DETECCIÓN DE LOS GENES QUE CODIFICAN SUPERANTÍGENOS …
113
DETECCIÓN DE LOS GENES QUE CODIFICAN SUPERANTÍGENOS EN Staphylococcus aureus AISLADOS DE MADRES COMUNITARIAS DEL INSTITUTO COLOMBIANO DE BIENESTAR FAMILIAR DE PEREIRA. GLADYS FERNANDA FLÓREZ RESTREPO UNIVERSIDAD TECNOLÓGICA DE PEREIRA FACULTAD CIENCIAS DE LA SALUD CENTRO DE BIOLOGÍA MOLECULAR Y BIOTECNOLOGÍA PEREIRA 2015
Transcript of DETECCIÓN DE LOS GENES QUE CODIFICAN SUPERANTÍGENOS …

DETECCIÓN DE LOS GENES QUE CODIFICAN SUPERANTÍGENOS EN Staphylococcus aureus AISLADOS DE MADRES COMUNITARIAS DEL
INSTITUTO COLOMBIANO DE BIENESTAR FAMILIAR DE PEREIRA.
GLADYS FERNANDA FLÓREZ RESTREPO
UNIVERSIDAD TECNOLÓGICA DE PEREIRA FACULTAD CIENCIAS DE LA SALUD
CENTRO DE BIOLOGÍA MOLECULAR Y BIOTECNOLOGÍA PEREIRA
2015

DETECCIÓN DE LOS GENES QUE CODIFICAN SUPERANTÍGENOS EN Staphylococcus aureus AISLADOS DE MADRES COMUNITARIAS DEL
INSTITUTO COLOMBIANO DE BIENESTAR FAMILIAR DE PEREIRA.
GLADYS FERNANDA FLÓREZ RESTREPO
TRABAJO DE GRADO PARA OPTAR AL TÍTULO DE MAGÍSTER
EN BIOLOGÍA MOLECULAR Y BIOTECNOLOGÍA
DIRECTOR JOSÉ IGNACIO MONCAYO ORTIZ
MSc. MICROBIOLOGÍA
UNIVERSIDAD TECNOLÓGICA DE PEREIRA FACULTAD CIENCIAS DE LA SALUD
CENTRO DE BIOLOGÍA MOLECULAR Y BIOTECNOLOGÍA PEREIRA
2015

Nota de aceptación:
___________________________________ ___________________________________ ___________________________________ ___________________________________ ___________________________________ ___________________________________
____________________________________ Firma del presidente del jurado
____________________________________
Firma del jurado
____________________________________ Firma del jurado
Pereira, __25__ de Septiembre de 2015

DEDICATORIA
A Jerónimo y Luis Gabriel, son la luz de mis ojos, el motor que me ayuda a seguir adelante, los amores de mi vida.
A mis padres por darme el apoyo y enseñanzas. A mis hermanos por creer en mí; y todos los que me acompañaron en este largo
proceso.

AGRADECIMIENTOS Quiero expresar mi profundo agradecimiento al Doctor José Ignacio Moncayo Ortiz, por su dedicación, paciencia y colaboración, para guiarme en este proceso. Doctor Álvaro Hernán Alegría, por su entereza en las clases, darme siempre los mejores consejos. Doctora Lucero Rengifo, por su cariño, ser una mujer intachable y un ejemplo a seguir. Doctora Adalucy Álvarez, Doctora Yina Marcela Guaca, por su orientación, paciencia en la etapa experimental y al Doctor Duverney Gaviria, por su apoyo en transcurso de la maestría A la Universidad Tecnología y sobre todo a CENBIOTEP por su apoyo incondicional. Al Laboratorio clínico del Hospital Universitario San Jorge en cabeza de su directora Doctora Carmen Elisa Llanos, a la bacterióloga Miriam Gómez, y a las directivas. A la Doctora Adriana Patricia Hoyos Sánchez y las directivas del Instituto Colombiano de Bienestar Familiar de Pereira, y especialmente a las madres comunitarias por haber aceptado participar en el estudio. A todas las personas que me apoyaron para alcanzar esta meta.

CONTENIDO Pág.
CONTENIDO
RESUMEN ............................................................................................................... 1
ABSTRACT ............................................................................................................. 2
INTRODUCCIÓN ..................................................................................................... 6
1. OBJETIVOS ...................................................................................................... 8
GENERAL ............................................................................................................ 8
ESPECÍFICOS ..................................................................................................... 8
2. MARCO TEÓRICO Y ESTADO DEL ARTE. ....................................................... 9
2.1 Reseña histórica. ....................................................................................... 9
2.2 Taxonomía, genoma y la diversidad genética. ................................... 9
2.3 Morfología y estructura antigénica. ..................................................... 9
2.4 Factores de virulencia. ....................................................................... 11
2.4.1 Enzimas. ........................................................................................ 11
2.4.2 Toxinas. ......................................................................................... 12
2.4.3 Los superantígenos. ..................................................................... 14
2.4.3.1 Clasificación estructural de los superantígenos. ................... 14
2.4.3.2 Clases de Superantígenos de S. aureus. ................................. 15
2.4.3.2.1 Toxina del síndrome de choque tóxico. ............................ 15
2.4.3.2.2 Las toxinas exfoliativas. ..................................................... 16
2.4.3.2.3 Enterotoxinas (SE, staphylococcal enterotoxin). ............. 17
2.4.3.2.3.1 Estructura de las Enterotoxinas. ................................. 20
2.4.3.2.3.2 Mecanismo de acción. .................................................. 21
2.4.4 Regulación de factores de virulencia en S. aureus. .................. 22
2.4.4.1 El sistema regulador Agr y otros sistemas reguladores. ....... 23
2.5 Epidemiología de la infección. ........................................................... 25
2.5.1 Epidemiología de las intoxicaciones alimentarias. ....................... 28
2.5.2 Epidemiología de portadores sanos: impactos de los portadores sanos de S. aureus en salud pública....................................................... 28

2.6 Patogénesis y enfermedades asociadas. .......................................... 30
2.7 Diagnóstico de la infección. ............................................................... 32
3. METODOLOGÍA. ............................................................................................... 33
3.1 Tipo de estudio. .......................................................................................... 34
3.2 Diseño maestral. ......................................................................................... 34
3.2.1 Población. ............................................................................................. 34
3.3 Técnicas y procedimientos ........................................................................ 34
3.3.1 Procedimiento para obtención de S. aureus. ..................................... 34
3.3.2 Diagnóstico Molecular. ........................................................................ 35
3.3.2.1 Extracción de ADN. ....................................................................... 35
3.3.2.2 Detección genotípica de superantígenos en los aislamientos de S. aureus. ................................................................................................... 36
3.4 Análisis estadístico. ................................................................................... 40
4. RESULTADOS. ................................................................................................. 40
4.1 Identificación de S. aureus. ....................................................................... 41
4.2 Prevalencia de estado de portador. .......................................................... 41
4.3.1 Prevalencia individual de los genes de SAg en los aislamientos de
S. aureus. ....................................................................................................... 43
4.3.2 Frecuencia de asociación de los genes de SAg (genotipos). .......... 43
4.4 Susceptibilidad a los antimicrobianos. .................................................... 44
4.5 Asociación entre la detección de los genes de SAgs y la resistencia
antimicrobiana. ................................................................................................. 45
5. DISCUSIÓN ....................................................................................................... 47
6. CONCLUSIONES. ............................................................................................. 51
7. RECOMENDACIONES ...................................................................................... 54
8. BIBLIOGRAFÍA. ................................................................................................ 55
ANEXOS. ............................................................................................................... 73

LISTA DE TABLAS Pág.
Tabla 1. Características generales y localización genómica de los………......18 genes de enterotoxinas (SEs – Sels) del S. aureus. Tabla 2. Clasificación de las enterotoxinas de acuerdo a sus………………....21 secuencias de aminoácidos.
Tabla 3. Cepas de referencia ATCC de S. aureus utilizadas……………………36 como control positivo. Tabla 4. Iniciadores específicos para las enterotoxinas clásicas……………..38 de S. aureus. Tabla 5. Iniciadores para la Toxina del Síndrome de Choque………………….39 Tóxico y las Toxinas Exfoliativas y para etd. Tabla 6. Número de genes de superantígenos detectados en los………….....43 aislamientos de S. aureus. Tabla 7. Prevalencia individual para los genes de enterotoxinas……………..43 clásicas, toxinas exfoliativas y toxina del síndrome del choque tóxico en los aislamientos de S. aureus. Tabla 8. Prevalencia de la asociación de genes de SAgs en…….…………….44
un mismo aislamiento.
Tabla 9. Porcentaje de aislamientos de S. aureus según la resistencia……..44 a uno o más antimicrobianos. Tabla 10. Prevalencia de resistencia en los aislamientos de…………….…….45 S. aureus a diferentes antimicrobianos incluyendo la producción de β-Lactamasa. Tabla 11. Asociación entre uno o más genes de SAgs y ………..……………..45 la prueba de β-Lactamasa positiva (β -L+) y la prueba de β-Lactamasa negativa (β -L-). Tabla 12. Asociación entre uno o más genes de SAgs y ………..……………..46 la susceptibilidad a cefoxitina-oxacilina. Tabla 13. Asociación entre uno o más genes de SAgs y la…………………….46 susceptibilidad a eritromicina, clindamicina y tetraciclina.

Tabla 14. Asociación entre la presencia o ausencia SAgs y la………….…….47 prueba de la β-Lactamasa. Tabla 15. Asociación entre la presencia o ausencia SAgs y la………….…….47 susceptibilidad a cefoxitina/ oxacilina. Tabla 16. Asociación entre la presencia o ausencia SAgs y la………….…….47 susceptibilidad a eritromicina. Tabla 17. Asociación entre la presencia o ausencia SAgs y la………….…….47 susceptibilidad a clindamicina. Tablas 18. Asociación entre la presencia o ausencia SAgs y la………….…...47 susceptibilidad a tetraciclina.

LISTA DE FIGURAS
Pág.
Figura 1. Funcionamiento del locus agr...…………………………………………25 Figura 2. PCR múltiple de la serie A. Detección de los genes sea, seb,..……41 sec, sed y see. Figura 3. PCR múltiple para la serie B. Detección de los genes tsst-1....……42 y eta. Figura 4. PCR simple para la detección de la toxina exfoliativa D…………....42 (gen etd).

ANEXOS Pág.
Anexo A. Número de aislamientos positivos para S. aureus,………………….73 aislados de Madres Comunitaria. Anexo B. Prevalencia de estado de portador para la población total………...78 Anexo C. Base de datos de genes de superantígenos en los………………….79 aislamientos de S. aureus. Anexo D. Base de datos de número de genes de superantígenos…………....82 detectados en los aislamientos de S. aureus.
Anexo E. Tabla de frecuencia de la prevalencia global de los genes………...84
de SAg.
Anexo F. Tabla de frecuencia de número de genes de SAg...………………….85
Anexo H. Tablas de frecuencia de asociación de los genes de SAg…………87
Anexo I. Base de datos de los antimicrobianos…………………………………..90 Anexo J. Total de genes en los aislamientos de S. aureus ……………………91 según la resistencia a uno o más antimicrobianos.
Anexo K. Tablas de frecuencia para la prevalencia de resistencia en……….93 los aislamientos de S. aureus a diferentes antimicrobianos. Anexo L. Tablas de contingencia para las asociaciones entre la……………..94 detección de los genes de SAgs y la resistencia antimicrobiana.

1
RESUMEN La intoxicación alimentaria es un importante problema de salud pública a nivel mundial, en la intoxicación alimentaria muchos manipuladores de alimentos pueden ser portadores sanos de cepas de S. aureus en sus fosas nasales o en sus manos, estos son considerados como una de las principales fuentes de contaminación en alimentos, a través del contacto manual o mediante las secreciones respiratorias. El propósito del presente estudio fue detectar los genes que codifican los superantígenos en S. aureus aislados de madres comunitarias del Instituto Colombiano de Bienestar Familiar de Pereira (ICBF) Seccional Risaralda. Se tomaron 406 muestras en fosas nasales y manos de las madres comunitarias. El aislamiento se realizó por métodos microbiológicos y la confirmación por métodos microbiológicos y moleculares. Se obtuvieron 95 aislamientos de S. aureus. A los aislamientos se les determinó el perfil de susceptibilidad antimicrobiana por sistema microbiológico automatizado. La detección de los genes de enterotoxinas clásicas (SEA a SEE), toxinas exfoliativas (ETA y ETB) y la toxina del síndrome de choque tóxico – 1 (TSST-1), se realizó por medio de la reacción en cadena de la polimerasa única y/o múltiple. La prevalencia del estado de portador entre fosas nasales y/o manos fue del 22%. El 17% fueron portadores en fosas nasales, 5% en manos y 1,5% tanto en fosas nasales y manos. La prevalencia global de los genes de fue 73,7%. La prevalencia individual determinó la mayor frecuencia del gen etd 47,4% y los de menor frecuencia see - eta con 4,2%. En los 95 aislamientos se encontró mínimo un gen con 34,7%, máximo cuatro genes con 4,2%. La frecuencia de asociación entre genes, permitió establecer 8 genotipos diferentes para dos o más genes. La mayor asociación encontrada fue sea y tsst-1 con 10%. La susceptibilidad antimicrobiana mostró que el 74,7% de los aislamientos eran resistentes al menos a un antibiótico y 3,2% eran resistentes a cuatro antibióticos. El 93,7% de los aislamientos fueron positivos para la producción de β–Lactamasa, seguido de la resistencia a tetraciclina con 11,6% y el de menor resistencia fue a clindamicina con 3,2%. La mayoría de las posibles asociaciones entre SAgs y la resistencia antimicrobiana, no mostraron diferencias significativas, la única asociación que mostró diferencia significativa fue con tetraciclina. Este es un riesgo potencial en la transferencia en

2
la comunidad de cepas enterotoxigénicas y resistentes. La detección de los genes que codifican los superantígenos, mostraron una alta prevalencia, indicando que S. aureus posee una gran variación genética; que se ha encontrado en aislamientos hospitalarios, en alimentos y en manipuladores de alimentos sobretodo en cepas de origen nasal. El gen de mayor prevalencia fue etd, siendo un hallazgo novedoso en aislamientos de portadores nasales o manos, a diferencia de los otros genes que han reportado con frecuencia en otros estudios, dicho gen puede interrumpir la barrera cutánea epitelial y de esta manera contribuir a la propagación bacteriana. Palabras clave: Staphylococcus aureus, superantígenos, toxina del síndrome del choque tóxico, toxina exfoliativa, enterotoxinas.
ABSTRACT

3
Food poisoning is a major public health problem worldwide, in the food poisoning, many food handlers can be healthy carriers of S. aureus in their nostrils or in their hands, and they are considered as one of the main sources food contamination through hand contact or through respiratory secretions. The purpose of this study was to detect genes encoding the superantigen in S. aureus isolated from community mothers of the Colombian Institute of Family Welfare Pereira (ICBF) Seccional Risaralda. Were taken 406 samples nostrils and hands of community mothers. The isolation was performed by microbiological methods and confirmation by microbiological and molecular methods. Were obtained 95 S. aureus isolates. The isolates were determined antimicrobial susceptibility profile for automated microbiology system. The isolates were determined antimicrobial susceptibility profile for automated microbiology system. Detecting genes classical enterotoxins (SEA to SEE), exfoliative toxins (ETA and ETB) and toxin toxic shock syndrome - 1 (TSST-1), it was performed by the chain reaction polymerase and / or multiple. The prevalence of carrier status among nostrils and / or hands was 22%. It was also established that 17%, were nostrils carriers in 5% held 1.5% of the total community mothers were carriers in nostrils with in hands. The overall prevalence of the genes was 73.7% and the prevalence individually determined most frequency of 47.4% etd gene and less frequently see - eta with 4.2%. At least 95 isolates found a gene with 34.7%, maximum than four genes with 4.2%. The frequency of association between genes, 8 different genotypes allowed establishing two or more genes. The largest association was found and TSST-1 is 10%. The antimicrobial susceptibility showed that 74.7% of the isolates were resistant to at least one antibiotic and 3.2% were resistant to four antibiotics. The 93.7% of the isolates were positive for β-lactamase production; and continued resistance to tetracycline was 11.6% and the lowest resistance to clindamycin was 3.2%. Most possible associations between SAgs and antimicrobial resistance, showed no significant difference, the only association that showed significant difference was tetracycline. This is a potential risk in the transfer of enterotoxigenic community and resistant strains. Detecting genes encoding superantigens, showed a high prevalence, indicating that S. aureus has a genetic variation, this event has also been found in hospitals isolates

4
in food and food handlers nasal strains especially in origin. The gene most prevalent was etd, showing a novel finding in isolates nasal carriers or hands, unlike other genes that have been reported frequently in other studies, the gene can disrupt the skin barrier epithelial and thus contribute to bacterial propagation. Keywords: Staphylococcus aureus, superantigen, toxic shock syndrome, exfoliative toxin, enterotoxin.

5

6
INTRODUCCIÓN El S. aureus es un comensal común de la piel y las membranas mucosas en los seres humanos, con estimaciones del 20 a 30% de persistencia y el 60% para la colonización intermitente. El S. aureus es considerado como la principal causa de enfermedad nosocomial e intoxicación alimentaria. Está asociado a enfermedades supurativas de la piel como foliculitis, impétigo, furúnculos, hidradenitis supurativa y celulitis. También produce otras enfermedades más severas como síndrome de choque tóxico, endocarditis, osteomielitis, meningitis y neumonía. La intoxicación alimentaria estafilocócica es un importante problema de salud pública a nivel mundial, la más frecuente de las intoxicaciones alimentarias es la alimentaria estafilocócica, que resulta del consumo de alimentos en los que el S. aureus, está presente por encima de 110 UFC/g o ml, (Según la normatividad colombiana- INVIMA), permitiendo sintetizar factores de virulencia como la familia de las exoproteínas o exotoxinas con actividad superantigénica y pirógenica. Muchos manipuladores de alimentos pueden ser portadores de cepas de S. aureus en sus fosas o en sus manos, estos son considerados como la principal fuente de contaminación de los alimentos, además de las deficientes buenas prácticas de manufactura (BPM), la poca identificación de puntos de control críticos en el proceso y la contaminación cruzada con alimentos contaminados, superficies y equipos. La intoxicación alimentaria estafilocócica, ocurre cuando se alcanza un rango 0,1 – 1,0 µg/kg, en los niveles de toxina, los datos epidemiológicos están en los boletines SIVIGILA, que ha reportado el 51% de los casos es producido por ETA (enfermedades transmitidas por alimentos), probablemente está subestimada por una serie de razones, que incluyen un diagnóstico equivocado, sobre portadores o intoxicaciones, no se publican las intoxicaciones menores, inadecuada e incorrecta recogida de muestras y exámenes de laboratorio. El diagnostico de portador sano es de gran importancia social y económica, para evitar futuras intoxicaciones, la diseminación en la población y para tener un control epidemiológico de dicha enfermedad; ya que las madres comunitarias tienen la responsabilidad de cuidar y alimentar niños de diferentes edades. Los superantígenos de S. aureus son exoproteínas, que incluye entre otras las enterotoxinas reconocidas como causantes de intoxicación alimentaria, dejando como secuela el síndrome de choque tóxico. Los genes que codifican para superantígenos, han sido localizados en bacteriófagos, plásmidos e islas de patogenecidad. Todos estos aspectos están siendo intensamente estudiados por diferentes grupos de investigación en el mundo. Sin embargo, en Colombia existen pocas investigaciones relacionadas con la genotipificación de estos factores de virulencia. El S. aureus posee la habilidad de producir una gran cantidad de factores de

7
virulencia. Estos factores de virulencia son los responsables de la alta patogenecidad del microorganismo, que sumado a su capacidad para evadir la respuesta inmune y la resistencia a un amplio rango de antibióticos y desinfectantes, lo convierten en un problema debido a su difícil tratamiento y a sus altos niveles de supervivencia en objetos inanimados. Entre los factores de virulencia producidos por las cepas más patógenas se encuentran los superantígenos. Estos producen la activación de un gran número de linfocitos T (alrededor del 20%), mientras que los antígenos convencionales activan sólo un 0.01%. Adicionalmente, desencadenan respuesta a muy bajas concentraciones, sin requerir la especificidad de unión como sucede con los antígenos convencionales e inducen la liberación de citocinas proinflamatorias, lo cual genera una fuerte respuesta inflamatoria que puede producir severas complicaciones como el choque tóxico. El propósito de esta investigación es continuar los estudios de detección y expresión de los genes de superantígenos en aislamientos de S. aureus diferentes a las de origen hospitalario, como son los asilamientos en madres comunitarias; lo cual permitirá hacia el futuro buscar relaciones entre el estado de portador y la intoxicación alimentaria. Además, en Colombia no existen muchas investigaciones relacionadas con superantígenos y el estado de portador, la mayoría de los estudios están enfocados a la determinación de la resistencia antimicrobiana y el seguimiento epidemiológico de infección nosocomial por S. aureus meticilino resistente.

8
1. OBJETIVOS
GENERAL Detectar genes que codifican los superantígenos en Staphylococcus aureus aislados de fosas nasales y manos de madres comunitarias del Instituto Colombiano de Bienestar Familiar de la seccional Pereira (ICBF).
ESPECÍFICOS Describir la prevalencia del estado de portador en fosas nasales y/o manos de S. aureus aislados en la población de estudio. Identificar la frecuencia de cada uno de los genes que codifican las enterotoxinas clásicas (SEA a SEE), toxinas exfoliativas (ETA y ETB) y la toxina del síndrome de choque tóxico – 1 (TSST-1) de S. aureus aislados en la población de estudio. Enumerar la presencia única o múltiple de los genes que codifican los superantígenos en S. aureus aislados en la población de estudio. Establecer la susceptibilidad antimicrobiana de los aislamientos de S. aureus en la población de estudio. Determinar asociaciones entre la detección de los genes que codifican los superantígenos y la resistencia antimicrobiana en los aislamientos de S. aureus de la población de estudio.

9
2. MARCO TEÓRICO Y ESTADO DEL ARTE.
2.1 Reseña histórica. S. aureus fue identificado por primera vez en el año 1880, en una ciudad escocesa llamada Aberdeen, por el cirujano Sir Alexander Ogston en el pus que drenaba a un absceso infectado (Lowy, 1998). El nombre binominal de esta especie fue asignado por Friederich Julius Rosenbach en 1884. Loeb en 1903 realizó el descubrimiento de la coagulasa (Hurtado et al., 2002); Loeb mezcló pequeños volúmenes de caldos bacterianos con 3 centímetros cúbicos de plasma de ganso, después lo incubo por seis horas, determinando que “Staphylococcus pyogenes aureus” (conocido en la actualidad como Staphylococcus aureus) era capaz de influir en el proceso de coagulación de la sangre y concluyendo que en diversas condiciones patológicas el Staphylococcus pyogenes aureus juega un papel importante en la causa de la trombosis y la formación de un exudado fibrinoso (Much, 1908). Por ello, se reconoce a S. aureus como “coco Gram positivo coagulasa positivo”. Las infecciones estafilocócicas en 1941 fueron controladas por la penicilina, pero después de poco tiempo, reporto Sprink Ferris que la cepa de S. aureus se había vuelto resistente a la penicilina por la acción de la enzima β-lactamasa que destruyó el antibiótico en 1945 (Hurtado, M. P. et al. 2002; Gil de M, 2000). En 1950 los estafilococos desplazaron los estreptococos como agentes de infección intrahospitalaria y para 1959 el 60% de las cepas ya eran resistentes a penicilina (Gil de M, 2000). Se realizó el primer reporte de la existencia de un Staphylococcus aureus resistente a meticilina por Jevons, cuando era una causa importante de infección nosocomial en Europa (Barrett et al., 1993, Barrett, 1993).
2.2 Taxonomía, genoma y la diversidad genética. El Staphylococcus aureus pertenece al dominio Bacteria, phylum Firmicutes, clase Bacilli, orden Bacillales, familia Staphylococcaceae y género Staphylococcus. El género tiene 51 especies y 24 subespecies1 (Vasconcelos et al., 2010).
2.3 Morfología y estructura antigénica. S. aureus es una bacteria anaerobia facultativa, cuagulasa positivo, se considera un coco Gram positivo, puede estar solo, en parejas, en tétradas o agrupado en forma de racimo (Hennekinne et al., 2010); es un no formador de esporas, inmóvil y se
1Consultado en el URL: http://www.ncbi.nlm.nih.gov. Taxonomy Browser para Staphylococcus aureus. Fecha de
consulta: Diciembre de 2014.

10
diferencia de los estreptococos por ser un catalasa positivo, puede crecer a altas concentraciones salinas en donde la mayoría de ellos crecen en medios con 10% de NaCl y con una actividad acuosa (Aw) mínima de 0,86 (Vasconcelos et al., 2010; Hennekinne et al., 2010). Las colonias del S. aureus presentan un color amarillo dorado característico debido a la producción de carotenoides durante su crecimiento (Rosenbach et al., 1984). Este microorganismo es considerado un patógeno, que coloniza principalmente la piel y membranas mucosas del hombre y de los animales (Murray, 2005). Los factores de virulencia del S. aureus abarca componentes de superficie tales como la cápsula, peptidoglicanos, ácido teicoico, proteína A, proteínas de unión a colágeno, enzimas tales como lipasas, esterasas, ácidos grasos, proteasas, lipasas, lecitinasas, hialuronidasa, coagulasa ligada, coagulasa libre, catalasa, β- lactamasas, termonucleasas (DNasa) y varias toxinas, tales como toxina exfoliativas, leucocidina, enterotoxinas, TSST-1 y hemolisinas α, β, γ, δ. La principal función de estas proteínas sería convertir tejidos del huésped en nutrientes requeridos para el crecimiento bacteriano (Vasconcelos et al., 2010). El componente básico, que confiere forma y estabilidad al microrganismo y representa el 50% de la pared, es el peptidoglicano (Lowy, 1998), lo que les permite resistir la presión osmótica (Capparelli et al., 2011). El andamiaje del peptidoglicano de la pared celular bacteriana está formado por repeticiones de disacaridos unidas por enlaces β-1-4 de N-acetilglucosamina (NAG) y N-acetil-murámico (Meroueh et al., 2006, Pietrocola et al., 2011, Plata et al., 2009). Entre sus propiedades biológicas presenta actividad endotóxica y se destaca por su capacidad para inducir la producción de interleucina-1 por monocitos humanos y desencadenar una reacción local capaz de atraer a leucocitos polimorfonucleares (PMN), activar el complemento y estimular la producción de anticuerpos opsónicos (Mandell et al., 2000). La pared celular está compuesta por polisacáridos, proteínas y otras sustancias importantes. Los ácidos teicoicos, son esenciales para el mantenimiento de la integridad estructural y forma de la célula bacteriana (Atilano et al., 2010), también se encargan de mediar la unión de los estafilococos a las superficies mucosas mediante uniones especificas a la fibronectina, induce anticuerpos y contribuyen a la carga negativa presente en la superficie de la célula que desempeña un papel en la adquisición de iones, también se ha descubierto que es un componente del receptor de fagos de S. aureus (Plata et al., 2009). Los ácidos teicoicos son glicopolimeros ricos en fosfatos que puede estar unidos covalentemente al peptidoglicano o ligados a los lípidos de la membrana citoplasmática (Atilano et al., 2010). Además la pared del S. aureus posee muchas proteínas de superficie: la proteína A (SpA) tiene un peso molecular de 56-KDa, es codificada por el gen spa; SpA presenta cinco dominios (E, D, A, B, C) de enlace a inmunoglobulinas (Ig) en su

11
extremo N-terminal que están anclados covalentemente al peptidoglicano de la pared celular a través del motivo LPXTG en su extremo carboxilo terminal (Widaa et al., 2012). La proteína A (SpA) se une a la porción del fragmento cristalizable (Fc) de la IgG, excepto IgG3, teniendo una importante expresión antifagocítica. Aunque las interacciones entre SpA y las cadenas inmunoglobulina humana han sido reconocidas, sólo recientemente hay una apreciación de la importancia SpA en la patogénesis de la infección por S. aureus (Gomez et al., 2006, Navarre and Schneewind, 1999, Widaa et al., 2012). La proteína A ayuda a inhibir la opsonización, fagocitosis y tiene la capacidad para adherirse al factor de von Willebrand, uniéndose a los sitios de daño al endotelio, y como consecuencia, puede desempeñar un papel en la adherencia y la inducción de enfermedades endovasculares (Plata et al., 2009). Cuando una proteína estafilocócica está anclada a la pared, se clasifica como un componente de la superficie microbiana que reconoce moléculas de la matriz extracelular. S. aureus utiliza estas proteínas para reconocer receptores de la matriz extracelular u otros elementos presentes en los tejidos del hospedero (Navarre and Schneewind, 1999, Plata et al., 2009).
2.4 Factores de virulencia.
2.4.1 Enzimas. Catalasa es una enzima que tiene un grupo hemo; durante el metabolismo celular la enzima cataliza la descomposición del peróxido de hidrogeno en agua y oxígeno. En S. aureus se ha observado una relación entre la actividad y la virulencia, lo que sugiere que la catalasa tiene un papel fundamental en los mecanismos de defensa contra los radicales de oxígeno producidos por los macrófagos (Blaiotta et al., 2010). Esta es útil para diferenciar los estafilococos catalasa positivos, de los estreptococos que son negativos (Brown et al., 2012). Las cepas de S. aureus producen factores de virulencia extracelulares; el gen coa de S. aureus produce coagulasa (Navarre and Schneewind, 1999), una enzima secretada que coagula el plasma oxalatado o citratado, razón por la cual se denomina el estafilococo coagulasa positivo. La coagulasa es una proteína secretada en forma extracelular, se une a la protrombina formando un complejo llamado estafilotrombina, para inducir la coagulación del plasma mediante la conversión del fibrinógeno en fibrina (Widaa et al., 2012). La coagulasa induce la formación de una fibrina alrededor de la célula bacteriana, impidiendo la acción de las células fagocíticas y su destrucción dentro de dichas células (Chevalier et al., 2010).

12
La hialuronidasa es un enzima que degrada el ácido hialurónico de la matriz del tejido conjuntivo y facilita la propagación de la infección. La penicilinasa es una β-lactamasa que inactiva la penicilina mediante hidrólisis de su anillo β-lactámico.
2.4.2 Toxinas. S. aureus cuenta con diferentes toxinas como hemolisinas, exotoxinas, leucocidinas, superantígenos, estos factores de virulencia le permiten evadir la respuesta inmune del huésped. Las hemolisinas conocidas como alfa (α), beta (β), gama (γ) Г y delta (δ), poseen capacidad hemolítica y citolítica dañando las membranas de las células eucariotas del huésped como leucocitos, macrófagos, plaquetas y fibroblastos. La hemolisina alfa (α), es una toxina de un peso molecular de 33 KDa, codificada por el gene hla; formadora de poros especialmente en eritrocitos, hepatocitos, leucocitos, miocitos y plaquetas. Esta toxina ejerce su acción citolítica formando poros en las membranas, alterando de esta forma la osmorregulación, el flujo de cationes y otras moléculas, llegando, incluso, a producir apoptosis. Además estas toxinas inducen la liberación de citocinas y quimiocinas como la IL-6, IL-1β, IL-1α, IL-8, TNF- α, KC, y MIP-2. Estas observaciones se enfatizan en la importancia de la toxina en infecciones por S. aureus (Burnside et al., 2010). La hemolisina Beta (β), es una toxina de 35 KDa, su importancia ha sido demostrada en infecciones del ser humano, en pulmón y córnea. Esta tiene la capacidad de inhibir la actividad ciliar de células epiteliales nasales. La hemolisina delta (δ), es un polipéptido pequeño con 26 aminoácidos, es capaz de causar daños en la membrana de una amplia variedad de células. La δ-toxina es producida por 97% de los aislamientos de S. aureus, esta toxina está codificada en el extremo 5’ de RNAIII, la molécula efectora del sistema regulador Agr, se encarga de lisar eritrocitos y una variedad de células de mamíferos, estructuras subcelulares, tales como orgánulos unidos a la membrana (esferoplastos y protoplastos) (Burnside et al., 2010). El S. aureus tiene la capacidad de producir numerosas exoproteínas que causan una amplia gama de manifestaciones clínicas. La leucocidina de Panton-Valentine (PVL, leucocidina de Panton-Valentine) es una proteína citotóxica que forma poros en la membrana plasmática de los leucocitos y tiene afinidad por los neutrófilos y macrófagos. La leucocidina de Panton-Valentine ocurre en menos del 5% de las cepas de S. aureus. La leucocidina es citotóxica para los monocitos, macrófagos y leucocitos polimorfonucleares del humano. La leucocidina es una proteína que forma poros en la membrana plasmática de los leucocitos, lo cual provoca un aumento en la permeabilidad y eventualmente produce la lisis de la célula. La lisis de los leucocitos produce la liberación de mediadores de la inflamación, con una

13
consecuente respuesta inflamatoria grave células responsables de la defensa contra las bacterias invasoras (Lipinska et al., 2011). PVL ha ganado reconocimiento por la neumonía necrotizante severa y por su virulencia asociada cepas resistentes a la meticilina (SARM, S. aureus resistente a meticilina), que se han extendido rápidamente a partes de América del Norte, Europa y Australia (Day, 2012). La habilidad del S. aureus para provocar enfermedades humanas depende de la producción en la superficie celular de adhesinas, factores antifagocíticos y exotoxinas. Entre los factores de virulencia secretados están los superantígenos (Schlievert et al., 2008). Es importante destacar que los factores de virulencia secretados también incluyen una gran grupo de exotoxinas tales como superantígenos, incluyendo enterotoxinas estafilocócicas (SEs): SEA, SEB, SEC1,2,3, SED, SEE, SEG, SEH, SEI, SElR, SElS, SElT y las enterotoxinas (staphylococcal enterotoxin like) (SELs): SElJ, SElK, SElL, SElM, SElN, SElO, SElP, SElQ, SElU2, SElV, SELW y las del síndrome de choque tóxico (TSST -1), y las toxinas exfoliativas A, B, C Y D (Schlievert et al., 2010, Ortega et al., 2010)
Genoma y diversidad genética.
El primer genoma secuenciado de S. aureus fue publicado en 2001 (Kuroda et al., 2001). En el 2008 se habían completado 12 secuencias genómica que están disponibles para el dominio público (Harris et al., 2010) y en el 2015, 36 grupos han reportado 72 genomas completos, 4532 genomas ensamblados2. S. aureus posee un cromosoma circular, de un tamaño promedio de genoma entre 2,73 - 3,1 Mpb, con contenido de GC 33 %, un número de genes de 2,545 - 2,998 y numero de proteínas de 2,433- 2,926. Las secuencias genómicas del S. aureus han mostrado que el 75% del genoma base es altamente conservado en todas las cepas y está compuesto por genes esenciales para el crecimiento y supervivencia (Schlievert et al., 2010, Lindsay and Holden, 2004). La comparación de las cepas secuenciadas de S. aureus reveló que el orden de los genes es conservados y hay similitud en la secuencias de aminoácidos en un 98 – 100% (Lindsay and Holden, 2004, Lindsay, 2010). La mayoría de los genes que comprenden el genoma base, son asociados al metabolismo central y otras funciones de mantenimiento. El genoma accesorio es el 25%, comprende los elementos genéticos móviles, que codifican proteínas relacionadas con la virulencia y la resistencia (Lindsay and Holden, 2006). Las cepas de S. aureus han evolucionado debido a la integración de elementos genéticos móviles que codifica la resistencia a la meticilina y factores de virulencia. Algunos genes o regiones genéticas muestran una variación considerable entre los
2Consultado en el URL: http://www.ncbi.nlm.nih.gov/genome/genomes/154. Genome Browser para
Staphylococcus aureus. Fecha de consulta: Agosto de 20152012

14
genomas o pueden estar ausentes en algunas cepas y muchas de estas variaciones ahora se saben cual es su linaje específico. Por último, los elementos genéticos móviles, o que alguna vez fueron móviles, están presentes o ausentes en el genoma y muestran una frecuente evidencia de transferencia genética horizontal y recombinación genética (Schlievert et al., 2010).
2.4.3 Los superantígenos. El término de superantígenos (SAgs) fue acuñado por White en 1989 (White et al., 1989) para designar a un grupo de proteínas virales y toxinas bacterianas proteicas. Los superantígenos (SAgs) son pequeñas proteínas secretadas de un tamaño de 20 a 29 KDa (Hennekinne et al., 2010), que comparten características similares bioquímicas, estructurales, funcionales, genéticas e inmunobiológicas, tienen la capacidad para estimular la proliferación y liberación de citocinas de las células T policlonales, de los subtipos CD4+, CD8+, algunos gamma delta (γ- δ), y macrófagos, para producir grandes cantidades de citocinas, que es como finalmente se va a desencadenar un gran proceso inflamatorio (Torres et al., 2001, Plata et al., 2009, Schlievert et al., 2010).
2.4.3.1 Clasificación estructural de los superantígenos. Los superantígenos bacterianos son producidos predominantemente por Staphylococcus aureus y Streptococcus pyogenes. Se cree que los superantígenos comparten una estructura terciaria conservada. No obstante, se han propuesto cinco grupos evolutivos diferentes (I al V) para estas toxinas. Se han clasificado de acuerdo con sus relaciones filogenéticas, y las diferencias importantes que exhiben los representantes característicos de cada grupo que interacciona con los receptores del hospedero (Sundberg et al., 2007, Gunther et al., 2007). Su estructura tridimensional consta de 2 dominios conservados unidos por una alfa hélice: un dominio pequeño N-terminal beta barril y un dominio mayor C-terminal con un motivo de agarre beta (Mitchell et al., 2000, Xu and McCormick, 2012, Wilson et al., 2011). Grupo I: Este grupo es evolutivamente distinto, conformado por la toxina del síndrome de shock tóxico 1 (TSST-1) porque no produce emesis (Schlievert et al., 2000) y es único en su modo de unirse al CMH II, por lo tanto no comparte este grupo con ningún otro superantígeno. La toxina del síndrome de shock tóxico 1 (TSST-1) se une al CMH II, a través de dominio de unión N-terminal, de baja afinidad, que es dependiente del péptido (Kim et al., 1994, Gunther et al., 2007, Wen
et al., 1997) y liga el RCT V por medio de dos regiones independientes, tanto en la CDR2, como en la región marco FR3, juntas exhiben una unión cooperativa positiva (Sundberg et al., 2007, Gunther et al., 2007).

15
Grupo II: Contiene la enterotoxina B y la C. También se ligan la cadena del CMH II a través de la unión N-terminal del dominio baja afinidad; sin embargo, esta unión es independiente del péptido (Gunther et al., 2007, Jardetzky et al., 1994). Los SAgs
sujetan el RCT V por mecanismos dependientes de la conformación, que son
independientes de cadenas laterales aminoacídicas específicas de la V (Li et al., 1998, Sundberg et al., 2002, Gunther et al., 2007, Fields et al., 1996). Grupo III: Posee superantígenos estafilocóccicos como la enterotoxina A. Estas toxinas son capaces de ligar cruzadamente moléculas de CMH II (Abrahmsen et al., 1995, Gunther et al., 2007, Hudson et al., 1995) por un dominio de unión de baja afinidad similar al del grupo II, como también por un dominio de alta afinidad, con una interfase de unión al CMH dependiente de zinc, localizada en el dominio de
agarre del SAgss (Gunther et al., 2007, Petersson et al., 2001). Existe muy poca
información sobre como los SAgs del grupo III ligan la V del RCT. Este grupo es asociado con la intoxicación alimentaria estafilocócica (Brouillard et al., 2007, Dinges et al., 2000, Xu and McCormick, 2012). Grupo IV: Sus miembros son SAgs estreptocócicos. Tienen un dominio de alta afinidad, como el del grupo III (Brouillard et al., 2007, Li et al., 2001). La estructura
del superantígeno en asocio con el dominio variable del RCT, reveló que estas
toxinas ligan las asas hipervariables del V, incluyendo CDR1 hasta CDR3 (Gunther et al., 2007). Grupo V: Se encuentran agrupados la exotoxina pirogénica estreptocócica y las enterotoxinas K (SEK), I (SEI), L (SEL), M (SEM), Q (SEQ). La estructura cristalina de SEI en complejo con HLA-DR1, reveló las similitudes de este grupo en el complejo péptido-CMH II (pCMH), con el grupo IV. La diferencia significativa con
otros grupos radica en una extensión en forma de horquilla entre la tercera -hélice
y la octava hoja-, llamada asa 3-8. Esta extensión de 15 residuos de aminoácidos no se encuentra en otros grupos de SAgs, y no está involucrada en las interacciones péptido-CMH II (Brouillard et al., 2007, Gunther et al., 2007, Sundberg et al., 2007).
2.4.3.2 Clases de Superantígenos de S. aureus. Los SAgs de S. aureus comprenden las toxinas exfoliativas (ET), las toxinas pirogénicas (PTSAgs), enterotoxinas (SE) y la toxina del síndrome de choque tóxico 1 (TSST-1). Las toxinas pirogénicas se caracterizan por producir fiebre, hipertensión, inmunosupresión y aumento de la sensibilidad al shock endotóxico.
2.4.3.2.1 Toxina del síndrome de choque tóxico.

16
El síndrome de choque tóxico (TSS, Toxic shock syndrome) es una exotoxina secretada por S. aureus, se identificó como un agente principal causante de los TSS hace unos 25 años (Bergdoll et al., 1981, Schlievert et al., 1981, Moza et al., 2007). La toxina es sintetizada como un precursor de 234 residuos de aminoácidos con los primeros 40 aminoácidos como secuencia señal, la cual es clivada para generar la proteína madura de 22 KDa. (Dinges et al., 2000, Blomster-Hautamaa et al., 1986). TSST-1 está codificada por el gene tstH presente en el cromosoma bacteriano dentro de las islas de patogenicidad SaPI-1, SaPI-2 y SaPIbov (Kreiswirth, 1989, Lindsay et al., 1998, Ruzin et al., 2001). El síndrome de choque tóxico TSS-1 produce una enfermedad aguda y potencialmente mortal, caracterizada por comenzar con una fiebre alta, erupción eritematosa difusa con posterior descamación de la piel una o dos semanas después de la aparición, hipotensión, y afecta muchos órganos sistémicos (Todd et al., 1978, Ortega et al., 2010, Davis et al., 1980). La enfermedad fue detectada inicialmente en la década de 1980, con un número significativo de mujeres jóvenes en ciclo menstrual y asociada al uso de tampones, de alta absorbencia. El TSST-1 puede ser responsable del 100% de los casos TSST- menstruales, probablemente relacionado con la capacidad particular del superantígeno para cruzar las barreras mucosas (Todd et al., 1978, Davis et al., 1980, Shands et al., 1980, Li et al., 2011).
2.4.3.2.2 Las toxinas exfoliativas. Algunas cepas de S. aureus producen toxinas exfoliativas (ET) o también conocidas como toxinas epidermolíticas y es causante de la enfermedad epidermolítica. Las características clínicas de estos trastornos de la piel desde el impétigo ampolloso hasta el síndrome de piel escaldada (SSSS, staphylococcal scalded skin syndrome) que ocurren especialmente en la piel de neonatos o niños pequeños (Ladhani et al., 1999), se manifiesta con fiebre, malestar general, letargo y falta de apetito, estos síntomas son seguidos por una erupción eritematosa y la formación de grandes ampollas (Bukowski et al., 2010). Las ampollas se rompen con la acción mecánica, dejando a las partes afectadas del cuerpo sin capa protectora de la epidermis. En Europa, Estados Unidos y África, la toxina más frecuente es ETA, que se expresa en más del 80% de las cepas (Bukowski et al., 2010). Las toxinas exfoliativas tienen un peso molecular aproximado de 30 KDa. Se han clasificado en tres serotipos. Las toxinas exfoliativas obtenidas de cepas de Staphylococcus aureus aislados de humanos, han mostrado que las más frecuentes son las toxinas ETA y ETB, y con menor frecuencia la toxina ETD, existe una cuarta toxina exfoliativa C (ETC) que se aisló de una cepa de S. aureus obtenida de un caballo, no se ha asociado con enfermedades humanas y todavía no se ha caracterizado genéticamente (Sato et al., 1994). Los genes que codifican las toxinas exfoliativas se encuentran localizados en diferentes regiones del cromosoma o en

17
elementos genéticos móviles; ETA codificada por el gene eta, que está en un fago (43.5 kb) (Yamaguchi et al., 2002, Bukowski et al., 2010, Endo et al., 2003), mientras que ETB es codificada por el gene etb, se encuentra en un plásmido pETB (38.2 kb) (Yamaguchi et al., 2001, Bukowski et al., 2010) y el gen etd (ETD) se ubica en una isla de patogenicidad (9 Kb) (Yamaguchi et al., 2002, Bukowski et al., 2010), su actividad proteolítica parece ser la responsable de la exfoliación. Las toxinas ETA, ETB y ETD, son serín proteasas, ETD tienen la capacidad de actuar también como glutamil endopeptidasa (ETD); estas clivan Dsg-1 desmogleína 1 (Dsg1, proteína de adhesión transmembranal de los desmosomas), separando las células granulosas de la epidermis superficial, lo cual hace que las células pierdan el contacto entre sí y se desarrolle un plano de clivaje por debajo del estrato córneo (Yamaguchi et al., 2002, Bukowski et al., 2010).
2.4.3.2.3 Enterotoxinas (SE, staphylococcal enterotoxin). La intoxicación alimentaria estafilocócica (SFP, staphylococcal food poisoning) produce gastroenteritis por ingestión de una o más enterotoxinas estafilocócicas (SE) que se encuentran en alimentos que ha sido contaminada con cepas enterotoxigénicas de S. aureus. Las enterotoxinas (SEs) son pirogénicas y comparten propiedades importantes como la capacidad de inducir emesis, gastroenteritis y superantigenicidad (Pinchuk et al., 2010). Estas son potentes exotoxinas gastrointestinales, sintetizados por S. aureus a lo largo de la fase logarítmica de crecimiento o durante la transición de la fase exponencial a la estacionaria (Betley et al., 1992, Argudin et al., 2010, Otero et al., 1990). SE son activas a dosis bajas (Larkin et al., 2009) y resistentes a tratamientos térmicos, ácidos, pH bajos, que fácilmente destruirían cualquier bacteria y enzimas proteolíticas (como pepsina, tripsina, quimotripsina, renina y papaína); por lo tanto, las enterotoxinas no son totalmente desnaturalizados por cocción suave de alimentos contaminados (Pinchuk et al., 2010) y puede que se mantenga su actividad en el tracto digestivo después de su ingestión (Argudin et al., 2010, Schantz et al., 1965). Las enterotoxinas estafilocócicas (SE) pertenecen a una gran familia de exotoxinas pirogénicas (PT) de estafilococos y estreptococos, compartiendo homología en sus secuencias (Ortega et al., 2010). Las SEs han sido clasificadas según criterios serológicos en 23 miembros de SEs/SEls, con exclusión de variantes moleculares (Ver tabla 1).
Inicialmente se caracterizaron cinco tipos de SEs denominadas enterotoxinas clásicas SEA, SEB, SEC (con la SEC1, SEC2 y SEC3, variantes de SEC ovina y SEC bovina), SED y SEE (Ben Nejma et al., 2006, Casman, 1960), que fueron descubiertos en los estudios de las cepas de S. aureus implicadas en intoxicaciones alimentarias (Argudin et al., 2010). Posteriormente se detectaron otras nuevas enterotoxinas SEG, SEH, SEI, SER, SES, SET (Ono et al., 2008, Munson et al., 1998), entre las cuales se han reportado variantes para SEG y SEI. El Comité

18
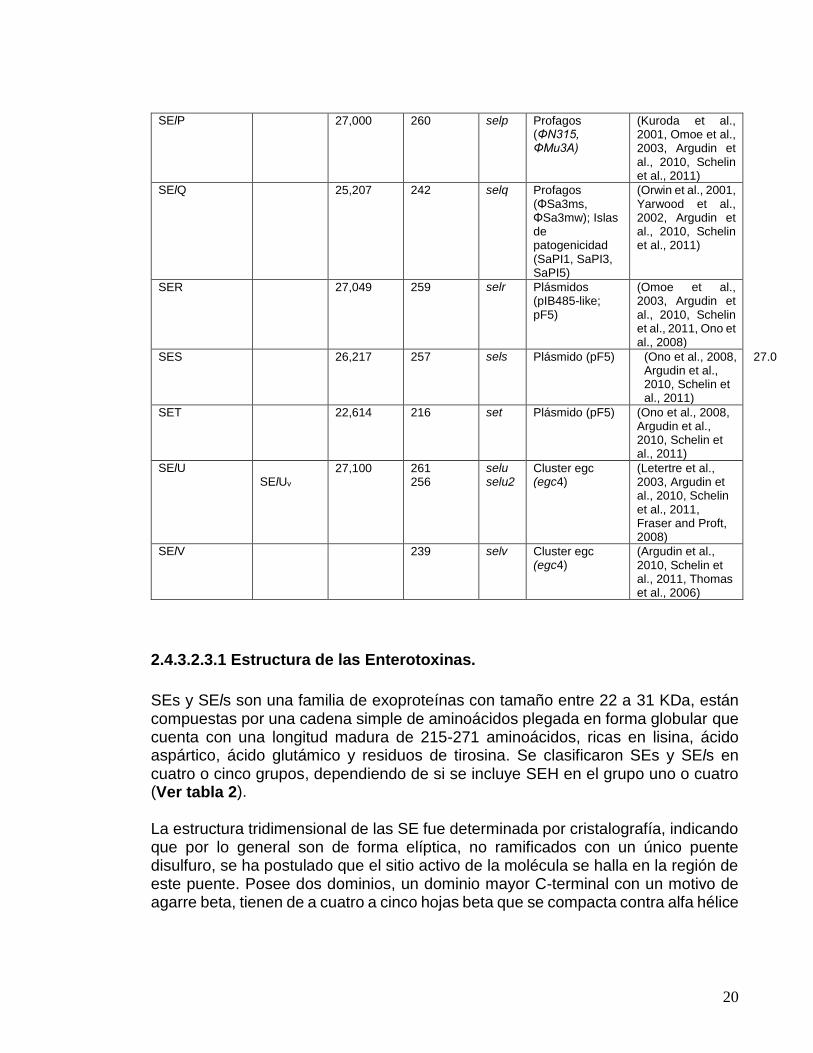
Internacional de nomenclatura para superantígenos estafilocóccicos plantea que solo los superantígenos estafilocócicos son capaces de inducir emesis después de su administración vía oral, por un modelo experimental con primates y la otras tóxinas similares que no presentan dicha actividad o que todavía no han sido aprobadas debe ser designadas como enterotoxina tipo “like” (Staphylococcal Enterotoxin-like; SEl) (Omoe et al, 2005), como SElJ, SElK, SElL, SElM, SElN, SElO, SElP, SElQ, SElU, SElV y SElW (Jarraud et al., 2001, Thomas et al., 2006, Zhang et al., 1998, Ono et al., 2008) y la toxina del síndrome choque tóxico (TSST-1), inicialmente designado como SEF, carece de actividad emética (Bergdoll et al., 1981, Reiser et al., 1983). Tabla 1. Características generales y localización genómica de los genes de enterotoxinas (SEs – SEls) del S. aureus.
Enterotoxinas Variantes Peso Molecular (Da)
Longitud (aa)
Gene Ubicación Referencias
SEA SEA1 SEA2
27.100 233 sea Profagos (ΦSa3m, ΦSa3mw, Φ252B, ΦNM3, ΦMu50a)
(Betley and Mekalanos, 1985, Gomez-Lucia et al., 1992, Le Loir et al., 2003, Wallin-Carlquist et al., 2010, Schelin et al., 2011).
SEB 28.336 239 seb Plásmido (pZA10); Isla de patogenicidad (SaPI3)
(Novick et al., 2001, Le Loir et al., 2003, Argudin et al., 2010, Schelin et al., 2011, Hennekinne et al., 2010)
SEC SEC1 SEC2 SEC3 SEC Bovina, SEC Ovejas
27.531 27.531 27.531 27,563 27,618 27,517
239 239 239 239 271
sec1 sec2 sec3 secbov secsheep
Islas de patogenicidad (SaPIn1, SaPIm1, SaPImw2, SaPIbov1)
(Fitzgerald et al., 2001a, Kuroda et al., 2001, Yarwood et al., 2002, Novick, 2003b, Argudin et al., 2010, Schelin et al., 2011, Marr et al., 1993)
SED
26,360 228 sed
Plásmido (pIB485-like)
(Le Loir et al., 2003, Argudin et al., 2010, Hennekinne et al., 2010, Schelin et al., 2011, Bayles and Iandolo, 1989)
SEE 26.4
230 see
Profago (ΦSa) (Le Loir et al., 2003, Argudin et al., 2010, Hennekinne et al., 2010, Schelin et al., 2011, Couch et al., 1988)

19
SElG SElG2 SElGV
27,043 26,985
233 242 233
seg Cluster egc egc1 (vSaβ I); egc2 (vSaβ III); egc3; egc4
(Jarraud et al., 2001, Baba et al., 2002, Le Loir et al., 2003, Sumby and Waldor, 2003, Argudin et al., 2010, Schelin et al., 2011, Abe et al., 2000)
SEH 25,210 218 seh SCCmec (Le Loir et al., 2003, Schelin et al., 2011)
SEI SEIv
24,298
218 242
sei Cluster egc egc1 (vSaβ I); egc2 (vSaβ III) ); egc3
(Jarraud et al., 2001, Argudin et al., 2010, Schelin et al., 2011, Letertre et al., 2003)
SElJ
28,565 245 selj Plásmido (pIB485-like; pF5)
(Omoe et al., 2003, Argudin et al., 2010, Schelin et al., 2011, Zhang et al., 1998)
SElK
SElK2
25,539 219 242
selk Islas de patogenicidad (SaPI1, SaPI3, SaPIbov1, SaPI5); Profago (ΦSa3ms, ΦSa3mw)
(Baba et al., 2002, Yarwood et al., 2002, Sumby and Waldor, 2003, Argudin et al., 2010, Schelin et al., 2011)
SElL
24,593 215 sell Islas de patogenicidad (SaPIn1, SaPIm1, SaPImw2, SaPIbov1, SaPI3)
(Fitzgerald et al., 2001a, Yarwood et al., 2002, Le Loir et al., 2003, Argudin et al., 2010, Schelin et al., 2011)
SElM
24,842 217 selm
Cluster egc egc1 (vSaβ I); egc2 (vSaβ III)
(Jarraud et al., 2001, Le Loir et al., 2003, Argudin et al., 2010, Schelin et al., 2011)
SElN SElNv
26,067 227 seln
Cluster egc egc1 (vSaβ I); egc2 (vSaβ III); egc3; egc4
(Le Loir et al., 2003, Letertre et al., 2003, Argudin et al., 2010, Schelin et al., 2011, Jarraud et al., 2001)
SElO
26,777 232 selo
Cluster egc egc1 (vSaβ I); egc2 (vSaβ III); egc3; egc4
(Jarraud et al., 2001, Le Loir et al., 2003, Argudin et al., 2010, Schelin et al., 2011)
20
SElP
27,000 260 selp Profagos (ΦN315, ΦMu3A)
(Kuroda et al., 2001, Omoe et al., 2003, Argudin et al., 2010, Schelin et al., 2011)
SElQ
25,207 242 selq
Profagos (ΦSa3ms, ΦSa3mw); Islas de patogenicidad (SaPI1, SaPI3, SaPI5)
(Orwin et al., 2001, Yarwood et al., 2002, Argudin et al., 2010, Schelin et al., 2011)
SER 27,049 259 selr
Plásmidos (pIB485-like; pF5)
(Omoe et al., 2003, Argudin et al., 2010, Schelin et al., 2011, Ono et al., 2008)
SES 26,217 257 sels Plásmido (pF5)
(Ono et al., 2008, Argudin et al., 2010, Schelin et al., 2011)
27.0 yes no ser pIB485-like; pF5
SET 22,614 216 set Plásmido (pF5) (Ono et al., 2008, Argudin et al., 2010, Schelin et al., 2011)
SElU
SElUv
27,100 261 256
selu selu2
Cluster egc (egc4)
(Letertre et al., 2003, Argudin et al., 2010, Schelin et al., 2011, Fraser and Proft, 2008)
SElV 239 selv Cluster egc (egc4)
(Argudin et al., 2010, Schelin et al., 2011, Thomas et al., 2006)
2.4.3.2.3.1 Estructura de las Enterotoxinas.
SEs y SEls son una familia de exoproteínas con tamaño entre 22 a 31 KDa, están compuestas por una cadena simple de aminoácidos plegada en forma globular que cuenta con una longitud madura de 215-271 aminoácidos, ricas en lisina, ácido aspártico, ácido glutámico y residuos de tirosina. Se clasificaron SEs y SEls en cuatro o cinco grupos, dependiendo de si se incluye SEH en el grupo uno o cuatro (Ver tabla 2).
La estructura tridimensional de las SE fue determinada por cristalografía, indicando que por lo general son de forma elíptica, no ramificados con un único puente disulfuro, se ha postulado que el sitio activo de la molécula se halla en la región de este puente. Posee dos dominios, un dominio mayor C-terminal con un motivo de agarre beta, tienen de a cuatro a cinco hojas beta que se compacta contra alfa hélice

21
muy conservada (Mitchell et al., 2000) y por ultimo un dominio pequeño N-terminal beta barril con una topología de llave griega (Argudin et al., 2010). Tabla 2. Clasificación de las enterotoxinas de acuerdo a sus secuencias de aminoácidos. SE codificadas por el Cluster egc, están en paréntesis. SEH se coloca en el grupo 1 o 5, según el autor.
Grupos Enterotoxinas Características Referencias
Grupo 1 SEA, SED, SEE, SElJ, (SElN, SElO), SElP, SES
Comprenden 53 a 81% de homología en la secuencia de aminoácidos
(Marrack and Kappler, 1990, Thomas et al., 2006, Larkin et al., 2009)
Grupo 2 SEB, SEC, (SEG), SER, (SElU, SElU2)
Cuentan con el 66 a 98% homología en la secuencia de aminoácidos
(Marrack and Kappler, 1990, Larkin et al., 2009, Argudin et al., 2010)
Grupo 3 (SEI), SElK, SElL, (SElM), SElQ, (SElV)
(Larkin et al., 2009, Argudin et al., 2010, Omoe et al., 2005b)
Grupo 4 SET
(Argudin et al., 2010)
Grupo 5 SEH
(Larkin et al., 2009, Argudin et al., 2010, Omoe et al., 2005)
2.4.3.2.3.2 Mecanismo de acción. Los superantígenos son toxinas de origen microbiana o virales, no son procesados por las células presentadoras de antígenos, sino que se unen a moléculas del complejo mayor de histocompatibilidad de clase II (MHC-II, major histocompatibility complex class II) de las células presentadoras de antígeno (APC, Antigen-presenting cells), fuera de la hendidura clásica de unión del antígeno produciendo una estimulación masiva (proliferación policlonal) de linfocitos (T y B) y la liberación de citoquinas, contribuyendo a la evasión de las defensas del hospedador y a la persistencia bacteriana (Ortega et al., 2010). El receptor de linfocitos T (TCR, T cell receptor) es el receptor para el antígeno de la célula T, su función es reconocer fragmentos peptídicos de proteínas antigénicas, que forman un complejo con moléculas del complejo mayor de histocompatibilidad, éste último ubicado en las células presentadoras de antígeno (Ortega et al., 2010).

22
El TCR es un heterodímero que está compuesto por dos cadenas glucoproteicas
paralelas, y (TCR) ó y (TCR), que se proyectan desde la membrana del
linfocito T. Cada una de las cadenas tiene una región interna constante (C y C) y
una fracción externa variable (V y V), donde la porción de unión al antígeno del TCR está formada por los dominios variables (Hong et al., 1996, Li et al., 1999). El extremo del superantígeno interacciona directamente con residuos conservados del complejo mayor de histocompatibilidad II (CMH II) (Schubert. M. S, 2009)
mientras que el otro extremo del superantígeno reacciona con la región V de los receptores para el antígeno de las células T (Schlievert et al., 2008, Wilson et al., 2011). Cuando la célula presentadora de antígenos expone a través del CMH, un antígeno exógeno convencional al TCR, éste reconoce la configuración de un antígeno específico y se activa un número escaso de células T. El superantígeno debe unirse al MHC de clase II al concentrarse sobre la superficie de la células presentadoras de antígeno; haciendo uso de las moléculas de adhesión y accesorias que forman la sinapsis inmunológica (Ortega et al., 2010), el
superantígeno se ajusta fuera de la hendidura de unión al péptido, al dominio V del TCR, regiones determinantes de la complementariedad 1 y 2 (CDR1 y CDR2), mientras que el antígeno convencional se uniría a la región regiones determinantes de la complementariedad (CDR3). Después la formación de puentes de unión entre
los TCR - V - específicos de las células T y el CMH II sobre las células B, inducen la activación policlonal de las células B que lleva a la formación de autoanticuerpos. Por último, la interacción entre las células presentadoras de antígeno y los linfocitos T, produciendo activación de más de 20% de estas (Ortega et al., 2010), las células dan lugar a la liberación de citoquinas, y la masiva producción de citocinas, pro-
inflamatorias como Factor de Necrosis Tumoral (TNF), interleucina 2 (IL-2) e
interferón gama (IFN) los cuales producen fiebre y choque (Muller-Alouf et al., 2001). Los superantígenos evitan la generación de una respuesta inmunológica eficaz, ocasionando evasión de las defensas del hospedador, haciendo al huésped más susceptible de padecer una infección microbiana (Ortega et al., 2010).
2.4.4 Regulación de factores de virulencia en S. aureus. S. aureus produce muchas infecciones debido a la expresión de varios factores de virulencia, que están asociadas a superficie o a secreción. Muchas de estas proteínas accesorias están implicadas en la colonización de los tejidos, lisis de las membranas de la célula, propagación bacteriana dentro de los tejidos y supervivencia a la acción microbicida de los fagocitos (Boyce JM., 1997, Lowy F., 1997., Ray et al., 2009).

23
El Quorum Sensing “QS” es el nombre que se le da al mecanismo de "Comunicación" entre las bacterias, por medio del cual una bacteria puede "percibir" la densidad de población en el medio. Este mecanismo es importante en S. aureus ya que algunas proteínas accesorias (tales como factores virulencia) sólo se expresan en determinadas fases de crecimiento (McCulloch., 2006). La producción de factores de virulencia es controlada en respuesta a la densidad celular (Quórum Sensing), disponibilidad de energía, señales medio-ambientales y a los superantígenos; así estos factores son producidos cuando son requeridos (Novick, 2003a). Se han descrito grandes familias de reguladores globales en S. aureus: los sistemas de dos componentes, el más conocido es el sistema Agr, que codifica el gen arg regulador accesorio (Peng et al., 1988), y la familia de proteínas homólogas a SarA (Arvidson and Tegmark, 2001, Novick, 2003a, Cheung et al., 2004), sar es el gen accesorio regulador estafilocócico, dividido en sarA, sarS, sarT y sarR (Cheung et al., 1992, McCulloch., 2006, Vasconcelos et al., 2010), rot (represor de toxinas) (McNamara et al., 2000), el cual puede afectar directamente la producción de enterotoxinas estafilocócicas. También cuenta con tres sistemas reguladores saeRS, σB (sigma-B), arlRS (McCulloch., 2006) y srrAB (Throup et al., 2001, Yarwood et al., 2001). Las proteínas accesorias se expresan de una manera coordinada dependiendo de la fase de crecimiento y son controlados por varios sistemas de regulación, tales como sistemas de regulación de dos componentes (AgrAC, ArlRS, SaeRS, SrrAB, LytRS, VraRS, YycFG) y los sistemas reguladores de la transcripción (por ejemplo, 10 familias, sigB) (Novick, 2003b, Bronner et al., 2004, Cheung et al., 2004, Cheung et al., 2008, Ray et al., 2009).
2.4.4.1 El sistema regulador Agr y otros sistemas reguladores. Los sistemas reguladores de dos componentes permiten a la bacteria detectar y responder a condiciones medioambientales incluso cuando los estímulos no penetran en el citoplasma. Estos sistemas constan de un sensor de membrana (histidín quinasa) y de un regulador de respuesta citoplásmico. En respuesta a una señal apropiada, se produce la autofosforilación de un residuo de histidina en el dominio citoplásmico del sensor y a continuación, el grupo fosfato es transferido a un residuo de aspartato en el regulador de respuesta, que al activarse, estimula o reprime la transcripción de sus genes diana, que elaboran una respuesta adecuada a la señal recibida (Yarwood et al., 2001). El locus agr (agr, accesory gen regulator) codifica un gen regulador accesorio (lo que indica que no es esencial para supervivencia), encargado de controlar la expresión de la mayoría de las exoproteínas, actuando como un regulador global de

24
virulencia, codificando el sistema de quórum sensing (Novick and Geisinger, 2008). El locus agr ha sido secuenciado y clonado, tiene dos operones diferentes, con al menos cinco genes: agrA, agrB, agrC, agrD y el gen para la hemolisina (hld) y que está controlado por dos promotores divergentes, P2 y P3 (Peng et al., 1988). El operón cuyo promotor es P2 contiene cuatro genes: agrA, agrB, agrC, agrD y el operón con promotor es P3 contiene un solo gen hld (McCulloch., 2006). El sistema agr consta de dos unidades transcripcionales divergentes, ARNII (agrA, agrB, agrC, agrD) y ARNIII (hld), que se transcriben a partir de promotores P2 y P3, respectivamente. ARNII codifica el sistema de quórum sensing, incluyendo la transcripción del operón agr (Reynolds and Wigneshweraraj, 2011). EL agrA y agrC son un sistema de transducción sensorial de dos componentes, AgrC el sensor y AgrA el regulador de respuesta; posee un casete de detección densidad (agrD y B) (Chevalier et al., 2010), donde AgrB es una endopeptidasa integral de membrana implicada en el procesamiento y la secreción de AgrD, dando lugar al péptido señal autoinducible (AIP), un receptor histidín quinasa (AgrC), y el regulador de respuesta de unión al ADN (AgrA). ARNIII es un ARN de 510 nucleótidos, es una molécula reguladora de un gran número de genes, en su mayoría a nivel de traducción (Peng et al., 1988, Novick et al., 1993, Novick, 2003a, Yarwood and Schlievert, 2003, Ray et al., 2009). La molécula efectora de este sistema regulador es RNAIII, que actúa a nivel transcripcional, inhibe la expresión de proteínas superficiales (proteína A, proteínas de unión a fibronectina, etc.) y actúa en primer lugar, con el fin de iniciar la transcripción de los factores de virulencia. La síntesis ARNIII es inducida cuando la concentración de medio específico genera la auto-inducción polipéptido (AIP), alcanzando ciertos niveles que son generalmente detectados en la transición de la fase de crecimiento exponencial a la fase estacionaria (Vasconcelos et al., 2010). La autoactivación agr, por medio del péptido AgrD precursor de AIP (AIP, Autoinducing Peptide), está compuesto de tres partes, alfa hélice anfipática N-terminal, una región media que contiene AIP y un extremo C-terminal; es procesado el extremo N-terminal por SpsB y el C-terminal por AgrB, secretada en forma de un anillo de tiolactona (AIP), que actúa en el dominio receptor transmembrana de AgrC, induciendo la fosforilación del dominio citoplásmico HPK. El fosfato se transfiere a AgrA, que activa la transcripción de los dos promotores de agr. El promotor P2 gobierna el circuito de autoactivación, y el promotor P3 dirige la transcripción de ARNIII, que es el efector regulador del sistema. La moléculas inhibitoria no cognados AIP (rojo) compiten con el cognados AIP (azul) para la unión al receptor (Novick and Geisinger, 2008) (Figura 1).

25
Figura 1. Funcionamiento del locus agr (Novick and Geisinger, 2008). SarA es un regulador global que coordina la síntesis de la pared celular y proteínas extracelulares de virulencia durante la fase exponencial. El locus sarA se transcribe desde tres promotores, sarP1, sarP2 y sarP3, que se activan en momentos distintos; los dos primeros se activan al comienzo de la fase exponencial de crecimiento y el último durante la fase postexponencial por acción del factor σB en respuesta a estímulos medioambientales (Arvidson and Tegmark, 2001). SarA es requerido para la activación de saeRS y arlRS, regula positivamente la transcripción de genes de exoproteínas desde el regulador global agr uniéndose a los promotores P2 y P3 (Novick, 2003a); es otro miembro de la familia de homólogos Sar, afecta la transcripción agr y su actividad de unión al ADN por P2, es dependiente de las condiciones redox y de pH, se ha demostrado controla positivamente seb y sec (Bronner et al., 2004, Fujimoto et al., 2009, Heinrichs et al., 1996). Se ha descrito que SarA promueve la síntesis de las proteínas de unión a fibrinógeno y fibronectina, α-, β- y δ-toxinas y biofilm, mientras que reprime las proteínas de unión a colágeno, proteína A y proteasas uniéndose directamente a los promotores de los genes diana, o indirectamente, por su efecto sobre otros reguladores (Heinrichs et al., 1996, Novick, 2003a, Cheung et al., 2004).
2.5 Epidemiología de la infección. Las intoxicaciones alimentarias son un riesgo permanente para salud pública, se conocen más de 200 enfermedades que se transmiten a través del consumo de alimentos. Cada año, Staphylococcus aureus, Salmonella, Clostridium perfringens, Campylobacter, Listeria monocytogenes, Vibrio parahaemolyticus, Bacillus cereus y Escherichia coli enteropatógena causan más de 90% de todas las intoxicaciones alimentarias transmitidas por alimentos (Moises and Schäferling, 2009); es posible que las intoxicaciones por S. aureus sean la causa principal de enfermedades alimentarias en todo el mundo, aunque el registro de ésta es aún más incompleto.

26
Diferentes alimentos puede ser un medio de crecimiento para S. aureus, y son causantes de intoxicación alimentaria estafilocócica, incluyendo productos lácteos, cárnicos, vegetales y avícolas. Los orígenes intoxicación alimentaria estafilocócica son muy diferentes entre los países y esto puede ser debido a diferencias en el consumo y los hábitos alimenticios de cada uno de ellos (Ortega et al., 2010). Se ha estimado que solo del 1 al 5% de todos los casos de intoxicación alimentaria estafilocócica (IAE o SFP, Staphylococcal food poisoning) que ocurren en EE.UU. son reportados al Departamento de Salud Pública; a su vez, se estima que la IAE corresponde al 14% del total de las ETA (ETA, enfermedades transmitidas por alimentos) siendo la tercera causa más común de tipo bacteriano en ese país. Los Centros para el Control y Prevención de Enfermedades (CDC, Centers for Disease Control and Prevention of USA) se encarga de recopilar los datos sobre brotes de enfermedades transmitidas por alimentos en EE.UU, indicando que el número estimado de enfermedades relacionadas con los alimentos causan aproximadamente 80 millones de intoxicaciones, 325.000 hospitalizaciones y 5000 muertes por patógenos conocidos como el Staphylococcus aureus (Mead et al., 1999, Kadariya et al., 2014). S. aureus causa un estimado 241.000 casos de intoxicaciones por alimentos contaminados en los EE.UU aureus (Mead et al., 1999, Kadariya et al., 2014). En Europa, la Autoridad Europea de Seguridad Alimentaria (EFSA) reporto que en el 2008, las toxinas bacterianas estuvieron involucradas en 525 de 5.332 brotes de intoxicaciones alimentarias (9,8%), ocupando el tercer lugar en patogenicidad, después de Salmonella spp. (35,4%) y los virus (13,1%). Entre las toxinas bacterianas, las enterotoxinas participaron en 291 de los 525 brotes de intoxicación alimentaria notificados (55,4%), o 5,5% de todos los brotes notificados en el 2008 (Hennekinne et al., 2010) y en el 2009, se informó que el queso seguido por las mezclas de comidas o “buffet” eran los dos principales causantes de intoxicaciones alimentarias por enterotoxinas de S. aureus (Schelin et al., 2011). Entre los años 1993 y 2000, en América Latina ocurrieron 191 brotes por intoxicación estafilocócica con 6,433 afectados y 2 muertes, no se conocen reportes más actuales (Valdiviezo et al., 2006). La intoxicación alimentaria por S. aureus se ha considerado, a nivel mundial y en Colombia, como la segunda causa de enfermedades transmitidas por alimentos (ETA), después de Campylobacter o Salmonella (Agrawal et al., 2012), los cuales ocupan los primeros lugares de los reportes anuales de la red de vigilancia en salud pública en Colombia (Vanegas et al., 2008). En Colombia, las autoridades encargadas de realizar acciones de inspección, vigilancia y control como el seguimiento epidemiológico a las ETA; detectan, notifican y reportan Staphylococcus coagulasa positiva, pero no se conoce la relación que existe entre el estado de portador y la intoxicación alimentaria (Boerema et al., 2006, El-Huneidi

27
et al., 2006), mientras que en el contexto internacional, la literatura científica se enfoca en S. aureus enterotoxigénico. Los datos suministrados por 11 de los 33 Laboratorios de las Direcciones Territoriales de Salud (DTS) de Colombia (Antioquia, Cundinamarca, Caldas, Córdoba, Distrito Capital, Nariño, Norte de Santander, Quindío, Risaralda, Valle del Cauca y Vichada), correspondientes al periodo 2000 – 2010 del sistema de Inspección, Vigilancia y Control (IVC), reportaron 23.009 alimentos con presencia de Staphylococcus coagulasa positiva3. De éstos, 2,779 (45,46%) de las muestras corresponden al grupo denominado alimentos preparados no industriales, los cuales fueron elaborados principalmente en restaurantes, colegios, establecimientos penitenciarios, casinos, hogares, clubes sociales, entre otros; de los 107 alimentos de preparados no industriales, los más relevantes por presencia de Staphylococcus coagulasa positiva fueron las preparaciones con pollo. En el año 2011, se notificó al Sistema nacional de vigilancia por archivos planos (colectivo) 13.961 casos de enfermedades transmitidas por alimentos, involucrados en 899 brotes; el 56,3% de los casos fueron confirmados por clínica, el 43,7% se encuentran asociados a la identificación de algún agente etiológico. Los alimentos que producen mayores intoxicaciones en Colombia son quesos, arroz con pollo. El lugar de consumo de mayor incidencia en la ocurrencia de brotes ETA fue el hogar (472 brotes) que representa el 53%, seguido de otros con 12% (108 brotes), restaurante comercial con 11,5% (104 brotes) y establecimiento educativo 11,3% (102 brotes). Dentro de la clasificación otros, las ventas ambulantes fueron las que tuvieron mayor frecuencia; las medidas sanitarias más aplicadas por parte de las autoridades fueron: medidas preventivas (174 brotes), decomiso de productos (54 brotes), suspensión total o parcial de trabajos o servicios (34 brotes), clausura temporal, total o parcial (8 brotes), entre otras. Los agentes etiológicos detectados en muestras biológicas, de alimentos o restos de alimentos y superficies, procedentes de brotes ETA en notificación colectiva fueron: Staphylococcus aureus coagulasa positivo, Salmonella spp, Escherichia coli, Listeria monocytogenes, Shigella spp, Bacillus cereus, entre otros y sustancias químicas como organofosforados y carbamatos4. Los alimentos son un importante factor de riesgo para la transferencia de resistencias a antibióticos. Esta transferencia puede ocurrir por residuos de antibióticos en los alimentos; los alimentos pueden transmitir patógenos resistentes o a través de la ingestión de cepas resistentes (Khan et al., 2000, Pesavento et al., 2007, Ortega et al., 2010). Las cepas de S. aureus son frecuentemente resistentes
3 MPS. Ministerio de la Protección Social de Colombia. Sistema de Inspección, Vigilancia y Control de las Direcciones
Territoriales de Salud (IVC). 2010.
4 MPS. Ministerio de la Protección Social de Colombia. Sistema de Inspección, Vigilancia y Control de las Direcciones
Territoriales de Salud (IVC). 2013.

28
a antibióticos debido a su capacidad para producir una barrera exopolisacárida que debido a su ubicación dentro de microabscesos limitando la acción de los antibióticos (Gündogan et al., 2000, Ortega et al., 2010).
2.5.1 Epidemiología de las intoxicaciones alimentarias. Entre 1969 y 1990, se produjeron en el Reino Unido 359 intoxicaciones que revelaron que el 79% de las cepas de S. aureus producían SEA (Argudin et al., 2010, Wieneke et al., 1993) y fue asociada a productos cárnicos y avícolas. Se detectó que SEA, estaba presente en el 56,9% de las intoxicaciones y se relacionó SEA con otras enterotoxinas SED, SEB, SEC o SEB y SED, obteniendo que las otras enterotoxinas causaban un número menor de intoxicaciones. Entre 1981 y 2002, 31 brotes fueron analizados, encontrando que SEA fue la enterotoxina más frecuente en Francia (69,7%), la intoxicación estaba asociados productos lácteos, diferentes tipos de carne y ensaladas (Kerouanton et al., 2007). SEA también fue la enterotoxina más común en brotes de intoxicación alimentaria en EE.UU. (77,8%), seguida de SED y SEB (Casman, 1965). Un estudio de S. aureus obtenido a partir de productos lácteos, responsables de 16 brotes en Brasil reveló que el gen de la enterotoxina más frecuente fue sea seguido por seb (Veras et al., 2008). En Austria, en el 2007 una intoxicación alimentaria por productos lácteos que afecto a 40 niños se atribuyó a las enteroxinas SEA y SED (Kitamoto et al., 2009). SEA también fue la enterotoxina más común asociado a las intoxicaciones por alimentos en Japón, en el año 2000 una intoxicación por leche baja en grasa, poseía la enterotoxina SEA, mientras que la intoxicación más reciente (2009) se debió a crepes contaminados con SEA y SEC (Kitamoto et al., 2009). En un estudio en Japón se demostró que el gen sea es el más frecuente con un 58% y seguido de seb con 44% (Omoe et al., 2005a).
2.5.2 Epidemiología de portadores sanos: impactos de los portadores sanos de S. aureus en salud pública. Entre las bacterias que originan enfermedades, S. aureus ocupa un papel destacado como agente etiológico de gastroenteritis que son frecuentes por consumo de alimentos contaminados y esta afección se conoce como intoxicación alimentaria estafilocócica (IAE) (Brizzio et al., 2011). S. aureus es un patógeno común asociado con la comunidad y enfermedades adquiridas en hospitales, se considera como un problema importante de Salud Pública (Ortega et al., 2010). Las fosas nasales especialmente los orificios son la colonización primaria de la cepas de S. aureus (Kreiswirth, 1989, Williams, 1963). El portador de S. aureus juega un papel clave en la epidemiología y la patogenia de la infección, y se asocia

29
como el mayor riesgo de complicaciones infecciosas después de una cirugía en pacientes con insuficiencia renal en fase terminal y en aquellos con dispositivos intravasculares (Kluytmans et al., 1997, Wenzel and Perl, 1995). Aproximadamente, el 80% sufre infecciones invasivas nosocomiales de origen endógeno en portadores nasales (Wertheim et al., 2005, Mertz et al., 2007). La intoxicación alimentaria por estafilococo se conoce como estafiloenterotoxicosis o estafiloenterotoxemia y se produce por consecuencia de la ingestión de alimentos que contienen toxinas preformadas. Las cepas de S. aureus una vez que llegan a los alimentos, si las circunstancias lo permiten, se multiplican y producen toxinas. La contaminación de los alimentos en la mayoría de los casos suele ocurrir después de ser cocinados, cuando no son conservados adecuadamente, lo que favorece su multiplicación (Argudin et al., 2010).
El estado de portador sano ha sido identificado como un factor de riesgo en el desarrollo de infecciones. Se ha estudiado extensamente en pacientes quirúrgicos, con hemodiálisis, diálisis peritoneal y otras condiciones (Kluytmans and Wertheim, 2005). Los portadores de cepas de S. aureus en fosas nasales y manos, cuando son manipuladores de alimentos, son la más importante fuente de intoxicación alimentaria estafilocócica, a través del contacto manual o mediante las secreciones respiratorias, en los restaurantes y puntos de comidas rápidas. De hecho, S. aureus es un comensal de la piel y las membranas mucosas de los seres humanos, del 20 al 30% presentan una colonización persistente y el 60% intermitente (Argudin et al., 2010, Kluytmans and Wertheim, 2005).
Históricamente, tres patrones de portadores se reconocen en los seres humanos: el portador persistente, el portador intermitente, y no persistente (Nouwen et al., 2004, Kluytmans and Wertheim, 2005, Manenschijn et al., 2012). Entre el 10 y el 35% de individuos sanos casi siempre llevan una cepa y son portadores persistentes, en una mayor proporción (20 a 75%) los individuos son portadores en forma intermitente de cepas de S. aureus, por último, entre el 5 y el 50% casi nunca son portadores de cepas (Kluytmans and Wertheim, 2005, Williams, 1963). El portador persistente tiene un mayor riesgo de desarrollar infecciones por S. aureus y una colonización mayor en comparación con los portadores intermitentes. Los portadores persistentes son generalmente colonizados por una cepa específica, mientras que los portadores intermitentes pueden tener diferentes cepas (Williams, 1963, Kluytmans and Wertheim, 2005). Sin embargo, recientemente se encontró que el intermitente y los no portadores comparten similitudes en la eliminación cinética y los perfiles de anticuerpos antiestafilocócicos, lo que sugiere que podría haber sólo dos tipos portadores, persistentes y otros (Manenschijn et al., 2012, van Belkum et al., 2009).

30
La patogénesis de la colonización nasal de S. aureus todavía no es clara, pero el sistema inmune en la determinación del estado del portador, así como los factores ambientales y humorales en las secreciones nasales son determinantes (Manenschijn et al., 2012, Wertheim et al., 2004). Otro factor que parece influir en el estado de portador es la sensibilidad de glucocorticoides (GC). Los polimorfismos del gen del receptor de glucocorticoides, son asociados con diferencias en la sensibilidad en el contenido de GC y han sido relacionadas con el estado de portador de S. aureus (van den Akker et al., 2006). El principal glucocorticoide en los seres humanos es el cortisol. El cortisol es capaz de suprimir el sistema inmunológico y los polimorfismos afectan la sensibilidad a GC, y parece que influyen en el estado de portador de S. aureus, los niveles de cortisol a largo plazo pueden variar entre los diferentes estados de portador (Manenschijn et al., 2012). La carga microbiana de S. aureus es mayor en portadores persistentes en comparación con los portadores intermitentes, lo que resulta en más dispersión y mayor riesgo de infección para la población (Kluytmans and Wertheim, 2005). Las tasas de portadores se aumentan cuando hay casos de sinusitis, faringitis y procesos gripales. Se ha reportado que un 34,4% de adultos de ambos sexos pueden tener S. aureus en la mucosa nasal y un 17,2% en la piel. Entre el 2003-2004, aproximadamente el 29% (78,9 millones de personas) de la población de los E.E.U.U. estaba colonizada en su mucosa nasal por S. aureus. Se ha demostrado que las cepas aisladas de manos pueden ser del mismo tipo que las aisladas de mucosa nasal (Gorwitz et al., 2008). En cualquier caso, las principales fuentes de contaminación son los seres humanos (los manipuladores de alimentos a través del contacto manual o por medio de las vías respiratorias al toser o estornudar), y la contaminación se puede producir después del tratamiento térmico de los alimentos (Ortega et al., 2010). El estado de portador de cepas de S. aureus en manipuladores de alimentos no es una enfermedad, sino más bien una condición no deseada que debe ser controlada para evitar la contaminación de los alimentos y las consecuencias a que ello conlleva.
2.6 Patogénesis y enfermedades asociadas. La patogenicidad de S. aureus es causada por la expresión de un arsenal de factores de virulencia contra las defensas del hospedero. Durante los últimos 100 años y más, S. aureus ha causado muchas infecciones en la comunidad y hospitales, desarrollando resistencia a todos los antibióticos utilizados en su contra, sin embargo, los mecanismos exactos que conducen a las

31
epidemiología de la infección virulenta no se conocen completamente (Ortega et al., 2010).
Este microorganismo es un patógeno versátil capaz de causar un amplio espectro de enfermedades humanas y en los animales, debido a su virulencia, toxicidad, invasión y resistencia a los antibióticos. La gama de infecciones por estafilococos oscila entre los granos y forúnculos, todas estas enfermedades dependen de numerosos factores de virulencia; el estafilococos produce muchas infecciones supurativas como absceso cutáneo, impétigo, foliculitis, ántrax, celulitis cuello y cara, hidradenitis supurada, mastitis, bacteriemia, endocarditis, neumonía, osteomielitis, artritis séptica, meningitis, peritonitis, pericarditis, piomiositis (Kumate et al., 2008) y enfermedades por producción de toxinas como el síndrome de la piel escaldada, intoxicación alimentaria y síndrome de choque tóxico estafilocócico.
Por otro lado, algunas infecciones, como la intoxicación alimentaria estafilocócica, dependen de un solo tipo de factor de virulencia: el SEs (Ortega et al., 2010).
La intoxicación alimentaria estafilocócica, se produce como consecuencia al consumir alimentos contaminados con cepas de S. aureus, se absorben las toxinas y causan gastroenteritis, mientras que las bacterias pasan a través del intestino sin causar efectos adversos sobre el huésped. Las toxinas estimulan el nervio vago en las terminaciones de las paredes estomacales y transmite la señal al centro del vómito que muestra una respuesta emética. La intoxicación alimentaria por SEs produce cambios inflamatorios en el tracto gastrointestinal con lesiones graves en el yeyuno e íleon. Estudios en animales demostraron que la enterotoxinas SEA y SEB están asociadas con el aumento del número de linfocitos y células polimorfonucleares en el yeyuno y el duodeno, produciendo una rápida respuesta emética y diarrea, lo que sugiere que el intestino es el sitio de acción de las enterotoxinas. A pesar de los avances en la comprensión de las SEs asociadas a la inflamación del tracto gastrointestinal, todavía no está claro cómo se inicia esta inflamación in vivo y cuál es la función exacta de las células inmunes o no, que contribuyen a la progresión de la enfermedad (Pinchuk, et al., 2010). Aún no se han identificado los receptores que intervienen y queda mucha incertidumbre con respecto a los eventos tempranos en la patogénesis de la intoxicación alimentaria estafilocócica (SFP, Staphylococcal food poisoning). Una de las principales hipótesis es que la emesis, que se produce en respuesta a la inflamación inducida por las enterotoxinas (Pinchuk, et al., 2010). Los síntomas de la SFP están relacionados con un número de citosinas inflamatorias, incluyendo la prostaglandina E2, leucotrieno B4 y Ácido 5-hidroxieicosatetraenoico (Jett et al., 1990). Cisteinil leucotrienos, como leucotrieno E4, también han sido implicado como mediador crítico en SFP (Scheuber et al., 1987).

32
No está claro si estos mediadores se generan directa o indirectamente, en respuesta a las SEs. En última instancia, la respuesta emética a la SEs es dependiente de la estimulación por los impulsos transmitidos a través de los nervios vago y simpático. Varios grupos han propuesto que los mastocitos son la fuente primaria de mediadores inflamatorios liberados durante la SFP (Reck et al., 1988, Komisar et al., 1992).
2.7 Diagnóstico de la infección. Los diagnósticos por intoxicaciones alimentarias por estafilococos, generalmente sólo evalúan los síntomas del paciente, realizando un examen físico y para construir la historia médica. En el diagnóstico de intoxicación alimentaria por S. aureus productor de toxinas pueden ser identificadas en las heces o vómitos utilizando técnicas especializadas. Las toxinas también pueden ser detectadas en los alimentos contaminados. Las técnicas de detección de bacterias productoras de toxinas o de sus toxinas usualmente no se hacen en pacientes individuales y es reservada a brotes que involucran varios individuos. La intoxicación alimentaria resulta del consumo de alimentos en los que S. aureus se ha multiplicado por encima de 105 UFC/g y producido enterotoxinas (SEs), que contienen cantidades suficientes de una (o más) enterotoxinas (Dinges et al., 2000, Le Loir et al., 2003). Las SEs son resistentes al calor (desde 5-10 minutos a 121 ◦C hasta varias horas a 80 ◦C - 100 ◦C), a la irradiación gamma (valores D entre 27-93 kGy) y la acción de las proteasas intestinales como la pepsina. La cantidad necesaria de enterotoxina requerida para causar la intoxicación es muy pequeña del orden de 0,5 µg kg-1 en monos y 90 ng en el hombre, siendo para éste la dosis emética del orden de 0,2 µg kg-1 (Balaban and Rasooly, 2000, Mossel et al., 2002). La sintomatología por intoxicación, tienen un inicio rápido (2-8 h) e incluyen náuseas, vómitos violentos, dolor abdominal, con o sin diarrea (Balaban and Rasooly, 2000, Argudin et al., 2010, Tranter, 1990), la intensidad de los síntomas depende de la cantidad de alimento contaminado ingerido, de la concentración de la toxina y de la susceptibilidad individual, la cual esta mediada por la edad y el estado inmunológico de la persona (Kim et al., 2011) El S. aureus no compite bien con la microbióta autóctona en los alimentos crudos, la contaminación se asocia principalmente con el manejo inadecuado de los alimentos cocinados o procesados, seguidos por el almacenamiento en condiciones que permitan el crecimiento de S. aureus y la producción de la enterotoxinas (Argudin et al., 2010). Sin embargo, S. aureus está presente también en animales productores de alimentos como ganado lechero, ovino y caprino, especialmente si están afectados por mastitis subclínica, que son los contaminantes más frecuentes de la leche. El aire, polvo, superficies en contacto con los alimentos también pueden

33
servir como vehículos para la transferencia de S. aureus a los alimentos (Argudin et al., 2010).
Los alimentos que han sido frecuentemente asociados a una intoxicación por S. aureus incluyen productos cárnicos, productos avícolas, productos lácteos, productos vegetales, productos de panificación (Wieneke et al., 1993, Tamarapu et al., 2001). Los productos alimenticios, tales como jamón (Qi and Miller, 2000), también han sido asociados a intoxicaciones alimentarias, de acuerdo con la capacidad de S. aureus para crecer a una actividad acuosa relativamente baja aw: 0,86 (Scott, 1953).
3. METODOLOGÍA.

34
3.1 Tipo de estudio.
Es un estudio Descriptivo - Analítico con una fase analítica experimental, donde se detectaron los portadores sanos de Staphylococcus aureus en madres comunitarias del ICBF de Pereira.
3.2 Diseño maestral. 3.2.1 Población. La población fue de 406 madres comunitarias del Instituto Colombiano de Bienestar Familiar de Pereira (ICBF), para este caso se dispuso de la base de datos del ICBF, facilitada por los nutricionistas del ICBF seccional Risaralda.
3.3 Técnicas y procedimientos
3.3.1 Procedimiento para obtención de S. aureus. Para los aislamientos de S. aureus se tomaron muestras de las fosas nasales y de manos, en el Laboratorio de Microbiología de la Universidad Tecnológica de Pereira a las madres comunitarias que aceptaron y firmaron el consentimiento informado. El procedimiento realizado fue el siguiente: con un escobillón estéril humedecido en una solución salina fisiológica estéril se realizó frotis de la parte interna anterior de las fosas nasales y en las manos entre la uña y los pulpejos de los dedos se cultivó en Agar Salino Manitol rojo de fenol (ASMRF) y se incubaron a 37°C por 24 - 48 h. Del cultivo primario se seleccionó colonias manitol positivo y luego se realizó la coloración de Gram para determinar microscópicamente la presencia de cocos Gram positivos, posteriormente estas colonias se sembraron en ASMRF para obtener un cultivo masivo y puro (37°C - 24 h). A partir del cultivo puro se inocularon los caldos BHI y caldo glucosado en anaerobiosis (prueba de fermentación de la glucosa en anaerobiosis), se incubaron por 24 h a 37°C. A partir de los cultivos de caldo BHI se realizaron las pruebas de coagulasa y termonucleasa. Prueba de coagulasa: se mezclaron 200 µl de plasma de conejo con 200 µl de caldo BHI inoculado el día anterior con cada una de las colonias, la mezcla fue incubada en baño maría a 37°C durante 4h, se observó la formación del coágulo a las 4 horas. Prueba de Termonucleasa: Se empleó agar DNasa con azul de toluidina, el cual, se sirvió en cajas de Petri. El caldo nutritivo inoculado con las cepas se hirvió por 10 min. Después, se adicionó a los pozos del agar y se incubaron a 37°C en por 24h. Cuando la prueba de coagulasa y/o termonucleasa fueron positivas; del cultivo puro se sembró en un caldo nutritivo (2 ml) y se incubó a 37°C en un tiempo de 16 h. Se inoculó 400 µl del caldo nutritivo con crecimiento en eppendorf con 100 µl de glicerol estéril y se guardó a -80°C.

35
Los aislamientos de S. aureus identificados en el laboratorio, se reconfirmaron por el sistema automatizado VITEK 2C, en el Laboratorio Clínico de microbiología del Hospital Universitario San Jorge, en un conjunto de paneles de microdilución que contiene medios de cultivo, diluciones de antibióticos en serie y productos bioquímicos seleccionados (Biomerieux). En el sistema automatizado (VITEX 2C) se determinó el S.aureus, basándose en la inoculación de una suspensión de microorganismos en tarjetas con determinados paneles de reacciones bioquímicas. La sensibilidad antimicrobiana se llevó a cabo en forma similar a través de tarjetas que contenían diluciones estandarizadas de distintos antibióticos correspondientes a los puntos de corte de sensibilidad. El control de calidad se realizó con la cepa de referencia ATCC 29213 de S.aureus, recomendada por ATCC (American Type Culture Collection, Manassas, Va, USA) y se siguió los parámetros bioquímicos en la identificación del microorganismo, con el rango de las Concentraciones Mínimas Inhibitorias (CMI).
3.3.2 Diagnóstico Molecular.
3.3.2.1 Extracción de ADN.
A los aislamientos de S. aureus se les realizó la extracción del ADN genómico. Se siguió el protocolo de extracción CTAB/Lisostafina, utilizando método CTAB “Bromuro de cetil –trimetil–amonio” (Ausubel et al; 1889), estandarizándolo con la enzima lisostafina “AMBI BL” (Corredor et al; 2011), para romper la pared del S. aureus, con algunas modificaciones. Se sembró densamente un cuarto de cultivo puro con un aplicador estéril en cajas de Petri incubándolo por 24 horas, luego se resuspendió en un 1 ml de amortiguador TE 1X (Tris-EDTA; pH: 8.0), agitándolo vigorosamente y se centrifugó a 14.000 rpm por 2 minutos, descartando el sobrenadante, quedando un pellet o botón. Posteriormente, el pellet se resuspendió
en 567 l de buffer fosfato salino (PBS) por pipeteo repetitivo y se le adicionó 10 l de la enzima lisostafina de Staphylococcus simulans (AMBI) a una concentración
de 10 mg/l incubándolo en un baño maría por una hora a 37°C, hasta observar una
lisis celular eficiente, aproximadamente una hora. Después, se le adicionaron 30 l
de SDS (10%) y 3 l de proteinasa K (20 mg/ml), se mezcló el contenido por inversión del tubo y se incubó por 1 hora a 37°C en un baño maría, en seguida se
añadieron 100 l de NaCl 5 M y se mezcló. Luego, se agregó 80 l de solución CTAB 1% / NaCl 0.7 M, mezclándolo por inversión del tubo e incubándolo a 65°C por 10 minutos. El ADN se extrajo con un igual volumen de cloroformo/alcohol isoamílico (24:1), vórtex durante 30 segundos y se centrifugó a 13.000 rpm por 15 minutos, llevándose la fase acuosa a un tubo estéril sin arrastrar la interfase. La precipitación del ADN se logró con 0,6 volúmenes de isopropanol e inversión repetida. Inmediatamente se centrifugó a 13.000 rpm durante 15 minutos a una temperatura de 4°C, descartando el sobrenadante y lavando el precipitado con 500

36
l de etanol al 70%, el botón se dejó secar a temperatura ambiente y se resuspendió
con 200 l de TE 1X durante.
La concentración de ADN fue medida en un espectrofotómetro a 260 nm, estimando la pureza y cantidad de ADN, se realizaron alícuotas de 20 ng/µl y se almacenaron a -20°C, para su posterior análisis molecular.
3.3.2.2 Detección genotípica de superantígenos en los aislamientos de S. aureus. La detección de los genes que codifican superantígenos, se efectuó por medio de una PCR múltiple en dos series: A y B. La técnica se estandarizó con las cepas de referencia de S. aureus (Tabla 3) y como control interno de la reacción se utilizó una pareja de iniciadores femA1 y femA2 (Tabla 4). El control positivo de la reacción fue una mezcla de ADN constituido por diferentes aislamientos toxigénicos de S. aureus que amplificaron los diferentes superantígenos estudiados y el control negativo fue agua desionizada estéril. Tabla 3. Cepas de referencia ATCC de S. aureus utilizadas como control positivo.
Cepa Designación Genes de S. aureus
Características
Referencia
ATCC 700679
Mu50 mecA Resistencia a Meticilina. Baja susceptibilidad
a Vancomicina.
(Hiramatsu et al., 1997)
Nucleotide (GenBank): BA000017 Staphylococcus aureus subsp. aureus Mu50 DNA, complete genome.
ATCC BAA-1707
USA 400 MW2[HIP0827
0]
mecA Resistencia a Meticilina.
(Hiramatsu et al., 1997)
Nucleotide (GenBank): BA000033 Staphylococcus aureus subsp. aureus MW2 DNA,

37
complete genome Nucleotide (GenBank): NC_003923 Staphylococcus aureus subsp. aureus MW2, complete genome.
ATCC 13565
FDA 196E sea, seb Producción de enterotoxinas.
(Casman et al., 1963)
PubMed: 14042955 92258: Electrophoretic and Immunoblot Analysis of Staphylococcal Enterotoxins in Food. Washington, DC: US Food & Drug Administration; Bacteriological Analytical Manual Online Chapt.13b, 2001.
ATCC 13566
FDA S6 tst, seb Producción de enterotoxinas
(De Boer and Chow, 1994) PubMed: 7930723.
ATCC 19095 ATCC 29213
FRI137
Wichita
sec
sak
Producción de enterotoxinas
Sensible a la meticilina y oxacilina
((Bergdoll et al., 1965, Marr et al., 1993) PubMed: 8406814.
(Bergdoll et al., 1965)

38
PubMed: 4954560. Nucleotide (GenBank): U77328 Staphylococcus aureus staphylokinase gene, partial cds.
PCR múltiple para la serie A. La Serie A se utilizó para detectar los genes de las enterotoxinas clásicas sea, seb, sec, sed y see, utilizando los iniciadores (primers) descritos en la Tabla 4. Tabla 4. Iniciadores específicos para las enterotoxinas clásicas de S. aureus.
Gen Secuencia del iniciador (5´- 3´) Tm Tamaño (pb)
sea GGT TAT CAA TGT GCG GGT GG CGG CAC TTT TTT CTC TTC GG
56,4 54,4
102
seb GTA TGG TGG TGT AAC TGA GC CCA AAT AGT GAC GAG TTA GG
53 50,1
164
sec AGA TGA AGT AGT TGA TGT GTA TGG CAC ACT TTT AGA ATC AAC CG
52,3 49,1
451
sed CCA ATA ATA GGA GAA AAT AAA AG ATT GGT ATT TTT TTT CGT TC
44,6 45,3
278
see AGG TTT TTT CAC AGG TCA TCC CTT TTT TTT CTT CGG TCA ATC
53,1 48,1
209
femA1 femA2
AAA AAA GCA CAT AAC AAG CG GAT AAA GAA GAA ACC AGC AG
55,2 54,3
132
Fuente: Mehrotra, et al 2000. La reacción de amplificación se efectuó en un volumen final de 25 µl, con buffer 1X
(KCl 50 M, tris-HCl 10 M (pH 8.0)), MgCl2 3 mM, dNTPs 200 µM, 20 pmoles de cada uno de los iniciadores de sea, seb, sec, see y sed, 1 U de Taq polimerasa (Invitrogen) y 3 µl del ADN a una concentración de 20 ng/µl. En un Termociclador Mastercycler® gradient, EPPENDORF; con un programa de temperaturas que consistió en una predesnaturalización a 96°C por un tiempo de 3 minutos, seguido por 35 ciclos de desnaturalización: 94°C durante 1 minuto, 53°C durante 30 segundos y 74°C por 1 min y un paso final de extensión por 7 min a 72°C.

39
PCR múltiple para la serie B. La Serie B se utilizó para detectar los genes de las toxinas exfoliativas eta, etb y la toxina del síndrome de choque tóxico tsst-1, utilizando los iniciadores (primers) descritos en la Tabla 5. Tabla 5. Iniciadores para la Toxina del Síndrome de Choque Tóxico y las Toxinas Exfoliativas y para etd.
Gen Secuencia del iniciador (5´- 3´) Tm(°C) Tamaño (pb)
tsst-1
eta
ACC CCT GTT CCC TTA TCA TC TTT TCA GTA TTT GTA ACG CC
GCA GGT GTT GAT TTA GCA TT AGA TGT CCC TAT TTT TGC TG
53,9 48,6
51,3 50,2
326
93
etb
ACA AGC AAA AGA ATA CAG CG GTT TTT GGC TGC TTC TCT TG
51,4 52,5
226
etd
femA1 femA2
AAC TAT CAT GTA TCA AGG CAC ACT TTT AGA ATC AAC CG
AAA AAA GCA CAT AAC AAG CG GAT AAA GAA GAA ACC AGC AG
52,3 49,1
55,2 54,3
376
132
Fuente: Mehrotra, et al 2000 y Ruzickova et al 2005. La reacción de amplificación para la serie B de la PCR múltiple, tuvo los mismos pasos que la serie A, solo difirió en la concentración de ADN; se amplificó con un
volumen final de 25 µl, con buffer 1X (KCl 50 M, tris-HCl 10 M (pH 9.0)), MgCl2 3 mM, dNTPs 200 µM, 20 pmoles de cada uno de los iniciadores de eta, etb, etd y tst-1, 1 U de Taq polimerasa (Invitrogen) y 1 µl del ADN a una concentración de 20 ng/µl. En un Termociclador Mastercycler® gradient, EPPENDORF, se sometió la amplificación bajo las siguientes condiciones: a 96°C por 3 minuto, 35 ciclos de 94°C por 1 minuto, 53°C por 30 segundos y 74°C por 1 minuto, por ultimo 72°C por 7 minutos.
Se obtuvieron 25 µl de los productos de ambas series y se corrieron en una electroforesis, con un marcador de peso molecular HyperLadderTM IV de Bioline en geles de agarosa al 2 %, teñidos con 10 µl de SYBR Safe, con un buffer TAE 1X (Tris-EDTA; pH: 8.0) a 45 voltios, durante 1 hora y 30 minutos. La observación de

40
los resultados se realizó a través de un fotodocumentador Gel Doc™ XR+ System de Bio-Rad con un software de ImageLab.
PCR simple para la identificación de la toxina exfoliativa D (gen etd).
Se amplifico siguiendo la metodología propuesta por Ruzickova et al 2005. Las condiciones de amplificación para la PCR, fueron idénticas a las de la serie A, usando el iniciador de la tabla 5.
3.4 Análisis estadístico. Se calculó la proporción de aislamientos positivos para los genes que codifican los superantígenos de enterotoxinas (SE) de Staphylococcus aureus de madres comunitarias utilizando métodos moleculares. Las variables de respuesta relacionadas con el estado de portador, presencia o ausencia de los genes en fosas nasales y manos, se organizaron en tablas de contingencia, y se comparó la prevalencia de los genes o los superantígenos versus resistencia. Para todos los análisis estadísticos se utilizó el software informático SPSS IBM 19, aplicando la prueba exacta de Fisher y los valores de probabilidad (p) ≤ 0,05 con un intervalo de confianza del 95%, se consideraron estadísticamente significativos.
4. RESULTADOS.

41
4.1 Identificación de S. aureus. Con la aplicación de los métodos de rutina microbiológicos y la ratificación por el sistema automatizado VITEX 2C, se lograron obtener 95 aislamientos para S. aureus.
4.2 Prevalencia de estado de portador. La prevalencia del estado de portador de S. aureus en las madres comunitarias del ICBF de Pereira fue del 22% (95/406) tanto en fosas nasales como en manos. Adicionalmente, se estableció que el 17% (69/406) fueron portadoras solamente en fosas nasales y 5% (20/406) en manos; y en el 1,5% (6/406) del total de madres comunitarias eran portadoras tanto en fosas nasales como en manos. Anexo A-B
Detección genotípica de los genes de superantígenos en los 95 aislamientos de S. aureus. Las PCRs aplicadas para detectar los genes sea, seb, sec, sed, see, eta, etb, etd y tsst-1 en los 95 aislamientos de S. aureus, amplificaron casi todos los genes estudiados, excepto para el gen etb. En las figuras 2 y 3, se muestran algunos resultados donde se puede observar los diferentes amplicones correspondientes a los genes detectados.
Figura 2. PCR múltiple de la serie A. Detección de los genes sea, seb, sec, sed y see. Los Carriles 1,5 y 15 gen sea (102 pb); carriles 3 y 11 gen seb (164 pb), carril 11 gen sec (451 pb), carril 3 gen sed (278 pb), y carril 11 gen see (209 pb), Control negativo (N) y control positivo (P). Control interno femA (132 pb). Carril 10 el marcador de peso molecular (HyperLadder II: 50bp-2000 pb).

42
Figura 3. PCR múltiple para la serie B. Detección de los genes tsst-1 y eta. Carriles 1, 6 y 10 gen tsst-1 (326pb); carril 7 gen eta (93 pb). Control negativo (N) y control positivo (P). Control interno femA. Carril 12 marcador de peso molecular (HyperLadder II: 50bp-2000 pb). En la figura 4, se muestra el patrón electroforético de los amplicones para la detección del gen etd.
Figura 4. PCR simple para la detección de la toxina exfoliativa D (gen etd). Los carriles de 1 al 3 y el carril 6 amplificaron para el gen etd (366 pb). Control negativo (N) y control positivo (P). Carril 4 marcador de peso molecular (HyperLadder II: 50bp-2000 pb). 4.3 Prevalencia Global de los genes de SAg. El número de aislamientos que fueron positivos para uno o más genes de superantígenos fue de 70, para una prevalencia de 73,7% (70/95) y los aislamientos negativos fueron de 26,3% (25/95). El mayor porcentaje en los aislamientos positivos fue para un solo gen con 34,7% (33/95) y la mayor asociación de genes fue de dos genes con 24,2% (23/95). El número máximo de genes detectados en un único aislamiento fue de cuatro (4,2%). Ver Tabla 6. Anexo C-D-E.F
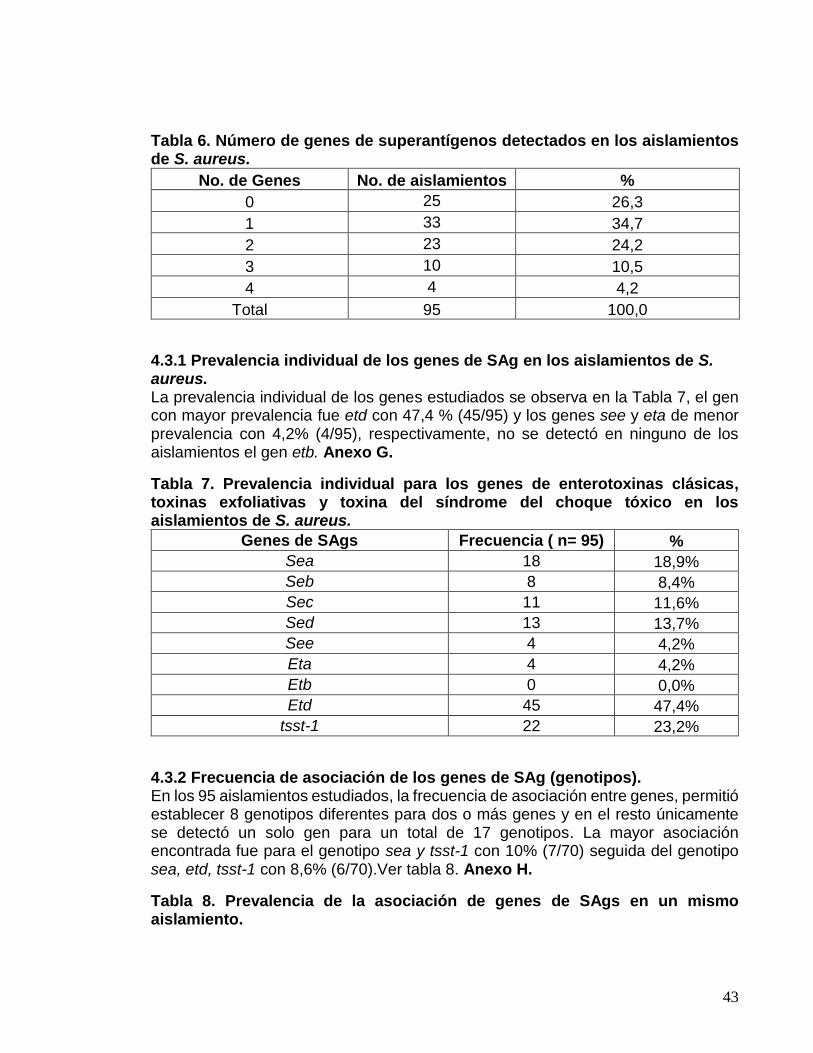
43
Tabla 6. Número de genes de superantígenos detectados en los aislamientos de S. aureus.
No. de Genes No. de aislamientos %
0 25 26,3
1 33 34,7
2 23 24,2
3 10 10,5
4 4 4,2
Total 95 100,0
4.3.1 Prevalencia individual de los genes de SAg en los aislamientos de S. aureus. La prevalencia individual de los genes estudiados se observa en la Tabla 7, el gen con mayor prevalencia fue etd con 47,4 % (45/95) y los genes see y eta de menor prevalencia con 4,2% (4/95), respectivamente, no se detectó en ninguno de los aislamientos el gen etb. Anexo G.
Tabla 7. Prevalencia individual para los genes de enterotoxinas clásicas, toxinas exfoliativas y toxina del síndrome del choque tóxico en los aislamientos de S. aureus.
Genes de SAgs Frecuencia ( n= 95) %
Sea 18 18,9%
Seb 8 8,4%
Sec 11 11,6%
Sed 13 13,7%
See 4 4,2%
Eta 4 4,2%
Etb 0 0,0%
Etd 45 47,4%
tsst-1 22 23,2%
4.3.2 Frecuencia de asociación de los genes de SAg (genotipos). En los 95 aislamientos estudiados, la frecuencia de asociación entre genes, permitió establecer 8 genotipos diferentes para dos o más genes y en el resto únicamente se detectó un solo gen para un total de 17 genotipos. La mayor asociación encontrada fue para el genotipo sea y tsst-1 con 10% (7/70) seguida del genotipo sea, etd, tsst-1 con 8,6% (6/70).Ver tabla 8. Anexo H.
Tabla 8. Prevalencia de la asociación de genes de SAgs en un mismo aislamiento.

44
Asociaciones de genes No. %
sea, tsst-1 7 10,0
sea, etd, tsst-1 6 8,6
sec, etd 5 7,1
seb,sed 3 4,3
seb, sed, etd 2 2,9
sed, etd 2 2,9
sed, tsst-1 2 2,9
etd, tsst-1 2 2,9
4.4 Susceptibilidad a los antimicrobianos. Los resultados de la susceptibilidad antimicrobiana (sensibles: S y resistentes: R) obtenidos por el sistema automatizado VITEK 2C se muestran en las Tablas 9 y 10, incluyendo en los resultados la prueba de la β–Lactamasa. En la Tabla 9, en general, 74,7% (71/95) de los aislamientos eran resistentes al menos a un antibiótico, y en tres aislamientos, 3,2% (6/95) fueron resistentes a cuatro antibióticos. Los aislamientos sensibles de S. aureus fueron 6,3% (6/95). Anexo I-JI.
Tabla 9. Porcentaje de aislamientos de S. aureus según la resistencia a uno o más antimicrobianos.
No. antimicrobianos No. aislamientos %
0 6 6,3
1 71 74,7
2 9 9,5
3 6 6,3
4 3 3,2
Total 95 100
La tabla 10 muestra los porcentajes de resistencia y la producción de β–Lactamasa; 93,7% de los aislamientos (89/95) fueron positivos para la producción de la β–Lactamasa y para los antimicrobianos probados, el de mayor resistencia fue para tetraciclina con 11,6% (11/95) y el de menor resistencia fue a clindamicina con 3,2% (3/95). Anexo L.
Tabla 10. Prevalencia de resistencia en los aislamientos de S. aureus a diferentes antimicrobianos incluyendo la producción de β- Lactamasa.
Antimicrobiano No. Aislamientos %
β- Lactamasa 89 93,7
Tetraciclina 11 11,6
Cefoxitina 6 6,3
Oxacilina 6 6,3

45
Eritromicina 4 4,2
Clindamicina 3 3,2
4.5 Asociación entre la detección de los genes de SAgs y la resistencia antimicrobiana. En las Tabla 11 a 16, se observa las diferentes asociaciones entre los genes de superantígenos y la susceptibilidad antimicrobiana. En la Tabla 11 se muestra la asociación entre presencia o ausencia de SAgs en los aislamientos β–Lactamasa+, donde la frecuencia de asociación fue mayor para un solo gen con 32,6% (31/95) seguido por la asociación de dos genes con 23,2% (22/95). No se detectó ningún gen de SAg en 23,2% (22/95) de los aislamientos productores de β-Lactamasa. También se muestra la asociación de 6 aislamientos β-Lactamasa negativos con los SAgs, donde el 3,2% (3/95) fueron negativos para 1 y 2 SAgs. Tabla 11. Asociación entre uno o más genes de SAgs y la prueba de β-Lactamasa positiva (β -L+) y la prueba de β-Lactamasa negativa (β -L-).
β -L+ No. Genes No. Resistentes %
89 0 22 23,2
1 31 32,6
2 22 23,2
3 10 10,5
4 4 4,2
β -L- 0 3 3,1
6 1 2 2,1
2 1 1,1
95 100
La tabla 12, muestra que el 6,3% (6/95) de los aislamientos fueron simultáneamente resistentes para cefoxitina, oxacilina y β-Lactamasa+; y 3,2% (3/6) de los aislamientos resistentes no expresaron genes de SAgs. Anexo L.
Tabla 12. Asociación entre uno o más genes de SAgs y la susceptibilidad a cefoxitina-oxacilina.

46
Cefoxitina-Oxacilina/ No. Genes R % S %
0 3 3,2 22 23,2
1 1 1,05 32 33,7
2 1 1,05 22 23,2
3 0 0,0 10 10,5
4 1 1,05 3 3,2
Total 6 6,3 89 93,7
La tabla 13 muestra que la resistencia a eritromicina se asoció con uno o más superantígenos en 4,2% (4/95); la resistencia a clindamicina se asoció con un 3,2% (3/95); y la resistencia a tetraciclina se asoció con un 11,6% (11/95).
Tabla 13. Asociación entre uno o más genes de SAgs y la susceptibilidad a eritromicina, clindamicina y tetraciclina.
Antimicrobiano No. Genes
R % S %
Eritromicina
0 0 0,0 25 26,3
1 2 2,1 31 32,6
2 0 0,0 23 24,2
3 2 2,1 8 8,4
4 0 0,0 4 4,2
Total 4 4,2 91 95,8
Clindamicina
0 0 0,0 25 26,3
1 1 1,1 32 33,7
2 0 0,0 23 24,2
3 2 2,1 8 8,4
4 0 0,0 4 4,2
Total 3 3,2 92 96,8
Tetraciclina
0 0 0,0 25 26,3
1 4 4,2 29 30,5
2 4 4,2 19 20,0
3 0 0,0 10 10,5
4 3 3,2 1 1,1
Total 11 11,6 84 88,4
En general, la asociación de la detección de los SAgs y la resistencia a cada uno de los antimicrobianos (producción de β-Lactamasa, cefoxitina/ oxacilina, eritromicina, clindamicina) no mostraron diferencias estadísticamente significativas. La única asociación que mostró diferencia significativa fue para la tetraciclina. Ver Tablas 14 a 18. Anexo L.

47
Tabla 14. Asociación entre la presencia o ausencia SAgs y la prueba de la β-Lactamasa.
B-L SAg+ SAg- Total
S 3 3 6
R 67 22 89
Total 70 25 95
p=0,1844
Tabla 15. Asociación entre la presencia o ausencia SAgs y la susceptibilidad a cefoxitina/ oxacilina.
Cefoxitina/ Oxacilina SAg+ SAg- Total
S 67 22 89
R 3 3 6
Total 70 25 95
p= 0,1844 Tabla 16. Asociación entre la presencia o ausencia SAgs y la susceptibilidad a eritromicina.
Eritromicina SAg+ SAg- Total
S 66 25 91
R 4 0 4
Total 70 25 95
p= 0,2880 Tabla 17. Asociación entre la presencia o ausencia SAgs y la susceptibilidad a clindamicina.
Clindamicina SAg+ SAg- Total
S 67 25 92
R 3 0 3
Total 70 25 95
p= 0,3954 Tablas 18. Asociación entre la presencia o ausencia SAgs y la susceptibilidad a tetraciclina.
Tetraciclina SAg+ SAg- Total
S 59 25 84
R 11 0 11
Total 70 25 95
p= 0,027
5. DISCUSIÓN El estado portador de Staphylococcus aureus en individuos sanos, juega un papel importante en la epidemiología y patogenicidad de la infección (Fosch et al., 2012),

48
siendo el ser humano uno de los reservorios naturales de esta bacteria. Se ha informado que el S. aureus puede colonizar cualquier parte del cuerpo, sin embargo, la mayor porción y la más frecuente son las fosas nasales con un rango del 20 al 40% en la población general (Kuehnert et al., 2006, Seybold et al., 2011). En la población estudiada de las madres comunitarias del ICBF de Pereira, resultaron portadoras de S. aureus en un 22% ( fosas nasales y/o manos), lo que correlaciona con lo publicado por diferentes autores en individuos sanos, donde establecen un rango entre el 20 al 40% (Cespedes et al., 2002, Lu et al., 2005, Kuehnert et al., 2006, Wattinger et al., 2012), representando un factor de riesgo en la diseminación de la bacteria y por ende puede causar intoxicaciones alimentarias por manipulación de alimentos u otro tipo de infecciones. El estudio reveló que el 17% eran portadoras en fosas nasales, este porcentaje es acorde con los resultados obtenidos por otros autores (Cespedes et al., 2002, Lu et al., 2005, Kuehnert et al., 2006, Wattinger et al., 2012); en estudios realizados en manipuladores de alimentos de muestras tomadas solo de fosas nasales reportan un rango de 18,8% al 43,5% (Figueroa et al., 2002, Acco et al., 2003, Mertz et al., 2007, Jorda et al., 2012, Wattinger et al., 2012), también en el estudio se obtuvo un bajo porcentaje de portadores en los pulpejos y uñas de las manos, en un 5%. Adicionalmente, el estudio mostró que el 1,5% del total de madres comunitarias eran portadoras simultáneas en fosas nasales como en manos, no se encontraron estudios donde hubiera datos comparativos de colonización simultánea de fosas y manos. La prevalencia global de la detección de los superantígenos en el estudio fue 73,7%, estos resultados se aproximan a los de un estudio realizado en personal de restaurantes de Kuwait con una prevalencia 71,0% obtenida de cepas de fosas nasales y manos aisladas de manipuladores de alimentos (Udo et al., 2009), y más baja que la prevalencia del 86,6% detectada en manipuladores de alimentos en los restaurantes de Kuwait en cepas de origen nasal (Al-Bustan et al., 1996) y similar a la prevalencia de 74,1% obtenido a partir de aislados de S. aureus de intoxicación alimentaria en Taiwán (Chiang et al., 2006). En estudios realizados en Latinoamérica la detección de superantígenos en portadores de S. aureus es superior al 50% (Figueroa et al., 2002, Mamprim et al., 2006, Jorda et al., 2012). El gen etd con 47,4 % tuvo la mayor frecuencia de todos los genes estudiados, pero no se encontraron reportes en aislamientos de S. aureus en portadores sanos, lo que arroja un importante hallazgo en este estudio. Se ha planteado que el gen etd puede interrumpir la barrera cutánea epitelial y de esta manera contribuir a la propagación bacteriana (Ladhani, 2003). Este gen ha sido reportado en aislamientos clínicos, con una frecuencia del 10,5% (Yamasaki et al., 2006).

49
Los genes tsst-1 y sea, siguieron en frecuencia con 23,2 % y 18,9 % respectivamente; estos resultados están de acuerdo con los hallazgos previos en individuos sanos portadores de S. aureus productores de las toxinas. Mehrotra y otros autores obtuvieron datos similares, el gen tsst-1 24,3% y el gen sea 19,6% (Chance, 1996, Mehrotra et al., 2000, Yamasaki et al., 2006, Aydin et al., 2011). Para el caso del gen tsst-1 es más frecuentemente reportado en aislamientos clínicos por su participación en el síndrome del choque tóxico, sin embargo, este gen también ha sido detectado en diferentes aislamientos de productos alimenticios contaminados por S. aureus. En Corea, en aislamientos de alimentos reportaron el gen en un 13,5% (Oh et al., 2007), 4,4% en manipuladores de quesos de cabra (Rapini et al., 2005), 12% en 1047 muestras de alimentos (Dallal et al. 2010). El gen tsst-1 fue detectado en leche de vacas con mastitis clínica en 58,1%, la subclínica 76,7% y en un tanque con leche 75,4% (Takeuchi et al., 1998) y en cambio se encontró en leche 47,4% de tsst-1 (Valle et al., 1991). Lo anterior indica que la distribución del gen es universal, se puede encontrar tanto en aislamientos clínicos, de alimentos y en portadores. El 23,2% de la detección de este gen en el estudio es superior a lo reportado por otros autores (Argudin et al., 2010, Sospedra et al., 2012) y casi igual a Mehrotra que obtuvo 24,3%. La prevalencia de gen de la enterotoxina sea 18,9 %, concuerda con lo reportado en la literatura. El gen sea es codificado por una familia de bacteriófagos atemperados cuyos genomas se incorporan y se replican con el cromosoma bacteriano de S. aureus. Sin embargo, su detección dependerá de la distribución de estos fagos, se ha demostrado que la distribución de los fagos es irregular (Le Loir et al., 2003, Casagrande Proietti et al., 2010). En este estudio se detectaron los genes de las toxinas exfoliativas. El gen eta es un factor de virulencia asociado al impétigo bulloso (Dagan, 1991, Nishi et al., 2002, Koning et al., 2003, Cha et al., 2006, Pragman et al., 2004) y fue detectado en un 4,2%, está dentro del rango descrito por otros autores 3% al 6 % (Peacock et al., 2002). Uno de los propósitos del estudio fue establecer las posibles asociaciones entre los genes de superantígenos para determinar los genotipos prevalentes. Así, el gen sea apareció principalmente asociado a tsst-1 y etd, con una prevalencia del 10% para sea, tsst-1 y el 8,6% sea, etd, tsst-1, demostrando que este gen es dominante en estas asociaciones y se puede asociar con cualquier tipo de genes de superantígenos y con diferentes orígenes de las cepas (Seo, 2010). Estas asociaciones son variantes aún no descritas, que pueden influir en la patogenicidad y se necesita hacer estudios complementarios para establecer su verdadero impacto. La información sobre asociaciones de enterotoxinas clásicas con los genes del síndrome de shock tóxico y toxinas exfoliativa, son escasos (Munson et al., 1998, Booth et al., 2001, Fitzgerald et al., 2001b, Jarraud et al., 2002, Musser et al., 1990); el estudio arrojó diferentes combinaciones y no se encontró información para dichas

50
asociaciones de genes. La importancia de este gen sea se ha demostrado en diferentes estudios que lo reportan como uno de los factores importantes en la intoxicación alimentaria (Argudin et al., 2010). El análisis de las asociaciones entre los genes de los superantígenos demostró heterogeneidad entre los perfiles de los genes, de hecho fue posible agruparlos en 8 combinaciones diferentes para dos o más genes, a diferencia de Morandi que obtuvo un número mayor de combinaciones, que pueden depender del número de aislamientos analizados 122 y determinó que los genes predominantes fueron sea y sed, concluyendo que los nuevos genes enterotoxigénicos (seg, seh, sei, sej y sel) se asocian a menudo con genes de clásicos (Morandi et al., 2009). En el análisis de la susceptibilidad a los antimicrobianos probados, se pudo evidenciar que 74,7% de los aislamientos mostraron resistencia a un antibióticos y el 3,2% fueron resistentes a cuatro antibióticos, lo cual concuerda con los reportes de Texeira y col (Teixeira et al., 1995). El S. aureus es una bacteria productora de β–Lactamasa, y teniendo en cuenta esta condición el 93,7% fueron positivas para β–Lactamasa, concordando con Seo que reporto 96%; la multiresistencia detectada de 3,2% a cuatro antibióticos difiere de una forma marcada a los datos obtenidos por Seo del 56% (Seo et al., 2010). La resistencia antimicrobiana de los aislamientos, sugiere que el estado de portador es una fuente de diseminación de cepas resistentes a antibióticos en la comunidad. Los diferentes estudios de resistencia antimicrobiana de S. aureus reportan que más del 90% de las cepas resultan resistentes a la penicilina y en diferentes tasas para otros antimicrobianos (Macfaddin, 1990). Los resultados para los 13 antimicrobianos estudiados indicaron que la mayor resistencia fue para tetraciclina con 11.6% cercano al 16% reportado por Morales (Morales et al, 2006). La tetraciclina es uno de los antibióticos más utilizado en animales (Sykes et al, 1976), para el caso humano estas cepas resistentes a tetraciclina se pueden transferir muy fácilmente a los alimentos sobre todo por los manipuladores de alimentos, dependiente del tipo de alimentos contaminados. En S. aureus, se ha reportado que los genes tetK y tetL intervienen en los mecanismos de resistencia a tetraciclinas en aislados de humanos y bovinos, los cuales, residen en plásmidos pequeños de la familia pT181 (Rivera et al., 2011). Por convención, la resistencia in vitro a oxacilina y/o cefoxitina, se asocia con cepas resistentes a meticilina (SARM: S. aureus resistente a meticilina). El sistema VITEK 2C utilizado en el estudio, detectó el 6,3% de SARM. Los 6 aislamientos fueron simultáneamente resistentes para cefoxitina, oxacilina y productores de β-Lactamasa, por ello, las personas portadoras de estos aislamientos se convierten en diseminadoras de bacterias multiresistentes. Adicionalmente, 3 de los 6 aislamientos resistentes expresaron genes de SAgs. Estos hallazgos son novedosos y no se encontraron artículos que referencien estos resultados.

51
Al determinar la relación entre asociaciones SAgs y resistencia antimicrobiana, no fue posible encontrar en la literatura revisada datos de dichas asociaciones. La asociación de los SAgs y el resultado positivo o negativo de la prueba de la β–Lactamasa no mostró diferencia significativa. Las asociaciones de la resistencia a eritromicina, clindamicina y tetraciclina con SAgs fueron bajas. La única diferencia estadísticamente significativa fue para la resistencia a tetraciclina, lo cual indica que este estudio se convierte en uno de los primeros en buscar una posible asociación entre SAgs y resistencia antimicrobiana en aislamientos obtenidos de portadores nasales y/o de manos. El análisis estadístico entre la asociación de presencia o ausencia de SAgs y la resistencia antibacteriana no mostraron diferencia estadísticamente significativa, aunque hay que resaltar que la resistencia a tetraciclina y la asociación a superantígenos si lo fue, esta significancia implicaría un factor de riesgo en el estado de portador. Por la multiresistencia que han adquirido los diferentes microrganismos, es muy complicada la elección de los antibióticos para tratar cualquier tipo de infección; diferentes autores reportan que las tetraciclinas son el tratamiento oral más rentable y eficiente (Jamali et al., 2015, Enoch et al., 2009). Siendo de infinita preocupación que el S. aureus ya está comenzando adquirir resistencia a este antibiótico (Jamali et al., 2015), como lo indican los resultados del estudio.
6. CONCLUSIONES. El estado de portador es un factor de riesgo para infecciones adquiridas en la comunidad, los datos obtenidos fueron similares a los de la población general,

52
representando un factor de riesgo debido a que las madres comunitarias por contacto manual o mediante secreciones respiratorias pueden contaminar los alimentos que preparan para los niños que tiene a su cuidado y por ende producir intoxicaciones alimentarias.
La intoxicación alimentaria estafilocócica es un importante problema de salud pública a nivel mundial, ya que muchos manipuladores de alimentos pueden ser portadores de S. aureus en fosas nasales o en manos; como se observó en el estudio, el mayor porcentaje fue en fosas nasales, considerándose como una de las principales fuentes de contaminación de alimentos. La normativa nacional (protocolo de vigilancia y control de ETA 2010 y 2014; ETA, enfermedades transmitidas por alimentos) específica la obligatoriedad de notificación, la metodología de diagnóstico de la intoxicación alimentaria, pero no del estado de portador de S. aureus. Esto puede generar que no se tengan datos acertados epidemiológicamente de dicha enfermedad.
Constantemente, las empresas procesadoras de alimentos experimentan problemas asociados con el elevado número de contaminación a causa de este microorganismo, el cual puede generar gastroenteritis en los consumidores de estos alimentos originados por la producción de enterotoxinas termoestables y la PCR Múltiple puede ser alternativa útil en la identificación de aislamientos enterotoxigenicos. La prevalencia de los superantígenos en los aislamientos fue alta, indicando que S. aureus posee una gran variación genética, tal vez por transferencia horizontal de elementos genéticos móviles que codifican los genes de virulencia; este evento también se ha encontrado en aislamientos hospitalarios, en alimentos y en manipuladores de alimentos sobretodo en cepas de origen nasal. El gen de mayor prevalencia fue etd, mostrándonos un hallazgo novedoso en aislamiento de portadores nasales o manos, a diferencia de los otros genes que han sido reportados con frecuencia en otros estudios, dicho gen puede interrumpir la barrera cutánea epitelial y de esta manera contribuir a la propagación bacteriana. El proyecto permitió hacer un estudio epidemiológico molecular dirigido a conocer la virulencia de los aislamientos obtenidos de madres comunitarias, encontrando a sea, etd y tsst-1 como los genes más frecuentes. Este estudio de epidemiología molecular mostró información determinante de la presencia en diferentes aislamientos de S. aureus que contienen resistencia antimicrobiana y virulencia por la detección de los superantígenos. Estos aislamientos codificaron no sólo enterotoxinas clásicas, sino también toxinas exfoliativas y TSST-1.

53
Las diferentes asociaciones mostraron dos o más genes y fueron similares a las reportadas por otros investigadores, demostrando que el gen sea es dominante y se puede asociar con cualquier tipo de genes de superantígenos independiente del origen de los aislamientos, sin embargo, algunas asociaciones encontradas en este estudio son variantes aun no descritas o reportadas que pueden influir en la patogenicidad de S. aureus. El análisis de las asociaciones entre los genes de los superantígenos demuestra la heterogeneidad genética siendo los genotipos sea, tsst-1 y sea etd los más prevalentes. La producción de la enzima β–Lactamasa por S. aureus fue alta y concuerda con lo reportado por otros investigadores descartando la posibilidad de utilizar beta lactámicos sensibles a β–Lactamasa, en tratamiento. La resistencia de los aislamientos de fosas nasales o manos, fue alta para uno o más antimicrobianos donde la resistencia a un antimicrobiano fue la de mayor frecuencia y la más baja a 4 antibióticos y estas cepas pueden ser transmitidas a los seres humanos por el consumo de alimentos que posean dichas bacterias resistentes o multirresistentes, mostrando alto riesgo para los consumidores, a falta de higiene y medidas preventivas para evitar la presencia de S. aureus y la producción de superantígenos en los alimentos, haciendo hincapié en la necesidad de mejorar las prácticas de inocuidad en la elaboración de alimentos, así como durante la distribución y el consumo de los productos terminados alimenticios. La susceptibilidad para los trece antimicrobianos, arrojó que la tetraciclina fue el que tuvo mayor resistencia seguido de cefoxitina u oxacilima. La tetraciclina es uno de los antibióticos más utilizados en animales; el uso inadecuado de tetraciclina para tratar enfermedades infecciosas en animales, empleando dosis subterapéuticas como profilaxis o promotores de crecimiento en los animales, han creando cepas resistentes a la tetraciclina, que se pueden transferir muy fácilmente a los alimentos y pasan al hombre por vía directa o por contacto con los manipuladores de alimentos. A pesar de que el número de aislamientos SARM fue bajo, esto debe considerarse como un índice de riesgo de diseminación de S. aureus multiresistente en la comunidad, estos hallazgos son novedosos y se requiere estudios adicionales para evaluar su impacto. Las diferentes asociaciones entre SAgs y la resistencia antimicrobiana, no mostraron diferencia significativas para la producción de β–Lactamasa, eritromicina y clindamicina; en cambio para la tetraciclina si mostró diferencia significativa y se debe tener cuidado porque por el abuso de este antimicrobiano en los animales hace posible la transferencia de esta bacteria a los seres humanos. Los alimentos son un factor importante para la transferencia de resistencias a antibióticos. Dicha transferencia puede ocurrir por medio de residuos de antibióticos

54
en alimentos, a través de la transferencia de patógenos por los alimentos resistentes o a través de la ingestión de cepas resistentes del alimento original. Las cepas de S. aureus son con frecuencia resistentes a la terapia de antibióticos debido a su capacidad para producir una barrera de exopolisacáridos y a su ubicación dentro de microabscesos, que limitan la acción de los medicamentos. La metodología molecular utilizada en el presente estudio, está habilitada para ser implementada en cualquier laboratorio clínico o de alimentos, estableciendo de forma rápida si los aislamientos de S. aureus son enterotoxigénicos.
7. RECOMENDACIONES
Complementar el estudio de la detección de los superantígenos con la expresión de los mismos para establecer si los genes detectados producen efectivamente sus correspondientes las proteínas y, buscar la relación con la virulencia,

55
Continuar estudios de epidemiología molecular en diferentes poblaciones de portadores, para caracterizar a nivel de aislamientos enterotóxicos los principales genes de enterotoxinas y otros superantígenos para establecer el verdadero papel en la virulencia de la bacteria. Para ello deberá determinarse la expresión de los superantigenos in vitro. Buscar posibles relaciones epidemiológicas entre el estado de portador y la producción de intoxicación alimentaria, mediante técnicas moleculares de detección de genes y sus productos. En hogares y establecimientos educativos hay mayor número de intoxicaciones alimentarias; sugiriendo que hay deficiencias en la infraestructura higiénico-sanitaria y capacitación en Buenas Prácticas de Manufactura, en hogares y establecimientos educativos del país, por eso es de importancia estudiar el estado del portador de S. aureus en poblaciones que de alguna manera tienen que ver con la manipulación y procesamiento de alimentos.
Implementar campañas permanentes de educación de las madres comunitarias en la manipulación y preparación de los alimentos con el fin de evitar la contaminación del alimento y así suministrar un alimento inocuo para los niños. Se debería tener una normativa nacional más rigurosa con la notificación de intoxicaciones alimentaria por S. aureus y se podrían implementar técnicas moleculares para rápido y fácil diagnóstico.
El uso de antibióticos como promotores del crecimiento en la cría de animales, debe ser revisado porque genera resistencia en los microrganismos que contamina los alimentos. Estudios futuros debería establecer una relación entre los genes que codifican las enterotoxinas clásicas – no y la producción de la enfermedad o la intoxicación alimentaria.
8. BIBLIOGRAFÍA.
Abe, J., Ito, Y., Onimaru, M., Kohsaka, T. & Takeda, T. (2000). Characterization and distribution of a new enterotoxin-related superantigen produced by Staphylococcus aureus. Microbiol Immunol, 44, 79-88.

56
Abrahmsen, L., Dohlsten, M., Segren, S., Bjork, P., Jonsson, E. & Kalland, T. (1995). Characterization of two distinct MHC class II binding sites in the superantigen staphylococcal enterotoxin A. Embo J, 14, 2978-86.
Acco M., Ferreira F, Henriques J &Tondo E. (2003). Identification of multiple strains of Staphylococcus aureus colonizing nasal mucosa of food handlers. Food Microbiol, 20, 489-93.
Agrawal, R., Singh, P. K., Sharma, S. K., Kamboj, D. V., Goel, A. K. & Singh, L. (2012). Highly Expressed Recombinant SEB for Antibody Production and Development of Immunodetection System. Indian J Microbiol, 52, 191-6.
Al-Bustan M. A., Udo E. E & Chugh T. D. (1996). Nasal carriage of enterotoxinproducing Staphylococcus aureus among restaurant Workers in Kuwait City. Epidemiol Infect, 116, 319-322.
Argudin, M. A., Mendoza, M. C. & Rodicio, M. R. (2010). Food poisoning and Staphylococcus aureus enterotoxins. Toxins (Basel), 2, 1751-73.
Arvidson, S. & Tegmark, K. 2001. Regulation of virulence determinants in Staphylococcus aureus. Int J Med Microbiol, 291, 159-70.
Atilano, M. L., Pereira, P. M., Yates, J., Reed, P., Veiga, H., Pinho, M. G. & Filipe, S. R. (2010). Teichoic acids are temporal and spatial regulators of peptidoglycan cross-linking in Staphylococcus aureus. Proceedings of the National Academy of Sciences, 107, 18991-18996.
Ausubel, F.M., Brent, R., Kingston, R.E., Moore, D.D., Seidman, J.G., Smith, J.A. & Struhl, K. (1987). Current Protocols in Molecular Biology, Vol. 1, New York: John Wiley and Sons.
Aydin, A., Sudagidan, M. & Muratoglu, K. (2011). Prevalence of staphylococcal enterotoxins, toxin genes and genetic-relatedness of foodborne Staphylococcus aureus strains isolated in the Marmara Region of Turkey. Int J Food Microbiol, 148, 99-106.
Baba, T., Takeuchi, F., Kuroda, M., Yuzawa, H., Aoki, K., Oguchi, A., Nagai, Y., Iwama, N., Asano, K., Naimi, T., Kuroda, H., Cui, L., Yamamoto, K. & Hiramatsu, K. (2002). Genome and virulence determinants of high virulence community-acquired MRSA. Lancet, 359, 1819-27.
Balaban, N. & Rasooly, A. (2000). Staphylococcal enterotoxins. Int J Food Microbiol, 61, 1-10.
Barrett, S. P. (1993). Australian MRSA. J Hosp Infect, 25, 218-9. Barrett, S. P., Teare, E. L. & Sage, R. (1993). Methicillin resistant Staphylococcus
aureus in three adjacent health districts of south-east England 1986-91. J Hosp Infect, 24, 313-25.
Bayles, K. W. & Landolo, J. J. (1989). Genetic and molecular analyses of the gene encoding staphylococcal enterotoxin D. J Bacteriol, 171, 4799-806.
Ben Nejma, M., Mastouri, M., Frih, S., Sakly, N., Ben Salem, Y. & Nour, M. (2006). Molecular characterization of methicillin-resistant Staphylococcus aureus isolated in Tunisia. Diagn Microbiol Infect Dis, 55, 21-6.
Bergdoll, M. S., Borja, C. R. & Avena, R. M. (1965). Identification of a new enterotoxin as enterotoxin C. J Bacteriol, 90, 1481-5.

57
Bergdoll, M. S., Crass, B. A., Reiser, R. F., Robbins, R. N. & Davis, J. P. (1981). A new staphylococcal enterotoxin, enterotoxin F, associated with toxic-shock-syndrome Staphylococcus aureus isolates. Lancet, 1, 1017-21.
Betley, M. J., Borst, D. W. & Regassa, L. B. (1992). Staphylococcal enterotoxins, toxic shock syndrome toxin and streptococcal pyrogenic exotoxins: a comparative study of their molecular biology. Chem Immunol, 55, 1-35.
Betley, M. J. & Mekalanos, J. J. 1(985) Staphylococcal enterotoxin A is encoded by phage. Science, 229, 185-7.
Blaiotta, G., Fusco, V., Ercolini, D., Pepe, O. & Coppola, S. (2010). Diversity of Staphylococcus species strains based on partial kat (catalase) gene sequences and design of a PCR-restriction fragment length polymorphism assay for identification and differentiation of coagulase-positive species (S. Aureus, S. Delphini, S. Hyicus, S. Intermedius, S. Pseudintermedius, and S. Schleiferi subsp. Coagulans). J Clin Microbiol, 48, 192-201.
Blomster-Hautamaa, D. A., Kreiswirth, B. N., Novick, R. P. & Schlievert, P. M. (1986). Resolution of highly purified toxic-shock syndrome toxin 1 into two distinct proteins by isoelectric focusing. Biochemistry, 25, 54-9.
Boerema, J. A., Clemens, R. & Brightwell, G. (2006). Evaluation of molecular methods to determine enterotoxigenic status and molecular genotype of bovine, ovine, human and food isolates of Staphylococcus aureus. Int J Food Microbiol, 107, 192-201.
Booth, M. C., Pence, L. M., Mahasreshti, P., Callegan, M. C. & Gilmore, M. S. (2001). Clonal associations among Staphylococcus aureus isolates from various sites of infection. Infect Immun, 69, 345-52.
Boyce J, M. (1997). Epidemiology and prevention of nosocomial infections. In: Crossley KB, Archer GL. The staphylococci in human disease. New York, NY, Churchill Livingstone, 309–29.
Brizzio, A. A., Tedeschi, F. A. & Zalazar, F. E. (2011). Description of a staphylococcal alimentary poisoning outbreak in Las Rosas, Santa Fe Province, Argentina. Rev Argent Microbiol, 43, 28-32.
Bronner, S., Monteil, H. & Prevost, G. (2004). Regulation of virulence determinants in Staphylococcus aureus: complexity and applications. Fems Microbiol Rev, 28, 183-200.
Brouillard, J. N., Gunther, S., Varma, A. K., Gryski, I., Herfst, C. A., Rahman, A. K., Leung, D. Y., Schlievert, P. M., Madrenas, J., Sundberg, E. J. & Mccormick, J. K. (2007). Crystal structure of the streptococcal superantigen spei and functional role of a novel loop domain in T cell activation by group V superantigens. J Mol Biol, 367, 925-34.
Brown, E. L., Nishiyama, Y., Dunkle, J. W., Aggarwal, S., Planque, S., Watanabe,
K., Csencsits-Smith, K., Bowden, M. G., Kaplan, S. L. & Paul, S. (2012). Constitutive production of catalytic antibodies to a Staphylococcus aureus virulence factor and effect of infection. J Biol Chem, 287, 9940-51.
Bukowski, M., Wladyka, B. & Dubin, G. (2010). Exfoliative Toxins of Staphylococcus aureus. Toxins (Basel), 2, 1148-65.

58
Burnside, K., Lembo, A., De Los Reyes, M., Iliuk, A., Binhtran, N. T., Connelly, J. E., Lin, W. J., Schmidt, B. Z., Richardson, A. R., Fang, F. C., Tao, W. A. & Rajagopal, L. (2010). Regulation of hemolysin expression and virulence of Staphylococcus aureus by a serine/threonine kinase and phosphatase. Plos One, 5, e11071.
Capparelli, R., Nocerino, N., Medaglia, C., Blaiotta, G., Bonelli, P. & Iannelli, D. (2011). The Staphylococcus aureus peptidoglycan protects mice against the pathogen and eradicates experimentally induced infection. Plos One, 6, e28377.
Casagrande Proietti, P., Coppola, G., Bietta, A., Luisa Marenzoni, M., Hyatt, D. R., Coletti, M. & Passamonti, F. (2010). Characterization of genes encoding virulence determinants and toxins in Staphylococcus aureus from bovine milk in Central Italy. J Vet Med Sci, 72, 1443-8.
Casman, E. P. (1960). Further serological studies of staphylococcal enterotoxin. J Bacteriol, 79, 849-56.
Casman, E. P. (1965). Staphylococcal enterotoxin. Ann N Y Acad Sci, 128, 124-31. Casman, E. P., Bergdoll, M. S. & Robinson, J. (1963). Designation of
Staphylococcal Exterotoxins. J Bacteriol, 85, 715-6. Cespedes, C., Miller, M., Quagliarello, B., Vavagiakis, P., Klein, R. S. & Lowy, F. D.
(2002). Differences between Staphylococcus aureus isolates from medical and nonmedical hospital personnel. J Clin Microbiol, 40, 2594-7.
Cha, J. O., Lee, J. K., Jung, Y. H., Yoo, J. I., Park, Y. K., Kim, B. S. & Lee, Y. S. (2006). Molecular analysis of Staphylococcus aureus isolates associated with staphylococcal food poisoning in South Korea. J Appl Microbiol, 101, 864-71.
Chance, T. D. (1996). Toxic shock syndrome: role of the environment, the host and the microorganism. Br J Biomed Sci, 53, 284-9.
Cheung AL, Koomeyy JM, Butler CA, Projan SJ, Fischetti VA. (1992). Regulation of Exoprotein expression in Staphylococcus aureus by a locus (sar) distinct from agr. Proc. Natl. Acad. Sci. USA, 89, 6462-6466.
Cheung, A. L., Bayer, A. S., Zhang, G., Gresham, H. & Xiong, Y. Q. (2004). Regulation of virulence determinants in vitro and in vivo in Staphylococcus aureus. Fems Immunol Med Microbiol, 40, 1-9.
Cheung, A. L., Nishina, K. A., Trotonda, M. P. & Tamber, S. (2008). The sara protein family of Staphylococcus aureus. Int J Biochem Cell Biol, 40, 355-61.
Chevalier, C., Boisset, S., Romilly, C., Masquida, B., Fechter, P., Geissmann, T.,
Vandenesch, F. & Romby, P. (2010). Staphylococcus aureus RNAIII binds to two distant regions of coa mrna to arrest translation and promote mrna degradation. Plos Pathog, 6, e1000809.
Chiang YC, Lin CW, Yang CY, Tsen HY. (2006). PCR primers for the detection of

59
staphylococcal enterotoxin K, L, and M and Survey of staphylococcal enterotoxin types in Staphylococcus aureus isolates from food poisoning cases in Taiwan. J Food Prot 2006, 69,1072-1079.
Couch, J. L., Soltis, M. T. & Betley, M. J. (1988). Cloning and nucleotide sequence of the type E staphylococcal enterotoxin gene. J Bacteriol, 170, 2954-60.
Corredor L. F. Genotipificación de superantígenos de toxinas pirogénicas y toxinas exfoliativas en aislamientos de Staphylococcus aureus obtenidos en el Hospital Universitario San Jorge de Pereira (Fase I). Tesis. Universidad Tecnológica de Pereira. Facultad de Medicina. Pereira, 2012.
Dagan, R. (1991). Staphylococcus aureus in impetigo. Am J Dis Child, 145, 1223. Dallal, MMS, Salehipour, Z., Eshraghi, S., Mehrabadi, JF, Bakhtiari, R., (2010).
Occurrence and molecular characterization of Staphylococcus aureus strains isolated from meat and dairy products by PCR RFLP.Ann.Microbiol.60, 189-196.
Davis, J. P., Chesney, P. J., Wand, P. J. & Laventure, M. (1980) Toxic-shock syndrome: epidemiologic features, recurrence, risk factors, and prevention. N Engl J Med, 303, 1429-35.
Day, N. P. (2012). Panton-Valentine leucocidin and staphylococcal disease. Lancet Infect Dis.
De Boer, M. L. & Chow, A. W. (1994). Toxic shock syndrome toxin 1-producing Staphylococcus aureus isolates contain the staphylococcal enterotoxin B genetic element but do not express staphylococcal enterotoxin B. J Infect Dis, 170, 818-27.
Dinges, M. M., Orwin, P. M. & Schlievert, P. M. (2000). Exotoxins of Staphylococcus aureus. Clin Microbiol Rev, 13, 16-34, table of contents.
El-Huneidi, W., Bdour, S. & Mahasneh, A. (2006). Detection of enterotoxin genes seg, seh, sei, and sej and of a novel aroa genotype in Jordanian clinical isolates of Staphylococcus aureus. Diagn Microbiol Infect Dis, 56, 127-32.
Endo, Y., Yamada, T., Matsunaga, K., Hayakawa, Y., Kaidoh, T. & Takeuchi, S. (2003). Phage conversion of exfoliative toxin A in Staphylococcus aureus isolated from cows with mastitis. Vet Microbiol, 96, 81-90.
Enoch, D. A., Karas, J. A. & Aliyu, S. H. (2009). Oral antimicrobial options for the treatment of skin and soft-tissue infections caused by methicillin-resistant Staphylococcus aureus (MRSA) in the UK. Int J Antimicrob Agents, 33, 497-502.
Escartin F. E. (2000). Microbiología e inocuidad de los alimentos. Universidad Autónoma de Querétaro, 322.
Fields, B. A., Malchiodi, E. L., Li, H., Ysern, X., Stauffacher, C. V., Schlievert, P. M., Karjalainen, K. & Mariuzza, R. A. (1996). Crystal structure of a T-cell receptor beta-chain complexed with a superantigen. Nature, 384, 188-92.
Figueroa G, Navarrete P, Caro M, Troncoso M, Faúndez G. (2002). Portación de Staphylococcus aureus enterotoxigénicos en manipuladores de alimentos. Rev Med Chile, 130:85964.
Fitzgerald, J. R., Monday, S. R., Foster, T. J., Bohach, G. A., Hartigan, P. J., Meaney, W. J. & Smyth, C. J. (2001a). Characterization of a putative

60
pathogenicity island from bovine Staphylococcus aureus encoding multiple superantigens. J Bacteriol, 183, 63-70.
Fitzgerald, J. R., Sturdevant, D. E., Mackie, S. M., Gill, S. R. & Musser, J. M. (2001b). Evolutionary genomics of Staphylococcus aureus: insights into the origin of methicillin-resistant strains and the toxic shock syndrome epidemic. Proc Natl Acad Sci U S A, 98, 8821-6.
Fosch S., Yones C., Trossero M., Grosso O & Nepote A. (2012). Portación nasal de Staphylococcus aureus en individuos de la comunidad: factores epidemiológicos. Acta bioquím. clín. latinoam, 46, 1, 59-68.
Fraser, J. D. & Proft, T. (2008). The bacterial superantigen and superantigen-like proteins. Immunol Rev, 225, 226-43.
Fujimoto, D. F., Higginbotham, R. H., Sterba, K. M., Maleki, S. J., Segall, A. M., Smeltzer, M. S. & Hurlburt, B. K. (2009). Staphylococcus aureus sara is a regulatory protein responsive to redox and ph that can support bacteriophage lambda integrase-mediated excision/recombination. Mol Microbiol, 74, 1445-58.
Gil M, Monica. (2000). Staphylococcus aureus Microbiology and molecular aspects of methicillin resistance. Chilean Journal of Infectious Diseases, 17, 2, 145-152.
Gomez-Lucia, E., Goyache, J., Orden, J. A., Domenech, A., Javier Hernandez, F., Ruiz-Santa Quiteria, J. A., Lopez, B., Blanco, J. L. & Suarez, G. (1992). Growth of Staphylococcus aureus and synthesis of enterotoxin during ripening of experimental Manchego-type cheese. J Dairy Sci, 75, 19-26.
Gomez, M. I., O'seaghdha, M., Magargee, M., Foster, T. J. & Prince, A. S. (2006). Staphylococcus aureus protein A activates TNFR1 signaling through conserved igg binding domains. J Biol Chem, 281, 20190-6.
Gorwitz, R. J., Kruszon-Moran, D., Mcallister, S. K., Mcquillan, G., Mcdougal, L. K., Fosheim, G. E., Jensen, B. J., Killgore, G., Tenover, F. C. & Kuehnert, M. J. (2008). Changes in the prevalence of nasal colonization with Staphylococcus aureus in the United States, 2001-2004. J Infect Dis, 197, 1226-34.
Gündogan, N.; Citak, S.; Turan, E. 2006 Slime production, dnase activity and antibiotic resistance of Staphylococcus aureus isolated from raw milk, milk and ice cream samples. Food Control, 17, 389–392.
Gunther, S., Varma, A. K., Moza, B., Kasper, K. J., Wyatt, A. W., Zhu, P., Rahman, A. K., Li, Y., Mariuzza, R. A., Mccormick, J. K. & Sundberg, E. J. (2007). A novel loop domain in superantigens extends their T cell receptor recognition site. J Mol Biol, 371, 210-21.
Harris, S. R., Feil, E. J., Holden, M. T., Quail, M. A., Nickerson, E. K., Chantratita,
N., Gardete, S., Tavares, A., Day, N., Lindsay, J. A., Edgeworth, J. D., De Lencastre, H., Parkhill, J., Peacock, S. J. & Bentley, S. D. (2010). Evolution of MRSA during hospital transmission and intercontinental spread. Science, 327, 469-74.

61
Heinrichs, J. H., Bayer, M. G. & Cheung, A. L. (1996). Characterization of the sar locus and its interaction with agr in Staphylococcus aureus. J Bacteriol, 178, 418-23.
Hennekinne, J. A., Ostyn, A., Guillier, F., Herbin, S., Prufer, A. L. & Dragacci, S. (2010). How should staphylococcal food poisoning outbreaks be characterized? Toxins (Basel), 2, 2106-16.
Hiramatsu, K., Aritaka, N., Hanaki, H., Kawasaki, S., Hosoda, Y., Hori, S., Fukuchi, Y. & Kobayashi, I. (1997). Dissemination in Japanese hospitals of strains of Staphylococcus aureus heterogeneously resistant to vancomycin. Lancet, 350, 1670-3.
Hong, S. C., Waterbury, G. & Janeway, C. A., Jr. (1996). Different superantigens interact with distinct sites in the Vbeta domain of a single T cell receptor. J Exp Med, 183, 1437-46.
Hudson, K. R., Tiedemann, R. E., Urban, R. G., Lowe, S. C., Strominger, J. L. & Fraser, J. D. (1995). Staphylococcal enterotoxin A has two cooperative binding sites on major histocompatibility complex class II. J Exp Med, 182, 711-20.
Hurtado, M. P., de la Parte, M. A., Brito, A. (2002). Staphylococcus aureus: Revisión de los mecanismos de patogenicidad y la fisiopatología de la infección estafilocócica. Rev Soc Ven Microbiol, 22, 2, 112–118.
Jamali, H., Paydar, M., Radmehr, B. & Ismail, S. (2015). Prevalence, characterization, and antimicrobial resistance of Yersinia species and Yersinia enterocolitica isolated from raw milk in farm bulk tanks. J Dairy Sci, 98, 798-803.
Jardetzky, T. S., Brown, J. H., Gorga, J. C., Stern, L. J., Urban, R. G., Chi, Y. I., Stauffacher, C., Strominger, J. L. & Wiley, D. C. (1994). Three-dimensional structure of a human class II histocompatibility molecule complexed with superantigen. Nature, 368, 711-8.
Jarraud, S., Mougel, C., Thioulouse, J., Lina, G., Meugnier, H., Forey, F., Nesme, X., Etienne, J. & Vandenesch, F. (2002). Relationships between Staphylococcus aureus genetic background, virulence factors, agr groups (alleles), and human disease. Infect Immun, 70, 631-41.
Jarraud, S., Peyrat, M. A., Lim, A., Tristan, A., Bes, M., Mougel, C., Etienne, J., Vandenesch, F., Bonneville, M. & Lina, G. (2001). Egc, a highly prevalent operon of enterotoxin gene, forms a putative nursery of superantigens in Staphylococcus aureus. J Immunol, 166, 669-77.
Jett, M., Brinkley, W., Neill, R., Gemski, P. & Hunt, R. (1990). Staphylococcus aureus enterotoxin B challenge of monkeys: correlation of plasma levels of arachidonic acid cascade products with occurrence of illness. Infect Immun, 58, 3494-9.
Jorda, G. B., Marucci, R. S., Guida A. M., Pires P. S. & Manfredi E. A. (2012) Portación y caracterización de Staphylococcus aureus en manipuladores de alimentos. Rev. Argent. Microbiol, 44, 2,101-104.

62
Kadariya, J., Smith, T. C. & Thapaliya, D. (2014). Staphylococcus aureus and staphylococcal food-borne disease: an ongoing challenge in public health. Biomed Res Int, 827965.
Kerouanton, A., Hennekinne, J. A., Letertre, C., Petit, L., Chesneau, O., Brisabois, A. & De Buyser, M. L. (2007). Characterization of Staphylococcus aureus strains associated with food poisoning outbreaks in France. Int J Food Microbiol, 115, 369-75.
Khan, S. A., Nawaz, M. S., Khan, A. A. & Cerniglia, C. E. (2000).Transfer of erythromycin resistance from poultry to human clinical strains of Staphylococcus aureus. J. Clin. Microbiol, 38, 1832–1838.
Kim, J., Urban, R. G., Strominger, J. L. & Wiley, D. C. (1994). Toxic shock syndrome toxin-1 complexed with a class II major histocompatibility molecule HLA-DR1. Science, 266, 1870-4.
Kim, N. H., Yun, A. R. & Rhee, M. S. (2011). Prevalence and classification of toxigenic Staphylococcus aureus isolated from refrigerated ready-to-eat foods (sushi, kimbab and California rolls) in Korea. J Appl Microbiol, 111, 1456-64.
Kitamoto, M., Kito, K., Niimi, Y., Shoda, S., Takamura, A., Hiramatsu, T., Akashi, T., Yokoi, Y., Hirano, H., Hosokawa, M., Yamamoto, A., Agata, N. & Hamajima, N. (2009). Food poisoning by Staphylococcus aureus at a university festival. Jpn J Infect Dis, 62, 242-3.
Kluytmans, J., Van Belkum, A. & Verbrugh, H. (1997). Nasal carriage of Staphylococcus aureus: epidemiology, underlying mechanisms, and associated risks. Clin Microbiol Rev, 10, 505-20.
Kluytmans, J. A. & Wertheim, H. F. (2005). Nasal carriage of Staphylococcus aureus and prevention of nosocomial infections. Infection, 33, 3-8.
Komisar, J., Rivera, J., Vega, A. & Tseng, J. (1992). Effects of staphylococcal enterotoxin B on rodent mast cells. Infect Immun, 60, 2969-75.
Koning, S., Van Belkum, A., Snijders, S., Van Leeuwen, W., Verbrugh, H., Nouwen, J., Op 'T Veld, M., Van Suijlekom-Smit, L. W., Van Der Wouden, J. C. & Verduin, C. (2003). Severity of nonbullous Staphylococcus aureus impetigo in children is associated with strains harboring genetic markers for exfoliative toxin B, Panton-Valentine leukocidin, and the multidrug resistance plasmid psk41. J Clin Microbiol, 41, 3017-21.
Kreiswirth, B. N. (1989). Genetics and expression of toxic shock syndrome toxin 1: overview. Rev Infect Dis, 11 Suppl 1, S97-100.
Kuehnert, M. J., Kruszon-Moran, D., Hill, H. A., Mcquillan, G., Mcallister, S. K., Fosheim, G., Mcdougal, L. K., Chaitram, J., Jensen, B., Fridkin, S. K., Killgore, G. & Tenover, F. C. (2006). Prevalence of Staphylococcus aureus nasal colonization in the United States, 2001-2002. J Infect Dis, 193, 172-9.
Kumate, Jesus; Gutiérrez, Gonzalo; Munoz Onofre; Santos, Ignacio; Solorzano
Fontino & Guadalupe Miranda. (2008). Chapter 41: Skin infections and soft tissue. Clinical Infectious Kumate-Gutierrez. Mexico: Mendez Editor, 455-467.

63
Kuroda, M., Ohta, T., Uchiyama, I., Baba, T., Yuzawa, H., Kobayashi, I., Cui, L., Oguchi, A., Aoki, K., Nagai, Y., Lian, J., Ito, T., Kanamori, M., Matsumaru, H., Maruyama, A., Murakami, H., Hosoyama, A., Mizutani-Ui, Y., Takahashi, N. K., Sawano, T., Inoue, R., Kaito, C., Sekimizu, K., Hirakawa, H., Kuhara, S., Goto, S., Yabuzaki, J., Kanehisa, M., Yamashita, A., Oshima, K., Furuya, K., Yoshino, C., Shiba, T., Hattori, M., Ogasawara, N., Hayashi, H. & Hiramatsu, K. (2001). Whole genome sequencing of meticillin-resistant Staphylococcus aureus. Lancet, 357, 1225-40.
Ladhani, S. 2003. Understanding the mechanism of action of the exfoliative toxins of Staphylococcus aureus. Fems Immunol Med Microbiol, 39, 181-9.
Ladhani, S., Joannou, C. L., Lochrie, D. P., Evans, R. W. & Poston, S. M. (1999). Clinical, microbial, and biochemical aspects of the exfoliative toxins causing staphylococcal scalded-skin syndrome. Clin Microbiol Rev, 12, 224-42.
Larkin, E. A., Carman, R. J., Krakauer, T. & Stiles, B. G. (2009). Staphylococcus aureus: the toxic presence of a pathogen extraordinaire. Curr Med Chem, 16, 4003-19.
Le Loir, Y., Baron, F. & Gautier, M. (2003). Staphylococcus aureus and food poisoning. Genet Mol Res, 2, 63-76.
Letertre, C., Perelle, S., Dilasser, F. & Fach, P. (2003). Identification of a new putative enterotoxin SEU encoded by the egc cluster of Staphylococcus aureus. J Appl Microbiol, 95, 38-43.
Li, H., Llera, A., Malchiodi, E. L. & Mariuzza, R. A. (1999). The structural basis of T cell activation by superantigens. Annu Rev Immunol, 17, 435-66.
Li, H., Llera, A., Tsuchiya, D., Leder, L., Ysern, X., Schlievert, P. M., Karjalainen, K. & Mariuzza, R. A. (1998). Three-dimensional structure of the complex between a T cell receptor beta chain and the superantigen staphylococcal enterotoxin B. Immunity, 9, 807-16.
Li, J., Wang, W., Xu, S. X., Magarvey, N. A. & Mccormick, J. K. (2011). Lactobacillus reuteri-produced cyclic dipeptides quench agr-mediated expression of toxic shock syndrome toxin-1 in staphylococci. Proc Natl Acad Sci U S A, 108, 3360-5.
Li, Y., Li, H., Dimasi, N., Mccormick, J. K., Martin, R., Schuck, P., Schlievert, P. M. & Mariuzza, R. A. (2001). Crystal structure of a superantigen bound to the high-affinity, zinc-dependent site on MHC class II. Immunity, 14, 93-104.
Lindsay, J. A. (2010). Genomic variation and evolution of Staphylococcus aureus. Int J Med Microbiol, 300, 98-103.
Lindsay, J. A. & Holden, M. T. (2004). Staphylococcus aureus: superbug, super genome? Trends Microbiol, 12, 378-85.
Lindsay, J. A. & Holden, M. T. (2006). Understanding the rise of the superbug: investigation of the evolution and genomic variation of Staphylococcus aureus. Funct Integr Genomics, 6, 186-201.
Lindsay, J. A., Ruzin, A., Ross, H. F., Kurepina, N. & Novick, R. P. (1998). The gene for toxic shock toxin is carried by a family of mobile pathogenicity islands in Staphylococcus aureus. Mol Microbiol, 29, 527-43.

64
Lipinska, U., Hermans, K., Meulemans, L., Dumitrescu, O., Badiou, C., Duchateau, L., Haesebrouck, F., Etienne, J. & Lina, G. (2011). Panton-Valentine leukocidin does play a role in the early stage of Staphylococcus aureus skin infections: a rabbit model. Plos One, 6, e22864.
Lowy, F. D. (1998). Staphylococcus aureus infections. N Engl J Med, 339, 520-32. Lu, P. L., Chin, L. C., Peng, C. F., Chiang, Y. H., Chen, T. P., Ma, L. & Siu, L. K.
(2005). Risk factors and molecular analysis of community methicillin-resistant Staphylococcus aureus carriage. J Clin Microbiol, 43, 132-9.
Macfaddin, J. (2000). 13-lactamase test. In: Biochemical test for identification of medical bacteria. 3rd Ed. Uníted State of Ameríca. Edítoríal Lawrence mcgrew, 254-272.
Manenschijn, L., Jetten, A. M., Van Wamel, W. J., Tavakol, M., Koper, J. W., Van Den Akker, E. L., Van Belkum, A. & Van Rossum, E. F. (2012). Long-term cortisol levels are not associated with nasal carriage of Staphylococcus aureus. Eur J Clin Microbiol Infect Dis, 31, 97-100.
Mandell G.L, J.E Bennett, R. Dolin. Bennett’s – Principaples and practice of infectious diseases. (2000). 5th ed Churchill Livigstone (Philadelphia, Pennsylvania).
Mamprim, F. A. (2006). Enterotoxinas de Staphylococcus coagulase positiva e negativa isoladas das fossas nasais e mäos de manipuladores de alimentos, Dissertagäo (mestrado). Universidade Estadual Paulista, Faculdade de Medicina Veterinària e Zootecnia, Botucatu.
Marr, J. C., Lyon, J. D., Roberson, J. R., Lupher, M., Davis, W. C. & Bohach, G. A. (1993). Characterization of novel type C staphylococcal enterotoxins: biological and evolutionary implications. Infect Immun, 61, 4254-62.
Marrack, P. & Kappler, J. (1990). The staphylococcal enterotoxins and their relatives. Science, 248, 1066.
Mcculloch, J, A. (2006). Avaliação da funcionalidade do locus acessory gene regulator (agr) em cepas de Staphylococcus aureus brasileiras com suscetibilidade reduzida aos glicopeptídeos. Faculdade de Ciências Farmacêuticas, São Paulo.
Mcnamara, P. J., Milligan-Monroe, K. C., Khalili, S. & Proctor, R. A. (2000). Identification, cloning, and initial characterization of rot, a locus encoding a regulator of virulence factor expression in Staphylococcus aureus. J Bacteriol, 182, 3197-203.
Mead, P. S., Slutsker, L., Dietz, V., Mccaig, L. F., Bresee, J. S., Shapiro, C., Griffin, P. M. & Tauxe, R. V. (1999). Food-related illness and death in the United States. Emerg Infect Dis, 5, 607-25.
Mehrotra, M., Wang, G. & Johnson, W. M. (2000). Multiplex PCR for detection of genes for Staphylococcus aureus enterotoxins, exfoliative toxins, toxic shock syndrome toxin 1, and methicillin resistance. J Clin Microbiol, 38, 1032-5.
Meroueh, S. O., Bencze, K. Z., Hesek, D., Lee, M., Fisher, J. F., Stemmler, T. L. & Mobashery, S. (2006). Three-dimensional structure of the bacterial cell wall peptidoglycan. Proc Natl Acad Sci U S A, 103, 4404-9.

65
Mertz, D., Frei, R., Jaussi, B., Tietz, A., Stebler, C., Fluckiger, U. & Widmer, A. F. (2007). Throat swabs are necessary to reliably detect carriers of Staphylococcus aureus. Clin Infect Dis, 45, 475-7.
Mitchell, D. T., Levitt, D. G., Schlievert, P. M. & Ohlendorf, D. H. (2000). Structural evidence for the evolution of pyrogenic toxin superantigens. J Mol Evol, 51, 520-31.
Moises, S, S & Schäferling M. (2009).Toxin immunosensors and sensor arrays for f Food quality control. Bioanal Rev, 1, 73–104.
Morandi, S., Brasca, M., Andrighetto, C., Lombardi, A. & Lodi, R. (2009). Phenotypic and Genotypic Characterization of Staphylococcus aureus Strains from Italian Dairy Products. Int J Microbiol, 501362.
Morales, M. G & Ruiz de Chávez, C. G. (2006). Diferencias en la resistencia a los antimicrobianos de cepas de Staphylococcus aureus obtenidas de diversas fuentes de aislamiento. Revista del Centro de Investigación, 7, 25, 45- 64.
Moza, B., Varma, A. K., Buonpane, R. A., Zhu, P., Herfst, C. A., Nicholson, M. J., Wilbuer, A. K., Seth, N. P., Wucherpfennig, K. W., Mccormick, J. K., Kranz, D. M. & Sundberg, E. J. (2007). Structural basis of T-cell specificity and activation by the bacterial superantigen TSST-1. Embo J, 26, 1187-97.
Much, H. (1908). Ober eine vorstufe des fibrinfermentes in kulturen von Staphylokokkus aureus. Biochemische Zeitschrift, 14,143–155.
Muller-Alouf, H., Carnoy, C., Simonet, M. & Alouf, J. E. (2001). Superantigen bacterial toxins: state of the art. Toxicon, 39, 1691-701.
Munson, S. H., Tremaine, M. T., Betley, M. J. & Welch, R. A. (1998). Identification and characterization of staphylococcal enterotoxin types G and I from Staphylococcus aureus. Infect Immun, 66, 3337-48.
Murray, R. J. 2005. Staphylococcus aureus infective endocarditis: diagnosis and management guidelines. Intern Med J, 35, 2, S25-44.
Musser, J. M., Schlievert, P. M., Chow, A. W., Ewan, P., Kreiswirth, B. N., Rosdahl, V. T., Naidu, A. S., Witte, W. & Selander, R. K. (1990). A single clone of Staphylococcus aureus causes the majority of cases of toxic shock syndrome. Proc Natl Acad Sci USA, 87, 225-9.
Navarre, W. W. & Schneewind, O. (1999). Surface proteins of gram-positive bacteria and mechanisms of their targeting to the cell wall envelope. Microbiol Mol Biol Rev, 63, 174-229.
Nishi, J., Yoshinaga, M., Miyanohara, H., Kawahara, M., Kawabata, M., Motoya, T., Owaki, T., Oiso, S., Kawakami, M., Kamewari, S., Koyama, Y., Wakimoto, N., Tokuda, K., Manago, K. & Maruyama, I. (2002). An epidemiologic survey of methicillin-resistant Staphylococcus aureus by combined use of mec-HVR genotyping and toxin genotyping in a university hospital in Japan. Infect Control Hosp Epidemiol, 23, 506-10.
Nouwen, J. L., Ott, A., Kluytmans-Vandenbergh, M. F., Boelens, H. A., Hofman, A., Van Belkum, A. & Verbrugh, H. A. (2004). Predicting the Staphylococcus aureus nasal carrier state: derivation and validation of a "culture rule". Clin Infect Dis, 39, 806-11.

66
Novick, R. P. (2003a). Autoinduction and signal transduction in the regulation of staphylococcal virulence. Mol Microbiol, 48, 1429-49.
Novick, R. P. (2003b) Mobile genetic elements and bacterial toxinoses: the superantigen-encoding pathogenicity islands of Staphylococcus aureus. Plasmid, 49, 93-105.
Novick, R. P. & Geisinger, E. (2008). Quorum sensing in staphylococci. Annu Rev Genet, 42, 541-64.
Novick, R. P., Ross, H. F., Projan, S. J., Kornblum, J., Kreiswirth, B. & Moghazeh, S. (1993). Synthesis of staphylococcal virulence factors is controlled by a regulatory RNA molecule. Embo J, 12, 3967-75.
Novick, R. P., Schlievert, P. & Ruzin, A. (2001). Pathogenicity and resistance islands of staphylococci. Microbes Infect, 3, 585-94.
Oh, S. K., Lee, N., Cho, Y. S., Shin, D. B., Choi, S. Y. & Koo, M. (2007). Occurrence of toxigenic Staphylococcus aureus in ready-to-eat food in Korea. J Food Prot, 70, 1153-8.
Omoe, K., Hu, D. L., Takahashi-Omoe, H., Nakane, A. & Shinagawa, K. (2003). Identification and characterization of a new staphylococcal enterotoxin-related putative toxin encoded by two kinds of plasmids. Infect Immun, 71, 6088-94.
Omoe, K., Hu, D. L., Takahashi-Omoe, H., Nakane, A. & Shinagawa, K. (2005a). Comprehensive analysis of classical and newly described staphylococcal superantigenic toxin genes in Staphylococcus aureus isolates. Fems Microbiol Lett, 246, 191-8.
Omoe, K., Imanishi, K., Hu, D. L., Kato, H., Fugane, Y., Abe, Y., Hamaoka, S., Watanabe, Y., Nakane, A., Uchiyama, T. & Shinagawa, K. (2005b). Characterization of novel staphylococcal enterotoxin-like toxin type P. Infect Immun, 73, 5540-6.
Ono, H. K., Omoe, K., Imanishi, K., Iwakabe, Y., Hu, D. L., Kato, H., Saito, N., Nakane, A., Uchiyama, T. & Shinagawa, K. (2008). Identification and characterization of two novel staphylococcal enterotoxins, types S and T. Infect Immun, 76, 4999-5005.
Ortega, E., Abriouel, H., Lucas, R. & Galvez, A. (2010) Multiple roles of Staphylococcus aureus enterotoxins: pathogenicity, superantigenic activity, and correlation to antibiotic resistance. Toxins (Basel), 2, 2117-31.
Orwin, P. M., Leung, D. Y., Donahue, H. L., Novick, R. P. & Schlievert, P. M. (2001). Biochemical and biological properties of Staphylococcal enterotoxin K. Infect Immun, 69, 360-6.
Otero, A., Garcia, M. L., Garcia, M. C., Moreno, B. & Bergdoll, M. S. (1990). Production of staphylococcal enterotoxins C1 and C2 and thermonuclease throughout the growth cycle. Appl Environ Microbiol, 56, 555-9.
Peacock, S. J., Moore, C. E., Justice, A., Kantzanou, M., Story, L., Mackie, K., O'neill, G. & Day, N. P. (2002). Virulent combinations of adhesin and toxin genes in natural populations of Staphylococcus aureus. Infect Immun, 70, 4987-96.

67
Peng, H. L., Novick, R. P., Kreiswirth, B., Kornblum, J. & Schlievert, P. (1988). Cloning, characterization, and sequencing of an accessory gene regulator (agr) in Staphylococcus aureus. J Bacteriol, 170, 4365-72.
Pesavento, G., Ducci, B., Comodo, N., Nostro, A. L. (2007). Antimicrobial resistance profile of Staphylococcus aureus isolated from raw meat: A research for methicillin resistant Staphylococcus aureus (MRSA). Food Control, 18, 196–200.
Petersson, K., Hakansson, M., Nilsson, H., Forsberg, G., Svensson, L. A., Liljas, A. & Walse, B. (2001). Crystal structure of a superantigen bound to MHC class II displays zinc and peptide dependence. Embo J, 20, 3306-12.
Pietrocola, G., Arciola, C. R., Rindi, S., Di Poto, A., Missineo, A., Montanaro, L. & Speziale, P. (2011). Toll-like receptors (tlrs) in innate immune defense against Staphylococcus aureus. Int J Artif Organs, 34, 799-810.
Pinchuk, I. V., Beswick, E. J. & Reyes, V. E. (2010). Staphylococcal enterotoxins. Toxins (Basel), 2, 2177-97.
Plata, K., Rosato, A. E. & Wegrzyn, G. (2009). Staphylococcus aureus as an infectious agent: overview of biochemistry and molecular genetics of its pathogenicity. Acta Biochim Pol, 56, 597-612.
Pragman, A. A., Yarwood, J. M., Tripp, T. J. & Schlievert, P. M. (2004). Characterization of virulence factor regulation by srrab, a two-component system in Staphylococcus aureus. J Bacteriol, 186, 2430-8.
Qi, Y. & Miller, K. J. (2000). Effect of low water activity on staphylococcal enterotoxin A and B biosynthesis. J Food Prot, 63, 473-8.
Rapini, L. S., Cerqueira, M. M.O. P., Carmo., L. S, Veras, J. F. & Souza, M. R. (2005).Presence of Staphylococcus strains producer of enterotoxins and toxic shock toxin syndrome isolated from goat's cheese handlers.Arq.bras.med.vet.zootec. 57, 825-829.
Ray, B., Ballal, A. & Manna, A. C. (2009). Transcriptional variation of regulatory and virulence genes due to different media in Staphylococcus aureus. Microb Pathog, 47, 94-100.
Reck, B., Scheuber, P. H., Londong, W., Sailer-Kramer, B., Bartsch, K. & Hammer, D. K. (1988). Protection against the staphylococcal enterotoxin-induced intestinal disorder in the monkey by anti-idiotypic antibodies. Proc Natl Acad Sci U S A, 85, 3170-4.
Reiser, R. F., Robbins, R. N., Khoe, G. P. & Bergdoll, M. S. (1983). Purification and some physicochemical properties of toxic-shock toxin. Biochemistry, 22, 3907-12.
Reynolds, J. & Wigneshweraraj, S. (2011). Molecular insights into the control of transcription initiation at the Staphylococcus aureus agr operon. J Mol Biol, 412, 862-81.
Rivera, J., Mujica, I., Aranaga, V., Navarro, C., Zabala, I & Atencio, L. (2011). Staphylococcus aureus procedentes de quesos: susceptibilidad a antibióticos y su relación con plásmidos. Rev Científica, 21, 3, 202-210

68
Rosenbach, T. O., Moshonov, S., Zor, U. & Yaron, M. (1984). Interferon triggers experimental synovitis and may potentiate auto-immune disease in humans. Clin Rheumatol, 3, 361-4.
Ruzickova, V., Voller, J., Pantucek, R., Petras, P. & Doskar, J. (2005). Multiplex PCR for detection of three exfoliative toxin serotype genes in Staphylococcus aureus. Folia Microbiol (Praha), 50, 499-502.
Ruzin, A., Lindsay, J. & Novick, R. P. (2001). Molecular genetics of sapi1--a mobile pathogenicity island in Staphylococcus aureus. Mol Microbiol, 41, 365-77.
Sato, H., Matsumori, Y., Tanabe, T., Saito, H., Shimizu, A. & Kawano, J. (1994). A new type of staphylococcal exfoliative toxin from a Staphylococcus aureus strain isolated from a horse with phlegmon. Infect Immun, 62, 3780-5.
Scott, W. J. (1953). Water relations of Staphylococcus aureus at 30 degrees C. Aust J Biol Sci, 6, 549-64.
Schantz, E. J., Roessler, W. G., Wagman, J., Spero, L., Dunnery, D. A. & Bergdoll, M. S. (1965). Purification of staphylococcal enterotoxin B. Biochemistry, 4, 1011-6.
Schelin, J., Wallin-Carlquist, N., Cohn, M. T., Lindqvist, R., Barker, G. C. & Radstrom, P. (2011). The formation of Staphylococcus aureus enterotoxin in food environments and advances in risk assessment. Virulence, 2, 580-92.
Scheuber, P. H., Denzlinger, C., Wilker, D., Beck, G., Keppler, D. & Hammer, D. K. (1987). Cysteinyl leukotrienes as mediators of staphylococcal enterotoxin B in the monkey. Eur J Clin Invest, 17, 455-9.
Schlievert, P. M., Case, L. C., Strandberg, K. L., Abrams, B. B. & Leung, D. Y. (2008). Superantigen profile of Staphylococcus aureus isolates from patients with steroid-resistant atopic dermatitis. Clin Infect Dis, 46, 1562-7.
Schlievert, P. M., Jablonski, L. M., Roggiani, M., Sadler, I., Callantine, S., Mitchell, D. T., Ohlendorf, D. H. & Bohach, G. A. (2000). Pyrogenic toxin superantigen site specificity in toxic shock syndrome and food poisoning in animals. Infect Immun, 68, 3630-4.
Schlievert, P. M., Shands, K. N., Dan, B. B., Schmid, G. P. & Nishimura, R. D. (1981). Identification and characterization of an exotoxin from Staphylococcus aureus associated with toxic-shock syndrome. J Infect Dis, 143, 509-16.
Schlievert, P. M., Strandberg, K. L., Lin, Y. C., Peterson, M. L. & Leung, D. Y. (2010). Secreted virulence factor comparison between methicillin-resistant and methicillin-sensitive Staphylococcus aureus, and its relevance to atopic dermatitis. J Allergy Clin Immunol, 125, 39-49.
Schubert. M. S., (2009). Superantigens and Allergic Disease. Allergy Frontiers: Classification and Pathomechanisms, 2, 59-77.
Seo Y. H., Jang J. H & Moon K, D. (2010). Occurrence and characterization of enterotoxigenic Staphylococcus aureus isolated from minimally processed vegetables and sprouts in Korea, 19, 2, 313-319.
Seybold, U., Schubert, S., Bogner, J. R. & Hogardt, M. (2011). Staphylococcus aureus infection following nasal colonization: an approach to rapid risk stratification in a university healthcare system. J Hosp Infect, 79, 297-301.

69
Shands, K. N., Schmid, G. P., Dan, B. B., Blum, D., Guidotti, R. J., Hargrett, N. T., Anderson, R. L., Hill, D. L., Broome, C. V., Band, J. D. & Fraser, D. W. (1980). Toxic-shock syndrome in menstruating women: association with tampon use and Staphylococcus aureus and clinical features in 52 cases. N Engl J Med, 303, 1436-42.
Sospedra, I., Manes, J. & Soriano, J. M. (2012). Report of toxic shock syndrome toxin 1 (TSST-1) from Staphylococcus aureus isolated in food handlers and surfaces from foodservice establishments. Ecotoxicol Environ Saf, 80, 288-90.
Sumby, P. & Waldor, M. K. (2003). Transcription of the toxin genes present within the Staphylococcal phage phisa3ms is intimately linked with the phage's life cycle. J Bacteriol, 185, 6841-51.
Sundberg, E. J., Deng, L. & Mariuzza, R. A. (2007). TCR recognition of peptide/MHC class II complexes and superantigens. Semin Immunol, 19, 262-71.
Sundberg, E. J., Li, H., Llera, A. S., Mccormick, J. K., Tormo, J., Schlievert, P. M., Karjalainen, K. & Mariuzza, R. A. (2002). Structures of two streptococcal superantigens bound to TCR beta chains reveal diversity in the architecture of T cell signaling complexes. Structure, 10, 687-99.
Sykes, R.B., M. Mathew. (1976). “The b-lactamase of Gram-negative Bacteria and Their Role in Resistance to b-lactam Antibiotics”. J. Antimicrob. Chemother, 2, 115-157.
Takeuchi, S., Ishiguro, K., Ikegami, M., Kaidoh, T. & Hayakawa, Y. (1998). Production of toxic shock syndrome toxin by Staphylococcus aureus isolated from mastitic cow's milk and farm bulk milk. Vet Microbiol, 59, 251-8.
Tamarapu, S., Mckillip, J. L. & Drake, M. 2001. Development of a multiplex polymerase chain reaction assay for detection and differentiation of Staphylococcus aureus in dairy products. J Food Prot, 64, 664-8.
Teixeira, L. A., Resende, C. A., Ormonde, L. R., Rosenbaum, R., Figueiredo, A. M., De Lencastre, H. & Tomasz, A. (1995). Geographic spread of epidemic multiresistant Staphylococcus aureus clone in Brazil. J Clin Microbiol, 33, 2400-4.
Thomas, D. Y., Jarraud, S., Lemercier, B., Cozon, G., Echasserieau, K., Etienne, J., Gougeon, M. L., Lina, G. & Vandenesch, F. (2006). Staphylococcal enterotoxin-like toxins U2 and V, two new staphylococcal superantigens arising from recombination within the enterotoxin gene cluster. Infect Immun, 74, 4724-34.
Throup, J. P., Zappacosta, F., Lunsford, R. D., Annan, R. S., Carr, S. A., Lonsdale, J. T., Bryant, A. P., Mcdevitt, D., Rosenberg, M. & Burnham, M. K. (2001). The srhsr gene pair from Staphylococcus aureus: genomic and proteomic approaches to the identification and characterization of gene function. Biochemistry, 40, 10392-401.
Todd, J., Fishaut, M., Kapral, F. & Welch, T. (1978). Toxic-shock syndrome associated with phage-group-I Staphylococci. Lancet, 2, 1116-8.

70
Torres, B. A., Kominsky, S., Perrin, G. Q., Hobeika, A. C. & Johnson, H. M. (2001). Superantigens: the good, the bad, and the ugly. Exp Biol Med (Maywood), 226, 164-76.
Tranter, H. S. (1990). Foodborne staphylococcal illness. Lancet, 336, 1044-6. Udo, E. E., Al-Mufti, S. & Albert, M. J. (2009). The prevalence of antimicrobial
resistance and carriage of virulence genes in Staphylococcus aureus isolated from food handlers in Kuwait City restaurants. BMC Res Notes, 2, 108.
Valdiviezo N, Villalobos L B, Martínez R. (2006). Evaluación microbiológica en manipuladores de alimentos de tres comedores públicos en Cumana – Venezuela. Rev. Soc. Ven. Microbiol, 26, 2.
Valle, J., Vadillo, S., Piriz, S. & Gomez-Lucia, E. (1991). Toxic shock syndrome toxin 1 (TSST-1) production by staphylococci isolated from goats and presence of specific antibodies to TSST-1 in serum and milk. Appl Environ Microbiol, 57, 889-91.
Van Belkum, A., Verkaik, N. J., De Vogel, C. P., Boelens, H. A., Verveer, J., Nouwen, J. L., Verbrugh, H. A. & Wertheim, H. F. (2009). Reclassification of Staphylococcus aureus nasal carriage types. J Infect Dis, 199, 1820-6.
Van Den Akker, E. L., Nouwen, J. L., Melles, D. C., Van Rossum, E. F., Koper, J. W., Uitterlinden, A. G., Hofman, A., Verbrugh, H. A., Pols, H. A., Lamberts, S. W. & Van Belkum, A. (2006). Staphylococcus aureus nasal carriage is associated with glucocorticoid receptor gene polymorphisms. J Infect Dis, 194, 814-8.
Vanegas M., González L., Martínez, A. & Buitrago, F. (2008). Aislamiento y Caracterización de cepas de Staphylococcus enterotoxigénicos aislados de quesos en Bogotá. Rev. MVZ Córdoba, 13, 1288-1293.
Vasconcelos N. G. & Cunha M .L. R. S. (2010). Staphylococcal enterotoxins: Molecular aspects and detection methods. J Public Health Epidemiol, 2, 29-42.
Veras, J. F., Do Carmo, L. S., Tong, L. C., Shupp, J. W., Cummings, C., Dos Santos, D. A., Cerqueira, M. M., Cantini, A., Nicoli, J. R. & Jett, M. (2008). A study of the enterotoxigenicity of coagulase-negative and coagulase-positive staphylococcal isolates from food poisoning outbreaks in Minas Gerais, Brazil. Int J Infect Dis, 12, 410-5.
Wallin, N., Cao, R., Marta, D., Da Silva, A. S., Schelin, J. & Radstrom, P. (2010). Acetic acid increases the phage-encoded enterotoxin A expression in Staphylococcus aureus. BMC Microbiol, 10, 147.
Wattinger, L., Stephan, R., Layer, F. & Johler, S. (2012). Comparison of Staphylococcus aureus isolates associated with food intoxication with isolates from human nasal carriers and human infections. Eur J Clin Microbiol Infect Dis, 31, 455-64.

71
Wen, R., Broussard, D. R., Surman, S., Hogg, T. L., Blackman, M. A. & Woodland, D. L. (1997). Carboxy-terminal residues of major histocompatibility complex class II-associated peptides control the presentation of the bacterial superantigen toxic shock syndrome toxin-1 to T cells. Eur J Immunol, 27, 772-81.
Wenzel, R. P. & Perl, T. M. (1995). The significance of nasal carriage of Staphylococcus aureus and the incidence of postoperative wound infection. J Hosp Infect, 31, 13-24.
Wertheim, H. F., Melles, D. C., Vos, M. C., Van Leeuwen, W., Van Belkum, A., Verbrugh, H. A. & Nouwen, J. L. (2005). The role of nasal carriage in Staphylococcus aureus infections. Lancet Infect Dis, 5, 751-62.
Wertheim, H. F., Vos, M. C., Ott, A., Van Belkum, A., Voss, A., Kluytmans, J. A., Van Keulen, P. H., Vandenbroucke-Grauls, C. M., Meester, M. H. & Verbrugh, H. A. (2004). Risk and outcome of nosocomial Staphylococcus aureus bacteraemia in nasal carriers versus non-carriers. Lancet, 364, 703-5.
White, J., Herman, A., Pullen, A. M., Kubo, R., Kappler, J. W. & Marrack, P. (1989). The V beta-specific superantigen staphylococcal enterotoxin B: stimulation of mature T cells and clonal deletion in neonatal mice. Cell, 56, 27-35.
Widaa, A., Claro, T., Foster, T. J., O'brien, F. J. & Kerrigan, S. W. (2012). Staphylococcus aureus protein A plays a critical role in mediating bone destruction and bone loss in osteomyelitis. Plos ONE, 7, e40586.
Wieneke, A. A., Roberts, D. & Gilbert, R. J. (1993). Staphylococcal food poisoning in the United Kingdom, 1969-90. Epidemiol Infect, 110, 519-31.
Wilson, G. J., Seo, K. S., Cartwright, R. A., Connelley, T., Chuang-Smith, O. N., Merriman, J. A., Guinane, C. M., Park, J. Y., Bohach, G. A., Schlievert, P. M., Morrison, W. I. & Fitzgerald, J. R. (2011). A novel core genome-encoded superantigen contributes to lethality of community-associated MRSA necrotizing pneumonia. Plos Pathog, 7, e1002271.
Williams, R. E. (1963). Healthy carriage of Staphylococcus aureus: its prevalence and importance. Bacteriol Rev, 27, 56-71.
Xu, S. X. & Mccormick, J. K. (2012). Staphylococcal superantigens in colonization and disease. Front Cell Infect Microbiol, 2, 52.
Yamaguchi, T., Hayashi, T., Takami, H., Ohnishi, M., Murata, T., Nakayama, K., Asakawa, K., Ohara, M., Komatsuzawa, H. & Sugai, M. (2001). Complete nucleotide sequence of a Staphylococcus aureus exfoliative toxin B plasmid and identification of a novel ADP-ribosyltransferase, EDIN-C. Infect Immun, 69, 7760-71.
Yamaguchi, T., Nishifuji, K., Sasaki, M., Fudaba, Y., Aepfelbacher, M., Takata, T., Ohara, M., Komatsuzawa, H., Amagai, M. & Sugai, M. (2002). Identification of the Staphylococcus aureus etd pathogenicity island which encodes a novel exfoliative toxin, ETD, and EDIN-B. Infect Immun, 70, 5835-45.

72
Yamasaki, O., Tristan, A., Yamaguchi, T., Sugai, M., Lina, G., Bes, M., Vandenesch, F. & Etienne, J. (2006). Distribution of the exfoliative toxin D gene in clinical Staphylococcus aureus isolates in France. Clin Microbiol Infect, 12, 585-8.
Yarwood, J. M., Mccormick, J. K., Paustian, M. L., Orwin, P. M., Kapur, V. & Schlievert, P. M. (2002). Characterization and expression analysis of Staphylococcus aureus pathogenicity island 3. Implications for the evolution of staphylococcal pathogenicity islands. J Biol Chem, 277, 13138-47.
Yarwood, J. M., Mccormick, J. K. & Schlievert, P. M. (2001). Identification of a novel two-component regulatory system that acts in global regulation of virulence factors of Staphylococcus aureus. J Bacteriol, 183, 1113-23.
Yarwood, J. M. & Schlievert, P. M. (2003). Quorum sensing in Staphylococcus infections. J Clin Invest, 112, 1620-5.
Zhang, S., Iandolo, J. J. & Stewart, G. C. (1998). The enterotoxin D plasmid of Staphylococcus aureus encodes a second enterotoxin determinant (sej). Fems Microbiol Lett, 168, 227-33.

73
ANEXOS.
Anexo A. Número de aislamientos positivos para S. aureus, aislados de
Madres Comunitaria.
Aislamiento positivo Positivo para
Fosas Nasales
Positivo para
Manos Origen de la muestra
6 1 0 Aislamientos de Madres
comunitarias
10 1 0 Aislamientos de Madres
comunitarias
12 1 0 Aislamientos de Madres
comunitarias
24 1 0 Aislamientos de Madres
comunitarias
28 1 0 Aislamientos de Madres
comunitarias
36 1 0 Aislamientos de Madres
comunitarias
37 1 0 Aislamientos de Madres
comunitarias
41 0 1 Aislamientos de Madres
comunitarias
45 1 0 Aislamientos de Madres
comunitarias
52 0 1 Aislamientos de Madres
comunitarias
65 1 0 Aislamientos de Madres
comunitarias
68 0 1 Aislamientos de Madres
comunitarias
94 1 0 Aislamientos de Madres
comunitarias

74
95 1 0 Aislamientos de Madres
comunitarias
104 1 0 Aislamientos de Madres
comunitarias
111 1 1 Aislamientos de Madres
comunitarias
115 1 0 Aislamientos de Madres
comunitarias
121 1 0 Aislamientos de Madres
comunitarias
128 0 1 Aislamientos de Madres
comunitarias
129 1 0 Aislamientos de Madres
comunitarias
130 1 0 Aislamientos de Madres
comunitarias
134 1 1 Aislamientos de Madres
comunitarias
135 1 0 Aislamientos de Madres
comunitarias
139 1 0 Aislamientos de Madres
comunitarias
148 1 0 Aislamientos de Madres
comunitarias
152 1 0 Aislamientos de Madres
comunitarias
153 1 0 Aislamientos de Madres
comunitarias
154 1 0 Aislamientos de Madres
comunitarias
159 1 0 Aislamientos de Madres
comunitarias
164 0 1 Aislamientos de Madres
comunitarias

75
166 1 0 Aislamientos de Madres
comunitarias
168 0 1 Aislamientos de Madres
comunitarias
169 1 0 Aislamientos de Madres
comunitarias
173 1 0 Aislamientos de Madres
comunitarias
174 1 0 Aislamientos de Madres
comunitarias
187 1 0 Aislamientos de Madres
comunitarias
189 0 1 Aislamientos de Madres
comunitarias
195 1 0 Aislamientos de Madres
comunitarias
196 1 0 Aislamientos de Madres
comunitarias
197 0 1 Aislamientos de Madres
comunitarias
198 1 0 Aislamientos de Madres
comunitarias
200 1 0 Aislamientos de Madres
comunitarias
206 1 0 Aislamientos de Madres
comunitarias
210 0 1 Aislamientos de Madres
comunitarias
212 1 0 Aislamientos de Madres
comunitarias
217 1 0 Aislamientos de Madres
comunitarias
231 1 0 Aislamientos de Madres
comunitarias

76
235 1 0 Aislamientos de Madres
comunitarias
243 1 0 Aislamientos de Madres
comunitarias
256 1 0 Aislamientos de Madres
comunitarias
257 1 0 Aislamientos de Madres
comunitarias
259 1 0 Aislamientos de Madres
comunitarias
269 0 1 Aislamientos de Madres
comunitarias
272 1 1 Aislamientos de Madres
comunitarias
274 1 0 Aislamientos de Madres
comunitarias
275 1 0 Aislamientos de Madres
comunitarias
283 1 0 Aislamientos de Madres
comunitarias
285 1 0 Aislamientos de Madres
comunitarias
290 0 1 Aislamientos de Madres
comunitarias
291 0 1 Aislamientos de Madres
comunitarias
298 0 1 Aislamientos de Madres
comunitarias
299 1 0 Aislamientos de Madres
comunitarias
300 1 0 Aislamientos de Madres
comunitarias
301 1 0 Aislamientos de Madres
comunitarias

77
302 1 0 Aislamientos de Madres
comunitarias
303 1 0 Aislamientos de Madres
comunitarias
306 1 0 Aislamientos de Madres
comunitarias
312 1 0 Aislamientos de Madres
comunitarias
313 1 0 Aislamientos de Madres
comunitarias
321 1 0 Aislamientos de Madres
comunitarias
324 1 0 Aislamientos de Madres
comunitarias
328 1 0 Aislamientos de Madres
comunitarias
339 1 1 Aislamientos de Madres
comunitarias
346 1 0 Aislamientos de Madres
comunitarias
348 1 0 Aislamientos de Madres
comunitarias
355 1 0 Aislamientos de Madres
comunitarias
359 1 0 Aislamientos de Madres
comunitarias
364 1 0 Aislamientos de Madres
comunitarias
370 1 0 Aislamientos de Madres
comunitarias
371 1 1 Aislamientos de Madres
comunitarias
376 1 0 Aislamientos de Madres
comunitarias

78
388 0 1 Aislamientos de Madres
comunitarias
390 1 1 Aislamientos de Madres
comunitarias
391 0 1 Aislamientos de Madres
comunitarias
398 1 0 Aislamientos de Madres
comunitarias
402 1 0 Aislamientos de Madres
comunitarias
403 1 0 Aislamientos de Madres
comunitarias
405 1 0 Aislamientos de Madres
comunitarias
(1) Positivo.
(2) Negativo.
Anexo B. Prevalencia de estado de portador para la población total.
Muestra Estado de
Portador
Fosa nasal 18,5%
Mano 3,5%
Total muestra 22%
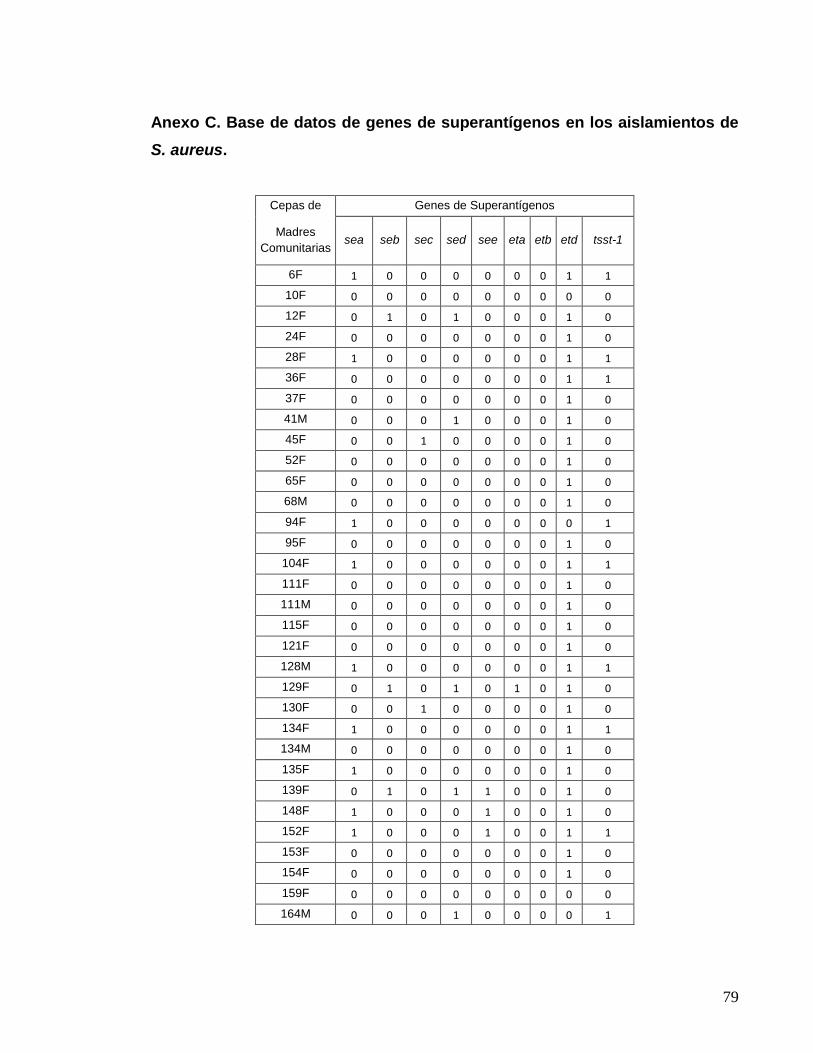
79
Anexo C. Base de datos de genes de superantígenos en los aislamientos de
S. aureus.
Cepas de Genes de Superantígenos
Madres
Comunitarias sea seb sec sed see eta etb etd tsst-1
6F 1 0 0 0 0 0 0 1 1
10F 0 0 0 0 0 0 0 0 0
12F 0 1 0 1 0 0 0 1 0
24F 0 0 0 0 0 0 0 1 0
28F 1 0 0 0 0 0 0 1 1
36F 0 0 0 0 0 0 0 1 1
37F 0 0 0 0 0 0 0 1 0
41M 0 0 0 1 0 0 0 1 0
45F 0 0 1 0 0 0 0 1 0
52F 0 0 0 0 0 0 0 1 0
65F 0 0 0 0 0 0 0 1 0
68M 0 0 0 0 0 0 0 1 0
94F 1 0 0 0 0 0 0 0 1
95F 0 0 0 0 0 0 0 1 0
104F 1 0 0 0 0 0 0 1 1
111F 0 0 0 0 0 0 0 1 0
111M 0 0 0 0 0 0 0 1 0
115F 0 0 0 0 0 0 0 1 0
121F 0 0 0 0 0 0 0 1 0
128M 1 0 0 0 0 0 0 1 1
129F 0 1 0 1 0 1 0 1 0
130F 0 0 1 0 0 0 0 1 0
134F 1 0 0 0 0 0 0 1 1
134M 0 0 0 0 0 0 0 1 0
135F 1 0 0 0 0 0 0 1 0
139F 0 1 0 1 1 0 0 1 0
148F 1 0 0 0 1 0 0 1 0
152F 1 0 0 0 1 0 0 1 1
153F 0 0 0 0 0 0 0 1 0
154F 0 0 0 0 0 0 0 1 0
159F 0 0 0 0 0 0 0 0 0
164M 0 0 0 1 0 0 0 0 1

80
166F 0 1 0 1 0 0 0 0 0
168M 0 1 0 1 0 0 0 0 0
169F 0 0 0 0 0 0 0 0 1
173F 0 0 0 0 0 0 0 0 1
174F 0 0 0 0 0 0 0 0 0
187F 0 0 0 0 0 0 0 0 0
189M 0 0 0 0 0 0 0 0 0
195F 0 0 0 0 0 0 0 0 0
196F 0 0 0 1 0 0 0 0 0
197M 0 0 0 1 0 0 0 1 0
198F 0 0 0 0 0 0 0 0 0
200F 0 0 0 0 0 0 0 1 0
206F 0 0 0 0 0 0 0 0 0
210M 1 0 1 0 0 0 0 0 0
212F 0 0 0 0 0 0 0 0 0
217F 0 0 0 0 0 0 0 0 0
231F 0 0 0 0 0 0 0 0 0
235F 0 0 0 0 0 1 0 0 0
243F 0 0 0 0 0 0 0 1 0
256F 0 0 0 0 0 0 0 1 0
257F 0 0 0 0 0 0 0 1 0
259F 0 0 0 0 0 0 0 1 0
269M 0 0 0 0 0 0 0 1 0
272F 0 0 0 0 0 0 0 1 0
272M 1 0 1 0 1 0 0 1 0
274F 0 0 0 0 0 0 0 0 0
275F 0 0 0 0 0 1 0 0 0
283F 0 0 0 0 0 0 0 0 0
285F 0 0 1 0 0 0 0 1 0
290M 0 0 0 0 0 0 0 0 0
291M 0 0 0 0 0 0 0 0 1
298M 0 0 1 0 0 0 0 0 0
299F 0 0 0 0 0 0 0 1 0
300F 0 0 0 0 0 0 0 0 0
301F 0 0 0 0 0 0 0 0 0
302F 0 0 1 0 0 0 0 1 0
303F 0 0 0 0 0 0 0 0 0
306F 0 0 0 0 0 0 0 0 0
312F 1 0 0 0 0 0 0 0 1
313F 1 0 0 0 0 0 0 0 1

81
321F 0 0 0 0 0 0 0 0 0
324F 1 0 0 0 0 0 0 0 1
326F 0 0 0 0 0 0 0 0 0
328F 0 0 0 0 0 0 0 0 0
339F 1 0 0 0 0 0 0 0 1
339M 1 0 0 0 0 0 0 0 1
346F 0 1 0 1 0 1 0 0 0
348F 0 0 0 0 0 0 0 0 1
355F 0 0 0 0 0 0 0 1 0
359F 0 1 0 1 0 0 0 0 0
364F 0 0 0 0 0 0 0 0 0
370F 0 0 0 0 0 0 0 0 0
371F 0 0 1 0 0 0 0 1 0
371M 0 0 1 0 0 0 0 0 0
376F 0 0 0 0 0 0 0 0 0
388M 1 0 0 0 0 0 0 1 1
390F 0 0 0 0 0 0 0 0 0
390M 0 0 1 0 0 0 0 0 0
391M 0 0 1 0 0 0 0 0 0
398F 0 0 0 1 0 0 0 0 1
402F 0 1 0 1 0 0 0 1 0
403F 0 0 0 0 0 0 0 1 1
405F 1 0 0 0 0 0 0 0 1
(1) Positivo.
(2) Negativo.
Anexo D. Base de datos de número de genes de superantígenos detectados
en los aislamientos de S. aureus.
Cepa 1 Gene 2 Gene 3 Gene 4 Gene Total
6F 0 0 1 0 3
10F 0 0 0 0 0
12F 0 0 1 0 3
24F 1 0 0 0 1
28F 0 0 1 0 3

82
36F 0 1 0 0 2
37F 1 0 0 0 1
41M 0 1 0 0 2
45F 0 1 0 0 2
52F 1 0 0 0 1
65F 1 0 0 0 1
68M 1 0 0 0 1
94F 0 1 0 0 2
95F 1 0 0 0 1
104F 0 0 1 0 3
111F 1 0 0 0 1
111M 1 0 0 0 1
115F 1 0 0 0 1
121F 1 0 0 0 1
128M 0 0 1 0 3
129F 0 0 0 1 4
130F 0 1 0 0 2
134F 0 0 1 0 3
134M 1 0 0 0 1
135F 0 1 0 0 2
139F 0 0 0 1 4
148F 0 0 1 0 3
152F 0 0 0 1 4
153F 1 0 0 0 1
154F 1 0 0 0 1
159F 0 0 0 0 0
164M 0 1 0 0 2
166F 0 1 0 0 2
168M 0 1 0 0 2
169F 1 0 0 0 1
173F 1 0 0 0 1
174F 0 0 0 0 0
187F 0 0 0 0 0
189M 0 0 0 0 0
195F 0 0 0 0 0
196F 1 0 0 0 1
197M 0 1 0 0 2
198F 0 0 0 0 0
200F 1 0 0 0 1
206F 0 0 0 0 0

83
210M 0 1 0 0 2
212F 0 0 0 0 0
217F 0 0 0 0 0
231F 0 0 0 0 0
235F 1 0 0 0 1
243F 1 0 0 0 1
256F 1 0 0 0 1
257F 1 0 0 0 1
259F 1 0 0 0 1
269M 1 0 0 0 1
272F 1 0 0 0 1
272M 0 0 0 1 4
274F 0 0 0 0 0
275F 1 0 0 0 1
283F 0 0 0 0 0
285F 0 1 0 0 2
290M 0 0 0 0 0
291M 1 0 0 0 1
298M 1 0 0 0 1
299F 1 0 0 0 1
300F 0 0 0 0 0
301F 0 0 0 0 0
302F 0 1 0 0 2
303F 0 0 0 0 0
306F 0 0 0 0 0
312F 0 1 0 0 2
313F 0 1 0 0 2
321F 0 0 0 0 0
324F 0 1 0 0 2
326F 0 0 0 0 0
328F 0 0 0 0 0
339F 0 1 0 0 2
339M 0 1 0 0 2
346F 0 0 1 0 3
348F 1 0 0 0 1
355F 1 0 0 0 1
359F 0 1 0 0 2
364F 0 0 0 0 0
370F 0 0 0 0 0
371F 0 1 0 0 2
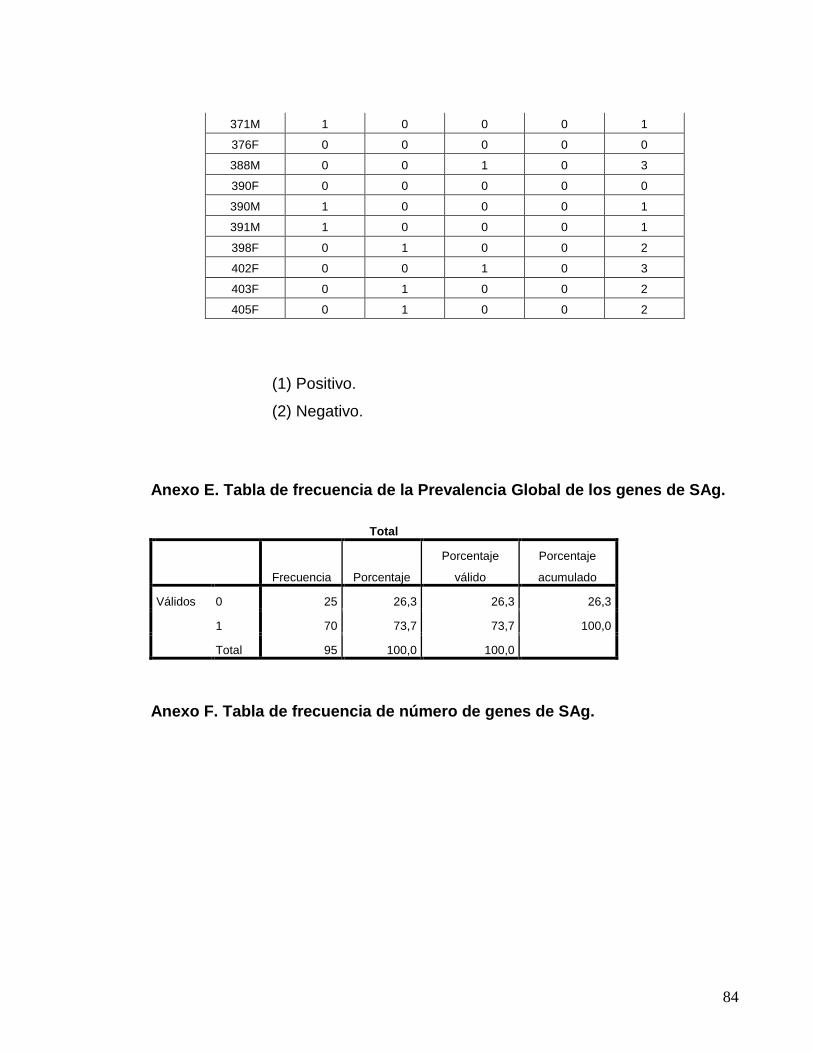
84
371M 1 0 0 0 1
376F 0 0 0 0 0
388M 0 0 1 0 3
390F 0 0 0 0 0
390M 1 0 0 0 1
391M 1 0 0 0 1
398F 0 1 0 0 2
402F 0 0 1 0 3
403F 0 1 0 0 2
405F 0 1 0 0 2
(1) Positivo.
(2) Negativo.
Anexo E. Tabla de frecuencia de la Prevalencia Global de los genes de SAg.
Total
Frecuencia Porcentaje
Porcentaje
válido
Porcentaje
acumulado
Válidos 0 25 26,3 26,3 26,3
1 70 73,7 73,7 100,0
Total 95 100,0 100,0
Anexo F. Tabla de frecuencia de número de genes de SAg.

85
Total
Frecuencia Porcentaje
Porcentaje
válido
Porcentaje
acumulado
Válidos 0 25 26,3 26,3 26,3
1 33 34,7 34,7 61,7
2 23 24,2 24,2 86,2
3 9 10 10 95,7
4 4 4,2 4,2 100,0
Total 94 100,0 100,0
Anexo G. Tablas de frecuencia para la prevalencia individual de los genes de
SAg en los aislamientos de S. aureus.
sea (102)
Frecuencia Porcentaje
Porcentaje
válido
Porcentaje
acumulado
Válidos 0 77 81,1 81,1 81,1
1 18 18,9 18,9 100,0
Total 95 100,0 100,0
seb (164)
Frecuencia Porcentaje
Porcentaje
válido
Porcentaje
acumulado
Válidos 0 87 91,6 91,6 91,6
1 8 8,4 8,4 100,0
Total 95 100,0 100,0

86
sec (451)
Frecuencia Porcentaje
Porcentaje
válido
Porcentaje
acumulado
Válidos 0 84 88,4 88,4 88,4
1 11 11,6 11,6 100,0
Total 95 100,0 100,0
sed (278)
Frecuencia Porcentaje
Porcentaje
válido
Porcentaje
acumulado
Válidos 0 82 86,3 86,3 86,3
1 13 13,7 13,7 100,0
Total 95 100,0 100,0
see (209)
Frecuencia Porcentaje
Porcentaje
válido
Porcentaje
acumulado
Válidos 0 91 95,8 95,8 95,8
1 4 4,2 4,2 100,0
Total 95 100,0 100,0
etd (376)
Frecuencia Porcentaje
Porcentaje
válido
Porcentaje
acumulado
Válidos 0 50 52,6 52,6 52,6
1 45 47,4 47,4 100,0
Total 95 100,0 100,0

87
tsst-1 (326)
Frecuencia Porcentaje
Porcentaje
válido
Porcentaje
acumulado
Válidos 0 73 76,8 76,8 76,8
1 22 23,2 23,2 100,0
Total 95 100,0 100,0
eta (93)
Frecuencia Porcentaje
Porcentaje
válido
Porcentaje
acumulado
Válidos 0 91 95,8 95,8 95,8
1 4 4,2 4,2 100,0
Total 95 100,0 100,0
Anexo H. Tablas de frecuencia de asociación de los genes de SAg.
Cepas de Genes de Superantígenos
Madres
Comunitarias sea seb sec sed see eta etb etd tsst-1 Combinaciones de genes
6F 1 0 0 0 0 0 0 1 1 sea-etd-tsst-1
10F 0 0 0 0 0 0 0 0 0 0
12F 0 1 0 1 0 0 0 1 0 seb-sed-etd
24F 0 0 0 0 0 0 0 1 0 etd
28F 1 0 0 0 0 0 0 1 1 sea-etd-tsst-1
36F 0 0 0 0 0 0 0 1 1 etd-tsst-1
37F 0 0 0 0 0 0 0 1 0 etd
41M 0 0 0 1 0 0 0 1 0 sed-etd
45F 0 0 1 0 0 0 0 1 0 sec-etd
52F 0 0 0 0 0 0 0 1 0 etd
65F 0 0 0 0 0 0 0 1 0 etd
68M 0 0 0 0 0 0 0 1 0 etd
94F 1 0 0 0 0 0 0 0 1 sea-tsst-1
95F 0 0 0 0 0 0 0 1 0 etd
104F 1 0 0 0 0 0 0 1 1 sea-etd-tsst-1

88
111F 0 0 0 0 0 0 0 1 0 etd
111M 0 0 0 0 0 0 0 1 0 etd
115F 0 0 0 0 0 0 0 1 0 etd
121F 0 0 0 0 0 0 0 1 0 etd
128M 1 0 0 0 0 0 0 1 1 sea-etd-tsst-1
129F 0 1 0 1 0 1 0 1 0 seb-sed-eta-etd
130F 0 0 1 0 0 0 0 1 0 sec-etd
134F 1 0 0 0 0 0 0 1 1 sea.etd.tsst-1
134M 0 0 0 0 0 0 0 1 0 etd
135F 1 0 0 0 0 0 0 1 0 sea-etd
139F 0 1 0 1 1 0 0 1 0 seb-sed-see-etd
148F 1 0 0 0 1 0 0 1 0 sea-see-etd
152F 1 0 0 0 1 0 0 1 1 sea-see-etd-tsst-1
153F 0 0 0 0 0 0 0 1 0 etd
154F 0 0 0 0 0 0 0 1 0 etd
159F 0 0 0 0 0 0 0 0 0 0
164M 0 0 0 1 0 0 0 0 1 sed-tsst-1
166F 0 1 0 1 0 0 0 0 0 seb-sed
168M 0 1 0 1 0 0 0 0 0 seb-sed
169F 0 0 0 0 0 0 0 0 1 tsst-1
173F 0 0 0 0 0 0 0 0 1 tsst-1
174F 0 0 0 0 0 0 0 0 0 0
187F 0 0 0 0 0 0 0 0 0 0
189M 0 0 0 0 0 0 0 0 0 0
195F 0 0 0 0 0 0 0 0 0 0
196F 0 0 0 1 0 0 0 0 0 sed
197M 0 0 0 1 0 0 0 1 0 sed-etd
198F 0 0 0 0 0 0 0 0 0 0
200F 0 0 0 0 0 0 0 1 0 etd
206F 0 0 0 0 0 0 0 0 0 0
210M 1 0 1 0 0 0 0 0 0 sea-sec
212F 0 0 0 0 0 0 0 0 0 0
217F 0 0 0 0 0 0 0 0 0 0
231F 0 0 0 0 0 0 0 0 0 0
235F 0 0 0 0 0 1 0 0 0 eta
243F 0 0 0 0 0 0 0 1 0 etd
256F 0 0 0 0 0 0 0 1 0 etd
257F 0 0 0 0 0 0 0 1 0 etd
259F 0 0 0 0 0 0 0 1 0 etd
269M 0 0 0 0 0 0 0 1 0 etd

89
272F 0 0 0 0 0 0 0 1 0 etd
272M 1 0 1 0 1 0 0 1 0 sea-sec-see-etd
274F 0 0 0 0 0 0 0 0 0 0
275F 0 0 0 0 0 1 0 0 0 eta
283F 0 0 0 0 0 0 0 0 0 0
285F 0 0 1 0 0 0 0 1 0 sec-etd
290M 0 0 0 0 0 0 0 0 0 0
291M 0 0 0 0 0 0 0 0 1 tsst-1
298M 0 0 1 0 0 0 0 0 0 sec
299F 0 0 0 0 0 0 0 1 0 etd
300F 0 0 0 0 0 0 0 0 0 0
301F 0 0 0 0 0 0 0 0 0 0
302F 0 0 1 0 0 0 0 1 0 sec-etd
303F 0 0 0 0 0 0 0 0 0 0
306F 0 0 0 0 0 0 0 0 0 0
312F 1 0 0 0 0 0 0 0 1 sea-tsst-1
313F 1 0 0 0 0 0 0 0 1 sea-tsst-1
321F 0 0 0 0 0 0 0 0 0 0
324F 1 0 0 0 0 0 0 0 1 sea-tsst-1
326F 0 0 0 0 0 0 0 0 0 0
328F 0 0 0 0 0 0 0 0 0 0
339F 1 0 0 0 0 0 0 0 1 sea-tsst-1
339M 1 0 0 0 0 0 0 0 1 sea-tsst-1
346F 0 1 0 1 0 1 0 0 0 seb-sed-eta
348F 0 0 0 0 0 0 0 0 1 tsst-1
355F 0 0 0 0 0 0 0 1 0 etd
359F 0 1 0 1 0 0 0 0 0 seb-sed
364F 0 0 0 0 0 0 0 0 0 0
370F 0 0 0 0 0 0 0 0 0 0
371F 0 0 1 0 0 0 0 1 0 sec-etd
371M 0 0 1 0 0 0 0 0 0 sec
376F 0 0 0 0 0 0 0 0 0 0
388M 1 0 0 0 0 0 0 1 1 sea-etd-tsst-1
390F 0 0 0 0 0 0 0 0 0 0
390M 0 0 1 0 0 0 0 0 0 sec
391M 0 0 1 0 0 0 0 0 0 sec
398F 0 0 0 1 0 0 0 0 1 sed-tsst-1
402F 0 1 0 1 0 0 0 1 0 seb-sed-etd
403F 0 0 0 0 0 0 0 1 1 etd-tsst-1
405F 1 0 0 0 0 0 0 0 1 sea-tsst-1

90

91
Anexo I. Base de datos de los antimicrobianos.
C epa β- Lactamasa C efo xit ina Oxacilina Gentamicina C ipro f lo xacina Levo flo xacina M o xif lo xacina Eritro micina C lindamicinaQuinuprist ina/
D alfo prist inaLinezo lid T eico planina Vanco micina M ino ciclina T etraciclina N itro furanto í na R ifampicina
Trimet rop in/
Sulf almet oxazo l
6F 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
10F 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
12F 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
24F 1 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0
28F 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
36F 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
37F 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
41M 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
45F 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
52F 1 1 1 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0
65F 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
68M 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
94F 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
95F 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
104F 1 0 0 0 0 0 0 1 1 0 0 0 0 0 0 0 0 0
111F 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
111M 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
115F 1 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0
121F 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
128M 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
129F 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
130F 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
134F 1 0 0 0 0 0 0 1 1 0 0 0 0 0 0 0 0 0
134M 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
135F 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
139F 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
148F 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
152F 1 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0
153F 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
154F 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
159F 1 1 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
164M 1 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0
166F 1 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0
168M 1 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0
169F 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
173F 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
174F 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
187F 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
189M 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
195F 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
196F 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
197M 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
198F 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
200F 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
206F 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
210M 1 1 1 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0
212F 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
217F 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
231F 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
235F 1 0 0 0 0 0 0 1 1 0 0 0 0 0 0 0 0 0
243F 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
256F 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
257F 1 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0
259F 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
269M 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
272F 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
272M 1 1 1 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0
274F 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
275F 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
283F 1 1 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
285F 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
290M 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
291M 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
298M 1 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0
299F 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
300F 1 1 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
301F 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
302F 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
303F 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
306F 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
312F 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
313F 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
321F 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
324F 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
326F 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
328F 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
339F 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
339M 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
346F 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
348F 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
355F 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
359F 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
364F 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
370F 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
371F 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
371M 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
376F 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
388M 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
390F 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
390M 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
391M 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
398F 1 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0
402F 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
403F 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
405F 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0

92
Anexo J. Número de genes totales en los aislamientos de S. aureus según la resistencia a uno o más antimicrobianos.
Cepas de
Madres
Comunitarias
1 Gene 2 Gene 3 Gene 4 Gene Ningún Gen
6F 1 0 0 0 0
10F 1 0 0 0 0
12F 1 0 0 0 0
24F 0 1 0 0 0
28F 1 0 0 0 0
36F 1 0 0 0 0
37F 1 0 0 0 0
41M 1 0 0 0 0
45F 1 0 0 0 0
52F 0 0 0 1 0
65F 1 0 0 0 0
68M 1 0 0 0 0
94F 1 0 0 0 0
95F 1 0 0 0 0
104F 0 0 1 0 0
111F 1 0 0 0 0
111M 1 0 0 0 0
115F 0 1 0 0 0
121F 1 0 0 0 0
128M 1 0 0 0 0
129F 1 0 0 0 0
130F 1 0 0 0 0
134F 0 0 1 0 0
134M 1 0 0 0 0
135F 1 0 0 0 0
139F 0 0 0 0 0
148F 1 0 0 0 0
152F 0 1 0 0 0
153F 1 0 0 0 0
154F 1 0 0 0 0
159F 0 0 1 0 0
164M 0 1 0 0 0
166F 0 1 0 0 0
168M 0 1 0 0 0

93
169F 1 0 0 0 0
173F 1 0 0 0 0
174F 1 0 0 0 0
187F 1 0 0 0 0
189M 1 0 0 0 0
195F 1 0 0 0 0
196F 1 0 0 0 0
197M 1 0 0 0 0
198F 1 0 0 0 0
200F 0 0 0 0 1
206F 1 0 0 0 0
210M 0 0 0 1 0
212F 1 0 0 0 0
217F 1 0 0 0 0
231F 1 0 0 0 0
235F 0 0 1 0 0
243F 1 0 0 0 0
256F 1 0 0 0 0
257F 1 0 0 0 0
259F 1 0 0 0 0
269M 0 0 0 0 1
272F 1 0 0 0 0
272M 0 0 0 1 0
274F 1 0 0 0 0
275F 1 0 0 0 0
283F 0 0 1 0 0
285F 1 0 0 0 0
290M 1 0 0 0 0
291M 1 0 0 0 0
298M 0 1 0 0 0
299F 1 0 0 0 0
300F 0 0 1 0 0
301F 1 0 0 0 0
302F 1 0 0 0 0
303F 0 0 0 0 1
306F 1 0 0 0 0
312F 1 0 0 0 0
313F 1 0 0 0 0

94
321F 1 0 0 0 0
324F 1 0 0 0 0
326F 1 0 0 0 0
328F 1 0 0 0 0
339F 1 0 0 0 0
339M 1 0 0 0 0
346F 1 0 0 0 0
348F 1 0 0 0 0
355F 1 0 0 0 0
359F 1 0 0 0 0
364F 1 0 0 0 0
370F 1 0 0 0 0
371F 1 0 0 0 0
371M 1 0 0 0 0
376F 1 0 0 0 0
388M 1 0 0 0 0
390F 0 0 0 0 1
390M 1 0 0 0 0
391M 1 0 0 0 0
398F 0 1 0 0 0
402F 1 0 0 0 0
403F 0 0 0 0 1
405F 1 0 0 0 0
Total de
antimicrobianos
positivos
71 8 6 3 5
% Total 71% 9% 6% 3% 5%
Anexo K. Tablas de frecuencia para la prevalencia de resistencia en los aislamientos de S. aureus a diferentes antimicrobianos.
β- Lactamasa
Frecuencia Porcentaje
Porcentaje
válido
Porcentaje
acumulado
Válidos 0 6 6,3 6,3 6,3
1 89 93,7 93,7 100,0
Total 95 100,0 100,0

95
Tetraciclina
Frecuencia Porcentaje
Porcentaje
válido
Porcentaje
acumulado
Válidos 0 84 88,4 88,4 88,4
1 11 11,6 11,6 100,0
Total 95 100,0 100,0
Cefoxitina
Frecuencia Porcentaje
Porcentaje
válido
Porcentaje
acumulado
Válidos 0 89 93,7 93,7 93,7
1 6 6,3 6,3 100,0
Total 95 100,0 100,0
Oxacilina
Frecuencia Porcentaje
Porcentaje
válido
Porcentaje
acumulado
Válidos 0 89 93,7 93,7 93,7
1 6 6,3 6,3 100,0
Total 95 100,0 100,0
Eritromicina
Frecuencia Porcentaje
Porcentaje
válido
Porcentaje
acumulado
Válidos 0 91 95,8 95,8 95,8
1 4 4,2 4,2 100,0
Total 95 100,0 100,0
Clindamicina
Frecuencia Porcentaje
Porcentaje
válido
Porcentaje
acumulado
Válidos 0 92 96,8 96,8 96,8
1 3 3,2 3,2 100,0
Total 95 100,0 100,0

96
Anexo L. Tablas de contingencia para las asociaciones entre la detección de los genes de SAgs y la resistencia antimicrobiana.
Tabla de contingencia
β - Lactamasa
Total S R
Total SAg- 3 22 25
SAg+ 3 67 70
Total 6 89 95
Valor gl
Sig. asintótica
(bilateral)
Sig. exacta
(bilateral)
Sig. exacta
(unilateral)
Chi-cuadrado de Pearson 1,853a 1 ,173
Corrección por continuidadb ,778 1 ,378
Razón de verosimilitudes 1,643 1 ,200
Estadístico exacto de Fisher ,184 ,184
Asociación lineal por lineal 1,833 1 ,176
N de casos válidos 95
a. 2 casillas (50,0%) tienen una frecuencia esperada inferior a 5. La frecuencia mínima esperada es 1,58.
b. Calculado sólo para una tabla de 2x2.

97
Tabla de contingencia
Cefoxitina/
Oxacilina
Total S R
Total SAg- 22 3 25
% dentro de Total 88,0% 12,0% 100,0%
% dentro de Cefoxitina 24,7% 50,0% 26,3%
SAg+ 67 3 70
% dentro de Total 95,7% 4,3% 100,0%
% dentro de Cefoxitina 75,3% 50,0% 73,7%
Total Recuento 89 6 95
% dentro de Total 93,7% 6,3% 100,0%
% dentro de Cefoxitina 100,0% 100,0% 100,0%
Valor gl
Sig. asintótica
(bilateral)
Sig. exacta
(bilateral)
Sig. exacta
(unilateral)
Chi-cuadrado de Pearson 1,853a 1 ,173
Corrección por continuidadb ,778 1 ,378
Razón de verosimilitudes 1,643 1 ,200
Estadístico exacto de Fisher ,184 ,184
Asociación lineal por lineal 1,833 1 ,176
N de casos válidos 95
a. 2 casillas (50,0%) tienen una frecuencia esperada inferior a 5. La frecuencia mínima esperada es 1,58.
b. Calculado sólo para una tabla de 2x2.

98
Tabla de contingencia
Eritromicina
Total S R
Total 0 SAg- 25 0 25
% dentro de Total 100,0% ,0% 100,0%
% dentro de Eritromicina 27,5% ,0% 26,3%
1 SAg+ 66 4 70
% dentro de Total 94,3% 5,7% 100,0%
% dentro de Eritromicina 72,5% 100,0% 73,7%
Total Recuento 91 4 95
% dentro de Total 95,8% 4,2% 100,0%
% dentro de Eritromicina 100,0% 100,0% 100,0%
Pruebas de chi-cuadrado
Valor gl
Sig. asintótica
(bilateral)
Sig. exacta
(bilateral)
Sig. exacta
(unilateral)
Chi-cuadrado de Pearson 1,491a 1 ,222
Corrección por
continuidadb
,411 1 ,521
Razón de verosimilitudes 2,505 1 ,113
Estadístico exacto de
Fisher
,570 ,288
Asociación lineal por lineal 1,476 1 ,224
N de casos válidos 95
a. 2 casillas (50,0%) tienen una frecuencia esperada inferior a 5. La frecuencia mínima esperada es
1,05.
b. Calculado sólo para una tabla de 2x2.
Tabla de contingencia
Clindamicina
Total S R
Total 0 SAg- 25 0 25
% dentro de Total 100,0% ,0% 100,0%
% dentro de Clindamicina 27,2% ,0% 26,3%
1 SAg+ 67 3 70

99
% dentro de Total 95,7% 4,3% 100,0%
% dentro de Clindamicina 72,8% 100,0% 73,7%
Total Recuento 92 3 95
% dentro de Total 96,8% 3,2% 100,0%
% dentro de Clindamicina 100,0% 100,0% 100,0%

100
Pruebas de chi-cuadrado
Valor gl
Sig. asintótica
(bilateral)
Sig. exacta
(bilateral)
Sig. exacta
(unilateral)
Chi-cuadrado de Pearson 1,106a 1 ,293
Corrección por continuidadb ,149 1 ,700
Razón de verosimilitudes 1,867 1 ,172
Estadístico exacto de Fisher ,564 ,395
Asociación lineal por lineal 1,095 1 ,295
N de casos válidos 95

101
a. 2 casillas (50,0%) tienen una frecuencia esperada inferior a 5. La frecuencia mínima esperada es ,79.
b. Calculado sólo para una tabla de 2x2.
Tabla de contingencia
Tetraciclina
Total S R
Total 0 SAg- 25 0 25
% dentro de Total 100,0% ,0% 100,0%
% dentro de Tetraciclina 29,8% ,0% 26,3%
1 SAg+ 59 11 70
% dentro de Total 84,3% 15,7% 100,0%
% dentro de Tetraciclina 70,2% 100,0% 73,7%
Total Recuento 84 11 95
% dentro de Total 88,4% 11,6% 100,0%
% dentro de Tetraciclina 100,0% 100,0% 100,0%
Pruebas de chi-cuadrado
Valor gl
Sig. asintótica
(bilateral)
Sig. exacta
(bilateral)
Sig. exacta
(unilateral)
Chi-cuadrado de Pearson 4,443a 1 ,035
Corrección por continuidadb 3,041 1 ,081
Razón de verosimilitudes 7,219 1 ,007
Estadístico exacto de Fisher ,062 ,028
Asociación lineal por lineal 4,396 1 ,036
N de casos válidos 95
a. 1 casillas (25,0%) tienen una frecuencia esperada inferior a 5. La frecuencia mínima esperada es 2,89.
b. Calculado sólo para una tabla de 2x2.

102
![ENSAYOS DEL BIOFILM (PSEUDOMONA SP. Y … final.pdf · agentes contaminantes nuevos en un ecosistema en particular [1,12]. X Además, poseen genes que codifican para enzimas, con](https://static.fdocuments.ec/doc/165x107/5be75bef09d3f2d66c8bd78c/ensayos-del-biofilm-pseudomona-sp-y-finalpdf-agentes-contaminantes-nuevos.jpg)