
CENTRO DE INVESTIGACIÓN Y DE ESTUDIOS AVANZADOS … · Prof. Gustavo de los Santos Guzmán ......
139
See discussions, stats, and author profiles for this publication at: https://www.researchgate.net/publication/248702584 ALTERNATIVAS AGRO-BIOTECNOLÓGICAS PARA LA OPTIMIZACIÓN DE LA CADENA DE VALOR DEL MANGO (Mangifera indica L.) EN MÉXICO Thesis · November 2012 CITATIONS 0 READS 558 1 author: Sergio de los Santos-Villalobos Instituto Tecnológico de Sonora 38 PUBLICATIONS 107 CITATIONS SEE PROFILE All content following this page was uploaded by Sergio de los Santos-Villalobos on 06 June 2014. The user has requested enhancement of the downloaded file.
Transcript of CENTRO DE INVESTIGACIÓN Y DE ESTUDIOS AVANZADOS … · Prof. Gustavo de los Santos Guzmán ......
See discussions, stats, and author profiles for this publication at: https://www.researchgate.net/publication/248702584
ALTERNATIVAS AGRO-BIOTECNOLÓGICAS PARA LA OPTIMIZACIÓN DE LA
CADENA DE VALOR DEL MANGO (Mangifera indica L.) EN MÉXICO
Thesis · November 2012
CITATIONS
0READS
558
1 author:
Sergio de los Santos-Villalobos
Instituto Tecnológico de Sonora
38 PUBLICATIONS 107 CITATIONS
SEE PROFILE
All content following this page was uploaded by Sergio de los Santos-Villalobos on 06 June 2014.
The user has requested enhancement of the downloaded file.

CENTRO DE INVESTIGACIÓN Y DE ESTUDIOS AVANZADOS
DEL
INSTITUTO POLITÉCNICO NACIONAL
UNIDAD IRAPUATO
DEPARTAMENTO DE BIOTECNOLOGÍA Y BIOQUÍMICA
ALTERNATIVAS AGRO-BIOTECNOLÓGICAS PARA LA OPTIMIZACIÓN DE LA
CADENA DE VALOR DEL MANGO (Mangifera indica L.) EN MÉXICO
TESIS QUE PRESENTA
IBQ. SERGIO DE LOS SANTOS VILLALOBOS
PARA OBTENER EL GRADO DE
DOCTOR EN CIENCIAS
EN LA ESPECIALIDAD DE
BIOTECNOLOGÍA DE PLANTAS
DIRECTOR DE TESIS
DR. JUAN JOSÉ PEÑA CABRIALES
IRAPUATO, GUANAJUATO NOVIEMBRE, 2012

El presente trabajo fue realizado gracias al apoyo económico brindado por el
Consejo Nacional de Ciencia y Tecnología (CONACYT) durante 5 años, bajo la
beca: 10512; así como por el apoyo económico para la obtención de grado
recibido por el Centro de Investigación y de Estudios Avanzados del Instituto
Politécnico Nacional.

El presente trabajo fue realizado en el Laboratorio de Microbiología Ambiental,
perteneciente al Departamento de Biotecnología y Bioquímica del CINVESTAV
Unidad Irapuato, bajo la dirección del Dr. Juan José Peña Cabriales.
Laboratorio de Microbiología Ambiental

DEDICATORIAS
A mis padres:
Profa. Olga Leticia Villalobos López
Prof. Gustavo de los Santos Guzmán
Por sus consejos y valores, por la motivación constante, por todo el amor y
apoyo incondicional a través del tiempo, sin importar calificaciones o experimentos
fallidos. Por enseñarme, con hechos, que la perseverancia y constancia con el
máximo esfuerzo, son claves para salir adelante.
Por ser los sólidos pilares de todo lo que soy, tanto en mi educación académica,
como de la vida.
A mi hermana:
IBQ. Fátima de los Santos Villalobos
Por su apoyo en aspectos familiares, personales y profesionales. Los cuales
fueron una pieza importante para la consecución de este trabajo.
A mi novia:
cDr. Fannie Isela Parra Cota
Por su comprensión, solidaridad y amor incondicional, esenciales en el
desarrollo y conclusión de este trabajo. Por sus valiosas contribuciones científicas
y por hacerme ver que sólo los logros profesionales no conducen a la felicidad
total.

AGRADECIMIENTOS
El presente trabajo fue el resultado de un esfuerzo en conjunto de muchas
personas, a quienes les agradezco sinceramente su apoyo.
Dr. Juan José Peña Cabriales. Por aceptarme y hacerme sentir parte de su gran
equipo de trabajo desde el día uno. Por su apoyo, confianza y por compartir sus
experiencias tanto académicas como de la vida. Por su firme trabajo y compromiso
para el crecimiento personal y profesional de todos sus estudiantes. Por lo cual,
además de ser mi director de tesis, lo considero un amigo.
Dra. Doralinda A. Guzmán Ortiz. Por sus valiosos consejos y apoyo a nivel
personal y académico, los cuales fueron parte fundamental para el desarrollo de
este trabajo.
Comité de sinodales: Dra. Doralinda A. Guzmán Ortiz, Dr. John P. Délano Frier,
Dr. Miguel Ángel Gómez Lim, Dr. Prometeo Sánchez García y Dr. Stefan De
Folter. Por sus valiosas aportaciones académicas y por compartir sus
conocimientos, los cuales condujeron a la consecución de este trabajo.
M.C. José Antonio Vera Núñez y Prof. Simón Rodríguez Castellanos. Por
compartir sus conocimientos, su fuerte compromiso de trabajo y amistad.
Con especial agradecimiento para IBQ. Fátima de los Santos Villalobos, IBQ.
Guadalupe Coyolxauhqui Barrera Galicia e IBQ. Luis Ernesto Hernández
Rodríguez. Por su gran apoyo y solidaridad para la realización del presente
trabajo; así como, por su confianza y verdadera amistad.
M.C. Enrique Ramírez y M.C. Susana Gómez. Por su apoyo y convivencia
dentro y fuera de los laboratorios.

A los integrantes del Laboratorio de Microbiología Ambiental. Juan José Peña
Cabriales, José Antonio Vera, Simón Rodríguez, Josué Altamirano, Juan Ramiro
Pacheco, David Alfonso Camarena, Ediel Pérez, John Larsen, Milton Senen
Barcos, Guadalupe Coyolxauhqui Barrera, Luis Ernesto Hernández, Daniel Torres,
Juan Colli, Oscar Ponce, Brenda Sánchez Montesinos, Xicoténcatl Martínez Ruiz,
Brenda Itzel Medrano, Víctor Laguna, Alma Lidia Aguiñaga, Yahir Maldonado,
Verónica Díaz, Claudia González, Oswaldo Castillo, Luis Fernando García, Ana
Lizbeth, Diana Cristina García, Luisa Elena Ayala, Oscar Manuel González, Luis
León, Abraham García, Sergio Abraham Hernández, María Elisa Paillie y Juan
Manuel Duque. Por su constante apoyo para el desarrollo del presente trabajo.
A mis amigos y compañeros de CINVESTAV Unidad Irapuato. Fannie Isela
Parra Cota, Fátima de los Santos Villalobos, Alicia Rodríguez Sixtos Higuera,
Kruskaia Caltzontzin Fernández, Meri Kokkonen, Jazmín Sánchez, Jose Luis
Hernandez Flores, Karla Iveth Pérez, Corina Elizabeth Díaz, Maria de Jesus
Ortega, Enrique Ibarra Laclette, Laura Hernández, Sara Raya, Soledad
Palomares, Juan Pablo Jaime, Dora Elia Anguiano Jaime, Yolanda Rodríguez,
Aurora Verver, Beatriz Vilchis, Diana Baca, Sergio Baltazar, Daniel Baltazar, Alicia
Navarro, Paul Blanquel, Alicia Chagoya, Antonio Cisneros, Tiburcio Corpus, Victor
Estrada, Ramón García, Alicia Gómez, Armando Guerrero, Mireya Hernández,
Salvador Hernández, Alma Rosa Hernández, Francisco Luna, Paulina Martínez,
Juan Carlos Ochoa, Germán Ortega, Raúl Pérez, Antonio Ramírez, José Ramírez,
Martín Ramírez, Cesar Raya, Mauro Razo, María del Carmen Reyes, Ramón
Salas, Antonio Sandoval, Humberto Torres, Gerardo Valladolid, integrantes del
CECIG y equipos de futbol/ voleibol de los cuales formé parte. Por su solidaridad y
sincera amistad.
Dr. Francisco Villaseñor, Dr. Mario Miranda, Dra. Lourdes Mondragón, Prof.
Gustavo de los Santos, Prof. Amador Barrera e Ing. Patricia Bravo. Por sus
valiosas aportaciones, facilidades de los sitios de estudio y uso de equipos
determinantes en la realización de este trabajo.

Al Laboratorio de Fisiología de la Defensa en Plantas, Laboratorio de
Micotoxinas, Laboratorio de Bioinsecticidas, Laboratorio de Química de Productos
Naturales, Laboratorio de Fitobioquímica, Laboratorio de Bioquímica Ecológica,
Laboratorio de Biotecnología de Alimentos, Laboratorio de Bioquímica y Biología
Molecular de Proteínas, Laboratorio de Bacteriología Molecular, Laboratorio de
Plantas Tropicales y Salud Humana, Laboratorio de Regulación
Postranscripcional, Laboratorio de Ecología de Plantas, Laboratorio de
Bacteriología Molecular, Laboratorio de Interacciones Microbianas, Laboratorio de
Virología, Laboratorio de Desarrollo y Diferenciación de Hongos, Laboratorio de
Marcadores Moleculares y Genética Molecular, Laboratorio de Evolución de la
Diveridad del Metabolismo, Laboratorio de Genómica Funcional del Desarrollo de
Plantas, Laboratorio de Expresión Génica y Desarrollo en Hongos. Por poner a la
disposición reactivos y equipos para el desarrollo de experimentos.

Los resultados de esta investigación fueron aceptados para su publicación en
los siguientes artículos:
de los Santos-Villalobos, S., de-Folter, S., Délano-Frier, J.P., Gómez-Lim, M.A.,
Guzmán-Ortiz, D.A., Sánchez-García, P., Peña-Cabriales, J.J. 2011. Critical aspects on
the integral management of mango: flowering, anthracnose and industrial waste. Revista
Méxicana de Ciencias Agrícolas. 2(2):221-234.
de los Santos-Villalobos, S., Barrera-Galicia, G.C., Miranda-Salcedo, M.A., Peña-
Cabriales, J.J. 2012. Burkholderia cepacia XXVI siderophore with biocontrol capacity
against Colletotrichum gloeosporioides. World Journal of Microbiology and Biotechnology.
28(8):2615-2623.
de los Santos-Villalobos, S., Hernández Rodríguez, L.E., Villaseñor-Ortega, F., Peña-
Cabriales, J.J. 2012. Production of Trichoderma asperellum T8a spores by a "home-made"
solid-state fermentation of mango industrial wastes. Bioresources 7(4):4938-4951.
de los Santos-Villalobos, S., Parra-Cota, F.I., de-Folter, S., Peña-Cabriales, J.J. 2012.
Primers to amplify Flowering Locus T (FT) transcript in mango (Mangifera indica) and their
potential use in other angiosperms. Plant Omics Journal. 5(5):453-457.
de los Santos-Villalobos, S., Guzmán-Ortiz, D.A., Gómez-Lim, M.A., Délano-Frier,
J.P., de-Folter, S., Sánchez-García, P., Peña-Cabriales, J.J. 2012. Potential use of
Trichoderma asperellum (Samuels, Liechfeldt et Nirenberg) T8a as a biological control
agent against anthracnose in mango (Mangifera indica L.). Biological Control. In press.
de los Santos-Villalobos, S., de-Folter, S., Délano-Frier, J.P., Gómez-Lim, M.A.,
Guzmán-Ortiz, D. A., Peña-Cabriales, J.J. Growth promotion and flowering induction in
mango by Burkholderia and Rhizobium inoculation: morphometric, biochemical and
molecular events. Journal of Plant Growth Regulation. Enviado.

CONTENIDO
I. RESUMEN ........................................................................................................................... 1
I.1. Abstract .............................................................................................................................. 2
II. INTRODUCCIÓN ................................................................................................................. 3
II.1. El mango ........................................................................................................................... 3
II.1.1. Origen y distribución ................................................................................................. 3
II.1.2. Descripción botánica ................................................................................................ 4
II.2. Importancia económica de la producción de mango .................................................... 6
II.2.1. Nivel mundial ............................................................................................................. 6
II.2.2. Nivel nacional ............................................................................................................ 8
II.3. Cadena de valor del mango en México ....................................................................... 10
II.3.1. La producción .......................................................................................................... 11
II.3.1.1.- El evento de floración......................................................................................... 11
II.3.1.2. Control de enfermedades ................................................................................... 16
II.3.2. La industrialización ................................................................................................. 20
II.3.2.1. Aprovechamiento de residuos industriales ........................................................ 20
III. JUSTIFICACIÓN ............................................................................................................... 23
IV. HIPÓTESIS ........................................................................................................................ 24
V. OBJETIVOS....................................................................................................................... 25
V.1. Generales ....................................................................................................................... 25
V.2. Específicos ..................................................................................................................... 25
VI. ESTRATEGIA EXPERIMENTAL ..................................................................................... 28
VII. MATERIALES Y MÉTODOS............................................................................................ 29
FASE I. Caracterización nutrimental y microbiológica de los sitios de estudio y
metabólica/molecular de los microorganismos aislados (promotores de crecimiento
vegetal, agentes de control biológico y el agente causal de la antracnosis). ...................... 29
VII.1. Selección de la variedad de mango e identificación de los sitios de estudio......... 29
VII.2. Procedimiento de muestreo........................................................................................ 29
VII.3. Análisis físico-químico y nutrimental de los sitios de estudio .................................. 30
VII.4. Aislamiento e identificación de los microorganismos de interés en los sitios de
estudio .................................................................................................................................... 30
VII.5. Caracterización metabólica de los microorganismos aislados ................................ 35

VII.6. Caracterización molecular de los microorganismos promisorios ............................ 41
VII.7. Microscopía electrónica de barrido de las cepas promotoras de crecimiento
vegetal y el agente de control biológico .............................................................................. 42
FASE II. Producción del agente de control biológico contra C. gloeosporioides usando los
residuos de industrialización del mango, bajo un sistema adoptable in situ por los
productores de mango. ............................................................................................................. 42
VII.8. Identificación de la fuente y caracterización proximal de los residuos de la
industrialización del mango .................................................................................................. 42
VII.9. Diseño de un sistema de fermentación para la producción del agente de control
biológico, seleccionado en la fase I, contra C. gloeosporioides........................................ 43
FASE III. Inoculación de microorganismos promotores de crecimiento vegetal a árboles de
mango, estudiando su impacto, así como el de las señales exógenas y endógenas, sobre
el evento de floración. ............................................................................................................... 45
VII.10. Estudio del ciclo fenológico de árboles de mango creciendo en invernadero ..... 45
VII.11. Diseño de un manejo de inoculación microbiana (seleccionadas en la Fase I)
para árboles de mango en invernadero .............................................................................. 45
VII.12. Cuantificación de parámetros morfométricos, bioquímicos y moleculares
involucrados en el evento de floración de árboles de mango bajo el manejo de la
inoculación microbiana ......................................................................................................... 46
VII.13. Análisis estadístico .................................................................................................... 49
VIII. RESULTADOS Y DISCUSIÓN ........................................................................................ 50
FASE I. Caracterización nutrimental y microbiológica de los sitios de estudio y
metabólica/molecular de los microorganismos aislados (promotores de crecimiento
vegetal, agentes de control biológico y el agente causal de la antracnosis). ...................... 50
VIII.1. Selección de la variedad de mango e identificación de los sitios de estudio ....... 50
VIII.2. Análisis físico-químico y nutrimental de los sitios de estudio ................................. 51
VIII.3. Aislamiento de microorganismos de interés en los sitios de estudio y su
caracterización metabólica - molecular ............................................................................... 54
FASE II. Producción del agente de control biológico contra C. gloeosporioides usando los
residuos de industrialización del mango, bajo un sistema adoptable in situ por los
productores de mango. ............................................................................................................. 70
VIII.4. Identificación de la fuente y caracterización proximal de los residuos de la
industrialización del mango .................................................................................................. 70
VIII.5. Diseño de un sistema de fermentación para la producción del agente de control
biológico, seleccionado en la fase I, contra C. gloeosporioides........................................ 74

FASE III. Inoculación de microorganismos promotores de crecimiento vegetal a árboles de
mango, estudiando su impacto, así como el de las señales exógenas y endógenas, sobre
el evento de floración. ............................................................................................................... 80
VIII.6. Manejo de la inoculación microbiana en árboles de mango creciendo en el
invernadero. ........................................................................................................................... 81
VIII.7. Cuantificación de parámetros morfométricos, bioquímicos y moleculares
involucrados en el evento de floración de árboles de mango bajo el manejo de la
inoculación microbiana ......................................................................................................... 83
IX. CONCLUSIONES .............................................................................................................. 96
X. PERSPECTIVAS ............................................................................................................... 98
XI. BIBLIOGRAFÍA ................................................................................................................. 99

ÍNDICE DE TABLAS
Tabla VIII.1. Características físico-químicas y principales nutrientes del suelo en los
diferentes sitios de estudios. ....................................................................................... 52
Tabla VIII.2. Contenido de nitrógeno y fósforo en hojas y frutos de árboles de mango
plantados en los sitios de estudio. ............................................................................... 53
Tabla VIII.3. Caracterización metabólica (actividades promotoras de crecimiento vegetal)
de las cepas bacterianas aisladas de suelo, hojas y frutos de árboles de mango en las
huertas localizadas en los sitios de estudio. ................................................................ 56
Tabla VIII.4. Grados de cobertura de los aislados de Trichoderma contra C.
gloeosporioides ATCC MYA 456 utilizando la técnica de confrontación in vitro. .......... 62
Tabla VIII.5. Capacidad de control biológico, caracterización metabólica (actividades
promotoras de crecimiento vegetal) y molecular de los aislados de Trichoderma
asociados a árboles de mango (suelo, hojas y frutos) ubicados en los sitios de estudio.
.................................................................................................................................... 63
Tabla VIII.6. Preparación de los extractos enzimáticos crudos producidos por Trichoderma
asperellum T8a para su evaluación sobre el control biológico de C. gloeosporioides
ATCC MYA 456. .......................................................................................................... 65
Tabla VIII.7. Caracterización proximal de los residuos provenientes de la industrialización
de mango “Tommy Atkins” (exocarpio y endocarpio). .................................................. 72
Tabla VIII.8. Producción de esporas y actividad celulolítica de T. asperellum T8a
creciendo bajo una fermentación en estado sólido, bajo diferentes relaciones C: N,
después de 6 días de incubación y un contenido de humedad del 88%. ..................... 75
Tabla VIII.9. Efecto de diferentes niveles de humedad sobre la producción de esporas de
T. asperellum T8a (relación C: N fijada en 26 y 6 días después de la inoculación) ..... 77
Tabla VIII.10. Efecto del tiempo de cosecha en la producción de esporas de T. asperellum
T8a (relación C: N fijada en 26 y un contenido de humedad del 78%). ........................ 78
Tabla VIII.11. Características físico-químicas del suelo utilizado para el crecimiento de los
árboles de mango en invernadero. .............................................................................. 81
Tabla VIII.12. Promoción del crecimiento y la inducción floral en el árbol de mango por el
manejo de la inoculación microbiana (B. caribensis XV y Rhizobium sp XXV) específica
a cada ciclo fenológico. ............................................................................................... 84
Tabla VIII.13. Secuencias de aminoácidos y de nucleótidos de las regiones altamente
conservadas (oligonucleótidos) identificadas en el alineamiento de las secuencias de
aminoácidos de la proteína FT. ................................................................................... 91

ÍNDICE DE FIGURAS
Figura II.1. Distribución pantropical del cultivo de mango. En la cual se observa sus zonas
de cultivo, las cuales están principalmente limitadas a las regiones comprendidas entre
el Trópico de Cáncer y de Capricornio [líneas punteadas arriba y debajo del Ecuador
(0o), respectivamente]. ............................................................................................................ 4
Figura II.2. Principales países productores de mango a nivel mundial. México se ubica en
el 4to. Lugar con una producción aproximada de 2 x 106 t. ................................................ 7
Figura II.3. Principales países exportadores de mango fresco a nivel mundial y sus
ingresos económicos generados por dicha comercialización. $: Valor de exportación en
millones de dólares. ................................................................................................................ 8
Figura II.4. Principales estados productores de mango en la República Mexicana y su
contribución (%) a la producción nacional. ........................................................................... 9
Figura II.5 Ingresos económicos, en millones de dólares, de EUA y México por la
exportación de mango fresco y sus productos industrializados. ....................................... 10
Figura II.6. Cadena de valor del mango en México y los puntos críticos que la
desestabilizan. ....................................................................................................................... 11
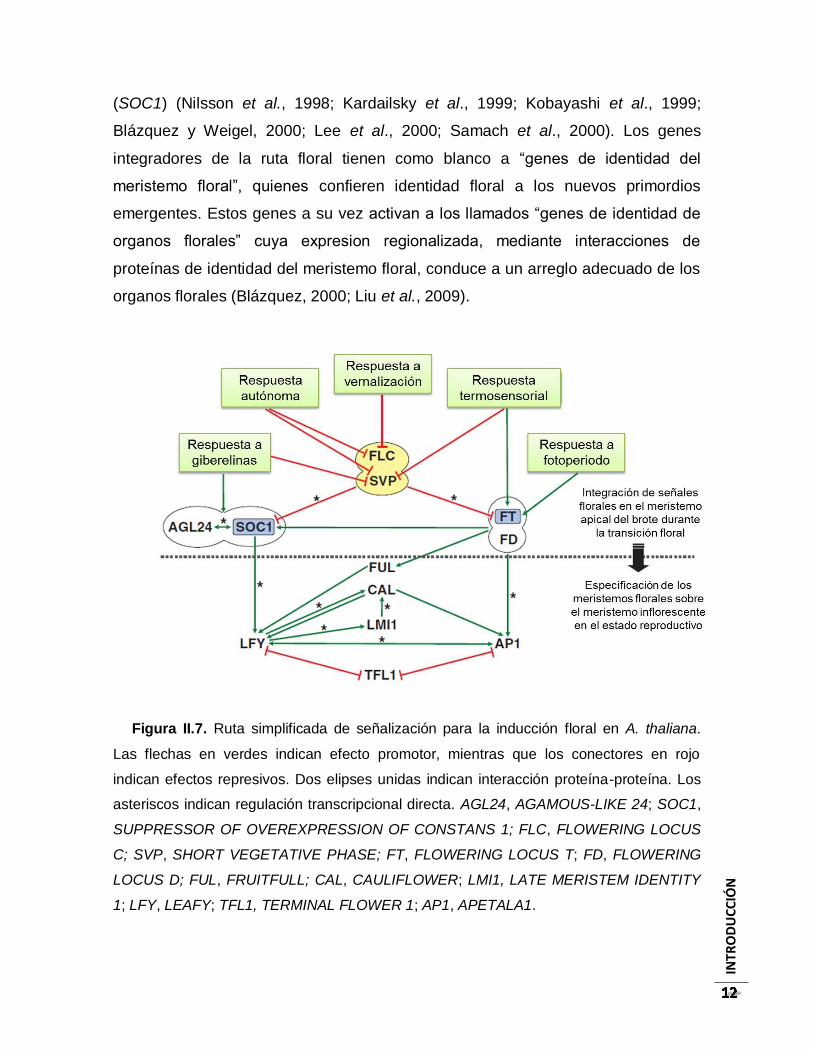
Figura II.7. Ruta simplificada de señalización para la inducción floral en A. thaliana. Las
flechas en verdes indican efecto promotor, mientras que los conectores en rojo indican
efectos represivos. Dos elipses unidas indican interacción proteína-proteína. Los
asteriscos indican regulación transcripcional directa. AGL24, AGAMOUS-LIKE 24;
SOC1, SUPPRESSOR OF OVEREXPRESSION OF CONSTANS 1; FLC, FLOWERING
LOCUS C; SVP, SHORT VEGETATIVE PHASE; FT, FLOWERING LOCUS T; FD,
FLOWERING LOCUS D; FUL, FRUITFULL; CAL, CAULIFLOWER; LMI1, LATE
MERISTEM IDENTITY 1; LFY, LEAFY; TFL1, TERMINAL FLOWER 1; AP1,
APETALA1. ............................................................................................................................ 12
Figura II.8. Modelo teórico de las etapas involucradas en el evento de floración en mango,
basado en Davenport (2007). Líneas continuas indican efecto promotor, mientras que,
líneas punteadas indican efecto represor, líneas sin cabeza de flecha indican efecto sin
ser determinado con certeza. ............................................................................................... 15
Figura II.9. Fases de colonización de C. gloeosporioides en el fruto del árbol de mango.17
Figura II.10. Síntomas de la antracnosis (causada por C. gloeosporioides) en diferentes
órganos del árbol de mango. ................................................................................................ 18
Figura VIII.1. Manejo de las malezas crecidas en las épocas de lluvias en los sitios de
estudio. Las malezas en las huertas de mango localizadas en Apatzingán, Michoacán
(19 ° 05'00 "N y 102 ° 21'00" W) y Tapachula, Chiapas (14° 56'00 "N y 092 ° 16'00" W)
son incorporadas naturalmente al suelo, mientras que aquellas crecidas en la huerta de
mango en Chahuites, Oaxaca (16° 15'00 "N y 094 ° 14'00" W) son removidas del
sistema. .................................................................................................................................. 51
Figura VIII.2. Curva de correlación entre el nitrógeno foliar y la producción relativa de
mango (%)………………………………………………………………………………………54
Figura VIII.3. Relaciones filogenéticas de las BPCV obtenidas en este estudio, inferidas
por el análisis del gen 16S RNAr mediante el método de neighbor-joining. Streptomyces
sp.-EF157833 fue utilizado como el grupo externo. Los números mostrados sobre las

ramas indican el porcentaje de reensamble de 1000 bootstrap (solamente son
mostrados aquellos valores > 50). Los colores indican el origen de los aislados, azul:
Chahuites y rojo: Apatzingán. .............................................................................................. 55
Figura VIII.4. Control biológico de Burkholderia cepacia XXVI contra C. gloeosporioides.
[a) inoculación de C. gloeosporioides (control negativo) y b) co-inoculación del patógeno
estudiado y B. cepacia XXVI (flechas), utilizando PDA como medio de cultivo], [c)
Inoculación de la cepa XXVI sobre una caja Petri conteniendo el medio CAS- agar (el
halo de coloración naranja evidencia la producción de sideróforo por esta cepa) y d) co-
inoculación de C. gloeosporioides y B. cepacia XXVI sobre cajas Petri conteniendo
CAS-agar]. La línea punteada indica el crecimiento radial del patógeno esperado en
ausencia de B. cepacia XXVI, [e) inoculación de C. gloeosporioides (control negativo) y
f) inoculación de C. gloeosporioides in cajas Petri conteniendo PDA suplementado con
0.64 µg ml-1 del sideróforo producido por B. cepacia XXVI]. ............................................. 58
Figura VIII.5. Relaciones filogenéticas de los aislados de Trichoderma obtenidos en este
estudio, inferidas por el análisis del ITS mediante el método de neighbor-joining.
Colletotrichum gloeosporioides-EU520114 fue utilizado como el grupo externo. Los
números mostrados sobre las ramas indican el porcentaje de reensamble de 1000
bootstrap (solamente son mostrados aquellos valores > 50). Los colores indican el
origen de los aislados, azul: Chahuites, rojo: Apatzingán y verde: Tapachula................ 61
Figura VIII.6. Observación microscópica de células viables [las mitocondrias, indicadas
por flechas, redujeron activamente el 2, 3, 5-trifeniltetrazolio (incoloro) a 1, 3, 5-
trifenilformazán (color rojo)] y lisis celular, indicada con flechas punteadas, de C.
gloeosporioides ATCC MYA 456 después de su incubación con diferentes actividades
enzimáticas producidas por T. asperellum T8a. Los tratamientos evaluados fueron
(Tabla VIII.6): a) 2, b) 4 c) 5, y d) 6. .............................................................................. 66
Figura VIII.7. Características macroscópicas y moleculares de los aislados de C.
gloeosporioides obtenidos en este trabajo, observando al menos dos cepas. a) C.
gloeosporioides XIV (las características macroscópicas fueron similares a las cepas X,
XI y XII); b) C. gloeosporioides VI (su característica microscópica fue diferentes a las
antes mencionadas); c) Relaciones filogenéticas de los aislados de C. gloeosporioides,
inferidas por el análisis del ITS mediante el método de neighbor-joining. Trichoderma
atroviride-JF502439 fue utilizado como el grupo externo. Los números mostrados sobre
las ramas indican el porcentaje de reensamble de 1000 bootstrap (solamente son
mostrados aquellos valores > 50). Los colores indican el origen de los aislados, azul:
Chahuites………………………………………………………………………………………..67
Figura VIII.8. Ensayo de patogenicidad de las cepas de C. gloeosporioides obtenidas en
este trabajo. a) Control negativo (agua), b) Inoculación de C. gloeosporioides XV,
observando lesiones necróticas típicas por la infección de este patógeno, causando la
antracnosis (flechas blancas). .............................................................................................. 68
Figura VIII.9. Severidad del daño antracnosis (indicado por flechas blancas) en un
tratamiento con 20% de frutos enfermos. a) Control (no inoculado con el patógeno); b)
Inoculación de C. gloeosporioides VI; c) Inoculación de C. gloeosporioides VI y T.
asperellum T8a)..................................................................................................................... 69

Figura VIII.10. Microscopias electrónicas de barrido de las cepas promotoras de
crecimiento vegetal y agente de control biológico utilizadas en la FASE II y FASE III,
respectivamente. a) Trichoderma asperellum T8a (1,100 x), b) Burkholderia caribensis
XV (45, 000 x), c) Rhizobium sp. XXV (43, 000 x). ............................................................ 70
Figura VIII.11. Actividad celulolítica de T. asperellum T8a crecimiento en medio líquido
con la celulosa añadida (línea continua) o residuos industriales de mango (línea
punteada) adicionados como única fuente de carbono. Una unidad de actividad (U) se
definió como la cantidad de enzima que libera 1 µmol equivalente glucosa o GlcNAc por
minuto bajo las condiciones de ensayo especificadas. ..................................................... 73
Figura VIII.12. Trichoderma asperellum T8a creciendo bajo diferentes relaciones de C: N
en una fermentación en estado sólido utilizando los residuos industriales de mango
como sustrato, seis días después de la inoculación y contenido de humedad del 88%.
Relaciones C: N a) 6, b) 14, c) 26 y d) 34. .......................................................................... 76
Figura VIII.13. Micrografías electrónicas de barrido de T. asperellum T8a después de 8
días de cultivo sobre: a) los residuos industriales de mango (r: residuos industriales de
mango, m: micelio de T. asperellum T8a y e: esporas de T. asperellum T8a) y b)
espuma de poliuretano (m1: micelio de T. asperellum T8a, m2: micelio de T. asperellum
T8a en los poros de la espuma de poliuretano, y e: espuma de poliuretano como matriz
de soporte inerte), en fermentación en estado sólido (relación C: N de 26 y contenido de
humedad 78%). ..................................................................................................................... 79
Figura VIII.14. Manejo de la inoculación microbiana (rectángulo gris) en árboles de mango
creciendo en el invernadero, de acuerdo con el ciclo fenológico de estos árboles y las
características metabólicas de cepas de bacterias seleccionadas. V: crecimiento
vegetativo, D: dormancia, F: floración y Fr: fructificación. ................................................. 82
Figura VIII.15. Niveles de sacarosa, glucosa y fructosa en las hojas de los árboles de
mango bajo inoculación microbiana o tratamiento sin inocular, cuantificados cada
semana a partir del 18 de diciembre del 2010 al 21 de enero del 2011. Las barras de
error no son visibles cuando éstas son más pequeñas que el tamaño del símbolo. ...... 88
Figura VIII.16. Alineamiento, por Clustal X, de 41 secuencias de aminoácidos de la
proteína FT (pertenecientes a 15 especies de angiospermas). Las regiones altamente
conservadas identificadas (FTf1, FTf2 y FTr2) fueron traducidos inversamente in silico
para la obtención de la secuencia de nucleótidos de las oligonucleótidos para la
amplificación de un fragmento del transcripto del gen FT. ................................................ 90
Figura VIII.17. Niveles de expresión de FT en las hojas de mango cuantificado cada
semana desde el 18 de diciembre de 2010 hasta el 21 de enero del 2011. Los datos se
muestran como la expresión relativa de FT, en ambos tratamientos, normalizado a ACT
o expresión relativa de FT normalizada al tratamiento no inoculado. Las barras de error
no son visibles cuando son más pequeñas que el tamaño del símbolo. ......................... 93
Figura VIII.18. Valores de temperatura, iluminación y humedad relativa en el invernadero,
registradas cada treinta minutos durante siete meses. El rectángulo amarillo indica el
período de inducción floral. Círculos verdes: crecimiento vegetativo; círculos azules:
crecimiento mixto, círculos naranja: crecimiento floral. ..................................................... 95

RES
UM
EN
I. RESUMEN
A nivel mundial, México se ubica como un participante importante en la
producción y comercio de mango fresco, destacando como productor (2 × 106 t
año-1) y exportador (2 × 105 t año-1), e importador de cantidades mínimas. La
actividad económica en torno al mango, está integrada por un conjunto de etapas
que van desde la producción del fruto hasta su consumo, la cual se le ha
denominado cadena de valor del mango. Con base en el conocimiento científico y
empírico sobre este cultivo, los puntos críticos de mayor impacto negativo que
hemos detectado sobre esta cadena en México son: el evento de floración, el
control de la antracnosis y el aprovechamiento de sus residuos industriales.
El objetivo central de este proyecto fue ofrecer alternativas agro-biotecnológicas
que permitan el manejo integral y sustentable del cultivo del mango. Dichas
alternativas se resumen a continuación:
El evento de floración. Se desarrolló un manejo de la inoculación microbiana
como alternativa para la inducción floral del mango. La inoculación, en ciclos
fenológicos determinantes, de dos cepas bacterianas aisladas de este cultivo en
campo condujo a una inducción tanto de la promoción del crecimiento vegetal,
como de la floración en mango (a nivel morfométrico, bioquímico y molecular),
como resultado de las características metabólicas de estas cepas.
El control de la antracnosis. Se identificaron y caracterizaron dos agentes
como una alternativa sustentable para el control in vitro e in vivo de C.
gloeosporioides (agente causal de la antracnosis), mediante la producción de
sideróforo (cepa bacteriana) y la actividad de enzimas líticas (cepa fúngica).
El aprovechamiento de los residuos industriales del mango. Se diseñó un
sistema de fermentación en estado sólido para la producción de esporas de la
cepa fúngica con capacidad de controlar la antracnosis, utilizando los residuos
industriales del mango, mediante la optimización de parámetros determinantes
para su adopción in situ, como son la relación C: N, contenido de humedad y el
tiempo para la cosecha de las esporas producidas. Este sistema mantuvo las
características metabólicas de dicha cepa.

AB
STR
AC
T
I.1. Abstract
Globally, Mexico ranks as an important participant in the production and trade of
fresh mango. It stands out as a major producer (2 × 106 t year-1) and exporter (2 ×
105 t year-1) of this fruit, with only a minimal percentage of its internal consumption
being covered by imported fruit. The economic activity in relation to the mango
consists of a set of stages from production to consumption of that fruit, which has
been called the mango value chain. Based on the scientific and empirical
knowledge on this fruit crop, we detected some critical points, which impact
negatively the economy of growers: the flowering event, the control of anthracnose
and the use of mango industrial wastes.
The aim of this work was to develop agro-biotechnological alternatives that allow
integral and sustainable management of mango production. Such alternatives are
summarized below:
The flowering event. We developed a microbial inoculation management as an
alternative to induce mango flowering. The inoculation, in determinant phenological
stages, of two bacterial strains isolated from mango orchards led to the induction of
both mango growth promotion and flowering (morphometric, biochemical and
molecular levels), as a result of metabolic characteristics of these strains.
The control of anthracnose. We identified and characterized two agents as
sustainable alternatives to control C. gloeosporioides (causal agent of
anthracnose) in vitro and in vivo, through siderophore production (bacterial strain)
and the lytic enzyme activity (fungal strain).
The use of mango industrial wastes. We designed a system to produce
spores of the fungal strain presetting biological control against C. gloeosporioides.
This system was a solid-state fermentation, using mango industrial wastes as
substrate and optimizing determinant parameters for its adoption in situ, such as
the C: N, moisture content and harvest time of spores produced. This system
remained the metabolic characteristics of this strain.

INTR
OD
UC
CIÓ
N
II. INTRODUCCIÓN
II.1. El mango
El mango (Mangifera indica), miembro de la familia Anacardiaceae
(http://plants.usda.gov), es una de las frutas tropicales más populares y
económicamente importantes alrededor del mundo. Esto es debido a sus altas
propiedades nutricionales, entre las que destacan: el contenido de carbohidratos,
fibra, proteínas, vitaminas y minerales (Bangerth y Carle, 2002). Por lo cual, se le
ha denominado “el Rey de las frutas” (Sareen y Shah, 2011).
II.1.1. Origen y distribución
El mango se ha cultivado desde tiempos prehistóricos, hace más de 6,000 años
a.C. (Galán-Saúco, 2009), en el sur y sureste de Asia. Sin embargo, actualmente
se desconoce su punto de origen exacto. Estudios morfológicos, fitogeográficos,
citológicos, anatómicos y de polen sugieren que el género Mangifera tuvo su
origen en los bosques montañosos bajos del este de la India, Bangladesh y
Myanmar, entre los 16° y 28° de latitud Norte (Opeke, 1982; Bodner y Gereau,
1988).
Bailey (1941) propone que la distribución natural del mango incluye a los cerros
boscosos del centro y suroeste de la India, Tailandia, Laos, Vietnam, Kampuchea
y la península Malaya.
Este cultivo fue introducido a México y Brasil antes del final del siglo XVII, y de
Brasil a las Indias Occidentales durante el siglo XVIII (Bailey, 1941). Desde
entonces, el mango ha sido cultivado y naturalizado tan extensamente que su
distribución se puede considerar como pantropical (Galán-Saúco, 1999), debido a
su área de ocurrencia geográfica en regiones tropicales de todos los continentes
(Fig. II.1).

INTR
OD
UC
CIÓ
N
Figura II.1. Distribución pantropical del cultivo de mango. En la cual se observa sus
zonas de cultivo, las cuales están principalmente limitadas a las regiones comprendidas
entre el Trópico de Cáncer y de Capricornio [líneas punteadas arriba y debajo del
Ecuador (0o), respectivamente].
II.1.2. Descripción botánica
Raíz. Las raíces del árbol de mango son pivotantes, alcanzando una
profundidad de 6 m y una extensión horizontal de hasta los 8 m del tronco. El
mayor porcentaje de las raíces absorbentes (pelos radicales) se encuentra
concentrado a 1.20 m de profundidad, con una distribución lateral radial de 1.50 m
(Avilán et al., 1979).
Tronco. El árbol de mango puede alcanzar una altura de 10 a 30 m. El tronco
generalmente es recto, cilíndrico, de 0.75 a 1 m de diámetro, cuya corteza es
gruesa y de color gris-café, presentando grietas longitudinales o surcos reticulados
poco profundos (Chávez et al., 2001).

INTR
OD
UC
CIÓ
N
Copa. Estructura densa y ampliamente oval o globular. Las ramificaciones son
gruesas y robustas, frecuentemente con grupos alternos de entrenudos largos y
cortos que corresponden al principio y final de cada crecimiento sucesivo (Chávez
et al., 2001).
Hojas. Son alternas, con una longitud de 12.5 a 37.5 cm y una anchura de 2.5 a
12.5 cm (Crane y Balerdi, 1991), espaciadas irregularmente a lo largo de las
ramificaciones, con una longitud del peciolo entre 1.5 a 12 cm, el cual se
encuentra hinchado en la base, liso en ambas superficies, de color verde-oscuro
por arriba y verde-amarillento por abajo, presentando un ápice acuminado. La
filotaxia es de 3/8, pero en las partes terminales las hojas se encuentran
acumuladas pareciendo verticilos (Chávez et al., 2001). Las hojas tienen
nervaduras visiblemente reticuladas, presentando entre 12 a 30 pares laterales
prominentes, mientras que una de ellas es robusta y conspicua. El color de las
hojas jóvenes es violeta-rojizo, posteriormente adquieren un color verde-oscuro.
Inflorescencia. También llamadas panículas, son muy ramificadas y
terminales, de aspecto piramidal, de 6 a 40 cm de largo y de 3 a 25 cm de
diámetro. Las raquis son de color rosa o morado, algunas veces verde-
amarillentas, redondeadas y densamente pubescentes. Las brácteas miden de 0.3
a 0.5 cm de largo, son oblongas-lanceoladas u ovadas–oblongas, intensamente
pubescentes, las cuales senescen en un corto periodo de tiempo (Crane y Balerdi,
1991).
Flores. Las flores son de dos tipos, hermafroditas y masculinas, estando ambas
presentes en la misma panícula (predominando las últimas). Éstas son de color
verde-amarillento, de 0.2 a 0.4 cm de largo y de 0.5 a 0.7 cm de diámetro. Los
sépalos son libres, caedizos, ovados u ovados-oblongos, un tanto agudos u
obtusos de color verde-amarillento o amarillo claro, cóncavos, densamente
cubiertos con pelos cortos visibles, especialmente en la parte exterior (Avilán et
al., 1998).

INTR
OD
UC
CIÓ
N
Semilla. Es ovoide, oblonga y alargada. Cubierta por un endocarpio grueso y
leñoso, con una capa fibrosa externa que se puede extender dentro del
mesocarpio. Ésta puede contener uno o más embriones (Chávez et al., 2001).
Fruto. Es una drupa carnosa, presentando un mesocarpio comestible de
diferente grosor y de color (amarillo o naranja), jugoso y con alto contenido de
carbohidratos, dependiendo de la variedad y las condiciones de cultivo. Su peso
varía desde 150 g hasta 2 kg, su forma también es variable, aunque generalmente
es ovoide-oblonga, notoriamente aplanada, redondeada u obtusa en ambos
extremos, de 4 a 25 cm de largo y 1.5 a 10 cm de grosor. El exocarpio es grueso,
frecuentemente con lenticelas blancas prominentes (Chávez et al., 2001).
II.2. Importancia económica de la producción de mango
La tendencia actual, cada vez más creciente, está dirigida hacia la
globalización. De esta manera, se induce una fuerte competencia entre los
distintos mercados nacionales debido a la apertura comercial, lo cual se ve
reflejado en un aumento de las exportaciones e importaciones en regiones
localizadas y sectores específicos como el de hortalizas y frutas tropicales (IPD,
2008).
II.2.1. Nivel mundial
La producción mundial anual de mango es aproximadamente 30 x 106 t,
localizándose el mayor consumo en los países productores (asiáticos y
latinoamericanos). Dicha producción se encuentra integrada de la siguiente
manera (FAO, 2010): India ocupa el primer lugar, al producir aproximadamente 11
x 106 t (36% de la producción mundial); inmediatamente después China con una
aportación del 11% y enseguida Pakistán con el 7% de la producción tota. No muy
atrás se encuentra México, con una aportación de más del 6%, lo que equivale
aproximadamente a 2 x 106 t (Fig. II.2).
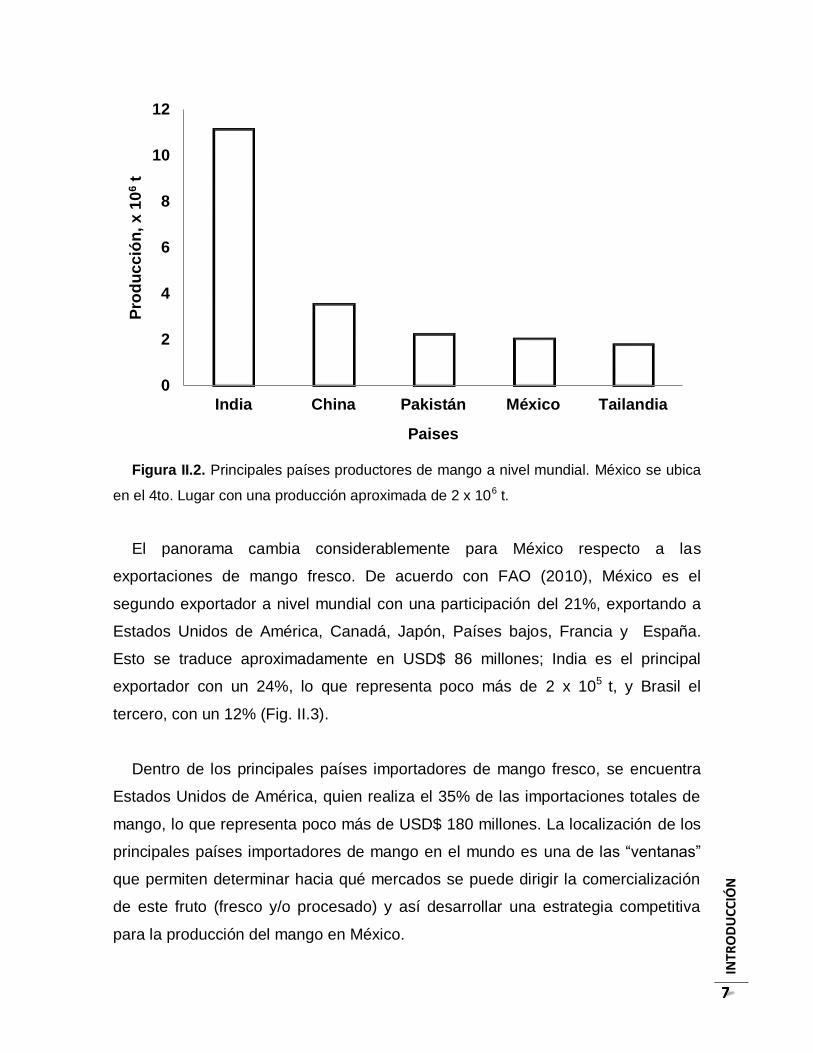
INTR
OD
UC
CIÓ
N
Figura II.2. Principales países productores de mango a nivel mundial. México se ubica
en el 4to. Lugar con una producción aproximada de 2 x 106 t.
El panorama cambia considerablemente para México respecto a las
exportaciones de mango fresco. De acuerdo con FAO (2010), México es el
segundo exportador a nivel mundial con una participación del 21%, exportando a
Estados Unidos de América, Canadá, Japón, Países bajos, Francia y España.
Esto se traduce aproximadamente en USD$ 86 millones; India es el principal
exportador con un 24%, lo que representa poco más de 2 x 105 t, y Brasil el
tercero, con un 12% (Fig. II.3).
Dentro de los principales países importadores de mango fresco, se encuentra
Estados Unidos de América, quien realiza el 35% de las importaciones totales de
mango, lo que representa poco más de USD$ 180 millones. La localización de los
principales países importadores de mango en el mundo es una de las “ventanas”
que permiten determinar hacia qué mercados se puede dirigir la comercialización
de este fruto (fresco y/o procesado) y así desarrollar una estrategia competitiva
para la producción del mango en México.
0
2
4
6
8
10
12
India China Pakistán México Tailandia
Pro
du
cc
ión
, x
10
6 t
Paises
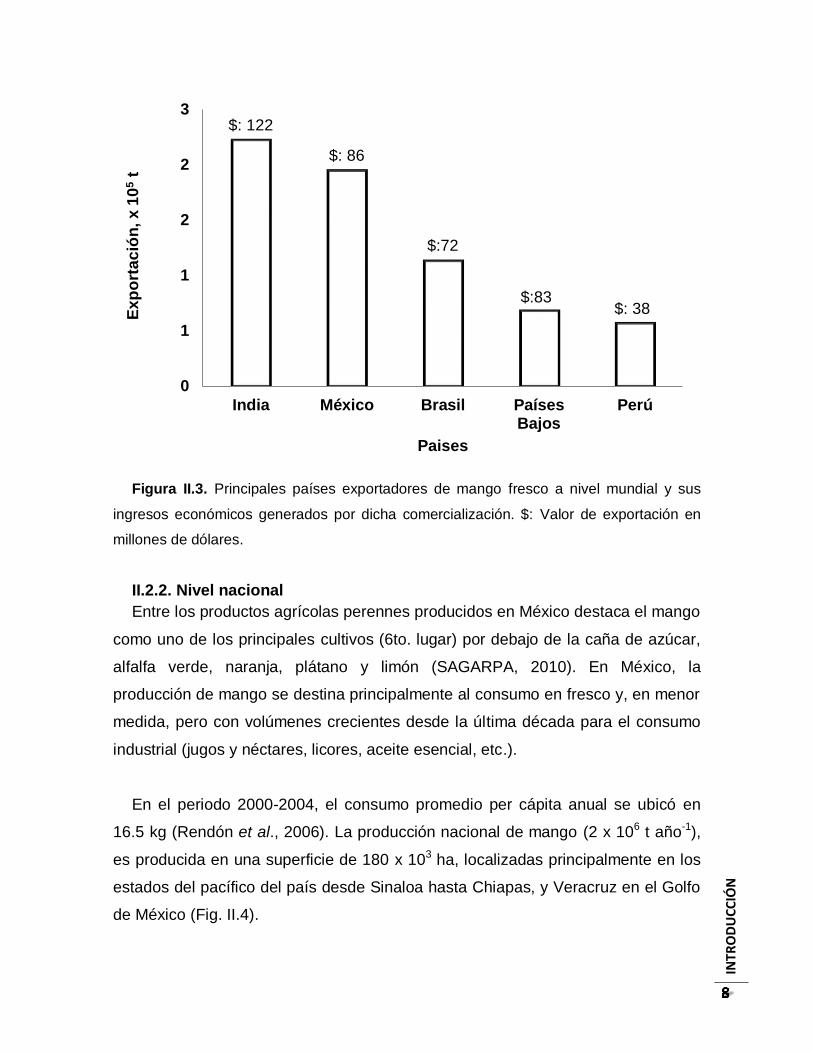
INTR
OD
UC
CIÓ
N
Figura II.3. Principales países exportadores de mango fresco a nivel mundial y sus
ingresos económicos generados por dicha comercialización. $: Valor de exportación en
millones de dólares.
II.2.2. Nivel nacional
Entre los productos agrícolas perennes producidos en México destaca el mango
como uno de los principales cultivos (6to. lugar) por debajo de la caña de azúcar,
alfalfa verde, naranja, plátano y limón (SAGARPA, 2010). En México, la
producción de mango se destina principalmente al consumo en fresco y, en menor
medida, pero con volúmenes crecientes desde la última década para el consumo
industrial (jugos y néctares, licores, aceite esencial, etc.).
En el periodo 2000-2004, el consumo promedio per cápita anual se ubicó en
16.5 kg (Rendón et al., 2006). La producción nacional de mango (2 x 106 t año-1),
es producida en una superficie de 180 x 103 ha, localizadas principalmente en los
estados del pacífico del país desde Sinaloa hasta Chiapas, y Veracruz en el Golfo
de México (Fig. II.4).
$: 122
$: 86
$:72
$:83 $: 38
0
1
1
2
2
3
India México Brasil PaísesBajos
Perú
Ex
po
rta
ció
n, x
10
5 t
Cie
nto
s
Paises
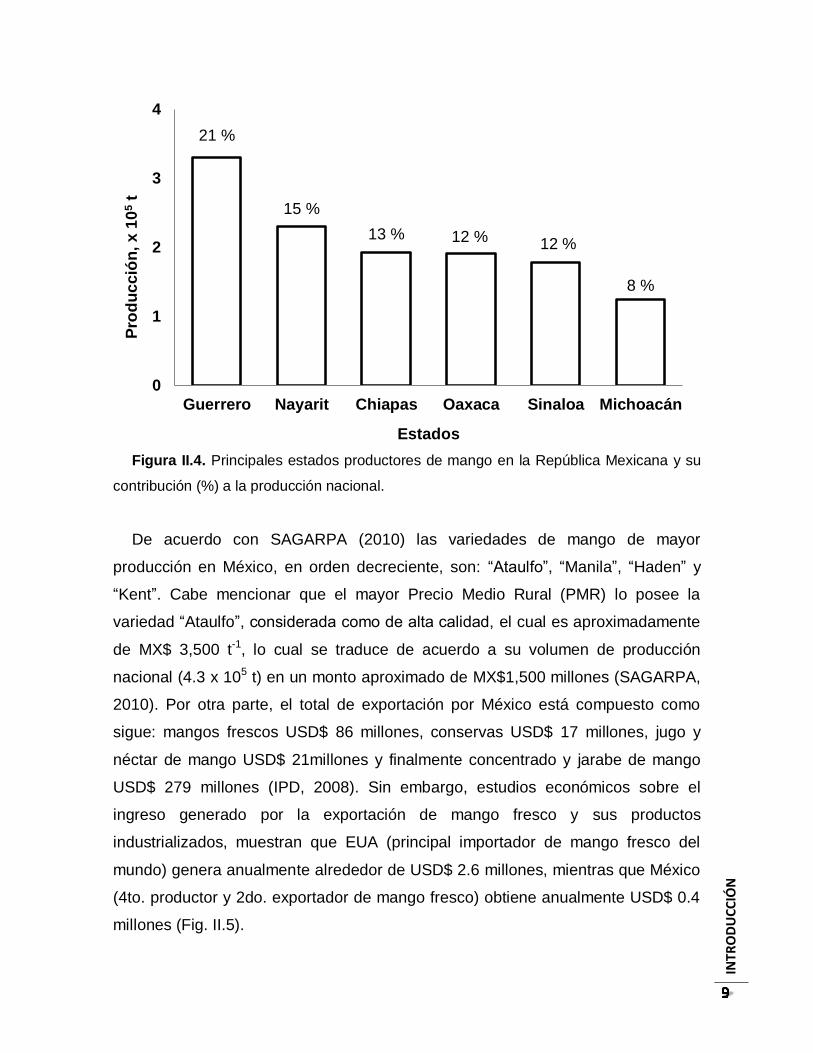
INTR
OD
UC
CIÓ
N
Figura II.4. Principales estados productores de mango en la República Mexicana y su
contribución (%) a la producción nacional.
De acuerdo con SAGARPA (2010) las variedades de mango de mayor
producción en México, en orden decreciente, son: “Ataulfo”, “Manila”, “Haden” y
“Kent”. Cabe mencionar que el mayor Precio Medio Rural (PMR) lo posee la
variedad “Ataulfo”, considerada como de alta calidad, el cual es aproximadamente
de MX$ 3,500 t-1, lo cual se traduce de acuerdo a su volumen de producción
nacional (4.3 x 105 t) en un monto aproximado de MX$1,500 millones (SAGARPA,
2010). Por otra parte, el total de exportación por México está compuesto como
sigue: mangos frescos USD$ 86 millones, conservas USD$ 17 millones, jugo y
néctar de mango USD$ 21millones y finalmente concentrado y jarabe de mango
USD$ 279 millones (IPD, 2008). Sin embargo, estudios económicos sobre el
ingreso generado por la exportación de mango fresco y sus productos
industrializados, muestran que EUA (principal importador de mango fresco del
mundo) genera anualmente alrededor de USD$ 2.6 millones, mientras que México
(4to. productor y 2do. exportador de mango fresco) obtiene anualmente USD$ 0.4
millones (Fig. II.5).
21 %
15 %
13 % 12 % 12 %
8 %
0
1
2
3
4
Guerrero Nayarit Chiapas Oaxaca Sinaloa Michoacán
Pro
du
cc
ión
, x
10
5 t
Estados

INTR
OD
UC
CIÓ
N
Figura II.5 Ingresos económicos, en millones de dólares, de EUA y México por la
exportación de mango fresco y sus productos industrializados.
Este análisis, muestra que la actividad económica en relación al mango, no
depende solamente de grandes superficies destinadas a su cultivo o de “altos”
volúmenes de producción, sino del desarrollo de nuevos e innovadores sistemas
de producción e industrialización, que permitan que la producción de mango en
México sea una actividad sustentable para la economía de los productores así
como para el ambiente.
II.3. Cadena de valor del mango en México
La cadena de valor del mango es un conjunto de eslabones interrelacionados
entre sí, que representan las etapas determinantes en el aprovechamiento óptimo
del mango, cuya aplicación repercute positivamente en la economía de la región
en la que se aplica (Fig. II.6). En el presente trabajo fueron identificados eslabones
débiles o críticos dentro de esta cadena, debido a la insuficiente investigación
científica entorno a ellos, los cuales son: la producción (el evento de floración y
control de enfermedades) y la industrialización (aprovechamiento de residuos
0
0.5
1
1.5
2
2.5
3
EUA México
Ex
po
rta
ció
n, ,
mil
lon
es
de
dó
lare
s
Paises
Fresco o seco
Conservas
Jugo y néctar
Concentrado y jarabe

INTR
OD
UC
CIÓ
N
industriales). Estos eslabones perjudican a la cadena completa y en consecuencia
la actividad económica de los productores, además de los problemas ambientales
que éstos ocasionan.
Figura II.6. Cadena de valor del mango en México y los puntos críticos que la
desestabilizan ( ).
II.3.1. La producción
II.3.1.1.- El evento de floración
Floración en Arabidopsis thaliana. Investigación sobre la floración en la
planta modelo A. thaliana ha generado conocimiento sobre la compleja red génica
que controla el tiempo de transición floral (Fig. II.7); estos genes participan en
diferentes rutas que integran señales ambientales y señales endógenas.
Básicamente, cinco rutas que interactúan entre sí han sido descritas: ruta de
respuesta a giberelinas, respuesta autónoma, respuesta a vernalización,
respuesta termosensorial y respuesta a fotoperiodo (Liu et al., 2009). Estas rutas
convergen en dos genes llamados “integradores de la ruta floral”: FLOWERING
LOCUS T (FT), y SUPPRESSOR OF OVEREXPRESSION OF CONSTANS
Punto crítico para la fortaleza del eslabón en la cadena de valor del mango en México. Punto crítico para la fortaleza del eslabón en la cadena de valor del mango en México. Punto crítico para la fortaleza del eslabón en la cadena de valor del mango en México. Punto crítico para la fortaleza del eslabón en la cadena de valor del mango en México.
INTR
OD
UC
CIÓ
N
(SOC1) (Nilsson et al., 1998; Kardailsky et al., 1999; Kobayashi et al., 1999;
Blázquez y Weigel, 2000; Lee et al., 2000; Samach et al., 2000). Los genes
integradores de la ruta floral tienen como blanco a “genes de identidad del
meristemo floral”, quienes confieren identidad floral a los nuevos primordios
emergentes. Estos genes a su vez activan a los llamados “genes de identidad de
organos florales” cuya expresion regionalizada, mediante interacciones de
proteínas de identidad del meristemo floral, conduce a un arreglo adecuado de los
organos florales (Blázquez, 2000; Liu et al., 2009).
Figura II.7. Ruta simplificada de señalización para la inducción floral en A. thaliana.
Las flechas en verdes indican efecto promotor, mientras que los conectores en rojo
indican efectos represivos. Dos elipses unidas indican interacción proteína-proteína. Los
asteriscos indican regulación transcripcional directa. AGL24, AGAMOUS-LIKE 24; SOC1,
SUPPRESSOR OF OVEREXPRESSION OF CONSTANS 1; FLC, FLOWERING LOCUS
C; SVP, SHORT VEGETATIVE PHASE; FT, FLOWERING LOCUS T; FD, FLOWERING
LOCUS D; FUL, FRUITFULL; CAL, CAULIFLOWER; LMI1, LATE MERISTEM IDENTITY
1; LFY, LEAFY; TFL1, TERMINAL FLOWER 1; AP1, APETALA1.

INTR
OD
UC
CIÓ
N
El impacto de algunos factores nutrimentales sobre la floración, debido a su
complejidad, aún no ha sido ubicado en las rutas antes mencionadas. Por ejemplo,
el papel de la disponibilidad de nutrientes sobre la floración es aún incierto; sin
embargo, se ha reportado que los niveles de carbohidratos, así como la actividad y
localización de las enzimas relacionadas con su biosíntesis, tienen un efecto sobre
este evento (Rolland et al., 2006), pudiendo ser estos factores parte de algunas de
las rutas descritas anteriormente o estar involucrados en nuevas rutas
completamente independientes.
Es posible que mecanismos similares a los descritos en A. thaliana se
presenten en especies vegetales de interés agronómico como lo es el árbol de
mango, pero con dinámicas de expresión génica diferentes, como lo sugieren las
distancia filogenética entre dichas especies, estrategias de inducción floral, etc.
Floración en mango. Se ha propuesto que la inducción floral en el árbol de
mango (Fig. II.8) comienza con el evento de iniciación, en el cual ocurren
crecimientos de los ápices o tallos en dormancia (Davenport y Núñez-Eliséa,
1997). Este evento involucra la división y elongación de células en dormancia en el
primordio de las hojas, meristemos axilares o ambos, y posteriormente del
meristemo apical (Davenport, 2007). Dicha iniciación es influenciada por la poda,
defoliación, irrigación bajo condiciones de sequía, fertilizantes nitrogenados y otros
factores como etileno o cambios de temperatura. Según Reece et al. (1949), el
destino de las yemas no está determinado hasta que su crecimiento es iniciado y
depende de los factores, no relacionados a la iniciación, que están presentes en el
momento que ésta ocurre. Lo cual conduce al siguiente evento que es la inducción
(vegetativa y/o floral), que difiere del modelo de floración en plantas herbáceas
propuesto por Bernier et al. (1993), debido a que la inducción en el árbol de
mango puede ser reversible dependiendo de la temperatura a la cual es sometido
durante el desarrollo temprano del ápice (Núñez-Eliséa et al., 1996). Se ha
propuesto que una alta relación citocininas: auxinas en las yemas determina el
crecimiento de los ápices. A su vez, el evento de inducción parece estar

INTR
OD
UC
CIÓ
N
gobernado, de acuerdo con Davenport y Núñez-Eliséa (1997), por la relación de
un promotor florigénico dependiente de temperatura (PF) (potenciado a bajas
temperaturas) y un promotor vegetativo dependiente de la edad (PV) (potenciado
a edades tempranas). Así, la inducción floral en el subtrópico es regulada
principalmente por la intensidad y duración de las bajas temperaturas, pero la
edad del crecimiento vegetativo anterior impacta la respuesta; mientras que en el
trópico, la floración es dictada por la duración del crecimiento vegetativo previo y
posiblemente modificada por las bajas temperaturas (Davenport, 2007). La
duración del crecimiento vegetativo así como el resto de los estadios fenológicos
presentes en el árbol de mango varían de acuerdo a su genotipo, nivel de
fertilización, humedad, y condiciones ambientales, entre otros (Avilán, 1971; Avilán
et al., 1995). Por lo cual, la aplicación del fertilizante se debe asociar con los
momentos de ocurrencia de los periodos de crecimiento vegetativos, floración y
fructificación, ya que estos son los periodos en los cuales existe una mayor
demanda de nutrimentos, como ha sido evidenciado por un marcado descenso en
las concentraciones de nitrógeno (N), fósforo(P) y potasio (K) durante las etapas
de formación del fruto (N: 10.4, P: 0.9 y K: 5.3 g kg-1 de materia seca) en relación a
la etapa de dormancia (N: 12.4, P: 1.1 y K: 7.5 g kg-1 de materia seca) (Avilán,
1971). La cantidad de fertilizantes a emplear durante un ciclo de producción, se
fracciona y aplica antes de la ocurrencia de los periodos críticos mencionados
(Avilán et al., 1993).
Zhang et al. (2005) y Davenport et al. (2006) aislaron un gen nombrado MiCOL
(CONSTANS-like) de la hoja del árbol de mango. CONSTANS es un gen de
expresión circadiana en la ruta de fotoperiodo en A. thaliana (Putterill et al., 2004).
Debido a que el mango no es sensible al fotoperiodo, el papel de este gen
permanece incierto. Así mismo, se han realizado esfuerzos -sin éxito- para aislar
el gen FT u otros ortólogos, probablemente responsable de la síntesis del PF
(Davenport, 2007). Por otra parte, se ha concluido que el PF posee un tiempo de
vida corto in situ (6 a 10 días) y se transporta hasta una distancia de 100 cm,
siendo acarreado por fotosintatos a través del floema, desde las hojas donadoras

INTR
OD
UC
CIÓ
N
a las yemas receptoras. El transporte del PF mediado por los fotosintatos es
apoyado por experimentos de sombreado (90 y 100% de hojas sombreadas), en
los cuales se detuvo la producción fotosintética completamente, observando sólo
crecimiento vegetativo, aún en condiciones de inducción floral (Davenport, 2007).
De igual manera, el proceso llamado anillado o constricción apoya esta idea, el
cual es usado para el control del flujo vascular basipétalo de los compuestos que
se desplazan a través del árbol mediante los haces vasculares. El anillado se
efectúa mediante un corte o aplicando una fuerte presión sobre éstos, lo cual
induce la floración del árbol de mango y por consiguiente, estimula la generación
de frutos (Avilán et al., 1995). Davenport (2007) sugiere que el PV está
estrechamente ligado a la ruta de biosíntesis de giberelinas, debido a que
inhibidores de esta ruta (e.g. triazoles), retrasan el crecimiento vegetativo
induciendo floración temprana en el árbol de mango.
Figura II.8. Modelo teórico de las etapas involucradas en el evento de floración en
mango. Líneas continuas indican efecto promotor, mientras que, líneas punteadas indican
efecto represor. Las líneas sin cabeza de flecha indican un efecto que aún no ha sido
determinado con certeza.

INTR
OD
UC
CIÓ
N
Las alternativas utilizadas actualmente para adelantar o retrasar la floración en
mango, presentan inconvenientes como inconsistencia en su efecto positivo,
elevados costo de producción por la adopción de estas tecnologías y problemas
de contaminación ambiental (Davenport y Núñez-Eliséa, 1991; Osuna-García et
al., 2001). El estudio detallado a nivel fisiológico, bioquímico y molecular del
evento de floración en el árbol de mango, es una herramienta que ayudará a
desarrollar alternativas biotecnológicas que permitan adelantar o retrasar dicho
evento, a través del entendimiento de los mecanismos involucrados, lo cual
conducirá a la generación de alternativas con mayor probabilidad de éxito,
disminuyendo los problemas de las prácticas actuales.
II.3.1.2. Control de enfermedades
Las enfermedades en cultivos agrícolas constituyen un grave problema a nivel
mundial (Gutiérrez-Alonso et al., 2002). Éstas son causadas por diversos
microorganismos, principalmente bacterias y hongos, que afectan las distintas
etapas fenológicas y órganos de la planta (Ploetz y Prakash, 1997), desde que se
encuentra en el vivero hasta el almacenamiento y transporte del fruto (Naqvi,
2004). El mango, como cualquier otra planta, es susceptible a enfermedades,
entre las cuales la antracnosis, causada por el hongo Colletotrichum
gloeosporioides, es la de mayor impacto negativo en la productividad y calidad del
fruto, estando distribuida en todas las zonas productoras en el mundo (Litz, 2000).
La clasificación taxonómica actual de este hongo fitopatógeno es CLASE:
Sordariomycetes, SUBCLASE: Hypocreomycetidae, ORDEN: Hypocreomycetidae
incertaesedis, FAMILIA: Glomerellaceae, GÉNERO: Glomerella, ESPECIE:
cingulata, siendo C. gloeosporioides su estado anamorfo (NCBI, 2010). El cual, es
un patógeno ubicuo, asociado a infecciones quiescentes y desarrollo de
antracnosis en muchas frutas tropicales incluyendo mango, plátano, aguacate,
papaya, café, pera, manzana y guayaba (Yenjit et al., 2004; Nelson, 2008). Este
patógeno produce hifas hialinas unicelulares, ovoides y alargadas, con conidios
ligeramente curvados (de 10 a 15 µm de longitud y de 5 a 7 µm de ancho). Los

INTR
OD
UC
CIÓ
N
acérvulos que se producen en el tejido infectado pueden ubicarse en forma
subepidermal, epidermal o subcuticular con conidióforos cortos y erguidos. Las
especies de Colletotrichum spp. causantes de la antracnosis en diversos cultivos,
presentan dos fases principales de nutrición durante la colonización de la planta
(Fig. II.9): la fase inicial biotrófica, en la cual el patógeno obtiene nutrientes de las
células vivas del hospedero, y la segunda fase tardía, necrotrófica, donde los
nutrientes se obtienen de las células hospederas muertas a causa del ataque del
patógeno (Bailey et al., 1992). La fase biótrofa es de corta duración y en ésta se
asegura el establecimiento del patógeno, sin daños severos en el tejido vegetal.
La actividad enzimática requerida para degradar la pared vegetal está
estrictamente limitada durante esta fase y la planta hospedera parece no
reconocer al patógeno, y en consecuencia, no se desencadenan respuestas de
defensa por la planta (Perfect et al., 1999). A la fase necrotrófica se asocia a la
aparición de los síntomas de la antracnosis, ya que existe una estrecha relación
entre su aparición, el incremento en la expresión enzimática para degradar la
pared celular vegetal y la virulencia del patógeno (Centis et al., 1997).
Figura II.9. Fases de colonización de C. gloeosporioides en el fruto del árbol de
mango.

INTR
OD
UC
CIÓ
N
Esta enfermedad se presenta en todas las etapas fenológicas y órganos de
árbol de mango, causando pérdidas en la producción, tanto en precosecha como
en postcosecha (Cook, 1975). Se ha reportado que la mayor concentración de
esporas de C. gloeosporioides está asociada con épocas de mayor humedad
relativa, lo que coincide con la fase fenológica del crecimiento vegetativo del
mango (Saldarriaga et al., 1997). Los síntomas se manifiestan con mayor
frecuencia en el ápice o en los puntos en que varios frutos de una misma
inflorescencia quedan en contacto, debido a que allí se presenta acumulación de
agua por un intervalo de tiempo mayor, favoreciendo el desarrollo inicial del hongo
(Girard, 1980; Pérez, 1993).
Los síntomas de esta enfermedad (Fig. II.10) sobre hojas jóvenes se
manifiestan como pequeñas manchas cafés-oscuras rodeadas de un halo clorótico
que pueden coalescer para formar lesiones irregulares de 0.3 a 1.0 cm de
diámetro. En muchas ocasiones, estas lesiones tienen como consecuencia el
desprendimiento del tejido. En la floración, los síntomas se inician como pequeñas
manchas negras sobre flores, pedúnculos, pedicelos y las raquis de la
inflorescencia, ocasionando la caída de flores (Espinosa et al., 2004) y pudiendo
afectar el porcentaje de retención de los frutos (Mendoza, 1977). En frutos en
desarrollo, puede ocasionar la caída de más del 90% cuando la enfermedad se
presenta en intensidades superiores al 80%; incluso cuando alcanza niveles de
daño del 40% la caída de fruto puede alcanzar magnitudes superiores al 50 o 60%
(Espinosa et al., 2004).
Figura II.10. Síntomas de la antracnosis (causada por C. gloeosporioides) en
diferentes órganos del árbol de mango.
Hoja Panícula Frutos

INTR
OD
UC
CIÓ
N
De acuerdo con Ploetz (1999), la caracterización de poblaciones de C.
gloeosporioides en todo el mundo, indican que las cepas que infectan al mango
comprenden una población genética y patológicamente distintiva de toda la
especie, debido a que la población predominantemente patógena en mango, no
fue encontrada infectando otras frutas tropicales.
El manejo de la antracnosis en mango requiere de un control constante debido
a que las condiciones meteorológicas promueven la infección y desarrollo de la
enfermedad. Recientemente, se ha propuesto como sistemas de control óptimo,
diferentes enfoques integrados que combinan las medidas más eficientes en
función de la variedad, la localidad de producción y el mercado final (Ploetz, 2009).
Básicamente, este manejo consiste en cinco enfoques o prácticas principales,
como son: selección del sitio y de variedades resistentes, prácticas culturales en el
campo -sanitización, espaciamiento entre plantas, intercalado de plantas-,
aplicación de fungicidas y tratamientos postcosecha –físicos y químicos- (Nelson,
2008).
Por décadas, la practica agrícola mayormente utilizada para el control de la
antracnosis ha sido la aplicación de fungicidas químicos, los cuales incluyen
benzimidazoles, ditiocarbamatos e inhibidores de la síntesis de ergosterol:
difenoconazol, miclobutanil, y procloraz (Ker, 2001; Peres et al., 2004; Arias y
Carrizales, 2007); sin embargo, el uso de estos fungicidas no es una práctica
económica y ambientalmente sustentable.
La antracnosis impacta negativamente a la cadena de valor del mango en
México, debido a que el daño que genera puede manifestarse en todas los
estadios fenológicos del árbol, disminuyendo así la productividad hasta en un
90%, teniendo como consecuencias perdidas económicas fuertes de los
productores (Espinosa et al., 2004). Además, en la actualidad, el uso excesivo de
fungicidas químicos para el control de esta enfermedad ha conducido a fuertes
problemas, tales como: contaminación ambiental, resistencia de los patógenos,

INTR
OD
UC
CIÓ
N
residuos tóxicos en los frutos, etc. (Siddiqui, 2005; Wu et al., 2009). Por lo cual, el
desarrollo de alternativas económicas y amigables con el ambiente, que controlen
eficientemente esta enfermedad, es determinante para disminuir el uso de
fungicidas químicos utilizados actualmente, así como los problemas que éstos
ocasionan (Michel et al., 2001; Guigón y González, 2004; Verma et al., 2007).
II.3.2. La industrialización
II.3.2.1. Aprovechamiento de residuos industriales
La agroindustria constituye una parte del sector industrial que se dedica a
producir y/o transformar, almacenar y comercializar productos provenientes del
campo (Saval, 2012). Dichos procesos generan residuos agroindustriales, los
cuales son materiales en estado sólido o líquido que se generan a partir del
consumo directo de productos primarios o de su industrialización, y que no son de
utilidad para el proceso que los generó, pero son susceptibles de aprovechamiento
o transformación para generar otros productos con valor económico y de interés
comercial y/o social (Mata y Martínez Carrera, 2007).
Actualmente, el principal problema asociado a la generación de los residuos
agroindustriales es la falta de una clara conciencia ambiental para su manejo, así
como la escases de alternativas tecnológicas y recursos económicos para su
aprovechamiento (Bustos et al., 2005). Datos recientes evidencian los grandes
volúmenes de residuos agroindustriales generados en diferentes procesos: la
industria de la cerveza, del aceite de palma y del café, solamente utilizan el 8.0,
9.0 y 9.5% del peso de la materia prima, respectivamente, por cual cantidades
superiores al 90% en peso son residuos generados (Saval, 2012).
La composición de los residuos agroindustriales es muy compleja, y dependen
de la materia prima y del proceso que los generó. No obstante, comparten una
característica principal que es el contenido de materia orgánica, constituida por
diferentes porcentajes de celulosa, lignina, hemicelulosa y pectina (Stamets,
2003). Estos residuos pueden ser aprovechados, generando beneficios

INTR
OD
UC
CIÓ
N
ambientales y económicos. Sin embargo, la mayoría de industrias no cuentan con
algún plan para el tratamiento de éstos, debido a los altos costos económicos para
su reutilización y por el contrario, los ubican junto con la basura en los vertederos
o rellenos sanitarios (Yepes et al., 2008). En el mejor de los casos, éstos son
utilizados en alimentación animal, lo que conduce a la subutilización del gran
potencial biotecnológico de estos residuos, debido a sus altos contenidos de
azúcares, pigmentos, fibra alimentaria, proteína, polifenoles, lignina, etc. (Sreenath
et al., 2001). Estas características, potencialmente útiles, pueden ser
aprovechadas a través de sistemas biotecnológicos que permitan imprimirles un
alto valor agregado (producción de enzimas, proteína unicelular, pigmentos,
antibióticos, biomasa microbiana, etc.).
Existen básicamente tres categorías tecnológicas para el aprovechamiento de
residuos agroindustriales: la valorización biológica y química, la obtención de
combustibles y la valorización térmica (Abraham et al., 2007; Tsai, 2008).
Valorización biológica y química. Este tipo de tecnología conduce a la
disposición final de los residuos orgánicos para la obtención de gases, líquidos o
sólidos que pueden ser comercializables. Entre los procesos biológicos más
comunes y más usados por las industrias se encuentran el compostaje y la
lombricultura.
Obtención de combustibles. El biogás es un producto gaseoso que se
obtiene de la descomposición de la materia orgánica mediante la acción
bacteriana en condiciones anaeróbicas, siendo considerado como un subproducto
del compostaje y de la pirólisis (Peters, 2003). Su principal uso es en la producción
de energía eléctrica, térmica y biocarburante (Abraham et al., 2007).
Valorización térmica. Desde el punto de vista físico-químico, los procesos de
conversión energéticos se basan en la degradación de las moléculas orgánicas
por la acción del calor (Elias, 2003). Las tecnologías que procesan térmicamente

INTR
OD
UC
CIÓ
N
los residuos buscan la reducción de su volumen y la recuperación de energía a
partir de los gases, líquidos y sólidos que se generan. Estos procesos térmicos
pueden clasificarse según los requerimientos de oxígeno: calderas/ incineradores
(presencia de oxígeno) y pirólisis/ termólisis (ausencia de oxígeno) (Castaño y
Londoño, 2002). En relación a la industrialización del mango, el valle de Cauca en
Colombia, genera sólo en el despulpado de este fruto entre 50 y 55% de residuos
(cáscara, semilla, resto de pulpa y fibra) Ordoñez (2002). El análisis proximal de
éstos presenta las siguientes características (en %): humedad, 68.55; materia
seca, 31.45; proteína, 7,03; extracto etéreo, 5.5; cenizas, 3.48 y carbohidratos,
15.44, con un pH alrededor de 4.2 (Mejía et al., 2007). Estos residuos son una
alternativa potencial para la aplicación de diversas estrategias para su
aprovechamiento. Por ejemplo, la producción de biocombustibles, proteína
celular, aceites esenciales, producción de jugos y remoción de contaminantes
fenólicos (Trejo-Márquez, 2010). Couto y Sanromán (2006) sugieren que el cultivo
en estado sólido es un sistema costo-efectivo para la utilización de residuos
agroindustriales, como lo son los residuos de uva y vinería. Otro uso potencial de
los residuos de industrialización del mango es la biosorción de Hg2+ y Cr3+ en la
biomasa muerta de mango, la cual se determinó mediante el uso de
radiotrazadores a partir de soluciones acuosas (Tiwari et al., 1999).
Para México, no se ha reportado el porcentaje de residuos que genera la
industrialización de mango fresco. Probablemente, es muy similar a lo reportado
para otros países, como Colombia, que es aproximadamente 50%; dichos
residuos presentan problemas graves de contaminación en su disposición final
debido a su alto contenido de azucares y elevados volúmenes de generación.
Los residuos de la industrialización del mango representan una oportunidad y
reto tecnológico para el aprovechamiento integral de dicho fruto en México, desde
la obtención de metabolitos de interés hasta su uso en sistemas de producción de
microorganismos de importancia agrícola.

JUST
IFIC
AC
IÓN
III. JUSTIFICACIÓN
La cadena de valor del mango en México se ve perjudicada por la existencia de
eslabones débiles. Con base en el análisis de la información científica y empírica
sobre la cadena de valor de este cultivo, se identificó que los eslabones con
escaso conocimiento científico y, por lo tanto, de mayor impacto negativo sobre
dicha cadena son: la producción (el evento de floración y control de antracnosis) e
industrialización (aprovechamiento de los residuos).
Por lo anterior, el manejo integral de este cultivo depende de la aplicación de
nutrientes limitantes para el desarrollo fisiológico de la planta, manejo adecuado
de las enfermedades e investigación/ innovación científica enfocada a la utilización
de los residuos generados en el proceso de industrialización del mango.
Por esta razón, la presente tesis fue orientada a generar conocimiento científico
sobre los puntos críticos de la cadena de valor del mango, lo cual permitió
desarrollar alternativas agro-biotecnológicas enfocadas a minimizar los efectos
adversos de las prácticas actuales en la producción de mango, las cuales
repercuten negativamente sobre el ambiente y la economía de los fruticultores.

HIP
ÓTE
SIS
IV. HIPÓTESIS
El empleo de microorganismos autóctonos (promotores de crecimiento vegetal y
agentes de control biológico) constituyen una alternativa integral tanto para la
sincronización floral del árbol, como para la utilización de los residuos generados
en la industrialización del fruto mediante la producción de un agente de control
biológico contra la antracnosis. Lo cual conducirá al fortalecimiento significativo de
la cadena de valor de mango en México.

OB
JETI
VO
S G
ENER
ALE
S Y
ESP
ECÍF
ICO
S
V. OBJETIVOS
V.1. Generales
Diseñar un manejo de inoculación microbiana para el árbol de mango, en
relación a su ciclo fenológico y las características metabólicas de las cepas
seleccionadas, que repercuta positivamente en su inducción floral; estudiando el
efecto de señales exógenas (fotoperiodo, intensidad luminosa, humedad relativa y
temperatura) y endógenas (parámetros morfométricos, bioquímicos y moleculares)
sobre dicha inducción.
Diseñar un sistema potencialmente adoptable in situ por los productores de
mango, que permita la producción de un agente de control biológico contra el
hongo causante de la antracnosis (Colletotrichum gloeosporioides), utilizando
como sustrato los residuos generados en el proceso de industrialización del
mango.
V.2. Específicos
FASE I. Caracterización nutrimental y microbiológica de los sitios de estudio y
metabólica/molecular de los microorganismos aislados (promotores de crecimiento
vegetal, agentes de control biológico y el agente causal de la antracnosis).
a) Seleccionar la variedad de mango a estudiar, en relación a su demanda en
el mercado, niveles de producción - exportación en México y calidad del fruto. Así
como la identificación de los sitos de estudio, produciendo la variedad
seleccionada, presentado manejos agronómicos contrastantes.
b) Determinar el status nutrimental de los sitios de estudio (suelo, hoja y
frutos), a través de la cuantificación de contenido de materia orgánica (suelo),
nitrógeno, fósforo, potasio (suelo) y capacidad de intercambio catiónico (suelo),
infiriendo el impacto de los manejos agronómicos específicos en cada uno de ellos
sobre dichos parámetros.

OB
JETI
VO
S G
ENER
ALE
S Y
ESP
ECÍF
ICO
S
c) Caracterizar microbiológicamente el suelo, hojas y frutos en los sitios de
estudio (potenciales microorganismos promotores de crecimiento vegetal y
agentes de control biológico), estudiando dichos aislados a nivel metabólico
(características mencionadas anteriormente) y molecular.
d) Aislar e identificar molecularmente aislados de C. gloeosporioides obtenidos
de los sitios de estudio seleccionados, evaluando el grado de patogenicidad de
éstos sobre frutos almacenados bajo condiciones controladas.
e) Identificar los metabolitos producidos por el agente de control biológico
involucrado en la inhibición del crecimiento del agente causal de la antracnosis (C.
gloeosporioides).
f) Cuantificar el nivel de control biológico de la cepa seleccionada sobre el
crecimiento de C. gloeosporioides in vivo (mangos almacenados bajo condiciones
controladas).
FASE II. Producción del agente de control biológico contra C. gloeosporioides
usando los residuos de industrialización del mango, bajo un sistema adoptable in
situ por los productores de mango.
g) Identificar una fuente cercana y de fácil acceso para la obtención de los
residuos de industrialización de mango.
h) Caracterizar la composición proximal de estos residuos, centrándose en las
características de mayor importancia para su uso como sustrato en la producción
de un agente de control biológico.
i) Diseñar y optimizar un sistema de fermentación que permita la producción
del agente de control biológico seleccionado en la FASE I de manera sustentable,
centrándose en aquellos parámetros determinantes para su potencial adopción por
productores in situ (huertas de mango).

OB
JETI
VO
S G
ENER
ALE
S Y
ESP
ECÍF
ICO
S
j) Evaluar el impacto del sistema de fermentación desarrollado sobre
características importantes del agente de control biológico producido, tales como
viabilidad inicial/ en almacenamiento y capacidad de control biológico sobre C.
gloeosporioides.
Fase III. Inoculación de microorganismos promotores de crecimiento vegetal a
árboles de mango, estudiando su impacto, así como el de las señales exógenas y
endógenas, sobre el evento de floración.
k) Determinar el ciclo fenológico de los árboles de mango creciendo en el
invernadero, mediante el registro de las fechas de ocurrencia de cada una de sus
etapas: crecimiento vegetativo, dormancia, floración, fructificación.
l) Diseñar y aplicar un manejo de inoculación microbiana a los árboles de
mango en invernadero, en relación a sus requerimientos nutrimentales en cada
etapa fenológica y las características metabólicas de las cepas seleccionadas.
m) Determinar el impacto del manejo de inoculación microbiana sobre el
evento de floración de los árboles de mango en invernadero, a través de la
cuantificación de parámetros morfométricos, bioquímicos y moleculares.
n) Monitorear cada treinta minutos, durante el periodo de tiempo del ensayo,
las condiciones ambientales en el invernadero (humedad relativa, iluminancia y
temperatura), estudiando su papel en la inducción floral de los árboles de mango.
o) Integrar el conocimiento generado sobre el papel del manejo de inoculación
microbiana y las condiciones ambientales en el evento de floración del árbol de
mango en invernadero (parámetros morfométricos, bioquímicos y moleculares).
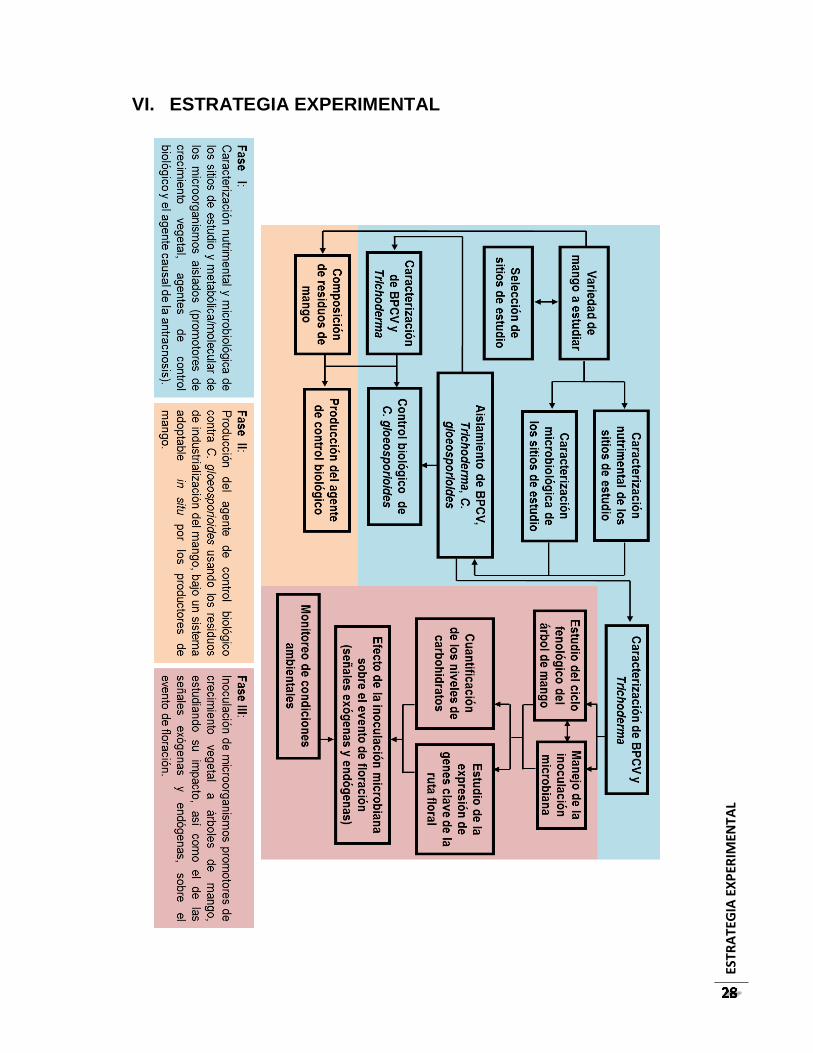
ESTR
ATE
GIA
EX
PER
IMEN
TAL
VI. ESTRATEGIA EXPERIMENTAL

MA
TER
IALE
S Y
MÉT
OD
OS
VII. MATERIALES Y MÉTODOS
FASE I. Caracterización nutrimental y microbiológica de los sitios de estudio y
metabólica/molecular de los microorganismos aislados (promotores de crecimiento
vegetal, agentes de control biológico y el agente causal de la antracnosis).
VII.1. Selección de la variedad de mango e identificación de los sitios de
estudio
La selección de la variedad de mango a estudiar fue analizada en relación al
impacto negativo de los puntos críticos antes mencionados sobre algunas de sus
ventajas competitivas de importancia, tales como: a) principal variedad producida
a nivel nacional, b) principal variedad de exportación y c) variedad con mayor
Precio Medio Rural.
Los sitios de estudio fueron ubicados en tres municipios productores de mango
de la variedad seleccionada, presentando características edáficas, climáticas, de
manejo y producción de mango contrastantes.
VII.2. Procedimiento de muestreo
Durante el ciclo de producción 2008-2009, aproximadamente 1 kg árbol-1 de
suelo, hojas y frutos fueron colectados de 10 árboles de mango “Ataulfo” de
huertas localizadas en los sitios de estudio seleccionados.
La colecta de las muestras se realizó de acuerdo con Avilán (2008), secciones
de 1 ha en cada uno de los sitios de muestreo fueron recorridos siguiendo un
trazado de dos diagonales en forma de “equis” (X), seleccionado 5 árboles en
forma aleatoria por cada diagonal, para un total de 10 individuos. Por cada árbol
seleccionado, se colectaron muestras de suelo, hojas y frutos en 4 puntos de
muestreos equidistantes (localizados en la dirección de los 4 puntos cardinales).
Las muestras de suelo fueron colectadas a una profundidad de 30 cm, en la
zona comprendida entre la mitad del radio de la copa y del perímetro de la misma.
Para el muestreo de material vegetal se siguieron las siguientes recomendaciones
(Kenworthy, 1964): hojas (el muestreo se realizó en hojas sanas y completas,

MA
TER
IALE
S Y
MÉT
OD
OS
entre la 3ra. y 6ta. desde la hoja terminal, durante la época de plena floración, en
brotes entre 5 y 6 meses de edad sin presentar dicho evento de crecimiento);
frutas (se colectaron aquellas que presentaron características similares de
tamaño, integridad y madurez).
Para el análisis nutrimental, las muestras fueron almacenadas en bolsas de
papel para su secado y posteriormente su análisis. Aquellas destinadas para el
análisis microbiológico fueron transferidas a una cámara húmeda y transportadas
a 4º C.
VII.3. Análisis físico-químico y nutrimental de los sitios de estudio
El análisis fisicoquímico y nutrimental de las muestras de suelo se llevó a cabo
por el Laboratorio Nacional de Fertilidad de Suelo y Nutrición Vegetal (Instituto
Nacional de Investigaciones Forestales, Agrícolas y Pecuarias; Campo
Experimental Bajío).
VII.4. Aislamiento e identificación de los microorganismos de interés en
los sitios de estudio
Dos estrategias fueron utilizadas para el aislamiento de microorganismos de
interés: (1) siembra directa y (2) siembra de diluciones.
En relación a la siembra directa, 1 g de suelo y de material vegetal (secciones
pequeñas de hojas y frutos) fueron directamente sembradas en cajas Petri
conteniendo medios de cultivos enfocados al aislamiento de los grupos
microbianos de interés. Para el aislamiento por siembra de diluciones, 10 g de
suelo, hojas o frutos fueron macerados en 90 ml de MgSO4 10mM (esterilizado por
autoclave a 121°C, 1 atm durante 15 min). El macerado fue homogenizado por 1 h
en un agitador rotatorio a 100 rpm. Una vez concluido el tiempo de
homogenización, diluciones seriales hasta 10-3 fueron preparadas y 1 ml de éstas
fueron sembradas en las cajas Petri que contenían medios de cultivo con la misma
composición a la utilizada para el aislamiento del microorganismo por siembra
directa, las cuales se citan a continuación.

MA
TER
IALE
S Y
MÉT
OD
OS
Aislamiento de cepas bacterianas. Medio Baz agar, con fuente de N* así
como libre de este elemento, fue utilizado para el aislamiento de microorganismos
promotores de crecimiento vegetal y fijadores de nitrógeno, respectivamente. Este
medio tuvo la siguiente composición (en g L-1): solución 1 (K2HPO4, 0.4; KH2PO4,
0.4; MgSO4, 0.2; CaCl2, 0.02; FeCl3, 0.01; Na2MoO4, 0.002; azul de bromotimol,
0.075; extracto de levadura*, 0.5; agar, 15) y solución 2 (arabinosa, succinato o
sacarosa, 5) (Estrada-de los Santos et al., 2001). Ambas soluciones fueron
ajustadas a pH de 5.7 y esterilizadas por autoclave (121°C, 1 atm durante 15 min)
separadamente. Posteriormente, las soluciones 1 y 2 fueron combinadas/
homogenizadas y 80 µg ml-1 de cicloheximida, esterilizada por filtración usando
una membrana hidrofílica Millipore con 0.22 µm de tamaño de poro, fueron
agregados.
Las cajas Petri inoculadas fueron incubadas a 28 ± 2° C durante 7 días. Todos
los aislamientos se llevaron a cabo por triplicado.
Aislamiento e identificación de cepas de Trichoderma. Dos medios de
cultivos fueron utilizados para el aislamiento de Trichoderma: Agar de Papa
Dextrosa (PDA), suplementado con carbencilina (80 µg ml-1), pH 6.0 y Medio
específico para Trichoderma (TSM), el cual tuvo la siguiente composición en g L-1:
MgSO4, 0.2; K2HPO4, 0.9; KCl, 0.15; KNO3, 2.0; glucosa, 3.0; rosa de bengala,
0.15, quintozeno, 0.65; agar, 20 y pH 6.0 (Askew y Laing, 1993).
Las cajas Petri inoculadas fueron incubadas a 28 ± 2° C durante 7 días. Todos
los aislamientos se llevaron a cabo por triplicado.
Las colonias fúngicas que presentaron características macroscópicas del
genero Trichoderma, tales como rápido crecimiento (Zadoks y Schein, 1979) y
formación de anillos concéntricos de color verde/ blanco en respuesta al
fotoperiodo, fueron seleccionadas y posteriormente analizadas
microscópicamente, en busca de las fialides típicas de este género (Barnet y
Hunter, 1978). Los aislados positivos para las características macroscópicas y

MA
TER
IALE
S Y
MÉT
OD
OS
microscópicas fueron crecidos en Caldo de Papa Dextrosa (PDB) para la
obtención de micelio, el cual fue utilizado para la extracción de ADN y
posteriormente para la identificación molecular de los posibles aislados del género
Trichoderma.
Extracción de ADN, modificado de Rader y Broda (1985):
1) Se colocaron ~ 100 mg de micelio fresco en un tubo Eppendorf de 1.5 ml y
se resuspendió con 500 µl de buffer de extracción. Posteriormente, la suspensión
se mezcló vigorosamente con ayuda de un Vórtex a máxima velocidad por 30 s.
Buffer de extracción: Tris HCL (pH 8.5), 200 mM; NaCl, 250 mM; EDTA, 25 mM;
SDS, 0.5%
2) 350 µl de fenol fueron agregados a la suspensión obtenida,
homogenizando vigorosamente utilizando un Vórtex a máxima velocidad por 30 s.
Posteriormente, 150 µl de cloroformo fueron adicionados, homogenizando
vigorosamente con un Vórtex a máxima velocidad por 30 s.
3) La suspensión obtenida fue centrifugada por 30 min a 13, 000 rpm. La fase
acuosa fue transferida inmediatamente a un nuevo tubo Eppendorf, y extraída con
1 volumen de cloroformo. Posteriormente, la mezcla fue centrifugada por 10 min a
13, 000 rpm.
4) La fase acuosa fue transferida a un nuevo tubo Eppendorf y 0.6 volúmenes
de isopropanol fueron agregados. La mezcla fue homogenizada 10 veces por
inversión y centrifugada por 5 min a 13, 000 rpm.
5) El pellet obtenido fue lavado dos veces utilizando 500 µl etanol 70% y
centrifugando por 2 min a 13, 000 rpm.
6) La integridad y concentración/pureza del ADN obtenido fueron analizadas
mediante un gel de agarosa 1% y NanoDrop, respectivamente.

MA
TER
IALE
S Y
MÉT
OD
OS
El ADN obtenido de los potenciales aislados pertenecientes al género
Trichoderma, fue utilizado para la confirmación molecular por la técnica de
Reacción en Cadena de la Polimerasa (PCR) usando oligonucleótidos específicos
para la identificación de este género. Esta identificación fue llevada a cabo de la
siguiente manera:
Oligonucleótidos específicos para el género Trichoderma (Liu et al., 2008)
Tf: 5´- TACCAAHCTGTTGCCTCGGCGG -3`
Tr: 5`- GATGAAGAAGGCAGCGAAATGCGATA -3`
Las reacciones de PCR fueron preparadas utilizando el kit PCR Master Mix -
K0171 de Fermentas, siguiendo las especificaciones del proveedor, bajo las
siguientes condiciones: 2 min a 94º C; 1 min a 94º C, 1 min a 56.6º C, 3 min at 72º
C para un total de 30 ciclos; y una extensión final de 10 min a 72º C). Utilizando
las condiciones anteriormente mencionadas, un aislado fue positivo para el género
Trichoderma si un amplicón de aproximadamente 450 pb fue observado en un gel
de agarosa al 1%.
Aislamiento e identificación de cepas de C. gloeosporioides. El aislamiento
de C. gloeosporioides fue llevado a cabo a partir de suelo, hojas y frutos
provenientes de los sitios de estudio, a través del aislamiento por siembra directa
o por siembra de diluciones, bajo las condiciones mencionadas con anterioridad.
El medio de cultivo utilizado para el aislamiento de este patógeno fue PDA
suplementado con 100 mg ml-1 de estreptomicina.
Además, el aislamiento fue llevado a cabo partiendo de lesiones características
generadas por la infección de hojas y frutos por C. gloeosporioides (antracnosis).
Para lo cual, hojas y mangos colectados de los tres sitios de estudio fueron
incubados a 28º C en una cámara húmeda conteniendo 0.1% de etileno, hasta
que la antracnosis fue observada. Posteriormente, secciones de los tejidos

MA
TER
IALE
S Y
MÉT
OD
OS
vegetales presentando dichas lesiones fueron desinfectados superficialmente con
etanol 70% por 1 min, eliminando el etanol con lavados de agua destilada estéril y
secando las secciones vegetales por flujo de aire. Las muestras secas fueron
inoculadas en cajas Petri conteniendo PDA suplementado con estreptomicina (100
mg ml-1) e incubadas a 25º C por 5 días. Después del periodo de incubación, las
colonias fúngicas fueron transferidas a nuevas cajas Petri conteniendo medio de
cultivo con la misma composición utilizada para el aislamiento de este hongo.
Todos los aislamientos se llevaron a cabo por triplicado.
Los potenciales aislados de C. gloeosporioides fueron identificados a través de
sus características macroscópicas y microscópicas, tales como: presencia de hifas
hialinas; coloración del micelio (esporas) rosa, salmón o gris y la forma ovoide de
las conidias (Smith y Black, 1990; Gunnel y Gubler, 1992).
Los aislados que presentaron características típicas de la especie C.
gloeosporioides fueron cultivados en PDB para la obtención de micelio, el cual fue
utilizado para la extracción de ADN, utilizando el protocolo mencionado
anteriormente (modificado de Rader y Broda, 1985) y posteriormente para la
identificación molecular utilizando oligonucleótidos específicos para dicha especie
Oligonucleótidos específicos para la especie C. gloeosporioides
C.gf: 5´- GGCCTCCCGCCTCCGGGCGG -3`
C.gr: 5`- TCCTCCGCTTATTGATATGC -3`
Las reacciones de PCR fueron preparadas utilizando el kit PCR Master Mix -
K0171 por Fermentas, siguiendo las especificaciones del proveedor, bajo las
siguientes condiciones: 5 min a 95°C; 30 s a 95°C, 30 s a 60°C, 1.5 min a 72°C
para un total de 30 ciclos; y una extensión final de 7 min a 72o C (Brown et al.,
1996). Utilizando las condiciones anteriormente mencionadas, un aislado fue
positivo para la especie C. gloeosporioides si un amplicón de aproximadamente
450 pb fue observado en un gel de agarosa al 1%.

MA
TER
IALE
S Y
MÉT
OD
OS
VII.5. Caracterización metabólica de los microorganismos aislados
Actividad de Reducción de Acetileno (ARA). Cada aislado bacteriano (5 x
103 UFC) fue inoculado en viales conteniendo 5 ml de medio Baz semisólido (2 g
L-1 de agar) libre de nitrógeno y con la fuente de carbono con la cual cada
microorganismo fue aislado. Estos viales fueron incubados a 28° C por 6 días,
posteriormente se sustituyó el 10% de aire de los viales por acetileno (Cavalcante
y Dobereiner, 1988). La concentración de etileno producido por cada aislado fue
estimada a las 2, 4 y 6 h después de la incubación, usando un cromatógrafo de
gases Hewlett Packard modelo 7890A, acoplado a un detector de ionización de
flama Hewlett Packard modelo 6890. Los experimentos de ARA fueron realizados
por triplicado.
Producción de Ácido Indol Acético (AIA). Cien mL de medio Baz líquido
suplementado con fuente de nitrógeno (0.5 g L-1 de extracto de levadura) y con
100 ppm de triptófano, fueron inoculados con 5 x 103 UFC de cada aislado
bacteriano o 1 x 105 esporas de cada aislado de Trichoderma. Estos cultivos
fueron incubados a 28° C durante 6 días a 100 rpm. Después de la incubación, 1
ml de cultivo se centrifugó a 13, 000 rpm por 3 min; al sobrenadante obtenido se le
agregaron 2 ml de reactivo de Salkowski (cloruro férrico, 0.4058 g; agua, 33 ml y
ácido sulfúrico, 20 ml). Dicha reacción fue incubada en oscuridad durante 20 min a
temperatura ambiente (Glickmann y Dessaux, 1995). La cuantificación de AIA fue
determinada a 540 nm en un espectrofotómetro TECAN modelo GENios. Este
ensayo fue realizado con tres réplicas independientes.
La cepa que produjo la mayor concentración de AIA, fue seleccionada para la
cuantificación de AIA, cinetina y ácido giberélico 3 (AG3), usando Cromatografía
Líquida de Alta Eficiencia (HPLC), bajo las condiciones de cultivo antes
mencionadas. Una vez que el tiempo de incubación del microorganismo concluyó,
1 mg de Hidroxi Tolueno Butilado (BTH) fue agregado a 100 ml del sobrenadante
del cultivo microbiano, para prevenir la oxidación de las fitohormonas (Ross et al.,
1987) y centrifugado a 1, 000 rpm por 10 min. La cuantificación de AIA, cinetina y

MA
TER
IALE
S Y
MÉT
OD
OS
AG3 fue determinada usando una columna de fase reversa LC-18 (25 cm x 4.6
mm, 5 µm). Las muestras fueron analizadas bajo condiciones isocráticas con
metanol 35% (en ácido acético 1%) para AIA, metanol (ajustado a pH 3 con H3PO4
0.1 M) para AG3 y metanol 70% para cinetina. Las longitudes de onda en el
detector ultravioleta fueron 280, 208, 254 para AIA, GA3 y cinetina,
respectivamente. El tiempo total de corrida para las separaciones fue de 15 min
con un flujo de 1 ml min-1. Las fitohormonas estudiadas fueron cuantificadas
utilizando estándares para cada una de ellas. La cuantificación de fitohormonas
fue realizada por duplicado.
Actividad 1-AminoCiclopropano-1-Carboxilato (ACC) desaminasa. Para
determinar la actividad ACC desaminasa de los aislados obtenidos, 5 x 103 UFC
de cada aislado bacteriano o 1 x 105 esporas de los aislados de Trichoderma
fueron inoculadas en tubos Falcon conteniendo 10 ml de Medio Sintético (MS)
(Yedidia et al., 1999) e incubados durante 4 días a 28º C y 150 rpm. La biomasa
total obtenida fue lavada dos veces utilizando agua desionizada estéril y
transferida a tubos Falcon conteniendo 5 ml de MS sin Amonio y con ACC 3 mM;
dichos tubos fueron incubados por 3 días, a 28º C y 150 rpm. Al finalizar el periodo
de incubación, los cultivos fueron resuspendidos y homogenizados en 0.5
volúmenes de Buffer Tris 0.1 M (pH 8.5). Posteriormente, 25 µl de Tolueno fueron
adicionados a una alícuota de 200 µl de suspensión celular, agitándose
vigorosamente usando Vórtex por 30 s. Posteriormente, 20 µl de una solución de
ACC 0.5 M fueron agregados a la suspensión celular. Esta reacción fue incubada
a 30° C por 15 min. Finalizado este periodo de incubación, 1 ml de HCl 0.56 N fue
agregado a la reacción. La suspensión celular lisada fue centrifugada a 10, 000
rpm por 10 min, y a 1 ml de sobrenadante se le agregó y homogenizó con 800 µl
de HCl 0.56 N y 300 µl de 2,4-dinitrofenilhidrazina (0.2 g en 100 ml de HCl 2 N). La
suspensión se incubó a 30° C por 30 min; posteriormente, 2 ml de NaOH 2N
fueron agregados y la absorbancia a 540 nm fue medida. La actividad ACC
desaminasa fue determinada por la cantidad de α-cetobutirato producida por la
desaminación del ACC (Viterbo Ada et al., 2010). La cantidad de proteína fue

MA
TER
IALE
S Y
MÉT
OD
OS
determinada con el Método de Bradford (Bradford, 1976). Los experimentos de
actividad ACC desaminasa se realizaron por triplicado.
Capacidad de control biológico in vitro de los microorganismos aislados.
Se realizaron ensayos de confrontación in vitro entre los aislados obtenidos y C.
gloeosporioides ATCC MYA 456. Para esto, cajas Petri conteniendo PDA (pH 6)
fueron inoculadas, con una distancia de 6 cm, con 1 x 105 esporas de C.
gloeosporioides ATCC MYA 456 y 1 x 105 UFC o esporas de la cepa [bacteriana
(en dos puntos) o de Trichoderma (en un punto), respectivamente]. Las cajas Petri
inoculadas fueron incubadas a 28 ± 2° C durante 7 días; posteriormente, el
porcentaje de inhibición de crecimiento de C. gloeosporioides ATCC MYA 456 fue
determinado (Rahman et al., 2007, Schmidt et al., 2009). La actividad antagónica
fue cuantificada utilizando tres réplicas independientes.
Ensayos de patogenicidad de los aislados de C. gloeosporioides y su
control biológico in vivo por la cepa seleccionada
Los ensayos de patogenicidad de los aislamientos de C. gloeosporioides se
realizaron en mangos verdes (cinco frutos por tratamiento) mediante la inoculación
de 1 x 106 esporas de cada aislado en cinco diferentes puntos por cada fruto. Los
frutos inoculados fueron incubados en una cámara húmeda a 28 ± 1° C, humedad
relativa de 85 ± 5%, fotoperiodo de 12 h (540 Lux) - 12h (20 Lux) durante 15 días.
El número de frutos que presentaron los síntomas característicos de la antracnosis
fueron registrados.
De manera similar, la cepa de Trichoderma que presentó la mayor capacidad de
control biológico contra C. gloeosporioides in vitro, se evaluó para estudiar su
capacidad de control biológico contra el patógeno in vivo (mangos verdes). Este
experimento se llevó mediante la inoculación de 1 x 106 esporas de cada uno de
los aislados de C. gloeosporioides y la cepa de Trichoderma estudiada. Las
esporas de cada hongo se colocaron a una distancia de 3 cm entre sí, en cinco
puntos diferentes por fruto (cinco frutos por tratamiento). Los frutos inoculados se

MA
TER
IALE
S Y
MÉT
OD
OS
incubaron en una cámara húmeda bajo las mismas condiciones descritas
anteriormente. Los ensayos de patogenicidad y de control biológico in vivo fueron
repetidos dos veces.
Metabolitos involucrados en la capacidad de control biológico de las
cepas seleccionadas
- Producción de compuestos volátiles. Este ensayo se llevó a cabo
inoculando, en cajas Petri conteniendo PDA pH 6.0, 1 x 104 UFC o esporas de la
cepa (bacteriana o de Trichoderma, respectivamente). Además, 1 x 106 esporas de
C. gloeosporioides ATCC MYA 456 fueron inoculadas en otra caja Petri
conteniendo el mismo medio de cultivo. Estas cajas (cepa bacteriana – C.
gloeosporioides y cepa de Trichoderma – C. gloeosporioides) fueron colocadas
una contra la otra y los extremos fueron sellados para aislar la atmósfera, evitando
la pérdida de compuestos volátiles. Las cajas fueron incubadas a 28 ± 2 °C por 7
días, posteriormente se determinó el porcentaje de inhibición de crecimiento de C.
gloeosporioides ATCC MYA 456 (Schmidt et al., 2009). Los experimentos de
producción de compuestos volátiles se realizaron por triplicado.
- Producción de sustancias difusibles. 1 x 104 UFC o esporas de las
cepas seleccionadas (cepa bacteriana y de Trichoderma, respectivamente) fueron
crecidas en 100 ml de medio PDB pH 6.0, e incubadas a 28 ± 2° C durante 4 días
a 100 rpm. Posteriormente, 1 ml de cultivo fue centrifugado a 10, 000 rpm durante
10 min. El sobrenadante obtenido fue filtrado a través de una membrana Millipore
hidrofílica con tamaño de poro de 0.45 µm. Cajas Petri conteniendo PDA pH 6.0
fueron inoculadas, en el centro, con 1 x 106 esporas de C. gloeosporioides ATCC
MYA 456 y 300 µl del sobrenadante filtrado en tres puntos alrededor del patógeno.
Las cajas Petri inoculadas fueron incubadas a 28 ± 2 °C por 7 días;
posteriormente, el porcentaje de inhibición de crecimiento de C. gloeosporioides
fue cuantificado (Schmidt et al., 2009). Este ensayo fue realizado utilizando tres
réplicas independientes.

MA
TER
IALE
S Y
MÉT
OD
OS
- Producción de sideróforos. Como ha sido establecido por Schwyn y
Neilands (1987), la técnica del Cromo Azurol S (CAS agar) fue utilizada para
determinar la producción de sideróforos por las cepas seleccionadas. 1 x 104 UFC
o esporas de las cepas (bacteriana o de Trichoderma, respectivamente) fueron
inoculadas en el centro de una caja Petri conteniendo el medio de cultivo antes
mencionado. El medio CAS fue preparado a partir de cuatro soluciones estériles:
La solución indicadora Fe-CAS (solución 1) se preparó con 10 ml de FeCl3 1 mM
(disuelto en HCl 1 mM) y 50 ml de CAS (1.21 mg ml-1). A la solución color azul
resultante se le agregaron 40 ml de CTAB (1.82 mg ml -1). La solución buffer
(solución 2) fue preparada disolviendo 30.24 g de PIPES en 750 ml de una
solución de sales que contenía 0.3 g KH2PO4, 0.5 g NaCl y 1.0 g NH4Cl, el pH fue
ajustado a 6.8 con KOH al 50%, el volumen se ajustó a 800 ml y se agregaron 15
g de agar. La solución 3 se preparó disolviendo 2 g glucosa, 2 g manitol, 493 mg
MgSO4, 11 mg CaCl2, 1.17 mg MnSO4, 1.4 mg H3BO3, 0.04 mg CuSO4, 1.2 mg
ZnSO4 y 1.0 mg Na2MoO4 en 70 ml de agua. Una solución de 30 ml de
casaminoácidos al 10% fue esterilizada por filtración (solución 4). Todas las
soluciones fueron mezcladas cuidadosamente adicionando al final la solución 1
(Alexander y Zuberer, 1991). Los experimentos de producción de sideróforos se
realizaron por triplicado.
- Identificación de la naturaleza química del sideróforo. La cepa
productora de sideróforo, 1 x 104 UFC, fue crecida en medio PDB a 28° C durante
4 días a 100 rpm; el cultivo fue centrifugado a 10, 000 rpm durante 10 min y
filtrado a través de una membrana Millipore con un tamaño de poro de 0.45 µm. La
naturaleza química del sideróforo producido fue determinada mediante un análisis
espectrofotométrico del sobrenadante. Estos ensayos fueron conducidos por
triplicado.
Hidroxamatos y catecolatos. La técnica del cloruro férrico (FeCl3) fue
empleada para la identificación de estas clases de sideróforo. Para ello, a 1 ml del
sobrenadante filtrado se le agregó 1 ml de FeCl3 2%. La presencia de un pico de

MA
TER
IALE
S Y
MÉT
OD
OS
absorbancia entre 400 y 450 nm indica la presencia de un sideróforo de naturaleza
hidroxamato, mientras que un pico entre 450 y 500 nm indica presencia de
catecolatos (Neilands, 1981). Las muestras fueron analizadas en un
espectrofotómetro de UV visible VARIAN modelo CARY 3E.
Carboxilatos. A 1 ml del sobrenadante filtrado se le agregó 1 ml de CuSO4
250 µM y 2 ml de buffer de acetato pH 4 (Na2HPO4 0.2 M, 19.3 ml y C6H8O7 0.1 M,
30.7 ml). El complejo formado con el cobre fue observado en un espectrofotómetro
de UV visible VARIAN modelo CARY 3E entre 190 y 280 nm (Shenker et al.,
1992).
- Cuantificación de la producción de sideróforo. La cantidad de sideróforo
producida fue cuantificada utilizando una curva de calibración (concentraciones de
10, 1 y 0.1 µg ml-1 de un sideróforo de la clase de hidroxamato, mesilato de
deferoxamina - Sigma Aldrich). Estas soluciones fueron sometidas a la prueba del
FeCl3, detectando un pico de absorbancia entre 400 y 450 nm en un
espectrofotómetro de UV visible VARIAN modelo CARY 3E.
- Actividades de enzimas líticas y su papel en el control biológico. La
actividad de enzimas líticas fue determinada en el sobrenadante del cultivo de la
cepa seleccionada en base a sus características de control biológico. Para ello, 1 x
104 esporas de Trichoderma fueron inoculadas en tubos conteniendo 25 ml de
medio sintético (MS) en g L-1: glucosa, 10; KNO3, 2.5; KH2PO4, 1; MgSO4, 0.5; el
pH fue ajustado 5.5 (Baig, 2005), sustituyendo la glucosa por carboximetilcelulosa
o residuos industriales de mango (composición descrita en la FASE II). Los tubos
fueron incubados durante 11 días a 24º C en un agitador rotatorio a 180 rpm.
Cada 24 h, los cultivos fueron filtrados a través de papel Whatman No. 1; el filtrado
se centrifugó a 10, 000 rpm durante 30 min y posteriormente fue filtrado a través
de fibra de vidrio. Los filtrados fueron utilizados como extractos enzimáticos crudos
para la cuantificación de las actividades enzimáticas estudiadas y para los
ensayos de control biológico contra C. gloeosporioides ATCC MYA 456.

MA
TER
IALE
S Y
MÉT
OD
OS
Las actividades de celulasas, quitinasas y glucanasas fueron cuantificadas de
acuerdo con Suresh y Chandrasekaran (1998), Kulminskaya et al. 2001 y Ghose
(1987), respectivamente. Los azúcares reductores liberados como resultado de las
actividades enzimáticas fueron medidos a 565 nm utilizando el método modificado
del ácido 3,5-dinitrosalicílico (DNS) (Miller, 1959). Una unidad de actividad fue
definida como la cantidad de enzima que libera 1 µmol equivalente de N acetil
glucosamina o de glucosa por minuto bajo las condiciones de reacción
especificadas. Estas actividades enzimáticas fueron expresadas como µmol g-1de
sustrato min-1. Los extractos enzimáticos crudos con las más altas actividades
específicas fueron utilizados para determinar sus papeles en el control biológico
contra C. gloeosporioides observado. Para esto, 1 x 104 esporas de C.
gloeosporioides ATCC MYA 456 fueron inoculados en tubos conteniendo 25 ml de
MS.
Los extractos enzimáticos crudos fueron esterilizados, utilizando un filtro
Millipore con 0.22 µm de tamaño de poro, y adicionados a los tubos inoculados
con las esporas del patógeno. Estos tubos fueron incubados durante 8 días a 24o
C en un agitador rotatorio a 180 rpm. Posteriormente, la viabilidad del micelio
producido fue determinada a través de su tinción debido a la reducción enzimática
(succinato deshidrogenasa) del cloruro de 2, 3, 5-trifeniltetrazolio (incoloro) al
0.15% disuelto en buffer de fosfato de potasio 50 mM (pH 7.0) a 1, 3, 5-
trifenilformazán (color rojo). La reacción fue incubada durante 24 h en la
oscuridad. Las células viables (específicamente las mitocondrias) redujeron
activamente el 2, 3, 5-trifeniltetrazolio a 1, 3, 5-trifenilformazán (color rojo), lo cual
se observó por microscopía de campo brillante (100x). Las células muertas
permanecieron incoloras. Además, el peso seco del micelio fue cuantificado. Estos
ensayos se llevaron a cabo por duplicado.
VII.6. Caracterización molecular de los microorganismos promisorios
La Identificación molecular de los aislados obtenidos, bacterias y hongos, fue
llevada a cabo por la amplificación por PCR del gen 16S ARNr (Weisburg et al.,
1991) y 5.8S ARNr (White et al., 1990), respectivamente. El ADN bacteriano fue

MA
TER
IALE
S Y
MÉT
OD
OS
extraído utilizando el Bacterial/Fungal DNA kit (Zymo Research) y el fúngico
utilizado el protocolo modificado de Rader y Broda (1985). Los amplicones
generados fueron purificados utilizando el GFX PCR DNA and Gel Band
Purification (Illustra) y secuenciados por plataforma Sanger. El programa
Sequence scanner fue utilizado para el análisis de los electroferogramas obtenidos
y el programa MEGA 5.0 para determinar las relaciones filogenéticas, utilizando el
método Neighbor-Joining. Las secuencias obtenidas fueron comparadas con
aquellas depositadas en el banco de genes del NCBI.
VII.7. Microscopía electrónica de barrido de las cepas promotoras de
crecimiento vegetal y el agente de control biológico
Las cepas promotoras de crecimiento vegetal y el agente de control biológico
seleccionados fueron cultivados (3 días, 28º C a 150 rpm) en medios líquidos con
la composición de los medios de los cuales fueron aislados. Las características
microscópicas fueron estudiadas utilizando microscopía electrónica de barrido
(SEM) [microscopio electrónico de barrido de bajo vacío (JEOL, JSM-5910 LV) a
30 Pa y kV 15)]. Las muestras fueron montadas sobre cilindros de cobre, y
posteriormente analizadas (Kaláb et al., 2008).
FASE II. Producción del agente de control biológico contra C. gloeosporioides
usando los residuos de industrialización del mango, bajo un sistema adoptable in
situ por los productores de mango.
VII.8. Identificación de la fuente y caracterización proximal de los residuos
de la industrialización del mango
La búsqueda de los residuos de la industrialización de mango fue realizada en
empresas dedicadas a esta actividad y localizadas en el municipio de Irapuato,
Guanajuato en México.
La caracterización proximal fue realizada a través de las siguientes
metodologías, contenido de sólidos totales y humedad [método 925.09, (AOAC,
1990)], cenizas [método 942.05, (AOAC, 1990)], nitrógeno [método 955.04,
(AOAC, 1990)], proteína [método 954.01, (AOAC, 1990)], grasas [método 920.39,
(AOAC, 1990)], materia orgánica/carbono (Walkley, 1947), fibra (Van Soest y

MA
TER
IALE
S Y
MÉT
OD
OS
Wine, 1967), y pH [método 943.02, (AOAC, 1990)]. El extracto libre de nitrógeno, o
carbohidratos más solubles, fue calculado por la resta al 100% de la suma de los
porcentajes de proteína, grasas, agua, cenizas, y fibra. Todas las determinaciones
fueron determinadas por tres réplicas independientes.
La caracterización proximal de los residuos de la industrialización del mango fue
utilizada como punto de partida para el diseño de un sistema de fermentación para
la producción de un agente de control biológico contra C. gloeosporioides.
VII.9. Diseño de un sistema de fermentación para la producción del agente
de control biológico, seleccionado en la fase I, contra C. gloeosporioides
La producción de esporas de Trichoderma cepa T8a, agente de biocontrol
contra C. gloeosporioides, fue estudiada bajo un sistema de fermentación,
potencialmente adoptable por los productores de mango in situ, en estado sólido
(espuma de poliuretano) utilizando los residuos de industrialización de mango
como sustrato.
La producción de esporas de la cepa T8a fue estudiada en cajas Petri de vidrio
conteniendo espuma de poliuretano como matriz de soporte inerte. Los
condiciones evaluadas fueron: relación carbono: nitrógeno, contenido inicial de
humedad y tiempo de colecta de las esporas.
Las relaciones carbono: nitrógeno (C: N) evaluadas estuvieron entre 6 y 34, las
cuales fueron obtenidas mediante la adición de sulfato de amonio comercial
(20.5% de pureza, la principal fuente de nitrógeno utilizada por los agricultores en
la producción de mango en México). El contenido de humedad varió desde 0, y de
64 hasta 94%. Los tiempos de colecta de esporas evaluados fueron de 0 a 9 días.
La fermentación en estado sólido consistió en 2 g de residuos industriales de
mango seco (ss), con un tamaño de partícula < 800 µm. Las cajas Petri de vidrio
conteniendo los residuos y espuma de poliuretano para todos los diferentes
tratamientos fueron esterilizadas a 121° C (1 atm) durante 15 min; después del

MA
TER
IALE
S Y
MÉT
OD
OS
proceso de esterilización, las cajas Petri fueron enfriadas a temperatura ambiente
antes de la inoculación de 2.5 x 106 esporas g-1 ss (28 ± 1 º C, 85 ± 5% de
humedad relativa, ciclos de 12 horas a 540 Lux seguido de 12 horas a 20 Lux).
Bajo estas condiciones de crecimiento, el impacto de diferentes relaciones C: N,
contenido de humedad, y tiempos de cosecha en la producción de esporas fueron
evaluados.
Las esporas fueron extraídas de los residuos industriales de mango y espuma
de poliuretano por lavado con agua conteniendo 1% de Triton X-100, seguido por
una homogenización vigorosa utilizando un Vórtex a máxima velocidad durante 5
min, y posteriormente una centrifugación a 5, 000 rpm durante 10 min a
temperatura ambiente. La extracción fue repetida tres veces, la concentración de
esporas se determinó utilizando una cámara de recuento al microscópico. Las
esporas colectadas de las condiciones que maximizaron su producción fueron
almacenadas a 4° C durante 7 días. Posteriormente, el porcentaje de viabilidad de
éstas fue evaluada como el número de unidades formadoras de colonias en cajas
Petri conteniendo PDA. Además, su capacidad de agente de control biológico fue
evaluada mediante ensayos de confrontación contra C. gloeosporioides ATCC
MYA 456, bajo las condiciones antes mencionadas. Todos los experimentos
fueron repetidos dos veces.
Además, muestras de la cepa de T. asperellum T8a creciendo sobre los
residuos industriales de mango y espuma de poliuretano (bajo las mejores
condiciones para la producción de esporas) fueron analizados por microscopía
electrónica de barrido (bajo las condiciones de preparación y análisis mencionadas
en la FASE I), con la finalidad de estudiar el comportamiento de esta cepa en
relación a la composición del material sobre el cual crece.

MA
TER
IALE
S Y
MÉT
OD
OS
FASE III. Inoculación de microorganismos promotores de crecimiento vegetal a
árboles de mango, estudiando su impacto, así como el de las señales exógenas y
endógenas, sobre el evento de floración.
VII.10. Estudio del ciclo fenológico de árboles de mango creciendo en
invernadero
Diez árboles de mango “Ataulfo” de 2 años de edad, injertados sobre una
variedad criolla de mango mexicano fueron utilizados. Estos árboles tuvieron
características morfométricas similares antes de iniciar el experimento, tales como:
número de hojas (de 15 a 20), altura (de 55 a 65 cm) y diámetro de tallo (de 3 a 4
cm). Dichos árboles fueron plantados en 10 kg de suelo proveniente de un huerto
de mango en producción activa ubicado en Tomatlán, Jalisco, México.
El ciclo fenológico de estos árboles de mango creciendo en el invernadero fue
determinado en la temporada junio 2009 - junio 2010, con la finalidad de conocer
la ocurrencia de cada etapa fenológica, así como la eficiencia de la floración bajo
éstas condiciones, identificando los períodos óptimos de riego y de inoculación de
los microorganismos promotores de crecimiento seleccionados previamente en
relación a sus características metabólicas. El ciclo fenológico se determinó como
el porcentaje del número total de árboles que presentan cada evento observado:
crecimiento vegetativo, latencia, floración y fructificación (Avilán, 2008).
VII.11. Diseño de un manejo de inoculación microbiana (seleccionadas en
la Fase I) para árboles de mango en invernadero
Una vez determinada la fenología de los árboles de mango en invernadero, el
efecto de la inoculación de las cepas seleccionadas sobre la inducción de la
floración del mango fue estudiado del 16 de julio de 2010 al 16 de febrero de 2011.
Para lo cual, dos tratamientos fueron diseñados: 1) no inoculados (control
negativo) y 2) manejo de la inoculación microbiana (1 x 106 UFC de cada cepa
seleccionada g-1 de suelo). Las cepas fueron aplicadas de acuerdo a los requisitos
nutrimentales del árbol en cada etapa fenológica y a las características
metabólicas de estos microorganismos. Un dispositivo HOBO U12 (Onset
Computer Corporation) fue colocado en el interior del invernadero con la finalidad
estudiar el impacto de condiciones climáticas (temperatura, la humedad relativa e

MA
TER
IALE
S Y
MÉT
OD
OS
iluminancia cuantificadas cada treinta minutos durante siete meses) en el evento
de floración.
VII.12. Cuantificación de parámetros morfométricos, bioquímicos y
moleculares involucrados en el evento de floración de árboles de mango
bajo el manejo de la inoculación microbiana
Parámetros morfométricos
Los parámetros morfométricos estudiados fueron peso seco de raíz, tallo y hoja;
así cómo, fecha de generación de yemas y su destino (vegetativo, reproductivo o
mixto), altura, número de hojas y flores, así como el área foliar [LI-3000C Portable
Leaf Area Meter (LI-COR)].
Parámetros bioquímicos
Hojas de cada árbol fueron colectadas e inmediatamente congeladas, usando
nitrógeno líquido, y almacenadas a -70º C. Este procedimiento fue repetido cada 7
días durante 6 semanas (del 18 de diciembre de 2010 al 21 de enero de 2011).
Estas muestras fueron utilizadas posteriormente para la cuantificación de
carbohidratos y perfiles de expresión del gen FT. Además, hojas al azar fueron
colectadas, secadas a 60º C y posteriormente el contenido de nitrógeno total fue
cuantificado (Axmann et al., 1990).
Cuantificación de carbohidratos en las hojas de mango. Muestras de tejido
foliar (150 mg) previamente colectadas y finamente molidas, usando nitrógeno
líquido, fueron colocadas en tubos de centrifuga de polietileno de 50 ml, a los
cuales fueron adicionados 10 ml de etanol al 80%. La mezcla fue homogenizada
vigorosamente utilizando un Vórtex a máxima velocidad durante 10 min,
posteriormente los tubos fueron incubados en un baño de agua a 80° C por 30 min
para extraer los carbohidratos solubles (Liu y Grieve, 2009). La suspensión fue
centrifugada a 10, 000 rpm durante 15 minutos y el sobrenadante fue transferido a
un tubo limpio de polietileno de 50 ml. El tejido húmedo se resuspendió en 10 ml
de etanol al 80% y el procedimiento de extracción fue repetido tres veces más. Los

MA
TER
IALE
S Y
MÉT
OD
OS
extractos etílicos obtenidos fueron combinados, congelados a -70º C y liofilizados
hasta que el volumen fue reducido a 1 ml, posteriormente éste fue evaporado con
nitrógeno gas hasta sequedad completa. Veinte µl de piridina y 100 µl de N, O-Bis
(trimetilsilil) trifluoroacetamida (BSTFA) fueron agregados a los extractos secos,
seguido de una incubación a 80° C durante 30 min. Posteriormente, las muestras
fueron inyectadas en un analizador de GC-MS (Agilent Technologies 7890A y
5975C, respectivamente). Los estándares (sacarosa, glucosa y fructosa), fueron
disueltos en agua destilada, y procesados simultáneamente con las muestras
(Medeiros y Simoneita, 2007). La cuantificación de carbohidratos se llevó a cabo
usando dos repeticiones independientes.
Parámetros moleculares
Extracción de ARN de las hojas de mango y cuantificación de la
expresión de FT por RT-PCR en tiempo real.
Los oligonucleótidos para la amplificación del transcrito de FT fueron diseñados
mediante el alineamiento de secuencias de aminoácidos de la proteína FT, las
cuales fueron obtenidas del banco de genes del NCBI, con el objetivo de identificar
regiones altamente conservadas, mediante el uso del programa Clustal X 2.0.12
(Thompson et al., 1997). Las secuencias de aminoácidos de las regiones
altamente conservadas fueron traducidas reversamente a nucleótidos (secuencia
de los oligonucleótidos) utilizando Sequence Manipulation Suite: Reverse
Translate (http://www.bioinformatics.org/sms2/rev_trans.html). El programa
Sequence scanner 1.0 y MEGA 5.0 fueron utilizados para el análisis de los
electroferogramas obtenidos por la secuenciación Sanger y para determinar las
relaciones filogenéticas utilizando el método Neighbor-Joining, respectivamente.
Una vez que los oligonucleótidos para la amplificación del transcrito del gen FT
fueron diseñados, la extracción de ARN total de las hojas de mango se llevó a
cabo utilizando Concert Plant RNA reagent (Invitrogen), de acuerdo a las
instrucciones del fabricante. Para la verificar la correcta amplificación de los
oligonucleótidos diseñados, una muestra del ARN obtenido fue utilizada para la

MA
TER
IALE
S Y
MÉT
OD
OS
síntesis de ADNc y amplificación de este transcrito por PCR, en el mismo tubo,
mediante el RobusT II RT-PCR Kit (Finnzymes). Las reacciones fueron llevadas a
cabo usando las siguientes condiciones de amplificación: 30 min a 40° C (1 ciclo);
2 min a 94° C (1 ciclo), 45 s a 94o C, 30 s a 53o C, 1 min a 72oC (30 ciclos) y una
extensión final a 72° C durante 5 min. Los productos de PCR fueron analizados
por electroforesis en un gel de agarosa al 2%. Los amplicones esperados fueron
purificados utilizando el GFX PCR DNA kit and Gel Band Purification (Illustra),
clonados usando el TOPO TA Cloning Kit (Invitrogen) y secuenciados por la
plataforma de Sanger. Las secuencias obtenidas fueron comparadas con aquellas
depositadas en el banco de genes del NCBI.
Una vez verificada la correcta amplificación de los oligonucleótidos diseñados,
los ARN obtenidos fueron utilizados para los estudiar los perfiles de expresión de
gel FT por RT-PCR en tiempo real. El ADNc de todas las muestras fue obtenido
utilizando el oligonucleótido inverso para FT (5'-
GAAACAACACAAATACAAACCTG -3 ') y SuperScript ™ III transcriptasa inversa
(Invitrogen) según las instrucciones del fabricante. Cada reacción fue preparada
ADNc generado a partir de 30 µg de ARN total, 1 × SYBR Green PCR Master Mix
(Applied Biosystems) y 500 nM del oligonucleótido directo (5'-
GTAATCCAAGTCTCAGAGAATAC-3 ') e inverso (5'-GAAACAACACAAATACAA
ACCTG-3') para la amplificación del transcrito de FT, los cuales fueron diseñados
utilizando el software AlleleID v. 7 y una secuencia parcial de ARNm del gen FT en
mango (obtenido anteriormente). El PCR en tiempo real se realizó en un ABI
PRISM 7500 Sequence Detection System (Applied Biosystems) bajo las siguientes
condiciones de ciclos térmicos: 10 min a 95 ° C seguido por un total de 40 ciclos
de 30 s a 95° C, 30 s a 60° C y 40 s a 72° C. En relación a qRT-PCR, la
abundancia relativa del transcripto se calculó y se normalizó con respecto al gen
de actina (ACT) [oligonucleótido directo (5'-CAATTTATGAGGGTTATGCACTC-3')
e inverso (5'-ATGTAACCTCTCTCAGTAA GAAT-3'), los cuales fueron diseñados
utilizando AlleleID v 7 software y una secuencia parcial de ARNm del gen de ACT
en mango (Singh et al., 2011)], así como con respecto al tratamiento no inoculado.

MA
TER
IALE
S Y
MÉT
OD
OS
Los datos mostrados representan los valores promedios obtenidos a partir de al
menos tres reacciones de amplificación independientes. Todos los cálculos y los
análisis fueron realizados utilizando el software 7500 v2.0.1 (Applied Biosystems)
y el método 2-ΔΔCt (Livak y Schmittgen, 2001). La eficiencia de amplificación
(0.9181 a 1.0071) para los pares de oligonucleótidos se determinó mediante
amplificación de diluciones seriales (1:5) del ADNc. La especificidad de los
productos de RT-PCR fue determinado por un análisis de curva de fusión con la
adquisición continua de datos de fluorescencia durante el evento de temperatura
de fusión entre 65-95° C.
VII.13. Análisis estadístico
Los datos obtenidos en el presente trabajo fueron analizados por la prueba de
análisis de varianza (ANOVA) de una vía y el método de Tukey-Kramer (p = 0.05),
utilizando el programa JMP-SAS 8.0.2.

RES
ULT
AD
OS
Y D
ISC
USI
ÓN
VIII. RESULTADOS Y DISCUSIÓN
FASE I. Caracterización nutrimental y microbiológica de los sitios de estudio y
metabólica/molecular de los microorganismos aislados (promotores de crecimiento
vegetal, agentes de control biológico y el agente causal de la antracnosis).
VIII.1. Selección de la variedad de mango e identificación de los sitios de
estudio
La variedad “Ataulfo” fue seleccionada como modelo de estudio debido a las
siguientes razones:
- Variedad de mayor producción a nivel nacional, alrededor de 4.3 x 105 t, lo
que representó en el 2010 un valor de la producción de MN$ 1,500 millones
aproximadamente (SAGARPA, 2010).
- Variedad con mayor Precio Medio Rural (MX$ 3,500 t-1), en otras palabras,
es la variedad que se vende a mejor precio en el mercado (SAGARPA, 2010).
- Se ubica dentro de las principales variedades de exportación (Chávez et al.,
2001).
Los sitios de estudio fueron ubicados en tres municipios productores de mango
“Ataulfo”, con características edáficas, climáticas, de manejo y producción de
mango contrastantes: Tapachula, Chiapas (6 x 104 t año-1, 14°56'00" N y
092°16'00" O, 179 MSNM, temperatura media anual de 26.4o C y 2,037 mm de
precipitación); Chahuites, Oaxaca (3 x 104 t año-1, 16°15'00" N y 094°14'00" O, 35
MSNM, temperatura media anual de 27.4o C y 1,010 mm de precipitación);
Apatzingán, Michoacán (3 x 101 t año-1, 19°05'00" N y 102°21'00" O, 320 MSNM,
temperatura media anual de 27.3o C y 809 mm de precipitación) (SMN, 2002;
SAGARPA, 2010).

RES
ULT
AD
OS
Y D
ISC
USI
ÓN
VIII.2. Análisis físico-químico y nutrimental de los sitios de estudio
El manejo agronómico de los sitios de estudio presentó contrastantes manejos
de las malezas crecidas durante la época de lluvias. Por ejemplo, éstas son
naturalmente incorporadas al suelo en las huertas de mango localizadas en
Apatzingán y Tapachula (menor densidad de malezas observadas comparada con
Apatzingán); mientras que en la huerta localizada en Chahuites éstas son
removidas del sistema (Fig. VIII.1).
Figura VIII.1. Manejo de las malezas crecidas en las épocas de lluvias en los sitios de
estudio. Las malezas en las huertas de mango localizadas en Apatzingán, Michoacán (19°
05'00 "N y 102 ° 21'00" W) y Tapachula, Chiapas (14° 56'00 "N y 092 ° 16'00" W) son
incorporadas naturalmente al suelo, mientras que aquellas crecidas en la huerta de
mango en Chahuites, Oaxaca (16° 15'00 "N y 094 ° 14'00" W) son removidas del sistema.
Esta incorporación de malezas, mostró una correlación positiva con la fertilidad
de los suelos en los sitos de estudio, siendo Apatzingán el más fértil (alto
contenido de materia orgánica, nitrógeno y capacidad de intercambio catiónico);
sin embargo, los suelos en Chahuites y Tapachula mostraron mayor contenido de
fósforo y potasio, respectivamente (Tabla VIII.1). El pH en los sitios de estudio fue
similar, con un rango entre 6.5 y 7.0. Además, el alto contenido de arcilla
observado en el suelo de Apatzingán comparado con Tapachula y Chahuites
(contenido de arcilla no detectado) sugiere una relación directa con la capacidad
de intercambio catiónico de los mismos (Tabla VIII.1). Observaciones similares
han sido reportadas por Otero et al. (1998), quienes observaron una fuerte
dependencia entre el contenido de arcilla y la capacidad de intercambio catiónico,
a diferentes niveles de profundidad de muestreo.
Chahuites Tapachula Apatzingán
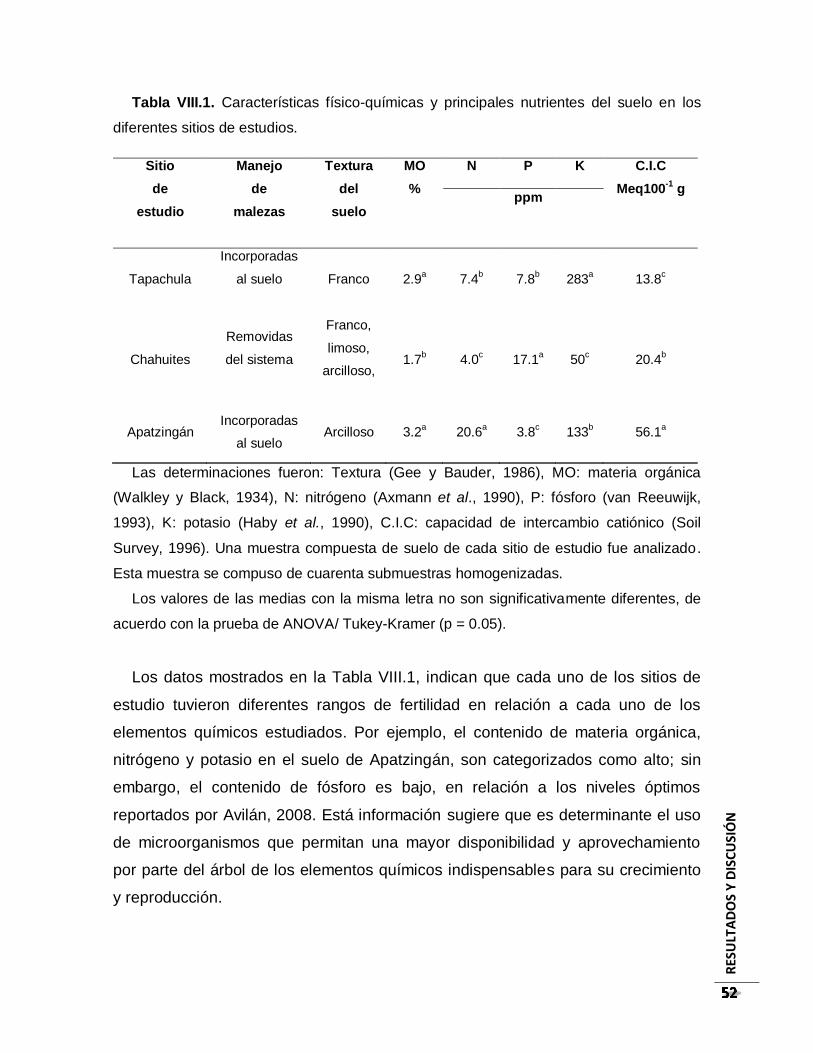
RES
ULT
AD
OS
Y D
ISC
USI
ÓN
Tabla VIII.1. Características físico-químicas y principales nutrientes del suelo en los
diferentes sitios de estudios.
Sitio
de
estudio
Manejo
de
malezas
Textura
del
suelo
MO
%
N P K C.I.C
Meq100-1
g ppm
Tapachula
Incorporadas
al suelo
Franco 2.9a 7.4
b 7.8
b 283
a 13.8
c
Chahuites
Removidas
del sistema
Franco,
limoso,
arcilloso,
1.7b 4.0
c 17.1
a 50
c 20.4
b
Apatzingán Incorporadas
al suelo Arcilloso 3.2
a 20.6
a 3.8
c 133
b 56.1
a
Las determinaciones fueron: Textura (Gee y Bauder, 1986), MO: materia orgánica
(Walkley y Black, 1934), N: nitrógeno (Axmann et al., 1990), P: fósforo (van Reeuwijk,
1993), K: potasio (Haby et al., 1990), C.I.C: capacidad de intercambio catiónico (Soil
Survey, 1996). Una muestra compuesta de suelo de cada sitio de estudio fue analizado.
Esta muestra se compuso de cuarenta submuestras homogenizadas.
Los valores de las medias con la misma letra no son significativamente diferentes, de
acuerdo con la prueba de ANOVA/ Tukey-Kramer (p = 0.05).
Los datos mostrados en la Tabla VIII.1, indican que cada uno de los sitios de
estudio tuvieron diferentes rangos de fertilidad en relación a cada uno de los
elementos químicos estudiados. Por ejemplo, el contenido de materia orgánica,
nitrógeno y potasio en el suelo de Apatzingán, son categorizados como alto; sin
embargo, el contenido de fósforo es bajo, en relación a los niveles óptimos
reportados por Avilán, 2008. Está información sugiere que es determinante el uso
de microorganismos que permitan una mayor disponibilidad y aprovechamiento
por parte del árbol de los elementos químicos indispensables para su crecimiento
y reproducción.
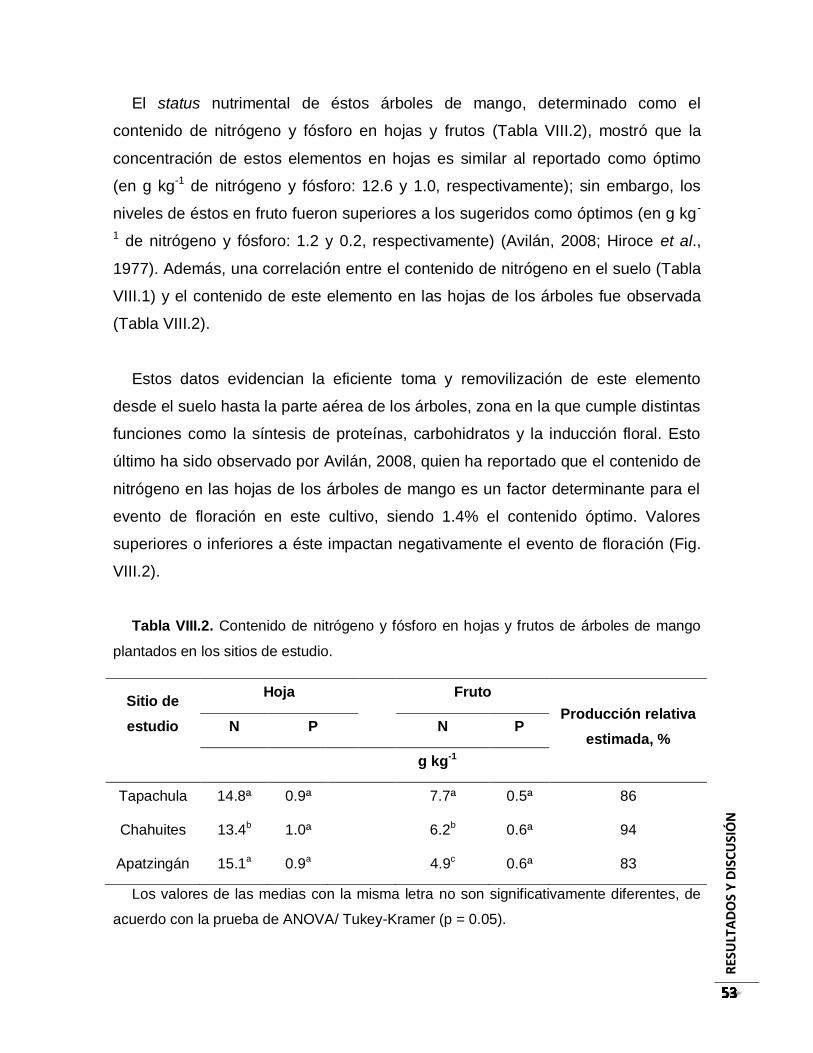
RES
ULT
AD
OS
Y D
ISC
USI
ÓN
El status nutrimental de éstos árboles de mango, determinado como el
contenido de nitrógeno y fósforo en hojas y frutos (Tabla VIII.2), mostró que la
concentración de estos elementos en hojas es similar al reportado como óptimo
(en g kg-1 de nitrógeno y fósforo: 12.6 y 1.0, respectivamente); sin embargo, los
niveles de éstos en fruto fueron superiores a los sugeridos como óptimos (en g kg -
1 de nitrógeno y fósforo: 1.2 y 0.2, respectivamente) (Avilán, 2008; Hiroce et al.,
1977). Además, una correlación entre el contenido de nitrógeno en el suelo (Tabla
VIII.1) y el contenido de este elemento en las hojas de los árboles fue observada
(Tabla VIII.2).
Estos datos evidencian la eficiente toma y removilización de este elemento
desde el suelo hasta la parte aérea de los árboles, zona en la que cumple distintas
funciones como la síntesis de proteínas, carbohidratos y la inducción floral. Esto
último ha sido observado por Avilán, 2008, quien ha reportado que el contenido de
nitrógeno en las hojas de los árboles de mango es un factor determinante para el
evento de floración en este cultivo, siendo 1.4% el contenido óptimo. Valores
superiores o inferiores a éste impactan negativamente el evento de floración (Fig.
VIII.2).
Tabla VIII.2. Contenido de nitrógeno y fósforo en hojas y frutos de árboles de mango
plantados en los sitios de estudio.
Los valores de las medias con la misma letra no son significativamente diferentes, de
acuerdo con la prueba de ANOVA/ Tukey-Kramer (p = 0.05).
Sitio de
estudio
Hoja Fruto
Producción relativa
estimada, % N P N P
g kg-1
Tapachula 14.8ª 0.9ª 7.7ª 0.5ª 86
Chahuites 13.4b 1.0ª 6.2b 0.6ª 94
Apatzingán 15.1a 0.9a 4.9c 0.6ª 83

RES
ULT
AD
OS
Y D
ISC
USI
ÓN
Figura VIII.2. Curva de correlación entre el nitrógeno foliar y la producción relativa de
mango (%).
VIII.3. Aislamiento de microorganismos de interés en los sitios de estudio
y su caracterización metabólica - molecular
- Cepas bacterianas
En este estudio, siete aislados bacterianos identificados mediante la
amplificación del gen 16S ARNr (Fig. VIII.3), presentaron potenciales actividades
de promoción de crecimiento vegetal in vitro de acuerdo con las características
metabólicas evaluadas (Tabla VIII.3). Estas cepas fueron identificadas como
pertenecientes a diferentes géneros previamente reportados como Bacterias
Promotoras del Crecimiento Vegetal (BPCV) (Requena et al., 1997; Compant et
al., 2005; Anandham et al., 2008; Magnani et al., 2010), a través de diversos
mecanismos tales como: la fijación biológica de nitrógeno, la producción de
fitohormonas y sideróforos, la actividad ACC desaminasa y la solubilización de
fósforo (Ahemad y Saghir, 2011).Estas cepas fueron aisladas de suelo (5 cepas),
hojas (1 cepa), y frutos (1 cepa). Dos de ellas fueron aislados de Chahuites y cinco
de Apatzingán, lo que mostró una tendencia positiva entre el número de BPCV y la
fertilidad del suelo en dicho sitio de estudio (en comparación con Chahuites).

RES
ULT
AD
OS
Y D
ISC
USI
ÓN
Figura VIII.3. Relaciones filogenéticas de las BPCV obtenidas en este estudio,
inferidas por el análisis del gen 16S ARNr mediante el método de neighbor-joining.
Streptomyces sp.-EF157833 fue utilizado como el grupo externo. Los números mostrados
sobre las ramas indican el porcentaje de reensamble de 1000 bootstrap (solamente son
mostrados aquellos valores > 50). Los colores indican el origen de los aislados, azul:
Chahuites y rojo: Apatzingán.
XV
Burkholderia caribensis GU144372 Burkholderia caribensis Y17009 Burkholderia caribensis Y17010
XVI
Burkholderia cepacia HM224392
XXVI
Burkholderia cepacia GU048852
Burkholderia cepacia U96927
Uncultured Dyella sp. FJ536902
X
Uncultured Dyella sp. GU569103
Uncultured Dyella sp. GU563745
Pseudomonas fluorescens FJ950618
Pseudomonas fluorescens FJ950667
Pseudomonas fluorescens FJ950665
XX
Agrobacterium tumefaciens GQ140317
Agrobacterium tumefaciens EU592041 Agrobacterium tumefaciens GQ140318 XI
Rhizobium sp. AY141981
XXV Rhizobium sp. AY822761 Rhizobium sp. FJ025120
Agrobacterium rhizogenes GU580895
Agrobacterium rhizogenes GU552895
Agrobacterium rhizogenes GU320290
IV
Brevibacillus sp. GU213173
Brevibacillus sp. EU730938
Brevibacillus sp.
XXIX
Streptomyces sp. EF157833
100
100
100
98
88
100
100
97
100
79
74
100
100
100
92
100
63
100
78
63
100
100
84
100
63
55
Burk
hold
eri
a
carib
ensis
B
urk
hold
eri
a
cepacia
Unculture
d
Dyella
sp.
Pseu
dom
on
as
flu
ore
sce
ns
Agro
bacte
rium
tum
efa
cie
ns
R
hiz
ob
ium
sp.
A
gro
bacte
riu
m r
hiz
oge
nes
Bre
vib
acill
us
sp.
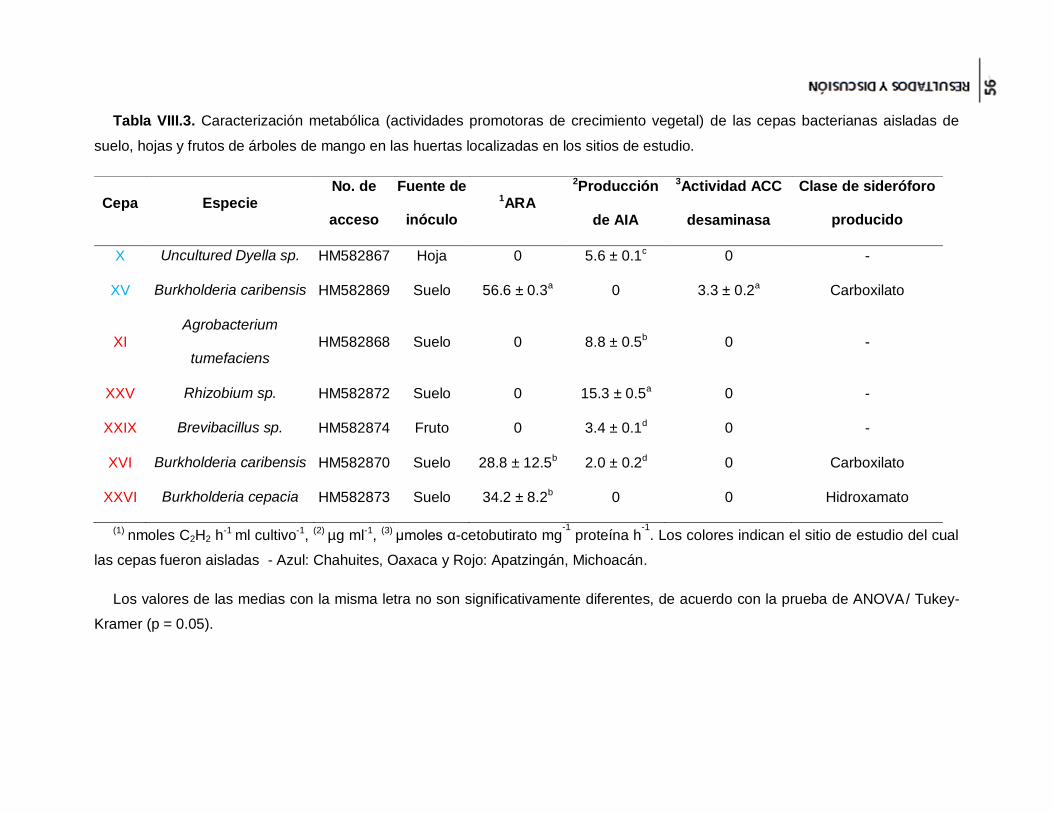
RES
ULT
AD
OS
Y D
ISC
USI
ÓN
Tabla VIII.3. Caracterización metabólica (actividades promotoras de crecimiento vegetal) de las cepas bacterianas aisladas de
suelo, hojas y frutos de árboles de mango en las huertas localizadas en los sitios de estudio.
Cepa Especie No. de
acceso
Fuente de
inóculo
1ARA
2Producción
de AIA
3Actividad ACC
desaminasa
Clase de sideróforo
producido
X Uncultured Dyella sp. HM582867 Hoja 0 5.6 ± 0.1c 0 -
XV Burkholderia caribensis HM582869 Suelo 56.6 ± 0.3a 0 3.3 ± 0.2a Carboxilato
XI
Agrobacterium
tumefaciens HM582868 Suelo 0 8.8 ± 0.5b 0 -
XXV Rhizobium sp. HM582872 Suelo 0 15.3 ± 0.5a 0 -
XXIX Brevibacillus sp. HM582874 Fruto 0 3.4 ± 0.1d 0 -
XVI Burkholderia caribensis HM582870 Suelo 28.8 ± 12.5b 2.0 ± 0.2d 0 Carboxilato
XXVI Burkholderia cepacia HM582873 Suelo 34.2 ± 8.2b 0 0 Hidroxamato
(1) nmoles C2H2 h-1 ml cultivo-1, (2) µg ml-1, (3) µmoles α-cetobutirato mg
-1 proteína h
-1. Los colores indican el sitio de estudio del cual
las cepas fueron aisladas - Azul: Chahuites, Oaxaca y Rojo: Apatzingán, Michoacán.
Los valores de las medias con la misma letra no son significativamente diferentes, de acuerdo con la prueba de ANOVA/ Tukey-
Kramer (p = 0.05).

RES
ULT
AD
OS
Y D
ISC
USI
ÓN
Dos cepas bacterianas Burkholderia caribensis XV y Rhizobium sp XXV,
mostraron promisorias capacidades de promoción de crecimiento vegetal, debido
a su actividad de reducción de acetileno, ACC desaminasa y producción de
sideróforo, así como la producción de ácido indol acético, respectivamente (Tabla
VIII.3). Estas actividades han sido reportadas como potenciadoras del crecimiento
vegetal cuando microorganismos que presentan estas características son
inoculadas a las plantas (Penrose y Glick, 2003; Onofre-Lemus et al., 2009).
La capacidad de control biológico de estas cepas contra el hongo causante de
la antracnosis fue explorada a través de ensayos de confrontación contra C.
gloeosporioides ATCC MYA 456, observando que solamente la cepa Burkholderia
cepacia XXVI presentó una significativa inhibición del crecimiento del patógeno
(Fig. VIII.4).
Los ensayos enfocados a conocer la naturaleza del metabolito responsable del
control biológico, evidenciaron que la producción de sideróforo (clase hidroxamato)
por la cepa XXVI fue el principal metabolito involucrado en dicho evento,
observando la inhibición del crecimiento del patógeno en la zona de acción del
sideróforo (halo naranja). De igual manera fue cuantificado el 91 % de inhibición
del crecimiento de C. gloeosporioides cuando este fue inoculado en cajas Petri
conteniendo PDA suplementado con 0.64 µg ml-1 del sideróforo producido por B.
cepacia XXVI (Fig. VIII.4).
El evento de control biológico contra C. gloeosporioides observado por la acción
del sideróforo producido por la cepa XXVI, puede ser explicada debido a que el
crecimiento del patógeno es dependiente de la disponibilidad de hierro en el
sistema. Por lo tanto, en un ambiente con limitación de este elemento, generado
por la acción de sideróforos, el crecimiento de C. gloeosporioides es inhibido, el
cual es restaurado cuando el hierro es añadido nuevamente al sistema (Santoyo et
al., 2010).

RES
ULT
AD
OS
Y D
ISC
USI
ÓN
Figura VIII.4. Control biológico de Burkholderia cepacia XXVI contra C.
gloeosporioides. [a) Inoculación de C. gloeosporioides (control negativo) y b) co-
inoculación del patógeno estudiado y B. cepacia XXVI (flechas), utilizando PDA como
medio de cultivo], [c) Inoculación de la cepa XXVI sobre una caja Petri conteniendo el
medio CAS- agar (el halo de coloración naranja evidencia la producción de sideróforo por
esta cepa) y d) co-inoculación de C. gloeosporioides y B. cepacia XXVI sobre cajas Petri
conteniendo CAS-agar]. La línea punteada indica el crecimiento radial del patógeno
esperado en ausencia de B. cepacia XXVI, [e) inoculación de C. gloeosporioides (control
negativo) y f) inoculación de C. gloeosporioides in cajas Petri conteniendo PDA
suplementado con 0.64 µg ml-1 del sideróforo producido por B. cepacia XXVI].
a) b)
c) d)
e) f)

RES
ULT
AD
OS
Y D
ISC
USI
ÓN
- Cepas de Trichoderma
Un total de veinte aislados pertenecientes al género Trichoderma [identificación
macroscópica, microscópica y molecular (utilizando oligonucleótidos específicos
para este género)] fueron obtenidos en el presente trabajo.
Estos aislados fueron identificados molecularmente por PCR, mediante la
amplificación del espaciador intergénico transcribible (ITS) (Fig. VIII.5), así como
caracterizados in vitro en relación a sus capacidades promotoras de crecimiento
vegetal y como agente de control biológico contra C. gloeosporioides (Tabla VIII.4
y Tabla VIII.5). El mayor número de aislados de Trichoderma fueron obtenidas de
suelo (80%), el 20% restante de hojas (obtenidos solamente en Chahuites); no
fueron obtenidos aislados de frutos, probablemente porque el suelo, a diferencia
de las hojas y frutos, es un hábitat complejo que permite el establecimiento de los
microorganismos, debido a los altos niveles de nutrientes y estrategias de
protección (estrés biótico y abiótico) que el éste presenta comparado con las hojas
y frutos (Garbeva et al., 2008). El mismo número de aislados fueron obtenidos
independientemente del medio de cultivo utilizado (PDA o TSM).
En este estudio, altos niveles de la fertilidad del suelo mostraron una tendencia
negativa en relación con el número de aislados de Trichoderma. Barabasz et al.
(2002) reportaron observaciones similares, sugiriendo que la generación de
compuestos tóxicos (nitrosamina) en el suelo cuando grandes cantidades de
nitrógeno son aplicadas tuvieron un impacto negativo en la población de hongos,
observando que la aplicación de 360 kg N ha-1 año-1 (65.3 g de nitrosaminas g-1
de suelo ) disminuyó la población de hongos en el suelo en comparación con la
aplicación de 240 kg de N ha-1 año-1 (18.4 g de nitrosamina g-1 de suelo), lo que
condujo a 27.5 y 32.4 x 103 UFC g-1 de suelo, respectivamente.
La identificación molecular de los aislados de Trichoderma basado en el ITS
(Fig. VIII.5), reveló que éstos pertenecen a las siguientes especies: Hypocrea lixii,
Hypocrea jecorina, Trichoderma asperellum, Trichoderma spirale y Trichoderma

RES
ULT
AD
OS
Y D
ISC
USI
ÓN
brevicompactum. La especie Hypocrea lixii fue aislada en todos los sitios de
estudio; sin embargo, algunas especies fueron específicas de un sitio de estudio,
por ejemplo, T. asperellum en Tapachula, T. spirale en Chahuites y T.
brevicompactum y H. jecorina en Apatzingán. El género Hypocrea es el estado
sexual (teleomorfo) del género Trichoderma (estado asexual, anamorfo). Este
género se ha reconocido por estar constituidos por hongos cosmopolitas del suelo,
colonizando una amplia gama de hábitats y nichos ecológicos (Schuster y
Schmoll, 2010). Además, el aislamiento de especies de Trichoderma específicas
de cada sitio de estudio, puede ser explicado por los parámetros ambientales tales
como temperatura, humedad, pH, materia orgánica y el contenido de nutrientes
presentes en los sistemas (Eastburn y Butler, 1991; Carreiro y Koske, 1992; Klein
y Eveleigh, 1998).
El género Trichoderma comprende un gran número de cepas que actúan como
agentes de control biológico a través de la activación de los mecanismos directos
o indirectos (Lo, 1998). En relación a las capacidades de control biológico de
Trichoderma contra C. gloeosporioides ATCC MYA 456, en el presente estudio
tendencias específicas del sitio de estudio fueron observadas: cepas aisladas de
Tapachula (T. asperellum T7a, T8a y T8c) fueron las únicas que se agruparon en
la clase 1 (Tabla VIII.4), debido a que estas cepas sobrecrecieron al patógeno
(Bell et al., 1982.), probablemente a través del micoparasitismo (el principal
mecanismo directo estudiado en esta especie) (Ramot et al., 2004; Sanz et al.,
2005; González et al., 2012.). Trichoderma asperellum ha sido reportado como un
agente de control biológico contra diversos hongos fitopatógenos, considerándose
un antagonista versátil (Watanabe, 2005; Guigón-López et al., 2010). Todas las
otras cepas obtenidas en este estudio, fueron agrupadas en la clase 2,
observando la competencia por nutrientes y espacio como su principal
mecanismo. Ninguna de las cepas fueron ubicadas en las clases 3, 4, y 5 (Tabla
VIII.4), esto puede ser explicado, como se muestra en la Tabla VIII.5, por la rápida
tasa de crecimiento de estas cepas (Schuster y Schmoll, 2010).
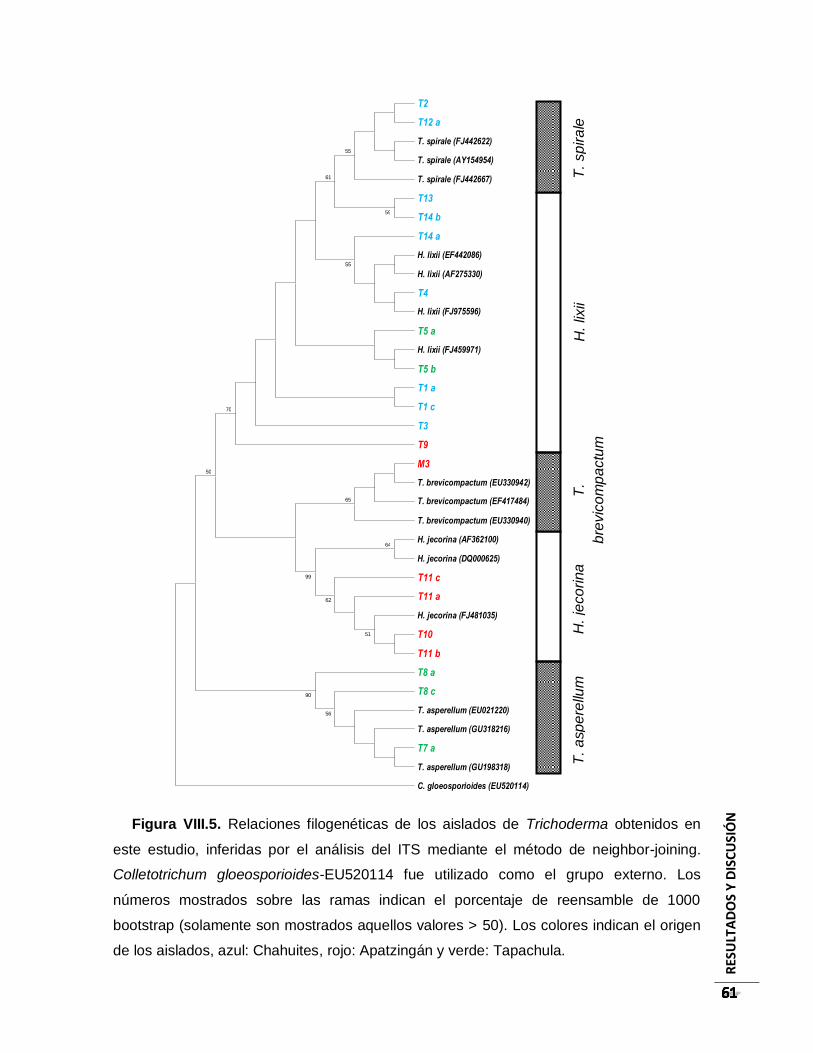
RES
ULT
AD
OS
Y D
ISC
USI
ÓN
Figura VIII.5. Relaciones filogenéticas de los aislados de Trichoderma obtenidos en
este estudio, inferidas por el análisis del ITS mediante el método de neighbor-joining.
Colletotrichum gloeosporioides-EU520114 fue utilizado como el grupo externo. Los
números mostrados sobre las ramas indican el porcentaje de reensamble de 1000
bootstrap (solamente son mostrados aquellos valores > 50). Los colores indican el origen
de los aislados, azul: Chahuites, rojo: Apatzingán y verde: Tapachula.
T2 T12 a
T. spirale (FJ442622)
T. spirale (AY154954)
T. spirale (FJ442667)
T13 T14 b T14 a H. lixii (EF442086)
H. lixii (AF275330)
T4 H. lixii (FJ975596)
T5 a H. lixii (FJ459971)
T5 b T1 a T1 c T3 T9 M3 T. brevicompactum (EU330942)
T. brevicompactum (EF417484)
T. brevicompactum (EU330940)
H. jecorina (AF362100)
H. jecorina (DQ000625)
T11 c T11 a H. jecorina (FJ481035)
T10 T11 b T8 a T8 c T. asperellum (EU021220)
T. asperellum (GU318216)
T7 a T. asperellum (GU198318)
C. gloeosporioides (EU520114)
56
90
64
51
62
99
65
50
59
70
55
61
55
T.
sp
ira
le
H.
lixii
T.
bre
vic
om
pa
ctu
m
H.
jeco
rina
T.
asp
ere
llum

RES
ULT
AD
OS
Y D
ISC
USI
ÓN
Tabla VIII.4. Grados de cobertura de los aislados de Trichoderma contra C.
gloeosporioides ATCC MYA 456 utilizando la técnica de confrontación in vitro.
Una clase es una escala del 1 al 5 que representan diferentes capacidades de
sobrecrecimiento de los aislados de Trichoderma, obtenidos en este trabajo, contra el
patógeno estudiado. La capacidad de sobrecrecimiento describe las características del
control biológico de los aislados de Trichoderma contra C. gloeosporioides. El número de
aislados indica la cantidad ellos que presentaron una capacidad sobrecrecimiento
específica, representada por un valor en la escala de clase.
Clase Capacidad de
sobrecrecimiento
Número de
aislados
1 Trichoderma sobrecreció completamente al
patógeno y la superficie del medio de cultivo
3
2 Trichoderma sobrecreció al menos 2/3 de la
superficie del medio
17
3 Trichoderma y el patógeno colonizaron
aproximadamente 1/2 de la superficie del medio
0
4 El patógeno colonizó al menos 2/3 de la
superficie del medio
0
5 El patógeno sobrecreció completamente a
Trichoderma y la superficie del medio de cultivo
0
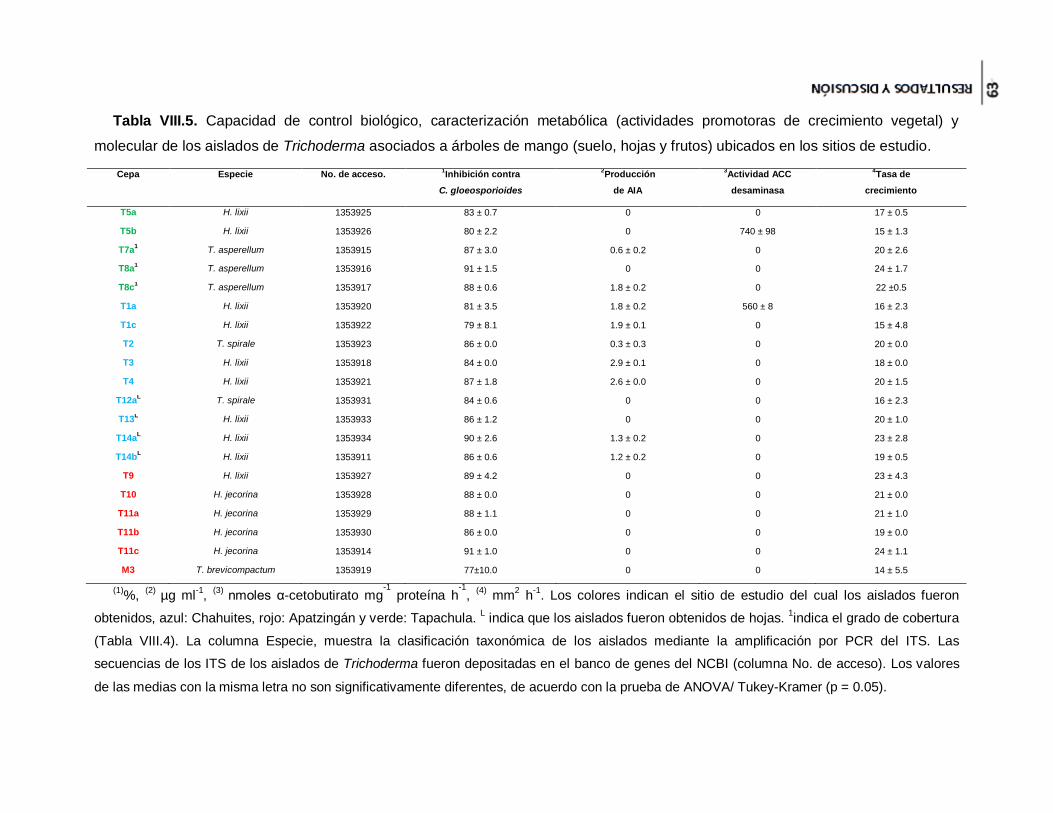
RES
ULT
AD
OS
Y D
ISC
USI
ÓN
Tabla VIII.5. Capacidad de control biológico, caracterización metabólica (actividades promotoras de crecimiento vegetal) y
molecular de los aislados de Trichoderma asociados a árboles de mango (suelo, hojas y frutos) ubicados en los sitios de estudio.
(1)%,
(2) µg ml
-1,
(3) nmoles α-cetobutirato mg
-1 proteína h
-1,
(4) mm
2 h
-1. Los colores indican el sitio de estudio del cual los aislados fueron
obtenidos, azul: Chahuites, rojo: Apatzingán y verde: Tapachula. L indica que los aislados fueron obtenidos de hojas.
1indica el grado de cobertura
(Tabla VIII.4). La columna Especie, muestra la clasificación taxonómica de los aislados mediante la amplificación por PCR del ITS. Las
secuencias de los ITS de los aislados de Trichoderma fueron depositadas en el banco de genes del NCBI (columna No. de acceso). Los valores
de las medias con la misma letra no son significativamente diferentes, de acuerdo con la prueba de ANOVA/ Tukey-Kramer (p = 0.05).
Cepa Especie No. de acceso. 1Inhibición contra
C. gloeosporioides
2Producción
de AIA
3Actividad ACC
desaminasa
4Tasa de
crecimiento
T5a H. lixii 1353925 83 ± 0.7 0 0 17 ± 0.5
T5b H. lixii 1353926 80 ± 2.2 0 740 ± 98 15 ± 1.3
T7a1 T. asperellum 1353915 87 ± 3.0 0.6 ± 0.2 0 20 ± 2.6
T8a1 T. asperellum 1353916 91 ± 1.5 0 0 24 ± 1.7
T8c1 T. asperellum 1353917 88 ± 0.6 1.8 ± 0.2 0 22 ±0.5
T1a H. lixii 1353920 81 ± 3.5 1.8 ± 0.2 560 ± 8 16 ± 2.3
T1c H. lixii 1353922 79 ± 8.1 1.9 ± 0.1 0 15 ± 4.8
T2 T. spirale 1353923 86 ± 0.0 0.3 ± 0.3 0 20 ± 0.0
T3 H. lixii 1353918 84 ± 0.0 2.9 ± 0.1 0 18 ± 0.0
T4 H. lixii 1353921 87 ± 1.8 2.6 ± 0.0 0 20 ± 1.5
T12aL T. spirale 1353931 84 ± 0.6 0 0 16 ± 2.3
T13L H. lixii 1353933 86 ± 1.2 0 0 20 ± 1.0
T14aL H. lixii 1353934 90 ± 2.6 1.3 ± 0.2 0 23 ± 2.8
T14bL H. lixii 1353911 86 ± 0.6 1.2 ± 0.2 0 19 ± 0.5
T9 H. lixii 1353927 89 ± 4.2 0 0 23 ± 4.3
T10 H. jecorina 1353928 88 ± 0.0 0 0 21 ± 0.0
T11a H. jecorina 1353929 88 ± 1.1 0 0 21 ± 1.0
T11b H. jecorina 1353930 86 ± 0.0 0 0 19 ± 0.0
T11c H. jecorina 1353914 91 ± 1.0 0 0 24 ± 1.1
M3 T. brevicompactum 1353919 77±10.0 0 0 14 ± 5.5

RES
ULT
AD
OS
Y D
ISC
USI
ÓN
Dos cepas, Trichoderma asperellum T8a e Hypocrea lixii, presentaron
características promisorias en relación al control biológico contra C.
gloeosporioides y la promoción de crecimiento vegetal observadas in vitro,
respectivamente (Tabla VIII.5).
Los experimentos enfocados a identificar el principal metabolito involucrado en
el evento de control biológico de la cepa T. asperellum T8a contra C.
gloeosporioides ATCC MYA 456, mostraron que el micelio del patógeno solamente
fue lisado en el tratamiento 4 (Fig. VIII.6), lo que sugiere que su muerte en este
tratamiento fue debida a la actividad de las celulasas (2.9 ± 0.7 U).
Las otras actividades enzimáticas (quitinasas o glucanasas) detectadas en los
tratamientos 1, 2, 3, y 5, no mostraron efecto sobre el micelio del patógeno. Los
datos muestran que el complejo celulolítico (2.9 ± 0.7 U) fue significativamente
efectivo en el control biológico de C. gloeosporioides ATCC MYA 456, comparado
con las otras actividades individuales estudiadas (Tabla VIII.6 y Fig. VIII.6).
Observaciones similares han sido descritas por Haran et al. (1996), quienes
reportaron que el mecanismo micoparasítico de Trichoderma está asociado a la
actividad de enzimas líticas de éste género.
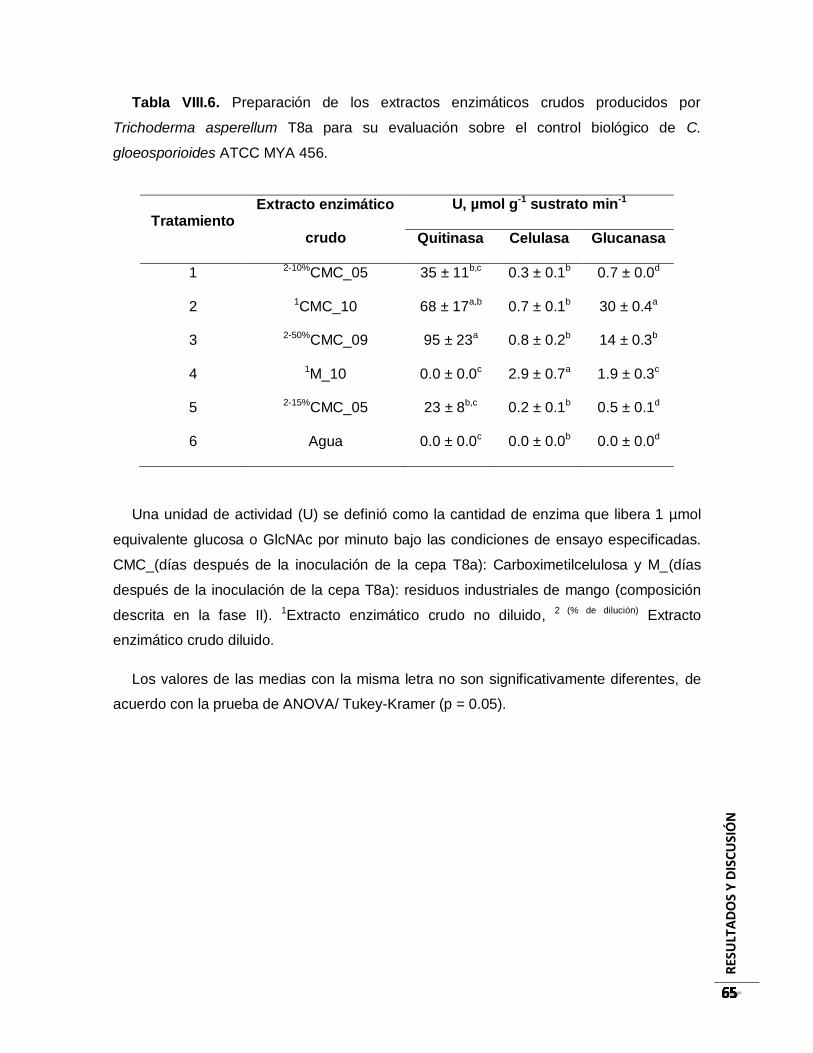
RES
ULT
AD
OS
Y D
ISC
USI
ÓN
Tabla VIII.6. Preparación de los extractos enzimáticos crudos producidos por
Trichoderma asperellum T8a para su evaluación sobre el control biológico de C.
gloeosporioides ATCC MYA 456.
Tratamiento Extracto enzimático
crudo
U, µmol g-1 sustrato min-1
Quitinasa Celulasa Glucanasa
1 2-10%CMC_05 35 ± 11b,c 0.3 ± 0.1b 0.7 ± 0.0d
2 1CMC_10 68 ± 17a,b 0.7 ± 0.1b 30 ± 0.4a
3 2-50%CMC_09 95 ± 23a 0.8 ± 0.2b 14 ± 0.3b
4 1M_10 0.0 ± 0.0c 2.9 ± 0.7a 1.9 ± 0.3c
5 2-15%CMC_05 23 ± 8b,c 0.2 ± 0.1b 0.5 ± 0.1d
6 Agua 0.0 ± 0.0c 0.0 ± 0.0b 0.0 ± 0.0d
Una unidad de actividad (U) se definió como la cantidad de enzima que libera 1 µmol
equivalente glucosa o GlcNAc por minuto bajo las condiciones de ensayo especificadas.
CMC_(días después de la inoculación de la cepa T8a): Carboximetilcelulosa y M_(días
después de la inoculación de la cepa T8a): residuos industriales de mango (composición
descrita en la fase II). 1Extracto enzimático crudo no diluido, 2 (% de dilución) Extracto
enzimático crudo diluido.
Los valores de las medias con la misma letra no son significativamente diferentes, de
acuerdo con la prueba de ANOVA/ Tukey-Kramer (p = 0.05).

RES
ULT
AD
OS
Y D
ISC
USI
ÓN
Figura VIII.6. Observación microscópica de células viables [las mitocondrias, indicadas
por flechas, redujeron activamente el 2, 3, 5-trifeniltetrazolio (incoloro) a 1, 3, 5-
trifenilformazán (color rojo)] y lisis celular, indicada con flechas punteadas, de C.
gloeosporioides ATCC MYA 456 después de su incubación con diferentes actividades
enzimáticas producidas por T. asperellum T8a. Los tratamientos evaluados fueron (Tabla
VIII.6): a) 2, b) 4 c) 5, y d) 6.
- Cepas de Colletotrichum gloeosporioides
Con base en las características macroscópicas, microscópicas y moleculares
(utilizando oligonucleótidos específicos para la especie estudiada) cinco aislados
pertenecientes a la especie C. gloeosporioides fueron identificadas. Éstas
solamente fueron aisladas de frutos colectados en Chahuites. Las características
macroscópicas y moleculares evidenciaron la existencia de al menos dos cepas
(Fig. VIII.7).
a)
b)
c)
d)
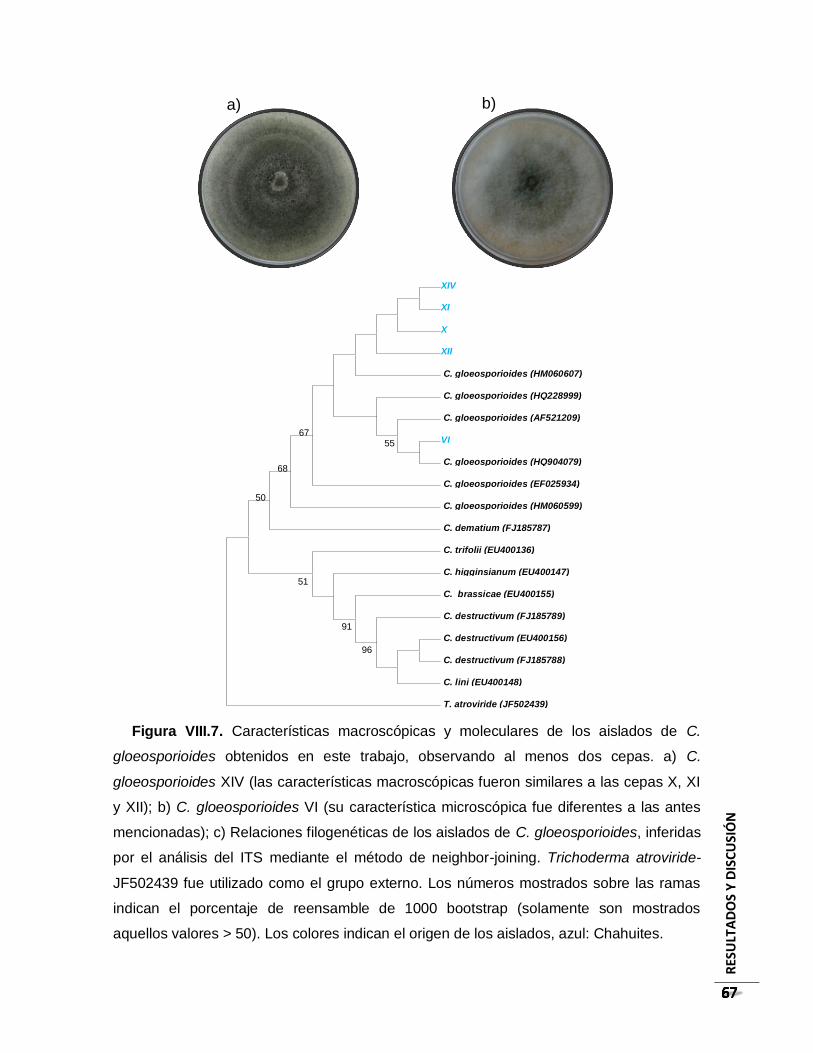
RES
ULT
AD
OS
Y D
ISC
USI
ÓN
Figura VIII.7. Características macroscópicas y moleculares de los aislados de C.
gloeosporioides obtenidos en este trabajo, observando al menos dos cepas. a) C.
gloeosporioides XIV (las características macroscópicas fueron similares a las cepas X, XI
y XII); b) C. gloeosporioides VI (su característica microscópica fue diferentes a las antes
mencionadas); c) Relaciones filogenéticas de los aislados de C. gloeosporioides, inferidas
por el análisis del ITS mediante el método de neighbor-joining. Trichoderma atroviride-
JF502439 fue utilizado como el grupo externo. Los números mostrados sobre las ramas
indican el porcentaje de reensamble de 1000 bootstrap (solamente son mostrados
aquellos valores > 50). Los colores indican el origen de los aislados, azul: Chahuites.
XIV
XI
X
XII
C. gloeosporioides (HM060607)
C. gloeosporioides (HQ228999)
C. gloeosporioides (AF521209)
VI
C. gloeosporioides (HQ904079)
C. gloeosporioides (EF025934)
C. gloeosporioides (HM060599)
C. dematium (FJ185787)
C. trifolii (EU400136)
C. higginsianum (EU400147)
C. brassicae (EU400155)
C. destructivum (FJ185789)
C. destructivum (EU400156)
C. destructivum (FJ185788)
C. lini (EU400148)
T. atroviride (JF502439)
96
91
55 67
68
51
50
a) b)

RES
ULT
AD
OS
Y D
ISC
USI
ÓN
Los ensayos de patogenicidad demostraron que todas las cepas aisladas
causaron antracnosis cuando fueron inoculadas en frutos de mango, 100% de
frutos infectados (Fig. VIII.8), sugiriendo su contribución en las importantes
pérdidas de la producción de mango por la antracnosis, cuando las condiciones en
el campo son las óptimas para su desarrollo (Arauz, 2000; Singh, 2011).
Figura VIII.8. Ensayo de patogenicidad de las cepas de C. gloeosporioides obtenidas
en este trabajo. a) Control negativo (agua), b) Inoculación de C. gloeosporioides XV,
observando lesiones necróticas típicas por la infección de este patógeno, causando la
antracnosis (flechas blancas).
Selección de los microorganismos usados en la FASE II y FASE III
Con base a las características promotoras de crecimiento vegetal de las cepas
bacterianas y de Trichoderma, Burkholderia caribensis XV y Rhizobium sp. XXV
fueron seleccionadas, debido a sus características metabólicas versátiles, tales
como: ARA-ACC desaminasa-sideróforo y producción de IAA, respectivamente
(Tabla VIII.3), para el diseño de un manejo de inoculación microbiana a aplicar a
los árboles de mango creciendo en el invernadero, con el objetivo de cuantificar el
impacto de dicho manejo sobre el evento de floración estos árboles (FASE I).
Además, la producción de IAA, así de cinetina y AG3 por la cepa XXV fue
confirmado por ensayos de HPLC, cuantificando una producción en g ml-1 de 18.9
± 0.1, 21.8 ± 0.1 y 274.4 ± 0.7, respectivamente.
En relación al agente de control biológico contra C. gloeosporioides para ser
producido utilizando los residuos industriales de mango (FASE II), tanto
a b

RES
ULT
AD
OS
Y D
ISC
USI
ÓN
Burkholderia cepacia XXVI como Trichoderma asperellum T8a, mostraron ser
eficientes en el control de este patógeno (cepa ATCC MYA 456 y los aislados de
C. gloeosporioides obtenidos en este trabajo), a través de la producción de
sideróforo y de enzimas líticas, respectivamente (Fig.VIII.4; Tabla VIII.6 y Fig.
VIII.6). Sin embargo, la cepa T. asperellum T8a fue seleccionada para la FASE II,
debido a que la cepa B. cepacia XXVI, debido a su alto porcentaje de similitud
(99%) de la secuencia del gen 16S ARNr, es un miembro del Complejo
Burkholderia cepacia, lo que sugiere que probablemente esta cepa sea patógena
de humanos (Coenye et al., 2001).
Ensayos enfocados a evaluar la capacidad de control biológico de T. asperellum
T8a contra C. gloeosporioides in vivo resultaron en 20, 0, 0, 20, 20, 0% de frutos
infectados por antracnosis cuando las cepas ATCC MYA 456, VI, X, XI, XII, XIV
del patógeno fueron inoculadas, respectivamente. El tratamiento control (sin
inoculación del patógeno) mostró 0% de frutos enfermos; sin embargo, el
tratamiento inoculado únicamente con las cepas de C. gloeosporioides mostró el
100% de frutos enfermos. Los tratamientos en los que el 20% de los frutos
desarrollaron antracnosis, la severidad de la enfermedad fue menor que el daño
ocasionado por la inoculación de las cepas de C. gloeosporioides (Fig. VIII.9).
Figura VIII.9. Severidad del daño antracnosis (indicado por flechas blancas) en un
tratamiento con 20% de frutos enfermos. a) Control (no inoculado con el patógeno); b)
Inoculación de C. gloeosporioides VI; c) Inoculación de C. gloeosporioides VI y T.
asperellum T8a).
a b c
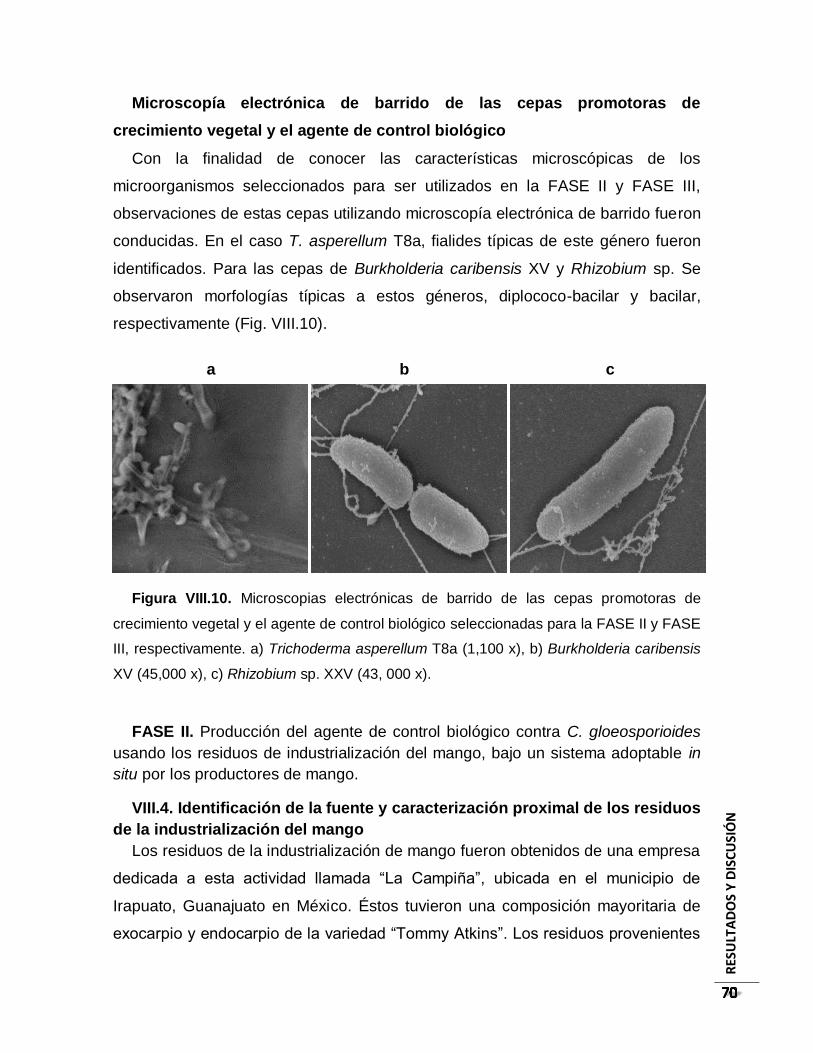
RES
ULT
AD
OS
Y D
ISC
USI
ÓN
Microscopía electrónica de barrido de las cepas promotoras de
crecimiento vegetal y el agente de control biológico
Con la finalidad de conocer las características microscópicas de los
microorganismos seleccionados para ser utilizados en la FASE II y FASE III,
observaciones de estas cepas utilizando microscopía electrónica de barrido fueron
conducidas. En el caso T. asperellum T8a, fialides típicas de este género fueron
identificados. Para las cepas de Burkholderia caribensis XV y Rhizobium sp. Se
observaron morfologías típicas a estos géneros, diplococo-bacilar y bacilar,
respectivamente (Fig. VIII.10).
Figura VIII.10. Microscopias electrónicas de barrido de las cepas promotoras de
crecimiento vegetal y el agente de control biológico seleccionadas para la FASE II y FASE
III, respectivamente. a) Trichoderma asperellum T8a (1,100 x), b) Burkholderia caribensis
XV (45,000 x), c) Rhizobium sp. XXV (43, 000 x).
FASE II. Producción del agente de control biológico contra C. gloeosporioides
usando los residuos de industrialización del mango, bajo un sistema adoptable in
situ por los productores de mango.
VIII.4. Identificación de la fuente y caracterización proximal de los residuos
de la industrialización del mango
Los residuos de la industrialización de mango fueron obtenidos de una empresa
dedicada a esta actividad llamada “La Campiña”, ubicada en el municipio de
Irapuato, Guanajuato en México. Éstos tuvieron una composición mayoritaria de
exocarpio y endocarpio de la variedad “Tommy Atkins”. Los residuos provenientes
a b c

RES
ULT
AD
OS
Y D
ISC
USI
ÓN
de la industrialización del mango fueron secados a temperatura ambiente y
molidos hasta un tamaño de partícula menor a 800 µm. Los cuales fueron
almacenados a 28º C en frasco cerrados hasta su utilización.
En la actualidad, la producción de agentes de control biológico es
económicamente no sustentable debido a: i) la tecnología limitada (las empresas
tienen que modificar sus sistemas de producción de fungicidas sintéticos a
sistemas de producción de microorganismos) y ii) el alto costo económico de los
sustratos utilizados en la producción de dichos agentes (Soccol y Vandenberghe,
2003). Por lo tanto, estudios centrados en la producción de agentes de control
biológico utilizando sustratos económicos y de fácil adquisición bajo sistemas
simples es determinante para el uso extensivo de estos microorganismos,
disminuyendo los problemas ambientales y económicos por el uso excesivo de
fungicidas sintéticos.
En el presente trabajo, los residuos generados en la industrialización del mango
fueron estudiados con la finalidad de estudiar su uso potencial como sustratos en
la producción de esporas, bajo una sencilla fermentación en estado sólido, de un
agente de control biológico como lo es Trichoderma asperellum T8a, el cual fue
aislado de la rizósfera de árboles de mango en huertas localizadas en Tapachula,
Chiapas (FASE I). El análisis proximal de los residuos industriales del mango
(Tabla VIII.7) reveló que éstos son sustratos adecuados y sostenibles para la
producción de microorganismos de importancia agrícola, debido a sus altos
contenidos de carbohidratos solubles y disponibles (extracto libre de nitrógeno),
los cuales pueden ser utilizados para el crecimiento de hongos de interés, tales
como T. asperellum T8a. Esta especie ha sido reconocida por su amplia
capacidad metabólica conduciendo al uso de varios residuos agrícolas para su
crecimiento (Trillas et al., 2006). Caracterizaciones proximales similares han sido
reportados para los residuos de paja de trigo, repollo, plátano, mazorca de maíz, y
las uvas (Chahal et al., 1996;. Ojumu et al., 2003;. Baig, 2005; Bai et al., 2008;.
Das y Ghosh, 2009).

RES
ULT
AD
OS
Y D
ISC
USI
ÓN
Tabla VIII.7. Caracterización proximal de los residuos provenientes de la
industrialización de mango “Tommy Atkins” (exocarpio y endocarpio).
Determinación Peso fresco, %
Humedad 79 ± 1
Sólidos totales 21 ± 1
Cenizas 0.6 ± 0.0
Proteína 1.9 ± 0.0
Grasas 0.3 ± 0.1
Extracto libre de nitrógeno 15 ± 2
Fibra 3.4 ± 0.2
Celulosa 2.1 ± 0.1
Hemicelulosa 0.7 ± 0.1
Lignina 0.6 ± 0.0
Materia orgánica 18 ± 0.5
Carbono 10 ± 0.3
Nitrógeno 0.3 ± 0.0
Relación C: N 33 ± 1
pH 4.0 ± 0.0
La celulosa, que está presente en estos residuos, es un importante sustrato, ya
que las celulasas excretadas por hongos pueden, a partir de este polisacárido,
liberar glucosa y consumirla para su crecimiento, cuando los carbohidratos
solubles y disponibles en este sistema han sido consumidos por el mismo
microorganismo (Jackson y Bothast, 1990). La actividad celulolítica en los
sobrenadantes de T. asperellum T8a fue observada a los dos días después de la
inoculación, pero se mantuvo baja cuando la celulosa fue la única fuente de
carbono. Dicha actividad alcanzó otro pico en el séptimo día (Fig. VIII.11),
pudiéndose explicar por la inhibición de la actividad celulolítica (del día 3 al 6)
debido a la glucosa liberada de la celulosa en el segundo día, detectando esta
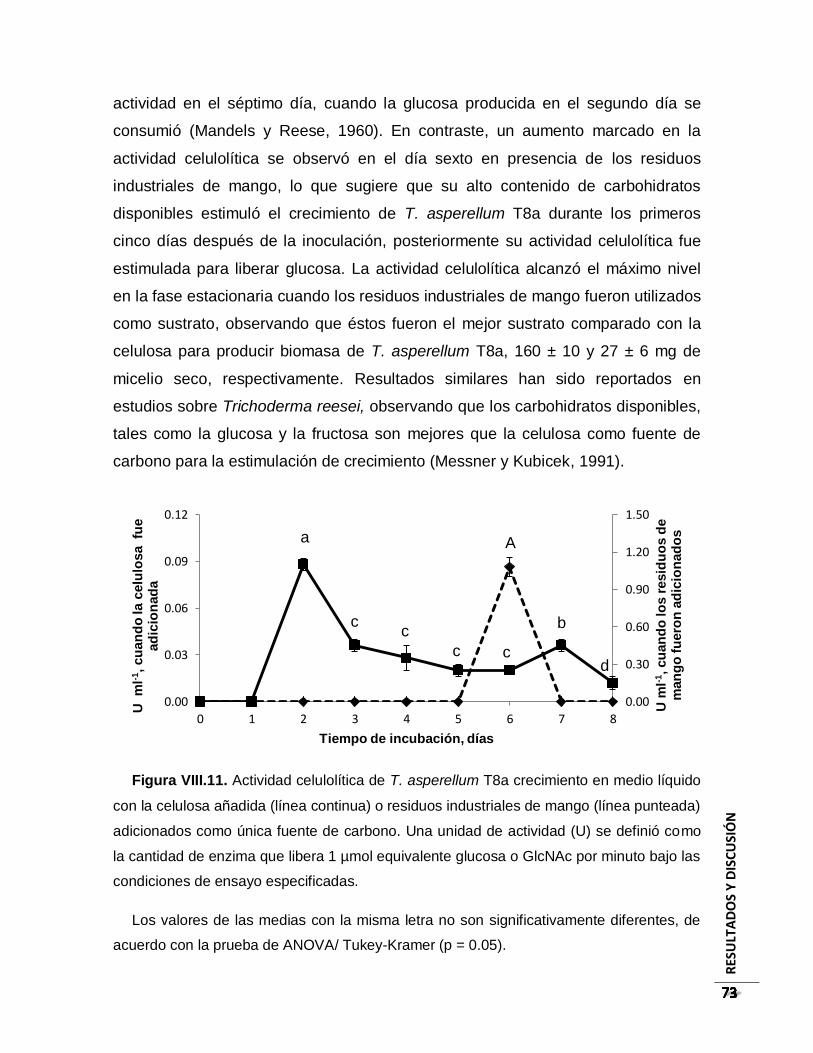
RES
ULT
AD
OS
Y D
ISC
USI
ÓN
actividad en el séptimo día, cuando la glucosa producida en el segundo día se
consumió (Mandels y Reese, 1960). En contraste, un aumento marcado en la
actividad celulolítica se observó en el día sexto en presencia de los residuos
industriales de mango, lo que sugiere que su alto contenido de carbohidratos
disponibles estimuló el crecimiento de T. asperellum T8a durante los primeros
cinco días después de la inoculación, posteriormente su actividad celulolítica fue
estimulada para liberar glucosa. La actividad celulolítica alcanzó el máximo nivel
en la fase estacionaria cuando los residuos industriales de mango fueron utilizados
como sustrato, observando que éstos fueron el mejor sustrato comparado con la
celulosa para producir biomasa de T. asperellum T8a, 160 ± 10 y 27 ± 6 mg de
micelio seco, respectivamente. Resultados similares han sido reportados en
estudios sobre Trichoderma reesei, observando que los carbohidratos disponibles,
tales como la glucosa y la fructosa son mejores que la celulosa como fuente de
carbono para la estimulación de crecimiento (Messner y Kubicek, 1991).
Figura VIII.11. Actividad celulolítica de T. asperellum T8a crecimiento en medio líquido
con la celulosa añadida (línea continua) o residuos industriales de mango (línea punteada)
adicionados como única fuente de carbono. Una unidad de actividad (U) se definió como
la cantidad de enzima que libera 1 µmol equivalente glucosa o GlcNAc por minuto bajo las
condiciones de ensayo especificadas.
Los valores de las medias con la misma letra no son significativamente diferentes, de
acuerdo con la prueba de ANOVA/ Tukey-Kramer (p = 0.05).
0.00
0.30
0.60
0.90
1.20
1.50
0.00
0.03
0.06
0.09
0.12
0 1 2 3 4 5 6 7 8
U m
l-1, cu
an
do
lo
s r
esid
uo
s d
e
man
go
fu
ero
n a
dic
ion
ad
os
U m
l-1, cu
an
do
la c
elu
losa fu
e
ad
icio
nad
a
Tiempo de incubación, días
a
b c c
c c d
A

RES
ULT
AD
OS
Y D
ISC
USI
ÓN
VIII.5. Diseño de un sistema de fermentación para la producción del agente
de control biológico, seleccionado en la fase I, contra C. gloeosporioides
La información mencionada anteriormente mostró que T. asperellum T8a fue
capaz de crecer utilizando los residuos industriales de mango como sustrato en
una fermentación en estado líquido, pero sin la producción de esporas (la principal
forma de aplicación de los agentes de control biológico). Por lo tanto, la
producción de esporas de esta cepa utilizando estos residuos industriales bajo una
fermentación en estado sólido, potencialmente adoptable por los productores de
mango, fue evaluada. Este sistema de fermentación conduce a una serie de
ventajas biotecnológicas. Por ejemplo, alta productividad de fermentación y
estabilidad de los productos comparado con otros sistemas, baja represión
catabólica y demanda de agua (Hölker et al., 2004).
Nuestro sistema de fermentación en estado sólido fue optimizado sobre todo en
aquellos parámetros determinantes para la aplicación potencial de esta alternativa
in situ (huertas de mango), con el objetivo de disminuir el elevado costo
económico, mencionado anteriormente, de la producción de esporas de agentes
de control biológico y de su alternativa sintética. Por esta razón, la relación C: N,
contenido de humedad, y el tiempo óptimo para la cosecha de esporas, en
relación a los resultados del análisis proximal de estos residuos, fueron
estudiados. El mayor número de esporas (de 2.5 x 106 hasta 57 ± 3 x 108 esporas
g-1 ss) fueron obtenidas con una relación C: N de 26, después de 6 días de
incubación y un contenido de humedad del 88% (Tabla VIII.8). Está relación C: N
ha sido reportada como un valor medio para el crecimiento y esporulación de
varias cepas de Trichoderma harzianum (Serrano-Carreón et al., 1992; Agosin et
al., 1997; Dahlan, 2007). Resultados similares a los obtenidos en nuestro
experimento indican que la relación C: N es un factor determinante afectando la
esporulación fúngica (Elson et al., 1998), siendo en muchos casos, el valor óptimo
de la relación C: N para la esporulación diferente al valor óptimo para el
crecimiento micelial, debido a que estos valores son dependientes de las especies
y/ o cepas de los hongos (Gao y Liu, 2009). Trichoderma asperellum T8a fue
incapaz de crecer, bajo las mismas condiciones mencionadas anteriormente

RES
ULT
AD
OS
Y D
ISC
USI
ÓN
usando la cáscara seca entera como sustrato, probablemente debido a que los
residuos utilizados, con tamaño de partícula menor a 800 µm, proporciona una
mayor superficie de contacto, incrementando la absorción de nutrientes por la
cepa T8a (Maurya et al., 2012). De manera similar, la actividad celulolítica más
alta fue observada en las relaciones C: N entre 22 y 30 (Tabla VIII.8), lo que
sugiere que la degradación de la celulosa es un factor determinante en la
inducción del crecimiento micelial y la producción de esporas, cuando el contenido
de carbohidratos disponibles ha sido disminuida por el crecimiento de la cepa T8a
(Ellouz et al., 1995; Xia y Shen, 2004) (Fig. VIII.12).
Tabla VIII.8. Producción de esporas y actividad celulolítica de T. asperellum T8a
creciendo bajo una fermentación en estado sólido, bajo diferentes relaciones C: N,
después de 6 días de incubación y un contenido de humedad del 88%.
Relación
C:N
Producción de esporas,
esporas x 108 g
-1 ss
Actividad celulolítica,
U g-1
ss
6 0.03 ± 0.01f 0.8 ± 0.1
b
10 14 ± 3e 0.2 ± 0.1
c
14 28 ± 1d 0.3 ± 0.1c
18 29 ± 3c,d
0.2 ± 0.0c
22 43 ± 0.3b 1.6 ± 0.1
a
26 57 ± 3a 1.0 ± 0.1
b
30 38 ± 4b,c
1.8 ± 0.2a
34 2.4 ± 0.8f 0.3 ± 0.0
c
Una unidad de actividad (U) se definió como la cantidad de enzima que libera 1 µmol
equivalente glucosa o GlcNAc por minuto bajo las condiciones de ensayo especificadas.
Los valores de las medias con la misma letra no son significativamente diferentes, de
acuerdo con la prueba de ANOVA/ Tukey-Kramer (p = 0.05).

RES
ULT
AD
OS
Y D
ISC
USI
ÓN
Figura VIII.12. Trichoderma asperellum T8a creciendo bajo diferentes relaciones de C:
N en una fermentación en estado sólido utilizando los residuos industriales de mango
como sustrato, seis días después de la inoculación y contenido de humedad del 88%.
Relaciones C: N a) 6, b) 14, c) 26 y d) 34.
Una vez que la relación C: N se fijó en 26, el contenido de humedad fue
optimizado en el sexto día. El contenido de humedad es un factor crítico para el
crecimiento celular y la producción de esporas en fermentaciones en estado
sólido. Además, en este tipo de fermentaciones, la cantidad de agua en el sistema
depende de la capacidad de retención de agua del sustrato utilizado. Esta
cantidad debe ser suficiente para el crecimiento de microorganismos a través de la
movilización de enzimas, nutrientes y metabolitos, así como en la solubilización
del oxígeno, sin destruir la estructura sólida o la reducción de la porosidad del
sustrato o del soporte (Oriol et al., 1988). En nuestro caso, no se observó un
impacto significativo sobre la producción de esporas, cuando diferentes contenidos
de humedad fueron evaluados (Tabla VIII.9), probablemente porque la máxima
capacidad de absorción de agua de los residuos de mango y de la espuma de
poliuretano fueron constantes en cada tratamiento, independiente del contenido de
agua inicial. Un contenido de humedad de 75% fue suficiente para saturar el
sistema de fermentación en estado sólido. Resultados similares han sido
reportados cuando el arroz, maíz o trigo fueron usados en este tipo de sistemas,
observando un estrecho rango de humedad independiente de la cantidad de agua
adicionada (Cavalcante et al., 2008). Además, Bai et al. (2008) reportaron que la
producción de esporas de T. viride en una fermentación en estado sólido de los
residuos de la producción de vinos, no fue incrementada en intervalo de humedad
entre el 66 y 72%.

RES
ULT
AD
OS
Y D
ISC
USI
ÓN
Tabla VIII.9. Efecto de diferentes niveles de humedad sobre la producción de esporas
de T. asperellum T8a (relación C: N fijada en 26 y 6 días después de la inoculación).
Contenido de Humedad,
%
Producción de esporas,
esporas x 108 g
-1ss
0 0.01 ± 0.002e
64 20 ± 2d,e
78 51 ± 7a,b,c
84 30 ± 7c,d
88 60 ± 11a,b
90 48 ± 3b,c
92 76 ± 11a
93 39 ± 11b,c,d
94 63 ± 6a,b
Los valores de las medias con la misma letra no son significativamente diferentes, de
acuerdo con la prueba de ANOVA/ Tukey-Kramer (p = 0.05).
Una vez fijadas una relación C: N de 26 y un contenido de humedad de 78% (la
más baja que indujo la mayor producción de esporas), se procedió a estudiar el
tiempo óptimo de cosecha de las esporas producidas. Como se observa en la
(Tabla VIII.10), ocho días después de la inoculación resultó ser el mejor tiempo
para la colecta de las esporas, lo cual coincidió con la cobertura completa del
sustrato por T. asperellum T8a. El incremento en la producción de esporas
después de 8 días de fermentación fue no significativo, lo que sugiere que la
fermentación puede ser completada en este momento. La producción de esporas
por T. asperellum T8a (76 x 108 esporas g-1 ss, después de 8 días) fue
significativamente mayor y en menor tiempo de cultivo en comparación con lo
reportado con T. harzianum utilizando salvado de arroz, paja de trigo, bagazo de
caña de azúcar en una fermentación en estado sólido, obteniendo 40, 2 y 3 x 108
esporas g-1ss, después de 15 días de inoculación (Lakshmi y Chandra, 2004). La
rápida producción de micelio y esporas por cepa T8a, puede ser atribuida a su
rápido crecimiento, así como la alta disponibilidad de nutrientes en los residuos

RES
ULT
AD
OS
Y D
ISC
USI
ÓN
industriales de mango (composición de estos residuos y la actividad celulolítica de
T. asperellum T8a) (Rossi-Rodrígues et al., 2009).
Tabla VIII.10. Efecto del tiempo de cosecha en la producción de esporas de T.
asperellum T8a (relación C: N fijada en 26 y un contenido de humedad del 78%).
Tiempo de colecta,
días
Producción de esporas,
esporas x 108 g
-1ss
1 0.02 ± 0.004e
2 0.03 ± 0.02e
3 0.08 ± 0.04e
4 3.9 ± 0.01d,e
5 12 ± 0.1d
6 41 ± 3c
7 60 ± 2b
8 76 ± 3a
9 77 ± 6a
Los valores de las medias con la misma letra no son significativamente diferentes, de
acuerdo con la prueba de ANOVA/ Tukey-Kramer (p = 0.05).
T. asperellum T8a fue incapaz de crecer en ausencia de la espuma de
poliuretano como una matriz de soporte inerte, probablemente debido a las bajas
tasas de transferencia de oxígeno y/ o la represión catabólica por el alto nivel de
carbohidratos solubles en los residuos industriales de mango (Viniegra-González
et al., 2003). Microscopías electrónicas de barrido de T. asperellum T8a utilizando
los residuos de mango para su crecimiento, en presencia de la espuma de
poliuretano y bajo las condiciones óptimas para la producción de esporas
(relación C: N de 26, contenido de humedad de 78% y 8 días después de la
inoculación) sugiere que la matriz de soporte inerte (espuma de poliuretano)
permite que el oxígeno y carbohidratos solubles se difundan a través de sus poros,
generando zonas de concentraciones óptimas de oxígeno y nutrientes para el
crecimiento y la esporulación de esta cepa (Fig. VIII.13).

RES
ULT
AD
OS
Y D
ISC
USI
ÓN
Figura VIII.13. Micrografías electrónicas de barrido de T. asperellum T8a después de 8
días de cultivo sobre: a) los residuos industriales de mango (r: residuos industriales de
mango, m: micelio de T. asperellum T8a y e: esporas de T. asperellum T8a) y b) espuma
de poliuretano (m1: micelio de T. asperellum T8a, m2: micelio de T. asperellum T8a en los
poros de la espuma de poliuretano, y e: espuma de poliuretano como matriz de soporte
inerte), en fermentación en estado sólido (relación C: N de 26 y contenido de humedad
78%).
La viabilidad de las esporas producidas bajo la fermentación en estado sólido
fue determinada sobre aquellas esporas colectadas en las condiciones que
maximizaron su producción utilizando como sustrato los residuos de
industrialización de mango, así como aquellas colectadas en medio PDA, después
de su almacenamiento durante 7 días a 4° C. El mayor número de esporas (94 ±
1%) colectadas de los residuos de mango sobrevivieron una semana de
almacenamiento a 4° C, mientras que sólo el 86 ± 4% permanecieron viables
cuando éstas fueron cosechadas del medio PDA y almacenadas bajo las
condiciones antes mencionadas. Esto sugiere que los altos niveles de
carbohidratos presentes en los residuos de mango sirvieron como
osmoprotectores (Thomas et al., 1994; Argüelles, 2000). Además, las esporas de
T. asperellum T8a producidas utilizando este sistema, pueden ser aplicadas al
campo utilizando como vehículo los residuos industriales mango (sustratos en
fermentación en estado sólido), incrementado la viabilidad y la germinación de las
esporas en el suelo, debido a que las especies del género Trichoderma son
b)
m1
m2
e
a)
e r
m

RES
ULT
AD
OS
Y D
ISC
USI
ÓN
hongos saprófitos, es decir, necesitan materia orgánica para germinar y crecer
(Gams y Bissett, 1998).
Ensayos de confrontación contra C. gloeosporioides ATCC MYA 456 fueron
realizados con la finalidad de cuantificar la capacidad antagónica de las esporas
producidas utilizando los residuos de mango o PDA. Similares porcentajes de
inhibicion y clase de control biológico (micoparasitismo) fueron obtenidos
utilizando los residuos de mango y PDA como sustrato (89 ± 2% y 91 ± 2% de
inhibición, respectivamente). En otras palabras, los residuos industriales de mango
ayudaron a prolongar la vida útil de las esporas de T. asperellum T8a (agentes de
control biológico contra la antracnosis) cuando éstos fueron utilizados para su
producción en fermentación en estado sólido. Además, este sistema de
producción de esporas (optimizado para ser utilizado in situ) sugiere una
alternativa económica que puede ser utilizada para el control biológico de
patógenos de plantas en el campo, así como el aprovechamiento de altos
volúmenes de residuos industriales de mango.
FASE III. Inoculación de microorganismos promotores de crecimiento vegetal a
árboles de mango, estudiando su impacto, así como el de las señales exógenas y
endógenas, sobre el evento de floración. En la actualidad, varias alternativas para inducir la promoción del crecimiento y
la floración en mango han sido desarrolladas; sin embargo, sus mecanismos de
acción todavía no están claros (Protacio, 2000; Urban y Alphonsout, 2007;.
Vázquez-Valdivia et al., 2009). Por otra parte, estas prácticas agrícolas presentan
inconvenientes como la baja eficiencia, altos costos económicos y la
contaminación del medio ambiente (Maganhotto et al., 2003). Por lo cual, el uso de
BPCV se ha estudiado en varios cultivos tales como alfalfa, manzana, plátano,
maíz, kiwi y trigo, en los cuales se ha observado un incremento en la absorción de
nutrientes y una mayor biomasa seca total, dando lugar a un incremento en la
producción.

RES
ULT
AD
OS
Y D
ISC
USI
ÓN
Este aumento de la producción agrícola se ha explicado a través de varios
mecanismos, tales como: actividad ACC desaminasa, la fijación biológica de
nitrógeno, solubilización de fósforo, la síntesis de fitohormonas y la producción de
sideróforos (Ercisli et al., 2003; Aslantas et al., 2007; Alikhani y Yakhchali, 2009;
Mia et al., 2010). Sin embargo, el uso de las BPCV no se ha explorado como una
alternativa para la producción de mango.
VIII.6. Manejo de la inoculación microbiana en árboles de mango creciendo
en el invernadero.
Las características fisicoquímicas y nutrimentales del suelo usado en este
experimento son mostrados en la Tabla VIII.11. Estos características indican que
este suelo tienen el nivel de fertilidad óptima para el desarrollo fenológico de los
árboles de mango en el invernadero (Avilán, 2008).
Tabla VIII.11. Características físico-químicas del suelo utilizado para el crecimiento
de los árboles de mango en invernadero.
Origen del Suelo Textura MO
%
N P K C.I.C
Meq 100-1 g ppm
Tomatlán, Jalisco, México Franco,
arcilloso
2.4 21.1 66.4 283 18.2
MO: Materia orgánica, N: nitrógeno, P: fósforo, K: potasio, C.I.C: capacidad de
intercambio catiónico.
El ciclo fenológico observado en árboles de mango creciendo en el invernadero
fue similar al comportamiento fenológico de éstos en huertos productivos (Avilán,
2008). Este ciclo fenológico mostró cuatro etapas bien definidas (Fig. VIII.13):
crecimiento vegetativo (de junio a septiembre del 2009), dormancia (de octubre a
noviembre del 2009), floración (de diciembre del 2009 hasta febrero del 2010) y
fructificación (abril del 2010).
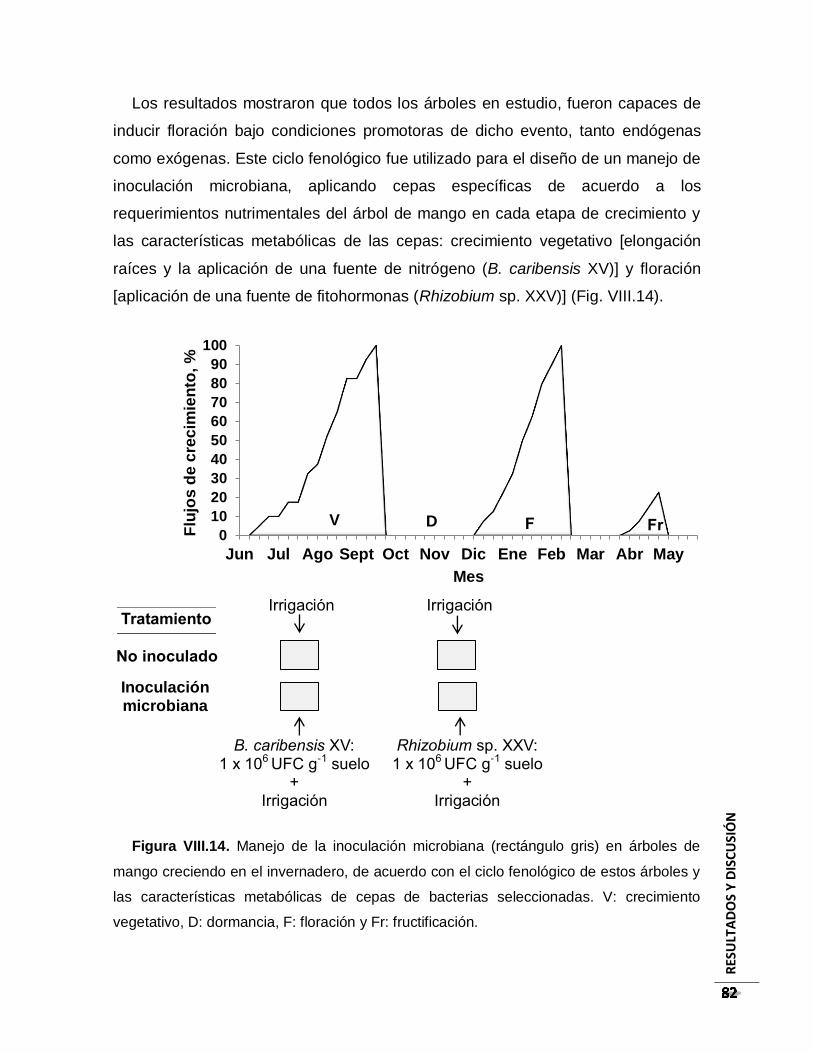
RES
ULT
AD
OS
Y D
ISC
USI
ÓN
Los resultados mostraron que todos los árboles en estudio, fueron capaces de
inducir floración bajo condiciones promotoras de dicho evento, tanto endógenas
como exógenas. Este ciclo fenológico fue utilizado para el diseño de un manejo de
inoculación microbiana, aplicando cepas específicas de acuerdo a los
requerimientos nutrimentales del árbol de mango en cada etapa de crecimiento y
las características metabólicas de las cepas: crecimiento vegetativo [elongación
raíces y la aplicación de una fuente de nitrógeno (B. caribensis XV)] y floración
[aplicación de una fuente de fitohormonas (Rhizobium sp. XXV)] (Fig. VIII.14).
Figura VIII.14. Manejo de la inoculación microbiana (rectángulo gris) en árboles de
mango creciendo en el invernadero, de acuerdo con el ciclo fenológico de estos árboles y
las características metabólicas de cepas de bacterias seleccionadas. V: crecimiento
vegetativo, D: dormancia, F: floración y Fr: fructificación.
0
10
20
30
40
50
60
70
80
90
100
Jun Jul Ago Sept Oct Nov Dic Ene Feb Mar Abr May
Flu
jos
de
cre
cim
ien
to,
%
Mes
V F Fr D
Inoculación microbiana
No inoculado
Tratamiento
B. caribensis XV: 1 x 106 UFC g-1 suelo
+ Irrigación
Rhizobium sp. XXV: 1 x 106 UFC g-1 suelo
+ Irrigación
Irrigación
Irrigación

RES
ULT
AD
OS
Y D
ISC
USI
ÓN
El manejo adecuado de la fertilización en cada ciclo fenológico ha sido
reportado como un factor determinante en la producción de mango. Por ejemplo,
la aplicación de nitrógeno promueve el crecimiento vegetativo de los árboles, lo
cual es determinante para la producción de biomasa foliar y en consecuencia un
aumento en la acumulación de carbohidratos, lo que conduce a un incremento en
la producción de frutos (Avilán, 2008; Ramírez y Davenport, 2010).
De la misma manera, la aplicación de fitohormonas tiene un papel importante
en la fisiología de este árbol. Se ha propuesto que en los yemas, la relación entre
fitohormonas endógenas, por ejemplo, alta concentración de citocininas y baja
concentración de auxinas, parece controlar el evento de iniciación de forma
independiente a otras influencias inductivas (Hegele et al., 2006).
Además, la evidencia del papel negativo de las giberelinas en el evento de
floración en mango se ha demostrado con la aplicación de triazoles, un promotor
floral que inhibe a la enzima kaureno oxidasa en la ruta de síntesis de giberelinas,
lo que sugiere que esta ruta puede estar relacionada con la inhibición de la
floración en mango (Turnbull et al., 1996; Davenport, 2007).
VIII.7. Cuantificación de parámetros morfométricos, bioquímicos y
moleculares involucrados en el evento de floración de árboles de mango
bajo el manejo de la inoculación microbiana
El manejo de la inoculación microbiana diseñado (Fig. VIII.14) condujo a la
promoción de crecimiento de los árboles de mango evidenciado por un incremento
en la biomasa seca de la raíz (89%), tallo (34%), y hojas (51%), así como aumento
del área foliar (53%), en comparación con el tratamiento no inoculado (Tabla
VIII.12).
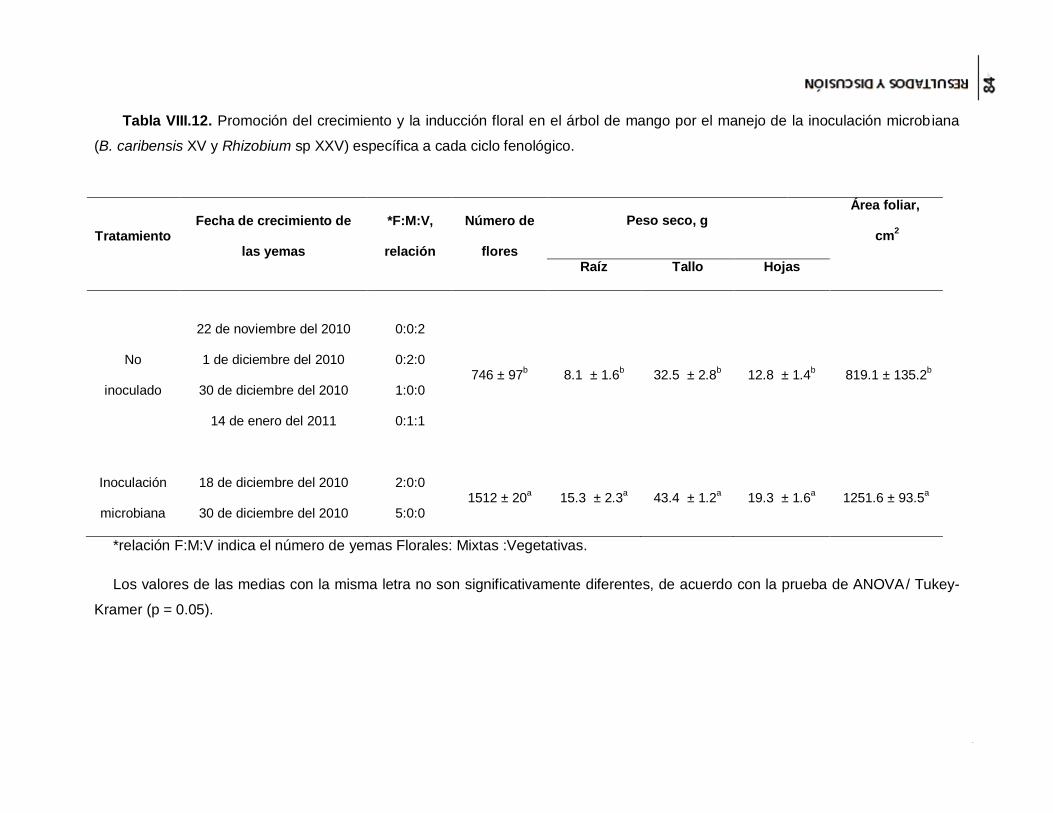
RES
ULT
AD
OS
Y D
ISC
USI
ÓN
Tabla VIII.12. Promoción del crecimiento y la inducción floral en el árbol de mango por el manejo de la inoculación microbiana
(B. caribensis XV y Rhizobium sp XXV) específica a cada ciclo fenológico.
*relación F:M:V indica el número de yemas Florales: Mixtas :Vegetativas.
Los valores de las medias con la misma letra no son significativamente diferentes, de acuerdo con la prueba de ANOVA/ Tukey-
Kramer (p = 0.05).
Tratamiento Fecha de crecimiento de
las yemas
*F:M:V,
relación
Número de
flores
Peso seco, g Área foliar,
cm2
Raíz Tallo Hojas
No
inoculado
22 de noviembre del 2010
1 de diciembre del 2010
30 de diciembre del 2010
14 de enero del 2011
0:0:2
0:2:0
1:0:0
0:1:1
746 ± 97b 8.1 ± 1.6
b 32.5 ± 2.8
b 12.8 ± 1.4
b 819.1 ± 135.2
b
Inoculación
microbiana
18 de diciembre del 2010
30 de diciembre del 2010
2:0:0
5:0:0 1512 ± 20
a 15.3 ± 2.3
a 43.4 ± 1.2
a 19.3 ± 1.6
a 1251.6 ± 93.5
a

RES
ULT
AD
OS
Y D
ISC
USI
ÓN
El incremento en los valores de estos parámetros morfométricos, los cuales son
determinantes en el ciclo de desarrollo del mango (Shaban, 2009), indican el
efecto positivo de las actividades promotoras del crecimiento vegetal de las cepas
inoculadas sobre los árboles de mango, lo que sugiere que la producción de
fitohormonas y actividad ACC desaminasa son las principales actividades
involucradas en los eventos observados. Estos efectos positivos pueden ser
explicados como las respuestas de las plantas a i) las auxinas (elongación y
división celular, iniciación radical, incremento de la tasa de crecimiento y
dominancia apical); ii) citocininas (elongación e inducción de la división celular y la
expansión del tejido), y iii) las giberelinas (modificación de morfología de la planta
por la extensión de sus tejidos, particularmente los tallos) (Verma et al., 2010).
Además, la actividad ACC desaminasa pudo haber contribuido a facilitar el
crecimiento y desarrollo de plantas al disminuir su concentración de etileno (Glick
et al., 1998).
Además, la producción de fitohormonas por la inoculación microbiana pudo
contribuir en la sincronía de crecimiento de las yemas, así como en la
determinación de su destino floral, en comparación con el tratamiento no
inoculado. El crecimiento de las yemas comenzó en dos fechas (18 y 30 de
diciembre del 2010) generando 100% de yemas con destino floral y cuatro fechas
(22 de noviembre, 11 y 30 de diciembre del 2010, y 14 de enero del 2011) lo cual
resultó en un 14% de yemas con desino floral, 43% mixto y 43% de destino
vegetativo, respectivamente (Tabla VIII.12).
Davies (2004) reportó que en las plantas, las auxinas son sintetizadas
principalmente en las hojas y en las semillas en desarrollo, y su transporte a la raíz
probablemente involucra el floema. La biosíntesis de citocininas ocurre en los
meristemos radicales y semillas en desarrollo, y se transporta a través del xilema
de la raíz a los brotes del tallo. Las giberelinas son sintetizadas en tejidos jóvenes
de los brotes del tallo y en semillas en desarrollo, y algunas son transportadas
probablemente vía el floema y xilema.

RES
ULT
AD
OS
Y D
ISC
USI
ÓN
Los resultados obtenidos sugieren que la sincronía e iniciación del crecimiento
de las yemas en el tratamiento bajo inoculación microbiana pudo ser el resultado
de una relación alterada de fitohormonas en éstas, generado por el incremento de
la producción de citocininas en las raíces (como fue observado en el aumento del
89% del peso seco de las raíces producida por el tratamiento de inoculación
microbiana) y de su translocación a las yemas, lo que generó una baja relación
auxina: citocinina, lo cual indujo la iniciación floral en mango (Ramírez y
Davenport, 2010).
En un acercamiento para explicar por qué los árboles bajo la inoculación
microbiana mostraron una inducción del evento de floración en comparación con
tratamientos no inoculado, varios factores fueron analizados.
El primero fue el contenido de nitrógeno foliar, el cual ha sido reportado como
un parámetro crítico implicado en la inducción floral del mango. Los árboles de
mango deben tener niveles nitrógeno en hoja de 1.4% para suprimir eventos
frecuentes de crecimiento vegetativo e inducir la floración (Avilán, 2008). En
nuestro caso, los tratamientos no inoculados y bajo inoculación microbiana
mostraron niveles de nitrógeno total en las hojas de 1.4% ± 0.1 y 1.5 % ± 0.1,
respectivamente, lo que indica que los árboles, en ambos tratamientos, tuvieron
las condiciones nutricionales óptimas, en relación al nitrógeno, para inducir la
floración. Sin embargo, el contenido de nitrógeno foliar total (contenido de
nitrógeno foliar x biomasa foliar total) en los árboles bajo la inoculación microbiana
incrementó significativamente en comparación con el tratamiento no inoculado,
283.8 ± 24.4 contra 181.8 ± 25.1 mg de N planta-1, respectivamente.
El contenido de nitrógeno total foliar ha sido reportado como un factor
importante en la síntesis de carbohidratos en la planta, lo cual repercute sobre
varios eventos fisiológicos, incluyendo la floración (Turnbull, 2011).

RES
ULT
AD
OS
Y D
ISC
USI
ÓN
El contenido de carbohidratos en hojas de los árboles de mango fue analizado
con el objetivo de estudiar su papel en este evento. Como se muestra en la Fig.
VIII.15, el contenido inicial (en mg planta-1) de sacarosa, glucosa y fructosa en el
tratamiento bajo inoculación microbiana fue significativamente mayor que el
tratamiento no inoculado, lo que se correlaciona con mayor área foliar y contenido
de nitrógeno total observado cuando las cepas de PCV fueron inoculadas (Tabla
VIII.12).
En relación al tratamiento bajo inoculación microbiana, los altos valores iniciales
de los carbohidratos estudiados disminuyeron con el tiempo, presentando una
dinámica similar en todas las fechas de muestreo, lo que sugiere que éstos fueron
consumidos para dar lugar a la formación de la panícula y el desarrollo de flores
(Pongsomboon et al., 1997).
Por otro lado, el contenido de sacarosa en los tratamientos no inoculados
presentó similares dinámica decreciente en comparación con los tratamientos bajo
inoculación microbiana, aunque la glucosa y la fructosa presentaron valores
mayores entre el 30 de diciembre del 2010 y el 14 de enero del 2011 (Fig. VIII.15).
Este incremento pudo ser debido a la falta de consumo de estos carbohidratos,
debido a la ausencia de floración (Tabla VIII.12).
Estos resultados sugieren que el contenido de carbohidratos en las hojas
fuentes está involucrado en el desarrollo de la panícula y la intensidad de la
inducción floral, mediante la movilización de la proteína FT a través del floema a
las yemas receptoras (Pongsomboon et al., 1997; Davenport, 2007).
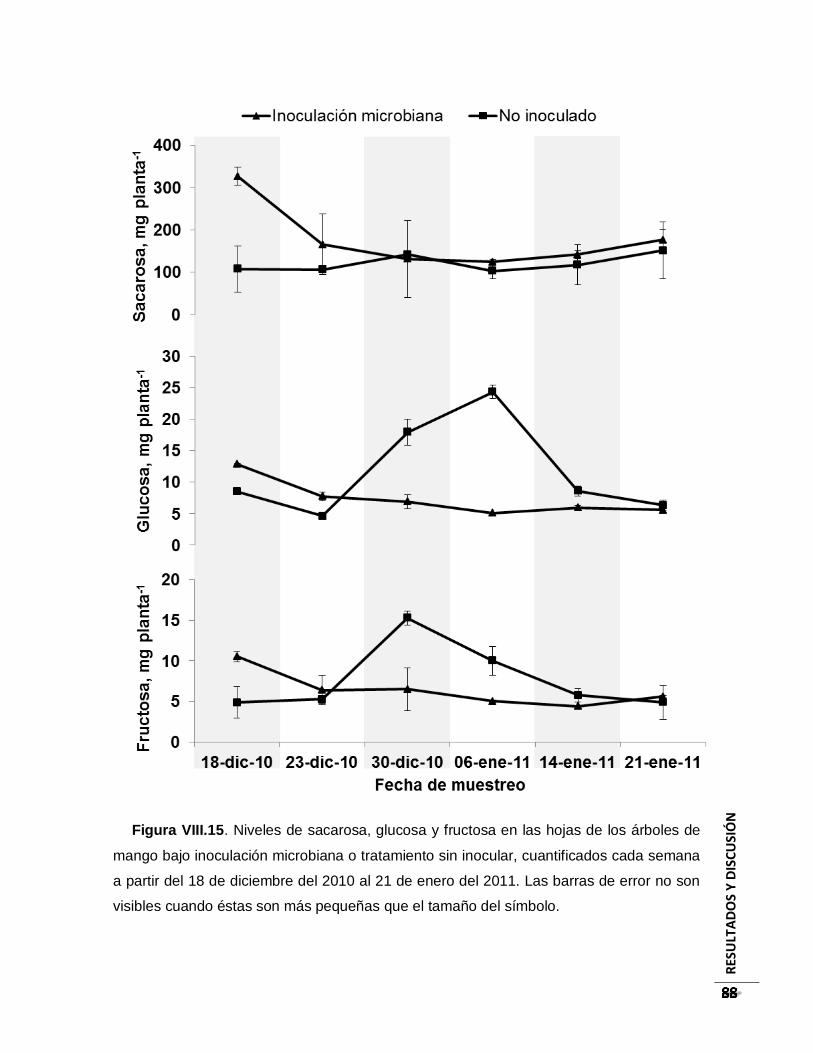
RES
ULT
AD
OS
Y D
ISC
USI
ÓN
Figura VIII.15. Niveles de sacarosa, glucosa y fructosa en las hojas de los árboles de
mango bajo inoculación microbiana o tratamiento sin inocular, cuantificados cada semana
a partir del 18 de diciembre del 2010 al 21 de enero del 2011. Las barras de error no son
visibles cuando éstas son más pequeñas que el tamaño del símbolo.

RES
ULT
AD
OS
Y D
ISC
USI
ÓN
Perfil de expresión del gen FT en las hojas de los árboles de mango
Debido a que la secuencia del ARNm del gen FT en mango no había sido
reportada, se procedió a la obtención de una fragmento de este transcrito con la
finalidad de ser utilizado para el diseño de los oligonucleótidos a usar en los
experimento de RT-PCR en tiempo real.
Estos oligonucleótidos fueron diseñados a partir de un alineamiento de 41
secuencias de aminoácidos de la proteína FT, las cuales pertenecieron a 15
especies de plantas (Fig. VIII.16), las cuales fueron obtenidas del banco de genes
del NCBI: Oncidium Gower Ramsey (ACC59806), Cymbidium goeringii
(ADI58462), Triticum aestivum (ACA25439), Triticum aestivum (ACA25437),
Triticum aestivum (AAW23034), Triticum aestivum (ACA25438), Citrus unshiu
(BAF96645), Citrus unshiu (BAF96644), Malus x domestica ( ADB12456), Malus x
domestica (ABF84010), Pyrus pyrifolia (BAI99731), Pyrus pyrifolia (BAJ11577),
Malus x domestica (ACL98164), Vitis vinifera (ACZ26523), Vitis vinifera
(ABL98120), Populus nigra (BAD02371), Populus nigra (BAG12904), Populus
nigra (BAG12902), Populus nigra (BAD08336), Populus nigra (BAG12901),
Populus nigra (BAD01576), Populus nigra (BAG12900), Populus nigra
(BAD08337), Populus nigra (BAD01561), Populus nigra (BAD02372), Populus
nigra (BAD08338), Populus nigra (BAD01612), Populus nigra (BAG12899), Carica
papaya (ACX85427), Petunia x hybrida (ADF42571), Brassica napus (ACY03403),
Brassica napus (ACY03404), Brassica oleracea (ACH86033), Brassica napus
(ACY03402), Brassica napus (ACY03405), Sinapis alba (ACM69284), Sinapis alba
(ACM69283), Brassica napus (ACY03400), Brassica napus (ACY03401),
Arabidopsis halleri (BAJ08316) y Arabidopsis thaliana (AF152096).
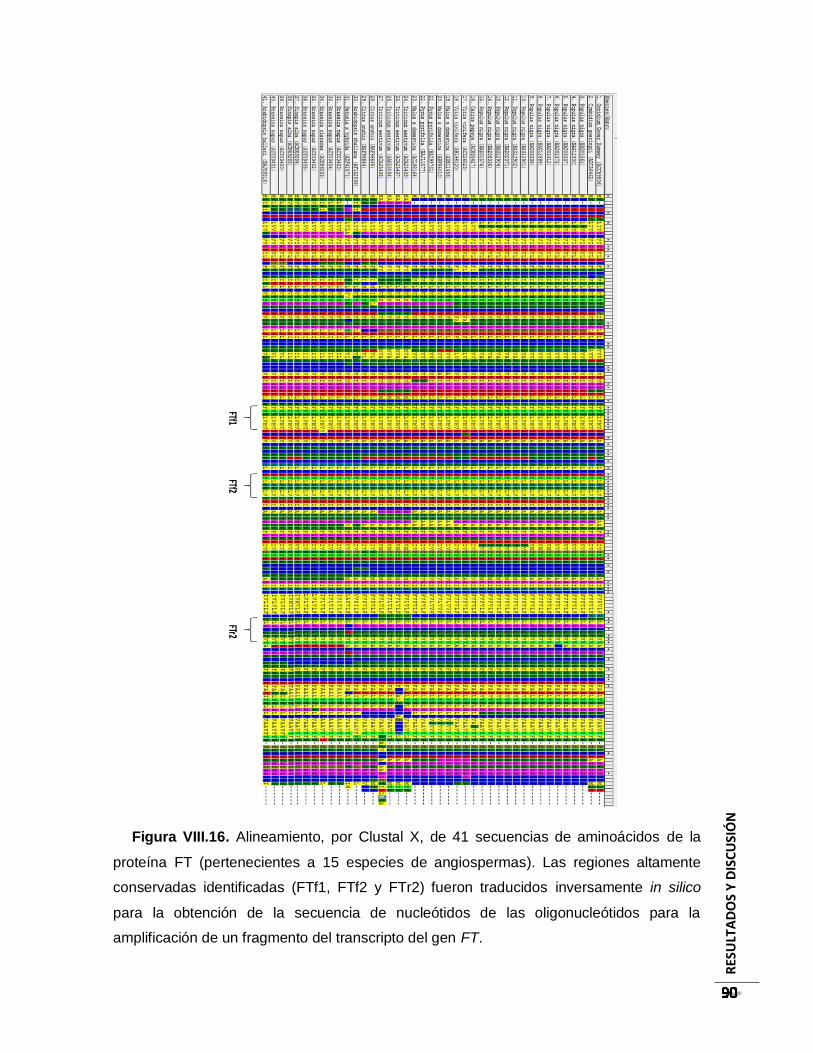
RES
ULT
AD
OS
Y D
ISC
USI
ÓN
Figura VIII.16. Alineamiento, por Clustal X, de 41 secuencias de aminoácidos de la
proteína FT (pertenecientes a 15 especies de angiospermas). Las regiones altamente
conservadas identificadas (FTf1, FTf2 y FTr2) fueron traducidos inversamente in silico
para la obtención de la secuencia de nucleótidos de las oligonucleótidos para la
amplificación de un fragmento del transcripto del gen FT.

RES
ULT
AD
OS
Y D
ISC
USI
ÓN
Tres regiones altamente conservadas, FTf1, FTf2 y FTr2, fueron identificadas a
partir del alineamiento de las secuencias de aminoácidos de la proteína de FT
(Fig. VIII.16), las cuales fueron traducidas inversamente para la obtención de las
secuencias de nucleótidos de los oligonucleótidos (Tabla VIII.13).
Tabla VIII.13. Secuencias de aminoácidos y de nucleótidos de las regiones altamente
conservadas (oligonucleótidos) identificadas en el alineamiento de las secuencias de
aminoácidos de la proteína FT.
Nombre Secuencia de aminoácidos
Secuencia de nucleotidos Tm, oC
FTf1 FYTLVMV 5`-TTCTACACTCTGGTTATGGTGG-3` 53.0
FTf2 EYLHWLV 5`-GAATACTTGCATTGGTTGGTGAC-3` 53.5
FTr2 QLGRQTV 5`-TACACTGTTTGCCTACCCAGTTG-3` 55.3
FTf1: oligonucleótido directo 1, FTf2: oligonucleótido directo 2, FTr2: oligonucleótido
reverso 2. Tm: Temperatura óptima de alineamiento.
Una vez que los oligonucleótidos fueron diseñados, el ARN total extraído de
hojas de los árboles de mango fue utilizado para la síntesis de ADNc y
posteriormente para la amplificación de un fragmento de transcrito del gen FT por
PCR. Los amplicones esperados (aproximadamente 210 pb y 150 pb utilizando
FTf1/FTr2 y FTf2/FTr2, respectivamente) fueron purificados, clonados y
secuenciados por la plataforma Sanger.
Las secuencias obtenidas fueron analizadas y comparadas con aquellas
depositadas en el banco de genes del NCBI, presentando un alto porcentaje de
identidad, 86 y 84% a Populus nigra, respectivamente.
Las secuencias obtenidas fueron depositadas en el banco de genes del NCBI
bajo los siguientes números de acceso: JX316911 (para la secuencia de 207 pb) y
JX316912 (para la secuencia de 147 pb).

RES
ULT
AD
OS
Y D
ISC
USI
ÓN
Con la obtención de oligonucleótidos capaces de la amplificación correcta de
un fragmento del gen FT, experimentos fueron llevados a cabo con la finalidad de
estudiar la expresión de este gen y su potencial papel como una señal floral en
este cultivo.
Como se muestra en la Fig. VIII 17, el perfil de expresión de FT normalizado a
ACT, presentó dinámicas similares en árboles de mango bajo inoculación
microbiana en comparación con el tratamiento no inoculado, aunque con distintos
niveles de expresión.
En el tratamiento no inoculado, el valor de la cuantificación relativa (RQ) no
excedió 0.03 durante el periodo de tiempo evaluado. Sin embargo, valores de RQ
mayores a 0.05 fueron observados en el tratamiento bajo inoculación microbiana.
Además, la expresión de FT en el tratamiento bajo inoculación microbiana
normalizado a el tratamiento no inoculado (RQ = 1) mostró una dinámica de
constante incremento (hasta RQ = 2.2). Sin embargo, una drástica disminución fue
observada en la muestra colectada el 21 de enero del 2011, cuando todos los
árboles presentaron panículas y flores desarrolladas.
Estos datos sugieren que la floración potencialmente reprime la expresión de
FT. Resultados similares han sido observados por Muñoz-Fambuena et al. 2011 y
Nakagawa et al. 2012, quienes reportaron que los frutos tienen un efecto negativo
en la expresión de FT en mandarina y mango, respectivamente.
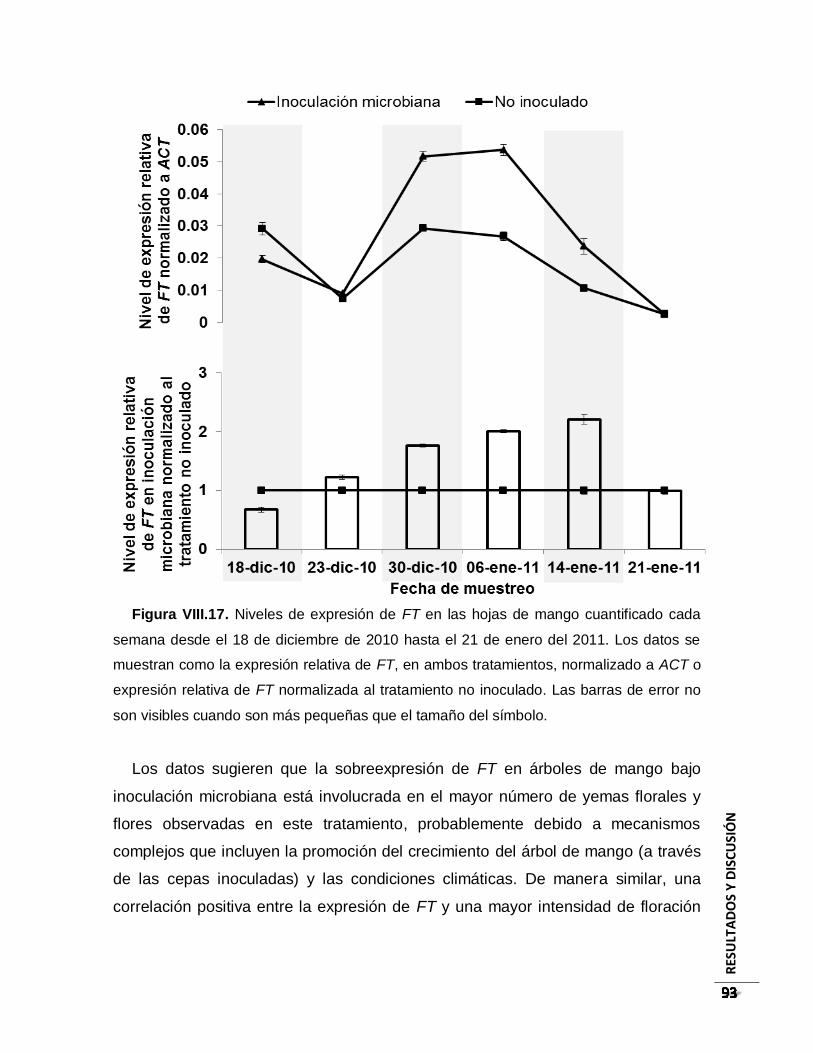
RES
ULT
AD
OS
Y D
ISC
USI
ÓN
Figura VIII.17. Niveles de expresión de FT en las hojas de mango cuantificado cada
semana desde el 18 de diciembre de 2010 hasta el 21 de enero del 2011. Los datos se
muestran como la expresión relativa de FT, en ambos tratamientos, normalizado a ACT o
expresión relativa de FT normalizada al tratamiento no inoculado. Las barras de error no
son visibles cuando son más pequeñas que el tamaño del símbolo.
Los datos sugieren que la sobreexpresión de FT en árboles de mango bajo
inoculación microbiana está involucrada en el mayor número de yemas florales y
flores observadas en este tratamiento, probablemente debido a mecanismos
complejos que incluyen la promoción del crecimiento del árbol de mango (a través
de las cepas inoculadas) y las condiciones climáticas. De manera similar, una
correlación positiva entre la expresión de FT y una mayor intensidad de floración

RES
ULT
AD
OS
Y D
ISC
USI
ÓN
ha sido reportada recientemente, sugiriendo la expresión FT como como una señal
potencial en la inducción floral en mango (Nakagawa et al., 2012).
El papel desempeñado por las condiciones climáticas en la floración del
mango
Las condiciones climáticas (temperatura, humedad relativa e iluminación) fueron
monitoreadas cada treinta minutos durante siete meses en el invernadero, con el
objetivo de analizar sus impactos sobre el evento de floración en el árbol de
mango (Fig. VIII.18). Los datos colectados mostraron un período de inducción
floral durante 1 mes (aproximadamente 10º C por la noche y 20º C durante el día)
desde el 1 diciembre de 2010 al 1 de enero del 2011. Observaciones similares han
sido observado por Davenport (2009), quien reportó que la floración del mango se
produce después o durante la exposición a temperaturas de inducción floral (por
debajo de 15° C durante la noche / por debajo de 20º C en el día). Además, el
crecimiento de yemas durante ese período de inducción resultará en yemas mixtas
o florales, dependiendo del tiempo de exposición a estas temperaturas inductivas,
de menos de dos semanas y más de tres semanas, respectivamente (Núñez-
Elisea y Davenport, 1995). Estos resultados apoyan la hipótesis que la inducción
de la floración mango está regulada por la interacción entre una señal florigénica
regulada por la temperatura (posiblemente la proteína FT) y una señal vegetativa
dependiente de la edad (posiblemente una giberelina) (Davenport, 2009). En
nuestro caso, el período inductivo coincidió con un período estable de iluminancia
y fotoperiodo mínimo, ~ 5500 lux y 10.5 horas de luz, respectivamente
(Fig.VIII.18). El papel del estos factores en la floración del mango aún no se ha
establecido, ya que el mango es un árbol que no responde al fotoperiodo. Esto ha
sido concluido a través de estudios enfocados en el papel de la temperatura y el
fotoperiodo sobre la floración de mango, observando que las temperaturas
inductivas causaron la inducción floral más que eventos de fotoperiodos cortos,
mientras que las temperaturas no inductivas inhibieron dicha inducción más que
eventos de fotoperiodo largo (Núñez-Elisea y Davenport, 1995). La humedad
relativa, en nuestro trabajo, no mostró una correlación con la inducción de la
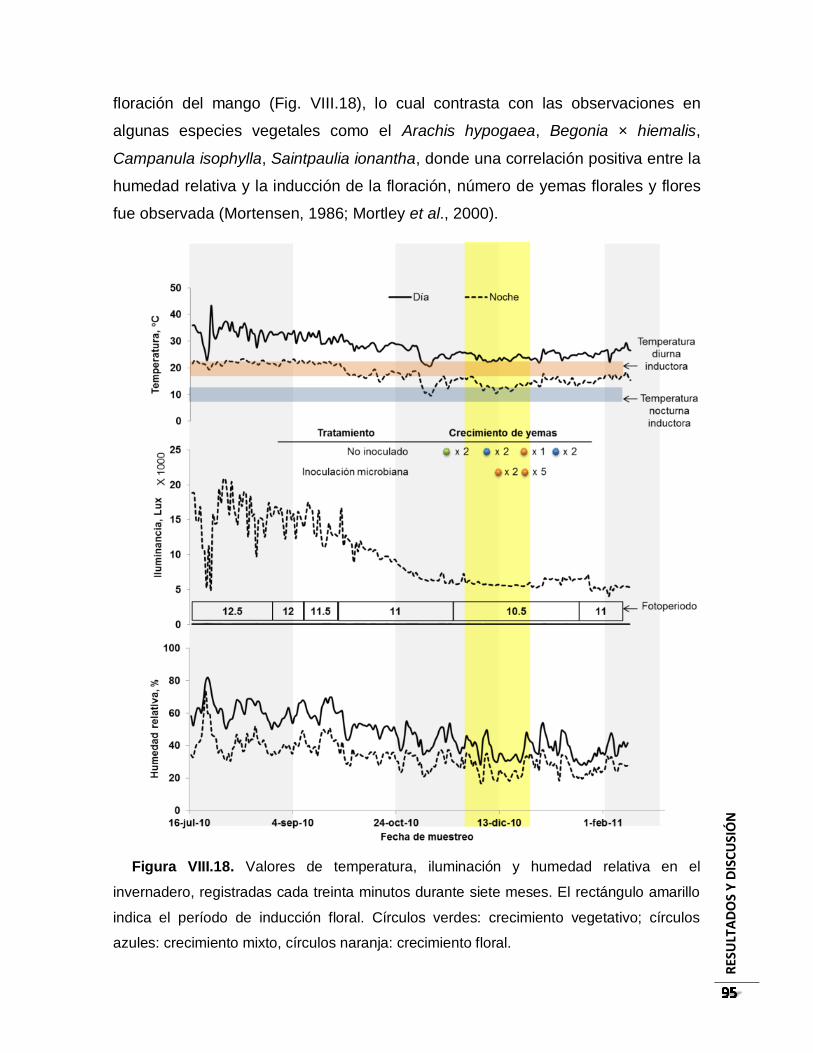
RES
ULT
AD
OS
Y D
ISC
USI
ÓN
floración del mango (Fig. VIII.18), lo cual contrasta con las observaciones en
algunas especies vegetales como el Arachis hypogaea, Begonia × hiemalis,
Campanula isophylla, Saintpaulia ionantha, donde una correlación positiva entre la
humedad relativa y la inducción de la floración, número de yemas florales y flores
fue observada (Mortensen, 1986; Mortley et al., 2000).
Figura VIII.18. Valores de temperatura, iluminación y humedad relativa en el
invernadero, registradas cada treinta minutos durante siete meses. El rectángulo amarillo
indica el período de inducción floral. Círculos verdes: crecimiento vegetativo; círculos
azules: crecimiento mixto, círculos naranja: crecimiento floral.

CO
NC
LUSI
ON
ES
IX. CONCLUSIONES
La cadena de valor del mango presenta etapas que demandan investigación
científica, la cual debe ser apoyada por los conocimientos acumulados en el
cultivo de este fruto a través de varias generaciones. Esto conducirá, además de
la generación de soluciones aplicables a corto plazo, a dilucidar los mecanismos
básicos de su acción, traduciéndose en soluciones cada vez más eficientes e
integrales.
En México, esta cadena de valor es negativamente impactada principalmente
en tres puntos críticos: el evento de floración, el control de antracnosis y el
aprovechamiento de residuos provenientes de la industrialización de mango.
El evento de floración. Las alternativas utilizadas actualmente para el control
de la floración, enfocadas a evitar la estacionalidad de la cosecha, presentan
inconvenientes tales como la inconsistente eficiencia en campo y problemas
económicos-ambientales por su uso. Como fue demostrado en el presente trabajo,
el manejo de la inoculación microbiana es una alternativa potencial para la
inducción floral del árbol de mango en campo; a través de la inoculación de dos
cepas bacterianas (aisladas en huertas de mango) en ciclos fenológicos
determinantes para este cultivo. Lo cual condujo a una inducción tanto de su
promoción del crecimiento, como de la floración (a nivel morfométrico, bioquímica
y molecular), como resultado de las características metabólicas de estas cepas.
El control de la antracnosis. Esta enfermedad impacta significativamente a la
cadena de valor del mango, debido a las grandes pérdidas generadas en
precosecha y postcosecha, de hasta un 90%. Su control en la actualidad conduce
a problemas económicos y ambientales, debido al uso excesivo de fungicidas
químicos, lo cual genera casos de resistencia del agente causal de esta
enfermedad. Como fue evidenciado en el presente trabajo, el uso de agentes de
control biológico contra esta enfermedad es una alternativa integral que permite el
control sustentable del patógeno. Lo cual condujo a la identificación y el estudio de

CO
NC
LUSI
ON
ES
dos cepas con capacidad de controlar in vitro e in vivo a C. gloeosporioides
(agente causal de la antracnosis) a través de la producción de sideróforo (cepa
bacteriana) y actividad de enzimas líticas (cepa fúngica).
El aprovechamiento de los residuos industriales del mango. Los problemas
generados por la inadecuada disposición de los residuos industriales generados
en los procesos de industrialización del mango impactan negativamente a esta
cadena de valor en México, debido a los problemas de contaminación ambiental
que estos generan, así como el desaprovechamiento de su potencial
biotecnológico para la obtención de productos finales con valor agregado. Como
resultado del presente trabajo, una alternativa para el aprovechamiento de estos
residuos es la producción de hongos con capacidad de controlar la antracnosis,
mediante la optimización de parámetros, determinantes para su adopción in situ,
bajo una fermentación en estado sólido, como son la relación C: N, contenido de
humedad y el tiempo para la cosecha de las esporas producidas (manteniendo las
características metabólicas de esta cepa).

PER
SPEC
TIV
AS
X. PERSPECTIVAS
Con base en la complejidad de los puntos estudiados en el presente trabajo y
los resultados generados hasta el momento sobre éstos, es necesaria la
generación de conocimiento científico que permita que las alternativas aquí
propuestas sean aplicadas al campo en un periodo corto de tiempo. Para lo cual,
es determinante el estudio de los siguientes aspectos:
a) El evento de floración
- Cuantificar el impacto del manejo de la inoculación microbiana, diseñado en
el presente trabajo, sobre la inducción de crecimiento y floración en árboles de
mango plantados en huertas productivas. Contrastando este impacto con las
alternativas de control de la floración aplicadas en la actualidad (anillado,
paclobutrazol, KNO3, etc.).
- Estudiar a nivel molecular (transcriptoma) el evento de floración en los
árboles de mango, así como la respuesta a este nivel por la aplicación del manejo
de inoculación microbiana, con la finalidad de diseñar estrategias de inducción
floral más eficientes.
- Determinar el impacto sobre la diversidad microbiana edáfica (RDP), en los
sitios de estudio, por la aplicación del manejo de inoculación microbiana propuesto
para la inducción de crecimiento y floración de los árboles de mango.
b) El control de la antracnosis
- Cuantificar el control de la antracnosis, a nivel de campo y de
almacenamiento, por la aplicación del agente de control biológico caracterizado en
este trabajo. Con la finalidad de conocer su potencial aplicación en estos sistemas
(condiciones ambientales, periodo de actividad, etc.).
c) El aprovechamiento de los residuos industriales del mango.
- Escalamiento del sistema de fermentación en estado sólido para su
adopción in situ, permitiendo un mayor volumen de residuos procesados.

BIB
LIO
GR
AFÍ
A
XI. BIBLIOGRAFÍA
Abraham, E.R., Ramachandran, S., Ramalingam, V. 2007. Biogas: Can it be an
important source of energy?. Env Sci Pollut Res 14:67-71.
Agosin, E., Volpe, D., Munoz, G., San Martin, R., Crawford, A. 1997. Effect of
culture conditions on spore shelf life of the biocontrol agent Trichoderma
harzianum. World J Microb Biot 13:225-232.24.
Ahemad, M., Saghir, M.K. 2011. Functional aspects of plant growth promoting
rhizobacteria: recent advancements. Insight Microbiology 1:39-54.
Alexander, D.B., Zuberer, D.A. 1991. Use of chrome azurol S reagents to
evaluate siderophore production by rhizosphere bacteria. Biol Fert Soils 12:39-45.
Alikhani, H.A., Yakhchali, B. 2009. Potential use of Iranian rhizobial strains as
plant growth promoting rhizobacteria (PGPR) and effects of selected strains on
growth characteristics of wheat, corn and alfalfa. Desert 14:27-35.
Anandham, R., Gandhi, P.I., Madhaiyan, M., Sa, T. 2008. Potential plant growth
promoting traits and bioacidulation of rock phosphate by thiosulfate oxidizing
bacteria isolated from crop plants. J Basic Microbiol 48:439-447.
AOAC. 1990. Official methods of analysis of the association of official analytical
chemists, 15th Edition, Washington, DC, Association of Official Analytical
Chemists.
Arauz, L.F. 2000. Mango anthracnose: economic impact and current options for
integrated management. Plant Dis 84:600–611.
Argüelles, J.C. 2000. Physiological roles of trehalose in bacteria and yeasts: a
comparative analysis. Arch Microbiol 174:217-224.

BIB
LIO
GR
AFÍ
A
Arias Rivas, B., Carrizales, L. 2007. Control químico de la antracnosis del
mango (Mangifera indica L.) en pre y postcosecha en el municipio Cedeño, estado
Monagas, Venezuela. Bioagro 19:19-25.
Askew, D.J., Laing, M.D. 1993. An adapted selective medium for the quantitative
isolation of Trichoderma species. Plant Pathol 42:686-690.
Aslantas, R., Cakmakci, R., Sahin, F. 2007. Effect of plant growth promoting
rhizobacteria on young apples trees growth and fruit yield under orchard
conditions. Sci Hortic 111:371-377.
Avilán, L., Rodríguez, M., Ruiz, J. 1998. Selección de cultivares de mango
(Mangifera indica L.) del Centro Nacional de investigaciones Agropecuarias,
Periodo 1952-1996. Proc Interamer Soc Trop Hort 42:191-214.
Avilán, R.L. 1971. Variación de los niveles de nitrógeno, fósforo, potasio y calcio
en las hojas del mango (Mangifera indica L.) a través de un ciclo de producción.
Agronomía 21:3-10.
Avilán, R.L. 2008. Nutrición y fertilización del mango. Quito Ecuador.
International Plant Nutrition Institute (IPNI).
Avilán, R.L., Meneses, L., Guadarrama, A. 1979. Estudio de los sistemas
radiculares del mango (Mangifera indica L.) y el grapefruit (Citrus paradisi
MacFadyen) cultivados en suelos de los llanos centrales de Venezuela.
Agronomía Tropical 29:173-183.
Avilán, R.L., Rengifo, C.A., Dorantes, I., Rodríguez, M. 1993. El cultivo del
manguero en Venezuela VI. Manejo agronómico del mango. FONAIAP Divulga
10:24-30.

BIB
LIO
GR
AFÍ
A
Avilán, R.L., Rodríguez, M., Ruiz, J.R., Rengifo, C.A. 1995. El manguero en
Venezuela. VIII. Manejo de las plantas de mango. FONAIAP Divulga 12:37-41.
Axmann, H., Sebastianelli, A., Arrillaga, L.J. 1990. Sample preparation
techniques of biological material for isotope analysis. In: Hardarson G (Ed.). Use of
nuclear techniques in studies of soil-plant relationship. Internactional Atomic
Energy Agency. Viena, Austria. p 146.
Bai, Z., Jin, B., Li, Y., Chen, J., Li, Z. 2008. Utilization of winery wastes for
Trichoderma viride biocontrol agent production by solid state fermentation. J
Environ Sci 20:353-358.
Baig, M.M.V. 2005. Cellulolytic enzymes of Trichoderma lignorum produced on
banana agro-waste: Optimisation of culture medium and conditions. J Sci Ind Res
India 64:57-60.
Bailey, J.A., O’connell, R.J., Pring, R.J., Nash, C. 1992. Colletotrichum: Biology,
Pathology and Control. Eds. Bailey, J.A. and Jeger M.J. CAB International
Wallingford.
Bailey, L. 1941. The standard cyclopedia of horticulture. Ed. Bailey, L.H. NY:
McMillan and Company, 3.
Bangerth, F., Carle, R. 2002. Physical, chemical and sensory properties of 9
Thai mango cultivars and evaluation of their technological and nutritional potential.
In: International Symposium 'Sustaining, Food Security and Managing Natural
Resources in Southeast Asia: Challenges for the 21st Century'; Chiang Mai,
Thailand.

BIB
LIO
GR
AFÍ
A
Barabasz, W., Albińska, D., Jaśkowska, M., Lipiec, J. 2002. Biological effects of
mineral nitrogen fertilization on soil microorganisms. Pol J Environ Stud 11:193-
198.
Barnet, H.L., Hunter, B.B. 1978. Illustrated genera of imperfect fungi. Fourth
Edition. MacMillan. Pub. Company New York, USA 217 p.
Bell, D.K., Wells, H.D., Markham, C.R. 1982. In vitro antagomisn of Trichoderma
species against six fungal plant pathogens. Phytopathology 72:279-382.
Bernier, G., Havelange, A., Houssa, C., Petitjean, A., Lejeune, P. 1993.
Physiological signals that induce flowering. Plant Cell 5:1147-1155.
Blázquez, M.A. 2000. Flower development pathways. Cell Science at a Glance
113:3547-3548.
Blázquez, M.A., Weigel, D. 2000. Integration of floral inductive signals in
Arabidopsis. Nature 404:889–892.
Bodner, C.C., Gereau, R.E. 1988. A contribution to Bontoc ethnobotany. Econ
Bot 42:307-369.
Bradford, M. 1976. A rapid and sensitive method for the quantitation of
microgram quantities of protein utilizing the principle of protein-dye binding. Anal
Biochem 72:248-254.
Brown, A.E., Sreenivasaprasad, S., Timmer, L.W. 1996. Molecular
characterization of slow-growing orange and key lime anthracnose strains of
Colletotrichum from Citrus as C. acutatum. Mol Plant Pathol 86:523-527.

BIB
LIO
GR
AFÍ
A
Bustos, G., Moldes, A.B., Cruz, J.M., Domínguez, J.M. 2005. Aprovechamiento
de residuos agroindustriales para la obtención de medios nutritivos económicos y
de ácido láctico. TURevista Digi.U@T 1:1-2.
Carreiro, M.M., Koske, R.E. 1992. Effect of temperature on decomposition and
development of microfungal communities in leaf litter microcosms. Can J Bot
70:2177-2183.
Castaño, A., Londoño, D. 2002. Sinergia de subproductos industriales en el
municipio de Itagüi. Trabajo de grado. Ingenieros de Producción. Escuela de
Ingeniería. Universidad EAFIT, Medellín, Colombia. 70 p.
Cavalcante, R.S., Lima, H.L.S., Pinto, G.A.S., Gava, C.A.T., Rodrigues, S. 2008.
Effect of moisture on Trichoderma conidia production on corn and wheat bran by
solid state fermentation. Food Bioprocess Technol 1:100-104.
Cavalcante, V.A., Dobereiner, J. 1988. A new acid-tolerant nitrogen-fixing
bacterium associated with sugarcane. Plant Soil 108:23-31.
Centis, S.I., Guillas, N., Séjalon, M.T., Esquerré, T. 1997.
Endopolygalacturonase genes from Colletotrichum: cloning of CLPG2 and
comparison of its expression to that of CLPG1 during saprophytic and parasitic
growth of the fungus. Mol Plant Microbe In 10:769-775.
Chahal, P.S., Chahal, D.S. Le, G.B.B. 1996. Production of cellulase in solid
state fermentation with Trichoderma resie MCG80 on wheat straw, Appl Biochem
Biotechnol 57:433-442
Chávez, C.X., Vega, P.A., Tapia, V.L., Miranda, S.M. 2001. Mango, su manejo y
producción en el trópico seco de México. En Libro Técnico Núm. 1. Campo

BIB
LIO
GR
AFÍ
A
Experimental Valle de Apatzingán, CIRPAC, INIFAP, Agosto. Michoacán, México.
p 110.
Coenye, T., Vandamme, P., Govan, J.R.W., Lipuma, J.J. 2001. Taxonomy and
Identification of the Burkholderia cepacia Complex. J Clin Microbiol 39:3427-3436
Compant, S., Reiter, B., Sessitch, A., Nowak, J., Clément, C., Ait Barka, E.
2005. Endophytic colonization of Vitis vinifera L. by plant growth-promoting
bacterium Burkholderia sp. strain PsJN. Appl Environ Microbiol 71:1685-1693.
Cook, A.A. 1975. Disease of tropical and subtropical fruits and nuts. 1st. edn.
New York Hafner Press. p 231.
Couto, S.R., Sanromán, M.A. 2006. Application of solid-state fermentation to
food industry–A review. J Food Eng 76:291–302.
Crane, J.H., Balerdi, C.F. 1991. El Mango en Florida. University of Florida, IFAS
Extension 1:1-7.
Dahlan, S.S. 2007. Spore production of biocontrol agent Trichoderma
Harzianum: effect of C/N ratio and glucose concentration. Jurnal Rekayasa Kimia
dan Lingkungan 6:35-40.
Das, A., Ghosh, U. 2009. Solid-state fermentation of waste cabbage by
Penicillium notatum NCIM NO-923 for production and characterization of
cellulases, J Sci Ind Res India 68:714-718.
Davenport, T., Núñez-Elisea, R. 1991. Is endogenous ethylene involved in
mango floral induction. Acta Hort 291:85-94.

BIB
LIO
GR
AFÍ
A
Davenport, T.L. 2007. Reproductive physiology of mango. Braz J Plant Physiol
19:363-376.
Davenport, T.L. 2009. Reproductive physiology. In: Litz, R.E. (Ed.), The Mango:
Botany Production and Uses, 2nd edition. CAB International, Wallingford, UK, pp.
97-169.
Davenport, T.L., Núñez-Eliséa, R. 1997. Reproductive physiology: The mango,
botany, production and uses. CAB International, pp. 69-146.
Davenport, T.L., Zhang, T., Ying, Z. 2006. Isolation of potentially regulating
mango flowering. Proceedings of the 33rd Annual Meeting of the Plant Growth
Regulation Society of America 109-110.
Davies, P.J. 2004. The plant hormones: their nature, occurrence and function.
In: Davies PJ (Ed.), Plant Hormones: Biosynthesis, Signal Transduction, Action!.
3th edition. Kluwer Academic Publisher, pp. 1-15.
Eastburn, D.M., Butler, E.E. 1991. Effects of soil moisture and temperature on
the saprophytic ability of Trichoderma harzianum. Mycologia 83:257-263.
Elias, X. 2003. Uso de combustibles alternativos. Tecnologías aplicables. Rev
Tec Residuos 13:68-82.
Ellouz, C.S., Belguith, H., Hassairi, I., M'Rad, K., Ellouz, R. 1995. Optimization
of cellulase production by Penicillium occitanis. Appl Microbiol Biotechnol 43:267-
269.
Elson, M.K., Schisler, D.A., Jackson, M.A. 1998. Carbon-to-nitrogen ratio,
carbon concentration, and amino acid composition of growth media influence
conidiation of Helminthosposition solani. Mycologia 98:406-413.

BIB
LIO
GR
AFÍ
A
Ercisli, S., Esitken, A., Cangi, R., Sahin, F. 2003. Adventitious root formation of
kiwifruit in relation to sampling date, IBA and Agrobacterium rubi inoculation. Plant
Growth Regul 41:133-137.
Espinosa, J., Arias, J., Rico, H., Miranda, M., Chávez, X. 2004. Dinámica del
daño y control de la antracnosis Colletotrichum gloeosporioides (Penz.) en mango
en Michoacán. INIFAP 5:1-28.
Estrada-de los Santos, P., Bustillos-Cristales, R., Caballero-Mellado, I. 2001.
Burkholderia, a genus rich in plant-asociated nitrogen fixers with wide
environmental and geographic distribution. Appl Environ Microbiol 67:2790-2798.
FAO. 2010. Recuperado el 11 de 04 de 2012, de FAOSTAT:
http://faostat.fao.org/
Galán-Saúco, V. 1999. El cultivo del mango. Mundi prensa. Madrid, 380.
Galán-Saúco, V. 2009. El cultivo del mango. 2º Edición. Ediciones Mundi-
Prensa, Madrid. p. 33-34, 251.
Gams, W., Bissett, J. 1998. Morphology and identification of Trichoderma,” In
Trichoderma and Gliocladium. Edited by C. P. Kubicek & G. E. Harman. London;
Bristol, PA: Taylor & Francis, pp. 3-31.
Gao, L., Liu, X.Z. 2009. A novel two-stage cultivation method to optimize carbon
concentration and carbon-to-nitrogen ratio for sporulation of biocontrol fungi. Folia
Microbiol 54:142-146.
Garbeva, P., van Veen, J.A., van Elsas, J.D. 2008. Rhizosphere microbial
community and its response to plant species and soil history. Plant Soil 302:19–32.

BIB
LIO
GR
AFÍ
A
Gee, G.W., Bauder, J.W. 1986. Particle size analysis. In A. Klute (ed.). Methods
of soil analysis Vol I Physical and mineralogical methods. Amer Soc Agron
Madison, Wisconsin, USA. pp. 383-411.
Ghose, T.K. 1987. Measurement of cellulase activities. Pure Appl Chem 59:257-
268.
Girard, E.1980. El tomate de árbol: las pestes más comunes. ICA-Informa,
programa de frutales. Medellín, 14:84-87.
Glick, B.R., Penrose, D.M., Li, J. 1998. A model for the lowering of plant
ethylene concentrations by plant growth promoting bacteria. J Theor Biol 190: 63-
68.
Glickmann, E., Dessaux, Y. 1995. A critical examination of the specificity of the
Salkowsky reagent for indolic compounds produced by phytopathogenic bacteria.
Appl Environ Microbiol 61:793-796.
González, I., Infante, D., Martínez, B., Arias, Y., González, N., Miranda, I.,
Peteira, B. 2012. Inducción de quitinasas y glucanasas en cepas de Trichoderma
spp. promisorias como agentes para el control biológico. Biotecnología Aplicada
29:7-11.
Guigón, C., González, P.A. 2004. Selección de cepas nativas de Trichoderma
spp. con actividad antagónica sobre Phytophthora capsici Leoniani y promotoras
de crecimiento en el cultivo de Chile (Capsicum annuum L.). Rev Mex Fitopatol
22:117-124.
Guigón-López, C., Guerrero-Prieto, V., Vargas-Albores, F., Carvajal-Millan, E.,
Ávila-Quezada, G.D., Bravo-Luna, L., Ruocco, M, Lanzuise, S., Woo, S., Lorito, M.
2010. Identificación molecular de cepas nativas de Trichoderma spp. su tasa de

BIB
LIO
GR
AFÍ
A
crecimiento in vitro y antagonismo contra hongos fitopatógenos. Rev Mex Fitopatol
28:87-96.
Gunnell, P.S., Gubler, W.D. 1992. Taxonomy and morphology of Colletotrichum
species pathogenic on strawberry. Mycologia 84:157-165.
Gutiérrez-Alonso, O., Nieto-Ángel, D., Gutiérrez-Alonso, J.G., Delgadillo-
Sánchez, F., Domínguez-Álvarez, J.L. 2002. Características morfológicas,
culturales y patogenicidad de aislamientos de Colletrotrichum spp. obtenidos de
frutos de guayaba (Psidium guajava L.). Rev Mex Fitopatol 20:24-30.
Haby, V.A., Russelle, M.P., Skogley, E.O. 1990. Testing soils for potassium,
calcium and magnesium. In R.L. Westerman (ed.). Soil Testing and Plant
Analysis, 3rd Edition. Soil Sci. Soc. of Amer. Book Series No. 3. SSSA, Madison,
WI.
Haran, S., Schikler, H., Chet, I. 1996. Molecular mechanisms of lytic enzymes
involved in the biocontrol activity of Trichoderma harzianum. Microbiology
142:2321-2331.
Hegele, M., Bangerth, F., Naphrom, D., Sruamsiri, P., Manochai, P. 2006.
Control of flower induction in tropical/subtropical fruit trees by phytohormones
using the example of longan and mango. Acta Hort 727:217-276.
Hiroce, R., Carvhalo, O., Bataglia, O., Furlani, P. Dos Santos, E., Gallo, J.
1977. Composiçao mineral de frutas tropicais na colheita. Bragantia 36:155-164.
Hölker, U., Höfer, M., Lenz, J. 2004. Biotechnological advantages of laboratory-
scale solid-state fermentation with fungi. Appl Microbiol Biotechnol 64:175-186 .

BIB
LIO
GR
AFÍ
A
IPD, A.C. 2008. Estrategia competitiva para el mango. Institución Científico-
Tecnológica.
Jackson, M.A., Bothast, R.J. 1990. Carbon concentration and carbon- to-
nitrogen ratio influence submerged-culture conidiation by the potential bioherbicide
Colletotrichum truncatum NRRL 13737. Appl Environ Microbiol 56:3435-3438.
Kaláb, M., Ann-Fook, Y., Chabot, D. 2008. Bacteria are present everywhere: in
the soil, deep in the rocks, in all bodies of water, in the atmosphere, including the
clouds, and also on and inside other living organisms. Conventional Scanning
Electron Microscopy of Bacteria. 10:43-61.
Kardailsky, I., Shukla, V.K., Ahn, J.H., Dagenais, N., Christensen, S.K., Nguyen,
J.T., Chory, J., Harrison, M.J., Weigel, D. 1999. Activation tagging of the floral
inducer FT. Science 286:1962-1965.
Kenworthy, A. 1964. Fruit nut and plantation crops. Desidues and everygreen (a
guide collecting foliar sample for nutrient analysis) Horticultural Department.
Michigan State University East lansing. p. 30.
Ker, C.K. 2001. Sensitivity of mango anthracnose pathogen, Colletotrichum
gloeosporioides, to the fungicide prochloraz in Taiwan. Proc Natl Sci Counc Repub
China B 25:174-179.
Klein, D., Eveleigh, D.E. 1998. Ecology of Trichoderma. In: Kubicek, C.P.,
Harman, G.E. (Eds.), Trichoderma and Gliocladium. Basic Biology, Taxonomy and
Genetics. Taylor & Francis, London, UK. pp. 57–74.
Kobayashi, Y., Kaya, H., Goto, K., Iwabuchi, M.A. 1999. A pair of related genes
with antagonistic roles in mediating flowering signals. Science 286:1960-1962.

BIB
LIO
GR
AFÍ
A
Kulminskaya, A.A., Tomsen, K.K., Shabalin, K.A., Sidorenko, I.A., Eneyskaya,
E.V., Savel’ev, A.N., Neustroev, K.N. 2001. Isolation, enzymatic properties, and
mode of action of an exo-1,3-b-glucanase from Trichoderma viride. Eur J Biochem
268:6123–6131.
Lakshmi, T., Chandra, B. 2004. Evaluation of agro-industrial wastes for conidia
based inoculum production of bio-control agent: Trichoderma harzianum. J Sci Ind
Res India 63:807-812.
Lee, H., Suh, S.S., Park, E., Cho, E., Ahn, J., Kim, S.G., Kwon, Y.M., Lee, I.
2000. The Agamous-Like 20 MADS domain protein integrates floral inductive
pathways in Arabidopsis. Gene Dev 14:2366–2376.
Litz, R.E. 2000. World mango breeding problems and perspectives: a
biotechnology overview. In: Proceedings Simposium Mango. Control de la floración
y mejoramieno genético. INIFAP 1-7.
Liu, B., Glenn, D., Buckley, K. 2008. Trichoderma communities in soils from
organic, sustainable, and conventional farms, and their relation with Southern
blight of tomato. Soil Biol Biochem 40:1124-1136.
Liu, C., Thong, Z., Yu, H. 2009. Coming into bloom: the specification of floral
meristems. Development. 136:3379-3391.
Liu, X., Grieve, C. 2009. Accumulation of chiro-inositol and other non-structural
carbohydrates in Limonium species in response to saline irrigation waters. J Amer
Soc Hort Sci 34:329-336.
Livak, K.J., Schmittgen, T.D. 2001. Analysis of relative gene expression data
using Real-Time quantitative PCR and the 2-ΔΔCt Method. Methods 25:402–408.

BIB
LIO
GR
AFÍ
A
Lo, C.T. 1998. General mechanisms of action of microbial biocontrol agents.
Plant Pathol Bull 7:155-166.
Maganhotto, S., Silva, C.M., Faria Vieira, R., Nicolella, G. 2003. Paclobutrazol
effects on soil microorganisms. Appl Soil Ecol 22:79-86.
Magnani, G.S., Didonet, C.M., Cruz, L.M., Picheth, C.F., Pedrosa, F.O., Souza,
E.M. 2010. Diversity of endophytic bacteria in Brazilian sugarcane. Genet Mol Res
9:250-258.
Mandels, M., Reese, E.T. 1960. Induction of cellulase in fungi by cellobiose. J
Bacteriol 79:816-826.
Maurya, D.P., Singh, D., Pratap, D., Maurya, J.P. 2012. Optimization of solid
state fermentation conditions for the production of cellulase by Trichoderma reesei,
J Environ Biol 33:5-8.
Medeiros, P.M., Simoneita, B.R.T. 2007. Analysis of sugars in environmental
samples by gas chromatography–mass spectrometry. J Chromatogr A 1141:271–
278.
Mejía, G.L., Martínez, C.H., Betancourt, G.J., Castrillón, C.C. 2007.
Aprovechamiento del residuo agroindustrial del mango común (Mangifera indica
L.) en la obtención de azúcares fermentables. Ingeniería y Ciencia 3:41-62.
Mendoza, W.S. 1977. Preharvest anthracnose control in mango. Philippine
Phytopathology 13:50-53.
Messner, R., Kubicek, C.P. 1991. Carbon source control of cellobiohydrolase I
and II formation by Trichoderma reesei. Appl Environ Microbiol 57:630-635.

BIB
LIO
GR
AFÍ
A
Mia, M.A.B., Shamsuddin, Z.H., Mahmood, M. 2010. Use of plant growth
promoting bacteria in banana: a new insight for sustainable banana production. Int
J Agric Biol 12:459-467.
Michel, A.A., Rebolledo, O., Lezama, R., Ochoa, M., Mesina, J.C., Samuels,
G.J. 2001. Especies de Trichoderma en suelos cultivados con mango afectado por
“escoba de bruja” y su potencial inhibitorio sobre Fusarium oxysporum y F.
subglutinans. Rev Mex Fitopat 19:154-160.
Miller. G. 1959. Use of dinitrosalicylic acid reagent for determination of reducing
sugar. Anal Chem 31:426-428.
Mora, V.M., Martínez-Carrera, D. 2007. Investigaciones básicas, aplicadas y
socioeconómicas sobre el cultivo de setas (Pleurotus) en México. Capítulo 1.1, pp.
7-26. In: El Cultivo de Setas Pleurotus spp. en México. J. E. Sánchez, D. Martínez-
Carrera, G. Mata & H. Leal (Eds.). ECOSUR, México, D.F.
Mortensen, L.M. 1986. Effect of relative humidity on growth and flowering of
some greenhouse plants. Sci Hortic 29:301-307
Mortley, D.G., Bonsi, C.K., Loretan, P.A., Hill, W.A., Morris, C.E. 2000. High
relative humidity increases yield, harvest index, flowering, and gynophore growth of
hydroponically grown peanut plants. HortScience 35:46-8.
Muñoz-Fambuena, N., Mesejo, C., Gonzalez-Mas, M.C., Primo-Millo, E., Agusti,
M., Iglesias, D.J. 2011. Fruit regulates seasonal expression of flowering genes in
alternate-bearing Moncada' mandarin. Ann Bot 108:511-519.
Nakagawa, M., Honsho, C., Kanzaki, S., Shimizu, K., Utsunomiya, N. 2012.
Isolation and expression analysis of flowering locus t-like and gibberellin
metabolism genes in biennial-bearing mango trees. Sci Hortic 139:108-117.

BIB
LIO
GR
AFÍ
A
Naqvi, S.A., M. H. Diseases of fruits and vegetables: diagnosis and
management. Kluner Academic Publisher. Netherlands 2004.
NCBI. 2010. National Center for Biotechnology Information. Recuperado el 18
de Octubre de 2010.
http://www.ncbi.nlm.nih.gov/Taxonomy/Browser/wwwtax.cgi?id=474922.
Neilands, J.B. 1982. Microbial envelope proteins related to iron. Annu Rev
Microbiol 36:285-309.
Nelson S.C. 2008. Mango Anthracnose (Colletotrichum gloeosporioides). Plant
Dis 48:1-9.
Nilsson, O., Lee, I., Blazquez, M. A. 1998. Flowering-time genes modulate the
response to LEAFY activity. Genetics 150:403-410.
Núñez-Eliséa, R., Davenport, T.L. 1995. Effect of leaf age, duration of cool
temperature treatment, and photoperiod on bud dormancy release and floral
initiation in mango. Sci Hort 62:63-73.
Núñez-Eliséa, R., Davenport, T.L., Caldeira, M.L. 1996. Control of bud
morphogenesis in mango (Mangifera indica L.) by girdling, defoliation and
temperature modification. J Hort Sci 71:25-40.
Ojumu, T.V., Solomon, B.O., Betiku, E., Layokun, S.K., Amigun, B. 2003.
Cellulase production by Aspergillus flavus isolate NSPR101 fermented in sawdust,
bagasse and corncob, African J Biotechnol 2:150-152.
Onofre-Lemus, J., Hernandez-Lucas, I., Girard, L., Caballero-Mellado, J. 2009.
ACC (1-Aminocyclopropane-1-Carboxylate) deaminase activity, a widespread trait

BIB
LIO
GR
AFÍ
A
in Burkholderia species, and its growth-promoting effect on tomato Plants. Appl
Environ Microbiol 75:6581-6590.
Opeke, L.K. 1982. Tropical tree crops. Chichester, UK.: John Wiley and Sons.
312.
Ordoñez, S.L. 2002. Proyecto, estudio, generación de residuos vegetales en el
valle de Cauca. Colombia: Universidad Nacional de Colombia.
Oriol, E., Raimbault, M., Roussos, S., Viniegra-Gonzalez, G. 1988. Water and
water activity in solid state fermentation of cassava starch by Aspergillus niger,
Appl Microbiol Biotechnol 27:498-503.
Osuna-García, J.A., Báez-Sañudo, R., Medina-Urrutia, V.M., Chávez-Contreras,
X. 2001. Residualidad de paclobutrazol en frutos de mango (Mangifera indica L.)
‘Tommy Atkins’. Revista Chapingo Serie Horticultura 7:275-282.
Otero, L., Ortega, F., Morales, M. 1998. Participación de la arcilla y la materia
orgánica en la capacidad de intercambio catiónico de vertisoles de la provincia
Granma. Terra latinoamericana 16:189-194.
Penrose, D.M. Glick, B.R. 2003. Methods for isolating and characterizing ACC
deaminase-containing plant growth-promoting rhizobacteria. Physiol Plant 118:10-
15.
Peres, N.A., Souza, N.L., Peever, T.L., Timmer, L.W. 2004. Benomyl sensitivity
of isolates of Colletotrichum gloesporioides from Citrus. Plant Dis 88:125-130.
Pérez, L. 1993. Enfermedades de las plantas. Medellín: Lealon, 46-51.

BIB
LIO
GR
AFÍ
A
Perfect, S.E., Hughes, H.B., O’connell, R.J., Green, J.R. 1999. Colletotrichum: a
model genus for studies on pathology and fungal-plant interactions. Fungal Genet
Biol 27:186-198.
Peters, T. 2003. Optimización del aprovechamiento energético de biogás de
vertederos. Rev Tec Residuos 13:34-37.
Ploetz, R.C. 1999. Malformation: a unique and important disease of mango,
Mangifera indica L. In: Summerell BA (eds) Paul E. Nelson Memorial Symposium.
APS Press, St. Paul, pp 1–8 ISBN 0890542686
Ploetz, R.C. 2009. Antracnosis en mango: Manejo de la enfermedad más
importante pre y postcosecha. Universidad de la Florida. Departamento de
Fitopatologia. Reporte. p. 1-11.
Ploetz, R.C., Prakash, O. 1997. Foliar, floral and soil borne diseases. In: The
mango: Botany, production and uses. CAB International, Willingfrod, UK. 281-326.
Pongsomboon, W., Subhadrabandhu, S., Stephenson, R.A. 1997. Some
aspects of the ecophysiology of flowering intensity of mango (Mangifera indica L.)
cv. Nam Dok Mai in a semi-tropical monsoon Asian climate. Sci Hortic 70:45-56.
Protacio, C.M. 2000. A model for potassium nitrate-induced flowering in mango.
Acta Hort 509:545-552.
Putterill, J., Laurie, R., Macknight, R. 2004. It´s time to flower the genetic control
of flowering time. Bioassays 26:363-373.
Rader, U., Broda, P. 1985. Rapid preparation of DNA from filamentous fungi.
Lett Appl Microbiol 1:17–20.

BIB
LIO
GR
AFÍ
A
Rahman, M.A., Kadir, J., Mahmud, T.M.M., Abdul Rahman, R., Begum, M.M.
2007. Screening of antagonistic bacteria for biocontrol activities on Colletotrichum
gloeosporioides in papaya. Asian J Plant Sci 6:12-20.
Ramirez, F., Davenport, T.L. 2010. Mango (Mangifera indica L.) flowering
physiology. Sci Hortic 126:65-72.
Ramot, O., Viterbo, A., Friesem, D., Oppenheim, A., Chet, I. 2004. Regulation of
two homodimer hexosaminidases in the mycoparasitic fungus Trichoderma
asperellum by glucosamine. Curr Genet 45:205–213.
Reece, P.C., Furr, J.R., Cooper, W.C. 1949. Further studies of floral induction in
the Haden mango (Mangifera indica L.). Am J Bot 36:734-740.
Rendón, M.R., Aguilar, A.J., Zarazúa, E.J., Bucio, P.I. 2006. Transferencia de
tecnología bajo la metodología de redes. Morelia, Michoacán.: SAGARPA.
Requena, N., Jimenez, I., Toro, M., Barea, J.M. 1997. Interactions between
plant-growth-promoting rhizobacteria (PGPR), arbuscular mycorrhizal fungi and
Rhizobium spp. in the rhizosphere of Anthyllis cytisoides, a model legume for
revegetation in mediterranean semi-arid ecosystems. New Phytol 136:667-677.
Rolland, F., Baena, E., Sheen, J. 2006. Sugar sensing and signaling in plants:
conserved and novel mechanisms. Annu Rev Plant Biol 57:675-709.
Ross, G.S., Elder, P.A., McWha, J.A., Pearce, D., Pharis, R.P. 1987. The
development of an indirect enzyme-linked immunoassay for abscisic acid. Plant
Physiol 85:46–50.
Rossi-Rodrigues, B.C., Brochetto-Braga, M.R., Tauk-Tornisielo, S.M., Cano
Carmona, E., Marques Arruda, V., Chaud Netto, J. 2009. Comparative growth of

BIB
LIO
GR
AFÍ
A
Trichoderma strains in different nutritional sources, using bioscreen c automated
system. Braz J Microbiol 40:404-410.
Ruzin, E. Plant microtechnique and microscopy. Oxford University press. United
States of America 1999.
SAGARPA. 2010. Servicio de Información Agroalimentaria y Pesquera.
Recuperado el 11 de 06 de 2012, de http://www.siap.gob.mx/
Saldarriaga, A., Bernal, J., Tamayo, P. 1997. Enfermedades del cultivo del
tomate de árbol en Antioquia: guía de reconocimiento y control. Boletín Técnico,
Corporación Colombiana de Investigación Agropecuaria (Corpoica); C.l. La Selva,
Rionegro (Antioquia), 8-10.
Samach, A., Onouchi, H., Gold, S., Ditta, G., Schwarz, Z., Yanofsky, M.F.,
Coupland, G. 2000. Distinct role of CONSTANS target genes in reproductive
development of Arabidopsis. Science 288:1613-1616.
Santoyo, G., Valencia-Cantero, E., Orozco-Mosqueda, M.C., Peña-Cabriales,
J.J., Farías-Rodríguez, R. 2010. Papel de los sideróforos en la actividad
antagónica de Pseudomonas fluorescens zum80 hacia hongos fitopatógenos.
Terra Latinoamericana 28:53-60.
Sanz, L., Montero, M., Redondo, J., Llobell, A., Monte, E. 2005. Expression of
an a-1, 3-glucanase during mycoparasitic interaction of Trichoderma asperellum.
FEBS Journal 272:493-499.
Sareen, R., Shah, A. 2011. Hypersensitivity manifestations to the fruit mango.
Asia Pac Allergy 1: 43-49.

BIB
LIO
GR
AFÍ
A
Saval, S. 2012. Aprovechamiento de Residuos Agroindustriales: Pasado,
Presente y Futuro. BioTecnología 16:14-46.
Schmidt, S., Blom, J.F., Pernthaler, J., Berg, G., Baldwin, A., Mahenthiralingam,
E., Eberl, L. 2009. Production of the antifungal compound pyrrolnitrin is quorum
sensing-regulated in members of the Burkholderia cepacia complex. Environ
Microbiol 11:1422-1437.
Schuster, A., Schmoll, M. 2010. Biology and biotechnology of Trichoderma. Appl
Microbiol Biotechnol 87:787-799.
Schwyn, B., Neilands, J.B. 1987. Universal chemical assay for the detection and
determination of siderophores. Anal Biochem 160:47-56.
Serrano-Carreon, L., Hathout, Y., Bensoussan, M., Belin, J.M. 1992. Production
of 6-pentyl-α-pyrone by Trichoderma harzianum from 18∶n fatty acid methyl esters.
Biotechnol Lett 14:1019- 1024.
Shaban, A.E.A. 2009 Vegetative growth cycles of some mango cultivars in
relation to flowering and fruiting. World J Agr Sci 5:751-759.
Shenker, M., Oliver, I., Helmann, M., Hadar, Y., Chen, Y. 1992. Utilization by
tomatoes of iron mediated by a siderophore produced by Rhizopus arrhizus. J
Plant Nutr 15:2173-2182.
Siddiqui, Z.A. 2005. PGPR: biocontrol and biofertilization. Springer 111-142.
Singh, P. 2011. Integrated management of storage anthracnose of mango. J
mycol pl pathol 41:63-66.

BIB
LIO
GR
AFÍ
A
Singh, R.K., Ali, S.A., Nath, P., Sane, V.A. 2011. Activation of ethylene-
responsive p-hydroxyphenylpyruvate dioxygenase leads to increased tocopherol
levels during ripening in mango. J Exp Bot 62:3375-3385.
Smith, B.J., Black, L.L. 1990. Morphological, cultural and pathogenic variation
among Colletotrichum species isolated from strawberry. Plant Dis 74:69-76.
SMN. 2002. Servicio Meteorológico Nacional. Recuperado el 15 de 06 de 2009,
de http://smn.cna.gob.mx/
Soccol, C.R., Vandenberghe, L.P.S. 2003. Overview of applied solid-state
fermentation in Brazil. Biochem Eng J 13:205-218.
Soil Survey, 1996. Laboratory Methods Manual. U. S. Department of Agriculture,
Soil Conservation Service. Washington D.C., USA. 693.
Sreenath, H., Koegel, R., Moldes, A., Straub, R. 2001. Lactic acid production
from agriculture residues. Biotechnol Lett 23:179-184.
Stamets, P. 2003. Mycomedicinal: an information booket on medicinal
mushroom. Editorial Olympia.
Suresh, P.V., Chandrasekaran, M. 1998. Utilization of prawn waste for chitinase
production by the marine fungus Beauveria bassiana by solid state fermentation.
World J Microbiol Biotechnol 14:655–660.
Thomas, K.C., Hynes, S.H., Ingledew, W.M. 1994. Effects of particulate
materials and osmoprotectants on very-high-gravity ethanolic fermentation by
Saccharomyces cerevisiae, Appl Microbiol Biotechnol 60:1519-1524.

BIB
LIO
GR
AFÍ
A
Thompson, J.D., Gibson, T.J., Plewniak, F., Jeanmougin, F., Higgins, D.G.
1997. The ClustalX windows interface: flexible strategies for multiple sequence
alignment aided by quality analysis tools. Nucleic Acids Res 25:4876-4882.
Tiwari, D., Shuddhodan, P.M., Manisha, M. Dubeyb, R.S. 1999. Biosorptive
behaviour of Mango (Mangifera indica) and Neem (Azadirachta indica) bark for
Hg2+, Cr3+ and Cd2+ toxic ions from aqueous solutions: a radiotracer study. Appl
Radiat Isotopes 50: 631-642.
Trejo-Márquez, M.A. 2010. Piel de mango, hueso de chocolate. Hypatia -
Revista de Divulgación Científico - Tecnológica del Estado de Morelos. 1-2
Trillas, I.M., Casanova, E., Cotxarrera, L., Orgovás, J., Borrero, C., Avilés, M.
2006. Compost from agricultural waste and the Trichoderma asperellum strain T-
34 suppress Rhizoctonia solani in cucumber seedlings. Biol Control 39:32-38.
Tsai, W.T. 2008. Management considerations and environmental benefit
analysis for turning food garbage into agricultural resources. Bioresour Technol
99:5309-5316.
Turnbull, C. 2011. Long-distance regulation of flowering time. J Exp Bot
62:4399-4413.
Turnbull, C.G.N., Anderson, K.L., Winston, E.C. 1996. Influence of gibberellin
treatment on flowering and fruiting patterns in mango. Aust J Exp Agr 36:603-611.
Urban, L., Alphonsout, L. 2007. Girdling decreases photosynthetic electron
fluxes and induces sustained photoprotection in mango leaves. Tree Physiol
27:345-352.

BIB
LIO
GR
AFÍ
A
Van Reeuwijk, L.P. 1993. Procedures for soil analysis. 4th Edition. Technical
paper No. 9, ISRIC, 102.
Van Soest, P.J., Wine, R.H. 1967. Use of detergents in the analysis of fibrous
feeds. IV. Determination of plant cell wall constituents. J Assoc Off Agric Chem
50:50-55.
Vázquez-Valdivia, V., Pérez-Barraza, M.H., Osuna-García, J.A. 2009. Effect of
gibberellic acid on flowering of 'Ataulfo' mangos. Acta Hort 820:413-418.
Verma, J.P., Yadav, J., Tiwari, K.N., Lavakush, Singh, V. 2010 Impact of plant
growth promoting rhizobacteria on crop production. Int J Agric Res 5:954-983.
Verma, M., Satinder, K.B., Tyagi, R.D., Surampalli, R.Y., Valéro, J.R. 2007.
Antagonistic fungi, Trichoderma spp.: Panoply of biological control. Biochem Eng J
37:1-20.
Viniegra-González, G., Favela-Torres, E., Aguilar, C.E., Rómero-Gomez, S.J.,
Díaz-Godínez, G., Augur, C. 2003. Advantages of fungal enzyme production in
solid state over liquid fermentation systems, Biochem Eng J 13:157-167.
Viterbo, A., Landau, U., Kim, S., Chernin, L., Chet, I. 2010. Characterization of
ACC deaminase from the biocontrol and plant growth-promoting agent
Trichoderma asperellum T203. FEMS Microbiol Lett 1–7.
Walkley, A. 1947. A critical examination of a rapid method for determining
organic carbon in soils: Effect of variations in digestion conditions and of organic
soil constituents. Soil Sci 63:251-263.

BIB
LIO
GR
AFÍ
A
Walkley, A., Black, I.A. 1934. An examination of Degtjareff method for
determining soil organic matter and a proposed modification of the chromic acid
titration method. Soil Sci 37:29-37.
Watanabe, S., Kumakura, K., Kato, H., Iyozumi, H., Togawa, M., Nagayama, K.
2005. Identification of Trichoderma SKT-1, a biological control agent against
seedborne pathogens of rice. J Gen Plant Pathol 71:351–356.
Weisburg, W.G., Barns, S.M., Pelletier, D.A., Lane, D.J. 1991. 16S Ribosomal
DNA Amplification for phylogenetic study. ASM 173:697-703.
White, T.J., Bruns, T., Lee, S., Taylor, J. 1990. Amplification and direct
sequencing of fungal ribosomal RNA genes for phylogenetics. In: Innis, M.A.,
Gelfand, D.H., Sninsky, J.J., White, T.J. (Eds.), PCR Protocols: A Guide to
Methods and Applications. Academic Press, San Diego. pp.315–322.
Wu, C.H., Bernard, S.M., Andersen, G.L., Chen, W. 2009. Developing microbe-
plant interactions for applications in plant-growth promotion and disease control,
production of useful compounds, remediation and carbon sequestration. Microb
Biotechnol 2:428-440.
Xia, L., Shen, X. 2004. High-yield cellulase production by Trichoderma reesei
ZU-02 on corn corb residue. Bioresour Technol 91:259-262.
Yedidia, I., Benhamou, N., Chet, I. 1999. Induction of defense responses in
cucumber plants (Cucumis sativus L.) by the biocontrol agent Trichoderma
harzianum. Appl Environ Microbiol 65:1061-1070.
Yenjit P., Intanoo W., Chamswarng C., Siripanich J., Itana W. 2004. Use of
promising bacterial strains for controling anthracnose on leaf and fruit of mango
caused by Colletotrichum gloeosporioides. Walailak J Sci Tech 1:56-59.

BIB
LIO
GR
AFÍ
A
Yepes, S.M., Montoya Naranjo, L.J., Orozco Sánchez, F. 2008. Valorización de
residuos agroindustriales –frutas- en Medellín y el sur del valle del Aburrá,
Colombia. Rev Fac Nal Agr Medellìn 61:4422-4431.
Zadoks, J.C, Schein, R.D. 1979. Epidemiology and plant disease management.
Oxford University Press. New York, USA. p. 427.
Zhang, T., Ying, Z., Davenport, T. L. 2005. Isolation and characterization of a
CONSTANS-like gene in mango. Plant Biology, p. 151.
View publication statsView publication stats


















