
EL PRESENTE TRABAJO SE DESARROLLÓ EN EL …148.206.53.84/tesiuami/UAMI12211.pdf · el presente...
36
Transcript of EL PRESENTE TRABAJO SE DESARROLLÓ EN EL …148.206.53.84/tesiuami/UAMI12211.pdf · el presente...


EL PRESENTE TRABAJO SE DESARROLLÓ EN ELDEPARTAMENTO DE BIOMEDICINA MOLECULAR DELCENTRO DE INVESTIGACIÓN Y DE ESTUDIOSAVANZADOS DEL I. P. N., BAJO LA DIRECCIÓN DE LA Men C. NATALY MANJARREZ ORDUÑO Y LA ASESORÍA DELM en C. HÉCTOR ROMERO RAMÍREZ.

AGRADECIMIENTOS I
En primer lugar quiero agradecer a Dios por otorgarme a los dos seres másimportantes en mi vida mi padre y mi madre.
A mi padre:
Sinceramente no tengo palabras para agradecerle, todo ese apoyo, consejos,comprensión, valores, fuerza y sobre todo el amor que me ha brindado y mebrinda día a día, simplemente Gracias por ser tal y como eres!
A mi madre:
Por ser mi pilar y motor de vida!
En forma especial a Nataly por permitirme aprender con ella, por supaciencia, apoyo y consejos.

AGRADECIMIENTOS II
Al CINVESTAV, y en particular al Dr. Leopoldo Santos Argumedo porbrindarme la oportunidad de ser parte de su equipo de trabajo.
Al M en C. Héctor Romero Ramírez y a la M en C. Gabriela López por sutiempo, recomendaciones y todo el apoyo brindado durante el desarrollo deeste proyecto.
Al M en B.E. Víctor Hugo Rosales García, por su disposición en el uso delos equipos de citometría de flujo.
A mis compañeros de laboratorio: Claudia, Juan Carlos, Isabel, Miguel y alSr. Manuel.
A mi amigo y compañero David ya que sin tu apoyo no hubiese podidoterminar esta historia.
A Karla, Tita, Moni, Toño por su tiempo y consejos.
A mis vecinos de laboratorio Oscar y Ricardo .
Al personal del bioterio.
Y a cada una de las personas que formaron parte de esta aventura.

ÍNDICE GENERAL
Página
INDICE DE FIGURAS Y TABLAS ii
LISTADE ABREVIATURAS iii
RESUMEN iv
INTRODUCCIÓN 1
OBJETIVOS 9
MATERIALES Y MÉTODOS 10
RESULTADOS 13
DISCUSIÓN 22
CONCLUSIONES 24
BIBLIOGRAFÍA 26

ÍNDICE DE FIGURAS Y TABLAS
Página
Figura 1. Eventos de linfopoyesis fetal y adulta. 7
Figura 2. Producción de anticuerpos monoclonales. 8
Figura3. Evaluación de la producción del anticuerpo 15monoclonal NIM-R3 crecido in vitro.
Figura 4. Evaluación de la producción del anticuerpomonoclonal NIM-R3 obtenido mediante la recolección 16del líquido de ascitis de ratones nu/nu.
Figura 5. Expresión del antígeno reconocido por el anticuerpo 17monoclonal NIM-R3 mediante el ensayo de Western Blot.
Figura6. Expresión de p30 sobre subpoblaciones 18de linfocitos en células de bazo y médula ósea.
Figura 7. Evaluación de la expresión de p30 sobre subpoblaciones 19de linfocitos en células de cavidad peritoneal y Placas de Peyer.
Figura 8. Evaluación de la expresión de p30 sobre subpoblaciones 20de linfocitos B en células de bazo.
Figura 9. Evaluación de la expresión de p30 sobre subpoblaciones 21de linfocitos B en células de médula ósea.
Figura 10. Evaluación de la expresión de p30 sobre subpoblaciones 22de linfocitos B en células de bazo.
Figura 11. Modelo de la expresión de p30 sobre subpoblaciones 26de linfocitos B en células de médula ósea y bazo.

LISTA DE ABREVIATURAS
AcM Anticuerpos Monoclonales
APC Aloficocianina
B FO Células B Foliculares
B INM Células B inmaduras
B ZM Células B de zona marginal
FITC Isotiocianato de fluoresceína
FIX – FACS Fijador para citometría de flujo
Ig Inmunoglobulina
LPS Lipopolisacárido
PBS Amortiguador salino de fosfatos
PE Ficoeritrina
SA Estreptovidina

RESUMEN
La identidad, características de expresión y función biológica de muchos antígenos es bienconocida, mientras que aún se desconoce para algunos otros. Una forma de estudiar antígenoses a través del uso de anticuerpos monoclonales y en este trabajo se reporta el patrón deexpresión en linfocitos B de una proteína de 30 kDa (p30), reconocida por el anticuerpomonoclonal NIM-R3.
La caracterización del anticuerpo monoclonal NIM-R3 en células linfoides mostró que p30solo se expresa en células B de diferentes compartimentos linfoides, sin embargo sólo seexpresa en una sola subpoblación. En el caso de células de médula ósea p30 se expresa enparticular en células B maduras e inmaduras y en menor grado células pro-B y pre-B. Por otraparte p30 se expresa en una subpoblaciòn que puede corresponder a las células B de ZonaMarginal y / o células B recién llegadas de la médula ósea.
Este antìgeno es inducido en células B después de la activación con LPS.
La expresión específica del antìgeno podría permitir su uso como blanco terapéutico; comoherramienta diagnóstico en linfomas.

INTRODUCCIÓN
Anticuerpos Monoclonales.
Los anticuerpos son proteínas denominadas también inmunoglobulinas (Ig), que tienen lafunción de unir moléculas denominadas antígenos a través de un fragmento específico de suestructura denominado sitio de unión del antígeno, a la zona del antígeno que se unedirectamente al anticuerpo se le denomina epítopo, en general, epítopes reconocidos por cadaanticuerpo, la afinidad de los anticuerpos, y su clase, son diferentes y pueden cambiar con elcurso del proceso de inmunización.
Los anticuerpos son producidos por los linfocitos B, cada linfocito B del organismo tiene ensu superficie múltiples moléculas de inmunoglobulina, y cada linfocito del organismo tiene lacapacidad de unir una molécula distinta. Cuando el linfocito B une a través de suinmunoglobulina de superficie a su antígeno específico, inicia un proceso de activación de lacélula que conduce a la producción de una gran cantidad de anticuerpo, lo que constituye laparte humoral de la respuesta inmunológica. La respuesta inmunológica tiene también unfuerte componente al azar, ya que en cada exposición con el antígeno, se emplea sólo unafracción, a veces mínima, de la gama de células B que poseen receptores específicos.
Todo lo anterior nos indica que en la práctica es imposible controlar la composición de laspreparaciones de anticuerpos policlonales que se obtienen de animales inmunizados. Estaspreparaciones son heterogéneas para los anticuerpos que contienen, en términos de isotipo,afinidad y epítope reconocido. La posibilidad de obtener preparaciones específicas, repetibles,e inagotables de anticuerpos con funciones predefinidas (AcM) se logró por primera vez en1974 y 1975 por Georges Kohler y Cesar Milstein en Cambridge, Reino Unido (Kohler, G. YC. Milstein.1975).
En sus experimentos Kohler y Milstein realizaron fusiones entre linfocitos provenientes deratones BALB/c inmunizados con antígenos definidos, y mielomas de la estirpe MOPC21,cuyas inmunoglobulinas aberrantes eran ya bien conocidas. Gracias a esto, dichosinvestigadores descubrieron que fusionar una célula B normal con una célula plasmáticamaligna inmortal puede dar origen a una línea celular híbrida que prolifera indefinidamente almismo tiempo que secreta la inmunoglobulina codificada por la célula B progenitora. Unaclona inmortal productora de anticuerpos híbridos obtenidos de esta manera se conoce comohibridoma y el anticuerpo que produce se denomina como anticuerpo monoclonal.
Un anticuerpo monoclonal es un anticuerpo producido a partir de una población de células;las cuales provienen todas de una célula individual. El primer paso en el desarrollo de losanticuerpos monoclonales es la inyección en un animal por lo general ratones, ratas o conejoscon un antígeno. Dentro del sistema inmunitario del ratón las células B específicas para elantígeno inyectado proliferan; las células B se obtienen de una muestra de sangre, del bazo oglándulas linfoides del ratón y se ponen en cultivo celular in vitro. Las células B solosobreviven un periodo corto fuera del cuerpo y para poder crecer las células B en el cultivocelular se unen a células de un mieloma que crecen vigorosamente cuando se les provee de

suficientes nutrientes, la mezcla que se obtiene contiene ahora células B, células del mielomay células unidas denominadas hibridomas, el cual produce un único tipo de anticuerpo, unanticuerpo monoclonal. El hibridoma que produce el anticuerpo monoclonal deseado seidentifica y esta nueva línea celular se utiliza para producir grandes cantidades del anticuerpo.
La producción masiva de anticuerpos monoclonales puede llevarse a cabo como se hamencionado anteriormente, tanto por el cultivo de los hibridomas y la recogida de lossobrenadantes que contienen los anticuerpos secretados, como mediante la inoculación delas células en la cavidad peritoneal de animales compatibles genéticamente con el linfocito yel mieloma parental (para hibridomas ratón-ratón y rata-rata). Este último procedimientogenera tumores ascíticos cuyos fluidos contienen entre 1-10 mg de anticuerpos por ml(aproximadamente 200-1000 veces más que la concentración alcanzada en el medio decultivo) (Cowley G. J. 1995)
Los AcM han revolucionado el estudio de la inmunología y de la biología celular, y hanencontrado múltiples aplicaciones en el diagnóstico. También se les estudia comoinstrumentos para formación de imágenes clínicas y en la terapéutica, en especial en cáncer.Por ejemplo, ciertos anticuerpos monoclonales marcados radiactivamente que reconocenantígenos de superficie tumoral pueden, si se inyectan en la corriente circulatoria, alojarse enel tumor y revelar su localización por emisión radioactiva. De manera similar, se ha mostradoque anticuerpos monoclonales acoplados a toxinas, como la ricina o la toxina diftérica (paraformar la llamada inmunotoxina) parecen prometedores como quimioterapéuticasantitumorales con capacidad de dirigir específicamente y de manera activa la toxina a lascélulas tumorales (Stites P.D; et al. 1998).
Los AcM se emplean para la definición de subpoblaciones celulares normales y tumorales, lacaracterización de antígenos tisulares y grupos sanguíneos, la detección de niveleshormonales y otros factores séricos circulantes, el diagnóstico y tratamiento de enfermedadesinfecciosas y parasitarias, la detección de tóxicos, mutágenos y drogas, la facilitación detransplante de órganos y tejidos, la determinación de la estructura de los antígenos, lafabricación de antivenenos y también ha permitido el desarrollo de potentes métodosinmunoquímicos de purificación de diversas moléculas.
Por desgracia, el uso de anticuerpos monoclonales de ratón en el ser humano con frecuenciase limita por su inmunogenicidad y puede ser difícil obtener las células B humanas necesariaspara producir hibridomas humanos útiles. Una solución posible consiste en aislar los genesde las cadenas pesada y ligera codificantes de un anticuerpo murino de interés, con el uso detécnicas de DNA recombinante para alterar sus secuencias de región C, con el propósito decodificar proteínas que semejen más de cerca de un anticuerpo humano y luego reintroducireste gen de anticuerpo humanizado a una célula plasmática para su expresión (Stites P.D; etal. 1998)
Existen numerosos procedimientos potencialmente utilizables para detectar la presencia deanticuerpos monoclonales secretados por los cultivos de hibridomas y para, posteriormente,caracterizar su reactividad. Se incluyen entre ellos la inmunofluorescencia indirecta, losradioinmunoensayos, las tinciones con inmunoperoxidasa, los inmunoensayos enzimáticos defase sólida y la citometría de flujo. Esta última es una técnica que analiza células ensuspensión que interfieren de forma individual con una fuente de luz. La intersección de cadacélula con la luz láser provoca la emisión de una serie de señales luminosas que permitediferenciar poblaciones celulares dentro de la muestra analizada, por su tamaño relativo, por

sus granulaciones o bien por su reactividad con fluorocromos previa incubación con diversosanticuerpos monoclonales. Es un método de lectura rápido, que permite analizar un elevadonúmero de células (habitualmente de 10.000 a 50.000 para cada anticuerpo monoclonal) yproporciona un registro computerizado de los resultados. La citometría de flujo permite ladeterminación de antígenos celulares de superficie, y por tanto, tiene utilidad en elinmunotipaje de leucemias agudas y síndromes linfoproliferativos crónicos. Así mismo,permite la cuantificación del ADN y la determinación de la actividad proliferativa de lapoblación celular. La cuantificación de ARN celular se aplica al recuento de reticulocitos. Lacitometría de flujo se ha beneficiado de los avances ocurridos en los últimos años eninformática, electrónica, óptica y tecnología láser. Así mismo, la producción de nuevosanticuerpos monoclonales con diferentes fluorocromos y los nuevos procedimientos detinción en citoquímica han permitido ampliar las áreas de estudio en diagnóstico clínico einvestigación biomédica. Actualmente, esta tecnología ha pasado de ser una herramienta útilen investigación básica a ser empleada en la practica habitual de muchos laboratorios dehematología y anatomía patológica.
Los anticuerpos monoclonales como marcadores.
Desde hace varios años mucho esfuerzo se ha dirigido hacia la caracterización de marcadores,los cuales como se ha mencionado anteriormente distinguen diferentes subclases delinfocitos. Si bien, tales estudios han establecido claramente subpoblaciones de células T;por consiguiente existe una investigación extensa sobre éste tipo de células, sin embargo elprogreso no ha sido tan rápido en el área de células B. A pesar de ello existe una variedad deestudios acerca de la heterogeneidad de células B.
En términos funcionales, existe evidencia sobre la sensibilidad de las células B haciaantígenos T-dependientes y T-independientes (Playfair, J. H.L y Purves, E.C. 1971) y enreactividad diferencial a mitógenos. Otro aspecto funcional variable es la susceptibilidaddiferencial de las células B maduras e inmaduras a la tolerogenésis. Las separacionescelulares basadas en el tamaño, adhesividad y carga, así como la habilidad de recircular esotro punto de la heterogeneidad de células B. Por otra parte, una variedad de marcadores desuperficie celular han sido mostrados para ser representados en células B, indicando así laheterogeneidad, igualmente en términos de subpoblaciones maduras o fases de desarrollo;esto incluye el isotipo Ig (Brandon, D. L, et al. 1979), receptores de segmentos Fc yreceptores del complemento, las moléculas Ia y las series Lyb de antígenos. Existen pruebaslas cuales mencionan que las células B de memoria difieren de las células B vírgenes. Apesar de tal evidencia de la heterogeneidad de células B, la limitación principal en esta áreade investigación ha sido una deficiencia general para asociar la heterogeneidad funcional conlos sistemas de marcadores bien definidos, usando reactivos fiables y reproducibles (Kohler,G y Milstein C. 1975). La técnica de anticuerpos monoclonales (AcM) proporcionan unacercamiento ideal a este problema.

Anticuerpo monoclonal NIM-R3.
En el año de 1982 se produjo el anticuerpo monoclonal, NIM-R3, usando el mieloma de ratade la línea celular 210RCY3-Ag 1.2.3 y células de bazo de ratas de Lou inmunizadas, conmembranas plasmáticas de células de bazo de ratón. Este anticuerpo identifica poblacionesdesconocidas de superficie de células Ig-positivas en bazo y una gran proporción de célulasde la médula (95%). También reconoce antígenos de diferenciación; la representación de lasuperficie de los marcadores cambia durante el desarrollo de la célula. La especificidad deNIM-R3 no aparece hasta las tres semanas de edad en el bazo y médula, y puede estar en unconjunto maduro de células. (Chayen A. y Parkhouse R. M. E. 1982).
Se han descrito varios anticuerpos monoclonales los cuales tienen propiedades similares aNIM-R3 hasta el punto que estos reaccionan con una población de células sIg+ en variostejidos y además reconocen una proporción de células de médula ósea que exceden el númerode células sIg+. Éstos incluyen a los anticuerpos RA3-2C2 (Yakura, H., Shen, F. et al.1981) yDNL 1.9 . Kincade et al. observaron la presencia de los primeros dos marcadores en célulaspre-B y encontró que ambos, se expresan en una fase posterior a la célula madrepluripotencial, pero antes de la aparición de sIg y antígenos Ia en las células en hígadoneonatal y médula ósea. Además, estos investigadores introdujeron varios anticuerposmonoclonales los cuales reconocen células pre-B en fase muy temprana, pero yacomprometidas en la fase de desarrollo. El anticuerpo monoclonal NIM-R3 no tienerelación con cualquiera de los que se habló anteriormente, según la comparación dedistribuciones del tejido.
Al parecer NIM-R3 parece reaccionar preferentemente con células B vírgenes y con célulasque se han relacionado con antígenos, cabe mencionar que las células que reaccionanextensivamente con anti-IgD se enriquecen en magnitud en subpoblaciones de células B enlas cuales hay células de memoria. En conclusión el anticuerpo monoclonal NIM -R3 defineun nuevo marcador para subpoblaciones de células B y células relacionadas. SegúnParkhouse y cols.
Por otra parte se Greenwood M. R et al., realizaron investigaciones acerca del anticuerpomonoclonal NIM-R3 de rata, y descubrieron que activa células B de ratón pero no activacélulas T. Por lo tanto se piensa que el AcM NIM-R3 podría reemplazar el factor decrecimiento de células-B (BCGF II) en cultivos de células B de murino, de linfoma BCL1;induciendo la proliferación pero no la diferenciación. Se ha sugerido que el AcM NIM-R3puede representar una nueva especificidad y puede dirigirse contra el receptor de la superficiecelular para BCGF II, y a su vez este receptor puede ser independiente de los receptores B15-TRF y BSF1 (Greenwood M. R. y Parkhouse R. M. E. 1986).

Origen de células B y Moléculas de Superficie.
El sistema inmunológico consiste de distintos tipos celulares con funciones específicas, deéstos, los linfocitos ocupan parte central, porque son las células que determinan laespecificidad de la inmunidad, y es su respuesta la que orquesta el brazo efector de larespuesta inmunológica.
Se distinguen dos clases de linfocitos, los linfocitos T, que tienen importantes funcionesregulatorias a través de la producción de citocinas, y efectoras a través de la lisis de células; ylos linfocitos B que son los precursores de las células productoras de anticuerpos.
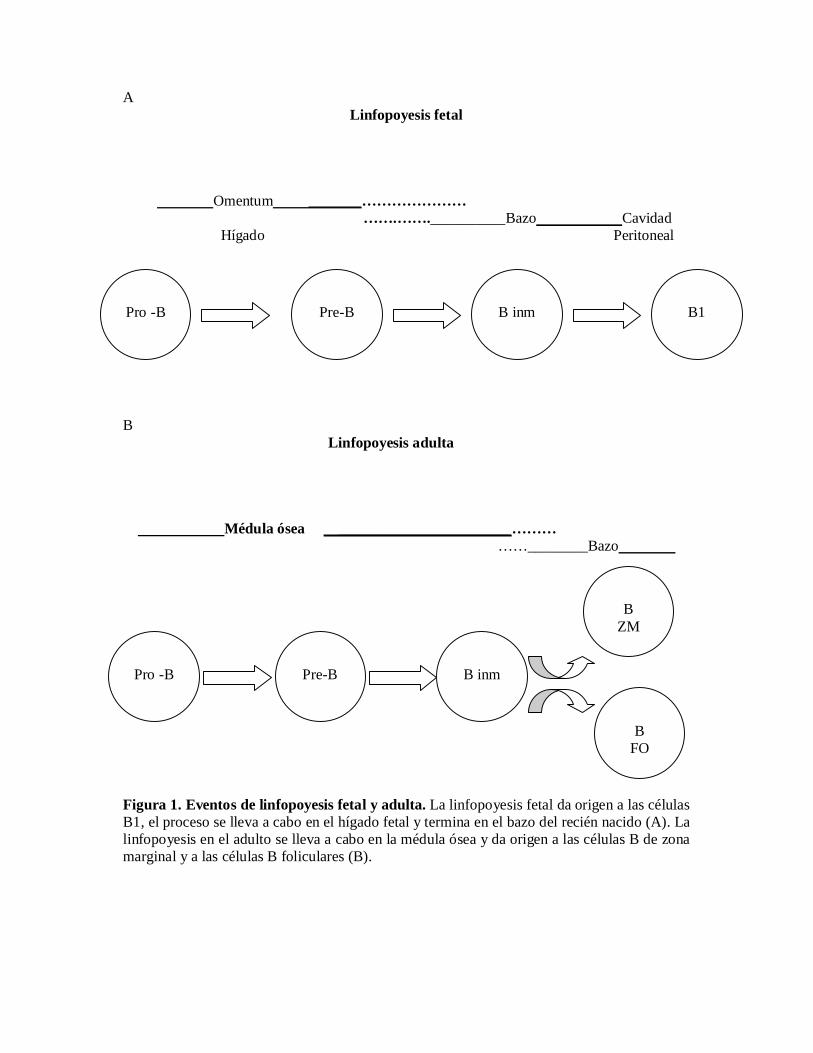
Los linfocitos B se diferencian a partir de células precursoras hematopoyéticas, mediante unproceso en el cual las células se van comprometiendo cada vez más hasta dar origen a unacélula B madura (Figura 1). Estos eventos de diferenciación se llevan a cabo en el hígadofetal, y durante la vida adulta en la médula ósea.
La diferenciación de los linfocitos B a partir de precursores hematopoyéticos se caracterizapor varios estadios intermedios. Durante el estadio de célula pro-B, el locus de cadenaspesadas de inmunoglobulina se vuelve accesible, y a través de un complejo enzimáticoconocido como recombinasa, los segmentos de la región de diversidad (D) se unen a uno delos segmentos de unión (J), y posteriormente a uno de los segmentos variables (V). Elrearreglo productivo de esos genes resulta en la expresión de una cadena de inmunoglobulinaen la superficie celular, lo que marca el inicio del estadio de célula pre-B. En esta etapa, serearreglan los genes V y J de los loci de cadenas ligeras, y su rearreglo y ensamble exitosocon la cadena pesada trae como consecuencia la expresión de una molécula deinmunoglobulina M en la superficie celular, para dar origen a una célula B inmadura, cuyaposterior expresión de inmunoglobulina D, la definirá como una célula B madura (Rolink A.G. et al. 2001). La molécula B220 (una isoforma de la fosfatasa CD45) caracteriza a loslinfocitos B de ratón desde que se encuentran en. el estadio pro-B, y debido a estacaracterística se utiliza como marcador de células B murinas, aunque se expresa también encélulas NK activadas.
Los eventos de linfopoyesis fetal y adulta dan origen a subpoblaciones de células B condistinta localización en el organismo y características funcionales diferentes, lo que confiereuna mayor diversidad a las respuestas mediadas por linfocitos B y las relaciona con otrosmecanismos de defensa del organismo(Hayakawa K. y Hardy R.R. 2000).
El proceso de linfopoyesis fetal, al parecer es el responsable del origen de las células B1(Hayakawa K. y Hardy R.R. 2000), que son la población predominante de células B en fetos yneonatos, mientras que en la vida adulta estas células constituyen solo una pequeña fracciónde las células B en bazo y en nódulos linfáticos, localizándose básicamente en las cavidadespleural y peritoneal, donde son la subpoblación de células B mayoritaria, y donde su númerose mantiene constante debido a su capacidad de autorregeneración.

Las células B1 se distinguen de las células B convencionales en que presentan un alto nivel deIgM y un bajo nivel de IgD; en forma semejante al resto de las células B, expresan elmarcador B220, pero en menor cantidad (Berland R. Y Wortis H. H. 2002).
El proceso de linfopoyesis adulta (en médula ósea), es al parecer, el responsable del origen dedos subpoblaciones de células B, las células B de la zona marginal (B ZM) y las células Bfoliculares (B FO).
Las células B foliculares, convencionales o B2, son las células B más extensivamenteestudiadas, se localizan en los folículos linfoides del bazo y nódulos linfáticos, además deque es ésta la población de linfocitos B que recircula predominantemente a través de la sangrey linfa.
Las células B de zona marginal, en cambio, se localizan en el bazo, entre el límite de la pulpablanca y el seno marginal, en estrecha vecindad con dos poblaciones de macrófagosrestringidos también a esta área: los macrófagos metalofílicos y los macrófagos de zonamarginal.
A diferencia de las células B foliculares, las células B de zona marginal no recirculan, sinoque al parecer, su función principal es la de funcionar como una primera línea de defensacontra antígenos sanguíneos particulados (Martín F. et al. 2001).Las células de zona marginalcapturan, procesan y presentan antígenos, además de enviar señales coestimulatorias a lascélulas T más rápida y eficientemente que las células B foliculares (Oliver A. M. et al. 1999).
Estas tres subpoblaciones de linfocitos B tienen pues, diferentes papeles dentro del sistemainmunológico, determinados por su especificidad hacia el antígeno, y la forma de responderante él, una característica que está relacionada con las moléculas que expresan en susuperficie.
La señalización que se obtiene a través de las moléculas de superficie es tan importante que lafalta de alguna de ellas, o de las moléculas intracelulares relacionadas con la transducción dela señal puede conducir a inmunodeficiencias o enfermedades autoinmunes (Lu P. et al.,1997), por esto el adecuado conocimiento de las moléculas de superficie y de los procesosinducidos por ellos tiene importancia diagnóstica (Khaw B., 1999) podría permitir lamanipulación del sistema inmunológico al modular los procesos regulados por ellos (TaylorP. C. et al., 2001); del mismo modo, la presencia de determinadas proteínas de membrana enpoblaciones celulares específicas podría permitir que éstas fueran sujetas de terapia dirigida através de anticuerpos o quimeras conjugados con agentes terapéuticos (DeNardo S. J. et al.,1999).
Uno de los enfoques en el estudio de las moléculas de superficie es a través del uso de AcM,que reconocen moléculas específicas y permiten su caracterización; con esta estrategia, en ellaboratorio se ha podido avanzar en el conocimiento de la biología del linfocito a través delestudio de diversas moléculas mediante los AcM.
ALinfopoyesis fetal
Omentum _______……………………….…….__________Bazo Cavidad
Hígado Peritoneal
BLinfopoyesis adulta
Médula ósea _________________________……… ……________Bazo
Figura 1. Eventos de linfopoyesis fetal y adulta. La linfopoyesis fetal da origen a las célulasB1, el proceso se lleva a cabo en el hígado fetal y termina en el bazo del recién nacido (A). Lalinfopoyesis en el adulto se lleva a cabo en la médula ósea y da origen a las células B de zonamarginal y a las células B foliculares (B).
Pro -B Pre-B B inm B1
Pro -B Pre-B B inm
BZM
BFO
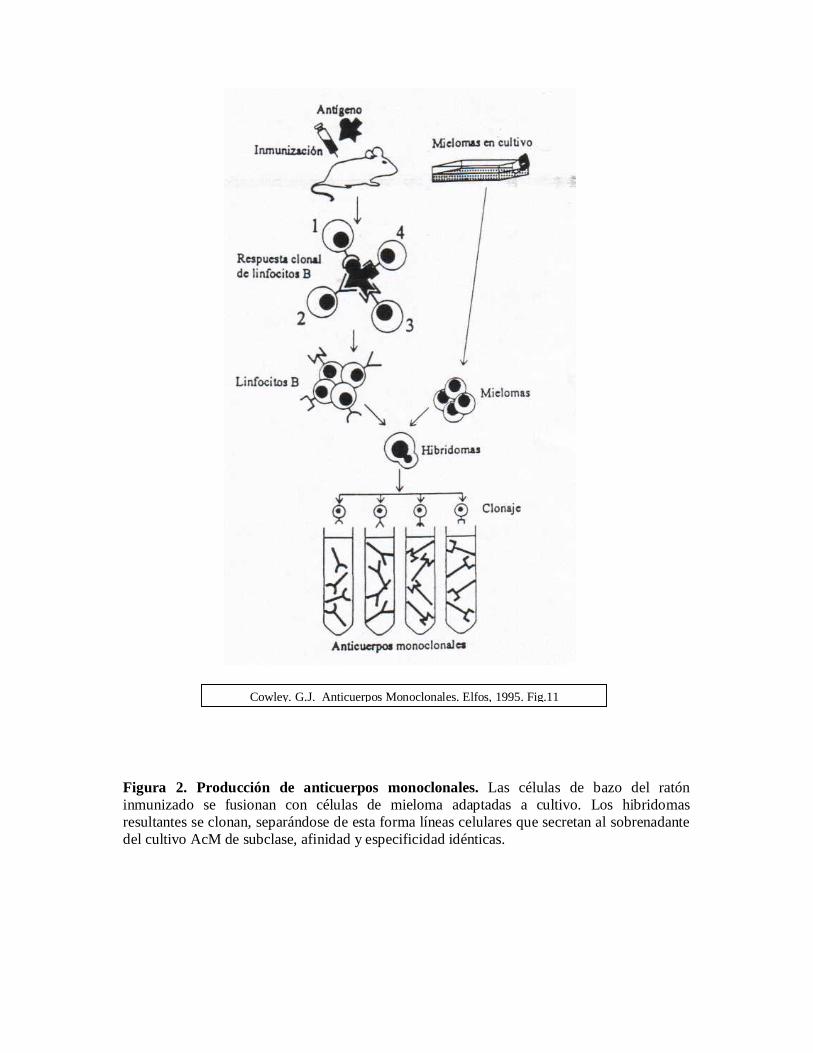
Figura 2. Producción de anticuerpos monoclonales. Las células de bazo del ratóninmunizado se fusionan con células de mieloma adaptadas a cultivo. Los hibridomasresultantes se clonan, separándose de esta forma líneas celulares que secretan al sobrenadantedel cultivo AcM de subclase, afinidad y especificidad idénticas.
Cowley. G.J. Anticuerpos Monoclonales. Elfos, 1995. Fig.11

OBJETIVOS:
Objetivo General:
Caracterizar la expresión del antígeno reconocido por el anticuerpo monoclonal NIM-R3sobre subpoblaciones de linfocitos murinos.
Objetivos particulares:
§ Analizar la expresión del antígeno reconocido por el anticuerpo monoclonal NIM-R3mediante ensayos de Western Blot.
§ Caracterizar la expresión del antígeno reconocido por el anticuerpo NIM-R3 sobresubpoblaciones de linfocitos.
§ Caracterizar la expresión del antígeno reconocido por el anticuerpo NIM-R3 sobresubpoblaciones de linfocitos B activados in vitro.

MATERIALES Y MÈTODOS
Animales.Se emplearon ratones BALB/c hembras, obtenidos del bioterio del CINVESTAV, con edadesde 6 a 8 semanas, así como ratones BALB/c hembras y machos nu/nu, obtenidos del bioteriodel Instituto Nacional de Nutrición.
Anticuerpos y conjugados.En las tinciones para citometría de flujo, se empleó el anticuerpo NIM-R3 como sobrenadantede cultivo del hibridoma o líquido de ascitis, sobrenadante de células XMMEN como controlde isotipo, puesto que los tres hibridomas producen Ig-M de rata se utilizó como anticuerposecundario anti-IgM de rata acoplada a FITC (Cappel,), así. como los siguientes anticuerpos:anti-IgM-PE (Southern Biotechnology Associates, Inc), anti-B220-PE, anti-B220-FITC, anti-CD21-FITC y anti-CD23-PE (Pharmingen).
Cultivo de hibridomas in vitro.Los hibridomas NIM-R3 y XMMEN fueron crecidos en medio RPMI 1640, suplementadocon 10% (v/v) de suero fetal bovino, 2mM de glutamina, 1000 µg/mL de estreptomicina y1000 µg/mL de penicilina (todos de Gibco) e incubadas a 37°C con 5% de C02.
Cultivo de hibridomas in vivo.El hibridoma NIM-R3 fue crecido in vivo, inoculando intraperitonealmente 5 x 106 célulasresuspendidas en 500µL de PBS en un ratón nu/nu. 10 a 15 días después, se extrajo ellíquido de ascitis con una aguja de 18 x 38 mm vía intraperitoneal. El sobrenadante serecuperó por centrifugación a 2500rpm durante 15 minutos.
Biotinación del anticuerpo monoclonal NIM-R3.Parte del líquido de ascitis de anticuerpo NIM-R3 obtenido se dializó con un buffer 0.1 MNaHCO3, 0.1 M NaCl, pH 7.4 durante dos días, a una temperatura de 4°C, realizando doscambios por día (200mL por cambio). Pasado el segundo día, al líquido de ascitis dializado sele adicionó 10 µL de 10 mg/ mL de biotina en DMSO por cada miligramo de anticuerpo y seincubó durante una hora a temperatura ambiente. Posteriormente para quitar la biotina noadherida se realizaron diálisis contra PBS, pH 7.4; realizando tres cambios por día (400mLpor cambio), a una temperatura de 4°C.
Suspensiones celulares.Los animales fueron sacrificados por dislocación cervical, a continuación se retiró la pielabdominal y las células de cavidad peritoneal se obtuvieron por lavados de la misma con PBS,mientras que las células de médula ósea se obtuvieron por perfusión de la cavidad del fémurcon PBS.Las células de nódulos linfáticos mesentéricos, placas de Peyer y timo fueron obtenidasmediante la extracción del órgano correspondiente, y posterior disgregación en una malla deacero inoxidable, manteniendo las células en PBS. Posteriormente las células fueroncentrifugadas durante 5 minutos a 1500 rpm, y el botón celular resuspendido en PBS; lascélulas de bazo fueron obtenidas mediante el mismo procedimiento anterior; al igual que lascélulas de médula ósea se les aplicó un tratamiento con 4 mL de regulador de lisis deglóbulos rojos (150 mM de cloruro de amonio, 10 mM de bicarbonato de potasio y 0. 1 mM

de EDTA) durante 4 minutos, tras los cuales, se lavaron con PBS, y las células fueronrecuperadas por centrifugación y resuspendidas en PBS.Las células fueron contadas al microscopio, utilizando azul tripano (0.4% de azul tripano enPBS) para excluir células muertas.
Tinción para citometría de flujoLas células de bazo, placas de Peyer, médula ósea y Cavidad Peritoneal fueron teñidas conanti-B220-PE. En todos los casos las células fueron teñidas con sobrenadante de cultivo decélulas NIM-R3 o XMMEN (control isotipo), seguidas de anti-IgM de rata acoplado a FITC.Con el fin de analizar subpoblaciones linfoides en bazo, se utilizaron las combinaciones anti-CD21-FITC y anti-CD23-PE, mientras que en las células de médula ósea fueron teñidas conanti-B220-FITC y anti-IgM-PE.El procedimiento consistió en incubar 1 x 106 células en 100 µL de NIM-R3 o XMMEN, másla mezcla de anticuerpos correspondiente, tras lo cual se lavaron con 2 mL de PBS y lascélulas se recuperaron por centrifugación a 1500 rpm durante 5 minutos. Posteriormente seempleó el anticuerpo anti-IgM de rata – FITC, se incubaron con 30µL de una dilución 1:2000durante 20 minutos en oscuridad. Cuando se empleó el anticuerpo NIM-R3 acoplado abiotina, las células se incubaron con 30µL de una dilución 1:30 de SA-APC, posteriormentelas células se lavaron como se explicó anteriormente y se fijaron en 500µL de FIX-FACS(PBS conteniendo 1% de formaldehído) antes de leerse.Los datos de al menos 104 células fueron obtenidos en un equipo FACSCalibur con elprograma CellQuest (Becton-Dickinson) y analizados con el programa WinMDI 2.8([email protected]).
Activación Celular in vitro.Una suspensión de células de bazo sin eritrocitos obtenida como se indicó anteriormente, fuecontada y un millón de células viables fueron incubadas en medio RPMI (10 % (v/v) de suerofetal bovino, 2mM de glutamina, 1000 µg/mL de estreptomicina y 1000 µg/mL de penicilina)con el siguiente estímulo: 20µg/mL de LPS (Sigma), durante 24, 48 o 72 horas a 37° C con5% de C02. Después de la incubación, las células fueron teñidas con 100 L de sobrenadantede NIM-R3, o bien de la línea celular XMMEN como control Isotipo y anti-B220-PE,utilizando como anticuerpo secundario un anti-IgM de rata –FITC.
Lisado de Células.Las células de bazo y médula ósea (10 8 células/ml) se incubaron durante 60 minutos en frío a4° C con el regulador de lisis (Tris-HCl 10 mM, pH 7.3, 150 mM de NaCl, 2mM de Na3VO4 ,10 mM de NaF y 0.4 mM de EDTA, 10 mM Paranitrofenilfosfato disódico (pNPP), PMSF1mM, 2.7 l / mL de aprotinina y 2.5 l / mL de leupeptina, conteniendo 1% del detergenteNP – 40. Transcurrido ese tiempo, los lisados se transfirieron a tubos Ependorf de 1.5 mL,poniendo 1 millón de células en 1 mL de lisado por tubo y se centrifugaron a 1400 rpm por 15minutos, para sedimentar el material insoluble y nuclear. Después de esto, los sobrenadantesse recuperaron y se les hizo la cuantificación de Proteínas mediante el Método de Lowrymidiendo la absorbancia a 680 nm. Se cargaron 60 g / mL de cada una de las muestrassobre geles de poliacrilamida al 10 % (SDS – PAGE) y se corrieron en una cámara deelectroforesis mini-protean 3 electrophoresis system (BIO – RAD).

Inmunoelectrotransferencia (Wester Blot):Las proteínas separadas en los geles de poliacrilamida se electrotransfirieron a papel denitrocelulosa (Hybond – ECL, Amersham) a 130 volts por 1.5 horas de baño en hielo según elmétodo descrito por Towbin y cols (1979) en una cámara de BIO – RAD. Las membranasde nitrocelulosa bloquearon en un amortiguador PBS-Tween 20 al 0.1% a temperaturaambiente con agitación , después se tiñeron con solución Rojo de Ponceau y enseguida selavaron tres veces durante diez minutos con PBS-T.
Las proteínas se revelaron por el siguiente método. A las membranas se les adicionó el primeranticuerpo (NIM-R3), se incubaron de 1 a 2 hrs a temperatura ambiente. Transcurrido eltiempo con el primer anticuerpo, las membranas se lavaron 3 veces por 7 minutos con PBS-T,las membranas se incubaron con el reactivo o anticuerpo secundario (anti-IgM rata marcadocon HRP) de 1 a 2 hrs a temperatura ambiente. Transcurrido ese tiempo, se lavaron otras tresveces con PBS-T durante diez minutos y se revelan por quimioluimiscencia (ECL-Amersham) usando placas sensibles a la luz desprendida en la reacción (Konica Medical FilmAX).

RESULTADOS
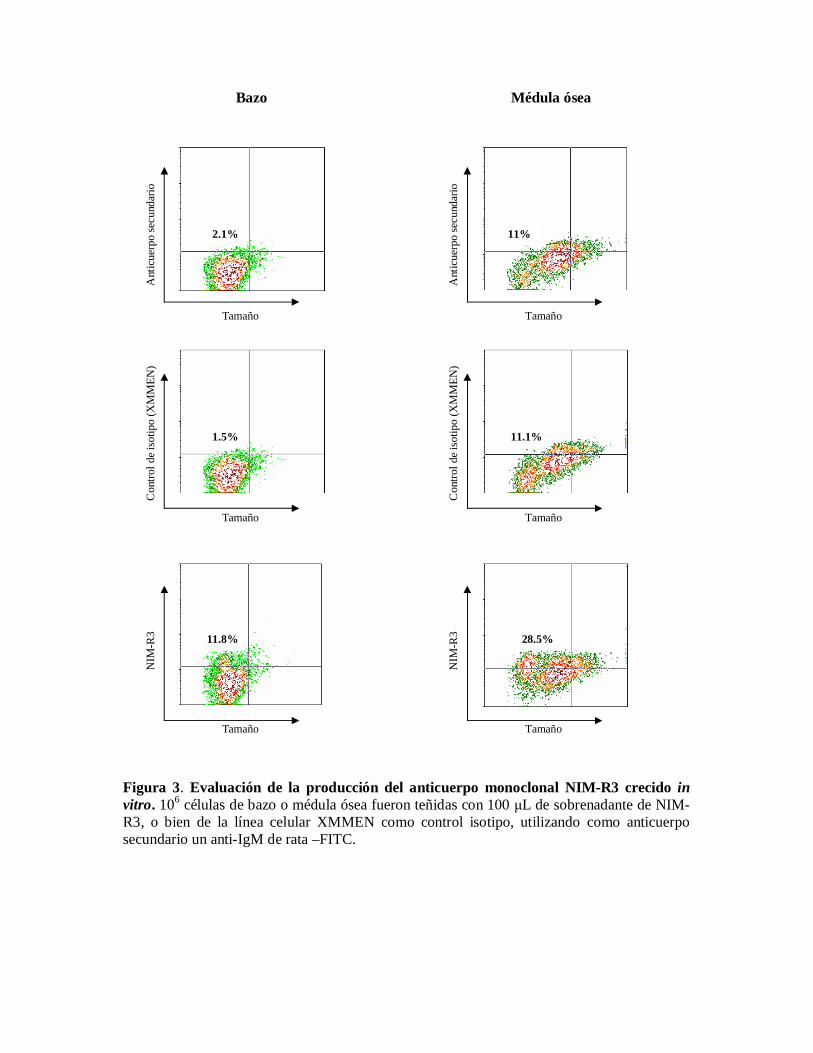
Evaluación de la producción del anticuerpo monoclonal NIM-R3 crecido in vitro.Para confirmar que el anticuerpo monoclonal se estuviera produciendo se realizaron tincionescon sobrenadantes de cultivo del hibridoma NIM-R3 en células de la línea celular A20, lacual, previamente había sido caracterizada en nuestro laboratorio. En la figura 3 puedeobservarse que si hay producción de anticuerpos.
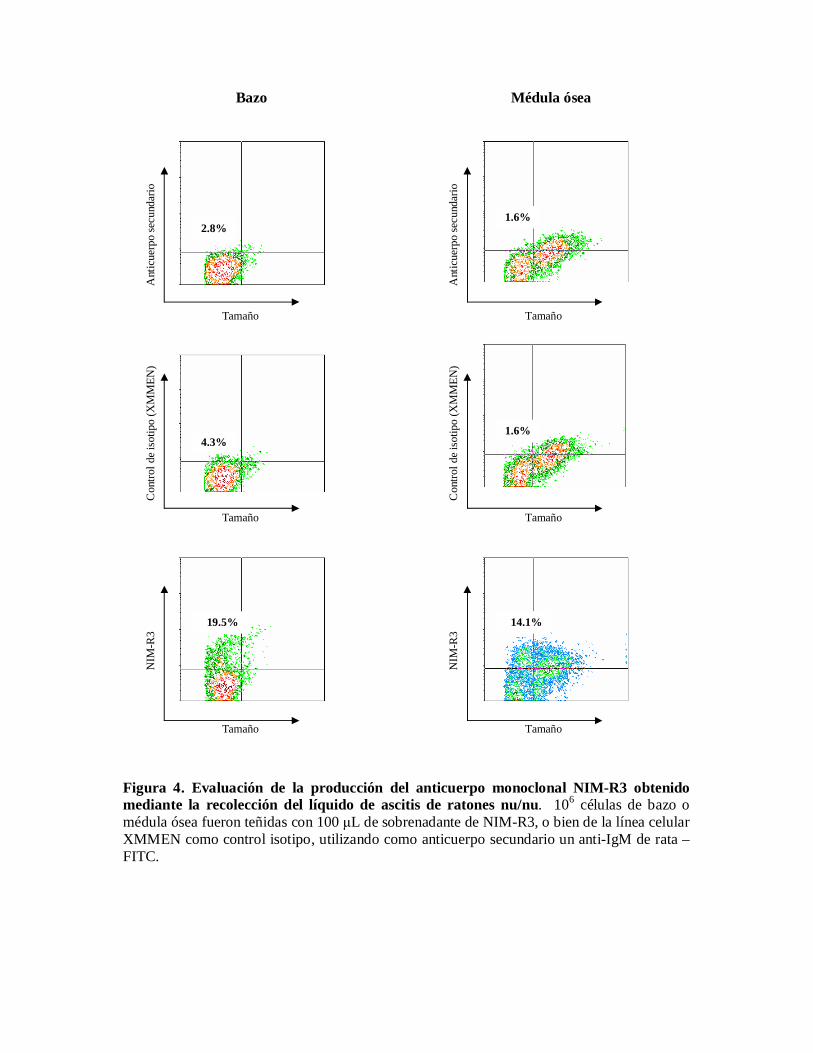
Evaluación de la producción del anticuerpo monoclonal NIM-R3 en líquido de ascitis.Una vez que se inocularon los ratones BALB/c nu/nu con el sobrenadante de NIM-R3positivo, para comprobar que el anticuerpo se produjo, se realizaron tinciones en células demédula ósea, utilizando líquido de ascitis recolectado del ratón. En la figura 4 se observaclaramente la tinción del anticuerpo sobre células de médula ósea, con respecto a los controlestanto el control del anticuerpo secundario (anti-IgM) como el control isotipo (línea celularXMMEN).
Reconocimiento de proteínas por el anticuerpo NIM-R3 en ensayos de Western-Blot.Con el fin de analizar la expresión del antígeno reconocido por el anticuerpo NIM-R3, serealizaron ensayos de Western-Blot; y como puede observarse en la figura 5 se observa unabanda en el carril 1, el cual corresponde a la línea celular BCL1; la cuál nos indica que elanticuerpo monoclonal NIM-R3 expresa una proteína de entre 26 y 36 kDa, aproximadamentede 30 kDa.
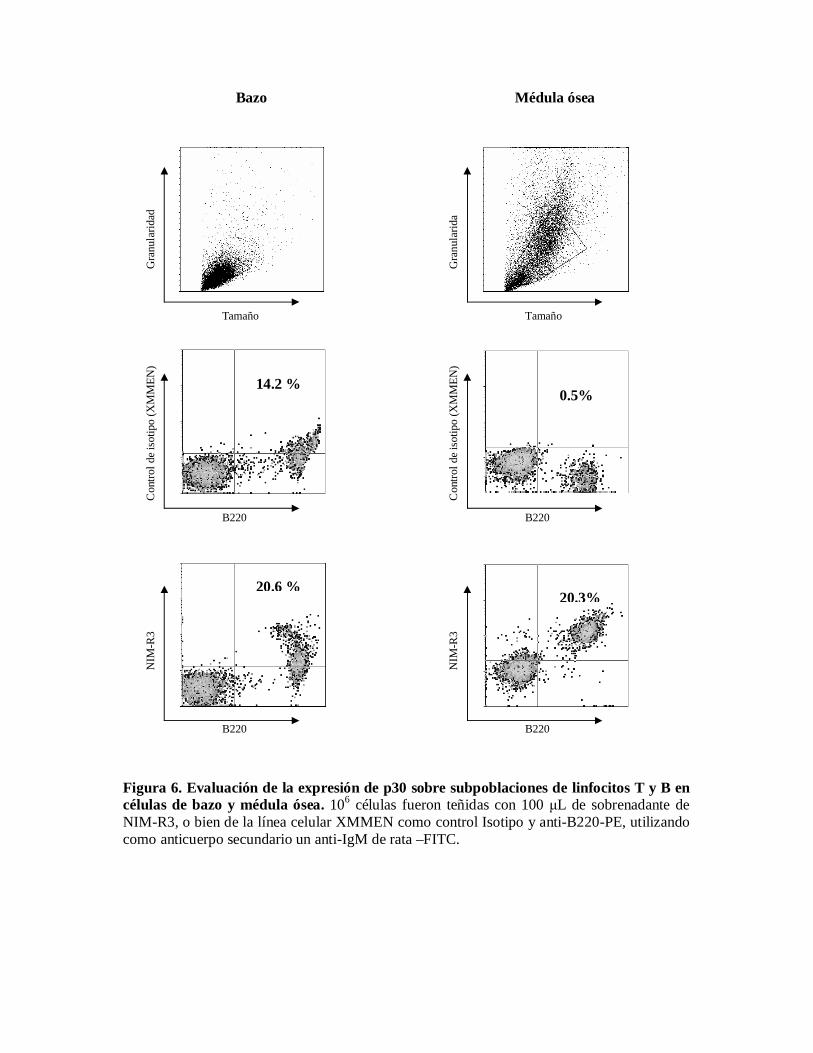
Expresión de p30 sobre subpoblaciones de linfocitos en células de diferentescompartimentos linfoides.Una vez que confirmamos que el hibridoma estaba produciendo el anticuerpo, para continuarcon el estudio de la expresión de p30, se realizaron tinciones de células de compartimentoslinfoides como bazo, médula ósea, placas de Peyer y cavidad peritoneal. En la figura 6 y 7puede verse que la expresión de p30 es muy alta y esta expresión sólo se observa en células B,sin embargo esta expresión no se presenta en toda la población de células B.
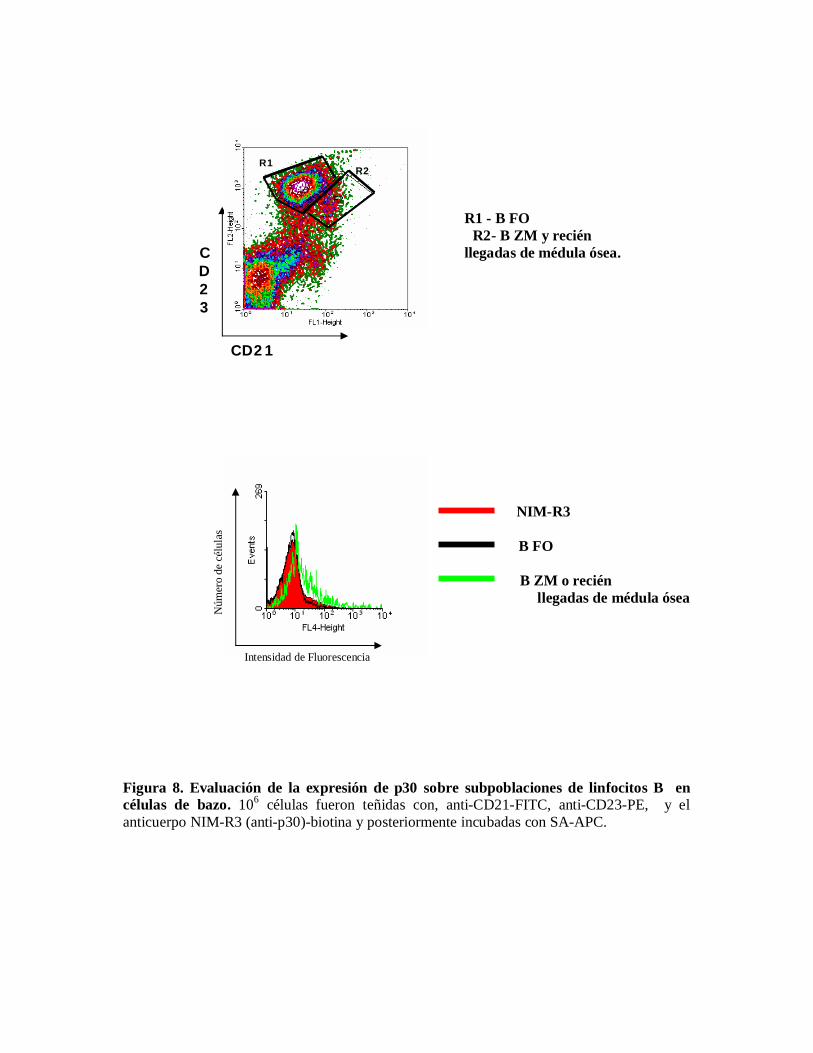
Evaluación de la expresión de p30 sobre subpoblaciones de linfocitos B en células debazo y médula ósea.p30 se expresa en células B tanto de bazo como de médula ósea, dado que en bazo haydiferentes poblaciones de células B, tales como B2 y células B de zona marginal, se procedióa definirlas por la expresión de CD23 y CD21; en la figura 8 puede verse que al separar laspoblaciones se aprecia que las células que corresponden a la subpoblación de células B dezona marginal y / o a células recién llegadas de médula ósea; son las que expresan la mayorcantidad de p30; en cambio la expresión de p30 sobre la subpoblación de células B1 es muybaja.
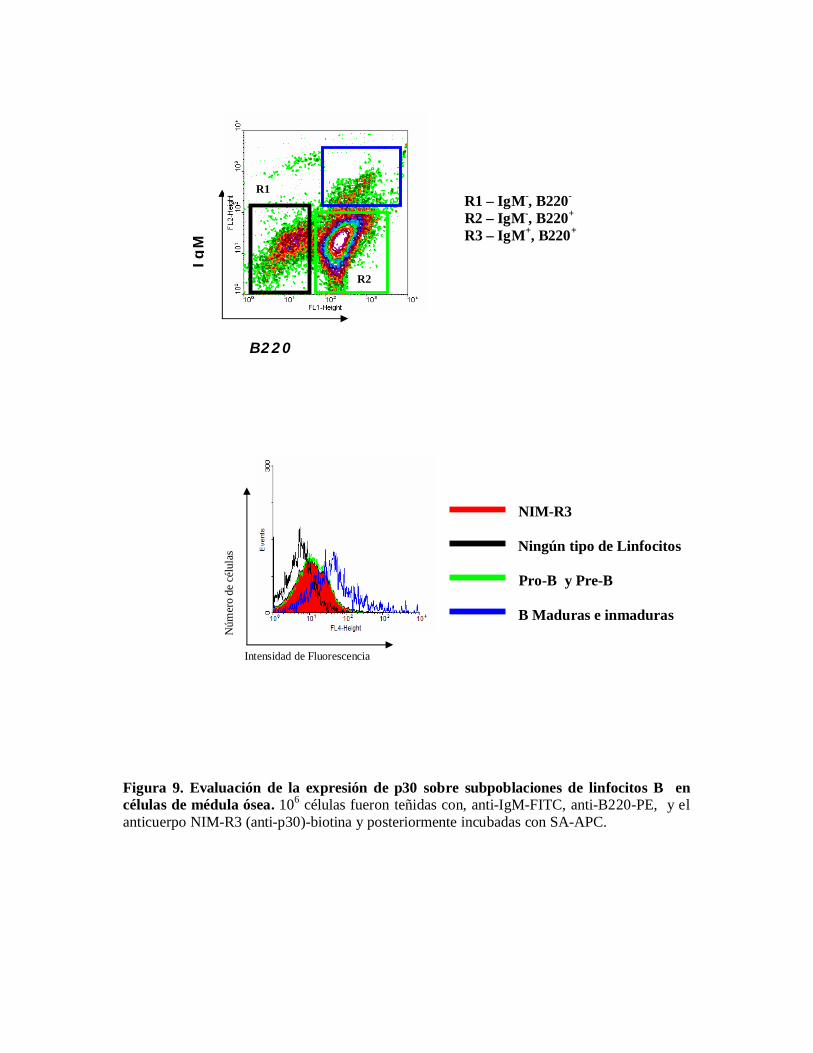
Por otra parte, para caracterizar la expresión de p30 sobre subpoblaciones de linfocitos B encélulas de médula ósea, se emplearon como marcadores los anticuerpos B220 e IgM. Elanálisis mostró que p30 se expresa en células B maduras e inmaduras (B220+, IgM+) y enmenor grado en células pro-B y pre-B (B220+, IgM-) (Figura 9).

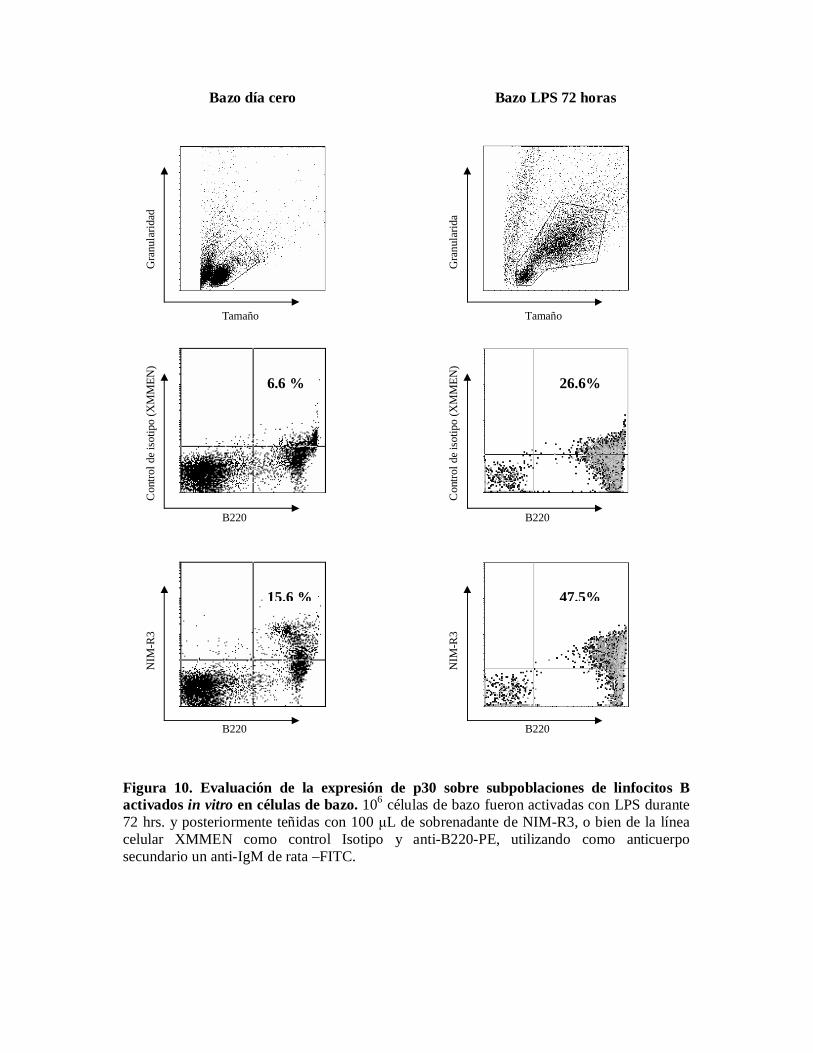
Evaluación de la expresión de p30 sobre subpoblaciones de linfocitos B activados in vitroen células de bazo.Puesto que uno de los objetivos de este trabajo consiste en conocer como se modifica laexpresión del antígeno reconocido por el anticuerpo monoclonal NIM-R3 durante laactivación, decidimos activar células de bazo con LPS durante 72 horas, tras lo cual, laexpresión basal de p30, que es aproximadamente del 16% se incrementó hasta un 47 %.(Figura 10).
Bazo Médula ósea
Figura 3. Evaluación de la producción del anticuerpo monoclonal NIM-R3 crecido invitro. 106 células de bazo o médula ósea fueron teñidas con 100 L de sobrenadante de NIM-R3, o bien de la línea celular XMMEN como control isotipo, utilizando como anticuerposecundario un anti-IgM de rata –FITC.
Tamaño
Ant
icue
rpo
secu
ndar
io
Tamaño
Con
trol d
e iso
tipo
(XM
MEN
)N
IM-R
3
Tamaño
Tamaño
Ant
icue
rpo
secu
ndar
io
Tamaño
Con
trol d
e iso
tipo
(XM
MEN
)N
IM-R
3
Tamaño
11.8%
1.5%
2.1%
11.1%
28.5%
11%
Bazo Médula ósea
Figura 4. Evaluación de la producción del anticuerpo monoclonal NIM-R3 obtenidomediante la recolección del líquido de ascitis de ratones nu/nu. 106 células de bazo omédula ósea fueron teñidas con 100 L de sobrenadante de NIM-R3, o bien de la línea celularXMMEN como control isotipo, utilizando como anticuerpo secundario un anti-IgM de rata –FITC.
Tamaño
Ant
icue
rpo
secu
ndar
io
Tamaño
Con
trol d
e iso
tipo
(XM
MEN
)N
IM-R
3
Tamaño
Tamaño
Ant
icue
rpo
secu
ndar
io
Tamaño
Con
trol d
e iso
tipo
(XM
MEN
)N
IM-R
3
Tamaño
2.8%
1.6%
14.1%
4.3%
19.5%
1.6%
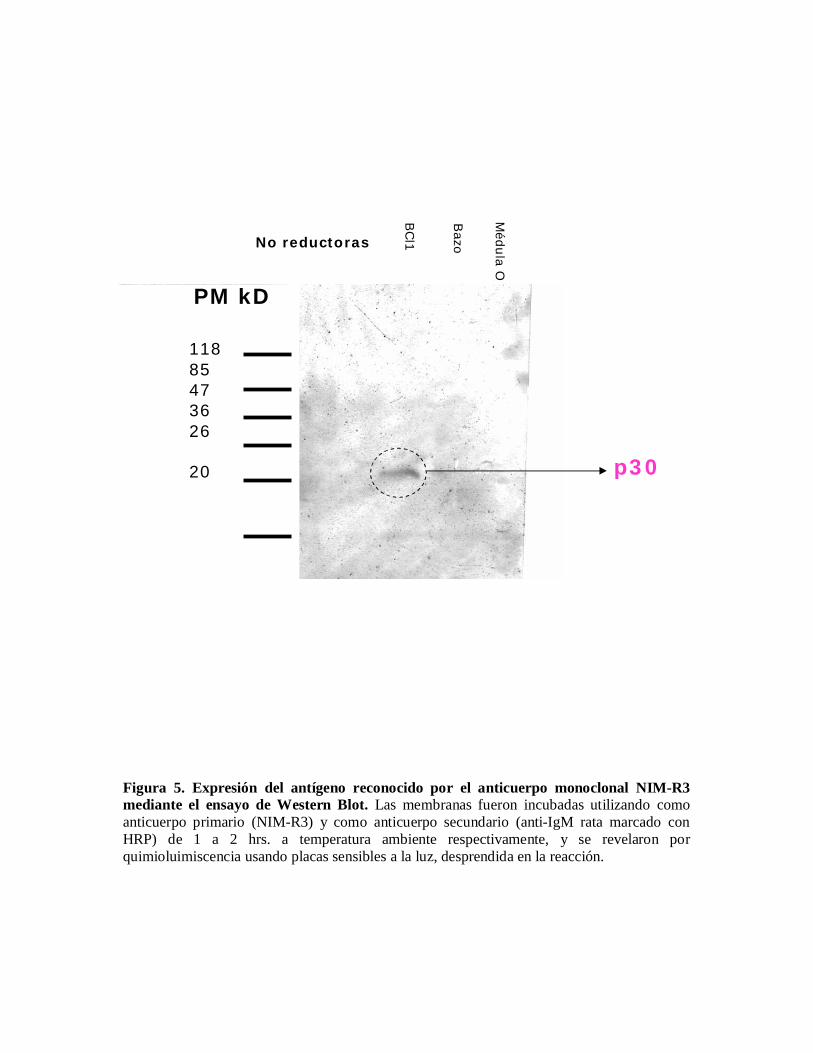
Figura 5. Expresión del antígeno reconocido por el anticuerpo monoclonal NIM-R3mediante el ensayo de Western Blot. Las membranas fueron incubadas utilizando comoanticuerpo primario (NIM-R3) y como anticuerpo secundario (anti-IgM rata marcado conHRP) de 1 a 2 hrs. a temperatura ambiente respectivamente, y se revelaron porquimioluimiscencia usando placas sensibles a la luz, desprendida en la reacción.
Méd
ula O
Bazo
BCl1
11885473626
20
PM kD
No reductoras
p30
Bazo Médula ósea
Figura 6. Evaluación de la expresión de p30 sobre subpoblaciones de linfocitos T y B encélulas de bazo y médula ósea. 106 células fueron teñidas con 100 L de sobrenadante deNIM-R3, o bien de la línea celular XMMEN como control Isotipo y anti-B220-PE, utilizandocomo anticuerpo secundario un anti-IgM de rata –FITC.
Tamaño
Gra
nula
rida
B220
Con
trol d
e iso
tipo
(XM
MEN
)N
IM-R
3
B220
Tamaño
Gra
nula
ridad
B220
Con
trol d
e iso
tipo
(XM
MEN
)N
IM-R
3
B220
14.2 %
20.6 %
0.5%
20.3%

Cavidad peritoneal Placas de Peyer
Figura 7. Evaluación de la expresión de p30 sobre subpoblaciones de linfocitos T y B encélulas de Cavidad Peritoneal y Placas de Peyer. 106 células fueron teñidas con 100 L desobrenadante de NIM-R3, o bien de la línea celular XMMEN como control Isotipo y anti-B220-PE, utilizando como anticuerpo secundario un anti-IgM de rata –FITC.
Tamaño
Gra
nula
rida
B220
Con
trol d
e iso
tipo
(XM
MEN
)N
IM-R
3
B220
Tamaño
Gra
nula
ridad
B220
Con
trol d
e iso
tipo
(XM
MEN
)N
IM-R
3
B220
21.2 %
42.2 %
4.1%
12.3%
R1 - B FO R2- B ZM y recién
llegadas de médula ósea.
NIM-R3
B FO
B ZM o recién llegadas de médula ósea
Figura 8. Evaluación de la expresión de p30 sobre subpoblaciones de linfocitos B encélulas de bazo. 106 células fueron teñidas con, anti-CD21-FITC, anti-CD23-PE, y elanticuerpo NIM-R3 (anti-p30)-biotina y posteriormente incubadas con SA-APC.
CD21
CD23
R1R2
Núm
ero
de c
élul
as
Intensidad de Fluorescencia
R1 – IgM-, B220-
R2 – IgM-, B220+
R3 – IgM+, B220+
NIM-R3
Ningún tipo de LinfocitosO
Pro-B y Pre-B
B Maduras e inmaduras
Figura 9. Evaluación de la expresión de p30 sobre subpoblaciones de linfocitos B encélulas de médula ósea. 106 células fueron teñidas con, anti-IgM-FITC, anti-B220-PE, y elanticuerpo NIM-R3 (anti-p30)-biotina y posteriormente incubadas con SA-APC.
B220
Intensidad de Fluorescencia
Núm
ero
de c
élul
as
R1
R22
IgM
Bazo día cero Bazo LPS 72 horas
Figura 10. Evaluación de la expresión de p30 sobre subpoblaciones de linfocitos Bactivados in vitro en células de bazo. 106 células de bazo fueron activadas con LPS durante72 hrs. y posteriormente teñidas con 100 L de sobrenadante de NIM-R3, o bien de la líneacelular XMMEN como control Isotipo y anti-B220-PE, utilizando como anticuerposecundario un anti-IgM de rata –FITC.
Tamaño
Gra
nula
rida
B220
Con
trol d
e iso
tipo
(XM
MEN
)N
IM-R
3
B220
Tamaño
Gra
nula
ridad
B220
Con
trol d
e iso
tipo
(XM
MEN
)N
IM-R
3
B220
6.6 %
15.6 %
26.6%
47.5%

DISCUSIÒN
El análisis de los resultados de la expresión del antìgeno reconocido por el anticuerpomonoclonal NIM-R3 mediante los ensayos de Western-Blot nos muestran que NIM-R3reconoce una proteína de aproximadamente 30 kDa (p30).
Como antecedentes se tenían que NIM-R3 teñía células B en bazo y una gran proporción decélulas de médula ósea, por lo que se proponía podría marcar subpoblaciones de células B.(Chayen y Parkhouse, 1982). Este resultado fue corroborado cuando se demostró queúnicamente las células B se tiñeron con el anticuerpo NIM-R3, y esto se logró con la ayuda deel marcador B220 (una isoforma de la fosfatasa CD45) el cual caracteriza a los linfocitos Bde ratón desde que se encuentran en el estadio pro-B, y debido a esta característica se utilizacomo marcador de células B murinas. Nuestros datos nos indicaron que efectivamente p30 seexpresaba sólo sobre células B de diferentes compartimentos linfoides como bazo, médulaósea, Cavidad Peritoneal y Placas de Peyer, sin embargo p30 solo se expresa en una solasubpoblaciòn. Por lo que para continuar con el estudio de la expresión de p30 se realizarontinciones dobles sobre células tanto de médula ósea como de bazo, con el fin de determinarsobre que subpoblaciones de células B se expresaba p30.
En el caso de las células de bazo se utilizaron como marcadores los anticuerpos CD21 yCD23.
Se sabe que el proceso de linfopoyesis adulta, es al parecer, el responsable del origen de dossubpoblaciones de células B, las células B de zona marginal (B ZM) y las célulasB Foliculares (B FO).
Fenotìpicamente, esta población cuando se encuentra en reposo se caracteriza por la presenciade altos niveles del marcador CD23 y baja expresión de CD21.
En cambio, las células B de zona marginal se localizan en el bazo, entre el límite de la pulpablanca y el seno marginal y se caracterizan por expresar gran cantidad de moléculas CD21.
La expresión de p30 que se observó sobre células B de zona fue mayor con respecto a célulasB Foliculares, por lo que podemos suponer que p30 en bazo se expresa predominantementesobre células B de zona marginal , dentro de las cuales podemos incluir células B inmadurasrecién ingresadas de médula ósea.
Y en el caso de células de médula ósea los marcadores que se emplearon fueron B220 quecomo ya se mencionó anteriormente es caracteriza a los linfocitos B de ratón desde que seencuentran en el estadio pro-B y el anticuerpo IgM el cual tiñe células B maduras einmaduras, los resultados de la expresión de p30 en la subpoblaciones de células B en médulaósea nos revelaron que p30 marca en particular linfocitos B maduras e inmaduras y en menorgrado células pro-B y pre-B.
Con el fin de avanzar en la caracterización de p30, en este trabajo se intentó la purificación deNIM-R3, pero durante el desarrollo de la investigación nos enfrentamos con grandesdificultades ya que el anticuerpo (NIM-R3) es una IgM la cual es muy inestable y sensible adiferentes condiciones desnaturalizantes razón por la que no fue posible purificarlo.

La expresión de p30 es muy interesante, y el hecho de que hasta el momento se desconozcaaún su identidad bioquímica, no demerita los resultados de este trabajo ya que es el procesopor el que se ha transitado en la identificación y caracterización de muchas otras moléculas deactivación linfocitaria, misma que han permitido que se avance en el conocimiento delproceso de activación de los B y de los eventos derivados de él.
Todos estos resultados nos señalan que la expresión de p30 es inducida en linfocitos in vitro,y su expresión está restringida sólo a células B.

CONCLUSIONES
§ El anticuerpo Monoclonal NIM-R3 reconoce una proteína de entre 26 y 36 kDa (p30).
§ p30 se expresa en células B de diferentes compartimentos linfoides, sin embargo sólose expresa en una sola subpoblación.
§ En médula ósea p30 marca en particular linfocitos B: B220+, IgM+; las cualesrepresentan células B maduras e inmaduras y en menor grado células pro-B y pre-B.
§ En bazo p30 se expresa en una subpoblación que puede corresponder a las células Bde Zona Marginal y / o a células B llegadas de la médula ósea.
§ La expresión de p30 se induce con la activación.
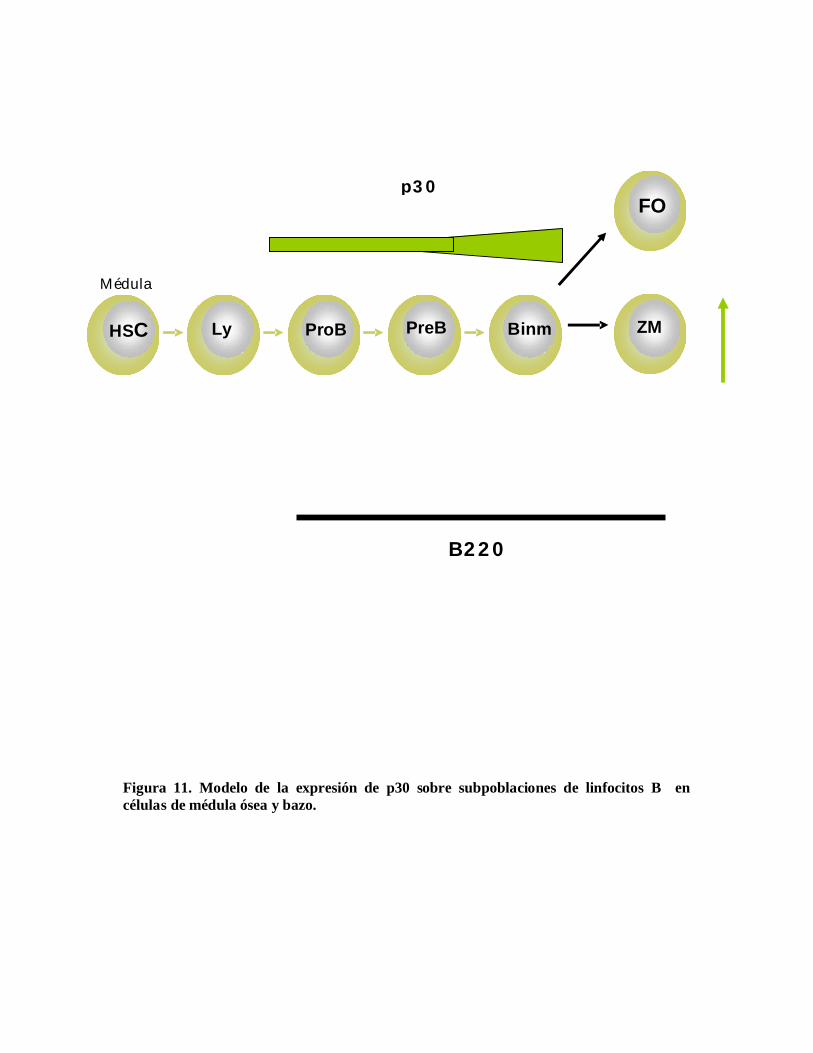
Figura 11. Modelo de la expresión de p30 sobre subpoblaciones de linfocitos B encélulas de médula ósea y bazo.
FO
ZM
Médula
Ly ProBHSC PreB Binm
p30
B220

BIBLIOGRAFÍA:
Berland R., Wortis H. H. 2002. Origins and functions of B-1 cells with notes on the role ofCD5. Annual Reviews in Immunology. 20:253-300.
Chapel H., Haeney M. 1992. Inmunología Clínica. Manual Moderno. México D.F 450-452.
Chayen A., Parkhouse R.M.E. 1982. B cell subpopulations in the mouse: analysis withmonoclonal antibodies NIM-R2 and NIM-R3. Eur. J. Immunol. 12: 725-732.
Cowley G. J. 1995. Anticuerpos Monoclonales. Elfos. 180 pp.
Greenwood M. R , Parkhouse R.M.E. 1986. Monoclonal antibody NIM-R3 substitutes for B-cell growth factor. Immunology 59: 7–13.
DeNardo S. J., Kroger L.A., DeNardo G. L. 1999. A new era for radiolabeled antibodies incancer?. Current Opinion in Immunology. 11:563-569.
Hayakawa K., Hardy R.R. 2000. development and function of B1 cells. Current Opinion inImmunology 12:346-353.
Khaw B. 1999. Antibodies as delivery systems for diagnostic functions. Asvanced DrugDelivery Reviews. 37:63-80.
Kohler, G. y Milstein C. 1975. Nature 256: 495.
Lu P., Wang Y.L., Linsley P.S. 1997. Regulation of self-tolerance by CD80/CD86interactions. Current Opinion in Immunology. 9:858-862.
Martin F., Oliver A.M. , Kearney J. F. 2001. Marginal zone and B1 B cells unite in the earlyresponse against T-independent blood-borne particulate antigens. Immunity 14:617-629.
Oliver A. M., Martín F., Kearney J.F. 1999. IgMhigh CD21high lymphocytes enriched in thesplenic marginal zone generate effector cells more rapidly than the bulk of follicular B cells.The journal of Immunology. 162:7198-7207.
Parkhouse R.M.E., Andrew E. M., Chayen A., Marshall-Clarke S. 1983. Heterogeneity of B-Cells. Annals New York Academy of Sciences. 215-229.
Parkhouse R.M.E., Askonas B. A. 1969. Biochem. J. 115:163-169.
Rolink A.G., Schaniel C., Andersson J., Melchers F.2001. Selection events operating atvarious stages in B cell development. Current Opinion in Immunology 13:202-207.
Snary D., Woods, F. R., Crumpton, M.J. 1976. Disruption of Solid Tissue for PlasmaMembrane Preparation.. Anal. Biochem. 74:457-465.

Stites P. D., Terr A. I. 1998. Inmunología básica y clínica. Manual Moderno. México D.F.117-127.
Taylor P.C., Williams R.O.Maini R.N.2001. Immunotherapy for rheumatoid arthritis.Current Opinion in Immunology.13:611-616.


















