
Ciencias Agronómicas - UNRXX).pdf · Los cultivos de cobertura en la horticultura familiar ... La...
86
Ciencias Agronómicas ISSN Nº 1853-4333 ISSN On-Line: 2250-8872 Facultad de Ciencias Agrarias - Universidad Nacional de Rosario Campo Experimental Villarino C.C. Nro. 14 (S 2125 ZAA) - Zavalla - Santa Fe - Argentina Telefax 0341 - 4970080 - 085 - [email protected] - www.fcagr.unr.edu.ar [email protected] Número XX Año 12 - 2012
Transcript of Ciencias Agronómicas - UNRXX).pdf · Los cultivos de cobertura en la horticultura familiar ... La...

Ciencias Agronómicas
ISSN Nº 1853-4333ISSN On-Line: 2250-8872
Facultad de Ciencias Agrarias - Universidad Nacional de RosarioCampo Experimental Villarino C.C. Nro. 14 (S 2125 ZAA) - Zavalla - Santa Fe - ArgentinaTelefax 0341 - 4970080 - 085 - [email protected] - www.fcagr.unr.edu.ar
Número XXAño 12 - 2012

Editor PrincipalDra. Susana Rosenstein
Secretario TécnicoIng. Agr. Néstor Di Leo
Conformación del Comité Asesor de la Revista Dr Christophe Jacques AlbaladejoINRA - Universidad Nacional del SurDr René BenavídezUniversidad Nacional de Entre RíosDra Elsa CamadroUniversidad Nacional de Mar del Plata Dra Silvia CloquellUniversidad Nacional de Rosario Dr Enrique CointryUniversidad Nacional de Rosario Dr Osvaldo FernándezUniversidad Nacional del SurDra María Teresa FontUniversidad Nacional de RosarioIng. Agr. (Ph D) Claudio GalmariniINTA - Universidad Nacional de CuyoDr Marcelo HelgueraINTA - EEA Marcos JuárezIng. Agr (Ph D) Daniel Ivan LeskovarUniversidad de TexasDr Sergio Montico Universidad Nacional de RosarioDr Juan Pablo Ortiz Universidad Nacional de RosarioDra Rossana PioliUniversidad Nacional de RosarioDr Eduardo Puricelli Universidad Nacional de RosarioIng. Agr (Ph D) Carlos F. QuirósUniversidad de California Lic. (Ms Sc) María Isabel Tort.INTA - Coordinación Nac. de Transferencia y ExtensiónDr Francisco SantiniUniversidad Nacional de Mar del Plata Dr Ivan TirantiUniversidad Nacional de Río CuartoDr Abelardo VegettiUniversidad Nacional del Litoral
Diseño y Diagramación:DG Juan Manuel VázquezÁrea Prensa y Difusión - Secretaría de Extensión FCA - UNR
Coordinadora Sra. María Andrea BarreraSecretaría de Investigaciones
Correcciones del InglésTrad. Gabriela M. VenturiCátedra de Inglés
Facultad de Ciencias Agrarias Universidad Nacional de RosarioCampo Experimental Villarino - C.C. Nro. 14 (S 2125 ZAA) Zavalla - Santa Fe - Argentina
Telefax 0341 - 4970080 - [email protected] - www.fcagr.unr.edu.ar
ro Año 12 - N 20 - 2012 - Páginas tipeadas por los Autores.
Impreso en Argentina - Printed in ArgentinaUNR EditoraEditorial de la Universidad Nacional de RosarioSecretaría de ExtensiónUrquiza 2050 - S2000AOB - Rosario - Santa Fe - ArgentinaAño 12 - Nº 20 - Julio - Diciembre de 20121º Edición de 150 ejemplaresPáginas tipeadas por los Autores.
Se prohibe la reproducción total o parcial, el almacenamiento, el alquiler, la transmisión o la transformación del contenido de esta Revista de Investigaciones, en cualquier forma o por cualquier medio, sea electrónico o mecánico, mediante fotocopias, digitalización u otros métodos, sin el permiso previo y escrito de los autores.Su infracción está penada por las leyes 11.723 y 25.446
ISSN Nº 1853-4333ISSN On-Line: 2250-8872

Autoridades de la Facultad de Ciencias Agrarias
DecanatoIng.Agr. MSc. Guillermo MONTERO
VicedecanatoIng.Agr. MSc. Teresa Mónica QÜESTA
Secretaría de Asuntos AcadémicosSecretaria:Ing.Agr. Silvana SETASubsecretaria:Med. Vet. MSc. Griselda MUÑOZ
Secretaría de InvestigacionesSecretaria:Ing. Agr. Marta PANELOSubsecretaria:Ing. Agr. MSc Miriam E. INCREMONA
Secretaría de Extensión UniversitariaSecretario:Ing. Agr. MSc. Daniel CAMPAGNASubsecretaria:Dra. Susana ROSENSTEINSecretario Técnico:Ing. Agr. José María ARREGUI
Secretaría de Asuntos FinancierosSecretario:C.P.N. Fernando AMELONGSubsecretaria:Ing.Agr. MSc. Susana ZULIANI
Secretaría de Asuntos EstudiantilesSecretario:Tec. Agr. Patricio MARENGHINISubsecretario:Sr. Federico LEON
Secretaría de Relaciones InternacionalesSecretario:Dr. Hugo PERMINGEATCoordinadora:Lic. Aneris CAO
Dirección del Campo ExperimentalDirector:Ing. Agr. Pablo PALAZZESIAsistente Técnico:Ing. Agr. Cecilia VIGNA
Dirección Escuela de PosgradoDirector:Dr. Juan Pablo ORTIZCoordinadora:Dra. Raquel BENAVIDEZAsitencia Técnica:Dra Juliana STEIN
Dirección general de AdministraciónSra. María Teresa SCARPELLO
Ciencias Agronómicas - Año 12 - 2012Revista XIX - -02-

Índice
Nota del Editor
Artículos Originales
Incidencia del ambiente y la revitalización en las pérdidas poscosecha de hortalizas de hoja, fruto y raízFIRPO, I.; ROTONDO, R.; FERRATTO, J.; GRASSO, R.; TREVISAN, A.; RODRIGUEZ, M.
Los cultivos de cobertura en la horticultura familiar urbana: una oportunidad para mejorar la formación del estudiante de agronomíaMILO VACCARO, M.; MUÑOZ, G.; TORRES, C.; COURETOT, J.
Desarrollo de una base de datos genéticos para la caracterización del germoplasma argentino de sojaOLSINA, C.; CAIRO, C.; PESSINO, S.
La cuestión curricular en la Facultad de Ciencias Agrarias de la UNRCAVALLI, A. y ROSENSTEIN, S.
Resúmenes
III Ciclo de Seminarios sobre avances en la caracterización genética y molecular de la apomixis en gramíneas forrajerasPESSINO, S.C.; ORTIZ, J.P.A.
Diplosporia concomitante con aposporia en Paspalum L.HONFI, AI; QUARIN, CL.
Relación entre el contenido de ADN y las características biológicas en PaspalumGALDEANO, F.; URBANI, M.; HONFI, A.; MATZK, F.; QUARIN, C.L.
Estudio de la diversidad contenida en poblaciones diploides y tetraploides de Paspalum simplex que crecen en forma simpátrica y alopátrica BRUGNOLI, E.A.; URBANI, M.H.; QUARIN, C.L.; MARTÍNEZ, E.J.; ACUÑA, C.A.
Caracterización del comportamiento reproductivo de citotipos diploides de Paspalum rufumDELGADO, L.; SARTOR, M.; QUARIN C.; ESPINOZA, F.; ORTIZ, J.P.
Mejoramiento genético de especies de Paspalum: una actualizaciónACUÑA, C.A.; URBANI, M.H.; QUARIN, C.L.
Transferencia generalizada de genes desde genotipos apomícticos a una población sexual en el grupo plicatula de PaspalumNOVO, P.E.; ESPINOZA, F.; QUARIN, C.L.
Obtención de variabilidad genética en los tetraploides sexuales de Paspalum notatum a partir de ecotipos tetraploides apomícticosZILLI, A.; BARONE, J.O.; RÍOS, E.F.; QUARIN, C.L.; ACUÑA, C.A.; MARTÍNEZ, E.J.
Avances en la caracterización genética de Acroceras macrum Stapf.FERRARI USANDIZAGA, S.C.; MARTÍNEZ, E.J.; QUARIN, C.L.; ZILLI, A.; BRUGNOLLI, E. A.; RANDAZZO, C.; PAGANO, E.N.; ACUÑA, C.A.
-03-
05
07
15
23
41
45
47
49
51
53
55
57
59
61
Ciencias Agronómicas - Año 12 - 2012Revista XX -
Caracterización de secuencias, mapeo in silico y análisis de metilación de secuencias ligadas a la aposporía en Paspalum notatumORTIZ, J.P.A.; PODIO, M.; RODRÍGUEZ, M.P.; FELITTI, S.; STEIN, J.; MARTÍNEZ, E.J.; SIENA, L.A.; QUARIN, C.L.; PESSINO, S.C.
Caracterización de la región genómica de la apomixis en especies del grupo plicatula de Paspalum L.AGUILERA, P.M.; QUARIN, C.L.; ORTIZ, J.P.A.; ESPINOZA, F.
Identificación de genes involucrados en el desarrollo de semillas de Paspalum notatumFELITTI, S.A.; ACUÑA, C.A.; ORTIZ, J.P.A.; QUARIN, C.L.
Caracterización de n19, un transcripto asociado con el desarrollo apospórico en Paspalum notatumSARTOR, M.; ESPINOZA, F.; SEIJO, G.; GONZÁLEZ, A.M.; PESSINO, S.
Análisis de expresión de genes exs en genotipos apomícticos y sexuales de Paspalum notatumPODIO, M.; DELGADO, L.; FELITTI S.; ORTIZ, J.P.A.
Caracterización de secuencias de metiltransferasas asociadas a la aposporía en Paspalum notatumSIENA, L.A.; SELVA, J.P.; ORTIZ, J.P.A.; ECHENIQUE, V.; LEBLANC, O.; PESSINO S.C.
Transformación biolística de Paspalum notatum tetraploide con un gen candidato para la apomixisMANCINI, M.; WOITOVICH, N.; PERMINGEAT, H.; ORTIZ, J.P.A.; QUARIN, C.; FELITTI, S.; PESSINO, S.C.
Apomixis en Eragrostis curvulaECHENIQUE, V.; PESSINO, S.C.; MEIER, M.; SELVA, J.P.; OCHOGAVÍA, A.; ZAPPACOSTA, D.; ROMERO, J.R.; RODRIGO, J.M.; GARBUS, I.; TERENTI, C.; DÍAZ, A.
Efectos del estrés abiótico sobre la expresión de la apomixis en pasto llorónRODRIGO, J.M.; ZAPPACOSTA, D.; BONAVENTURE, G.; ECHENIQUE, V.
Poblaciones de mapeo en pasto llorón - éxitos y fracasosZAPPACOSTA, D.; MEIER, M.; TERENTI, C.; RODRIGO, J.M.; ECHENIQUE, V.
Normas para la presentación de manuscritos
63
65
67
69
71
73
75
77
79
81
83


Estimados Lectores:
La Revista Ciencias Agronómicas sale a la luz con el número XX, correspondiente al segundo semestre del 2012, cumpliendo de esta manera en tiempo y forma con la periodicidad de la publicación. Contiene 4 artículos originales y una serie de resúmenes que se presentaron en el encuentro “III Ciclo de Seminarios sobre Avances en la Caracterización Genética y Molecular de la Apomixis en Gramíneas Forrajeras”, organizado por la Carrera de Doctorado en Ciencias Agrarias de nuestra Facultad, en el mes de octubre del corriente año.
El primer artículo aborda el problema del deterioro de la calidad durante la comercialización de hortalizas frescas. En este sentido, Firpo, I. y col evalúan la incidencia del ambiente y la revitalización en las pérdidas poscosecha de distintas hortalizas de hojas, fruto y raíz, considerando las siguientes variables: pérdida de peso por descarte, pérdida de agua y pérdidas totales en %. Los autores concluyen que las técnicas propuestas permiten disminuir las pérdidas poscosecha en la mayoría de las especies.
En el segundo, Milo Vaccaro, M., Muñoz, G., Torres, C. y Couretot, J., dan cuenta del trabajo conjunto entre nuestra Facultad y la Municipalidad de Rosario a través del Programa de Agricultura Urbana, mostrando una experiencia cuyos objetivos fueron: evaluar la eficiencia de los cultivos de cobertura para recuperar suelos urbanos, caracterizar la comunidad de huerteros que realiza las prácticas agroecológicas sobre esos suelos e identificar las competencias sociales y agronómicas desarrolladas por los estudiantes en un escenario de aprendizaje real y diverso. Los resultados muestran que los cultivos de cobertura son eficientes para recuperar suelos urbanos y la comunidad huertera posee conocimientos y experiencias que les posibilitan su incorporación como práctica agroecológica. Asimismo, los estudiantes mejoran su formación profesional al desarrollar competencias sociales y asumir un compromiso ético frente a situaciones de desigualdad social y baja calidad ambiental.
En el artículo siguiente, Olsina C., Cairo C., y Pessino S., proponen desarrollar un banco de datos genéticos basado en marcadores moleculares de SSR (Repeticiones de Secuencia Simple, o Microsatélites) y AFLP (polimorfismos en el largo de los fragmentos amplificados) para caracterizar el germoplasma argentino de soja, facilitar la identificación de variedades y asistir al mejoramiento genético. Los autores afirman que lograron seleccionaron un subconjunto de 20 marcadores SSR altamente informativos que permite discriminar cualquier
accesión del banco de manera simple y eficiente y que pueden ser utilizados en estudios rápidos de identificación o diferenciación de nuevos cultivares. Asimismo, los resultados sugieren que los marcadores AFLP reflejan fielmente las relaciones de parentesco entre las variedades, resultando apropiados para seleccionar materiales genéticamente divergentes.
En la cuarta contribución y desde un abordaje que rescata la investigación educativa, Cavalli, A y Rosenstein, S., analizan el modo en el que la Facultad de Ciencias Agrarias de la UNR, llevó adelante un cambio curricular tendiente a enfrentar el desafío que implicaba para las universidades atender a las crecientes exigencias sociales en un marco de restringidas políticas de financiamiento por parte del Estado a fines de la década de los 90. Y a la vez, el modo en el que se intentaron conciliar las contradicciones y los diferentes puntos de vista de los grupos de docentes y alumnos involucrados en el proceso de cambio. Las autoras concluyen que el nuevo Plan de estudios de la Facultad de Ciencias Agrarias constituye una síntesis que contiene elementos no sólo del grupo dominante sino también de los otros grupos o sectores presentes y que, aunque de manera contradictoria, incorpora innovaciones con respecto al curriculum anterior. Precisamente son estos cambios los que constituyen el eje de la polémica alrededor de los cuales se expresan las resistencias.
Finalmente, consideramos de suma importancia que esta Revista sea, además, un canal de difusión de los encuentros científicos que tienen lugar en nuestra y otras universidades, contribuyendo así a enriquecer el debate con nuevos aportes. En este caso, los resúmenes que se publican fueron presentados en el “III Ciclo de Seminarios sobre Avances en la Caracterización Genética y Molecular de la Apomixis en Gramíneas Forrajeras”, organizado con el objetivo de discutir los resultados del trabajo de distintos grupos de investigación argentinos en relación con la elucidación de las bases genéticas y moleculares de la apomixis en las gramíneas. Dicho encuentro contó con la participación de investigadores de las Universidades Nacionales del Nordeste, Misiones, del Sur, Rosario y del Instituto de Genética Vegetal de Perugia (Italia)
Agradeciendo nuevamente a los autores y colaboradores por sus aportes y a todo el equipo abocado a la realización de esta Revista por el tiempo y esfuerzo dedicados, nos despedimos hasta el próximo número, confiando en que seguiremos avanzando para brindar una Revista cada vez más excelente.
Cordialmente
Dra Susana RosenteinEditora Principal
-05-
Nota del Editor
Ciencias Agronómicas - Año 12 - 2012Revista XX -


-07-
Artículo Original / Recibido 21/12/2011 - Aceptado 25/10/2012
Incidencia del ambiente y la revitalización en laspérdidas poscosecha de hortalizas de hoja, fruto y raíz
Impact of environmental conditions and revitalization procedures onpost-harvest losses of leaf, fruit, and root vegetables
1 1 1-3 1 2 1Firpo, I. ; Rotondo, R. ; Ferratto, J. ; Grasso, R. ; Trevisan, A. ; Rodriguez, M.
1Cátedra de Cultivos Intensivos. Horticultura.2Cátedra de Estadística.3Investigador del CIUNR.
Facultad de Ciencias Agrarias.Universidad Nacional de Rosario.CC. Nº 14 (S2125ZAA). Zavalla. Santa Fe. Argentina
Las hortalizas frescas son productos perecederos, con importantes pérdidas y deterioros de la calidad durante la comercialización. La baja temperatura y alta humedad relativa lentifican los procesos fisiológicos. El objetivo fue evaluar la incidencia del ambiente y la revitalización en las pérdidas poscosecha de hortalizas de hojas, fruto y raíz. Se realizó en la Facultad de Ciencias Agrarias UNR (Zavalla, Santa Fe Argentina). Se evaluaron 2 factores con 2 niveles cada uno: 1) Ambientes de almacenaje: Cámara frigorífica 3,4 ºC; 97 % HR y ambiente natural (13 ºC media; 81 % HR; 2) Revitalización (inmersión en agua clorinada); con y sin revitalización. Se realizaron 4 repeticiones extraídas de cajones diferentes. Las muestras fueron: 5 plantas para cada tipo de lechuga (mantecosa, crespa y arrepollada); 1 planta en apio; 6 frutos en zapallito redondo de tronco y 3 raíces en zanahoria. Las variables medidas fueron: pérdida de peso por descarte, pérdida de agua y pérdidas totales, en %. En el análisis se aplicó un modelo de medidas repetidas en el tiempo con procedimiento MIXED de SAS. El almacenamiento en cámara redujo el porcentaje de pérdida en todas las variables y hortalizas evaluadas. La revitalización: en la mayoría de las especies, disminuyó la pérdida de agua en ambos ambientes; en el natural presentó significativamente menor descarte que el no revitalizado (10 % y 17% respectivamente). La lechuga arrepollada presentó mayor % de descarte con la revitalización, dado que se favoreció la presencia de alteraciones fisiológicas. Las técnicas propuestas permiten disminuir las pérdidas poscosecha en la mayoría de las especies.
Palabras clave:refrigeración-calidad posrecolección
Resumen
Fresh vegetables are perishable products, with significant loss and deterioration of quality during marketing. Low temperature and high relative humidity slow down physiological processes. The objective of this work was to evaluate the impact of environmental conditions and revitalization on post-harvest losses of leaf, fruit, and root vegetables. It was carried out in the College of Agricultural Sciences UNR (Rosario National University), in Zavalla, Santa Fe, Argentina. Two factors with 2 levels each were evaluated: 1) storage environments: 3,4 °C cold store at 97 % RH ;and natural environment (13 °C average temperature, 81 % RH; and 2) revitalization (immersion in chlorinated water): with and without revitalization. Four repetitions were performed with samples from different wooden boxes. The samples were: 5 plants for each type of lettuce (butterhead, crisp and iceberg), 1 celery plant, 6 round squash fruits and 3 carrot roots. The variables measured were: weight loss by discarding, water loss and total losses, in%. A model of repeated measures in time with the MIXED procedure of SAS was used for the analysis. Cold store reduced the percentage of loss in all variables and vegetables evaluated. Revitalization reduced water loss in most species in both environments; for vegetables stored in the natural environment, the percentage of loss by discarding was significantly lower for revitalized vegetables than for non-revitalized ones (10 % and 17 % respectively). Iceberg lettuce had a higher % of discarding with revitalization, since it favored the presence of physiological alterations. The techniques proposed are useful for reducing post-harvest losses in most species.
Keywords:cooling- post-harvest quality
Abstract
Introducción
Ciencias Agronómicas - Revista XX - Año 12 - 2012 / 007 - 013
se utilicen para reducir la velocidad de estos procesos. La cadena de distribución generalmente carece de instalaciones ideales para almacenar cada producto en condiciones adecuadas y requiere que todos los integrantes vean la necesidad de mejorar el manipuleo, la temperatura y la humedad relativa para limitar las pérdidas de calidad. Esta situación puede llevar al estrés fisiológico, pérdida de calidad y menor vida en estante (Paull, 1999).
La respiración es un proceso metabólico de fundamental importancia durante el período de la posrecolección. La velocidad a que respira un producto constituye un índice de la actividad metabólica de sus tejidos y es una guía útil para
Las frutas y hortalizas frescas son productos muy perecederos y por tal motivo, ocurren importantes pérdidas desde la cosecha hasta el consumidor final, principalmente en el proceso de acondicionamiento y comercialización (Kader, 2002). Las mismas, en países en vías de desarrollo, pueden representar entre el 20 y 50 %. En la región de Rosario (Pcia de Santa Fe) se realizaron estudios que indican que las pérdidas totales de lechuga son del 40,25 % (Mondino et al, 2007).
Los procesos fisiológicos más importantes durante la poscosecha son la respiración y la transpiración. La disminución de las pérdidas depende en gran parte de la capacidad y efectividad del sistema de comercialización, o de prácticas que

calcular la duración de su vida comercial (Wills et al, 1999; Robinson et al, 1975). Algunos órganos toman un color amarillento (degradación de la clorofila) y otros sufren cambios composicionales (contenidos de azúcares, sustancias nitrogenadas, etc.), haciendo que los productos pierdan valor comercial y aumente su descarte. El almacenamiento refrigerado se recomienda para muchos productos perecederos ya que retarda el envejecimiento causado por maduración, ablandamiento o cambios de textura, cambios metabólicos indeseables, producción de calor por respiración y la pérdida de humedad con el consiguiente marchitamiento (Handenburg et al., 1988).
La pérdida de agua por transpiración es otra de las causas del deterioro que se genera en el período de poscosecha. El principal componente de estos productos es el agua (80-95 % de su peso) y la pérdida de la misma no sólo trae como consecuencia una disminución del peso, sino también la formación de un producto flácido, poco atractivo y con disminución notable de la calidad
comercial (Nunes et al., 2009). La intensidad de la transpiración puede reducirse aumentando la humedad relativa, disminuyendo la temperatura del aire y .usando envolturas protectoras. La humedad relativa óptima, para disminuir la pérdida de agua por transpiración de la mayoría de los productos hortícolas para diferentes temperaturas es del 100 % (Handenburg et al., 1988). La inmersión de los productos en agua, durante un período determinado (que en el presente es llamado revitalización), antes de ser colocados en góndola es una técnica aconsejada para mantener la turgencia de los productos.
La evaluación del comportamiento de estas técnicas es importante a los fines de ofrecer alternativas de interés a nuestro sistema de comercialización.
El objetivo del ensayo fue evaluar la incidencia del almacenamiento refrigerado y la revitalización en las pérdidas poscosecha de lechuga mantecosa, de hoja y arrepollada, apio, zapallito y zanahoria.
El ensayo se realizó en la Facultad de Ciencias Agrarias de la UNR (Zavalla, Santa Fe, 33º01´LS y 60º53´LW). La mercadería evaluada fue traída directamente de un establecimiento productivo situado a 10 km de la ciudad de Rosario y solamente la zanahoria fue comprada en el Mercado de Concentración de Fisherton. La cosecha realizada en período otoño-invernal, según las técnicas habituales, fue transportada al Campo Experimental de la Facultad, donde al día siguiente se procedió a preparar las muestras y a la aplicación de los tratamientos (día 1). Las mediciones se realizaron día por medio, durante los días 3, 5, 7, 9 y 11 de almacenamiento.
Se evaluaron dos factores con dos niveles cada uno en un arreglo factorial 2x2 con mediciones repetidas en el tiempo, siendo los factores en estudio:
A) Ambientes de almacenaje:Cámara frigorífica (3,4 ºC; 97 % HR)Condiciones ambientales naturales (13 ºC; 81 % HR)
B) Revitalización, que consistió en sumergir las hortalizas en agua clorinada (50 ppm), durante 5 minutos, realizada cada día posteriormente a la medición:
Con revitalizaciónSin revitalización
La temperatura y humedad relativa fue monitoreada cada hora en cada ambiente con dispositivos electrónicos que registran mediciones ordenadas en el tiempo (Hobo ® U10 Temp/RH data loggers, Onset computer Corporation, Pocasset, MA USA).
Las especies evaluadas en hortalizas de hoja fueron: lechuga mantecosa, de hoja (crespa), arrepollada y apio; de fruto, zapallito y de raíz, zanahoria.
La mercadería evaluada respetó los parámetros de calidad según Trevor y Cantwell, (2011) En todos los casos las hortalizas estaban limpias, sin daños mecánicos, libres de plagas y enfermedades.
Lechuga: plantas de tamaño medio, hojas color verde brillantes, túrgidas y tiernas. Apio: pecíolos gruesos, compactos, poco curvados, verdes claro brillantes, túrgidos y tiernos. Sin pecíolo esponjoso, partido; tallos florales, y corazón negro.
Materiales y Métodos
-08-
Zapallito: frutos firmes al tacto; brillantes, uniformidad de color y forma, tamaño de 5 a 7 cm.Zanahoria: raíces firmes, rectas, cilíndricas y crujientes, color naranja brillante, sin raicillas laterales; ausencia de hombro y corazón verde.
Las variables medidas fueron: a) Pérdida de peso por descarte (en % respecto al peso del
día anterior). Para hortalizas de hoja, se eliminaron hojas rotas, amarillas, deshidratadas, enfermas, y base de tallo oxidado. Para zapallito se eliminaron frutos dañados, enfermos y sobremaduros. Para zanahoria, raíces dañadas, enfermas, rajadas, brotadas, con amarronamiento y se realizó despuntado en ambos extremos. Se aplicaron los mismos criterios de calidad mencionados anteriormente.
b) Pérdida o ganancia de peso por agua (en %, respecto al peso del día anterior). El resultado para todas las especies puede ser positivo o negativo, en función a la evaporación, transpiración y agua libre presente en la superficie de las hortalizas, debido al mojado luego de la cosecha o por la revitalización.
c) Pérdida de peso total (%). Suma de las dos anteriores, para todas las especies.
En todos los tratamientos se realizaron cuatro repeticiones para cada hortaliza, con muestras extraídas cada una de diferentes cajones o bolsas. En los tres tipos de lechuga, cada muestra tenía 5 plantas; en apio 1 planta; en zapallito 3 frutos y en zanahoria 3 raíces.
Para registrar la diferencia de peso de las variables se utilizó balanza digital modelo Systel Bumer (± 1g)
Para analizar este conjunto de datos se ajustó un modelo de medidas repetidas en el tiempo utilizando el procedimiento MIXED de SAS. Este tipo de análisis permite determinar si el efecto de los factores es similar a lo largo del tiempo o existe algún tipo de interacción con él, que no podría detectarse analizando por separado cada momento. Se determinó la estructura de covariancia correspondiente a cada una de las variables analizadas y se incluyeron en el modelo los efectos del tiempo, el ambiente de almacenaje, la revitalización y la interacción entre ellos. En los casos en que una interacción
Ciencias Agronómicas - Revista XX - Año 12 - 2012 / 007 - 013
Incidencia del ambiente y la revitalización en las pérdidas poscosecha de hortalizas de hoja, fruto y raíz

Resultados y Discusión
resultó significativa, se consideraron los factores intervinientes en la misma, y se compararon los niveles de uno de los factores en cada nivel del otro factor. El análisis estadístico se realizó en
función al tratamiento de menor duración poscosecha (día 3 al 9 inclusive, excepto en lechuga mantecosa y zapallito que fue hasta el día 7).
Pérdida por descarte (%): el ambiente con cámara frigorífica, mostró un descarte menor en todas las especies, excepto para zanahoria (Tabla 1). Las diferencias estadísticas (p < 0,05) en lechuga arrepollada fueron a partir del tercer día, en lechuga mantecosa y apio del quinto; desde el séptimo en lechuga de hoja y zapallito. La zanahoria, con menor tasa respiratoria que las otras especies, en condiciones invernales en la que se realizó el ensayo no mostró diferencia entre ambos ambientes.
La revitalización disminuyó significativamente las pérdidas por descarte en todas las especies, excepto para zanahoria y lechuga arrepollada. En esta última, dicha práctica favoreció la presencia de alteraciones fisiológicas como nervadura rosada y pudriciones acuosas, principalmente en el ambiente natural (Lipton, 1972; Namesny, 1993). Las pérdidas difirieron a partir del séptimo día en lechuga mantecosa, apio y zapallito, a partir del noveno en lechuga crespa o de hoja (p < 0,05). La técnica de revitalización, en general, tuvo un impacto menor que el
ambiente con cámara, manifestándose al final del almacenamiento.
En la mayoría de las especies el efecto de la interacción de los ambientes, revitalización y el tiempo fue significativo, es decir que el efecto del ambiente y la revitalización depende del tiempo de almacenaje de los productos evaluados. El uso de la cámara mostró un efecto más relevante.
En lechugas mantecosas, de hoja y arrepolladas, la aplicación de frío, produjo un descarte similar, en los tratamientos con y sin revitalización.
En zapallito, el descarte es similar con y sin revitalización en el ambiente natural y en cámara frigorífica, la revitalización disminuye el descarte. Esto podría explicarse porque el comportamiento de estas especies presenta una tasa respiratoria y una superficie expuesta menor. (Kader, 2007).
Tabla 1: Pérdidas por descarte (%) por día, aplicación de frío y revitalizado en todas las especies
T1: Cámara y Revitalización 6,6 6,0 12,5 18,7
T2: Cámara y sin Revitalización 11,4 9,3 14,0 32,7
T3: Cond ambi naturales y Revitalización 12,6 16,7 21,6
T4: Cond ambi naturales y sin Revitalización 13,8 22,2 95,2
T1: Cámara y Revitalización 7,1 8,3 12,0 28,0
T2: Cámara y sin Revitalización 4,0 4,7 7,0 21,8
T3: Cond ambi naturales y Revitalización 13,4 17,1 20,9 46,4
T4: Cond ambi naturales y sin Revitalización 15,3 15,4 23,0 88,8
T1: Cámara y Revitalización 8,1 6,5 22,5 11,1
T2: Cámara y sin Revitalización 7,7 6,3 8,9 8,6
T3: Cond ambi naturales y Revitalización 16,6 23,5 33,0 97,9
T4: Cond ambi naturales y sin Revitalización 16,2 11,1 13,5 16,0
T1: Cámara y Revitalización 0,0 0,2 0,0 1,5
T2: Cámara y sin Revitalización 0,0 0,1 0,0 11,3
T3: Cond ambi naturales y Revitalización 0,0 8,2 0,0 99,4
T4: Cond ambi naturales y sin Revitalización 6,9 4,6 15,1 98,6
T1: Cámara y Revitalización 0,0 0,0 0,0 5,1
T2: Cámara y sin Revitalización 0,0 0,0 23,2 73,8
T3: Cond ambi naturales y Revitalización 0,0 0,0 99,2
T4: Cond ambi naturales y sin Revitalización 0,0 0,0 98,4
T1: Cámara y Revitalización 0,0 0,0 0,0 0,0
T2: Cámara y sin Revitalización 0,0 0,0 0,0 0,0
T3: Cond ambi naturales y Revitalización 0,0 0,0 0,0 0,0
T4: Cond ambi naturales y sin Revitalización 0,0 0,0 0,0 0,0
**
** ns **
** **
** ** **
** ** ** **
R-D
ns ** **
p>F
A-D
** *
**
* **
ns
- - - - -
** ** * **
-
**
**
**
**
Día 3 Día 5p>F
R
p>F
A-RDía 7 Día 9
Zapallito
Zanahoria
p>F
A
p>F
D
Lechuga
Mantecosa
Lechuga deHoja
Lechuga
Arrepollada
Apio
Especie Tratamiento
**: Diferencias estadísticas altamente significativas (1%); *: Diferencias estadísticas significativas (5%); ns: sin diferencias estadísticasFactores: A (ambientes); D (dias); R (revitalización)Interacciones: A-R (ambiente-revitalización); A-D (ambiente-día); R-D (revitalización-día)
-09-Ciencias Agronómicas - Revista XX - Año 12 - 2012 / 007 - 013
Firpo, I.; Rotondo, R.; Ferratto, J.; Grasso, R.; Trevisan, A.; Rodriguez, M.
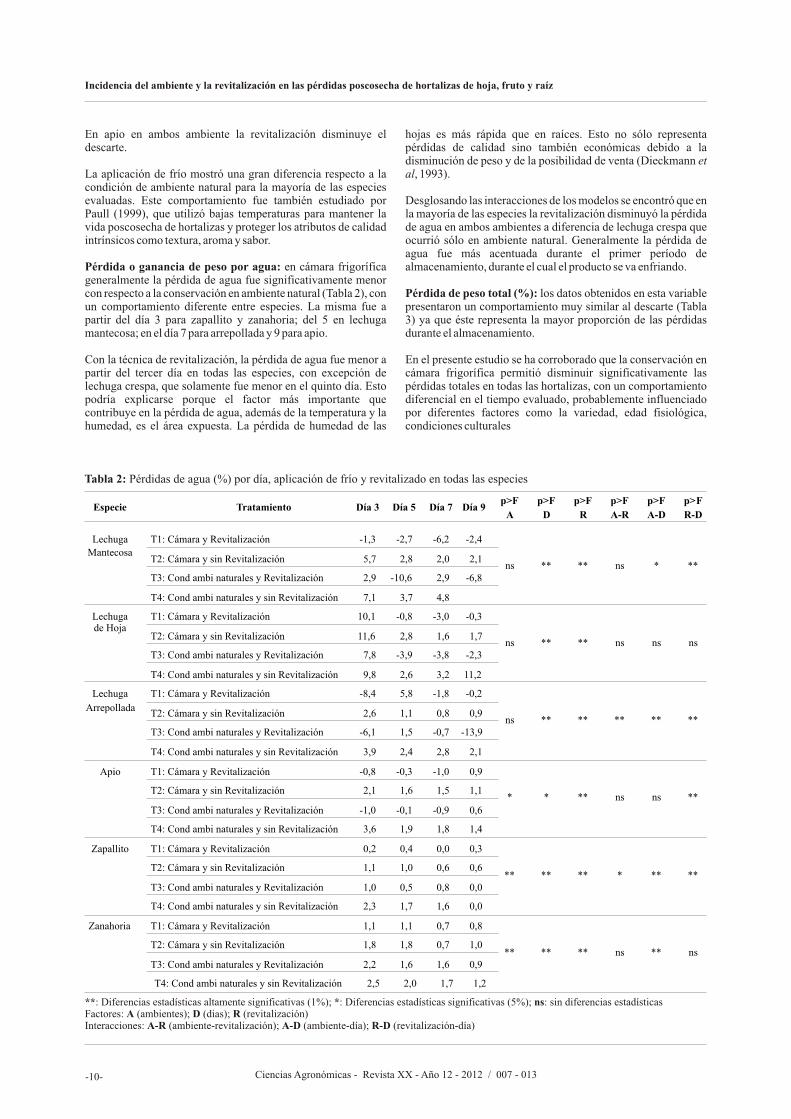
En apio en ambos ambiente la revitalización disminuye el descarte.
La aplicación de frío mostró una gran diferencia respecto a la condición de ambiente natural para la mayoría de las especies evaluadas. Este comportamiento fue también estudiado por Paull (1999), que utilizó bajas temperaturas para mantener la vida poscosecha de hortalizas y proteger los atributos de calidad intrínsicos como textura, aroma y sabor.
Pérdida o ganancia de peso por agua: en cámara frigorífica generalmente la pérdida de agua fue significativamente menor con respecto a la conservación en ambiente natural (Tabla 2), con un comportamiento diferente entre especies. La misma fue a partir del día 3 para zapallito y zanahoria; del 5 en lechuga mantecosa; en el día 7 para arrepollada y 9 para apio.
Con la técnica de revitalización, la pérdida de agua fue menor a partir del tercer día en todas las especies, con excepción de lechuga crespa, que solamente fue menor en el quinto día. Esto podría explicarse porque el factor más importante que contribuye en la pérdida de agua, además de la temperatura y la humedad, es el área expuesta. La pérdida de humedad de las
hojas es más rápida que en raíces. Esto no sólo representa pérdidas de calidad sino también económicas debido a la disminución de peso y de la posibilidad de venta (Dieckmann et al, 1993).
Desglosando las interacciones de los modelos se encontró que en la mayoría de las especies la revitalización disminuyó la pérdida de agua en ambos ambientes a diferencia de lechuga crespa que ocurrió sólo en ambiente natural. Generalmente la pérdida de agua fue más acentuada durante el primer período de almacenamiento, durante el cual el producto se va enfriando.
Pérdida de peso total (%): los datos obtenidos en esta variable presentaron un comportamiento muy similar al descarte (Tabla 3) ya que éste representa la mayor proporción de las pérdidas durante el almacenamiento.
En el presente estudio se ha corroborado que la conservación en cámara frigorífica permitió disminuir significativamente las pérdidas totales en todas las hortalizas, con un comportamiento diferencial en el tiempo evaluado, probablemente influenciado por diferentes factores como la variedad, edad fisiológica, condiciones culturales
Tabla 2: Pérdidas de agua (%) por día, aplicación de frío y revitalizado en todas las especies
T1: Cámara y Revitalización -1,3 -2,7 -6,2 -2,4
T2: Cámara y sin Revitalización 5,7 2,8 2,0 2,1
T3: Cond ambi naturales y Revitalización 2,9 -10,6 2,9 -6,8
T4: Cond ambi naturales y sin Revitalización 7,1 3,7 4,8
T1: Cámara y Revitalización 10,1 -0,8 -3,0 -0,3
T2: Cámara y sin Revitalización 11,6 2,8 1,6 1,7
T3: Cond ambi naturales y Revitalización 7,8 -3,9 -3,8 -2,3
T4: Cond ambi naturales y sin Revitalización 9,8 2,6 3,2 11,2
T1: Cámara y Revitalización -8,4 5,8 -1,8 -0,2
T2: Cámara y sin Revitalización 2,6 1,1 0,8 0,9
T3: Cond ambi naturales y Revitalización -6,1 1,5 -0,7 -13,9
T4: Cond ambi naturales y sin Revitalización 3,9 2,4 2,8 2,1
T1: Cámara y Revitalización -0,8 -0,3 -1,0 0,9
T2: Cámara y sin Revitalización 2,1 1,6 1,5 1,1
T3: Cond ambi naturales y Revitalización -1,0 -0,1 -0,9 0,6
T4: Cond ambi naturales y sin Revitalización 3,6 1,9 1,8 1,4
T1: Cámara y Revitalización 0,2 0,4 0,0 0,3
T2: Cámara y sin Revitalización 1,1 1,0 0,6 0,6
T3: Cond ambi naturales y Revitalización 1,0 0,5 0,8 0,0
T4: Cond ambi naturales y sin Revitalización 2,3 1,7 1,6 0,0
T1: Cámara y Revitalización 1,1 1,1 0,7 0,8
T2: Cámara y sin Revitalización 1,8 1,8 0,7 1,0
T3: Cond ambi naturales y Revitalización 2,2 1,6 1,6 0,9
T4: Cond ambi naturales y sin Revitalización 2,5 2,0 1,7 1,2
Día 3 Día 5p>F
R
p>F
A-R
p>F
A-D
Lechuga
Mantecosa
Día 7 Día 9p>F
A
p>F
DEspecie Tratamiento
R-D
ns
Zanahoria
*
ns **
Lechugade Hoja
Lechuga
Arrepollada
Apio
Zapallito
**
ns ** ** ns ns ns
** ** ns
** ** ** **
* * ** ns
** ns
** ** ** *
ns **
** **
** ** ** ns
**: Diferencias estadísticas altamente significativas (1%); *: Diferencias estadísticas significativas (5%); ns: sin diferencias estadísticasFactores: A (ambientes); D (dias); R (revitalización)Interacciones: A-R (ambiente-revitalización); A-D (ambiente-día); R-D (revitalización-día)
-10- Ciencias Agronómicas - Revista XX - Año 12 - 2012 / 007 - 013
Incidencia del ambiente y la revitalización en las pérdidas poscosecha de hortalizas de hoja, fruto y raíz
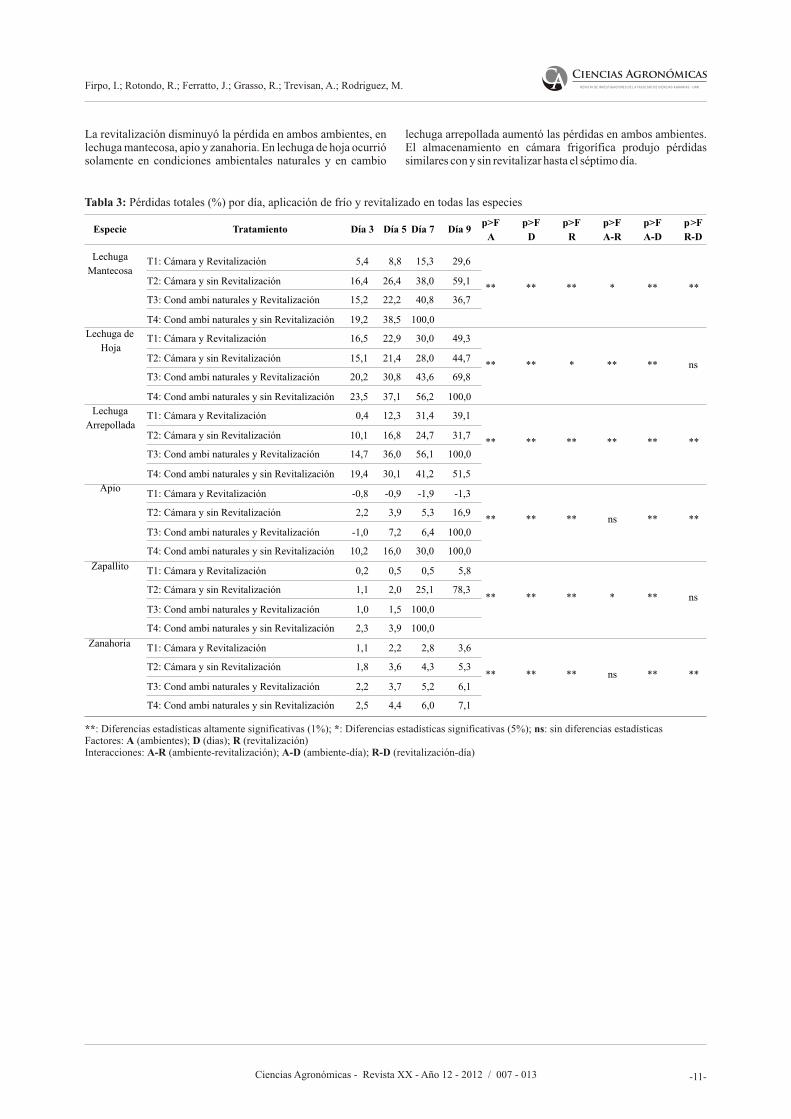
Tabla 3: Pérdidas totales (%) por día, aplicación de frío y revitalizado en todas las especies
T1: Cámara y Revitalización 5,4 8,8 15,3 29,6
T2: Cámara y sin Revitalización 16,4 26,4 38,0 59,1
T3: Cond ambi naturales y Revitalización 15,2 22,2 40,8 36,7
T4: Cond ambi naturales y sin Revitalización 19,2 38,5 100,0
T1: Cámara y Revitalización 16,5 22,9 30,0 49,3
T2: Cámara y sin Revitalización 15,1 21,4 28,0 44,7
T3: Cond ambi naturales y Revitalización 20,2 30,8 43,6 69,8
T4: Cond ambi naturales y sin Revitalización 23,5 37,1 56,2 100,0
T1: Cámara y Revitalización 0,4 12,3 31,4 39,1
T2: Cámara y sin Revitalización 10,1 16,8 24,7 31,7
T3: Cond ambi naturales y Revitalización 14,7 36,0 56,1 100,0
T4: Cond ambi naturales y sin Revitalización 19,4 30,1 41,2 51,5
T1: Cámara y Revitalización -0,8 -0,9 -1,9 -1,3
T2: Cámara y sin Revitalización 2,2 3,9 5,3 16,9
T3: Cond ambi naturales y Revitalización -1,0 7,2 6,4 100,0
T4: Cond ambi naturales y sin Revitalización 10,2 16,0 30,0 100,0
T1: Cámara y Revitalización 0,2 0,5 0,5 5,8
T2: Cámara y sin Revitalización 1,1 2,0 25,1 78,3
T3: Cond ambi naturales y Revitalización 1,0 1,5 100,0
T4: Cond ambi naturales y sin Revitalización 2,3 3,9 100,0
T1: Cámara y Revitalización 1,1 2,2 2,8 3,6
T2: Cámara y sin Revitalización 1,8 3,6 4,3 5,3
T3: Cond ambi naturales y Revitalización 2,2 3,7 5,2 6,1
T4: Cond ambi naturales y sin Revitalización 2,5 4,4 6,0 7,1
** **** ** ** ns
** ns
** **
** ** ** *
** ns
** ns
** **
** **
** ** ** **
** ** * **
Día 9p>F
A R-D
** **** ** ** *
Día 5 Día 7
Zapallito
Zanahoria
Lechuga de
Hoja
Lechuga
Arrepollada
Apio
p>F
D
p>F
R
p>F
A-R
p>F
A-DEspecie Tratamiento Día 3
Lechuga
Mantecosa
**: Diferencias estadísticas altamente significativas (1%); *: Diferencias estadísticas significativas (5%); ns: sin diferencias estadísticasFactores: A (ambientes); D (dias); R (revitalización)Interacciones: A-R (ambiente-revitalización); A-D (ambiente-día); R-D (revitalización-día)
La revitalización disminuyó la pérdida en ambos ambientes, en lechuga mantecosa, apio y zanahoria. En lechuga de hoja ocurrió solamente en condiciones ambientales naturales y en cambio
lechuga arrepollada aumentó las pérdidas en ambos ambientes. El almacenamiento en cámara frigorífica produjo pérdidas similares con y sin revitalizar hasta el séptimo día.
-11-Ciencias Agronómicas - Revista XX - Año 12 - 2012 / 007 - 013
Firpo, I.; Rotondo, R.; Ferratto, J.; Grasso, R.; Trevisan, A.; Rodriguez, M.
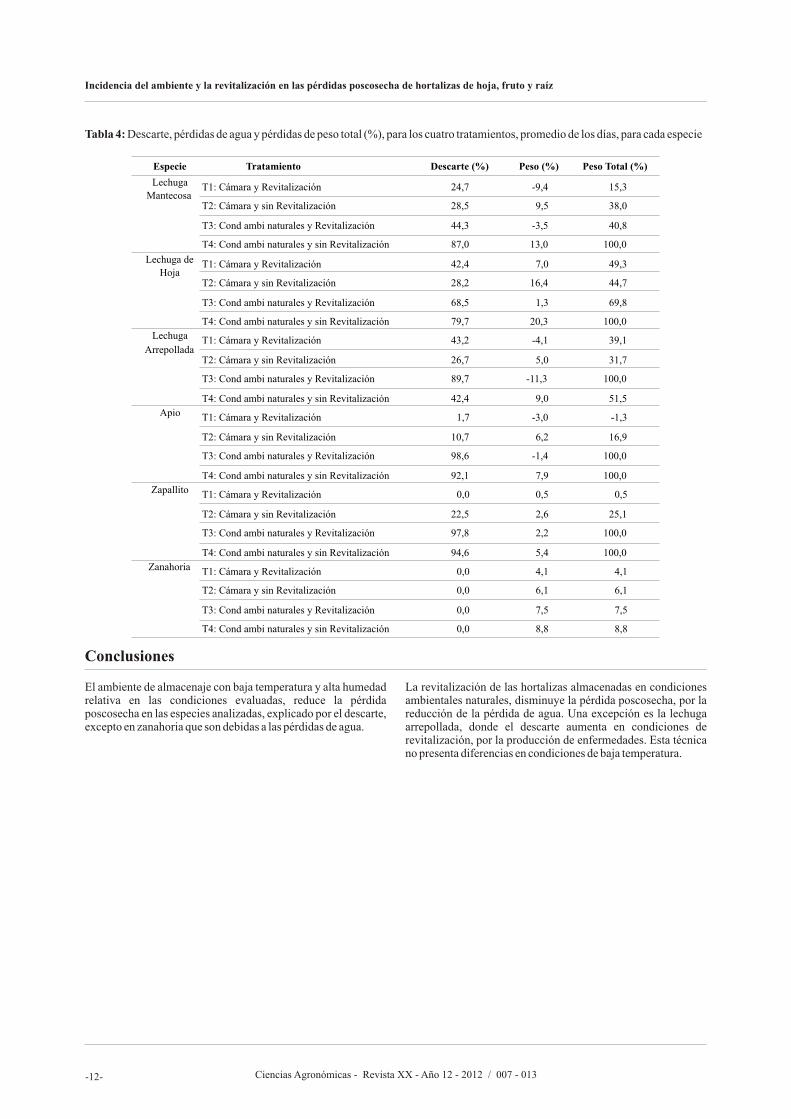
Tabla 4: Descarte, pérdidas de agua y pérdidas de peso total (%), para los cuatro tratamientos, promedio de los días, para cada especie
T1: Cámara y Revitalización 24,7 -9,4 15,3
T2: Cámara y sin Revitalización 28,5 9,5 38,0
T3: Cond ambi naturales y Revitalización 44,3 -3,5 40,8
T4: Cond ambi naturales y sin Revitalización 87,0 13,0 100,0
T1: Cámara y Revitalización 42,4 7,0 49,3
T2: Cámara y sin Revitalización 28,2 16,4 44,7
T3: Cond ambi naturales y Revitalización 68,5 1,3 69,8
T4: Cond ambi naturales y sin Revitalización 79,7 20,3 100,0
T1: Cámara y Revitalización 43,2 -4,1 39,1
T2: Cámara y sin Revitalización 26,7 5,0 31,7
T3: Cond ambi naturales y Revitalización 89,7 -11,3 100,0
T4: Cond ambi naturales y sin Revitalización 42,4 9,0 51,5
T1: Cámara y Revitalización 1,7 -3,0 -1,3
T2: Cámara y sin Revitalización 10,7 6,2 16,9
T3: Cond ambi naturales y Revitalización 98,6 -1,4 100,0
T4: Cond ambi naturales y sin Revitalización 92,1 7,9 100,0
T1: Cámara y Revitalización 0,0 0,5 0,5
T2: Cámara y sin Revitalización 22,5 2,6 25,1
T3: Cond ambi naturales y Revitalización 97,8 2,2 100,0
T4: Cond ambi naturales y sin Revitalización 94,6 5,4 100,0
T1: Cámara y Revitalización 0,0 4,1 4,1
T2: Cámara y sin Revitalización 0,0 6,1 6,1
T3: Cond ambi naturales y Revitalización 0,0 7,5 7,5
T4: Cond ambi naturales y sin Revitalización 0,0 8,8 8,8
Zanahoria
Lechuga
Mantecosa
Lechuga de
Hoja
Apio
Lechuga
Arrepollada
Zapallito
Especie Descarte (%) Peso (%) Peso Total (%)Tratamiento
Conclusiones
El ambiente de almacenaje con baja temperatura y alta humedad relativa en las condiciones evaluadas, reduce la pérdida poscosecha en las especies analizadas, explicado por el descarte, excepto en zanahoria que son debidas a las pérdidas de agua.
La revitalización de las hortalizas almacenadas en condiciones ambientales naturales, disminuye la pérdida poscosecha, por la reducción de la pérdida de agua. Una excepción es la lechuga arrepollada, donde el descarte aumenta en condiciones de revitalización, por la producción de enfermedades. Esta técnica no presenta diferencias en condiciones de baja temperatura.
-12- Ciencias Agronómicas - Revista XX - Año 12 - 2012 / 007 - 013
Incidencia del ambiente y la revitalización en las pérdidas poscosecha de hortalizas de hoja, fruto y raíz

Bibliografía
1. DIECKMANN, A.; LIST, D.; ZACHE, U. 1993. Cold water mist humedification to preserve the quality of fresh vegetables during retail sale. Lebensmittel-Wissenschaft und-Technologie.Volume 26, Issue 4, August, p. 340-346.
2. HANDENBURG, R.; WATADA, A. Y YI WANG, C. 1988. Almacenamiento comercial de frutas, legumbres y existencias de floristerías y viveros, p. 149.
3. KADER, A. 2002. Postharvest biology and technology: An overview. En: Postharvest Technolology of Horticultural Crops (3º edición). Kader, A. (ed). Publicación 3311. División of Agriculture and Natural Resources. Univ of California, EE.UU., p. 535.
4. LIPTON, W, STEWART, J. 1972. An ilustrated guide to the identification of some market disorders of head lettuce, Marketing Research Report Nº 950. Agricultural Research Service U.S Department of Agriculture, Beltsville, md., p. 27.
5. MONDINO, M.; FERRATTO, J.; FIRPO, I.; ROTONDO, R.; ORTIZ MACKINSON, M.; GRASSO, R.; CALANI, P.; LONGO, A. 2007. Pérdidas poscosecha de lechuga, en la región de Rosario, Argentina. Horticultura Argentina 26 (60) Ene.-Jun., p. 17-24.
6. NUNES, M; EDMOND, J; RAUTH, M; DEA, S. AND CHAU, K. 2009. Enviromental condictions encountred during typical consumer retail display afecct fruit andvegetable quality and waste. Postharvest Biol. Technol.51:232-241.
7. NAMESNY VALLESPIR, A. 1993. Post-recolección de hortalizas. Vol. I- Hortaliza de hoja, tallo y flor. Compendio de Horticultura 1. Ediciones de Horticultura S.L., p. 330.
8. PAULL, R. 1999. Effect of temperature and relative humidity on fresh commodity quolity. Department of Horticulture, College of Tropical Agriculture and Human Resources, University of Hawaii. Postharvest Biology and Technology Vol. 15 issue, March 1999, p. 263-277.
9. ROBINSON, J.E; BROWNE, K AND BURTON, W. 1975. Storage characteristics of some vegetables and soft fruits. Ann.appl. Biol. 81: 399-408.
10. T R E V O R , V. & C A N T W E L L , M . 2 0 1 1 . Recommendations for maintaining postharvest quality. A c c e s o e n e r o 2 0 1 1 . D i s p o n i b l e e n http://postharvesttechnology.ucdavis.edu/Produce/Produce Facts/Veg.
11. WILLS, R; MC GLASSON, B; GRAHAM, D.; JOYCE, D. 1999. Introducción a La fisiologia y manipulación poscosecha de frutas, hortalizas y plantas ornamentales. Segunda edición. Editorial Acribia S.A.. Zaragoza, España. ISBN: 84-200-0892-3. p. 240.
-13-Ciencias Agronómicas - Revista XX - Año 12 - 2012 / 007 - 013
Firpo, I.; Rotondo, R.; Ferratto, J.; Grasso, R.; Trevisan, A.; Rodriguez, M.


-15-
Artículo Original / Recibido 03/10/2012 - Aceptado 08/11/2012
Los cultivos de cobertura en la horticultura familiarurbana: una oportunidad para mejorar
la formación del estudiante de agronomíaCover crops in familiar urban horticulture:
an opportunity to improve the training of agricultural engineering students
1-2 2 2 3MILO VACCARO, M. ; MUÑOZ, G. ; TORRES, C. ; COURETOT, J.
1 INTA EEA Oliveros2 Taller de Integración I: La Investigación en las Ciencias Naturales y Sociales.Facultad de Ciencias Agrarias UNR3 Programa de Agricultura Urbana-Secretaría de Promoción Social-Municipalidad de Rosario. [email protected]
La investigación se realizó en el Taller de Integración I: La investigación en las Ciencias Naturales y Sociales, espacio curricular interdisciplinario ubicado en el segundo año de la carrera de Ingeniería Agronómica de la Facultad de Ciencias Agrarias-UNR. La experiencia se desarrolló en el Parque Huerta Molino Blanco durante el 2012, en el marco del Programa de Agricultura Urbana de la Municipalidad de Rosario, con la colaboración del Programa PROHUERTA de la Estación Experimental Agropecuaria del INTA Oliveros. Los objetivos fueron evaluar la eficiencia de los cultivos de cobertura para recuperar suelos urbanos, caracterizar la comunidad de huerteros que realiza las prácticas agroecológicas sobre esos suelos e identificar las competencias sociales y agronómicas desarrolladas por los estudiantes en ese contexto, entendido como un escenario de aprendizaje real y diverso. La complejidad de la problemática requirió un abordaje multidimensional. La dimensión agronómica implicó un diseño experimental de parcelas donde se midió biomasa y propiedades físico-químicas del suelo; la social comprendió el análisis interpretativo de la opinión de la comunidad huertera sobre sus prácticas y formas de vida. Por último, la dimensión educativa se focalizó en observar y registrar las competencias sociales desarrolladas por los estudiantes. Los resultados demuestran que los cultivos de cobertura son eficientes para recuperar suelos urbanos y la comunidad huertera posee conocimientos y experiencias que les posibilitan su incorporación como práctica agroecológica. Asimismo, los estudiantes mejoran su formación profesional al desarrollar competencias sociales y asumir un compromiso ético frente a situaciones de desigualdad social y baja calidad ambiental.
Palabras clave:competencias-agroecología-cultivos de cobertura
Resumen
The research was carried out in the Integration Workshop I: Research in the Natural and Social Sciences, which is an interdisciplinary learning program delivered in Year 2 of the Agricultural Engineering career at the College of Agricultural Sciences - UNR. The experience was developed in 2012 at the “Molino Blanco” Vegetable Garden Park, under the 'Urban Agriculture' programme of the Rosario Municipality, in collaboration with the “PROHUERTA” Programme of the INTA Oliveros Agricultural Experimental Station. The objectives were to evaluate the efficiency of cover crops to recover urban soils, to characterize the community of workers performing agro-ecological practices on these soils, and to identify the social and agronomic skills developed by students in this context, which makes a real and diverse learning scenario. The complexity of the problem required a multidimensional approach. The agronomic dimension required an experimental plot design that allowed us to measure biomass and physico-chemical properties of the soil; the social dimension covered the interpretive analysis of the agricultural workers' views on their practices and lifestyles; and the educational dimension focused on observing and recording the social skills developed by students. The results show that cover crops are efficient to recover urban lands and that vegetable producers have the knowledge and experience to incorporate cover crops as an agro-ecological practice. Also, this experience was shown to improve the professional training of students by developing their social skills and ethical commitment in situations of social inequalities and low environmental quality.
Key words:skills - agroecology - cover crops
Abstract
Introducción
Ciencias Agronómicas - Revista XX - Año 12 - 2012 / 015 - 021
El Parque Huerta Molino Blanco (PHMB) se ubica en el extremo sur de la ciudad de Rosario (Santa Fe) lindando con la Avenida Circunvalación y el Arroyo Saladillo, en un barrio carenciado formado por asentamientos precarios. Sobre un total de 4,5 hectáreas, alrededor de 25 familias afectan 4 hectáreas a la producción agroecológica en el marco del Programa de Agricultura Urbana (PAU) desarrollado como política pública por la Municipalidad de Rosario.
El PAU integra varones y mujeres en la generación de emprendimientos sociales de producción y elaboración de alimentos mediante técnicas ecológicas, destinados al consumo
familiar, comunitario y al mercado. Contribuye a la integración social, la superación de la pobreza, el mejoramiento del hábitat y el ambiente urbano. Posibilita, además, redes solidarias de
1producción, circulación y consumo de alimentos sanos .
Si bien la agricultura urbana basada en la agroecología se presenta como una alternativa en todo el país, en el caso del PHMB adquiere características particulares vinculadas a la migración de un campesinado que busca oportunidades
1Para más información sobre fundamento y funciones del PAU se puede consultar el sitio http://rosario.gov.ar

-16-
laborales ligadas a la propia experiencia y al acervo cultural. En este sentido, Lattuca (2012) señala que el peso relativo de los pobladores, con experiencia en horticultura o en otras formas de agricultura intensiva, es una de las diferencias sustantivas del Gran Rosario, en comparación con otras grandes ciudades del país. Es un factor que contribuyó positivamente al Programa, que se reflejó en la multiplicación de huertas familiares y grupales, y en los resultados obtenidos en términos de cantidad y calidad de la producción. Esta situación ha posibilitado el despliegue de las capacidades de los pobladores, así como la apropiación y adaptación de un enfoque técnico novedoso, como es el agroecológico.
Prácticas y técnicas campesinas tienden a ser intensivas en conocimiento y no intensivas en insumos, pero es evidente que no todas son aplicables, por lo tanto, pueden ser necesarias algunas modificaciones y adaptaciones. Es aquí donde la agroecología ha jugado un papel clave en la revitalización de la productividad de los pequeños sistemas agrícolas (Altieri y Nicholls, 2012). Para el caso de la recuperación de los suelos, se ha demostrado que el uso de cultivos de cobertura es eficiente al mejorar la macroporosidad y estabilidad estructural, fundamentalmente, mediante la acción de las raíces y la incorporación de materia orgánica (Varela, et.al., 2011). Un cultivo de cobertura es una especie que no es cultivo principal y que se siembra en el campo o parcela para proveer cobertura al suelo. Los cultivos de cobertura aumentan la cantidad de materia orgánica, estimulan la actividad biológica del suelo, reducen la erosión del suelo, pueden contribuir a la fijación biológica del nitrógeno y ser hospedantes alternos para enemigos benéficos de los insectos plaga de los cultivos. Cuando las leguminosas son usadas como coberturas, tanto solas como en combinación con especies no leguminosas, la calidad de la biomasa mejora. La biomasa resultante puede ser incorporada al suelo o dejada sobre la superficie como un acolchado o cobertura protectora hasta su posterior descomposición
Frente a esta problemática se planteó como posibilidad que la comunidad huertera incorpore a sus prácticas los cultivos de cobertura como herramienta para mejorar el suelo y transformar la producción en sustentable, a partir de un proceso dialógico-colaborativo entre huerteros, estudiantes y docentes. Proceso que debía sustentarse en la investigación y la transferencia de los resultados desde la comunidad científica hacia la comunidad huertera y viceversa.
En este sentido, Sarandón (1995) reclama que las instituciones de educación superior deben asumir la responsabilidad de formar profesionales capaces de transformar la agricultura y la implementación de los contenidos de agricultura sustentable o agroecológica en las Facultades de Ciencias Agrarias, que deben tener en cuenta las distintas situaciones ecológicas, sociales y económicas de las distintas regiones de nuestro país y adaptarse a las mismas. Para el caso de Zavalla se advierte que los Parques Huertas ofrecen un contexto social y cultural que los posiciona como potenciales escenarios educativos susceptibles de ser incorporados a las currículas como espacios de integración de enfoques agroecológicos y propuestas de enseñanza basadas en el desarrollo de competencias.
La articulación entre el conocimiento científico y el saber popular constituye un desafío para el docente en su rol de formador y las comunidades huerteras, como contextos de
aprendizaje, generan oportunidades inagotables para que los estudiantes se involucren y comprometan con la problemática ambiental. Al respecto, se ha demostrado que las prácticas de agricultura urbana constituyen mecanismos colectivos a escala familiar y comunitaria, adquiriendo una dimensión de educación popular medioambiental (Ottmann, et.al., 2009).
Es así que, desde la articulación entre un ensayo sobre el uso de cultivos de cobertura y la caracterización de la comunidad huertera como fenómeno social complejo e idiosincrásico, surge la posibilidad de enriquecer la formación del estudiante de agronomía desarrollando estrategias educativas basadas en la adquisición de competencias.
La competencia no es una disposición previa a la acción, sino que se adquiere, se desarrolla y consolida en ella. Ya generada, se constituye en un recurso para futuras acciones y se suma al capital profesional y laboral del sujeto, de acuerdo a ello, los docentes universitarios deben generar instancias de formación que coloquen al estudiante en un papel protagónico y dinámico (Coronado, 2009).
Para transformar la educación y posibilitar su enseñanza es necesario partir del análisis de las situaciones y de la acción y de ahí derivar los conocimientos. Para desarrollarlas el profesor debe trabajarlas particularmente por problemas y proyectos, es decir, proponer tareas complejas, retos, que inciten a los alumnos a movilizar sus conocimientos y habilidades con una pedagogía activa, cooperativa, abierta (Tenutto, et.al., 2010).
Se trata, entonces, de ser creativos y privilegiar la adquisición de competencias por sobre el aprendizaje de conocimientos desvinculado de la realidad, aun cuando la propuesta de enseñanza implique asumir riesgos. En una línea de coherencia, se asume la necesidad de implementar una evaluación continua seleccionando un instrumento que recoja a lo largo del tiempo evidencias de comprensión y aplicación de los conocimientos. Así, la evaluación se transforma en una instancia más de aprendizaje, en la cual estrategias como: resolución de problemas, estudio de casos, presentación de informes, exposiciones orales, elaboración de un portafolio, entre otras, generan un proceso de retroalimentación para el estudiante y para el docente que en función de los resultados podría reorientar la experiencia.
Es importante comprender que incidir en el sistema de conocimiento del alumno no depende sólo de la actividad de enseñar en sí misma, sino de la actividad de diseñar y emprender propuestas. Por ello, es fundamental utilizar estrategias de aprendizaje que le permitan transitar por el saber cómo, saber qué y saber para qué (Rué, 2009).
Tomando como punto de partida el marco referencial expuesto, donde se conjugan la dimensión agronómica, social y educativa que atraviesan la problemática abordada, surgen algunos interrogantes: ¿Es posible mejorar la formación del estudiante de agronomía desarrollando una propuesta educativa en una huerta comunitaria de producción orgánica? ¿Qué competencias desarrollan los estudiantes en este contexto tan particular? ¿Son eficientes los cultivos de cobertura para recuperar suelos urbanos? ¿Qué características tiene la comunidad huertera del PHMB? ¿En qué medida se transfieren los conocimientos científicos? ¿Cómo se articulan con los saberes populares?
Los cultivos de cobertura en la horticultura familiar urbana: una oportunidad para mejorar la formación del estudiante de agronomía
Ciencias Agronómicas - Revista XX - Año 12 - 2012 / 015 - 021

-17-
Analizar el PHMB como potencial escenario para mejorar la formación del estudiante de agronomía a través del desarrollo de competencias sociales y vinculadas a la profesión.
Objetivos
Evaluar la capacidad de los cultivos de cobertura para recuperar suelos urbanos en una huerta comunitaria.
Caracterizar la comunidad huertera y la aplicación de prácticas agroecológicas.
MILO VACCARO, M.; MUÑOZ, G.; TORRES, C.; COURETOT, J.
La experiencia se desarrolló en el PHMB localizado en la intersección de la calle Ayacucho y el Arroyo Saladillo, en el extremo sur de Rosario (Santa Fe) en el límite con la localidad de Villa Gobernador Gálvez. Gran parte del predio está rodeado por un barrio humilde, bastante urbanizado, en el que predominan los asentamientos precarios o “villas”.
Históricamente, el PHMB tuvo diversos usos y ocupantes. Hace aproximadamente 30 años, funcionó un tambo, luego, por un período de 10 años hubo una chatarrería. Más tarde fue un espacio de asentamientos irregulares, pero luego sus habitantes fueron re-localizados por la Municipalidad en otra zona de la ciudad. Por último, se utilizó como depósito de basura doméstica hasta que en el año 2002 comenzó a utilizarse como huerta.
Para el caso del PHMB, es necesario desarrollar trabajos científicos que evalúen las prácticas agroecológicas implementadas por la comunidad huertera y que caractericen a esta última como fenómeno social surgido de cara a la crisis económica del 2001.
Diseño de la experienciaLa experiencia fue interdisciplinaria e interinstitucional y se extendió de marzo del 2011 a junio del 2012. El equipo de investigación estuvo constituido por 3 docentes del Taller de Integración I, 1 profesional del PAU, 1 profesional del INTA Oliveros; y participaron 6 Ayudantes Alumnos y 10 estudiantes que desarrollaron Prácticas Pre-Profesionales. Las actividades comenzaron en el marco de un Proyecto de Extensión anual acreditado por la Universidad Nacional de Rosario, pero al tiempo, el involucramiento del equipo de trabajo con las necesidades del PHMB, condujo al desarrollo de la presente investigación.
Con el propósito de lograr una aproximación lo más comprensiva posible de la problemática abordada, se diseñó una estrategia metodológica que articulara tres dimensiones: una agronómica enfocada en los cultivos de cobertura, una social orientada a la comunidad huertera, y la educativa centrada en la formación del estudiante de agronomía. A continuación, para cada dimensión, se presentan las variables o categorías seleccionadas, los instrumentos y el tratamiento de los datos.
Dimensión agronómicaAplicando un diseño experimental ajustado a la situación de manejo
Metodología
y condición socioeconómica-cultural de los huerteros, se evaluaron las siguientes especies para cultivos de cobertura: avena, trigo, cebada, centeno y triticale (gramíneas) combinados con vicia (leguminosa) en parcelas de 3m de largo por 1,5m de ancho ubicadas en un sector con suelo degradado respecto de su estructura, textura y contenido de nutrientes. Se hicieron 2 repeticiones de cada parcela y se midió:
- Cultivos: stand de plantas, cantidad de macollos, materia 2verde (g/m ) y materia seca (g/m2) medidas en el estadio de hoja
embuchada o comienzo de espigazón, mediante la obtención de muestras con cuadros de alambre de 0,25cm por 0,25cm.
- Propiedades químicas del suelo: mediante el análisis químico de muestras de suelo, determinando el porcentaje de materia orgánica y cantidad de macronutrientes (N, P y K), nitratos totales por el método Harper, fósforo por el método de Bray 1, sulfatos con fosfato de Ca, PH y conductividad eléctrica. Determinación de cationes en solubilidad y retención: sodio (Na); calcio (Ca); magnesio (Mn).
- Propiedades físicas del suelo: observación en calicatas para caracterizar el perfil y el desarrollo/distribución radicular
Ay
ac
uch
o
Arroyo LudueñaAv. 25 de Mayo (Av. Circunvalación)
O
N
S
Av 1 Cb 2 Ce 3
Tr 4 Tt 5 Av 6
ECe 8 Tr 9
Test 10 Test 11 Tt 12
Cb 7
Figura 1: Distribución de las parcelas en el PHMB.
Dimensión social El enfoque cualitativo aplicado en esta dimensión articuló la realización de entrevistas semiestructuradas a los integrantes de la comunidad huertera con la observación de sus prácticas agronómicas. A partir de la información obtenida se construyeron las siguientes categorías de análisis:
- Contexto socioeconómico: influencia de la situación socioeconómica imperante en el inicio del emprendimiento productivo.
- Organización familiar del trabajo: cómo se distribuyen las actividades de la huerta entre los integrantes de la familia de los huerteros y quién o quiénes asumen el rol de dirigir para la realización de las tareas.
- Sistema de conocimientos prácticos/aprendizaje: circulación e internalización del conocimiento agronómico entre los huerteros y demás actores.
Ciencias Agronómicas - Revista XX - Año 12 - 2012 / 015 - 021

-18-
- Tenencia de la tierra: refiere a la forma de ocupación del predio por parte de los huerteros
- Sentimientos por la actividad y la tierra: expresiones manifiestas (libertad, felicidad y gusto) de los huerteros/as con respecto a su tarea.
- Expectativas sobre los hijos: deseos por el futuro de sus hijos.- Manejo agroecológico/prácticas: conocimientos y habilidades para la realización de las prácticas de manejo agroecológicas.
- Actitud de liderazgo: relación de poder entre los huerteros que influye en la comunicación.
- Autodeterminación: nivel de autogestión de las propuestas y toma de decisiones en distintos ámbitos que dan continuidad al emprendimiento parque huerta.
- Institucionalidad: nivel de organización local/regional de los huerteros.
Dimensión educativaSe aplicó una encuesta de opinión anónima y voluntaria a los estudiantes que participaron en la experiencia. Con la intención de no orientar las respuestas y menos aún limitar las opiniones, se eligió un diseño abierto con una única pregunta sobre lo que habían experimentado en relación al proyecto del PHMB. La información obtenida fue procesada y organizada en categorías conceptuales que posibilitaban contextualizar las expresiones y otorgarles un significado:
- Organización y planificación de la experiencia: involucra la formación del equipo de trabajo y la dinámica de trabajo grupal. También incluye la elaboración del cronograma de actividades y su
cumplimiento; formas de comunicación; división de las tareas; organización de traslados y tratamiento de situaciones imprevistas.
- Estilos de enseñanza: cómo enseña el docente, cómo utiliza los recursos, qué actividades propone y de qué manera orienta y acompaña al estudiante en su proceso de aprendizaje. Involucró el desocultamiento de las teorías y creencias que subyacen a la práctica docente. Se enfatizó la búsqueda de una enseñanza orientada a la adquisición de competencias a partir de estrategias que motivan y desafían al estudiante a involucrarse con un problema real que requiere compromiso, actitudes y valores, además de conocimientos y habilidades técnicas.
- Modos de aprendizaje: cómo los estudiantes aprenden, qué estrategias desarrollan y de qué recursos se valen para aprender. Si bien se reconocen diferentes modos de aprender, este tipo de propuesta favorece el protagonismo, la autonomía y la metacognición. Se constituyen en escenarios que generan un clima de aprendizaje donde los estudiantes no solo construyen los saberes sino que además reflexionan sobre cómo aprendieron.
- Metodología de evaluación: refiere a las estrategias e instrumentos que el docente utiliza para evaluar la adquisición de competencias durante el proceso y al finalizar la propuesta. La evaluación, entendida como una oportunidad de revisar qué se aprende, cómo se aprende y para qué se aprende, incluye instancias de evaluación no formales que retroalimentan la propuesta de enseñanza sin afectar las emociones y la autoestima de los estudiantes.
El trabajo analítico-interpretativo estuvo orientado a la caracterización de los procesos de enseñanza, aprendizaje y evaluación para reflexionar sobre las potencialidades educativas de la propuesta para lograr la adquisición de competencias..
Resultados y Discusión
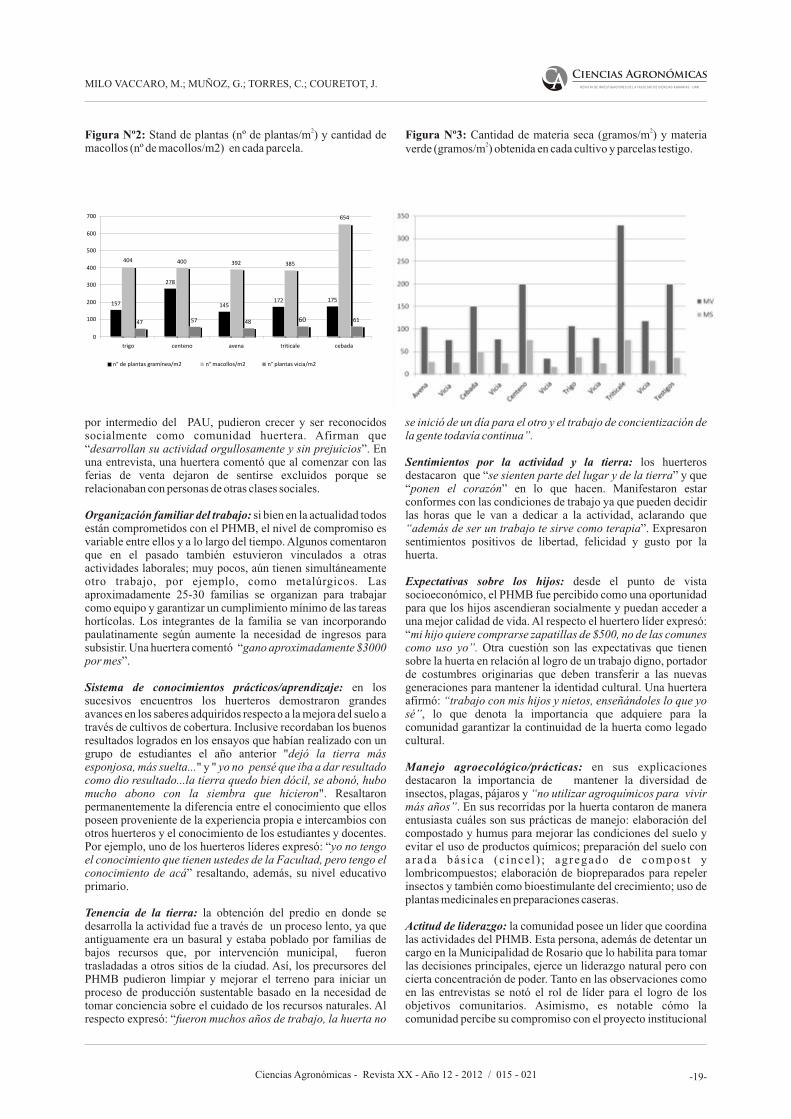
Dimensión agronómicaEn la Figura Nº2 se muestra una mayor formación de macollos en la cebada seguida por el trigo, centeno, avena y triticale, los que presentaron casi la misma cantidad de macollos por metro cuadrado. En relación al stand de plantas se observa que la combinación centeno-vicia presenta ventajas comparativas.
Como se puede observar en la Figura Nº3, comparativamente hubo una importante producción de materia verde de triticale en primer lugar y de centeno en segundo orden, respecto de las restantes gramíneas, mientras que ambas especies -si bien no mostraron diferencias notables entre ellas- superaron al resto en cuanto a la producción de materia seca.
En ambos cuadros se observa que la cebada si bien presentó el mayor número de macollos por metro cuadrado y parecía una especie promisoria para cultivo de cobertura (de hecho la utilizan ampliamente los productores para tal fin), no resultó así en cuanto a la producción de materia verde y materia seca, parámetros relevantes para la selección de una gramínea como cultivo de cobertura. Resultaron ventajosos el centeno y el triticale.
Los resultados de los análisis químicos del suelo mostraron un pH alto, cercano al valor 8, lo cual indica alcalinidad, esto se debe, probablemente, a la presencia de calcáreos aportados por los escombros y de carbonatos debido a la proximidad del arroyo Saladillo. Los niveles de nutrientes resultaron óptimos para Nitratos, Fosfatos y Potasio y deficientes en Azufre. También, la proporción de materia orgánica es adecuada, aunque es
importante señalar que se debe al aporte continuo de compost que realizan los huerteros.
Los análisis de agua de riego develaron que es alcalina o salina, lo cual sumado al pH elevado implica riesgo de alcalinidad del suelo, lo cual es perjudicial para el desarrollo de las especies hortícolas. Esto se podría remediar con el uso de agua de lluvia y con elevados aportes de materia orgánica.
Las evaluaciones de la estructura del suelo demostraron que los cultivos de cobertura la mejoraron por la mayor producción de biomasa radicular sobre la base de materia seca. Con respecto al desarrollo y distribución radicular, se observó una mejora estructural debido a la presencia de un sistema radicular denso que incrementó la materia orgánica y mejoró los agregados de suelo.
Dimensión socialLa información recabada a partir de las entrevistas y de las observaciones, nos permitió caracterizar la comunidad huertera en relación al contexto histórico de surgimiento y conocer las creencias y supuestos que subyacen a sus prácticas agronómicas.
Contexto socioeconómico: la huerta surgió como consecuencia de la crisis económica, social y política ocurrida en el 2001. Las actividades que se desarrollaban en ésta constituían el único ingreso económico para numerosas familias, aun cuando su trabajo era desvalorizado socialmente. A través del paso del tiempo y gracias al apoyo de instituciones que brindaron capacitación e insumos, como ser la Municipalidad de Rosario
Ciencias Agronómicas - Revista XX - Año 12 - 2012 / 015 - 021
Los cultivos de cobertura en la horticultura familiar urbana: una oportunidad para mejorar la formación del estudiante de agronomía
-19-
por intermedio del PAU, pudieron crecer y ser reconocidos socialmente como comunidad huertera. Afirman que “desarrollan su actividad orgullosamente y sin prejuicios”. En una entrevista, una huertera comentó que al comenzar con las ferias de venta dejaron de sentirse excluidos porque se relacionaban con personas de otras clases sociales.
Organización familiar del trabajo: si bien en la actualidad todos están comprometidos con el PHMB, el nivel de compromiso es variable entre ellos y a lo largo del tiempo. Algunos comentaron que en el pasado también estuvieron vinculados a otras actividades laborales; muy pocos, aún tienen simultáneamente otro trabajo, por ejemplo, como metalúrgicos. Las aproximadamente 25-30 familias se organizan para trabajar como equipo y garantizar un cumplimiento mínimo de las tareas hortícolas. Los integrantes de la familia se van incorporando paulatinamente según aumente la necesidad de ingresos para subsistir. Una huertera comentó “gano aproximadamente $3000 por mes”.
Sistema de conocimientos prácticos/aprendizaje: en los sucesivos encuentros los huerteros demostraron grandes avances en los saberes adquiridos respecto a la mejora del suelo a través de cultivos de cobertura. Inclusive recordaban los buenos resultados logrados en los ensayos que habían realizado con un grupo de estudiantes el año anterior "dejó la tierra más esponjosa, más suelta..." y " yo no pensé que iba a dar resultado como dio resultado...la tierra quedo bien dócil, se abonó, hubo mucho abono con la siembra que hicieron". Resaltaron permanentemente la diferencia entre el conocimiento que ellos poseen proveniente de la experiencia propia e intercambios con otros huerteros y el conocimiento de los estudiantes y docentes. Por ejemplo, uno de los huerteros líderes expresó: “yo no tengo el conocimiento que tienen ustedes de la Facultad, pero tengo el conocimiento de acá” resaltando, además, su nivel educativo primario.
Tenencia de la tierra: la obtención del predio en donde se desarrolla la actividad fue a través de un proceso lento, ya que antiguamente era un basural y estaba poblado por familias de bajos recursos que, por intervención municipal, fueron trasladadas a otros sitios de la ciudad. Así, los precursores del PHMB pudieron limpiar y mejorar el terreno para iniciar un proceso de producción sustentable basado en la necesidad de tomar conciencia sobre el cuidado de los recursos naturales. Al respecto expresó: “fueron muchos años de trabajo, la huerta no
se inició de un día para el otro y el trabajo de concientización de la gente todavía continua”.
Sentimientos por la actividad y la tierra: los huerteros destacaron que “se sienten parte del lugar y de la tierra” y que “ponen el corazón” en lo que hacen. Manifestaron estar conformes con las condiciones de trabajo ya que pueden decidir las horas que le van a dedicar a la actividad, aclarando que “además de ser un trabajo te sirve como terapia”. Expresaron sentimientos positivos de libertad, felicidad y gusto por la huerta.
Expectativas sobre los hijos: desde el punto de vista socioeconómico, el PHMB fue percibido como una oportunidad para que los hijos ascendieran socialmente y puedan acceder a una mejor calidad de vida. Al respecto el huertero líder expresó: “mi hijo quiere comprarse zapatillas de $500, no de las comunes como uso yo”. Otra cuestión son las expectativas que tienen sobre la huerta en relación al logro de un trabajo digno, portador de costumbres originarias que deben transferir a las nuevas generaciones para mantener la identidad cultural. Una huertera afirmó: “trabajo con mis hijos y nietos, enseñándoles lo que yo sé”, lo que denota la importancia que adquiere para la comunidad garantizar la continuidad de la huerta como legado cultural.
Manejo agroecológico/prácticas: en sus explicaciones destacaron la importancia de mantener la diversidad de insectos, plagas, pájaros y “no utilizar agroquímicos para vivir más años”. En sus recorridas por la huerta contaron de manera entusiasta cuáles son sus prácticas de manejo: elaboración del compostado y humus para mejorar las condiciones del suelo y evitar el uso de productos químicos; preparación del suelo con a rada bás ica (c ince l ) ; agregado de compos t y lombricompuestos; elaboración de biopreparados para repeler insectos y también como bioestimulante del crecimiento; uso de plantas medicinales en preparaciones caseras.
Actitud de liderazgo: la comunidad posee un líder que coordina las actividades del PHMB. Esta persona, además de detentar un cargo en la Municipalidad de Rosario que lo habilita para tomar las decisiones principales, ejerce un liderazgo natural pero con cierta concentración de poder. Tanto en las observaciones como en las entrevistas se notó el rol de líder para el logro de los objetivos comunitarios. Asimismo, es notable cómo la comunidad percibe su compromiso con el proyecto institucional
157
278
145172 175
404 400 392 385
654
47 57 48 6160
0
100
200
300
400
500
600
700
trigo centeno avena triticale cebada
n° de plantas gramínea/m2 n° macollos/m2 n° plantas vicia/m2
2Figura Nº2: Stand de plantas (nº de plantas/m ) y cantidad de macollos (nº de macollos/m2) en cada parcela.
2Figura Nº3: Cantidad de materia seca (gramos/m ) y materia 2verde (gramos/m ) obtenida en cada cultivo y parcelas testigo.
Ciencias Agronómicas - Revista XX - Año 12 - 2012 / 015 - 021
MILO VACCARO, M.; MUÑOZ, G.; TORRES, C.; COURETOT, J.

-20-
y su habilidad para llevar adelante las gestiones de manera autónoma.
Autodeterminación: los resultados beneficiosos que logran cotidianamente se deben en gran parte a la autodeterminación que los caracteriza como comunidad autogestionada capaz de elegir libremente cómo asociarse y comercializar sus productos. La comunidad presenta un alto nivel de autonomía e iniciativa propia y esto le posibilita integrarse con otros actores sin perder su identidad social y cultural. Por ejemplo, cuando tuvieron que afrontar algunas circunstancias adversas, tomaron la decisión de crear una Red de huerteros con personería jurídica. Así, lograron el reconocimiento de sus derechos sociales y políticos, y un mecanismo para proteger la producción y la comercialización de sus productos.
Institucionalidad: la Municipalidad de Rosario, a través del PAU, fomentó la agricultura familiar urbana y les brindó capacitación e insumos. Para la comunidad, esta política pública fue un gran avance ya que les permitió obtener un ingreso capaz de solventar el estudio de sus hijos y otros gastos. Perciben el PHMB como un proyecto de la municipalidad que “sirve como ejemplo para cambiar la sociedad” y asumen la tarea de recibir y guiar a los visitantes, proporcionándoles información. Generalmente, el líder de la huerta brinda su discurso agroecológico, basado en el protocolo de trabajo comunitario. El PAU les facilitó la comercialización a través de las ferias y por un mecanismo centralizado de pedido domiciliario de un bolsón de productos orgánicos. Asimismo, la organización en Red les posibilitó realizar viajes de intercambio con otras regiones, autofinanciados a través de almuerzos solidarios, y superar adversidades climáticas, como por ejemplo, cuando en el 2006 perdieron todos los cultivos por la granizada. En relación a este fenómeno de gran impacto para la producción hortícola, un huertero recordaba: "se vendió un bono de $25 en aquel entonces y se lo devolvíamos a cuatro o cinco meses y nos comprometimos siempre a devolverlo en bolsón de mercadería".
Dimensión educativaEl análisis de la información recabada en las encuestas tomó como pivote las categorías conceptuales para, en relación a las mismas, poner en discusión la percepción del estudiante y otorgarle un significado contextual.
Organización de la experiencia: los estudiantes reconocieron las dificultades que surgen cuando se trabaja con una dinámica grupal en una situación de aprendizaje compleja y dinámica. Esto queda claramente evidenciado cuando dicen: “trabajar en grupo representa todo un reto” y “nuestra forma de aprender fue buena pero comenzó luego de haber superado nuestras propias dificultades como grupo”. Algunos comentarios sugirieron la percepción de inconvenientes en la organización: “los procedimientos fueron un poco arrebatados quizás por falta de tiempo, falta de organización de parte nuestra o desacuerdos con los docentes”, sin embargo, hubo opiniones que mostraban claramente que los estudiantes visualizaron las ventajas que ofrece un proceso de aprendizaje grupal, que propicia una mirada holística e integradora. Desde el punto de vista operativo, la heterogeneidad del grupo de trabajo complejizó la concreción de algunas tareas según lo planificado.
Estilos de enseñanza: la mayoría mencionó como aspecto positivo el posicionamiento del docente en la manera de enseñar, que asumió un rol de mediador y orientador muy diferente al rol tradicional: “mucho que enseñar no había ya que era un proyecto en el que nosotros teníamos que aprender por nuestra cuenta, lo cual fue muy bueno ya que nos enfrentamos a problemáticas reales y eso, a mi criterio, te prepara mucho para
lo que va a venir” y “aprendí el trato con los productores, como hacer una entrevista, como preparar un trabajo para una exposición, la manera de exponer en sí”. En este sentido, la propuesta se centró en promover la autogestión de los grupos y la autonomía en el aprendizaje. Los docentes desarrollaron estrategias de comunicación fluidas que favorecieron la construcción conjunta del conocimiento, a través de la contextualización y confrontación con la realidad. Esto se evidencia cuando dicen: “con respecto a los docentes fue bueno haber establecido un contacto más cercano […] un ida y vuelta más dinámico”. El PHMB y la comunidad de huerteros constituyeron un escenario de aprendizaje fértil, colmado de diversos recursos que los docentes aprovecharon para generar actividades integradoras que fueron valoradas por los estudiantes. Al respecto, opinaron: “se tocan varios temas de la agronomía […] vimos acerca de gramíneas y leguminosas, tipos de raíces, fijación biológica del nitrógeno, materia seca y su medición, reconocimiento de malezas, insectos, preparación de un experimento, análisis estadístico, forma de trabajo agroecológico, producción orgánica, clima, análisis de suelo, comercialización de la producción, trato con los productores, también aprendimos a trabajar en equipo […] cómo hacer una entrevista, cómo preparar un trabajo para una exposición ... por nombrar algunas.”
Modos de aprendizaje: todos los estudiantes destacaron las oportunidades que ofrece la experiencia de construir los saberes a partir de la integración de los conocimientos disciplinares con la experiencia de los huerteros: “se dio otra forma de aprender fuera de la facultad relacionándonos con gente diferente […] de la gente del parque también se aprende” y “fue una forma de aplicar algunos de los conocimientos aprendidos en las diferentes materias durante estos años y poder ponerlos en práctica”. Se valoró el aprendizaje colaborativo y la adquisición de competencias en una situación de aprendizaje real: “aprendimos a tratar de cerca con productores, tutores y profesores, tanto para compartir experiencias como también para tratar las problemáticas que de a poco fueron surgiendo.” Señalaron la necesidad de trascender las paredes de la institución para articular la teoría con la práctica en un contexto cierto: “tuvimos un trato con la gente -que generalmente se da cuando uno se recibe- que no solo nos sirvió para enseñarles, sino también aprender mucho de ellos [...] creo que se aprende mucho más de esta forma, aplicando los conocimientos en la práctica, como hicimos, que estando en un aula y leyendo un libro”. En algunos casos, expresaron haber logrado la adquisición de competencias de comunicación: “sirvió mucho para darse cuenta cómo llegar a la gente, para sacarnos la vergüenza ya que no fue fácil hablar para una cámara o en un congreso ¡tuvimos que aprender!”, señalando que la experiencia “me enseñó los verdaderos procesos por los que pasa un proyecto, como tantos que se hacen en la facultad todos los años […]me sirvió para conocer otras formas de vida, realidades distintas y pensamientos nuevos.”
Metodología de evaluación: en esta categoría es interesante mencionar que el Plan de Estudios vigente propugna la evaluación continua y desde el Taller 1 se adhiere a través de la elección de instrumentos de evaluación que la garantizan, como por ejemplo, la elaboración de un portafolios que evidencia tanto los logros como los obstáculos de aprendizaje durante todo el año. En este marco, la propuesta en el PHMB incluyó dentro del portafolios numerosas instancias de evaluación no formales que fueron percibidas por los estudiantes como oportunidades de autoevaluarse y mejorar sus modos de aprender: “no sé cuándo evaluaron, no me di cuenta qué evaluaban” y “los docentes nos preguntaban cuestiones prácticas sobre cómo ayudar a los huerteros y teníamos que buscar una solución, pensar…”. Se
Ciencias Agronómicas - Revista XX - Año 12 - 2012 / 015 - 021
Los cultivos de cobertura en la horticultura familiar urbana: una oportunidad para mejorar la formación del estudiante de agronomía

-21-
evidencia el logro de una evaluación de la adquisición de competencias por medio de la confrontación con los problemas que realmente iban surgiendo y donde ellos tenían que demostrar “saber, saber hacer y saber ser”. En especial, se destaca la adquisición de competencias transversales vinculadas a la sensibilización frente a los problemas sociales: “me gustó salir de las 'cuatro paredes´ y acercarnos, en este caso, a productores hortícolas de bajos recursos económicos, con los cuales intercambiamos ideas y opiniones, fue muy productivo y me hizo abrir la cabeza”.
En síntesis, a partir de la observación de los estudiantes en el contexto y de las expresiones de los encuestados se verificó la adquisición de competencias pre-profesionales vinculadas al ejercicio de la agronomía y al desarrollo de un compromiso ciudadano frente a situaciones de emergencia social. Por otra parte, es importante destacar la valoración de los saberes culturales provenientes de una comunidad no científica muchas veces relegados y desestimados por los profesionales en el ámbito laboral: “aprendí cómo los huerteros a partir de sus propios conocimientos y experiencia, desarrollan técnicas de compostado [...] aprenden a mezclar hormigueros, a intercalar cultivos… ¡para evitar usar agroquímicos!”.
Conclusiones
Los cultivos de cobertura protegen al suelo de la pérdida de humedad y contienen altos contenidos de materia verde que aportan gran cantidad de nutrientes y oligoelementos, beneficiosos para el desarrollo de las plantas. Los resultados demuestran que la técnica de manejos de cultivos de cobertura es apropiada para mejorar las condiciones físico-químicas de suelos degradados en un parque huerta periurbano.
Para el caso de la comunidad huertera del PHMB, destacamos su idiosincrasia vinculada al contexto de surgimiento y el acompañamiento del Estado a través de una política pública basada en la agricultura familiar urbana con un enfoque agroecológico. El patrimonio cultural de los huerteros se pudo articular con el desarrollo urbano, dando origen a un proyecto que trasciende lo económico adquiriendo una dimensión socioeducativa singular, vinculada al logro de una alimentación sana y a la preservación de los recursos naturales. En este contexto, la adopción de cultivos de cobertura para recuperar los suelos es factible y su apropiación por parte de los huerteros denota que fue percibido como beneficioso.
Por otra parte, los estudiantes estimaron la experiencia como valiosa y superadora de las propuestas educativas más tradicionales destacando la importancia de “salir de las aulas” para integrar, aplicar y significar los aprendizajes. Valoraron la experticia de una comunidad no científica y la oportunidad de establecer un vínculo solidario en la búsqueda de soluciones a problemas agronómicos. Los ensayos con cultivos de cobertura para su aplicación en la agricultura familiar urbana fueron ventajosos para lograr entornos de aprendizaje colaborativos y estilos de enseñanza orientados a la comprensión, donde el estudiante es el protagonista y, además de adquirir conocimientos, reflexiona sobre su forma de aprender.
Finalmente, se advierte que esta investigación en el PHMB significó un desafío en tanto requirió de una mirada integradora de lo heterogéneo, capaz de alcanzar un conocimiento multidimensional a partir de una relación “sujeto investigador/objeto de estudio” racional y emocional. Aun cuando, por cuestiones estrictamente metodológicas, el diseño requiriera de una escisión en las posibles dimensiones del problema.
Bibliografía
1. LATTUCA A (2012) La agricultura urbana como política pública: el caso de la ciudad de Rosario, Argentina. Revista Agroecología. Facultad de Biología. Universidad de Murcia. http://www.spta.org.br-wp-content-uploads-2012-04-Agroecologia6-2012-CTP.pdf Acceso: 17 de julio de 2012.
2. VARELA M, FERNANDEZ P, RUBIO, G, TABOADA M (2011) Cultivos de cobertura: efectos sobre la macroporosidad y la estabilidad estructural de un suelo franco-limoso. . Ciencia del suelo [online] vol.29, nº1, pp. 9 9 - 1 0 6 . D i s p o n i b l e e n : http://www.scielo.org.ar/scielo.php?script=sci_arttext&pid=S1850-20672011000100011&lng=es&nrm=iso. ISSN 1850-2067. Acceso: 12 de julio de 2012.
3. ALTIERI M, NICHOLS C (2012). Agroecología: única esperanza para la soberanía alimentaria y la resiliencia socioecológica. Una contribución a las discusiones de Rio+20 sobre temas en la interface del hambre, la agricultura, y la justicia ambiental y social http://www.wae-energy.com/job/lapluma/es/images/stories/documents_periodicos_app/Agroecologia_para_la_resiliencia-1.pdf2012. Acceso: 12 de julio de 2012.
4. OTTMANN G, SEVILLA GUZMÁN E, LATTUCCA A (2009) La Agroecología como Propuesta de Educación Ambiental para la Sustentabilidad Social en Rosario. Argentina. Revista Brasileira de Agroecologia. Resumos do VI CBA e II CLAA. Vol. 4 Nº 2 p.800-804.
5. SARANDON S, HONG, G (1995) El Rol de la Universidad en la Incorporación de un Enfoque Agroecológico para el Desarrollo Sostenible. Revista de CLADES Agroecología y Desarrollo. Numero Especial 8/9. http://www.clades.cl Acceso: 12 de julio de 2012.
6. RUÉ J. (2009). El aprendizaje Autónomo en Educación Superior. Narcea. Madrid, España. p. 54-59.
7. CORONADO, M. (2009). Competencias docentes. Ampliación, enriquecimiento y consolidación de la práctica profesional. Noveduc. Buenos Aires, Argentina. p. 95-106.
8. TENUTTO M, BRUTTI C, ALGARAÑÁ S. (2010) Planificar, enseñar, aprender y evaluar por competencias. Conceptos y propuestas. La Imprenta Ya para Digital & Papel. Buenos Aires, Argentina. p. 24.
Ciencias Agronómicas - Revista XX - Año 12 - 2012 / 015 - 021
MILO VACCARO, M.; MUÑOZ, G.; TORRES, C.; COURETOT, J.


-23-
Artículo Original / Recibido 20/07/2012 - Aceptado 24/11/2012
Desarrollo de una base de datos genéticos para lacaracterización del germoplasma argentino de soja
Construction of a genetic database to characterize the argentine soybean germplasm
1 2 2OLSINA, Cesar ; CAIRO, Carlos ; PESSINO, Silvina1 Bolsa de Comercio de RosarioRosario, Santa Fe, Argentina, [email protected] Facultad de Ciencias Agrarias, UNRParque Villarino, S2125ZAA, Zavalla, Argentina,[email protected], [email protected]
Argentina es actualmente el primer exportador mundial de aceites y harinas de soja y el tercer productor mundial de semillas de soja. El objetivo de este trabajo fue desarrollar un banco de datos genéticos basado en marcadores moleculares de SSR (Repeticiones de Secuencia Simple, o Microsatélites) y AFLP (polimorfismos en el largo de los fragmentos amplificados) para caracterizar el germoplasma argentino de soja, facilitar la identificación de variedades y asistir al mejoramiento genético. Se estudiaron 51 loci SSR y 15 combinaciones de cebadores AFLP en los 86 cultivares de soja más utilizados en Argentina. Los SSR revelaron en promedio 5 alelos por locus, con una diversidad génica media de 0,523. Los AFLP detectaron en promedio 17 bandas polimórficas por combinación de oligonucleótidos, con una diversidad génica media de 0,1924. Los 86 cultivares estudiados mostraron coeficientes de similitud genética medios de 0,292 y 0,921 para SSR y AFLP, respectivamente. La base de datos de SSR se validó en forma exitosa a través de la identificación de muestras incógnitas remitidas al laboratorio. Además, se seleccionó un subconjunto de 20 marcadores SSR altamente informativos que permite discriminar cualquier accesión del banco de manera simple y eficiente, y que pueden ser utilizados en estudios rápidos de identificación o diferenciación de nuevos cultivares. Los marcadores AFLP produjeron dendrogramas basados en la distancia genética donde los materiales agruparon de acuerdo al obtentor. Estos resultados sugieren que los marcadores AFLP reflejan fielmente las relaciones de parentesco entre las variedades, resultando apropiados para seleccionar materiales genéticamente divergentes.
Palabras clave:Glycine max-marcadores moleculares-huellas genéticas
Resumen
Argentina is currently the leading country in terms of soybean oil and flour exports, and ranks third in soybean seed production. The aim of this work was to develop a genetic database based on SSR (Simple Sequence Repeats, or Microsatellites) and AFLP (Amplified Fragment Length Polymorphisms) markers, in order to characterize the argentine soybean germplasm, allow cultivar identification and provide assistance to breeding. Fifty-one SSR loci and 15 AFLP primer combinations were analyzed on the 86 soybean cultivars most used in Argentina. The SSR markers revealed an average of 5 alleles per locus and a genic diversity of 0.523. The AFLP markers showed an average of 17 polymorphic bands per primer combination and a genic diversity of 0.1924. The 86 cultivars under analysis showed coefficients of mean genetic similarities of 0.292 and 0.921 for SSR and AFLP, respectively. The SSR database was successfully validated for the identification of unknown samples. A subset of 20 highly-informative SSR markers was selected to achieve an effective discrimination of any databank accession by means of a simple and efficient procedure, in order to be used for rapid identification analysis and differentiation of new cultivars. The AFLP markers produced dendrograms based on genetic distance in which the plant materials grouped according to the identity of the breeder. These results suggest that AFLP markers accurately reflect the parentage relationship among cultivars, and are appropriate to the selection of genetically divergent materials.
Keywords:Glycine max - molecular markers - genetic fingerprints
Abstract
Introducción
La soja (Glycine max L. Merrill) es una leguminosa nativa del Asia Oriental. Las evidencias arqueológicas más antiguas de su uso en China, Corea y Japón datan de circa 1000 AC, aunque se calcula que su utilización como alimento y en la composición de medicamentos comenzó hace aproximadamente 5000 años (Hymowitz & Newell, 1981; Dong et al., 2004; Zhao & Gai, 2004). En 1765 fue trasladada desde China a los Estados Unidos, observándose una notable expansión del cultivo a partir de 1840. El primer registro de su ingreso en Argentina data de 1862 (Martínez, 2010).
A principios de 1960 la producción argentina de soja comenzó a crecer sostenidamente, incrementándose desde 136.700 hasta 52.700.000 toneladas en el periodo 1971-2010 (Sistema Integrado de Información Agropecuaria, //www.siia.gov.ar/).
Pasó de ocupar 37.700 hectáreas en la campaña 1970/71 a 18.343.000 hectáreas en 2010 (informe GEA-BCR 2010, www.bcr.com.ar). Los principales factores que ocasionaron el asombroso aumento observado en la producción de soja en Argentina fueron el uso de la siembra directa, los altos precios internacionales del complejo soja debido a la importante demanda asiática, el desarrollo de un gran número de variedades adaptadas a las distintas zonas de producción argentinas y la introducción de cultivares transgénicos resistentes a herbicidas (Olsina, 2012). Argentina es actualmente: 1) el primer exportador mundial de aceites y harinas de soja, con un volumen de exportación de 5,4 millones de toneladas de aceite y 27,3 millones de toneladas de harina de soja en el año 2010; 2) el tercer productor mundial de soja, con 52,7 millones de toneladas, luego de Estados Unidos y Brasil, según datos de la
Ciencias Agronómicas - Revista XX - Año 12 - 2012 / 023 - 039

-24-
campaña 2009/2010 (Sistema Integrado de Información Agropecuaria, //www.siia.gov.ar/). Los granos, harinas y aceites representan el 25 % del valor total de las exportaciones argentinas (Leavy & Dewes, 2009). Considerando la producción conjunta de Argentina y Brasil, los países del MERCOSUR ocupan el primer lugar en la producción de soja a nivel mundial.
En Argentina existe un número significativo de semilleras locales que se dedican al mejoramiento de este cultivo. En el país está vigente una legislación que regula los derechos de propiedad intelectual de las obtenciones vegetales, y controla su propagación y comercialización. Se trata de la Ley de Semillas y Creaciones Fitotécnicas Nº 20.247, que establece que un cultivar puede ser objeto de título de propiedad cuando reúna condiciones de distinguibilidad, homogeneidad y estabilidad que permitan la caracterización del mismo (Bonamico et al., 2004). El organismo responsable de su aplicación es el Instituto Nacional de Semillas (INASE), dependiente del Ministerio de Agricultura, Ganadería, Pesca y Alimentos de la Nación. Por su parte, el país está adherido a la Unión Internacional para la Protección de Nuevas Variedades de Plantas (UPOV/TG/1/2, 1979), lo que permite extender la confianza nacional e internacional en el ámbito de la concesión de propiedad al creador de una nueva variedad (Olsina, 2012).
Tradicionalmente la determinación de las condiciones de distinguibilidad, homogeneidad y estabilidad de un cultivar se realizaban a través de análisis morfológicos y bioquímicos. Pero en las últimas décadas, la implementación de las técnicas de marcadores moleculares basadas en el análisis de la estructura del ADN permitieron un estudio directo sobre el genotipo en lugar del fenotipo (Powell et al., 1996). Los marcadores moleculares tienen la ventaja de revelar la estructura de numerosos loci en la secuencia de bases del ADN, facilitando la identificación de polimorfismos genéticos, que en su mayoría no se reflejan en el fenotipo. En este contexto, la Oficina de Protección de Variedades Vegetales del Departamento de Agricultura de los Estados Unidos (USDA) acepta actualmente los perfiles alélicos de marcadores moleculares de secuencias simples repetitivas (SSR) como evidencia adicional de la singularidad de un nuevo cultivar (Diwan & Cregan, 1997; Song et al., 1999). Asimismo, diferentes organismos internacionales como la Organización Internacional para la Estandarización (ISO), la Asociación Internacional para el Análisis de Semillas (ISTA) y la Unión Internacional para la Protección de Nuevas Variedades de Plantas (UPOV) están realizando permanentes validaciones y estandarizaciones de marcadores moleculares aplicados a la identificación de variedades de los cultivos mayores, entre ellos la soja (Olsina, 2012).
Es un hecho ampliamente aceptado que G. max posee un bajo nivel de diversidad morfológica y genética. Por ello, la utilización de marcadores moleculares es particularmente útil en lo que se refiere a la diferenciación de cultivares de esta especie. Inicialmente se reportaron estudios basados en marcadores de RFLP (restriction fragment length polymorphisms) aplicados a su caracterización (Kein et al., 1992; Shoemaker et al., 1992). Luego se hizo más común el uso de métodos basados en la reacción de amplificación en cadena de la polimerasa (PCR) (Mullis et al., 1986), entre ellos los de RAPD (Random Amplified Polymorphic DNA) (Williams et al., 1990; Welsh & McClelland 1990), AFLP (Amplified Fragment Length Polymorphism) (Vos et al., 1995) y SSR (Simple Sequence Repeats), también conocidos como microsatélites (Weber, 1990; Akkaya et al., 1992).
El método de RAPD es simple, necesita mínimas cantidades de ADN y no requiere información previa de secuencia. Consiste en
la amplificación del ADN al azar, usando cebadores decámeros únicos de secuencia variable (Williams et al., 1990; Welsh & McClelland 1990). Sin embargo, se ha demostrado que en la soja los RAPDs tienen poca utilidad, debido el bajo nivel de polimorfismo detectado (Giancola et al., 2002; Powell et al., 1996; Brown-Guedira et al., 2000). Por otro lado, los AFLP (Vos et al., 1995) son muy efectivos porque revelan una gran cantidad de marcadores por unidad experimental, son muy reproducibles, no necesitan información de secuencia previa y requieren una baja cantidad de ADN de partida. La técnica de AFLP se basa en la digestión del ADN genómico simultáneamente con dos endonucleasas de restricción, una de ellas de corte frecuente y la otra de corte poco frecuente. Luego se ligan adaptadores específicos para cada uno de los extremos cohesivos generados por estas enzimas y se pre-amplifican los fragmentos utilizando cebadores complementarios a los adaptadores, que pueden o no presentar bases adicionales al azar en el extremo 3'. Finalmente, una porción seleccionada de los fragmentos se re-amplifican utilizando cebadores complementarios a los adaptadores, que poseen una, dos o tres bases de secuencia al azar en el extremo 3' (Vos et al., 1995). La utilización de diferentes combinaciones de cebadores con distintas bases al azar asegura la amplificación de diferentes grupos de fragmentos genómicos. Los AFLP son una buena opción cuando se quiere obtener una caracterización rápida de genotipos, ya que se puede generar una cantidad considerable de marcadores a partir de una única combinación de cebadores (Mueller & LaReesaWolfenbarger, 1999). Se han utilizado exitosamente para caracterizar el germoplasma en cultivares comerciales de soja como así también en poblaciones naturales (Maughan et al., 1996; Ude et al., 2003; Bonato et al., 2006). Por último, los SSR son marcadores con características únicas que los hacen valiosos en los experimentos de identificación, debido a su naturaleza multialélica y codominante, su abundancia relativa y su amplia cobertura genómica. Requieren solo pequeñas cantidades de ADN de partida y son muy reproducibles entre laboratorios (Powell et al., 1996; Jones et al., 1997). Sin embargo, deben ser desarrollados a partir del conocimiento de las secuencias específicas que rodean un sector genómico de repeticiones cortas, de entre dos y seis bases, localizadas en tándem, ya que para su obtención se diseñan cebadores que amplifiquen la región completa donde está incluida la repetición. Este requerimiento de conocimiento de secuencia previo puede representar un inconveniente en la caracterización de especies poco estudiadas, pero actualmente no representa un obstáculo para los investigadores que estudian las especies mayores, ya que en las bases de datos públicas suelen estar disponibles numerosas secuencias de oligonucleótidos para amplificar marcadores SSR desarrollados por otros equipos de investigación. Respecto al caso particular de la soja, el grupo del Dr. Cregan (Servicio de Investigación en Agricultura dependiente del Departamento de Agricultura de los Estados Unidos, ARS-USDA, Beltsville) llevó a cabo la identificación de numerosos loci de microsatélites y diseñó cebadores específicos para SSR. Finalmente se determinó la posición en el genoma de estos marcadores mediante mapeo genético (Akkaya et al. 1992; Cregan et al. 1994). Las secuencias de cada locus SSR, cebadores, condiciones experimentales para su amplificación y ubicación en el mapa de ligamiento de soja, se pueden encontrar en forma libre en internet en la página web: www.soybase.org. Se ha reportado un elevado nivel de polimorfismo en los loci de SSR de soja (Akkaya et al. 1992; Cregan et al. 1994; Rongwen et al. 1995; Diwan & Cregan 1997; Song et al. 1999; Brown-Guerida et al. 2000; Narvel et al. 2000; Meesang et al. 2001). Analizando 20 SSR, Diwan and Cregan (1997) fueron capaces de distinguir cultivares de soja comerciales que eran considerados idénticos sobre la base de análisis de RFLP, morfología y marcadores de pigmentación. Brown-Guerida et al. (2000) pudieron identificar grupos
Desarrollo de una base de datos genéticos para la caracterización del germoplasma argentino de soja
Ciencias Agronómicas - Revista XX - Año 12 - 2012 / 023 - 039

-25-
relacionados de germoplasma de soja, observando mayor diversidad con tres SSR que con 46 RAPDs. Priolli et al. (2002) diferenciaron grupos morfológicamente similares de 186 cultivares de soja brasileña utilizando únicamente 12 marcadores SSR.
En Argentina existe un único antecedente de caracterización molecular del germoplasma de soja. Se trata del estudio publicado por Giancola et al. (2002), donde se utilizaron distintos tipos de marcadores moleculares y descriptores clásicos con el fin de caracterizar y diferenciar el conjunto de las 100 variedades de soja de uso comercial más utilizadas en ese momento. Los autores concluyeron que la variabilidad genética de los cultivares comerciales de soja argentina era baja. Aun así, se pudieron obtener huellas genéticas (fingerprints) discriminatorias para las 100 variedades analizadas. De los marcadores moleculares utilizados (AFLP, SSR y RAPD), los marcadores SSR mostraron ser los más efectivos, además de manifestar la correlación más alta con los descriptores clásicos (Giancola et al, 2002). Según estos autores, los marcadores moleculares de SSR podrían aplicarse eficazmente para la clasificación del germoplasma de soja y su protección intelectual (Giancola et al. 2002).
Las variedades utilizadas en el estudio de Giancola et al. (2002) habían sido inscriptas antes de la aparición del evento GTS 40-3-2 de resistencia al herbicida glifosato, por lo que su participación en la siembra actual es muy baja o nula. La mayoría de estas variedades han caducado su derecho de propiedad hace ya algunos años, por lo que la base genética de los cultivares de soja argentina utilizados en la actualidad puede haber cambiado. Es por eso que resulta de interés realizar una actualización de la caracterización de los cultivares argentinos de soja, para proporcionar herramientas moleculares que puedan ser aplicadas a la protección de propiedad intelectual vegetal y la implementación de programas de mejoramiento. Este trabajo tuvo como objetivo desarrollar una metodología basada en matrices de datos genéticos de SSR y AFLP que pueda utilizarse para clasificar en forma inequívoca las variedades comerciales de soja más utilizadas actualmente en nuestro país, para contribuir a la protección de la propiedad intelectual, asegurar la distinguibilidad de nuevos materiales y aportar información aplicable a la selección eficaz de variedades para el mejoramiento.
Materiales y Métodos
Material VegetalSe investigaron 86 cultivares comerciales de soja liberados para su comercialización en Argentina (Tabla 1). El material genético fue obtenido a partir de los diferentes criaderos originadores (ACA, A. L. Marchionni, Don Mario, FN Semillas, La Tijereta, Nidera, Pioneer, Relmó, SPS, Syngenta). En forma adicional, se analizaron muestras incógnitas provenientes de multiplicadores o semilleros, cuya identidad genética probable o cultivar a contrastar fue indicada por los mismos (Tabla 2).
Extracción y cuantificación de ADN genómicoPara cada cultivar o muestra, se molieron 50 semillas en un molino a cuchillas A11 Basic (IKA-Works, Inc) con el objeto de representar en forma homogénea la constitución alélica (Diwan & Cregan, 1997). Una fracción de 200 mg de semillas molidas se procesó con el equipo de extracción de ADN genómico DNeasy Plant Extraction kit (Qiagen) de acuerdo a las instrucciones del fabricante. El ADN obtenido fue cuantificado utilizando un espectrofotómetro de fluorescencia Modulus (Turner Biosystems, Sunnyvale) a una longitud de onda de absorción y emisión de 360 y 415 nm, respectivamente. A 3 mL de una solución conteniendo 10 mM Tris (Promega), 1 mM EDTA (Promega), 200 mM NaCl (Merck) pH 7.4, Hoesch 33258 0,15 μg/mL (Sigma-Aldrich) se le adicionaron 5 μL de extracto de ADN. La curva de calibración del fluorómetro se realizó con 1000; 750; 500; 250 y 100 ng de ADN del fago Lambda (Promega). Los ADN se diluyeron con agua ultrapura hasta alcanzar una concentración de 10 ng/μL para los análisis de SSR y de 150 ng/μL para los análisis de AFLP, y se conservaron a -20°C hasta su uso. Estas diluciones y la posterior preparación de los cócteles de reacción de PCR fueron llevadas a cabo en una campana de flujo laminar NuAire® (Plymouth, Minnesota), de manera de minimizar la posibilidad de contaminaciones.
Experimentos de SSRLa técnica de SSR se implementó en base al polimorfismo y la cobertura genómica reportados previamente para soja (Cregan et al., 1999; Song et al., 2004). Se seleccionaron 51 marcadores SSR obtenidos a partir de Soybase (www.soybase.org) (Tabla 3). Las reacciones de PCR se llevaron a cabo según se describe en Akkaya et al. (1995), con pequeñas modificaciones. Las mezclas
de reacción incluyeron 60 ng de ADN genómico, solución amortiguadora de Platinum Taq 1 X, 1.5 mM MgCl2, 100 nM de cada cebador, 0,5 U de Platinum Taq ADN Polimerasa (Invitrogen) y 0,2 μM de dNTP (Promega), en un volumen final de 25 μL. Las condiciones para la amplificación fueron: desnaturalización inicial del ADN a una temperatura de 95ºC durante 3 minutos, 40 ciclos de desnaturalización a una temperatura de 95ºC durante 50 segundos, hibridación a 48°C durante 40 segundos y extensión a 72ºC durante 60 segundos, en un ciclador térmico programable modelo ABI 9600 (Applied Biosystems). A los productos de amplificación obtenidos se les agregaron 5 μL de solución de siembra (formamida 95% P/V, NaOH 10 mM y azul de bromofenol 2 mg/ml) y se los almacenó a -20°C hasta su uso. Luego fueron resueltos en geles de 6 % de acrilamida (BioRad), 0,3 % de metilen-bisacrilamida (BioRad) y 7,5 M de urea (Invitrogen), 360 μL de persulfato de amonio al 10 % (APS, Sigma Aldrich), 36 μL de N,N,N2,N1-tetrametil-etilendiamino (TEMED, Sigma Aldrich) y corridos en una cuba vertical Sequigen GT (BioRad) utilizando como tampón de corrida 0,5X TBE [50 mM de Tris, 50 mM de ácido bórico y 1 mM de EDTA (pH 8)] en el reservorio superior y 1X TBE en el inferior. Previo a la corrida electroforética, se realizó una precorrida de la matriz a una potencia constante de 90 W por aproximadamente 1 hora hasta que el gel alcanzó una temperatura homogénea de 50 ºC. Se cargaron 5μL de cada producto de amplificación por calle del gel y al menos 3 calles con marcador de peso molecular de 10 pares de base de separación entre bandas (10 bp DNA Ladder, Invitrogen). La corrida electroforética se realizó a una potencia constante de 85 W por 2 horas, con una temperatura del gel de 50°C. Para visualizar los marcadores se utilizó el método de tinción con nitrato de plata descripto en el manual “DNA Silver Staining System” (Promega). Los pasos que se siguieron se detallan a continuación: fijación del gel en ácido acético al 10 % por 20 minutos con agitación suave; 3 lavados de 2 minutos con agua destilada con agitación suave; tinción por incubación de 30 minutos con agitación suave en una solución conteniendo: 1 g/L de nitrato de plata (MERCK) y 0,05% de formaldehído; lavado con agua destilada por 10 segundos; revelado con 2 L de solución reveladora conteniendo 30 g/l de carbonato de sodio (MERCK), 0,05 % de formaldehído y 2,5 mg/L de tiosulfato de sodio hasta
OLSINA, Cesar; CAIRO, Carlos; PESSINO, Silvina
Ciencias Agronómicas - Revista XX - Año 12 - 2012 / 023 - 039

-26-
la observación de las bandas; finalización de la reacción de revelado por el agregado de 1 L de ácido acético al 10 % por 2 minutos; 2 lavados de 2 minutos con agua destilada; secado en estufa a 37 ºC. Los patrones de bandas se analizaron observando los vidrios sobre un transiluminador de luz blanca. Finalmente, los geles teñidos fueron registrados para su documentación utilizando un escáner DCP-8065 DN (Brother).
Experimentos de AFLPLa generación de marcadores moleculares de AFLP se realizó según lo descrito en Vos et al. (1995), con las modificaciones
pertinentes para la utilización de tinción con nitrato de plata en lugar del empleo de radioactivo (Olsina, 2012). Los oligonucleótidos utilizados en los experimentos de AFLP fueron diseñados por Key Gene (AFLP Protocol for Public Release, Version 2.0, Netherlands, 1994) y sintetizados por Invitrogen Life Technologies. El ADN genómico de las líneas analizadas fue digerido con dos enzimas de restricción simultáneamente: EcoRI (G/AATTC), de corte poco frecuente y MseI (T/TAA), de corte frecuente. Las mezclas de digestión enzimática incluyeron: 600 ng de ADN genómico, 1X de amortiguador NEBEcoRI 1X (50 mM NaCl 100, mM,Tris-HCl, 10 mM MgCl2, 0.025 %
Cultivar Obtentor Grupo de madurez Cultivar Obtentor Grupo de madurez
ACA360 ACA III TJS2068 La Tijereta VII
ACA420 ACA IV TJS2170 La Tijereta VII
ACA470 ACA IV TJS2171IR La Tijereta VII
ACA480 ACA IV TJS2130 La Tijereta III
ACA530 ACA V A3901RG Nidera III
ACA570 ACA V A6445RG Nidera VI
ALM3530 A. L.Marchionni III A5417RG Nidera V
ALM4200 A. L.Marchionni IV A5520RG Nidera V
ALM4500 A. L.Marchionni IV A4910RG Nidera IV
ALM4650 A. L.Marchionni IV A4725RG Nidera IV
DM2000 Don Mario II A5901RG Nidera V
DM2200 Don Mario II A4303RG Nidera IV
DM3100 Don Mario III A8000RG Nidera VIII
DM3700 Don Mario III A5409RG Nidera V
DM4200 Don Mario IV A6019RG Nidera VI
DM4400 Don Mario IV A4505RG Nidera IV
DM4600 Don Mario IV A5009RG Nidera IV
DM4670 Don Mario IV A7636RG Nidera VII
DM4800 Don Mario IV A4613RG Nidera IV
DM4870 Don Mario IV 94B73 Pioneer IV
DM5800 Don Mario V Rosario65 Relmó VI
DM6200 Don Mario VI ANTA82 Relmó VIII
DM50048 Don Mario V Mercedes70 Relmó VII
DM5.1i Don Mario V Ayelen22 Relmó II
DM8002 Don Mario VIII Azul35 Relmó III
DM4250 Don Mario IV Maravilla45 Relmó IV
DM3500 Don Mario III Natalia49 Relmó IV
DM5.2i Don Mario V NvaMaría55 Relmó V
DM4970 Don Mario IV Rafaela58 Relmó V
DM7.0i Don Mario VII SPS3900 SPS III
DM5.8i Don Mario V SPS4X0 SPS IV
DM5.5i Don Mario V SPS4X4 SPS IV
FN360 FN semillas III SPS4500 SPS IV
FN410 FN semillas IV SPS500 SPS V
FN485 FN semillas IV SPS5X5 SPS V
TJS2136 La Tijereta III SPS6X0 SPS VI
TJS2138 La Tijereta III SPS6X2 SPS VI
TJS2145 La Tijereta IV SPS8X0 SPS VIII
TJS2049 La Tijereta IV SPS9800 SPS IX
TJS2156 La Tijereta V NKCoker68 Syngenta VI
TJS2164 La Tijereta VI NK37-00 Syngenta III
TJS2165 La Tijereta VI NKCoker75 Syngenta VII
TJS2139 La Tijereta III NKMireya Syngenta IV
Tabla 1: Cultivares de soja utilizados en el presente estudio. Se indica el grupo de madurez y el obtentor.
Desarrollo de una base de datos genéticos para la caracterización del germoplasma argentino de soja
Ciencias Agronómicas - Revista XX - Año 12 - 2012 / 023 - 039

-27-
a identidad putativa declarada por el solicitante, quien requirió su comparación con cultivares testigo.b Marcadores de SSR utilizados para el análisis particular de cada muestra
Tabla 2: Muestras incógnitas de soja utilizadas en el presente estudio
Muestra incógnita Identidad putativa a Marcadores utilizados en el análisis b
11209 A5009RG Satt 324; 173; 30; 175; 294; 242; 373; 009; 147; 141; 197; 191; 458; 308; 237; 436;
453; 511; 281; 311; 181; 180; 94 y 005
11210 DM4870 Satt 324; 173; 30; 175; 294; 242; 373; 009; 147; 141; 197; 191; 458; 308; 237; 436;
453; 511; 281; 311; 181; 180; 94 y 005
11995 A4613RG Satt 534; 324; 173; 30; 70; 231; 175; 242; 373; 09; 147; 38; 197; 243; 475; 308 y
237
1023 DM4870 Satt 534; 324; 173; 30; 42; 70; 231; 175; 242; 414 009; 147; 038; 141; 197; 253;
191; 114; 117; 577;
DM4200
DM3700
1355 A6019RG Satt 534; 324; 173; 231; 294; 242; 373; 191; 030; 577; 147; 141; 453; 197; 237;
175; 009; 038; 253; 308
1357 A7636RG Satt 534; 324; 173; 231; 294; 242; 373; 197; 191; 030; 577; 147; 141; 453; 197;
237; 175; 009; 038; 253; 308
DM4600
A4303RG
1269 Satt 534; 324; 173; 231; 294; 242; 373; 197; 191; 030; 577; 147; 141; 453; 197;
237; 175; 009; 038; 253; 308
1262 Satt 242; 141; 475; 197; 458; 308; 177; 114; 577; 175; 42; 324; 173; 30; 373; 42
Triton X-100 pH 7.5, New England Biolab, Inc), 5 U de cada enzima de restricción, 100 μg/mL de albúmina sérica bovina (New England Biolabs, Inc) y agua tridestilada estéril hasta completar un volumen final de 50 μL. La mezcla de reacción fue incubada a 37 ºC durante 2 horas. Posteriormente, se procedió a ligar de los adaptadores de AFLP a los fragmentos digeridos. La mezcla de reacción de ligación incluyó: 5 pmol del adaptador EcoRI, 50 pmol del adaptador MseI, 1X de solución tampón de T4 DNA Ligase (50 mM Tris-HCl, 10 mM MgCl2, 1 mM ATP,10 mM DTT pH 7.5 New England Biolabs, Inc), 1 U de enzima T4 DNA Ligasa (New England Biolabs, Inc) y agua ultrapura hasta completar un volumen de 10 μL. La mezcla de ligación se agregó directamente a los productos de la reacción de corte, se incubó a 37 ºC por 1 hora, y luego se inactivó la enzima T4 DNA Ligasa, incubando a 65°C por 10 minutos. Las reacciones de amplificación se realizaron en dos pasos: (i) reacción de pre-amplificación usando cebadores de AFLP EcoRI/MseI con un nucleótido selectivo en el extremo 3' (+1), y (ii) reacciones de amplificación selectiva usando cebadores alternativos de AFLP EcoRI/MseI con tres nucleótidos selectivos en el extremo 3' (+3). Una vez realizado el ligado de los adaptadores, se llevó a cabo la primera reacción de PCR o pre-amplificación en un volumen final de 25 μL conteniendo: 2 μl de la mezcla de corte y ligación, 1X de amortiguador Taq (Invitrogen), 0,2 M de cada dNTP (Promega), 1,5 mM de MgCl , 1 U de enzima Taq 2
polimerasa (Invitrogen), 1,2 ng/μL de cada cebador EcoRI/MseI +1 (Tabla 1) y agua ultrapura estéril hasta completar el volumen final. Las condiciones de pre-amplificación fueron las siguientes: una etapa de desnaturalización inicial de 30 segundos a 94 ºC seguida de 20 ciclos de 30 segundos a 94 ºC, 30 segundos a 60 ºC y 1 minuto a 72 ºC. Luego se realizaron reacciones de amplificación selectiva, utilizando 15 combinaciones de los cebadores de AFLP EcoRI/MseI (+3). El producto de pre-amplificación se diluyó 10 veces en amortiguador TE [10 mM Tris-HCl y 0,1 mM EDTA (pH 8)] y en estas condiciones se utilizó como molde en las amplificaciones subsiguientes. Las reacciones de amplificación selectiva se realizaron en un
volumen final de 10 μL conteniendo: 2 μL del producto de amplificación diluido, 1X de amortiguador Taq (Invitrogen), 0,2 M de cada dNTP (Promega), 1,5 mM de MgCl , 1 U de enzima 2
Taq polimerasa (Promega), 1.2 ng/μL de cada cebador EcoRI/MseI +3 (Tabla 2) y agua ultrapura hasta completar el volumen final. Las condiciones de amplificación fueron las mismas que las descriptas en Vos et al. (1995): un primer ciclo de 30 segundos a 94 ºC, 30 segundos a 65 ºC y 1 minuto a 72 ºC, seguido de 12 ciclos donde la temperatura de hibridación (65 ºC) se disminuyó en 0,7 ºC por ciclo, y finalmente 23 ciclos de 30 segundos a 94 ºC, 30 segundos a 56 ºC y 1 minutos a 72 ºC. Los productos de amplificación fueron suplementados con 5 μL de solución tampón de siembra (formamida 95% p/v, NaOH 10 mM y azul de bromofenol 2 mg/mL), denaturalizados a 96 ºC por 5 min, y resueltos en geles de poliacrilamida, en una cuba vertical Sequigen GT (BioRad). La matriz del gel se preparó usando 4,75 % de acrilamida (BioRad), 0,25 % de metilen-bisacrilamida (BioRad) y 7,5 M de urea (Invitrogen) en presencia de 0,5X de amortiguador TBE [50 mM de Tris, 50 mM de ácido bórico y 1 mM de EDTA (pH 8)]. A 80 mL de esta solución se agregaron 360 μl de persulfato de amonio al 10 % (APS, Sigma Aldrich) y 36 μL de N,N,N2,N1-tetrametil-etilendiamino (TEMED, Sigma Aldrich) para permitir la polimerización. Se realizó una precorrida de la matriz a una potencia constante de 60 W por aproximadamente 1 hora, hasta que el gel alcanzó una temperatura homogénea de 50 ºC. Se utilizó amortiguador 0,5X TBE en el reservorio superior de la cuba y 1X TBE en el reservorio inferior. Se sembraron 5 μL de cada muestra en los correspondientes pocillos y se realizó la corrida electroforética a una potencia constante de 60 W por 1 hora 30 minutos. Los marcadores se revelaron utilizando el procedimiento de tinción con plata descripto anteriormente. Los marcadores moleculares fueron identificados de acuerdo a la combinación de cebadores de AFLP utilizada, empleando la denominación alfa-numérica establecida por los proveedores seguido por un número en orden de mayor a menor peso molecular en forma sucesiva (Olsina, 2012).
OLSINA, Cesar; CAIRO, Carlos; PESSINO, Silvina
Ciencias Agronómicas - Revista XX - Año 12 - 2012 / 023 - 039
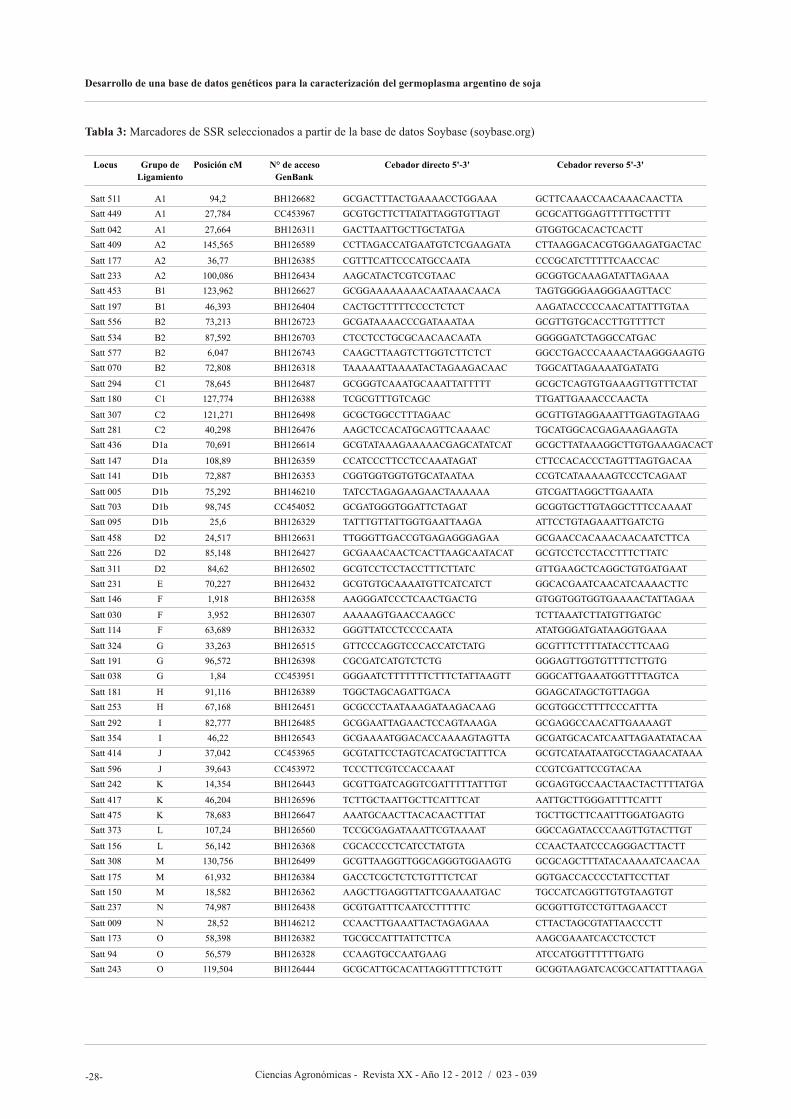
-28-
Locus Grupo de
Ligamiento
Posición cM N° de acceso
GenBank
Cebador directo 5'-3' Cebador reverso 5'-3'
Satt 511 A1 94,2 BH126682 GCGACTTTACTGAAAACCTGGAAA GCTTCAAACCAACAAACAACTTA
Satt 449 A1 27,784 CC453967 GCGTGCTTCTTATATTAGGTGTTAGT GCGCATTGGAGTTTTTGCTTTT
Satt 042 A1 27,664 BH126311 GACTTAATTGCTTGCTATGA GTGGTGCACACTCACTT
Satt 409 A2 145,565 BH126589 CCTTAGACCATGAATGTCTCGAAGATA CTTAAGGACACGTGGAAGATGACTAC
Satt 177 A2 36,77 BH126385 CGTTTCATTCCCATGCCAATA CCCGCATCTTTTTCAACCAC
Satt 233 A2 100,086 BH126434 AAGCATACTCGTCGTAAC GCGGTGCAAAGATATTAGAAA
Satt 453 B1 123,962 BH126627 GCGGAAAAAAAACAATAAACAACA TAGTGGGGAAGGGAAGTTACC
Satt 197 B1 46,393 BH126404 CACTGCTTTTTCCCCTCTCT AAGATACCCCCAACATTATTTGTAA
Satt 556 B2 73,213 BH126723 GCGATAAAACCCGATAAATAA GCGTTGTGCACCTTGTTTTCT
Satt 534 B2 87,592 BH126703 CTCCTCCTGCGCAACAACAATA GGGGGATCTAGGCCATGAC
Satt 577 B2 6,047 BH126743 CAAGCTTAAGTCTTGGTCTTCTCT GGCCTGACCCAAAACTAAGGGAAGTG
Satt 070 B2 72,808 BH126318 TAAAAATTAAAATACTAGAAGACAAC TGGCATTAGAAAATGATATG
Satt 294 C1 78,645 BH126487 GCGGGTCAAATGCAAATTATTTTT GCGCTCAGTGTGAAAGTTGTTTCTAT
Satt 180 C1 127,774 BH126388 TCGCGTTTGTCAGC TTGATTGAAACCCAACTA
Satt 307 C2 121,271 BH126498 GCGCTGGCCTTTAGAAC GCGTTGTAGGAAATTTGAGTAGTAAG
Satt 281 C2 40,298 BH126476 AAGCTCCACATGCAGTTCAAAAC TGCATGGCACGAGAAAGAAGTA
Satt 436 D1a 70,691 BH126614 GCGTATAAAGAAAAACGAGCATATCAT GCGCTTATAAAGGCTTGTGAAAGACACT
Satt 147 D1a 108,89 BH126359 CCATCCCTTCCTCCAAATAGAT CTTCCACACCCTAGTTTAGTGACAA
Satt 141 D1b 72,887 BH126353 CGGTGGTGGTGTGCATAATAA CCGTCATAAAAAGTCCCTCAGAAT
Satt 005 D1b 75,292 BH146210 TATCCTAGAGAAGAACTAAAAAA GTCGATTAGGCTTGAAATA
Satt 703 D1b 98,745 CC454052 GCGATGGGTGGATTCTAGAT GCGGTGCTTGTAGGCTTTCCAAAAT
Satt 095 D1b 25,6 BH126329 TATTTGTTATTGGTGAATTAAGA ATTCCTGTAGAAATTGATCTG
Satt 458 D2 24,517 BH126631 TTGGGTTGACCGTGAGAGGGAGAA GCGAACCACAAACAACAATCTTCA
Satt 226 D2 85,148 BH126427 GCGAAACAACTCACTTAAGCAATACAT GCGTCCTCCTACCTTTCTTATC
Satt 311 D2 84,62 BH126502 GCGTCCTCCTACCTTTCTTATC GTTGAAGCTCAGGCTGTGATGAAT
Satt 231 E 70,227 BH126432 GCGTGTGCAAAATGTTCATCATCT GGCACGAATCAACATCAAAACTTC
Satt 146 F 1,918 BH126358 AAGGGATCCCTCAACTGACTG GTGGTGGTGGTGAAAACTATTAGAA
Satt 030 F 3,952 BH126307 AAAAAGTGAACCAAGCC TCTTAAATCTTATGTTGATGC
Satt 114 F 63,689 BH126332 GGGTTATCCTCCCCAATA ATATGGGATGATAAGGTGAAA
Satt 324 G 33,263 BH126515 GTTCCCAGGTCCCACCATCTATG GCGTTTCTTTTATACCTTCAAG
Satt 191 G 96,572 BH126398 CGCGATCATGTCTCTG GGGAGTTGGTGTTTTCTTGTG
Satt 038 G 1,84 CC453951 GGGAATCTTTTTTTCTTTCTATTAAGTT GGGCATTGAAATGGTTTTAGTCA
Satt 181 H 91,116 BH126389 TGGCTAGCAGATTGACA GGAGCATAGCTGTTAGGA
Satt 253 H 67,168 BH126451 GCGCCCTAATAAAGATAAGACAAG GCGTGGCCTTTTCCCATTTA
Satt 292 I 82,777 BH126485 GCGGAATTAGAACTCCAGTAAAGA GCGAGGCCAACATTGAAAAGT
Satt 354 I 46,22 BH126543 GCGAAAATGGACACCAAAAGTAGTTA GCGATGCACATCAATTAGAATATACAA
Satt 414 J 37,042 CC453965 GCGTATTCCTAGTCACATGCTATTTCA GCGTCATAATAATGCCTAGAACATAAA
Satt 596 J 39,643 CC453972 TCCCTTCGTCCACCAAAT CCGTCGATTCCGTACAA
Satt 242 K 14,354 BH126443 GCGTTGATCAGGTCGATTTTTATTTGT GCGAGTGCCAACTAACTACTTTTATGA
Satt 417 K 46,204 BH126596 TCTTGCTAATTGCTTCATTTCAT AATTGCTTGGGATTTTCATTT
Satt 475 K 78,683 BH126647 AAATGCAACTTACACAACTTTAT TGCTTGCTTCAATTTGGATGAGTG
Satt 373 L 107,24 BH126560 TCCGCGAGATAAATTCGTAAAAT GGCCAGATACCCAAGTTGTACTTGT
Satt 156 L 56,142 BH126368 CGCACCCCTCATCCTATGTA CCAACTAATCCCAGGGACTTACTT
Satt 308 M 130,756 BH126499 GCGTTAAGGTTGGCAGGGTGGAAGTG GCGCAGCTTTATACAAAAATCAACAA
Satt 175 M 61,932 BH126384 GACCTCGCTCTCTGTTTCTCAT GGTGACCACCCCTATTCCTTAT
Satt 150 M 18,582 BH126362 AAGCTTGAGGTTATTCGAAAATGAC TGCCATCAGGTTGTGTAAGTGT
Satt 237 N 74,987 BH126438 GCGTGATTTCAATCCTTTTTC GCGGTTGTCCTGTTAGAACCT
Satt 009 N 28,52 BH146212 CCAACTTGAAATTACTAGAGAAA CTTACTAGCGTATTAACCCTT
Satt 173 O 58,398 BH126382 TGCGCCATTTATTCTTCA AAGCGAAATCACCTCCTCT
Satt 94 O 56,579 BH126328 CCAAGTGCCAATGAAG ATCCATGGTTTTTTGATG
Satt 243 O 119,504 BH126444 GCGCATTGCACATTAGGTTTTCTGTT GCGGTAAGATCACGCCATTATTTAAGA
Tabla 3: Marcadores de SSR seleccionados a partir de la base de datos Soybase (soybase.org)
Desarrollo de una base de datos genéticos para la caracterización del germoplasma argentino de soja
Ciencias Agronómicas - Revista XX - Año 12 - 2012 / 023 - 039

-29-
Análisis de perfiles genéticos y establecimiento de matrices binariasLa lectura de cada gel se llevó a cabo en forma manual de manera independiente por dos operadores. Las bandas intensas, claramente diferenciables de las espurias, y confirmadas por ambos operadores, fueron utilizadas en el análisis. Las bandas que mostraron igual movilidad electroforética fueron consideradas como idénticas, y se asumió que representaban el mismo alelo. El tamaño en pares de bases (pb) de los alelos de cada uno de los marcadores SSR fue estimado mediante la aplicación de una curva de logaritmo del peso molecular vs distancia de migración desde el pocillo hasta el centro de la banda, utilizando como referencia un marcador de peso molecular de 10 pb de separación, y procesando los datos con el sistema Excel®. El cálculo del tamaño de los alelos SSR se realizó por duplicado. Las bandas de ADN obtenidas del análisis de los marcadores de SSR y AFLP fueron registradas como 1 (presencia) o 0 (ausencia) y los resultados fueron almacenados en una hoja de cálculo de Excel® en forma de matriz numérica binaria.
Análisis EstadísticosLa comparación de la eficiencia discriminatoria de cada marcador se realizó en base a los siguientes parámetros calculados por unidad de ensayo (U), según Powell et al. (1996).: 1) Número de bandas polimórficas (np); 2) Número de bandas monomórficas (nnp); 3) Promedio de bandas polimórficas por unidad de ensayo (np/U); 4) Número de loci (L); 5) Número de loci por unidad de ensayo: nu = L/U; 6) Índice de Diversidad Genética (ID) de los loci polimórficos para un marcador genético: ID = 1-Σpi2, donde pi es la frecuencia alélica para el i-ésimo alelo (Botstein et al, 1980); 7) Media aritmética del Índice de Diversidad Genética de los loci polimórficos: ICp = Σ IC/np; 8) Fracción de loci polimórficos: β = np/(np + nnp); 9) Relación múltiplex efectiva: E = nu x β; 10) Índice de marcador: MI = E x Hep. Para el cálculo de las matrices de similitud genética se utilizaron los coeficientes recomendados por Kosman & Leonard (2005), de acuerdo a la ploidía del organismo en estudio, tipo de marcador molecular y tipo de dominancia. En el caso de los SSR, para calcular la similitud entre pares de cultivares i y j se utilizó el coeficiente de similitud de Jaccard (1908), mientras que en el caso de marcadores AFLP se empleó el coeficiente de emparejamiento simple descripto en Sneath &
Sokal (1973). Con los datos se realizaron análisis de agrupamientos utilizando el método de apareamiento de grupos no ponderado mediante media aritmética (UPGMA) y se representaron como dendrogramas. Para determinar con cuánta exactitud los agrupamientos representaban las estimaciones de similitud genética entre los genotipos, se generó una matriz cofenética a partir de cada uno de los agrupamientos, la cual se comparó con la matriz de similitud correspondiente por la prueba de correspondencia de matrices de Mantel (Mantel, 1967). La misma estadística de Mantel fue utilizada para comparar las matrices de similitud, así como las matrices cofenéticas producidas por las técnicas de AFLP y SSR. Adicionalmente, para ambos marcadores moleculares se llevó a cabo un análisis de ordenamiento mediante la técnica de Análisis de Coordenadas Principales (Gower, 1966) y se construyó un árbol de mínima distancia no sesgada (Nei, 1978). Todos estos procedimientos se realizaron por rutinas adecuadas en el programa NTSYSpc versión 2.17 (Exeter Software, Setauket, NY). Para comparar la media y varianza de un subconjunto seleccionado de 20 marcadores SSR en relación al conjunto de 51 marcadores iniciales, se realizó un test-t de comparación de medias y un test F para la comparación de varianzas. Además se realizaron histogramas para analizar las distribuciones de frecuencias de los coeficientes de similitud genética calculados para el subgrupo de 20 loci SSR versus el conjunto de 51. Todos estos cálculos, gráficos y test de hipótesis fueron realizados utilizando Excel® (Olsina, 2012).
Identificación de muestras incógnitasPara cada comparación se eligió un número particular de marcadores SSR procesables en un único gel, y que además presentasen alelos que permitiesen la diferenciación de los cultivares de referencia indicados como identidad putativa. Cada muestra fue comparada contra la base de datos para determinar la capacidad de los marcadores seleccionados para diferenciar correctamente los cultivares evaluados. Para ello se confeccionó una matriz de datos con los 86 cultivares previamente caracterizados, teniendo en cuenta los marcadores efectivamente evaluados en cada muestra y agregando la/s muestras reales analizadas. A cada una de estas matrices se le calculó la similitud genética y se obtuvo el correspondiente dendrograma.
Resultados
Caracterización genética con marcadores SSRLa caracterización de 86 cultivares seleccionados de soja fue realizada mediante la selección de 51 marcadores SSR descriptos para esta especie. Se seleccionaron entre 2 y 5 marcadores por cada uno de los grupos de ligamiento, con el objeto de lograr una buena cobertura genómica. Los marcadores mostraron perfiles electroforéticos claros y con buena resolución, amplificando casi siempre una sola banda por locus SSR. Esta observación está en total acuerdo con la estructura homocigota esperada para la mayoría de los loci en este cultivo (Diwan y Cregan, 1997), aunque algunos resultaron heterocigotas. Para lograr la caracterización de los cultivares analizados en este estudio con los 51 loci SSR, se realizaron más de 100 geles de poliacrilamida. La Figura 1, muestra un ejemplo correspondiente a una de esas corridas electroforéticas, donde se observa el perfil polimórfico de las bandas obtenidas.
En la Tabla 4 se muestran los resultados de la caracterización de los cultivares mediante marcadores SSR. Se obtuvieron un total de 242 bandas polimórficas, con un promedio de 5 bandas por locus, variando de 2 a 9 bandas. La frecuencia de ocurrencia observada de los 242 alelos varió de 0,01 a 0,95 con un promedio
de 0,22. Cuatro alelos mostraron una frecuencia de ocurrencia de 0,8 o mayor en los cultivares analizados y 27 alelos una frecuencia mayor a 0,5, mientras que 32 alelos mostraron frecuencias menores a 0,02. Estos alelos que muestran una muy baja frecuencia pueden ser útiles como marcadores diagnósticos para la identificación inequívoca de algunos de los cultivares de soja analizados, ya que poseen un alto poder de discriminación. Los valores de índice de diversidad génica (ID), que indican la probabilidad de que al tomar dos genotipos al azar a partir de una población de individuos ensayados estos sean polimórficos, variaron de 0,081 a 0,753 con un promedio de 0,523. Los marcadores SSR Satt 141 y 373 mostraron el ID más alto, con valores de 0,753 y 0,730 respectivamente, reflejando el alto número de alelos observados y una distribución de frecuencias alélicas uniformes (Figura 2). Por otro lado, los marcadores Satt 150 y Satt 475 presentaron los ID más bajos con valores de 0,111 y 0,081, respectivamente, mostrando un bajo número de alelos y frecuencias alélicas poco uniformes (Figura 2). Como es de esperarse, se observa que la variación del ID está significativa-mente asociada con el número de alelos detectados por locus (Figura 3).
OLSINA, Cesar; CAIRO, Carlos; PESSINO, Silvina
Ciencias Agronómicas - Revista XX - Año 12 - 2012 / 023 - 039
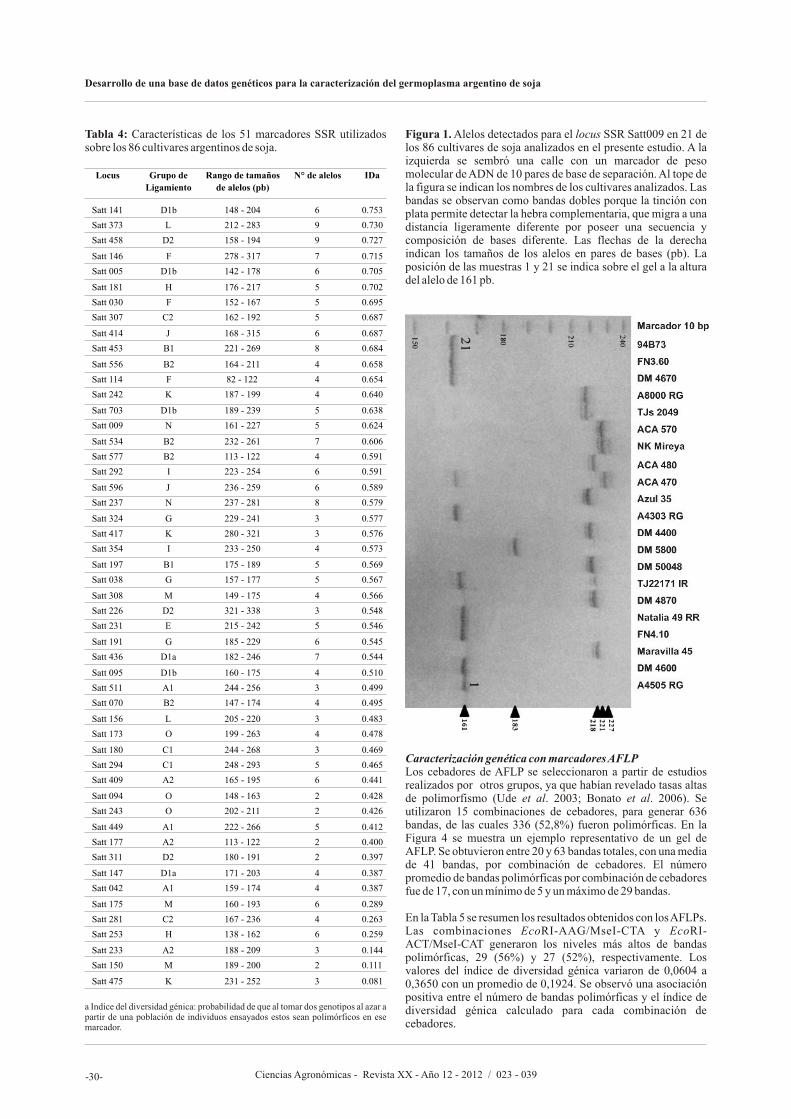
-30-
Caracterización genética con marcadores AFLPLos cebadores de AFLP se seleccionaron a partir de estudios realizados por otros grupos, ya que habían revelado tasas altas de polimorfismo (Ude et al. 2003; Bonato et al. 2006). Se utilizaron 15 combinaciones de cebadores, para generar 636 bandas, de las cuales 336 (52,8%) fueron polimórficas. En la Figura 4 se muestra un ejemplo representativo de un gel de AFLP. Se obtuvieron entre 20 y 63 bandas totales, con una media de 41 bandas, por combinación de cebadores. El número promedio de bandas polimórficas por combinación de cebadores fue de 17, con un mínimo de 5 y un máximo de 29 bandas.
En la Tabla 5 se resumen los resultados obtenidos con los AFLPs. Las combinaciones EcoRI-AAG/MseI-CTA y EcoRI-ACT/MseI-CAT generaron los niveles más altos de bandas polimórficas, 29 (56%) y 27 (52%), respectivamente. Los valores del índice de diversidad génica variaron de 0,0604 a 0,3650 con un promedio de 0,1924. Se observó una asociación positiva entre el número de bandas polimórficas y el índice de diversidad génica calculado para cada combinación de cebadores.
Locus Grupo de
Ligamiento
Rango de tamaños
de alelos (pb)
N° de alelos IDa
Satt 141 D1b 148 - 204 6 0.753
Satt 373 L 212 - 283 9 0.730
Satt 458 D2 158 - 194 9 0.727
Satt 146 F 278 - 317 7 0.715
Satt 005 D1b 142 - 178 6 0.705
Satt 181 H 176 - 217 5 0.702
Satt 030 F 152 - 167 5 0.695
Satt 307 C2 162 - 192 5 0.687
Satt 414 J 168 - 315 6 0.687
Satt 453 B1 221 - 269 8 0.684
Satt 556 B2 164 - 211 4 0.658
Satt 114 F 82 - 122 4 0.654
Satt 242 K 187 - 199 4 0.640
Satt 703 D1b 189 - 239 5 0.638
Satt 009 N 161 - 227 5 0.624
Satt 534 B2 232 - 261 7 0.606
Satt 577 B2 113 - 122 4 0.591
Satt 292 I 223 - 254 6 0.591
Satt 596 J 236 - 259 6 0.589
Satt 237 N 237 - 281 8 0.579
Satt 324 G 229 - 241 3 0.577
Satt 417 K 280 - 321 3 0.576
Satt 354 I 233 - 250 4 0.573
Satt 197 B1 175 - 189 5 0.569
Satt 038 G 157 - 177 5 0.567
Satt 308 M 149 - 175 4 0.566
Satt 226 D2 321 - 338 3 0.548
Satt 231 E 215 - 242 5 0.546
Satt 191 G 185 - 229 6 0.545
Satt 436 D1a 182 - 246 7 0.544
Satt 095 D1b 160 - 175 4 0.510
Satt 511 A1 244 - 256 3 0.499
Satt 070 B2 147 - 174 4 0.495
Satt 156 L 205 - 220 3 0.483
Satt 173 O 199 - 263 4 0.478
Satt 180 C1 244 - 268 3 0.469
Satt 294 C1 248 - 293 5 0.465
Satt 409 A2 165 - 195 6 0.441
Satt 094 O 148 - 163 2 0.428
Satt 243 O 202 - 211 2 0.426
Satt 449 A1 222 - 266 5 0.412
Satt 177 A2 113 - 122 2 0.400
Satt 311 D2 180 - 191 2 0.397
Satt 147 D1a 171 - 203 4 0.387
Satt 042 A1 159 - 174 4 0.387
Satt 175 M 160 - 193 6 0.289
Satt 281 C2 167 - 236 4 0.263
Satt 253 H 138 - 162 6 0.259
Satt 233 A2 188 - 209 3 0.144
Satt 150 M 189 - 200 2 0.111
Satt 475 K 231 - 252 3 0.081
Tabla 4: Características de los 51 marcadores SSR utilizados sobre los 86 cultivares argentinos de soja.
a Indice del diversidad génica: probabilidad de que al tomar dos genotipos al azar a partir de una población de individuos ensayados estos sean polimórficos en ese marcador.
Figura 1. Alelos detectados para el locus SSR Satt009 en 21 de los 86 cultivares de soja analizados en el presente estudio. A la izquierda se sembró una calle con un marcador de peso molecular de ADN de 10 pares de base de separación. Al tope de la figura se indican los nombres de los cultivares analizados. Las bandas se observan como bandas dobles porque la tinción con plata permite detectar la hebra complementaria, que migra a una distancia ligeramente diferente por poseer una secuencia y composición de bases diferente. Las flechas de la derecha indican los tamaños de los alelos en pares de bases (pb). La posición de las muestras 1 y 21 se indica sobre el gel a la altura del alelo de 161 pb.
Desarrollo de una base de datos genéticos para la caracterización del germoplasma argentino de soja
Ciencias Agronómicas - Revista XX - Año 12 - 2012 / 023 - 039
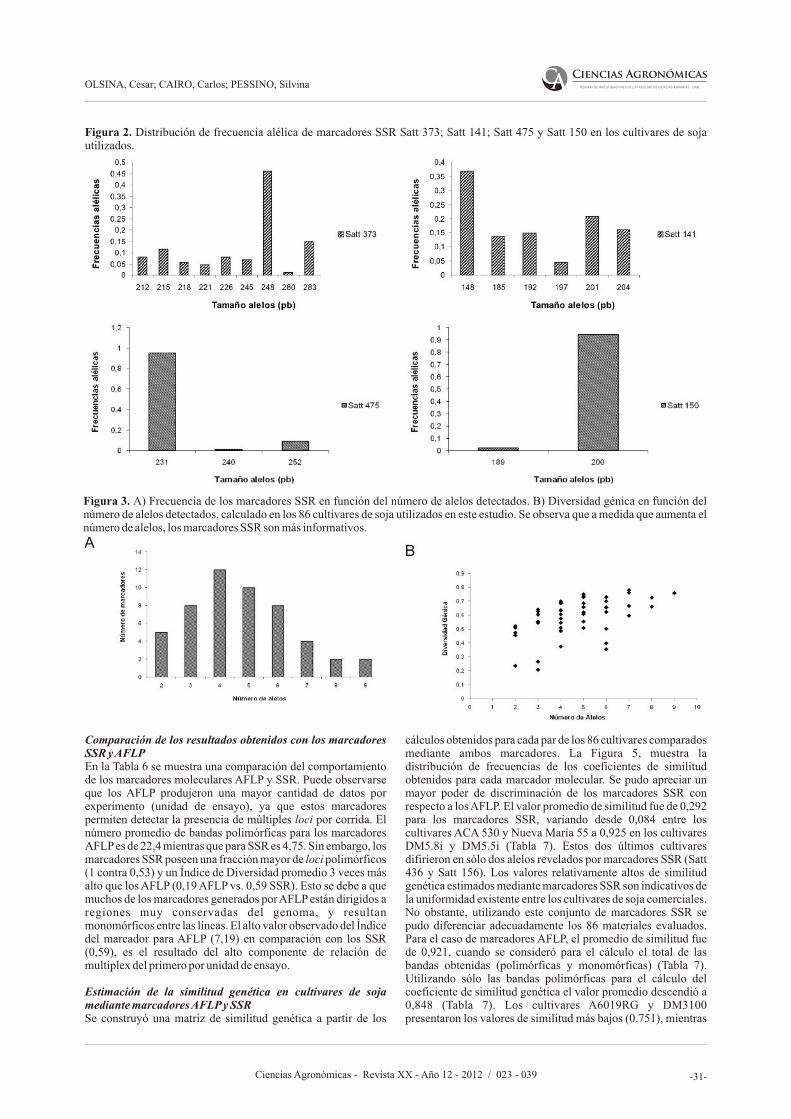
-31-
Comparación de los resultados obtenidos con los marcadores SSR y AFLPEn la Tabla 6 se muestra una comparación del comportamiento de los marcadores moleculares AFLP y SSR. Puede observarse que los AFLP produjeron una mayor cantidad de datos por experimento (unidad de ensayo), ya que estos marcadores permiten detectar la presencia de múltiples loci por corrida. El número promedio de bandas polimórficas para los marcadores AFLP es de 22,4 mientras que para SSR es 4,75. Sin embargo, los marcadores SSR poseen una fracción mayor de loci polimórficos (1 contra 0,53) y un Índice de Diversidad promedio 3 veces más alto que los AFLP (0,19 AFLP vs. 0,59 SSR). Esto se debe a que muchos de los marcadores generados por AFLP están dirigidos a regiones muy conservadas del genoma, y resultan monomórficos entre las líneas. El alto valor observado del Índice del marcador para AFLP (7,19) en comparación con los SSR (0,59), es el resultado del alto componente de relación de multiplex del primero por unidad de ensayo.
Estimación de la similitud genética en cultivares de soja mediante marcadores AFLP y SSRSe construyó una matriz de similitud genética a partir de los
cálculos obtenidos para cada par de los 86 cultivares comparados mediante ambos marcadores. La Figura 5, muestra la distribución de frecuencias de los coeficientes de similitud obtenidos para cada marcador molecular. Se pudo apreciar un mayor poder de discriminación de los marcadores SSR con respecto a los AFLP. El valor promedio de similitud fue de 0,292 para los marcadores SSR, variando desde 0,084 entre los cultivares ACA 530 y Nueva María 55 a 0,925 en los cultivares DM5.8i y DM5.5i (Tabla 7). Estos dos últimos cultivares difirieron en sólo dos alelos revelados por marcadores SSR (Satt 436 y Satt 156). Los valores relativamente altos de similitud genética estimados mediante marcadores SSR son indicativos de la uniformidad existente entre los cultivares de soja comerciales. No obstante, utilizando este conjunto de marcadores SSR se pudo diferenciar adecuadamente los 86 materiales evaluados. Para el caso de marcadores AFLP, el promedio de similitud fue de 0,921, cuando se consideró para el cálculo el total de las bandas obtenidas (polimórficas y monomórficas) (Tabla 7). Utilizando sólo las bandas polimórficas para el cálculo del coeficiente de similitud genética el valor promedio descendió a 0,848 (Tabla 7). Los cultivares A6019RG y DM3100 presentaron los valores de similitud más bajos (0.751), mientras
Figura 2. Distribución de frecuencia alélica de marcadores SSR Satt 373; Satt 141; Satt 475 y Satt 150 en los cultivares de soja utilizados.
Figura 3. A) Frecuencia de los marcadores SSR en función del número de alelos detectados. B) Diversidad génica en función del número de alelos detectados, calculado en los 86 cultivares de soja utilizados en este estudio. Se observa que a medida que aumenta el número de alelos, los marcadores SSR son más informativos.
OLSINA, Cesar; CAIRO, Carlos; PESSINO, Silvina
Ciencias Agronómicas - Revista XX - Año 12 - 2012 / 023 - 039
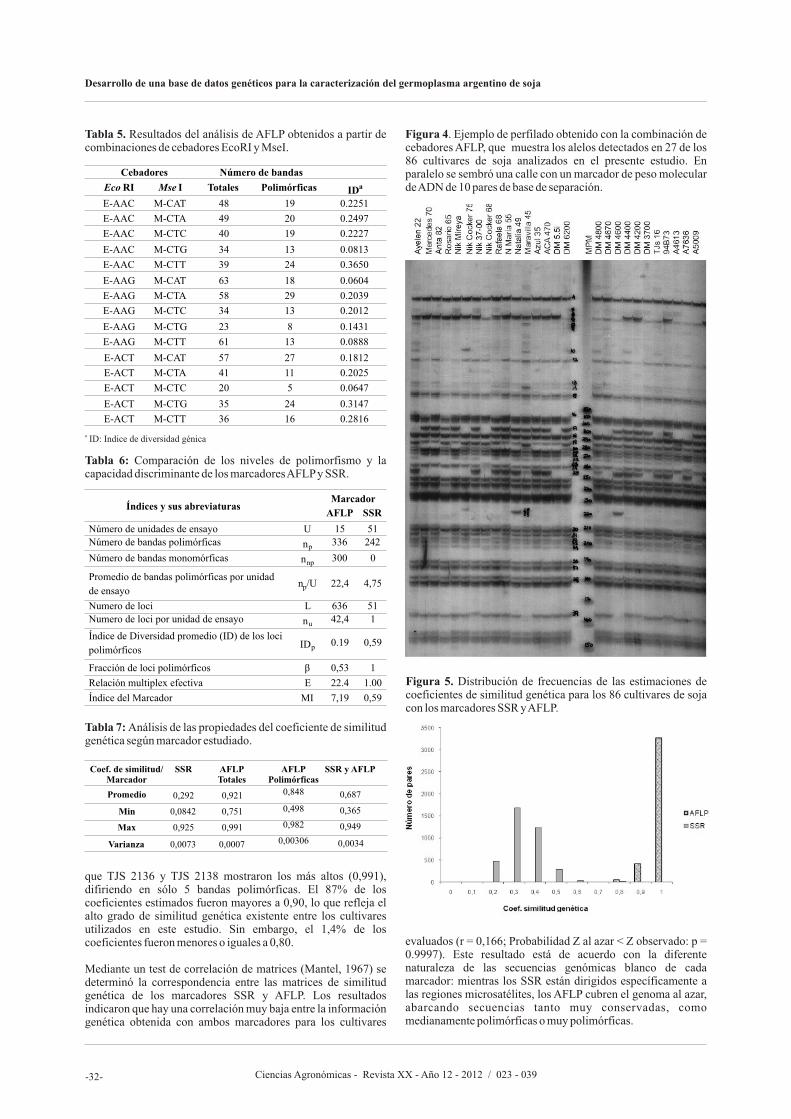
-32-
Tabla 5. Resultados del análisis de AFLP obtenidos a partir de combinaciones de cebadores EcoRI y MseI.
Eco RI Mse I Totales Polimórficas IDa
E-AAC M-CAT 48 19 0.2251
E-AAC M-CTA 49 20 0.2497
E-AAC M-CTC 40 19 0.2227
E-AAC M-CTG 34 13 0.0813
E-AAC M-CTT 39 24 0.3650
E-AAG M-CAT 63 18 0.0604
E-AAG M-CTA 58 29 0.2039
E-AAG M-CTC 34 13 0.2012
E-AAG M-CTG 23 8 0.1431
E-AAG M-CTT 61 13 0.0888
E-ACT M-CAT 57 27 0.1812
E-ACT M-CTA 41 11 0.2025
E-ACT M-CTC 20 5 0.0647
E-ACT M-CTG 35 24 0.3147
E-ACT M-CTT 36 16 0.2816
Cebadores Número de bandas
a ID: Indice de diversidad génica
AFLP SSR
Número de unidades de ensayo U 15 51
Número de bandas polimórficas np 336 242
Número de bandas monomórficas nnp 300 0
Promedio de bandas polimórficas por unidad
de ensayo np/U 22,4 4,75
Numero de loci L 636 51
Numero de loci por unidad de ensayo nu 42,4 1
Índice de Diversidad promedio (ID) de los loci
polimórficosIDp 0.19 0,59
Fracción de loci polimórficos β 0,53 1
Relación multiplex efectiva E 22.4 1.00
Índice del Marcador MI 7,19 0,59
Índices y sus abreviaturasMarcador
Tabla 6: Comparación de los niveles de polimorfismo y la capacidad discriminante de los marcadores AFLP y SSR.
Tabla 7: Análisis de las propiedades del coeficiente de similitud genética según marcador estudiado.
Coef. de similitud/Marcador
SSR AFLPTotales
AFLPPolimórficas
SSR y AFLP
Promedio 0,292 0,921 0,848 0,687
Min 0,0842 0,751 0,498 0,365
Max 0,925 0,991 0,982 0,949
Varianza 0,0073 0,0007 0,00306 0,0034
que TJS 2136 y TJS 2138 mostraron los más altos (0,991), difiriendo en sólo 5 bandas polimórficas. El 87% de los coeficientes estimados fueron mayores a 0,90, lo que refleja el alto grado de similitud genética existente entre los cultivares utilizados en este estudio. Sin embargo, el 1,4% de los coeficientes fueron menores o iguales a 0,80.
Mediante un test de correlación de matrices (Mantel, 1967) se determinó la correspondencia entre las matrices de similitud genética de los marcadores SSR y AFLP. Los resultados indicaron que hay una correlación muy baja entre la información genética obtenida con ambos marcadores para los cultivares
evaluados (r = 0,166; Probabilidad Z al azar < Z observado: p = 0.9997). Este resultado está de acuerdo con la diferente naturaleza de las secuencias genómicas blanco de cada marcador: mientras los SSR están dirigidos específicamente a las regiones microsatélites, los AFLP cubren el genoma al azar, abarcando secuencias tanto muy conservadas, como medianamente polimórficas o muy polimórficas.
Figura 4. Ejemplo de perfilado obtenido con la combinación de cebadores AFLP, que muestra los alelos detectados en 27 de los 86 cultivares de soja analizados en el presente estudio. En paralelo se sembró una calle con un marcador de peso molecular de ADN de 10 pares de base de separación.
Figura 5. Distribución de frecuencias de las estimaciones de coeficientes de similitud genética para los 86 cultivares de soja con los marcadores SSR y AFLP.
Desarrollo de una base de datos genéticos para la caracterización del germoplasma argentino de soja
Ciencias Agronómicas - Revista XX - Año 12 - 2012 / 023 - 039

-33-
Análisis de agrupamientoCon las matrices de similitud genética obtenidas, se realizó un análisis de agrupamiento de pares no ponderado utilizando la media aritmética (UPGMA). Este procedimiento reveló que la similitud genética promedio entre genotipos era dependiente del tipo de marcador molecular empleado. En las Figuras 6 y 7, se muestran los resultados obtenidos para cada tipo de marcador molecular (SSR o AFLP).
Los dendrogramas permitieron la diferenciación de los cultivares en estudio, sin embargo las asociaciones observadas en cada uno son características del sistema de marcadores
empleados. En el de SSRs (Figura 6) se pueden distinguir tres grupos principales a una similitud genética de 0,26. El primero de ellos (Grupo 1) está constituido por los cultivares ACA 530 y ACA 570, que se muestran muy diferentes al resto de los materiales en estudio y similares entre sí (coeficiente ~ 0,70). El coeficiente de similitud genético de estos cultivares respecto al resto es de 0,191. El resto de los genotipos se distribuyen en los dos grupos restantes (Grupos 2 y 3). No se observa una asociación entre los agrupamientos formados y los grupos de madurez de los cultivares evaluados, ni tampoco con la identidad del obtentor de las variedades. Los cultivares DM5.5i y DM5.8i, pertenecientes al grupo 3, son los más semejantes entre sí,
Figura 6. Dendrograma obtenido utilizando marcadores SSR en los cultivares de soja seleccionados en este estudio. La línea trazada a un similitud genética de 0,24 permite diferenciar tres grupos principales.
OLSINA, Cesar; CAIRO, Carlos; PESSINO, Silvina
Ciencias Agronómicas - Revista XX - Año 12 - 2012 / 023 - 039
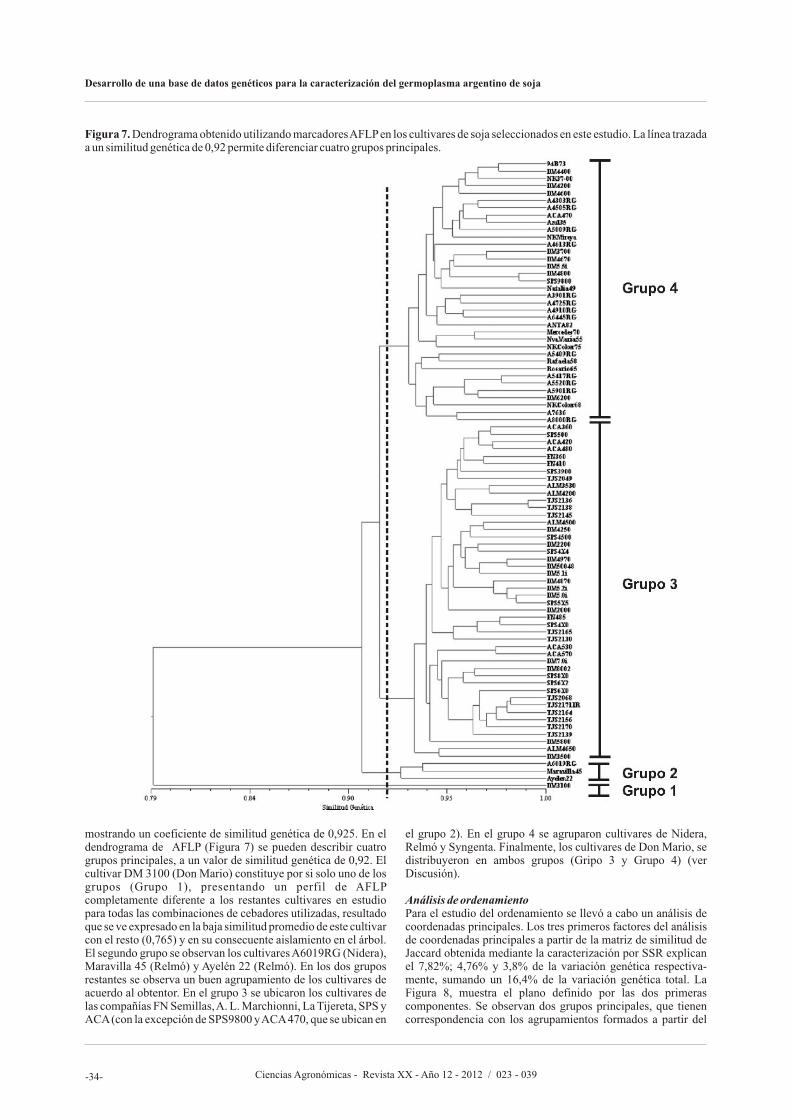
-34-
mostrando un coeficiente de similitud genética de 0,925. En el dendrograma de AFLP (Figura 7) se pueden describir cuatro grupos principales, a un valor de similitud genética de 0,92. El cultivar DM 3100 (Don Mario) constituye por si solo uno de los grupos (Grupo 1), presentando un perfil de AFLP completamente diferente a los restantes cultivares en estudio para todas las combinaciones de cebadores utilizadas, resultado que se ve expresado en la baja similitud promedio de este cultivar con el resto (0,765) y en su consecuente aislamiento en el árbol. El segundo grupo se observan los cultivares A6019RG (Nidera), Maravilla 45 (Relmó) y Ayelén 22 (Relmó). En los dos grupos restantes se observa un buen agrupamiento de los cultivares de acuerdo al obtentor. En el grupo 3 se ubicaron los cultivares de las compañías FN Semillas, A. L. Marchionni, La Tijereta, SPS y ACA (con la excepción de SPS9800 y ACA 470, que se ubican en
el grupo 2). En el grupo 4 se agruparon cultivares de Nidera, Relmó y Syngenta. Finalmente, los cultivares de Don Mario, se distribuyeron en ambos grupos (Gripo 3 y Grupo 4) (ver Discusión).
Análisis de ordenamientoPara el estudio del ordenamiento se llevó a cabo un análisis de coordenadas principales. Los tres primeros factores del análisis de coordenadas principales a partir de la matriz de similitud de Jaccard obtenida mediante la caracterización por SSR explican el 7,82%; 4,76% y 3,8% de la variación genética respectiva-mente, sumando un 16,4% de la variación genética total. La Figura 8, muestra el plano definido por las dos primeras componentes. Se observan dos grupos principales, que tienen correspondencia con los agrupamientos formados a partir del
Figura 7. Dendrograma obtenido utilizando marcadores AFLP en los cultivares de soja seleccionados en este estudio. La línea trazada a un similitud genética de 0,92 permite diferenciar cuatro grupos principales.
Desarrollo de una base de datos genéticos para la caracterización del germoplasma argentino de soja
Ciencias Agronómicas - Revista XX - Año 12 - 2012 / 023 - 039

-35-
análisis de UPGMA, excepto por el cultivar DM 7.0i. El análisis mediante un árbol de mínima distancia muestra que hay una buena concordancia entre los agrupamientos propuestos en el espacio bidimensional y las distancias entre los cultivares en el espacio de N-Dimensiones original.
El análisis de coordenadas principales obtenido a partir de la caracterización por AFLP mostró una variación de 17,85%; 6,80% y 5,54% en las tres primeras dimensiones respectivamente, sumando un 30,21% de la variación genética total. En la Figura 9 se observa la proyección de los cultivares analizados en el plano definido por las dos primeras componentes. Los cultivares tienden a agruparse en dos grupos principales siguiendo un gradiente a través del primer eje. Estos agrupamientos tienen una muy buena concordancia con los agrupamientos obtenidos a partir del análisis de UPGMA. El árbol de mínima distancia superpuesto, muestra una buena asociación entre las proyecciones en las dos dimensiones del plano y las distancias obtenidas de la matriz de similitud original. Al igual que con los dendrogramas, se observa un buen agrupamiento de los cultivares de acuerdo al obtentor a través del gradiente formado en la componente 1. En el grupo 1 se encuentran ordenados los cultivares de las compañías FN semillas; A. L. Marchionni, La Tijereta, SPS y ACA (menos SPS9800 y ACA 470 respectivamente, que se ubican en el grupo 2). En el grupo 2 por otra parte se encuentran ordenados los cultivares de Nidera, Relmó y Syngenta. Finalmente, los cultivares de Don Mario, se ordenaron entre ambos grupos con los cultivares DM4670, DM4400, DM3700, DM5.5i, DM4800 y DM6200 en el grupo 2 y los 16 cultivares restantes en el grupo 1.
Uso de la base de datos de SSR para la identificación de muestras incógnitas. Prueba de conceptoCon el objeto de poner a prueba la base de datos de SSR de cultivares de soja se analizaron un total de 12 muestras enviadas al laboratorio por diferentes semilleros o multiplicadores, los cuales a su vez designaron una identidad putativa del cultivar que debería ser contrastado con el banco. Los resultados obtenidos se muestran en la Tabla 8. En 10 de los 12 casos los marcadores SSR seleccionados permitieron concluir eficazmente sobre la identidad las muestras ensayadas y diferenciarlas de los restantes cultivares del banco. Esto demuestra el poder discriminatorio de los marcadores SSR como herramienta para la diferenciación de cultivares de soja argentina. En los restantes dos casos no se detectó similitud genética con el cultivar propuesto como identidad putativa, pero además no se encontró similitud con ningún otro cultivar del banco.
Diseño de una metodología eficiente para ensayos de diferenciación de cultivaresLa caracterización de muestras incógnitas comentada en la sección anterior se basó en una selección específica de marcadores de acuerdo a la identidad putativa (propuesta) de la muestra, que había sido proporcionada por el solicitante de la identificación. Sin embargo, pensamos que resultaría conveniente contar con un subconjunto selecto y fijo de marcadores que permitiera diferenciar cualquier variedad caracterizada en el banco. Así, nos propusimos seleccionar el mínimo subconjunto de marcadores cuya aplicación, independientemente de la identidad de la muestra, nos permitiera diferenciarla inequívocamente.
Figura 8. Ordenamiento en dos dimensiones de las 86 cultivares de soja luego de un Análisis de Coordenadas Principales y la construcción de un árbol de mínima distancia (MST) para marcadores SSR. Se muestran las dimensiones 1 y 2. Los símbolos indican los criaderos obtentores: (1) Pionner; (2) Nidera; (3) ACA; (4) A. L. Marchionni; (5) Relmó; (6) Don Mario; (7) FN Semillas; (8) Syngenta; (9) SPS y (10) La Tijereta.
OLSINA, Cesar; CAIRO, Carlos; PESSINO, Silvina
Ciencias Agronómicas - Revista XX - Año 12 - 2012 / 023 - 039

-36-
A partir de la caracterización realizada sobre 86 cultivares de soja se seleccionaron 20 loci SSR localizados en los diferentes grupos de ligamiento y que además presentaban los valores de índice de diversidad génica más altos. La utilización de este número reducido de marcadores comunes permitiría el análisis simultáneo de hasta tres muestras cualesquiera en un único gel. Para determinar la efectividad del conjunto de 20 marcadores SSR seleccionados versus el conjunto completo de 51 loci, se compararon los dos conjuntos en términos de su habilidad para distinguir los 86 cultivares de soja analizadas en este estudio. La similitud media calculada sobre los 20 loci fue numéricamente más baja, que la obtenida utilizando los 51 loci SSR (0,214 con 20 loci versus 0,293 para el caso de los 51 loci). La diferencia entre las medias fue estadísticamente significativa (P<0.001). La varianza obtenida fue de 0,0093 mediante el análisis de los 20
loci, y de 0,0073 basándose en los 51 loci. Estas estimaciones de la varianza no difieren significativamente en su magnitud.
Estos resultados indican que el procedimiento utilizado para seleccionar los loci fue efectivo para identificar un subconjunto más informativo. Este subconjunto se comportó de manera similar al conjunto completo de 51 loci en términos de habilidad para distinguir cultivares de soja comerciales argentinas. El valor máximo de similitud genética obtenido para el subconjunto de 20 SSR corresponde a los cultivares DM 4800 y DM 50048, con un valor obtenido de 0,909. En el caso de los valores mínimos, para el subconjunto de 20 loci corresponden a los cultivares ACA470 y A5417 RG con un valor de similitud genética de 0,00, indicando que en los 20 loci SSR seleccionados estos cultivares no tienen ningún alelo en común.
Discusión
La caracterización mediante marcadores moleculares de los cultivares de soja que se utilizan actualmente en la producción argentina es necesaria, debido a la importancia que tiene la diferenciación e identificación de los mismos para el mejoramiento, así como también para el control del comercio y de la calidad de lotes de semilla. Esta caracterización permite contar con una base de datos de huellas genéticas de las variedades locales, determinar la variabilidad genética existente y seleccionar parentales divergentes para la obtención de nuevas variedades mediante cruzamiento.
En el presente trabajo se realizó una caracterización de 86 cultivares de soja argentina resistentes a glifosato, utilizadas en la actualidad y provenientes de las compañías semilleras más importantes del país, por medio de dos tipos de marcadores moleculares, SSR y AFLP. Se construyó una base de datos que fue luego validada mediante el ensayo de muestras incógnitas enviadas por semilleros o multiplicadores de semilla de soja, invirtiendo un tiempo de ensayo corto y utilizando un método robusto y reproducible. Se diseñó además un procedimiento simplificado para una clasificación eficaz y estandarizada de cualquier muestra incógnita bajo estudio.
Figura 9. Ordenamiento en dos dimensiones de las 86 cultivares de soja luego de un Análisis de Coordenadas Principales y la construcción de un árbol de mínima distancia (MST) para marcadores AFLP. Se muestran las dimensiones 1 y 2. Los símbolos indican los criaderos obtentores: (1) Pionner; (2) Nidera; (3) ACA; (4) A. L. Marchionni; (5) Relmó; (6) Don Mario; (7) FN Semillas; (8) Syngenta; (9) SPS y (10) La Tijereta.
Desarrollo de una base de datos genéticos para la caracterización del germoplasma argentino de soja
Ciencias Agronómicas - Revista XX - Año 12 - 2012 / 023 - 039

-37-
En la caracterización con marcadores AFLP se utilizaron 15 combinaciones de cebadores que produjeron un total de 636 bandas de las cuales un 52% fueron polimórficas. Este valor alto de bandas polimórficas se debió principalmente a la presencia del cultivar DM3100, que presenta la mayoría de dichas bandas. El número promedio de bandas polimórficas por marcador fue de 17, resultado que está en concordancia con estudios realizados en cultivares de soja brasileña (Bonato et al., 2006) y cultivares chinos y norteamericanos (Ude et al., 2003) con 13 y 18 respectivamente. Las combinaciones de cebadores más informativos fueron EcoRIAAC/MseI-CTT y EcoRI-ACT/MseI-CTG, con 24 bandas polimórficas y los valores de diversidad génica más altos. Estos resultados contrastan con los obtenidos por otros grupos en donde las combinaciones más polimórficas fueron otras. Por ejemplo, Bonato (2006) y Zhu et al. (1999) encontraron que la combinación EcoRI-AAC/MseI-CTC fue la más polimórfica, mientras que Ude et al. (2003) describieron que los marcadores EcoRI-ACC/MseI-CAA producían la mayor cantidad de bandas polimórficas. Esta diferencia entre los resultados de varios autores podría estar indicando variaciones genéticas subyacentes en los germoplasmas caracterizados.
Los resultados obtenidos en Giancola et al. (2002) en cultivares de soja argentino mediante la utilización de marcadores AFLP no son comparables en forma directa con este trabajo, dado que estos autores utilizaron cebadores selectivos +4 (con cuatro bases de secuencia al azar en el extremo 3' del cebador) mientras que aquí utilizamos +3 (con sólo 3 bases de secuencia al azar), generando diferentes números y tipos de bandas. Los resultados obtenidos en este trabajo confirman resultados previos en los que se demuestra que los marcadores AFLP son una técnica molecular que detecta polimorfismos en múltiples loci simultáneamente, generando un gran número de marcadores reproducibles en un periodo corto de tiempo, siendo además una herramienta poderosa para el análisis de genotipos altamente relacionados (Powell et al., 1996).
El presente estudio demuestra que la diversidad génica esperada para los loci polimórficos utilizando marcadores SSR es mayor que para AFLP. Esto se relaciona con la variación del número medio de bandas polimórficas por unidad de ensayo, que varía de un 21% a 61% para AFLP a un máximo de 100% para SSR. El nivel de polimorfismo más alto detectado de los marcadores de SSR con respecto a AFLP destaca la capacidad discriminatoria de los primeros. Sin embargo, la comparación de la eficacia total de los dos sistemas de marcadores proporcionada por el índice del marcador (MI), indica que los marcadores AFLP poseen un MI 12 veces más alto con respecto a loci SSR. Esto confirma la naturaleza altamente discriminatoria del análisis de AFLP, que está en concordancia con informes anteriores en varias especies de plantas (Powell et al., 1996; Belaj et al., 2003; Saini et al., 2004; Medini et al., 2005). El valor distintivo del MI para los marcadores AFLP radica en el cociente multiplex eficaz. Es decir, en el alto número de bandas polimórficas obtenidas por experimento.
Los resultados comentados nos permiten inferir que los marcadores SSR permiten detectar un alto grado de polimorfismos, siendo muy útiles para estudios de diferenciación de cultivares (Diwan & Cregan, 1997). Pero, por otro lado, los marcadores AFLP permiten realizar estudios de comparación de cultivares muy rápidos, dado el gran número de bandas obtenidas por unidad de ensayo, de las cuales un número importante son polimórficas (Powell et al., 1996). El direccionamiento de los marcadores AFLP hacia cualquier región del genoma al azar garantiza una cobertura genómica completa, lo que potencialmente permitiría realizar análisis de
El nivel de polimorfismos obtenidos para los 51 loci SSR analizados arrojó una media de 4,7 alelos por locus variando entre 2 y 9 alelos, y fue más bajo que el informado en otros trabajos como los de Rongwen et al. (1995) y Diwan & Cregan (1997). En el estudio de la diversidad genética en soja de Rongwen et al. (1995), realizado sobre 96 genotipos, se amplificaron entre 11 y 26 alelos por cada locus de microsatélites, aunque este número se vio reducido entre 5 y 10 alelos por locus en los 26 cultivares provenientes de programas de mejoramiento de soja norteamericano. Una posible razón para esta diferencia es que los materiales utilizados en el presente estudio provienen justamente de cultivares comerciales inscriptos, los cuales poseen una base genética estrecha. Sin embargo, los resultados de nuestro trabajo están en consonancia con los obtenidos por Priolli et al. (2002), quienes estudiaron 186 cultivares de soja brasileños por medio de 12 loci SSR y amplificaron entre 4 y 8 alelos por locus, con un promedio de 5,3 alelos. También concuerda con el trabajo de Giancola et al. (2002), quienes analizaron 30 loci SSR en 100 cultivares argentinos, y observaron entre 2 y 8 alelos por loci con un promedio de 4,4 alelos por locus.
El índice de diversidad génica (ID), que describe el potencial del locus para la discriminación de dos variedades tomadas al azar, tiene en cuenta para su cálculo tanto el número como la frecuencia de aparición de alelos en los cultivares analizados. En este trabajo el valor promedio de ID obtenido para marcadores de SSR fue de 0,523, mostrando el alto grado de polimorfismo de estos marcadores. Valores de diversidad génica similares fueron obtenidos por Maughan et al. (1995) en 62 cultivares se soja con un valor medio de ID de 0,55 o Narvel et al. (2000) con un valor medio de ID de 0,50 en 39 cultivares elite norteamericanos. Sin embargo, en los trabajos de Priolli et al. (2002) y Giancola et al. (2002), los valores medios de diversidad génica fueron levemente mayores que el obtenido en el presente trabajo (0,64 y 0,60, respectivamente). Esto puede deberse tanto a una disminución de la variabilidad genética, como así también a que en nuestro estudio se incluyó un número mayor de loci SSR que poseen alelos de muy baja frecuencia, conduciendo a valores de ID muy bajos (por ejemplo Satt 475 con un ID de 0,082). Estos alelos al ser poco frecuentes son útiles para la identificación de algún cultivar en particular, pero son poco polimórficos y por lo tanto hacen que el valor promedio obtenido en el cálculo de la diversidad genética sea menor.
La presencia de estos alelos de baja frecuencia en los marcadores SSR, como los encontrados en los loci Satt 534; 70; 231; 175; 294; 373; 414; 038; 197; 253; 191; 475; 458; 596; 292; 436; 409; 449; 703; 146; 556 y 095, podría reflejar la tasa de mutación de
-5 -4los microsatélites de soja, estimada en 10 a 10 por generación (Diwan & Cregan, 1997). Esta tasa es similar a la encontrada en humanos, y no implicaría una restricción para el uso de los microsatélites en la identificación de cultivares (Diwan & Cregan, 1997). Se ha determinado que para la identificación de cultivares de soja se debe utilizar un conjunto de entre 30 y 50 individuos, ya que los alelos mutantes que pudiesen surgir no serían detectados y de esta manera, las mutaciones en una planta aislada no alterarían la constitución alélica del cultivar (Diwan & Cregan, 1997). Por ejemplo, Song et al. (1999) encontraron 10 alelos nuevos en 66 cultivares de soja, los cuales no estuvieron presentes en 35 líneas ancestrales, mientras que Narvel et al. (2000) registraron 32 alelos específicos de cultivares elite, dentro de un total de 397 alelos que han sido detectados en 40 líneas y 39 cultivares de soja. Dado que en este trabajo no se analizaron las líneas progenitoras o ancestrales que dieron lugar a los cultivares actuales, no se puede concluir que pudiesen haber surgido alelos de baja frecuencia por este mecanismo.
OLSINA, Cesar; CAIRO, Carlos; PESSINO, Silvina
Ciencias Agronómicas - Revista XX - Año 12 - 2012 / 023 - 039

-38-
soja evaluados en este trabajo es muy baja (r = 0,166), aunque significativa. Valores comparables fueron encontrados en soja por Giancola et al. (2002), que observaron una correlación entre las estimaciones de similitud genética de marcadores AFLP y SSR de 0,210. Asimismo, Powell et al. (1996) encontraron valores de correlación bajos entre estos marcadores. Las estimaciones de la similitud genética se ven influenciadas por una variedad de factores entre los que se pueden contar la distribución de los marcadores en el genoma, la naturaleza de los mecanismos que generan la variación medida y errores en la clasificación de bandas no homólogas de loci AFLP de movilidad similar. Por una parte, los marcadores AFLP se originan al azar, a partir de regiones que pueden resultar hipervariables, medianamente variables o muy conservadas. Sin embargo, dado que la mayor parte de las regiones del genoma de soja están conservadas entre las variedades, un mayor número de marcadores presenta perfiles monomórficos, por lo que las tasas de variación son más bajas que las observadas en los loci SSR, los cuales están dirigidos específicamente a regiones altamente polimórficas (Powell et al., 1996). Es de interés mencionar que aunque en general se considera que la distribución de los marcadores de AFLP en el genoma es azarosa, se han notificado casos de sobrerepresentación de ciertas secuencias genómicas, como por ejemplo regiones centroméricas (Menz et al. 2004).
parentesco más precisos. En los dendrogramas y análisis de ordenamientos generados a partir de la matriz de AFLP se observó que los genotipos agruparon según el obtentor. Sin embargo, dado que carecíamos de datos de pedigrí, no pudimos analizar la asociación entre los perfiles de marcadores y el parentesco entre variedades.
En este trabajo los marcadores SSR arrojaron estimaciones de similitud genética más bajas con respecto a los marcadores de AFLP, obteniéndose valores medios de 0,292 y 0,921 para estas técnicas, respectivamente. Estos resultados eran esperables, ya que los SSR están dirigidos a regiones altamente variables, mientras los AFLPs detectan loci que presentan un amplio rango en su nivel de polimorfismo, desde los que son completamente conservados hasta los hipervariables, y la posibilidad de detectar este tipo de loci está relacionada con la frecuencia de su ocurrencia en el genoma. Nuestros resultados están en completo acuerdo con otros estudios que comparan el nivel del polimorfismo detectado con los marcadores de AFLP y SSR en la soja (Powell et al., 1996), el maíz (Pejic et al., 1998), el sorgo (Menz et al., 2004), y el triticale (Tams et al., 2005).
La correlación existente entre las estimaciones de la similitud genética entre marcadores AFLP y SSR para los cultivares de
Conclusiones
selección de genotipos divergentes. Estas bases de datos están siendo utilizadas en la Bolsa de Comercio de Rosario para prestar un servicio de clasificación y caracterización de genotipos. Su utilización puede contribuir al mejoramiento del germoplasma argentino de soja y a la protección de la propiedad intelectual.
En este trabajo se informa el desarrollo de una base de datos genéticos de SSR y AFLP que permitió la caracterización de las 86 variedades de soja más cultivadas actualmente en Argentina. Mientras los marcadores de SSR resultaron útiles para la rápida identificación de variedades, los de AFLP fueron más apropiados para la caracterización global del genoma y la
Agradecimientos
Agradecemos a los semilleros ACA, AL Marchionni, Don Mario, FN Semillas, La Tijereta, Nidera, Pioneer, Relmó, SPS y Syngenta, por proveernos gentilmente el material vegetal utilizado en este análisis. Este estudio fue financiado por la Bolsa de Comercio de Rosario. Silvina C. Pessino es miembro de la Carrera de Imvestigador Científico del Consejo Nacional de Investigaciones Científicas y Tecnológicas (CONICET).
Bibliografía
1. AKKAYA MS, BHAGWAT AA, CREGAN PB (1992) The use of microsatellite DNA markers for soybean genotype identification. Genetics, 132: 1131—1139.
2. AKKAYA MS, SHOEMAKER RC, SPECHT JE, BHAGWAT AA, CREGAN PB (1995) Integration of simple sequence repeat DNA markers into a soybean linkage map. Crop Sci, 35: 1439-1445.
3. BELAJ A, SATOVIC Z, CIPRIANI G, BALDONI L, TESTOLIN R, RALLO L, TRUJILLO I (2003) Comparative study of the discriminating capacity of RAPD, AFLP and SSR markers and of their effectiveness in e s t a b l i s h i n g g e n e t i c r e l a t i o n s h i p s i n o l i v e . Theor.Appl.Genet, 107: 736-744.
4. BONAMICO N, AIASSA J, IBAÑEZ M, DI RENZO M, DÍAZ D, SALERNO J (2004) Caracterización y clasificación de híbridos simples de maíz con marcadores SSR. Revista de Investigaciones Agropecuarias, INTA, Argentina, 33: 129-144.
5. BONATO A, CALVO E, GERALDI I, ARIAS C (2006) Genetic similarity among soybean (Glycine max L. Merrill) cultivars released in Brazil using AFLP markers. Genet Mol Biol, 29: 692-704.
6. BOTSTEIN D, WHITE R, SKOLNICK M, DAVIS RW (1980) Construction of a genetic linkage map in man using restriction fragment length polymorphisms. Am. J. Hum. Genet, 32: 314-331.
7. BROWN-GUEDIRA G, THOMPSON J, NELSON R, WARBURTON M (2000) Evaluation of genetic diversity of soybean introductions and North American ancestors using RAPD and SSR markers. Crop Sci, 40: 815-823.
8. CREGAN PB, BHAGWAT AA, AKKAYA MS, RONGWEN J (1994) Microsatellite fingerprinting and mapping of soybean. Methods Mol Cell Biol, 5: 49-61.
9. CREGAN PB, JARVIK T, BUSH AL, SHOEMAKER RC, LARK KG, KAHLER AL, KAYA N, VAN TOAI TT, LOHNES DG, CHUNG J, SPECHT JE (1999). An integrated genetic linkage map of the soybean genome. Crop Sci, 39: 1464–1490.
10. DIWAN N, CREGAN PB (1997) Automated sizing of fluorescent-labeled simple sequence repeat (SSR) markers to assay genetic variation in soybean. Theor Appl Genet, 95: 723-733.
11. DONG YS, ZHAO LM, LIU B, WANG ZW, JIN ZQ, SUN H (2004) The genetic diversity of cultivated soybean grown in China. Theor Appl Genet, 108: 931-936.
12. GIANCOLA S, MARCUCCI POLTRI S, LACAZE P, HOPP HE (2002) Feasibility of integration of molecular markers and morphological descriptors in a real case study of a plant variety protection system for soybean. Euphytica, 127: 95-113.
Desarrollo de una base de datos genéticos para la caracterización del germoplasma argentino de soja
Ciencias Agronómicas - Revista XX - Año 12 - 2012 / 023 - 039

-39-
31. PEJIC I, AJMONE-MARSAN P, MORGANTE M, KOZUMPLICK V, CASTIGLIONI P, TARAMINO G, MOTTO M (1998) Comparative analysis of genetic similarity among maize inbredlines detected by RFLPs, RAPDs, SSRs, and AFLPs. Theor. Appl. Genet. 97: 1248-1255.
32. POWELL W, MORGANTE M, ANDRE C, HANAFEY M, VOGEL J, TINGEY S, RAFALSKY A (1996) The comparison of RFLP, RAPD, AFLP and SSR (microsatellite) markers for germoplasm analysis. Mol Breed, 2: 255-238.
33. PRIOLLI RHG, MENDES-JUNIOR CT, ARANTES NE, CONTEL EPB (2002) Characterization of brazilian soybean cultivars using microsatellite markers. Genet Mol Biol, 25: 185-193.
34. RONGWEN J, AKKAYA MS, BHAGWAT AA, LAVI U, CREGAN PB (1995) The use of microsatellite DNA markers for soybean genotype identification. Theor. Appl. Genet. 90: 43-48.
35. SAINI N, JAIN N, JAIN S, JAIN RK (2004) Assessment of genetic diversity within and among Basmati and non-Basmati rice varieties using AFLP, ISSR and SSR markers. Euphytica 140: 133-146.
36. SHOEMAKER RC, GUFFY RD, LORENZEN LL, SPECHT JE (1992) Molecular mapping of soybean: Map utilization. Crop Sci, 32: 1091-1098.
37. SNEATH PHA, SOKAL RR (1973) En: Numerical Taxonomy, W.H. Freeman and Company, San Francisco, California, USA, pp. 230-234.
38. SONG QJ, QUIGLEY CV, NELSON RL, CARTER TE, BOERMA HR, STRACHAN JL, CREGAN PB (1999) A selected set of trinucleotide simple sequence repeats markers for soybean cultivar identification. Plant Var and Seed, 12: 207-220.
39. SONG QJ, MAREK LF, SHOEMAKER RC, LARK KG, CONCIBIDO VC, DELANNAY X, SPECHT JE, CREGAN PB (2004) A new integrated genetic linkage map of the soybean. Theor Appl Genet, 109:122-128.
40. TAMS SH, MELCHINGER AE, BAUER E (2005) Genetic similarity among European winter triticale elite germplasms assessed with AFLP and comparisons with SSR and pedigree data. Plant Breeding, 124: 154-160.
41. UDE GN, KENWORTHY WJ, COSTA JM, CREGAN PB ALVERNAZ J (2003) Genetic diversity of soybean cultivars from China, Japan, North America and North American ancestral lines determined by amplified fragment length polymorphism. Crop Sci, 43: 1858-1867.
42. VOS P, HOGERS R, BLEEKER M, REIJANS M, VAN DE LEE T, HORNES M, FRIJTERS A, POT J, KUIPER M, ZABEAU M (1995) AFLP: A new technique for DNA fingerprinting. Nucleic Acids Res, 23: 4407-4414.
43. WEBER JL (1990) Informativeness of human (dC-dA)n(dG-dT)n polymorphims. Genomics, 7 : 524-530.
44. WELSH J, MCCLELLAND M (1990) Fingerprinting genomes using de PCR with arbitrary primers. Nucl Acid Res, 18: 7213-7218.
45. WILLIAMS JGK, KUBELIK AR, LIVAK KJ, RAFALSKI JA, TINGEY SV (1990) DNA polymorphisms amplified by arbitrary primers are useful as genetic markers. Nucl Acid Res, 18: 6531-6535.
46. ZHAO TJ, GAI JY (2004) The origin and evolution of cultivated soybean [Glycine max (L) Merr]. SciAgricult Sin, 37: 954-962.
47. ZHU SL, MONTI LM, AVITABILE A, RAO R (1999) Evaluation of genetic diversity in Chinese soyabean germplasm by AFLP. Plant Genet Res Newsletter, 119: 10- 14.
13. GOWER JC (1966) Some distance properties of latent root and vector methods used in multivariate analysis. Biometrika, 53: 325-338.
14. HYMOWITZ T, NEWELL CA (1981) Taxonomy of the genus Glycine, domestication and uses of soybeans. Econ Bot, 35: 272-288.
15. JACCARD P (1908) Nouvelles recherches sur la distribution florale. Bull. Soc. Vaud. Sci. Nat., 44:223-270.
16. JONES CJ, EDWARDS KJ, CASTAGLIONE S, WINFIELD MO, SALA F, VAN DEWIEL C, BREDEMEIJER G, VOSMAN B, MATTHES M, DALY A, BRETTSCHNEIDER R, BETTINI P, BUIATTI M, MAESTRI E, MALCEVSCHI A, MARMIROLI N, AERT R, VOLCKAERT G, RUEDA J, LINACERO R, VAZQUEZ A, KARP A (1997) Reproducibility testing of RAPD, AFLP and SSR markers in plants by a network of European laboratories. Mol Breed, 3: 381–390.
17. KEIM P, BEAVIS WD, SCHUPP J, FREESTONE R (1992) Evaluation of soybean RFLP marker diversity in adapted germplasm.Theor Appl Genet, 85: 205-212.
18. KOSMAN E, LEONARD KJ (2005) Similarity coefficients for molecular markers in studies of genetic relationships between individuals for haploid, diploid, and polyploid species. Molecular Ecology 14: 415-424.
19. LEAVY S, DEWES H (2009) Drivers y tendencias en los escenarios futuros de la cadena de la soja argentina. Anais do Congresso da SOBER (Sociedade Brasileira de Economia, Administraçao e Sociologia Rural), Porto Alegre, pp. 21.
20. MANTEL NA (1967) The detection of disease clustering and a generalized regression approach. Cancer Res, 27: 209-220.
21. MARTINEZ F (2010) Crónica de la soja en la región pampeana argentina. Para mejorar la producción, INTA EEA OLIVEROS, pp 141.
22. MAUGHAN PJ, SAGHAI MAROOF MA, BUSS GR, HUESTIS GM (1996) Amplified fragment length polymorphism (AFLP) in soybean: Species diversity, inheritance, and near-isogenic line analysis. Theor Appl Genet 93: 392–401.
23. MEDINI M, HAMZA S, REBAI A, BAUM M (2005) Analysis of genetic diversity in tunisian durum wheat cultivars and related wild species by SSR and AFLP markers. Genetic Resources and Crop Evolution, 52: 21-31.
24. MEESANG N, RANAMUKHAARACHCHI SL, PETERSEN MJ, ANDERSEN SB (2001) Soybean cultivar identification and genetic purity analysis using microsatellite DNA markers. Seed Science and Technology, 29: 637-645.
25. MENZ MA, KLEIN RR, UNRUH NC, ROONEY WL, KLEIN PE, MULLET JE (2004) Genetic diversity of public inbreds of Sorghum determined by mapped AFLP and SSR markers. Crop Sci 44: 1236-1244.
26. MUELLER UG, LAREESA WOLFENBARGER L (1999) AFLP genotyping and fingerprinting. Trends Ecol Evol, 14: 389-394.
27. MULLIS K, FALOONA F, SCHARF S, SAIKI R, HORN G, ERLICH H (1986) Specific Enzymatic Amplification of DNA in vitro: The Polymerase Chain Reaction. Proceedings of the Cold Spring Harbor Symposia on Quantitative Biology, pp. 263-273.
28. NARVEL JM, FEHR WR, CHU WS, GRANT D, SHOEMAKER RC (2000) Simple sequence repeat diversity among soybean plant introductions and elite genotypes. Crop Sci 40: 1452-1458.
29. NEI M (1978) Estimation of average heterozygosity and genetic distances from a small number of individuals. Genetics, 89: 583- 590.
30. OLSINA C (2012) Desarrollo de un banco de datos genéticos del germoplasma de soja argentino, Tesis de Maestría, Maestría en Genética, Facultad de Ciencias Agrarias, Universidad Nacional de Rosario, Rosario, Argentina.
OLSINA, Cesar; CAIRO, Carlos; PESSINO, Silvina
Ciencias Agronómicas - Revista XX - Año 12 - 2012 / 023 - 039


-41-
Artículo Original / Recibido 04/08/2012 - Aceptado 29/10/2012
La cuestión curricular en la Facultad deCiencias Agrarias de la UNR
Curricular issues at the College of Agricultural Sciences (UNR - Rosario National University)
Cavalli, A. y Rosenstein, S.Facultad de Ciencias Agrarias, UNRParque Villarino, S2125ZAA, Zavalla, Argentina,[email protected]
En la última década del siglo XX, en la Argentina se pusieron en tela de juicio los currícula universitarios bajo el justificativo de no responder a las necesidades sociales de un fin de siglo que se presentaba signado por la revolución científico tecnológica, y por un nuevo modelo de desarrollo y de concentración económica.Los cambios operados en el contexto socioeconómico nacional y regional repercutieron en la Universidad enfrentándola al desafío de atender a las crecientes exigencias sociales en un marco de restringidas políticas de financiamiento por parte del Estado.Las Universidades intentaron enfrentar las crisis que ponían en tela de juicio su hegemonía, legitimidad y autonomía institucional mediante procesos de reformas curriculares. De este modo “las contradicciones se mantuvieron bajo control a través de la gestión de las tensiones que ellas provocan recurriendo a mecanismos de dispersión”. (de Souza Santos1997: 228).En este trabajo se analiza de qué modo la Facultad de Ciencias Agrarias de la UNR realizó un cambio curricular tendiente a enfrentar las crisis intentando conciliar las contradicciones que surgían en torno a la cuestión curricular, en la polémica sostenida por los diversos grupos involucrados en el proceso.
Palabras clave:currículum - cambio - tensiones
Resumen
In the last decade of the twentieth century, the university curricula in Argentina were brought into question on the grounds that they failed to meet the social needs of the end of a century that appeared marked by scientific and technological revolution, and by a new model of development and economic concentration.The changes in the national and regional socio-economic context affected the University by urging it to face the challenge of meeting the increasing social demands within a framework of restricted public funding policies.Universities attempted to address the crisis that brought into question their hegemony, legitimacy and institutional autonomy through curriculum reform processes. Thus "the contradictions are kept under control through management of the tensions that lead them through mechanisms of dispersion." (de Souza Santos1997: 228).This paper analyzes how the College of Agricultural Sciences at UNR conducted a curriculum change aimed at addressing the crisis by trying to reconcile the contradictions that arose around the curricular issue in the debate held by the various groups involved in the process.
Key words:curriculum - change - tensions
Abstract
Introducción
En la última década del siglo XX, en la Argentina se pusieron en tela de juicio los currículos universitarios bajo el justificativo de no responder a las necesidades sociales de un fin de siglo que se presentaba signado por la revolución científico tecnológica, y por un nuevo modelo de desarrollo y de concentración económica.
Los cambios operados en el contexto socioeconómico nacional y regional repercutieron en la Universidad enfrentándola al desafío de atender a las crecientes exigencias sociales en un marco de restringidas políticas de financiamiento por parte del Estado.
La sanción de la Ley de Educación Superior en 1995, -en el marco de las concepciones neoliberales dominantes que asociaron calidad educativa con rentabilidad medida en términos de eficacia, rendimiento y productividad- señaló el rumbo de la nueva política educativa impulsada desde el Estado para las universidades. Esta ley constituye un instrumento regulatorio que afecta a los distintos actores con prohibiciones o mandatos e instituye nuevas reglas de juego en las relaciones entre el Estado y la universidad.
Se estableció la evaluación institucional, se fijaron requisitos para ciertos planes de estudio y se normó sobre la acreditación de carreras de interés público (entre las que se hallan las Ciencias Agrarias). Se creó la CONEAU (Comisión Nacional de Evaluación y Acreditación Universitaria), encargada de coordinar la gestión de los procesos de evaluación interna y externa de las instituciones universitarias y la acreditación de carreras de grado y de posgrado. Asimismo y como parte de las acciones que se desarrollaron desde el Programa de Reforma de la Educación Superior (PRES) -cuyo objetivo fue 'el mejoramiento de la eficiencia y calidad de la gestión universitaria y también de la reforma académica, administrativa y de gestión'- se puso en marcha el programa FOMEC (Fondo para el Mejoramiento de la Calidad) destinado a financiar programas de reforma al nivel de grado y postgrado con recursos provenientes de créditos otorgados por el Banco Mundial.
La presión ejercida desde el Estado por la 'adecuación' de los currículos al progreso de la ciencia y la tecnología en los distintos ámbitos del conocimiento y de la cultura, junto con la demanda social de una profesionalización que atendiera a la creciente exigencia de una formación articulada con el mundo de la producción y del trabajo, tuvo como respuesta procesos de
Ciencias Agronómicas - Revista XX - Año 12 - 2012 / 041 - 044

-42-
función (la de la Comisión) era coordinar el proceso que emprendiera la comunidad en su conjunto para determinar si era
4necesario o no realizar una reforma curricular .
Se manifestaba de este modo la contradicción latente entre las exigencias de realizar cambios en los tiempos establecidos según los compromisos contraídos ante el FOMEC y la necesidad de llevar a cabo un proceso institucional reflexivo cuyos tiempos no respondían a la lógica externa del corto plazo. Se ponía en evidencia la tensión que provoca la contradicción entre “la reivindicación de la autonomía en la definición de los valores y de los objetivos institucionales y la sumisión creciente a criterios de eficacia y productividad de origen y naturaleza empresarial” (de Souza Santos:229).
Pero, cuando una institución universitaria se interroga acerca de su currículum, se está cuestionando acerca del proyecto educativo que, de manera explícita o no, pone de manifiesto la intencionalidad con la que orienta la concreción de las funciones que estima que le son propias. En ese sentido el currículum es la 'arena' en la que se dirimen proyectos políticos, sociales y epistemológicos acerca de la acción educativa. Las decisiones que se toman alrededor del currículum son decisiones valorativas, no neutrales, que implican opciones ideológicas y epistemológicas. Como señala Contreras Domingo (1990: 230) “el currículum no es una idea inmaterial, sino el reflejo material, socialmente construido, de ciertas pretensiones que se tienen para la escuela [...] cualquier intento de innovación supone un conflicto con aquella práctica curricular en funcionamiento y con las condiciones de realidad en las cuales y por las cuales mantiene su existencia”.
¿Qué significados adquiría entonces el 'análisis integral' del currículum al que se refería la Comisión de Reforma Curricular?. ¿Qué se entendía por currículum?, ¿De qué modo se intentó compatibilizar las contradicciones?
IIEn el proceso de cambio curricular en la Facultad convergían la 'exigencia' y la 'necesidad' y esta tensión expresaba además perspectivas diferentes con respecto al currículo y con respecto al cambio.
Tal como ya ha afirmado, la Facultad se propuso 'analizar integralmente' el currículum. Pero este análisis adquirió diferentes significados según el alcance que diferentes grupos asignaban a la cuestión curricular. Para algunos el problema curricular era un problema técnico racional. Por lo tanto, había que detectar, mediante un 'buen' diagnóstico realizado con bases científicas, cuáles eran los problemas que impedían lograr los resultados esperados, luego se debía considerar los factores que intervenían en el problema para elegir los procesos adecuados que permitieran alcanzar las soluciones necesarias, mediante la aplicación práctica de los conocimientos científicos a las necesidades particulares de la Facultad. Además se debían diseñar los instrumentos que permitieran 'monitorear' la implementación de la propuesta de modo de asegurar el cumplimiento de lo prescripto. Para este grupo, el cambio curricular consistía en llevar a cabo un proceso técnico que suponía la necesidad de 'expertos' en currículum que analizaran el plan de estudio vigente para 'detectar las fallas' y diseñar un nuevo plan.
Desde esta perspectiva, el docente -visto como “un adaptador racional del cambio [...] adoptaría, por convicción intelectual, la
reformas curriculares a través de los cuales las Universidades intentaron enfrentar las crisis que ponían en tela de juicio su hegemonía, legitimidad y autonomía institucional (de Souza Santos, 1997a). De este modo “las contradicciones se mantuvieron bajo control a través de la gestión de las tensiones que ellas provocan recurriendo a mecanismos de dispersión”. (de Souza Santos: 228).
En este trabajo se analiza de qué modo la Facultad de Ciencias Agrarias de la UNR realizó un cambio curricular tendiente a enfrentar las crisis intentando conciliar las contradicciones que surgían en torno a la cuestión curricular, en la polémica sostenida por los diversos grupos involucrados en el proceso.
Si bien se trata de un proceso iniciado hace ya más de diez años, creemos importante dar a conocer esta reflexión para que, a la luz de lo ocurrido, podamos rediseñar el trabajo presente y futuro, reformulando los errores y valorizando los logros. Sobre todo, si partimos de la concepción de que la currícula es precisamente un proceso en permanente revisión.
IA partir del advenimiento de la democracia, en la Facultad de Ciencias Agrarias de la Universidad Nacional de Rosario (UNR) se problematizó la cuestión curricular a veces en forma amplia y en otras limitándose al plan de estudios.
Pero por lo general, las reformas quedaron reducidas a la ubicación o reubicación de materias, a la incorporación o reducción de contenidos, sin cuestionar en profundidad la estructura curricular y mucho menos, los contextos de desarrollo y de significación. Los sucesivos cambios de Planes de Estudio (1986, 1991 con reajustes 'coyunturales' en 1996) parecían 'no dar respuestas' a las dificultades que se presentaban para el avance regular de los alumnos en la carrera. Asimismo existía una fuerte crítica de los graduados hacia una formación 'desvinculada del medio rural', que desconocía la problemática del sector impartiendo conocimientos abstractos y 'desarticulados'.
Esta situación se unía a la preocupación de las Facultades de 1Agronomía del país –expresada en AUDEAS - por cuanto sus
carreras registraban uno de los mayores índices de demora en el egreso de sus estudiantes, en un contexto de políticas universitarias tendientes al acortamiento de las carreras de grado de acuerdo con las 'sugerencias' emanadas de los organismos internacionales de financiamiento.
En ese marco, el proyecto institucional presentado a FOMEC por la Facultad de Ciencias Agrarias de la UNR en lo que hace específicamente al plan de estudios, planteaba su revisión para lograr “la reducción de la duración de la carrera de grado para el
2título de Ingeniero Agrónomo a un período de 5 años” .
3Sin embargo, en 1997 la Comisión de Reforma Curricular -creada por el Consejo Directivo de la Facultad- planteó que era necesaria una evaluación curricular y no sólo una revisión del Plan de Estudios. Señalaba que esto requería 'analizar integralmente el Currículo en vigencia' y que su principal
1AUDEAS es la Asociación Universitaria de Educación Agropecuaria Superior.2UNR. Fac. Cs. Agr. Proyecto FOMEC 126-127. “Mejoramiento de la calidad de la enseñanza agropecuaria de nivel universitario”. Formularios para la presentación de Proyectos. Rosario, noviembre 1995.3Esta Comisión estaba integrada por docentes representantes de los Departamentos, dos graduados, dos estudiantes de las agrupaciones que integraban el Consejo Directivo y una no docente del Área Alumnado. La coordinación estuvo a cargo del Área pedagógica de la Facultad. Posteriormente se integró en algunas reuniones, un representante del Colegio de Ingenieros Agrónomos.
4 “La evaluación curricular en la Facultad de Ciencias Agrarias”. Documento elaborado por la Comisión para la Evaluación Curricular. Setiembre, 1997
Ciencias Agronómicas - Revista XX - Año 12 - 2012 / 041 - 044
La cuestión curricular en la Facultad de Ciencias Agrarias de la UNR

-43-
De allí en más, la confrontación entre los diversos grupos con intereses opuestos y contradictorios, se puso en evidencia en la determinación de la selección de los contenidos que integrarían la síntesis de elementos culturales que conformarían la propuesta curricular (de Alba: 95). ¿Qué cuestiones debían dirimirse? ¿Cómo se intentó diluir las nuevas tensiones?
IIILas dicotomías teoría-práctica y educación-trabajo, habían quedado evidenciadas en el diagnóstico resultante del análisis realizado por la comunidad, en el que se señalaba la existencia de “un currículum rígido y frondoso, de estructura disciplinar sin instancias de articulación, con escasas y aisladas prácticas profesionales”, con “procesos de enseñanza basados fundamentalmente en la transmisión de contenidos, con una marcada separación entre teoría y práctica”, resaltándose que en consecuencia se brindaba una formación excesivamente abstracta desligada de la realidad social y productiva”.
Pero había que decidir qué quedaba dentro del currículum y qué quedaba afuera; si la diferenciación entre los contenidos curriculares respondería a una lógica de clasificación fuerte (esto es, con nítidas fronteras entre unos y otros) o si habría espacios compartidos; qué peso relativo tendrían los espacios curriculares tanto en lo referido a su ubicación como a la carga horaria. Y todo esto implicaba dirimir cuestiones de poder tanto en lo referido a concepciones epistemológicas acerca de la formación del Ingeniero Agrónomo como a cuestiones institucionales relativas al lugar que ocuparían los sujetos docentes en función de la relevancia que se le otorgara a 'su asignatura'. Ninguno de los grupos contaba con una relación de fuerzas que le permitiera imponer su posición. Por lo cual, la negociación fue el mecanismo al que apeló la Comisión de Cambio Curricular para lograr consensos de modo de dar cabida a las materias existentes y al mismo tiempo incluir espacios integradores de práctica profesional y cursos electivos (de acuerdo con las 'nuevas tendencias' de formación profesional).
Y en pos 'del consenso', durante la negociación se intentó nuevamente eludir los conflictos buscando equilibrar la carga horaria –reduciendo proporcionalmente el tiempo destinado a cada asignatura- y proponiendo la reubicación de los contenidos 'de profundización' en los cursos electivos. De este modo, no sólo se lograba llevar la carrera a los cinco años exigidos, sino también ubicar los nuevos espacios curriculares.
En el nuevo diseño curricular se procuró diferenciar los contenidos obligatorios -incluidos en las distintas materias-, de los opcionales –adaptados a las orientaciones personales-; se replanteó la división del saber en disciplinas, al incluir los Talleres de integración como una progresiva aproximación entre especialidades que, sin perder la especificidad de cada una de ellas, propiciara la coordinación interdisciplinaria en función de la resolución de problemas propios de la actividad profesional.
De manera que, como resultado de la lucha entre los diferentes sectores intervinientes en el proceso curricular, la síntesis cultural que se enuncia en el Currículum de la Facultad de Ciencias Agrarias, “contiene de diversas maneras no sólo elementos del grupo dominante o hegemónico, sino también de los otros grupos o sectores sociales” (de Alba: 63). Incorpora –aún de manera contradictoria- innovaciones con respecto al currículum anterior, pero son justamente estos cambios los que constituyen el eje de la polémica alrededor de los cuales se expresan las resistencias.
Es elocuente al respecto, el documento que la Comisión de Cambio Curricular elevó al Consejo Directivo al momento de
lógica que impregnara todo el proyecto y aprendería a solucionar problemas y tomar decisiones racionales que facilitaran el cambio” (Contreras Domingo: 232)
Para otros –que asumían una perspectiva política del cambio curricular- lo que se estaba dirimiendo era un conflicto de intereses entre sectores que confrontaban ideas diferentes acerca de la educación y de lo que 'debía ser' el currículum. El currículum debía ser el resultado de una decisión –producto de la deliberación- que guiara y seleccionara la acción posible. Estas decisiones, lejos de tener un carácter prescriptivo, se refrendarían en la práctica y a partir de sus resultados podrían y deberían revisarse y modificarse. En ese sentido los cambios que se realizasen debían atender al análisis de las reflexiones, críticas y sugerencias de los docentes que tendrían a su cargo la aplicación de las modificaciones, de los alumnos que vivenciaban 'los problemas' curriculares y de los graduados.
Este último grupo era el que predominaba en la Comisión de Cambio Curricular y es por eso que el acento de su accionar estuvo puesto en la participación de todos los claustros, en la negociación y en la búsqueda de consensos. En tal sentido el documento ya mencionado en el apartado I de este trabajo, señalaba que “el aspecto central de la filosofía de esta propuesta, consensuado por esta Comisión, es que todos los miembros de la comunidad universitaria deben ser partícipes en este proceso” y más adelante agregaba que “una propuesta hecha sin la participación activa de todos los miembros de una comunidad universitaria es, a lo sumo, un buen documento”y que “para que no sea sólo un buen documento, sino que sirva para generar cambios superadores de la realidad actual, es necesario un compromiso de todos los actores de la Institución, compromiso que se generará a partir de que todos sintamos que el cambio nos pertenece, porque lo hemos hecho nosotros mismos”.
Para cumplir ese cometido, generaron intensas actividades que abarcaron consultas a especialistas y graduados, encuentros y jornadas tanto internas como con otras Facultades de Ciencias Agrarias del país, que convocaron a docentes, graduados, estudiantes y no docentes. Y en este proceso, la cuestión curricular se convertía en el emergente de los diversos conflictos que existían al interior de la Facultad y se ponían en evidencia las pujas de poder entre diferentes sectores. Sin embargo, en pos 'del consenso', los conflictos se omitían, las decisiones se diluían y no existía una instancia de convalidación de las conclusiones de los debates.
Poco a poco, la aspiración deliberativa inicial en torno a los fines del currículum, fue quedando limitada al diseño curricular. Frente a las presiones ejercidas por el vencimiento de los plazos establecidos por FOMEC para la puesta en marcha de un nuevo plan a partir del año 2000, los tiempos de discusión política se dieron por finalizados por entenderse que se habían logrado los acuerdos marco necesarios para realizar los cambios. La Comisión de Cambio Curricular debió abocarse a la elaboración del plan, reduciéndose así la aspiración inicial de abordar la problemática curricular en todas sus dimensiones, a lo que de Alba (1998) denomina “plano estructural formal del currículum”. De este modo, la tensión entre necesidad y exigencia de cambio curricular, se dispersó en tanto se entendió que, como producto de la deliberación, las contradicciones iniciales se habían subsanado. No sólo se había visto la necesidad de introducir cambios, sino que se había 'acordado' el acortamiento de la carrera. Así la exigencia pasa a ser considerada como necesidad.
Ciencias Agronómicas - Revista XX - Año 12 - 2012 / 041 - 044
Cavalli, A. y Rosenstein, S.

-44-
IVEl Plan de estudios de la Facultad de Ciencias Agrarias de la UNR, expresa “la síntesis de la polémica que en torno a él han sostenido los diversos sujetos, grupos y sectores sociales con intereses específicos en su formulación” (de Alba,1991: 141), constituye el proyecto político, cultural, de los sujetos y sectores sociales que impulsaron la propuesta curricular en su intento de afrontar las crisis de legitimidad y hegemonía. Pero, “una nueva propuesta curricular no se instaura en un espacio vacío sino que emerge en el seno de prácticas más o menos establecidas e intereses encontrados” (Terigi, 1999: 111). Si bien el diseño curricular constituye un instrumento que incide en las prácticas a partir de sus determinaciones, son los sujetos sociales, concretos, situados, con intereses y vinculaciones también concretas y situadas, quienes en sus acciones cotidianas dan significado a lo prescripto. “[...] son las transacciones únicas, concretas y particulares de la práctica (cotidiana) las que determinan su sentido” (Angulo Rasco, 1994: 114)
En ese sentido, es en la dinámica del desarrollo curricular que se pueden observar los alcances de los cambios introducidos.
presentar el nuevo Plan de Estudios: “Vivimos en este tiempo el desafío de elaborar una propuesta curricular que resulta de las confrontaciones y negociaciones. [...] el proyecto que presentamos [...] es una radiografía de lo que hoy somos y pudimos acordar, es lo que hoy parece ser lo posible [...] Se nos pidió que elaboráramos un Plan de Estudio en el que no quedaran afuera ninguna de las asignaturas existentes, pero que se redujera la duración de la carrera a cinco años. [...]que los alumnos tuvieran un contacto con las actividades propias de la profesión a lo largo de la carrera con una visión global y que además se incluyera la posibilidad de acceder a los nuevos conocimientos que demanda la actividad profesional. Como verán, se nos pidió que rompiéramos con una ley de la física. Esto es que hiciéramos que más de un cuerpo ocupara el mismo espacio o aún menor. Obviamente no logramos hacerlo, pero sí buscamos una alternativa posible en las condiciones existentes, [...] que permitiera atender a los requerimientos y que fuera medianamente factible de acuerdo con quiénes somos y qué estamos más o menos dispuestos a aceptar”.
Bibliografía
1. ANGULO RASCO, J F y BLANCO, N. (coord.), (1994). Teoría y desarrollo del currículo. Málaga. Aljibe.
2. De ALBA, A. (1991); Evaluación curricular. Conformación conceptual del campo. México. UNAM.
3. De ALBA, A. (1995); Currículo, mito y perspectivas. Buenos Aires: Miño y Dávila.
4. De SOUZA SANTOS, B. (1997); Por las manos de Alicia. Lo social y lo político en la posmodernidad. Bogotá: Ediciones Uniandes.
5. GIMENO SACRISTÁN, J. y PÉREZ GÓMEZ, Á. (1993); Comprender y transformar la enseñanza. Madrid. Morata.
6. GIMENO SACRISTÁN, J. (1988); El currículum: una reflexión sobre la práctica. Madrid: Morata.
7. ROSENSTEIN, S.; CAVALLI, A.; FARINA, J.; ROSBACO, I; TREVIZÁN, A (2003) El cambio del Plan de Estudios en la Facultad de Ciencias Agrarias: construyendo nuevos significados y prácticas”. Revista de Investigaciones de la Facultad de Ciencias Agrarias. UNR. Nº 3. ISSN 1515-9116. pp 47-65
8. ROSENSTEIN, S.; CAVALLI, A. (2007) Viejas y nuevas prácticas en un contexto de cambio. El caso de la Facultad de Ciencias Agrarias del la UNR. Revista de Educación Superior Nº 141. Enero-marzo 2007. Asociación Nacional de Universidades e Instituciones de Educación Superior. Méjico. ISSN 0185-2760. pp145-166
9. TERIGI, F. (1999); Currículo. Itinerarios para aprehender un territorio. Buenos Aires: Santillana.
Ciencias Agronómicas - Revista XX - Año 12 - 2012
La cuestión curricular en la Facultad de Ciencias Agrarias de la UNR

-45-
Durante los días 25 al 27 de Octubre de 2012 tuvo lugar el encuentro científico “III Ciclo de Seminarios sobre Avances en la Caracterización Genética y Molecular de la Apomixis en Gramíneas Forrajeras”, organizado por la Carrera de Doctorado en Ciencias Agrarias de la Facultad de Ciencias Agrarias, Universidad Nacional de Rosario. Esta reunión tuvo como objetivo retomar los ciclos de seminarios previamente organizados sobre esta temática, que se iniciaron en el año 2008 en el Departamento de Agronomía de la Universidad Nacional de Sur, Bahía Blanca y continuaron en el año 2010 en la Facultad de Ciencias Agrarias de la Universidad Nacional de Rosario. El objetivo de estos encuentros fue discutir los resultados del trabajo que realizan varios grupos de investigación argentinos respecto a la elucidación de las bases genéticas y moleculares de la apomixis en las gramíneas. En esta oportunidad asistieron investigadores de las Universidades Nacionales del Nordeste, Misiones, del Sur y Rosario. Como investigador extranjero invitado participó el Dr. Fulvio Pupilli, del CNR-Istituto di Genetica Vegetale, Perugia, Italia. El ciclo contó con una conferencia plenaria presentada por el Dr. Pupilli y seminarios a cargo de investigadores, post-doctorandos y doctorandos argentinos, donde se comunicaron avances importantes en el conocimiento de la apomixis, a saber: a) se discutió la presencia de aposporía y diplosporía conjunta en los mismos individuos para dos especies de Paspalum ; b) se realizaron estudios sobre la relación entre el contenido de ADN genómico y características biológicas en especies de Paspalum;c) se confirmó que existe flujo genético desde el nivel diploide al poliploide en especies de Paspalum; d) se constató la ocurrencia de apomixis residual en especies diploides de Paspalum; e) se desarrollaron herramientas de gran importancia en el mejoramiento de forrajeras apomícticas nativas del género Paspalum, como el establecimiento de poblaciones de centenares de individuos sexuales a los cuales se les transfirieron masivamente genes provenientes del germoplasma apomíctico, la inscripción de cultivares mejorados por selección y la caracterización de especies con potencial forrajero para áreas específicas; f) se avanzó en la realización de mapeo genético y caracterización de la región que determina la apomixis en dos especies de Paspalum; g) se identificaron genes involucrados en el desarrollo del endosperma en la misma especie; h) se analizó la expresión de genes que participan del proceso de apomixis en Paspalum y se obtuvieron las primeras transformantes donde la actividad de esos genes ha sido modificada; i) se profundizó la caracterización de la apomixis diplospórica en Eragrostis curvula, la relación de su expresividad con el estrés, y el establecimiento de poblaciones de mapeo genético en esta especie. A continuación se publican los resúmenes de los seminarios presentados en la mencionada reunión.
III Ciclo de Seminarios sobre avances en lacaracterización genética y molecular de la apomixis
en gramíneas forrajeras
Coordinadores:
Dra. Silvina C. PessinoFacultad de Ciencias Agrarias, Universidad Nacional de Rosario, CONICET – FCA [email protected]
Dr. Juan Pablo A. OrtizFacultad de Ciencias Agrarias, Universidad Nacional de Rosario, CONICET – [email protected]
Resumen
Ciencias Agronómicas - Revista XX - Año 12 - 2012
Resúmenes


-47-
Se estudiaron genotipos pentaploides (2n=5x=50) de P. arundinellum y P. mandiocanum, que poseen un sistema genético excepcional para el género y las gramíneas en general. Consiste en la existencia de pentaploidía, seguramente de origen alopoliploide, asociada a un comportamiento cromosómico principalmente asináptico durante la meiosis, con formación de núcleos de restitución de primera división (RPD), producción de gametos masculinos y femeninos 2n, reproducción apomíctica combinada (diplosporia coexistente con aposporia en el mismo genotipo) y seudogamia. También se estudió un tetraploide (2n=4x=40) de P. arundinellum conespecífico y simpátrico de los pentaploides, que posee un comportamiento meiótico con 20I + 10 II, forma núcleos de restitución (RPD), gametos 2n y se reproduce también por apomixis combinada (diplosporia y aposporia). Los resultados permiten establecer como hipótesis la posibilidad de que el origen de los pentaploides de P. arundinellum se explique mediante la formación de híbridos 2n+n, a partir de un tetraploide como el mencionado y un progenitor diploide sexual estrechamente emparentado que haya contribuido con un gameto reducido. Los aspectos más relevantes de este sistema genético y reproductivo en Paspalum están relacionados con los modos de formación de gametos 2n y la existencia de diplosporia en coexistencia con aposporia en el mismo óvulo. Este comportamiento posiblemente contribuyó a que estas especies, que seguramente se originaron por hibridaciones interespecíficas, lograran escapar a la esterilidad.
Resúmenes
Diplosporia concomitante con aposporia en Paspalum L.
1 2Honfi AI , Quarin CL
1IBS (Universidad Nacional de Misiones- CONICET)2 Instituto de Botánica del Nordeste (IBONE-CONICET)Facultad de Ciencias Agrarias UNNE. Corrientes, [email protected]
Resumen
Ciencias Agronómicas - Revista XX - Año 12 - 2012


-49-
La estimación del contenido de ADN mediante citometría de flujo ha facilitado los estudios sobre su relación con la historia de vida, los sistemas genéticos, y la evolución en diferentes especies de plantas. Se ha sugerido usar el término “valor C” en referencia al contenido de ADN del complemento haploide cromosómico entero (n), y el término “valor Cx” en referencia al contenido de ADN del juego cromosómico monoploide, es decir, del número básico de cromosomas. En plantas superiores de los géneros Veronica, Hieracium y Hypericum se ha documentado la existencia de correlación entre el tamaño del genoma y diferentes características biológicas. El objetivo de este trabajo fue determinar si el tamaño del genoma y algunos caracteres biológicos están correlacionados en el género Paspalum, especialmente en referencia al modo de reproducción, el nivel de ploidía, y los conceptos actuales sobre la clasificación taxonómica infragenérica. Se determinó el contenido relativo de ADN en picogramos (pg) por citometría de flujo en 46 especies del género Paspalum. Se analizaron 75 entidades (especies, variedades, citotipos o biotipos) de las cuales 31 eran de reproducción sexual y 44 apomícticas; 16 diploides y 59 poliploides. Se consideró el contenido relativo de ADN del juego de cromosomas monoploide (valor Cx) de cada entidad. El contenido relativo de ADN promedio para las entidades sexuales fue de 0,68 pg, mientras que para las apomícticas fue de 0,69 pg, no existiendo diferencias significativas en el valor de contenido relativo de ADN entre entidades sexuales y apomícticas. Cuando se analizaron las entidades de acuerdo con su nivel de ploidía, el valor Cx varió en promedio de 0,57 pg en entidades pentaploides a 0,74 pg en entidades diploides, pero las diferencias no resultaron estadísticamente significativas. Se observaron variaciones en el contenido relativo de ADN (valor Cx) entre las cuatro categorías infragenéricas, seleccionadas por estar mejor representadas en número de entidades en este estudio. El grupo Dilatata (8 entidades), reflejó menor valor Cx con un promedio de 0,52 pg. Este valor Cx fue estadísticamente diferente de los valores de las otras categorías infragenéricas analizadas: los grupos Notata y Plicatula y el subgénero Anachyris. El grupo Notata (14 entidades en cuestión) mostró un Cx promedio de 0,65 pg, estadísticamente diferente de Dilatata, Plicatula y Anachyris; mientras que Anachyris, con 7 entidades, presentó el mayor valor Cx con un promedio de 0,84 pg; sin embargo el contenido de ADN del subgénero Anachyris no fue estadísticamente diferente del observado en el grupo Plicatula (16 entidades representadas) con un valor Cx promedio de 0,82 pg. Estos resultados nos permiten concluir que el tamaño del genoma en el género Paspalum no está relacionado con el modo reproductivo ni el nivel de ploidía de las entidades que lo integran; mientras que sí está relacionado con eventos que en la historia evolutiva del género llevaron a las diferencias morfológicas en las que están basados las distintas agrupaciones infragenéricas consideradas actualmente.
Resúmenes
Relación entre el contenido de ADN y las característicasbiológicas en Paspalum
1 1 2 3 1Galdeano F , Urbani M , Honfi A , Matzk F , Quarin CL
1Instituto de Botánica del Nordeste (IBONE)Facultad de Ciencias Agrarias, Universidad Nacional del Nordeste, Pcia. de Corrientes2IBS (Universidad Nacional de Misiones- CONICET)3 Institut für Pflanzengenetik und Kulturpflanzenforschung (IPK)06466, Gatersleben, [email protected]
Resumen
Ciencias Agronómicas - Revista XX - Año 12 - 2012


-51-
Paspalum simplex es una gramínea nativa de la región chaqueña y se caracteriza por la presencia de varios niveles de ploidía. El citotipo tetraploide predomina en toda la región y es de reproducción apomíctica, mientras que el diploide es sexual alógamo y habita en el suroeste de la región. El objetivo del trabajo fue evaluar la diversidad presente dentro y entre poblaciones diploides y tetraploides de P. simplex, mediante el estudio de variabilidad fenotípica y polimorfismos del ADN. Veintitrés caracteres morfo-fisiológicos y 10 cebadores de ISSR fueron utilizados para evaluar la diversidad en seis poblaciones naturales de P. simplex. Dos de las poblaciones evaluadas fueron diploides y cuatro tetraploides. Una de las poblaciones tetraploides es simpátrica con una de las diploides, mientras que el resto de las poblaciones son alopátricas. De las 23 características fenotípicas evaluadas, 16 mostraron gran diversidad, en especial para la producción de biomasa estacional, presencia de racimo terminal y pubescencia foliar. El estudio de la diversidad a través de ambas técnicas mostró que la variabilidad intrapoblacional fue superior en las diploides que en las tetraploides. Además dentro de las poblaciones tetraploides, la simpátrica con la diploide fue más variable que las demás. Se destaca que la diversidad interpoblacional fue superior a la intrapoblacional, y que como era de esperar, las poblaciones diploides y tetraploides simpátricas tuvieron la menor distancia genética observada. El índice de correlación de Pearson para diversidad medida por marcadores moleculares y caracteres morfo-fisiológicos fue alto (r=0.6 para diversidad entre poblaciones y r=0.9 para diversidad dentro de poblaciones). La variabilidad genotípica a nivel tetraploide sería generada mayormente a partir de las poblaciones diploides altamente variables.
Resúmenes
Estudio de la diversidad contenida en poblaciones diploides ytetraploides de Paspalum simplex que crecen
en forma simpátrica y alopátrica
Brugnoli EA, Urbani MH, Quarin CL, Martínez EJ, Acuña CA
Instituto de Botánica del Nordeste (IBONE)-CONICETFacultad de Ciencias Agrarias, Universidad Nacional del [email protected]
Resumen
Ciencias Agronómicas - Revista XX - Año 12 - 2012


-53-
La apomixis se encuentra generalmente asociada a la poliploidía. El carácter está ampliamente difundido en el género Paspalum (Poaceae), cuyas especies están organizadas como un complejo agámico. Los citotipos diploides son sexuales y la mayoría de los poliploides son apomícticos pseudógamos. En el caso de Paspalum rufum existen dos citotipos naturales: el diploide (2n=2x=20) sexual y auto-incompatible y el tetraploide (2n=4x=40) apomíctico, pseudógamo y auto-compatible. Estudios anteriores revelaron que un individuo diploide (Q3754) producía sacos embrionarios no reducidos y progenies por apomixis en baja proporción (apomixis residual). Estas observaciones indican que el carácter apomixis puede expresarse a nivel diploide, aunque en niveles residuales. Los objetivos de este trabajo fueron: a) investigar cuán difundida está la apomixis residual en los citotipos diploides naturales de esta especie, y b) seleccionar citotipos diploides contrastantes, algunos con un mayor grado de expresión de la apomixis y otros completamente sexuales, para utilizarlos en posteriores estudios genéticos. Se colectaron plantas diploides de tres poblaciones naturales. Se indujo la autofecundación de cinco plantas de cada una de las tres poblaciones, mediante la polinización conjunta con un citotipo tetraploide, utilizado como polen mentor. El comportamiento reproductivo se analizó en principio por citometría de flujo, ya que este método permite determinar si la semilla se originó por sexualidad o por apomixis, considerando la relación de contenido de ADN embrión/endospermo de cada semilla. Los primeros resultados indicaron que dos de las poblaciones sólo producían semillas por autopolinización de sacos reducidos. Sin embargo, una de las poblaciones mostró tres plantas que producían semillas a partir de sacos no reducidos. Estas plantas fueron nuevamente analizadas por citometría de flujo y se corroboró la producción de sacos no reducidos (entre 11% y 35%) identificándose además, en uno de los individuos, la formación de cariopses por apomixis. Posteriormente el individuo con mayor proporción de sacos no reducidos y uno 100% sexual, según la clasificación por citometría, se seleccionaron para realizar observaciones citoembriológicas. Estos análisis confirmaron la presencia de sacos apospóricos (17%) en el individuo con apomixis residual y también, aunque en menor proporción (6%), en el individuo sexual. Los resultados indican que los citotipos diploides presentan reproducción sexual, pero tienen la capacidad de generar sacos no reducidos en diferentes proporciones y en algunos casos generar progenies por apomixis. Este hallazgo confirma la presencia del los factores genéticos necesarios para la expresión del carácter al nivel diploide. Los estudios citoembriológicos indican que la capacidad de producir sacos apospóricos no implica la producción de semillas por apomixis sugiriendo un desacoplamiento entre los diferentes componentes de la apomixis. Estos resultados apoyan la teoría acerca del origen autoploide de los individuos poliploides apomícticos.
Resúmenes
Caracterización del comportamiento reproductivode citotipos diploides de Paspalum rufum
1-2 2 2 2 1-2Delgado, L. , Sartor M. , Quarin C. , Espinoza F. , Ortiz J.P.
1Laboratorio de Biología MolecularFacultad de Ciencias AgrariasUniversidad Nacional de RosarioZavalla, Santa Fe; 2Instituto de Botánica del Nordeste (IBONE)Facultad de Ciencias AgrariasUniversidad Nacional del Nordeste, Corrientes, [email protected]
Resumen
Ciencias Agronómicas - Revista XX - Año 12 - 2012


-55-
El mejoramiento genético de especies de Paspalum en Corrientes, Argentina, comenzó hace alrededor de 22 años en la Facultad de Ciencias Agrarias de la Universidad Nacional del Nordeste, aunque hace más de 30 años que se vienen realizando estudios genéticos básicos con miras al mejoramiento. El programa actual tiene como principales objetivos 1) la generación de información en genética y ecofisiología de especies del género con interés agronómico, 2) la generación y evaluación de técnicas de mejoramiento genético, 3) el desarrollo de plantas con bondades desde el punto de vista de la producción de forraje o césped, y 4) la liberación de cultivares comerciales para distintos ambientes. Tres cultivares forrajeros se destacan como los principales productos del programa hasta el presente. El primer cultivar se denomina Cambá FCA (P. atratum) y fue inscripto en el INASE en 1997. Posteriormente fueron desarrollados los cultivares Chané FCA (P. guenoarum) y Boyero UNNE (P.notatum) los que fueron inscriptos en 2002 y 2012, respectivamente. En los últimos tres años el programa se ha expandido a través de convenios con el INTA para la evaluación de ecotipos de P. simplex, con la Universidad de Florida para el desarrollo conjunto de cultivares de P. notatum y con la empresa PGG WrigthsonSeeds para la comercialización de los cultivares resultantes del programa y para el desarrollo conjunto de cultivares forrajeros y céspedes, incluyendo un número importante de híbridos y variantes naturales de diferentes especies del género. En la actualidad, se está trabajando mayormente en la caracterización genética y agronómica de ecotipos e híbridos tetraploides (2n=4x=40) de P. simplex, P. notatum, P. guenoarum, P. plicatulum, P. atratum, y P. lenticulare. Los híbridos tetraploides son generados mediante la intervención de tetraploides sexuales, obtenidos artificialmente, que son utilizados como progenitores femeninos en cruzamientos controlados. El proceso de hibridación es seguido por la identificación de los híbridos apomícticos mediante el uso de marcadores moleculares ligados a la apomixis, observaciones de sacos embrionarios o a partir de citometría de flujo en semillas. Las principales características que se tienen en consideración para la selección de los materiales son: la capacidad de crecimiento estacional, en particular durante los meses más fríos, la producción de semillas, la resistencia al frío, el valor nutritivo, y la extensión del período reproductivo. Varias líneas genéticas ya han sido identificadas como promisorias en las distintas especies en evaluación y se espera poder inscribir un cultivar que permita extender el período de producción de forraje en sistemas de producción de carne ubicados en el subtrópico, así como un cultivar a ser usado como césped de bajo mantenimiento que pueda ser establecido por medio de semillas.
Resúmenes
Mejoramiento genético de especies de Paspalum:una actualización
Acuña CA, Urbani MH, Quarin CL
Facultad de Ciencias AgrariasUniversidad Nacional del Nordeste, IBONE (CONICET-UNNE)[email protected]
Resumen
Ciencias Agronómicas - Revista XX - Año 12 - 2012


-57-
El grupo Plicatula es una categoría botánica informal infragenérica que agrupa alrededor de 30 especies afines a P. plicatulum, la especie inicialmente descrita entre las del grupo. El grupo se caracteriza por poseer el antecio de color pardo-negruzco, brillante y lemma estéril con pliegues transversales. Todos los representantes poliploides del grupo que han sido analizados en sus sistemas reproductivos son apomícticos y mayoritariamente tetraploides. Para algunas de estas especies poliploides apomícticas se conocen también citotipos diploides sexuales (2xS), mientras que para otras sólo se conocen representantes 4x apomícticos (4xA). Hasta hace muy poco, la posibilidad de mejoramiento genético basada en cruzamientos y recombinación génica era prácticamente nula, porque la mayoría de las especies son del tipo 4xA. Algunas especies 4xA se introdujeron al cultivo y se registraron cultivares, pero esto se hizo exclusivamente partiendo de la selección de biotipos destacados por sus condiciones agronómicas. Para las especies que contienen citotipos 2xS y 4xA queda la posibilidad de duplicar los cromosomas de los 2xS para generar madres sexuales 4xS y cruzarlas con individuos 4xA. El objetivo de este trabajo fue transferir genes de citotipos 4xA de distintas especies del grupo Plicatula hacia P. plicatulum, para formar, mediante selección por sistema reproductivo y fertilidad, una población 4xS de amplia variabilidad genética. Una planta 4xS de P. plicatulum obtenida al ser inducida con colchicina fue utilizada como progenitor femenino en cruzamientos con nueve genotipos 4xA (polinizadores), pertenecientes a nueve especies del grupo Plicatula. Dadas las condiciones de autoincompatibilidad de la planta 4xS no fue necesaria la castración y se supuso que todos los descendientes serían de origen híbrido. De hecho, todos los descendientes mostraron características fenotípicas indicativas de su origen híbrido. Se determinó el modo reproductivo de los híbridos por citometría de flujo, en base a la relación del contenido de ADN entre el embrión y el endospermo. La fertilidad de los híbridos se consideró en relación al porcentaje de espiguillas que producen grano. Se determinó la producción de semilla de algunos híbridos en condición de autopolinización y polinización libre. Teniendo en cuenta el modo reproductivo y la fertilidad, se seleccionaron los híbridos de reproducción sexual más vigorosos de cada familia, los cuales se usaron para formar una población 4xS. Se permitió la polinización libre entre todos los miembros de esta población 4xS y de este policruzamiento se cosecharon las semillas, se sembraron y se formó una población 4xS sintética, que se supone que tiene una amplia base genética debido a su origen. Se pretende que esta población sea la base para el mejoramiento genético de especies tetraploides apomícticas del grupo Plicatula. El próximo paso será seleccionar en esta población unas pocas plantas, por características agronómicas destacables y por mayor grado de autoincompatibilidad de cada una de ellas, pero con alto grado de fertilidad en cruzamientos. Estas plantas constituirán las madres en futuros cruzamientos entre plantas sexuales por plantas apomícticas dentro del grupo Plicatula.
Resúmenes
Transferencia generalizada de genes desde genotiposapomícticos a una población sexual en el grupo
plicatula de Paspalum
Novo PE, Espinoza F, Urbani M, Quarin CL.
Instituto de Botánica del Nordeste (IBONE-CONICET)Facultad de Ciencias Agrarias , UNNE. Corrientes, [email protected]
Resumen
Ciencias Agronómicas - Revista XX - Año 12 - 2012


-59-
Paspalum notatum Flüggé es una gramínea forrajera de origen americano con una amplia distribución geográfica en el continente. Posee razas diploides sexuales y poliploides apomícticos; siendo los tetraploides los que prevalecen. La apomixis es un mecanismo de clonación natural por medio de semillas donde no se produce variación por falta de recombinación meiótica en la parte femenina y ausencia de singamia. El mejoramiento genético por hibridación en especies apomícticas requiere de genotipos sexuales con igual nivel de ploidía para poder generar variación. En la naturaleza, nunca se encontraron individuos tetraploides sexuales de P. notatum y los pocos que se obtuvieron experimentalmente poseen una base genética muy estrecha para iniciar un programa de mejoramiento genético. El objetivo de este trabajo fue incrementar el pool génico del germoplasma tetraploide sexual de P. notatum transfiriendo, por medio de hibridación, la alta variación genética existente en los ecotipos tetraploides apomícticos. En esta primera etapa se obtuvieron 12 combinaciones diferentes (familias), a partir de cruzamientos controlados entre 3 madres tetraploides sexuales y 10 padres tetraploides apomícticos. Se obtuvieron entre 39 y 50 descendientes por cada familia. Se confirmó su origen híbrido, en una muestra del 20%, mediante el análisis de co-segregación de marcadores de ISSR específicos del padre apomíctico por parte de los descendientes. Los híbridos de las diferentes familias fueron clasificados por su modo de reproducción, a partir del empleo de un marcador de RAPD 100% ligado a la apomixis. Los resultados moleculares fueron confirmados, en una muestra al azar del 15 %, por clarificado de ovarios con metilsalicilato y posterior observación de los sacos embrionarios maduros en un microscopio con dispositivo Nomarski. En 10 familias se confirmó el origen híbrido de los descendientes; mientras que en dos de ellas se detectó un descendiente que no se originó por hibridación. La clasificación reproductiva de las familias fue coincidente por ambas metodologías y mostró rangos de segregación sexual vs. apomíctico de 1:1 hasta 8,6:1. En la mayoría de las familias hubo supremacía de descendientes sexuales sobre los apomícticos lo que concuerda con resultados previos obtenidos en la especie. En una familia se obtuvo una proporción de 1:1 entre sexuales y apomícticos lo cual resulta inédito para esta especie. La alta proporción de híbridos sexuales obtenidos en la mayoría de las combinaciones garantiza la generación de una población tetraploide sexual sintética con una base genética amplia. Un posterior análisis de diversidad genética en esta población sintética podrá establecer el avance genético logrado con respecto a los progenitores originales y de esa forma confirmar la eficacia de la transferencia de variabilidad desde los ecotipos tetraploides apomícticos.
Resúmenes
Obtención de variabilidad genética enlos tetraploides sexuales de Paspalum notatum a
partir de ecotipos tetraploides apomícticos
Zilli A., Barone JO., Ríos EF, Quarin CL, Acuña CA, Martínez EJ
Facultad de Ciencias AgrariasUNNE, IBONE-CONICET, Sargento Cabral 2131. C.C. 209(3400) Corrientes, [email protected]
Resumen
Ciencias Agronómicas - Revista XX - Año 12 - 2012


-61-
Acroceras macrum es una gramínea perenne de origen sudafricano que posee interesantes características forrajeras por ser una especie C de clima cálido, adaptada a terrenos bajos y anegadizos, con una excelente adaptación y producción en ambientes bajos del 3
norte argentino. La ausencia de semilla fértil para su implantación es una limitante para su amplia difusión. Existe escasa información sobre esta especie en lo referente a sus características genéticas y reproductivas, lo que dificulta el inicio de un proceso de mejora genética. El objetivo de este trabajo fue determinar los niveles de ploidía, el modo de reproducción y la diversidad genética de una colección de Acroceras macrum de gran parte del germoplasma presente en el NEA. Se realizaron recuentos cromosómicos por técnicas de citogenética clásica y citometría de flujo. Un total de 30 individuos resultaron ser tetraploides (2n=4x=36) y 27 pentaploides (2n=5x=45). Cuando se analizaron las asociaciones cromosómicas en la meiosis masculina, en la mayoría de las células madres del polen (CMP) de los individuos tetraploides se observaron 18 II; mientras que en los pentaploides los cromosomas se asociaron formando 18 II + 9 I. En anafase I se observaron cromosomas rezagados en ambos citotipos y en los pentaploides se observó la formación de puentes citoplasmáticos, aunque no se evidenció la formación de micronúcleos. La maduración de las microsporas fue homogénea en los tetraploides pero muy heterogénea en los pentaploides. Se observaron los sacos embrionarios maduros a partir de clarificado de ovarios y posterior observación de los mismos bajo un microscopio óptico con dispositivo Nomarski. En todos los ovarios analizados de los individuos tetraploides se observó la presencia de un sólo saco embrionario similar al tipo Poligonum. La diversidad genética se evaluó mediante marcadores ISSR y posterior análisis con los programas InfoGen y Genotype sobre 30 plantas tetraploides y 17 pentaploides. Un total de 91 loci ISSR fueron detectados en los 4x y 44 en los 5x. La distancia genética mostró un rango de variación de 0 a 0,70 siendo los 4x los que mayor variación mostraron. La mayoría de los 5x no se distanciaron por lo que resultaron ser un único genotipo. Los resultados preliminares obtenidos demuestran que la colección evaluada está compuesta por plantas 4x, de origen alopoliploide y un posible modo de reproducción sexual, y plantas 5x que poseen dos genomios homólogos y un tercero no homólogo de origen desconocido. Futuros estudios sobre fertilidad e hibridación en A. macrum nos permitirán saber si es posible la producción de semilla a partir de un proceso sexual y la obtención de nuevos genotipos con características superiores.
Resúmenes
Avances en la caracterización genética deAcroceras macrum Stapf.
1,2 1 1 1 1Ferrari Usandizaga SC , Martínez EJ , Quarin CL ; Zilli A , Brugnolli EA ,3 3 1Randazzo C , Pagano EN , Acuña CA
1Instituto de Botánica del Nordeste (IBONE)-CONICET,Facultad de Ciencias Agrarias, Universidad Nacional del Nordeste2EEA-INTA Corrientes3IGEAF-INTA [email protected]
Resumen
Ciencias Agronómicas - Revista XX - Año 12 - 2012


-63-
En estudios previos informamos la identificación de varios marcadores moleculares de AFLP, RAPD y RFLP completamente ligados al locus que controla la aposporía en Paspalum notatum. El objetivo de este trabajo fue caracterizar las secuencias de estos marcadores, estimar el número de copias de los mismos en el genoma, obtener las regiones flanqueantes de algunos de ellos por medio de estrategias de caminata cromosomal y realizar un análisis de mapeo in silico sobre arroz y maíz. Asimismo, el estado de metilación de dos secuencias asociadas a este locus fue determinado mediante experimentos RFLP utilizando enzimas sensibles a metilación. Como material vegetal se emplearon los genotipos tetraploides de P. notatum Q4188 (sexual), Q4117 (apomíctico) y una población segregante por el modo de reproducción derivada del cruzamiento entre ambos. Los marcadores moleculares de AFLP y RAPD ligados a la apomixis fueron ensayados sobre los genotipos parentales y su progenie a fin de confirmar su cosegración con el carácter aposporía. Las bandas correspondientes a cada uno fueron aisladas a partir del genotipo apomíctico (Q4117) y clonadas en el sistema pGEM-Teasy (Promega) y secuenciadas por Macrogen (Corea). Aproximadamente 5 kb correspondiente a diferentes marcadores fueron caracterizados por este procedimiento. El análisis de 14 secuencias de este tipo reveló que la mayoría de las secuencias no presentó similitudes significativas en la bases de datos. Sólo dos fragmentos correspondieron a ESTs expresados en Panicum virgatum. La estimación del número de copias mostró la presencia de elementos repetitivos como también de copia simple. En particular los fragmentos derivados de los marcadores PnMAC5 y PnMAI3 mostraron 1 y 2-4 copias por genoma, respectivamente. El marcador PnMAC5 resultó monomórfico entre los progenitores de la población de mapeo, mientras que PnMAI3 mostró un fragmento ligado a la aposporía. La extensión de este marcador por caminata cromosomal posibilitó la identificación de secuencias adicionales, una de las cuales correspondió a una proteína del tipo AMT-70. El análisis de mapeo in silico determinó que las secuencias asociadas al locus aposporía en P. notatum se localizan principalmente en los cromosomas 2 y 12 de arroz y 3 y 5 de maíz. Dos loci asociados a la apomixis fueron analizados en experimentos RFLP utilizando enzimas sensibles a metilación. Los resultados experimentales mostraron que ambas secuencias se encuentran metiladas en los genotipos sexuales y apomícticos y no fue posible detectar polimorfismos de metilación asociados con el modo de reproducción.
Resúmenes
Caracterización de secuencias, mapeo in silicoy análisis de metilación de secuencias
ligadas a la aposporía en Paspalum notatum
1,2 1,2 1 1 2 1Ortiz JPA , Podio M , Rodríguez MP , Felitti S , Stein J , Martínez EJ ,2 2 1
Siena LA , Quarin CL y Pessino SC
1Laboratorio de Biología MolecularFacultad de Ciencias AgrariasUniversidad Nacional de RosarioZavalla, Santa Fe, Argentina2Instituto de Botánica del NordesteFacultad de Ciencias Agrarias, Universidad Nacional del Nordeste,Corrientes, [email protected].
Resumen
Ciencias Agronómicas - Revista XX - Año 12 - 2012


-65-
Plicatula es una categoría botánica infragenérica informal, que incluye aproximadamente 30 especies del género Paspalum L. con marcadas afinidades morfológicas con la especie emblemática del grupo, Paspalum plicatulum Michx.
La mayoría de estas especies son tetraploides (2n = 4x = 40) y de reproducción apomíctica (4xA), y algunas de ellas tienen una contraparte coespecífica diploide (2n = 2x = 20) y de reproducción sexual (2xS). N
Los objetivos de este trabajo fueron: I) Generar una población segregante para el modo reproductivo mediante cruzamiento interespecífico de P. plicatulum x P. guenoarum; II) Establecer la estructura básica de los grupos de ligamientos de especies del grupo Plicatula a nivel tetraploide y localizar la región responsable de la apomixis en P. guenoarum. Se realizaron cruzamientos controlados utilizando como madre a una planta 4xS experimental y como parental masculino a un genotipo natural 4xA. La población segregante para el modo de reproducción estuvo compuesta por 206 individuos F . 1
El análisis del contenido relativo de ADN embrión:endospermo en semillas de estos híbridos por citometría de flujo mostró una relación 2:3 para 127 híbridos indicando que los mismos se reproducen por sexualidad, mientras que 79 individuos presentaron una relación embrión:endospermo de 2:5, indicando que los mismos se reproducen por apomixis. Del total de individuos de la población segregante, 89 F (55 sexuales y 34 apomícticos) formaron parte de la población de mapeo. Se ensayaron 18 combinaciones de 1
cebadores de AFLP.
Muchas especies del grupo son componentes de las pasturas naturales de América del Sur. Algunas de ellas han sido introducidas al cultivo como forrajeras en Argentina y otros países.
uestro grupo de trabajo ha logrado obtener recientemente plantas tetraploides artificiales completamente sexuales (4xS) a partir de un citotipo diploide de P. plicatulum, luego de un tratamiento con colchicina.
El mapa marco de ligamiento genético del genotipo tetraploide sexual de P. plicatulum consistió en 89 marcadores que fueron asignados a 11 grupos de cosegregación cubriendo una distancia total de 819 cM. El mapa marco de ligamiento genético de P. guenoarum tetraploide apomíctico consistió en 128 marcadores que se distribuyeron en 22 grupos de cosegregación, en una distancia total de aproximadamente 1393 cM. Mediante la combinación de datos de citometría de flujo y moleculares de cada individuo de la población de mapeo, se identificó en el mapa paterno al grupo de ligamiento en el que mapeó el carácter apomixis (grupo apo, P10). El grupo apo P10 quedó constituido por 7 loci: el locus del carácter apomixis, 5 marcadores paternos y un marcador biparental. Éstos son los primeros mapas de ligamiento genético para especies de Plicatula y podrán ser empleados como marco genético en estudios básicos y aplicados. Fue posible identificar el grupo de ligamiento en el que mapea la apomixis y, si bien no se identificaron marcadores que cosegreguen estrictamente con el carácter, esto podría ser de utilidad en futuras investigaciones concernientes al modo reproductivo.
Resúmenes
Caracterización de la región genómica dela apomixis en especies del grupo Plicatula de Paspalum L.
Aguilera PM, Quarin CL, Ortiz JPA, Espinoza F
Instituto de Botánica del Nordeste (IBONE-CONICET)Facultad de Ciencias Agrarias, Universidad Nacional del Nordeste.Sargento Cabral 2131, Corrientes, Argentina. [email protected]
Resumen
Ciencias Agronómicas - Revista XX - Año 12 - 2012


-67-
Paspalum notatum es utilizado como modelo en estudios de genética reproductiva vegetal. La especie es multiploide, incluyendo citotipos diploides y poliploides (3x, 4x, 5x, 6x y 8x). El citotipo tetraploide es el más frecuente, mientras que los demás poliploides son muy raros o han sido obtenidos en forma experimental. El citotipo diploide es sexual y autoincompatible mientras que los poliploides son apomícticos apospóricos, pseudógamos y autofértiles. La formación del endospermo en genotipos apomícticos no depende del aporte genómico 2:1 (materno:paterno), típico de la mayoría de las angiospermas. Este es un aspecto crucial en la perspectiva de incorporar el carácter apomixis a los cereales, ya que estos cultivos no producen granos si esa relación 2m:1p se desvía. En este trabajo analizamos la expresión génica durante la formación de semillas de P. notatum. Se realizaron cruzamientos entre biotipos con distintas ploidías y sistemas reproductivos. Se extrajo ARN de ovarios a distintos intervalos después de la polinización y se comenzó una caracterización del transcriptoma utilizando la metodología de cDNA-AFLP. Los resultados preliminares permitieron identificar transcriptos asociados con la producción de semillas en esta especie. Los transcriptos que se secuenciaron se hallan relacionados con: el transporte de aminoácidos, un factor de transcripción y dos proteínas de función desconocida expresadas en diversos tejidos de gramíneas. Los resultados derivados de este proyecto contribuirán a elucidar los mecanismos genéticos que, a nivel molecular, le permiten a este sistema prescindir de la estricta relación genómica 2m:1p para formar el endospermo, y por ende las semillas.
Resúmenes
Identificación de genes involucrados enel desarrollo de semillas de Paspalum notatum
1 2 1,2 2Felitti SA, Acuña CA , Ortiz JPA , Quarin CL
1Laboratorio de Biología MolecularFacultad de Ciencias AgrariasUniversidad Nacional de Rosario Pcia. de Santa Fe2Instituto de Botánica del Nordeste (IBONE)Facultad de Ciencias AgrariasUniversidad Nacional del Nordeste, Pcia. de [email protected]
Resumen
Ciencias Agronómicas - Revista XX - Año 12 - 2012


-69-
La apomixis apospórica es considerada una desviación de la vía canónica de reproducción sexual de las angiospermas, que conduce a la formación de progenies clonales. En las plantas apospóricas se observa una diferenciación de algunas células somáticas de la nucela del óvulo, las cuales adquieren un destino gamético sin atravesar un proceso de meiosis. En trabajos previos dirigidos a la identificación de transcriptos diferencialmente expresados en tejidos reproductivos de plantas apospóricas y sexuales de Paspalum notatum aislamos un candidato (código experimental N19) cuyo nivel de expresión superaba en tres órdenes de magnitud al de otros genes. La secuencia asociada mapeaba in silico en un sector del genoma de arroz que guarda sintenia con la región que controla la aposporía en Paspalum notatum. Este trabajo estuvo dirigido a caracterizar en detalle la secuencia de dicho transcripto y su expresión en tejidos reproductivos en distintos estadíos de desarrollo. Se realizaron experimentos de RACE (Rapid Amplification of cDNA Ends) que permitieron ampliar la secuencia conocida hasta un total de 714 nucleótidos. Una búsqueda Blastn en el NCBI reveló que la secuencia expandida presentaba máxima homología con el RNAm de una proteína hipotética de Sorghum bicolor (E val: 0.0, ID: 98%). El análisis de Blastx no mostró dominios putativos conservados, pero arrojó máxima homología en un sector de 70
-29aminoácidos de una proteína no caracterizada de Glycine max (E val: 5 e , ID: 80%). Mediante una serie de experimentos de PCR en tiempo real se cuantificó en forma relativa la expresión del transcripto en flores de plantas apomícticas y sexuales en cuatro estadíos de desarrollo: premeiosis, meiosis, postmeiosis y antesis. Se observaron patrones de expresión heterocrónicos, con un nivel de representación ligeramente mayor en la planta apomíctica en premeiosis (relación 1.6) y ligeramente menor en postmeiosis (relación 2.5). La localización in situ de la expresión se estudió mediante hibridización de tejidos reproductivos con sondas de ARN marcadas con digoxigenina, en los estadíos de premeiosis tardía y antesis, que resultan particularmente interesantes porque en ellos se inducen dos etapas claves de la aposporía: la aparición de células iniciales en la nucela y la partenogénesis, respectivamente. Los patrones de hibridización en premeiosis tardía resultaron contrastantes: mientras que en la planta apomíctica la hibridación se produjo tanto en la nucela como en la célula madre de la megáspora, en la planta sexual se observó señal únicamente en la célula madre de la megáspora. Nuestros resultados permiten concluir que N19 codifica para una proteína no caracterizada, cuyo transcripto se expresa en la célula madre de la megáspora en las plantas sexuales, pero tiene una representación ectópica en la nucela de las plantas apospóricas, justamente en el sitio donde se diferencian las células iniciales de la aposporia que adquieren destino gamético.
Resúmenes
Caracterización de N19, un transcripto asociadocon el desarrollo apospórico en Paspalum notatum
1 1 1 1 2Sartor M , Espinoza F , Seijo G , González AM , Pessino S
1Instituto de Botánica del Nordeste (IBONE),Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET),Facultad de Ciencias Agrarias, Universidad Nacional del Nordeste, Corrientes, Argentina2Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET),Facultad de Ciencias Agrarias, Universidad Nacional de RosarioZavalla, Provincia de Santa Fe, [email protected]
Resumen
Ciencias Agronómicas - Revista XX - Año 12 - 2012


-71-
Una secuencia homóloga al dominio EXS fue identificada por el Dr. Fulvio Pupilli, IGV, Perugia, Italia, en un clon de BAC que contenía marcadores moleculares completamente ligados al locus que controla la aposporía en Paspalum simplex. Dado que esta secuencia es una de las pocas que muestran capacidad codificante entre aquellas asociadas posicionalmente con este locus, decidimos investigar una hipotética relación funcional con la apomixis. El objetivo de este trabajo fue aislar la secuencia completa y caracterizar la expresión del gen EXS de Paspalum notatum. Siguiendo una estrategia de caminata cromosomal a partir del ADN genómico, se aislaron fragmentos de 2298 pb con alta homología con el gen EXS de arroz y maíz a partir del genotipo sexual Q4188 y del genotipo apomíctico Q4117. Los análisis de homología determinaron que ambos fragmentos resultaron prácticamente idénticos (% ID), por lo cual en lo sucesivo se trabajó con una secuencia consenso. Estudios de filogenia molecular mostraron que PnEXS presenta alta similitud con varios genes EXS y que agrupa junto a aquellos pertenecientes al género de las gramíneas. El mismo contendría los exones 4 al 10, interrumpidos por los correspondientes intrones. El análisis por Southern blot utilizando esta secuencia como sonda reveló la presencia de 2 a 4 copias del mismo en el genoma de P. notatum. Los estudios de expresión por PCR en tiempo real sobre inflorescencias en distintos estadios del desarrollo muestran un comportamiento similar entre los genotipos sexuales y apomícticos, no encontrándose diferencias significativas en ningún estadio del desarrollo reproductivo. El análisis de mapeo in silico, indicó que PnEXS es ortólogo a la copia del gen EXS de arroz y maíz presente en una posición distal del brazo largo del cromosoma 12 de arroz y en una región pericentromérica en los cromosomas 1 y 3 de maíz, lo cual es coincidente con regiones que ya han sido reportadas como asociadas a la aposporía en otras especies. Estudios de mapeo genético en P. notatum, lograron localizar una de las copias del gen PnEXS en el grupo de ligamiento Lg6a del genotipo apomíctico Q4117, que sería una de las copias no relacionadas a la aposporía.
Resúmenes
Análisis de expresión de genes EXS engenotipos apomícticos y sexuales de Paspalum notatum
1,2 1,2 1 3 1,2Podio M , Delgado L , Felitti S , Pupilli F , Ortiz JPA
1Laboratorio de Biología Molecular,Facultad de Ciencias Agrarias, Universidad Nacional de RosarioZavalla, Santa Fe, Argentina;2Instituto de Botánica del Nordeste, Facultad de Ciencias AgrariasUniversidad Nacional del NordesteCorrientes, Argentina. 3CNR-Istituto di Genetica Vegetale, Perugia, [email protected]
Resumen
Ciencias Agronómicas - Revista XX - Año 12 - 2012


-73-
La apomixis se refiere a diversos comportamientos reproductivos que resultan en una clonación vía semillas. Las plantas apomícticas omiten tanto la reducción meiótica como la fecundación de la ovocélula, y producen descendientes que son réplicas genéticas exactas de la planta madre. Esta reproducción clonal única aparece naturalmente en numerosos taxones de plantas, pero está ausente en las especies de gran cultivo. Su introducción en especies de consumo masivo representaría un beneficio potencial enorme para la agricultura, ya que permitiría la fijación inmediata del vigor híbrido. En los últimos años se ha generado una evidencia creciente acerca del rol que desempeñan ciertas vías de metilación de ácidos nucleicos en la expresión de varios tipos de apomixis gametofítica en las gramíneas. En Zea mays se identificaron dos mutantes de pérdida de función para genes de metiltransferasas de la vía de metilación del ADN dirigida por ARN (RdDM)(dmt102 y dmt103) que eran capaces de formar gametas femeninas no reducidas. En Paspalum notatum se identificaron dos genes homólogos a metiltransferasas (N69 y PnMT-A70) que están asociados por diferencia de expresión o posicionalmente con la apomixis. El objetivo de este trabajo fue caracterizar en detalle la secuencia y la expresión de estos genes candidatos en plantas apomícticas y sexuales de P. notatum. Se realizaron experimentos de RACE sobre bibliotecas no clonadas, los cuales permitieron extender las secuencias de los fragmentos N69 y PnMT-A70 en 918 y 565 pb adicionales, respectivamente. Se realizaron experimentos de PCR en tiempo real en tejidos reproductivos de plantas apomícticas y sexuales en diferentes estadíos de desarrollo. El transcripto PnMT-A70 presentó un aumento significativo de la expresión en antesis, sin embargo no se observó una expresión diferencial significativa entre plantas apomícticas y sexuales en ningún estadio ensayado. El transcripto N69 mostró diferencias de expresión significativa entre plantas apomícticas y sexuales en todos los estadios de desarrollo, presentando un aumento significativo de expresión en antesis sólo en la planta sexual. La diferencia de expresión en antesis se repitió en 6 individuos (3 apomícticos y 3 sexuales). Además se amplificaron mediante cebadores heterólogos otros dos genes de la vía de RdDM (chr106 y ago104) que habían mostrado indicios de expresión diferencial entre plantas apomícticas y sexuales en P. notatum en experimentos previos. Actualmente estamos caracterizando la secuencia completa y la expresión de estos genes en distintos estadíos de desarrollo.
Resúmenes
Caracterización de secuencias de metiltransferasasasociadas a la aposporía en Paspalum notatum
1 2 3 2 4 1Siena LA , Selva JP , Ortiz JPA , Echenique V , Leblanc O , Pessino SC
1Laboratorio de Biología MolecularFacultad de Ciencias Agrarias, Universidad Nacional de Rosario, Argentina2CERZOS-CONICET, Bahía Blanca, Argentina3IBONE-CONICET, Corrientes, Argentina4Institut de Recherche pour le Développement, Montpellier, [email protected]
Resumen
Ciencias Agronómicas - Revista XX - Año 12 - 2012


-75-
En trabajos previos, nuestro grupo identificó 65 genes diferencialmente expresados en genotipos sexuales y apomícticos de Paspalum notatum. Uno de estos genes (código experimental N46) resultó homólogo a MAP3Ks. Fue aislado del genotipo sexual y su secuencia mapeó in silico en el genoma de arroz en una región sinténica con la región genómica que controla la aposporía en P. notatum. El objetivo de nuestro trabajo fue caracterizar la secuencia y la expresión del gen candidato N46 y desarrollar una plataforma de transformación para modificar su expresión en órganos reproductivos de plantas apomícticas y sexuales. Se utilizó la técnica de RACE para aislar la secuencia codificante completa (1390 nucleótidos), que en los análisis de Blastx arrojó la mayor similitud con un gen de maíz con anotación MAP3K (E-value: 0.0, % ID: 96%). Los experimentos de hibridización in situ de tejidos reproductivos indicaron que la hebra sentido del transcripto se expresa en óvulos inmaduros de la planta apomíctica, y la antisentido en planta sexual. Los experimentos de PCR en tiempo real confirmaron una expresión diferencial en plantas apomícticas y sexuales en estadío meiótico. Para iniciar los experimentos de transformación, se desarrolló un sistema de cultivo de tejidos sobre el genotipo Q4117 (tetraploide apomíctico), ensayando tres tipos de explantos: embriones maduros, meristemas de vástagos y semillas maduras. Los porcentajes de regeneración obtenidos fueron similares en los tres casos (~45%), por lo que se eligió trabajar con semillas maduras. Los experimentos de transformación transiente fueron realizados utilizando un acelerador de partículas de tungsteno, presiones de helio comprimido de 600, 800 y 1100 psi, y vectores de transformación con genes de la vía de antocianinas o EGFP (green fluorescent protein), clonados bajo el promotor constitutivo act1 de arroz. Se establecieron curvas de selección con el herbicida glufosinato de amonio y el antibiótico kanamicina. Los experimentos de transformación estable se realizaron utilizando una mezcla de dos plásmidos en cantidades equimolares, uno de ellos con una construcción en horquilla del gen N46 (MAP3K) bajo el protomor act1 de arroz, junto con el selector bar (tolerancia a glufosinato de amonio), y el otro con GFP clonado bajo el promotor act1. El porcentaje de regeneración obtenido fue de 71 plantas regenerantes/600 callos bombardeados. La introducción de los transgenes fue analizada por PCR, obteniéndose los siguientes resultados: 23.3% gfp, 3,3% bar, 30% co-transformación (bar + gfp) y 26.7% escapes. Actualmente se están realizando experimentos de Southern Blot para confirmar la integración de los transgenes al genoma. Además los fenotipos de las inflorescencias de las plantas transgénicas están siendo analizadas y se encontró expresión de gfp en polen de plantas reportadas como gfp positivas según los experimentos de PCR.
Resúmenes
Transformación biolística de Paspalum notatum tetraploidecon un gen candidato para la apomixis
1 1 1 1,2 2 1 1Mancini M , Woitovich N , Permingeat H , Ortiz JPA , Quarin C. , Felitti S , Pessino S.
1Laboratorio de Biología Molecular,Facultad de Ciencias Agrarias, Universidad Nacional de Rosario,Parque Villarino, S2125ZAA Zavalla, Santa Fe, Argentina;2Instituto de Botánica del Nordeste (IBONE)-CONICET,Facultad de Ciencias Agrarias, UNNE,3400 Corrientes, [email protected]
Resumen
Ciencias Agronómicas - Revista XX - Año 12 - 2012


-77-
En los últimos años nuestro grupo de trabajo se ha dedicado a investigar la apomixis en el pasto llorón, Eragrostis curvula, especie que incluye citotipos de diferentes ploidías (2x a 8x). Los diploides son sexuales y poco frecuentes. Los poliploides se reproducen principalmente por apomixis obligada., del tipo diplosporía mitótica pseudógama. El desarrollo del saco embrionario es de tipo Eragrostis, donde el megasporocito sufre dos rondas de mitosis para formar un saco embrionario no reducido, tetranucleado, con una célula huevo, dos sinérgidas y un núcleo polar. La obtención de una serie euploide con diferentes ploidías y modos reproductivos (4x apo-2x sex -4x altamente sex, obtenida por duplicación con colchicina del diploide) en un mismo fondo genético, fue el punto de partida para el estudio de la apomixis en esta especie. Se analizó la estructura del genoma y los perfiles transcriptómicos de estas plantas así como los niveles de metilación utilizando la técnica de MSAP. Se secuenciaron 12.295 ESTs de inflorescencias y hojas y se obtuvieron 8.884 unigenes de los cuales 112 (1,26%) resultaron diferencialmente expresados por ploidía y/o modo reproductivo. Estos genes fueron mapeados in silico sobre los cromosomas de maíz, con especial foco en los cromosomas 6, 8 y 3, donde se informó la presencia de una región sinténica a la región de Tripsacum dactyloides asociada a la diplosporía. Las observaciones indicarían que la expresión de los genes localizados en esa región podrían estar afectados por ploidía y se encuentran silenciados en plantas apomícticas. Un cambio en el nivel de expresión de la apomixis observado en la planta colchiploide entre 2003 y 2007, desde vestigial a casi el 90%, nos llevó a estudiar el efecto del cambio de ploidía sobre el estado de metilación y la estructura genética de estas plantas en el tiempo. Se determinó que durante estos años se produjeron modificaciones genéticas y epigenéticas en la planta control (4x apo) y la colchiploide. Mientras las variaciones genéticas condujeron a la diversificación, la variación epigenética resultó en un incremento en la similitud. Procesos de metilación prevalecieron en la planta colchiploide en relación al control. La mayoría de las secuencias involucradas en estos cambios fueron similares a elementos transponibles (TEs) y/o pseudogenes. El análisis de previo de ESTs reveló la presencia de secuencias con similitud a varios elementos repetitivos en las genotecas de inflorescencias. Las familias más representada fueron los CACTA (transposones), no encontrándose diferencias entre plantas sexuales y apomícticas ylos Gypsy (retrotransposones), especialmente en la genoteca de la planta colchiploide. Estos resultados, junto con otras observaciones de plantas en condiciones de estrés por déficit de agua y cultivo de tejidos sugieren un control epigenético sobre la expresión de la sexualidad en plantas apomícticas de pasto llorón, pudiendo estar los retrotransposones, particularmente los Gypsy involucrados. Se puso a punto un sistema de transformación mediado por Agrocaterium tumefaciens a fin de validar, en el futuro cercano, los genes candidatos a afectar este modo reproductivo.
Resúmenes
Apomixis en Eragrostis curvula
1 2 1 1 2 1 1Echenique V , Pessino SC , Meier M , Selva JP , Ochogavía A , Zappacosta D , Romero JR1 1 1 1
Rodrigo JM , Garbus I , Terenti C , Díaz A
1Departamento de AgronomíaUniversidad Nacional del SurCERZOS (CONICET), CCT Bahía Blanca, Buenos Aires2Facultad de Ciencias Agrarias,Universidad Nacional de Rosario,Zavalla, [email protected]
Resumen
Ciencias Agronómicas - Revista XX - Año 12 - 2012


-79-
El pasto llorón es una gramínea apomíctica diplospórica. El carácter apomixis está genéticamente controlado y presenta una expresividad variable que puede verse afectada por factores ambientales que generan estrés genómico. El estrés se ha definido como el efecto producido por un factor ambiental externo que dista del óptimo y actúa sobre la planta generando una respuesta. Varios autores proponen que las plantas poseen mecanismos para responder a situaciones de estrés que estarían relacionados a reestructuraciones en su genoma, incluyendo cambios estructurales y activación de transposones. El objetivo de este trabajo fue evaluar el nivel de expresión de la apomixis en condiciones controladas durante el período de floración y ante condiciones de estrés. Para ello se colectaron periódicamente espiguillas de plantas del cultivar Tanganyika (2n=4x=40) y se analizaron los estadíos de desarrollo que comprendieron desde célula madre de la megáspora hasta saco embrionario octanucleado, de manera de identificar procesos sexuales o apomícticos. El análisis de 565 sacos permitió evidenciar estabilidad en la expresión de la apomixis durante el periodo de floración en condiciones control, con una sexualidad promedio del 1,4%. En plantas sometidas a situaciones de estrés por déficit hídrico durante tres meses previos al proceso de floración, el análisis de sacos mostró un incremento del 12% en la proporción de procesos sexuales respecto al control. Estos resultados indican que las plantas del cv. Tanganyika podrían incrementar la expresión de procesos sexuales cuando se enfrentan a un estrés hídrico durante un periodo prolongado. Sin embargo, las pruebas de progenie efectuadas en plántulas utilizando RAPDs evidenciaron porcentajes menores de sexualidad (0% en el control y 5% en los tratamientos de estrés hídrico) y escasos polimorfismos. La diferencia en los porcentajes de sexualidad entre el estudio de sacos y los test de progenie podría deberse a defectos en el desarrollo de los embriones o en la germinación de las semillas producto de procesos sexuales. Se decidió analizar el contenido endógeno de ácido abscísico (ABA) y evaluar su correlación con la expresión del modo reproductivo. Se analizaron muestras provenientes de hoja y panoja de dos cultivares tetraploides apomícticos, uno tetraploide sexual y dos diploides sexuales, en plantas control y bajo tratamiento de supresión de riego. En todos los casos las muestras de hoja mostraron una menor concentración de ABA que las de panoja. La mayor concentración de ABA se observó en el cultivar tetraploide sexual, que triplicó los valores de los otros cultivares. En panoja, se encontraron valores similares en ambos tetraploides apomícticos y menores y muy similares en los diploides sexuales. Los valores en las muestras bajo régimen de sequía aumentaron considerablemente, y se redujeron las diferencias relativas, es decir, desapareció la diferencia entre hoja/panoja y entre variedades. Actualmente se está realizando un nuevo ensayo para corroborar estos resultados en un mayor número de genotipos.
Resúmenes
Efectos del estrés abiótico sobre la expresiónde la apomixis en pasto llorón
1 1 2 1Rodrigo JM , Zappacosta D , Bonaventure G , Echenique V
1Laboratorio de Biotecnología Vegetal,Centro de Recursos Naturales de la Zona Semiárida (CERZOS),Departamento de Agronomìa,Universidad Nacional del Sur, CCT-Bahía Blanca, Argentina;2Department of Molecular Ecology,Max Planck Institute for Chemical Ecology,Hans-Knoell-Str. 8 D-07745 Jena, Alemania. [email protected]
Resumen
Ciencias Agronómicas - Revista XX - Año 12 - 2012


-81-
El pasto llorón (Eragrostis curvula) es una forrajera originaria de Sudáfrica, ampliamente distribuida en ambientes áridos y semiáridos de nuestro país y que presenta apomixis diplospórica con pseudogamia. El modo reproductivo se puede estudiar desde un punto de vista estructural (mapeo de la/s región/es genómica/s) o funcional (identificación y caracterización de genes y/o mecanismos epigenéticos que definen el carácter). Para la construcción de un mapa de ligamiento se requiere de una población segregante, es decir una población derivada de un cruzamiento y los parentales seleccionados deben diferir en el carácter de interés, que en este caso es la apomixis. En la mayoría de las plantas que se reproducen por apomixis gametofítica, el desarrollo partenogenético del embrión requiere de la fertilización de los núcleos polares, lo cual es denominado pseudogamia. Un hecho importante que se ha comprobado en las especies apomícticas de los géneros Brachiaria, Paspalum y Panicum, es que los genotipos sexuales son alógamos, con un alto grado de autoincompatibilidad, mientras que los apomícticos son autocompatibles. El objetivo del presente trabajo fue la obtención de un mapa genético y marcadores ligados a la apomixis en Eragrostis curvula. En el Laboratorio de Biotecnología Vegetal (Departamento de Agronomía-UNS, CERZOS-CONICET) se cuenta con una colección de materiales de pasto llorón con diferentes ploidías (desde 2x hasta 8x, con x=10). Los primeros cruzamientos hacia la obtención de la población de mapeo se realizaron entre una madre tetraploide (genotipo UNST1131, 2n=4X=40) con alta expresión de la sexualidad (evaluado en el año 2003) y un donante de polen de los cultivares tetraploides Morpa y Ermelo. Al evaluar las progenies obtenidas se observó una alta proporción de plantas clonales maternas (90%), lo que indicó una elevada expresión de la apomixis en el progenitor femenino al realizarse los cruzamientos (año 2007). Frente a esta situación, se utilizó como progenitor femenino a OTA-S (PI 574506 USDA, 2n=4X=40), que presentó 100% de sacos embrionarios sexuales. Como donante de polen se utilizaron plantas del cultivar tetraploide apomíctico Tanganyika (PI 234217 USDA). Al evaluar la progenie utilizando marcadores AFLPs, se encontró que, de las 310 plantas analizadas, sólo seis presentaban marcadores paternos, indicando la existencia de autofecundación. La disrupción en el sistema de autoincompatibilidad en el genotipo sexual podría deberse a la presencia del polen del donante apomíctico. Este fenómeno es conocido como efecto mentor y fue observado en otras especies apomícticas. Actualmente se están evaluando técnicas de emasculación (tratamientos químicos, térmicos y de castración) para superar la alta proporción de autofecundación y la utilización de otros parentales en los cruzamientos, como por ejemplo plantas híbridas sexuales provenientes de la F obtenida.1
Resúmenes
Poblaciones de mapeo en pasto llorón - Éxitos y fracasos
Zappacosta D, Meier M, Terenti C, Rodrigo JM, Echenique V
Lab. Biotecnología Vegetal (CERZOS),Departamento de Agronomìa (UNS), CCT-Bahía Blanca, [email protected]
Resumen
Ciencias Agronómicas - Revista XX - Año 12 - 2012


La Revista de Investigaciones de la Facultad de Ciencias Agrarias es una publicación bianual que acepta artículos y trabajos originales de investigación y experimentación según se detalla más abajo. Los trabajos enviados para su publicación deben ser inéditos, si bien pueden haber sido comunicados en reuniones científicas y técnicas. Las contribuciones serán enviadas al Editor Principal, quien se reserva el derecho de juzgar los trabajos y/o remitirlos a los árbitros, así como de introducir, con conocimiento de los autores, todos los cambios exigidos por las normas gramaticales y las necesidades de compaginación y edición. Los idiomas oficiales son el español, el portugués y el inglés.
La Revista incluye las secciones Artículos Originales (Trabajos de investigación, Notas breves, Resúmenes de Tesis), Artículos de Revisión, Notas del Editor y Cartas. En determinados números se incluirá la sección Visión conteniendo el enfoque de una personalidad destacada en un campo particular y la de Comentarios Bibliográficos con un análisis breve de publicaciones recientemente editadas y relacionadas con el enfoque de la Revista.
Los Artículos Originales -tanto teóricos como experimentales- deben estar caracterizados por su calidad científica. Los Trabajos completos no pueden ocupar más de 20 páginas, a simple espacio y separación entre párrafos, incluyendo Tablas y Figuras. Las Notas Breves no excederán de 10 hojas, también a simple espacio y separación entre párrafos, incluyendo Tablas y Figuras. En ambos casos, estarán organizados en secciones: Resumen, Palabras clave, Introducción, Materiales y Métodos, Resultados y Discusión, Conclusiones, Agradecimientos y Bibliografía. El Resumen no deberá superar las 250 palabras. Estará escrito en el idioma del artículo y se sumará otro en idioma inglés, precedidos por los correspondientes títulos, con sus palabras clave (3 separadas por guiones) en ambos idiomas, excluyendo las contenidas en el título. Si el idioma del artículo es el inglés, deberá contener un resumen en español. En el caso de artículos originales de las Ciencias Sociales, la sección Materiales y Métodos puede ser reemplazada por Metodología. Las secciones Resultados y Discusión podrán desarrollarse en conjunto.
Los Artículos de Revisión serán solicitados por el Editor principal o el Comité Editorial a personalidades con trayectoria en un campo del conocimiento en particular y deberán fundamentarse en una actualizada revisión bibliográfica.
La sección Notas del Editor contendrá una breve introducción del escenario del momento y presentará sucintamente el contenido de cada número. Las Cartas estarán referidas a comentarios de naturaleza editorial, relacionadas con los artículos publicados en la Revista. No excederán de 3 páginas, pudiendo incluir hasta 6 referencias y 1 Tabla o Figura.
En todos los casos, cada autor debe haber participado suficientemente en el trabajo como para asumir responsabilidad de su contenido. Cuando no corresponda ese criterio, serán mencionados en Agradecimientos si correspondiese esa sección. Cada autor contendrá una llamada en formato superíndice que remita a su lugar de trabajo y la dirección postal de la Institución. El correo electrónico corresponderá al del primer autor. Los autores irán en el orden de mérito surgido de su participación en el trabajo, a menos que haya responsabilidades equivalentes, en cuyo caso corresponderá agregar al final el vocablo latino ex-aequo.
Los vocablos latinos al igual que los nombres científicos deben ir en itálico incluyendo en el segundo caso la o las iniciales del nomenclador (Ej. Glycine max. L. Merr.) sólo en la primera cita (sin incluirlo en el título ni en los resúmenes). Los nombres de principios activos o drogas deben seguir las normas internacionales ISO ó BSI o bien la nomenclatura IUPAC. Se pueden utilizar los símbolos químicos en el texto. Las cifras se expresarán en números arábigos, excepto cuando aquellas inicien una oración o cuando se trate de una sola unidad. Las unidades serán las correspondientes al Sistema Internacional. Toda referencia o comentario adicional deberá realizarse con nota a pie de página.
En Materiales y Métodos se incluirán los detalles de las técnicas estadísticas utilizadas. En la Bibliografía sólo deberán incluirse las fuentes citadas en el texto, ordenadas alfabéticamente. Los trabajos de un mismo autor serán ordenados cronológicamente. Las referencias bibliográficas tendrán el siguiente formato:
KAPLUM M (1987) El comunicador popular. Buenos Aires. Editorial Humanitas.OGREN WL, RINNE RW (1973) Photosynthesis and seed metabolism. En: Caldwell, BE (Ed.) Soybeans: improvement, production and uses. Wisconsin USA. American Society of Agronomy, Inc. Publisher. p. 285-294.LEWIS RW (1988) Biology: a hypothetic-deductive science. The American Biology Teacher, 50:362-366.PEREZ JF (1999) Influencia del fuego sobre la composición del bosque septentrional. I Jornadas Nacionales de Zonas Áridas y Semiáridas. Santiago del Estero. p. 225-228. NIELSEN RL (1997) Stand establishment variability in corn. http://www.agry.purdue.edu/ext/pubs/AGRY-91-01_v5.pdf. Acceso: 15 de septiembre de 2003.
Se enviarán dos archivos digitales: uno en formato Word para el cuerpo del texto y las leyendas de Tablas y Figuras y otro en Excel para las Tablas y Figuras (véase más adelante). El archivo en formato Word deberá tener tamaño de hoja A4, margen izquierdo y derecho 3 cm, margen superior e inferior 2.5 cm, numeradas en la parte inferior (pie de página) centrada. Para todo el documento se utilizará interlineado simple, con separación de un espacio entre párrafos y tipo de letra Time New Roman. Para el Título: tamaño 12, mayúscula, Resumen y Summary: tamaño 10, Cuerpo del Artículo: tamaño 12.
a) Títulos que acompañan a las Tablas y Figuras:Deben ser suficientemente explicativos per se sin que sea necesario remitirse para su comprensión al cuerpo del artículo, incluyendo si es necesario el tipo de análisis estadístico y/o la significancia. Tipo de letra Arial, tamaño 10.
Normas para la presentación de manuscritos
-83-Ciencias Agronómicas - Revista XX - Año 12 - 2012 / 083 - 084

b) Tablas (no deben denominarse «Cuadros»):Serán construidas en Excel. Las cifras tendrán dos decimales como máximo. El tamaño no deberá superar las dimensiones de una página A4 con los márgenes indicados más arriba. El tipo de letra será Arial, estilo normal (sin negrita), tamaño máximo 8. La tabla deberá mostrar solo las líneas horizontales, sin verticales ni bordes exteriores. Los valores numéricos estarán alineados a la derecha y los alfanuméricos centrados. Los encabezados de columnas irán centrados horizontal y verticalmente en negrita. Se ingresarán los datos con 1º letra mayúscula (tipo oración). Se utilizará el punto como separadores de miles y la coma como separador de decimales.
c) Figuras (Gráficos):En formato Excel o cualquier otro formato compatible. Normalmente todos los programas estadísticos o de graficación científica (Statgraphics, SPSS, Sigma Plot) generan gráficos en formatos de fácil exportación y de gran compatibilidad. No deben poseer bordes externos y el tipo de letra debe ser Arial normal. No deben poseer título, solamente debe incluirse los correspondientes a los ejes.
d) Ilustraciones, imágenes y/o fotografías: insertas en el archivo formato Word, en formato JPG, TIFF o BMP de 300 dpi de resolución.
e) Ecuaciones: Las ecuaciones serán elaboradas con Editor de Ecuaciones. Si no están incluidas dentro de un párrafo, deberán colocarse alineadas a la izquierda, separadas del párrafo anterior y subsiguiente por un espacio. En caso de ser necesario deberán enumerarse correlativamente contra el margen derecho y entre corchetes, como se muestra en el siguiente ejemplo:
f) Citas textuales: si la cita no supera las dos o tres líneas, puede insertarse en el párrafo entre comillas inglesas (“ ”). Si es más extensa, se colocará en párrafo aparte sangrado, sin entrecomillar, con interlineado sencillo y tipografía tamaño 12. La supresión de una parte de la cita se indicará mediante puntos suspensivos encerrados en corchetes: […]. Asimismo, la inclusión de una segunda cita dentro de la primera se indicará entre comillas simples (' ').
g) Las citas bibliográficas deben incluirse en el texto de la siguiente manera:-Citas bibliográficas de un autor: (Zurada, 1992) cuando se trate de una cita textual o de una idea enunciada por el autor. Irá al final del párrafo. Si forma parte del texto: Krepper (1987). Si son dos autores en ambos casos el conector será “&”. Si son más de dos autores: (Landgrebe et al., 1989).-Si se cita más de un trabajo para un mismo autor y año, se utilizará las letras a, b, c después del año: (Huete & Myneni, 1996a, 1996b).-Las comunicaciones personales deberán ser citadas de la siguiente forma: (Ríos, comunicación personal) e indicar en las referencias bibliográficas: Apellido y nombre, año, lugar de trabajo y país, v.gr. RIOS, A. 2006. Sección Maquinaria Agrícola. INTA Manfredi. Argentina.
La publicación en la Revista de Investigaciones de la Facultad de Ciencias Agrarias es gratuita para los autores, siempre que el 2formato papel sea impresión en blanco y negro (escala de grises) y papel obra de 80 gr/m . La publicación de artículos completos,
secciones, gráficos y/o figuras a color tendrá un cargo adicional que deberá ser presupuestado específicamente por el personal técnico. Por cada artículo publicado, se remitirán dos ejemplares de la revista al primer autor.
Los artículos originales deberán ser remitidos a la siguiente dirección de correo electrónico: [email protected].
Normas para la presentación de manuscritos
∑=
+=n
i
ixy
1
)5( [1]
-84- Ciencias Agronómicas - Revista XX - Año 12 - 2012 / 083 - 084



















