
Distribucion y ecologia de la comunidad de hemipteros...
93
UNIVERSIDAD MAyaR DE SAN ANDRÉS FACULTAD nE CIENCIAS PURAS Y NATURALES CARRERr\ DE BIOLOG[A DISTRIBUCiÓN Y ECOLOGíA DE LA COMUNIDAD DE HEMíPTEROS AcuA.TiCOS (SUBORDEN: HETERÓPTERA), EN LAS LAGUNAS DE LA ZONA DE INUNDACiÓN DEL Río MAMORÉ (TRINIDAD- SEN!) Tesis para optar al grado de Licenciado en Biología: Univ. Osear Carrasco Arias Tutor: Rubén Marin Pantoja Asesor: Franc;ois-Marie Giban La Paz - Bolivia 2005
Transcript of Distribucion y ecologia de la comunidad de hemipteros...

UNIVERSIDAD MAyaR DE SAN ANDRÉSFACULTAD nE CIENCIAS PURAS Y NATURALES
CARRERr\ DE BIOLOG[A
DISTRIBUCiÓN Y ECOLOGíA DE LA COMUNIDAD DE HEMíPTEROSAcuA.TiCOS (SUBORDEN: HETERÓPTERA), EN LAS LAGUNAS DE LA
ZONA DE INUNDACiÓN DEL Río MAMORÉ (TRINIDAD- SEN!)
Tesis para optar al grado de Licenciado en Biología:
Univ. Osear Carrasco Arias
Tutor: Rubén Marin Pantoja
Asesor: Franc;ois-Marie Giban
La Paz - Bolivia
2005

¡~N AGRADECIMIENTO A:
... MI FAMILIA, POR LA AYUDA y PERSEVERANCIAEN MOMENTOS DIFÍCILES, POR LA COMPAÑÍA
EN TRIZTEZAS y ALEGRJAS ...
.. .MI ASESOR, EL DR. FRAN~OIS GIBON ...
... LA PERSONA QUE ME AYUDO Y ME DIO ENSEÑANZA ENMIS PRIMEROS PASOS DE PROFESIONAL. ..
'.' MIS DOCENTES, JULIO, RUBEN Y ROBERTO...
' ..DR. AXEL BACHMANN, DRA. SILVIA MAZZUCONI,DRA. MONICA LOPEZ RUF Y LA LIC. PAULINA.
...EL PERSONAL DEL INSTITUTO DEECOLOGÍA, EL PERSONAL DEL LR.D.
Y EL PERSONAL DE LA CARRERA DE BIOLOGÍA.
...MARCELO, KAREN, GABRIEL, ISABEL,CECILIA, JORGE, NOEMI, SERGIO, MAURICIO,
MOORY, NICANED, POLITO, ABDUL Y A TODASLAS PERSONAS Y AMIGOS QUE HAN APORTADO
UN POCO DE CADA UNO A MI FORMACIÓN ...
... A LA PERSONA QUE ME ACOMPAÑAY HACE DIFERENTES MIS DIAS.

,~iNDICE GENERALIl' .=-! ntroducción 111-. ¡ Objetivos 2i1.1.1. Objetivo general 211. \ .2. Objetivos específicos 21, F I ' . 4'_o .uneamento teonco
12-~ I Funcionamiento ecológico de ecosistemas acuáticos tropicales 4I12. \.1 El régimen pulsátil de los ecosistemas 512.1.2 Consecuencias ecológicas del régimen pulsátil 612.1 A Teorías de perturbación 7I
12.2 Generalidades del orden Hemiptera 8i2.~ Características anatómicas internas en Hemiptera 13¡2A Consideraciones ecológicas 14r...,' d di 16jo). Arca e estu 10
fl .Cuenca Mamoré 16L
[3.1.1 Clima y régimen hidrológico 17!~.1.3 Características fisicoquímicas de las aguas 18L
13.1 A Gradientes medioambientales de las lagunas de bosque de galería y de sabana 19I
[3.2. Macrófitas 201--
1).3. Ubicación de las lagunas en estudio 22i..L Materiales y métodos 24P.I. Diseño de muestreo 241::2. Metodología para la colecta e identificación de invertebrados 25I~U Tratamientos de datos 25¡...J..3.1 Análisis multivariado 25~~.: Análisis factorial de correspondencia (AFC) 25¡4.,) .." Prueba T de Student 2715. Resultados 28I
Is. 1 Determinación de especies 29r2 Distribución y ecología de hemípteros acuáticos 48r5.3 Análisis estadísticos 505.3.1 Análisis factorial de correspondencia .. 505.3.2. Análisis discriminante 515.4. Comparación de abundancia por factores 5615.4. J Prueba T de Student 565.4.1.1 Factor época 5615.4.1.2 Factor laguna 595.4.1.3 Factor vegetación 60

Discusión 62Conclusiones 69[3 ibliografía 71
leí .1-
"I! .18.
ii

INDICE DE FIGURAS
! Fi<Jura 1. Sucesión hipotética en un espacio vacíoI h
1--
i Fisura 2. Relación teórica entre la diversidad biótica y varios medios de perturbaciónI h
I Fioura 3. Vista general un hemíptero acuático. A: Vista dorsal B: Vista Ventral, h
iFigura 4. A: Estructura del aparato bucal; B: Esquema en vista dorsal de las medidas de laI cabeza. V: Vertex, S: Sintlipsis; O: Borde posterior del ojo; A: Ancho del disco pronotal; L:Ll-ollgitud del disco pronotal.! Figunl 5. Hemielitro derecho. Abreviaciones: CL: Clavo; el: longitud del area pruinosa de
lb sutura clava]; CO: Cario; E: Surco embolar; M: Membrana; N: Nodo; P: Longitud del,
I arca pruinosa postnodal; SM: Sutura membranalIr
6. Hábitat de los hemípteros acuáticos: A) Superficiales, B) Verdaderamente! Finura¡ h
1 acuáticos, C) De orilla y fondo.
Figura 7. Cuenca del Río Mamaré
Figura 8. Ubicación de las lagunas en estudio
Figura 9. Diagrama de flujo del estudio
Figura 10. Estimación del número de ejes importantes. Histogramas de los valores propios
1 con dos ejes importantes
~r·I '¡gura 11. Cápsula genital en macho y hembra de Neoplea maculosa; A: Séptimo!i urosternito de la hembra, B: 8vo urito macho, C: Pigóforo, parámero derecho y parámero!
¡ izquierdo, O: Valva del ovopositor
rFigura 12. Neoplea Semipicta; A: Séptimo urostemito de la hembra, B: 8vo urito macho,1
I C: Pigóforo, parárnero derecho y parámero izquierdo, O: Valva del ovopositor
!Figura 13. Cápsula genital en macho y hembra de la especie Neoplea absona; A: SéptimoI
I
Iurosternito de la hembra, B: 8vo mito macho, C: Pigóforo, parámero derecho y parámero
I
r izquierdo, O: Valva del ovopositor
Figura 14. Estructuras para la determinación de la especie Tenagobia incerta. A:
Parárncros izquierdo y derecho; B: Pata anterior de la hembra; C: Uña pata posterior de la
hembra ..Figura 15. Tenagobia socialis. A: urosterguitos 5 y 8vo del macho; B: Parámeros izquierdo
y derecho; C: Urosternito 7mo y 8vo de la hembra
Figura 16. Estructuras para determinar la especie Tenagobia selecta. A: Parámeros derecho
iii

(l) e izquierdo (2); B: Apéndice mito 8vo macho; C: pata anterior (1) y antena en la
hCi11bra (2)
Fioura 17. A: Vista dorsal de la larva del género Heterocorixa; B: Antena en larva y adulto1 "
II;:ig~u-r-a-1-8-.-B-l-le-n-o-a-.-)G-l:-u-tl-·s-A-:-P-r:-im-er-p-a--r----:-d-e-p-a-ta-s-en-e-=-l -m-a-c""":"h-o-,-e-s-tr-u-c-tu-r-a-e-s-t:-r i:-:d:-u-=-l a-t-o-r¡:-a-; -=B:-1:I
I Rostrurn
II~igura 19. Martarega chinai A: Hemielitro B: Vista dorsalf-Ir· 20. Belostoma A: Abdomen pilosidad extendida hasta los esternitos; B:1 '¡gura conI! Abdomen vista ventral!
:i'Tgura 21. Lethocerus truncatus A: genitalia vista lateral; B: Pilosidad abdominal~i Finura 22 A: Vista dorsal ápice abdomen, Neogerris sp; B: ápice abdomen Neogerris1 h •
f
i lubricusII1, Figura 23. Limnogonus aduncus. A: Abdomen vista dorsal; B: Genitales vista lateral
! Figura 24. Rheutnatobates crassifemur crassifemur A: Antena del macho, B: Vista general
i Figura 25. A: Vista general de Mesovelia mulsanti B: Tarso con garras apicales
¡ C: HcmielitroI
ir"igura 26. A: Vista general de Stridulivelia B: Pata posterior vista lateral C: Vista lateralI
I del tóraxi! FigUIoa 27. Gelastocoris A: Vista general. B: Tarso anterior no fusionado a la tibia¡-1 Figura 28. Pelocoris binotulatus binotulatus A: Vista general; B: Estructuras: 1. Pata 1 2.
1 Antena 3. Genitalia femenina 4. Aedeago,,-! Figura 29. Ctenipocoris shadei A: Vista general; B: Estructuras genitales: 1. Genitalia
'¡masculina, 2. Genitalia femenina, 3. Parámero, 4. Aedeago (vista ventral y lateral)i
i Figura 30. Ranatra signoreti A: Vista general; B: Paraesternitos abdominales; C: Antenas;
1
1
D: Genitalia
Figura 31. Abundancia de los géneros y especies mas representativos de hemípteros
acuáticos
IFigura 32. Análisis factorial de correspondencia (AFC) de 8 géneros de hemípteros
acuáticos. A) Valor propio, B) Mapa factorial de géneros, C) Agrupación de las lagunas
Figura 33. Análisis factorial de la tabla faunística de hemípteros acuáticos. Proyección delas épocas clasificadas en función a la abundancia
l
iv

¡-Fi:!,Unl 34. Test por simulaciones aleatorias: época¡_._--.-
iFigura 35. Análisis factorial de la tabla faunística de hemípteros acuáticos. Proyección de I
1 las lagunas clasificadas en función a la abundancia~--_._----_._------------------------------¡
[Figura 36. Test por simulaciones aleatorias: laguna
¡ Figu ra 37. Análisis factorial de la tabla faunística de hemípteros acuáticos. Proyección deI
: la vegetación clasificada en función a la abundancia de hemípteros acuáticos.I _. -'- ---1
¡-Figura 38. Test por simulaciones aleatorias: Vegetación1----------------------------------------1I Figura 39. Comparación de abundancias en géneros de hemípteros acuáticosI •
i~;'~g~;'a 40. Abundancia de hemípteros acuáticos por época
. Figura 41. Abundancia de hemípteros acuáticos por lagunaI --------c-----------------------------ji Figura 42. Abundancia de hemípteros acuáticos por vegetación.1._
v

iNDICE DE CUADROS
IClladr~o ]. Familias que comprenden los grupos Gerromorpha y Nepomorphai.f~adro 2. Características fisicoguímicas de tres tipos de aguas de la Cuenca AmazónicaICuadro 3. Resumen de las lagunas en estudio~uadro 4. Lista de las especies de hemípteros acuáticos identificadosCuadro 5. Tabla de géneros y especies identificadas en las 144 muestrasCuadro 6. Prueba T de Student: Época~~!:~dro 7. Test de Student: Laguna
[!:uadro 8. Test de Student: Vegetación
vi

L- In troducción
La cuenca amazónica es una de las más importantes redes hídricas del mundo, ocupa el
(j()";, de la superficie total del territorio boliviano, se encuentra caracterizada por una gran
llanura central con un clima tropical y una estación de lluvias concentradas entre los meses
de octubre y marzo (Montes de Oca 1997).
F:-;t;l:) extensas llanuras constituyen sistemas complejos que involucran generalmente varios
ecosistemas, de ahí que es necesario considerarlos como macrosistemas que comprenden
ambicntes acuáticos temporarios y sectores de tierra firme (Neiff 1996).
En este tipo de ambiente se denotan épocas marcadas de inundación, transición y estiaje,
siendo los organismos de las planicies inundables condicionados por las sequías y en menor
gr:ldo por las inundaciones extremas (Neiff 1990).
1\11":1 un mejor estudio de estos ambientes, es importante determinar los factores que
influyen sobre sus comunidades bióticas, en este caso la composición cualitativa y
cuantitativa de macro invertebrados bentónicos. La interpretación de la distribución de estos
corno componentes de la red trófica, permitirá obtener información y generará propuestas
sobre estrategias de conservación y manejo sostenible de los recursos disponibles.
El trabajo realizado por el equipo mOCAB (Pouilly el al 2004), sobre la biodiversidad
acuática en la cuenca amazónica boliviana, ha permitido obtener información inicial sobre
la ecología de las comunidades bióticas y su interrelación con los factores abióticos. Al
interior de este programa, los macroinvertebrados bentónicos fueron el grupo menos
estudiado. Con este trabajo se presentarán datos complementarios respecto a la taxonomía y
el papel ecológico, en función a la vegetación y su variación en el tiempo y espacio.
Este estudio como también el proyecto actual que ejecuta el programa "Estudio del proceso
de concentración y bioaculumación del Mercurio y Metilmercurio en la estructura trófica
. mediante la utilización de isótopos estables de 813C y 815N en las principales subcuencas
1

de la Amazonia boliviana", forman parte de una línea base de investigación en la
composición de la entomofauna acuática de esta región, y principalmente en particular de
los hemípteros acuáticos, relacionándolo con la temporalidad y los factores físicos y
químicos.
Por tanto, surgen las siguientes preguntas: ¿Existe influencia en la composición faunística
cle la comunidad de hemípteros acuáticos respecto de la ubicación espacial de las lagunas
en estudio", ¿El factor época (inundación, transición y estiaje) influye en la composición y
distribución de la comunidad de hemípteros acuáticos?, ¿Existe asociación de las familias,
los géneros y especies acuáticas del orden Hemiptera con la vegetación acuática existente
en las lagunas en estudio?
Este trabajo de investigación está dentro el Proyecto "Variabilidad Medio Ambiental y
Estrategias en las Comunidades Acuáticas en la llanura de inundación del Beni," que se
desarrolla en coordinación con la Unidad de limnología IE-UMSA y el Instituto de
Investigación para el Desarrollo (rRD) de Francia.
2

1.1 objetivos
1.1.1. Objetivo general
Estudiar la ecología y distribución de la comunidad de hemípteros acuáticos en cuatro
lagunas de la llanura de inundación del Beni.
1.1.2. Obj etivos específicos
• Elaborar una lista faunística de la comunidad de hemípteros acuáticos de las
llanuras de inundación de Trinidad-Beni.
• Determinar si existe relación entre la época y la abundancia de la comunidad de
hemípteros acuáticos en los diferentes ambientes de la llanura de inundación de
Trinidad-Beni.
• Determinar si existe relación entre las lagunas (por su ubicación espacial) y la
abundancia de la comunidad de hemípteros acuáticos en los diferentes ambientes de
la llanura de inundación de Trinidad-Beni.
• Determinar si existe asociación entre la vegetación del entorno de las lagunas
estudiadas con la distribución y ecología de la comunidad de hemípteros acuáticos.
3

2. Fundamento teórico
2.1 Funcionamiento ecológico de ecosistemas acuáticos tropicales
Los sistemas con pulsos de inundación (como. ríos y humedales) se comportan como
sistemas en permanente desequilibrio debido al incremento y decremento de aguas, en los
L111e la variabilidad temporal es una compleja función dependiente de las entradas y salidas
de energía y materiales en distintos sitios de la cuenca y de las oportunidades combinatorias
de las poblaciones en la misma para ajustar su distribución y abundancia (Neiff 1990).
1.~lS car-acterísticas de los grandes humedales están esencialmente descritas por la capacidad
de transformación interna de estímulos que ellos tienen, en el espacio geográfico que
ocupan los humedales y sobre los sistemas de los cuales reciben su influencia (Holland
1(88).
la 1lanura de inundación corresponde a un tipo de gran humedal. En ella el encharcamiento
del suelo proviene en gran medida de desbordes fluviales, de cauces cuyos caudales se
origi nan en otras regiones. En consecuencia, el cambio energético del sistema se origina
rundameotalmcntc de procesos ocurridos en sectores alejados, por lo cual, el cambio de
estado del sistema puede tener uno o varios meses de destiempo o desfasaje respecto del
área de origen del estímulo meteorológico (Neiff 1996).
Los sedimentas son dominantemente finos (arenas muy finas, limos y arcillas); esto resulta
en una disponibilidad claramente mayor de nutrientes y otros solutos, y en una mayor
salinidad total. Las inundaciones y la mayor movilidad del agua genera el ingreso y/o
translocación importante de materiales, solutas, sólidos suspendidos y materia orgánica
coloidal (Lesack 1993).
4

2.1.1 El régimen pulsátil de los ecosistemas
Iunk el al. (1989), explican que los pulsos de inundación representan la principal fuerza
que determina y controla la bioproductividad de un sistema, desarrollada fundamentalmente
en las planicies de inundación y son responsables en gran medida de la organización biótica
en lagunas o ríos con llanura de inundación. Se encuentran eventos periódicos de
inundación que producen situaciones de estrés biótico que se reflejan en un nuevo
comienzo del sistema. Bonetto (1976) indica que las inundaciones producen "procesos de
rcjuvenecirniento" de los ecosistemas.
[n ríos y humedales con movimiento horizontal del agua, los flujos de energía y de
materiales ocurren como pulsos con fases de inundación y de sequía (Neiff 1996).
Durante la fase de inundación, los cuerpos de agua de la planicie (lagunas o meandros
abandonados) se interconectan por el flujo del río, se producen flujos horizontales (agua,
sedimentos, minerales, organismos) desde el curso del río hacia la planicie de inundación,
se recibe de esta inundación materiales en suspensión y a su vez aportan al agua materia
orgánica y minerales del suelo.
La fase de sequía tiene flujo de materiales desde la planicie hacia el río y se produce el
aislamiento paulatino de los cuerpos de agua de la planicie y aún su extinción temporal,
hasta una nueva fase de inundación. El patrón de variabilidad de estas ondas en una
secuencia temporal conforman el régimen pulsátil (Neiff 1990).
Los requerimientos para que algunos organismos se encuentren presentes, están en relación
con el tiempo de vida. Como consecuencia, es habitual encontrar agrupaciones de
organismos que tienen adaptaciones individuales y patrones poblacionales mejor ajustados
él la variabilidad hidrométrica en diferentes sectores de la planicie de inundación (Drago
1994; Orfeo 1995).

/\lguI10S organismos están predominantemente condicionadas por la época de inundación,
otros por la época de estiaje (son los denominados estrategas de fase o época), mientras que
otros han sido favorecidos por su capacidad de adecuación a una amplia gama de
condiciones del régimen pulsátil (Neiffel al. 1994).
2.1.2 Consecuencias ecológicas del régimen pulsátil
En los ríos que presentan planicie de inundación es posible encontrar complejidad creciente
de organización en las comunidades, desde el curso del río al borde externo de la planicie.
Se describe una zonación típica con incremento en la complejidad (cantidad de especies,
diversidad específica, nichos tróficos) desde el curso principal del río a los canales
secundarios de escurrimiento (Marchese & Ezcurra de Drago 1992).
Las biocenosis de los grandes ríos están reguladas por la hidrodinámica de pulsos, pero las
fases de aguas bajas son tan importantes como las inundaciones (Neiff 1990; Neiff el al.
¡994). Durante esta fase seca, las plantas sufren estrés que produce la interrupción del
crecimiento y la abscisión de las hojas (Neiff y Poi de Neiff 1990). La mayoría de las
poblaciones de peces no pueden sobrevivir debido a la carencia de organismos para
alimentarse como también sufren importantes pérdidas durante las sequías prolongadas o la
diferencia de aguas.
Las inundaciones representan el mayor factor de cambio en la estructura biótica. Es preciso
enfatizar que la fase seca de los pulsos constituye un poderoso factor de selección que
condiciona la distribución y abundancia de animales, plantas e invertebrados acuáticos.
La vegetación arraigada de hojas flotantes que crecen en los humedales fluviales tiene
ecofenos propios de las fases de inundación y de suelo seco (Junk 1970; Neiff 1978).
Durante el período crítico de inundación las plantas aceleran el crecimiento y se adaptan,
pero las plantas desaparecen si la sequía es prolongada.
6

La información disponible para las llanuras de inundación de la región amazónica es
lragmentarin y no permite suficientes generalizaciones, además que la productividad
primaria y la descomposición de la materia orgánica están reguladas principalmente por el
régimen hidrológico que condiciona la velocidad de procesos, las condiciones de máxima
circulación de la energía y el balance entre acumulación y transferencia de elementos
dentro del sistema.
2.1.4 Teorías de perturbación
Las perturbaciones son eventos discretos en el tiempo, que remueven a los organismos
abriendo nuevos espacios que pueden ser colonizados por las mismas o diferentes especies.
Las comunidades ecológicas no son estables porque se encuentran continuamente sujetas a
cambios aleatorios o perturbaciones. Los efectos de perturbación hacen que una comunidad
vuelva a su estado anterior de sucesión, donde el espacio abierto es colonizado por una o
más especies oportunistas (figura 1A), en las primeras etapas de sucesión, a medida que
pasa el tiempo invaden mas especies ese espacio abierto (Figura 1B Y 1C), convirtiéndose
estas ultimas en dominantes al avanzar su estado de madurez (Figura 10). En esta
secuencia la diversidad empieza con un bajo nivel, aumenta hacia las etapas intermedias de
la sucesión y disminuye de nuevo cuando se produce la exclusión competitiva ejercida por
las nuevas especies (Begon el al. 1988 cit en Rocabado 2004).
a b e d
Figura 1. Sucesión hipotética en un espacio vacío. (Begon el al. 1988 cit. en Recabado
2004).
7

Existe una hipótesis de perturbación intermedia que plantea que la alta diversidad biótica
resulta de cierto nivel de perturbación. Es decir, la diversidad mas elevada es mantenida por
los niveles intermedios de perturbación y se incrementa si existe el tiempo necesario para
que las otras especies invadan el espacio (Connel\ 1978, cito en Ward 1992, Figura 2).
Diversidadbiólica
Frecuencia de perturbaciónIntensidad de perturbación
Prediclibilidad de perturbaciónInestabilidad de la predación
Variabilidad del recurso
Figura 2. Relación teórica entre la diversidad biótica y varios medios de perturbación
(modificado de Ward & Stanford 1983)
2.2 Generalidudes del orden Hemiptera
Según Schuh (1986), los hemípteros acuáticos y semi acuáticos se encuentran comprendidos
en dos grandes grupos: Neoheteroptera y Euheteroptera (Cuadro. 1).
NEOHETEROPTERAGerromorpha
MesoveliidaeHebridae
ParaphrynoveiidaeMacroveliídaeHydrometridae
HermatobatidaeVeliidaeGerridae
EUHETEROPTERANepomorpha
BelostomatidaeNepidae
GelastocoridaeOchteridaeCorixidae
PotamocoridaeNaucoridae
AphelocheridaeNotonectidae
PleidaeHelotrephidae
Cuadro 1. Familias que comprenden los grupos Gerromorpha y Nepomorpha.
8

La talla es muy variable, encontrándose desde 1 mm en la familia Pleidae, hasta más de 100
a 120 cm como Belostomatidae (De Carla 1964; Schnack 1976). Su coloración puede se..
parda o negruzca con manchas irregulares ylo puntiformes. También puede existir un
patrón formado por pequeñas manchas oscuras, castañas o negras, que alterna con un fondo
parduzco o amarillento más claro (López Ruf 1994).
Presentan un cuerpo dividido en tres partes, cabeza, tórax y abdomen, el cuerpo es aplanado
dorsoventralmente y presenta en conjunto aspecto deprimido. La mayoría es de contorno
ovalado (Figura 3 A Y B), con la cabeza acuminada anteriormente como Naucoridae.
.'\Igunos son muy alargados y cilíndricos con patas muy largas y finas como Ncpidae,
lIydrometridae, Veliidae y Mesoveliidae (Pennak 1953; Nieto Nafria & Mier Durante
10X5; Richards & Davies 1983).
PRONOTO [
fiEMIEUTRO
A
\\\ /\ ¡
\1
\.-,
ESTERNITOSABDOMINALES
B
TORAX
ABDOMEN
Figura 3. Vista general de un hemíptero acuático. A: Vista dorsal B: Vista Ventral
(Bachrnann 1981).
9

a) Cabeza
Los hemípteros acuáticos presentan ojos compuestos y su gran mayoría carecen de ocelos,
pero en algunos semiacuáticos los ocelos están presentes solo en individuos macrópteros
(l.ópcz Ruf 1994).
Tienen transformado el aparato bucal en el rostro, el cual está constituido por el labio
formando un tubo, segmentado, donde se mueven las maxilas y las mandíbulas
transformadas en estiletes y por el cual atraviesa la hipofaringe (Figura 4A). En la figura
~B, se observa un esquema de la cabeza con las palies que diferencian en medida o forma
algunas familias o géneros.
En adultos las antenas no tienen nunca más de cinco artejos. En las familias subacuáticas
(Nepornorpha) están escondidas en fosetas y ubicadas en un reborde inferior a los lados de
la cabeza; los artejos son tan cortos que la mayoría dan aspecto baciliforme. En algunos
grupos semiacuáticos (Gerromorpha), cuyos miembros no efectúan incursiones por debajo
ele la superficie del agua, las antenas son más largas y expuestas con un aspecto lineal
ESTILETEMANDIBULAR
quebrado (Hungerford 1959).
..-------<,
/..<J ... \YNTENAti (1 (~i'<¿./~V··~./ CLlPEO
\." !?'"" ~/ LABR\/~
f .. II )
tI)VESTILETE MAXILAR
1j1 A B
Figura 4. A: Estructura del aparato bucal; B: Esquema en vista dorsal de las medidas
de la cabeza. V: Vcrtcx, S: Sintlipsis; O: Borde posterior del ojo; A: Ancho del disco
pronotal; L: Longitud del disco pronotaI.
10

h) Tórax
1] prenoto es grande, el mesotórax esta más desarrollado que el metatórax. Al nivel del
mesotórax se observa una pieza triangular, el escudete. Las patas son de formas variadas
según los diferentes grupos y según su posición. El tarso comprende de uno a tres
segmentos en los adultos y siempre de un solo segmento en las larvas, con una o dos uñas
tarsalcs,
Algunas familias presentan el tercer par de patas aplanadas y provistas de franjas ele setas
1:1Ci Iitando esto la natación. En algunos el primer par de patas son raptoras dependiendo
mucho esto del hábitat y la dieta. En otros el tarso del primer par de patas esta constituido
por un solo segmento, formando una paleta que permite la recolecta de rnicropartículas
orgánicas o algas microscópicas. (Bachmann ]98]).
Los Gcrromorpha no predadores presentan el primer par de patas no raptor y los dos pares
restantes caminadores. Algunas familias presentan los últimos pares de patas no adaptados
para la caminata, sino para patinar sobre la superficie del agua, los Gerromorpha
predadores presentan el primer par de patas raptor y los demás pares de patas caminadores
no estando adaptados para patinar sobre la película de agua pero si para la natación (Lopez
Ruf 1994).
El primer par de alas se encuentra casi siempre modificado en hemielitros. Se pueden
distinguir varias áreas, pero básicamente se trata de una porción proximal esclerotizada y
una distal, casi siempre mas pequeña y membranosa. En ningún caso se pliegan, solo se
rebaten sobre el abdomen estando en reposo y sus porciones membranosas se superponen
más o menos extensamente (Schuh 1986, Figura 5).
11

M
Figura 5 Hcmiclitro derecho. Abreviaciones: CL: Clavo; el: longitud del arca
pruinosa de la sutura elaval; CO: Corio; E: Surco embolar; M: Membrana; N: Nodo;
P: Longitud del arca pruinosa postnodal; SM: Sutura membranal.
Las alas del segundo par son membranosas y presentan una nerviación muy simplificada,
con fusión y desaparición de nervaduras longitudinales. Las nerviaciones transversales son
muy escasas, las alas se pliegan por debajo de los hemielitros (Richards & Davies 1984).
En la mayor parte de los grupos aparecen especies con polimorfismo alar, hecho que parece
ser desencadenado por los factores ambientales. Pueden ser macrópteros, braquípteros,
micróptcros o ápteros. Los macrópteros son capaces de realizar vuelos casi siempre cortos,
10 cual es de mucho valor en la dispersión y la supervivencia cuando las lluvias escasean o
son muy estacionales (Tachet el al. 2000).
12

e) Abdomen
Los hemípteros acuáticos en su mayoría presentan de diez segmentos abdominales, pero
exteriormente se observan solo cinco o seis, ya que dos o tres se invaginan para constituir la
. armadurn genital. El primer urito completo y el esternito del segundo segmento, suelen
estar reducidos y no se observan con facilidad. Los órganos genitales externos masculinos
son de configuración diversa, siendo utilizados en las clasificaciones taxonómicas. El
noveno urosternito forma la mayor parte de la cápsula genital o pigóforo, en el fondo del
cual se encuentra la inserción del falo y a cada lado los parámeros (Lopez Ruf 1994).
Los genitales femeninos tienen una simplificación del ovopositor teniéndolo diferenciado,
comprimido con tres pares de valvas y el ovopositor en placa deprimido y con uno o dos
Ixm:s de valvas membranosas (Schuh 1986).
2.3 Características anatómicas internas en Hemiptera
Los hemípteros acuáticos en el sistema digestivo presentan dos glándulas salivales labiales
produciendo una saliva tóxica que actúa en conjunto con los estiletes en el caso de
depredadores. Se observan glándulas maxilares, ponzoñosas en algunos casos y lubricantes
en otros. Por ser fluidofilos realizan la succión por acción de la bomba cibarial, la faríngea
o ambas, las que presentan potentes músculos para tal fin.
Presentan un mesodco tubular pero sin zonas diferenciadas en el estomodeo. En la parte
posterior del mesodeo se encuentran cuatro pares de tubos de Malpighi. El proctodeo es
corto y termina en una bolsa rectal (Jansson 1972).
El sistema reproductor en machos presenta de dos a ocho folículos testiculares agrupándose
en dos o más paquetes a cada lado, las hembras presentan ovarios con la misma cantidad de
ovariolas acrotróficas que en machos (Schuh 1986).
13

En el sistema nervioso de los hemípteros se observa un cerebro relativamente retrasado,
debido al desarrollo del cibario (relacionado a la succión), estando los ganglios de la cadena
nerviosa ventral muy condensados. El sistema nervioso es diferente entre familias (Lopez
Ruf 1994).
La respiración en hemípteros acuáticos es muy simple, presentando un sistema abierto en el
cual entra aire en las tráqueas a través de uno o varios espiráculos, habiéndose perdido el
mecanismo de cierre y apertura de espiráculos en muchos grupos, reduciéndose simples
aberturas. Algunos pueden poseer estructuras sumamente complicadas habiendo cámaras
I1lUY ramificadas en forma de rosetas o tramas de trabéculas (Jansson 1972).
En Ncpomorpha se observan 5 ciclos larvarios. En estadios 1 y 2 la respiración se realiza
directamente a través del tegumento, en los otros estadios por medio de espiráculos
presentes en diferentes partes del cuerpo. Los Gerromorpha igualmente cercanos al agua
viven en la superficie (neuston) o cerca de ella y no fuera del agua. La respiración en los
diferentes estadios es similar a Nepomorpha (Tachet el al. 2000).
l.os heterópteros acuáticos son insectos paurometábolos con coexistencia de larvas y
adultos (Tachet el al 2000), tienen estadios juveniles denominados ninfas o larvas, los
cuales viven y se alimentan del mismo modo que los adultos (Lopez Ruf 1994). Estos no
tienen desarrollo de alas (ápteros) como también carecen de órganos genitales (Hungerford
1919).
2.4 Consideraciones ecológicas
Los Hemípteros acuáticos son un grupo muy diverso, algunos ocupan exclusivamente la
superficie del agua o pueden estar posados en las hojas que se encuentran por encima cerca
de la película de agua (Figura 6A), otros son estrictamente acuáticos, habitando entre las
plantas existentes (Figura 6B) y los últimos habitan en bordes barrosos o arenosos, en las
orillas como en el fondo de los cuerpos de agua (Figura 6C, Bachmann 1981).
14

- -
~c -- ~ _--~-M~
Figura 6. Hábitat de los hemípteros acuáticos: A) Verdaderamente acuáticos,
B),Superficiales C) De orilla y fondo.
La familias verdaderamente acuáticas o bentonicas son Pleidae, Notonectidae, Corixidae,
l lclotrcphidae, Nepidae, Naucoridae, Belostomatidae (Figura 6A), las familias:
llydrometridae, Mesoveliidae y Veliidae son superficiales o patinadoras encontrándose en
la superficie de la película de agua (Figura 6B); Ochteridae, Gelastocoridae y Hebridae son
hemípteros semiacuáticos encontrándose entre algas flotantes, en el fondo del sustrato o
rondando la interfase tierra-agua (Figura 6C, Lopez Ruf 1994).
Los hemípteros acuáticos pueden abandonar temporalmente los cuerpos de agua, son
mayormente depredadores sobre diversas presas, peces, artrópodos, moluscos, anfibios,
como oligoquetos y pequeños invertebrados, pero ciertos géneros son detritívoros o
consumen únicamente algas microscópicas (Mazzuconi & Bachmann 1995).
15
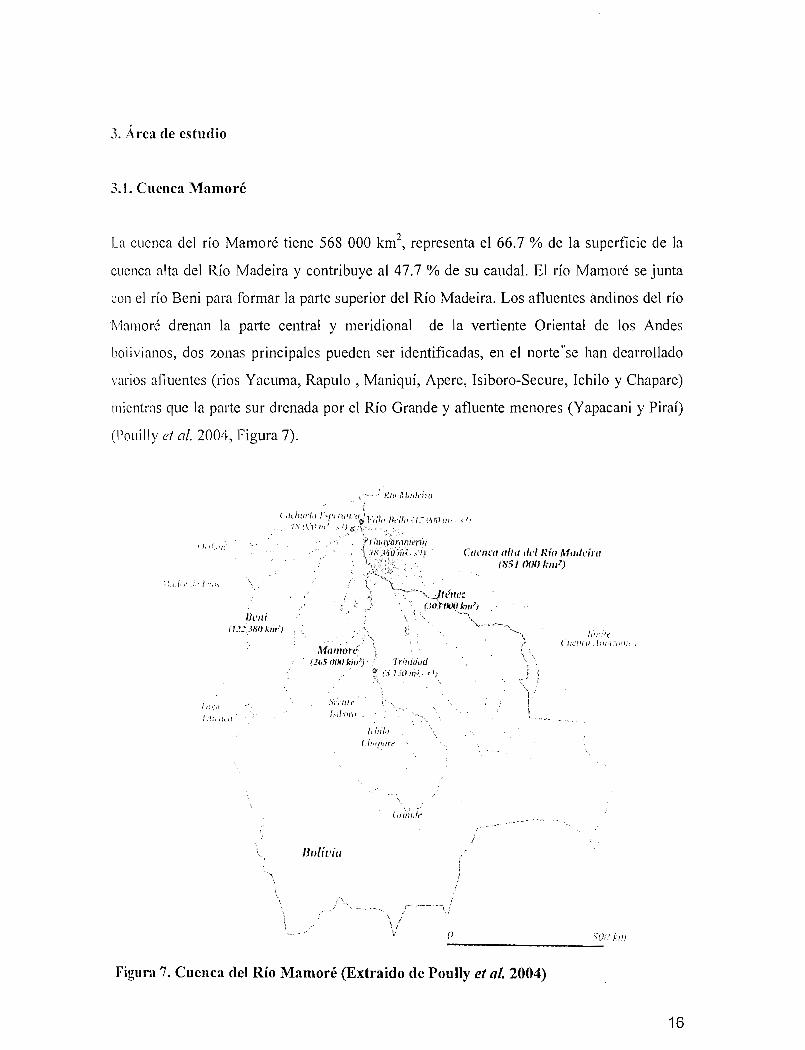
3. Área de estudio
3.1. Cuenca Mamoré
La cuenca del río Mamaré tiene 568 000 km", representa el 66.7 % de la superficie de la
cuenca alta del Río Madeira y contribuye al 47.7 % de su caudal. El río Mamaré se junta
con el río Beni para formar la parte superior del Río Madeira. Los afluentes andinos del río
Mamaré drenan la parte central y meridional de la vertiente Oriental de los Andes
bolivianos, dos zonas principales pueden ser identificadas, en el nortese han dearrollado
varios afluentes (rios Yacuma, Rapulo , Maniquí, Apere, Isiboro-Secure, Ichilo y Chapare)
mientras que la parte sur drenada por el Río Grande y afluente menores (Yapacani y Piraí)
(Pouilly el a!. 2004, figura 7).
! ,;<":'>1
lIclli(1.~:'.38()km.')
MOl/la,.,,:: •. (lr,S ()r~) k;,;') cÓ. Trinidad
? (31:ftJ nj·;. f"1/{
t, luloI.IMPorr"
'.
Bolio¡«
Cuenca alta dr! Rio Miutcira(S,'; I n(Jo (",,2)
lP::;"r:'( uctu « .'~m'!:·('¡¡¡: j
l·"
5f tn 1:111-------_.:......Figura 7. Cuenca del Río Mamoré (Extraido de Poully el al. 2004)
16

3.1.1 Clima y régimen hidrológico
El clima de la zona se encuentra clasificado como tropical, isotérmico y húmedo
presentando temperaturas medias anules entre 24 y 26 o e y una humedad relativa ambiente
ele 76 % promedio anual (Salm & Flores 1994), la estación de lluvias se encuentra
concentrada en verano ocurriendo el 60 - 80 % de las precipitaciones entre diciembre y
marzo como también las temperaturas altas (Hanagarth 1993; Montes de Oca 1997).
Los niveles de agua mas importantes son registrados entre los meses de.. enero y marzo, sin
embargo las precipitaciones y las temperaturas decrecen en un eje noreste-sureste paralelo
al eje principal de los Andes.
El patrón de distribución de las lluvias depende de los movimientos de masas de aire,
puesto que durante el verano los vientos provienen del noreste llegando cargados de
humedad del Atlántico y del bosque tropical de la Amazonia central. Durante el invierno
10s vientos del norte son menos intensos, pero algunas veces vienen superados por los
vientos polares procedentes del sur del continente y canalizado en los Andes. Estos vientos
fríos y secos generan los fenómenos de "surazos", caracterizados por durar poco y por un
descenso abrupto de la temperatura (Pouilly el al. 2004).
3.1.2 Habitats acuáticos de la región del río Mamoré
Los medios acuáticos del Mamaré se distinguen tanto por sus propiedades abióticas como
por la forma de sus habitats y la calidad de sus aguas (Loubens el al. 1992), como por sus
características de vegetación que también están influenciadas de manera directa o indirecta
porel régimen hidrológico.
Los medios acuáticos lóticos de la región del Mamaré corresponden en particular a dos
tipos de ríos (Loubens el al. 1992):
17

• Río Mamaré, eje central y componente principal del sistema que drena las aguas
blancas provenientes de los Andes.
o Otros ríos que confluyen con el Mamaré drenando las aguas locales de la llanura,
como ser: Ibare, Tejamuchi, Matiquipiri, presentando aguas de calidad variable,
turbias, con un pl-l y una conductividad menores que el Mamaré.
Los medios acuáticos lénticos corresponden a un amplio rango de condiciones (cuerpos de
aguas temporales y permanentes), de esto Loubens el al. (1992) distingue tres tipos
principales de habitats lénticos: las lagunas de depresión, formadas por diferencias de
. acumulación de los sedimentos en el bosque galería, las lagunas de meandro, antiguos
cauces abandonados por el río Mamaré asociados al bosque galería y las lagunas de sabana,
correspondiendo a una depresión topográfica probablemente debidas a causas geológicas y
antrópicas. Las lagunas de sabana se encuentran en contacto con la capa freática superficial
(0.5 a 2 m de profundidad), lo que asegura un nivel de agua estable en el transcurso del año,
a pesar de la gran estacionalidad de las precipitaciones.
3. t.3 Características fisicoquímicas de las aguas
De acuerdo con la clasificación de Sioli (1968) para los sistemas acuáticos de la Amazonia
central se diferencian tres tipos de aguas (Cuadro 2):
Cuadro 2. Características fisicoquímicas de tres tipos de aguas de la Cuenca
Amazónica (Sioli 1968 y Moyle & Cech 1996, cit. en PouilIy el al. 2004)
Parámetro Aguas Blancas Aguas Claras Aguas Negras
pH 6.2 -7.2 4.5 -7.8 3.8 -4.9
Transparencia (m) 0,1 -0,5 1.0 - 4.3 1.3 - 2.3
Turbidez Alta Baja Baja
O2 disuelto Moderado Alto Bajo
Nutrientes Alto Bajo Escaso
18

Aguas claras, provenientes de las zonas cristalinas de los escudos precámbricos brasileño y
guyanerise, aguas que contienen alta transparencia, pH y una tasa de oxigeno variable.
Aguas blancas, reconocidas por un color café mezclado con blanco, aguas drenadas de los
ríos de los Andes. El color de las aguas está relacionado con el proceso de erosión, ya que
éstas vienen cargadas de una gran cantidad de materia en suspensión y nutrientes. Baja
transparencia, alta turbidez y pH neutro.
Aguas negras, estás aguas arrastran ácidos húmicos procedentes de la descomposición de
la materia orgánica dándole una coloración característica de café a negro, pH ácido,
transparencia intermedia y pobre en nutrientes que provienen de la degradación de la
materia alóctona.
Corbin el al. (1988), en un estudio en el Mamaré central, hizo un aporte importante sobre
las características fisicoquímicas de varios cuerpos de agua, proponiendo una
categorización en tres grupos:
• Aguas de planicie, presentando importantes cantidades de hierro y potasio, poco
mineralizadas de tipo bicarbonato sódico.
• Aguas de ongen andino, son aguas blancas mineralizadas por bicarbonatos de
calcio, magnesio y sulfatos.
• Aguas intermedias, éstas se encuentran influenciadas por aguas blancas yaguas de
planicie.
3.1.4 Gradientes medioambientales de las lagunas de bosque de galería y de sabana
Los cambios de los caudales del Río Mamaré influyen sobre su forma, así los brazos
abandonados generalmente se encuentran cerca del cauce principal, presentando una
19

importante profundidad (10 a 15 m en época seca), en el transcurso de años estas lagunas
sufren cambios morfológicos relacionados al proceso de co1matación, por la deposición de
sedimentos y favoreciendo a la colonización de la vegetación (Pouilly el al. 2004).
Mientras el lecho principal sigue migrando, las lagunas abandonadas envejecen y se
encuentran sometidas a fluctuaciones del nivel del agua de menor importancia, esto
asociado a la dinámica hidrológica local genera también variación en la calidad de aguas,
esto depende si son alimentadas todavía por las aguas blancas del lecho principal o aguas
provenientes de llanura por escorrentía. (Pouil1y el al. 2004).
Así, en el sistema de llanura de inundación, la alimentación de una laguna puede cambiar
ocurriendo inundaciones por dos procesos complementarios: proceso exógeno y endógeno.
El proceso exógeno se inicia con la llegada de la onda de crecida de las aguas blancas del
Mamaré, estas aguas blancas se expanden en la llanura de inundación. El proccso endógeno
ocurre con el desborde de las capas freáticas alimentadas por las precipitaciones locales
(Bourrel el al. 1999).
Por esto, la calidad de las aguas de las lagunas cercanas al Mamaré está ligada a la
intensidad de ambos procesos y el balance entre las precipitaciones de las cuencas altas
andinas (aguas de origen andino) y las lluvias locales (aguas de planicie).
3.2. Macrófitas
Existen muchas comunidades acuáticas y palustres que se desarrollan en los márgenes de
las lagunas, algunas forman extensos colchones de vegetación arraigada o Dotante, estas
especies vegetales que flotan libremente se encuentran adaptadas a aguas ricas en nutrientes
no apareciendo en condiciones ácidas ni pobres de nutrientes (Junk el al. 1989).
Así, cuando los niveles de nutrientes son suficientes (ríos de aguas blancas), gran cantidad
de macrófitas puede establecerse en los canales de los ríos y las lagunas, por 10 tanto, en
lagunas conectadas directamente con el río Mamaré, se desarrollan comunidades flotantes
20

dominadas por Eichhornia crassipes en áreas donde la velocidad de la corriente es baja
(Sanjinés & Beck 2004 cito en Pouilly el al. 2004).
En las lagunas de bosque galería, es probable que la profundidad pueda afectar a las
comunidades de plantas acuáticas. Según Junk el al. (1984), ésto puede deberse por niveles
bajos de nutrientes, en lagunas aisladas y no influenciadas por las aguas blancas del
Marnoré. Por 10 tanto, no reciben ningún tipo de aporte de nutrientes que favorezca al
desarrollo de comunidades acuáticas más diversas.
Las lagunas inundadas por aguas blancas pueden presentar cambios estacionales en la
vegetación ya que las crecidas favorecen al desarrollo de la vegetación flotante, en el
periodo de estiaje, esta vegetación acuática muere y especies terrestres ocupan S\.l lugar.
Cuando el nivel de agua asciende vuelven las especies acuáticas a colonizar el ambiente
desarrollándose a partir de semillas o individuos que han sobrevivido a la época seca.
. Muchas de estas especies presentan gran capacidad de reproducción y tolerancia a las
fluctuaciones del agua (Puhakka & Kalliola cit. en Pouilly el al. 2004).
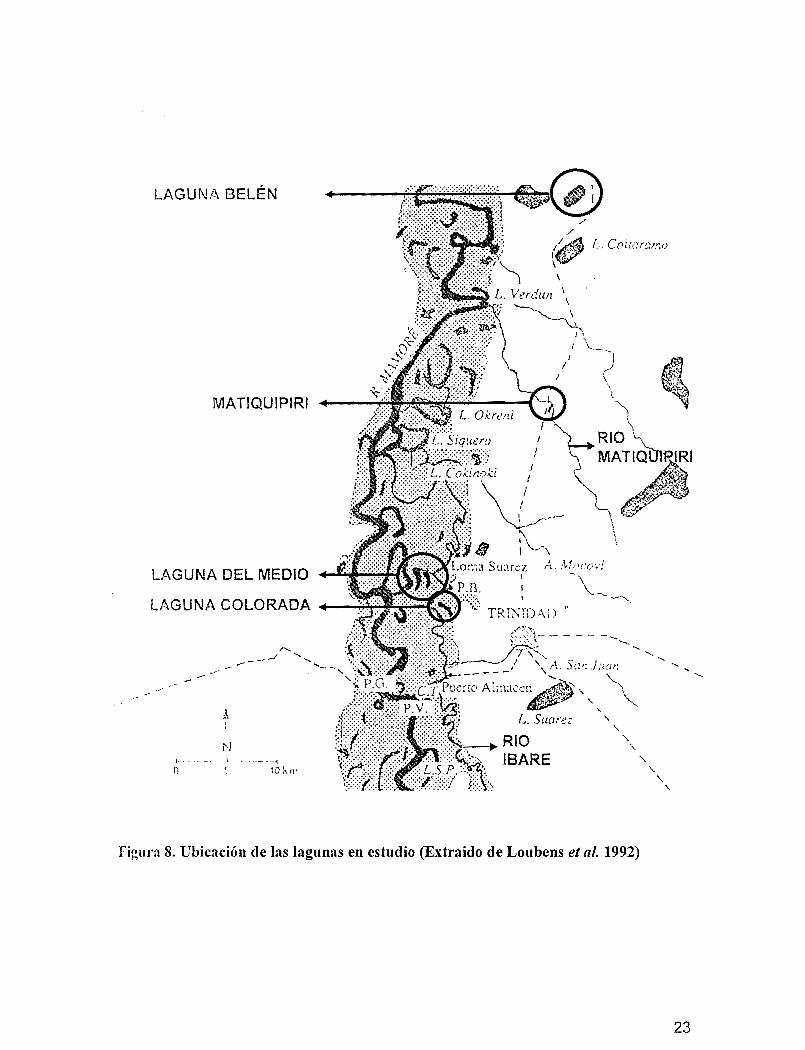
3.3. Ubicación de las lagunas en estudio
Las lagunas seleccionadas para la presente investigación son representativas por SllS
caractertsticas físicas y químicas, como también su situación geográfica (Figura 8). Así se
tiene:
• La laguna del Medio (laguna de bosque), situada cerca del lecho principal y en
conexión directa y casi permanente con el río Mamaré, sin la influencia del hombre,
presenta vegetación reducida y moderada profundidad.
• La laguna Colorada (laguna de bosque), situada en el bosque galería. Se encuentra
alejada del canal activo. Esta laguna es influenciada por la inundación del río
Mamaré, pero también mantiene relación por canales de conexión entre lagunas yel
21

flujo de agua proveniente de la llanura. Está ubicada a 3 kilómetros
aproximadamente de Puerto Ballivián.
• La laguna Belén (laguna de sabana), con un área de 2 krrr', se encuentra situada a 15
km de Trinidad, desconectada del Mamoré, presenta aguas de" planicie que
corresponde a lluvias locales, presenta actividad ganadera en sus alrededores.
• Matiquipiri (sabana), es una zona pantanosa que alcanza aproximadamente poca
profundidad durante la época de aguas altas y mínima profundidad en la époea de
aguas bajas, vegetación permanente e influencia ganadera; recibiendo el aporte de
aguas de la planicie por acumulo de las lluvias.
Cuadro 3. Resumen de las lagunas en estudio
BELEN MATIQUIPIRI COLORADA DEL MEDIO
Sabana Sabana Bosque galería Bosque galería
Desconectada del Desconectada del Influenciada por el Influenciada por el
Mamoré Mamoré Mamoré Mamoré
Poca profundidad Poca profundidad Profunda 1 a 5 m Profunda 2 a 7 m
22
MATIQUIPIRI
LAGUN¡\ BELÉN
\.\.
\.\.
:~~RIOIBAREl .. -----1'
~ :0 km
!.J•••
n
LAGUNA DEL MEDIO
LAGUNA COLORADA ...---,;¡.;¡,
Figura 8. Ubicación de las lagunas en estudio (Extraido de Loubens el al. 1992)
23

-l. Materiales y métodos
4.1. Diseño de muestreo
Para el presente estudio se realizaron 144 colectas en diferentes épocas. La colecta de
muestras fue realizada en los meses de noviembre del 2002 (M. Gutierrez & F. Giban),
época de aguas bajas (seca), enero-febrero 2003 (O. Carrasco & F. Giban), época de aguas
altas (inundación), y abril-mayo (M. Gutierrez & F. Giban), época de transición.
/\ continuación, en el siguiente diagrama de flujo, se presenta el protocolo de muestreo y
trabajo realizado en laboratorio considerados en esta tesis (Figura 9).
DPARÁMETROS
B10TICOS
SEPARACiÓN E IDENTIFICACIONDE MUESTRAS
COLECTA DE INVERTEBRADOSMUESTREOS CUANTITATIVOS
CUALITATIVOSRED DE MANO 250 MICRAS
PARÁMETROSABIOTICOS
/ -.~C'S'CO~
l lPROFUNDIDAD
PH TRANSPARENCIACONDUCTIVIDAD TEMPERATURA
"-
\ 1/
IANÁLISIS ESTADISTICOS: MULTlVARIADOS:PROGRAMA ESTADISTICO ADE-4, TEST DE
'1
STUDENT
Figura 9. Diagrama de flujo del estudio
1ABUNDANCIA
RIQUEZA
1CARACTERíSTICAS:
BIOLOGICASECOLOGICAS
UBICUIDADESTACIONALlDADPREFERENCIAS
24

·U. Metodología para la colecta e identificación de invertebrados
Se realizaron muestreos cuantitativos-cualitativos mediante una red manual 250 micras y
colectas manuales para la vegetación acuática flotante. La conservación fue en alcohol al
70%. con la adición de tres gotas de glicerina. Se precauteló no estropear las alas ni algunas
estructuras frágiles de los hemípteros acuáticos hasta su determinación taxonómica en
laboratorio.
La separación de invertebrados de la materia orgánica se realizó a partir de tamices y pinzas
manualmente bajo un estereomicroscopio. Se utilizó la solución de Faurer para el montaje y
preservación de estructuras en placas microscópicas, mediante las cuales se determinó a
nivel especie. Las muestras fueron identificadas y corroboradas en los laboratorios de
entomología de las universidades de Buenos Aires y La Plata (Argentina) con la
colaboración del Dr. Axel Bachmann, la Dra. Silvia Mazzuconi y la Dra. Mónica Lopez
Rur.
. .t.3 Tratamientos de datos
4.3.1 Análisis multivariado
Para el presente trabajo el análisis multivariado de datos se realizó a partir del programa
ADEA para el estudio y representación gráfica de datos ecológicos (Chessel & Doledec
1996).
4.3.2 Análisis factorial de correspondencia (AFC)
En este análisis se clasifican Jos resultados en dos categorías particulares, una categoría son
las especies encontradas y la otra categoría son las estaciones o sitios de muestreo y la
época de colecta. Este análisis permite describir las relaciones entre las variables
considerando las diferencias existentes.
2S

Las contribuciones relativas se expresan en 111 000 e indican cuales son los valores que
construye el eje. La suma en estas contribuciones absolutas permite elegir los grupos más
importantes para cada eje, tomando en cuenta un limite, en nuestro caso el 3 % (Fossati el
al. 2002).
I~I valor propio representa el número de ejes principales en el anál isis factorial de
correspondencia siendo éste representado en un grafico de histogramas ordenado en forma
decreciente, son significativos únicamente los que decrecen linealmente, ya no es necesario
estudiar los ejes después del cambio de pendiente (Figura 10).
I---------~-----~~ --------~~---~I
! 17%15 %
Eje Valores Inerciapropios
1 8.8648 0.16552 8.0701 0.15073 7.0286 0.13124 6.2167 0.1161
~~~~~D;:JFigura 10. Estimación del número de ejes importantes. Histogramas de los valores
propios con dos ejes importantes.
Se clasificó a los individuos en estudio caracterizados por un número de variables
numéricas o nominales. Es un método descriptivo y predictivo que puede ser considerado
como una extensión de la regresión múltiple (Lebart el al. 1995)
Si se dispone de n individuos u observaciones descritas por un conjunto de p variables (XI,
X], ... , xr) y repartidas en q clases definidas a priori por la variable nominal q modalidades.
Con el análisis factorial de correspondencia se propone separar al nivel de clases q con la
ayuda de p variables explicativas. Además de resolver el problema de la afectación de los
individuos nuevos caracterizados por las p variables (Lebart el al. 1985).
26

En orden descriptivo podemos decir que las funciones lineares discriminantes sobre el
muestreo de tamaño n, son las combinaciones lineales de las variables explicativas (XI,
xc ..... xrJ donde los valores se separan a nivel de q clases. La clase de afectación de n
nuevos individuos esta descrita por las variables explicativas (Xl, X2, .. ·, xrJ. Ellas muestran
un problema de clasificación que consiste en construir las clases más homogéneas posibles
dentro de una muestra.
4.3.3. Prueba T de Student
La prueba t de Student como todos los estadísticos de contraste se basa en el cálculo de
estadísticos descriptivos previos: el número de observaciones, la media y la desviación
típica en cada grupo. A través de estos estadísticos previos se calcula el estadístico de
contraste experimental. Con la ayuda de unas tablas se obtiene a partir de dicho estadístico
el p-valor. Si p<O,OI se concluye que hay diferencia entre los dos tratamientos.
Las hipótesis o asunciones para poder aplicar la t de Student son que en cada grupo la
variable estudiada siga una distribución Normal y que la dispersión en ambos grupos sea
homogénea (hipótesis de homocedasticidad=igualdad de varianzas).
5. Resultados
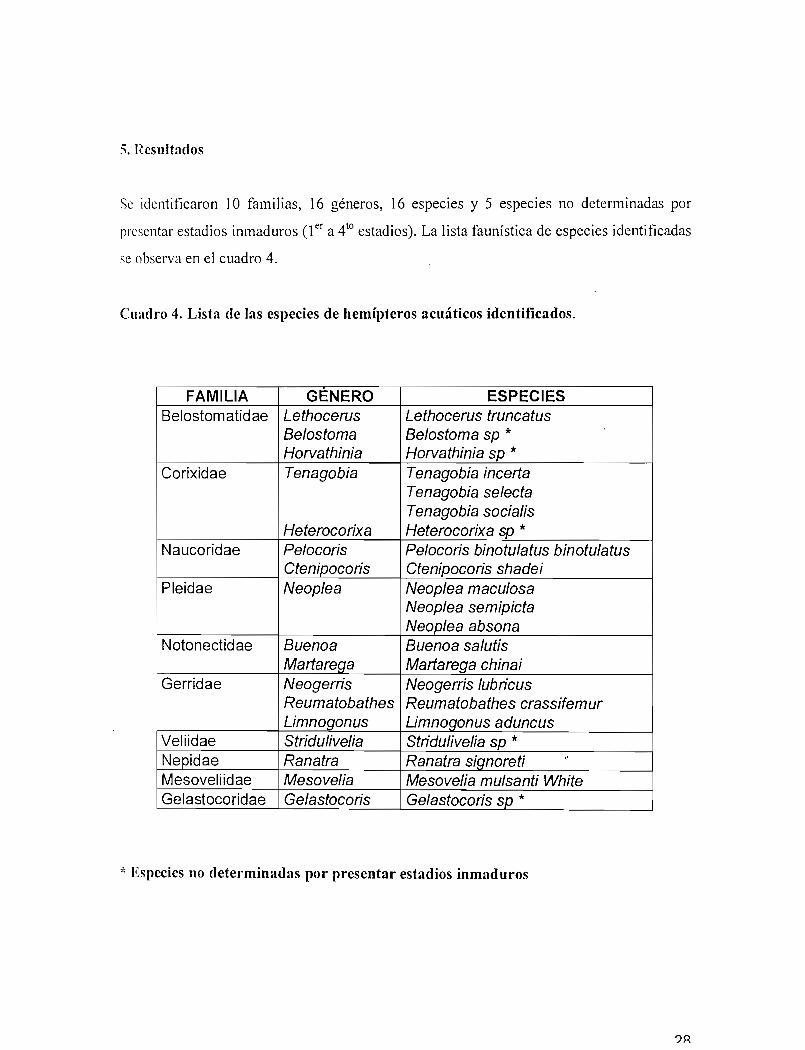
Se identificaron 10 familias, 16 géneros, 16 especies y 5 especies no determinadas por
presentar estadios inmaduros (1er a 410 estadios). La lista faunística de especies identificadas
se observa en el cuadro 4.
Cuadro 4. Lista de las especies de hemípteros acuáticos identificados.
FAMILIA GENERO ESPECIESBelostomatid ae Lethocerus Lethocerus truncatus
Be/ostoma Be/ostoma sp *Horvathinia Horvathinia sp *
Corixidae Tenagobia Tenagobia incertaTenagobia selectaTenagobia socia/is
Heterocorixa Heterocorixa sp *Naucoridae Pe/ocoris Pe/ocoris binotu/atus binotu/atus
Gtenipocoris Gtenipocoris shadeiPleidae Neop/ea Neop/ea macu/osa
Neop/ea semipictaNeop/ea absona
Notonectidae Buenoa Buenoa sa/utisMartarega Martarega chinai
Gerridae Neogerris Neogerris /ubricusReumatobathes Reumatobathes crassifemurLimnogonus Limnogonusaduncus
Veliidae Stridulive/ia Stridulivelia sp *Nepidae Ranatra Ranatra signoreti ..Mesoveliidae Mesovelia Mesovelia mu/santi WhiteGelastocoridae Ge/astocoris Ge/astocoris sp *
* Especies no determinadas por presentar estadios inmaduros
?R

5.1 Determinación de especies
El presente estudio contiene partes de claves sistemáticas para la separación de géneros y
especies. Puesto que mucha de la bibliografía referente no es de fácil acceso hasta la
identificación de especie, se ha reproducido en algunos casos algunas ilustraciones previa
aclaración.
a) Neop/ea maculosa (Berg 1879) FAM. PLEIDAEParámero derecho engrosado
con punta sinuosa
~,{ 1i~\•
Figura 11. Cápsula genital en macho y hembra de Neop/ea maculosa; A: Séptimo
urosternito de la hembra, B: 8vo urito macho, C: Pigóforo, parámero derecho y
parámcro izquierdo, D: Valva del ovopositor (Bachmann et al. 1994)
Presenta la longitud del cuerpo desde 1.9 a 2.6 milímetros presentando manchas difusas
castañas más o menos oscuras con carenas externales oscuras. La superficie de su cuerpo es
punteada principalmente sobre los hemielitros, se observa también que en estadios
inmaduros la puntuación y color difiere mucho. La determinación taxonómica más exacta
en machos es a partir del 8vo urito y pigóforo del macho (Figura IIA) y menos precisa
mediante el séptimo urosternito en la hembra (Figura l1B) con unaedeago curvo en vista
lateral, concavidad dorsal (antero-ventral en reposo), parámero derecho robusto terminado
en una punta sinuosa (Figura 11 C), en hembras las valvas del ovopositor con púas robustas
(Figura II O).
?q

Neoplea inaculosa fue primeramente descrita por Berg (1879), en Argentina; Kirkaldi
(1899) en Bolivia, Horvarth (1918) en Argentina y Paraguay como Plea maculosa y
redescrita en 1958 por Drake & Chapman. Fue descrita como Neoplea maculosa por por
Bachmann (1962) para Argentina, Paraguay y Bolivia.
b) Neoplea semipicta (Horvath 1918) FAM. PLEIDAE
A
Edeago
~L~
delgadas
B e D
Figura 12. Neoplea semipicta; A: Séptimo urosternito de la hembra, B: 8vo uritomacho, C: Pigóforo, parámero derecho y parámero izquierdo, D: Valva del ovopositor(Bachmann 1994).
Presenta una longitud pequeña de 1.5 a 1.8 milímetros, tiene como ancho máximo 0,8
milímetros, el color es oscuro con partes ventrales mas oscuras, con una forma ancha mas
que larga. Presenta la cabeza punteada, con el pronoto y los hemielitros profundamente
punteados, con un abdomen con carenas en decremento hacia atrás, 7mo urosternito y 8vo
urito y pigóforo para determinación mas precisa (Figura 12 A Y B), con aedeago grueso,
globoso, parárnero derecho alargado y el izquierdo en hoz con poca punta (Figura 12C),rt. valvas con púas delgadas en las hembras (Figura 12D) (Bachmann y Lopez Ruf 1994).~
f¡ Se encuentra distribuida al este de Bolivia, siendo el primer registro de esta especie descrito
t en el departamento de Santa Cruz, distribuida también en Argentina, Colombia y Paraguay.~
r

e) Neoplea absona (Drake & Chapman 1953) FAM. PLEIDAE
Pueslargas pocorobustas
':4
e
Edeago largo
~
&- .~{ '<; 2 .. -:.~-
-- ._-'"
BA
\
-:::::::::::::::::7.. ... .-.
'\~>
Figura 13. Cápsula genital en macho y hembra de la especie Neoplea absona; A:Séptimo urosternito de la hembra, B: 8vo urito macho, C: Pigóforo, parámeroderecho y parámero izquierdo, D: Valva del ovopositor (Bachmann 1994).
Presenta una longitud de 1.6 a 1.75 milímetros y un milímetro de ancho máximo, forma
angosta, convexa poco arqueada, se observa en las muestras colectadas en este trabajo un
mayor presencia de hembras (por confirmar con posteriores trabajos en machos), presentan
un 7mo urosternito característico en hembras (Figura 13 A), un 8vo urito en el macho poco
simétrico (Figura 13 B), presentando un aAaedeago delgado y largo con una base robusta
(Figura 13 C) y con las valvas del ovopositor púas largas poco robustas (Figura 13D,
Bachmann y Lopez Ruf 1994).
Según Bachmann & Lopez Ruf (1994), la descripción realizada es únicamente para la
Argentina siendo la especie todavía por confirmar para Bolivia.
31

ti) Tenagobia incerta (Lundblad 1929) FAM. CORIXIDAE
Parte inferior ensanchada
Uña bifida
A B eFigura 14. Estructuras para la determinación de la especie Tenagobia incerta. A:Parámeros izquierdo y derecho; B: Pata anterior de la hembra; C: Uña pata posteriorde la hembra (Bachmann 1981).
Presentan un tamaño de 1 a 3 mm, de color parduzco reticulado sobre los hemielitros,
formando bandas difusas y sinuosas que forman mallas coordenadas longitudinalmente,
sintlipsis un poco mas ancha que el borde posterior del ojo, disco pronotal muy corto, borde
posterior del disco pronotal con una excavación ancha poco profunda mal definida. Vertex
algo prominente entre los ojos en vista dorsal, en los machos hemielitros con una banda
hialina, longitudinal, en la membrana derecha, que parece continuar la línea sutural de los
clavos, metaxifo muy corto y ancho, mal definido, redondeado. Parámero derecho presenta
la parte inferior ensanchada y el parámero izquierdo con la punta engrosada, característico
ele la especie (Figura 14A), paleta anterior de la hembra ancha con 13 cerdas y 3 pelos
delgados basales (Figura 14B), pata posterior con uña bifida con dos puntas de diferente
grosory en la bifurcación un tubérculo romo (Figura 14C). Todos son macrópteros.
Sudistribución es Brasil, Perú, Paraguay, Argentina, Colombia, Venezuela y Bolivia.
32

e) Tenagobia socialis (White 1879) FAM. CORIXIDAE
e
Figura 15. Tenagobia socialis. A: urosterguitos 5 y 8vo del macho; B: Parámeros
izquierdo y derecho; C: Urosternito 7mo y 8vo de la hembra (Bachmann 1981).
De tamaño pequeño 2.35 a 2.80 mm en machos y 2.60 a 2.95 mm en hembras, color
generalmente parduzco a claro, los hemielitros suelen ser suavemente acanalados, sintlipsis
tan ancha o un poco mas ancha que el borde posterior del ojo, disco pronotal con
excavación poco profunda y relativamente bien delimitada, lobulillo izquierdo del 8vo urito
de los machos muy poco expandido atrás, con pocos pelos cortos y con una proyección
glabra hacia la linea media (Figura ISA).
Parámero derecho con cabeza bien marcada con punta aguda larga y ligeramente curvada,
borde inferior casi recto y a veces ligeramente cóncavo, finamente serrulado, a veces con
doble serrulación, parámero izquierdo con extremo expandido(Figura 15B) 7mo urosternito
de la hembra poco expandido atrás, muy abierto y redondeado (Figura 15C). Vertex algo
prominente entre los ojos en vista dorsal en los machos; hemielitro derecho con una banda
clara traslucida longitudinalmente, parece seguir la línea sutural de los clavos, Metaxifo
mal definido muy corto y ancho.
Distribución en la Cuenca Amazónica, desde las Guyanas hasta Colombia, Argentina,
Brasil y Bolivia.
33

1) Tenagobia selecta (White 1879) FAM. CORIXIDAE
2
A B eFigura 16. Estructuras para determinar la especie Tenagobia selecta. A: Parárnerosderecho (1) e izquierdo (2); B: Apéndice urito 8vo macho; C: pata anterior (1) yantena en la hembra (2) (Bachmann 1981).
Tiene un color parduzco grisáceo, manchas difusas poco mas oscuras que el tono de fondo,
sintlipsis algo mas ancha que el borde posterior de un ojo, vertex redondeado, poco
prominente en vista dorsal, disco pronotal muy corto, escudete muy ancho y corto,
redondeado, pala del macho con unas 10 cerdas son braquípteros (Figura 16A), variando su
longitud entre 3 a 4 mm, parámero derecho grueso, la parte basal muy abultada seguida de
una parte subcilíndrica compleja con estrías y extremo dilatado en un lóbulo subtriangular
de ángulos redondeados como una punta de flecha, parámero izquierdo con la paleta
compleja, recorrida por una cresta lateral(Figura 16B), 7mo urosternito de la hembra muy
ancho con el borde posterior expandido en un lóbulo semicircular flanqueado por
escotaduras casi semicirculares, 8vo urito en machos angosto con un lóbulo robusto
proyectándose en la mitad proximal (Figura 16C).
Su distribución es amplia en Sudamérica encontrándose los primeros registros en Paraguay.
34

Larva
g) flelerocorixa (White 1879) FAM. CORIXIDAE
A B
Figura 17. A: Vista dorsal de la larva del género Heterocorixa; B: Antena en larva yadulto (Bachmann 1981).
Esta especie es exclusivamente neotropical resulta muy característica por los escleritos y las
suturas cefálicas presenta sintlipsis mucho mas angosta que el borde posterior de un ojo, en
vista dorsal; disco pronotal corto (Figura 17A), los hemielitros al igual que el disco
pronotal con diseño vermiculado coriáceos y pigmentados excepto en la membrana del
izquierdo que es clara, antenas en adultos con 4 artejos (Figura 17B).
Se encuentran dos especies: Heterocorixa boliviensis y Heterocorixa hintoni descritas
únicas especies para Bolivia. El genero se encuentra distribuido desde Corumbá en el Brasil
hasta Argentina y Bolivia (Bachmann 1981).
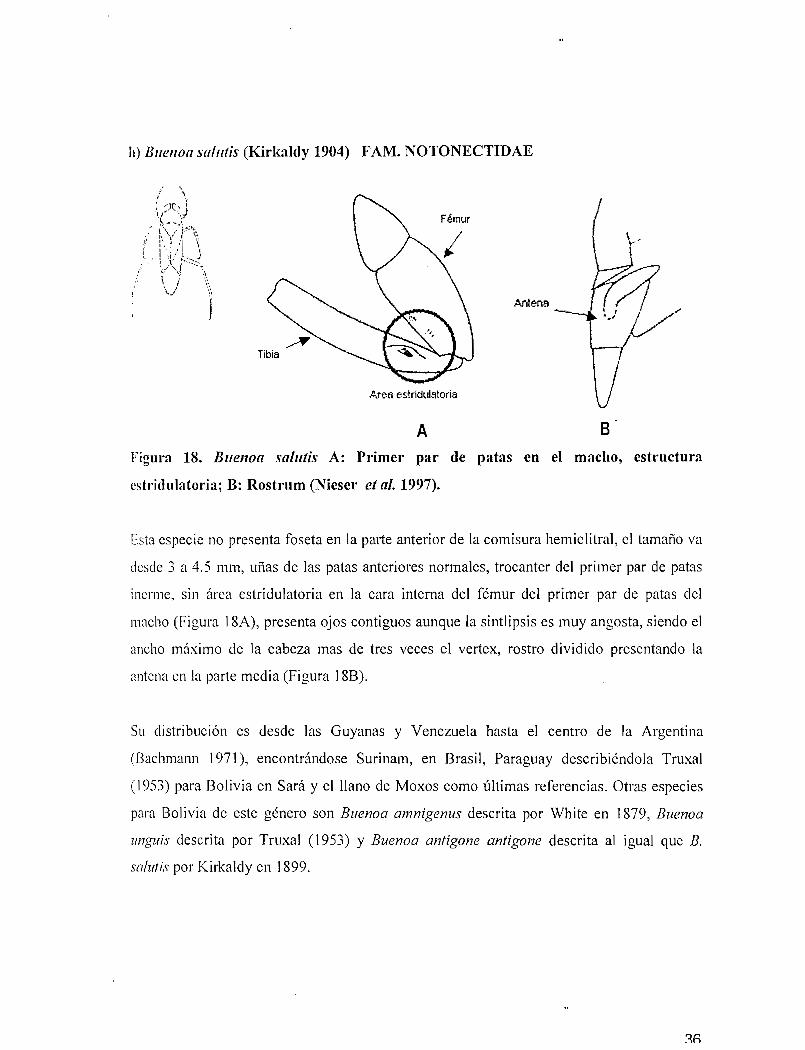
11) BlIe110a salutis (Kirkaldy 1904) FAM. NüTüNECTIDAE
Area estridulatorie
A
Antena
vB
Figura 18. Buenoa salutis A: Primer par de patas en el macho, estructura
estridulatoria; B: Rostrum (Nieser et al. 1997).
Esta especie no presenta foseta en la palie anterior de la comisura hemielitral, el tamaño va
desde 3 a 4.5 mm, uñas de las patas anteriores normales, trocanter del primer par de patas
inerme, sin área estridulatoria en la cara interna del fémur del primer par de patas del
macho (Figura 18A), presenta ojos contiguos aunque la sintlipsis es muy angosta, siendo el
ancho máximo de la cabeza mas de tres veces el vertex, rostro dividido presentando la
antena en la parte media (Figura 18B).
Su clistribución es desde las Guyanas y Venezuela hasta el centro de la Argentina
(Bachmann 1971), encontrándose Surinam, en Brasil, Paraguay describiéndola Truxal
(1953) para Bolivia en Sará y el llano de Moxas como últimas referencias. Otras especies
para Bolivia de este género son Buenoa amnigenus descrita por White en 1879, Buenoa
unguis descrita por Truxa1 (1953) y Buenoa antigone antigone descrita al igual que B.
salutis por Kirkaldy en 1899.

i) Martarega cltinai (White 1879) FAM. NOTONECTIDAE
/Ausencia de comisurahemielitral
A
Banda opaca long~udinal bifurcada
B
Figura 19. Martarega chinai A: Hemielitro B: Vista dorsal (Nieser & Melo 1997).
Este género no presenta foseta en la parte anterior de la comisura hemielitral (Figura 19A),
presenta los ojos unidos a diferencia de Buenoa y Notonecta, con antenas de 3 a 4 artejos,
son de tamaño pequeño (menor a 3 mm) presenta una banda opaca longitudinal bifurcada
en el hemielitro, formas braquípteras (Figura 19B).
Presenta tres especies, Martarega uruguayensis por Berg (1879), Martarega meatesi
descrita por Jaczewski (1929), encontrándose en el noreste del Brasil, Uruguay y al este de
la Argentina y la especie Martarega chínai descrita por Hynes (1948) registrada en Bolivia.
37

j) Belostoma (De CarIo 195) FAM. BELOSTOMATIDAE
A B
Fig;ura 20. Belostoma A: Abdomen con pilosidad extendida hasta los esternitos; B:
Abdomen vista ventral (Nieser & Melo 1997).
Presenta un cuerpo oval, elíptico o alargado, tamaño comprendido entre 0,9 cm a 6 cm,
rostro con el tercer segmento muy corto con 3 o 4 artejos en las antenas. Fémures en el
primer par de patas anchos, respecto a los posteriores con una pequeña excavación en estos
para la recepción de la tibia. Con protarsos dímeros con dos uñas, uno rudimentario y el
otro bien desarrollado característica importante para la determinación de géneros.
Esicrnitos abdominales con pubescencia en los márgenes, falobase bifurcada dorsalmente
con los brazos laterales que se extienden hasta el ápice del divertículo ventral. Se observa el
abdomen con pi losidad extendida hasta los esternitos (Figura 20A), o en algunos casos
solamente en el borde (Figura 208).
Esta distribuido en casi todos los países de América del Sur y Centroamérica como en
México y Estados Unidos, En Bolivia se encuentra las especies B. martinezi descrita por De
Carla (1957), B. Aurivillianum descrito por Montandon (1899), B. dilatatum descrito por
Dufour (1863), Belostoma elegans descrito por Mayr (1871), B. elongatum descritas por
Montandon (1908) Y B. gestroi también por Montandon (1900) (cit. Schnack 1976), todas
las descripciones para Bolivia corresponden a los departamentos de Santa Cruz de la Sierra
y Cochabamba.
38

k) Lethocerus truncatus (Curnminus 1933) FAM. BELOSTOMATIDAE
A B
Figura 21. Lethocerus truncatus A: genitalia vista lateral; B: Pílosidad abdominal(Schnuck 1976)
1'\l:l especie tiene un largo del cuerpo entre 4.5 hasta 8 cm, presenta uñas del primer par de
patas un poco mas largas que los artejos de su tarso medidos por el borde externo, fémures
anteriores robustos, metatibias de ancho apenas mayor, casi igual al diámetro del ojo con el
extremo de la cara inferior redondeado. Metaxifo con una pequeña punta aguzada en la
parte caudal, en algunos individuos presenta como variación una punta redondeada. La
armadura genital para determinar especies es idéntica a la figura 21A Y presenta
pubescencia abdominal bordeando los laterotergitos (Figura 21B).
La distribución de esta especie es subtropical y subandina, presentándose también en Brasil
y Argentina. Otras especies descritas para Bolivia son Lethocerus dilatus descrito también
por Cummings (1933), encontrándose los holotipos y paratipos en Museo de la Universidad
ele Kansas, procedentes de Buena Vista, Departamento de Santa Cruz de la Sierra,
Lethocerus maximus descritas por De Carla en 1938 provenientes también de Santa Cruz de
la Sierra y Lethocerus mazzai descritas por De Carla en 1961, registrada también en Bolivia
en los Yungas del Palmar por Schnack en 1976.
39

1) Neogerris (White 1879) FAM. GERRIDAE
Una característica general de este género que presenta menor tamaño que Limnogonus, un
poco más oscuro, caracterizado principalmente por la gran extensión del pronoto hacia atrás
con un rostro largo alcanzando el mesosterno. Ápice del abdomen característico para
Nl'IJgerris (Figura 22A).
A
m) Neogerris lubricus (White 1879)r-
.'\./ ,/,/.1:;" , )
T;T ),,¡ ! .~··,·.vil!;wvt
.. ¡", 'h~
/,/11 ¡:'\' I 1'1 1:;;;
,j ¡l d
:,: .....~. \
\ \\ \
\ ¡
j 1, \
.' ,Figura 22. A: Vista dorsal ápice abdomen, Neogerris sp; B: ápice abdomen Neogerris
Inbricus (Aristizabal 2001).
Esta especie encontrada únicamente en época de transición en la laguna Colorada presenta
como característica única el pronoto prolongado posteriormente mas largo que ancho,
cubriendo enteramente el mesonoto, presentaba sinonimia con Limnogonus lubricus según
\Vhite (1879). su color es café dorsal y ventral mente. Prosterno, propleura y proacetabula
café claro. Las antenas y las patas cafés más claras que el cuerpo, en seco presenta una
delgada línea bordeando la cabeza con una pubescencia blanca a plateada. Ápice del
abdomen pequeño característico de esta especie (Figura 22B).
Especie presente en Argentina según trabajos de Bachmann (1966), Guyana, Surinam,
Brasil, Perú por trabajos de Nieser (1970), en Colombia en trabajos de Roback (1974) (cit.
en Aristizábal 2002), en Ecuador nombrado como primer registro por Aristizábal (2002) y
en Bolivia por Nieser en 1970.
40

11) Lil11110g01111S aduncus (Drakc & Harris 1941) FAM. GERRIDAE
Proyección apical en terma de espre
A
" \\
B
Figura 23. LiI11110go1111S aduncus. A: Abdomen vista dorsal; B: Genitales vista lateral(Niescr & Melo 1997).
El género Limnogonus se observan tamaños de mediano a grande 2 a 3 cm, según
Aristizábal (2002) presentan dimorfismo sexual en cuanto a tamaño, siendo la hembra más
grande. La especie Limnogonus aduncus presenta una mesopleura coloreada, en vista dorsal
en el último segmento se observa el ápice proyectado hacia abajo distinguiéndose poco la
genitalia (Figura 23A), presenta solamente el macho proyección apical en forma de espina,
siendo el séptimo segmento abdominal proyectada un poco hacia atrás (Figura 23B).
La distribución según Nieser (1970), desde Panamá, Venezuela, Guyana, Ecuador, Perú,
Brasil, Bolivia y Paraguay.
41

o) Rlteumatobates crassifemur crassifemur (Esaki 1926) FAM. GERRIDAE
2clo segmentocorto con 2 cerdas l~
A
[<,igllra 24. Rlteumatobutes crassifemur crassifemur A: Antena del macho, B: Vistagcncnll (Nieser & Melo 1997).
bta especie presenta ojos no emarginados, pronoto con el borde anterior no cóncavo y el
borde posterior sinuoso, Antena del macho modificada con el primer segmento engrosado,
el segundo segmento corto con dos cerdas largas ubicadas en la cara ventrolateral externa,
tercer segmento con dos cerdas en la cara interna, cuarto segmento muy modificado con el
icrc io basal delgado engrosándose en el segundo tercio, lleva en la base un grupo de pelos
fusionados que se dirigen hacia la parte externa (Figura 24A). El fémur anterior modificado
en machos ligeramente engrosado siendo en hembras delgado (Figura 24B).
Su distribución fue primeramente descrita en Bolivia y Paraguay por (Hungerford 1954),
posteriormente descrito por Bachmann en 1966 y para Colombia por Aristizábal (2002).
42
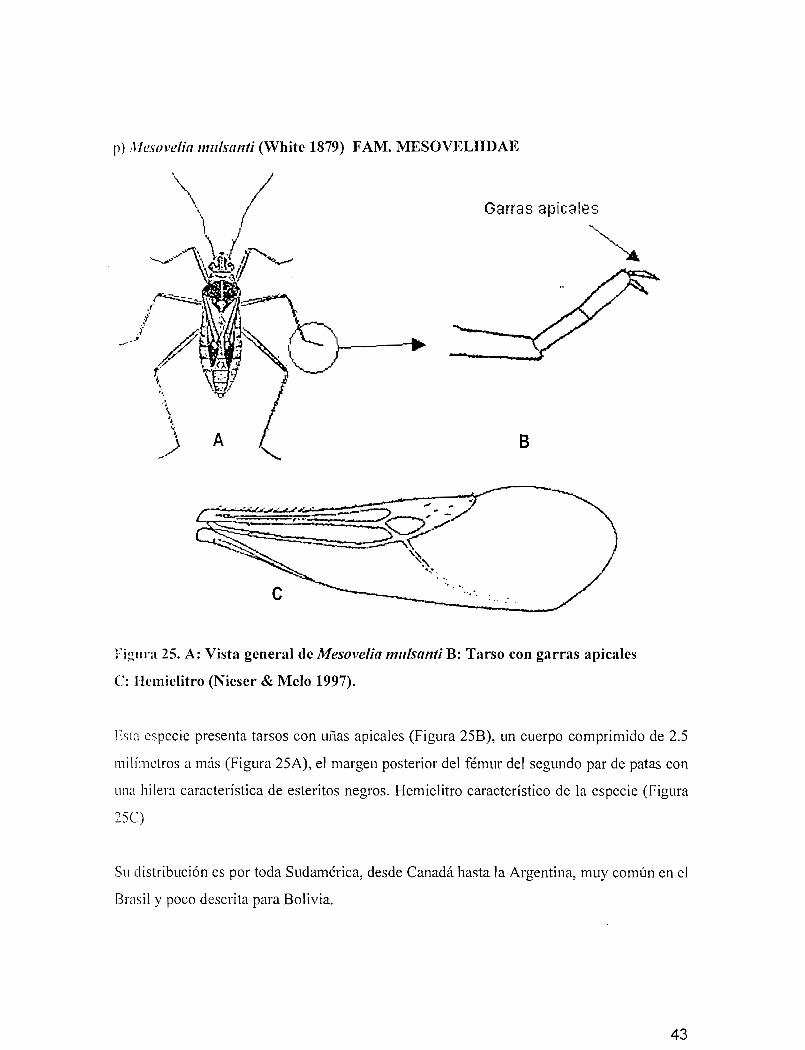
p) Mesovelia mulsttnti (vVhite 1879) FAM. MESOVELIIDAE
Garras aptcales
~~
B
FiglInl 25. A: Vista general de Mesovelia mulsanti B: Tarso con garras apicales
C: Hcmiclitro (Nieser & Melo 1997).
Esta especie presenta tarsos con uñas apicales (Figura 25B), un cuerpo comprimido de 2.5
milímetros a más (Figura 25A), el margen posterior del fémur del segundo par de patas con
una hilera característica de esteritos negros. Hemielitro característico de la especie (Figura
25C)
Su distribución es por toda Sudamérica, desde Canadá hasta la Argentina, muy común en el
Brasil y poco descrita para Bolivia.
43
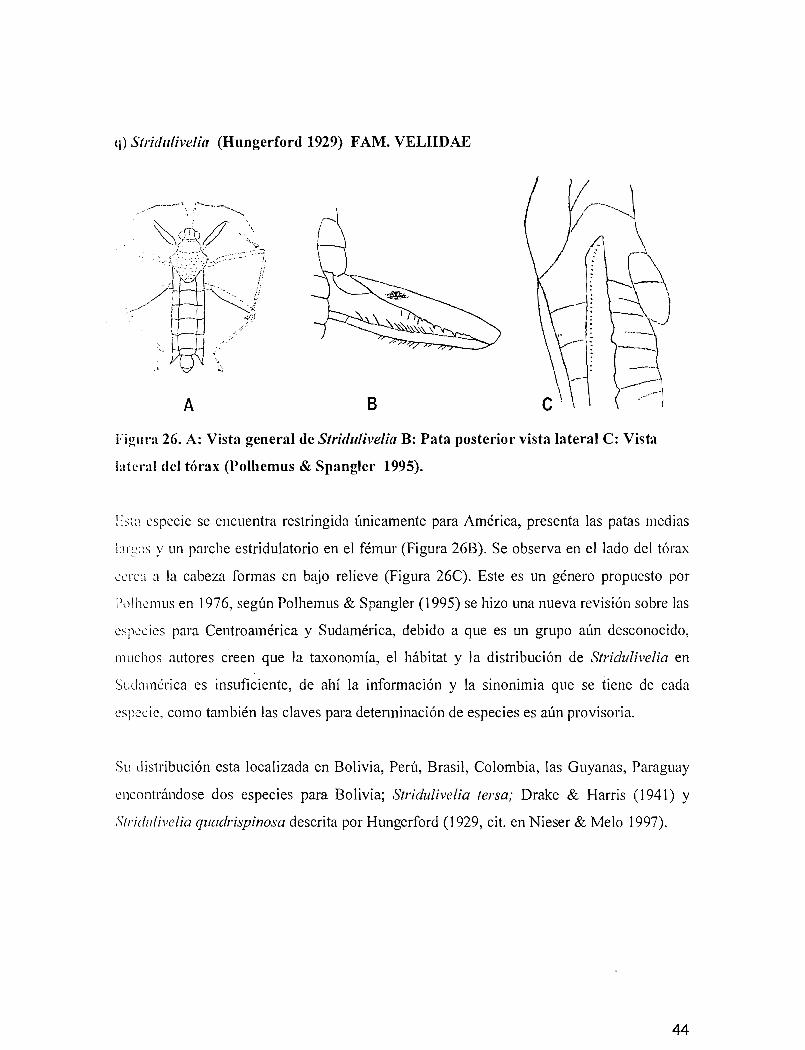
q) Stridttlivelia (Hungerford 1929) FAM. VELIIDAE
-----._--~..~.~ ,;.._- •. - '<;<,
A B
Figura 26. A: Vista general de Stridutivelia B: Pata posterior vista lateral C: Vista
lateral del tórax (Polhemus & Spangler 1995).
1::>t~\ especie se encuentra restringida únicamente para América, presenta las patas medias
i:lr~-,;ls y un parche estridulatorio en el fémur (Figura 26B). Se observa en el lado del tórax
L"-?I"Cl a la cabeza formas en bajo relieve (Figura 26C). Este es un género propuesto por
PlJlh-?l11l1S en 1976, según Polhemus & Spangler (1995) se hizo una nueva revisión sobre las
especies para Centroamérica y Sudamérica, debido a que es un grupo aún desconocido,
muchos autores creen que la taxonomía, el hábitat y la distribución de Stridulivelia en
Sudamérica es insuficiente, de ahí la información y la sinonimia que se tiene de cada
especie, como también las claves para determinación de especies es aún provisoria.
Su distribución esta localizada en Bolivia, Perú, Brasil, Colombia, las Guyanas, Paraguay
encontrándose dos especies para Bolivia; Stridulivelia tersa; Drake & Harris (1941) Y
Stridulivelia quadrispinosa descrita por Hungerford (1929, cit. en Nieser & Mela 1997).
44

r) Gelastocoris (Kirkaldv 1899) FAM. GELASTOCORIDAE
"'\\\
>,l,
";. AFigura 27. Gelastocoris A: Vista general. B: Tarso anterior no fusionado a la tibia
(Nicscr & lVIelo 1997).
El género Gelastocoris presenta un cuerpo ensanchado y corto, con ojos prominentes, las
patas anteriores son raptoras; los hemielitros son separados y desarrollados completamente
(Figura 27A). Es predador, con los tarsos anteriores no fusionados a las tibias, las patas
anteriores con dos uñas bien desarrolladas (Figura 27B). La parte posterior del pronoto sin
carena longitudinal.
Distribuido en parte de Sudamérica, Gelastocoris no se encuentra presente en el continente
Ncartico (Nieser & Mela 1997).
s) Pelocoris binotulatus bino/lila/lis (StaI1876) FAM. NAUCORIDAE
Esta especie presenta el rostro corto, OJos globosos, superficie dorsal lisa, placas
propleuricas cubriendo el prosterno (Figura 28A). Fémur mediano estrecho, presenta cerdas
pequeñas en la tibia, tarso dividido (Figura 28B 1) antena con el 2do antenito
subcuadrangular, los siguientes alargados terminando el ultimo en punta roma (Figura 28B
2, Neiser & Mela 1997). Presenta el abdomen un borde de forma aserrada, puesto que cada
laterotergito termina en punta. El ángulo posterior de cada laterotergito ostenta una mancha
oscura que le confiere al conexivo un aspecto característico. Estructura genitales como en
las figuras 28B 3 Y4, (López Ruf] 994).
45

2
41
Figura 28. Pelocoris binotulatus binotulatus A: Vista general; B: Estructuras: 1. Pata 1
2. Antena, 3. Genitalia femenina, 4. Aedeago (López Ruf 1994).
t) Ctenipocoris shadei (StaI1876) FAM. NAUCORIDAE
Cuerpo deprimido dorsoventralmente, de contorno suboval, mas piloso dorsal que
ventral mente (figura 29A). Cabeza sin manchas, borde anterior suavemente convexo, tarso
bisegmentado con un par de uñas distales. Estructuras genitales características (figura 29B
1 Y B2). Aedeago característico hasta género (Figura 29B 4) Parameros acintados,
siguiendo desde su base un recorrido helicoidal entrecruzandose cerca de la base del
aedeago, terminados en un ensanchamiento redondeado y aplanado cubierto de puas cortas.
Fig 29B 3 (López Ruf2004).
3
B
/\I lI I, I
I !¡l i
! J1
(U?~/
4
Figura 29. Ctenipocoris sltadei A: Vista general; B: Estructuras genitales: 1. GenitaJia
masculina, 2. Genitalia femenina, 3. Parámero, 4. Aedeago (vista ventral y lateral
(López Ruf 2004).
46
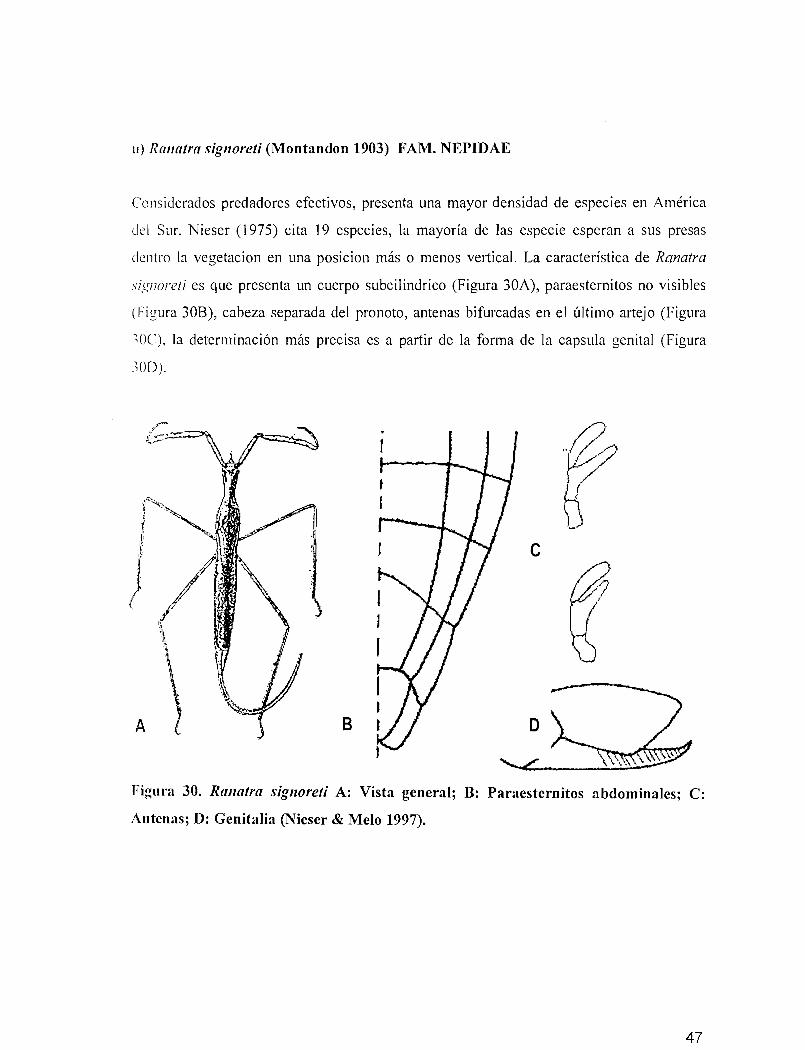
ti) Ranatra signoreti (Montandon 1903) FAM. NEPIDAE
Considerados predadores efectivos, presenta una mayor densidad de especies en América
del Sur, Nieser (1975) cita 19 especies, la mayoría de las especie esperan a sus presas
dentro la vegetacion en una posicion más o menos vertical. La característica de Ranatra
signoreti es que presenta un cuerpo subcilindrico (Figura 30A), paraesternitos no visibles
(Figura 30B), cabeza separada del pronoto, antenas bifurcadas en el último artejo (Figura
-'OC). la determinación más precisa es a partir de la forma de la capsula genital (Figura
-'OD).
,- .:"~'/1/
~e
a~/~
8
Figura 30. Ranatra signoreti A: Vista general; B: Paraesternitos abdominales; C:
Antenas; D: Genitalia (Nieser & Melo 1997).
47

3.2 Distribución y ecología de hemípteros acuáticos
l. a fi gura 31 muestra la abundancia de los géneros y especies más representativos en este
estudio, se observa la mayor abundancia en los géneros Neoplea, y Tenagobia , como
también en las especies Buenoa salutis y Neoplea maculosa.
Lelhocerus larva
_ ____ _ _ _____ _ _ N:..:-e=oplea larva _
Helerocorixa larva
Tenagobia larva
Tenagobia socielis I
I
IIJ
Tenag obia incerta
..Pelocoris larva
Buenoa salulis
Buenoa larva
Neoplea maculosa
Figura 31. Abundancia de los géneros y especies más representativos de hem ípteros
acu úticos.
l.os organismos que presentaban menor abundancia fueron separados asumie ndo corno
mínimo 6 organismos para la realización del análisis factorial de correspondencia (AFC).
En el cuadro 5 se observa las especies y géneros presentes en el análisis como también los
géneros y especies descartadas para el análisis.
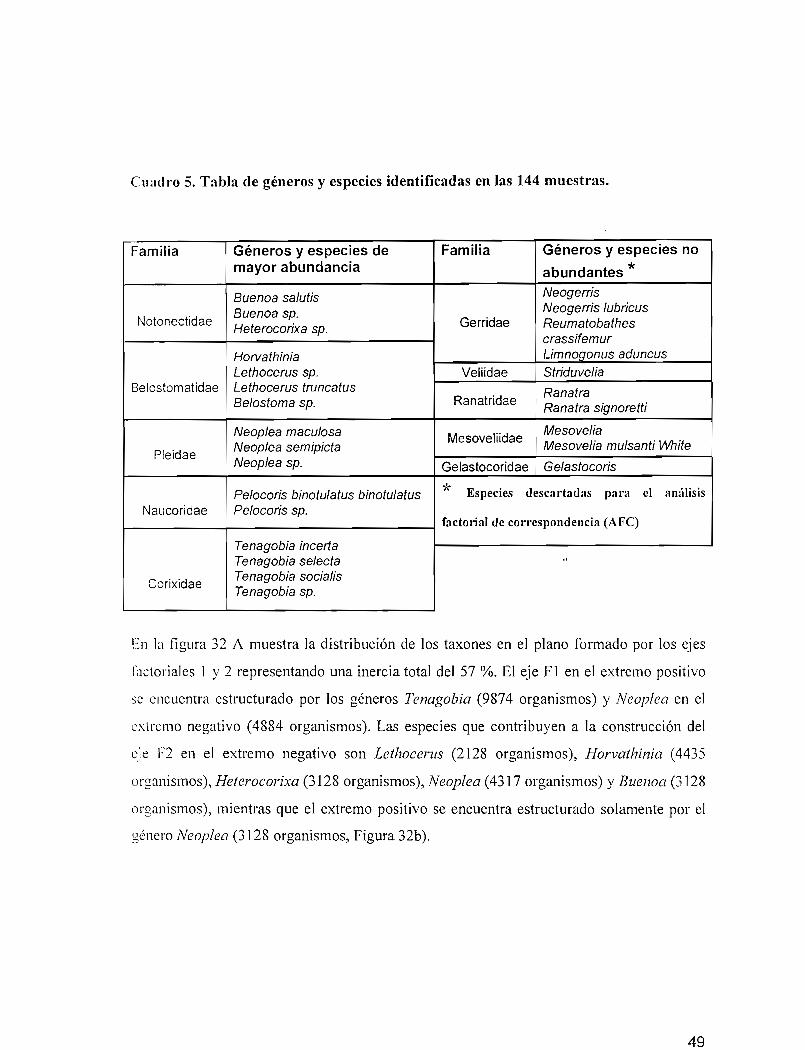
Cuadro 5. Tabla de géneros y especies identificadas en las 144 muestras.
Familia Géneros y especies de Familia Géneros y especies nomayor abundancia abundantes *Buenoa salutis Neogerris
Buenoa sp. Neogerris lubrieusNotonectídae
Heteroeorixa sp. Gerridae Reumatobatheserassifemur
Horvathinia Limnogonus aduneusLethoeerus sp. Veliidae Striduvelia
Belostomatidae Lethoeerus truncatus RanatraBelostoma sp. Ranatridae
Ranatra signoretti
Neoplea maeulosa MesoveliidaeMesovelia
PleídaeNeoplea semipieta Mesovelia mulsanti WhiteNeoplea sp. Gelastocoridae Gelastoeoris
Peloeoris binotulatus binotulatus * Especies descartadas para el análisisNaucoridae Peloeoris sp.
factorial de correspondencia (AFC)
Tenagobia incerteTenagobia selecta "
CorixidaeTenagobia socielisTenagobia sp.
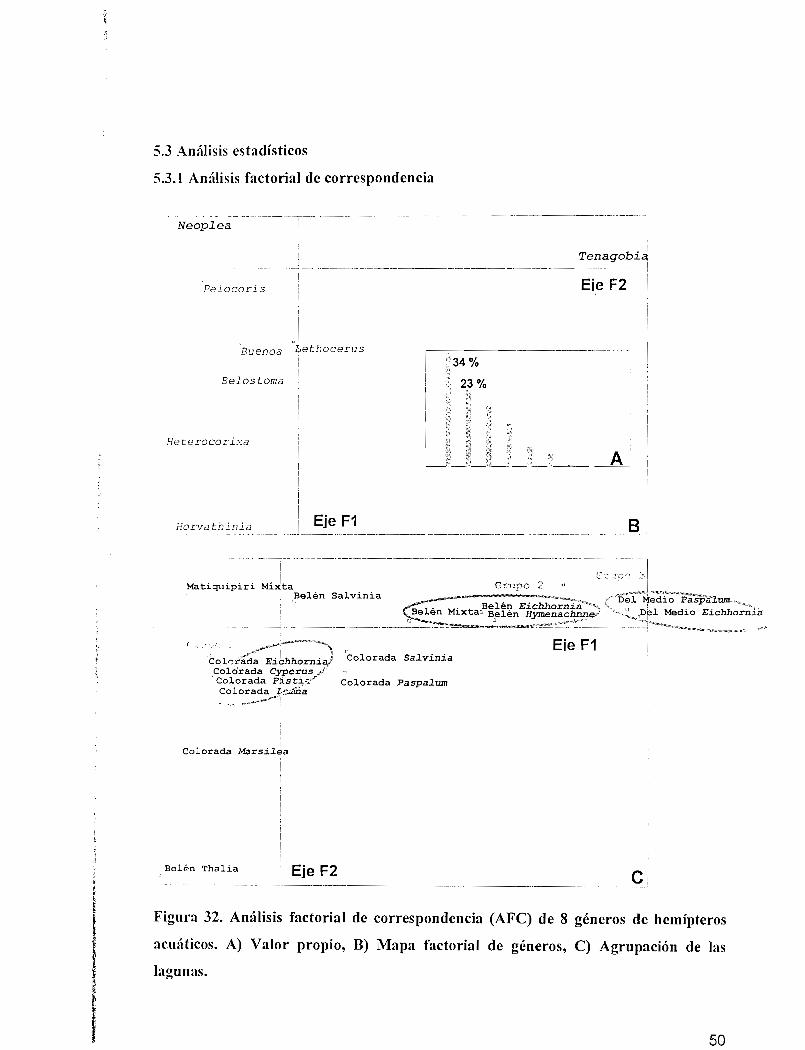
En la figura 32 A muestra la distribución de los taxones en el plano formado por los ejes
lactoriales 1 y 2 representando una inercia total del 57 %. El eje F 1 en el extremo positivo
se encuentra estructurado por los géneros Tenagobia (9874 organismos) y Neoplea en el
extremo negativo (4884 organismos). Las especies que contribuyen a la construcción del
eje [72 en el extremo negativo son Lethocerus (2128 organismos), Horvathinia (4435
organismos), Heterocorixa (3128 organismos), Neoplea (4317 organismos) y Buenoa (3128
organismos), mientras que el extremo positivo se encuentra estructurado solamente por el
género Neoplea (3] 28 organismos, Figura 32b).
49
5.3 Análisis estadísticos
5.3.1 Análisis factorial de correspondencia
---- ---- ---~----
Neoplea
..... ITena qoos.s;-- ---~- - ----------------------- ---- - ---\
'Pelocoris
Buenoa
Belostoma
He terocor .íxa
Horvathinia
Eie F2
I
"lethocerus
III
I
iII
I Eje F1_1 . ,, . B
Matiquipiri
!. I
M~xta
I Beléni L;
SalviniaGrn.po 2
""_~1>.•:<'~.•""'''''';~~--'''~>\
Col~tada EihhhorniaJColorada C~ez;ps,,/,/Colorada Pist;¡;.,:"Colorada t.~rña
,.,,-,,-'''''+·...~''';#'''i
Colorada Marsi~Ja
Colorada Sa~vinia
Colorada Paspa~um
Eie F1
e
Figura 32. Análisis factorial de correspondencia (AFC) de 8 géneros de hemípteros
acuáticos. A) Valor propio, B) Mapa factorial de géneros, C) Agrupación de las
lagunas.
50

En la figura 32 sc observa la distribución de las lagunas en relación con los géneros. Sobre
el eje Fl se muestra la asociación de estaciones de las lagunas Belén (laguna de sabana) y
Del Medio (laguna de bosque), lagunas de diferentes características por su ubicación pero
"i milares por presentar alta abundancia del género Tenagobia (Corixidae), se observa una
contribución significativa de Matiquipiri para el género Neoplea.
Sobre el eje F2 en el plano negativo se observa la asociación de las estaciones de la laguna
Co [orada (laguna de bosque), con la máxima contribución para construcción de este eje, por
presentar la mayor abundancia del género Buenoa (Notonectidae).
1] mapa factorial muestra solamente estaciones con altas contribuciones, descartando las
menos significativas, mostrando asociación de las estaciones respectivas a cada laguna
(grupos 1,2 y 3). Se puede observar que la distribución en el plano factorial de abundancia
hemípteros acuáticos respecto a la vegetación Eichhornia sobresale en las estaciones de las
lagunas del Medio, Colorada y Belén.
5.3.2. Análisis discriminante
La figura 33 representa las proyecciones en el plano factorial x 2, son las estaciones
clasificadas en función a la época.
r CV :-('I,)~~j~= Época
Ii Noviembre 1
I
~brero 23Abril
! ,Eje 2 M::lVO 4
Figura 33. Análisis factorial de la tabla faunística de hemípteros acuáticos. Proyecciónde las épocas clasificadas en función a la abundancia.
51

Los centros de clase están alineados en el eje 1, no mostrando una superposición clara de
las clases excepto en los meses de abril y mayo (misma época respecto a las demás). La
inercia interclase muestra un 21 % de la inercia total, se observa una correlación no
significativa (Figura 34).
Número de permutaciones aleatorias: 1000
Valor observado = 451
1****** Mínimo = 118Ij.,.*************
1********************
\***************************************1**************************************************1*********************************************
~I*****************************************
1***********************************1*******************************1*********************1************1*******1*****1***
Inercia interclase: 451 (21 %)Inercia intrac!ase: 1717 (79 %)
Número de simulaciones X<Obs: 571Número de simulaciones X>=Obs: 429
1*
Figura 34. Test por simulaciones aleatorias: época.

(2)
r----T-------~---·---
I (~) 1 Eje 1~--~- -~~~~~~-~--~~--
I
IIi
IEje 2
Lagunas
Belén 1Colorada 2Del Medio 3Matiquipiri 4
Figu ra 35. Análisis factorial de la tabla faunística de hemípteros acuáticos. Proyección
de las lagunas clasificadas en función a la abundancia
LJ figura 35 nos muestra las proyecciones en el plano factorial 1 x 2, son las estaciones
elasificadas según las lagunas.
Número de permutaciones aleatorias: 1000
Mínimo = 67
Valor observado = 630
Inercia interclase: 630 (29 %)Inercia intraclase: 1538 (71 %)
Número de simulaciones X<Obs: 929Número de simulaciones X>=Obs: 71
\****1**************
1*********************1***********************************1*************************************1**************************************1**************************************************1*******************************************1**************************************1************************************1*****************************1****************************1*****************
~I*************
/*********
1*****1***I
Figura 36. Test por simulaciones aleatorias: laguna
53

Se observa las clases agrupadas más cercanas al eje 1, al observar las proyecciones tan
separadas se muestra las diferencias existentes entre lagunas respecto a la abundancia.
1.~1 inercia interclase representa el 29 % de la inercia total, el test de permutaciones
aleatorias muestra que la correlación no es significativa.
.-_._--_._~- -------------_._--- - ---- ------_._------ Plantas
Cyperus 1
@Eichhornia 2
(6) ( 7Hymenachne 3
(~) (2) Eje 1 Polygonum y4
11\ LemnaI ~4 ~ Marsilea 5
Mixta 618 1 Paspalum 7
Pistia 8,5\
Salvinia 9(10' Eje 2 Thalia 10
Fignra 37. Análisis factorial de la tabla faunística de hemípteros acuáticos. Proyecciónde la vegetación clasificada en función a la abundancia de hemípteros acuáticos.
La figura 37 representa las proyecciones en el plano factorial 1 x 2, estas estaciones se
encuentran clasificadas en función de la vegetación identificada y la figura 38 muestra los
resultados del test de permutaciones aleatorias.
Fi4

Número de permutaciones aleatorias: 1000
Mínimo = 334
Número de simulaciones X<Obs: 949Número de simulaciones X>=Obs: 51
1**
/*****1*********1****************
1***********************
1*****************************************[*********************************************
1***********************************************1**************************************************
1**********************************************
\************************************
I************************************~**~*
1*********************** Inerc~a ~nterclase: 1277 (5~ %)1********** Inercia lntraclase: 891 (41 Yo)
=>1************
1****1****1**
/*1*
Valor observado = 1277
Figura 38. Test por simulaciones aleatorias: Vegetación
La inercia interc1ase es el 59 % de la inercia total y el test muestra que la correlación no es
significativa.
55

:'04 . Comparación de abundancia por factores
1:" el cuadro 7 se observa la comparación de abundancias de géneros donde Neop lea
(Plcidae) es el género que presenta la mayor abundancia de organismos (99 1), Tenagobia
( () 11 -t G9 organismos y Buenoa con 384 organismos.
- - ---- ---- - - ---- - - -------
Buen aa
LethocerusBe/as/a ma
___---r-~ Harvathinia
Tenagabia
Pelocoris
Lethocerus
98Belostoma
6
Horvat hinia22
Tenagobi a469
Heterocorixa
70Buenoa
384
fignra 39. Comparación de abundancias en géneros de hemípteros acuáticos.
5..+.1 Prueba T de Student
5.4.1.1 Factor época
1::1 análisis estadístico T de Student nos muestra en el cuadro 6, la relación de la abundancia
COIl In época asumiendo homogene idad y normalidad en los datos, donde el género
Tenagob ia (Corixidae) muestra un estadístico de p = 0,01626 < 0,05 existiendo diferencias
de abundancia entre los meses de noviembre y abril, mostrando incremento en abundancia
en el mes de abril (transición) respecto a noviembre (estiaje), Cuadro 5.
56
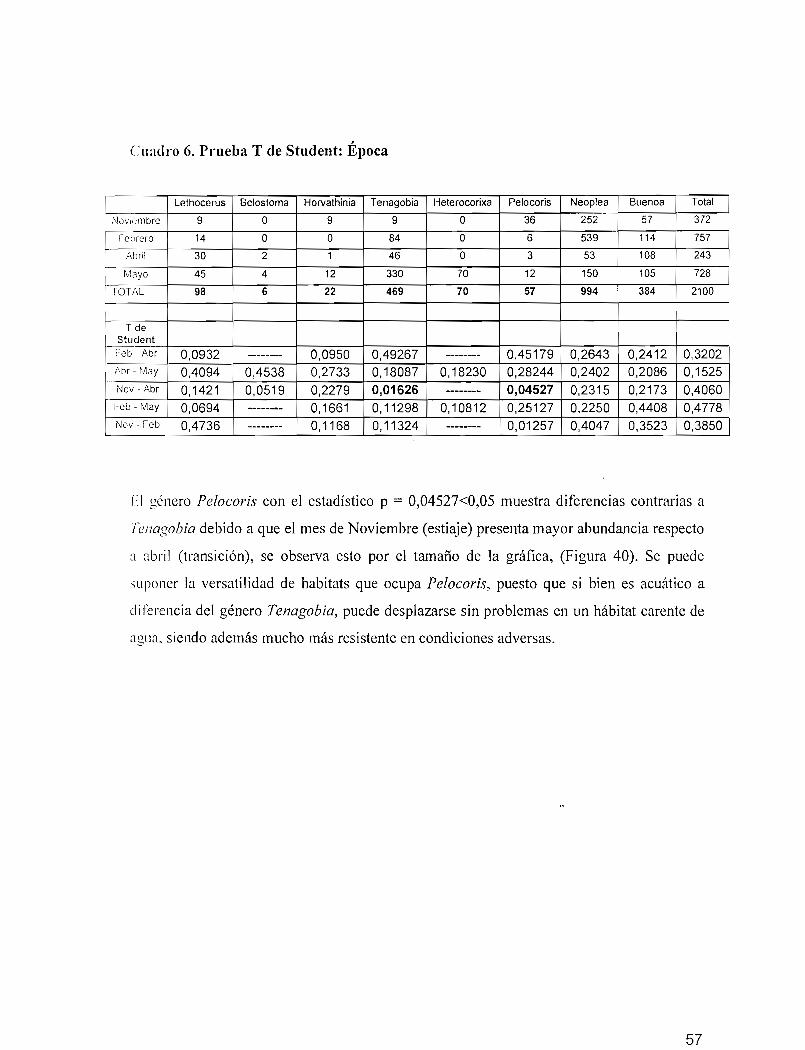
Cuadro 6. Prueba T de Student: Época
r-'-
Lethocerus Belostoma Horvathinia Tenagobia Heterocorixa Pelocoris Neoplea Buenoa Total
'"N"ov,,·,mbre 9 O 9 9 O 36 252 57 372
¡·ellrero 14 O O 84 O 6 539 114 757
!Illril 30 2 1 46 O 3 53 108 243
Mnyo 45 4 12 330 70 12 150 105 728
TOT/IL 98 6 22 469 70 57 994 384 2100f-.
T deStucJent
I-"~.
leb . Abr 0,0932 -------- 0,0950 0,49267 -------- 0,45179 0,2643 0,2412 0,3202I\br· May 0,4094 0,4538 0,2733 0,18087 0,18230 0,28244 0,2402 0,2086 0,1525Nov· Abr 0,1421 0,0519 0,2279 0,01626 -------- 0,04527 0,2315 0,2173 0,4060¡":eb· May 0,0694 ------- 0,1661 0,11298 0,10812 0,25127 0,2250 0,4408 0,4778Nov· Feb 0,4736 -------- 0,1168 0,11324 -------- 0,01257 0,4047 0,3523 0,3850
L
[1 género Pelocoris con el estadístico p = 0,04527<0,05 muestra diferencias contrarias a
Tenagobia debido a que el mes de Noviembre (estiaje) presenta mayor abundancia respecto
~l abril (transición), se observa esto por el tamaño de la gráfica, (Figura 40). Se puede
suponer la versatilidad de habitats que ocupa Pelocoris, puesto que si bien es acuático a
diferencia del género Tenagobia, puede desplazarse sin problemas en un hábitat carente de
agua. siendo además mucho más resistente en condiciones adversas.
57

NOVIEMBRE
Neoplea
ABRIL
FEBREROLethocerus
MAYO
Tenagobia
Lethocerus
Tenagobia
Neoplea
Buenoa
Neoplea
Helerocorixa
Figura 40 Abundancia de hemípteros acuáticos por época. El tamaño de cada gráfi ca
indica la abundancia de organismos colectados por mes.
En la figura 40 sc muestra la abundancia por época, se observa mayor abundancia en los
meses de Febrero (inundación) y Mayo (transición) y menor abundan cia en los meses de
Noviembre (estiaje) y Abril (transición), esta disminución de abundancia en época de
transición se puede la baja temperatura registrada el momento de la colecta.
Existen diferencias entre los meses de noviembre de 2002 y abril de 2003 (p = 0,0 16) para
el género Y enagobia (Corixidae) y entre noviembre y abril también de los mismos años (p
= 0,045) para Pelocoris (Naucoridae), con un intervalo de confianza del 90 %.
58
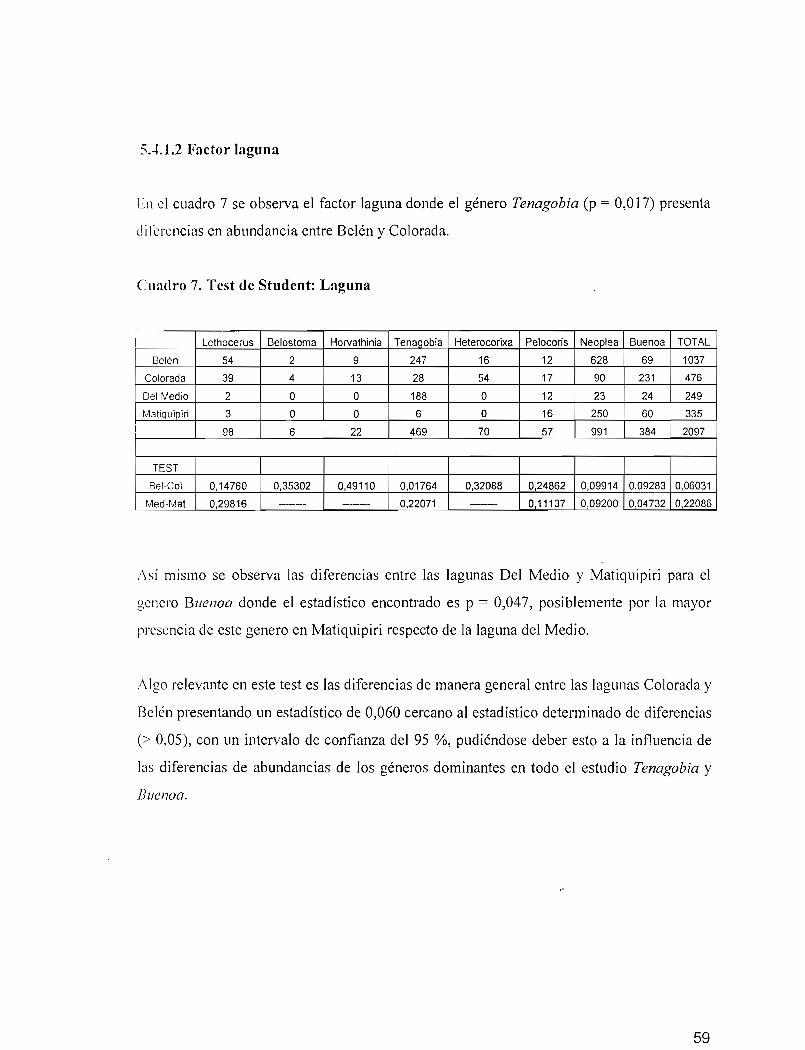
5.-U.2 Factor laguna
En el cuadro 7 se observa el factor laguna donde el género Tenagobia (p = 0,017) presenta
el i ícrcncias en abundancia entre Belén y Colorada.
Cuadro 7. Test de Student: Laguna
Lethocerus Belostoma Horvathinia Tenagobia Heterocorixa Pelocoris Neoolea Buenoa TOTAL
Belén 54 2 9 247 16 12 628 69 1037
Colorada 39 4 13 28 54 17 90 231 476
Del Medio 2 O O 188 O 12 23 24 249
Matiquipiri 3 O O 6 O 16 250 60 335I 98 6 22 469 70 57 991 384 2097
TEST
Bel-Col 0,14760 0,35302 0,49110 0,01764 0,32088 0,24862 0,09914 0,09283 0,06031
Med-Mat 0,29816 ----- ------ 0,22071 ---- 0,11137 0,09200 0,04732 0,22086
i\sí mismo se observa las diferencias entre las lagunas Del Medio y Matiquipiri para el
genero BlIcnoa donde el estadístico encontrado es p = 0,047, posiblemente por la mayor
presencia de este genero en Matiquipiri respecto de la laguna del Medio.
Algo relevante en este test es las diferencias de manera general entre las lagunas Colorada y
Belén presentando un estadístico de 0,060 cercano al estadístico determinado de diferencias
(> 0,05), con un intervalo de confianza del 95 %, pudiéndose deber esto a la influencia de
las diferencias de abundancias de los géneros dominantes en todo el estudio Tenagobia y
Bucnoa.
59

1I
Heteroc arixa
DEL MEDIOLelh acerus
BuenaaNeaplea
Ten agabia
LethacerusBelastama
Horve íninisTenagab ia
Bue naa
COLORADA
MATIQUIPIRI
Heterocarixa
Pelacaris
-- - --- - - -
BELÉN-
NeO:I~
Figu ra 41. Abundancia de hemípteros acuáticos por laguna. El tamaño de cada
~ rúli l: a indica la abundancia organismos colectados por laguna.
[11 I ~ figura 41 se muestra la abundancia de hemípteros acuáticos por laguna, ex iste mayor
abundancia de organ ismos en las lagunas Belén y Colorada, y menores abundancias entre
Matiquipiri y laguna Del Medio.
504.1.3 Factor vegetación
el cuadro 8 muestra el factor vegetación por abundancia. Se observa diferencias para
Ncoplea (p = 0.016) entre Eichhornia y Mixta, debido a la mayor presencia de Neoplea
para la vegetac ión M ixta respec to a Eichhornia.
60

("U :1l ro 8. Test de Student: Vegetac ión
Lelhoc eru s Belostoma Horvalhinia Tenaqobia IHeterocorixa Pelocoris Neoplea Buenoa TOTAL
Cvcerus 7 o o I 3 o 3 27 75 115
Eichhornia 44 3 I 1 226 o 8 60 121 463
I tvrnenachne 1 o o 18 I o 2 4 1 26
Lemna o o o o o o o 6 6
Mar silea 19 2 12 o 54 3 35 19 144
Pa5palum o o o 167 o o 2 2 171
I Pi5lia o o o o o 1 o 17 18
Pohqonum o o o 15 o 3 o 30 48
Salvinia 3 o o 5 o 12 22 12 54
Tha lia 12 1 9 o 16 8 48 32 126
1II1ixta 12 o o 35 o 17 793 69 926
TOTAL 98 6 22 469 70 57 991 384 2097
TEST
Cvp - Salv 0,22091 I --- - - - - - - - -- 0,32166 - ---- 0,25358 0,41741 0,09296 0,18747
Eich-Mixta 0,2643 3 0,09805 I 0,24998 0,19773 I - ------ 0,12432 0,01695 0,44477 0,02901
Existe también un claro factor de la vegetación de manera general entre Eichhornia y Mixta
(p =0,029), siendo estas además las que mayor abundancia de hemípteros acuáticos
presentan. El intervalo de confianza fue del 90 % (Cuadro 7)--- - ---- - - -- - - ---- _.
EICHHORNIALe/hocerus
MIXTA
Neoplea
Figura 42 . A bunda ncia de hemípteros ac uáticos por vegetació n. E l tam año de cada
gnífica indica su rna tor ia total de los organism os col ectados por vegetac ion.
Respecto al factor vegetació n se observa en la figura 42 la abundancia de organ ismos por
vegetación, Eichhornia presenta mayor abundancia para él género"Tenagobia y Buenoa,
mientras que vegetación Mixta presentan mayor abundancia para el género Neoplea.
61

(¡. Discusión
I :1" :¡~lIas del río Mamaré según Corbin el al. (1988) siguen procesos eventuales regulados
por el ciclo hidrológico que determina directamente la calidad del agua. Es posible que la
~'¡,c)(:a ele inundación provoque homogenización en la calidad de las aguas ubicadas en la
;',\11:¡ de influencia del río Mamaré, en nuestro caso en las lagunas Del Medio y Colorada,
C\...rcanas al río. Las lagunas de sabana (Belén y Matiquipiri) muestran características
dilcrcntes variando las aguas de estas últimas solo en función de las lluvias locales, así
:11 h,-'r~ando pequeños habitats que crean su propia diversidad ambiental.
"e~(¡n .Iunk el al. (1989), la diversidad ambiental en las lagunas de las llanuras fluviales, es
reconocida como compleja, particularmente por la alternancia de inundación yaguas bajas.
L;¡ Ilunura de inundación del Mamaré es un claro ejemplo de este funcionamiento, por sus
variaciones morfológicas y las diferencias tanto físicas como químicas en sus lagunas,
sumado a esto el grado de inestabilidad de las condiciones ambientales.
ln este estudio, los hemípteros acuáticos muestran riqueza y abundancia indistinta en la
mayoría de los sitios de muestreo. Se ha precisado observar y tratar de entender mejor a la
comunidad de hemípteros acuáticos separando tres tipos factores que pueden influir en
riqueza y abundancia respecto a su distribución: el factor época, referido a las épocas
estiaje e inundación, el factor laguna, por la ubicación espacial de las lagunas respecto al
río Mamaré y el factor vegetación, referido a la preferencia de los hemípteros por alguna
especie vegetal acuática.
Factor época
El factor época puede causar cambios en los factores físicos y químicos en las lagunas de la
llanura de inundación del Mamaré (Pouilly el al. 2004), lo que influenciaría el
comportamiento de la comunidad de hemípteros acuáticos. En un estudio sobre la ecología
de hemípteros acuáticos, Ward (1992) pone como ejemplo el cambio en la conductividad
eléctrica (ocurrido por la época) con la abundancia de algunos géneros, esta conductividad
62

define la concentración de sestón, acompañado del incremento de comunidades de
vooplancton y fitoplancton, dando lugar a una mayor abundancia de géneros predadores
como Buenoa (Fam. Notonectidae) y Tenagobia (Fam. Corixidae), pero no inl1uycndo en la
riqueza de hemípteros, puesto que otros géneros no presentan la misma dieta.
De la misma manera, Jansson el al. (1972) indican que en el período de inundación se
observan temperaturas medias (20 a 24 "C) y valores altos de conductividad (mayor a 300
'-IS). se presume que la gran concentración de nutrientes caracteriza una gran productividad
l'ri maria. esto aumentaría el numero de géneros de hemípteros afines a la dieta de plancton
atrayendo estos como parte de la cadena trófica a peces pequeños o alevinos. De acuerdo al
estudio de Ribeiro el al. (1998), los predadores efectivos de la comunidad de hemípteros
acuáticos como Lethocerus (Fam. Belostomatidae) y Pelocoris (Fam. Naucoridae) se ven
lavorccidos, puesto que presentan entre su dieta anfibios como también peces pequeños.
En época de estiaje, según Pouilly et al. (1999) cada laguna es característica y distinta de
las otras por la evolución propia de sus aguas, entonces, cada laguna debería albergar una
variedad de organismos característicos durante esta época, lo cual difiere en época de
inundación cuando las comunidades tienen oportunidad de mezclarse. Pero, según el
estudio realizado por Lino el al. cit. en Pouilly el al. (2004), sobre familias de
macroinvertebrados bentónicos, no se observan diferencias en la riqueza pero si en la
abundancia de hemípteros acuáticos encontrados en época de estiaje, de la misma manera
que en nuestro estudio, mostrando que la composición de la comunidad hemípteros es
similar a lo largo de la llanura de inundación del río Mamaré, lo que puede indicar que los
organismos tienen un amplio rango de tolerancia, debido a las características extremas de
su ambiente.
En época de estiaje, según Bachmann (1981) ocurriría migración de algunos géneros, esto
explicaría los cambios observados en abundancia en algunos géneros por época, pero
siempre manteniendo la misma riqueza. Por otro lado, un trabajo sobre abundancias en
cinco familias de hemípteros acuáticos realizado por Ribeiro el al. (1988), indica que en
época de inundación, los hemípteros presentan un decremento continúo en abundancia y no
63

en riqueza, argumentando que a mayor nivel de agua existe menor concentración de
:\1 imento o presas por tanto menor abundancia. En nuestro caso es posible que suceda lo
mismo. pero también se puede argumentar que el decremento en abundancia es
<implemente un efecto de la dilución debido a la mayor expansión que sufre la laguna al
elevarse el nivel de agua, creando nuevos habitats y dando lugar a una menor abundancia
de organismos.
I:Y; hemípteros acuáticos son claramente influenciados por la mayor expansión o retracción
Lk úrea y por la profundidad de un cuerpo de agua, como se indicó anteriormente, pero
también se verían influenciados por las características físicas y químicas del medio, esto
sumaria otro factor, el factor laguna, por el cual existe variación en riqueza y abundancia
entre los diferentes lagunas.
Factor laguna
l.lli, & Borden cit. en Bachmann (1981), en un estudio de diferentes tipos de cuerpos de
agua. indican que los géneros Tenagobia y Helerocorixa (Fam. Corixidae) al igual que el
género Neoplea (Fam. Pleidae), se presentan en gran abundancia en aguas lénticas, el
género Tenagobia (Fam. Corixidae) no abandona su ambiente acuático aún cuando éste se
hace inhabitable, como sería el caso de la Laguna Del Medio (laguna caracterizada por
disminuir sus aguas en época de estiaje), puesto que tiene la desventaja este género de estar
incapacitado para la locomoción terrestre según Bachmann (1981), como también presentar
una alta tolerancia a los cambios bruscos en su medio según Usinger (1956). Lino el al.
(2004) muestra una tendencia de distribución del género Neoplea en lagunas de bosque
galería, en nuestro caso las lagunas Colorada y Del Medio, más que en lagunas de sabana
(laguna Belén) pudiéndose deber a las características de las aguas y la influencia en estas
por el Mamaré.
Macan (1938) indica que es raro observar al género Tenagobia (Fam. Corixidae) en
ambientes acuáticos con más de dos metros de profundidad como los que presentan las
lagunas Colorada y Del Medio, esto puede deberse a que Tenagobia por ser una especie de
menor tamaño prefiere lugares poco profundos con poca materia orgánica, así los
64

ambientes de las lagunas de sabana como Belén y Matiquípiri hacen que esas especies sean
más frecuentes .
.'\L!elllús de la profundidad, la gran abundancia de Tenagobia (Fam. Corixidae) en la laguna
13e lén (laguna de sabana), puede también deberse a la buena visibilidad que presenta esta
laguna por encontrarse en medio de la sabana. Según Macan (1962) las lagunas que
prcxcntun buena visibilidad atraen hemípteros que tienden a migrar y que presentan
colonización masiva. Por otro lado, la familia Corixidae se caracteriza por la rapidez con la
que colonizan un ambiente (Bachmann 1962), otra razón por la cual se encontrarían en gran
abundancia en la laguna Belén.
f)opham (1964) indica que la atractividad de un ambiente acuático particularmente para los
géneros de la familia Pleidae puede estar dada por el brillo de la superficie del cuerpo de
agua. Los vuelos de migración de esta familia los realizan por la noche (Bachman, 1981),
así que el brillo nocturno que presentan algunos cuerpos de agua hace que estos géneros
sean afines a ocupar ese hábitat. Por esta razón se presume que las características físicas de
la laguna Belén con sus aguas transparentes y poca vegetación cubriendo la superficie,
hacen que Neoplea sea afín a ese hábitat, razón de la presencia de este género para esa
laguna.
Según Ribeiro el al. (1998), el comportamiento de los hemípteros acuáticos es también
influenciado por la profundidad del medio. Se presume que en cuerpos de agua poco
profundas, el sustrato sufre procesos muy rápidos de descomposición causados por la
cstacioualidad presentando así gran cantidad de alimento para muchos invertebrados
bentónicos y filtradores. Así el género Buenoa (Fam. Notonectidae), en el estudio de
Ribeiro, acompaña ese patrón debido a que su población aumenta en períodos más secos
donde ha ocurrido descomposición de materia orgánica de fondo. Además, se debe subrayar
que los organismos de la familia Notonectidae son resistentes a temperaturas altas respecto
a los demás hemípteros (Gittelman 1975 ), las que presentan estas lagunas de sabana en
época de estiaje.

l.l sustrato puede ser otro condicionante para los cambios en abundancia y riqueza de la
comunidad de hemípteros acuáticos, puesto que forma hábitat y protección contra
depredadores y ofrece un recurso alimenticio para los géneros filtradores como Buenoa
(Riheiro et al. 1998) y Tenagobia (Bachmann 1981). Esto corrobora Friday (1987)
indicando que un sustrato arenoso y pobre en materia orgánica se presenta una fauna de
hemípteros acuáticos reducido en riqueza y abundancia.
\\';ml (1992) también indica que en sustratos arcillosos formados en lagunas de aguas
blancas se dificulta la presencia de especies bentónicas. Esto puede deberse al sedimento
íino. el cual retiene menor cantidad de materia orgánica, alterando la estructura trófica de
ioda la comunidad, y modificando la relación predador-presa debido a la poca visibilidad en
las aguas turbias, reducción de la penetración de la luz, disminución térmica, y por ende
reducción de la producción primaria, características similares en las lagunas de bosque
galería las cuales presenta menor presencia de hemípteros acuáticos.
l.a teoría de MacArthur & Wilson según el trabajo de Friday (1987) indica que el área de
un cuerpo de agua es un factor importante de colonización de las especies. Probablemente
esto explica la baja diversidad de hemípteros acuáticos en épocas de estiaje en las lagunas,
también puede estar relacionado a los pocos recursos ofrecidos y los altos índices de
competición y predación con otros órdenes de la fauna béntonica.
Según Ribciro el al. (1998), la disminución del hábitat acuático provocado por la época de
estiaje, en asociación con las características de la laguna (diámetro, profundidad), hace que
la capacidad de dividir recursos aumente y consecuentemente el número de especies de la
comunidad de hemípteros acuáticos se reduzca. Estos cambios motivan a que las especies
que permanecen, tiendan a ser diferentes para coexistir y no presenten la necesidad de
partición de un mismo recurso, de acuerdo al concepto de Begon el al. (1988) sobre la
teoría de la perturbación. Así, las especies intermedias y algunas altamente tolerantes son
favorecidas, explicando esto la variación de abundancia pero no de riqueza en todas las
lagunas de nuestro estudio.
66

Probablemente estos procesos de cambio existentes en las lagunas ya sea por la época o por
I~\ ubicación de la laguna respecto al Mamaré, generan diferenciación en riqueza y
abundancia de hemípteros, como la distribución del género Buenoa (característico de la
laguna Colorada).
l.xistcn muchos estudios referidos a biomas acuáticos y/o la preferencia de algunas especies
de hemípteros, los cuales comprueban que la distribución de las especies principalmente
depende del equilibrio entre la preferencia de un ambiente y la tolerancia de la especie a ese
ambiente (Jansson & Scuddre 1972).
Lictor vegetación
Respecto al factor vegetación, en este trabajo no se observa una preferencia especifica a
nivel comunidad pero si a nivel genérico respecto a la vegetación, en el trabajo de
lvlcl.achlan (1969), una especie vegetal característica, puede aumentar la diversidad de
hemípteros acuáticos, ofreciendo al igual que el sustrato, recursos y protección. Esto se
demostrarla en las preferencias de vegetación de Eichhornia y vegetación mixta del género
Ncoplea (Fam. Plcidae), principalmente porque, al ser de tamaño pequeño, utiliza este tipo
de vegetación aferrado a las hojas y/o tallo como protección de sus predadores y camuflaje
para atacar a sus presas (crustáceos entomostracos y larvas de dípteros) (Bachman 1994).
l.a vegetación determina la diversidad según el trabajo de Ward (l992). La presencia de
Salvinia, Pistia y Eichhornia, en nuestro estudio, parece generar poca diversidad,
observado esto en la laguna Colorada. Estas especies vegetales, generando un colchón
vegeta! en la pe!ícula de agua, bloquean gran parte de la luz, disminuyendo la producción
autótrofa, mientras que otro tipo de vegetación como Cyperus, no bloquea la luz,
aumentando la diversidad y ofreciendo además recursos como también protección
(Mclachlan 1969).
Ribeiro el al. (1988) indican que las especies de Neoplea (Pleidae) se encuentran también
en el pleuston de Azolla y Lemna. Según Poi de Neiff (1977) y Neiff el al. (1994) se
67

observaría a Ncoplea asociada también a Pistia y Lemna. Bruquetas (1981) mencionan a
\'cojllea maculosa y Neoplea semipieta asociados a Eiehhornia como también a Salvinia y
.L:o!/o .
l.:IS especies de Tenagobia (Fam. Corixidae) son indicadoras de lagunas con grandes
concentraciones de sustrato orgánico proveniente de materia vegetal en descomposición,
pues muchas especies utilizan este sustrato como fuente de alimento y esa vegetación
muerta como protección (Hungerford 1948). Ward (1992) indica que muchas especies de
hemípteros buscan un hábitat con sustratos con mucha vegetación en descomposición.
1'\ll existe un estudio específico sobre los factores época, laguna o vegetación relacionados
«(\11 riqueza o abundancia en hemípteros acuáticos. Se observa que las especies encontradas
tienen un amplio espectro de colonización por la capacidad de varias familias de volar
(( 'orixidae, Notonectidae, Pleidae) y/o desplazarse por el medio terrestre (Belostomatidae,
Youcoridaei , y la capacidad de algunas familias de una rápida reproducción (Corixidae,
I'lcidae]. Así, en un ambiente tan dinámico e inestable como las lagunas de inundación del
Mamoré, la diversidad de hemípteros acuáticos generalmente no cambia sino las diferencias
entre las lagunas se expresan en la abundancia de las especies de este orden. Bruquetas et
al. (1981) también resalta lo versátil de su ecología respecto a los demás ordenes de los
organismos bentónicos.
68

7. Conclusiones
• Se han identificado 12 géneros con 16 especies, se presentan 5 géneros Sin
determinación de especie por encontrarse en estadios inmaduros, haciendo un total
de 17 géneros y 16 especies determinadas. No se ha podido registrar nuevas
especies para Bolivia.
• Las familias que presentan mayor abundancia son Pleidae con el género Neoplea y
la especie Neoplea maculosa, la familia Notonectidae con el género Buenoa y la
especie Buenoa salutis y la familia Corixidae con el género Tenagobia.
o El sitio de estudio que presenta más riqueza y mayor abundancia de hemípteros
acuáticos es la laguna Belén (laguna de sabana), la laguna del Medio (laguna de
bosque galería) presenta menor riqueza y abundancia de hemípteros acuáticos.
e Se encontró al género Buenoa asociado a la laguna Colorada por la riqueza y la
abundancia encontrados de acuerdo al análisis factorial de correspondencia (AFC).
• El género Tenagobia presenta alta riqueza y abundancia en las lagunas Belén y del
Medio (lagunas de sabana y bosque galería respectivamente), encontrándose esa
asociación en el análisis factorial de correspondencia (AFC).
• No se encontró relación entre la época y la abundancia de la comunidad hemípteros
acuáticos, descartando la relación entre la abundancia y la época, verificando esto el
test de Montecarlo, el cual no muestra significancia alguna.
• No se encontró relación entre las lagunas estudiadas y la abundancia de la
comunidad hemípteros acuáticos, puesto que es similar en todas las lagunas,
verificando esto el test de Montecarlo el cual no muestra significancia, descartando
la hipótesis de preferencia por alguna laguna en particular.
69

• No se encuentra asociación entre la vegetación del entorno de las lagunas y la
distribución de la comunidad de hemípteros acuáticos, descartando la preferencia a
nivel comunidad por alguna vegetación especifica, verificándose esto según el test
ele Montecarlo en cual no muestra significancia.
• Se encontró relación del género Tenagobia (Fam. Corixidae) y del género Pelocoris
(Fam. Naucoridae) con las épocas estiaje y transición, esto puede deberse al amplio
rango de supervivencia que presentan ambos géneros, además que Tenagobia es un
género característico por encontrarse en ambientes con poca cantidad de agua. Estas
diferencias son significativas según la prueba T de Student.
e En la laguna Colorada se encuentra la mayor abundancia de Buenoa por ser
característico de aguas profundas, mientras que la laguna Belén muestra gran
abundancia del género Tenagobia por ser un género característico por encontrarse
en cuerpos de agua de poca profundidad. Estas diferencias son significativas según
la prueba T de Student.
o En vegetación Mixta existe mayor abundancia de Neoplea lo cual se observa en las
diferencias encontradas con la prueba T de Student. También se comprobó
diferencias de abundancia para Eichhornia y vegetación Mixta mostrando
significancia según la prueba T de Student.
• Los géneros y especies identificadas en este estudio constituyen un primer aporte
para el conocimiento taxonómico y ecológico en esta zona, no pudiéndose
caracterizar habitats específicos para géneros o para la comunidad en general.
70

8. Bibliografía
o Aristizabal 1-1. 200 l. Los hemipteros de la película superficial del agua en
Colombia. Palie 1: Gerridae. Guadalupe Ltda. Bogota Colombia. 250 p.
• Bachmann, A. O. 1962. Clave para determinación de las subfamilias, géneros, y
especies de los Corixidae de la Republica Argentina. Physis 23'(64): 21-25 pp.
o Bachmann, A. O. 1966. Catalogo sistemático y clave para la determinación de las
subfamilias, géneros y especies de las Gerridae de la Republica Argentina (lnsecta.
Herniptera). Physis 26(71): 207-218 pp.
• Bachmann, A. O. 1971. Catalogo sistemático y clave para la determinación de las
subfamilias, géneros y especies de las Notonectidae de la Republica Argentina.
Physis 30(81): 600-617 pp.
" Bachmann, A. O. 1981. Insecta: Hemiptera Corixidae. Buenos Aires - Argentina.
270 p.
• Bachmann, A. O., Lopez Ruff M. 1994. Los pleoidea de la Argentina. La Plata.
Profadu (Conicet). 32 p.
• Berg, C. 1879. Hemiptera argentina. Ensayo de una onografia delos hemípteros
heteropteros y homopteros de la Republica Argentina. Anls. Soe. Cient. Argent, 8:
77-78 pp.
• Bonetto, A.A., 1976. Calidad de las aguas del río Paraná. Introducción a su estudio
ecológico. Dir. Nac. Constr. Port. y Vías Navegables. INCYTH-PNUD-ONU.
Buenos Aires. 202 p.
71

• Bourrel, L., S. Moreau & L. Phillips. 1999. Dinámica de las inundaciones en la
Cuenca amazonica Boliviana. Revista Boliviana de Ecología y Conservación
Ambiental 6: 5-17 pp.
• Bruquetas, 1.Y. Y1.J. Neiff, 1981. Decomposition and colonization by invertebrates
of Typha latifolia L. Iitter in Chaco cattail swamp (Argentina). Aquatic Botany, 40:
185-193 pp.
• Chessel D. & Dolédec S. 1996. Programmatéque ADE: Analyses multivariées et
rcpresentations graphiques de données écologiques. v 4.0 Université de Lyon.
Francc.
• Corbin D., Guyot J., Calle L. & Quintanilla 1. 1988 - Datos físico-químicos de los
medios acuáticos de la zona del Mamoré Central. Región de Trinidad - Amazonía
Bolivia. ORSTOM - UTB - CORDEBEN1. Bolivia. 57 p.
• Cummings, C. 1933. The giant water-bugs. Univ. Kansas Sci. Bul!. 21(2): 197-219.
tab. Xviii-xix. Lawrence Kansas.
• De Cario, 1. A. 1938. Los belostomatidos americanos. An. Mus. Argent. Cien. Nat.
39 (Entom.155): 189-260 pp.
• De Carlo, 1. A. 1956. Especies nuevas de la familia Belostomatidae (l-lem.) y
algunas consideraciones sobre otras poco conocidas. Rev. Soco Ent. Arg. 19 (3-4):
51-56 pp.
• De Carlo, J. A. 1961. 1. Una nueva especie del genero Ranatra y referencias de
Ranatra magna Kuitert (Ranatridae) Il. Una nueva especie del genero Belostoma
Latreille (Belostomatidae). Rev. Soco Ent. Arg, 24: 19 - 22 pp.
72

• De Carla, J. A. 1964. Genero Lcthocerus Mayr (Hemiptera, Belostomatidae).
PI-IYSIS 24 (68): 337-350 pp.
• Drago, E.e., 1994. The physical limnology of the river-lake systems of the Paraná
River floodplain. In: Sustaining the Ecological Integrity ofLarge Floodplain Rivers.
Internat, Conference, U.S. Dcp. of Interior, Nat. Biol. Survey, Univ. of Wisconsin.
La Crosse WI, July 12-15, 1994. (In press).
• Drake, C. J., Harris H. M. 1941. Notes sorne South American Gerridae (Hemiptera).
235-239 pp.
• Drake, C. J., Chapman, H. e. 1953. Preliminary report on the Pleidae (Herniptera)
ofthe Americas. Proc. Boil. Soco Washington 66: 53-60 pp.
• Drakc, C. J., Chapman, H. C. 1958. "The subfamily Saldoidinae ( Hemiptera:
Saldidae). Annas EntomologicaJ Society of America. 51: 480-485 pp.
o Dufour. 1863 Zaitha dilata. Ann. Soco Ent. France. (4) 3: 387 p.
• Ellis, R. A., Borden J. H. 1970. Predation by Notonecta undulata (Heteroptera:
Notonectidae) on larvae of yellow - fever mosquito. Annals of the Entomological
Socíety of America. 63 (4): 963 - 973 pp.
• Esaki, T. 1926. The water striders of the subfamily Halobatinae in the Hungarian
National Museurn. Annales Musei Nationalis Hungarici 23: 117-164 pp.
• Fernando, e.H. 1959. The colonization of small freshwater habitats by aquatic
insects. 2. Hemiptera (the water bugs) CeylonJ. Sci. (BioI.Sci) Colombo 2(1):
5-32 pp.
73

• Fossati O., Dumas P., Archairnbault Y., Fernandez B., Rocabado G., Wasson 1. G.
& Usseglio -Polatera P. 2002. Deriving life traits from habitat charactcristics : and
initial applications for neotropical invertebrates. 1. Rech. Oceanogr.
• Friday, L.E. 1987. The diversity of macroinvertebrate and macrophyte cornmunities
in ponds. Freshwater Biology. 18: 87-104 pp.
• Gittelman, S.H. 1975. The ecology of sorne Costa Rican backswimmers
(Hemiptera: Notonectidae). Annals of the Entomological Society of America. 68
(3): 511-518 pp.
• Hanagarth, W. 1993. Acerca de la geoecología de las sabanas del Beni en el Noreste
de Bolivia. Instituto de Ecología, La Paz, Bolivia. 186 p.
1) Holland, M.M. (Compiler), 1988. SCOPE/MAB technical consultations on
landscape boundaries. Report of a SCOPEIMAB workshop on ecotones. In: F. di
Castri, AJ. Hansen y M.M. Holland (eds.): A New Look at Ecotones: Emcrging
lnternational Projects on Landscape Boundaries. 47-106. Biology International.
Special Issue 17. IUBS. Paris.
o Horvarth, G. 1918. De hydrocorisis nonnulis extraeuropaeis. Annls. Mus. Nation.
Hungar. 6 : 140-146 pp.
• Hungerford, RB., 1919. The Biology and Ecology of Aquatic and Serniaquatic
Herniptera. Univ. Kansas Sc. Bul1. 11: 1-341 pp.
o Hungerford, H.B. 1948. The Corixidae of the Western Hemisphere (Hemiptera).
Scientific Bulletin ofthe University ofKansas. 21(1): 5 -195 pp.
• Hungerford H.B., 1954. The genus Reumatobates Bergtroth (Hemiptera- Gerridae)
J. Kansas. Ent. Soco 37 (7) : 529-588 pp.
74

.. Hungcrford, H.B., 1959. En Edmonsdson, W.T. (Ed). Freshwater Biology. 2da
edición. 958-972 pp.
• Hynes, H. B. N. (1948). "Notes on the aquatic Hemiptera -Heteroptera of Trinidad
and Tobago, B. W. L, with description of a new species of Martarega B. White
(Notonectidae). Trans. R. Ent. Soco Lond, 99(10): 341-361 pp.
• Jansson, A. & G.G. E. Scudder. 1972. Corixidae (Herniptera) as predators: rearing
on brozen brine shrimp. Journal of the Entomological Society of British Columbia,
69: 44-45 pp.
o Junk, W.J., 1970. Investigations on the ecology and production biology of thc
"floating meadows" (paspalo-Echinochloeturn) on the Middlc Amazon. !. The
íloating vegetation and its ecology. Amazoniana 2: 449-495 pp.
o Junk W. L, BayJey P. B. & Sparks R. E. 1989. The fiood pulse conccpt in rivcr
floodplain systems. In : International large river symposium, Can. Spcc. Pub!. Fish.
Aquat. Sci., 110-127 pp.
o Kirkaldy, G. W. 1899. On sorne aquatic Rhynchota from South America in the
col1ections of the museum of Zoology and Comparative Anatomy of the Royal
Univcrsity ofTurin. Bol1. Mus. Zoo!. Anat. Comp. Univ. Torino 14 (347); 1-2 pp.
• Kirkaldy, G. W. 1904. Buenoa salutis.Wiener entom. Z. XXIII: 124 p.
• Lebart, L., Morineau A. & Fénelon, 1. P. 1995. Tratamiento estadístico de datos.
Métodos y programas. Marcornbo. Barcelona. 593 p.
• Lebart, L., Morineau A. & Piron, M. 1985. Statisttique exploratoire
multidimensionnelle. Dunod. Paris. 439 p.
75

• Lcsack, L. F. W. 1993. Water balance and hydrologic characteristics 'of a rain forest
catchment in the Central Amazon Basin. Water Resources Research. 29 (3): 759
773 pp.
• Lopcz Ruf, M. 1994. "El genero Pelocoris en la Argentina (Heteroptera
Limnocoridae). 1. Introduccion, diagnosis, clave de especies y redescripcion de P.
(P.) Binoculatus nigriculus Berg, P. (P.) Binoculatus binoculatos (Stal) y P. (P.)
impicticollis Stal. PHYSIS 49 (116-117): 13-22 pp.
• Loubens,G., Lauzanne, L. & Le Guennec, 1992. Les milieux aquatiques de la région
de Trinidad (Beni, Amazonie bolivienne). Rev. Hydrobiol. trop., 25 (1) : 3-21 pp.
• Lundblad, O. 1929. Zur Kenntnis der Gattung Tenagobia Bergroth. Ark. Zool
Stockholm 20A (7): 1-22 pp.
• Macan. T. T. 1938. Evolution of aquatic habitats with special reference to the
distribution of Corixidae. 1.animo Ecol. London 7: 1-19 pp.
• Macan T. T. 1962. Why do sorne pieces ofwater have more Corixidae than others.
Arch. I-Iydrobiol. Stuttgart 58 (2): 224 - 232 pp.
• Marchese, M. and 1. Ezcurra de Drago, 1992. Benthos ofthe lotic environmcnts in
the Middle Paran á River System: transverse zonation. Hydrobiología 237: 1-13 pp.
• Mayr. 1871. Zaitha elegans. Verh. Zool. Bo1. Wien. 21: 415-416 pp.
• McLachlan, A. J. 1969. The effect of aquatic macrophytes on the variety and
abundante of benthic fauna in a newly created lake in the tropics (Lake Kariba).
Archiv für Hydrobiologie. 66: 212-231 pp.
76

• Mazzuconi, S., Bachmann A. 1995. "Geograpic distribution of the Gerridae m
Argentina (lnsecta, Heteroptera). Insecta Mundi 9: 363-369 pp.
41 Montandon, A., 1899. Zaitha martín. Bul!. Mus. Hist. Nat. Paris. 5 : 170 p.
• Montandon, A., 1900. Belostoma gestroi. Ann. Mus. Civ. Genova. 20 (40): 537
539 pp.
• Montandon, A., 1903. Hemiptéres aquatiques. Notes synonimyques et
geographiques, descriptions déspeces nouvelles.- Bull. Sci. Soco Bucarest, XII: 97
121 pp.
• Montandon, A., 1908. Belostoma elongatum. Ann. Hist. Nat. Mus. Hung. 7: 299 p.
41 Montes de Oca, 1. 1997. Geografia y recursos naturales de Bolivia. Academia
Nacional de Ciencias de Bolivia. La Paz. 614 pp.
• Neiff, J.1., 1978. Fluctuaciones de la vegetación acuática en ambientes del valle de
inundación del Paraná Medio. Physis, B, Buenos Aires, 85(38): 41-53 pp.
41 Neiff, J.1., 1990. Aspects of primary productivity in the lower Paraná and Paraguay
riverinc system. Acta Limnol. Bras. Vol. III. Tomo 1: 77-113 pp.
41 Nciff, J.1. y Poi de Neiff, A., 1990. Litterfall, leaf decomposition and litter
colonization of Tessaria integrifolia in the Paraná River floodplain. Hidrobiología
203( 1-2): 45 - 52 pp.
• Neiff, J.J., lriondo, M.H. and Carignan, R., 1994. Large tropical south american
wetlands: an overvicw. Pp. 156-165. In: Link, G.L. and Naiman, R.J. (eds.): The
Ecology and Management of Aquatic-terrestrial Ecotones. Proccedings book, Univ.
of Washington. 225 p.
77

O) Neiff J.J. 1996. Large rivers of South America: toward the new approach. Verh.
Internat. Verein. Limnol., 26: 167-180 pp. Stuttgart.
lO Nieser, N., 1975. The water bugs (Heteroptera: Nepomorpha) ofthe GutanaRegion.
Stud. Fauna Suriname 16: 1-308 pp.
<1 Nieser, N. Melo, A. L.; Pelli, A. & Campos, N. D. 1997. A new species of Buenoa
(Heteroptera: Notonectidae) from Minas Gerais (Brazil). Ent. Ber Amst. 57: 129
135 pp.
• Nieto Nafria, J.M. y M.P. Mier Durante., 1985. Tratado de Entomologia. Omega.
599 p.
" Orfeo, O., 1995. Aumento de carga sedimentaria por erosión de taludes en ríos
chaqueños, En: Neiff, J (Ed.): Contaminación en cursos de agua del Chaco oriental.
Convenio Gobierno de la Provincia del Chaco (COFEA)-Consejo Nacional de
Investigaciones Científicas y Técnicas (CECOAL), VII: 117-121 pp.
e Pennak, R.W., 1953. Freshwater Invertebrates ofthe United State. The Ronald Press
Company . 541 - 559 pp.
• Poi de Nciff, A., 1977. Estructura de la fauna asociada a tres hidrófitos flotantes en
ambientes leníticos del nordeste argentino. Comunicaciones Científicas del
CECOAL N° 6: 1-16 pp.
• Polhemus, D. A. 1976. A reconsideration of the status of the genus Paravelia
Breddin, with othcr notes and check list of species (Veliidae: Heteroptera). J.
Kansas Entomol. Soco 49: 509-513 pp.
78

• Polhernus, J. T., Spangler, P. J. (1995). A review of the genus Stridulivclia
f-Iungerford and two new species (Heteroptera: Veliidae) frorn South América.
Proc. Entomol. Soco Wash. 97(1): 128-152 pp.
• Popham, E. J. 1964. The migration of aquatic bugs with special reference to the
Corixidae (Hemiptera: Heteróptera). Archiv für Hydrobiologie, 60: 450-496 pp.
• Pouilly, M., Ibañez, C.,Gutierrez, M. & Yunoki, T. 1999 - Funcionamiento
ecológico de las lagunas de la zona de inundación del río Mamoré (Beni - Bolivia).
Rcv. Bol. de Ecol. 6: 41-54 pp.
• Pouilly, M., Beck, S., Moraes, M., & Ibañez, C. 2004. Diversidad Biologica en la
llanura de inundacion del río Mamoré: Importancia y ecología de la dinamica
fluvial. Ed. Centro de Ecología Simon & Patiño. Santa Cruz. Bolivia. 383 p.
• Puhakka, M. & R. Kallioma. 1993. La vegetación en áreas de inundación en la selva
baja de la amazonía peruana. Proyecto arnazonía, Universidad de Turku, Finlandia.
HONREN. Lima-Perú. 113-]38 pp.
• Ribeiro, J. R. L., Nessimian, J. L. & E. C. Mendoc;a. 1998. Aspectos da distribuicao
dos Nepomorpha (Hemiptera: Heteroptera) em corpos d'agua na Restinga de
Maricá, Estado do Río de Janeiro. Vol. V. ] 13-]28 p.
• Richards, O.W. y R. G. Davies., 1984. Tratado de Entomología. Imms. 2. Omega.
998 p.
• Roback, S.S., ]974. Pollution Ecology ofFreshwater Invertebrates. 333-336 p.
• Rocabado, G. 2004. Ecología y distribución de invertebrados en ríos andinos de la
cuenca Amazónica Boliviana. Tesis Doctoral. Tucumán Argentina. ]50 p.
79

• Roche, M. A. & Fernandez Jauregui, C. 1988. Water rcsources, salinity and salt
exportations of rivers ofthe Bolivian Amazon. J Hydrol., 101 : 305-331pp.
• Salm, H. & X. Flores. 1994. Potencialidades y limitantes para un desarrollo
sostenible del departamento del Beni como base para su plan de ordenamiento
territorial. Propuesta preliminar del Plan de Ordenamiento Territorial del Beni.
FUNDECO/fEIPROADE/GTZ. Lapaz, Bolivia. 107 p.
• Sanjines A. & S. Beck. 2004. Vegetación acuática y ribereña de las lagunas. En:
PouiJly, M., Beck, S., Moraes, M., & Ibañez, C. 2004. Diversidad Biologica en la
llanura de inundacion del río Mamoré: Importancia y ecología de la dinamica
fluvial. Ed. Centro de Ecología Simon & Patiño. Santa Cruz. Bolivia. 193-232 pp.
• Schnack, 1. A. (1976). "Estimación de parámetros en una población de Corixidae
(Hemiptera) en base a un modelo estocástico de captura y recaptura. Limnobios
1(1): 1-7 pp.
• Schuh, R. T. 1986. The influence of cladistics on heteropteran classification. Ann.
Rev. Entornol. 31: 67-93 pp.
• Sioli H. 1968 - Hydrochemistry and Geology in the Brasilian Amazon region.
Amazoniana. 1: 267-277 pp..
• Stal, c., 1876. Enumeratio Hemipterorum. 5 Kongl, Svensk. Akad. Handl., 14 (4):
1-143 pp.
• Tachet B., Richoux, P. Bournaud, M. & Usseglio-Polatera P. 2002. Invertébrés
d'eau douce systematique, biologié, ecologié. Ed CNRS. Paris. 588 p.
• Truxal. 1953. Buenoa salutis. Contr. Sci. Los Angeles County Mus. Brasil. (12) :
17 p.
80

• Usinger, R., 1956. Hemiptera en R. Usinger (ed). Aquatic Insects of California:
182-227 pp. univ. OfCalifornia Press.
• Ward, 1. V. 1992. Aquatic insect ecology. Biology. Biology and habitat. 1. Wiley &
Sons lnc., New York. 438 p.
• White, F. B. 1879. List of the Hemiptera collected in the Amazons by Prof. 1. M.
Trail, M. A., M. D., in 19873-1875, with description ofthe new species. Trans. Roy.
Ent. Soco London 1879 (4) : 267-276 pp.
81

METODOLOGIA DE COLECTA
,,;, ,

ZONA DE ESTUDIO
Mezcla de aguas del río Mamar é (color café) y del río Ibare (color oscuro)
Río Mocovi: Aguas negras yaguas blancas

SITIOS DE MUESTREO
La guna Belén
M atiq uipir i
30 S: 7P

Laguna Del Medio
0: '" A
Laguna Colorada


















