Quercus duriflora 0,0191 0,1541 0,0029 Quercus crassifolia 0,0000 0,0000 0,0000 Prunus serotina 0,0000 0,0000 0,0000 Pinus ayacahuite 0,0446 0,0226 0,0010 Pinus durangensis 0,0127 0,0377 0,0005 Pinus cooperi 0,3376 0,4069 0,1374 Juniperus deppeana 0,1783 0,1049 0,0187 Suma 1,0000 1,0000 1,0000
Tabla 4-4. Frecuencia relativa del número de pies y de la proporción del área basimétrica para la parcela de ensayo “Coa” en un rodal mixto de El Salto, Durango, México (Aguirre et al., 2003).
Tal y como era de esperar los valores de importancia de las especies Pinus cooperi
(0,1374), Pseudotsuga menziesii (0,0124), Juniperus deppeana (0,0187), Cupressus lindleyi
(0,0077) y Picea chihuahuana (0,0072) son especialmente elevados.
Otra forma de caracterizar la estructura de los rodales es mediante el empleo de los
conceptos de riqueza de especies y riqueza de dimensiones. La riqueza de especies

Estructura y Diversidad 161
hace referencia a la cantidad de especies presentes en el rodal y se suele caracterizar
mediante el empleo de índices como el definido por Shannon y Weaver (1949):
( )∑=
−=S
iiiS pnp)p,....,p,p('H
121 l
4-12
donde S es el número de especies existentes y pi es la proporción de individuos de esa
especie, es decir Nnp ii = siendo ni el número de individuos de la especie i y N el
número total de individuos.
El índice de Shannon-Weaver cumple tres condiciones (Pielou, 1977): a) la
riqueza de especies alcanza su máximo valor cuando todas las especies están
representadas en la misma proporción ( ) )(),....,,(' 21 pnpnpppppH S ll −=−== ∑ , b) si
las especies se reparten de forma homogénea entre dos poblaciones, entonces la
población con el mayor número de especies es la de mayor riqueza y c) si se emplea
una clasificación adicional, por ejemplo la división en clases de altura, y ambas
clasificaciones son independientes, (es decir, el conocer la especie de un individuo no
tiene ninguna implicación sobre su altura), entonces la riqueza conjunta de especies y
alturas es igual a la suma de la riqueza de especies y la riqueza de alturas: H’(especie,
altura)=H’(especie)+H’(altura).
La riqueza de dimensiones se suele caracterizar mediante la distribución de
diámetros, de volumen, de longitud de copa o de superficie lateral de la copa. En la
figura 4-10 se muestran cuatro ejemplos de rodales con diferentes estructuras
caracterizados por su riqueza de especies y riqueza de dimensiones.

Estructura y Diversidad 162
baja alta Variable
Riqueza de
especies
Índice de
Shannon-
Weaver
Riqueza de
dimensiones
Distribución
diamétrica
Figura 4-11. Características de la estructura forestal sin considerar la distribución espacial de los atributos de los árboles. Diferentes colores hacen referencia a diferentes especies. Los tamaños de los círculos son proporcionales a los diámetros de cada árbol.
El concepto de estructura hace referencia a la distribución y el orden de los elementos
de un sistema. Por tanto, la estructura forestal debe describir la distribución y orden de
los atributos del árbol dentro de un rodal y para ello no basta con caracterizar las
dimensiones de los árboles o la presencia de diferentes especies, sino que también es
necesario tener en cuenta la distribución en el espacio de tales características. Por esta
razón, para la descripción completa de la estructura de un rodal es necesario emplear
tres grupos diferentes de variables o índices que tengan en cuenta las especies
presentes, sus dimensiones y su distribución sobre el terreno (ver figura 4-12).

Estructura y Diversidad 163
Estructura forestal
Diversidad deposición
Diversidad de especies
Diversidad de dimensiones
Índices independientes de la posición para caracterizar el rodal en su conjunto
Índices dependientes de la posición para caracterizar el rodal en su conjunto
Variables de vecindad para describir las diferencias de estructura a pequeña escala
Figura 4-12. Para la descripción de la estructura forestal se recomienda el empleo de tres grupos de variables o índices.
Los índices dependientes e independientes de la posición en el terreno de los
individuos se emplean para caracterizar al rodal en su conjunto. Para analizar las
diferencias estructurales a pequeña escala se adaptan los índices generales empleando
relaciones de vecindad (Albert y Gadow, 1998).
Parámetros de la estructura espacial Una forma de definir la estructura espacial consiste en el empleo de funciones de
correlación como las denominadas pair-correlation, mark correlation o mark connection
(Stoyan y Penttinen, 2000; Pommerening, 2002), aunque su uso es limitado puesto que
sólo se pueden emplear cuando se conozcan las distribuciones espaciales de los
árboles y sus coordenadas. Otro índice muy empleado es el índice de agregación de
Clark y Evans (1954), aunque también presenta limitaciones para caracterizar la
estructura de rodales en los que la distribución espacial de los árboles es muy irregular
(Albert, 1999), por lo que sólo pueden aportar una impresión general sobre la
estructura forestal (Zenner y Hibbs, 2000).

Estructura y Diversidad 164
Gadow (1999) y Pommerening (2002) consideran que la diversidad estructural de los
rodales forestales se puede caracterizar considerando tres factores: la agregación o
distribución espacial de los árboles (contagion), el grado de mezcla de las especies (species
mingling) y el grado de diferenciación espacial por dimensiones de los árboles (size
differentiation). En la figura 4-13 se muestran esquemáticamente los tres factores más
importantes de la estructura espacial de los rodales forestales: la agregación o grado de
regularidad en la distribución espacial, el grado de mezcla entre diferentes especies y el
grado de diferenciación en el espacio por tamaños.
baja alta
Agregación
(contagion)
Mezcla
(species mingling)
Diferenciación
(size differentiation)
Figura 4-13. Dos ejemplo de rodales con alta y baja diversidad en la distribución espacial de los árboles, de las especies y de sus dimensiones.
La distribución espacial de los árboles refleja si el patrón de disposición de los árboles
sobre el terreno es regular, aleatorio, se tienden a agrupar o es una mezcla de todas

Estructura y Diversidad 165
estas situaciones. El grado de mezcla analiza si los árboles de la misma especie tienden
a formar pequeños grupos o por el contrario existe una mezcla espacial de especies.
Por ultimo, el grado de diferenciación espacial por dimensiones tiene en cuenta si
árboles del mismo diámetro y/o altura tienden a agruparse o se distribuyen de forma
aleatoria. Estos autores consideran que el uso de un conjunto de índices que tengan en
cuenta estos tres factores puede ser suficiente para definir, e incluso permitir simular,
la estructura espacial de un rodal forestal.
Agregación: el índice Winkelmass
La distribución espacial de los árboles es uno de los factores que definen la estructura
del rodal. Uno de los índices empleados para medir y describir dicha distribución
espacial es el denominado Winkelmass. El índice Winkelmass requiere un tipo de
información que es posible obtener sin necesidad de realizar laboriosas mediciones de
distancias y además, permite representar los resultados en forma de distribuciones de
los valores para cada árbol o con el valor medio (Albert, 1999; Staupendahl, 2001). El
valor de este índice se calcula como:
∑=
=n
jiji v
nW
1
1 siendo y
⎩⎨⎧
=<
contrario casoen ,0 ,1 0αα
v jkij 1W0 i ≤≤ 4-13
Donde Wi describe la regularidad o la irregularidad de la distribución espacial de los n
árboles más próximos al árbol de referencia i. El proceso de obtención del valor de Wi
comienza seleccionando el árbol más próximo al árbol de referencia (árbol j), a
continuación se selecciona el árbol siguiente según el sentido de las agujas del reloj
(árbol k) y luego se mide desde el árbol de referencia el ángulo αjk que forman el árbol j
y el árbol k (ver figura 4-14). El ángulo αjk es siempre el más pequeño de los dos
ángulos que se forma entre los árboles i, j y k (αjk y βjk), por lo tanto αjk es siempre ≤
180° (figura 4-14)

Estructura y Diversidad 166
i
αjk < α0
j
k
βjk
Figura 4-14. Ángulo entre dos vecinos desde el árbol de referencia i. La suma de ambos ángulos (αjk y βjk) es de 360º.
El valor del ángulo αjk se compara con un ángulo de referencia o ángulo estándar α0
que es el ángulo esperado en una distribución regular. Si αjk es menor que el ángulo de
referencia se le asigna un 1 a la variable binaria vij y en caso contrario se le asigna un 0.
Si se consideran cuatro árboles vecinos se pueden obtener cinco valores diferentes del
índice Wi (figura 4-15).
i
α≥α0
i
α<α0
i
0αα <0αα <
i α α> 0
i
α α< 0
Wi = 0 Wi = 0,25 Wi = 0,5 Wi = 0,75 Wi = 1
Ninguno de los ángulos αij es menor que α0 (muy regular)
Uno de los ángulos αij es menor que α0 (regular)
Dos de los ángulos αij son menores que α0 (aleatorio)
Tres de los ángulos αij son menores que α0 (irregular)
Los cuatro ángulos α- j son menores que α0 (muy irregular)
Figura 4-15. Posibles valores del índice considerando cuatro vecinos y un ángulo estándar α0 =90º.
Si Wi=0 existe una regularidad local muy fuerte en torno al árbol de referencia,
mientras que si Wi=1, entonces la distribución a escala local es irregular o agregada.
Como estimador del índice para el rodal completo se emplea la media aritmética de los
valores obtenidos para todos los árboles del rodal (El valor del índice Winkelmass
referido a la superficie se calcula como una media ponderada obtenida a partir de
todos los valores Wi de los árboles de una cierta superficie, ver Staupendahl, 2001):

Estructura y Diversidad 167
∑=
=N
1iiW
N1W 4-14
Siendo Wi el valor del índice para el i-ésimo árbol de referencia y N el número de
árboles de árboles de referencia.
El parámetro clave para la asignación de un rodal a un tipo de distribución
espacial determinada es el ángulo estándar utilizado (α0). El ángulo escogido como
referencia define el valor del índice para cada árbol y, por tanto, también el valor
medio del índice, aunque, sea cual sea el valor de α0 siempre es válida la siguiente
relación agregadoaleatorioregular WWW << . Originalmente, se consideró como valor del ángulo
estándar el cociente α0 =360º/n (Gadow et al., 1998). Sin embargo, la mejor manera de
determinar el valor más adecuado del ángulo estándar es emplear datos de
distribuciones espaciales regulares, aleatorias y agregadas de los árboles en rodales
forestales y clasificarlas en función del índice calculado con diferentes valores del
ángulo estándar. Hui y Gadow (2002) estimaron que el mejor valor para este ángulo
estándar es de 72º. La figura 4-16 muestra las distribuciones de los valores del índice
Winkelmass para 2000 rodales simulados con tres tipos diferentes de distribuciones de
árboles empleando un ángulo estándar de 72º. Se observa que se pueden clasificar las
distribuciones espaciales de los árboles en los rodales forestales en tres grupos (regular,
aleatorio y agregado) sin que exista un gran solape entre categorías.
0
100
200
300
400
0.45 0.46 0.47 0.49 0.50 0.52 0.53 0.54 0.56
Valor medio del Winkelmass
Fre
c. r
elat
iva
aleatorio regular agregado
Figura 4-16. Distribuciones de los valores medios del índice para 2000 rodales simulados para tres tipos diferentes de distribuciones empleando un ángulo estándar de 72º.

Estructura y Diversidad 168
El índice Winkelmass también se puede emplear para estimar la distancia entre árboles
vecinos sin necesidad de realizar costosas mediciones en campo (Gadow et al., 2003).
Esta distancia es la base para el empleo de modelos de crecimiento de árbol individual
dependientes de la distancia, también es imprescindible para poder simular o
reproducir la estructura espacial del rodal y para la visualización del desarrollo de dicho
rodal.
La distribución de las distancias entre árboles se puede describir mediante una
función de distribución de probabilidad, como por ejemplo la función de Weibull de
dos parámetros:
c
bx
e-1=F(x)⎟⎠⎞
⎜⎝⎛
4-15
Donde x es la distancia al vecino más próximo y b y c son los parámetros de escala y
forma, respectivamente.
Conocidos los valores de b y c se puede reconstruir la distribución de distancias
entre árboles empleando la ecuación 4-15. Estos parámetros (b y c) están directamente
relacionados con el valor medio de la distancia entre árboles y con la desviación típica
de esas distancias. En la figura 4-18 se muestra un ejemplo de la relación entre b y la
distancia media al primer vecino (D01) y c y la desviación típica de las distancias al
primer vecino (S01).

Estructura y Diversidad 169
0.00 0.50 1.00 1.50 2.00 2.50 3.00
0.00 1.00 2.00 3.00
D01
b
0.00
1.00
2.00
3.004.00
5.006.00
7.008.00
0.00 2.00 4.00 6.00 8.00
D01/S01
c
Figura 4-18. Relaciones lineales entre los parámetros b y c de la función Weibull y la distancia media al primer vecino (D01) o el cociente entre (D01) y la desviación típica de las distancias al primer vecino (s01).
Como se observa en la figura anterior, las relaciones entre b, c y D01 o D01/S01 son
lineales siendo para este ejemplo: b =-0,0277 + 1,129 (D01) y c =-0,3215 + 1,2065
(D01/S01).
Por otro lado, el valor medio y la desviación típica de las distancias se pueden
relacionar con el valor del índice Winkelmass y la densidad del rodal (Gadow et al.,
2003):
28308250804101
.-W.T
.- DWD ⋅⋅= 0.260 4-16
521001
1770694101
4131 .W.T
. DDWS.-⋅⋅= 1.672 4-17
Donde D01 es el valor medio de la distancia la vecino más próximo (m); S01 es la
desviación típica de la distancia media al vecino más próximo (m); W es el valor
medio del índice; DT es la distancia esperada para una distribución aleatoria (m), es
decir )10000/(5.0 NDT = y N es la densidad del rodal (número de pies por ha).

Estructura y Diversidad 170
De este modo, sustituyendo los valores de las ecuaciones 4-16 y 4-17, en las
relaciones lineales de los parámetros b y c de la función de Weibull se puede
reconstruir la distribución de frecuencias de distancias entre árboles del rodal.
El valor del índice Winkelmass se puede relacionar con otro índice de agregación
muy utilizado, el índice de Clark y Evans (RCE). Si se reorganizan los términos de la
ecuación (4-16) se llega a la siguiente expresión matemática que relaciona los dos
índices y la densidad del rodal para el ejemplo analizado:
( ) 1825.0804.101283.0
10000/5.0260.0−⋅−
−
⋅⋅==W
TCE NW
DDR 4-18
Mezcla de especies
La mezcla de especies es una medida de la segregación espacial de los individuos de
diferentes especies y se puede describir mediante el índice denominado grado de
mezcla Mi. Füldner (1995) definió el valor del índice del árbol de referencia i como la
proporción de vecinos que pertenecen a especies diferentes a dicho árbol de
referencia:
∑=
=n
jji v
n1M
1 4-19
siendo y vj es igual a 0 si el árbol j es de la misma especie que el árbol de
referencia i y 1 en caso contrario. De este modo, si se analizan los n vecinos más
próximos el grado de mezcla puede tomar n+1 valores discretos.
10 ≤≤ iM
En el ejemplo de la figura 4-19 se muestra la distribución de los valores del grado de
mezcla de especies para haya y fresno en un rodal mixto de haya y otras frondosas
considerando los 3 vecinos más cercanos.

Estructura y Diversidad 171
Haya Fresno
Prop
orci
ón
0,00
0,20
0,40
0,60
0,00 0,33 0,67 1,00
M
Anteil
0,00
0,20
0,40
0,60
0,00 0,33 0,67 1,00
M Figura 4-19. Representación de los valores de grado de mezcla de especies en un rodal mixto de haya y
otras frondosas. A la izquierda la distribución de valores obtenida para haya y a la derecha para fresno.
La mayoría de las hayas están rodeadas por árboles de la misma especie (Mi=0)
mientras que los fresnos aparecen solitarios entre las hayas (Mi=1) y solo existe una
pequeña proporción de fresnos con tendencia a agruparse (Mi<1). En determinadas
ocasiones también puede resultar de interés para extraer conclusiones sobre la
distribución espacial de las especies el analizar la relación que existe entre el grado de
mezcla de una especie y su proporción dentro del rodal Mp , siendo p la proporción
del número de pies o del área basimétrica de la especie analizada y M el valor medio
del grado de mezcla de la especie. Otras posibles aplicaciones del grado de mezcla de
especies se pueden encontrar en el trabajo de Aguirre et al. (2003).
Diferenciación espacial por dimensiones
La diferenciación de dimensiones es una medida de la proximidad espacial que existe
entre árboles de diferentes dimensiones en un rodal. Una medida simple para la
dispersión de dimensiones entre árboles, considerando como variable representativa
de la dimensión el diámetro normal es el coeficiente de variación en el entorno del
árbol de referencia i:
100⋅j
jiCV
d=%
σ 4-20

Estructura y Diversidad 172
Siendo σj y jd la desviación típica y el diámetro medio de los j (j = 1,…n) árboles
vecinos del árbol de referencia i.
Otra forma de describir la diferenciación espacial por dimensiones es el empleo
de índices como los denominados grado de diferenciación (Füldner, 1995), medida del
entorno (Hui et al., 1998) o dominancia de dimensiones (Albert, 1999).
El grado de diferenciación es un índice que se puede aplicar a cualquier variable
que represente el tamaño de un árbol. Por ejemplo, si se consideran los diámetros su
valor es:
∑=
−=n
1ji )max(
)min(n11
ji
ji
,dd,dd
T 4-21
Donde Ti es el grado de diferenciación en diámetros considerando los n árboles más
cercanos al árbol de referencia i, 0 ≤ Ti ≤ 1; di es el diámetro normal del árbol de
referencia y dj es el diámetro normal del árbol j (j = 1...n).
El valor de Ti se incrementa al aumentar la diferencia media de los tamaños de
los árboles cercanos. Un valor de cero se corresponde con la situación en la que todos
los árboles tienen el mismo tamaño. La sencillez del índice y el hecho de contar con un
valor de referencia (cero) para comparar diferentes situaciones aconseja su empleo
para describir la estructura de una masa desde el punto de vista de las dimensiones de
los árboles que la constituyen.
El índice denominado medida del entorno describe la dominancia relativa de una
especie en su entorno directo. Este índice se define como la proporción de los n
vecinos más próximos al árbol de referencia que son de menor tamaño que él:
∑=
n
jjv
1n1=Ui 4-22

Estructura y Diversidad 173
siendo y vj es igual a 1 si el árbol j es menor que el árbol de referencia i y 0 en
caso contrario. Considerando cuatro vecinos Ui puede tomar cinco valores. Para el
cálculo de este índice se puede emplear como dimensión el diámetro normal, la altura,
o cualquier otra variable de interés.
10 ≤≤ iU
En la figura 4-20 se muestra la distribución de la medida del entorno de tres
especies en la parcela de ensayo Chichimoco en El Salto, Durango, México (Aguirre et
al., 2003).
Cupressus lindleyi 3.0=U
Abies durangensis 26.0=U
Pseudotsuga menziesii 65.0=U
0
0,1
0,2
0,3
0,4
0,5
0 0,25 0,5 0,75 1
U
p
0
0.05
0.1
0.15
0.2
0.25
0.3
0.35
0.4
0.45
0 0.25 0.5 0.75 1
U
p
0
0,1
0,2
0,3
0,4
0,5
0 0,25 0,5 0,75 1
U
p
Figura 4-20. Dominancia específica de tres especies en la parcela de ensayo Chichimoco en El Salto, México. U es el valor medio de la medida del entorno de la especie.
La distribución de la medida del entorno de la especie Cupressus lindleyi está desplazada
hacia la izquierda, lo que indica que pocos de los árboles analizados son dominantes en
su entorno local. Una interpretación similar puede hacerse para la especie Abies
durangensis aunque con una mayor dominancia que la especie anterior. Por el contrario,
la especie es frecuentemente dominante en su entorno local. Cada una de estas tres
especies tiene árboles dominantes, codominantes y dominados en su entorno, aunque
las proporciones son muy diferentes en cada caso. .
Albert (1999, página 51 y siguientes) empleó la dominancia de dimensiones, que
es un índice que considera tanto la posición relativa del árbol en relación a sus vecinos
(medida del entorno) como la información cuantitativa de la diferencia de tamaño
(grado de diferenciación).

Estructura y Diversidad 174
Estructuras espaciales esperadas y observadas
Todos los índices que se han analizado en los apartados anteriores para describir la
estructura del rodal a pequeña escala son muy fáciles de calcular y se basan en variables
que son fáciles de medir sobre el terreno sin costosos inventarios. Por ejemplo, la
figura 4-21 muestra un grupo de 5 árboles (el árbol de referencia i y sus cuatro vecinos
más próximos) con sus correspondientes diámetros.
i
α<α0
20 cm
30 cm
10 cm
50 cm
20 cm
Figura 4-21. Las variables para describir la estructura espacial son fáciles de medir sobre el terreno.
Se miden el árbol de referencia más próximo respecto al punto de referencia y sus n vecinos más próximos
Los diferentes colores de los árboles indican especies distintas. Sólo un ángulo α es
menor que el ángulo estándar entre dos vecinos α0 (Cómo ya se comentó
anteriormente, el valor más adecuado para α0 cuando se consideran cuatro vecinos es
de 72º). Para el grupo de cinco árboles presentado en la figura 4-21 se obtienen los
siguientes valores del índice Winkelmass (Wi), del grado de mezcla (Mi); del
coeficiente de variación local de los diámetros (CVi%; el diámetro medio es de
110/4=27.5 cm.); del grado de diferenciación de diámetros (Ti) y de la medida del
entorno (Ui) para el árbol de referencia i:

Estructura y Diversidad 175
25,04
1000=
+++=iW ; 50,0
41100
=+++
=iM ;
%651005,27
)5,2710()5,2720()5,2730()5,2750(%
2222
=⋅−+−+−+−
=iCV ;
36,05020
2020
3020
2010
411 =⎟
⎠⎞
⎜⎝⎛ +++⋅−=iT ; 25,0
40001
=+++
=iU
La trasformación a valores por unidad de superficie (hectárea) de los resultados
obtenidos con estos índices a escala local se basa en la probabilidad de selección de un
árbol como árbol de referencia, y esa probabilidad depende de la superficie topológica
o superficie de terreno que, teóricamente, ocupa cada árbol (Albert, 1999, página 78 y
siguientes; Staupendahl, 1997).
Los parámetros estructurales facilitan el análisis de los estados de desarrollo de
los rodales y de sus cambios como consecuencia de la realización de tratamientos
selvícolas. Dos rodales con el mismo número de pies, la misma distribución de
especies y la misma distribución diamétrica pueden presentar estructuras muy
diferentes. Esas diferencias pueden ser observadas mediante los valores de los índices
Winkelmass, grado de mezcla de especies y diferenciación de dimensiones (Füldner,
1995; Schmidt et al., 1997; Pommerening, 1997). Por lo tanto, es posible comparar los
rodales entre sí, pero cuando se pretende conocer la situación de un rodal concreto es
necesario compararlo con un estándar absoluto, es decir conviene establecer una
referencia de comparación. Una posible referencia de comparación es el valor
esperado de los índices obtenido a partir de simulaciones. Por ejemplo, a partir de los
diámetros observados en un rodal real con N pies/ha, en el que se conocen las
coordenadas espaciales de los árboles, se pueden generar N! rodales simulados, es
decir, N! permutaciones o formas diferentes de asignar los diámetros a cada
coordenada. El análisis de los valores de los índices estructurales en estos rodales
simulados permite establecer un valor de referencia para realizar comparaciones (ver,

Estructura y Diversidad 176
por ejemplo los trabajos de Lewandowski y Pommerening, 1996; Pommerening 1997,
página 15 y siguientes o Schröder, 1998).
En la figura 4-22 se muestran los valores observado y esperado del índice grado
de diferenciación de diámetros en un bosque natural de Sudáfrica (Schröder, 1998).
Los valores esperados se obtuvieron mediante la generación de 1000 rodales y se
observa que hay una gran coincidencia entre ambos valores tanto por especie como
para el conjunto del rodal.
0
0,2
0,4
Diferenciación mediade diámetros
Oleacapensis
Podocarpuslatifolius
Todas lasespecies
Goniomakamassi
Figura 4-22. Valores del grado de diferenciación en diámetros observada (columna de la derecha) y esperada (columna de la izquierda en azul) para tres especies y para todo el conjunto de especies en un bosque natural de Sudáfrica considerando para el cálculo del índice sólo el árbol vecino más cercano (Schröder, 1998).
El empleo de rodales simulados permite calcular el valor de cualquier índice estructural
para cada uno de ellos y, por tanto, conocer no sólo la media sino también la
distribución de dichos valores y su variabilidad. Esto permite que al analizar el valor
del índice en un rodal real se pueda asignar una probabilidad de ocurrencia al valor real
observado.

Crecimiento del Rodal 177
Crecimiento del Rodal
Un árbol necesita para su crecimiento luz, dióxido de carbono, agua y nutrientes
minerales. Todas estas sustancias elementales se transforman en moléculas
complejas como resultado de reacciones químicas específicas. Por lo tanto, el
crecimiento del árbol es más que un mero incremento del diámetro y de la altura.
En el árbol se producen numerosos procesos biológicos: las células germinales se
van diferenciando en diferentes tipos de células que forman tejidos por agregación
y estos a su vez constituyen diferentes órganos. Gracias a estos procesos el árbol
tiene la capacidad, dentro de determinados límites fijados por su amplitud
ecológica, de reaccionar ante cambios en su entorno y adaptarse para sobrevivir.
Esta capacidad se pone de manifiesto en la dinámica del crecimiento del árbol
(Mitscherlich, 1971, 1975; Raven et al., 1987).

178 Crecimiento del Rodal
Teóricamente es posible pronosticar el estado de un sistema biológico al final de un
corto periodo de tiempo si se conocen su estado inicial y el proceso de desarrollo
del sistema. Ese nuevo estado del sistema sirve a su vez como punto de partida
para el siguiente período de tiempo y así sucesivamente. El resultado de uno de
estos pronósticos depende de la bondad de la estimación del modelo y de la
exactitud con la que puede ser descrita la situación inicial. La determinación de
ambos valores es casi imposible de realizar en un sistema macroscópico como es el
árbol. Sin embargo, es posible utilizar modelos operacionales en los que el mundo
real se simplifica, reduciéndolo a unas variables de estado escogidas que se
considera que contienen la información mínima para poder simular el desarrollo del
sistema. La modelización del crecimiento forestal es un intento de construir ese
tipo de modelos operacionales basados en la comprensión de los procesos reales
que acontecen en los árboles o en los rodales.
Los modelos forestales de crecimiento proporcionan una perspectiva sobre las
complejas interacciones entre las estructuras y los procesos en los ecosistemas
forestales y posibilitan una mejor comprensión de la dinámica forestal natural
(Pretzsch, 2001). En el campo de la investigación los modelos de crecimiento
sirven para simular experimentos, que no sería posible realizar en la realidad debido
a los largos períodos de observación necesarios. En el campo de la formación y la
consultoría los modelos de crecimiento proporcionan información sobre las
consecuencias económicas y ecológicas de una determinada decisión. En la gestión
práctica los modelos de crecimiento son herramientas imprescindibles de apoyo a la
toma de decisiones para un rodal, un monte o incluso a escala regional.
Los modelos de crecimiento para la práctica forestal se basan en la experiencia
acumulada del desarrollo de rodales y/o árboles. El grado de agregación o de escala
es muy diferente para los distintos tipos de modelos forestales. Por ejemplo, los
modelos orientados a la fisiología del árbol analizan detalles morfológicos como el
ángulo de las ramas, la forma del tronco o el crecimiento de las raíces. Los modelos

Crecimiento del Rodal 179
agregados suministran información sobre el desarrollo de una población bajo unas
condiciones determinadas y sus reacciones específicas a cambios de la estructura
del rodal debidos a la influencia humana y/o a fenómenos naturales. Una finalidad
de la modelización del crecimiento es generar un sistema que pueda aclarar los
procesos básicos de crecimiento y que al mismo tiempo pueda generar como
resultado adicional información sobre la producción que sea relevante desde el
punto de vista económico.
La mayoría de los modelos se conciben para fines concretos y se suelen
diferenciar en función de su ámbito de validez, de su posible aplicación para una
serie de situaciones determinadas, de su exactitud o de su error de estimación
(Sharpe, 1990). Una elevada exactitud se consigue a costa de una validez reducida
por lo tanto la elección de un modelo adecuado suele requerir un compromiso
entre estos dos condicionantes.
Obtención de datos Un aprovechamiento forestal adecuado requiere una comprensión de los procesos
biológicos del sistema. En términos de modelización forestal se podría decir que el
conocimiento de las tasas de crecimiento, relevantes desde el punto de vista
biológico, desde un estado inicial determinado constituye la base para esa
comprensión. Estos conocimientos nos permiten crear programas informáticos con
cuya ayuda los gestores forestales pueden juzgar los efectos de diferentes
tratamientos selvícolas. Una de las herramientas más importantes son los modelos
de crecimiento creados a partir de datos empíricos, es decir, de observaciones
reales.
La base de datos empírica para estos modelos procede en su mayor parte de
parcelas de ensayo. Frecuentemente se trata de ensayos de procedencias, ensayos de
densidades o ensayos de claras cuyas finalidades son diferentes. En los ensayos de
procedencias se analiza la adecuación o el crecimiento de una especie o una

180 Crecimiento del Rodal
procedencia en una estación determinada. En los ensayos de densidades y de claras
se analizan los efectos de diferentes marcos de plantación y diferentes intensidades
de clara sobre el crecimiento de los árboles individuales y del rodal. Todas estas
parcelas de ensayo suelen ser mantenidas por centros de investigación estatales,
universidades o instituciones privadas de investigación.
La financiación privada de la investigación forestal se da en países donde
existe un aprovechamiento forestal y maderero que produzca elevados
rendimientos, como por ejemplo Australia, Sudáfrica, Nueva Zelanda, Chile, los
países escandinavos o USA. Un ejemplo de investigación forestal cooperativa con
la participación de Universidades y Empresas son las denominadas “Research
Cooperatives”, como por ejemplo la de la Universidad de Georgia (“Plantation
Management Research Cooperative”) o la del Instituto Politécnico de Virginia (“Loblolly
Pine Growth and Yield Research Cooperative”). Los institutos públicos de investigación
forestal son más frecuentes en Europa, como por ejemplo el INIA en España, el
INRA en Francia, el METLA en Finlandia, el WSL en Suiza, el Instituto de
Investigación Forestal en Austria y los Institutos de Investigación de los Estados
Alemanes.
Los limitados medios de investigación y la complejidad creciente de las
cuestiones que se plantean obligan a una reflexión continua sobre el
establecimiento de parcelas de ensayo. Según la duración del establecimiento de las
parcelas de ensayo se pueden distinguir entre: parcelas permanentes, parcelas
temporales y parcelas de intervalo. Las parcelas permanentes se observan durante
un período de tiempo largo y se miden en intervalos de tiempo regular. Las parcelas
temporales son parcelas que se miden una única vez y se distribuyen en un amplio
espectro de edades y calidades de estación. Estas parcelas evitan el largo tiempo de
espera para la obtención de información característico de las parcelas permanentes.
Las parcelas de intervalo son un compromiso entre ambos tipos de parcelas. Estas

Crecimiento del Rodal 181
parcelas se distribuyen en un amplio espectro de estados iniciales como las parcelas
temporales, pero la diferencia es que se miden como mínimo dos veces.
Parcelas permanentes
Nuestros conocimientos actuales sobre el desarrollo de diferentes ecosistemas
forestales se basan en gran parte en datos procedentes de parcelas permanentes que
han sido medidas repetidamente durante un largo período de tiempo. Los datos de
las parcelas permanentes constituyen una base importante para el desarrollo de
modelos para una amplia variedad de tratamientos selvícolas. Un ejemplo sencillo
del empleo de este tipo de información son las primeras tablas de producción
construidas para caracterizar el crecimiento con programas de claras estándar.
La figura 5-1 muestra una parcela permanente con tres mediciones de altura
consecutivas a las edades t1, t2 y t3.
tt 3 t 2 t 1
0
5
1 0
1 5
2 0
2 5
3 0
3 5
4 0
0 2 0 4 0 6 0 8 0 1 0 0 1 2 0
H [m ]
P a r c e la 1 P a r c e la 2 P a r c e la 3
t
Figura 5-1. Izquierda: una parcela permanente con tres mediciones sucesivas de altura consecutivas (t indica el eje temporal, los árboles en color blanco fueron cortados durante las claras). Derecha: un ejemplo de series de datos de altura dominante procedentes de tres parcelas permanentes.
Un modelo empleado frecuentemente para describir el desarrollo de la altura dominante con la edad es la función de Chapman-Richards:
( ) 2110ataeaH ⋅−−⋅= 5-1
Donde H es la altura dominante, t es la edad del rodal y a0, a1 y a2 son los parámetros empíricos del modelo.

182 Crecimiento del Rodal
Los datos de parcelas permanentes tienen la ventaja de que permiten describir con
facilidad crecimientos polimórficos, es decir, curvas de evolución de una variables
dasométrica o dendrométrica (en este caso la altura) que no son proporcionales
entre sí. En el caso de la ecuación 5-1, el modelo es polimórfico cuando alguno de
los parámetros que determinan la forma de la curva a1 y a2 varía en función de la
calidad de la estación, ya sea representada por el índice de sitio o por valores de
complejos factores de la estación como la temperatura, la humedad o el contenido
de nutrientes (Kahn, 1994). Ejemplos de estos modelos se encuentran en los
trabajos de Jansen et al. (1996). Para cada parcela se puede desarrollar una curva
diferente de evolución de altura, de este modo es posible desarrollar curvas de
altura polimórficas que pueden llegar a cruzarse (“non-disjoint”) (Clutter et al., 1983).
Numerosas tablas de producción se han elaborado a partir de datos
procedentes de parcelas permanentes (ver por ejemplo Schober, 1987; Jansen et al.,
1996 o Rojo y Montero, 1996). También modelos de árbol individual como por
ejemplo BWin (Nagel, 1994), Silva (Pretzsch, 1992), Prognaus (Sterba y Monserud,
1997), y Moses (Hasenauer et al., 1995) se basan en datos de parcelas permanentes.
Ya que en el futuro los medios de investigación serán también limitados, es de
prever que no será posible cubrir la gran diversidad de estados iniciales posibles y
de variantes de clara sólo con datos procedentes de parcelas permanentes. Sin
embargo, las parcelas permanentes seguirán siendo imprescindibles para resolver
ciertas cuestiones aunque haya que esperar mucho tiempo para obtener datos útiles
para la construcción de modelos y aunque, a veces, no se pueda alcanzar el objetivo
marcado por la perdida de las antes de tiempo debido a circunstancias imprevistas.
Parcelas temporales
Una solución rápida cuando no se dispone de ningún dato sobre el desarrollo
forestal es la implantación de parcelas temporales. Una parcela temporal sólo se
mide una vez, por lo tanto para obtener información relevante empleando este tipo

Crecimiento del Rodal 183
de parcelas se requiere que, en su conjunto, se cubra un amplio espectro de estados
de desarrollo y calidades de estación. De este modo se sustituye la medición
sucesiva en el tiempo propia de las parcelas permanentes por una distribución
espacial de parcelas que se miden una única vez.
Este método ya se empleó en el siglo XIX (Assmann, 1953; Kramer, 1988,
página 97; Wenk et al., 1990 página 116; Sterba, 1991). Por ejemplo, en el
denominado método de franjas se tomaron datos de numerosas parcelas de
densidad normal de diferentes edades para elaborar tablas de producción (Baur,
1877). En el denominado método del indicador de Hartig (1868) se reconstruyó el
crecimiento de árboles individuales mediante análisis de tronco y así se pudo
obtener rápidamente información sobre el crecimiento.
El principio se representa de manera esquemática en la figura 5-2. Las parcelas
de diferentes edades se separan mediante líneas verticales. El eje x describe la
posición de los árboles. El símbolo t indica el eje temporal. Los datos procedentes
de parcelas temporales desempeñan un papel muy importante en la construcción de
modelos forestales, especialmente en aquellos casos en los que no se dispone de
datos de parcelas permanentes (ver por ejemplo Lee, 1993 o Biber, 1996).
Naturalmente, otra posible forma de obtener datos de manera rápida es realizar un
análisis de tronco que permite reconstruir el desarrollo de una variable de estado
(como por ejemplo la altura dominante). Sin embargo, este método tiene el
inconveniente de que la reconstrucción de la constelación de los árboles de un
rodal en el pasado lleva consigo errores. Por ejemplo, en la parcela 3 representada
en la figura 5-2 no se dispone de información sobre el árbol competidor con el
signo de interrogación, únicamente se conoce su existencia por el tocón.

184 Crecimiento del Rodal
?
t
x
1
2
3
+
0
10
20
30
40
0 20 40 60 80 100 120 140
t
H[m]
+
+
+
+
+
+
++
+ ++ +
+
+ ++ +
+ ++
+
+ ++ +
++ +
+
++
++
++
Figura 5-2. Izquierda: tres parcelas temporales de diferentes edades; el eje x describe la posición de los árboles; t define el eje temporal (ver Biber, 1996, página 27). Derecha: Datos de edades y alturas procedentes de parcelas temporales.
En la segunda mitad del siglo XX se construyeron numerosas tablas de producción
en base a datos de parcelas temporales, por ejemplo los modelos de Hamilton y
Christie (1971) o Madrigal et al. (1992). Estas tablas de producción únicamente
reflejan el desarrollo forestal de alternativas selvícolas estándar y no se pueden
emplear para simular diferentes opciones de tratamiento selvícola (Alder, 1980).
La limitación del método de franjas es que con una única medición no se
puede obtener datos de tasas de crecimiento, por ejemplo, en la figura anterior se
obtienen datos independientes de edades y alturas dominantes. Con esta
información no es posible desarrollar un modelo que permita proyectar hacia el
futuro el estado de una masa partiendo de un estado inicial definido por unas
variables de estado. En el caso de los modelos de árbol, la información de las
parcelas temporales se puede asociar con análisis de tronco y emplear para simular
el crecimiento de árboles individuales, sin embargo, la reconstrucción de la
competencia es muy insegura porque no se conoce el entorno directo del árbol
analizado. Además, los análisis de tronco tampoco permiten calcular determinadas
tasas de crecimiento como por ejemplo los cambios en la copa.
Aunque las parcelas temporales no son las más adecuadas para la
modelización forestal pueden proporcionar información muy valiosa sobre las
relaciones entre diferentes variables de crecimiento. Por ejemplo, en la tabla 5-1 se

Crecimiento del Rodal 185
muestran las relaciones entre el contenido en biomasa y nutrientes de rodales de
pino de diferentes edades (Rademacher, 2002). Rodal de pino de 25 años (Fuhrberg)
Fracciones BM (t/ha) N (kg/ha) P (kg/ha) K (kg/ha) Ca (kg/ha) Mg (kg/ha) Acículas 3,2 44,7 4,4 14,8 7,3 2,3 Ramas+ramillas+punta<7cm 31,1 79,3 6,6 36,2 42,7 12,2 Corteza>7cm 3,3 11,1 1,1 3,5 21,3 1,1 Tronco>7cm 16,3 14,9 0,5 10,6 30,3 3,3 Tocón 2,2 2,9 0,2 1,6 5,7 0,5 Raíces gruesas 4,9 7,1 0,9 5,4 3,7 1,3 Raíces finas 4,1 24,1 2,7 7,2 3,4 1,3 Biomasa total 65,0 184,1 16,2 79,2 114,5 22,0 Rodal de pino de 46 años
Fracciones BM (t/ha) N (kg/ha) P (kg/ha) K (kg/ha) Ca (kg/ha) Mg (kg/ha) Acículas 6,5 99,2 8,9 35,6 19,9 5,4 Ramas+ramillas+punta<7cm 27,5 126,2 10,6 46,7 99,2 14,3 Corteza>7cm 8,0 22,8 2,2 9,9 47,9 3,7 Tronco>7cm 77,5 68,2 2,1 22,7 54,8 14,6 Tocón 9,5 10,1 0,5 3,6 11,4 2,0 Raíces gruesas 18,9 32,6 3,6 21,8 14,4 4,7 Raíces finas 4,1 24,1 2,7 7,2 3,4 1,3 Biomasa total 152,0 383,2 30,5 147,4 251,1 46,1 Rodal de pino de 115 años
Fracciones BM (t/ha) N (kg/ha) P (kg/ha) K (kg/ha) Ca (kg/ha) Mg (kg/ha) Acículas 2,9 50,0 3,3 12,6 9,2 2,1 Ramas+ramillas+punta<7cm 15,2 55,9 3,6 15,4 30,1 5,0 Corteza>7cm 7,9 31,7 2,4 10,6 66,2 3,8 Tronco>7cm 104,3 47,9 2,7 27,3 71,4 16,7 Tocón 11,9 8,2 0,5 3,9 14,3 2,1 Raíces gruesas 22,2 39,7 2,3 19,0 18,8 5,4 Raíces finas 4,1 24,1 2,6 1,2 3,4 1,3 Biomasa total 168,4 257,3 17,5 96,1 213,3 36,4
Tabla 5-1. Distribución de la biomasa y de los elementos químicos en rodales de pino de diferentes edades (Rademacher, 2002).
El análisis de los datos de la tabla anterior procedentes de rodales de diferentes
edades proporciona una visión de conjunto del reparto y la evolución de la biomasa
y los nutrientes en las diferentes partes del árbol. En base a estos datos se pueden
desarrollar relaciones útiles entre la edad del rodal, la proporción de biomasa y el
contenido en elementos químicos (Figura 5-3).

186 Crecimiento del Rodal
mg(K)/g = -0,2035Ln(age) + 1,28
mg(Mg)/g = -0,059Ln(age) + 0,42
mg(P)/g = -0,026Ln(age) + 0,16
0,0
0,2
0,4
0,6
0,8
1,0
0 50 100
Edad (años)
mg
elem
ento
por
g b
iom
asa
150
0,00
0,20
0,40
0,60
0,80
1,00
0 50 100 150 200
Edad
Prop
orci
ón d
e bi
omas
a
Madera
Ramas
Hojas
Figura 5-3. Izquierda: relaciones entre la edad del rodal y el contenido de elementos químicos.
Derecha: proporción de biomasa de las diferentes partes del árbol.
Debido a la laboriosad de la toma y el análisis de datos de biomasa y nutrientes, este
tipo de estudios se realizan muy raramente, de ahí que los resultados sean
especialmente valiosos.
Parcelas de intervalo
Las parcelas de intervalo constituyen un compromiso entre los dos tipos anteriores.
Estas parcelas se miden como mínimo dos veces, al principio y al final de un
intervalo de tiempo durante el que no se realizan intervenciones humanas ni existe
otro tipo de perturbaciones. El intervalo de tiempo entre las dos mediciones
consecutivas debe ser lo suficientemente largo para suavizar el efecto que pueden
tener las oscilaciones climatológicas. Las características principales del concepto de
las parcelas de intervalo se presentan en la figura 5-4.
Las parcelas de intervalo tienen algunas de las ventajas de las parcelas
temporales puesto que permiten abarcar un amplio espectro de estados iniciales y
además los datos están disponibles en un corto período de tiempo; y a su vez tienen
algunas de las ventajas de las parcelas permanentes ya que permiten obtener tasas
de crecimiento para los diferentes estados iniciales, como por ejemplo el
crecimiento de la copa. Como ya se ha comentado, el intervalo entre las dos
mediciones debe coincidir con un período de tiempo de crecimiento sin
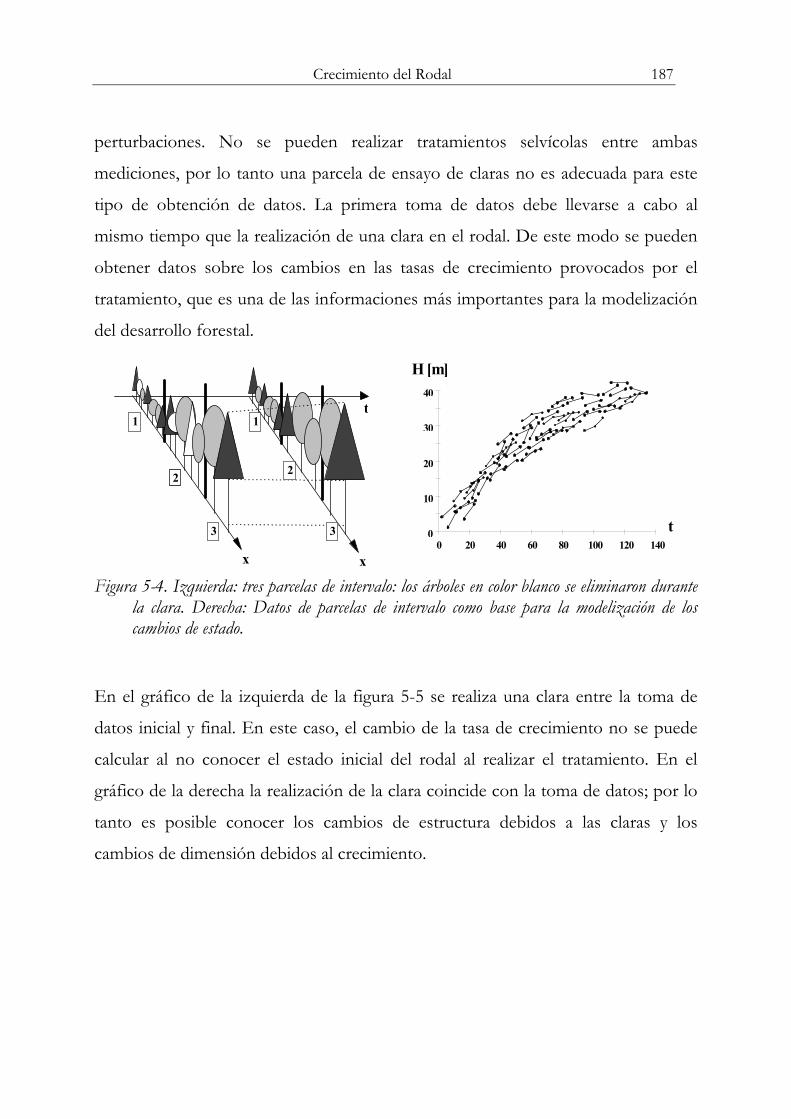
Crecimiento del Rodal 187
perturbaciones. No se pueden realizar tratamientos selvícolas entre ambas
mediciones, por lo tanto una parcela de ensayo de claras no es adecuada para este
tipo de obtención de datos. La primera toma de datos debe llevarse a cabo al
mismo tiempo que la realización de una clara en el rodal. De este modo se pueden
obtener datos sobre los cambios en las tasas de crecimiento provocados por el
tratamiento, que es una de las informaciones más importantes para la modelización
del desarrollo forestal.
t
x x
1
2
3 3
2
1
0
10
20
30
40
0 20 40 60 80 100 120 140 t
H [m]
Figura 5-4. Izquierda: tres parcelas de intervalo: los árboles en color blanco se eliminaron durante la clara. Derecha: Datos de parcelas de intervalo como base para la modelización de los cambios de estado.
En el gráfico de la izquierda de la figura 5-5 se realiza una clara entre la toma de
datos inicial y final. En este caso, el cambio de la tasa de crecimiento no se puede
calcular al no conocer el estado inicial del rodal al realizar el tratamiento. En el
gráfico de la derecha la realización de la clara coincide con la toma de datos; por lo
tanto es posible conocer los cambios de estructura debidos a las claras y los
cambios de dimensión debidos al crecimiento.

188 Crecimiento del Rodal
W
t
*
Δt
ΔW
*
*
t1 t2
W2
W1
W
t
*
Δt
ΔW
*
*
t1 t2
W2
W1
*a
a b
Figura 5-5. Dos tomas de datos consecutivas para calcular las tasas de cambio de una variable de estado W. Izquierda: incorrecta aplicación. Las consecuencias de la clara son desconocidas, por lo tanto ΔW no ofrece ninguna información útil. Derecha: aplicación correcta. Se toman los cambios como consecuencia de la clara y el crecimiento natural.
Una de las ventajas de contar con datos que permiten obtener tasas de crecimiento,
como por ejemplo los datos de parcelas de intervalo, es la posibilidad de poder
emplear funciones de crecimiento en forma de diferencias algebraicas (ver por ejemplo
Clutter et al. 1983; Ramírez-Maldonado et al., 1988; Forss et al., 1996). Este tipo de
ecuaciones tiene, entre otras ventajas, la de ser modelos invariantes con respecto al
camino de proyección. Esto quiere decir que si se simula la evolución de un rodal
desde la edad t1 hasta la edad t2 y después desde t2 hasta la edad t3, se llega a la
misma estimación que si se va directamente desde la edad t1 hasta la edad t3. A
partir de la ecuación 5-1 se pueden obtener tres ecuaciones en diferencias
algebraicas, puesto que la ecuación tiene tres parámetros (a0, a1 y a2). Si se cuenta
con dos mediciones de altura dominante de un mismo rodal H1 y H2 obtenidas a las
edades t1 y t2, respectivamente, se puede obtener la siguiente ecuación en diferencias
algebraicas despejando el parámetro a0:
2
11
21
11
12
a
ta
ta
eeHH ⎟⎟
⎠
⎞⎜⎜⎝
⎛−−
⋅= ⋅−
⋅−
5-2
El concepto de la parcela de intervalo ofrece la ventaja de que dentro de un espacio
de tiempo relativamente corto se pueden obtener las tasas de cambio para una gran

Crecimiento del Rodal 189
diversidad de estados iniciales. Otra ventaja es su flexibilidad: las parcelas de
intervalo pueden ser abandonadas en cualquier momento después de que hayan
sido medidas dos veces, a veces, incluso puede ser ventajoso abandonar una parcela
ya medida e instalar una nueva, puesto que aunque los costes de instalación son
superiores a los de mantenimiento de una parcela ya existente, al instalar una nueva
parcela se aumenta la diversidad de estados iniciales.
Pese a la utilidad de este tipo de parcelas, hay preguntas que sólo se pueden
responder con precisión en base a parcelas permanentes. Por ejemplo, los cambios
a largo plazo en el crecimiento debido a influencias climáticas o como consecuencia
de cambios en las condiciones de la estación sólo pueden ser analizados en rodales
cuyo desarrollo se ha venido observando durante un largo período de tiempo. Por
tanto, no se debe renunciar a las parcelas permanentes aunque su número nunca
podrá ser muy elevado debido a los costes de mantenimiento.
De todo esto se deduce que las parcelas permanentes, por si solas, no son
suficiente fuente de información debido a la amplia variedad de estados iniciales y
de tratamientos posibles. Tampoco el inventario forestal continuo proporciona
buenos datos para la modelización del crecimiento forestal ya que no se realiza la
medición coincidiendo con las claras. Algo parecido ocurre con la información
obtenida del análisis de troncos cuyos datos no proporcionan ninguna conclusión
sobre el entorno histórico y desarrollo de la copa del árbol analizado. Por tanto, en
vista de la limitación de los medios disponibles y de la creciente diversidad de las
estructuras forestales y formas de mezcla es necesaria una nueva orientación en la
planificación de los ensayos forestales. Sería conveniente una mayor integración y
coordinación de todos los centros de investigación forestal con sus experiencias
para la obtención de bases de datos más completas y fiables.

190 Crecimiento del Rodal
Modelos de producción regional Los modelos de producción regional permiten realizar una estimación de la
producción maderera en amplias regiones forestales. Se trata de estimaciones muy
generales basadas en hipótesis muy simples por las que las condiciones de la
estación y del aprovechamiento forestal se consideran constantes. Un ejemplo de
este tipo de modelos son las ecuaciones obtenidas por Shvidenko et al. (1995) para
varias especies en Rusia. La ecuación obtenida por estos autores para una clase de
producción III de Pinus sylvestris es la siguiente:
( ) ( ) 93,20231,013,205 tetV ⋅−−⋅= 5-3
Donde V(t) es el volumen del rodal (m³/ha) a la edad t. Con la ayuda de la ecuación
5-3 se puede crear una sencilla tabla de evolución del volumen con la edad:
Centro de la clase de edad (años)
10 30 50 70 90 110 130 150 170 190 Volumen (m³/ha) 2,0 26,9 67,7 107,4 138,8 161,4 176,8 187,1 193,7 197,9
Tabla 5-2. Tabla de producción para un rodal de Pinus sylvestris de la clase de producción III en el norte de Rusia.
Estos modelos se pueden aplicar a grandes área geográficas conociendo la
distribución real de la superficie por clases de edad y son una base para la
planificación de la producción regional. Se trata de modelos altamente agregados, es
decir, que aportan información general del rodal, sin especificar la situación de cada
árbol o de una serie de árboles representativos, por lo que ofrecen la posibilidad de
comprobar fácilmente diferentes alternativas de aprovechamiento en relación a su
sostenibilidad. Sin embargo, hay que tener en cuenta que el desarrollo de un rodal
forestal puede ser muy diferente según la calidad de estación y los tratamientos
selvícolas aplicados.

Crecimiento del Rodal 191
Una condición para la utilización de estos modelos de producción es la
comprensión de las relaciones entre la masa total, el crecimiento medio y el
crecimiento corriente.
La masa total y el incremento de crecimiento
El rendimiento de una determinada variable, como por ejemplo el volumen,
referido a la edad de un rodal se describe mediante las variables masa total, crecimiento
medio y crecimiento corriente. La masa total a a la edad t (MTt) es igual al volumen de la
masa residual del rodal a la edad t (Vt) más la suma del volumen extraído en las
claras (Dfi) hasta la edad t:
∑=
+=t
1iitt DfVMT 5-4
El crecimiento medio a la edad t (CMt) es igual al valor de la masa total MTt dividido por la edad del rodal.
tMTCM t
t = 5-5
El crecimiento corriente (CCt) es igual a la pendiente de la tangente a la curva de
crecimiento, es decir, por definición es la derivada de la ecuación de masa total
(MTt) con respecto a la edad (t) y describe la tasa de crecimiento a la edad t:
tMTTMCCt Δ
Δ=′= 5-6
El crecimiento corriente culmina en el punto de máxima pendiente, es decir en el
punto de inflexión de la curva de crecimiento. El crecimiento medio alcanza su
máximo en el punto donde la tangente de la curva de crecimiento pasa por el
origen, es decir cuando el crecimiento medio iguala al crecimiento corriente. Hasta
este momento el crecimiento corriente es mayor que el crecimiento medio y pasado
este punto es inferior.

192 Crecimiento del Rodal
Por ejemplo, suponiendo que la función de crecimiento en volumen con la
edad para un rodal hipotético de pino es )exp(-100/t1800MTt ⋅= , las ecuaciones del
crecimiento medio y el crecimiento corriente serán t/)t/100exp(1800CMt −⋅= y
, respectivamente. 2tt t/100)t/100exp(1800MTCC ⋅−⋅=′=
Por tanto, la edad a la que se alcanza el máximo del crecimiento medio (CM) se obtendrá como:
⎟⎠⎞
⎜⎝⎛−⎟
⎠⎞
⎜⎝⎛−
⋅−⋅⋅==′ t100
22t
100
t et
1800t
100et
18000CM 100tt1
t100
2 =→=→
Por último, la edad a la que se alcanza el máximo del crecimiento corriente se
obtendrá del siguiente modo:
50t t1
t50 0e
t360000
t100e
t180000TM 34
t100
32t
100
2 =→=→=⋅−⋅⋅=′′⎟⎠⎞
⎜⎝⎛−⎟
⎠⎞
⎜⎝⎛−
Un ejemplo de este tipo de modelos es la ecuación de estimación del volumen de
rodal en plantaciones de Pinus elliottii en el Sur del estado de Georgia propuesta por
Pienaar et al. (1990):
( )3,187tNP0.00821.70 0.349
e1,0581SI0,043V ⋅⋅−⋅−⋅⋅= 5-7
Siendo V el volumen sin corteza (cunits/acre); SI es el índice de sitio definido
como la altura dominante (pies) a la edad de referencia de 25 años; NP es el
número de pies supervivientes por acre a la edad de 2 años y t es la edad del rodal
(años).
Las relaciones esenciales entre estas tres variables de crecimiento antes
descritas se presentan en la figura 5-6. El crecimiento corriente en el período t1 - t2
(CCt1-t2) es igual al cociente entre el crecimiento de la masa total y el tiempo
transcurrido:

Crecimiento del Rodal 193
ΔtΔMT
ttMTMT
CC12
tttt
12
21=
−
−=− 5-8
0
100
200
300
400
500
600
700
800
900
1000
0 50 100 150 Edad [años]
MT
[m3/ha]
0
10
20
30
40 CM, CC
[m3/ha año]MT
CCCM
Figura 5-6. Relación entre masa total (MT), crecimiento medio (CM) y crecimiento corriente
(CC). Como se observa en la ecuación 5-8, el crecimiento corriente se puede medir
independientemente de la edad, con sólo conocer el intervalo de tiempo
transcurrido entre mediciones. El cálculo del crecimiento corriente se representa en
el ejemplo de la figura 5-7.
1 32
Figura 5-7. Rodal con tres árboles en los que se esquematiza su crecimiento en volumen en tres años (Sterba, 1991).
El centro blanco representa el volumen del tronco a la edad t1. Hasta la edad t2 los
árboles han crecido la superficie punteada. En este momento se extrae el árbol

194 Crecimiento del Rodal
central. Hasta la edad t3 los árboles han crecido nuevamente en volumen rayado. El
incremento periódico total es igual a la suma de ambas superficies:
CC t1 - t3= (V13-V11) + (V22-V21) + (V33-V31) = VE - VA + Vext
donde VE es el volumen al final de período; VA es el volumen inicial y Vext es el
volumen extraído en claras.
Ya que la madera extraída, que suele permanecer en el suelo después de haber
efectuado la clara, no siempre se inventaría, en la práctica es necesario para calcular
el crecimiento periódico métodos de inventario especiales, como por ejemplo el
inventario de la masa extraída antes de la corta.
Tablas de producción
Con la construcción de las primeras tablas de producción al final del siglo XVIII se
crearon los primeros modelos de crecimiento, que reconstruyen el desarrollo del
rodal en base a valores medios (ver Paulsen, 1795). Estas tablas de producción
presentan las variables más importantes en rodales puros con un tratamiento
selvícola definido y unidades temporales fijas (normalmente 5 años). Las tablas de
producción publicadas en la mayor parte del mundo hasta la actualidad han sido
principalmente tablas de producción regionales en base a datos de parcelas
permanentes de los centros de investigación. En Europa, la construcción de tablas
de producción tiene su origen en centro-Europa, por ejemplo, en Alemania las
primeras tablas de producción vieron la luz entre finales del siglo XIX y el primer
tercio del siglo XX (Baur, 1881; Schwappach, 1890; Eberhard, 1902; Grundner,
1904; Schwappach, 1911; Wimmenauer, 1914; Wiedemann, 1931).
Las tablas de producción se derivan de las pautas de crecimiento medidas en
numerosas parcelas de ensayo; son modelos comprensibles y fáciles de manejar que
reflejan el crecimiento de las especies más importantes y fueron ampliamente
aplicadas en la planificación forestal tradicional. Las tablas de producción estiman el
desarrollo de variables importantes desde el punto de vista productivo para la masa

Crecimiento del Rodal 195
residual y la masa extraída para un número limitado de tratamientos típicos. Su
función es describir operacionalmente las claras planeadas, estimar los volúmenes
extraídos y pronosticar el desarrollo del rodal tras la clara (Tabla 5-3).
La representación tabular de estos modelos apenas ha cambiado en los
últimos 200 años (ver por ejemplo Paulsen, 1795; Hartig, 1847; Weise, 1880;
Schwappach, 1890; Gehrhardt, 1930; Wiedemann, 1949; Schober, 1972, 1995).
Esto es sin duda un signo de que las tablas de producción cumplen los
requerimientos de la gestión forestal a medio y largo plazo.
Los valores de las variables de masa que aparecen reflejados en las tablas de
producción son válidos para una clara cuyo tipo e intensidad coincida con los de las
parcelas de las que se obtuvieron los datos.
Masa residual Masa extraída Edad (años)
N° de pies
Diámetro(Pulgada)
Altura (Pies)
Volumen(Klafter)
N° de pies
Volumen (Klafter)
20 3644 2 12 5,95 - - 28 911 4 24 11,92 2733 4,46 36 405 6 33 16,39 506 6,62 44 228 8 40 19,86 177 7,17 52 146 10 45 22,35 82 7,15 60 101 12 49 24,34 45 6,83 68 74 14 53 26,32 27 6,41 76 57 16 56 27,81 17 6,16 84 45 18 59 29,30 12 5,83 92 36 20 61 30,30 9 5,56
100 30 22 63 31,18 6 5,34 110 25 24 64 31,78 5 4,89 120 21 26 64 31,78 4 4,71
Tabla 5-3. Tablas de producción de haya para una buena calidad de estación realizadas por Paulsen (1795). El número de pies y el volumen se refieren a 1 Morgen, el diámetro y la altura son los valores medios del rodal.
Para caracterizar esas claras se han venido utilizando códigos como el definido por
la Asociación Alemana de Centros de Investigación Forestal (Vereins Deutscher
Forstlicher Versuchsanstalten, 1902):

196 Crecimiento del Rodal
Árbol dominado Árbol dominante Grado 5 4 3 2 1
residual A
extraído
residual B
extraído
residual
C extraído
residual
D extraído
residual
E extraído
Figura 5-8. Representación simplificada de los grados de clara (Schober, 1994). El color amarillo indica que el árbol no se extrae y el color blanco indica que el árbol se extrae. Las secciones indican parcialmente extraído.
Las variables de entrada para las tablas de producción suelen ser el índice de sitio o
la clase de producción. Para cada clase de producción hay una tabla con los datos
de la masa residual y la masa extraída. Por ejemplo, en la Tabla 5-4 se muestra un
extracto de una tabla de producción para pícea.
En general, los rodales no se desarrollan conforme a las tablas de producción
debido a las diferencias en la calidad de estación y en los tratamientos selvícolas
llevados a cabo. Esta circunstancia se debe tener en cuenta al emplear las tablas
mediante una calibración de las estimaciones de la propia tabla de producción.

Crecimiento del Rodal 197
Picea Clara mediaMasa residual Masa extraída
Edadr N/ha Hm Rango calidad Altura dom. G/ha Diam f V/ha N/ha Hm G/ha Diam V/ha
Hm de - a Weise h100
años N°pies m m m m qm cm 0,... fm Stück m qm cm fm
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
I. Clase de producción20 5917 7,1 6,0- 8,2 7,5 8,5 26,0 7,5 212 39 - - - - -
25 4260 9,2 7,9-10,6 10,2 11,1 30,1 9,5 373 103 1657 6,6 5,0 6,2 3
30 3110 11,5 10,0-13,1 12,8 13,7 32,5 11,5 462 172 1150 8,3 5,9 8,1 13
35 2382 14,1 12,4-15,8 15,8 16,4 34,2 13,5 497 240 728 9,8 5,4 9,7 19
40 1886 16,6 14,8-18,4 18,3 19,0 35,5 15,5 516 304 496 11,4 5,1 11,4 23
45 1548 19,0 17,0-20,9 21,1 21,4 37,1 17,5 518 365 338 13,6 4,2 12,6 24
50 1326 21,2 19,0-23,1 23,5 23,7 38,7 19,3 516 423 222 15,4 3,3 13,8 25
55 1148 23,1 20,9-25,0 25,2 25,5 40,3 21,1 513 478 178 17,6 3,0 14,6 26
60 1007 24,7 22,6-26,7 26,9 27,1 41,9 23,0 511 529 141 19,6 2,7 15,6 27
65 887 26,1 24,0-28,1 28,3 28,4 43,4 25,0 507 574 120 21,3 2,6 16,6 29
70 787 27,4 25,3-29,5 29,6 29,7 44,7 26,9 502 615 100 22,7 2,7 18,5 32
75 702 28,6 26,5-30,6 30,7 30,8 45,8 28,8 496 650 85 24,3 2,8 20,5 35
80 631 29,7 27,6-31,7 31,7 31,8 46,7 30,7 491 681 71 25,3 2,9 22,8 37
85 571 30,7 28,6-32,8 32,7 32,7 47,4 32,5 485 706 60 26,4 3,0 25,2 40
90 520 31,6 29,6-33,7 33,5 33,5 47,9 34,2 479 725 51 28,0 3,0 27,4 42
95 475 32,5 30,5-34,6 34,4 34,4 48,2 35,9 473 741 45 29,5 3,0 29,1 43
100 435 33,3 31,3-35,4 35,3 35,1 48,3 37,6 469 754 40 31,0 3,0 30,9 46
105 399 34,1 32,1-36,1 36,4 35,8 48,2 39,2 463 761 36 32,2 3,0 32,6 47
110 366 34,8 32,8-36,8 36,8 36,4 48,0 40,9 458 765 33 33,4 3,0 34,0 48
115 336 35,4 33,4-37,4 37,4 36,9 47,7 42,5 454 767 30 34,0 3,0 35,7 48
120 308 35,9 34,0-37,8 37,6 37,3 47,4 44,3 451 767 28 34,6 2,9 36,3 48 Tabla 5-4. Extracto de la tablas de producción para picea de Wiedemann-Schober, clara media
(N/ha = Número de pies por ha; Hm = altura media; G/ha = área basimétrica m²/ha, Diam = diámetro medio cm; f: factor de forma; V/ha = volumen m³/ha).
Ejemplos de tablas de producción regional para el haya
Como ejemplo de tablas de producción construidas en diferentes regiones para una
misma especie común se muestran en la tabla 5-5 los datos más importantes de las
tablas de producción existentes para haya en 16 países europeos recopiladas gracias
a la cooperación de varios institutos de investigación.

198 Crecimiento del Rodal
País Año Región Autor Referencia
Albania 1998 Gadow K.v., Postoli A.
Waldwachstum: Modelle der Wald-entwicklung (en albano). GTZ Publicación: 201 p.
Alemania 1972 Schober R. Die Rotbuche. J. D. Sauerländer's Verlag, Frankfurt am Main.
Alemania 1986 DDR Dittmar O., Knapp E.,
Lembcke G.
DDR-Buchenertragstafel. IFE-Berichte aus Forschung und Entwicklung Heft 4 1986.
Dinamarca 1996 Skovsgaard J.P.; Mosing M.
Bogefoyngelser i Ostjylland. Danish forest and Landscape Research Institute
Eslovaquia 1998 Halaj J., Petras R.
Rastove tabulky hlavnych drevin
España 1992 Navarra Madrigal A., Puertas F., Martinez J.
Tablas de producción para Fagus sylvatica L. en Navarra. Gobierno de Navarra, Dep. de Agricultura, Ganadería y Montes
Grecia 1999 (Norte de Grecia) Gatzojannis S.
Yield tables for beech stands in the forest A. Brontou Serres Prefecture, N. Greece (en griego). ΑΝΨΕΡΕ ΗΚΙΣΑΔ,12: 91-104.
Gran Bretaña 1971 Hamilton G.J., Christie J M.
Forest Management Tables (Metric). Her Majesty's stationary office.
Hungría 1983 Mendlik G. Beech Yield Table (en Hungaro). Erdeszeti-Kutatasok 75:157-162
Italia 1974 Gargano Gualdi V. Ricerche auxometriche sulle faggete del Gargano. L'Italia Forestale e Montana 29:85-116.
Países Bajos 1996 Jansen J.J.,
Sevenster J., Faber P.J.
Opbrengst Tabellen voor belangrijke boomsoorten in Nederland. Landbouwuniversiteit Wageningen; IBN-Rapport 221; Hinkeloord-Report No 17.
Polonia 1998 Bruchwald A., Dudzinska M., Wirowski M.,
Model wzrostu dla lisciastych gatunkow drzew lesnych. Model wzrostu buka. Forest Research Institute in Warsaw.
República Checa 1996 CernyM., Parez J., Malik Z.
Rustove a taxacni tabulky hlavnich drevin Ceske Repbliky Ministerstva zemedelství.
Rumanía 1972 Giurgiu V., Decei I., Armasescu S.
Biometria arborilor si arboretelor din Romania (The biometry of trees and stands in Rumania:forest mesuration tables). Ceres Verlag.
Suecia 1971 Sur de Suecia Carbonnier C. Studia Forestalia Suecica. Nr. 90. Royal College of Forestry, Stockholm.
Suiza 1983 WSL Birmensdorf
Ertragstafeln 3. Auflage
Tabla 5-5. Datos de las tablas de producción de haya de diferentes países europeos.
Para analizar las diferencias regionales se parametrizaron separadamente los datos
de las tablas de producción para las variables número de pies, altura dominante y
área basimétrica. En la figura 5-9 se muestra la evolución en el tiempo de dos

Crecimiento del Rodal 199
variables de rodal para dos de las tablas de producción de las incluidas en la tabla
anterior.
0 5
10 15 20 25 30 35 40
0 20 40 60 80 100 120 140
Edad
Skovsgaard y Mosing
Schober
H (m)
0
5
10
15
20
25
30
35
40
45
0 10 20 30 40 50 60 70
Edad
G (
m²/
ha)
Döbbeler y Spellmann
Skovsgaard y Mosing
Schober
Figura 5-9. Comparación del desarrollo de la altura dominante (izquierda) y del área basimétrica (derecha) según Schober (1972) y Skovsgaard y Mosing (1996) para una calidad de estación representada por una altura dominante de 32 metros a la edad de referencia de 100 años. En la figura de la derecha aparece también la evolución del área basimétrica correspondiente a un aprovechamiento en densidad máxima (Döbbeler y Spellmann, 2002).
El peso de la clara se puede describir mediante el área basimétrica extraída relativa
(rG%) y el tipo de clara se puede describir mediante la relación entre el número de
pies y el área basimétrica (NG). Los datos de la tabla 5-6 se corresponden con los
gráficos en la figura 5-9. Schober Skovsgaard y Mosing Edad rG (%) NG rG (%) NG
20 - - 0,10 1,60 25 - - 0,11 1,59 30 - - 0,22 1,38 35 0,15 2,44 0,08 1,56 40 0,14 2,10 0,20 1,38 45 0,13 1,85 0,14 1,40 50 0,12 1,78 0,17 1,36 55 0,11 1,71 - - 60 0,10 1,68 - - 65 0,10 1,67 - -
Tabla 5-6. Peso de la clara (rG%) y tipo de clara (NG) para las variantes representadas en la figura 5-9.
La relación NG obtenida para las dos tablas de producción indica que se trata de
claras débiles en la que se extrajeron los árboles de los estratos inferiores. El peso
de las claras de Skovsgaard y Mosing (1996) es variable. Como se observa en la

200 Crecimiento del Rodal
gráfica de área basimétrica, en ambos casos el valor de las tablas permanece por
debajo de los valores de la densidad máxima según Döbbeler y Spellmann (2002).
Parametrización de los modelos de tablas de producción
En base a los valores de las tablas de producción de haya en Europa se desarrolló
un modelo de crecimiento de rodal dependiente de la edad. El modelo se puede
emplear para determinar la evolución de un rodal para una alternativa silvícola
similar a la descrita en la tabla de producción pero no se puede emplear, o sólo de
manera limitada, para analizar diferentes tratamientos.
Para la determinación de la altura dominante o del índice de sitio de un rodal
de haya se parametrizó la ecuación exponencial de Sloboda (1971):
( ) ( ) ⎟⎟⎠
⎞⎜⎜⎝
⎛
⋅−+
⋅−−
−−
⎟⎠⎞
⎜⎝⎛⋅=
11 11001exp ac Aa
ba
b
cSIcH 5-9
Donde H es la altura dominante en metros; SI es el índice de sitio, definido como
la altura dominante a una edad de referencia de 100 años; t es la edad del rodal y a,
b y c son parámetros empíricos. En la tabla 5-7 se incluyen los valores de estos
parámetros para los diferentes países con tablas de producción de la especie. País n a b C R2
Albania 135 0,1364 1,6407 1,986·10-4 0,99 Alemania 335 0,0145 4,3698 1,092·10-4 0,99 Dinamarca 141 0,4416 0,7131 4,643·10-4 0,98 Eslovaquia 374 0,2761 1,1287 2,262·10-4 0,99 España 94 0,6699 0,2561 5,751·10-4 0,99 Francia 75 0,1019 3,1119 1,409·10-4 0,99 Grecia 91 0,6158 0,2740 3,138·10-4 0,99 Gran Bretaña 105 0,0660 2,1057 3,306·10-4 0,99 Hungría 78 0,3660 0,8282 2,328·10-4 0,99 Italia 39 -0,1748 5,1838 1,490·10-4 0,99 Países Bajos 137 0,2646 1,4974 9,530·10-4 0,99 Polonia 401 0,3762 0,8456 6,799·10-6 0,99 Rep. Checa 111 0,4258 0,5276 5,381·10-4 0,99 Rumanía 127 0,1900 1,3108 3,333·10-4 0,99 Suecia 122 0,0033 6,5685 1,207·10-4 0,99 Suiza 52 0,0507 3,6291 3,360·10-4 0,99
Tabla 5-7. Coeficientes para la estimación de la altura dominante/índice de sitio (n es el número de datos empleado y R2 es el coeficiente de determinación del ajuste).

Crecimiento del Rodal 201
Para la modelización del desarrollo del área basimétrica se empleó la ecuación en
diferencias algebraicas propuesta por Hui y Gadow (1993c):
cHaHa
HHNNGG
bb
⎟⎟⎠
⎞⎜⎜⎝
⎛⋅⋅⋅= −⋅⋅−
1
211
1212
12 5-10
Donde G1 y G2 son el área basimétrica (m2/ha) a las edades inicial y final de la
proyección, H1 y H2 son las alturas dominantes (m) a esas mismas edades y a, b y c
son los parámetros del modelo. Los coeficientes empíricos obtenidos a partir de las
tablas de producción de cada uno de los países se incluyen en la tabla 5-8. País N a b c R2
Albania 130 1,0040 -0,0981 1,6315 0,99 Alemania 322 1,0177 -0,0069 0,4985 0,98 Dinamarca 133 0,1159 0,4957 3,2284 0,97 Eslovaquia 361 1,0057 -0,0024 0,4404 0,91 España 88 0,4062 -0,3146 2,4437 0,99 Grecia 84 1,6743 -0,1511 -0,8258 0,99 Gran Bretaña 101 0,2795 0,4029 3,2957 0,99 Hungría 72 0,2544 0,3452 0,3217 0,97 Italia 36 0,5142 0,1289 2,1465 0,99 Países Bajos 132 0,3847 0,1873 2,0798 0,99 Polonia 389 -2,3307 -1,4479 2,9257 0,99 Rep. Checa 100 1,0592 0,0146 0,4252 0,99 Rumanía 118 1,4166 -0,0968 -0,0691 0,99 Suecia 114 0,0493 0,0501 3,7126 0,95 Suiza 46 1,4751 -0,1000 -0,4777 0,98
Tabla 5-8. Coeficientes empíricos para estimar el área basimétrica (n es el número de datos empleados y R2 es el coeficiente de determinación del ajuste).
El desarrollo del número de pies se acopló al crecimiento de la altura dominante,
basándose en el método de Clutter et al. (1983):
( )a
cca
HHbNN
1
121
2 000.1000.1
⎥⎥⎦
⎤
⎢⎢⎣
⎡−⋅+⎟
⎠⎞
⎜⎝⎛⋅= 5-11
Donde N1 y N2 son el número de pies por hectárea a las edades inicial y final de la
proyección, H1 y H2 son las alturas dominantes (m) a esas mismas edades y a, b y c
son los parámetros del modelo. En la tabla 5-9 se incluyen los coeficientes
empíricos obtenidos para cada uno de los países cuyas tablas de producción figuran
en la tabla se incluyen en la tabla 5-5.

202 Crecimiento del Rodal
País n a b c R2
Albania 130 0,0247 -0,0429 0,3898 0,99 Alemania 318 0,0115 -3,0966 0,0082 0,99 Dinamarca 133 0,0728 -0,0002 2,2683 0,99 Eslovaquia 361 -0,8224 -53,6177 -0,0024 0,99 España 88 -0,2491 -47,7015 -0,0048 0,99 Grecia 84 0,0360 -0,0015 1,2283 0,99 Gran Bretaña 101 0,3373 -0,1435 0,8369 0,99 Hungría 72 -0,0056 -5,0719 -0,0017 0,99 Italia 36 0,0805 -0,0797 0,5008 0,99 Países Bajos 132 0,0284 -0,0048 0,9458 0,99 Polonia 389 -0,0020 -0,2685 -0,0120 0,99 Rep. Checa 100 0,1905 -0,9984 0,2581 0,99 Rumanía 118 0,0051 -0,0087 0,4101 0,99 Suecia 114 0,0035 -0,2706 0,0333 0,99 Suiza 46 -0,2210 -66,6699 -0,0056 0,99
Tabla 5-9. Coeficientes empíricos para estimar el número de pies (n es el número de datos empleados y R2 es el coeficiente de determinación del ajuste).
Modelos matemáticos de producción La mayoría de los modelos de producción matemáticos se derivan de datos
empíricos y son, por tanto, una representación simplificada de dichas
observaciones. Una excepción a esta regla es el modelo de crecimiento medio
máximo de Murray que se deriva del valor del incremento máximo en volumen de
un rodal y de los valores de crecimiento de la especie observados en los rodales del
área de estudio.
Para la modelización de la producción potencial regional es especialmente
importante conocer el valor del crecimiento medio, la edad a la que se alcanza su
máximo y cuál es el valor de dicho máximo. Una función que por sus características
es muy adecuada para la descripción del crecimiento es la función de Chapman-
Richards:
{ }( )mtotk-e1AV(t) −−= 5-12
donde V(t) es el volumen del rodal a la edad t (en m³/ha). A se puede interpretar
como la asíntota o punto de culminación y tiene las mismas unidades que V(t). El
parámetro k es el factor de escala del eje temporal y sus unidades son la inversa del
tiempo, es decir, si el tiempo se mide en años entonces las unidades de k son años-1.

Crecimiento del Rodal 203
El parámetro t0 es el límite inferior de edad a partir del cual es apreciable el
crecimiento en volumen del rodal. El parámetro m esta relacionado con la forma de
la curva de crecimiento.
Dividiendo la ecuación 5-12 por la edad se obtiene la expresión del
crecimiento medio anual CM.
[ ]
t
mttkeA
tV ⎟
⎠⎞
⎜⎝⎛ −⋅−−⋅
==
01CM 5-13
El valor máximo del crecimiento medio anual (CMmax), que se supone conocido, se
alcanzará a una edad tmax, también conocida, para la cual la primera derivada de la
ecuación 5-13 es igual a cero:
[ ] ( ) [ ]0
11
1CM' 2
000
=⎟⎠⎞
⎜⎝⎛ −⋅−−−−−−
⎟⎠⎞
⎜⎝⎛ −⋅−−
=⋅⋅⋅⋅⋅⋅
t
mttkeAtttkemttkeA km
Resolviendo se obtiene la siguiente relación:
( ) ( ) 10 0maxe1 max −⋅⋅⋅+= −⋅− ttkkmt 5-14
Murray y Gadow (1993) consideraron que el valor de t0 para especies de
crecimiento rápido se puede asimilar a 1 y que el parámetro de forma (m) toma
valores muy cercanos a 3; por lo que la ecuación 5-14 permite obtener el valor del
parámetro k conocida la edad (tmax) a la que se alcanza el máximo del crecimiento
medio anual. Dicho valor no se puede despejar directamente de la ecuación anterior
por lo que debe ser estimado por procedimientos iterativos.
Una vez determinados tres parámetros de la función de Chapman-Richards, el
valor de la asíntota se obtiene haciendo pasar la ecuación 5-12 por el punto de
coordenadas (CMmax , tmax). Cuando sólo se conoce la edad a la que se alcanza el
máximo pero no su valor se puede obtener la evolución del crecimiento medio
anual relativo asignando un valor de 1 al CMmax.

204 Crecimiento del Rodal
[ ]
[ ] mttket
mttket
⎟⎠⎞
⎜⎝⎛ −⋅−−⋅
⎟⎠⎞
⎜⎝⎛ −⋅−−
=⋅
0
0
max
max
1
1CMrelativo 5-15
La Figura 5-10 muestra tres ejemplos de curvas de crecimiento medio anual relativo
para diferentes edades de obtención del máximo y considerando t0 igual a 1 y m
igual a 3.
0,0
0,2
0,4
0,6
0,8
1,0
1,2
0 20 40 60 80 100 120 Edad [años]
rel. CM
a b c
Figura 5-10. Tres curvas del crecimiento medio relativo siendo a: tmax = 20, k = 0,1043; b: tmax = 50, k = 0,0394 y c: tmax = 80, k = 0,02434.
Las curvas de crecimiento medio en valores absolutos se obtienen multiplicando los
valores relativos por el valor conocido por la experiencia de campo del crecimiento
medio máximo. Murray y Gadow (1993) observaron que las estimaciones de este
modelo matemático se adecuaban con bastante exactitud a las estimaciones de
modelos de crecimiento empíricos. En caso de que se conozcan por medio de la
experiencia los valores del crecimiento medio máximo (CMmax) y la edad a la que se
alcanza (tmax) estos modelos pueden proporcionar estimaciones más fiables que las
de modelos empíricos obtenidos a partir de bases de datos de observaciones
incompletas.

Crecimiento del Rodal 205
Porcentaje de incremento Otra herramienta útil para la planificación de los tratamientos selvícolas es el
empleo de los denominados “porcentajes de incremento”. Los porcentajes de
incremento son las tasas de crecimiento relativas de una variable dasométrica o
dentrométrica, como por ejemplo el volumen del rodal. Si V0 es el volumen de un
rodal (m3/ha) tras la realización de una clara y p es el porcentaje de incremento del
volumen, entonces tras un período entre claras de “cc” años el volumen del rodal
Vcc (m3/ha) vendrá dado por la siguiente expresión:
cc
ccp=VV ⎟
⎠⎞
⎜⎝⎛ +⋅
10010 5-16
La tabla 5-10 muestra los porcentajes de incremento en función del volumen para
bosques mixtos y para bosques de coníferas en Finlandia (Lähde et al., 2002). Porcentaje de incremento
Volumen del rodal (m³/ha)
Tipo de rodal
Irregular Regular Coníferas 5,1 ± 1,8 4,4 ± 0,7
80-120 Mixtos 5,5 ± 1,5 4,9 ± 1,1 Coníferas 4,2 ± 1,1 3,9 ± 0,5
120-160 Mixtos 4,6 ± 1,0 3,8 ± 1,0 Coníferas 3,3 ± 0,9 2,8 ± 0,7
>160 Mixtos 3,6 ± 0,8 2,9 ± 0,8
Tabla 5-10. Valores del porcentaje de incremento en volumen (valor medio ± desviación típica) en bosques de coníferas y en bosques mixtos de estructura irregular y regular.
Si se conoce el volumen inicial, la tasa de crecimiento y el volumen que se quiere
obtener antes de realizar la siguiente corta, entonces se puede calcular el tiempo que
debe transcurrir entre tratamientos según la siguiente fórmula:
( ) ( )
⎟⎠⎞
⎜⎝⎛ +
=
1001
- 0
pn
VnVncc cc
l
ll 5-17

206 Crecimiento del Rodal
Esta metodología se aplica, por ejemplo, para calcular el ciclo de claras para
bosques de pino en la región de El Salto, Durango, México.
Modelos de crecimiento dependientes de la densidad Una de las metas de la investigación forestal actual es el desarrollo de modelos que
posibiliten comparar diferentes tratamientos selvícolas con vistas a distintos
objetivos finales. Las tablas de producción y otros modelos de producción
regionales no se pueden emplear con este fin porque estiman el crecimiento y la
producción de rodales que han sido tratados de una forma determinada y
específica. En estos casos sería necesaria una calibración de las tablas de
producción existentes, que será tanto más complicada cuanto más se desvíen las
nuevas prácticas selvícolas de las prescripciones de las tablas de producción.
La relación entre la densidad del rodal y el crecimiento
Numerosos estudios empíricos han indicado que el crecimiento en altura no se ve
fuertemente influido por la densidad del rodal, mientras que el crecimiento en
diámetro reacciona de manera sensible a los cambios en dicha densidad. Los
trabajos de Craib (1939) son los primeros que indican, en parte, la rápida respuesta
del crecimiento en diámetro a la realización de diferentes tratamientos selvícolas. La
figura 5-11 es una representación original de la tesis doctoral de Craib. Los troncos
analizados provienen de tres árboles de la especie Pinus patula de la misma edad
pero extraídos de parcelas de ensayo de diferente densidad. Craib (1939) pudo
determinar en base a sus observaciones de sus parcelas de ensayo que el
crecimiento en diámetro de los árboles dominantes apenas se ve influido por la
densidad del rodal. Esto provoca que, además de que la varianza del diámetro
aumente con la edad, aparezca una típica inclinación o asimetría de la distribución
diamétrica.

Crecimiento del Rodal 207
1
2 3 Figura 5-11. Muestras de rodajas tomadas a la altura del pecho en tres árboles de Pinus patula
de 9 años de edad. El árbol 1 creció sin competidores, el árbol 2 por el contrario, creció en un rodal muy denso. El árbol 3 creció primeramente en un rodal denso y reaccionó tras una clara fuerte a la edad de 6 años, aumentando notoriamente su crecimiento en diámetro.
El gran incremento en diámetro provocado por las claras, unido al aumento del
valor de los árboles individuales ligado a ella y a la mejora de la estabilidad del
rodal, llevo a que ya en el primer tercio del siglo XX se produjese un alejamiento de
la selvicultura tradicional centroeuropea de mantener áreas basimétricas elevadas.
Assmann (1961) formuló el principio del “mantenimiento del área basimétrica
óptima”1 (figura 5-12) en base a sus investigaciones sobre la influencia de diferentes
áreas basimétricas sobre el crecimiento del rodal. El área basimétrica máxima
corresponde a la máxima densidad que puede soportar el rodal en función de la
estación y la especie. El área basimétrica óptima es aquella para la cual el
crecimiento en volumen en función de la superficie disponible por árbol alcanza su
valor máximo. Según Assmann es crítico mantener un área basimétrica para la cual
se alcanza el 95% del crecimiento máximo en volumen. A causa del elevado riesgo
de derribamiento por el viento en rodales muy densos el área basimétrica óptima es
una variable objetivo que raramente se alcanza.
1 Para más detalle véase Kramer (1988) y Pretzsch (2001).

208 Crecimiento del Rodal
90
120
110
100
80 70 60
95
100
Incremento en volumen
%
%
“máximo“
“óptimo“
“crítico“
área basimétrica media relativa Figura 5-12. Representación esquemática del área basimétrica máxima, óptima y crítica (según
Assmann, 1961; ver Krammer, 1988 página 86). Según Thomasius y Thomasius (1976, 1978) el incremento en volumen en función
del espacio disponible por árbol en rodales de picea alcanza su máximo para una
densidad del rodal por debajo de la densidad máxima posible (Figura 5-13,
derecha). La relación entre el espacio y el crecimiento en volumen de árbol muestra,
como era de esperar, una tendencia asintótica (Figura 5-13, izquierda).
0,00 0,01 0,02 0,03 0,04 0,05 0,06 0,07
0 20 40 60 80 100 120
Espacio disponible m²/árbol
Incr
emen
to m
³/añ
o ár
bol
( )[ ]2,5-disponible espacio-0,075e-10,06=iv ⋅⋅
0,00
5,00
10,00
15,00
20,00
25,00
30,00
0 20 40 60 80 100 120
espacio m²/árbol
Incr
emen
to m
³/ha
año
( )[ ]2,5-espacio×-0,075e-1×espacio600=iV
Figura 5-13. Relaciones entre el espacio disponible y el incremento en volumen en un rodal de picea de 39 años; a la izquierda el crecimiento del árbol individual, a la derecha el incremento referido a la superficie según Thomasius y Thomasius (1976; ver también Schmidt-Vogt, 1986, página 142).
Un área basimétrica elevada supone un incremento en volumen elevado por unidad
de superficie, sin embargo disminuye la estabilidad del rodal y reduce el valor
individual de cada árbol (Kramer, 1988). Esta situación se observó ya hace tiempo
en plantaciones por lo que, aún asumiendo una pérdida de volumen considerable

Crecimiento del Rodal 209
puede ser preferible aplicar claras fuertes para mejorar a la larga la estabilidad del
rodal.
Ejemplo de modelos de crecimiento a nivel rodal
La diferencia fundamental entre un modelo dependiente de la densidad y unas
tablas de producción clásicas no está en la forma de representar la información,
puesto que una tabla de producción también se puede implementar en un programa
de ordenador, sino que se trata de una diferencia básica de concepto. Un modelo
de gestión dependiente de la densidad no está limitado a una única alternativa de
gestión, por el contrario, debido a su estructura es posible analizar diferentes
tratamientos selvícolas para un rodal real. Debido a la flexibilidad que deben tener
los componentes del modelo para poder simular diferentes situaciones, cada vez
adquiere más importancia la correcta utilización de los conocimientos existentes en
el proceso de desarrollo de estos modelos de simulación (Pretzsch y Bossel, 1988).
El desarrollo lógico de las tablas de producción se dirige a la construcción de
modelos que no estén condicionados a un número limitado de tratamientos
selvícolas predefinidos. Según Levins (1966) la bondad de un modelo biológico
viene determinada, entre otras cosas, por los atributos validez general y precisión. En el
desarrollo de modelos de crecimiento y de producción normalmente se sacrifica la
validez general en favor de la precisión, se trata de encontrar un modelo cuyas
simulaciones se asemejen lo máximo posible a los datos. Esta es una aspiración
lógica que conduce a descubrir nuevos modelos que en casos especiales pueden
alcanzar una extraordinaria precisión pero que sin embargo, debido a la falta de
validez general, no son fácilmente aplicables (Gadow, 1996).
En cada caso, el modelo a desarrollar debe adaptarse a las condiciones del
ecosistema objeto de estudio, por ejemplo, debido a la complicada dinámica de los
bosques mixtos y a la gran diversidad de tipos de rodales, en estos casos los
modelos más adecuados son los de árbol individual. Por otro lado, la ordenación de

210 Crecimiento del Rodal
montes requiere de modelos flexibles dependientes de la densidad que se puedan
emplear de forma sencilla en programas de optimización y para ello se suele
adoptar como modelo más sencillo el de rodal. De todos modos, sean cuales sean
las condiciones del ecosistema forestal, cuando el objetivo es la gestión el modelo
debe de permitir simular diferentes estados iniciales y tratamientos y, además sería
deseable que la producción de madera estimada se clasificara según su calidad y
destinos finales.
Las variables de rodal más importantes que deben ser consideradas en estos
modelos son la altura dominante, el número de pies y el área basimétrica (en
rodales mixtos la proporción correspondiente a cada especie) así como el tipo, la
intensidad y el momento de aplicar las claras. De este modo, los modelos
posibilitan no solamente pronosticar el desarrollo de un caso determinado, sino
que suministran información general sobre la relación entre el crecimiento del área
basimétrica, la densidad del rodal y la altura dominante. Por ejemplo, Hui y Gadow
(1993c) desarrollaron un modelo de área basimétrica para rodales puros de la
especie Cunninghamia lanceolata, que es una de las especies de mayor importancia
económica en China y que relaciona las tres variables antes comentadas.
4.2921H0.142
1H0.1421
2121
20.601
10.601
2
HH
NNGG ⎟⎟⎠
⎞⎜⎜⎝
⎛= −⋅⋅−
5-18
Donde G1 ,G2 ; N1, N2 y H1 y H2 son el área basimétrica (m²/ha), el número de
árboles por hectárea y la altura dominante (m) a las edades t1 y t2, respectivamente.
Ecuaciones como la anterior permiten pronosticar el área basimétrica futura. La
expresión del modelo como una ecuación en diferencias algebraicas garantiza el
cumplimiento de una serie de propiedades fundamentales entre las que destaca la
invarianza con respecto al camino de proyección, ya explicada al principio de este
capítulo.

Crecimiento del Rodal 211
Este tipo de modelos deben incluir en su formulación tanto el crecimiento natural
de una variable, es decir, el crecimiento cuando el rodal no se ve afectado por
perturbaciones externas y el efecto de los tratamientos en el crecimiento. Por
ejemplo, en la figura 5-14 se representan los dos procesos básicos que determinan
la evolución del área basimétrica y el número de pies por hectárea, por una parte el
crecimiento natural y por otra parte el cambio condicionado por la clara.
N e x t( t 1 )
t
G ( t 1 ) G ( t 2 )
G ( m ² / h a )
G e x t( t 1 )
)t-G ( t 21Δ
)t-N ( t 21Δ
N ( t 1 ) N ( t 2 )
N / h a
t 1 t 2
Figura 5-14. Secuencia esquemática de crecimiento y clara (Gext es el área basimétrica extraída en
la clara (m²/ha); G(t1) y G(t2) son el área basimétrica (m²/ha) a la edad t1 y t2,respectivamente; ΔG(t1-t2) es el incremento de área basimétrica debido al crecimiento (m²/ha); Nest es el número de pies por ha extraídos en la clara, N(t1) y N(t2) son el número de pies por ha a la edad t1 y t2, respectivamente e ∆N(t1-t2) es la reducción del número de pies debida a la mortalidad natural. En determinadas ocasiones se puede asumir que en aprovechamientos forestales intensos donde el período de tiempo entre claras es corto, ∆N(t1-t2) ≈ 0).
Un modelo como el 5-18 no se puede emplear para proyectar entre dos edades
cuando en el intervalo se realiza una clara. En estos casos se debe proyectar hasta la
edad de la clara, determinar como afecta el tratamiento al área basimétrica y a la
densidad y, partiendo de la nueva situación del rodal volver a proyectar hacia el
futuro. En el caso de que al especificar la clara sólo se indique el número de pies a
extraer o el área basimétrica a extraer es necesario establecer relaciones entre ambas

212 Crecimiento del Rodal
variables para estimar como afecta la clara ambas. Relaciones sencillas de estas
características se pueden obtener a partir de unas tablas de producción o de datos
de ensayos de claras. Por ejemplo, la ecuación 5-19 relaciona el área basimetrica y el
número de pies extraídos con los valores antes de la clara y un parámetro específico
del tipo de clara que debe estimarse.
δ
⎟⎠⎞
⎜⎝⎛=
NN
GG extext 5-19
Donde G es el área basimétrica antes de clara (m2/ha); Gext el área basimétrica
extraída (m2/ha); N el número de pies por ha antes de clara; Next el número de pies
por ha extraídos y δ un parámetro cuyo valor viene determinado por el tipo de clara
Por ejemplo, para picea en una clara de intensidad media resulta un valor de δ de
1,42 y para una clara de intensidad fuerte δ = 1,25 según las tablas de producción
de Wiedemann-Schober.
En los trabajos de Gurjanov et al. (2000) y Sánchez Orois et al. (2001) se
desarrolló un modelo de crecimiento para picea empleando los datos de 19 parcelas
permanentes del Centro de Investigación Forestal de Braunschweig2. En total se
contó con 228 períodos de observación con datos de las variables dasométricas
edad, altura dominante, área basimétrica y número de pies entre dos mediciones.
En la figura 5-15 se presenta el desarrollo de la altura dominante, el número de pies
y el área basimétrica en función de la edad así como el desarrollo del área
basimétrica en función de la altura dominante.
2 Los datos empleados ya por Schübeler (1997) fueron cedidos por el Centro de Investigación Forestal de la Baja
Sajonia.

Crecimiento del Rodal 213
Desarrollo de la altura dominante Desarrollo del área basimétrica
0 5
10 15 20 25 30 35 40
0 50 100 150
Edad(años)
Altu
ra d
omin
ante
(m)
0
10
20
30
40
50
60
70
0 20 40 60 80 100 120 140
Edad(años)
G(m
²/ha)
Desarrollo del número de pies Desarrollo del área basimétrica en función de la
altura dominante
0 1000 2000 3000 4000 5000
0 50 100 150
Edad(años)
Den
sida
d (p
ies/
ha)
0
10
20
30
40
50
60
70
0 5 10 15 20 25 30 35 40
Altura dominante (m)
G (m
2 /ha)
Figura 5-15. Representación gráfica de los datos de 228 intervalos de crecimiento obtenidos de 19 parcelas permanentes de ensayo de picea.
Las ecuaciones que estiman el crecimiento de una variable dasométrica o
dendrométrica se pueden clasificar en dos grandes grupos: i) modelos de
diferencias algebraicas basados en ecuaciones de crecimiento (por ejemplo la
propuesta por Schumacher, 1939), que determinan el valor de la variable a la edad
t2 en función del valor de esa misma variable a la edad t1 y de otro conjunto de
variables de rodal o de árbol a las edades inicial y final de la estimación (ver por
ejemplo Souter, 1986 o Forss, 1994); ii) ecuaciones diferenciales que estiman el
incremento anual (o periódico) de la variable analizada. Ejemplos de este tipo son
los modelos que se basan en el denominado “state space approach” propuesto por
García (1994) (ver por ejemplo los modelos de Rodríguez Soalleiro et al., 1995;
Kvist Johannsen, 1999 o Castedo et al., 2007).

214 Crecimiento del Rodal
En el modelo de picea se optó por analizar la adecuación de diferentes modelos en
diferencias algebraicas para rodales y las ecuaciones obtenidas se presentan en la
tabla 5-11.
Área basimétrica ⎟⎟
⎠
⎞
⎜⎜
⎝
⎛
⎟⎟⎠
⎞⎜⎜⎝
⎛⋅⎟
⎟⎠
⎞⎜⎜⎝
⎛−⋅+⎟
⎟⎠
⎞⎜⎜⎝
⎛−⋅⋅⎟
⎟⎠
⎞⎜⎜⎝
⎛−⋅+⋅⎟
⎟⎠
⎞⎜⎜⎝
⎛= 1
2
12
2
1
2
11
2
1 lnNlnN00681-01SI0112-0153575lnGexpGtt
tt
tt
tt ,,,2
Altura dominante 507.1
,,
⎟⎟⎠
⎞⎜⎜⎝
⎛⋅⋅−−
⋅⋅−−⋅=
A)10000060exp(1A)SI00060exp(1SIH
Altura media 12061m H59810H ,, ⋅=
Factor de forma ( ) ( )201063,09401,0258,4 mm HHFH ⋅−⋅+−=
Volumen FHGV ⋅=
Mortalidad 618365-0
1
34198321
34198322
618365-01 )H(H0002640
1000N1000N
,,,
,
2 ,⎥⎥⎦
⎤
⎢⎢⎣
⎡−⋅+⎟
⎠⎞
⎜⎝⎛⋅=
Tabla 5-11. Modelo de crecimiento para rodales regulares de picea desarrollado en base de los trabajos de Gurjanov et al. (2000) y Sánchez Orois et al. (2001); t=edad del rodal; SI=índice de sitio (edad de referencia 100; m); G=área basimétrica (m2/ha); N=árboles por ha; H=altura dominante (m); Hm=altura media (m); FH=factor de forma (m); V=volumen por ha (m3/ha).
Este modelo estima el área basimétrica, el número de pies vivos y la altura
dominante a partir de los datos iniciales y se adecua muy bien para generar
diferentes opciones selvícolas; además, tiene una ventaja sobre los modelos de
árbol individual que es su mayor robustez3.
Volumen y clasificación de la calidad
El volumen no es una variable originaria del inventario sino que es un valor que se
puede calcular a partir de otras variables como el área basimétrica, el número de
pies y la altura dominante. En ocasiones también se calcula el volumen a partir del
área basimétrica y el factor de forma (Franz et al., 1973). El volumen es, por así
3 Véase por ejemplo la evaluación de modelos de Windhager (1999) que demuestra las incertidumbres en el pronósti-
co de las alturas individuales.

Crecimiento del Rodal 215
decirlo, un producto secundario de la información proporcionada por el inventario.
Por lo tanto es especialmente importante una estimación exacta de las variables
originales área basimétrica, número de pies y altura dominante.
Para la toma de decisiones desde el punto de vista económico es
imprescindible la estimación del volumen y la clasificación de la calidad y de los
productos finales. Con este fin se pueden emplear funciones de perfil, que permiten
estimar la forma del tronco de un árbol concreto y, por integración obtener su
volumen, o se pueden emplear funciones de razón, que permiten obtener el
volumen hasta un cierto diámetro en punta delgada en función del volumen total.
Una de las diferencias entre las funciones de perfil y las tarifas de razón es que estas
últimas se pueden construir tanto para árbol individual como para rodal. Un
ejemplo de función de razón para rodal que permite obtener el volumen de un
rodal hasta un determinado diámetro en punta delgada m es la propuesta por
Amateis et al. (1986):
⎟⎟⎟
⎠
⎞
⎜⎜⎜
⎝
⎛⋅=
⎟⎟⎠
⎞⎜⎜⎝
⎛+⎟⎟
⎠
⎞⎜⎜⎝
⎛ 4
3
2
1
γγ
γγdgd
dgm
m,d eVV 5-20
donde Vm,d es el volumen del rodal hasta un diámetro en punta delgada m; V es el
volumen total del rodal; dg es su diámetro medio cuadrático; d es el diámetro medio
cuadrático mínimo para que en le rodal haya trozas con un diámetro mínimo en
punta delgada igual a m y γ1, γ2, γ3 y γ4 son los parámetros del modelo.
La ecuación 5-20 es válida tanto para la clasificación de rodales como de
árboles individuales (ver por ejemplo Trincado y Gadow, 1996). Para cada especie
resultaron dos conjuntos de parámetros, un conjunto para su empleo en el modelo
de árbol individual y un segundo conjunto para su empleo en el modelo de rodal.
En general se puede estimar el diámetro medio cuadrático mínimo (d) que debe
tener el rodal para obtener trozas con un diámetro en punta delgada igual a m
mediante un modelo lineal simple.
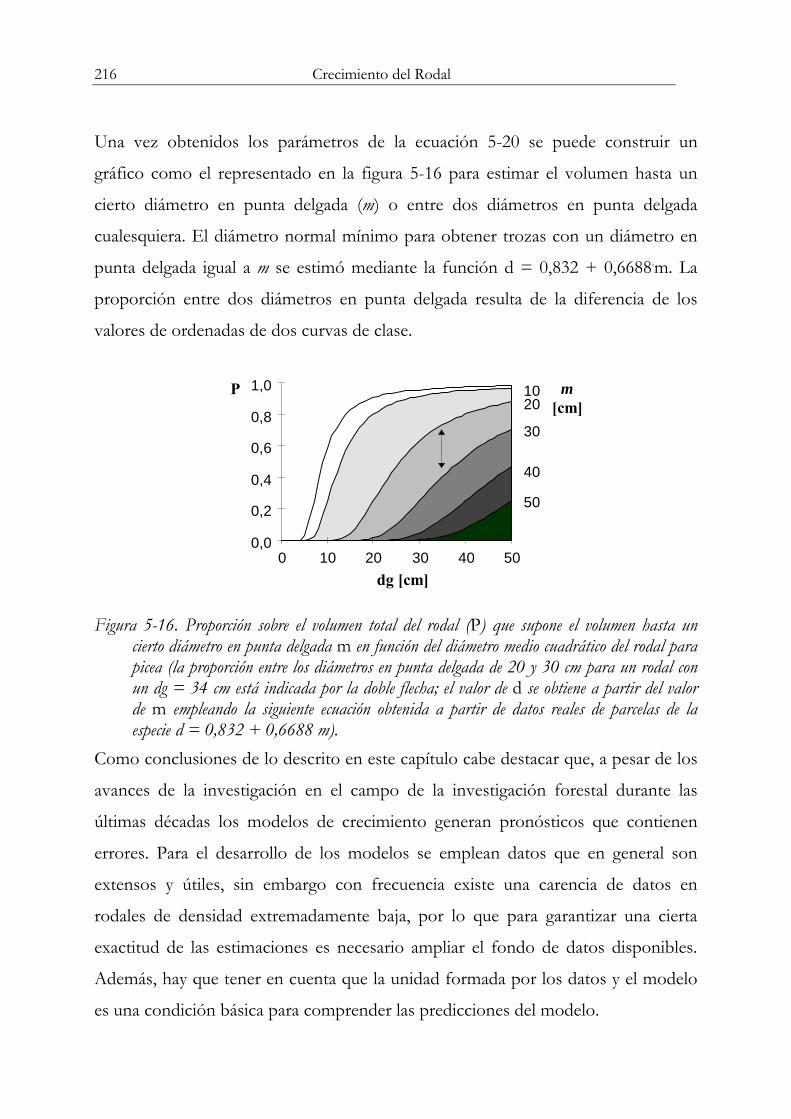
216 Crecimiento del Rodal
Una vez obtenidos los parámetros de la ecuación 5-20 se puede construir un
gráfico como el representado en la figura 5-16 para estimar el volumen hasta un
cierto diámetro en punta delgada (m) o entre dos diámetros en punta delgada
cualesquiera. El diámetro normal mínimo para obtener trozas con un diámetro en
punta delgada igual a m se estimó mediante la función d = 0,832 + 0,6688.m. La
proporción entre dos diámetros en punta delgada resulta de la diferencia de los
valores de ordenadas de dos curvas de clase.
0,0
0,2
0,4
0,6
0,8
1,0
0 10 20 30 40 50dg [cm]
P 10 20
30
40
50
m [cm]
Figura 5-16. Proporción sobre el volumen total del rodal (P) que supone el volumen hasta un
cierto diámetro en punta delgada m en función del diámetro medio cuadrático del rodal para picea (la proporción entre los diámetros en punta delgada de 20 y 30 cm para un rodal con un dg = 34 cm está indicada por la doble flecha; el valor de d se obtiene a partir del valor de m empleando la siguiente ecuación obtenida a partir de datos reales de parcelas de la especie d = 0,832 + 0,6688 m).
Como conclusiones de lo descrito en este capítulo cabe destacar que, a pesar de los
avances de la investigación en el campo de la investigación forestal durante las
últimas décadas los modelos de crecimiento generan pronósticos que contienen
errores. Para el desarrollo de los modelos se emplean datos que en general son
extensos y útiles, sin embargo con frecuencia existe una carencia de datos en
rodales de densidad extremadamente baja, por lo que para garantizar una cierta
exactitud de las estimaciones es necesario ampliar el fondo de datos disponibles.
Además, hay que tener en cuenta que la unidad formada por los datos y el modelo
es una condición básica para comprender las predicciones del modelo.

Crecimiento del Rodal 217
Los modelos dependientes de la densidad suponen una mejora importante de la
información aportada por las tablas de producción y al mismo tiempo asumen su
función como modelos de referencia. Son flexibles y además son fáciles de manejar
por los usuarios finales como las tablas de producción. Sus pronósticos se pueden
comprobar de forma relativamente fácil, y las claras se pueden definir y simular de
forma más sencilla que con los modelos de árbol individual, permitiendo realizar
evaluaciones para la selección de la mejor alternativa silvícola en función de uno o
varios objetivo de producción (Cañellas y Montero, 2002; Cañellas et al., 2004;
Rodríguez Soalleiro et al., 2000, 2002 y 2007 y Balboa et al., 2006). Por tanto, se
puede afirmar que un modelo de rodal dependiente de la densidad aúna las ventajas
de las tablas de producción y de los modelos de árbol individual. Sin embargo, aún
es necesario un esfuerzo para conseguir reducir el error de estimación de estos
modelos mediante la ampliación continua y bien dirigida de los datos y la
cooperación entre centros de investigación para obtener información sobre el
crecimiento natural para intervalos de n períodos vegetativos donde no se hayan
realizado claras y el efecto de las claras sobre el crecimiento del rodal en contraste
con el crecimiento natural.

218 Crecimiento del Rodal

Crecimiento del Árbol 219
Crecimiento del Árbol
Los árboles tienen la capacidad de reaccionar a los cambios que se producen en su
entorno. Esta capacidad se muestra en la dinámica de su crecimiento, es decir, en la
reacción a los cambios desde el estado inicial determinados por las condiciones de
la estación y la competencia. Las estimaciones sobre el crecimiento de los árboles
deben ser fiables y a su vez tan detalladas como sea posible. Estos requisitos no son
fáciles de cumplir ya que los estados iniciales son muy diversos y las reacciones
posibles son también muy numerosas. El plantear como objetivo obtener una
elevada precisión en un modelo puede llevar a una situación que Kimmins (1997)
denomino reduccionista en la que la construcción de modelos cada vez de mayor
resolución puede conducir a un resultado final que tenga muy difícil aplicación
práctica (Figura 6-1).

Crecimiento del Árbol 220
Por lo tanto, es lógico que el modelo empleado armonice con la información
disponible. Si se dispone por ejemplo de datos de árboles individuales incluidas sus
posiciones, entonces es razonable utilizar esta información para simular el
crecimiento empleando un modelo de árbol individual dependiente de la posición.
Si por el contrario sólo se dispone de datos de rodales, entonces se debe emplear
un modelo de rodal y si la información de que se dispone es altamente agregada,
por ejemplo datos muy globales sobre un monte o un área geográfica muy amplia,
entonces la única posibilidad de realizar un pronóstico es mediante un modelo de
producción regional.
Modelo original
Modelo ampliado y mejorado
Modelo ampliado poco práctico
Modelo ampliado y mejorado
Modelo ampliado inútil
Modelo de rodal
(Área basimétrica; altura dominante; n° de pies)
Modelo de árbol individual
(Diámetro y altura de los árboles)
Modelo de árbol representativo
(Distribución diamétrica; regresión d/h)
Modelo de producción regional
(Volumen de madera)
Figura 6-1. Izquierda: Típico desarrollo de modelos reduccionista (Kimmins, 1997). Derecha: Sistema modular estructurado jerárquicamente de los modelos de crecimiento según su resolución.
Un conjunto de modelos jerárquizados que, en función de la resolución de los
datos aporta diferentes informaciones, se puede comparar con un microscopio con
el que se pueden reconocer más detalles a medida que se van intercalando más
aumentos. La característica fundamental de este conjunto de modelos jerarquizados
es que los modelos de diferente resolución tienen que generar el mismo resultado
de una variable concreta, es decir, los modelos deben ser compatibles (Burkhart,
1987). En los puntos de solape entre las estimaciones de modelos de diferente

Crecimiento del Árbol 221
resolución debe haber una coincidencia entre las estimaciones. Aunque este
principio jerárquico es fundamental, en la práctica no se ha podido aplicar de forma
generalizada pese a los avances en las técnicas de modelización.
En este principio de jerarquía hay que tener en cuenta que los procesos
biológicos de un nivel jerárquico determinado se rigen por los procesos de los
niveles inferiores de más resolución y están también limitados por las condiciones
de los niveles superiores. Por ejemplo, el crecimiento de un árbol concreto
simulado por un modelo de árbol individual esta regulado por una serie de
procesos fisiológicos que pueden ser simulados con un modelo de proceso pero, a
su vez las condiciones del rodal, simuladas mediante un modelo más general de
rodal, también limitan las posibilidades de desarrollo del árbol.
Modelos de árbol representativo Como ya se ha comentado en ocasiones anteriores, los modelos de crecimiento se
pueden clasificar en diferentes categorías según el elemento base de la simulación,
así por ejemplo los modelos de crecimiento de rodal aporta información sobre
valores medios de las principales variables del rodal referidos a superficies (área
basimétrica, número de pies por ha, volumen). Existe otro grupo de modelos en los
que el elemento base de la simulación es un árbol que representa las carácterísticas
medias de un grupo de árboles del rodal. Estos modelos, denominados modelos de
árbol representativo se pueden emplear cuando se dispone de una distribución de
los árboles en función de alguna caracteríostica como el diámetro, la sección, la
altura o el volumen. Si la variable de la que se conoce la distribución es el diámetro,
se puede considerar como árbol representativo de cada clase diamétrica al árbol
cuyo diámetro es el centro de la clase. Como los diámetros son variables sencillas
de medir, los modelos de árbol representativo suelen basarse en esta variable para
agrupar los árboles del rodal. Estos modelos de árbol representativo tienen mayor

Crecimiento del Árbol 222
resolución que los modelos de rodal y desempeñan un papel central en la
investigación del crecimiento orientado a la práctica.
Estimación futura de las distribuciones diamétricas
Es muy frecuentemente que se conozcan los diámetros de los árboles de un rodal,
pero en muchas ocasiones no se cuenta con datos de su edad. Este es el caso por
ejemplo de los rodales irregulares en los que existen árboles de diferentes edades.
En estos casos es necesario emplear metodologías de estimación del crecimiento
diamétrico que sean independiente de la edad del árbol. Un método que se emplea
frecuentemente en rodales irregulares se basa en la estimación de la distribución
diamétrica futura mediante matrices de transición. La estructura de estos modelos
es muy sencilla y se esquematiza como:
⎥⎥⎥⎥⎥⎥⎥⎥
⎦
⎤
⎢⎢⎢⎢⎢⎢⎢⎢
⎣
⎡
+
⎥⎥⎥⎥⎥⎥⎥⎥
⎦
⎤
⎢⎢⎢⎢⎢⎢⎢⎢
⎣
⎡
⋅
⎥⎥⎥⎥⎥⎥⎥⎥
⎦
⎤
⎢⎢⎢⎢⎢⎢⎢⎢
⎣
⎡
=
⎥⎥⎥⎥⎥⎥⎥⎥
⎦
⎤
⎢⎢⎢⎢⎢⎢⎢⎢
⎣
⎡
0..00
.
.
...0..
0...00...0...0
.
.
1
13
12
11
2
21
1
2
23
22
21 C
n
nnn
a
bab
a
n
nnn
mmm
Donde n2i es el número de árboles que en el momento t2 pertenecen a la clase
diamétrica i; a i es la probabilidad de que un árbol permanezca en la clase i en el
intervalo de tiempo analizado; b i es la probabilidad de paso o probabilidad de que
un árbol de la clase i pase a la clase diamétrica siguiente en el mismo intervalo de
tiempo; n1i es el número de árboles que en el momento t1 pertenecen a la clase
diamétrica i, y C es la incorporación o número de árboles jóvenes que entran a
formar parte de la clase diamétrica menor en el intervalo de tiempo analizado. En
este modelo también se puede tener en cuenta la mortalidad, de modo que cuando
dentro de la matriz central la suma de probabilidades no sea igual a 1 es que existe
mortalidad.

Crecimiento del Árbol 223
La probabilidad de paso de los árboles de la clase diamétrica i a la siguiente clase
diamétrica j en un período de tiempo determinado mrij (en inglés movement ratio) se
puede calcular a partir del crecimiento diamétrico medio en la clase i (Δdi cm) y de
la amplitud de la clase diamétrica (∆ en cm) empleando la siguiente expresión
Δ⋅= 100iij Δdmr . Por ejemplo, si el crecimiento diamétrico medio en la clase i
cuya amplitud de clase es 4 cm, es de 1,5 cm en 2 años, entonces resulta que, bajo la
condición de que los diámetros en la clase i se reparten homogéneamente mrij =
1,5·100/4 = 37,5, es decir, el 37,5% del total de árboles de la clase diamétrica i
pasan en dos años a la clase diamétrica siguiente.
Como ejemplo de aplicación de los modelos matriciales descritos considerese
la siguiente tabla en la que se muestran los datos de un rodal hipotético con 21
árboles que se distribuyen en 5 clases diamétricas con unos crecimientos conocidos
(∆di), donde n1 es el número de pies inicial y n2 es el número de pies después de un
intervalo de tiempo determinado:
di n1 ∆di mrij n2
14 4 0,5 0,25 3 16 8 1,0 0,50 5 18 5 0,8 0,40 7 20 4 0,5 0,25 5 22 0 0,4 0,20 1 21 21
La tabla anterior también se puede presentar en forma de matriz de transición en la
que no existe incorporación al clase inferior ni mortalidad (la suma de todas las
columnas de la matríz es igual a 1).
⎥⎥⎥⎥⎥⎥
⎦
⎤
⎢⎢⎢⎢⎢⎢
⎣
⎡
=
⎥⎥⎥⎥⎥⎥
⎦
⎤
⎢⎢⎢⎢⎢⎢
⎣
⎡
⋅
⎥⎥⎥⎥⎥⎥
⎦
⎤
⎢⎢⎢⎢⎢⎢
⎣
⎡
15753
04584
80,025,0000075,040,0000060,050,0000050,025,0000075,0

Crecimiento del Árbol 224
En el caso de que exista una mortalidad de un 5% en cada una de las clases
diamétricas y haya una incorporación de 4 árboles a la clase diamétrica inferior
entonces el resultado sería el siguiente:
⎥⎥⎥⎥⎥⎥
⎦
⎤
⎢⎢⎢⎢⎢⎢
⎣
⎡
=
⎥⎥⎥⎥⎥⎥
⎦
⎤
⎢⎢⎢⎢⎢⎢
⎣
⎡
+
⎥⎥⎥⎥⎥⎥
⎦
⎤
⎢⎢⎢⎢⎢⎢
⎣
⎡
⋅
⎥⎥⎥⎥⎥⎥
⎦
⎤
⎢⎢⎢⎢⎢⎢
⎣
⎡
18,47,66,48,6
00004
04584
75,025,0000070,040,0000055,050,0000045,025,0000070,0
Algunas de las primeras contribuciones al desarrollo de modelos matriciales de
transición son los trabajos de Rudra (1968), Suzuki (1971), Moser (1974) y Sloboda
(1976). Un ejemplo de ecuación que relaciona la probabilidad de paso de una clase
diamétrica a otra con variables del rodal es la propuesta por Kolström (1992, ver
Pukkala y Kolström, 1988) para rodales irregulares de pícea en Finlandia. La
ecuación obtenida por este autor estima la probabilidad bi de que un árbol que se
encuentra en la clase diamétrica i pase a la clase siguiente en un intervalo de tiempo
de cinco años y para una amplitud de clase de 4 cm.
( ) ( ) ii dG,G,d,,i=eb ⋅⋅−⋅−⋅+− 00070ln550ln86012
6-1
La probabilidad de paso depende del diámetro al principio del período de
crecimiento (di en cm) y del área basimétrica del rodal (G en m2/ha).
El diámetro a la altura de pecho es una variable que se mide frecuentemente y
que se correlaciona con el valor y el volumen de un árbol; el diámetro constituye
por lo tanto una base importante para las decisiones selvícolas y económicas. Por
este motivo la información sobre la distribución diamétrica que se utiliza para
pronosticar el desarrollo forestal es muy anhelada y al mismo tiempo relativamente
fácil de obtener (Kennel, 1972a; Gadow, 1987; Gerold, 1990; Puumalainen, 1996).

Crecimiento del Árbol 225
Un ejemplo de China A parte de la descripción estadística de las distribuciones diamétricas el pronóstico
del desarrollo forestal mediante funciones de distribución continuas es un método
muy empleado en la investigación del crecimiento forestal1. En primer lugar se
determinan los parámetros de la distribución para el mayor número posible de
rodales con diferentes calidades de estación y diferentes tratamientos. En un
segundo paso se buscan las relaciones estadísticas entre los valores de los
parámetros de la distribución y los atributos del rodal. Este procedimiento se aclara
mediante la función Weibull.
c
bax
eF(x)⎟⎠⎞
⎜⎝⎛ −
−−=1 6-2
Donde F(x) es la probabilidad de que un árbol tenga un diámetro igual o inferior a
x y a, b y c son los parámetros de posición, escala y forma de la distribución.
Una de las primeras aplicaciones del método de estimación de parámetros es el
trabajo de Smalley y Bailey (1974). Las funciones para estimar los parámetros de la
función Weibull son las siguientes:
⎪⎩
⎪⎨
⎧
>
≤⋅+−=
PiesH,
PiesHH,a
260
260757,09492,1 6-3
(H)/N)(Nab 103 log2046,6101955,10003,02352,5 ++⋅−−−= 6-4
NHc ⋅−⋅−= 0006,00391,00560,6 6-5
Siendo H la altura dominante del rodal (pies) y N el número de pies por acre. Un
método similar es el empleado por Nagel y Biging (1995). Una variante especial del
método de estimación de parámetros es el denominado método de recuperación de
parámetros, según el cual los parámetros se derivan directamente de los valores
1 Las funciones de distrubicion diamétrica más empleadas son la función Beta-(Swindel et al., 1987; Maltamo et al.
1995); Weibull-(Bailey y Dell, 1973) y la función Sb de Johnson (Hafley y Schreuder, 1977).

Crecimiento del Árbol 226
medios de la distribución y éstos de relaciones con variables de masa (Hyink, 1979;
Hyink y Moser, 1983). Hui y Gadow (1996) emplearon para ello la función logística
bxaex)P(XF(x) −+=≤=
11
6-6
Con b = 2,1972/(XF=0,9 - XF=0,5); a = -2,1972+b(XF=0,9) siendo XF=0,9 y XF=0,5 los
percentiles del 90 y el 50% de la distribución diamétrica, respectivamente y cuyos
valores se obtienen empleando las siguientes expresiones empíricas que dependen
de la altura dominante (H en metros) y el diámetro medio cuadrático (dg en cm) del
rodal : y 1403,0504,12762,050 4043,0
−⋅= ⋅= oH
g.F dHX00668,07888,01671,0
90 2963,1−⋅
= ⋅= oHg.F dHX
La figura 6-2 muestra dos distribuciones diamétricas generadas en base a los
valores medios del rodal. La forma de la distribución diamétrica no solo viene
determinada por el crecimiento de los árboles sino también por las operaciones
forestales. Por ese motivo la estimación de los parámetros de la distribución tras
una clara es muy importante para pronosticar la distribución diamétrica. Un
ejemplo son las funciones de estimación de Álvarez González (1997) para
pronosticar la modificación del parámetro b y c después de clara en rodales de Pinus
pinaster en España.2
ac
ext
ac
extacdc G
GNNbb 617,7335,850205,17067,4 −+⋅+−= 6-7
ac
ext
ac
extacdc G
GNNcc 255,5170,8178,1059,1 −+⋅+−= 6-8
Donde bdc, cdc, bac y cac son los valores de los parámetros de escala y forma de la
función de Weibull antes (ac) y después de clara (dc), respectivamente; Next y Nac son
los valores de la densidad extraída y antes de la clara, respectivamente y Gext y Gac
son los valores del área basimétrica extraída y antes de la clara, respectivamente.
2 Metodos similares para describir las consecuencias de las claras se encuentran en los trabajos de Römisch (1983),
Faber (1987), Murray y Gadow (1991), Chikumbo et al. (1992), Kassier (1993).

Crecimiento del Árbol 227
Un ejemplo de Chile Los modelos de árbol representativo no han tenido hasta ahora tanto protagonismo
en Alemania como los modelos de árbol individual. Su aplicación por ejemplo en el
análisis de tratamientos selvícolas en la ordenación de montes es sin embargo muy
prometedora.
Entre los métodos más acreditados del pronóstico de distribuciones
diamétricas se encuentra el de Pienaar y Harrison (1988); Nepal y Somers (1992);
Cao y Baldwin (1999); Trincado et al. (2002). La fig. 6-3 muestra el ejemplo del
pronóstico de la distribución diamétrica de la especie Eucalyptus nitens en Chile.
0
40
80 120
160
200
2 6 10 14 18 22 26 30 34 38 42 46
Clase diamétrica (cm)
Nº d
e pi
es p
or h
a
8 años 13 años
Figura 6-3. Ejemplo del pronóstico de la distribución diamétrica para un rodal de Eucalyptus
nitens en Chile (Trincado et al., 2002). La utilización de este método requiere la estimación de la tasa de supervivencia y
del incremento diamétrico para las diferentes clases diamétricas. Después de
realizar la predicción se corrigen las frecuencias de diámetros hasta que el resultado
sea congruente con el valor del área basimétrica proyectada. El método se adapta
por lo tanto para rodales con distribución diamétrica multimodal.
Un ejemplo de Sudáfrica Sánchez et al. (2003) siguieron un método similar. Los datos para este análisis
provinieron de cuatro parcelas de ensayo de la especie Eucalyptus grandis en
Sudáfrica que se midieron 8 veces en un intervalo de tiempo de 36 años. El período
de tiempo entre dos mediciones varió entre 2 y 5 años. Se dispone de la

Crecimiento del Árbol 228
distribución diamétrica para cada medición. Los datos dasométricos más
importantes se muestran en la tabla 6-1. La aplicación del método de Nepal y
Somers (1992) requiere estimar la tasa de supervivencia y el crecimiento en
diámetro para cada clase diamétrica. Después de predecir el crecimiento diamétrico
se corrigen las frecuencias diamétricas hasta que el resultado sea congruente con el
área basimétrica proyectada. En la figura 6-4 se aclara este método paso a paso.
Parcela Edad(años) Nº de pies por ha
Altura dominante (m)
Área basimétrica
(m2/ha) 1 2,08 6091 6,5 13,9 1 5,17 3998 21,1 40,1 1 10,17 3379 32,1 55,0 1 14,17 2984 38,2 60,0 1 20,17 1846 42,3 56,3 1 24,58 1451 48,9 60,8 1 27,83 1352 51,1 64,8 1 38,90 874 57,1 73,0 3 2,08 2734 6,5 8,5 3 5,17 2471 21,1 35,9 3 10,17 2026 32,1 47,6 3 14,17 1960 38,2 56,7 3 20,17 1639 42,3 60,6 3 24,58 1310 48,9 60,8 3 27,83 1244 51,1 63,5 3 38,90 799 57,1 69,2 4 2,08 1474 6,5 4,5 4 5,17 1441 21,1 29,7 4 10,17 1400 32,1 45,3 4 14,17 1391 38,2 53,8 4 20,17 1202 42,3 58,1 4 24,58 1112 48,9 63,2 4 27,83 1037 51,1 62,9 4 38,90 659 57,1 63,8 5 2,08 988 6,5 2,8 5 5,17 988 21,1 26,0 5 10,17 856 32,1 37,1 5 14,17 856 38,2 46,1 5 20,17 840 42,3 55,5 5 24,58 799 48,9 60,3 5 27,83 790 51,1 64,3 5 38,90 576 57,1 63,6
Tabla 6- 1. Datos dasométricos de las parcelas de ensayo. En un primer paso se estiman los parámetros de la distribución Weibull. A partir de
la distribución Weibull se deriva una función para estimar el crecimiento en

Crecimiento del Árbol 229
diámetro (Bailey, 1980); así se puede calcular la distribución diamétrica en un
momento futuro t2. Mediante un algoritmo se ajusta la distribución estimada para
garantizar la coincidencia entre los valores de área basimétrica y número de pies
estimados y observados (o calculados mediante un modelo de crecimiento).
Los parámetros de la distribución Weibull se calculan mediante el método de
recuperación de parámetros. Ejemplos de aplicación de este método se encuentran en
Hyink (1980), Hyink y Moser (1983), Yang y Feng (1989), Meng (1991), Hui et al.
(1994), Trincado et al. (2003). La metodología presentada en este trabajo fue
propuesta por Cao et al. (1982). Según este método se estiman los parámetros de la
función Weibull mediante el diámetro medio y el diámetro medio cuadrático.
Estimación de los parámetros Weibull en el momento t1 y t2
(método de recuperación de parámetros)
Estimación del crecimiento en diámetro
Algoritmo para ajustar el crecimiento
y la mortalidad
Distribución diamétrica estimada
Figura 6-4. Diagrama de flujo del método de Nepal y Somers (1992) para la predicción
compatible de distribuciones diamétricas. El parámetro de localización a se obtiene a partir del diámetro mínimo de la
distribución (dmin en cm) y del tamaño de clase diamétrica (∆ en cm) según la
siguiente ecuación:
2/min Δ−= da 6-9
El parámetro de escala b se despeja de la ecuación 6-10 y se sustituye en la ecuación
6-11 de tal manera que se puede calcular el parámetro de forma c :

Crecimiento del Árbol 230
)/11()( cbad +Γ⋅=− 6-10
))/21(())/11(2( 22 cbcbaadg +Γ⋅++Γ⋅⋅+⋅= 6-11
Siendo d y dg los diámetros medio aritmético y medio cuadrático de la distribución
y es la función gamma, cuyos valores están tabulados. )(xΓ
Para estimar el crecimiento diamétrico se empleó la función propuesta por
Bailey (1980). Suponiendo que la distribución diamétrica en los momentos t1 y t2 se
aproxima a la distribución Weibull y bajo el supuesto de que la dimensión relativa
de un árbol en relación a los demás permanece invariable se puede calcular el
crecimiento individual en diámetro mediante la ecuación 6-12.
21
1
11222
cc
badbad ⎟⎟⎠
⎞⎜⎜⎝
⎛ −⋅+= 6-12
Donde d1 y d2 son los diámetros del árbol en los momentos t1 y t2 y ai, bi y ci son los
valores de los parámetros de la función de Weibull en esos dos mismos instantes.
Por motivos de cálculo es más práctico realizar el cálculo en sentido inverso
para calcular el número de árboles en cada clase diamétrica futura. La ecuación 6-13
se puede expresar de la siguiente forma:
12
2
22111
cc
badbad ⎟⎟⎠
⎞⎜⎜⎝
⎛ −⋅+= 6-13
Ya que las distribuciones diamétricas se aproximan a la distribución de Weibull se
puede admitir que la distribución dentro de una clase diamétrica se puede
aproximar mediante la distribución de Weibull truncada. Por lo tanto el número de
árboles que pasa a la clase diamétrica futura se puede obtener de la ecuación 6-14.
( )[ ]( ) ( )[ ]( )⎟⎟⎠
⎞⎜⎜⎝
⎛
−−
= ∑ )()(),(max),(min
1111
21121112
jj
ijiiji
jji LFUF
LLdFUUdFNN
6-14

Crecimiento del Árbol 231
Siendo N2i el número de pies estimados en el momento 2 en la clase diamétrica i;
N1j el número de pies observado en el momento 1 en la clase diamétrica j; L2i el
límite inferior de la clase diamétrica i en el momento 2; U2i el límite superior de la
clase diamétrica i en el momento 2; L1j el límite inferior de la clase diamétrica j en el
momento 1; U1j el límite superior de la clase diamétrica j en el momento 1; d1() es el
diámetro en el momento 1 que se corresponde con el diámetro d2()en el momento
2 y F1() es la función de distribución de Weibull en el momento 1.
Si no se considera la mortalidad, el resultado es una distribución diamétrica
estimada que tiene el mismo número de pies en el momento t1 y en el momento
futuro t2. Esa distribución diamétrica tiene que ser ajustada, por lo tanto es
necesario multiplicar cada diámetro por un factor Pi, para garantizar la
compatibilidad del número de pies y área basimétrica estimados con los valores
observados (o calculados mediante un modelo de crecimiento).
21
NnP i
k
ii =⋅∑
=
6-15
22
1GdnPg ii
k
ii =⋅⋅⋅∑
=
6-16
Donde ni es el número de pies predichos en la clase diamétrica i; di es el centro de la
clase diamétrica i; k es el número de clases diamétricas; N2 y G2 son el número pies
y el área basimétrica en el momento t2, respectivamente y Pi es una función del
diámetro que se puede describir de la siguiente manera:
idi eP ⋅⋅= 1
0αα 6-17
Pi en la ecuación 6-17 solo puede tomar valores positivos (valores negativos
conducirían a valores negativos en la distribución ajustada). Además se trata de una
función monótona creciente, de tal manera que durante el proceso de ajuste la
estructura de la distribución original se mantiene constante.

Crecimiento del Árbol 232
El parámetro α0 solo puede tomar valores positivos, ya que sino Pi sería negativo.
El valor del parámetro α1 depende del ajuste. Si no se produce mortalidad el factor
Pi solo modifica el crecimiento diamétrico. En este caso α0 es mayor (o menor) que
1 y α1 es mayor (o menor) que 0 si el área basimétrica debe de ser ajustada hacia
arriba (o hacia abajo). Un valor positivo del parámetro α1 implica en rodales con
mortalidad, un mayor número de árboles que pasan de las clases diamétricas
menores a las superiores. Un valor negativo del parámetro α1 implica que solo los
árboles de las clases diamétricas superiores pasen a la siguiente clase diamétrica. Si
α1=0 la proporción de árboles que pasan a la clase siguiente es la misma para todas
las clases diamétricas. Si se sustituye la ecuación 6-17 en las ecuaciones 6-16 y 6-15
resulta:
( ) 21
01 Nne i
k
i
d i =⋅⋅∑=
⋅αα 6-18
( ) 22
10
1 Gdneg ii
k
i
d i =⋅⋅⋅⋅∑=
⋅αα 6-19
De las ecuaciones 6-18 y 6-19 resulta la ecuación siguiente:
( )( )
21
1
22
0
1
1
Gne
Ndnegk
ik
ii
d
iid
i
i
=⋅
⋅⋅⋅⋅⋅∑∑=
=
⋅
⋅
α
αα 6-20
Para calcular los parámetros α1 y α0 se resuelve la ecuación 6-20 por un
procedimiento iterativo.
Mediante programas estadísticos se resuelve la ecuación 6-20 y se obtienen los
parámetros α0 y α1. Un ejemplo de las distribuciones observadas y estimadas y el
factor Pi y las correspondientes distribuciones diamétricas corregidas para la parcela
3 se muestran en la tabla 6-2. El rodal estimado tiene un área basimétrica de 2,9 m2
y un número de pies de 224 (la superficie de la parcela es de 600 m2). La
distribución estimada tiene que ser ajustada para obtener un valor de área
basimétrica de 2,7 m2 y un número de pies de 206. Los valores α1=0,0095 y

Crecimiento del Árbol 233
α0=0,818627 son la solución del sistema 10-11. A partir de estos dos parámetros se
calcula el factor Pi para cada una de las clases diamétricas.
Clase
diamétrica (cm)
Distribución diamétrica observada
Distribución diamétrica estimada
PiDistribución
diamétrica corregida
1 0 0,0 0,83 0,0 3 7 4,5 0,84 3,8 5 12 11,5 0,86 9,8 7 10 12,7 0,87 11,1 9 38 36,6 0,89 32,7
11 29 41,6 0,91 37,8 13 36 41,9 0,93 38,8 15 30 39,6 0,94 37,4 17 27 24,2 0,96 23,3 19 12 6,3 0,98 6,1 21 4 4,3 1,00 4,3 23 1 0,5 1,02 0,5 25 0 0,4 1,04 0,4 27 0 0,0 1,06 0,0
206 224 206
Tabla 6-2. Distribuciones diamétricas observadas y estimadas para la parcela nº 3. La predicción de la distribución diamétrica se ejemplifica en la figura 6-5 en base a
dos mediciones consecutivas de la parcela 3.
0
5
10
15
20
25
30
1 3 5 7 9 11 13 15 17 19 21 23 25 27 29 31 33 35 37 39 Clase diamétrica (cm)
Nº
de
pie
s
Distribución observada a los 10 añosDistribución estimada A los 14 años
Figura 6-5. Ejemplo de una predicción de la distribución diamétrica para la parcela nº 3 de
Eucalyptus grandis. Una ventaja de este método es la compatibilidad de los modelos de crecimiento de
distinta resolución, por una parte los modelos de rodal en forma de sumas y valores

Crecimiento del Árbol 234
medios y los modelos de árbol individual en forma de distribuciones diamétricas,
que se combinan en un método conjunto.
Un elemento esencial de los modelos de árbol representativo es la
representación de la relación entre los diámetros y las alturas de los árboles
individuales. En general esto se realiza mediante curvas de altura unitarias.
Predicción sencilla del diámetro
Un método muy bien valorado para pronosticar el diámetro de árboles individuales
el la predicción sencilla mediante una ecuación de crecimiento de diferencias
algebraicas. La siguiente función denominada ecuación de Mitscherlich fue empleada
por Saborowski (1982) y Lemm (1991):
( )
( )01
02
ttk
ttk
i1i2 e1e1dd −−
−−
−−
= 6-21
Donde t1 y t2 son las edades del rodal al comienzo y al final del período; d1i y d2i son
los diámetros a la altura de pecho del árbol i a las edades t1 y t2; t0 es la edad a la que
se alcanza la altura de 1,3 m y k es un parámetros a estimar a partir de datos reales.
Ejemplo: En un inventario forestal se calculó la distribución diamétrica de un rodal de haya de 50 años. La altura dominante es de 19 m. Según Lemm (1991) los parámetros k y t0 se calculan a partir de la altura dominante mediante las ecuaciones siguientes:3
00356,0)19(000016,0003257,0 =⋅+=k y . 56,0469 )19(35379,00 =⋅= ⋅−et
El diámetro a la altura de pecho de dos árboles del rodal con un factor de cabida cubierta de 1 debe calcularse a la edad de 55 años. El diámetro medio cuadrático (árbol 1) es actualmente de 13 cm, el límite inferior es de 9 cm (árbol 2). Según la ec. 6-8 resulta:
( )
( ) cme
ed 2,14
1
113 56,05000356,0
56,05500356,0
21 =−
−−−
−−
= y ( )
( ) cme
ed 8,9
1
19 56.05000356,0
56.05500356,0
22 =−
−= −−
−−
El ejemplo demuestra una desventaja de este método sencillo de predicción de
diámetros, y es que se mantiene el rango absoluto. La relación entre el diámetro de
3 El modelo seria compatible si t0 se derivade directamente del modelo de altura

Crecimiento del Árbol 235
dos árboles no ha variado a lo largo del tiempo puesto que 8,9
2,14
9
13= . El método se
amplía en ocasiones con un componente estocástico y con ello se evita el problema
del manetimiento albsoluto del ranogo (Sloboda, 1984; Gaffrey, 1996).
El hecho de que el crecimiento diamétrico para una misma edad disminuye en
parte a medida que el diámetro aumenta fue demostrado por Schwappach (1905) en
rodales de roble de edad joven y media (figura 6-6). Esta experiencia se pudo
constatar también para rodales de pino (ver Gadow, 1984).
0
1
2
3
4
5
0 20 40 60 80 100
d [cm]
id [mm/ano]
23 anos
81 anos 230 anos
Figura 6-6. Relación entre el diámetro y el incremento en diámetro (id) en rodales de roble de
diferentes edades en las parcelas de ensayo Bordesholm y Freienwalde según Schwappach (1905).
El cambio del área basimétrica relativa
La desventaja del método de predicción del diámetro simple, el mantenimiento del rango absoluto, se soluciona cuando se utiliza un método que no mantenga la posición relativa del árbol de manera constante durante el período de tiempo a pronosticar.
El área basimétrica relativa (rg) se define como la relación entre el área basimétrica del árbol i con respecto al área basimétrica media.
1
i11 g
grg = y 2
i22 g
grg = 6-22
siendo g1i, g2i = Área basimétrica del árbol i a la edad 1 y 2 [cm²]
y g g1 , 2 = área basimétrica media del rodal a la edad 1 y 2 [cm²]

Crecimiento del Árbol 236
El método parte de la suposición de que el área basimétrica relativa de un árbol i
cambia con el tiempo. La dimensión relativa de un árbol pequeño se irá reduciendo
mientras que el área basimétrica relativa de un árbol grande se irá incrementando.
Una posibilidad de entender este desarrollo es el método propuesto por Clutter y
Jones (1980, ver Forss et al., 1996):
β
1
2tt
12 rgrg ⎟⎟⎠
⎞⎜⎜⎝
⎛
= 6-23
siendo t1, t2 = Edad del rodal al inicio y al final del período a pronosticar. El área
basimétrica del árbol i a la edad t2 (g2i) se estima a partir del área basimétrica del
árbol i a la edad t1 y del área basimétrica media g1 y g2 .
Ejemplo: El área basimétrica de dos árboles de un rodal hipotético a la edad de 50 años es: g11 = 132,7 cm² und g12 = 254,5 cm², El área basimétrica media a la edad de 50 años es de g1 = 201,1 cm², Mediante un modelo de crecimiento a nivel rodal se estima que el área basimétrica media a la edad de 55 años es g2 = 283,5 cm², Siendo β = 0,3 resulta que dentro de 5 años el área basimétrica de ambos árboles es:
( )
( )2
5055
22
5055
21
23611201
52545283
81841201
71325283
30
30
cm..
..g
cm..
..g
.
.
/
/
²
=⎥⎦⎤
⎢⎣⎡=
⎥⎦⎤
⎢⎣⎡= =
En este método no se mantiene el rango de los árboles ya que 3612184 8
254 5132 7
.
...
> .
Según Pienaar et al. (1990) el modelo del cambio relativo del área basimétrica
puede ser configurado de manera compatible (según el principio del telescopio) si la suma
del área basimétrica de todos los árboles individuales coincide exactamente con el
área basimétrica calculada por el modelo a la edad t2. La compatibilidad se consigue
mediante la siguiente limitación.
( )( )aj1
k
1jj
aj
2j2j
rgN
rgNGgN j1
∑=
= 6-24

Crecimiento del Árbol 237
Siendo Nj = Número de árboles de la clase diamétrica j (j = 1,2,...,k) que
sobreviven hasta la edad t2 y β
⎟⎟⎠
⎞⎜⎜⎝
⎛=
tta
1
2
El modelo del cambio relativo del área basimétrica es más plausible que el
modelo simple de pronóstico del crecimiento en diámetro porque el rango absoluto
de los árboles no se mantiene. Una desventaja de ambos métodos es la dependencia
de la edad, ya que no siempre se dispone de la edad de los árboles por ejemplo en
bosques irregulares mixtos.
Estimación directa del crecimiento El crecimiento del diámetro se puede estimar mediante una función de crecimiento,
donde dn = F(d0, t) + e1, o bien como función de incremento de diámetro Δd =
f(d) + e2 , siendo e1 y e2 los errores de estimación. No se dispone de datos directos
sobre incrementos de diámetro dd/dt, por lo tanto se tiene que estimar estos datos
a partir del crecimiento periódico medio.
Métodos paramétricos
El incremento en diámetro (Δd) se estima frecuentemente a partir del diámetro (d)
por lo tanto se consigue que este cálculo sea independiente de la edad. Como
alternativa al cálculo del incremento diamétrico mediante el cambio relativo del área
basimétrica Hessenmöller (2002) estimó el crecimiento en diámetro para varias
especies de frondosas mediante la función de crecimiento de KORSUN. El
científico checo Korsun (1935) empleó por primera vez una función con tres
parámetros que se adapta bien para estimar el incremento diamétrico:
⎥⎦⎤
⎢⎣⎡ ⋅+⋅+
=2)d(lnkdlnkk 210edΔ und 2
210 )d(lnkdlnkkdln ⋅+⋅+=Δ 6-25
con k0, k1, k2 = Parámetros empíricos, d = Diámetro a la altura de pecho (cm) Δ d = Crecimiento medio anual (cm)
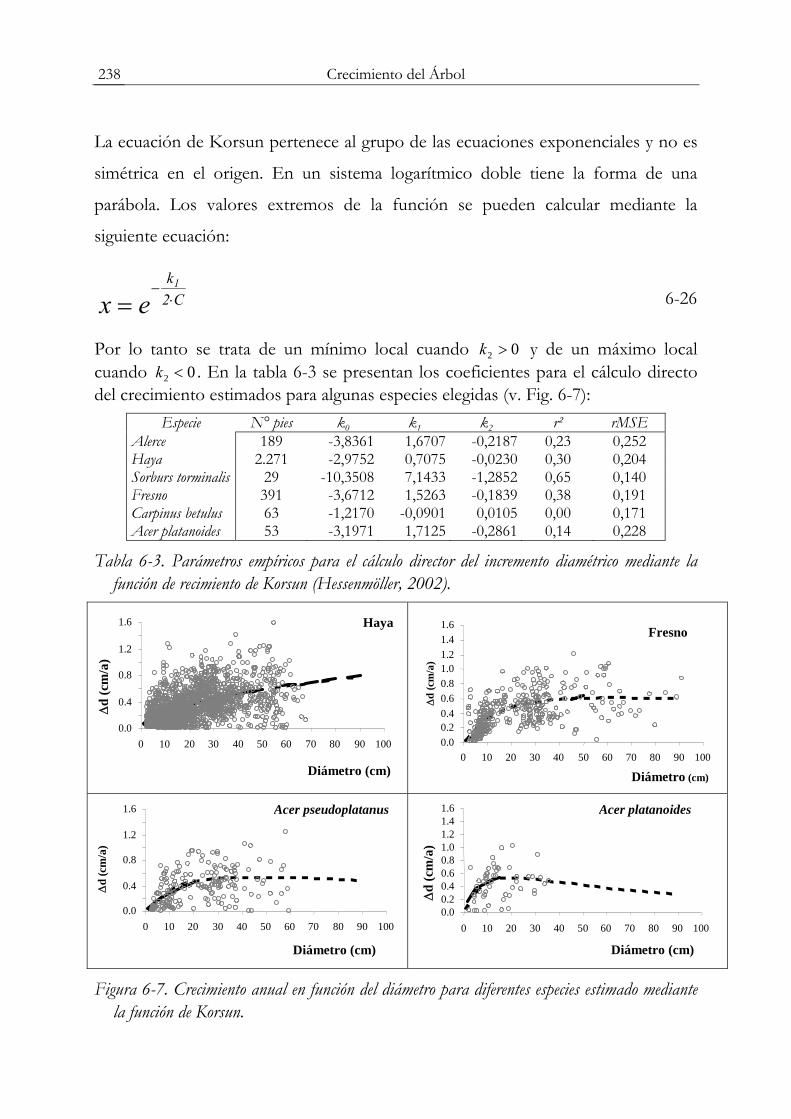
Crecimiento del Árbol 238
La ecuación de Korsun pertenece al grupo de las ecuaciones exponenciales y no es
simétrica en el origen. En un sistema logarítmico doble tiene la forma de una
parábola. Los valores extremos de la función se pueden calcular mediante la
siguiente ecuación:
C2k1
ex ⋅−
= 6-26
Por lo tanto se trata de un mínimo local cuando y de un máximo local cuando . En la tabla 6-3 se presentan los coeficientes para el cálculo directo del crecimiento estimados para algunas especies elegidas (v. Fig. 6-7):
02 >k02 <k
Especie N° pies k0 k1 k2 r² rMSE Alerce 189 -3,8361 1,6707 -0,2187 0,23 0,252 Haya 2.271 -2,9752 0,7075 -0,0230 0,30 0,204 Sorburs torminalis 29 -10,3508 7,1433 -1,2852 0,65 0,140 Fresno 391 -3,6712 1,5263 -0,1839 0,38 0,191 Carpinus betulus 63 -1,2170 -0,0901 0,0105 0,00 0,171 Acer platanoides 53 -3,1971 1,7125 -0,2861 0,14 0,228
Tabla 6-3. Parámetros empíricos para el cálculo director del incremento diamétrico mediante la función de recimiento de Korsun (Hessenmöller, 2002).
Haya
0.0
0.4
0.8 1.2
1.6
0 10 20 30 40 50 60 70 80 90 100
Diámetro (cm)
Δd
(cm
/a)
Fresno
0.00.20.40.60.81.01.21.41.6
0 10 20 30 40 50 60 70 80 90 100
Diámetro (cm)
Δd
(cm
/a)
Acer pseudoplatanus
0.0 0.4 0.8 1.2 1.6
0 10 20 30 40 50 60 70 80 90 100
Diámetro (cm)
Δd
(cm
/a)
Acer platanoides
0.00.20.40.60.81.01.21.41.6
0 10 20 30 40 50 60 70 80 90 100
Diámetro (cm)
Δd
(cm
/a)
Figura 6-7. Crecimiento anual en función del diámetro para diferentes especies estimado mediante la función de Korsun.

Crecimiento del Árbol 239
La función de crecimiento de Korsun indica una típica culminación temprana del
crecimiento diamétrico, por lo tanto el desarrollo de un árbol en función del
diámetro queda bastante bien representada. Las diferencias más claras en el
transcurso de la función se encuentran en las especies Acer platanoides, Acer
pseudoplatanus y Sorbus torminalis. Es significativo el transcurso casi idéntico de las
funciones para el haya y el carpinus betulus. En el caso del carpinus betulus la
correlación entre el incremento medio anual y el diámetro es bastante pequeña.
Esto se aclara por el hecho de que ambas funciones muestra un transcurso casi
lineal. En el caso del haya ambos parámetros empíricos k1 y k2 de la función de
KORSUN son casi cero.
Vanclay (1994, p. 165) empleó un método para la estimación del incremento
diamétrico en árboles de bosques tropicales que condujo a resultados similares, por
ejemplo:
a) Bertalanffy: d0147.0d245.0d 44.0 ⋅−⋅=Δ
b) Botkin: 2
22
d00668.0d527.174.2611677
d167.0d9.50137ddd
⋅−⋅+
⋅−⋅+⋅−
=Δ
6-27
6-28
Estos parámetros se estimaron a partir de datos correspondientes a la especie Acer
saccharum (Hahn y Leary, 1979). La figura 6-8 muestra que las funciones de
Bertalanffy (1948) y Botkin (1993) describen curvas de incremento similares. Un
problema de este método es que el incremento en diámetro depende solo del
diámetro. Esta es una simplificación extrema puesto que el crecimiento en
diámetro depende entre otras cosas de la densidad del rodal.
Ya que entre el incremento en diámetro y la superficie de copa existe una clara
relación (ver Kramer, 1988) se puede utilizar esta última a partir del incremento
obtenido mediante la función que emplea el diámetro, como factor para estimar el
crecimiento del árbol representativo. Una medida de la competencia a la que están
sometidos los árboles individuales es el factor de cobertura C66, tal y como fue

Crecimiento del Árbol 240
definido por Wensel et al. (1987). Este método fue utilizado por Nagel (1994) para
pronosticar el crecimiento de bosques en el noroeste de Alemania. El C66 es igual
a la suma de la proyección de las copas (ks66) de todos los árboles, que cortan la
copa del árbol de referencia a una altura del 66 % de su longitud de copa, relativo a
la superficie del rodal. Para calcular el índice de competencia C66 se necesita
información sobre la forma de la copa del árbol (ver Capítulo 3).
0 50 100 150
d [cm]
Δd [cm] 0.7
0.4
0.5
0.6
0
0.1
0.2
0.3
Figura 6-8. Dos funciones de incremento de diámetro según Vanclay (1994). Sin embargo en un modelo independiente de la posición no es tan importante la
descripción exacta de la copa como en un modelo dependiente de la posición ya
que se emplea un mayor grado de abstracción. El incremento en diámetro y en
altura disminuye cuanto mayor es la cobertura, es decir cuanto mayor es el valor del
índice C66.
Métodos no paramétricos
Para estimar el incremento anual partiendo de los valores de una base de datos
Hessenmöller (2002) empleó la regresión del k-esimo vecino más cercano (k-nearest-
neighbour). El incremento del árbol de referencia j ( jdΔ ) se estima a partir de los
incrementos conocidos y ponderados de los k árboles vecinos:
∑∑==
⎟⎠
⎞⎜⎝
⎛⋅Δ=Δ
k
ii
k
iiij GGdd
11 6-29
donde idΔ
= Incremento anual del árbol vecino i (cm)
N = Número de árboles vecinos considerados

Crecimiento del Árbol 241
Gi = Peso del vecino i.
Para calcular Gi se emplea la distancia Wi entre el árbol de referencia j y su vecino i.
Cada árbol posee unas características que en este caso se denominan “Instancias”.
La distancia Wi se calcula a partir de las distancias de las instancias w y de sus
ponderaciones g:
kki wgwgW ⋅++⋅= ...11 6-30
Por ejemplo la distancia Wi empleando las instancias diámetro D y altura H de un
árbol vecino i en comparación con el árbol de referencia j se puede calcular de la
siguiente manera:
( ) ( 2jiH
2jiDi HHgDDgW −⋅+−⋅= ) 6-31
Hay muchas maneras de determinar el peso Gi de un árbol vecino i; por ejemplo
mediante una función exponencial . Frecuentemente se emplea la
siguiente función:
iWi eG −=
ii W1
1G+
= 6-32
Para poder comparar mejor la influencia de las instancias es razonable normalizar
las distancias. Hessenmöller (2002) empleó para ello cuatro funciones de distancia
diferentes en función de la instancia considerada. Estas fueron a su vez
normalizadas mediante la varianza de la instancia. Dos funciones de distancia son:
)(PVar
PPw ji
iII
−= 6-33
5,2
5,2
))(( PVar
PPw ji
iIV
−= 6-34

Crecimiento del Árbol 242
En la tabla 6-4 se encuentran los resultados de la regresión knn (knn = k nearest
neighbors) para 2271 datos independientes de la posición para el haya. Todos los
modelos muestran una bondad del ajuste similar. N = 2271 W g SSQ rMSE r²
Diámetro IV 1.275 90,917 0,200 0,32 Altura II 625 90,542 0,200 0,34 Volumen de copa IV 1.000 91,830 0,201 0,32 Superficie de copa IV 2.050 90,471 0,200 0,34 Diámetro y altura IV / II 1.750/330 88,282 0,197 0,35
Tabla 6-4. Los parámetros y las funciones de distancia para instancias independientes de la posición del modelo knn para haya (w = función de distancia óptima, g = peso parcial optimizado). Las columnas de la derecha se refieren al test KS sobre la distribución normal de los residuos.
Si existen numerosas instancias es recomendable antes de emplear el método knn
compararlas y dado el caso reducirlas. Un método adecuado para esto es el análisis
de los componentes principales.
0.0 0.2 0.4 0.6 0.8 1.0 1.2 1.4 1.6
0 10 20 30 40 50 60 70 80 90 100
Diámetro (cm)
Δd
(cm
/ano
)
0,00,20,40,60,81,01,21,41,6
0 10 20 30 40 50 60 70 80
Diámetro (cm)
Δd
(cm
/ano
)
Figura 6-9. El crecimiento estimado de hayas en comparación con el crecimiento observado en función del diámetro. A la izquierda modelo paramétrico y a la derecha modelo no paramétrico.
Hessenmöller (2002) pudo reducir el error de estimación a la mitad mediante el
método knn en comparación a otros modelos paramétricos para la estimación del
crecimiento. Sin embargo este resultado no es ningún indicio de la superioridad
general de los métodos no paramétricos. La validez general de la estimación viene
determinada por la cantidad de observaciones y la representatividad de los valores

Crecimiento del Árbol 243
observados. Las observaciones empíricas son decisivas para la exactitud de la
estimación; menos importante es el tipo de modelo.
Actualización de datos de inventario
Un ámbito de aplicación de los modelos de crecimiento es la actualización
periódica de datos de inventario. Para esta finalidad Schröder et al. (2002)
desarrollaron un modelo de incremento de área basimétrica independiente de la
edad para la especie Pinus pinaster.
Ejemplo: En el rodal “As Neves” en el noroeste de España se midió una parcela de ensayo de 0,05 ha. En la parcela hay un total de 77 árboles. La altura media de los 5 árboles más gruesos es de 8,8 m. La profundidad media del suelo (ESD) calculada mediante tres mediciones aleatorias es de 40, 83 cm. En la siguiente lista se incluye el diámetro a la altura de pecho de los 77 árboles.
9.0 11.0 11.8 12.5 13.3 14.0 14.8 15.5 9.5 11.0 11.8 12.5 13.3 14.0 14.8 16.0
10.0 11.0 12.0 12.5 13.5 14.0 14.8 16.0 10.0 11.0 12.0 12.5 13.5 14.3 15.0 16.0 10.3 11.0 12.3 12.8 13.5 14.3 15.0 16.5 10.3 11.3 12.3 12.8 13.8 14.3 15.0 17.5 10.3 11.8 12.3 13.0 13.8 14.5 15.3 18.0 10.5 11.8 12.3 13.0 14.0 14.5 15.3 10.5 11.8 12.3 13.0 14.0 14.5 15.3 10.8 11.8 12.5 13.0 14.0 14.8 15.5
La finalidad de este ejemplo es estimar el diámetro del árbol j (marcado en negrita en la tabla, dj = 16,5 cm) dentro de cinco anos. Para cuantificar la situación de competencia actual del árbol j hay que calcular en primer lugar para la parcela de ensayo las variables de masa, estas son:
Área basimétrica(G) = 21,3 m2 / ha
Número de pies(N) = 1540 árboles por ha
Distancia relativa (RS) = (10000/1540)0.5 / 8,8 = 0,29
En un segundo paso se calcula el índice BAL del árbol j. Referido a la parcela de superficie 0,05 ha se obtiene un valor del ( ) 222
PLOT m 904,0180,0175,0
4=+⋅=
πjBAL
Referido a una hectárea se obtiene un valor 2m 89,005,0049,0 ==jBAL . Con este resultado intermedio se puede calcular el índice BALMOD: ( )
16,029,0
3,2198,01===
−=
RSGBAL
RSp
BALMOD jjj
La competencia actual a la que está sometido el árbol j es relativamente pequeña. Interesa no solo conocer la competencia actual sino también la vitalidad histórica. Esta última se puede cuantificar

Crecimiento del Árbol 244
mediante el denominado crown spread ratio (csr) que proporciona la relación entre la anchura de la copa y la altura del árbol. La altura del árbol j es hj = 8,3 m. El radio medio de la copa se calculó mediante un espejo de copa y es de krj = 1,3 m. A partir de estos datos se calcula el crown spread ratio (csr) 95,03,86,2 5.05,0 =⋅=⋅= −−
jjj hcwcsr
95,03,86,2 5.05,0 =⋅=⋅= −−jjj hcwcsr
La competencia histórica del árbol j se considera como la competencia media. Se dispone de toda la información para pronosticar el diámetro del árbol j. El incremento anual de área basimétrica estimado se obtiene de la siguiente ecuación (Schröder, 2001): ln(Δgj) = + 0,6266 Intercepto
+ 0,6088 ln(16,5) Diámetro (dj) - 0,00027 16,52 Diámetro al cuadrado (dj
2) + 0,8776 0,95 vitalidad histórica (csrj) - 0,2041 0,16 competencia actual (BALMODj) + 0,0030 40,83 Productividad de la estación (ESD)
----------------------------------------------------------------------------------------- = 3,183 cm2
El incremento de área basimétrica en cinco años es por lo tanto: . El
área basimétrica del árbol j dentro de 5 años será:
2183,35 j c 595,1205 meg =⋅=Δ ⋅
225 j c 42,334595,1205,16
4mg =+⋅=+
π . El diámetro
del árbol j será por lo tanto: cm 6,2042,33445 j =⋅=+ π
d .
Modelos dependientes de la posición La ventaja de los modelos de árbol representativo independientes de la posición es
que, generalmente, solo se necesita una lista de los diámetros para calcular el
desarrollo de los árboles. La desventaja es que se posee información insuficiente
sobre la situación de competencia de los árboles individuales. Los modelos de árbol
individual dependientes de la posición se caracterizan porque las coordenadas y al
menos el diámetro de los árboles son conocidos (Figura 6-10).
Este [m]
Norte [m] Figura 6-10. En los modelos de árbol individual dependientes de la posición se conocen las coordenadas de los árboles. A partir de las coordenadas se pueden derivar la situación de competencia en la vecindad de un árbol.

Crecimiento del Árbol 245
Las coordenadas de los árboles se pueden generar bajo determinadas condiciones
de manera automática a partir de fotos aéreas (Dralle, 1997). Una segunda
posibilidad es generar las posiciones de los árboles mediante simulación (Pretzsch,
1995; Lewandowski y Gadow, 1997).
Reproducción simulada de las posiciones de los árboles
La finalidad de la reproducción simulada de los rodales es describir mejor las
operaciones forestales y emplear modelos de crecimiento que puedan utilizar la
posición de los árboles. Todo ello depende naturalmente de que el rodal generado
artificialmente y el rodal real sean lo más parecidos posible. En primer lugar se
plantea la pregunta de en que forma se deben emplear las distribuciones de los
árboles vecinos, que son conocidas, para generar un rodal similar. Para ello hay que
clarificar que atributos del rodal original se deben mantener en la copia y que
características pueden perderse.
Una reproducción en relación a la distribución de las posiciones de los árboles
del rodal simulado se califica como perfecta cuando a cada árbol del rodal real le
corresponde un árbol del rodal simulado que mantiene exactamente la misma
distancia a sus tres vecinos más próximos que en el rodal original. Además para la
reproducción del rodal debe conocerse la distribución empírica de la mezcla de
especies. La constelación de especies que rodean a un árbol se calcula según
Füldner y Gadow (1994) mediante la proporción relativa de los tres vecinos que
pertenecen a una misma especie. Si se sabe, por ejemplo, que la constelación de
especies de un árbol es 1/3 se puede deducir que uno de los tres árboles pertenece
a una especie diferente que los otros dos. Una reproducción se define como
perfecta en relación a la mezcla de especies cuando en el rodal simulado el valor de
las constelaciones de especies aparece con las mismas frecuencias que en rodal
original. Lo mismo sucede con la variable diferenciación de tamaños. Esta variable
toma el valor cero cuando los tres árboles vecinos tienen el mismo diámetro que el
árbol de referencia. En relación a la diferenciación de tamaños la reproducción del

Crecimiento del Árbol 246
rodal es óptima cuando todos los valores del rodal original coinciden con los del
rodal simulado. Finalmente se utiliza la distribución de diámetros y especies.
El método para reproducir un rodal se puede resumir de la siguiente manera:
los árboles del rodal original descritos mediante su diámetro y especie se deben
distribuir en la superficie del rodal, cuyos límites son conocidos, de tal manera que
las distribuciones empíricas de la diferenciación de tamaños y de las constelaciones
de especies coincidan con las distribuciones del rodal original. Partiendo de una
situación cualquiera de los árboles dentro del rodal se consigue una similitud con
las principales variables del rodal original mediante la realización de cambios
sistemáticos de la posición de los árboles. Este método de optimización se realiza
en fases independientes separadas unas de otras. Se ha demostrado que el algoritmo
descrito proporciona resultados satisfactorios considerando las variables
estructurales mezcla de especies y diferenciación de tamaños. Otra alternativa es el
generador de estructuras de rodal STRUGEN (Pretzsch, 1993), que también
persigue la finalidad de reproducir estructuras de rodal. La ventaja de este método
es que las estructuras espaciales se pueden reproducir fácilmente en base a
descripciones del rodal ya existentes4. La reproducción de estructuras de rodal a
partir de mediciones de parcelas de ensayo es una condición importante para la
aplicación de modelos de árbol individual.
Los modelos WASIM y MOSES
Sterba (1983) desarrolló un modelo de árbol individual para masas mixtas de pícea
y pino similar al de Ek y Monserud (1974). La estructura de este modelo se
representa de manera simplificada en la figura 6-11. El incremento en altura o en
diámetro de un árbol es igual al incremento potencial (PotZuw) multiplicado por
4 Un proceso estocástico puntual tiene la ventaja de que está apoyado por una base teórica (ver por ejemplo Tompo,
1986; Penntinen et al, 1994 o Degenhardt, 1995), pero este método no es adecuado para la aplicación práctica. La mayoría de los trabajos tratan solamente el tema de la distribución espacial de los árboles. Los procesos puntuales marcados ofrecen la posibilidad de asignar un atributo a una posición, sin embargo una condición para la reproducción exitosa de los rodales es que a una posición se le puedan asignar varios atributos simultaneamente.

Crecimiento del Árbol 247
los factores de reducción (MGO) y MOS. ETZuw es igual al incremento en altura
del árbol dominante de las tablas de producción o en el caso del diámetro es igual al
incremento en diámetro calculado de la relación diámetro-altura dominante para un
árbol solitario (Regla C-D de Sterba).
Crecimiento = PotZuw × MGO ×
MGO CR= γ
MOS
Mos e Konk= −1δ
( )βα HETZuwPotZuw ⋅+⋅= 1
Figura 6-11. Estructura del modelo de árbol individual WASIM para masas mixtas de pícea y
pino según Sterba (1983, 1990). La variable MGO es una medida de la competencia histórica en el rodal y viene
determinada por el grado de cobertura de copas (CR). La variable MOS describe la
competencia actual en base a un índice de competencia (Konk).
Ejemplo: Pícea, altura=20m; ETZuw=5mm/año; α= 1.16E-12; β = 8.54; γ = 0.4001; δ = -4.81
CR Konk id
0,6 1,5 1,9
0,6 0,5 3,3
0,6 1,0 2,5
0,3 1,0 1,3
Para un mismo grado de cobertura de copas aumenta el incremento en diámetro a
medida que disminuye la competencia (3,3 >1,9). Los árboles que se encuentran en
una misma situación de competencia tienen un mayor incremento en diámetro
cuanto mayor es la copa (2,5 > 1,3). El modelo MOSES (Hasenauer, 1994;
Hasenauer et al., 1994) muestra la misma estructura que el modelo WASIM. Solo se
estimaron de nuevo los parámetros para otros tipos de rodal. La ecuación 6-35
muestra la estructura básica del modelo MOSES para calcular el incremento en

Crecimiento del Árbol 248
altura. Como ya se ha citado anteriormente la estructura del modelo MOSES es
idéntica a la presentada en la figura 6-11.
)e1(CRhh CIpot
βαΔΔ
−
−⋅⋅= 6-35
siendo α = 0,0845 y β = 6,158 para haya y α = 0,241 y β = -3,953 para pícea. El modelo para calcular el incremento diamétrico tiene la misma estructura. El
siguiente procedimiento CR_CI5 calcula el grado de cobertura de copas y el índice
de competencia para todos los árboles de un rodal cuyas coordenadas son
conocidas. Procedure CR_CI const {constantes y variables globales } maxN = 1000; var Nr,BA : array [1..maxN] of byte; Diam,H,KAH,X,Y, ih_pot,id_pot,kd_pot, CR,CI,ih,id,delta_KAH : array [1..maxN] of single; device1 : text; Superficie,s,incremento: string; j,numeropies : integer; periodocrecimiento, OH_Bon_Bu,OH_Bon_Fi : byte; Alter_Bu,Alter_Fi : integer; Procedure CR_CI; {calcula el crown ratio (CR) y competition index (CI) para cada árbol} var ca,S_j,S_k,CI_sum,dist, solapamiento, help1,help2,help3,help4, help5,help6,help7,help8, angulo1,angulo2, CI_jk : real; k : byte; begin for j:=1 to numeropies do {bucle externo} begin CR[j]:=(H[j]-KAH[j])/H[j]; {crown ratio} {para calcular el CI se necesita la superficie de copa (CA), la superficie de superposicion potencial (O) y parametros del arbol (S) } ca:=PI*sqr(kd_pot[j]/2); S_j:=h[j]*kd_pot[j]; CI_sum:=0; {Restablecer la suma de CI para el siguiente arbol j} for k:=1 to numeropies do {bucle interno para solapamiento} begin if j<>k then begin {eliminacion de competidores} dist:=sqrt(sqr(X[j]-X[k])+sqr(Y[j]-Y[k])); {consulta, si existe solapamiento} if dist<(kd_pot[j]/2+kd_pot[k]/2) then begin {cuando una copa esta totalmente solapada} if dist<=abs(kd_pot[j]/2-kd_pot[k]/2) then begin if kd_pot[j]>=kd_pot[k] then solapamiento:=PI*sqr(kd_pot[k]/2) else solapamiento:=PI*sqr(kd_pot[j]/2); end else begin {begin of else, d.h. dist>abs( )} help3:=(sqr(dist)+sqr(kd_pot[j]/2)-sqr(kd_pot[k]/2)) /(2*dist*(kd_pot[j]/2)); help4:=(sqr(dist)+sqr(kd_pot[j]/2)-sqr(kd_pot[k]/2)) /(4*sqr(dist)*sqr(kd_pot[j]/2));
5 Extraído de un programa de M. Albert (1997).

Crecimiento del Árbol 249
help5:=sqrt((4*sqr(dist)*sqr(kd_pot[j]/2)) -sqr(sqr(dist)+sqr(kd_pot[j]/2)-sqr(kd_pot[k]/2))); {conversion arccos en arctan, Bronstein, p.185} angulo1:=PI/2-arctan(help3/sqrt(1-sqr(help3))); help1:=angulo1-help4*help5; help6:=(sqr(dist)+sqr(kd_pot[k]/2)-sqr(kd_pot[j]/2)) /(2*dist*(kd_pot[k]/2)); help7:=(sqr(dist)+sqr(kd_pot[k]/2)-sqr(kd_pot[j]/2)) /(4*sqr(dist)*sqr(kd_pot[k]/2)); help8:=sqrt((4*sqr(dist)*sqr(kd_pot[k]/2)) -sqr(sqr(dist)+sqr(kd_pot[k]/2)-sqr(kd_pot[j]/2))); angulo2:=PI/2-arctan(help6/sqrt(1-sqr(help6))); help2:=angulo2-help7*help8; angulo:=sqr(kd_pot[j]/2)*help1+sqr(kd_pot[k]/2)*help2; end; {fin de else, es decir dist>abs( )} S_k:=h[k]*kd_pot[k]; CI_jk:=(solapamieno/ca)*(S_j/S_k); CI_sum:=CI_sum+CI_jk; end; end; end; {Fin del bucle interno} CI[j]:=CI_sum; {Indice de competencia para el árbol j} end; {Fin del bucle externo} end;
En la tabla 6-5 se presentan los datos iniciales y el pronóstico para cinco años
realizados con el modelo MOSES para una parte de un rodal con 15 árboles. En las
publicaciones citadas anteriormente y en la representación realizada por Albert
(1997) se pueden encontrar mas detalles sobre el modelo MOSES Datos iniciales en 5 años
NR BA BHD
H KAH X Y Diam H KAH
1 2 12,1 14,2 4,7 12,50 2,10 16,83 17,81 5,702 2 12,8 14,2 6,5 9,40 3,10 15,61 17,34 7,19 3 2 5,7 9,2 4,6 5,20 3,30 8,43 12,59 4,82 4 2 9,8 14,2 7,6 6,20 3,70 10,58 16,10 8,17 5 2 12,4 14,2 8,2 2,90 4,20 13,51 16,40 8,65 6 5 12,8 12,5 8,4 12,40 4,30 13,93 14,25 8,99 7 2 6,3 10,0 3,5 5,90 4,70 7,50 12,47 4,02 8 2 8,8 12,7 6,5 5,40 4,80 9,37 14,39 6,99 9 2 5,5 9,9 6,2 5,10 5,00 6,51 12,14 6,36 10 2 4,4 8,2 5,9 14,00 5,50 10,63 12,18 5,94 11 2 6,2 11,1 8,3 4,90 5,60 6,82 12,85 8,41 12 5 19,7 15,6 9,2 8,30 5,80 19,97 16,48 9,88 13 5 19,4 14,7 7,3 10,60 5,80 20,49 16,33 8,18 14 2 9,3 16,1 8,0 13,10 5,80 11,04 18,65 8,89 15 2 9,7 14,0 5,4 6,00 6,10 10,01 15,27 6,45
Tabla 6-5. Datos iniciales y resultados obtenidos empleando el modelo MOSES en una parte de un rodal con 15 árboles

Crecimiento del Árbol 250
El modelo SILVA
Es ya conocido que el crecimiento de un árbol se ve influido por el espacio
disponible para la copa y por la cobertura y la sombra proporcionada por otros
árboles, es decir en el crecimiento influye la disposición espacial de los árboles
vecinos. Estos factores se pueden considerar en un método espacial tridimensional,
conocido como el método de las celdas matriz (Pretzsch, 1992 página 129, ver figura 6-
12). En base a las coordenadas de los árboles, los radios de las copas, las alturas y
las alturas donde comienzan las copas así como en base a la forma de las copas se
representa la expansión espacial de los árboles en coordenadas cartesianas y se
localizan en una matriz espacial tridimensional. Para cada punto medio de las celdas
de la matriz se determinan los árboles que localizan en este punto. El resultado se
almacena en una matriz espacial que modeliza la estructura del rodal y proporciona
información sobre la extensión de las copas y la presencia de árboles en diferentes
niveles de altura. Según Pretzsch (1992 página 199) el incremento en altura de un
haya se puede estimar mediante la ecuación 6-36.
KonkCRΔhΔh 088.0pot ⋅⋅= 6-36
siendo Δhpot = incremento en altura potencial CR =grado de cobertura de copa Konk =Índice que describe la cobertura y la limitación de la expansión lateral de las copas
Biber (1996) desarrolló las simulaciones de Fisheye en rodales de pícea y haya para
mejorar la descripción de la situación de competencia de un árbol. Si se conocen las
posiciones de los árboles, la longitud y ancho de las copas se puede generar una
foto Fisheye hemisférica. La competencia a la que está sometida el árbol de
referencia se obtiene de las proporciones de cobertura en la foto hemisférica
discretizada (ver Courbaud, 1995).
Debido a la escasa disponibilidad de datos espaciales los modelos de árbol
individual solo son adecuados en casos muy concretos para realizar pronósticos del
desarrollo forestal. Sin embargo el perfeccionamiento de estos métodos es muy

Crecimiento del Árbol 251
importante ya que el crecimiento de los árboles bajo condiciones complejas – como
por ejemplo rodales mixtos irregulares – solo se puede similar en base a estos
modelos de árbol individual.
Modelos de pequeñas superficies (gap models)
Los ecosistemas forestales que se extienden por grandes superficies se componen
de pequeñas superficies en mosaico. Mediante el análisis de las pequeñas superficies
se llega a comprender el ecosistema en su conjunto. Ejemplos de modelos de
desarrollo de pequeñas superficies (en inglés gaps, en francés chablis) se encuentran
los trabajos de Botkin et al. (1972), Shugart (1984), Kienast y Kuhn (1989) y Botkin
(1993). Estos ejemplos tratan, entre otras cuestiones, de entender los procesos
naturales en los bosques nativos (Falinski, 1998), que se apoyan de cierta manera
en una base teórica denominada el concepto ciclo-mosaico (Schmidt, 1989; Remmert,
1991). La figura 6-13 muestra la dinámica de desarrollo natural de una pequeña
superficie de bosque mixto de eucaliptos en Australia (Shugart, 1984 página 105).
El ciclo de la pequeña superficie se caracteriza por tener cuatro fases típicas:
Dominancia de un árbol adulto en la pequeña superficie (a); fase de regeneración
natural y muerte del árbol dominante (b), crecimiento y diferenciación a
consecuencia de la competencia (c) y fase de envejecimiento (d).
El tamaño de las pequeñas superficies se define de manera diferente. Los
árboles A y B de la figura 6-13 (derecha.) se han caído y han dañado a los árboles C,
D y E (ver Van der Meer, 1995). Según Brokaw (1982) el límite exterior de una
pequeña superficie se define mediante una apertura en las copas determinada por
los árboles vivos, se trata por lo tanto de la zona central destruida. Según Rièra
(1982) una pequeña superficie es la zona de apertura de copas donde todavía es
posible la regeneración.

Crecimiento del Árbol 252
Dinámica natural en pequeñas superficies Definición de una pequeña superficie
a
b
c
d
B
A
D
C
Brokaw
Riera
Figura 6-13. Izquierda: Representación esquemática de la dinámica de desarrollo natural de una pequeña superficie de un bosque mixto de eucaliptos en Australia (Shugart, 1984 página 105). Derecha: Diagrama de perfil de una pequeña superficie imaginaria con diferentes definiciones del tamaño de la pequeña superficie.
En la figura 6-14 (la parte derecha) se muestra la vista en planta de una pequeña
superficie imaginaria con una zona central (delimitada por la vegetación de hasta 20
m de altura), una zona exterior (delimitada por los árboles dominantes) y la zona
limítrofe del bosque. En inventarios de vegetación y de la estación Van der Meer
(1995, página 84) recopiló datos sobre la especie y la densidad de aparición de la
misma en parcelas de forma cilíndrica de 25 cm de radio y niveles de altura 1 m.
Segmento de muestra
Figura 6-14. Método de recopilación de datos sobre la vegetación en una pequeña superficie. Partiendo del nuevo establecimiento de un bosque en Europa central, Thomasius
diferenció cuatro estadios: el estadio herbáceo y arbustivo, el bosque pionero, el
bosque de transición y el bosque cerrado. Si se destruye el bosque cerrado, el
bosque de transición o el bosque pionero se repite el ciclo (figura 6-15).

Crecimiento del Árbol 253
Zona virgen
Estadio de bosque cerradoFase de envejecimiento
Fase de Fase de maduración destrucción
Fase de regeneración
Fase de formación del bosque cerrado
Estadio bosque pionero
- Fase juvenil - Fase madura
Estadio de transición
Estadio herbáceo
Inmigración de especies arbóreas pioneras
Inmigración de especies climax Eliminación de las
especies pioneras
Destrucción del bosque cerrado por influencias exógenas
Inmigración de plantas herbáceas
Destrucción del bosque pionero por influencias
exógenas
Destrucción del bosque de transición por influencias
exógenas
Figura 6-15. Desarrollo natural del bosque según Thomasius (1988) Dentro del estadio de bosque cerrado se repite un subciclo interno de menor
duración que comprende la maduración, el envejecimiento, la destrucción y la
regeneración. Para bosques de haya Jenssen y Hofmann (1997) distinguieron dentro
de este subciclo los estadios de crecimiento juvenil y engrosamiento (dura unos 20
años), formación del fuste (dura unos 40 años), estadio de superficie arbolada (dura
unos 90 años), estadio de superficie arbolada envejecida (dura de 80 a 120 años) y
estadio de desintegración y nueva formación (dura de 40 a 60 años).
Otras reflexiones sobre la sucesión del bosque en Europa central se
encuentran en los trabajos de Otto (1995), Kenk y Weise (1998) y Perpeet (1999).
Regeneración La modelización de la regeneración pasa por varias fases características desde la
floración hasta el enraizamiento y finalmente el crecimiento juvenil de donde se
recluta el Ingrowth (ver Vanclay, 1994, página 193). La formación de las flores,
polinización, fructificación, enraizamiento y la distribución espacial de las semillas
son elementos importantes dentro de la dinámica de las pequeñas superficies. Estos
procesos se intentan reproducir mediante distribuciones empíricas de la

Crecimiento del Árbol 254
distribución de semillas, de la dirección del viento, de las condiciones
meteorológicas y en base a las condiciones edáficas conocidas. Liu y Hytteborn
(1991) pudieron comprobar en bosque nativo de pícea que los abedules
principalmente colonizan las elevaciones y las hondonadas del terreno. La pícea sin
embargo se reparte mayoritariamente sobre viejos troncos caídos mientras que las
semillas de pino se distribuyen aleatoriamente sobre la superficie. Tipo de modelo Elementos de los procesos
Modelos de enraizamiento Floración, Polinización, Fructificación, Liberación de semilla, enraizamiento
Modelos de regeneración Enraizamiento hasta la altura de pechoModelos de reclutamiento Altura de pecho hasta el reclutamiento, p. ej. 10
cm de diámetro
El desarrollo de la vegetación natural en parcelas con árboles derribados por el
viento o parcelas de corta a hecho es objeto de análisis similares. En una superficie
de 1 ha cortada a hecho sobre una estación de Luzulo-Fagetum observaron Gregor
y Seidling (1997) el desarrollo de la vegetación durante un período de 50 años
(Figura 6-16).
0 10 20 30 40 50
0
Tiempo (años)
Rubus 100
% d
e su
perf
icie
Instalación de la parcela permanente
Bosque pionero
Bosque de transición
Bosque cerrado
Corta a hecho
Lluvia helada
%
Sambucus
Vias de saca
Rubes- gestrüpp
Figura 6-16. Desarrollo de la vegetación en una superficie de corta a hecho en Luzulo-Fagetum
Durante los primeros años dominaron las especies ruderales, hasta el año 10
dominaron las especies de Rubus, del año 10 al 15 predominan matorrales de
sambucus-frangula, después en una proporción similar el bosque pionero, bosque
de transición y el bosque cerrado.

Crecimiento del Árbol 255
Moeur (1993) y Golser y Hasenauer (1997) analizaron la modelización del
crecimiento de la regeneración natural en diferentes condiciones de radiación solar.
El crecimiento en altura de la regeneración se ve influido por la radiación solar
directa e indirecta considerando diferentes índices de competencia que describen
las zonas de influencia y las zonas de sombra en la superficie de regeneración. La
radiación adicional debido al efecto de borde se considera cuando la distancia al
borde de la masa forestal es como máximo dos veces la altura dominante. En este
caso el ángulo de radiación α se determina mediante la altura del rodal y la distancia
al borde. La tan(α) como medida de la radiación difusa adicional se calcula para
todas las direcciones excepto el norte (Figura 6-17).
α
Absti > Hdom Absti < Hdom Figura 6-17. Modelización de la radiación disponible para la regeneración según Golser y
Hasenauer (1997).
Mortalidad Una de las cuestiones más complicadas dentro de la modelización forestal es
comprender los procesos de mortalidad de los árboles. Los datos referidos a las
direcciones en las que se caen los árboles constituyen una base importante para
modelizar la dinámica en las pequeñas superficies. La figura 6-18 muestra la
dirección en la que se han caído 35 árboles en un bosque en la Guayana francesa
según Van der Meer (1995, página 64). El vector medio de la dirección de caída
señala N-NW. En base a un análisis realizado mediante el test aleatorio de Raleigh
se descubrió que la dirección de caída en este ejemplo no se desvía de manera

Crecimiento del Árbol 256
significativa de la dirección de caída aleatoria. En cambio Liu y Hytteborn (1991)
demostraron que en un bosque de pícea nativo el 62% de los árboles muertos (gaps
makers) se habían caído en las direcciones SW, S y SO.
N
S
Media
Figura 6-18. Dirección de caída de 35 árboles en un bosque de la Guayana francesa.
Representación espacial Las situaciones de partida para simular la dinámica de una pequeña superficie
pueden ser reales o hipotéticas. Un ejemplo de una situación de partida real es
mosaico estructural inventariado por Namikawa y Kawai (1998) en una parcela de
50 x 200 m en un bosque mixto de coníferas y frondosas en Hokkaido (Japón;
figura 6-19).
140 120 100 80 : Pequeña superficie : Periferia de la pequeña superficie60 : Cubierta de copas interrumpida : Cubierta de copas cerrada 40 20 0 m 50 m
Figura 6-19. Mosaico espacial de diferentes estadios de desarrollo en un bosque mixto de coníferas y frondosas en Hokkaido, Japón (parte de un gráfico de Namikawa y Kawai, 1998).

Crecimiento del Árbol 257
La mayoría de los modelos de pequeñas superficies simulan la dinámica de un rodal
como un vector de pequeñas superficies independientes unas de otras. A cada una
de las pequeñas superficies se le asigna una situación inicial. Para ello se fijan el
número de pies y la distribución espacial de las coordenadas de los árboles, de las
especies, de los diámetros, de las alturas, de las alturas del punto donde comienza la
copa etc. En base a experiencias y mediciones y con la ayuda de un software
adecuado se puede simular el desarrollo de cualquier situación inicial.

Crecimiento del Árbol 258

Bibliografía 259
Bibliografía Aguirre, O., Hui G.Y., Gadow, K.v. y Jiménez, J., 2003: An analysis of spatial forest structure using
neighbourhood-based variables. For. Ecol. Manage. 183, 137-145. Aguirre, O., Kramer, H. y Jiménez, J., 1998: Strukturuntersuchungen in einem Kiefern-
Durchforstungsversuch Nordmexikos. Allgemeine Forst- Und Jagdzeitung 169 (12): 213-219. Akça, A., Gadow, K.v., Mench, A., Mann P., Pahl, A y Setje-Eilers, U., 1994: Überprüfung des
Formquotienten q7 (d7/BHD) in der Bundeswaldinventur für die Hauptbaumarten Buche, Eiche, Fichte und Kiefer im landeseigenen Wald Nordrhein-Westfalens. Project report.
Albert, M., 1999: Analyse der eingriffsbedingten Strukturveränderung und Durchforstungsmodellierung in Mischbeständen. Dissertation, Universität Göttingen. Hainholz-Verlag, Band 6.
Albert, M. y Gadow, K.v., 1998: Assessing Biodiversity with New Neighborhood-based Parameters. En: Laumonier Y., King B., Legg C. y Rennolls K. (eds.): Data Management and Modelling Using Remote Sensing and GIS for Tropical Forest Land Inventory. Rodeo International Publishers, Jakarta.
Albert, M., 1997: Positionsabhängige Einzelbaummodellierung am Beispiel des Simulationsansatzes von Hasenauer. Unveröffentlichtes Manuskript, Institut für Forsteinrichtung u. Ertragskunde, Universität Göttingen.
Albert, M., 1994: Erhebung von Strukturinformationen in einem Buchen-Edellaubholz Mischbestand. Diplomarbeit am Institut für Forsteinrichtung und Ertragskunde, Universität Göttingen.
Alder, D., 1980: Estimación del volumen forestal y predicción del rendimiento. Estudios FAO. Montes 22/2. Roma.
Alisov, B.P., Drosdov, O.A. y Rubinstein, E.S., 1956: Lehrbuch der Klimatologie. VEB Deutscher Verlag der Wissenschaften. Berlin.
Altherr, E., 1963: Untersuchungen über Schaftform, Berindung und Sortimentsanfall bei der Weißtanne. Allgemeine Forst- und Jagdzeitung 134 (5/6), 140-151.
Altherr, E., 1971: Wege zur Buchen-Starkholzproduktion. Bericht zur 15. Hauptversammlung des Baden-Württembergischen Forstvereins: 123-127.
Álvarez González, J.G., 1997: Análisis y caracterización de las distribuciones diametricas de Pinus Pinaster Ait en Galicia. Tesis Doctoral. Univ. Politécnica de Madrid.
Álvarez González, J.G., Schröder, J., Rodríguez Soalleiro, R. y Ruiz González, A.D., 2002: Modelling the effects of thinnings on the diameter distribution of even-aged Maritime pine stands. For. Ecol. Manage. 165(1-3): 57-65.
Álvarez González, J.G., Ruiz González, A.D., Rodriguez Soalleiro, R. y Barrio Anta M., 2005: Ecoregional site index models for Pinus pinaster in Galicia (Northwestern Spain) Ann. For. Sci. 62(2), 115-127.
Amateis, R.L., Burkhart, H.E. y Burk, T.E., 1986: A ratio approach to predicting merchantable yields of unthinned loblolly pine plantations. For. Sci. 32, 187-296.
Assmann, E., 1953: Zur Bonitierung süddeutscher Fichtenbestände. AFZ 10, 61-64. Assmann, E., 1961: Waldertragskunde. Bayrischer Landwirtschaftsverlag, München. Assmann, E. y Franz, F., 1963: Vorläufige Fichtenertragstafel für Bayern. Institut für Ertragskunde der
Forstlichen Forschungsanstalt München. 2. Auflage 1972. Atkins, P. W., 1987: Physikalische Chemie. Verlag VCH, Weinheim. 890 S. Avery, T.E. y Burkhart, H.E., 1994: Forest measurements, 4th Edition, McGraw-Hill. Badoux, E., 1939 : De l'influence de divers modes et degrés d'éclaircie dans les Hetraies pures. Mitt.
Schweiz. Anst. f. d. forstl. Vers. Wesen 21, 59-146. Bailey R.L., 1980: Individual tree growth derived from diameter distribution models. For. Sci. 26: 626–
632. Bailey, R.L. y Dell, T.R., 1973: Quantifying diameter distributions with the Weibull function. For. Sci. 19:
97-104.

Bibliografía 260
Baker, J.B. y Broadfoot, W.M., 1977: A practical field method for site evaluation for eight important Southern hardwoods. USDA For. Serv., Southern Forest Experiment Station, New Orleans, LA, Gen. Tech. Rep. SO-14.
Balboa Murias, M.A., Rodríguez Soalleiro, R., Merino, A. y Álvarez González, J.G., 2006: Temporal variations and distribution of carbon stocks in aboveground biomass of radiata pine and maritime pine pure stands under different silvicultural alternatives. For. Ecol. Manage. 237, 29-38.
Barnes, B.V., Zak, D.R., Denton, S.R. y Spurr, S.H., 1997: Forest Ecology. 4th Edition. John Wiley and sons. New York.
Barrio Anta, M., Balboa Murias, M, Castedo Dorado, F., Dieguez Aranda, U. y Álvarez González, J.G., 2006: An ecoregional model for estimating volume, biomass and carbon pools in maritime pine stands in Galicia (northwestern Spain). For. Ecol. Manage. 223, 24-34.
Barrio Anta, M. y Álvarez González, J.G., 2005: Development of a stand density management diagram for even-aged pedunculate oak stands and its use in designing thinning schedules. Forestry 78(3), 209-216.
Bauhus, J. y Bartsch, N., 1996: Fine-root growth in beech (Fagus sylvatica) forest gaps. Can. J. For. Res. 26, 2153-2159.
Baur, F.v., 1877: Die Fichte in Bezug auf Ertrag, Zuwachs und Form. Berlin. Baur, F.v., 1881: Über Durchforstungen und Durchforstungsversuche. In Ganghofer: Das Forstliche
Versuchswesen, Bd. II, Augsburg. Becker, G. y Seeling, U., 1998: Holzqualität der Fichte. AFZ 53 (8), 434 - 435. Bella, I.E., 1971: A New Competition Model for Individual Trees. For. Sci. 17 (3): 364-372. Bergel, D., 1985: Douglasien-Ertragstafel für Nordwestdeutschland. Niedersächsische Forstliche
Versuchsanstalt, Göttingen. Bertalanffy, L.v., 1948: Quantitative laws in metabolism and growth. Quart. Rev. Biol. 32: 217-230. Biber, P., 1996. Konstruktion eines einzelbaumorientierten Wachstumssimulators für Fichten-Buchen-
Mischbestände im Solling. Diss. Forstw. Fak. d. Ludwig-Maximilians-Univ. München. Biederbick, K.H., 1992: Die direkte Sonnenstrahlung als Standortfaktor. Forstarchiv 63, 131-136. Biging, G.S. y Dobbertin, M., 1992: A comparison of distance-dependent competition measures for height
and basal area growth of individual conifer trees. For. Sci. 38 (3): 695-720. Biging, G.S. y Dobbertin, M., 1995: Evaluation of competition indices in Individual Tree Growth Models.
For. Sci 41 (2), 360-337. Biging, G.S. y Gill, S.J., 1997: Stochastic models for conifer tree crown profiles. For. Sci. 43(1), 25-33. Bolte, A., Rahmann, T., Kuhr, M., Spanuth, P., Murach, D. y Gadow, K. v., 2003: Beziehungen zwischen
Baumdimension und Wurzelstruktur in Fichten-Buchenmischbeständen. Tagungsbericht der Jahrestagung 2002 der Sektion Waldbau im DFFV, Berlin: 85-96.
Bonnemann, A., 1939: Der gleichaltrige Mischbestand von Kiefer und Buche. Mitt. a. Forstwirt. u. Forst-wiss. (5): 439-483.
Bonnemann, A., 1956: Eichen-Buchen-Mischbestände. Allgemeine Forst- und Jagdzeitung 127: 118-126. Bossel, H., 1992: Das Modell FORMIX. Vieweg Verlag, Braunschweig. Bossel, H., 1994a: Modellbildung und Simulation. Konzepte, Verfahren und Modelle zum Verhalten
dynamischer Systeme. Vieweg Verlag, Braunschweig/Wiesbaden. Bossel, H., 1994b: TREEDYN3 Forest Simulation Model. Mathematical model, program documentation
and simulation results. Berichte des Forschungszentrums Waldökosysteme, Reihe B, Bd. 35. Göttingen.
Botkin, D.B., 1993: Forest Dynamics. An ecological model. Oxford University Press, New York. Botkin, D.R., Janak, J.F. y Wallis, J.R., 1972: Some ecological consequences of a computer model of forest
growth. The Journal of Ecology 60: 849-872. Bravo-Oviedo, A. y Montero, G., 2005: Site index in relation to edaphic variables in stone pine (Pinus pinea
L.) stands in south west Spain. Ann. For. Sci. 62 (1): 61-72. Brink, C. y Gadow, K.v., 1986: On the use of growth and decay functions for modelling stem profiles.
EDV in Medizin und Biologie 17(1/2), 20-27. Brokaw, N.V.L., 1982: The definition of treefall gap and its effect on measures of forest dynamics.
Biotropica 14 (2): 158-160. Brown, G. S., 1965: Point density in stems per acre. Forestry Research Notes No. 38, New Zealand, 1965.

Bibliografía 261
Bruchwald, A., Dudzinska, M. y Wirowski, M., 1998: Model wzrostu dla lisciastych gatunkow drzew lesnych. Model wzrostu buka. Typescript in Forest Research Institute in Warsaw.
Bundesministerium für Ernährung, Landwirtschaft und Forsten, 1990: Bundeswaldinventur. Inventur-bericht und Übersichtstabellen für das Bundesgebiet nach dem Gebietsstand vor dem 3.10.1990 ein-schließlich Berlin (West). Gitschel-Werbung, Hamburg.
Burger, H., 1939a: Kronenaufbau gleichaltriger Nadelholzbestände. Mitt. Schweiz. Anst. f. d. forstl. Vers. Wesen 21: 5-58.
Burger, H., 1939b: Baumkrone und Zuwachs in zwei hiebsreifen Fichtenbeständen. Mitt. Schweiz. Anst. f. d. forstl. Vers. Wesen 21: 147-176.
Burkhart, H.E., 1987: Data collection and modelling approaches for forest growth and yield prediction. En: Predicting Forest Growth and Yield - Current Issues, Future Prospects. Inst. of Forest Resources. Univ. of Washington. Contribution Nr. 58, 3-16.
Büttner, V., Leuschner, C., 1994: Spatial and temporal patterns of fine root abundance in a mixed oak beech forest. For. Ecol. Manag. 70, 11-21.
Cajander, A. K., 1909: Über Waldtypen. Acta For. Fenn. Helsingfors. 1 (1): 1-175. Cajander, A. K., 1926: The theory of forest types. Acta For. Fenn. 29 (3): 1-108. Calama, R., Cañadas, N. y Montero, G., 2003: Inter-regional variability in site index models for even-aged
stands of stone pine (Pinus pinea L.) in Spain. Ann. For. Sci. (3): 259-269. Cañellas, I., del Rio, M., Roig, S. y Montero, G.,. 2004: Growth response to thinning in Quercus pyrenaica
Willd. coppice stands in Spanish central mountain. Ann. For. Sci. 61 (3): 243-250. Cañellas, I. y Montero, G., 2002: The influence of cork oak pruning on the yield and growth of cork. Ann.
For. Sci. 59 (7): 753-760. Cao, Q.V., Burkhart, H.E. y Lemin, Jr., 1982: Diameter distributions and yields of thinned loblolly pine
plantations. VPI & SU, Sch. For. & Wildl. Resour. Publ FWS-1-82. Cao Q.V. y Baldwin V.C., 1999: A new algorithm for stand table projection models. For. Sci. 45: 506 –
511. Carbonnier, C., 1971: Yield of beech in Southern Sweden. Studia Forestalia Suecica Nr. 91. Royal College
of Forestry, Stockholm. Carlyle, J.C., 1986: Nitrogen cycling in forested ecosystems. For. Abstracts 47, 307-336. Carmean, W. H., 1973: Forest Soils Bibliography for the North Central Region. USDA For. Serv. Tech.
Rep. NC-5. Castedo Dorado, F., Barrio Anta, M., Parresol, B.R. y Álvarez González, J.G., 2005: A stochastic height-
diameter model for maritime pine ecoregions in Galicia (northwestern Spain). Ann. For. Sci. 62, 455-465.
Castedo Dorado, F., Diéguez Aranda, U. y Álvarez González, J.G., 2007: A growth model for Pinus radiata D. Don stands in north-western Spain. Ann. For. Sci. 64(4), 453-466.
Černý, M., Parez, J. y Malik, Z., 1996: Rustové a taxační tabulky hlavních drevin. Ministerstva zemedelstiví.
Chen S.J. y Hwang C.L., 1992. Fuzzy multiple attribute decision making - methods and applications. Lecture Notes in Economics and Mathematical Systems, Vol. 375. Springer Verlag, Berlin.
Chikumbo, O., Mareels, I.M.Y. y Turner, B.J., 1992: Integrating the Weibull into a dynamic model to predict future diameter distributions. In: Wood G.B. y Turner, B.J., 1992: Integrating forest informa-tion over space and time. Proc. IUFRO Conf., Australian National Univ., Canberra: 94-102.
Chroust, L., 1968: Das Temperaturregime in verschieden durchforsteten Eichen-Stangenhölzern. Allgemeine Forst –und Jagdzeitung 139: 163-173.
Chung, D.J., 1996: Konkurrenzverhältnisse und Struktur natürlicher Pinus densiflora - Quercus variabilis - Mischwälder in Korea. Diss. Fak. f. Forstw. u. Waldökologie, Universität Göttingen.
Cieszewski, C.J., 2000: Analytical solution to the generalized log-logistic equation. For. Sci. 46: 291-296. Cieszewski, C.J., 2001: Three methods of deriving advanced dynamic site equations demonstrated on
inland Douglas-fir site curves. Can. J. For. Res. 31: 165-173. Cieszewski, C.J., 2002: Comparing fixed- and variable-base-age site equations having single versus multiple
asymptotes. For. Sci. 48: 7-23. Cieszewski, C.J., 2003: Developing a well-behaved dynamic site equation using a modified Hossfeld IV
function Y3 = (axm)/(c + xm-1), a simplified mixed-model and scant subalpine fir data. For. Sci. 49: 539-554.

Bibliografía 262
Cieszewski, C.J., 2004: GADA derivation of dynamic site equations with polymorphism and variable asymptotes from Richards, Weibull, and other exponential functions. University of Georgia PMRC-TR 2004-5.
Cieszewski, C.; y Bella, J., 1989: Polimorphic height and site index curves for lodgepole pine in Alberta. Canadian Journal of Forest Research 19: 1151-1160.
Clark P.J. y Evans F.C., 1954: Distance to nearest neighbour as a measure of spatial relationships in populations. Ecology 35, 445-453.
Clutter, J.L., 1980: Development of taper functions from variable-top merchantable volume equations. For. Sci. 26(1), 117-120.
Clutter, J. L. y Jones, E. P., 1980: Prediction of Growth after Thinning in Old-Field Slash Pine Plantations. USDA For. Serv. Res. Paper 217 S.
Clutter, J.L., Fortson, J.C., Pienaar, L.V., Brister, G.H. y Bailey, R.L., 1983: Timber management - a quantitative approach. John Wiley, New York.
Condes, S. 1997: Simulación de parcelas arboladas con datos del 2 Inventario Forestal Nacional. Tesis Doctoral. Escuela Técnica Superior de Ingenieros de Montes de Madrid.
Colin, F., Houllier, F., 1991: Branchiness of Norway spruce in north-eastern France: modelling vertical trends in maximum nodal branch size. Annales des Sciences Forestieres 48, 679 - 693.
Colin, F., Houllier, F., 1992: Branchiness of Norway spruce in north-eastern France: predicting the main crown characteristics from usual tree measurements. Annales des Sciences Forestieres 49, 511-538.
Corral Rivas, J.J., Álvarez González, J.G., Ruíz González, A.D. y Gadow, K.v., 2004: Compatible height and site index models for five pine species in El Salto, Durango (Mexico). Forest Ecology and management, 201(2-3), 145-160.
Corral Rivas, J.J., Álvarez González, J.G., Aguirre, O. y Hernández, F.J., 2005: The effect of competition on individual tree basal area growth in mature stands of Pinus cooperi Blanco in Durango (Mexico). Eur. J. Forest Res. 124, 133-142.
Courbaud, B., 1995: Modélisation de la croissance en forêt irrégulière, - perspectives pour les pessières irrégulières de montagne. Rev. For. Fr. XLVII No. sp.: 173.
Coutts, M. P., 1983: Root architektur and tree stability. Plant and Soil 56; 171-188.Cowley, J. M., 1975: Diffraction Physics. North Holland.
Craib I.J., 1939.:Thinning, Pruning and management studies on the main exotic conifers grown in South Africa. Govt. Printer, Pretoria.
Daniel, T., Helms, J.A. y Bakeer, F.S., 1979. Principles of Silviculture. 2nd Edition. McGraw-Hill. Daniels, R. F., 1976: Simple Competition Indices and their Correlation with Annual Loblolly Pine Tree
Growth. Forest Science 22(4): 454-456. Degenhardt, A., 1995: Analyse der Entwicklung von Bestandesstrukturen mit Hilfe des Modells der
zufälligen Punktprozesse in der Ebene. Tagungsbericht, Dt. Verb. Forstl. Forschungsanstalten - Sektion Biometrie u. Informatik, 8.-10.9.93 in Freising: 93-105.
Demaerschalk, J.P., 1973: Integrated systems for the estimation of tree taper and volume. Can. J. For. Res. 3 (1), 90-94.
Dengler, A., 1982: Waldbau auf ökologischer Grundlage. Bearbeitet von E. Röhrig und H. A. Gussone. Verlag Paul Parey, Hamburg und Berlin, 275 S.
Dhôte, J. F., 1996: A model of even-aged beech stands productivity with process-based interpretations. Ann. Sc. For. 53, 1-20.
Dieguez Aranda, U., Álvarez González, J.G., Barrio Anta, M. y Rojo Alboreca A., 2005: Site quality equations for Pinus sylvestris L. plantations in Galicia (northwestern Spain). Ann. For. Sci. 62(2), 143-152.
Diéguez Aranda, U., Castedo Dorado, F., Álvarez González, J.G. y Rojo Alboreca, A., 2006a: Compatible taper function for Scots pine (Pinus sylvestris L.) plantations in northwestern Spain. Can. J. For. Res. 36, 1190-1205.
Diéguez Aranda, U., Grandas Arias, J.A., Álvarez González, J.G. y Gadow K.v., 2006b: Site quality curves for Birch stands in North-Western Spain. Silva Fennica 40(4), 631-644.
Dittmar, O., Knapp, E. y Lembcke, G., 1986: DDR-Buchenertragstafel 1983. IFE-Berichte. Döbbeler, H. y Spellmann H., 2002: Methodological approach to simulate and evaluate silvicultural
treatments under climate change. Fw. Cbl. 121, Supplement 1, 52-69.

Bibliografía 263
Dralle, K., 1997: Locating trees by digital image processing of aerial photos. Dina Research Rapport No. 58: 116 p.
Drexhage M., 1994: Die Wurzelentwicklung 40 jähriger Fichten (Picea abies [L.]) in der Langen Bramke (Harz). Berichte des Forschungszentrums Waldökosysteme, Reihe A, Band 111, Göttingen.
Ďurský J. 2000: Einsatz von Waldwachstumssimulatoren für Bestand, Betrieb und Großregion. Dissertation, Technische Universität München.
Eberhard J., 1902: Tafeln zur Bonitierung und Ertragsbestimmung nach Mittelhöhen [Höhen-Ertragskurven] für Tanne, Fichte, Forche, Buche und Eiche. Selbstverlag d. Verfassers.
Eichhorn, F., 1904: Beziehungen zwischen Bestandeshöhe und Bestandesmasse. Allgemeine Forst –und Jagdzeitung 80: 45-49.
Eichhorn, J., 1992: Wurzeluntersuchungen an sturmgeworfenen Bäumen in Hessen. Forst und Holz 47(18): 555-559.
Ek, A. R. y Monserud, R. A., 1974: Trials with program Forest: Growth and reproduction simulation for mixed species even- or uneven-aged forest stands. In: Fries, J. (ed.): Growth models for tree and stand simulation, Royal College of Forestry, Research Notes, Nr. 30: 56-73.
Ekö, P.M. y Ågestam, E., 1994: A comparison of naturally regenerated and planted Scots pine (Pinus sylvestris L.) on fertile sites in southern Sweden. For u. Landsc. Res. 1:III-126.
Ellenberg, H., 1982: Ökosystemforschung. Ergebnisse des Sollingprojekts: 1966-1986. Verlag Ulmer. Stuttgart.
Ellenberg, H., 1996: Vegetation Mitteleuropas mit den Alpen in ökologischer Sicht. 6. Auflage. Ulmer, Stuttgart.
Ellenberg, H., Weber, H. E., Düll, R., Wirth, V., Werner, W. y Paulissen, D., 1991: Zeigerwerte von Pflanzen Mitteleuropas. Scripta Geobotanica XVIII. Goltze, Göttingen.
Embrog, J., Christensen, M. y Heilmann-Clausen, J., 2000: The structural dynamics of Suserup Skov, a near-natural temperate deciduous forest in Denmark. For. Ecol. Manage. 126, 173-189.
Evers, F. H. y Moosmayer, H.-U., 1980: Zusammenhänge zwischen Standortseinheiten, Nährstoffverhältnissen des Bodens und Wachstum von Fichtenbeständen im regionalen Vergleich. Fw. Cbl. 99: 137-146.
Faber, R. J., 1987: The Japanese Larch in the Netherlands - a new growth prediction. Ned. Bosbouw-tijdskrift 59 (1/2): 13-27.
Fähser, L., 1998: Naturnahe Waldnutzung – das Beispiel Lübeck. Handbuch Kommunale Politik Raabe: S. 1-17.
Falinski, J. B., 1988: Succession, regeneration and fluctuation in the Bialowieza Forest. Vegetatio 77: 115-128.
FAO, 1997: State of the World’s Forests. Rom. FAO, 2001: Global Forest Resources Assessment – Main Report, Rom. Fehrmann, L., Kuhr, M. y Gadow, K.v., 2003: Zur Analyse der Grobwurzelsysteme großer Waldbäume an
Fichte [Picea abies (L.) Karst.] und Buche [Fagus sylvatica]. Forstachiv 74(3), 96-102. Fiedler, H. J., Nebe, W. y Hoffmann, F., 1973: Forstliche Pflanzenernährung und Düngung. Verlag G.
Fischer, Jena. Figueiredo-Filho, A., Borders, B.E. y Hitch, K.L., 1996: Taper equations for Pinus taeda plantations in
Southern Brazil. For. Ecol. Manage. 83: 39-46. Flury, Ph., 1929: Über den Aufbau des Plenterwaldes. Mitteilungen der schweizerischen Centralanstalt für
das Versuchswesen, Band 15: 305-357. Fogel, R., 1983: Root turnover and productivity of coniferous forests. Plant and soil 71, 75-85. Forslund, R.R., 1982: A geometrical tree volume model based on the location of center of gravity of the
bole. Can. J. For. Res. 12, 215-221. Forss, E., 1994: Das Wachstum der Baumart Acacia mangium in Südkalimantan, Indonesien. Magister Diss.,
Forstw. Fak., Univ. Göttingen. Forss, E., Gadow, K.v. y Saborowski, J., 1996: Growth models for unthinned Acacia mangium plantations
in South Kalimantan, Indonesia. J. of Trop. For. Sci. 8 (4), 449-462. Franz, F., Bachler, J., Deckelmann, B., Kennel, E., Kennel, R., Schmidt, A., Wotschikowski, U., 1973:
Bayrische Waldinventur 1970/71 - Aufnahme und Auswertungsverfahren. Forstl. Forschungsanstalt München. Forschungsbericht Nº. 11.
Franz, F., 1972: Ertragskundliche Prognosemodelle. FwCbl 91: 65-80.

Bibliografía 264
Frauendorfer, 1959: Fichtenertragstafel. In: Marschall, J., 1975: Hilfstafeln für die Forsteinrichtung. Österr. Forstverein, 2. Aufl., Wien.
Freise, C. y Spieker, H., 1999: Konkurrenzfreies Wachstum der Buche. AFZ/Der Wald, 25: 1346-1349 Freist, H., 1962: Untersuchungen über den Lichtungszuwachs der Rotbuche und seine Ausnutzung im
Forstbetrieb. Forstwiss. Forschungen Nr. 17. Paul Parey, Berlin. Fritz, H.W., 1999: Feinwurzel- Verteilung, -Vitalität, -Produktion und -Umsatz von Fichten (Picea abies [L.]
Karst.) auf unterschiedlich versauerten Standorten (ein Beitrag zur ökosystemorientierten Waldschadensforschung). Berichte des Forschungszentrums Waldökosysteme, Reihe A, Band 165, Göttingen.
Früh, T., 1992: C-Bilanz auf der Basis der Pipe-Modell-Theorie, Produktionsmodell. Berichte des For-schungszentrums Waldökosysteme (Göttingen), Band 31, 355-367.
Fujimori, T. y Kiyono Y., 1986: Dynamics of crown structure and effects of crown control on tree growth in Cryptomeria japonica. En Fujimori T.,. Whitehead (eds). Crown and canopy structure in relation to productivity. Published by the Forestry and Forest Products Institute, Ibaraki, Japan.
Füldner, K. y Gadow, K. v., 1994: How to Define a Thinning in a Mixed Deciduous Beech Forest. Proc. IUFRO Conference in Lousa, Portugal: Mixed stands - research plots, measurements and results, models, 1994: 31-42.
Füldner, K., 1995: Strukturbeschreibung von Buchen-Edellaubholz-Mischwäldern. Diss., Fak. f. Forstw. u. Waldökologie, Universität Göttingen. Cuvillier Verlag, Göttingen.
Gadow, K.v., 1987: Untersuchungen zur Konstruktion von Wuchsmodellen für schnellwüchsige Plantagenbaumarten. Forstl. Forschungsberichte, Universität München Nr. 77.
Gadow, K.v., 1996: Modelling growth in managed forests - realism and limits of lumping. The Science of the Total Environment 183, 167-177.
Gadow, K.v., 1999: Waldstruktur und Diversität. Allg. Forst- u. J.- Zeitung 170, 117-122. Gadow, K.v., 1992: Ein Wachstums- und Ertragsmodell für die Fortschreibung von Bestandesparametern.
In: Preuhsler, T., Röhle, H., Utschig, H. y Bachmann, M. (Hrsg.): Festschrift zum 65. Geburtstag von Prof. Franz; Lehrstuhl f. Waldwachstumskunde, Universität München, 75-83.
Gadow, K.v., 1984: The relationship between diameter and diameter increment in Pinus patula. Proceedings of the IUFRO Conference "Site and Productivity of Fast-growing Plantations", held at Pretoria 1984, Vol. 2: 741-751.
Gadow, K.v. y Heydecke, H., 2000: Wachstum und Durchforstung in einem Buchenmischbestand. Forst und Holz 56 (3): 86-88.
Gadow, K.v., Heydecke, H. y Riemer, Th., 1996: Zur Beschreibung der Schaftprofile stehender Wald-bäume. Festschrift A. Akça, Inst. f. Forsteinrichtung u. Ertragskunde, Universität Göttingen: 31-44.
Gadow, K.v. y Hui, G. Y., 1993: Stammzahlentwicklung und potentielle Bestandesdichte bei Cunninghamia lanceolata. Cbl. ges. Forstw. 110 (2): 41-48.
Gadow, K.v. y Hui, G. Y., 1998: Modelling forest development. Kluwer Academic Publishers. Gadow, K.v., Hui, G.Y. y Albert, M., 1998: Das Winkelmaß - ein Strukturparameter zur Beschreibung der
Individualverteilung in Waldbeständen. Cbl. ges. Forstw. 115, 1- 10. Gadow, Kv., Hui, G.Y., Chen, B.W. y Albert, M., 2003: Beziehungen zwischen Winkelmaß und
Baumabständen. Forstw. Cbl. 122, 127-137. Gadow, K.v. y Postoli, A., 1998: Waldwachstum: Modelle der Waldentwicklung. Skript aus dem Institut
für Forsteinrichtung und Ertragskunde. Georg-August-Universität Göttingen, 201 S. Gadow, K.v. y Schmidt, M., 1998: Periodische Inventuren und Eingriffsinventuren. Forst und Holz, 22,
667-671. Gadow, K.v. y Stüber, V., 1994: Die Inventuren der Forsteinrichtung. Forst und Holz 49 (5): 129-131. Gaffrey, D., 1988: Forstamts- und bestandesindividuelles Sortimentierungsprogramm als Mittel zur
Planung, Aushaltung und Simulation. Diplomarbeit, Fachbereich Forstwissenschaft, Universität Göttingen.
Gaffrey, D., 1996: Sortenorientiertes Bestandeswachstums-Simulationsmodell auf der Basis intraspezifischen, konkurrenzbedingten Einzelbaumwachstums - insbesondere hinsichtlich des Durchmessers- am Beispiel der Douglasie. Diss. Forschungszentrum Waldökosysteme d. Uni. Göttingen.

Bibliografía 265
García, O., 1988: Experience with an advanced growth modelling methodology. In: Ek, A.R., Shifley, S.R. y Burke, T.E. (eds): Forest growth modelling and prediction. USDA For. Serv. Gen. Techn. Rep. NC-120: 668-675.
García, O., 1994: The state space approach in growth modelling. Can. J. For. Res. 24, 1894-1903. Gates, D. M., 1980: Biophysical ecology. Springer Verlag, New York. Gatzojannis, S., 1999: Yield tables for beech stands in the forest A. Brontou Serres Prefecture, N. Greece
(en griego). ΔΑΣΙΚΗ ΕΡΕΨΝΑ12: 91-104. Gehrhardt, E., 1930: Ertragstafeln für reine und gleichaltrige Hochwaldbestände von Eiche, Buche,
Tanne, Fichte, Kiefer, grüner Douglasie und Lärche. 2. Aufl. Julius Springer Verlag, Berlin. Gehrke, J.T., Schlaghamersky, A. y Sterzik, H.K., 1992: Tragfähigkeit und Befahrbarkeit von Waldböden.
Forst und Holz 8, 205-207. Gehrmann, J., 1984: Einfluss von Bodenversauerung und Kalkung auf die Entwicklung von
Buchenverjüngungen im Wald. Bericht des Forschungszentrums Waldökosysteme/Waldsterben 1. Gerold, D., 1990: Modellierung des Wachstums von Waldbeständen auf der Basis der
Durchmesserstruktur. Dissertation, B. TU Dresden. Gerrard, D. J., 1969: Competition quotient - a new measure of the competition affecting individual forest
trees. Mich. State Univ. Agr. Exp. Stn. Res. Bull. No. 20. Giurgiu, V., Decei, I. y Armasescu, S., 1972: Biometria arborilor ¸si arboretelor din România : Tabele
dendrometrice. Ceres Verlag. Golser, M. y Hasenauer, H., 1997: Predicting juvenile tree height growth in uneven-aged mixed species
stands in Austria. For. Ecol. Manage. 97: 133-146. Göttsche, D., 1972: Verteilung von Feinwurzeln und Mykorrhizen im Bodenprofil eines Buchen- und
Fichtenbestandes im Solling. Mitt. Bundesforschungsanstalt f. Forst- u. Holzwirtschaft 88, 102. Goulding, C. J., 1972: Simulation technique for a stochastic model of growth of Douglas-fir. Ph. D. thesis.
Univ. of Brit. Col. Vancouver. Grace, J.C. y Pont, D., 1997: Modelling Branch Development in Radiata Pine. En: Amaro, A. y Tomé, M.
(ed.), Empirical and process-based models for forest tree and stand growth simulation, 21-27 September 1997, Oeiras, Portugal, 173-184.
Grammel, R., 1989: Forstbenutzung. Verlag Paul Parey, Hamburg. Gregar, T., 1992: Flora und Vegetation des Schlitzerlandes. Diss. TU Berlin. Gregoire, T.G., Valentine, H.T. y Furnival, G.M., 1986: Estimation of bole volume by importance
sampling. Can. J. For. Res. 16, 554-557. Gregorius, H.R., 1974: Genetischer Abstand zwischen Populationen. Zur Konzeption der genetischen
Abstandsmessung. Silvae Genetica 23, 22-77. Groll, M., 1996: Boden-, Bestandes- und Wurzelschäden beim Einsatz eines Harvestersystemes. Forst und
Holz 12, 409. Gruber, F., 1992: Dynamik und Regeneration der Gehölze. Berichte des Forschungszentrums
Waldökosysteme, Reihe A, Band 86, Göttingen. Grundner, F., 1904: Untersuchungen im Buchenhochwalde über Wachstumsgang und Massenertrag.
Springer Verlag, Berlin. Grundner, F. y Schwappach, A., 1942: Massentafeln zur Bestimmung des Holzgehaltes stehender
Waldbäume und Waldbestände. Berlin, 9. Auflage. Gualdi, V., 1974: Ricerche auxometriche sulle faggete del Gargano. L’Italia Forestale e Montana. 29:85-
116. Gundersen, P., 1995: Impacts of nitrogen deposition - scientific background. UN ECE Convention on
Long Range Transboundary Air Pollution. Annual Synoptic Report, 9-18. Gurjanov, M., Sánchez Orois, S. y Schröder, J. 2000: Grundflächenmodelle für gleichaltrige
Fichtenreinbestände - Eine vergleichende Analyse. Cbl. ges. Forstwes. 117 3/4, 187-198. Háckel, H., 1993: Meteorologie. Verlag Ulmer, Stuttgart. Hafley, W. L. y Schreuder, H. T., 1977: Statistical distributions for fitting diameter and height data in even-
aged stands. Canadian Journal of Forest Research 7: 481-487. Hagglund, B., 1981: Evaluation of forest site productivity. Review article. For. Abstr. 42(11): 515-527. Hahn, J. T. y Leary, R. A., 1979: Potential diameter growth functions. In: A generalized forest growth
projection system applied to the lake states region. USDA For. Serv., Gen. Tech. Rep. NC-49: 22-26.

Bibliografía 266
Haith D. A., Tubbs, L. J. y Pickering, N. B., 1984: Simulation of pollution by erosion and soil nutrient loss. Pudoc. Centre for Agriculture Publishing and Documentation, Wageningen.
Hakes, W., 1994: On the predictive power of numerical and Braun-Blanquet classification: an example from beechwoods. Journal of Vegetation Science 5: 153-160.
Halaj, J. y Petráš, R., 1998: Rastové tabuľky hlavných drevín. SAP – Slovac Academic Press, Bratislava. Hamilton, G.J. y Christie, J.M., 1971: Forest management tables (metrics). For. Comm. booklet n° 34.
London. Hánninen, H., 1990: Modelling bud dormancy release in trees from cool and temperate regions. Acta
Forestalia Fennica 213, 1-47. Hapla, F., 1986: Beeinflussen unterschiedliche Durchforstungsmaßnahmen die Holzeigenschaften der
Douglasie? Forstarchiv 57 (3), 99-104. Hartig, R., 1868: Die Rentabilität der Fichtennutzholz- und Buchenbrennholzwirtschaft im Harze und im
Wesergebirge. Stuttgart. Hartig, TH., 1847: Vergleichende Untersuchungen über den Ertrag der Rotbuche. Verlag A. Förstner.
Berlin. Hasenauer, H. y Monserud, R.A., 1997: Biased predictions for tree height increment models developed
from smoothed data. Ecol. Modelling 98, 13-22. Hasenauer, H., 1994: Ein Einzelbaumwachstumssimulator für ungleichaltrige Fichten-, Kiefern- und
Buchen-Fichtenmischbestände. Forstl. Schriftenreihe d. Universität f. Bodenkultur, Wien, Bd. 8. Hasenauer, H., Moser, M. y Eckmüllner, O., 1994: Programmbeschreibung. Distanzabhängiger Wachs-
tumssimulator für Mischbestände, MOSES 2.0. Inst. f. Waldwachstumsforschung, Universität f. Bodenkultur, Wien.
Hasenauer, H., Moser, M. y Eckmüllner, O., 1995: MOSES - a computer simulation program for modelling stand response. En: Pinto da Costa M.E., Preuhsler T. (Eds.): Mixed stands, research plots, and results, models. Instituto Superior de Agronomia, Universidade Tecnica de Lisboa, Portugal.
Hauhs, M., Rost-Siebert, K., Kastner-Maresch, A. y Lange, H., 1993: A New Model Relating Forest Growth to Input and Output Fluxes of Energy and Matter of the Corresponding Ecosystem (TRAGIC). Final Report of Project EV4V-0032-D(B), Faculty of Forestry, Univ. of Göttingen.
Hegyi, F., 1974: A simulation model for managing jack-pine stands. 74-76p in Growth models for tree and stand simulation, Fries, J. (ed.). Royal Coll. of For., Stockholm.
Heinrich, D. y Hergt, M., 1990: dtv-Atlas zur Ökologie. Deutscher Taschenbuchverlag, München. Henderson, G. S., Hammer, R. D. y Grigal, D. F., 1990: Can measurable soil properties be integrated into
a framework for characterizing forest productivity? In: Gessel, S. P., Lacate, D. S., Weetman, G. F. and Powers, R. F. (eds): Sustained productivity of forest soils. Proc. 7th North Am. Forest soils conf. Vancouver: 137-154.
Hendriks, C.M.A. y Bianchi, F.J.J.A., 1995: Root density and root biomass in pure and mixed forests of Douglas fir and beech. Netherlands Journal of agricultural Science 43 (3), 321-331.
Hengst, E., 1959: Allgemeine Bemerkungen zur Weihmouthskiefer und ihrer Form. Archiv für Forstwesen 8, 781-811.
Hertel, D., 1999: Das Feinwurzelsystem von Rein- und Mischbeständen der Rotbuche: Struktur, Dynamik und interspezifische Konkurrenz. Diss. Bot. 317.
Hertstein, U. y Jáger, H.J., 1995: Wirkungen der sich ändernden Atmosphäre (CO2, O3, verstärkte UV-Strahlung) auf die Vegetation. In: Klimaänderungen und Naturschutz. Angewandte Landschaftsöko-logie, Heft 4. Bundesamt für Naturschutz, Bonn - Bad Godesberg.
Hessen, 1993: Richtlinien für die Bewirtschaftung des Staatswaldes (RIBES 93). Hrsg.: Ministerium für Landesentwicklung, Wohnen, Landwirtschaft, Forsten und Naturschutz.
Hessenmöller, D., 2001: Modelle zur Wachstums- und Durchforstungssimulation im Göttinger Kalkbuchenwald. Diss., Fak. f. Forstw. u. Waldökologie, Universität Göttingen. Logos Verlag, Berlin.
Hessenmöller, D. Y Gadow, K.v., 2001: Beschreibung der Durchmesserverteilung von Buchenbeständen mit Hilfe der bimodalen Weibullfunktion. Allgemeine Forst- und Jagdzeitung 172 (3), 46-50.
Hessische Forsteinrichtungsanstalt, 1985: Hessische Anweisung zu Forsteinrichtungsarbeiten (HAFEA). Staatsanzeiger für das Land Hessen Nr. 32.

Bibliografía 267
Holmes, M. J. y Reed, D. D., 1991: Competition Indices for Mixed Species Northern Hardwoods. Forest Science 37(5): 1338-1349.
Hubbell, S.P., 2001: The unified Neutral Theory of Biodiversity and Biogeography. Princeton University Press. Princeton.
Hui, G.Y. y Gadow, K.v., 1993a: Zur Entwicklung von Einheitshöhenkurven am Beispiel der Baumart Cunninghamia lanceolata. Allgemeine Forst- und Jagdzeitung 164, 218-220.
Hui, G.Y. y Gadow, K.v., 1993b: Zur Modellierung des Oberhöhenwachstums bei Cunninghamia lanceolata. FA 64: 311-313.
Hui, G.Y., Gadow, K.v., 1993c: Zur Modellierung der Bestandesgrundflächenentwicklung - dargestellt am Beispiel der Baumart Cunninghamia lanceolata. Allgemeine Forst- und Jagdzeitung 164, 144-149.
Hui, G.Y. y Gadow, K.v., 1996: Ein neuer Ansatz zur Modellierung von Durchmesserverteilungen. Cbl. ges. Forstw. 113 (¾): 101-113.
Hui, G.Y., 1997: Wuchsmodelle für die Baumart Cunninghamina lanceolata. Diss., Fak. f. Forstw. u. Waldökologie, Universität Göttingen. Cuvillier Verlag, Göttingen, 121 S.
Hui, G.Y., Gadow, K.v., 1997: Entwicklung und Erprobung eines Einheitsschaftmodells für die Baumart Cunninghamia lanceolata. Forstw. Cbl. 116, 315-321.
Hui, G.Y., Albert, M. y Gadow, K.v., 1998: Das Umgebungsmaß als Parameter zur Nachbildung von Bestandesstrukturen. Forstw.Cbl. 117(5), 258 - 266.
Hui, G. Y. y Gadow, K. v., 2002: Das Winkelmaß – Herleitung des optimalen Standardwinkels. Allgemeine Forst u. Jagdzeitung. 173(10).
Husch, B., Miller, C.I. y Beers, T.W., 1982: Forest Mensuration. John Wiley & Sons, New York. Hussein, K.A., Albert, M., Gadow, K.v., 2000: The crown window - a simple device for measuring tree
crowns. Forstw. Cbl. 119, 43-50. Hussein, K.A., 2001: Parameter-parsimonious models for crown and stem profiles. Diss. Univ. of
Göttingen. Hutchison, A. y Matt, D.R., 1977: The distribution of solar radiation within a deciduous forest. Ecol.
Monogr. 47, 185-207. Hyink, D. M., 1979: A generalized method for the projection of diameter distributions applied to uneven-
aged forest stands. Unpublished Ph. Diss., Purdue University. Hyink, D. M., 1980: Diameter distributions approaches to growth and yield modelling. Proceedings of a
Workshop on Forecasting stand dynamic. Lakehead University, Thunder Bay, Ontario, June 24-25. Hyink, D. M., y Moser, J. W., 1983: A generalized framework for projecting forest yield and stand
structure using diameter distributions. For. Sci. 29 (1): 85-95. Itô H., Sumida, A., Isagi, Y. y Kamo, K., 1997: The crown shape of an evergreen Oak, Quercus glauca, in a
hardwood community. J. For. Res. 2, 85-88. Jack, W. H., 1967: Single tree sampling in even-aged plantations for survey and experimentation. IUFRO-
Tagungsband, Sektion 25, München: 379-403. Jäde, K., 1995: Ein Algorithmus zur quantitativen Beschreibung der Kronenstruktur von Nadelbäumen.
Diplomarbeit. Inst. f. Numerische u. Angewandte Mathematik (unter Mitwirkung d. Inst. f. Forstein-richtung u. Ertragskunde). Universität Göttingen.
Jansen, J.J., Sevenster, J. y Faber, P.J., 1996: Opbrengsttabellen voor belangrijke boomsoorten in Nederland. Hinkeloord Report Nr. 17, Landbouwuniversiteit, Wageningen.
Jansen, M. y Martin, G., 1995: Anwendung des Bestandessimulationsmodells TREEDYN3 auf die Fichten-Versuchsfläche Solling F1. Berichte d. Forschungszentrums Waldökosysteme d. Universität Göttingen, Reihe B, Bd. 45, 1-79.
Jensen, M. y Hofmann, G., 1997: Entwicklungszyklen des baltischen Buchenwaldes. AFZ/Der Wald 19: 1012-1018.
Jenssen, M. y Hofmann, G., 2002: Pflanzenartenvielfalt, Naturnähe und ökologischer Waldumbau. AFZ/Der Wald 8, 402-405.
Johnson, N.L., 1949: Systems of frequency curves generated by methods of translation. Biometrika 36, 149-176.
Kahn, M., 1994: Modellierung der Höhenentwicklung ausgewählter Baumarten in Abhängigkeit vom Standort. Dissertation, Forstwissenschaftliche Fakultät, Universität München.
Kahn, M., 1995: Quasikausale Modellierung des Standort-Leistungsbezuges als Voraussetzung zum Aufbau flexibler Mischbestands-Modelle. Forstw. Cbl. 114, 175-187.

Bibliografía 268
Kändler, G., 1986: Ermittlung von Bestandesparametern als Eingangsgrößen für Interzeptionsmodelle mit Hilfe aerophotogrametrischer Verfahren. Mitteilungen der Forstlichen Versuchs- und Forschungsanstalt Baden-Württemberg 127.
Kassier, H. W., 1993: Dynamics of Diameter and Height Distributions in Commercial Timber Plantations, PhD dissertation, Faculty of Forestry, Univ. of Stellenbosch, South Africa.
Keller, W., 1978: Einfacher ertragskundlicher Bonitätsschlüssel für Waldbestände in der Schweiz. Mitt. d. Eidgen. Anst. f. d. Versuchswesen, Bd. 54 (1): 1-98.
Kellomäki, S., 1986: A model for the relationship between branch number and biomass in Pinus sylvestris crowns and the effect of crown shape and stand density on branch and stem biomass. Scand. J. For. Res. 1, 455-472.
Kellomäki, S. y Kurttio, O., 1991: A model for the structural development of a Scots pine crown based on modular growth. For. Ecol. Manage. 43, 103-123.
Kellomäki, S. y Strandman, H., 1995: A model for the structural growth of young Scots pine crowns based on light interception by shoots. Ecol. Modelling 80, 237-250.
Kellomäki, S., Väisänen, H. y Strandman, H., 1993: FINNFOR: - A model for calculating the response of a boreal forest ecosystem to climate change. Univ. of Joensuu. Faculty of Forestry. Research Notes 6.
Kenk, G. y Weise, U., 1998: Zu: Beobachtungen zur Bestandesstruktur undurchforsteter Fichtenbestände. AFZ/Der Wald 53: 937-939.
Kennel, R., 1972a: Die Buchendurchforstungsversuche in Bayern von 1870 bis 1970. Forschungsberichte der forstl. Forschungsanstalt München Nr. 7.
Kennel, R., 1972b: Die Bestimmung des Ertragsniveaus bei der Buche. Dt. Verb. Forstl. Forsch. Anst. 48-50, Neustadt a. d. W.
Kern, K.G., Moll, W. y Braun, H.J., 1961: Wurzeluntersuchungen in Rein- und Mischbeständen des Hochschwarzwaldes (Vfl. Todtmoos 2/I-IV). Allgemeine Forst- und Jagdzeitung , 132, 241-260.
Kienast, F. y Kuhn, N., 1989: Simulating forest succession along ecological gradients in Southern Central Europe. Vegetatio 79: 7-20.
Kimmins J.P., 1987. Forest Ecology. Macmillan Publishing Company, New York. Kimmins, J.P., 1990: Modeling the sustainability of forest production and yield for a changing and
uncertain future. For. Chron. 66, 271-280. Kimmins, J.P., 1997: Forest Ecology 2nd edition. Macmillan Publishing Company, New York. Kiviste, A., Álvarez, J. G., Rojo, A., González, A. D., 2002: Funciones de crecimiento de aplicación en el
ámbito forestal. Comunicaciones I.N.I.A. Serie: Recursos Naturales. No. 4. Klädtke, J., 1992: Konstruktion einer Z-Baum-Ertragstafel am Beispiel der Fichte. Diss., Forstl. Fak.,
Univ. Freiburg. Klier, G., 1969: Beitrag zu den Baumabstandsmethoden in der Forstwirtschaft. Arch. Forstwes., Berlin 18
Jg. Heft 12, 1257-1264. Knoke, T. y Plusczyk, N., 2001: On economic consequences of transformation of a spruce (Picea abies (L.)
Karst.) dominated stand from regular into irregular age structure. For. Ecol. Manage. 151: 163-179. Kolström, T., 1992: Dynamics of uneven-aged stands of Norway Spruce - a Model Approach. Diss., Univ.
Joensuu. Finnland. Konitzer, A., 2000: Waldumbaumassnahmen und ihre betriebswirtschaftlichen Auswirkungen – eine GIS-
gestützte Untersuchung am Beispiel des Niedersächsischen Harzes. Diss., Univ. Göttingen. Koop, P H., 1989. Forest Dynamics. Springer Verlag, Berlin. Korol, M. y Gadow, K.v., 2002: Zur quantitativen Beschreibung von Fichtenkronen im Gebiet der
ukrainischen Karpaten. Unveröff. Manuskript, Inst. f. Waldinventur u. Waldwachstum, Univ. Göttingen.
Korol, M. y Gadow, K.v., 2003: Ein Einheitsschaftmodell für die Baumart Fichte. Forstw. Cbl. 122, 175-182.
Korotaev, A., 1994: Untersuchungen der Wurzelsysteme von forstlich wichtigen Baumarten im Nordwesten Rußlands. Dissertation, Fakultät für Forstwissenschaften und Waldökologie, Universität Göttingen.
Korsun, F., 1935: Zivotnrmalniho porostu ve vzorcich. Lesnicka prace (in tschechisch). Korpel, S., 1992: Ergebnisse der Urwaldforschung für die Waldwirtschaft im Buchenökosystem.
Allgemeine Forstzeitschrift 47 (21), 1148-1152.

Bibliografía 269
Košir Z., 1966: Beitrag zur Erforschung der Urwaldstruktur reiner Buchenwälder. In: Gesellschaftsmorphologie, Ber. Int. Symp. Int. Ver. Für Vegetationskunde, Rinteln. 1966, 306-314.
Köstler, J.N., 1956: Waldbauliche Beobachtungen an Wurzelstöcken sturmgeworfener Waldbäume. Forstw. Cbl. 75, 65-91.
Köstler, J.N., Brückner, E. y Bibelriether H., 1968: Die Wurzeln der Waldbäume. Verlag Paul Parey, Hamburg.
Kotar, M., 1993: Bestimmung des Verteilungsmusters von Bäumen im Wald. Manuscript. Kouba, J., 1989: The theory of an estimate of the development of calamities and of management of the
process of forest adjustment to normal forest. Lesnictvi 35 (10): 925-944. Kozak, A., 1988. A variable exponent taper equation. Can. J. For. Res. 18, 1363-1368. Kraft, G., 1884: Beiträge zur Lehre von den Durchforstungen, Schlagstellungen und Lichtungshieben.
Klindworth, Hannover. Krajicek, J.E., Brinkman, K.A. y Gingrich S.F., 1961: Crown competition. A measure of density. For. Sci.
7, 35-42. Kramer, H., 1988: Waldwachstumslehre. Verlag Paul Parey. Hamburg. Kramer, H., 1990a: Nutzungsplanung in der Forsteinrichtung. 2. Auflage. J. D. Sauerländer´s Verlag.
Frankfurt a.M. Kramer, H., 1990b: Zur Nutzungsplanung bei geänderter Waldstruktur. Forst und Holz, 45 (21): 621-623. Kramer, H. y Kätsch, C.H., 1994: Individuelles Wachstum von Waldbäumen in Abhängigkeit von
natürlichen und anthropogenen Einflüssen. Schr. a. d. Forstl. Fak. d. Univ. Göttingen und der Nds. Forstl. Vers. Anst. 117. Sauerländer, Frankfurt.
Kramer, H. y AKÇA A., 1995: Leitfaden zur Waldmesslehre. J. D. Sauerländer´s Verlag, Frankfurt. Kramer, K., 1996. Phenology and growth of European trees in relation to climate change. PhD Diss.
Landbouw-Universität Wageningen. Kramer, P. J. y Kozlowski, T. T., 1979: Physiology of woody plants. Academic press, New York. Kranigk, J. y Gravenhorst, G., 1993: Ein dreidimensionales Modell für Fichtenkronen. Allgemeine Forst-
und Jagdzeitung, 164: 146-149. Kublin, E. y Scharnagl, G., 1988: Verfahrens- und Programmbeschreibung zum BWI-Unterprogramm
BDAT. Abschlußbericht, Forstliche Versuchs- und Forschungsanstalt Baden-Württemberg, 7800 Freiburg.
Kuhr, M., 1999: Grobwurzelarchitektur in Abhängigkeit von Baumart, Alter, Standort und sozialer Stellung. Dissertation, Fak. f. Forstw. u. Waldökologie, Universität Göttingen.
Kurth, H., Gerold, D. y Ulbricht, R., 1994: Forsteinrichtung. Deutscher Landschaftsverlag Berlin GMBH: 233-322.
Kurth, W., 1994a: Morphological models of plant growth: Possibilities and ecological relevance. En: Ecological Modelling, International Journal on Ecological Modelling and Systems Ecology. Volumes 75/76:299-308. Verlag Elsevier.
Kurth, W., 1994b: Growth Grammer Interpreter GROGRA 2.4: A software tool for the 3-dimensional interpretation of stochastic, sensitive growth grammars in the context of plant modelling. Berichte des Forschungszentrums Waldökosysteme, Reihe B, Bd. 38.
Kurth, W., 1998: Some new formalisms for modelling the interactions between plant architecture, competition and carbon allocation. Bayreuther Forum Ökologie Bd. 52, 53-98.
Kurth, W., 1999: Die Simulation der Baumarchitektur mit Wachstumsgrammatiken. Habilitation, Univ. Göttingen.
Kutschera, L. y Lichtenegger, E., 2002: Wurzelatlas mitteleuropäischer Waldbäume und Sträucher. Leopold Stocker Verlag, Graz – Stuttgart.
Kvist Johannsen, V., 1999: A growth model for oak in Denmark. Ph. D. Dissertation, Danish Forest and Landscape Research Institute, Hørsholm, Denmark .
Láhde, E., Laiho, O., Norokorpi, Y. y Saska, T., 2002: Zuwachs von Fichte und Birke in gleich und ungleich strukturierten Beständen. Forst und Holz, 15/16, 483.
Lamprecht, H. 1986. Waldbau in den Tropen. Verlag Paul Parey, Hamburg. Larson, P.R., 1963: Stem form development of forest trees. For. Sci. Monograph 5. Lássig, R., 1991: Zum Wachstum von Fichtensolitären [Picea abies (L.) Karst.] in Südwestdeutschland.
Dissertation, Univ. Freiburg.

Bibliografía 270
Last, F.T., Mason, P.A., Wilson, J. y Deacon, J.W., 1983: Fine roots and sheathing mycorrhizas: their formation, function and dynamics. Plant and Soil 71, 9-21.
Lee, D.H., 1998: Architektur der Wurzelsysteme von Fichten (Picea abies L.) auf unterschiedlich versauerten Standorten. Diss., Fak. f. Forstw. u. Waldökologie, Universität Göttingen.
Lee, W.K. y Gadow, K.v., 1997: Iterative Bestimmung der Konkurrenzbäume in Pinus densiflora - Beständen. Allgemeine Fost- und Jagdzeitung, 186 (3/4): 41-44.
Lee, W.K., 1993: Wachstums- und Ertragsmodelle für Pinus densiflora in der Kangwon-Provinz, Korea. Diss., Forstwiss. Fachbereich, Univ. Göttingen. Cuvillier Verlag.
Lee, W.K., Seo, J.H., Son, Y.M., Lee, K.H. y Gadow, K.v., 2003: Modelling stem profiles for Pinus densiflora in Korea. For. Ecol. Manage. 172, 69-77.
Leemans, K., 1992: The biological component of the simulation model for boreal forest dynamics. In: A systems analysis of the global boreal forest: 428-445.
Leibundgut, H., 1978: Die Waldpflege. Paul Haupt, Bern und Stuttgart. Lemay, V., Kozak, A., Muhairwe, K. y Kozak, R.A., 1993: Factors affecting the performance of Kozak's
(1988) variable-exponent taper function. En: Wood, G.B. y Wiant, H.V. (Eds.) Modern Methods of Estimating Tree and Log Volume. IUFRO Conference, Morgantown, West Virginia, 34-53.
Lembcke, G., Knapp, E. y Dittmar, O., 1977: DDR-Kiefern-Ertragstafel 1975. Institut für Forstwissen-schaft, Eberswalde.
Lemm, R., 1991: Ein dynamisches Forstbetriebs-Simulationsmodell. Dissertation, ETH Zürich. Leslie, P. H., 1945: On the use of matrices in certain population mathematics. Biometrika 33: 183-212. Leuschner, C., 2002: Lebensraum Baumkrone - Schatzkiste der Biodiversität. Georgia Augusta,
Wissenschaftsmagazin der Georg-August-Universität Göttingen; No. 1, 18-25. Levins, R., 1966: The strategy of model building in population biology. American Scientist 54, 421-431. Lewandowski, A. y Gadow, K. v., 1997: Ein heuristischer Ansatz zur Reproduktion von Waldbeständen.
Allgemeine Forst- und Jagdzeitung , 168.Jg., 9: 170-174. Lewandowski, A. y Pommerening, A., 1997: Zur Beschreibung der Waldstruktur - Erwartete und
beobachtete Artendurchmischung. Forstw. Cbl. 116, 129-139. Lindenmayer, A., 1975: Developmental algorithms for multicellular organisms: A survey of L-Systems. J.
Theor. Biol. 54, 3-22. Liu, C., Zhang, L., Davis, C.J., Solomon, D.S. y Gove, J.H., 2002: A Finite Mixture Model for
Characterizing the Diameter Distributions of Mixed-Species Forest Stands. For. Sci. 48 (4), 653-661. Liu, Q. y Hytteborn, H., 1991: Gap structure, disturbance and regeneration in a primeval Picea abies forest.
J. Veg. Sci. 2: 391-402. López Sánchez, C.A., Gorgoso, J.J., Castedo, F., Rojo, A., Rodríguez, R., Álvarez González, J.G. y
Sánchez F., 2003: A height-diameter model for Pinus radiata D.Don in Galicia (Northwest Spain). Ann. For. Sci. 60, 237-245.
Lorimer, C.G., 1983: Test of age-independent competition indices for individual trees in natural hardwood stands. For. Ecol. Manage. 6, 343-360.
Lundegårdh, H., 1957: Klima und Boden in ihrer Wirkung auf das Pflanzenleben. Fischer Verlag, Jena. Lüpke, B. v., 1992: Waldbau ohne Kahlschlag – Möglichkeiten und Risiken. Forstarchiv 63 (1): 10-15. Luxmore, R.J., Oren, R., Sheriff, D.W. y Thomas, R.B., 1995: Source - sink - storage relationships of
conifers. En: Smith, W.K. y Hinckley, T.M. (eds): Resource physiology of conifers. Academic Press, 179-216.
Madrigal, A., Puertas, F. y Martínez Millán,, F. J., 1992: Tablas de producción para Fagus sylvatica en Navarra. Gobierno de Navarra. Dpto. de Agric. Ganadería y Montes. Serie Agraria n° 3, Pamplona.
Maguire, D.A., Moeur, M. y Bennett, W.S., 1994: Models for describing basal diameter and vertical distribution of primary branches in young Douglas-Fir. For. Ecol. Manage. 63, 23-55.
Maguire, D. A. y Hann, D. W., 1989: The relationship between cross crown dimensions and sapwood area at crown base in Douglas-fir. Can. J. For. Res. 19(5), 557-565.
Maier, C.A. y Teskey, R.O., 1992: Internal and external control of net photosynthesis and stomatal con-ductance of mature eastern white pine (Pinus strobus). Can. J. For. Res. 22, 1387-1394.
Maltamo, M., Puumalainen, J. y Päivinen, R., 1995: Comparison of beta and Weibull functions for modelling the basal area diameter distribution in stands of Pinus silvestris and Picea abies. Scand. J. For. Res. 10: 284-295.

Bibliografía 271
Marques, P., 1991: Evaluating site quality of even-aged stands in northern Portugal using direct and indirect methods. For. Ecol. Manage. 41: 193-204.
Martin, G. L. y Ek, A. R., 1984: A Comparison of Competition Measures and Growth Models for Predicting Plantation Red Pine Diameter and Height Growth. For. Sci. 30 (3): 731-743.
Matèrn, B., 1986: Spatial Variation. 2nd edition. Lecture Notes in Statistics. Springer-Verlag, Berlin. Matsuda, M. y Baumgartner, A., 1975: Ökosystemare Simulation des Nutzeffektes der Sonnenenergie für
Wälder. Forst. Cbl. 94, 89-104. Matsue, K., Iwagami, S., Yamamoto, T. y Shibayama, Z., 1999: Modelling crown form of Hinoki
(Chamaecyparis obtusa) based on branch measurement. J. For. Plann 5, 13-18. Matsumura, N., 1988: Entwicklung und Anwendungsmöglichkeiten eines einzelbaumorientierten
Simulationsmodells für Douglasie. Diss. Georg-August-Universität Göttingen. Matthies, D., Weixler, H. y Hess, U., 1995: Befahrungsbedingte Strukturveränderungen von Waldböden.
AFZ/Der Wald (22), 1218-1221. Matyssek, R., 2003: Kosten und Nutzen von Raumbesetzung und –ausbeute. AFZ/Der Wald 17: 862-863. Max, T.A. y Burkhart, H.E., 1976: Segmented polynomial regression applied to taper equations. For. Sci.,
22(33): 283-289. Mayer, H., 1984: Waldbau auf sozio-ökologischer Grundlage. 3. Auflage, Fischer-Verlag Stuttgart. Mayer, K.E., Laudenslayer, W.F., 1988: A Guide to Wildlife Habitats of California. California Department
of Forestry and Fire Protection. Mehldahl, R. S. y Beltz, R. C., 1990: Summary of modelling efforts for SET: Southeastern TWIGS, School
of Forestry. Auburn University Alabama. Mendlik, G., 1983: Beech Yield Table (en hungaro). Erdeszeti-Kutatasok, 75:157-162. Meng, X. Y., 1991: The significance of taper equations and stand diameter structures in developing
merchantable volume tables. Journal of Beijing Forestry University, 13(2): 14-20. Meusel, H., Jäger, E. y Weinert, E., 1965: Vergleichende Chorologie der zentraleuropäischen
Flora.Karten/Text. Fischer Verlag, Jena. Michailoff, I., 1943: Zahlenmäßiges Verfahren für die Ausführung der Bestandeshöhenkurven. Forstw.
Cbl. u. Thar. Jahrb. 6: 273-279. Mitscherlich, G., 1939a: Sortenertragstafel für Kiefer, Buche und Eiche. Mitt. Forstwirtsch. u. Forstwiss.
10, 1-86. Mitscherlich, G., 1939b: Sortenertragstafel für Fichte. Mitt. Forstwirtsch. u. Forstwiss. 10, 87-101. Mitscherlich, G., 1942: Der Einfluß des Seewindes auf das Wachstum von Kiefer, Fichte, Buche, Birke,
Erle. Thar. Forstl. Jahrb. 93: 346-373. Mitscherlich, G., 1961: Untersuchungen in Plenterwäldern des Schwarzwaldes. Allgemeine Forst- und
Jagdzeitung, 132: 61-73 + 85-95. Mitscherlich, G., 1970: Wald, Wachstum und Umwelt. 1. Band: Form und Wachstum von Baum und
Bestand. J.D. Sauerländer´s Verlag, Frankfurt. Mitscherlich, G., 1971: Wald, Wachstum und Umwelt. 2. Bd: Waldklima und Wasserhaushalt. J.D.
Sauerländer's Verlag, Frankfurt. Mitscherlich, G., 1975: Wald, Wachstum und Umwelt. 3. Bd: Boden, Luft und Produktion. J.D.
Sauerländer's Verlag, Frankfurt. Mitscherlich, G., 1978: Wald, Wachstum und Umwelt. 1. Band: Form und Wachstum von Baum und
Bestand. 2., überarbeitete Auflage. J.D. Sauerländer´s Verlag, Frankfurt. Mitscherlich, G., Kern, K.G. y Künstle, E., 1963: Untersuchungen über den Kohlensäuregehalt in
Plentergehalt und Fichtenreinbestand. Allgemeine Forst- und Jagdzeitung 134: 281-290. Moeur, M., 1981: Crown width and foliage weight of northern Rocky mountain conifers. USDA For.
Serv. Res. Pap. INT.-283. Moeur, M., 1993: Crown width and foliage weight of northern Rocky mountain conifers. USDA For. Serv.
Res. Pap. INT.-283. Mohr, H. y Schöpfer, P., 1992: Lehrbuch der Pflanzenphysiologie. Springer-Verlag, Berlin. Mohren, G. M. J., Van Gerwen, C. P. y Spitters, C. J. T., 1984: Simulation of primary production in even-
aged stands of Douglas-fir. For. Ecology Manage. 9: 27-49. Möhring, B., 1986: Dynamische Betriebsklassensimulation. Berichte d. Forschungszentrums
Waldökosysteme, Univ. Göttingen, Bd 20.

Bibliografía 272
Møller, C. M., 1933: Bonitetsvise Tilvaekstovesigter for BØg, Eg og RØdgran in Danmark. Sammendrag fra Dansk Skovf. Tidsskr. 18.
Monserud, R.A. y Sterba, H., 1996: A basal area increment model for individual trees growing in even-aged forest stands in Austria. For. Ecol. Manage. 80: 57-80.
Monsi, M. y Saeki, T., 1953: The light factor in plant association and its importance in dry matter production. Japan. J. Bot. 14, 22-52.
Mooney, H.A., 1986: Photosynthesis. En: Crawley, M.J. (ed.). Plant Ecology. Blachwell Scientific Publications, Oxford, 345-373.
Moosmayer, H.U. y Schöpfer, W., 1972: Beziehungen zwischen Standortsfaktoren und Wuchsleistung der Fichte. Allgemeine Forst- und Jagdzeitung, 143(10): 203-215.
Moosmayer, U., 1957: Zur ertragskundlichen Auswertung der Standortsgliederung im Ostteil der schwäbi-schen Alb. Mitt. d. Vereins f. forstl. Standortskunde, Forstpflanzenzüchtung 7: 1-41.
Moser J.W., 1974: A system of equations for the components of forest growth. En: Fries, J. (Ed.): Growth models for tree and stand simulation; Royal College of Forestry. Research Notes Nr. 30: 56-76.
Moser, M., 1964: Die Mykorrhizafrage bei der Anzucht von Forstpflanzen für das Hochgebirge. En Schmidt-Vogt, H., 1964: Forstsamengewinnung und Forstpflanzenzucht für das Hochgebirge, 225-231.
Moser, W. G., 1793: Einige Actenstücke über die Forst-Einrichtung. In: Moser’s Forst-Archiv, Ulm: 147-243.
Muhairwe, C. K., 1999: Taper equations for Eucalyptus pilularis and Eucalyptus grandis for the north coast in New South Wales, Australia. Forest Ecology and Management 113: 251-269.
Murach, D., 1984: Die Reaktion der Feinwurzeln von Fichten auf zunehmende Bodenversauerung. Diss., Forstwissen. Fachbereich, Universität Göttingen.
Murach, D., 1991: Feinwurzelumsätze auf bodensauren Fichtenstandorten. Forstarchiv 62, 12-17. Murray, D. M. y Gadow, K. v., 1991: Relationships between the diameter distributions before and after
thinning. For. Sci. 37 (2): 552-559. Murray, D.M. y Gadow, K.v., 1993: A flexible yield model for regional timber forecasting. Southern
Journal of Applied Forestry 17 (2), 112-115. Nagashima, I. y Kawata, N., 1994: A stem taper model including butt swell. J. Jpn. For. Soc. 76(4), 291-
297. Nagel, J. y Biging, G. S., 1995: Schätzung der Parameter der Weibull-Funktion zur Generierung von
Durchmesserverteilungen. Allgemeine Forst- und Jagdzeitung 166 (9/10): 185-189. Nagel, J., 1994: Ein Einzelbaumwachstumsmodell für Roteichenbestände. Forst und Holz. 49 (3), 69-75. Nagel, J., 1998: Möglichkeiten der Einschätzung von Artendiversität und Struktur im Rahmen von
Betriebsinventuren. Vortrag am 14. 7. 1998, Fak. f. Forstw. u. Waldökologie, Univ. Göttingen. Nagel, J., 1999: Konzeptionelle Überlegungen zum schrittweisen Aufbau eines waldwachstumskundlichen
Simulationssystems für Nordwestdeutschland. Schriften aus d. Forstl. Fak. d. Univ. Göttingen u. d. Nieders. Forstl. Vers. Anst., Bd 128.
Nagel, J., Albert, M. y Schmidt, M., 2002: Das waldbauliche Prognose- und Entscheidungsmodell BWINPro 6.1. Forst und Holz. 57 (15/16):486-493.
Nagel, P., 1976: Die Darstellung der Diversität von Biozönosen. Schriftenreihe Vegetationskunde 10: 381-391. Bonn-Bad Godesberg.
Namikawa, K. y Kawai, Y., 1998: Stand structure and establishment process of an old-growth stand in the mixed deciduous broadleaf/conifer forest of Mt. Moiwa forest reserve, Central Hokkaido, Northern Japan. J. For. Res. Japan 3: 205-211.
Nepal, S. K. y Somers, G. L., 1992: A generalized approach to stand table projection. For. Sci. 38: 120 – 133.
Newberry, J. D. y Burkhart, H. E., 1986: Variable-form stem profile models for loblolly pine. Can. J. For. Res. 16: 109-114.
Newnham, R.M., 1992: Variable-form taper functions for four Alberta tree species. Can. J. For. Res. 22, 210-223.
Niedersachsen, 1991: Langfristige ökologische Waldentwicklung in den Landesforsten – Programm der Landesregierung Niedersachsen. Niedersächsisches Ministerium f. Ernährung, Landw. u. Forsten, 49 S.

Bibliografía 273
Nielsen, C.C.N., 1990: Einflüsse von Pflanzenabstand und Stammzahlhaltung auf Wurzelform, Wurzel-biomasse, Verankerung sowie auf die Biomasseverteilung im Hinblick auf die Sturmfestigkeit der Fichte. Schriften aus der Forstl. Fakultät der Universität Göttingen und der Niedersächsischen Forstl. Versuchsanstalt, Band 100. J. D. Sauerländer´s Verlag, Frankfurt.
Nielsen, C.C.N., 1995. Report of project “Development of roots and root/shoot-rations”. Air3-CT93-1269. Unveröffentlicht.
Niggemeyer, P., 1999. Schätzung von Durchmesserverteilungen mit Hilfe von nicht-parametrischen Methoden. Diplomarbeit, Fakultät für Forstwissenschaften und Waldökologie d. Univ. Göttingen.
Nordrhein-Westfalen, 1991: Waldwirtschaft 2000 - Gesamtkonzept für eine ökologische Waldbewirtschaftung des Staatswaldes in Nordrhein-Westfalen. Hrsg.: Ministerium für Umwelt, Raumordnung und Landwirtschaft, Landesforstverwaltung.
Oheimb, G.v., Westpahl, C., Tempel, H. y Härdtle, W., 2005. Structural pattern of a near-natural beech forest (Fagus sylvatica) (Serrahn, North-east Germany). For. Ecol. Manage. 212, 253-263.
Oohata, S.I., 1986: Some quantitative analysis of tree form. Further extensions to the pipe model theory. En: Fujimori, T., Whitehead (eds). Crown and canopy structure in relation to productivity. Published by the Forestry and Forest Products Institute, Ibaraki, Japan.
Ortloff, W. y Schlaepfer, R., 1997: Stickstoff und Waldschäden - eine Literaturübersicht. Allgemeine Forst- und Jagdzeitung, 167 (9-10): 184-201.
Otto, H.J., 1989: Langfristige ökologische Waldbauplanung für die Niedersächsischen Landesforsten. Aus dem Walde, Heft 42.
Otto, H.J., 1991: Langfristige ökologische Waldbauplanung für die Niedersächsischen Landesforsten. Aus dem Walde, Heft 43.
Otto, H.J., 1994: Waldökologie. Verlag Eugen Ulmer, Stuttgart. Otto, H.J., 1995: Zielorientierter Waldbau und Schutz sukzessionaler Prozesse – eine Diskussion. Forst u.
Holz 50: 203-209. Otto, H.J., 2002: Antriebskräfte natürlicher Buchenwalddynamik in Europa. Forst und Holz 57 (21): 649-
653. Ottorini, J.M., 1991. Growth and development of individual Douglas-fir stands for applications to simula-
tion in silviculture. Annales des sciences Forestieres 48, 651-666. Pardé, J. y Bouchon, J., 1990: Dendrométrie. Ecole Nationale du Genie Rural des Eaux et des Forêts,
Nancy. Pardé, J., 1961: Dendrométrie. Editions de l’École nationale des Eaux et des Forêts, Nancy. 350 S. Paterson, S.S., 1962: Introduction to Phytochorology of Norden. Medd. Stat. Skogsforskininginstitut 50:
1-145. Paulsen, J.C., 1795: Praktische Anweisung zum Forstwesen. Detmold. Pearson, S. M., Turner, M. G., Gardner, R. H. y O’Neill, R. V., 1996: An organism-based perspective of
habitat fragmentation. En: Szaro y Johnston, 1996: 41-57. Penning de Vries, F.W.T. y Laar H.H.v, 1982 : Simulation of plant growth and crop production. Pudoc
Centre for Agriculture Publishing and Documentation, Wageningen. Penttinen, A., Stoyan, D. y Henttonen, H. M., 1994: Marked Point Processes in Forest Statistics. For. Sci.
38 (4): 806-824. Perpeet, M., 1999: Weniger wäre mehr – waldbauliche Illusion oder Chance? Forst u. Holz 54 (3): 71-74. Petterson, N., 1954: Die Massenproduktion des Nadelwaldes. Medd. Stat. Skogsforskninginst. 45. Philip, M.S., 1994: Measuring Trees and Forests. Wallingford, Cab International. Pielou, E.C., 1977: Mathematical ecology. John Wiley, New York. Pienaar, L.V. y Harrison, W.M., 1988: A Stand Table Projection Approach to Yield Prediction in
Unthinned Even-Aged Stands. For. Sci., 34(3): 804-808. Pienaar, L.V., Harrison, W.M. y Rheney, J.W., 1990: PMRC Yield Prediction System for Slash Pine
Plantations in the Atlantic Coast Flatwoods. Plantation Management Research Cooperative. School of Forest Resources. University of Georgia.
Polomski, J. y Kuhn, N., 2001: Wurzelhabitus und Standfestigkeit der Waldbäume. Forstw. Cbl. 120, 303-317.
Pommerening, A., 1997: Eine Analyse neuer Ansätze zur Bestandesinventur in strukturreichen Wäldern. Diss., Fak. f. Forstw. u. Waldökologie, Göttingen. Cuvillier Verlag Göttingen.
Pommerening, A., 2002: Approaches to quantifying forest structures. Forestry 75(3), 305-324.

Bibliografía 274
Pressler, M. R., 1865: Das Gesetz der Stammbildung und dessen forstwirtschaftliche Bedeutung für den Waldbau höchsten Reinertrags. Arnold Verlag, Leipzig.
Pretzsch, H. y Bossel, H., 1988: Rahmenbedingungen für die Konstruktion forstlicher Wachstumsmodelle und Simulationsprogramme. AFZ (22), 615- 617.
Pretzsch, H., 1991a: Konzeption und Konstruktion von Wuchsmodellen für Rein- und Mischbestände. Habilitationsschrift. Universität München.
Pretzsch, H., 1991b: Konzeption einer modellorientierten Mischbestandsforschung. Vortrag anläßlich der Jahrestagung der Sektion Ertragskunde des Dt. Verb. Forstl. Forschungsanstalten in Treis/Mosel am 13. Mai 1991.
Pretzsch, H., 1992: Konzeption und Konstruktion von Wuchsmodellen für Rein- und Mischbestände. Schriftenreihe der Forstwissenschaftlichen Fakultät, Universität München, No. 115.
Pretzsch, H., 1993: Analyse und Reproduktion räumlicher Bestandesstrukturen - Versuche mit dem Strukturgenerator STRUGEN. Schriften aus der Forstl. Fak. d. Univ. Göttingen u. d. Nieders. Forstl. Vers. Anst., Band 114, J.D. Sauerländer's Verlag, Frankfurt a.M.
Pretzsch, H., 1994: Waldwachstumsforschung in strukturreichen Rein- und Mischbeständen - Unter-suchungsansatz, Arbeitsschwerpunkte, Kooperationsbedarf. Tagungsbericht der Sektion Ertragskunde des Dt. Verb. Forstl. Forschungsanstalten: 6-26. Habilitationsschrift.
Pretzsch, H., 1995: Perspektiven einer modellorientierten Waldwachstumsforschung. FwCbl 114: 188-209. Pretzsch, H. y Kahn, M., 1998: Forschungsvorhaben „ Konzeption und Konstruktion von Wuchs- und
Prognosemodellen für Mischbestände in Bayern“: Abschlußbericht Projekt W28 Teil 2. Konzeption und Konstruktion des Wuchsmodells SILVA 2.2 – Methodische Grundlagen. Lehrstuhl für Waldwachstumskunde der Ludwig-Maximilians-Universität München, Freising.
Pretzsch, H., 2001: Modellierung des Waldwachstums. Parey Buchverlag Berlin. Prodan, M., 1965: Holzmeßlehre. J.D. Sauerländer's Verlag, Frankfurt. Prusinkiewicz, P. y Lindenmayer, A., 1990: The Algorithmic Beauty of Plants. Verlag Springer, New York. Puhe, J., 1994: Die Wurzelentwicklung der Fichte (Picea abies [L.] Karst.) bei unterschiedlichen chemischen
Bodenbedingungen. Berichte des Forschungszentrums Waldökosysteme, Reihe A, Band 108, Göttingen.
Pukkala T. y Kolström T., 1988: Simulation of the Development of Norway Spruce Stands using a Tran-sition Matrix. For. Ecol. Manage. 25, 255-267.
Puumalainen, J., 1996: Die Beta-Funktion und ihre analytische Parameterbestimmung für die Darstellung von Durchmesserverteilungen. Arbeitspapier 15-96, Inst. f. Forsteinrichtung, Univ. Göttingen.
Rademacher, P., 2002: Ermittlung der Ernährungssituation, der Biomasseproduktion und der Nährelementakkumulation mit Hilfe von Inventurverfahren sowie Quantifizierung der Entzugsgrößen auf Umtriebsebene in forstlich genutzten Beständen. Habilitationsschrift, Univ. Göttingen.
Ramírez-Maldonado, H., Bailey, R.L. y Borders, B.B., 1988: Some implications of the algebraic difference form approach for developing growth models. En: Ek, A.R., Shifley, S.R. y Burke, T.E. (Eds): Forest growth modelling and prediction. USDA For. Serv. Gen. Techn. Rep. NC-120, 731-738.
Rapp, C., 1991: Untersuchungen zum Einfluss von Kalkung und Ammoniumsulfat-Düngung auf Feinwurzeln und Ektomykorrhizen eines Buchenaltbestandes im Solling. Ber. Forschungszentrum Waldökosysteme Göttingen A 72.
Raunkiaer, C., 1907: Planterigets Livsformer og deres Betydning for Geografien. Kjöbenhavn, Kristiana. (Engl. Übersetzung von Gilbert-Carter, H., 1937: Plant life forms. Clarendon press, Oxford.)
Rautiainen, O., 1999: Growth Dynamics and Management of Shorea robusta Forests in Southern Nepal. Dissertation, Univ. Joensuu.
Raven, P.H., Evert, R.F. y Eichhorn S.E., 1987: Biology of plants. Worth Publishers Inc., New York. Reed, D.D. y Green, E.J., 1984. Compatible stem taper and volume ratio equations. For. Sci. 30 (4), 977-
990. Reinecke, L. H., 1933: Perfecting a Stand Density Index for Even-Aged Forests. J. Agric. Res. 46: 627-638. Remmert, H., 1991: Das Mosaik-Zyklus-Konzept und seine Bedeutung für den Naturschutz – eine
Übersicht. Tagungsbericht „Das Mosaik-Zyklus-Konzept der Ökosysteme und seine Bedeutung für den Naturschutz“, 6.-9. Sept. 1989; Bayer. Akademie f. Naturschutz u. Landschaftspflege: 5-15.
Rennie, P. J., 1963: Methods of assessing site capability. Commonw. For. Rev. 42: 306-317.

Bibliografía 275
Rennols, K y Smith, W. R., 1993: Zone of Influence Models for Inter Tree Forest Competition. En: Rennols, K. (ed) 1993: Stochastic Spatial Models in Forestry. Proc of a IUFRO S4.11 Conf. held in Thessaloniki, Greece; published by The University of Greenwich: 27-36.
Richardson, C.W., 1981: Stochastic simulation of daily precipitation, temperature and solar radiation. Water Resources Research 17: 182-190.
Riemer, T., Gadow, K.v. y Sloboda, B., 1995: Ein Modell zur Beschreibung von Baumschäften. Allgemeine Forst- und Jagdzeitung. 166(7), 144-147.
Rièra, B., 1982: Observations sur les chablis, Piste de St. Elie, Guyane. Bulletin de liaison de groupe de travail sur l’écosystème forestier guyanais, ORSTOM, Cayenne, 6: 165-183.
Rodríguez Soalleiro, R.J., Álvarez González, J.G. y Vega Alonso, G., 1995: Pineiro do Pais - Modelo dinamico de crecemento de masas regulares de Pinus pinaster Aiton en Galicia. Xunta de Galicia.
Rodríguez Soalleiro, R., Álvarez González, J.G. y Schröder, J., 2000: Simulation and comparison of silvicultural alternatives for even-aged Pinus pinaster Ait. stands in Galicia (Northwestern Spain). Ann. For. Sci. 57: 747-754.
Rodríguez Soalleiro, R., Sánchez Rodríguez, F., Gorgoso, J., Castedo, F., López Sánchez, C.A. y Gadow, K.v., 2002: Evaluating standard treatment options for Pinus radiata plantations in Galicia (north-western Spain). Forestry 75(3): 273-284.
Rodríguez Soalleiro, R., Balboa Murias, M.A., Álvarez González, J.G. y Merino González, A., 2007: Evaluation through a simulation model of nutrient exports in fast-growing southern European pine stands in relation to thinning intensity and harvesting operations. Ann. For. Sci. 64(4), 375-384.
Röhe, P., 1985: Untersuchungen über das Wachstum der Buche in Baden-Württemberg. Schriftenreihe der Landesforstverwaltung Baden-Württemberg 61.
Rojo, A. y Montero, G., 1996: El Pino Silvestre en la Sierra de Guadarrama. Ministerio de Agriculture, Pesca y Alimentación, Madrid.
Rojo, A., Perales, X., Sánchez Rodríguez, F., Álvarez González J.G. y Gadow K.v., 2005: Stem taper functions for maritime pine (Pinus pinaster Ait.) in Galicia (Northwestern Spain). Eur. J. Forest Res. 124(3), 177-186.
Römisch, K., 1983: Ein mathematisches Modell zur Simulation von Wachstum und Durchforstung gleichaltriger Reinbestände. Dissertation, Universität Tharandt.
Römisch, K., 1995: Durchmesserwachstum und ebene Bestandesstruktur am Beispiel der Kiefernversuchsfläche Markersbach. DVFFA-Sektion Forstliche Biometrie und Informatik, 8. Tagung Grillenburg, S. 84-103.
Roschak, C., 1999: Schnellwuchsuntersuchung Lensahn - Eine Untersuchung zur Bedeutung schnellwachsender Baumarten im naturnahen Waldbau. Projektbericht für Gesellschaft zur Förderung schnellwachsender Baumarten in Norddeutschland e.V.: 37 S.
Rothe, A., 1997: Einfluss des Baumartenanteils auf Durchwurzelung, Wasserhaushalt, Stoffhaushalt und Zuwachsleitung eines Fichten-Buchen-Mischbestandes am Standort Höglwald. Forstl. Forschungsber. München 163.
Rubner, K., 1960: Die pflanzengeographischen Grundlagen des Waldbaus. Neumann, Radebeul. Rudra, A. B., 1968: A stochastic Model for the Prediction of Diameter Distributions of Even-aged Forest
Stands. OPSEARCH. Journal of the operational Society of India. 5 (2): 59-73. Rysavy, T. y Roloff, A., 1994: Ursachen der Vereschung in Mischbeständen und Vorschläge zu ihrer
Vermeidung. Forst und Holz 49, 392-395. Saborowski, J., 1982: Entwicklung biometrischer Modelle zur Sortimentenprognose. Dissertation,
Universität, Göttingen. Saeki, T., 1963: Light relations in plant communities. En: Evans, L.T. (ed.): Environmental control of
plant growth. Academic Press, New York. Sánchez Orois, S., Gurjanov, M., Schröder, J., 2001. Analyse des Grundflächenzuwachses gleichaltriger
Fichtenreinbestände. Allgemeine Forst- und Jagdzeitung, 172 (3), 51-59. Sánchez Orois, S., Kotzé, H. y Corral Rivas, J. J., 2003: Ein kompatibles Modell für die Fortschreibung
von Durchmesserverteilungen. Vortrag anlässlich der Jahrestagung der Sektion Ertragskunde des Deutschen Verbandes Forstlicher Forschungsanstalten in Torgau am 2. – 4. Juni 2003.
Sánchez-Orois, S. y Vilčko, F., 2002: Generierung, Bewertung und Optimierung von Managementoptionen für die Vornutzung von Fichtenbeständen. Vortragsmanuskript Ertragskundetagung im Mai 2002.

Bibliografía 276
Sauter, U.H. y Fahrback, M., 1993: Ästigkeitsverhältnisse und Schnittholzqualität in einem weitständig begründeten Fichtenbestand. AFZ 48 (17), 875 - 878.
Scheffer, F. y Schachtschabel, P., 1976: Lehrbuch der Bodenkunde. Fischer Verlag, Stuttgart. Schmid I. y Kazda M., 2001: Vertical distribution and radial growth of coarse roots in oure and mixed
stands of Fagus sylvatica and Picea abies. Can. J. For. Res. 31, 539-548. Schmidt, M. G. y Carmean, W. H., 1988: Jack pine site quality in relation to soil and topography in North
Central Ontario. Can. J. For. Res. 18: 297-305. Schmidt, M., 2001: Prognosemodelle für ausgewählte Holzqualitätsmerkmale wichtiger Baumarten. Diss.,
Fak. f. Forstwiss. u. Waldökologie, Univ. Göttingen. Schmidt, M., Schütz, J.P. y Gadow K.v., 1997: Strukturanalyse in vier Plenterüberführungsbeständen.
Schweiz. Zeitschr. f. Forstw. 148(5), 335-352. Schmidt, M. y Gadow, K.v., 1999: Baumhöhenschätzung mit Hilfe der bivariaten Johnson’s SBB-
Funktion. Forstw. Cbl. 118, 355-367. Schmidt, W., 1991: Die Bodenvegetation im Wald und das Mosaik-Zyklus-Konzept. Tagungsbericht „Das
Mosaik-Zyklus-Konzept der Ökosysteme und seine Bedeutung für den Naturschutz“, 6.-9. Sept. 1989; Bayer. Akademie f. Naturschutz u. Landschaftspflege: 16-29.
Schmidt-Vogt, H., 1986: Die Fichte. Paul Parey, Berlin. Schober, R., 1952: Ausbauchungsreihen. In: Grundner, F., Schwappach, A., 1952: Massentafeln zur
Bestimmung des Holzinhaltes stehender Waldbäume und Waldbestände. 10. völlig neu bearbeitete Auflage; Hrsg. R. Schober. Paul Parey, Berlin.
Schober, R., 1953: Ertragstafeln und Forsteinrichtung. Forstwissenschaftliches Centralblatt 72: 1-13. Schober, R., 1972: Die Rotbuche 1971. J. D. Sauerländer's Verlag, Frankfurt. Schober, R., 1987: Ertragstafeln wichtiger Baumarten. J. D. Sauerländer´s Verlag. Frankfurt. Schober, R., 1991: Eclaircies par le Haut et Arbres D´avenir. Rev. For. Fr. XLIII: 385-402. Schober, R., 1994: Darstellung von Durchforstungsgraden. Unveröffentl. Vorlesungsmanuskript. Institut
für Forsteinrichtung und Ertragskunde, Universität Göttingen. Schober, R., 1995: Ertragstafeln wichtiger Baumarten. J. D. Sauerländer´s Verlag, Frankfurt. Schönau, A. P. G. y Aldworth, W. J. K., 1991: Site evaluation in black wattle with special reference to soil
factors. S. Afr. For. J. 156: 35-43. Schönau, A. P. G., 1988: A re-appraisal of a site evaluation study in Black Wattle with special reference to
soil factors. En: Gadow, K.v. (ed.) Research Contributions to Plantation Forestry. Faculty of Forestry, Stellenbosch: 83-102.
Schönhar, S., 1993: Die Waldbodenvegetation als Standortsweiser. Allgemeine Forst- und Jagdzeitung 164: 173-180.
Schröder, J. y Gadow, K. v., 1999: Testing a new competition index for maritime pine in North-Western Spain. Can. J. For. Res. 29: 280-283.
Schröder, J., 1998: Beschreibung von Bestandesstrukturen im Knysna-Wald, Südafrika. Diplomarbeit, Fak. f. Forstwissen. u. Waldökologie, Univ. Göttingen.
Schröder, J., 2001: Analyse der Wuchsdynamik von Pinus pinaster Ait. in Nordwest-Spanien. Cuvillier Verlag, Göttingen.
Schröder, J., Oliver-Villanueva, J. V., Rojas-Briales, E. y Gadow, K. v., 1995: Zum Höhenwachstum von Pseudotsuga menziesii (MIRB.) FRA. in Katalonien. FA 66: 214-217.
Schröder, J., Rodríguez, R., y Vega Alonso, G., 2002: An age-independent basal area increment model for Maritime Pine trees in North-Western Spain. For. Ecol. Manage., 157: 55-64.
Schübeler, D., 1997: Untersuchungen zur standortabhängigen Wachstumsmodellierung bei der Fichte. Diss. Fak. f. Forstwiss. u. Waldökologie d. Univ. Göttingen.
Schübeler, D., Nagel, J., Pommerening, A. y Gadow, K.v., 1995: Modellierung des standortbezogenen Wachstums der Fichte. Unveröffentlichtes Manuskript, Institut f. Forsteinrichtung u. Ertragskunde, Univ. Göttingen.
Schubert, R., 1991: Lehrbuch der Ökologie. Gustav Fischer Verlag, Jena. Schulze, E.D., 1970: Der CO2 - Gaswechsel der Buche (Fagus sylvatica L.) in Abhängigkeit von den
Klimafaktoren im Freiland. Flora 159, 177-232. Schumacher, F.X., 1939: A new growth curve and its application to timber yield studies. Journal of
Forestry 37, 819-820. Schumann, K., 2002: Eingriffsanalyse im Forstamt Bovenden. Unveröff. Manuskript.

Bibliografía 277
Schütz, J.-Ph., 1989: Der Plenterbetrieb. Unterlage zur Vorlesung Waldbau III, ETH Zürich. Schwappach, A., 1890: Wachstum und Ertrag normaler Fichtenbestände. Julius Springer Verlag, Berlin. Schwappach, A., 1905: Untersuchungen über die Zuwachsleistungen von Eichen-Hochwaldbeständen in
Preussen. Neumann Verlag., Neudamm. 131 S. Schwappach, A., 1911: Die Rotbuche. Neumann Verlag, Neudamm. Seidling, W., 1997: 50 Jahre Vegetationsentwicklung auf einer Schlagfläche im osthessischen Bergland. Fw.
Cbl. 116, 218-231. Shafer, G. N., 1988: A Site Growth Model for Pinus eliottii in the Southern Cape. S. Afr. For. J. 146: 12-17. Shafer, G. N., 1989: Site Indicator Species for Predicting Productivity of Pine Plantations in the Southern
Cape. S. Afr. For. J. 148: 7-17. Shannon, C.E., Weaver, W., 1949: The mathematical theory of communication. University of Illinois
Press, Urbana. Sharpe, P.J.H., 1990: Forest modelling approaches - compromises between generality and precision. En:
Dixon, Mehldal, Ruark, Warren (Eds.): Process Modelling of Forest Growth Responses to Environmental Stress; Timber Press, Portland, Oregon: 21-32.
Shugart, H.H., 1984: A theory of forest dynamics: the ecological implication of forest successsion models. Springer, New York.
Shumway, R. H., 1988: Applied statistical time series analysis. Prentice-Hall, Englewood Cliffs, NJ. Shvidenko, A., Venevsky, S., Raille, G. y Nilsson, S., 1995. A system for evaluation of growth and mor-
tality in Russian forests. Water, Air and Soil Pollution 82, 333-348. Sievänen, R., 1993: A process-based model for the dimensional growth of even-aged stands. Scand. J. For.
Res. 8: 28-48. Skovsgaard, J.P. y Mosing, M., 1996: Bogefoyngelser i Ostjylland. Danish Forest and Landscape Research
Institute. Sloboda, B., 1971: Zur Darstellung von Wachstumsprozessen mit Hilfe von Differntialgleichungen erster
Ordnung. Mitt. Bad.-Württemb. Forstl. Vers. u. Forsch. Anstalt. Heft 32. Sloboda, B., 1976: Mathematische und stochastische Modelle zur Beschreibung der Statik und Dynamik
von Bäumen und Beständen – insbesondere das bestandesspezifische Wachstum als stochastischer Prozeß. Habil.-Schrift, Univ. Freiburg. 310 S.
Sloboda, B., 1984: Bestandesindividuelles biometrisches Schaftformmodell zur Darstellung und zum Ver-gleich von Formigkeit und Sortimentausbeute sowie Inventur. Tagungsbericht d. Sektion Ertragskunde, Neustadt.
Smalley, G. W. y Bailey, R. L., 1974: Yield tables and stand structure for shortleaf pine plantations in Tennessee, Alabama and Georgia highlands. USDA For. Service Res. Paper SO-97.
Smaltschinski, T., 1981: Bestandesdichte und Verteilungsstruktur. Dissertation, Universität Freiburg. Sonntag, M., 1998: Klimaveränderungen und Waldwachstm – Treedyn Simulationen mit einer Analyse
modellstruktureller Unsicherheiten. Verlag Mainz, Wissenschaftsverlag Aachen. Soares, P. y Tomé, M., 2002: Height-diameter equation for first rotation eucalypt plantations in Portugal.
For. Ecol. Manage. 166, 99-109. Souter, R.A., 1986: Dynamic stand structure in thinned stands of naturally regenerated loblolly pine in the
Georgia Piedmont. Ph. D. Thesis. University of Georgia. Athens, GA. Späth, H., 1973: Spline-Algorithmen zur Konstruktion glatter Kurven und Flächen. Oldenbourg,
München. Speidel, G., 1972: Planung im Forstbetrieb. Grundlagen und Methoden der Forsteinrichtung. Verlag
Parey, Hamburg und Berlin. 267 S. Spellmann, H., 1998: Überführung als betriebliche Aufgabe. Forst und Holz, 54 (4): 110-116. Spiecker, H., 1995: Ein Vergleich natürlicher und pflegebedingter Selektionsprozesse. Mitt. d. Forstl. Vers.
Anstalt Rhld-Pfalz, Nr. 34 161-179. Spiecker, M., 1994: Wachstum und Erziehung wertvoller Waldkirschen. Mitt. d. Forstl. Vers. u.
Forschungsanstalt Baden-Württemberg, Heft 181. Sprugel, D.G., 1991: Disturbance, Equilibrium, and Environmental Variability - what is Natural
Vegetation in a Changing Environment. Biological Conserv. 58: 1-18. Spurr, S. H., 1962: A measure point density. For. Sci. 8: 85-96.

Bibliografía 278
Staupendahl, K., 1997: Ein neues Stichprobenverfahren zur Erfassung und Beschreibung von Naturverjüngung. Vortrag anlässlich der Jahrestagung der Sektion Forstliche Biometrie und Informatik des Deutschen Verbandes Forstlicher Versuchsanstalten am 24. 9. 97 in Freiburg.
Staupendahl, K., y Puumalainen, J., 1999: Modellierung des Einflusses von Durchforstungen auf die Durchmesserverteilung von gleichaltrigen Fichtenreinbeständen. Cbl. ges. Forstw. 116 (4): 249-262.
Staupendahl, K., 2001: Das flächenbezogene Winkelmaß Wf. En: Akça, A, Hoffmann, B., Schumann, K. y Staupendahl K. (Eds.): Waldinventur, Waldwachstum und Forstplanung – Moderne Technologien, Methoden und Verfahrensweisen. Festschrift K. v. Gadow. Zohab Verlag, Göttingen: 101-116.
Steingaß, F., 1995: Beschreibung der Schaftprofile von Douglasien. Diplomarbeit, Fachbereich Forstwissenschaft, Universität Göttingen.
Sterba, H., 1975: Assmann´s Theorie der Grundflächenhaltung und die ”Competition-Density-Rule” der Japaner Kira, Ando und Tadaki. Cbl. ges. Forstw. 92 (1): 46-62.
Sterba, H., 1981: Natürlicher Bestockungsgrad und Reinecke's SDI. Cbl. ges. Forstw. 98 (2): 101-116. Sterba, H., 1983: Single stem models from inventory data with temporary plots. Mitt. D. Forstl.
Bundesversuchsanstalt Wien Nr. 147: 87-101. Sterba, H., 1990: Validitätsprüfung des Einzelbaumsimulators für Fichten-Kiefern-Mischbestände.
Tagungsbericht. Dt. Verb. Forstl. Forschungsanstalten - Sektion Ertragskunde: 101-117. Sterba, H., 1991: Forstliche Ertragslehre. Lecture Notes. Sterba, H., 1997: Waldwachstumsforschung – zwischen Erkenntnisdrang und Praxisanspruch. Allgemeine
Forst- und Jagdzeitung 168 (11-12), 227-230. Sterba, H., 1998: Das Randproblem bei der Erfassung von Strukturparametern. Vortrag am 25. 6. 1998,
Fak. f. Forstw. u. Waldökologie, Univ. Göttingen. Sterba, H. y Monserud, R.A., 1997: Applicability of the forest stand growth simulator Prognaus for the
Austrian part of the Bohemian Massif. Ecol. Mod. 98: 23-34. Stöhr, F. K., 1963: Erweiterungsmöglichkeiten der Winkelzählprobe. Diss., Albert-Ludwig-Univ.
Freiburg/Br. Stoyan, D. y Penttinen, A., 2000: Recent applications of point process methods in forestry statistics.
Statistical Science 15, 61-78. Struck, G., 1999: Effekte von Pflanzverband, Kulturdüngung und Schlagräumung am Beispiel junger
Traubeneichenbestände. Diss., Fak. f. Forstwiss. u. Waldökologie, Univ. Göttingen. Sturm, K., 1994: Naturnahe Waldnutzung in Mitteleuropa. Studie im Auftrag von Greenpeace
Deutschland. Duvensee. Suzuki, T., 1971: Forest Transition as a Stochastic Process. Mitt. FBVA Wien. Heft 91: 137-150. Suzuki, T., Ohsaki, K., Sato, H. y Yamamoto, Y., 1992: A representation method for todo-fir Shapes
Using Computer Graphics. J. of the Jap. For. Soc. 74 (6), 504-508. Swindel, B. F., Smith, H. D. y Grosenbaugh, L. R., 1987: Fitting diameter distributions with a hand-held
programmable calculator. Scand. J. For. Res. 2: 325-334. Szaro, R. C. y Johnston, D. W., 1996: Biodiversity in Managed Landscapes. Oxford University Press. Teepe, R., Brumme, R. y Beese, F., 2000: Der Einfluß der Bodenverdichtung bei der Holzernte auf den
Austausch der Spurengase CO2, CH4 und N2O. Abschlußbericht zum Forschungsvorhaben der Deutschen Bundesstiftung Umwelt.
Temesgen, H. y Gadow, K.v., 2003: Generalized Height-diameter Models for Major Tree Species in Complex Stands of Interior British Columbia. Europ. J. For. Res. 123(1), 45-51.
Thomasius, H., 1963: Untersuchungen über die Brauchbarkeit einiger Wachstumsgrößen von Bäumen und Beständen für die qualitative Standortsbeurteilung. Archiv für Forstwesen 12 (12): 1267-1323.
Thomasius, H., 1988: Stabilität natürlicher und künstlicher Waldökosysteme sowie deren Beeinflußbarkeit durch forstliche Maßnahmen. AFZ No. 43: 1037-1043, 1064-1068.
Thomasius, H., 1996: Geschichte, Anliegen und Wege des Waldumbaus in Sachsen. Schriftenreihe d. Sächs. Landesanst. f. Forsten 6/96: 11-52.
Thomasius, H.O. y Thomasius, H., 1976: Anwendungsbeispiel zu einem Verfahren der Berechnung ertragskundlich optimaler Bestandesdichtewerte und Diskussion dieses Verfahrens. Informationen TU Dresden, Sektion Forstwirtschaft, Bereich Waldbau und Forstschutz, Tharandt, 21-76.
Thomasius, H.O. y Thomasius, H., 1978. Ableitung eines Verfahrens zur Berechnung der ertragskundlich optimalen Bestandesdichte. Beiträge f. d. Forstwirtschaft 12 (79).

Bibliografía 279
Thomasius, H., 1978: Modell des gleichaltrigen Reinbestandes. TU Dresden, Sektion Forstwirtschaft, Tharandt, II Vorträge: 122-148.
Thomasius, H., 2001: 50 Jahre Arbeitsgemeinschaft Naturgemässe Waldwirtschaft - Dauerwaldbewirtschaftung. Der Dauerwald, 24: 15-39.
Tomé, M. y Burkhart, H. E., 1989: Distance-Dependent Competition Measures for Predicting Growth of Individual Trees. For. Sci. 35(3): 816-831.
Tomppo, E., 1986: Models and Methods for Analysing Spatial Patterns of Trees. Communicationes Instituti Forestalis Fenniae No. 138, Helsinki.
Torres Rojo, J. M. y Brodie, J. D., 1990: Adjacency constraints in harvest scheduling. Can. J. of For. Res. 20: 978-986.
Trincado, G. y Gadow, K.v., 1996: Zur Sortimentschätzung stehender Laubbäume. Clbl. ges. Forstw. 1, 27-38.
Trincado, G., Gadow, K.v. y Tewari, V.P., 1996: Comparison of three stem Profile Equations for Quercus robur L. South African Forestry Journal 177, 23-29.
Trincado, G., Quezada, P. R. y Gadow, K.v., 2003: A Comparison of two stand table projection methods for young Eucalyptus nitens (Maiden) plantations in Chile. For. Ecol. Manage. 180: 443-451.
Trincado, G., Kiviste, A. y Gadow, K.v., 2003: Preliminary site index models for native Roble (Nothofagus obliqua) and Raulí (N. alpina) in Chile. New Zealand J. of Forestry Science 32 (3): 322-333.
Tsubo, M., Walkers, S., 2005: Relationships between photosynthetically active radiation and clearness index at Bloemfontein, South Africa. Theor. Appl. Climatol. 80, 17–25.
Turner, J., Thompson, C. H., Turvey, N. D., Hopmans, P. y Ryan, P. J., 1990: A soil technical classifi-cation for Pinus radiata (D. Don) plantations. I. Development. Aust. J. Soil Res. 28: 797-811.
Turton, S.M., 1985: The relative distribution of photosynthetically active radiation within four tree cano-pies, Craigieburn Range, New Zealand. Austr. For. Res. 15, 363-394.
Turvey, N. D., Booth, T. H. y Ryan, P. J., 1990: A soil technical classification for Pinus radiata (D. Don) plantations. II. A basis for estimation of crop yield. Aust. J. Soil Res. 28: 813-824.
Ulrich, B., 1986: Stoffhaushalt von Waldökosystemen. Bioelement-Haushalt. Institut für Bodenkunde und Waldernährung der Universität Göttingen.
Ulrich, B., Pirouzpanah, D., Murach, D., 1984: Beziehungen zwischen Bodenversauerung und Wurzelentwicklung von Fichten mit unterschiedlich starken Schadsymptomen. Forstarchiv 55, 127-134.
Ung, C.-H., Raulier, F., Oullet, D. y Dhôte, J.-F., 1997: L’indice de compétition interindividuelle de Schütz. Can. J. For. Res. 27: 521-526.
Ung, C.H., 1993: Growth and branchiness of Picea mariana (Mill.) BSP. En: Canadian Forest Service Modelling Working Group. Proc. of the seventh annual meeting and workshop.
Upton, G. y Fingleton, B., 1990: Spatial Data Analysis by Example. John Wiley, New York. Van der Meer, P. J., 1995: Canopy dynamics of a tropical rain forest in French Guiana. Diss.,
Landbouwuniversiteit Wageningen. Van Laar, A. y Akça, A., 1997: Forest Mensuration. Cuvillier Verlag Göttingen, 418 pp. ISBN 3-89588-874-5.
Vanclay, J. K., 1989: Site productivity assessment in rainforests: an objective approach using indicator species. In: Wan Razali Mohd, H.T. Chan, S. Appanah (Editors), Proceedings of the Seminar on Growth and Yield in Tropical Mixed / Moist Forests, 20-24 June 1988, Kuala Lumpur. Forest Research Institute, Malaysia: 225-241.
Vanclay, J. K., 1992: Assessing site productivity in tropical moist forests: a review. Forest Ecology and Management 54: 257-287.
Vanclay, J. K., 1994: Modelling forest growth - applications to mixed tropical forests. CAB International, Wallingford, UK.
Vanclay, J., 1998: Towards more rigorous assessment of biodiversity. En: Bachmann, P., Köhl, M. y Päivinen, R., (eds), 1998: Assessment of biodiversity for improved forest planning. Kluwer Academic Publishers: 211-232.
Van Laar, A. y Akça, A., 1997: Forest mensuration. Cuvillier Verlag. Göttingen. Verein Deutscher Forstlicher Versuchsanstalten, 1902: Beratungen der vom Vereine deutscher Forstlicher
Versuchsanstalten eingesetzten Kommission zur Feststellung des neuen Arbeitsplanes für Durchforstungs- und Lichtungsversuche. Allg. Forst- und Jagdzeitung, 78, 180-184.

Bibliografía 280
Walther, H., 1960: Einführung in die Phytologie - III. Grundlagen der Pflanzenverbreitung - 1. Teil: Standortslehre. Stuttgart.
Wang, Y.P. y Jarvis, P.G., 1990: Description and validation of an array model - MAESTRO. Agricultural and Forest Meteorology 51: 257-280.
Waring, R.H. y Running, S.W. 1998: Forest ecosystems. Analysis at multiples scales. 2nd Edition. Academic Press. London.
Webb, L. J., 1959: A physiognomic classification of Australian rainforests. J. Ecol. 47: 551-570. Wegener, G. 1995: Perspektiven der Holznutzung. FwCbl. 114(2): 97-106. Weise, W., 1880. Ertragstafeln für die Kiefer. Julius Springer Verlag. Wenk, G., Antanaitis,S V. y Smelko, S., 1990. Waldertragslehre. Deutscher Landwirtschaftsverlag. Berlin. Wenk, G., Römisch, K. y Gerold, D., 1985: DDR-Fichtenertragstafel 1984. Technische Universität
Dresden, Sektion Forstwissenschaft, Tharandt. Wenk, G., 1996: Durchmesserverteilungen im Buchenplenterwald (als Bestandteil des
Forschungsprojektes “Sortenorientiertes Wachstumsmodell”). Tagungsband des Deutschen Verbandes Forstlicher Versuchs-anstalten. Sektion Ertragskunde, Mai 1996 in Neresheim.
Wensel, L., Meerschaert, W. y Biging, G., 1987: Tree Heigh and Diameter Growth Models for Northern California Conifers, Hilgardia, University of California, Volume 55, No. 8.
Wiant, H.V., Wood, G.B. y Gregoire, T.G., 1992: Practical guide for estimating the volume of a standing sample tree using either importance or centroid sampling. For. Ecol. Manage. 49, 333-339.
Wiedemann, E., 1931: Die Rotbuche. Mitteilung aus Forstwirtschaft und Forstwissenschaft. Wiedemann, E., 1936: Die Fichte. Schaper Verlag, Hannover. Wiedemann, E., 1942: Der gleichaltrige Buchen-Fichten-Mischbestand. Mitt. a. Forstwirtschaft und Forst-
wissensschaft 13: 1-88. Wiedemann, E., 1949: Ertragstafeln der wichtigsten Holzarten. Schaper Verlag, Hannover. Wimmenauer, E., 1914: Ertrags- und Sortimentuntersuchungen im Buchenhochwald. Mitteilungen aus
dem forstlichen Versuchswesen Badens. Windhager, M., 1999: Evaluierung von vier verschiedenen Waldwachstumssimulatoren. Diss., Universität
für Bodenkultur Wien. Yang, Y.Ch.U. y Feng, F.L., 1989: The application of parameter prediction models and parameter recovery
methods to the analysis of tree growth and stand structure in Taiwan. Q. Journ. Chin. For., 22(1):53-71.
Zeide, B., Gresham, C.A., 1991: Fractal dimensions of tree crowns in three loblolly pine plantations of coastal South Carolina. Can. J. For. Res. 21(8), 1208-1212.
Zeide, B. y Pfeifer, P., 1991: A method for estimation of fractal dimension of tree crowns. For. Sci. 37(5), 1253-1265.
Zenner, E.K. y Hibbs, D.E., 2000: A new method for modelling the heterogeneity of forest structure. For. Ecol. Manage. 129(1), 75-87.
Zoth, R. y Block, J., 1992: Untersuchungen an Wurzelballen sturmgeworfener Bäume in Reinland-Pfalz. Forst und Holz 47(18), 566-571.
Zucchini, W., Schmidt, M. y Gadow, K.v., 2001. A model for the diameter-height distribution in an uneven-aged beech forest and a method to assess the fit of such models. Silva Fennica. 35 (2), 168-183.


















