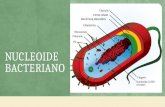
Estructura del dna bacteriano
28
Estructura del DNA bacteriano Por Oloarte R., Delgado B., Coyotl R., Estrada M. y Estrada S.
-
Upload
rebeca-alejandra-oloarte-pulido -
Category
Education
-
view
86 -
download
1
Transcript of Estructura del dna bacteriano

Estructura del DNA bacteriano
Por Oloarte R., Delgado B., Coyotl R., Estrada M. y Estrada S.

Introducción
El ADN es una molécula enormemente larga que no es recta ni homogénea en toda su longitud. Se puede curvar según las circunstancias y además se puede enrollar o desenrollar respecto al modelo canónico definido por Watson y Crick.
La variación en la helicidad se traduce en una tensión, tanto positiva como negativa, que se libera formando espirales de doble hélice en un sentido u otro

• El ADN circular cerrado es capaz de adoptar una estructura terciaria denominada superenrollamiento, que implica el enrollamiento del eje de la doble hélice sobre si mismo

Superenrollamiento Es el retorcimiento o giro sobre
si misma de una hélice, de forma que su eje no sigue una línea recta, sino otra héliceabrir glosario. Puesto que la hélice de partida ya lleva implícito un enrollamiento, a la nueva hélice descrita por el eje de la primera se la llama una "superhélice".
Un ejemplo muy intuitivo es el cordón del teléfono, que tiene un enrollamiento helicoidal y con el uso alcanza un superenrollamiento

Superenrollamiento
El superenrollamiento sólo permanece si los extremos se mantienen retenidos o sujetos, pues de no ser así el libre giro "deshace" las superhélices.
Tal sujeción puede ser externa o bien debida a la unión de ambos extremos entre sí, de modo que la cuerda (o la molécula de DNA) forme un círculo (no en el sentido geométrico, sino en el de línea cerrada).
En este caso la restricción de movimiento es permanente.

Superenrollamiento
En 1963 se descubrió que el DNA bicatenario del virus de polioma existe en forma de círculo cerrado.
En la actualidad, se admite que ésta es la forma típica del DNA en las bacterias.
Además, las grandes moléculas del DNA nuclear en los organismos superiores, aun siendo lineales y no circulares, forman grandes bucles retenidos mediante proteínas, de modo que cada bucle es bastante análogo al DNA circular cerrado.
Por todo ello, el DNA en las células de todo tipo experimenta constantemente fenómenos de superenrollamiento.

Topoisomerasas
Son enzimas capaces de actuar sobre la topología del ADN, ya sea enredándolo para permitir que se almacene de manera más compacta o desenredándolo para que controle la síntesis de proteínas y para facilitar la replicación del mismo.
Las topoisomerasas son capaces de cortar y pegar ya sea una o dos de las hebras de azúcar fosfato que forman el armazón o esqueleto del ADN.
Estas enzimas son necesarias debido a los inherentes problemas causados por la configuración estructural del ADN

Tipos de topoisomerasas
Las topoisomerasas se clasifican en las de tipo I, que rompen una sola hebra, y las de tipo II, que rompen las dos hebras al mismo tiempo.
Las topoisomerasas I actúan realizando un corte transitorio en una de las hebras de un duplex de ADN superenrollado que cambia el numero de enlace en incrementos de una unidad y que da como resultado la relajación del DNA superenrollado

Mecanismo de Acción
Para relajar el DNA las topoisomerasas 1(a) se unen al DNA y separan localmente las hebras complementarias; (b) a continuación, cortan una de las hebras y se unen a los extremos generados; © hacen pasar la hebra intacta atreves del corte y finalmente restablecen el enlace fosfodiester. El resultado es una estructura relajada (d).

Mecanismo de Acción de Topoisomerasas II
Las topoisomerasas II cambian el numero de enlace del DNA uniéndose a la molécula de DNA y pasando un segmento de DNA a través de un corte reversible formando en un segmento diferente de la misma molécula de DNA.
Las topoisomerasas II actúan fijándose a una molécula de DNA, de tal forma que se generan dos bucles súper enrrollados.

Mecanismo de accion de la Girasa
Este se ilustra utilizando como ejemplo la conversión de una molécula de DNA relajado en una molécula que contiene dos superenrollamientos, uno positivo y otro negativo (primer paso).
El paso de un segmento de DNA a través del superenrollamiento positivo mostrando en la parte derecha de la figura (tercer paso) cambia el numero de enlace, dando lugar a una molécula que contiene dos superenrollamientos negativos.

Topoisomerasas tipo III
Pueden relajar vueltas superhelicoidales sin necesidad de una fuente de energía, tal como la hidrolisis de ATP.
Estas topoisomerasas pueden estar especializadas en la resolución de los productos encadenados de DNA circular (eslabonados) que se generan justo antes de que se complete la replicación de DNA
Una clase poco usual de topoisomerasas, las girasas inversas, se han aislado de varias especies de arquebacterias. Estos enzimas tienen la notable propiedad de introducir vueltas superhelicoidales positivas en el DNA.
El súper enrollamiento positivo podría proteger el DNA de las condiciones desnaturalizantes de alta temperatura y acidez del hábitat de las bacterias

Molécula Modelo
Para entender mejor los conceptos y terminología relacionados con el superenrollamiento, emplearemos como ejemplo una molécula bicatenaria de DNA de 104 pb (pares de bases; es decir, la asociación de dos hebras con 104 nucleótidos cada una).
Si adopta la conformación más habitual, la doble hélice de tipo B, habrá exactamente 10 vueltas de hélice.
Decimos que la torsión de la molécula tiene un valor de 10 (T=10, o Tw=10, del inglés twist).

Ahora supongamos que flexionamos la molécula, aproximando sus extremos para formar un círculo. Puesto que el número de vueltas de hélice en la molécula es entero (10 en este ejemplo), los extremos de ambas hebras quedarán correctamente alineados con posibilidad de unirse por enlaces fosfodiéster (3' con 5' y 5' con 3'). Si se realiza esa unión, tendremos una molécula circular cerrada.Al ser esta molécula plana y sin tensión estructural, tiene una conformación relajada (tiene superenrollamiento cero) (Estrictamente, lo que es plano no es la molécula, sino la trayectoria del eje de la doble hélice.)Como consecuencia del cierre de la molécula, las hebras ya no se pueden separar físicamente (por ejemplo, tirando de ellas en sentidos contrarios), sino que están trabadas, ligadas, entrelazadas.Concretamente, en este ejemplo están entrelazadas 10 veces, y se dice que la molécula tiene un índice de ligazón de 10 (L=10, o Lk=10, del inglés linking number).

Restricción topológica
Un valor Lk = 0 significa que las dos hebras no están ligadas, es posible separarlas. Cuando Lk es distinto de cero las dos hebras están ligadas físicamente, es decir, no se pueden separar; esto es así a pesar de que no hay enlaces covalentes entre ellas; para referirnos a esta unión física sin enlace covalente se dice que hay un enlace topológico.
En una molécula con enlace topológico sólo se pueden separar las hebras mediante ruptura de un enlace covalente (fosfodiéster) o, dicho de otro modo, cortando una hebra. Lo mismo es aplicable para cualquier proceso que cambie el valor Lk (por ejemplo, de Lk=10 a Lk=9). Por la misma razón, la deformación, plegamiento, etc. de las hebras, sin cortarlas, no altera el índice de ligazon.

Generación de superenrollamiento
Si, en lugar de unir los extremos de la molécula lineal tal y como quedaron enfrentados, antes rotamos uno de ellos sobre sí mismo, de modo que se reduzca el enrollamiento de la doble hélice en una vuelta (la torsión es menor, Tw=9), y entonces unimos los extremos, tendremos una molécula con sus hebras menos entrelazadas, con menor índice de ligazón, Lk=9.
Al mismo tiempo, los 104 pb ahora se reparten en sólo 9 vueltas de hélice, por lo que en lugar de una vuelta cada 10,4 pb habrá una cada 11,56 pb (=104/9) y la conformación no será ya de tipo B.
Esto supone una inestabilidad para la molécula, una tensión estructural (similar a la que tiene el cable retorcido del teléfono si intentamos aplanarlo sobre la mesa sin que forme bucles).
La tensión se puede liberar si la molécula forma un bucle sobre sí misma, una vuelta superhelicoidal; así recupera la torsión más estable (Tw=10, con 10,4 pb/vuelta).
Al número de vueltas superhelicoidales se le denomina retorcimiento (W o Wr, del inglés writhe). En este caso, debe considerarse de signo negativo (Wr=–1), pues suple la disminución de torsión inicial. El giro de esta superhélice es dextrorso, hacia la derecha, como se puede comprobar con un modelo de cuerda


Topoisomerasas
En las células, existen enzimas que catalizan este proceso de superenrollamiento.
Se les denomina topoisomerasas, pues convierten unos topoisómeros en otros.
Hay topoisomerasas de tipo I (hidrolizan un enlace fosfodiéster en una hebra, hacen pasar la otra hebra a través del corte y vuelven a unir los extremos de la primera, el indice de ligazón varía en una unidad)
Topoisomerasas tipo II (hidrolizan sendos enlaces fosfodiéster en ambas hebras y hacen pasar otra doble hebra a través del corte, resellando éste de nuevo; de este modo, el índice de ligazón varía en dos unidades de una sola vez)

Funciones
Mediante el superenrollamiento, las moléculas de DNA se compactan y ocupan un volumen inferior en varios órdenes de magnitud.
El superenrollamiento permite que se acerquen regiones de la molécula alejadas en su estructura primaria, y esto es clave en procesos como la regulación de la replicación y la transcripción.
Del grado de compactación local conseguido depende la regulación de la accesibilidad a la información genética y, por consiguiente, la regulación de la expresión de los genes (transcripción).
Para que puedan tener lugar los procesos de replicación (copiado del DNA) y transcripción (copiado de la información del DNA en una molécula de RNA) es necesario que se separen las dos hebras complementarias, lo cual supone un desenrollamiento local de la doble hélice. Éste (mediado por proteínas específicas, principalmente polimerasas y helicasas) provoca sobreenrollamiento en las regiones adyacentes. Además, dificulta el desenrollamiento adicional, necesario para la progresión de los citados procesos a lo largo de la molécula.


Características del superenrollamiento
Positivo Negativo
Superhélice a derechas, dextrorsa.
Se encuentra de forma natural en las eubacterias.
En eucariotas es más frecuente en las zonas codificantes (facilita que se abra la doble hebra para la transcripción).
En eucariotas, transitoriamente por detrás de la RNApol durante la transcripción.
Une preferentemente intercalantes que desenrollan la doble hélice (p.ej., bromuro de etidio y cloroquina)
Superhélice a izquierdas, sinistrorsa.
Se puede encontrar en arqueobacterias.
En eucariotas, transitoriamente por delante de la RNApol durante la transcripción Une preferentemente ligandos que aprietan la doble hélice (p.ej., netropsina)

Torsión: Tw – TNumero o Índice de enroscamiento (Twist)
Retorcimiento: Wr – WNumero o Índice de retorcimiento (Writhe)Índice de ligazón Lk – LNumero o Índice de enlace (Linking)

Agentes que modifican el superenrollamiento
La geometría de una molécula superenrollada se puede ver afectada por cualquier agente que altere la torsión intrínseca de la doble hélice. Por ejemplo, un aumento de temperatura reduce la torsión y un aumento de fuerza iónica puede aumentarla.
Otro factor importante es la presencia de los denominados agentes intercalantes, moléculas planas e hidrófobas que se insertan entre los pares de bases apilados de la doble hélice, por lo común forzando ésta a una apertura, un desenrollamiento parcial, una reducción de la torsión

Bromuro de etidio
El sistema aromático de anillos condensados es responsable del intercalamiento. Su unión reduce localmente la torsión de la doble hélice en aprox. 26°, lo que habitualmente se compensa con un aumento del retorcimiento (superenrollamiento negativo).
Por otro lado, la fluorescencia propia del bromuro de etidio aumenta mucho al unirse al DNA. Por esta razón, se utiliza habitualmente para detectar la posición del DNA en los geles de electroforesis.

Cloroquina
Se utiliza a menudo en estudios de superenrollamiento de plásmidos.
Además, es uno de los fármacos clásicos para el tratamiento de la malaria, aunque su principal mecanismo de acción no depende de su capacidad intercalante.

Amsacrina
4'-(9-acridinilamino)-3'-metoximetanosulfonanilida
El intercalante es el anillo de acridina; el anillo de metanosulfonanilida se sitúa en el surco menor.
Es inhibidor de la topoisomerasa II.

Analisis Experimental
Las diferencias de superenrollamiento de un DNA circular suponen un volumen diferente de la molécula, pues el superenrollamiento la compacta. Debido a esto, se pueden detectar tales diferencias separando los topoisómeros mediante electroforesis en gel. Por ejemplo:
Análisis por electroforesis del superenrollamiento de un plásmido de 3000 pb, en gel de agarosa al 1,75%, revelado con bromuro de etidio bajo luz ultravioleta. A = plásmido intacto.B = plásmido abierto por corte con una endonucleasa.C...H = plásmido tratado con concentraciones de topoisomerasa y tiempos decrecientes.

Propiedades de las topoisomerasas