
Efecto del Quitosano en la Funcionalidad de la Membrana ...
64
INSTITUTO POLITÉCNICO NACIONAL SECRETARÍA DE INVESTIGACIÓN Y POSGRADO ESCUELA NACIONAL DE CIENCIAS BIOLÓGICAS SECCIÓN DE ESTUDIOS DE POSGRADO E INVESTIGACIÓN Efecto del Quitosano en la Funcionalidad de la Membrana Plasmática de Rhizopus stolonifer T E S I S QUE PARA OBTENER EL GRADO DE: M A E S T R O EN C I E N C I A S Q U I M I C O B I O L Ó G I C A S P R E S E N T A Q. B. P. JOSAM VEGA PÉREZ DIRECTORAS DRA. MA. GUADALUPE GUERRA SÁNCHEZ DRA. ANA N. HERNÁNDEZ LAUZARDO MÉXICO, D.F. 2009
Transcript of Efecto del Quitosano en la Funcionalidad de la Membrana ...

INSTITUTO POLITÉCNICO NACIONAL
SECRETARÍA DE INVESTIGACIÓN Y POSGRADO ESCUELA NACIONAL DE CIENCIAS BIOLÓGICAS
SECCIÓN DE ESTUDIOS DE POSGRADO E INVESTIGACIÓN
Efecto del Quitosano en la Funcionalidad de la Membrana
Plasmática de Rhizopus stolonifer
T E S I S QUE PARA OBTENER EL GRADO DE:
M A E S T R O EN C I E N C I A S
Q U I M I C O B I O L Ó G I C A S
P R E S E N T A
Q. B. P. JOSAM VEGA PÉREZ
DIRECTORAS DRA. MA. GUADALUPE GUERRA SÁNCHEZ
DRA. ANA N. HERNÁNDEZ LAUZARDO
MÉXICO, D.F. 2009


El presente trabajo fue realizado en el Laboratorio de Bioquímica Microbiana del Departamento de Microbiología de la Escuela Nacional de Ciencias Biológicas del Instituto Politécnico Nacional bajo la dirección de la Dra. Ma. Guadalupe Guerra Sánchez* y de la Dra. Ana Niurka Hernández Lauzardo**.
Algunos de los experimentos de este trabajo fueron realizados en el Laboratorio 306 ote. del Instituto de Fisiología Celular de la Universidad Nacional Autónoma de México contando con el apoyado del Dr. Antonio Peña Díaz, de la QFB. Norma Silvia Sánchez y de la Dra. Martha Calahorra Fuertes.
Este trabajo fue financiado por la Secretaria de Investigaciones y Posgrados del IPN mediante los proyectos SIP 20080561*, 20080476**, 20090565* y 20090224**.
El sustentante recibió beca por parte del Consejo Nacional de Ciencia y Tecnología (CONACyT) de Agosto 2007 a Junio 2009, por el Programa Institucional de Formación de Investigadores (PIFI) Agosto 2007 a Diciembre 2009 y por parte del IPN a través de la Beca Tesis de Agosto 2009 a Diciembre 2009.

AGRADECIMIENTOS PROFESIONALES
En primer lugar a la Dra. Ma. Guadalupe Guerra Sánchez y a la Dra. Ana Niurka
Hernández Lauzardo, por la confianza, el apoyo y la paciencia que me brindaron
durante todo el desarrollo de esta tesis.
A los integrantes del Comité Tutoral
Dra. Ma. Guadalupe Guerra Sánchez
Dra. Ana Niurka Hernández Lauzardo,
Dra. Enriqueta Amora Lazcano
Dra. Ethel García Latorre
Dra. Lourdes Villa Tanaca
Dr. Miguel G. Velázquez del Valle
Por dedicar parte de su tiempo y su entusiasta colaboración en la revisión y
corrección del presente trabajo.
Al Dr. Antonio Peña Díaz, por faci l i tarme las instalaciones de su
laboratorio en Insti tuto de Fisiología Celular de la UNAM, también a
la QFB. Norma Silvia Sánchez y a la Dra. Martha Calahorra Fuertes,
por el apoyo que me brindaron durante la estandarización de las
técnicas que uti l icé en dicho laboratorio.
Al CONACyT, al PIFI y a la Beca Tesis, por la beca recibida durante
la maestría.

AGRADECIMIENTOS PERSONALES
A mi familia
A mis padres Carmen y Rafael, por no dejarme de apoyar nunca, por todo el cariño que me han brindado, por haberme hecho una persona responsable, por todo esto y más este nuevo logro lo comparto con ustedes esperando que vengan muchos más para poderlos seguir disfrutando juntos.
A Bianca por la compañía, la comprensión, el amor, la bonita relación que hemos disfrutado durante todo este tiempo. A Niam por demostrarme que con perseverancia y esfuerzo todos los retos se pueden superar y por ser mi aliciente más grande para culminar esta etapa.
A Jonadab por ser un apoyo constante, aun y cuando no sé diera cuenta, por las platicas que teníamos en las que algunas ocasiones terminamos discutiendo, pero de las cuales aprendía mucho y por supuesto por aguantar la convivencia de los últimos meses.
A mis amigos
Al Klan aunque el tiempo pase y ya no nos reunamos con tanta frecuencia, nuestra amistada se mantiene tan inquebrantable como desde el primer día: Trinidad, Sinuhe, Lorena, Brenda, Carlos, Francisco, Alejandro, Ricardo, Erica, Jaime, Octavio, Cesar, Miguel Alejandro, Perla, Viridiana, Paul, Carmen, etc., por escucharme, apoyarme, estar con migo en los buenos y malos momentos que vivimos y sobre todo por la “sana” diversión que tuvimos durante todo este tiempo.
A mis compañeros de laboratorio
A Leo, Karina, Brenda, Ariana, Selene, Oliver, Josuhe, Karolina, Alejandra, Antonio, Verónica, Griselda, Armando, por su ayuda al momento de resolver dificultades, por las largas pláticas que teníamos y por haber hecho más ameno el trabajo en el laboratorio.

i
ÍNDICE
ÍNDICE………………...…………………………………………………………………………….……… i
ÍNDICE DE ABREVIATURAS...………………………………………………………………….…….... iii
ÍNDICE DE FIGURAS……………………………………………………………………………….......... iv
ÍNDICE DE CUADROS.…….……………………………………………………………………….…... v
RESUMEN………….……………………………………………………………………………….….….. vi
ABSTRACT………..……………………………………………………………………………………..... vii
I INTRODUCCIÓN……………………………………………………………………………..... 1
1.1 Rhizopus stolonifer agente causal de pudriciones postcosecha……………………………. 1
1.2 Quitosano, alternativas de control natural…………………………………………….……….. 4
1.3 Mecanismos de acción del quitosano……………………..…………………………………… 6
1.4 Membrana celular.…………………………………………………………..……..................... 8
1.5 Las ATPasas…………………………………………………………………………………….. 10
1.6 Las ATPasas tipo-P…………………………………………………………………………….. 11
ANTECEDENTES……………………………………………………………………………… 14
JUSTIFICACIÓN………………………………………………………………………………… 15
HIPÓTESIS.…………………………………………………………………………………….. 16
OBJETIVO GENERAL………………………………………………………………………… 17
Objetivos particulares…………………………………………………………………..……….. 17
II MATERIAL Y MÉTODOS……………………………………………………………………… 18
2.1 Muestra biológica………………………………………………………………………………… 18
2.2 Solución de quitosano…………………………………….……………………………………... 18
2.3 Medios de cultivo………………………….……………………………………………………. 18
2.4 Condiciones de cultivo y obtención del inóculo fúngico ……………………………………... 19
2.5 Cinética de germinación…………………………………………………………………………. 19
2.6 Variación del pH durante el crecimiento de Rhizopus stolonifer………….......................... 19
2.7 Determinación de la integridad de la membrana de las esporas y micelio ……………….. 19

ii
2.8 Potencial de la membrana plasmática…………………………………………………………. 20
2.9 Salida de Potasio …………….……………………………………………………………......... 20
2.10 Obtención de membranas plasmáticas de R. stolonifer....…………………………………… 21
2.11 Cuantificación de fosfolípidos en la membrana plasmática…………………………………. 22
2.12 Observación de las membranas plasmáticas en el microscopio electrónico…………........ 22
2.13 Concentración de proteínas presentes en las membranas plasmáticas...………............... 22
2.14 Actividad de H+ ATPasa…………………………………………………………………..…….. 23
2.15 Cinética enzimática de la actividad de H+ ATPasa………………………………………........ 24
2.16 Diseño de experimentos y análisis estadístico…………………………………………..……. 24
2 III RESULTADOS……………………………………………………………………………………. 25
3.1 Cinética de germinación…………………………………………………………………………. 25
3.2 Variación del pH durante el crecimiento de Rhizopus stolonifer…………………..………... 26
3.3 Determinación de la integridad de la membrana de las esporas y del micelio……………. 27
3.4 Potencial de la membrana plasmática de esporas de R. stolonifer………………………… 30
3.5 Salida de potasio del micelio provocado por la presencia de quitosano…………………… 31
3.6 Fosfolípidos presentes en la membrana plasmática…………………………………………. 31
3.7 Observación de membranas plasmáticas en microscopio electrónico de transmisión………………………………………………………………………………….......... 32
3.8 Proteínas presentes en las membranas plasmáticas………………...……………………… 33
3.9 Actividad de H+ATPasa………………………………………………………………………….. 33
3.10 Cinética enzimática de la H+ATPasa de la membrana plasmática…………………………. 34
3 IV DISCUSION…...……………………………………………………………………………….….. 36
V CONCLUSIONES..……………………...…………………………………………………….….. 42
4 PERSPECTIVAS……………………………………………………………………………….…. 43
ANEXOS………………………………………………………………………………………..….. 44
REFERENCIAS…...………………………………..………………………………………….….. 47

iii
ÍNDICE DE ABREVIATURAS
AMPc Adenosín monofosfato cíclico
ATP Adenosín trifosfato
BSA Albumina sérica bovina
°C Grados centigrados
CCCP Carbonil cianida m-clorofenilhidrazona
DEAE-D Dietilaminoetil-Dextran
DHS Dihidroestreptomicina
DTT Ditiotreitol
EB Bromuro de etidio
EDTA Ácido etilendiaminotetraacético
EGTA Ácido etilenglicoltetraacético
h Horas
IM Integridad de membrana
M Molar
mg Miligramos
MM Medio mínimo
mM Minimolar
mL Mililitros
min Minutos
NADP Nicotinamida adenina dinucleótido fosfato oxidado
NADPH Nicotinamida adenina dinucleótido fosfato reducido
μg Microgramos
μl Microlitros
PDA Agar papa dextrosa
Pi Fosforo inorgánico
PI Ioduro de propidio
PMSF Fenilmetilsulfonil fluoruro
ROS Especies reactivas del oxigeno
RPM Revoluciones por minuto
SIDA Síndrome de inmunodeficiencia adquirida
TEA Trietanolamina
Tris Trishidroximetilaminometano
UV Luz ultra violeta

iv
ÍNDICE DE FIGURAS
Figura 1 Ciclo de vida de Rhizopus stolonifer............................................................................... 2
Figura 2 Esquema simplificado del proceso de desacetilación química de la quitina para obtener quitosano……………………………………………………………………………...
5
Figura 3 Representación esquemática de la estructura de la membrana celular……………. 8
Figura 4 Topología de la H
+ATPasa de la membrana plasmática de S. cerevisiae, basado en
cristalografía de segunda y tercera dimensión. …………………………………………… 12
Figura 5 Cinética de germinación…………………………………………….…………..................... 25
Figura 6 Microfotografía de la cinética de germinación……………………………………………... 26
Figura 7 Efecto del quitosano sobre el pH del medio de cultivo ………………………………..…. 27
Figura 8 Microfotografía del efecto del quitosano sobre la integridad de membrana plasmática de las esporas de Rhizopus stolonifer…………………………………………………….
28
Figura 9 Efecto del quitosano sobre la integridad de membrana plasmática de las esporas de Rhizopus stolonifer. ……………..…………………………………………………..……….
28
Figura 10 Micrografía del efecto del quitosano sobre la integridad de la membrana plasmática del micelio de Rhizopus stolonifer………………………………………………………….
29
Figura 11 Efecto del quitosano sobre el potencial de la membrana plasmática de las esporas de Rhizopus stolonifer. ………………………………………………………………………
30
Figura 12 Efecto del quitosano sobre la salida de K+ de las esporas de Rhizopus stolonifer…… 31
Figura 13 Microfotografía electrónica de transmisión de las membranas plasmáticas (MP) de R. stolonifer………………………………………………………………………………………...
32
Figura 14 Efecto del quitosano sobre la cinética enzimática de la H+ATPasa de membrana plasmática de Rhizopus stolonifer…………………………………………………………...
35

v
ÍNDICE DE CUADROS
Cuadro1 Aplicaciones del quitosano en diferentes áreas……………………………………..…... 6
Cuadro 2 Cuantificación de los mayores complejos lipídicos y esteroles de la membrana celular de dos diferentes hongos (µg mg
-1 proteína)…………………………………….
9
Cuadro 3 Comparación de la sensibilidad a los principales inhibidores de las ATPasas……………………………………………………………………………..............
11
Cuadro 4 Concentración de fosfolípidos calculada a las diferentes muestras de membranas plasmáticas de Rhizopus stolonifer crecido en MM durante 12 h………………………
32
Cuadro 5 Concentración de proteínas cuantificadas a las diferentes muestras obtenidas de Rhizopus stolonifer crecido en MM durante 12 h………………………………………....
33
Cuadro 6 Actividad de la H
+ATPasa calculada a las diferentes muestras de membranas
plasmáticas de Rhizopus stolonifer crecido en MM durante 12 h…………………….... 34
Cuadro 7
Valores de la Velocidad máxima y de la Km obtenidos del curso temporal de la actividad de la H
+ATPasa de membrana plasmática de Rhizopus
stolonifer……................................................................................................................. 35

vi
RESUMEN
Rhizopus stolonifer es considerado uno de los principales hongos fitopatógenos
que causan enfermedades postcosecha, es el agente causal de la pudrición
blanda de frutas y hortalizas, ocasionando importantes pérdidas económicas. El
control de este fitopatógeno se ha realizado utilizando fungicidas químicos
sintéticos. Sin embargo, los efectos nocivos que éstos han causado al medio
ambiente, los riesgos que representan para la salud humana y la resistencia que
han generado las cepas fúngicas motivan la búsqueda de alternativas naturales de
control. El quitosano, compuesto natural, biodegradable y no tóxico ha sido
evaluado para controlar las enfermedades potscosecha. El mecanismo de la
actividad antimicrobiana de este polímero no está del todo elucidado. El objetivo
de este trabajo fue estudiar el efecto del quitosano en la funcionalidad de la
membrana plasmática de Rhizopus stolonifer para lo cual se evaluó el efecto del
quitosano en la integridad de la membrana plasmática, se determinaron los
cambios en el pH del medio, potencial de membrana y en la salida de K+.
Adicionalmente, se cuantificaron los fosfolípidos totales y el contenido proteico de
la membrana plasmática en presencia de quitosano y se evaluó el efecto del
polímero en la actividad de la H+ATPasa de la membrana. Los resultados
obtenidos demuestran que el quitosano causa la pérdida de la integridad de la
membrana plasmática de R. stolonifer, afecta el pH del medio de cultivo, el
potencial de membrana y causa la salida K+ del interior celular. Sin embargo,
también se evidencia que no afecta el contenido de fosfolípidos totales pero
disminuye el contenido proteico de la membrana plasmática y la actividad de la H+
ATPasa. Por otra parte, se expone que el polímero disminuye los parámetros
cinéticos (Vmax y Km) de la H+ATPasa. Los resultados del presente estudio
demuestran que el quitosano afecta la funcionalidad de la membrana plasmática
de Rhizopus stolonifer y contribuyen a esclarecer los mecanismos de acción de
este compuesto.

vii
ABSTRACT
Rhizopus stolonifer is considered one of the main pathogens causing postharvest
diseases, is the causative agent of soft rot of fruits and vegetables, causing major
economic losses. Control of this plant pathogen has been conducted using
synthetic fungicides. However, the harmful effects they have caused to the
environment, the risks posed to human health and resistance to fungal strains
generated motivates the search for natural alternatives of control. Chitosan, is a
natural compound, biodegradable and nontoxic has been evaluated for postharvest
disease control. The mechanism of the antimicrobial activity of this polymer is not
fully elucidated. The aim of this work was to study the effect of chitosan on the
functionality of the plasma membrane of Rhizopus stolonifer for which we
evaluated the effect of chitosan on plasma membrane integrity, were determined
pH changes in the culture medium, potential membrane and the K+ efflux.
Additionally, were quantified the total phospholipids and protein content of the
plasma membrane in the presence of chitosan and evaluated the effect of the
polymer in the activity of membrane H+ATPase. The results demonstrated that
chitosan causes loss of plasma membrane integrity of R. stolonifer, affected the pH
of the culture medium, the membrane potential and causes the K+ efflux of the cell.
However, also demonstrated does not affect the total phospholipid content but
decreased the protein content of the plasma membrane and the activity of
H+ATPase. On the other hand, were showed that the polymer decreases the
kinetic parameters (Vmax and Km) of the H+ATPase. The results of this study
show that chitosan affects the functionality of the plasma membrane of Rhizopus
stolonifer and help to elucidate the mechanisms of action of this compound.

Efecto del quitosano en la funcionalidad de la membrana plasmática Rhizopus stolonifer INTRODUCCIÓN
1
INTRODUCCIÓN
1.1 Rhizopus stolonifer, agente causal de pudriciones postcosecha
Rhizopus stolonifer (Ehrenb.:Fr.) Vuill es considerado uno de los principales
hongos fitopatógenos que provocan enfermedades postcosecha, es el agente
causal de la pudrición blanda de frutas y hortalizas ocasionando importantes
pérdidas económicas. Este hongo pertenece a la Subdivisión: Zygomycotina,
Clase: Zygomycetes, Orden: Mucorales, Género: Rhizopus y Especie: Rhizopus
stolonifer (Agrios, 2001). Esta especie se encuentra ampliamente distribuida en la
naturaleza sobreviviendo de manera saprófita en el suelo y en residuos orgánicos
invadiendo potencialmente los tejidos vegetales. Sus principales características
son: crecimiento rápido, desarrolla un micelio aéreo cenocítico, forma
esporangióforos que presentan en sus puntas esporangios esféricos donde se
alojan las esporangiosporas, las cuales presentan diferentes formas: globosas,
elipsoidales y angulares con superficies lisas o estrías distintivas (Schipper, 1984;
Hernández-Lauzardo et al., 2006).
Las esporas de R. stolonifer pueden sobrevivir largos periodos sin agua y soportar
temperaturas elevadas, germinando sobre tejidos vegetales dañados y generando
rápidamente la maceración de los tejidos y la pudrición de los frutos, provocando
pérdidas considerables en un corto periodo de tiempo (Northover and Zhou, 2002;
Holmes and Stange, 2002).
R. stolonifer tiene la capacidad de reproducirse de forma asexual y sexual (figura
1). La reproducción asexual en los Zygomycetes tiene lugar después de que el
micelio ha experimentado un crecimiento extensivo y es por medio de
esporangiosporas. El esporangióforo es una estructura larga, en cuyo ápice se
forma el esporangio. El esporangio es el productor de esporangiosporas que
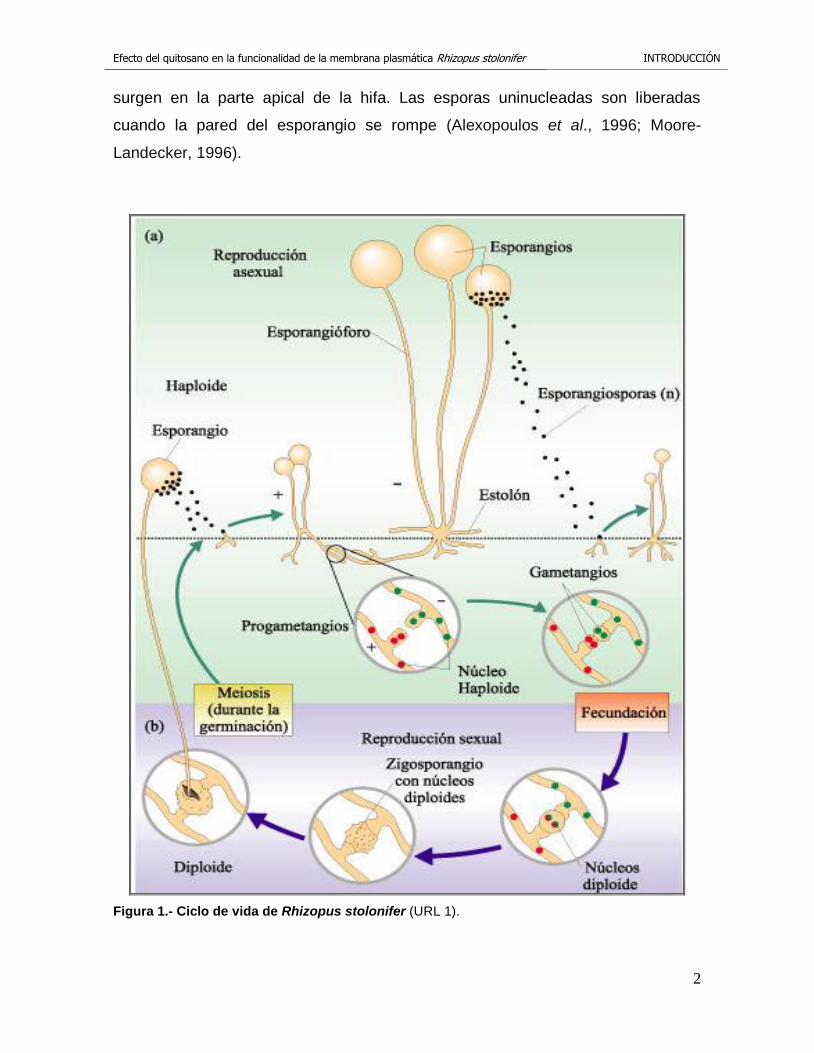
Efecto del quitosano en la funcionalidad de la membrana plasmática Rhizopus stolonifer INTRODUCCIÓN
2
surgen en la parte apical de la hifa. Las esporas uninucleadas son liberadas
cuando la pared del esporangio se rompe (Alexopoulos et al., 1996; Moore-
Landecker, 1996).
Figura 1.- Ciclo de vida de Rhizopus stolonifer (URL 1).

Efecto del quitosano en la funcionalidad de la membrana plasmática Rhizopus stolonifer INTRODUCCIÓN
3
La reproducción sexual se caracteriza por la fusión de gametos morfológicamente
idénticos contenidos en diferentes hifas, una hifa (+) y una hifa (-). La atracción de
las mismas es debida a la producción de hormonas volátiles. Estas sustancias
inducen el contacto de los ápices que se hinchan hasta que se fusionan los
gametangios multinucleados. Primero, el cigoto se rodea de una delgada pared y
dentro del cigoto comienza el desarrollo de la zigospora con una nueva pared más
gruesa. El material que es depositado en la misma, causa que la pared de la
zigospora se engruese y comience a pigmentarse. Las zigosporas maduras son
típicamente oscuras (negra o café). Una vez que empieza la germinación de la
zigospora, al esporangióforo le emerge en la punta un esporangio (Alexopoulos et
al., 1996; Moore-Landecker, 1996).
En la etapa de postcosecha es cuando los hongos provocan el mayor daño a los
cultivos, penetran los frutos dañados y generan la pudrición (González-Aguilar et
al., 2005). R. stolonifer aprovecha las heridas, magulladuras o aberturas naturales
que presentan los tejidos vegetales vivos para infectar, después las hifas secretan
enzimas pectinolíticas que degradan las sustancias pécticas de la lámina media de
las células, ocasionando la pérdida de cohesión entre las células, dado que se
rodean de una sustancia líquida y al momento de aplicar cierta presión pueden
moverse, esto provoca una “pudrición blanda” (Agrios, 2001).
Para controlar la pudrición blanda causada por R. stolonifer usualmente se
emplean diversas estrategias, como cambios físicos en el medio ambiente (bajas
temperaturas o tratamientos de calor), el empleo de atmósferas controladas y las
radiaciones [gamma (γ) y luz ultra violeta (UV)] (Velázquez-del Valle et al, 2008),
además del uso de productos químicos sintéticos como Dicloran, Iprodione,
Fludioxonil y Tebuconazole (Adaskaveg et al., 2002). Sin embargo, esta última
práctica ha provocado diversos problemas como son la resistencia generada por

Efecto del quitosano en la funcionalidad de la membrana plasmática Rhizopus stolonifer INTRODUCCIÓN
4
R. stolonifer, daños al medio y a la salud humana, entre otros, conllevando a la
búsqueda de alternativas naturales para controlar las pudriciones ocasionadas por
este fitopatógeno, entre las que se encuentra el control biológico y el empleo del
quitosano (Bautista-Baños et al., 2006).
1.2 Quitosano, alternativa de control natural
La quitina es el segundo biopolímero más abundante en la naturaleza y está
compuesto de monómeros de 2-acetil-amido-2-desoxi-β-D-glucopiranosa (Seyfarth
et al., 2008), es parte de los componentes que constituyen los caparazones de los
crustáceos (cangrejos, camarones, jaibas, etc.), así como de la pared celular de
los hongos, en donde es el componente estructural más importante, debido a que
es el responsable de mantener la forma e integridad de la pared celular (Allan and
Hadwiger, 1979; Martínez and Gonzalbo, 2001).
El quitosano es un derivado de la quitina, es un polímero que está compuesto por
monómeros de 2-acetil-amido-2-desoxi-β-D-glucopiranosa y monómeros
desacetilados de 2-amino-2-desoxi-β-D-glucopiranosa (Seyfarth et al., 2008),
también está presente en la pared celular de algunos hongos (Allan and Hadwiger,
1979). Por medio de una reacción de desacetilación se eliminan al menos el 50%
de sus grupos acetilo y la quitina se convierte en quitosano (figura 2) (Lárez,
2006). El grado de desacetilación del quitosano casi nunca es completo y de esto
dependerán las propiedades químicas, físicas y biológicas del polímero
(Hernández-Lauzardo et al., 2005; Seyfarth et al., 2008). Sus principales ventajas
son que es un producto inocuo, natural, biodegradable y no tóxico (Bautista-Baños
et al., 2006; Lárez, 2006).
La presencia de grupos aminos en la cadena polimérica ha hecho que el quitosano
sea uno de los materiales más versátiles que se estudian desde hace algún
tiempo, por la posibilidad de realizar una amplia variedad de modificaciones, tales

Efecto del quitosano en la funcionalidad de la membrana plasmática Rhizopus stolonifer INTRODUCCIÓN
5
como la reacciones de anclaje de enzimas, reacciones de injerto, obtención de
películas entrecruzadas, etc., de las cuales se obtienen materiales con
propiedades adecuadas para aplicaciones inmediatas y futuras en biotecnología,
biomedicina y agricultura (Cuadro 1) (Lárez, 2003).
Figura 2.- Esquema simplificado del proceso de desacetilación química de la quitina para obtener quitosano.
En la agricultura, se ha utilizado quitosano con diversos grados de polimerización
para cubrir frutos, verduras o raíces y prevenir el ataque por hongos y bacterias
durante la fase postcosecha con el propósito de alargar la vida de anaquel del
producto (Gonzalez-Aguilar et al., 2005; Hernández-Muñoz et al., 2006). El
carácter fungicida y/o fungistático que presenta el quitosano se ha demostrado en
una gran variedad de hongos entre los cuales se encuentran: Alternaria alternata,
Aspergillus flavus, Aspergillus parasiticus, Botrytis cinérea, Colletotrichum
gloeosporioides, Fusarium acuminatum, Fusarium graminearum, Fusarium
oxysporum, Micronectriella navils, Mucor spp. Penicillum sp., Pyricularia grisea,
Rhizotonia solani y Rhizopus stolonifer (Ait-Barka et al., 2004; Bautista-Baños et
al., 2004; Hernández-Lauzardo et al., 2005; 2007 Pacheco et al., 2008; Rabea et
al., 2005)
NaOH (40%) Temperaturas altas

Efecto del quitosano en la funcionalidad de la membrana plasmática Rhizopus stolonifer INTRODUCCIÓN
6
Cuadro 1.- Aplicaciones del quitosano en diferentes áreas.
Área Aplicación
Química analítica
En cromatografías, intercambiadores de iones, absorción
de iones de metales pesados y absorción de ácidos,
fabricación de electrodos específicos para metales, etc.
Biomedicina
Membrana de hemodiálisis, suturas biodegradables,
sustitutos artificiales de piel, agente cicatrizante en
quemaduras, sistemas liberadores de fármacos (insulina),
transporte de agentes anticancerígenos, tratamiento de
tumores, control del virus del SIDA, etc.
Industrias
Del papel, textil, alimentaría (soporte para inmovilización de
enzimas en la producción de maltosa, espesante en
alimentos, agente de oxidación controlada, agente
preservante).
Agricultura
Recubriendo semillas para su conservación durante el
almacenamiento, sistemas liberadores de fertilizantes, en
formulación de pesticidas, como agente bactericida y
fungicida para la protección de plántulas.
Tratamiento de agua
Como agente floculante, agente coagulante, tratamientos
de flotación para la remoción de aceite de pescado en
agua, remoción de metales, remoción de surfactantes.
Tomado de Lárez, 2003.
1.3 Mecanismo de acción del quitosano
El mecanismo de la actividad antimicrobiana del quitosano y sus derivados no está
completamente elucidado, pero se cuenta con una amplia información del efecto
que tienen las soluciones acuosas de quitosano frente a una diversa variedad de
microorganismos, incluyendo bacterias, levaduras y hongos filamentosos (Allan
and Hadwiger, 1979; Guo et al., 2006; Seyfarth et al., 2008). Lo que sugieren la

Efecto del quitosano en la funcionalidad de la membrana plasmática Rhizopus stolonifer INTRODUCCIÓN
7
mayoría de las investigaciones es que las cargas positivas en el grupo NH3+ del
monómero de glucosamina a pH <6.3 permite la interacción con las cargas
negativas de la membrana celular formando complejos polielectrolitos, lo cual
altera la permeabilidad de la membrana, interfiriendo con el crecimiento, las
funciones fisiológicas normales y provocando la fuga de constituyentes
intracelulares (Rabea et al., 2003; Kim and Rajapakse, 2005).
Otros estudios atribuyen el efecto antifúngico del quitosano a diferentes causas
(concentración, polimerización, acetilación, tiempo de incubación, etc.) pero de
todas formas existe un factor en común alrededor del cual versan todas las
explicaciones y es el relacionado con la naturaleza policatiónica de la molécula de
quitosano, la cual le confiere propiedades biológicas y fisiológicas únicas (Liu et
al., 2004). Como esta es una de las propuesta más aceptadas, existen grupos de
investigadores que se han dado a la tarea de utilizar compuesto derivados de la
quitina o similares en estructura pero con una diferencia principal no tener grupos
NH3+ en el monómero, obteniendo como resultados actividades antifúngicas muy
bajas, lo que sugiere que el carácter policatión del quitosano es crucial para tener
una alta actividad antifúngica (Seyfarth et al., 2008).
Estudios recientes avalan que el quitosano penetra a la célula fúngica mediante un
proceso dependiente de energía y permeabiliza y afecta diferentes tipos de células
de Neurospora crassa, resultando las esporas más sensibles al quitosano que las
hifas (Palma-Guerrero et al., 2009). Adicional a estos efectos, estudios por
microscopía electrónica han evidenciado efectos del quitosano sobre las
estructuras de hongos filamentosos, como lesiones, distorsiones y retracción de
las hifas, con alteración morfológica de organelos (Liu et al., 2007). Además
existen otro reporte donde se estudió por microscopía electrónica de barrido y de
transmisión las esporas de R. stolonifer obtenidas de diferentes tratamientos con
quitosano y las imágenes de los resultados mostraron que el quitosano provocó
cambios en la ornamentación y en la profundidad de la crestas de las esporas
(Hernández-Lauzardo et al., 2008).

Efecto del quitosano en la funcionalidad de la membrana plasmática Rhizopus stolonifer INTRODUCCIÓN
8
1.4 Membrana Celular
La membrana celular de los organismos eucariotas es la parte externa de la célula
que envuelve al citoplasma, es la que permite el intercambio entre la célula y el
medio que la rodea. La célula no puede funcionar apropiadamente y permanecer
viva a menos que su membrana regule el intercambio de compuestos. Las
membranas biológicas están compuestas por una bicapa lipídica de
aproximadamente 7.5 nm de grosor y está constituidas básicamente por lípidos,
proteínas y carbohidratos (proporciones aproximadas 50%, 40% y 10%,
respectivamente) (figura 3).
Figura 3.- Representación esquemática de la estructura de la membrana celular (URL2).
La mayor parte de los lípidos que se encuentran en las membranas de eucariotas
son fosfoglicerolípidos (fosfolípidos), esfingolípidos, glicoglicerolípidos y esteroles.
Los fosfolípidos son los elementos estructurales más importantes de las
membranas biológicas. Las membranas plasmáticas también poseen
carbohidratos unidos a proteínas (glicoproteínas) y a los lípidos (glicolípidos)
(Daum et al., 1998). Cada uno de estos componentes, es característico de las
membranas biológicas, y las proporciones en que están presentes varían
Esterol

Efecto del quitosano en la funcionalidad de la membrana plasmática Rhizopus stolonifer INTRODUCCIÓN
9
enormemente. La relación existente entre los lípidos y las proteínas de membrana
suele variar dependiendo del tipo celular estudiado (Cuadro 2). Por tanto, se
considera que la mayor parte de la estructura de las membranas depende de los
fosfolípidos y las funciones de las proteínas (Hernández et al., 1994).
Cuadro 2.- Cuantificación de los mayores complejos lipídicos y esteroles de la membrana
celular de dos diferentes hongos.
Penicillium cyclopium
(µg mg-1 proteína)
Ustilago maydis
(µg mg-1 proteína)
Fosfatidiletanolamida 74.07 ± 3.93 43.92 ± 5.18
Fosfatidilcolina 26.85 ± 0.31 40.08 ± 7.19
Ergosterol 84.37 ± 4.13 73.82 ± 7.00
Tomado de Hernández et al., 1994.
Las proteínas de membrana realizan ciertas actividades y de acuerdo a su función
pueden ser divididas en: estructurales, para unirse al citoesqueleto, receptores de
membrana, para recibir y transducir señales y transportadoras para acarrear iones
y nutrientes. Otra de las funciones importantes de la membrana es la de
compartamentalizar las células originándose así los diferentes tipos de organelos.
Cada uno de éstos se puede identificar por medio de enzimas específicas o
marcadoras.
En la membrana plasmática de los hongos la cantidad de proteínas puede ser
variable. Entre las enzimas más fácilmente reconocidas se encuentra la
H+ATPasa. Existen un gran número de proteínas de membrana en hongos
relacionados tanto con la señalización como con el transporte de electrones. Por
ejemplo la adenilato ciclasa que produce AMPc a partir de ATP es un componente

Efecto del quitosano en la funcionalidad de la membrana plasmática Rhizopus stolonifer INTRODUCCIÓN
10
importante en las cascadas de transducción de señales (Jain et al., 2003), la
oxidasa de NADPH en la mitocondria, oxida al NADP con el oxígeno molecular y
participa en la formación de peróxido de hidrógeno generando especies reactivas
de oxígeno (ROS) (Paraje et al., 2009).
1.5 Las ATPasas
Las ATPasas de membrana son esenciales para la captura de nutrientes como
amino ácidos y azucares, además participan de manera importante en la
regulación del pH interno, hidrolizan el 25% del adenosín-trifosfato (ATP)
producido por la célula y acoplan la energía liberada con el transporte activo de
iones, generando un gradiente iónico que es aprovechado para el transporte de
nutrientes y de otros iones. El ATP es la forma de energía química rápidamente
utilizable, se sintetiza en la glucólisis y en la fosforilación oxidativa, se hidroliza en
procesos como la síntesis de proteínas, ácidos nucleicos y como ya se menciono
en el transporte activo (Bowman et al., 1988; Guerra-Sánchez et al., 1995).
Las ATPasas se distinguen por su composición de subunidades, la presencia o
ausencia de un intermediario fosforilado, por la sensibilidad a inhibidores
específicos, por la estequiometria ión/ATP y por la fisiología de sintetizar o
hidrolizar ATP (Pedersen and Carafoli, 1987). Las ATPasas de membrana de
acuerdo a su composición proteica, localización celular, función y mecanismo de
acción se han clasificado en tres grupos: F-ATPasas (F1/F0 o mitocondriales), V-
ATPasas (vacuolares) y P-ATPasas (fosforilables) (Van Der Rest et al., 1995).
Una de las principales herramientas que más han servido para caracterizar a las
diferentes ATPasas, son los inhibidores específicos de cada tipo de ATPasa
(Cuadro 3).
Efecto del quitosano en la funcionalidad de la membrana plasmática Rhizopus stolonifer INTRODUCCIÓN
11
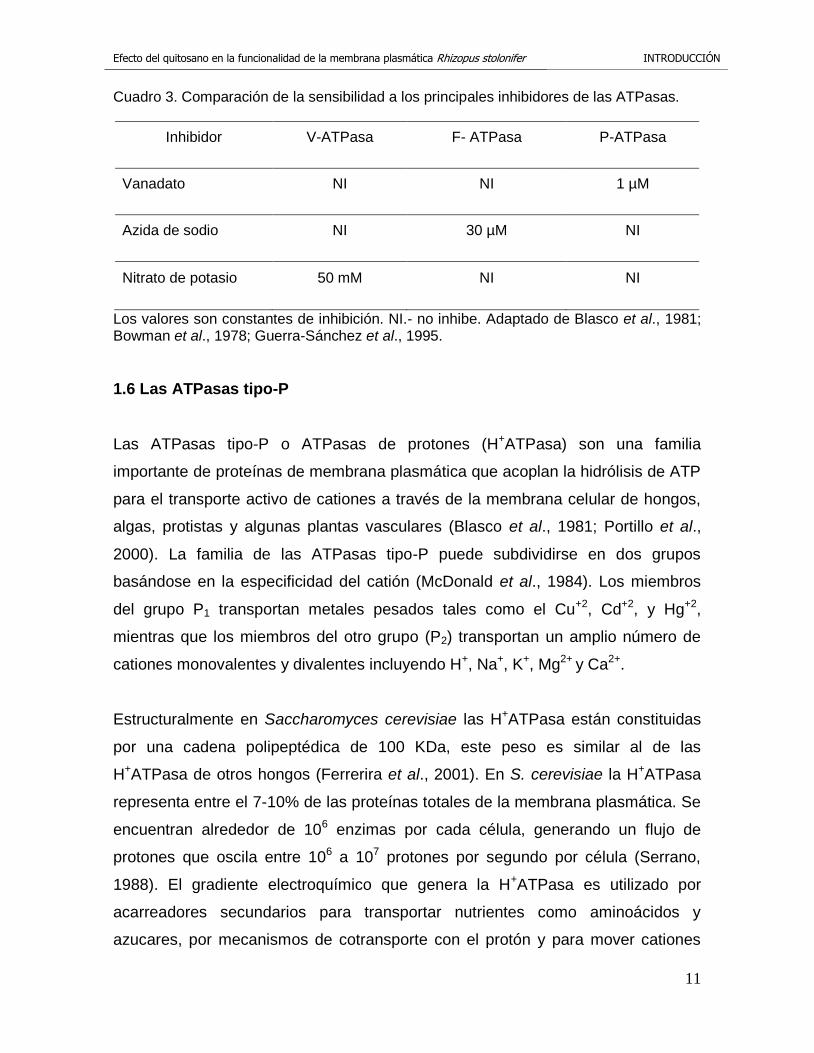
Cuadro 3. Comparación de la sensibilidad a los principales inhibidores de las ATPasas.
Inhibidor V-ATPasa F- ATPasa P-ATPasa
Vanadato NI NI 1 µM
Azida de sodio NI 30 µM NI
Nitrato de potasio 50 mM NI NI
Los valores son constantes de inhibición. NI.- no inhibe. Adaptado de Blasco et al., 1981; Bowman et al., 1978; Guerra-Sánchez et al., 1995.
1.6 Las ATPasas tipo-P
Las ATPasas tipo-P o ATPasas de protones (H+ATPasa) son una familia
importante de proteínas de membrana plasmática que acoplan la hidrólisis de ATP
para el transporte activo de cationes a través de la membrana celular de hongos,
algas, protistas y algunas plantas vasculares (Blasco et al., 1981; Portillo et al.,
2000). La familia de las ATPasas tipo-P puede subdividirse en dos grupos
basándose en la especificidad del catión (McDonald et al., 1984). Los miembros
del grupo P1 transportan metales pesados tales como el Cu+2, Cd+2, y Hg+2,
mientras que los miembros del otro grupo (P2) transportan un amplio número de
cationes monovalentes y divalentes incluyendo H+, Na+, K+, Mg2+ y Ca2+.
Estructuralmente en Saccharomyces cerevisiae las H+ATPasa están constituidas
por una cadena polipeptédica de 100 KDa, este peso es similar al de las
H+ATPasa de otros hongos (Ferrerira et al., 2001). En S. cerevisiae la H+ATPasa
representa entre el 7-10% de las proteínas totales de la membrana plasmática. Se
encuentran alrededor de 106 enzimas por cada célula, generando un flujo de
protones que oscila entre 106 a 107 protones por segundo por célula (Serrano,
1988). El gradiente electroquímico que genera la H+ATPasa es utilizado por
acarreadores secundarios para transportar nutrientes como aminoácidos y
azucares, por mecanismos de cotransporte con el protón y para mover cationes

Efecto del quitosano en la funcionalidad de la membrana plasmática Rhizopus stolonifer INTRODUCCIÓN
12
como Na+ y Ca2+ por antitransporte con el protón (Goffeau and Slayman, 1981).
Además, este gradiente de potencial electroquímico se utiliza en la regulación del
pH citosólico, el balance osmótico, la tolerancia a los metales pesados, etc.
(Weissman, 2000).
Las H+ATPasas están unidas a la membrana plasmática por 8-10 segmentos
transmembranales. La mayor parte de la cadena polipeptídica se encuentra en el
citosol incluyendo los extremos amino y carboxilo terminal y dos asas
citoplasmáticas de gran tamaño (figura 4) (Ferrerira et al., 2001).
Figura 4.- Topología de la H
+ATPasa de la membrana plasmática de S. cerevisiae, basado en
cristalografía de segunda y tercera dimensión. N y C terminales se encuentran en el citoplasma. Las líneas en zig-zag representan las regiones que se consideran estructuras secundaria del tipo α-hélice. Tomado de Ferrerira et al., 2001.
La región amino terminal es la que menos conserva la estructura primaria y
también es la que más varía en el número de residuos de aminoácidos que la
integran, hasta 115 en las ATPasas de protones de los hongos (Serrano, 1988).
Exterior
Interior

Efecto del quitosano en la funcionalidad de la membrana plasmática Rhizopus stolonifer INTRODUCCIÓN
13
Estudios cinéticos detallados han confirmado que las ATPasas presentan dos
estados conformacionales (E1 y E2) durante su ciclo de reacción (Nakamoto et al.,
1989), además de que forman un intermediario fosforilado con el fosfato gamma
del ATP, llamado aspartil-fosfato (Bowman et al., 1978; Hubbard et al., 1986);
Cabe resaltar que el vanadato puede sustituir al fosfato uniéndose a la enzima con
gran afinidad por lo que la ATPasa queda inhibida.
Las H+ATPasas de hongos y plantas se regulan tanto a nivel transcripcional como
a nivel postraduccional (Portillo, 2000). En S. cerevisiae y S. pombe la actividad
enzimática depende de las condiciones de crecimiento (Serrano, 1988). Algunos
factores que incrementan la actividad de la enzima son las fuentes de carbono
fermentables, también existen agentes estresantes como las temperaturas
supraóptimas, los choques térmicos, la disminución de las fuentes de carbono o
de nitrógeno, las altas concentraciones de etanol, de metales, de sales o la
acidificación del medio de cultivo, que se han reportado como activadores de la
enzima (Eraso and Gancedo, 1987; Benito et al., 1992; Hernández et al., 2002).

Efecto del quitosano en la funcionalidad de la membrana plasmática Rhizopus stolonifer ANTECEDENTES
14
ANTECEDENTES
El efecto antifúngico del quitosano en el desarrollo in vitro de Rhizopus stolonifer
ha sido demostrado mediante la inhibición del crecimiento micelial, la esporulación
y germinación de este fitopatógeno. En ese mismo trabajo se demostró que
quitosanos de diferentes pesos moleculares afectan la forma y la ornamentación
de las esporas de R. stolonifer (Hernández-Lauzardo et al., 2008).
Investigaciones previas han demostrado que la inhibición de crecimiento que
genera el quitosano en contra de R. stolonifer depende del medio de cultivo,
siendo mayor en medios ricos como PDA, además de que el fenómeno es
proporcional al aumento en la concentración que se utiliza (datos no publicados).
Otros estudios realizados con R. stolonifer en medio mínimo líquido evidencian
que el quitosano incrementa la liberación de proteínas y de compuestos que
absorben a 260 y 280 nm, aumenta el consumo de glucosa y la actividad
hexocinasa alterando el metabolismo celular (Guerra-Sánchez et al., 2009).
Investigaciones recientes indican que el quitosano afecta a R. stolonifer
incrementado el consumo de oxigeno, lo cual está relacionado con el aumento en
el numero de mitocondrias que presentan de forma normal las esporas. Otros
cambios que se identificaron en este trabajo fueron que hay un aumento en las
actividades de la isocitrato deshidrogenasa, la citocromo c oxidasa y la ATP
sintetasa, componentes importantes de la cadena respiratoria mitocondrial, por lo
que también se ve modificada (datos no publicados).

Efecto del quitosano en la funcionalidad de la membrana plasmática Rhizopus stolonifer JUSTIFICACIÓN
15
JUSTIFICACIÓN
Los hongos fitopatógenos causan pérdidas cuantiosas durante el transporte y
almacenamiento de los productos hortofrutícolas. El uso de compuestos
antifúngicos biodegradables y no tóxicos como el quitosano representa una
alternativa natural para extender la vida de anaquel de estos productos, sin causar
daños al ambiente y a la salud humana. Se ha demostrado que el quitosano afecta
e inhibe el desarrollo de diferentes hongos fitopatógenos, entre ellos Rhizopus
stolonifer. El mecanismo de acción de este polímero no ha sido del todo elucidado.
La propuesta más apoyada es la que involucra el carácter policatiónico de la
molécula, que provoca alteraciones en la permeabilidad de la membrana
plasmática generando la salida de componentes del interior de la célula. Sin
embargo, son escasos los estudios que involucran el efecto del quitosano en la
estructura y función de los principales componentes de la membrana. Por lo
anterior, en esta investigación se propone estudiar el efecto del quitosano en la
funcionalidad de la membrana plasmática de R. stolonifer.

Efecto del quitosano en la funcionalidad de la membrana plasmática Rhizopus stolonifer HIPÓTESIS
16
HIPÓTESIS
El quitosano podría afectar la funcionalidad de la membrana plasmática de
Rhizopus stolonifer provocando cambios en la concentración de fosfolípidos y
proteínas de la membrana y en la actividad de la H+ATPasa.

Efecto del quitosano en la funcionalidad de la membrana plasmática Rhizopus stolonifer OBJETIVOS
17
OBJETIVOS
OBJETIVO GENERAL
Estudiar el efecto del quitosano en la funcionalidad de la membrana plasmática de
Rhizopus stolonifer.
OBJETIVOS PARTICULARES
Evaluar el efecto del quitosano en la integridad de la membrana plasmática en
esporas de R. stolonifer.
Determinar los cambios en el pH del medio, potencial de membrana y salida de
potasio por efecto del quitosano.
Comparar la concentración de fosfolípidos totales de la membrana plasmática en
ausencia y presencia de quitosano.
Cuantificar el contenido de proteínas en la membrana plasmática del hongo
cultivado con y sin quitosano.
Determinar los cambios en la actividad de la H+ATPasa de la membrana
plasmática por la presencia de quitosano.

Efecto del quitosano en la funcionalidad de la membrana plasmática Rhizopus stolonifer MATERIAL Y MÉTODOS
18
MATERIAL Y MÉTODOS
2.1 Muestra biológica
Se utilizó un aislado de Rhizopus stolonifer (R3) la cual fue obtenida de frutos de
jitomate (Lycopersicon esculentum Mill) del tipo “Saladette” en el Estado de
Morelos (Hernández-Lauzardo et al., 2006).
2.2 Solución de quitosano
Se preparó una solución concentrada de quitosano [10 mg mL-1] de bajo peso
molecular (Sigma-Aldrich, St. Louis, MO, USA) pesando 2 g de quitosano en una
balanza analítica, se disolvió en 100 mL de agua destilada más 2 mL de ácido
acético y se dejó homogenizar por agitación constante durante 24 h. Transcurrido
el tiempo se ajustó el pH en un potenciómetro a 5.6 adicionando NaOH al 1 N. Se
filtró la solución con una gasa de algodón y se aforó a 200 mL con agua destilada.
Se esterilizó en autoclave a 121ºC durante 15 min. A partir de la solución madre
de quitosano se obtuvo la concentración evaluada [2 mg mL-1] (Guerra et al.,
2009).
2.3 Medios de cultivo
Se empleó agar papa dextrosa (PDA) (Bioxon) (anexo) para mantener y propagar
el aislado utilizado en este estudio. El medio mínimo (MM) líquido ajustado a pH
5.6 se usó para determinar el efecto del quitosano sobre el crecimiento de R.
stolonifer (Guerra et al., 2009).
Medio mínimo: glucosa 10 g L-1; NH4NO3 3 g L-1; solución de sales y elementos trazas 62.5 mL L-1.
Solución de sales: KH2PO4 16 g L-1; Na2SO4 4 g L-1; KCl 8 g L-1; MgSO4 2 g L-1; CaCl2 1 g L-1; Elementos traza 8 mL L-1.
Elementos trazas: H3BO3 60 mg L-1; MnCl2-4H2O 140 mg L-1; ZnCl2 400 mg L-1; Na2MoO4-2H2O 40 mg L-1; FeCl3-6H2O 100 mg L-1; CuSO4-5H2O 400 mg L-1.

Efecto del quitosano en la funcionalidad de la membrana plasmática Rhizopus stolonifer MATERIAL Y MÉTODOS
19
2.4 Condiciones de cultivo y obtención del inóculo fúngico
El aislado de Rhizopus stolonifer (R3) se cultivó en medio PDA a 25 ± 2 °C
durante 96 h. Para obtener el inóculo se tomaron discos de 5 mm de diámetro o se
prepararon suspensiones de esporas (1X106 esporas mL-1) (anexo) según fuera el
caso.
2.5 Cinética de germinación
Se inoculó 1 mL de las suspensiones de esporas (1X106 esporas mL-1) del aislado
R3 en 100 mL de MM. Se incubaron durante 0, 3, 4, 5 y 6 h en agitación constante
(140 rpm) a 25 ± 2 °C de forma independiente. Las células se observaron en un
microscopio óptico a 40X. Se observaron varios campos al azar y se contaron 100
esporas. Se consideró como espora germinada en el momento que su tubo
germinal fuera igual o mayor al diámetro de la espora (El Ghaouth et al., 1992).
2.6 Variación del pH durante el crecimiento de Rhizopus stolonifer
Se inoculó 1 mL de las suspensiones de esporas (1X106 esporas mL-1) del aislado
R3 en 100 mL de MM en presencia de quitosano [2 mg mL-1]. Los testigos no
contenían quitosano. Se incubaron durante 0, 3, 6 y 12 h en agitación constante
140 rpm a 25 ± 2 °C de forma independiente. Se colectó el medio de cultivo y se
centrifugó a 8000 rpm, para eliminar las esporas y se determinó el pH del medio
de cultivo, en un potenciómetro previamente calibrado con dos estándares de pH
(3 y 7).
2.7 Determinación de la integridad de la membrana de las esporas y micelio
Se inoculó 1 mL de las suspensiones de esporas (1X106 esporas mL-1) del aislado
R3 en MM en presencia de quitosano [2 mg mL-1]. Los testigos no contenían
quitosano. Se incubaron durante 4, 5 y 6 h en agitación constante (140 rpm) a 25
± 2 °C de forma independiente. Se recolectaron las esporas y se centrifugó a 8000
rpm, se lavó con un regulador de fosfatos (50 mM Na2HPO4) pH 7.0 para eliminar

Efecto del quitosano en la funcionalidad de la membrana plasmática Rhizopus stolonifer MATERIAL Y MÉTODOS
20
residuos del medio de cultivo. A las esporas se les adicionó ioduro de propidio a
10 µg mL-1, se incubó durante 5 min a 25 ± 2°C. Las esporas se lavaron con el
mismo regulador para eliminar restos del colorante. Las células se observaron en
un microscopio de fluorescencia (40X). Se observaron 3 campos aleatorios y se
contaron el total de esporas y la cantidad de esporas teñidas (Liu et al., 2007).
La integridad de la membrana se calculó con la siguiente fórmula:
MI (%) = No. de esporas totales – No. de esporas teñidas × 100
No. de esporas totales
2.8 Potencial de la membrana plasmática
Se inoculó 10 mL de las suspensiones de esporas (1X106 esporas mL-1) del
aislado R3 en 100 mL de MM en presencia de quitosano [2 mg mL-1]. Los testigos
no contenían quitosano. Se incubaron durante 6 h en agitación constante (140
rpm) a 25 ± 2 °C. Las esporas se lavaron en 2 ocasiones con agua destilada
estéril, al final las esporas se resuspendieron en agua destilada. El potencial de la
membrana plasmática de las esporas de R. stolonifer se estableció siguiendo la
fluorescencia del DiSC3(3) (Peña et al., 1984). La determinación se realizó en 1.8
mL de regulador (MES 10 mM pH 6.0/TEA), 0.02 mL (glucosa 1 M), 0.05 mL
(CaCl2 10 mM), 0.002 mL (CCCP 10 mM). El trazo se inició por la adición de
0.01mL (1X106 esporas) de una suspensión de esporas germinadas.
Posteriormente, a los 50 segundos de trazo se añadió 0.005 mL de cianina 0.1
mM, por último se adicionó 0.03 mL de KCl2 2 M a los 300 segundos. La
fluorescencia se registró a 540-590 nm.
2.9 Salida de Potasio
Se inoculó 1 mL de las suspensiones de esporas (1X106 esporas mL-1) del aislado
R3 en 100 mL de MM. Se incubaron durante 12 horas a 25 ± 2°C en agitación
constante (150 rpm). El micelio se lavo 2 veces con agua destilada estéril, al final

Efecto del quitosano en la funcionalidad de la membrana plasmática Rhizopus stolonifer MATERIAL Y MÉTODOS
21
el micelio se resuspendió en agua destilada (mL g-1). En la cámara de ensayo se
colocó un electrodo selectivo de iones potasio, 3.95 mL de regulador (MES 2 mM
pH 6.6/Tris, glucosa 10 mM) y 0.05 mL de la suspensión celular. El trazo se inició
y se dejó que se estabilizara, después de 40 segundos se adicionó 1 mL de
quitosano de una solución concentrada [10 mg mL-1] para obtener una
concentración final de 2 mg mL-1. En el caso del testigo, se inició el trazo y nunca
se adicionó quitosano, se dejo trascurrir el mismo tiempo que en el otro ensayo.
Las lecturas se registraron en una computadora en donde se observó la salida del
ion potasio.
2.10 Obtención de membranas plasmáticas de R. stolonifer
Suspensiones de esporas de 1 mL del aislado R3 fueron inoculadas en matraces
con 100 mL de MM con quitosano [2 mg mL-1]. Los testigos no contenían
quitosano. Se incubaron a 25 ± 2 °C en agitación constante (140 rpm) durante 12
h. Se colectó la biomasa y se sometió a una digestión con enzimas líticas durante
3h a 30 °C. El rompimiento mecánico se realizó en presencia de inhibidores de
proteasas y estabilizadores de proteína para asegurar la integridad de la
membrana (1 M Sorbitol pH 7.4/Tris base, 50 mM KH2PO4, 1 mM EDTA, 1mM
PMSF, 5 mM DTT). El homogenado se centrifugó a 4000 rpm a 4 °C durante 5 min
y el extracto crudo se sometió a una centrifugación diferencial, se centrifugó a
8000 rpm a 4 °C durante 15 min y se continuó con una ultracentrifugación a 35000
rpm a 4 °C durante 1 h, para obtener una fracción de membranas plasmáticas
crudas, las cuales se resuspendieron en 0.5 mL de regulador (1 mM EGTA pH
7.2/Tris base). Para la purificación de las membranas plasmáticas crudas se utilizó
un gradiente discontinuo de glicerol (80, 70, 55 y 40%), preparado en un regulador
(10mM Tris pH 7, 1mM EDTA, 0.5% Azolectina) y se centrifugó a 35000 rpm a 4
°C durante 3 h. Se recuperaron las bandas y se lavaron utilizando el regulador de
EGTA. El marcador de membranas plasmáticas que se utilizó fue la actividad de la
H+ ATPasa.

Efecto del quitosano en la funcionalidad de la membrana plasmática Rhizopus stolonifer MATERIAL Y MÉTODOS
22
2.11 Cuantificación de fosfolípidos en la membrana plasmática.
La cuantificación de los fosfolípidos presentes en las membranas plasmáticas
comenzó con la extracción de los fosfolípidos en la cual se utilizó con una mezcla
de cloroformo/etanol (2:1), con la que re realizaron tres lavados, se recuperó la
fase orgánica de cada uno de los lavados en un solo vial, después el total de la
fase orgánica se sometió a una evaporación con nitrógeno hasta sequedad (Folch
et al., 1957). Posteriormente el residuo se resuspendió en metanol y se cuantificó
el fosforo total presente en los fosfolípidos utilizando el método de Fiske–
Subbarow modificado (Dryer et al., 1957).
2.12 Observación de las membranas plasmáticas en el microscopio
electrónico.
La observación en el microscopio electrónico de transmisión (Transmission
Electron Microscope JEM-1010), se realizó utilizando las membranas plasmáticas
purificadas, se colocó aproximadamente 20-25 µg de proteína de membrana en
unas rejillas de cobre que contenían una membrana de Formvar como soporte, se
incubaron las rejillas durante 5 min a 25 ± 2°C en ambiente húmedo.
Posteriormente se agregó ácido fosfotúngstico al 0.5% y se incubaron las rejillas
durante 30 seg, se retiró la solución del ácido y se dejaron secar la muestras
durante 90 min. Para posteriormente observarse al microscopio electrónico de
transmisión
2.13 Concentración de proteínas presentes en las membranas plasmáticas
La concentración de proteínas se determinó por el método de Lowry (Kit-comercial
500-0114 BioRad) (Lowry et al., 1951), utilizando albúmina sérica bovina (BSA)
como proteína estándar. Para determinar la concentración de proteínas se
construyó una curva tipo con una solución concentrada de BSA (0.5 mg mL-1).
Para elaborar la curva tipo (anexos) diferentes volúmenes de la solución de BSA y
los reactivos A y B del kit comercial fueron mezclados en un Vortex y se incubaron
a 25 ± 2 °C durante 10 min. Para realizar la lectura se utilizó un espectrofotómetro

Efecto del quitosano en la funcionalidad de la membrana plasmática Rhizopus stolonifer MATERIAL Y MÉTODOS
23
a una longitud de onda de 655 nm, se ajustó a 0 con el blanco de reactivos.
Posteriormente, se interpolaron las lecturas de absorbencia obtenidas para cada
muestra en la curva tipo y se convirtieron las absorbencias en g L-1. La curva
tipo se realizó por duplicado para aumentar el grado de confianza.
Para cuantificar las proteínas presentes en las muestras de membranas se utilizó
0.002 mL de cada una de las muestras en diferentes tubos. Se adicionaron los
reactivos A y B del kit comercial y se mezclaron en Vortex, se dejaron incubando
durante 10 min. Se leyeron en un espectrofotómetro a 655 nm, donde se ajustó a
0 con el blanco de reactivos. Finalmente, se realizaron los cálculos para convertir
las concentraciones en mg mL-1.
2.14 Actividad de H+ ATPasa
La actividad de la enzima se realizó utilizando membranas plasmáticas purificadas
en presencia de azida de sodio para inactivar a las ATPasas mitocondriales. La
mezcla de reacción constaba de 5 mM ATP, 5 mM MgCl2, 10mM PIPES, 5 mM
Azida de sodio, 5 mM PEP, 0.005mL de Piruvato cinasa (por cada 1mL de mezcla
de reacción que se preparó), se ajustó a pH 6.7/Tris base y 28 – 30 µg de proteína
de membrana. La reacción se inició con la adición de la proteína, se incubó por 10
min a 30 °C y se detuvo con 0.01 mL de TCA 30% en un baño de hielo para evitar
la hidrólisis del ATP residual. El fosfato liberado por la enzima se midió utilizando
el método de Fiske–Subbarow modificado (Dryer et al., 1957), en el que el Pi en el
medio reacciona con el molibdato de amonio formando un complejo fosfo-
molibdato de amonio, produciendo el complejo azul de molibdeno, que absorbe a
660 nm. Se utilizó fosfato de sodio monobásico (NaH2PO4•H20) como estándar (1
µmol Pi/0.5 mL).Para elaborar la curva tipo diferentes volúmenes de una solución
de fosfato de sodio monobásico y los reactivos se mezclaron en Vortex y se
incubaron en hielo durante 10 min. Para realizar la lectura se utilizó un
espectrofotómetro a una longitud de onda de 660 nm, se ajustó a 0 con blanco de
reactivos. Posteriormente, se interpolaron las lecturas de absorbencia obtenidas
para cada muestra en la curva tipo y se convirtieron las absorbencias en nmolPi
L-1. La curva tipo se realizó por duplicado para aumentar el grado de confianza.

Efecto del quitosano en la funcionalidad de la membrana plasmática Rhizopus stolonifer MATERIAL Y MÉTODOS
24
2.15 Cinética enzimática de la actividad de H+ ATPasa
La cinética enzimática se realizó utilizando membranas plasmáticas purificadas.
Se utilizó una solución madre de ATP-Mg a concentración de 24 mM, el cual se
varió dependiendo del ensayo, también se empleó un regulador de reacción (100
mM MES pH 5.7/Tris base, 10 mM MgCl2, 10 mM PEP y 10 mM NaN3) y 28 – 30
µg de proteína de membrana. La reacción se inició con la adición de la enzima, se
incubó por 10 min a 30 °C y se detuvo con 0.01 mL de TCA 30% en un baño de
hielo. El fosfato liberado por la enzima se medió utilizando el método de Fiske–
Subbarow modificado (Dryer et al., 1957).
2.16 Diseño de experimentos y análisis estadístico
Los experimentos se realizaron mediante un diseño completamente al azar. Los
datos se procesaron mediante un análisis de varianza de una vía con el paquete
estadístico Sigma Stat versión 2.0.

Efecto del quitosano en la funcionalidad de la membrana plasmática Rhizopus stolonifer RESULTADOS
25
RESULTADOS
3.1 Cinética de germinación
Los resultados obtenidos para el aislado R3 se muestran en la figura 5. Se
observa que existe un periodo en el que las esporas no germinan (0 a 3 h). A las
4 h inició la germinación, observándose un 15 % de esporas germinadas, después
a las 5 h este porcentaje aumentó hasta un 70 %. Posteriormente, a las 6 h se
evidenció un porcentaje de germinación del 88 %. Este porcentaje se incrementó
cuando se realizó la cuantificación a las 7 h, mostrándose un máximo del 91 % de
germinación.
Figura 5.- Cinética de germinación. Porcentaje de germinación de las esporas Rhizopus stolonifer cultivado en medio mínimo con agitación constante a temperatura de 25 ± 2 °C. La barra representa la desviación estándar de 3 repeticiones.

Efecto del quitosano en la funcionalidad de la membrana plasmática Rhizopus stolonifer RESULTADOS
26
En la figura 6 se observa la germinación de esporas de R. stolonifer, la elongación
del tubo germinal (4, 5 y 6 h) y la ramificación del micelio después de 7 h de
incubación en medio mínimo.
Figura 6.- Microfotografía de la cinética de germinación. Germinación de esporas de Rhizopus stolonifer cultivadas en medio mínimo durante 7 h en agitación constante a temperatura de 25 ± 2 °C. 400X.
3.2 Variación del pH durante el crecimiento de Rhizopus stolonifer
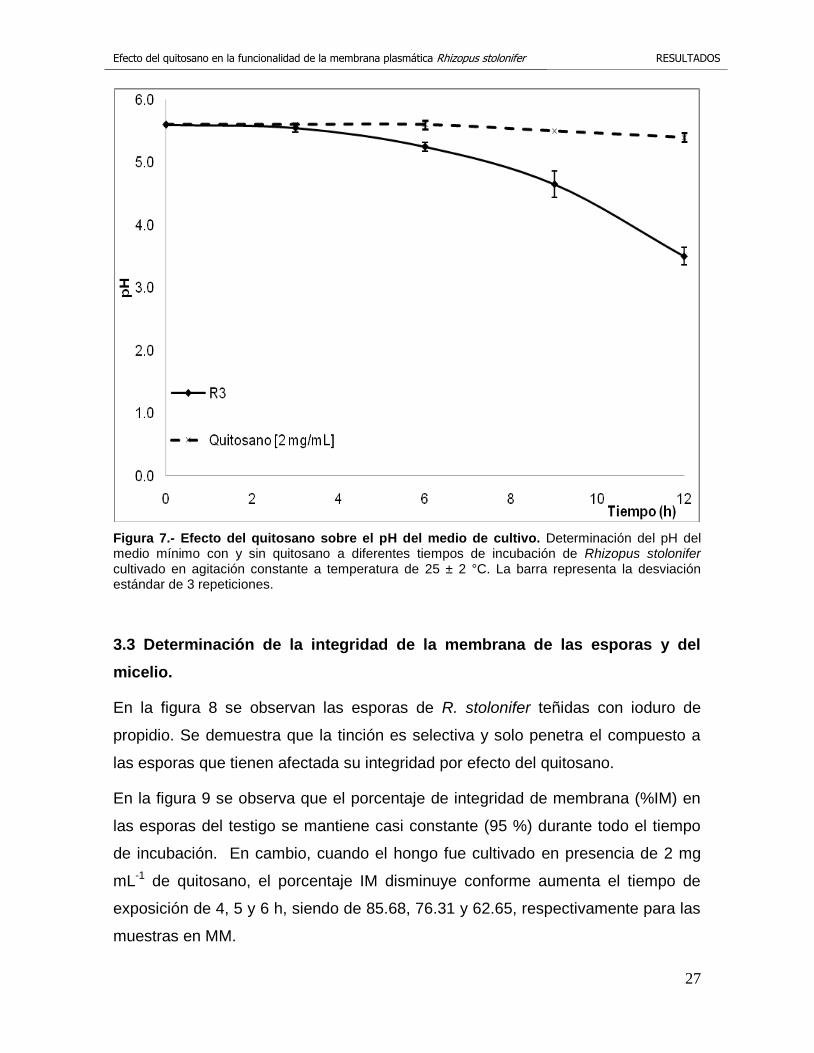
Los resultados obtenidos con el aislado R3 se muestran en la figura 7. Se observa
que el testigo provocó una acidificación gradual del medio de cultivo durante el
periodo de incubación. Cuando se cultivó al hongo en presencia de 2 mg mL-1 de
quitosano se observó que no hay una acidificación del medio, a las 12 h de
incubación el pH disminuyó a 5.4, prácticamente se mantiene el pH original de 5.6.
En cambio, en el tratamiento testigo el pH disminuyó a partir de las 6 h de
incubación obteniéndose un pH de 5.3. A las 9 h de incubación el pH disminuyó a
4.7 y a las 12 h de incubación el valor de pH fue de 3.5.
Efecto del quitosano en la funcionalidad de la membrana plasmática Rhizopus stolonifer RESULTADOS
27
Figura 7.- Efecto del quitosano sobre el pH del medio de cultivo. Determinación del pH del medio mínimo con y sin quitosano a diferentes tiempos de incubación de Rhizopus stolonifer cultivado en agitación constante a temperatura de 25 ± 2 °C. La barra representa la desviación estándar de 3 repeticiones.
3.3 Determinación de la integridad de la membrana de las esporas y del
micelio.
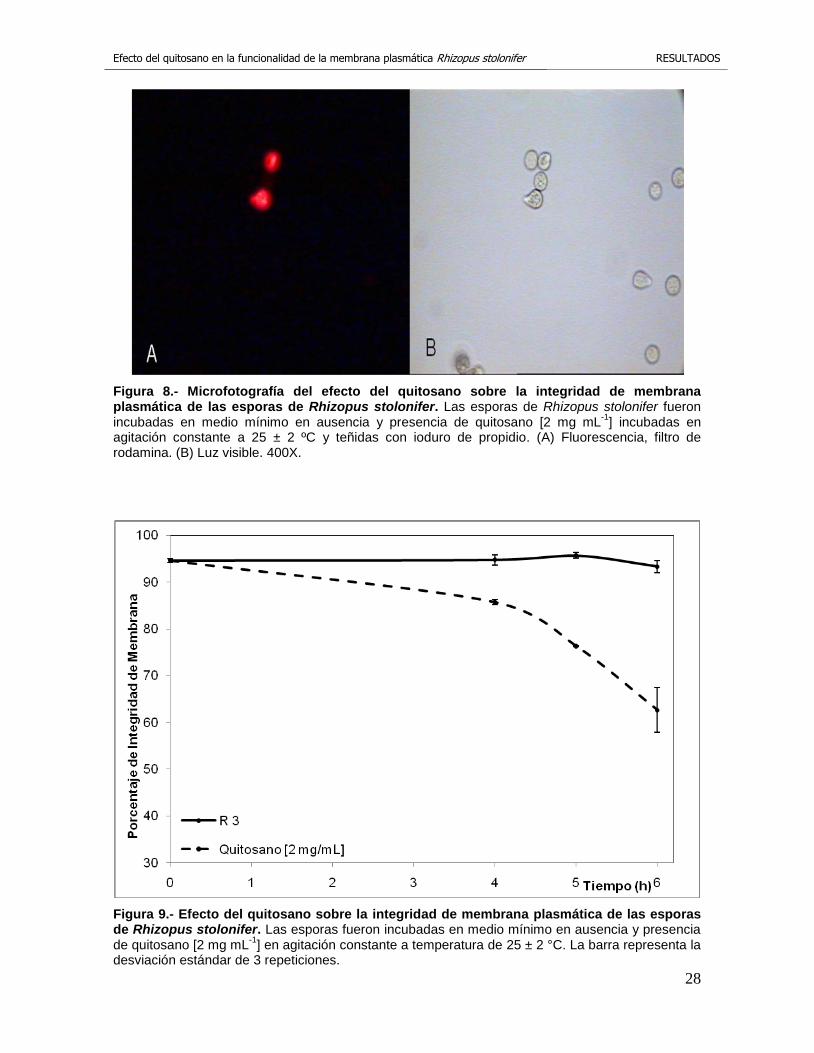
En la figura 8 se observan las esporas de R. stolonifer teñidas con ioduro de
propidio. Se demuestra que la tinción es selectiva y solo penetra el compuesto a
las esporas que tienen afectada su integridad por efecto del quitosano.
En la figura 9 se observa que el porcentaje de integridad de membrana (%IM) en
las esporas del testigo se mantiene casi constante (95 %) durante todo el tiempo
de incubación. En cambio, cuando el hongo fue cultivado en presencia de 2 mg
mL-1 de quitosano, el porcentaje IM disminuye conforme aumenta el tiempo de
exposición de 4, 5 y 6 h, siendo de 85.68, 76.31 y 62.65, respectivamente para las
muestras en MM.
Efecto del quitosano en la funcionalidad de la membrana plasmática Rhizopus stolonifer RESULTADOS
28
Figura 8.- Microfotografía del efecto del quitosano sobre la integridad de membrana plasmática de las esporas de Rhizopus stolonifer. Las esporas de Rhizopus stolonifer fueron incubadas en medio mínimo en ausencia y presencia de quitosano [2 mg mL
-1] incubadas en
agitación constante a 25 ± 2 ºC y teñidas con ioduro de propidio. (A) Fluorescencia, filtro de rodamina. (B) Luz visible. 400X.
Figura 9.- Efecto del quitosano sobre la integridad de membrana plasmática de las esporas de Rhizopus stolonifer. Las esporas fueron incubadas en medio mínimo en ausencia y presencia de quitosano [2 mg mL
-1] en agitación constante a temperatura de 25 ± 2 °C. La barra representa la
desviación estándar de 3 repeticiones.

Efecto del quitosano en la funcionalidad de la membrana plasmática Rhizopus stolonifer RESULTADOS
29
En la figura 10 se observa el micelio de R. stolonifer teñido con ioduro de propidio.
Esta figura demuestra que la penetración del colorante al micelio es selectiva y
que este efecto se mantiene por lo menos hasta las 12 horas de incubación.
Figura 10.- Micrografía del efecto del quitosano sobre la integridad de la membrana plasmática del micelio de Rhizopus stolonifer. El micelio de Rhizopus stolonifer crecido en presencia (A y B) y ausencia (C y D) de quitosano [2 mg mL
-1] durante 12 h en medio mínimo con
agitación constante a 25 ± 2 ºC teñido con ioduro de propidio. (A y C) Fluorescencia, filtro de rodamina. (B y D) Luz visible. 400X.

Efecto del quitosano en la funcionalidad de la membrana plasmática Rhizopus stolonifer RESULTADOS
30
3.4 Potencial de la membrana plasmática de esporas de R. stolonifer
En la figura 11 se observan los cambios de fluorescencia de DiSC3(3) de las
esporas incubadas en presencia y ausencia de quitosano. En los resultados
obtenidos con el aislado R3 testigo se observó un aumento en la fluorescencia, lo
que indica la existencia de un potencial de membrana plasmática producido por la
H+ATPasa.
En cambio, las esporas tratadas con quitosano mostraron un comportamiento
diferente al de las esporas testigo. Se observó un incremento en la fluorescencia,
aunque de menor intensidad, también se comprobó la existencia del potencial
adicionando KCl y se observó como disminuye la fluorescencia aunque en menor
proporción comparado con el testigo.
Figura 11.- Efecto del quitosano sobre el potencial de la membrana plasmática de las esporas de Rhizopus stolonifer. Las esporas de Rhizopus stolonifer fueron incubadas en medio mínimo durante 6 h en ausencia y presencia de quitosano [2 mg mL
-1] en agitación constante a 25
± 2 ºC. El trazo representa el promedio de 3 repeticiones.
Efecto del quitosano en la funcionalidad de la membrana plasmática Rhizopus stolonifer RESULTADOS
31
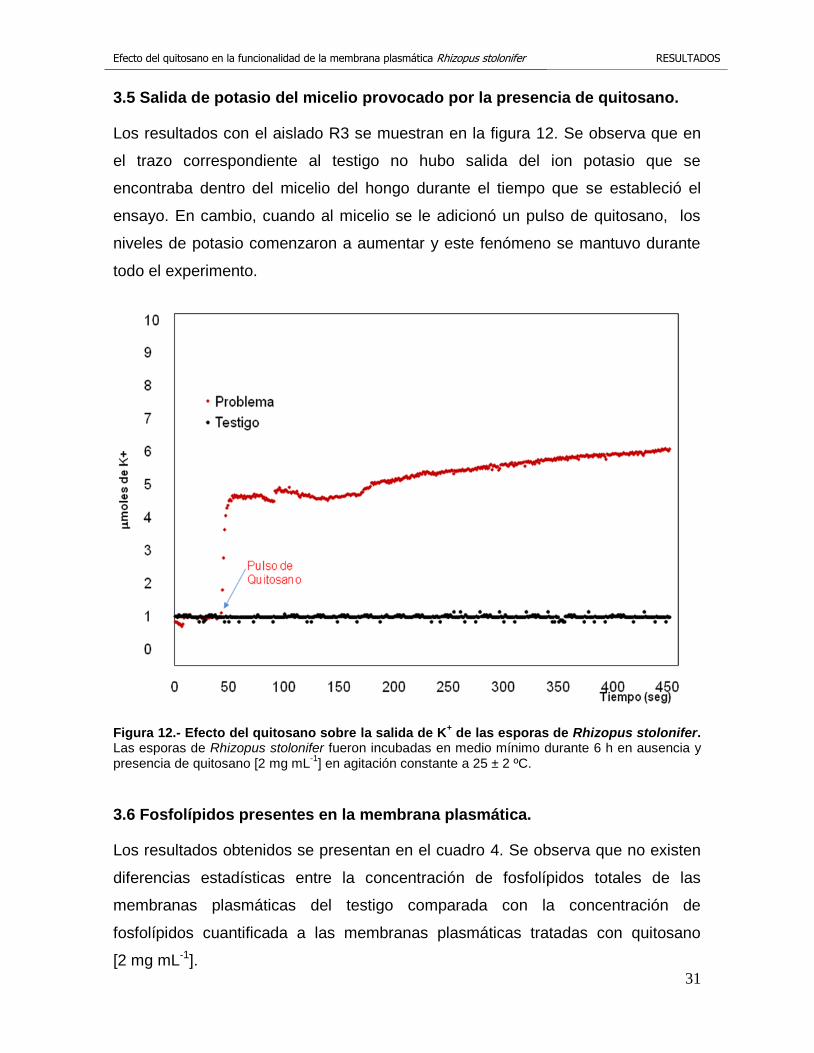
3.5 Salida de potasio del micelio provocado por la presencia de quitosano.
Los resultados con el aislado R3 se muestran en la figura 12. Se observa que en
el trazo correspondiente al testigo no hubo salida del ion potasio que se
encontraba dentro del micelio del hongo durante el tiempo que se estableció el
ensayo. En cambio, cuando al micelio se le adicionó un pulso de quitosano, los
niveles de potasio comenzaron a aumentar y este fenómeno se mantuvo durante
todo el experimento.
Figura 12.- Efecto del quitosano sobre la salida de K+ de las esporas de Rhizopus stolonifer.
Las esporas de Rhizopus stolonifer fueron incubadas en medio mínimo durante 6 h en ausencia y presencia de quitosano [2 mg mL
-1] en agitación constante a 25 ± 2 ºC.
3.6 Fosfolípidos presentes en la membrana plasmática.
Los resultados obtenidos se presentan en el cuadro 4. Se observa que no existen
diferencias estadísticas entre la concentración de fosfolípidos totales de las
membranas plasmáticas del testigo comparada con la concentración de
fosfolípidos cuantificada a las membranas plasmáticas tratadas con quitosano
[2 mg mL-1].

Efecto del quitosano en la funcionalidad de la membrana plasmática Rhizopus stolonifer RESULTADOS
32
Cuadro 4.- Concentración de fosfolípidos calculada a las diferentes muestras de
membranas plasmáticas de Rhizopus stolonifer crecido en medio mínimo durante 12 h.
Muestra Concentración de Fosfolípidos (µmolPi mg proteína -1)*
Membranas sin quitosano 1.111 ±0.520 a
Membranas con quitosano 1.185 ±0.635 a
*Los valores están seguidos por la desviación estándar de la media de 3 repeticiones. Letras diferentes indican diferencias estadísticas significativas de acuerdo a la prueba de rangos múltiples de Tukey p< 0.05
3.7 Observación de membranas plasmáticas en microscopio electrónico de
transmisión.
Se observó en el microscopio electrónico de transmisión las membranas
plasmáticas utilizando una tinción negativa, lo que confirma que nuestro método
de obtención de membranas es efectivo (figura 13).
Figura 13.- Microfotografía electrónica de transmisión de las membranas plasmáticas (MP) de R. stolonifer. El hongo fue incubado en medio mínimo durante 12 h en (a) ausencia y (b) presencia de quitosano [2 mg mL
-1] en agitación constante a 25 ± 2 ºC.

Efecto del quitosano en la funcionalidad de la membrana plasmática Rhizopus stolonifer RESULTADOS
33
3.8 Proteínas presentes en las membranas plasmáticas.
Los resultados obtenidos con el aislado R3 se exponen en el cuadro 5. El testigo
presentó una concentración de proteína mayor (1.608 mg mL-1) comparado con la
concentración de proteínas cuantificada en las diferentes muestras del problema
(0.980 mg mL-1). Lo que indica que existe un 49 % menos de la proteína que
normalmente se presenta en la membrana plasmática del hongo.
Cuadro 5.- Concentración de proteínas cuantificadas a las diferentes muestras obtenidas
de Rhizopus stolonifer crecido en medio mínimo durante 12 h.
Muestra Concentración de Proteínas
(mg mL-1)*
Membranas sin quitosano 1.608 ±0.274 a
Membranas con quitosano 0.980 ±0.364 b
*Los valores están seguidos por la desviación estándar de la media de 4 repeticiones. Letras diferentes indican diferencias estadísticas significativas de acuerdo a la prueba de rangos múltiples de Tukey p< 0.05
3.9 Actividad de H+ATPasa.
Los resultados obtenidos se muestran en el Cuadro 6. Estos resultados indican
que la actividad de la enzima H+ATPasa es superior en el testigo comparado con
las actividades de los tratamientos con quitosano [2 mg mL-1]. Cabe resaltar, que
en las muestras de membrana que estuvieron en contacto con el polímero, la
actividad específica (0.479 μmolPi min-1 mg-1) se redujo aproximadamente en un
25 % comparado con el testigo (0.636 μmolPi min-1 mg-1) y la actividad total de los
membranas plasmáticas problemas (0.247 μmolPi min-1 mL-1) también se redujo
en un 50 % aproximadamente, comparado con la actividad total cuantificada a las
membranas plasmáticas testigo (0.514 μmolPi min-1 mL-1).

Efecto del quitosano en la funcionalidad de la membrana plasmática Rhizopus stolonifer RESULTADOS
34
Cuadro 6.- Actividad de la H+ATPasa calculada a las diferentes muestras de membranas
plasmáticas de Rhizopus stolonifer crecido en medio mínimo durante 12 h.
Muestra Actividad específica
(μmolPi min-1 mg-1)*
Actividad total
(μmolPi min-1 mL-1)*
Membranas sin quitosano 0.636 ±0.063 a 0.514 ±0.112 a
Membranas con quitosano 0.479 ±0.095 b 0.247 ±0.120 b
*Los valores están seguidos por la desviación estándar de la media de 4 repeticiones. Letras diferentes indican diferencias estadísticas significativas de acuerdo a la prueba de rangos múltiples de Tukey p< 0.05
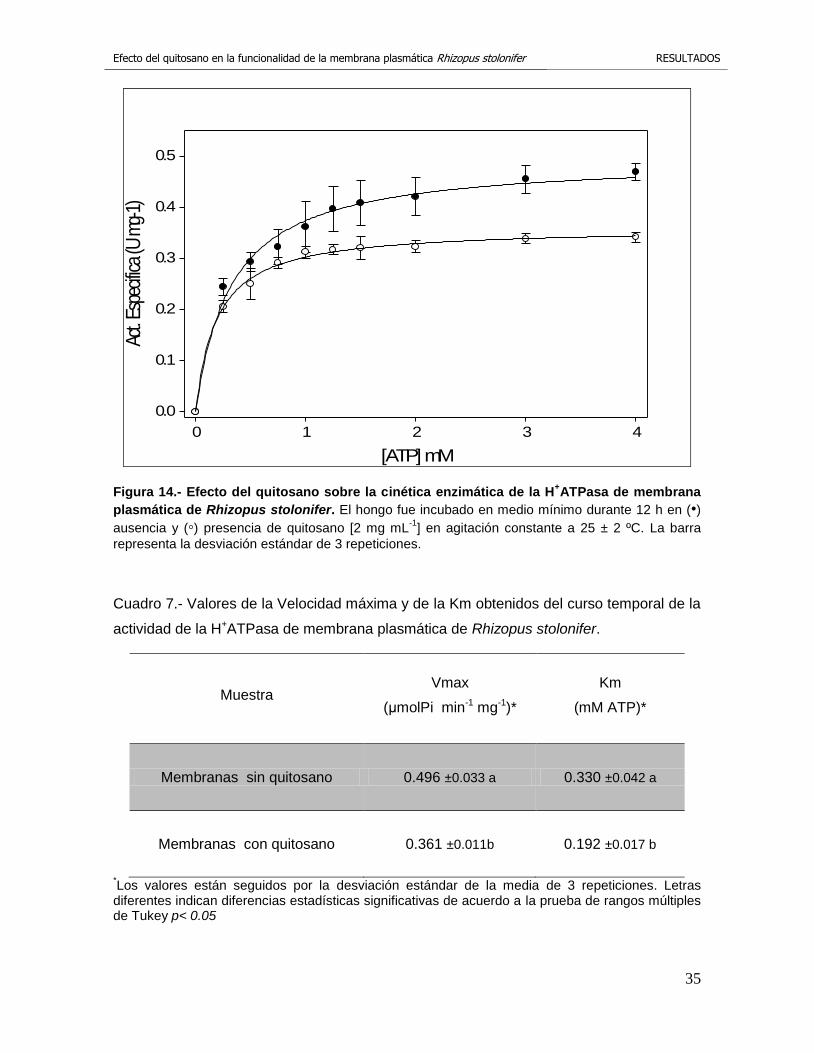
3.10 Cinética enzimática de la H+ATPasa de la membrana plasmática.
En la figura 14 se muestran los resultados obtenidos con el aislado R3 donde se
comparan las dos cinética enzimática de las diferentes H+ATPasa, con y sin
tratamiento de quitosano [2 mg mL-1]. Se puede observar que cuando se utilizaron
las membranas testigo la cinética alcanzo una Vmax de 0.496 μmolPi min-1 mg-1,
en cambio, cuando se utilizaron las membranas tratadas con quitosano la Vmax
disminuyó a 0.361μmolPi min-1 mg-1 (Cuadro 7), lo que indica que existe una
reducción del 25 % de la actividad específica de la H+ATPasa, provocada por la
presencia del quitosano. Un fenómeno similar ocurrió con la Km, las membranas
plasmáticas testigo presentaron un valor de Km de 0.330 mM ATP, sin embargo,
las membranas plasmáticas problemas presentaron un valor de Km de 0.192 mM
ATP (Cuadro 7), lo que refleja una reducción del 42 % aproximadamente.
Efecto del quitosano en la funcionalidad de la membrana plasmática Rhizopus stolonifer RESULTADOS
35
[ATP] mM
0 1 2 3 4
Act.
Espe
cific
a (U
mg-
1)
0.0
0.1
0.2
0.3
0.4
0.5
Figura 14.- Efecto del quitosano sobre la cinética enzimática de la H+ATPasa de membrana
plasmática de Rhizopus stolonifer. El hongo fue incubado en medio mínimo durante 12 h en (•) ausencia y (◦) presencia de quitosano [2 mg mL
-1] en agitación constante a 25 ± 2 ºC. La barra
representa la desviación estándar de 3 repeticiones.
Cuadro 7.- Valores de la Velocidad máxima y de la Km obtenidos del curso temporal de la
actividad de la H+ATPasa de membrana plasmática de Rhizopus stolonifer.
Muestra Vmax
(μmolPi min-1 mg-1)*
Km
(mM ATP)*
Membranas sin quitosano 0.496 ±0.033 a 0.330 ±0.042 a
Membranas con quitosano 0.361 ±0.011b 0.192 ±0.017 b
*Los valores están seguidos por la desviación estándar de la media de 3 repeticiones. Letras diferentes indican diferencias estadísticas significativas de acuerdo a la prueba de rangos múltiples de Tukey p< 0.05

Efecto del quitosano en la funcionalidad de la membrana plasmática Rhizopus stolonifer DISCUSIÓN
36
DISCUSIÓN
Rhizopus stolonifer es un fitopatógeno que produce pérdidas económicas debido
a que causa la pudrición blanda en productos hortofrutícolas. Tradicionalmente, el
control de este hongo se ha realizado empleando fungicidas químicos sintéticos,
pero los riesgos que implica su uso para el medio ambiente y la salud humana
unido a la resistencia que se ha generado a los químicos sintéticos han conllevado
a la búsqueda de diferentes alternativas naturales para su control entre las que se
encuentra el quitosano (Triphathy and Dubey, 2004; Velázquez-del Valle et al.,
2008).
En esta investigación, se estudió el efecto del quitosano sobre la integridad de la
membrana plasmática de las esporas y previamente se determinó la cinética de
germinación de las mismas, encontrándose resultados coincidentes con los
reportados en la literatura (Van-Etten et al., 1974). La cinética de germinación que
se estableció bajo estas condiciones de cultivo e incubación para R. stolonifer
evidenció que las esporas comenzaron a germinar a las 4 h de incubación,
obteniéndose un porcentaje máximo de germinación (91%) a las 7 h de
incubación. Los resultados del pH del medio de cultivo reflejaron que disminuye en
el medio donde se cultiva el hongo sin quitosano, mostrándose la acidificación del
mismo, por el contrario, la adición del polímero abatió esta tendencia de
acidificación, manteniéndose estable el pH durante las 12 h de incubación. Este
efecto podría deberse a que el quitosano induce la salida de compuestos que
neutralizan la acidez del medio o también podría afectar la salida de protones al
medio de cultivo, estas hipótesis no han sido demostradas en estudios previos
incluso este efecto por acción del quitosano no se ha reportado a la fecha.
Con relación a la integridad de la membrana de R. stolonifer, se observó que la
misma se afectó en presencia de quitosano [2 mg mL-1], obteniéndose un

Efecto del quitosano en la funcionalidad de la membrana plasmática Rhizopus stolonifer DISCUSIÓN
37
porcentaje de integridad de membrana de 62.65 a las 6 h de incubación, lo que
sugiere que hay cambios en la permeabilidad de la membrana. Estos resultados
concuerdan con los ya reportado por Liu et al., (2007), donde ellos observan como
la integridad de membrana también disminuye en los hongos Botrytis cinerea y
Penicillium expansum, siendo más sensible P. expansum (aún comparando a los
tres hongos). Adicionalmente, otras investigaciones demostraron que el quitosano
permeabilizó y penetró la membrana plasmática de diferentes células fúngicas
(hifas, esporas y tubos germinales) mediante un proceso dependiente de energía
(ATP), los autores sugirieron que el polímero interactuó con los componentes
externos de la membrana provocando cambios conformacionales que implicaron la
formación de poros (Palma-Guerrero et al., 2008; 2009). En investigaciones
recientes se propone a la membrana plasmática como el principal blanco de
acción del quitosano en hongos (Park et al., 2008), otros autores como Rabea et
al., (2003) y Eikenes et al., (2005) plantean que el quitosano interacciona con la
membrana plasmática de las célula alterando la permeabilidad de dicha estructura.
Además, existen resultados que aportan evidencia acerca de que el quitosano [2.5
mg mL-1] permeabilizó diferente tipos de células fúngicas (levaduras, conidios y
micelio) con las que estuvo en contacto durante 24 h (Park et al., 2008).
Una vez demostrado el cambio en la permeabilidad de la membrana plasmática
del hongo, se estudiaron las variaciones en el potencial de la membrana
plasmática de dicho hongo, esto por la relación que hay entre la permeabilidad, el
bombeo de protones y la generación del potencial de membrana. Los resultados
demostraron que existieron variaciones en el potencial de la membrana plasmática
cuando el hongo estuvo en contacto con el quitosano, la intensidad del potencial
de la membrana que se generó es menor comparado con el testigo, este resultado
puede deberse a que la H+ATPasa que es la principal encargada del bombeo de
protones y generación del potencial de membrana no esté funcionando de forma
normal, existen reportes en los que se demostró que la actividad de la H+ATPasa
de plantas fue inhibida por el quitosano (Amborabé et al., 2008). En este estudio
se debe tener en cuenta que la integridad de la membrana está disminuida y la

Efecto del quitosano en la funcionalidad de la membrana plasmática Rhizopus stolonifer DISCUSIÓN
38
permeabilidad está alterada, lo que puede provocar que no se genere un potencial
de membrana óptimo. Es importante destacar que no existen reportes previos en
hongos filamentosos del efecto del quitosano sobre el potencial de membrana, en
este estudio se demuestra su afectación.
Con este trabajo se demuestra que el quitosano afecta funciones básicas de la
membrana plasmática de R. stolonifer, por lo que se evaluó si esté fenómeno
estaba provocando la salida de moléculas propias del citosol del hongo. El K+ es
un elemento casi exclusivo del interior de las células, está involucrado en el
balance osmótico entre la célula y el medio que la rodea, también juega un papel
en el control del pH intracelular, participa en el potencial de membrana, funciones
enzimáticas, entre otros. Su medición es aparentemente sencilla, gracias a la
existencia de electrodos específicos para el K+. Los resultados evidenciaron la
salida de K+ de las esporas íntegras de R. stolonifer después de haberles
adicionado una cantidad conocida de quitosano. Esto sugiere que el polímero
afecta la membrana plasmática de tal forma que provocó la liberación de K+, lo
que podría explicarse por la activación de distintos canales, por los cuales se da la
salida de K+ de la célula, también existe la posibilidad de que los canales que se
pudieran estar activando sean canales de potasio dependientes de voltaje. Estos
resultados se pueden apoyar con los reportados por Enríquez-Freire et al., (1999)
donde este grupo de trabajo demostró la salida de K+ de la levadura S. cerevisiae
cuando estuvo en contacto con 4 compuestos catiónicos (dietilaminoetil-dextran
DEAE-D, dihidroestreptomicina DHS, cloruro de terbio y bromuro de etidio EB), los
autores afirmaron que la salida de K+ inducida por los compuestos catiónicos no
se deriva de un efecto generalizado debido a su interacción con la cargas
negativas de la membrana plasmática de la levadura, sino que parece depender
de diferentes mecanismos propios de cada agente, aunque para la mayoría de
ellos el fenómeno parece implicar la interacción de las cargas negativas de la
membrana plasmática con las cargas positivas de los compuesto. De igual forma,
se explica la que la salida de K+ se debe dar por la activación de canales
dependientes o no de voltaje. Existen otros trabajos (Thevissen et al., 1996; 2003)

Efecto del quitosano en la funcionalidad de la membrana plasmática Rhizopus stolonifer DISCUSIÓN
39
donde se demuestra que otro tipo de compuestos catiónicos como son las
defensinas producidas por plantas, tienen el mismo efecto sobre los células
fúngicas (Neurospora crassa), provocando la liberación de K+. Las defensinas
afectan la permeabilidad de la membrana plasmática, debido a una interacción
entre las defensinas y los fosfolípidos de la membrana, resaltando que las
interacciones más fuertes se dan específicamente con los esfingolípidos, que son
los fosfolípidos que presentan la mayor carga negativa en la membrana
plasmática. Estos autores también coinciden en que la salida de K+ tiene que estar
mediada por la activación de canales de potasio y de sistemas de antiporte K+/H+
(Thevissen et al., 1996). Todos estos efectos en los cuales está involucrado la
salida incontrolada de K+ a lo que conlleva es a inducir alteraciones en el
crecimiento celular y en el desarrollo de los organismos (Zhao et al., 2008), lo cual
se pudo observar microscópicamente y macroscópicamente en R. stolonifer
después de haber sido cultivado en presencia de quitosano (resultados no
mostrados).
En este trabajo, al cuantificar los fosfolípidos totales de la membrana plasmática
de R. stolonifer no se encontraron cambios en la concentración de los mismos. Sin
embargo, en reportes previos se demostró que en bicapas lipídicas sintéticas, el
quitosano logró disgregar a los fosfolípidos al grado de generar poros de un gran
tamaño, debido a las fuertes interacciones hidrofóbicas, electrostáticas y dipolos
que se establecieron entre el quitosano y las colas de los fosfolípidos (Fang et al.,
2001; Pavinatto et al., 2007). En este estudio, aunque la membrana está siendo
afectada por la presencia del quitosano, se podría inferir que esta estructura se
mantiene en una biosíntesis constante y reparándose continuamente, para
permitirle al hongo seguir sobreviviendo a pesar del estrés que el quitosano le está
causando.
Una vez que se demostró que el quitosano estaba alterando la membrana
plasmática in situ e in vivo, se decidió trabajar con las membranas aisladas para
determinar algunos otros parámetros que podrían estar afectados por la presencia

Efecto del quitosano en la funcionalidad de la membrana plasmática Rhizopus stolonifer DISCUSIÓN
40
del quitosano. Resultados previos demostraron que se liberan proteínas al medio
de cultivo cuando el hongo crece en presencia de quitosano (Guerra-Sánchez et
al., 2009). Sin embargo, se desconoce si estas proteínas son propias o no de la
membrana plasmática. Los resultados de este estudio mostraron que
efectivamente existe una disminución en la concentración de proteínas presentes
en la membrana plasmática de R. stolonifer cuando el hongo se incubó en
presencia de quitosano. Este fenómeno pudo deberse a la desestabilización o
disminución en la integridad de la membrana plasmática provocando la liberación
no controlada de péptidos y/o proteínas membranales, otra posible explicación es
que la mayoría de las proteínas transmembranales (H+ATPasa, Gas1p) se
encuentran asociadas a las “Balsas lipídicas” del Inglés “lipid rafts” ricas en
esfingolípidos y ergosterol (Ferrerira et al., 2001), las cuales podrían ser afectadas
por las interacciones con el quitosano provocando el desprendimiento de péptidos
y/o proteínas que estaban ancladas a la membrana plasmática. Adicionalmente,
una tercera explicación podría ser que la presencia del polímero inhiba la síntesis
de algunas proteínas de la membrana, se desconoce la causa de este fenómeno
pero el efecto del quitosano es evidente.
Estos resultados justificaron que se realizara la determinación de la actividad
enzimática de una de las proteínas más importantes de la membrana plasmática,
la H+ATPasa. Se realizó la determinación de la actividad de la H+ATPasa de
membrana plasmática de R. stolonifer, los resultados mostraron que tanto la
actividad específica como la actividad total de la H+ATPasa se ven disminuidas un
25 y 50 % respectivamente, cuando el hongo es cultivado en presencia de
quitosano. Estos resultados están muy relacionados con la disminución en la
concentración de proteínas presentes en la membrana celular y con la explicación
que se propuso del porque podría ocurrir ese fenómeno. Estos resultados
concuerdan con los estudios que ya existen en los cuales se evidenció que el
quitosano inhibió la actividad de la H+ATPasa, demostrándose una vez más que la
membrana plasmática, aunque de la células vegetales en este caso, también es el
principal blanco de acción de este polímero (Amborabé et al., 2008). Otros

Efecto del quitosano en la funcionalidad de la membrana plasmática Rhizopus stolonifer DISCUSIÓN
41
reportes señalan que la desorganización de lípidos de membrana de S. cerevisiae
provocados por altas concentraciones de cobre están involucrados en la disipación
del gradiente electroquímico producido por la H+ATPasa, afectando la
funcionalidad y disminuyendo la actividad de la enzima, generando la acidificación
citosolica del hongo (Fernandes et al., 1998). Lo que podría estar ocurriendo en el
caso de este estudio, aclarando que aquí el quitosano es el agente que provoca la
desorganización en la membrana plasmática.
Otros resultados interesantes que se obtuvieron de la H+ATPasa, fueron sus
valores de Vmax y de Km. Ambos parámetros fueron inhibidos por la presencia del
quitosano durante la incubación del hongo. Los valores de Vmax y de Km
disminuyeron en un 25 y 42 % respectivamente, lo que podría deberse a los
cambios en la permeabilidad, a los cambios en el potencial de membrana, a la
disminución del contenido de proteínas de la membrana, a la desorganización de
los fosfolípidos, entre otros factores. Existe un reporte previo que señala que en
mutantes de S. cerevisiae a las que les eliminaron los 18 últimos amino ácidos del
extremo carboxilo terminal de la proteína H+ATPasa, la mutación provocó que las
levaduras presentaran una disminución en el valor de la Km comparada con las
levaduras silvestres. En ese mismo trabajo lograron generar una mutante a la que
le eliminaron los primeros 27 aminoácidos del extremo amino terminal, de la
proteína H+ATPasa. Esta mutación provocó que la levadura presentara una Vmax
menor comparada con las levaduras silvestres (Mason et al., 1998). Lo que
podría sugerir que en este caso el quitosano esté afectando la integridad de la
membrana plasmática, sobre todo a las balsas lipídicas y esto a su vez afectaría a
la H+ATPasa, liberándolas, fragmentándola y generando enzimas que no trabajen
adecuadamente.
Los resultados obtenidos en esta investigación, demuestran el efecto antifúngico
del quitosano sobre el hongo fitopatógeno R. stolonifer expresado a través de las
afectaciones a la funcionalidad de la membrana plasmática del hongo
fitopatógeno, aspecto que contribuye a esclarecer el mecanismo de acción del
quitosano y que podría repercutir en desarrollar mejores estrategias de control del
fitopatógeno.

Efecto del quitosano en la funcionalidad de la membrana plasmática Rhizopus stolonifer CONCLUSIONES
42
CONCLUSIONES
La presencia del quitosano causa la pérdida de integridad de la
membrana plasmática de las esporas de R. stolonifer.
El pH del medio, el potencial de membrana y la salida de K+ de las
esporas se afectan por la presencia del quitosano.
El quitosano no afecta el contenido de fosfolípidos totales de la
membrana plasmática del micelio de R. stolonifer.
El quitosano ocasiona la disminución de las proteínas de la
membrana plasmática del micelio de R. stolonifer.
La actividad de la H+ ATPasa y sus parámetros cinéticos (Vmax y
Km) disminuyen por efecto del quitosano.

Efecto del quitosano en la funcionalidad de la membrana plasmática Rhizopus stolonifer Perspectivas
43
PERSPECTIVAS
Estudiar el efecto del quitosano y de agentes permeabilizantes conocidos
(digitonina, TritonX-100, tolueno, etc.) en la permeabilización de las células
fúngicas.
Investigar la actividad de enzimas claves que estén involucradas en la síntesis y el
recambio de fosfolípidos como lo son las fosfolipasas (A, C y D), debido a que
podría estar acelerada la síntesis o el recambio, lo cual contribuiría a explicar que
las membranas no presentan alteraciones en los resultados de fosfolípidos totales.
Cuantificar los fosfolípidos de la membrana plasmática del hongo, en condiciones
naturales y bajo la presencia de quitosano. Esto ayudaría a comprobar si existen
cambios en la cantidad y variedad de los fosfolípidos a causa del quitosano, lo que
podría estar provocando interacciones electrostáticas más fuertes entre el
quitosano y la membrana.
Comprobar si el quitosano penetra a las células de R. stolonifer (hifas y/o esporas)
y demostrar su localización celular. Esto se podría realizar utilizando microscopia
confocal y marcando al quitosano con algún compuesto fluorescente, lo que
permitiría profundizar las investigaciones sobre alguna de las estructuras a la cual
el quitosano se uniera.
Con respecto al estudio de la H+ATPasa, se podría continuar con la búsqueda de
la proteína en los sobrenadantes (medio de cultivo), para lo cual se tendría que
implementar un método de purificación y concentración. Esto ayudaría a
comprobar si esta proteína de membrana está siendo liberada al medio como
consecuencia de la presencia del quitosano. Adicionalmente, se podría demostrar
si se altera la expresión de la proteína por la presencia de quitosano.
Efecto del quitosano en la funcionalidad de la membrana plasmática Rhizopus stolonifer Anexos
44
ANEXOS
Agar Papa Dextrosa (PDA) (BIOXON) PDA en polvo 39 g Agua destilada 1000 mL Preparación: Pesar 39 g de agar papa dextrosa en polvo en una balanza analítica,
disolver en 500 mL de agua destilada, calentar a ebullición por 1 minuto con
agitación periódica, después aforar a 1000 mL con agua destilada y esterilizar en
autoclave a 121ºC durante 15 minutos. Esperar a que enfríen un poco y en
condiciones de esterilidad, verter el medio a las placas Petri estériles, dejar
solidificar a temperatura ambiente y almacenarlas en refrigeración hasta su uso.
Curva tipo de albumina sérica bovina por el método de Lowry
BSA ( L) Reactivo A
( L)
Reactivo B
( L)
Concentración
final ( g/ L)
0 100 800 0
10 100 800 5
20 100 800 10
40 100 800 20
60 100 800 30
Solución de Albúmina Sérica Bovina (BSA) 0.5 mg/mL
BSA 0.005 g
Agua destilada 10 mL
Preparación: Pesar 0.0005 g de BSA en una balanza analítica, disolver
perfectamente en 10 mL de agua destilada. Separar en viales en volúmenes de 1
mL y congelar hasta su uso, cada vial se utiliza una sola ocasión.

Efecto del quitosano en la funcionalidad de la membrana plasmática Rhizopus stolonifer Anexos
45
Curva tipo de fosforo para la actividad de H+ATPasa por el método de Fiske–Subbarow.
NaH2PO4•H20
( L)
TCA 8.3%
( L)
Molibdato de
Amonio ( L)
Elon 0.05%
( L)
Concentración
final (nmolPi L-1)
0 250 250 250 0
20 250 250 250 40
40 250 250 250 80
50 250 250 250 100
60 250 250 250 120
Concentración de ATP empleado para la cinética enzimática. Solución madre de ATP-Mg a concentración de 24 mM
ATP
( L)
Concentración de
ATP (mM)
Regulador de
reacción ( L)
0.0 0.00 250.0
2.6 0.25 247.4
5.2 0.50 244.8
7.8 0.75 242.2
10.4 1.00 239.6
13.0 1.25 237.0
15.6 1.50 234.4
20.8 2.00 229.2
31.2 3.00 218.8
41.6 4.00 208.4
Hidróxido de Sódio 1 N NaOH 10 g Agua destilada 250 mL Preparación: Pesar 10 g de NaOH en una balanza analítica, disolver con agua destilada y aforar a 250 mL.

Efecto del quitosano en la funcionalidad de la membrana plasmática Rhizopus stolonifer Anexos
46
Suspensión de Esporas
Para preparar la suspensión de esporas se necesitan una caja de Petri que tenga
un crecimiento del hongo entre el 90-100%, este se logra con 4 días de
incubación, adicionar 10 mL de agua destilada estéril con el fin de empapar las
esporas de Rhizopus stolonifer, con una varilla de vidrio flameada previamente,
frotar toda la superficie de la caja de Petri, recuperar el volumen directamente en
un vaso de precipitados y agregar otros 10 mL de agua destilada estéril, para
tener una suspensión de esporas en 20 mL. Una vez preparada utilizarse
inmediatamente. Para conocer la cantidad de esporas es necesario realizar el
conteo de esporas en una cámara de conteo (Neubauer), de esta manera se sabe
cantidad de esporas que se están inoculando por mililitro de suspensión utilizada.

Efecto del quitosano en la funcionalidad de la membrana plasmática Rhizopus stolonifer BIBIOGRAFÍA
47
BIBLIOGRAFÍA
Adaskaveg, J.E., Förster, H. and Sommer, N.F. 2002. Principles of postharvest pathology and management of decays of edible horticultural crops. pp. 163-195. In: A. kader (ed.) Postharvest Technology of Horticultural Crops. University of California. Oakland, California, USA. 535 p.
Agrios, G.N. 2001. Fitopatología. Segunda edición. Noriega Editores, México. p 332-336.
Amborabé, B., Bonmort, J. Fleurat-Lessard, P. and Roblin, G. 2008. Early events induced by chitosan on plant cells. J Exp Bot. 59:2317-2324
Ait-Barka, E., Eullaffroy, P., Clément, C. and Vernet, G. 2004. Chitosan improves development, and protects Vitis vinifera L. against Botrytis cinerea. Plant Cell Rep. 22:608-614.
Alexopoulos, C.J., Mims C. W. and Blackwell, M. 1996. Introductory Mycology. Cuarta edición. John Wiley & Sons editores, New York.
Allan, C. and Hadwiger, L. 1979. The fungicidal effect of chitosan on fungi of vaying cell with competition. Exp Mycol. 3:285-287.
Bautista-Baños, S., Hernández-Lauzardo, A.N., Velázquez del Valle, M.G., Hernández-López, M., Ait-Barka, E., Bósquez-Molina, E. and Wilson, C.L. 2006. Chitosan as a potencial natural compound to control pre and postharvest diseases of horticultural commodities. Crop Protection. 25:108-118.
Bautista-Baños, S., Hernández-López, M. and Bósquez-Molina, E. 2004. Growth inhibition of selected fungi by chitosan and plant extracts. Rev Mex Fitop. 22:178-186.
Benito, B. Portillo, F. and Laguna, R. 1992. In vivo activation of the yeast plasma membrane ATPase during nitrógeno starvation. FEBS Lett. 300:271-274.
Blasco, F., Chapuis, J.P. and Giordani, R. 1981. Characterization of the plasma. membrane ATPase of. Candida tropiealis. J Biochimie. 63:507- 514
Böttcher C.J.F., Van Gent, C.M. and Pries, C. 1961. A rapid and sensitive sub-micro phosphorus determination. Anal Chim Acta 24:203-204.
Bowman, B.J., Mainzer, S.E., Allen, K.E. and Slayman, C.W. 1978. Effects of inhibitors on the plasma membrane and mitochondrial adenosine triphosphatases of Neurospora crassa, Biochim Biophys Acta. 512:13-28.
Bowman E.J., Siebers A. and Altendorf K. 1988. Bafilomycins: a class of inhibitors of membrane ATPases from microorganisms, animal cells, and plant cells. Proc. Natl. Acad. Sci. USA 85:7972-7976.
Daum, G., Lees, N.D., Bard, M. and Dickson, R. 1998. Biochemistry, cell biology and molecular biology of lipids of Saccharomyces cerevisiae. Yeast. 14:1471-1510.
Dryer, R.L., Tammes, A.R. and Routh, J.I. 1957. The determination of phosphorus and phosphatase with N-phenyl-p-phenylenediamine. J Biol Chem. 225:177-183.
Eikenes, M., Alfredsen, G., Christensen, B., Militz, H. and Solheim, H. 2005. Comparison of chitosan with different molecular weights as possible wood preservative. J Wood Sci. 51:387–394.

Efecto del quitosano en la funcionalidad de la membrana plasmática Rhizopus stolonifer BIBIOGRAFÍA
48
El Ghaouth, A., Arul, J., Asselin, A. and Benhamou, N. 1992. Antifungal activity of chitosan on post-harvst pathogens: introduction of morphological and cytological alterations in Rhizopus stolonifer. Mycol Res. 96:769-779.
Enriquez-Freire, E., López, R. and Peña, A. 1999. Potassium ion efflux induced by cationic compounds in yeast. Biochim Biophys Acta. 1418:147-157.
Eraso, P. and Gancedo, C. 1987. Activation of yeast plasma membrane ATPase by acidpH during growth. FEBS Lett. 224:187-192.
Fang, N., Chan, V., Mao, H.Q. and Leong, K.W. 2001 Interactions of phospholipid bilayer with chitosan: effect of molecular weight and pH. Biomacromolecules. 2:1161-1168.
Fernandes, A.R., Peixoto, F.P. and Sá-Correia, I. 1998. Activation of the H+-ATPase in the plasma membrane of cells of Saccharomyces cerevisiae grown under mild copper stress. Arch Microbiol. 171:6–12
Ferreira, T., Mason, A.B. and Slayman C.W. 2001. The yeast Pma1 proton pump: a model for understanding the biogenesis of plasma membrane proteins. J Biol Chem. 276:29613-29616.
Folch, J., Lees, M. and Sloane-Santley, G.H. 1957. A simple method for the isolation and purification of total lipides from animal tissues. J Biol Chem. 226:497-509.
Goffeau, A. and Slayman, C.W. 1981. The proton-translocating ATPase of the fungal plasma membrane. Biochim Biophys Acta. 639:197–223.
Guerra-Sánchez, M.G., Uribe-Carvajal, S. and Pardo.Vásquez, J.P. 1995. Las ATPasas de protones de los hongos. Ciencias. 46:483-503.
Guerra-Sánchez, M.G., Vega-Pérez, J., Velázquez-del Valle, M.G. and Hernández-Lauzardo, A.N. 2009. Antifungal activity and release of compounds on Rhizopus stolonifer (Ehrenb.:Fr.) Vuill. by effect of chitosan with different molecular weights. Pestic Biochem Physiol. 93:18-22.
Gonzalez-Aguilar G., Gardea A. y Cuamea-Navarro F. 2005. Nuevas tecnologías de conservación de productos vegetales frescos cortados. CIAD, CYTED, CONACYT y COFUPRO. Jalisco, México.
Guo, Z., Chen, R., Xing, R., Liu, S., Yu, H., Wang, P., Li, C. & Li, P. 2006. Novel derivatives of chitosan and their antifungal activities in vitro. Carbohyd Res. 341:351-354
Hernández, A., Cooke, D.T. and Clarkson, D.T. 1994. Lipid composition and proton transport in Penicillium cyclopium and Ustilago maydis plasma membrane vesicles isolated by two-phase partitioning. Biochim Biophys Acta. 1195:103-109.
Hernández, A., Cooke, D.T. and Clarkson, D.T. 2002. In vivo activation of plasma membrane H+ATPase hydrolytic activity by complex lipid-bound unsaturated fatty acids in Ustilago maydis. Eur J Biochem. FEBS. 269:1006-1011.
Hernández-Lauzardo, A.N., Bautista-Baños, S. Velázquez-del Valle, M.G., Méndez-Montealvo, M.G., Sánchez-Rivera M.M. and Bello-Pérez L.A. 2008. Antifungal effects of chitosan with different molecular weights on in vitro development of Rhizopus stolonifer (Ehrenb.:Fr.) Vuill. Carbohydrate Polymers. 73:541-547.
Hernández-Lauzardo, A.N., Bautista-Baños, S., Velázquez-del Valle, M.G., Rodríguez-Ambriz, S.L., Corona-Rangel, M.L., Solano-Navarro, A. and Bósquez-Molina, E. 2005. Potencial del quitosano en el control de las enfermedades postcosecha. Rev Mex Fitop. 23:198-205

Efecto del quitosano en la funcionalidad de la membrana plasmática Rhizopus stolonifer BIBIOGRAFÍA
49
Hernández-Lauzardo, A.N., Bautista-Baños, S., Velázquez-del Valle, M.G. and Trejo-Espino, J.L. 2006. Identification of Rhizopus stolonifer (Erenb.:Fr.) Vuill., causal agent of Rhizopus rot disease of fruits and vegetables. Rev Mex Fitop. 24:65-69.
Hernández-Lauzardo, A.N., Hernández-Martínez, M., Velázquez-del Valle, M.G., Guerra-Sánchez, M.G. and Melo-Giorgana, G.E. 2007. Actividad antifúngica del quitosano en el control de Rhizopus stolonifer (Erenb.:Fr.) Vuil. y Mucor spp. Rev Mex Fitop. 25:109-113.
Hernández-Muñoz, P., Amenar, E., Ocio, M.J. and Gavara, R. 2006. Effects of calcium dips and chitosan coatings on postharvest life of strawberries (Fragaria x ananassa). Postharvest Biol Technol. 39:247-253.
Holmes, G. and Stange, R. 2002. Influence of wound type and storage duration on susceptibility of sweetpotatoes to Rhizopus soft rot. Plant Disease. 86:345-348.
Hubbard, M.J., Surarif, R., Sullivan, P.A. and Shepard M.G. 1986. The isolation of plasma membrane and characterisation of the plasma membrane ATPase from the yeast Candida albicans. Eur J Biochem. 154:375-381.
Jain P., Akula I. and Edlind T. 2003. Cyclic AMP signaling pathway modulates susceptibility of Candida species and Saccharomyces cerevisiae to antifungal azoles and other sterol biosynthesis inhibitors. Antimicrob Agents Chemother. 47:3195-320.
Kim, S.K. and Rajapakse, N. 2005. Enzymatic production and biological activities of chitosan oligosaccharides (COS): a review. Carbohydrate Polymers. 62:357-368.
Lárez, V.C. 2003. Algunos usos del quitosano en sistemas acuosos. Rev Iberoam Polím. 4:91-108.
Lárez, V.C. 2006. Quitina y quitosano: materiales del pasado para el presente y el futuro. Rev Avan Quím. 1:15-21.
Liu, J., Tian, S., Meng, X. and Xu, Y. 2007. Effects of chitosan on control of postharvest diseases and physiological responses of tomato fruit. Postharvest Biol Technol. 44:300-306.
Liu, H., Du, Y., Wang, X. and Sun, L. 2004. Chitosan kills bacteria through cell membrane damage. Int J Food Microbiol. 95:147-155.
Lowry, O.H., Rosebrough N,J., Farr A.L. and Randall R.J. 1951. Protein measurement with the Folin phenol reagent. J Biol Chem. 193:265-275.
Martínez, J.P. and Gozalbo, D. 2001. Chitin. Encyclopedia of life sciences. Nature publishing group.
Mason, A.B., Kardos, T.B. and Monk, B.C. 1998. Regulation and pH-dependent expression of a bilaterally truncated yeast plasma membrane H+ATPase. Biochim Biphys Acta. 1372:261-271.
McDonald, J.I.S. and Weeks, G. 1984. A plasma membrane Mg2+-ATPase in the cellular slime mold Dictyostelium discoideum. Arch Biochem Biophys. 235:1-7.
Moore-Landecker, E. 1996. Fundamentals of the Fungi. Cuarta edición. Prentice Hall, Inc., New Jersey.
Nakamoto, R.K., Roa, R., and Slayman, C.W. 1989. Transmembrane segments of the P-type cation-transporting ATPases. A comparative study. Ann. N. Y. Acad. Sci. 574:165-179

Efecto del quitosano en la funcionalidad de la membrana plasmática Rhizopus stolonifer BIBIOGRAFÍA
50
Northover, J. and Zhou, T. 2002. Control of rhizopus rot of peaches with postharvest treatments of tebuconazole, fludioxonil, and Pseudomonas syringae. Can J Plant Pathol. 24:144-153.
Pacheco, N., Larralde-Corona, C-P., Sepulveda, J., Trombotto, S., Domard, A. and Shirai, K. 2008. Evaluation of chitosans and Pichia guillermondii as growth inhibitors of Penicillium digitatum. Int J Biol Macromol. 43:20-26.
Palma-Guerrero, J., Huang, I.C., Jansson, H.B., Salinas, J., Lopez-Llorca, L.V. and Read, N.D. 2009. Chitosan permeabilizes the plasma membrane and kills cells of Neurospora crassa in an energy dependent manner. Fungal Genet Biol. 46:585-594.
Palma-Guerrero, J., Jansson, H.B., Salinas, J. and Lopez-Llorca, L.V. 2008. Effect of chitosan on hyphal growth and spore germination of plant pathogenic and biocontrol fungi. J Appl Microbiol. 104:541-553.
Paraje, M.G., Correa, S.G., Albesa I. and Sotomayor, C.E. 2009. Lipase of Candida albicans induces activation of NADPH oxidase and L-arginine pathways on resting and activated macrophages. Biochem Biophys Res Commun. Article in Press.
Pavinatto, F.J., Pavinatto, A., Caseli, L., Santos, D.S., Nobre, T.M., Zaniquelli, M.E. and Oliveira, O.N. 2007. Interaction of chitosan with cell membrane models at the air-water interface. Biomacromolecules. 8:1633-1640.
Park, Y., Kim, M-H., Park, S-C., Cheong, H., Jang, M-K., Nah, J-W. and Hahm K-S. 2008. Investigation of the antifungal activity and mechanism of action of LMWS-Chitosan. J Microbiol Biotechnol. 18:1729-1734.
Peña, A., Uribe, S., Pardo, J.P. and Borbolla M. 1984. The use of a cyanine dye in measuring membrane potential in yeast. Arch Biochem Biophys. 231:217-225.
Pedersen, P.L. and Carafoli, E. 1987. Ion motive ATPases. I. Ubiquity, properties and significance to cel function. Trends Biochem Sci. 12:146-150.
Portillo, F. 2000. Regulation of plasma membrane H+ATPase in fungi and plantas. Biochem Biphys Acta. 1469:31-42.
Rabea, E.I., Badawy, M.E-T., Rogge, T.M., Stevens C., Hofte, M. Steurbaut W. and Smagghe, G. 2005. Insecticidal and fungicidal activity of new synthesized derivates. Pest Manag Sci. 61:951-960.
Rabea, E.I., Badawy, M.E-T., Stevens, C.V., Smagghe, G. and Steurbaut, W. 2003. Chitosan as antimicrobial agent: applications and mode of action. Biomacromolecules. 4:1457–1465.
Schipper, M.A. 1984. A revision of the genus Rhizopus. Studies in Mycology. Serie No. 25. Centraalbureau voor Schimmelcultures. Baarn, Netherlands. pp. 34.
Serrano, R. 1988. Structure and function of proton translocating ATPase in plasma membranes of plants and fungi. Biochim Biophys Acta. 947:1-28.
Seyfarth, F., Schliemann, S., Elsner, P. and Hipler, U.-C. 2008 Antifungal effect of high- and low- molecular-weight chitosan hydrochloride, carboxymethyl chitosan, chitosan oligosaccharide and N-acetyl-d-glucosamine against Candida albicans, Candida krusei and Candida glabrata. Int J Pharm. 353:139.148.
Thevissen, K., Ferket, K.K.A., Francois, I.E.J.A. and Cammue, B.P.A. 2003. Interaction of antifungal plant defensins with fungal membrane components. Peptides. 24:1705-1712.

Efecto del quitosano en la funcionalidad de la membrana plasmática Rhizopus stolonifer BIBIOGRAFÍA
51
Thevissen,K. Ghazi, A. De Samblanx, G.W., Brownlee, C., Osborn, R.W. and Broekaert, W.F. 1996. Fungal membrane response induced by plants defensins and thionins. J Biol Chem. 271:15018-15025.
Triphathy, P. and Dubey, N.K. 2004. Exploitation of natural products as an alternative strategy to control postharvest fungal rotting of fruit and vegetables. Postharvest Biol Technol. 32:235-245.
Van Der Rest, M.E., Kamminga, A.H., Nakano, A., Anraku, Y., Poolman, B. and Konings, W.N. 1995. The plasma membrane of Saccharomyces cerevisiae: structure, function and biogenesis. M. Reviews. 59:304-322.
Van-Etten, J.L., Bulla, L.A.Jr. and Julian, G.S. 1974. Physiological and Morphological Correlation of Rhizopus stolonifer Spore Germination. J. Bacteriol. 117:882-887
Velázquez-del Valle M. G., Bautista-Baños S., Hernández-Lauzardo A. N., Guerra-Sánchez M. G. and Amora-Lazcano E. 2008. Estrategias de control de Rhizopus stolonifer (Ehrenb.:Fr.) Vuill. agente causal de pudriciones postcosecha en productos agrícolas. Rev Mex Fitop. 26:49-55.
Weissman, Z., Berdicevsky, I., Cavari, B.Z. and Kornitzer, D. 2000. The high copper tolerance of Candida albicans is mediated by a P-type ATPase. Proc Natl Acad Sci. 27:3520-3525.
Zhao, J., Cheng, N.-H., Motes, C.M., Blancaflor, E.B., Moore, M., Gonzales, N., Padmanaban, S., Sze, H., Ward, J.M. and Hirschi K.D. 2008. AtCHX13 is a Plasma membrane K+ transporter. Plant Physiol. 148:796–807.
PÁGINAS WEB SELECCIONADAS
URL1.http://iescarin.educa.aragon.es/depart/biogeo/varios/BiologiaCurtis/Seccion%205/5%20-%20Capitulo%2029.htm
URL2. http://www.puc.cl/sw_educ/biologia/bio100/html/portadaMIval2.5.html

Efecto del quitosano en la funcionalidad de la membrana plasmática Rhizopus stolonifer BIBIOGRAFÍA
52


















