Centro de Investigación Científica y de Educación ... · como un grado de diferenciación...
102
Centro de Investigación Científica y de Educación Superior de Ensenada, Baja California Maestría en Ciencias en Acuicultura Diversidad genética y éxito reproductivo del Jurel de Castilla Seriola lalandi (Valenciennes, 1833) en cautiverio Tesis para cubrir parcialmente los requisitos necesarios para obtener el grado de Maestro en Ciencias Presenta: Eduardo Martínez Matus Ensenada, Baja California, México 2016
Transcript of Centro de Investigación Científica y de Educación ... · como un grado de diferenciación...

Centro de Investigación Científica y de Educación Superior de Ensenada, Baja California
Maestría en Ciencias
en Acuicultura
Diversidad genética y éxito reproductivo del Jurel de Castilla Seriola lalandi (Valenciennes, 1833) en cautiverio
Tesis para cubrir parcialmente los requisitos necesarios para obtener el grado de
Maestro en Ciencias
Presenta:
Eduardo Martínez Matus
Ensenada, Baja California, México
2016

Tesis defendida por
Eduardo Martínez Matus
y aprobada por el siguiente Comité
_________________________________ Dra. Fabiola Lafarga De La Cruz
Codirectora de tesis
__________________________________ Dra. Blanca Claudia Farfán
Codirectora de tesis
Eduardo Martínez Matus © 2016 Queda prohibida la reproducción parcial o total de esta obra sin el permiso formal y explícito del autor y director de la tesis.
Dr. Benjamín Barón Sevilla
Dra. Clara Elizabeth Galindo Sánchez
Dr. Benjamín Barón Sevilla Coordinador del Posgrado en Acuicultura
Dra. Rufina Hernández Martínez Directora de Estudios de Posgrado

ii
Resumen de la tesis que presenta Eduardo Martínez Matus como requisito parcial para la obtención del grado de Maestro en Ciencias en Acuicultura.
Diversidad genética y éxito reproductivo del Jurel de Castilla Seriola lalandi (Valenciennes, 1833) en cautiverio
Resumen aprobado por:
_________________________________ Dra. Fabiola Lafarga De La Cruz
Codirectora de tesis
__________________________________ Dra. Blanca Claudia Farfán
Codirectora de tesis
El Jurel de Castilla, Seriola lalandi, es una especie marina de distribución cosmopolita de importancia para la pesca y la acuicultura. Australia y Nueva Zelanda contribuyen con el 95% del total de la producción mundial (~ 5,000 tons/año); mientras que otros países como Chile, México y EUA se encuentran desarrollando su cultivo comercial. El primer criadero de S. lalandi en México, está basado en el uso de reproductores silvestres, y las progenies F1 serán en el futuro sus reproductores domesticados. Sin embargo, aún se desconoce el estatus genético de las poblaciones silvestres mexicanas por lo que los reproductores no están genéticamente caracterizados. Además, la diferenciación genética entre las poblaciones del Pacífico noroeste, Pacífico noreste y hemisferio sur, sugieren la reclasificación taxonómica de S. lalandi por lo que la translocación de estas especies puede repercutir en la riqueza ecológica y genética de nuestras especies locales. En este trabajo se caracterizó genéticamente a las poblaciones silvestres y en cautiverio de S. lalandi a nivel regional y global, evaluando el grado de diferenciación entre ellas; así como el éxito reproductivo de la especie en cautiverio. A partir de nueve loci microsatélites, se analizaron seis poblaciones silvestres (cuatro mexicanas, una de Chile y una de Australia) y varias poblaciones cultivadas de diversos orígenes (MEX, EUA, CHL, AUS). No se identificaron diferencias significativas en la diversidad genética entre poblaciones silvestres ni entre las poblaciones cultivadas. La riqueza alélica (Ra) de la población F1 de AUS fue significativamente menor en comparación con la población de Bahía de los Ángeles, México. En la F1 en cautiverio de las poblaciones de Bahía de Todos Santos, Bahía Magdalena, Chile y Australia, se cuantificó una pérdida de la variabilidad genética del 23, 46, 28 y 38%, respectivamente. Se observó ausencia de estructura genética en las localidades mexicanas, así como un grado de diferenciación genética significativo entre las poblaciones de ambos hemisferios. Por otro lado, la contribución parental de hembras y machos a la descendencia fue desigual entre sexos. El 100% de los machos y el 55% de las hembras en el lote uno contribuyeron a la descendencia, mientras que en el lote dos solo el 64% de los machos y el 23% de las hembras participaron. El grado moderado de endogamia (> 0.1) en tres poblaciones silvestres de la Península de Baja California, aunado a la pérdida de la diversidad genética durante la primera generación, sugieren la necesidad de implementar un manejo reproductivo adecuado en los núcleos de producción del Jurel de Castilla en México.
Palabras clave: microsatélites, Seriola lalandi, éxito reproductivo, diversidad genética.

iii
Abstract of the thesis presented by Eduardo Martínez Matus as a partial requirement to obtain the Master of Science degree in Aquaculture. Genetic Diversity and Reproductive Success of yellowtail kingfish Seriola lalandi (Valenciennes, 1833)
in Captivity
Abstract approved by:
_________________________________ PhD. Fabiola Lafarga De La Cruz
Thesis Co-director
__________________________________ PhD. Blanca Claudia Farfán
Thesis Co-director
The Yellowtail kingfish, Seriola lalandi, is a cosmopolitan marine species supporting fishery and aquaculture industries worldwide. Yellowtail kingfish aquaculture industry from Australia and New Zealand accounts 95% of the world total production (~ 5,000 tons/year), while other countries like Chile, Mexico and USA have been recently started developing its commercial mariculture. Although hatchery production of S. lalandi is growing rapidly, unfortunately, in Mexico the genetic status of wild and captive S. lalandi populations is unknown. The first S. lalandi hatcheries are based on wild broodstock and F1 progenies will be, in the future, their domesticate broodstock. Recently, a taxonomic reclassification of S. lalandi has been suggested, based on the genetic differentiation among the Northwest Pacific, the Northeast Pacific and the Southern Hemisphere. This is of our concern because translocation of these species may have an important impact on the ecological and genetic richness of our local populations. The purpose of this study was to evaluate the genetic diversity of wild and culture S. lalandi populations, at global and regional level; and to assess the S. lalandi reproductive success in captivity. Using nine microsatellites loci, six wild S. lalandi populations (four from Mexico, one from Chile and one from Australia) and some other cultured populations (MEX, USA, CHL, AUS) were evaluated. In terms of genetic diversity, no significant differences were identified among wild populations and neither among cultivated stocks. Allelic richness (Ra) of the F1 stock from AUS was significant lower than in the Bahia de los Angeles wild population. A loss of genetic variability of 23, 46, 28 and 38% was recorded on the F1 stocks from Bahia Todos Santos, Bahia Magdalena, Chile and Australia, respectively. Among the Mexican locations evaluated a lack of genetic structure were observed, whereas between the northern and the southern hemispheres a strong genetic divergence was assess. Female and male contribution to progeny were unequal from two tanks evaluated. In tank one, 100% of males and only 55% of females contribute to the F1 progeny, while in tank two just 64% of males and 23% of females participate. A moderate inbreeding coefficient (>0.1) of three of the four wild populations in Baja California Peninsula and the loss of genetic diversity of first generation progeny produce in captivity suggests than an adequate reproductive management in the Yellowtail kingfish culture in México should will be implemented. Keywords: Microsatellites, Seriola lalandi, parental assignment, genetic diversity.

iv
Dedicatoria
A mi madre, Ma. Del Rosario Matus, por fomentar mi creatividad, imaginación y curiosidad desde
pequeño, que indirectamente desarrollaron mi vocación científica; por su inmenso apoyo y cariño durante
todo este tiempo.
A mis hermanas, Karen y Aby, por hacerme reír y por los buenos momentos.
A mi sobrino, J. Isaac, por alegrar nuestras vidas.

v
Agradecimientos
Al Centro de Investigación Científica y de Educación Superior de Ensenada (CICESE), por permitirme cursar mis estudios de maestría. Al Consejo Nacional de Ciencia y Tecnología (CONACYT), por administrar los recursos de la sociedad mexicana, y otorgarme una beca de maestría, además de un complemento económico (beca mixta) durante mi estancia en el extranjero. Al CPP del Departamento de Acuicultura, por su respaldo académico antes, durante y después de mi estancia académica. Así como, por el apoyo económico otorgado. A Ocean Baja Labs, por las muestras y compartir información valiosa de la empresa. A Roberto Flores, Mauricio y Jazmín por su colaboración en el proyecto. A mis directoras de tesis, por sus contribuciones académicas en el transcurso de la investigación, y por el apoyo económico durante los tiempos difíciles. A la Dra. Fabiola Lafarga De La Cruz por transmitirme su optimismo y espíritu científico, por compartir su motivación en el proyecto, por sus colaboraciones que me permitieron realizar una estancia en Australia, por las sugerencias en la organización de los resultados y por sus observaciones. A la Dra. Blanca Claudia Farfán, por su realismo y honestidad, por sus valiosas clases de redacción y gramática, por sus importantes sugerencias en la estructura de la discusión, y por sus dos grandes consejos de vida “leer y torturar a los datos”. A mi comité de tesis, por las sugerencias en los avances de tesis, por su disponibilidad, agilidad y observaciones en las revisiones. Al Dr. Benjamín por la sugerencia de incluir las tallas de los reproductores. A la Dra. Clarissa, por hacerme dudar de los parámetros genéticos. I would like to thank Dr. Wayne Knibb, for the time, patience and support I received during my visit at the University of the Sunshine Coast, AU. I also want to thank to genetics group: Dra. Abigail Elizur, Ajith, Luu, Chontida, Giang, Jane, Dan, for your hospitality, and for assisting me in genotyping. Al grupo de genética de acuicultura del CICESE. Al Dr. Miguel Ángel del Río por invitarme a pertenecer al grupo, por las recomendaciones de la literatura científica, por sus aportaciones respecto al tema y en general, por cuestionar todo. A los integrantes del grupo, por compartir su trabajo y ampliar mi perspectiva de la genética y sus aplicaciones en la acuicultura. A mis compañeros de maestría por su increíble generosidad durante el tiempo crítico de mi posgrado, por las historias que compartieron conmigo, los momentos y el apoyo moral. A Ana, Benito, Bety (Irving), César, Enrique, Erik, Gabo, Jonathan, Josué, Juan, Laura, Lili, Noemí, Omar, Romy, Sergio ¡Gracias a todos!. A los profesores del CICESE que contribuyeron con mi aprendizaje. A la Dra. Beatriz Cordero, por brindarme la oportunidad de participar en el proceso de selección, aun cuando no estaba titulado, y por el apoyo en la maestría. Al técnico titular del laboratorio de genética, Carmen Elvira Vargas Peralta por su apoyo en la parte experimental del proyecto, sus enseñanzas y en su paciencia.

vi
A mi familia y amigos, por estar ahí apoyándome en todo momento. A Zoila Guadalupe Martínez, gracias por el apoyo y los patrocinios. A Yara y Ulises por su amistad y por el invaluable apoyo durante mi convalecencia. A Reez Lucas por su asistencia en los mapas. A todos los que contribuyeron directa e indirectamente en la culminación de este proyecto. ¡Muchas gracias!.

vii
Tabla de contenido Página
Resumen en español ....................................................................................................................... ii
Resumen en inglés .......................................................................................................................... iii
Dedicatoria ...................................................................................................................................... iv
Agradecimientos ............................................................................................................................. v
Lista de figuras ................................................................................................................................ x
Lista de tablas .................................................................................................................................. xiii
Capítulo 1. Introducción
1.1 Antecedentes ............................................................................................................................ 2
1.1.1 Biología y distribución de Seriola lalandi ......................................................................... 2
1.1.2 Importancia económica de la especie ............................................................................... 4
1.1.3 Programas de selección genética: domesticación ............................................................ 6
1.1.4 Marcadores moleculares tipo microsatélites.................................................................... 8
1.1.5 Diversidad y estructura genética ...................................................................................... 9
1.1.6 Éxito reproductivo en cautiverio ....................................................................................... 10
1.2 Justificación ............................................................................................................................... 10
1.3 Hipótesis .................................................................................................................................... 11
1.4 Objetivos ................................................................................................................................... 12
1.4.1 Objetivo general ................................................................................................................ 12
1.1.1 Objetivos específicos ........................................................................................................ 12
Capítulo 2. Metodología
2.1 Origen de las muestras .............................................................................................................. 13
2.2 Muestreo de reproductores ...................................................................................................... 15
2.3 Muestreo de juveniles ............................................................................................................... 16
2.4 Extracción de DNA genómico .................................................................................................... 16
2.5 Amplificación de los marcadores genéticos tipo microsatélites ............................................... 17
2.6 Genotipado ................................................................................................................................ 19
2.7 Análisis de diversidad genética ................................................................................................. 19
2.8 Análisis de estructura genética ................................................................................................. 20
2.9 Análisis de asignación parental ................................................................................................. 21

viii
Capítulo 3. Resultados
3.1 Muestreos genéticos ................................................................................................................. 22
3.2 Extracción de DNA genómico .................................................................................................... 22
3.3 Amplificación de microsatélites ................................................................................................ 24
3.4 Genotipado ................................................................................................................................ 24
3.5 Análisis de diversidad genética ................................................................................................. 25
3.5.1 Poblaciones de cultivo ...................................................................................................... 25
3.5.1.1 F1 de Bahía de Todos Santos ....................................................................................... 25
3.5.1.2 F1 de Bahía Magdalena ............................................................................................... 26
3.5.1.3 F1 de Estados Unidos de América ............................................................................... 26
3.5.1.4 F1 de Chile ................................................................................................................... 27
3.5.1.5 F1 de Australia ............................................................................................................. 27
3.5.2 Poblaciones silvestres ....................................................................................................... 28
3.5.2.1 Bahía de Todos Santos, BC, México ............................................................................. 28
3.5.2.2 Ejido Eréndira, BC, México .......................................................................................... 28
3.5.2.3 Bahía Magdalena, BCS, México ................................................................................... 29
3.5.2.4 Bahía de los Ángeles, BC, México ................................................................................ 29
3.5.2.5 Antofagasta, Chile ....................................................................................................... 30
3.5.2.6 Nueva Gales del Sur, Australia .................................................................................... 30
3.5.3 Análisis estadístico ............................................................................................................ 30
3.6 Análisis de estructura genética ................................................................................................. 33
3.6.1 Análisis de grupos y asignación de individuos .................................................................. 33
3.6.2 Análisis discriminante de componentes principales ......................................................... 36
3.6.3 Similitud genética entre poblaciones ................................................................................ 37
3.6.4 Diferenciación genética poblacional ................................................................................. 39
3.6.5 Análisis de varianza molecular (AMOVA) .......................................................................... 41
3.6.6 Aislamiento por distancia .................................................................................................. 42
3.7 Análisis de asignación parental ................................................................................................. 43
3.7.1 Contribución parental general .......................................................................................... 43
3.7.2 Aporte parental individual ................................................................................................ 44
3.7.2.1 Lote uno ....................................................................................................................... 44
3.7.2.2 Lote dos ....................................................................................................................... 45
3.7.3 Fenotipo y contribución parental ..................................................................................... 46
3.7.4 Grado de parentesco genético entre reproductores ........................................................ 48

ix
3.7.5 Deformidades .................................................................................................................... 49
Capítulo 4. Discusión
4.1 Generalidades de los microsatélites ......................................................................................... 52
4.2 Diversidad genética ................................................................................................................... 53
4.2.1 Caracterización genética regional de las poblaciones silvestres de Seriola lalandi .......... 53
4.2.2 Diversidad genética en poblaciones silvestres extranjeras .............................................. 54
4.2.3 Variabilidad genética en el cultivo de S. lalandi................................................................ 55
4.3 Estructura genética ................................................................................................................... 57
4.3.1 Estructura genética de las poblaciones mexicanas ........................................................... 57
4.3.2 Diferenciación genética en poblaciones mundiales de Seriola ......................................... 58
4.4 Éxito reproductivo en cautiverio (BC) ....................................................................................... 60
4.4.1 Relación entre el fenotipo y la contribución parental ...................................................... 63
4.4.2 Deformidades en el cultivo de S. dorsalis ......................................................................... 64
Capítulo 5. Conclusiones
Literatura citada .................................................................................................................... 67
Anexos .................................................................................................................................. 76

x
Lista de figuras
Figura
Página
1. Ejemplar de Seriola lalandi. Fuente: Australian National Fish Collection, CSIRO ........ 2
2. Distribución geográfica de Seriola lalandi. Los puntos indican probabilidades de ocurrencia. Puntos rojos: 80-100%, puntos lilas: 60-79%, puntos verdes: <20%. Fuente: www.aquamaps.org ....................................................................................... 4
3. Resultados a través del tiempo, de las tres posibilidades de manejo genético en los núcleos de producción. Modificado de Graham, 2009 ................................................ 7
4. Representación simplificada de los microsatélites en los cromosomas de un individuo ...................................................................................................................... 8
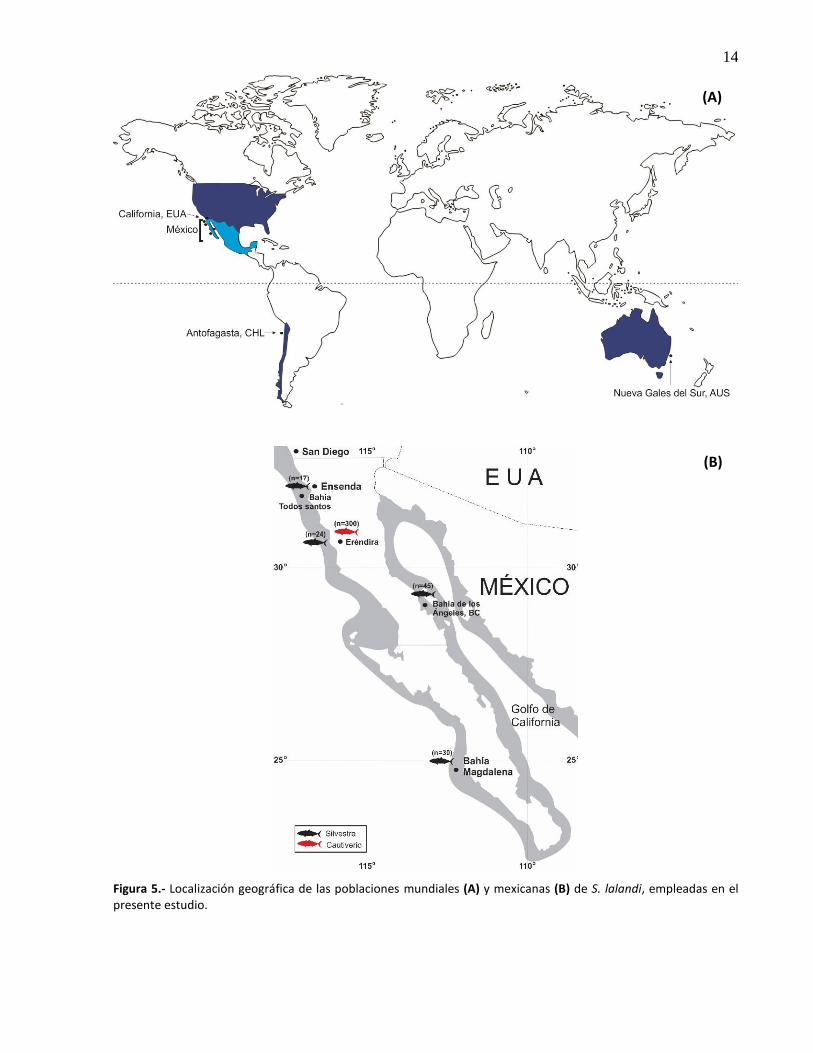
5. Localización geográfica de las poblaciones mundiales (A) y mexicanas (B) de S. lalandi, empleadas en el presente estudio .................................................................. 14
6. Representación del muestreo a los 30 días después de la eclosión en Seriola Sp ...... 15
7. Gel virtual generado para el grupo uno (A) y el grupo dos (B) ................................... 19
8. Electroforesis en gel de agarosa al 1% durante 45 min a 90 volts con buffer TBE 1X de muestras de S. lalandi provenientes del lote 2 (n= 20) Se observan fragmentos de DNA de alto peso molecular (> 12,000 pb) y en algunas muestras de bajo peso molecular, se incluyen las concentraciones de DNA obtenidas, mediante el protocolo de sales modificado, en unidades de ng/µl. Esc: marcador de referencia de 1 Kb ........................................................................................................................ 23
9. Concentración e índices de calidad del DNA obtenido (A. relación 260/280 y B. relación 260/230) con el protocolo de sales (puntos azules) y el protocolo comercial (puntos rojos) .............................................................................................................. 23
10. Ejemplo de la visualización de productos de PCR en gel de agarosa al 2% durante 80 min a 60 volts en buffer TBE 1X del grupo uno de microsatélites (Sel008, Sel011, Sel017, Sel019) de la población de cultivo de Bahía de los Ángeles (BA, n=14). Esc: marcador de referencia de 100 pb, Pos: Control positivo, Neg: Control negativo ...... 24
11. Gel virtual y electroferograma de un organismo heterocigoto (A) y un organismo homocigoto (B) para el locus Sel008 (fluorocromo FAM, azul), obtenido mediante análisis de fragmentos en secuenciador ...................................................................... 25

xi
12.
Representación del número de alelos (Na) y riqueza alélica (Ra) en los nueve loci microsatélites por población, las barras representan los valores máximos y mínimos, la línea dentro de las cajas representa la mediana. Las comparaciones múltiples entre las poblaciones según valor de significancia [*(p<0.05), **(p<0.01), ***(p<0.001)], están representados por la línea exterior ...........................................
31
13. Representación de la frecuencia de alelos nulos (Fn) y coeficiente de endogamia (Fis) en los nueve loci microsatélites por población, las barras representan los valores máximos y mínimos. la línea dentro de las cajas representa la mediana. Las comparaciones múltiples entre las poblaciones según el valor de significancia [*(p<0.05), **(p<0.01), ***(p<0.001)], están representados por la línea exterior.* en las localidades representa que su Fis difiere de cero ............................................. 32
14. Estructura genética de las poblaciones de Seriola lalandi mexicanas para dos (A), tres (B) y cuatro (C) componentes genéticos (valor de K asignado). Poblaciones de cultivo: F1BT= F1 de Bahía de Todos Santos, F1BM= F1 de Bahía Magdalena, y silvestres: BT= Bahía de Todos Santos, EE= Ejido Eréndira, BM= Bahía Magdalena y BA= Bahía de los Ángeles ............................................................................................. 34
15. Estructura genética de las poblaciones de Seriola lalandi mexicanas y extranjeras para dos (A), tres (B) y cuatro (C) componentes genéticos (valor de K asignado). Poblaciones de cultivo: F1BT= F1 de Bahía de Todos Santos, F1BM= F1 de Bahía Magdalena, F1EUA= F1 de Estados Unidos, F1CHL= F1 de Antofagasta, Chile, F1AUS= F1 de Nueva Gales del Sur, Australia, y silvestres: BT= Bahía de Todos Santos, EE= Ejido Eréndira, BM= Bahía Magdalena, BA= Bahía de los Ángeles, CHL= Chile y AUS= Australia ....................................................................................................................... 35
16. Plano cartesiano del análisis discriminante de componentes principales de las poblaciones empleadas del noreste del pacífico. Poblaciones de cultivo: F1BT= F1 de Bahía de Todos Santos, F1BM= F1 de Bahía Magdalena, F1EUA= F1 de Estados Unidos; y silvestres: BT= Bahía de Todos Santos, EE= Ejido Eréndira, BM= Bahía Magdalena y BA= Bahía de los Ángeles ....................................................................... 36
17. Plano cartesiano del análisis discriminante de componentes principales de las poblaciones del noroeste del Pacífico (poblaciones mexicanas y de EUA) y del hemisferio sur (Chile y Australia). Poblaciones de cultivo: F1BT= F1 de Bahía de Todos Santos, F1BM= F1 de Bahía Magdalena, F1EUA= F1 de Estados Unidos, F1CHL= F1 de Antofagasta, Chile, F1AUS= F1 de Nueva Gales del Sur, Australia, y silvestres: BT= Bahía de Todos Santos, EE= Ejido Eréndira, BM= Bahía Magdalena, BA= Bahía de los Ángeles, CHL= Chile y AUS= Australia ..................................................................... 37
18. Dendrograma inferido por el método del vecino más cercano a través de la Distancia genética de Nei (Da) en las poblaciones mexicanas, anclado a un grupo externo genéticamente diferente (AUS). .................................................................................. 38
19. Dendrograma inferido por el método del vecino más cercano a través de la Distancia genética de Nei (Da) en las poblaciones a nivel global ................................................ 39

xii
20. Correlación entre la distancia geográfica (km) y la distancia genética (Fst) de las poblaciones obtenidas ................................................................................................. 42
21. Porcentaje de reproductores que contribuyen a la descendencia por desove y total 44
22. Contribución parental individual del lote uno durante los tres eventos de desove. La letra M representa a los machos mientras que la letra H a las hembras. * = Reproductor con mayor contribución. NA = porcentaje de crías no asignadas .......... 45
23. Contribución parental individual del lote dos durante los tres eventos de desove. La letra M representa a los machos mientras que la letra H a las hembras. * = Reproductor con mayor contribución.......................................................................... 46
24. Relación entre la longitud estándar (cm) y la contribución parental en reproductores del lote uno (A) y dos (B). ........................................................................................... 47
25. Relación entre la longitud estándar (cm) y la contribución parental total (%) considerando la totalidad de reproductores disponibles ............................................ 48
26. Contribución parental (%) a la descendencia normal y deforme durante el mismo evento de desove. *Reproductor con mayor contribución, NA = no asignados. ....... 50
27. Variabilidad genética en organismos deformes y normales ........................................ 51
28. Barrera térmica ecuatorial. Fuente: NASA's Goddard Space Flight Center, http://svs.gsfc.nasa.gov/10953 ................................................................................... 60

xiii
Lista de tablas
Tabla Página
1. Muestreo genético e información adicional de las poblaciones empleadas ................ 13
2. Detalles de los marcadores moleculares tipo microsatélites que fueron utilizados en el presente estudio ........................................................................................................ 18
3. Master mix para PCR multiplex empleado en la amplificación de microsatélites ........ 18
4. Origen del muestreo y número de muestras empleadas para los análisis de diversidad genética y parental ...................................................................................... 22
5. Tamaño en número de bases de los fragmentos de PCR obtenidos para cada locus microsatélite amplificado .............................................................................................. 24
6. Valores de Fst (debajo de la diagonal) y Rst (arriba de la diagonal) obtenidos por pares de poblaciones ............................................................................................................... 40
7. Resultados del análisis de varianza molecular (AMOVA) basado en microsatélites en tres grupos de poblaciones: grupo uno (F1BT, BT), grupo dos (F1BM, BM), y grupo tres (EE, BA) .................................................................................................................. 41
8. Resultados del análisis de varianza molecular (AMOVA) basado en microsatélites entre dos grupos de poblaciones: Hemisferio norte (F1BT, F1BM, F1EUA, BT, EE, BM, BA), y Hemisferio sur (F1CHL, F1AUS, CHL, AUS) ......................................................... 42
9. Resultados de la Prueba de Mantel para la correlación lineal entre la distancia genética (FST) y la distancia geográfica expresada en kilómetros (km) y transformada a su logaritmo (Log) ....................................................................................................... 43
10. Grado de parentesco entre reproductores de cada lote ............................................... 49

1
Capítulo 1. Introducción
La pesca y la acuicultura contribuyen sustancialmente a preservar la seguridad alimentaria, debido a que
aportan anualmente 167 millones de toneladas de alimento para la humanidad (FAO, 2016). Sin embargo,
el inminente incremento en la pérdida de la biodiversidad marina por contaminación, sobreexplotación o
alteraciones debidas al cambio climático, podría llevar a un colapso de los recursos marinos que
comprometería la disponibilidad de alimentos para millones de personas en el mundo (Worm et al., 2006;
Thorpe et al., 1981). En un panorama en el que la pesca ha permanecido estática desde la década de los
años 80, la acuicultura, por su rápido crecimiento se ha convertido en una alternativa para enfrentar los
retos alimentarios que se avecinan. Sin embargo, es necesario optimizar los sistemas de producción e
incrementar la participación de la maricultura, actividad que actualmente contribuye con el 36% del total
de la producción acuícola (FAO, 2016).
Desde esta perspectiva, Seriola lalandi es una especie con alto potencial acuícola por su facilidad de
adaptarse al cautiverio, rápida tasa de crecimiento, así como por la excelente calidad de su carne (Symonds
et al., 2014; Sicuro y Luzzana, 2016). En México, el primer cultivo experimental de esta especie en jaulas
marinas inició en el año 2000, y estuvo basado principalmente en la captura de organismos silvestres
(Avilés y Castelló, 2004). Mientras que el año 2014, se estableció el primer laboratorio a nivel comercial
de producción de larvas de Seriola lalandi en el Ejido Eréndira, Baja California, el primero en Norteamérica
(Luening, 2014; Scarratt, 2014).
Una de limitantes para la expansión del cultivo de Seriola es la ausencia de programas de selección
genética, por lo que las poblaciones silvestres que se mantienen en cautiverio carecen de programas de
manejo reproductivo eficientes (Knibb, 2000; Lind et al., 2012; Sicuro y Luzzana, 2016;). Para evitar influir
negativamente en la capacidad adaptativa de las especies, es importante promover al mismo tiempo, la
conservación de los recursos genéticos acuáticos de las poblaciones silvestres, mediante el conocimiento
de su compatibilidad genética, evitando la introducción de especies y/o poblaciones exóticas que podrían
perturbar la biodiversidad local (Kapuscinski, 2008; ICES, 2005; Charles y Leith, 1999; FAO, 1995; Allendorf,
1991). Además, para mantener la viabilidad de los cultivos a largo plazo, es de vital importancia
implementar un manejo reproductivo en los sistemas acuícolas, para controlar los niveles de
consanguinidad dentro de los núcleos genéticos e identificar reproductores superiores, en términos de
fecundidad, crecimiento y/o resistencia a parásitos, con los cuales se pueden establecer linajes que
permitirían incrementar la producción (Graham, 2009).

2
En virtud de lo anterior, el objeto principal de esta investigación se centró en describir el estado genético
de poblaciones silvestres y cultivadas del Jurel de Castilla Seriola lalandi de diversas partes del mundo, y
evaluar el éxito reproductivo de reproductores silvestres mexicanos mantenidos en cautiverio, con base
en el uso de marcadores genéticos moleculares tipo microsatélites. Todo ello con la finalidad de optimizar
el aprovechamiento futuro del acervo genético de este recurso acuático.
1.1 Antecedentes
1.1.1 Biología y distribución de Seriola lalandi
Seriola lalandi (Fig. 1), conocido en México como Jurel de Castilla, es una de las especies de la familia
Carangidae más importantes para la pesca comercial y deportiva, así como para la acuicultura (Gillanders
et al., 1999; Avilés y Castelló, 2004; Rimmer y Ponia, 2007). Generalmente se encuentran en aguas cuyas
temperaturas oscilan entre los 18 y 24°C, se considera una especie pelágica y demersal, capturándose en
profundidades que van de 20 a 70 m (Avilés & Castelló, 2004). La talla máxima registrada es de 2.5 m con
un peso de 97 kg (Ottolenghi et al., 2004). A pesar de que aún no se define su longevidad, Baxter (1960)
encontró peces de 12 años en el océano Pacífico. Su alimentación se basa principalmente en sardina,
anchoveta, macarela, cangrejo rojo y calamar; sin embargo, su dieta está determinada por la prevalencia
del alimento en el área en que se encuentre (John L Baxter, 1960).
Figura 1. Ejemplar de Seriola lalandi. Fuente: Australian National Fish Collection, CSIRO.

3
Los machos alcanzan la madurez sexual al año, mientras que las hembras a los dos o tres años. Las hembras
son capaces de desovar a los tres años de edad, durante su primer evento de desove producen
aproximadamente 450,000 huevos, en comparación con las hembras de mayor edad (7 años) que llegan a
producir cerca de 4,000,000 de huevos en un periodo reproductivo anual en el medio silvestre. Son
organismos que tienen la capacidad de producir desoves múltiples en un periodo reproductivo,
generalmente durante primavera y verano (Baxter, 1960; Crooke, 1983; Ottolenghi et al., 2004; Shiraishi
et al., 2010). En cautiverio se ha reportado una producción anual que va de 1,400,000 a 4,000,000 de
huevos durante 16 a 43 eventos de desoves, en temperaturas entre 16 a 22°C (Stuart y Drawbridge, 2013).
En su hábitat natural, durante el periodo reproductivo pueden formar grandes agrupaciones, de hasta 80
individuos, en lugares alejados de las costas. En México, la Isla Ángel de la Guarda, dentro del Golfo de
California, es considerada un punto de agregación para desovar (Sala et al., 2003). El centro de la península
de Baja California tiene un 96% de abundancia de larvas de S. lalandi, Isla Cedros e Isla Guadalupe son los
lugares de mayor abundancia de S. lalandi durante todo el año. Mientras que en el área comprendida
entre Punta Abreojos y Punta San Juanico, se han encontrado organismos jóvenes (<1.5 años), lo que
sugiere que esta región podría ser importante para el crecimiento (Baxter, 1960; Sumida et al., 1985). En
1932 en EUA, un pescador reportó agrupaciones de centenares de jureles formando pequeños círculos y
desovando en la Isla Santa Cruz, California (Baxter, 1960).
La migración de esta especie ocurre desde Isla Cedros en México hasta el sur de California en Estados
Unidos (EUA) en época de primavera, retornando en invierno a las costas mexicanas; la mayoría de los
jureles migran alrededor de 50 a 80 km, y algunos pueden recorrer una distancia de hasta 3,000 km
(Baxter, 1960; Gillanders et al., 2001). Durante los primeros dos años de vida migran distancias cortas, y a
partir del tercer años de vida forman cardúmenes e inician su comportamiento migratorio en un intervalo
más amplio de su distribución. Sin embargo, al llegar a la edad adulta (ocho años) forman agrupaciones de
peces con hábitos sedentarios (John L Baxter, 1960).
Seriola lalandi es una especie que se distribuye en el Océano Pacífico a lo largo de la costa este de Estados
Unidos hasta Chile, y de Japón a Australia; sin embargo, se ha reportado también en el Atlántico en las
costas de Brasil y en el Océano Índico en las costas de Sudáfrica (FAO, 1998; Kolkovski y Sakakura, 2004;
Ottolenghi et al., 2004; Fig. 2). Por otra parte, estudios recientes demuestran que las poblaciones de S.
lalandi corresponden a tres especies genéticamente distintas que habitan en diferentes áreas geográficas,
por lo cual se propone se retome la clasificación hecha inicialmente por sus descriptores como: Seriola
lalandi (Valenciennes, 1833) en el hemisferio sur, Seriola dorsalis (Gill, 1863) en el Pacífico Noreste y

4
Seriola aureovittata (Temminck y Schlegel, 1845) en el Pacífico Noroeste (Martinez-Takeshita et al., 2015;
Purcell et al., 2015; Swart, 2014).
Desafortunadamente, debido a que aún no se llega a un concenso sobre su clasificación taxonómica, para
los fines del presente trabajo se ha decidido emplear el nombre taxonómicamente válido de Seriola lalandi
(Bailly, 2015), para referirnos a la especie que se distribuye en el Océano Pacífico en ambos hemisferios.
Figura 2. Distribución geográfica de Seriola lalandi. Los puntos indican probabilidades de ocurrencia. Puntos rojos: 80-100%, puntos lilas: 60-79%, puntos verdes: <20%. Fuente: www.aquamaps.org.
1.1.2 Importancia económica y producción de la especie
Por la calidad de su carne, S. lalandi está posicionada como una especie con alto valor comercial en el
mercado internacional. Por ejemplo en Italia, el costo promedio del Jurel es de $400 pesos MXN/kg,
mientras que en Japón el precio al consumidor puede alcanzar los $550 pesos MXN/kg (Nakada, 2002;
Ottolenghi et al., 2004). En México, el panorama es distinto, el valor de S. lalandi proveniente de las
pesquerías no supera los $100 pesos MXN/kg en los supermercados, y el precio de venta promedio a “pie
de playa” (venta que realiza un pescador a un intermediario) es de $11 pesos MXN/kg (SEPESCABC, 2014b).

5
Por otra parte, su rápida tasa de crecimiento, adaptabilidad a condiciones de cultivo, así como la existencia
de un mercado nacional e internacional bien definido, lo posicionan como una especie con alto potencial
acuícola (Symonds et al., 2014).
En Asia, el cultivo de S. lalandi, registrado desde 1960, se ha basado en la captura de organismos silvestres
para su posterior engorda en jaulas marinas o estanques (Nakada, 2002; Ottolenghi et al., 2004). Sin
embargo, Japón en 1980 comenzó las primeras investigaciones para inducir la maduración en cautiverio
en S. lalandi, y en 2004 sus estadísticas reportan la producción de 150 mil toneladas de todas las especies
de Seriola cultivadas con un valor aproximado de $1.334 billones de USD (Nakada, 2008). Por otra parte,
en Corea del Sur se logró el primer evento de desove en cautiverio de esta especie en el año 2014 (Yang
et al., 2016).
En Australia y Nueva Zelanda, el cultivo de jurel inicio en el año 2000 (Kolkovski y Sakakura, 2004) y
estimaciones recientes indican que Australia produce cerca de 4 mil toneladas equivalentes a 20 millones
de AUD (Whatmore et al., 2013). En el caso de Chile, con la finalidad de diversificar la acuicultura, en el
año 2009 se cerró el ciclo de vida de esta especie, y se comenzó la producción comercial de alevines en
sistemas de recirculación acuícola (Orellana et al., 2014). En Holanda, el cultivo de S. lalandi en sistemas
de recirculación acuícola se encuentra en fase experimental, y esta actividad es considerada de alta
prioridad, debido a las presiones ambientales y económicas sobre la industria de cultivo de peces marinos
(Abbink et al., 2012).
En California, Estados Unidos, desde el año 2003 el Instituto de investigación Hubbs-SeaWorld en San
Diego ha mantenido un cultivo experimental de S. lalandi, y a partir del 2007 se han documentado los
desoves de S. lalandi producidos en cautiverio, provenientes de una primera generación de reproductores
silvestres capturados en la Isla Santa Catalina y las costas de San Diego, California (Stuart y Drawbridge,
2013).
En México, en marzo de 2014 ocurrió el primer desove en cautiverio de S. lalandi producido por la empresa
Ocean Baja Labs S. de R.L de C.V, dando así inicio a sus actividades enfocadas en la producción de alevines
de jurel para su posterior engorda en jaulas marinas en Bahía Magdalena, BCS (Scarratt, 2014; Luening,
2014). Se estima que la producción de este laboratorio será de 2,000,000 de larvas anuales (Luening,
2014).

6
En el 2012, de acuerdo con la Secretaría de Agricultura, Ganadería, Desarrollo Rural, Pesca y Alimentación
(SAGARPA) , se tenían registradas cuatro concesiones de acuicultura comercial en Baja California Sur y una
concesión en Baja California; además, de tres permisos para la acuicultura de fomento en los estados de
Sonora, Baja California y Baja California Sur, respectivamente. No obstante, según la Carta Estatal Acuícola
de Baja California (SEPESCABC, 2014a) en el estado se tienen dos concesiones de acuicultura comercial en
Bahía de Todos Santos y un permiso para la acuicultura de fomento en Bahía Salsipuedes. En el 2011, la
producción anual por acuicultura reportada en Baja California fue de 4.5 ton, mientras que en Baja
California Sur la producción alcanza las 70 ton anuales (Roberto Flores, Baja Seas, 2016. Com. Pers.). En
términos pesqueros, en Baja California el jurel es considerado una especie objetivo en su nivel máximo
sustentable, sin embargo, a nivel nacional se considera una especie de acompañamiento.
Desafortunadamente, los volúmenes de producción de S. lalandi en términos pesqueros, están mezclados
con los volúmenes de producción de otras especies de la misma familia Carangidae; por lo que no sé puede
determinar con precisión la aportación de esta especie respecto al total registrado oficialmente
(CONAPESCA, 2013; SEPESCABC, 2014b).
1.1.3 Programas de selección genética: domesticación
La inversión en los programas de selección genética debería ser prioritaria, ya que del total de la
producción acuícola mundial, cerca del 90% proviene de empresas que utilizan reproductores silvestres
que no han sido seleccionados para la mejora de un rasgo importante, por lo que, al no promover
incrementos sustanciales en las características fenotípicas de importancia económica, el potencial
productivo de las especies no se aprovecha, en el corto y largo plazo (Gjedrem, 2012; Gjedrem y Kolstad,
2012). Lo anterior, representa un enorme potencial de crecimiento económico no explotado en su
totalidad, en el sector acuícola (Gjedrem y Kolstad, 2012).
Gracias a los programas de selección genética es posible incrementar el crecimiento, la supervivencia o la
resistencia a enfermedades, de esta forma los beneficios económicos podrían duplicarse; además, la
ganancia genética puede ser sostenible en el tiempo, siempre y cuando, la endogamia se controle y los
individuos seleccionados no se crucen con individuos inferiores (Knibb, 2000). Por esta razón, se han
desarrollado algunos programas de selección genética que han sido muy exitosos para la acuicultura, e.g.,
en trucha, en carpa, en bagre, en salmón y en tilapia. En los peces, el incremento en los valores fenotípicos
7
de interés económico (ganancia genética) es alrededor del 15 % por generación, lo que sugiere por
ejemplo, que en siete generaciones la tasa de crecimiento se duplicaría (Gjedrem y Kolstad, 2012).
El principal objetivo a largo plazo de los programas de selección genética es la domesticación, es decir, el
desarrollo de organismos con una mayor adaptabilidad al cautiverio para mejorar su bienestar, al mismo
tiempo que se mejora la conversión alimenticia y la calidad de sus productos (Price, 1984; Gjedrem, 2005).
Para que el desarrollo de estos programas pueda lograr una respuesta a la selección, es indispensable que
la población cuente con variabilidad genética y fenotípica (Lind et al., 2012; Falconer y Mackay, 1996).
En general, los programas de selección genética constan de cinco fases: 1) el establecimiento de una
población base, 2) el planteamiento del objetivo de producción (qué y cuánto se quiere mejorar), 3) el
registro de las características fenotípicas, 4) la selección y el cruzamiento, para concluir con la quinta fase
de diseminación y evaluación del progreso genético y fenotípico (Dekkers et al., 2004; Gjedrem y Baranski,
2009). Otro factor de suma importancia a considerar es la captura de la mayor parte de la diversidad
genética disponible en el medio silvestre, y subsecuentemente, administrarla a lo largo de las generaciones
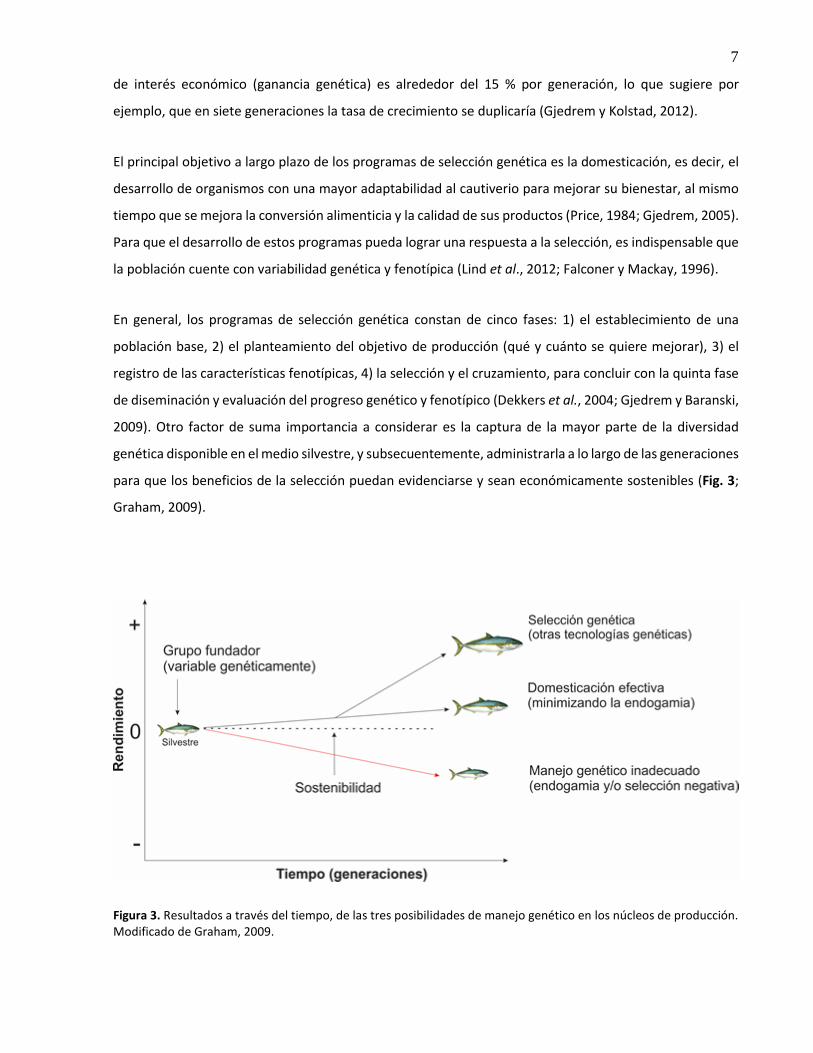
para que los beneficios de la selección puedan evidenciarse y sean económicamente sostenibles (Fig. 3;
Graham, 2009).
Figura 3. Resultados a través del tiempo, de las tres posibilidades de manejo genético en los núcleos de producción. Modificado de Graham, 2009.

8
1.1.4 Marcadores moleculares tipo microsatélites
El genoma de los teleósteos esta constituido por aproximadamente 25,000 genes; e.g., en el pez zebra
Danio rerio se han reportado un total de 25,748 genes (Collins et al., 2012). De este modo, secuenciar,
evaluar e interpretar la diversidad genética de todo el genoma resultaría una tarea demandante y costosa.
Una de las herramientas para evaluar la variabilidad genética son los marcadores moleculares, que son
una representación reducida del genoma del individuo. Existen varios tipos de marcadores genéticos
moleculares, uno de ellos son las secuencias simples repetidas ó SSRs por sus siglas en inglés (Simple
Sequence Repeats), denominados más comúnmente microsatélites, que consisten en repeticiones
consecutivas de secuencias nucleotídicas de entre 2-6 pb, presentes en todas las regiones del genoma de
los cromosomas (Tautz, 1989; Liu y Cordes, 2004). Los microsatélites se heredan bajo siguiendo un modelo
mendeliano, por lo que son codominantes, es decir presentan un alelo materno y otro paterno; y es posible
discriminar los individuos heterocigotos de los homocigotos (Fig.4). Además, son altamente polimórficos,
lo que significa que pueden presentar un gran número de alelos (variantes) evaluado por el tamaño de los
fragmentos; debido a esta variabilidad, es posible obtener genotipos diferenciables entre individuos (Liu
y Cordes, 2004).
Figura 4. Representación simplificada de los microsatélites en los cromosomas de un individuo.

9
A la fecha, existen cerca de 20,000 estudios en la acuicultura que involucran este tipo de marcador
molecular (https://www.ncbi.nlm.nih.gov/genbank/). Los microsatélites, por sus características
intrínsecas, son considerados los marcadores de elección en estudios que evalúan la variabilidad genética
y el comportamiento reproductivo en cautiverio de especies de importancia socio-económica y ecológica
(Aranguren-Méndez et al., 2005; Porta et al., 2006).
1.1.5 Diversidad y estructura genética
A la totalidad de las variaciones en secuencias o tamaños en los fragmentos de DNA (alelos) presentes en
los genomas de una población, se le conoce como diversidad genética; generalmente se cuantifica con
base en el número de alelos (Na), que a su vez se traduce en niveles de heterocigosidad (número de
heterocigotos en la población); una mayor diversidad genética le confiere a los organismos una mayor
capacidad de respuesta a sobrevivir a cambios ambientales o factores estresantes (FAO/UNEP, 1981).
La diversidad genética ha sido uno de los principales objetivos a preservar como parte de la estrategia
mundial de la conservación desde 1980 (IUCN et al., 1980). Actualmente sigue teniendo relevancia debido
a la crisis en la pérdida de la biodiversidad biológica por actividades humanas que enfrenta el planeta
(Savolainen, et al., 2006; Frankham et al., 2002). Conservar la variabilidad genética en las especies
silvestres es fundamental debido a que se asegura el potencial evolutivo que les permitirá adaptarse a
condiciones ambientales cambiantes, mientras que, para las especies y/o poblaciones en cautiverio es
indispensable para lograr el éxito en los programas de crianza selectiva, en donde se busca potenciar la
productividad y viabilidad de los animales, como resultado de la domesticación (Lind et al., 2012; Toro y
Caballero, 2005; Frankham et al., 2002). La importancia de evaluarla y cuantificarla en los programas en
cautiverio, radica en que en las etapas tempranas del proceso es la única oportunidad en la que se puede
capturar esa diversidad. Permitir la entrada de organismos silvestres después de años de selección puede
repercutir en las metas logradas.
Además, el conocimiento del estado genético de las especies puede ser aplicado en la conservación de las
mismas, en el desarrollo de planes de selección y para diferenciar organismos cultivados de los silvestres
(Chauhan & Rajiv, 2010).

10
1.1.6 Éxito reproductivo en cautiverio
En el cultivo de especies marinas el estudio del éxito reproductivo a través de la asignación parental es
imprescindible debido al alto grado de complejidad en los apareamientos de las especies acuáticas, su uso
ofrece la posibilidad de monitorear el coeficiente de endogamia ocasionado por la selección artificial (Liu
& Cordes, 2004).
Por ejemplo, algunos de los problemas que enfrenta la industria acuícola son el resultado de una reducción
en los valores fenotípicos que suele ocasionarse por el apareamiento entre organismos emparentados a
través de uno o varios ancestros comunes (endogamia), esto ocasiona un incremento de homocigotos que
fenotípicamente puede expresarse como una pérdida en la capacidad reproductiva, alteraciones en la tasa
de fecundación y de supervivencia, alteraciones en la tasa de eclosión y sobre la tasa de crecimiento
(Falconer & Mackay, 1996). La probabilidad de que en un cultivo de organismos acuáticos se presente
depresión endogámica, es alta, debido principalmente al desove masivo de los organismos, así como al
reducido tamaño de reproductores efectivos en comparación con el medio silvestres (McDonald et al.,
2004; Yousefian & Nejati, 2008). Por lo que conocer, las relaciones genéticas entre los individuos en
ausencia de genealogía es fundamental para evitar la pérdida de la diversidad genética, mediante la
aplicación de un adecuado manejo reproductivo (Porta et al., 2006).
La asignación parental tuvo un auge a mediados de los noventas gracias a la disponibilidad de marcadores
moleculares tipo microsatélites (Herbinger et al., 1995). Ha sido implementado ya en algunas especies
acuícolas como en salmón del Atlántico (Salmo salar), lenguado japonés (Paralichthys olivaceus), lenguado
senegalés (Solea senegalensis) y bacalao del Atlántico (Gadus morhua; Norris et al., 2000; Sekino et al.,
2004; Porta et al., 2006; Herlin et al., 2008).
1.2 Justificación
En México, la competitividad de la industria acuícola del Jurel de Castilla Seriola lalandi se ve limitada por
la falta de disponibilidad de larvas durante diferentes periodos del año para cubrir así, sus demandas
durante la etapa de engorda. Debido a ello y a pesar de que se ha logrado la reproducción y el desarrollo
larval de la especie en México, las empresas se ven obligadas a importar organismos de diversas partes
del mundo para mantener la producción. Esto se realiza sin evaluar la compatibilidad genética con la de la

11
población local. Esta evaluación puede desarrollarse a partir del análisis de la diversidad genética, que
permitirá vislumbrar el potencial de riesgo, al introducir especies y/o poblaciones diferentes. Mantener la
diversidad genética de las poblaciones cultivadas en una condición similar a las del medio silvestre,
contribuye a disminuir el impacto genético de la incorporación accidental de organismos cultivados a las
poblaciones silvestres, a preservar el potencial evolutivo de las especies, así como a garantizar la respuesta
a la selección en los programas genéticos.
A pesar de la ausencia de información genética en las poblaciones de S. lalandi a lo largo de la Península
de Baja California, estudios recientes confirman que S. lalandi es una agrupación de tres especies. Esto
refleja la necesidad de caracterizar no solo las poblaciones silvestres mexicanas sino también las de otras
regiones del mundo, para evaluar el riesgo de translocación y poder conservar el acervo regional.
Por otra parte, el control de la reproducción en cautiverio es un factor primordial que asegura el éxito en
el cultivo. Sin embargo, dado que los jureles son reproductores asincrónicos con desoves múltiples y
fecundación externa, el aporte de cada parental a la descendencia obtenida en cautiverio se desconocía
hasta hace poco. En el país existen trabajos acerca del efecto de las condiciones de cultivo, tales como
temperatura, manejo larval y alimentación de la especie. Dichos componentes ambientales juegan un
papel fundamental en la expresión del genotipo, determinando el fenotipo de los organismos en cultivo
bajo condiciones particulares. Dado que, teóricamente existirán genotipos mejor adaptados a las
condiciones particulares de cultivo, los efectos del ambiente se verán reflejados en una mayor
supervivencia y crecimiento de la progenie que herede estas características genéticas. Así, la identificación
de los reproductores con mayor éxito reproductivo, es decir aquellos que generen progenies más viables
desde el punto de vista productivo, permitirá el establecimiento de líneas genéticas específicas a fin de
controlar los apareamientos entre los reproductores F1, optimizando los programas de selección genética.
1.3 Hipótesis
La diversidad genética de las poblaciones cultivadas no difiere significativamente a la de sus contrapartes
silvestres, ya que se trata de la primera generación filial.
Debido al amplio rango de distribución de la especie, los individuos silvestres de S. lalandi de las distintas
localidades de México corresponden a una sola población genéticamente homogénea.

12
El grado de aislamiento geográfico entre las poblaciones de otras regiones del mundo de ambos
hemisferios ha producido diferencias genéticas, suficientes para diferenciarlas como especies distintas.
La estrategia reproductiva del Jurel de Castilla, de desoves múltiples por lotes, con un sistema de
apreamiento promiscuo garantiza que todos los reproductores contribuyan genéticamente en igual
proporción a la progenie.
1.4 Objetivos
1.4.1 Objetivo general
Evaluar la diversidad genética y el éxito reproductivo del Jurel de Castilla (Seriola lalandi) en cultivo, a
través de marcadores moleculares tipo microsatélites.
1.4.2 Objetivos específicos
1. Caracterizar el genotipo de S. lalandi en organismos adultos provenientes de seis poblaciones silvestres
y cinco lotes de crías producidos en cautiverio, mediante nueve marcadores moleculares tipo
microsatélites.
2. Estimar los parámetros genéticos para cuantificar la diversidad genética de las poblacionales silvestres
y cultivadas de S. lalandi.
3. Inferir la estructura genética poblacional de S. lalandi de las localidades mexicanas muestreadas.
4. Determinar el grado de diferenciación genética entre las poblaciones del hemisferio norte y hemisferio
sur.
5. Evaluar el éxito reproductivo de reproductores silvestres de S. lalandi mantenidos en cautiverio en
México, a través del aporte parental en la progenie cultivada.
13
Capítulo 2. Metodología
2.1 Origen de las muestras
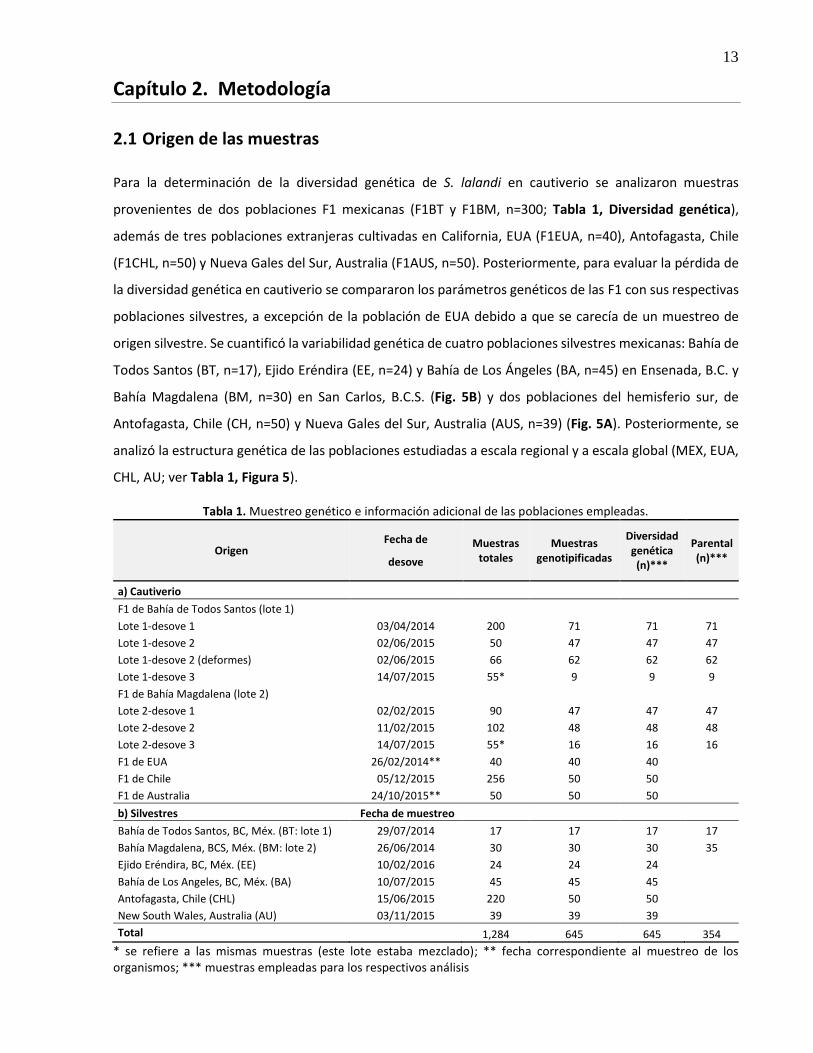
Para la determinación de la diversidad genética de S. lalandi en cautiverio se analizaron muestras
provenientes de dos poblaciones F1 mexicanas (F1BT y F1BM, n=300; Tabla 1, Diversidad genética),
además de tres poblaciones extranjeras cultivadas en California, EUA (F1EUA, n=40), Antofagasta, Chile
(F1CHL, n=50) y Nueva Gales del Sur, Australia (F1AUS, n=50). Posteriormente, para evaluar la pérdida de
la diversidad genética en cautiverio se compararon los parámetros genéticos de las F1 con sus respectivas
poblaciones silvestres, a excepción de la población de EUA debido a que se carecía de un muestreo de
origen silvestre. Se cuantificó la variabilidad genética de cuatro poblaciones silvestres mexicanas: Bahía de
Todos Santos (BT, n=17), Ejido Eréndira (EE, n=24) y Bahía de Los Ángeles (BA, n=45) en Ensenada, B.C. y
Bahía Magdalena (BM, n=30) en San Carlos, B.C.S. (Fig. 5B) y dos poblaciones del hemisferio sur, de
Antofagasta, Chile (CH, n=50) y Nueva Gales del Sur, Australia (AUS, n=39) (Fig. 5A). Posteriormente, se
analizó la estructura genética de las poblaciones estudiadas a escala regional y a escala global (MEX, EUA,
CHL, AU; ver Tabla 1, Figura 5).
Tabla 1. Muestreo genético e información adicional de las poblaciones empleadas.
Origen Fecha de
desove
Muestras totales
Muestras genotipificadas
Diversidad genética (n)***
Parental (n)***
a) Cautiverio
F1 de Bahía de Todos Santos (lote 1)
Lote 1-desove 1 03/04/2014 200 71 71 71
Lote 1-desove 2 02/06/2015 50 47 47 47
Lote 1-desove 2 (deformes) 02/06/2015 66 62 62 62
Lote 1-desove 3 14/07/2015 55* 9 9 9
F1 de Bahía Magdalena (lote 2)
Lote 2-desove 1 02/02/2015 90 47 47 47
Lote 2-desove 2 11/02/2015 102 48 48 48
Lote 2-desove 3 14/07/2015 55* 16 16 16
F1 de EUA 26/02/2014** 40 40 40 F1 de Chile 05/12/2015 256 50 50 F1 de Australia 24/10/2015** 50 50 50 b) Silvestres Fecha de muestreo
Bahía de Todos Santos, BC, Méx. (BT: lote 1) 29/07/2014 17 17 17 17
Bahía Magdalena, BCS, Méx. (BM: lote 2) 26/06/2014 30 30 30 35
Ejido Eréndira, BC, Méx. (EE) 10/02/2016 24 24 24 Bahía de Los Angeles, BC, Méx. (BA) 10/07/2015 45 45 45 Antofagasta, Chile (CHL) 15/06/2015 220 50 50 New South Wales, Australia (AU) 03/11/2015 39 39 39 Total 1,284 645 645 354
* se refiere a las mismas muestras (este lote estaba mezclado); ** fecha correspondiente al muestreo de los organismos; *** muestras empleadas para los respectivos análisis
14
Figura 5.- Localización geográfica de las poblaciones mundiales (A) y mexicanas (B) de S. lalandi, empleadas en el presente estudio.
(B)
(A)

15
Los organismos para el análisis parental (Tabla 1, Parental) fueron proporcionados por la empresa Ocean
Baja Labs S. de R.L. de C.V., dedicada a la producción y engorda de Jurel (Seriola lalandi) que está ubicada
en el Ejido Eréndira en Ensenada, Baja California, México. Se realizó el muestreo genético de dos lotes de
reproductores de origen mexicano, y se muestrearon los juveniles producidos en tres desoves de cada lote
de reproductores para realizar el análisis parental y los estudios de diversidad genética. Asimismo, dentro
del desove número dos producido por reproductores provenientes de Bahía de Todos Santos, se muestreó
un subgrupo de organismos deformes (n=62) con la finalidad de cuantificar la variabilidad genética y
compararla con un subgrupo considerado normal (sin deformidades aparentes) del desove
correspondiente (Fig. 6).
Figura 6 Representación del muestreo a los 30 días después de la eclosión en Seriola Sp.
2.2 Muestreo de reproductores
Para evitar el estrés en los reproductores durante el muestreo, los peces fueron anestesiados siguiendo
los protocolos establecidos por la empresa. Se extrajeron de 30 a 50 mg de la aleta caudal o dorsal con
pinzas y tijeras estériles, las muestras se etiquetaron con base al sistema de marcaje de los reproductores
definido por la empresa y fueron conservadas en tubos Eppendorf de 1.7 ml con etanol al 96% y
almacenadas a -80°C hasta su análisis. Posterior al muestreo, los peces recibieron un tratamiento
profiláctico para evitar la infección de la herida, y una vez recuperados fueron transferidos a un estanque
de mantenimiento.

16
Se colectaron 17 muestras de reproductores (nueve machos y ocho hembras) de Bahía de Todos Santos
que conformaron el lote uno (BT). El lote dos estuvo conformado por 35 reproductores (14 machos, 14
hembras y siete de sexo indefinido); 26 originarios de Bahía Magdalena (BM) y nueve individuos F1
provenientes de Hubbs-Sea World Research Institute, California, EUA (Hubbs) (Anexo 1, Tablas 1 y 2
relación de reproductores).
2.3 Muestreo de juveniles
Debido a que no es posible extraer suficiente DNA de buena calidad a partir de ovocitos recién fecundados
de jurel (Comm. Pers. Estrella Cordova), se tomaron muestras de juveniles de entre 30 y 40 días de edad.
Para su conservación, las larvas fueron mantenidas en un tubo Eppendorf de 1.7 ml con etanol al 96% y
almacenadas a -80°C hasta su análisis. Las muestras se etiquetaron de acuerdo al registro de la hoja de
muestreo para monitoreo de parámetros morfométricos que realiza la empresa como control; en donde
se registran la longitud total, peso, altura, ancho, la presencia o ausencia de deformidades y otros datos
que se consideren relevantes. (lote 1 y lote 2),
Se obtuvieron 300 larvas F1 en cautiverio producidas por ambos grupos reproductores (lote 1 y 2) en los
tres eventos de desove, con las cuales se determinó el aporte parental de S. lalandi en cautiverio (Tabla1).
2.4 Extracción de DNA genómico
Para la extracción de DNA genómico se utilizaron dos protocolos: un kit comercial y un método tradicional.
El kit DNeasy Blood & Tissue de QIAGEN® (Valencia, Ca. USA) se empleó para el aislamiento y purificación
del DNA siguiendo las instrucciones del fabricante . Respecto al método tradicional se usó una
modificación del protocolo de sales publicado por Aljanabi y Martinez, (1997), consistente en: lisis de la
membrana celular y nuclear de las muestras con 5 µl de proteinasa K y 500 µl del Buffer A (Tris-HCl 100
mM; pH 7.5) e incubación a 65°C por 10 horas para facilitar la ruptura de lípidos de la membrana celular;
adición de 570 µl de LiCl 6 M y 230 µl de KAc 5 M y centrifugación a 10,000 rpm por 10 min; posterior a
este proceso, se agregó 200 µl de Fenol-Cloroformo a las muestras para aislar el DNA de proteínas, lípidos
y RNA. Finalmente, el DNA se precipitó con 550 µl de Isopropanol frío (-20°C), el precipitado se lavó con

17
500 µl de Etanol frío al 70% (-20°C); el DNA fue re-suspendido en 50 µl de agua para PCR y se incubó con
RNAsas a 37°C durante 40 minutos.
Para verificar la calidad del DNA extraído, en términos de la integridad (grado de fragmentación) de las
moléculas de DNA, se realizó una electroforesis en gel de agarosa al 1% por 60 min a 80 volts con buffer
TBE 1X, usando como referencia un marcador de peso molecular de 1 Kb (New England BioLabs ®, Reino
Unido). A continuación, el gel se tiñó por inmersión en bromuro de etidio por 15 min y se visualizó en un
sistema digital de fotodocumentación Gel Doc™ XR+ (BIO-RAD®, Estados Unidos) bajo luz UV, las imágenes
se capturaron mediante el programa Quantity One® (BIO-RAD®, Estados Unidos). La concentración y
pureza (contenido proteínas y solventes) se evaluó con la ayuda de un espectrofotómetro ND1000
(NanoDrop Technologies ®, Estados Unidos). La concentración de DNA se midió a 260 nm, a escala de 50
nm y en unidades de ng/μL; además se registraron las razones 260/280 nm que refleja la relación
DNA/proteínas y 260/230 que refleja la relación DNA/solventes, el criterio para considerar un DNA de
buena calidad fue de 1.6-2.1 (Wasko et al., 2003). El DNA obtenido se diluyó a 50 ng/µl con agua para PCR
y fue mantenido a -40°C hasta su posterior análisis.
2.5 Amplificación de los marcadores genéticos tipo microsatélites
La amplificación de los microsatélites, así como su genotipado, se realizó en el GeneCology Research
Centre de la University of the Sunshine Coast, en Queensland, Australia. Se amplificaron nueve marcadores
moleculares tipo microsatélites (Tabla 2) mediante la técnica de reacción en cadena de la polimerasa
(PCR), de los cuales, seis marcadores son especie-específicos Sel001, Sel002, Sel008, Sel011, Sel017 y
Sel019 desarrollados a través de un análisis transcriptómico de tejido de hígado y sistema digestivo
(Whatmore et al., 2013) y tres marcadores heterólogos, desarrollados para la especie Seriola dumerili:
Sdu21 (Renshaw et al., 2006) y Sdu32, Sdu46 (Renshaw et al., 2007).
18
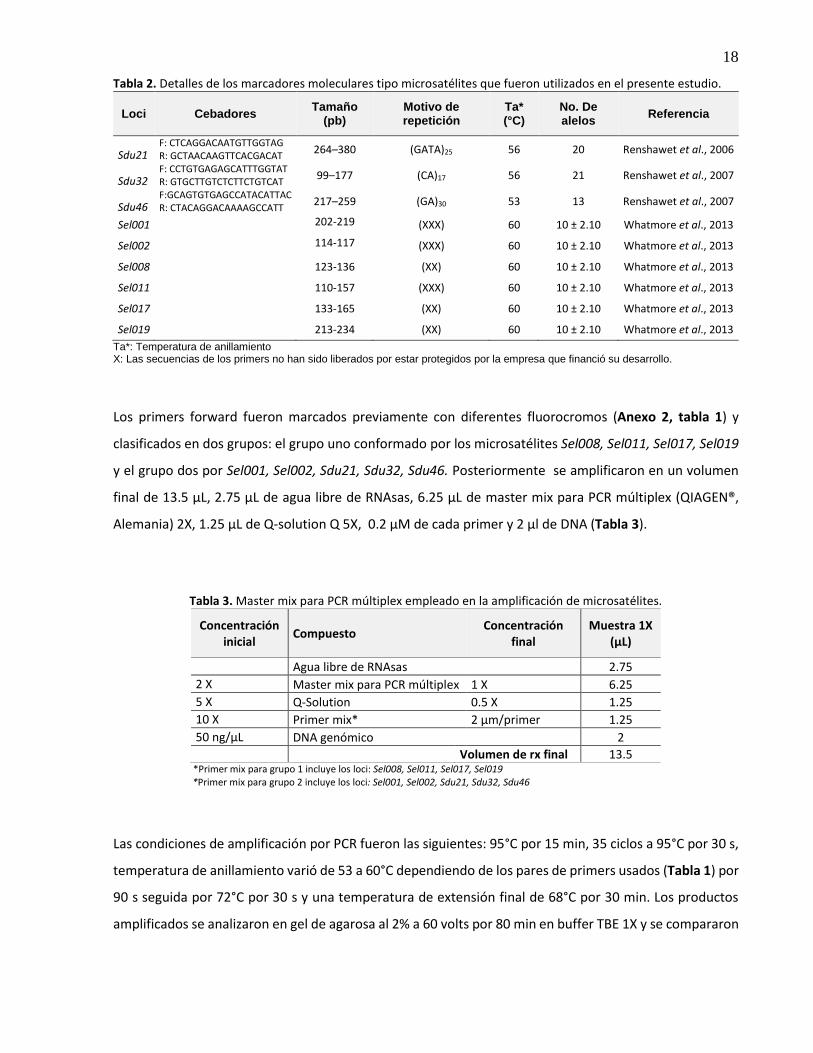
Tabla 2. Detalles de los marcadores moleculares tipo microsatélites que fueron utilizados en el presente estudio.
Loci Cebadores Tamaño
(pb) Motivo de repetición
Ta* (°C)
No. De alelos
Referencia
Sdu21 F: CTCAGGACAATGTTGGTAG R: GCTAACAAGTTCACGACAT
264–380 (GATA)25 56 20 Renshawet et al., 2006
Sdu32 F: CCTGTGAGAGCATTTGGTAT R: GTGCTTGTCTCTTCTGTCAT
99–177 (CA)17 56 21 Renshawet et al., 2007
Sdu46 F:GCAGTGTGAGCCATACATTAC R: CTACAGGACAAAAGCCATT
217–259 (GA)30 53 13 Renshawet et al., 2007
Sel001 202-219 (XXX) 60 10 ± 2.10 Whatmore et al., 2013
Sel002 114-117 (XXX) 60 10 ± 2.10 Whatmore et al., 2013
Sel008 123-136 (XX) 60 10 ± 2.10 Whatmore et al., 2013
Sel011 110-157 (XXX) 60 10 ± 2.10 Whatmore et al., 2013
Sel017 133-165 (XX) 60 10 ± 2.10 Whatmore et al., 2013
Sel019 213-234 (XX) 60 10 ± 2.10 Whatmore et al., 2013
Ta*: Temperatura de anillamiento X: Las secuencias de los primers no han sido liberados por estar protegidos por la empresa que financió su desarrollo.
Los primers forward fueron marcados previamente con diferentes fluorocromos (Anexo 2, tabla 1) y
clasificados en dos grupos: el grupo uno conformado por los microsatélites Sel008, Sel011, Sel017, Sel019
y el grupo dos por Sel001, Sel002, Sdu21, Sdu32, Sdu46. Posteriormente se amplificaron en un volumen
final de 13.5 μL, 2.75 μL de agua libre de RNAsas, 6.25 μL de master mix para PCR múltiplex (QIAGEN®,
Alemania) 2X, 1.25 μL de Q-solution Q 5X, 0.2 μM de cada primer y 2 μl de DNA (Tabla 3).
Tabla 3. Master mix para PCR múltiplex empleado en la amplificación de microsatélites.
Concentración inicial
Compuesto Concentración
final Muestra 1X
(µL)
Agua libre de RNAsas 2.75
2 X Master mix para PCR múltiplex 1 X 6.25
5 X Q-Solution 0.5 X 1.25
10 X Primer mix* 2 µm/primer 1.25
50 ng/µL DNA genómico 2
Volumen de rx final 13.5 *Primer mix para grupo 1 incluye los loci: Sel008, Sel011, Sel017, Sel019 *Primer mix para grupo 2 incluye los loci: Sel001, Sel002, Sdu21, Sdu32, Sdu46
Las condiciones de amplificación por PCR fueron las siguientes: 95°C por 15 min, 35 ciclos a 95°C por 30 s,
temperatura de anillamiento varió de 53 a 60°C dependiendo de los pares de primers usados (Tabla 1) por
90 s seguida por 72°C por 30 s y una temperatura de extensión final de 68°C por 30 min. Los productos
amplificados se analizaron en gel de agarosa al 2% a 60 volts por 80 min en buffer TBE 1X y se compararon

19
con un marcador de peso molecular de 100 pb (New England BioLabs®) para verificar su concentración y
tamaño.
2.6 Genotipado
El genotipado se realizó mediante análisis de fragmentos por electroforesis capilar de alta resolución en
un secuenciador automatizado de la marca Applied Biosystems 3500 (Life Technologies). A partir del
programa GeneMarker® (SoftGenetics, USA) se generó un gel virtual con el que posteriormente se
obtuvieron las matrices de datos de genotipado en formato Excel (*.xls). En la figura 7 se observan los
geles virtuales obtenidos para el grupo uno de microsatélites (Sel008, Sel011, Sel017, Sel019;Fig 7A) y el
grupo dos (Sel001, Sel002, Sdu21, Sdu32, Sdu46; Fig 7B).
Figura 7.- Gel virtual generado para el grupo uno (A) y el grupo dos (B).
2.7 Análisis de diversidad genética
Los datos del genotipado se ajustaron a los formatos necesarios utilizando el programa CREATE 1.35
(Coombs et al., 2008) y revisados con respecto a posibles errores de asignación de alelos con el software
MICROCHECKER v2.2.3 (Oosterhout et al., 2004). La frecuencia de alelos nulos se estimó mediante el
método de máxima verosimilitud en el programa ML-nullFreq (Kalinowski y Taper, 2006).
Los parámetros de diversidad genética tales como número de alelos por locus (Na), alelos privados (Np),
heterocigosidad esperada y observada (He, Ho), además del equilibrio de Hardy-Weinberg (EHW) se

20
calcularon con el programa GenAlex 6.5 (Peakall y Smouse, 2006, 2012). Los coeficientes de endogamia
(Fis), así como la riqueza alélica (Ra) fueron calculados con FSTAT 2.9.3 (Goudet, 1995). Se realizó la
corrección de Bonferroni (Rice, 1989) por el efecto de comparaciones múltiples (nueve loci) en los
resultados del coeficiente de endogamia (Fis), y el equilibrio de Hardy-Weinberg (EHW).
Para determinar las diferencias en los parámetros genéticos entre las poblaciones silvestres y la F1, se
llevó a cabo la prueba de Kruskall-Wallis ya que no se cumplieron las condiciones de normalidad y
homocedasticidad de varianzas para la aplicación de un análisis de varianza (ANDEVA). Posteriormente, se
aplicó la prueba de Dunn para determinar los grupos en los que hubo diferencias significativas con un nivel
de confianza del 95%. Los análisis se realizaron con el programa Statistica 10 (Quest Software Inc).
Para el análisis de variabilidad genética entre los subgrupos normal y deforme, se hizo la prueba U de
Mann Whitney en el programa Statistica 10, ya que los datos no cumplían con el principio de normalidad
y/o homocedasticidad de varianzas.
2.8 Análisis de estructura genética
Para determinar la estructura genética de las poblaciones estudiadas, se empleó el programa STRUCTURE
2.3.4 (Pritchard et al., 2000) que emplea un modelo bayesiano que agrupa a los individuos de acuerdo a
sus frecuencias alélicas. Posteriormente, se implementó la prueba de Evanno (Evanno, et al., 2005), en el
programa STRUCTURE HARVESTER (Earl y vonHoldt, 2012) para determinar delta K (ΔK) que se refiere al
número más probable de componentes genéticos (ΔK). Se realizó un análisis discriminante de
componentes principales en el programa estadístico R (Jombart, 2008; Jombart et al., 2010), cuyo
propósito es disminuir las variables (loci), identificando aquellos que tienen un efecto mayor en la
estructura poblacional, de esta forma las diferencias entre poblaciones pueden ser observadas con
facilidad.
Con las frecuencias alélicas obtenidas con el programa GenAlex 6.5 (Peakall y Smouse, 2006) se infirieron
las distancias genéticas de Nei (Da), a partir de las cuales, se construyó un dendrograma de similitud
genética usando el programa Poptree2 (Takezaki et al., 2010) mediante el método del vecino más cercano
(Neighboor-joining). La confiabilidad de las ramas en el dendrograma fue probada estadísticamente
mediante 1000 repeticiones con la prueba de Bootstrapping.

21
La diferenciación genética se cuantificó por pares de poblaciones considerando siete de los nueve loci
debido a la frecuencia de alelos nulos, mediante el índice de fijación tradicional (FST) y el desarrollado para
microsatélites (RST). Subsecuentemente, se realizó un análisis de permutaciones en donde se determinó
si los valores eran significativamente diferentes de cero, considerando el ajuste al p valor crítico de la
corrección de Bonferroni (Rice, 1989). Se empleó un análisis de varianza molecular (AMOVA) para
determinar diferencias significativas entre los grupos de poblaciones estudiadas. Estos análisis se
desarrollaron en el programa Arlequín (Excoffier et al., 2005).
2.9 Análisis de asignación parental
La contribución de los reproductores se determinó mediante la asignación parental inferida por parejas
empleando un método de máxima probabilidad en el programa COLONY 2.0.5.8 (Jones y Wang, 2010).
Por otra parte, se clasificó a los organismos dependiendo de su longitud estándar en tres categorías (chico,
mediano y grande) para determinar la relación entre el fenotipo y la contribución parental. La clasificación
anterior se realizó para cada lote en forma independiente, y después se usó la información de la totalidad
de los reproductores para evaluar la relación fenotipo - contribución parental de manera global.
Adicionalmente, se determinó el coeficiente de coancestría por pares de reproductores para evaluar el
parentesco entre las posibles cruzas, también se reevaluó el coeficiente de endogamia utilizando este
estimador ya que es más preciso para identificar la endogamia cuando los alelos son idénticos por
descendencia. Esta evaluación se desarrolló con el programa COANCESTRY 1.0.1.2 (Wang, 2011).
22
Capítulo 3. Resultados
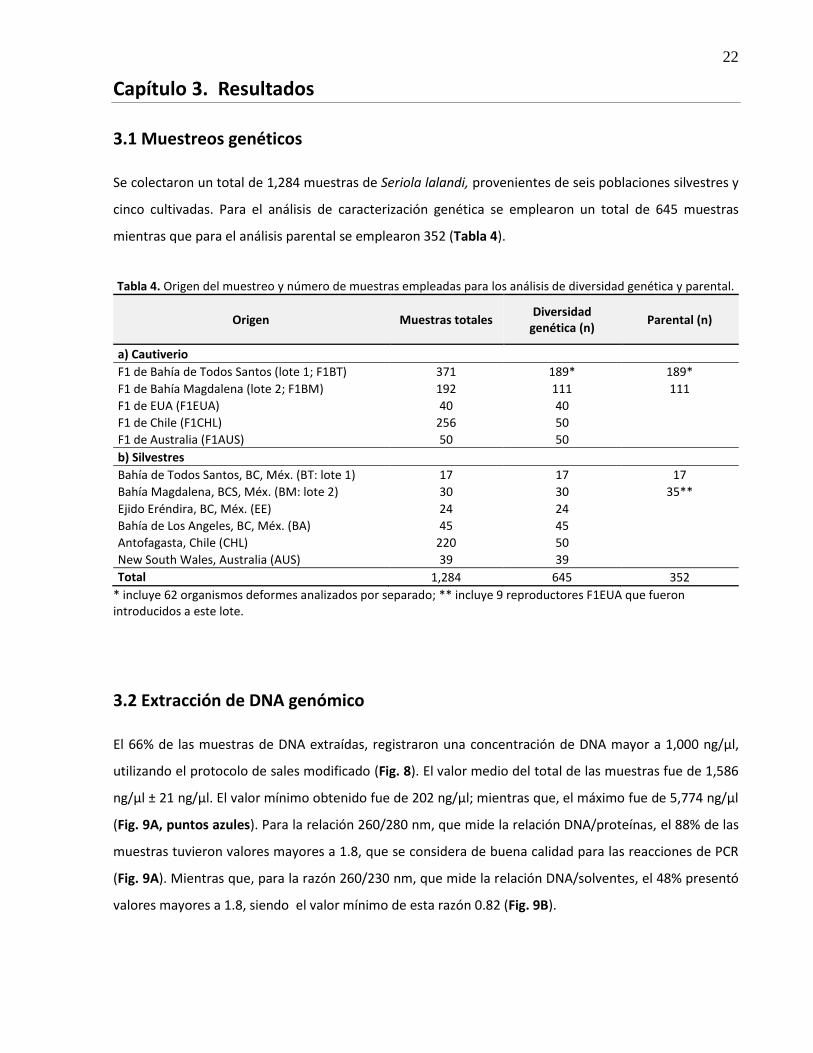
3.1 Muestreos genéticos
Se colectaron un total de 1,284 muestras de Seriola lalandi, provenientes de seis poblaciones silvestres y
cinco cultivadas. Para el análisis de caracterización genética se emplearon un total de 645 muestras
mientras que para el análisis parental se emplearon 352 (Tabla 4).
Tabla 4. Origen del muestreo y número de muestras empleadas para los análisis de diversidad genética y parental.
Origen Muestras totales Diversidad
genética (n) Parental (n)
a) Cautiverio
F1 de Bahía de Todos Santos (lote 1; F1BT) 371 189* 189*
F1 de Bahía Magdalena (lote 2; F1BM) 192 111 111
F1 de EUA (F1EUA) 40 40 F1 de Chile (F1CHL) 256 50 F1 de Australia (F1AUS) 50 50 b) Silvestres
Bahía de Todos Santos, BC, Méx. (BT: lote 1) 17 17 17
Bahía Magdalena, BCS, Méx. (BM: lote 2) 30 30 35**
Ejido Eréndira, BC, Méx. (EE) 24 24 Bahía de Los Angeles, BC, Méx. (BA) 45 45 Antofagasta, Chile (CHL) 220 50 New South Wales, Australia (AUS) 39 39 Total 1,284 645 352
* incluye 62 organismos deformes analizados por separado; ** incluye 9 reproductores F1EUA que fueron introducidos a este lote.
3.2 Extracción de DNA genómico
El 66% de las muestras de DNA extraídas, registraron una concentración de DNA mayor a 1,000 ng/µl,
utilizando el protocolo de sales modificado (Fig. 8). El valor medio del total de las muestras fue de 1,586
ng/µl ± 21 ng/µl. El valor mínimo obtenido fue de 202 ng/µl; mientras que, el máximo fue de 5,774 ng/µl
(Fig. 9A, puntos azules). Para la relación 260/280 nm, que mide la relación DNA/proteínas, el 88% de las
muestras tuvieron valores mayores a 1.8, que se considera de buena calidad para las reacciones de PCR
(Fig. 9A). Mientras que, para la razón 260/230 nm, que mide la relación DNA/solventes, el 48% presentó
valores mayores a 1.8, siendo el valor mínimo de esta razón 0.82 (Fig. 9B).
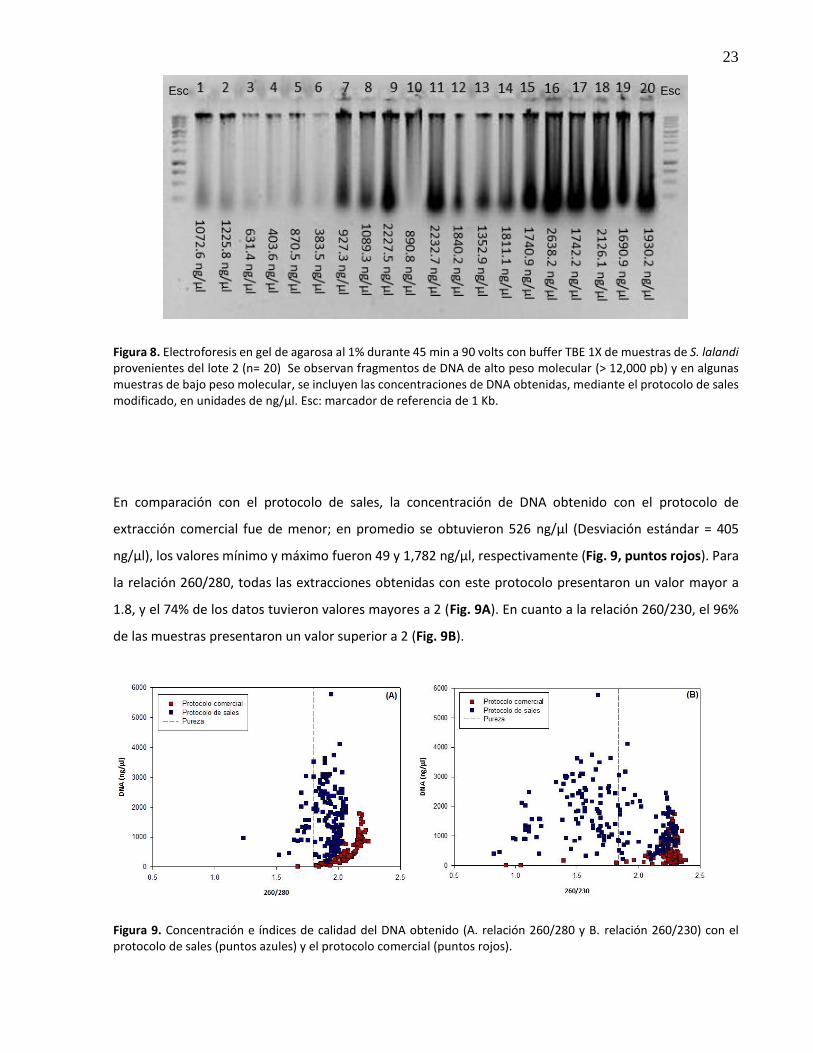
23
Figura 8. Electroforesis en gel de agarosa al 1% durante 45 min a 90 volts con buffer TBE 1X de muestras de S. lalandi provenientes del lote 2 (n= 20) Se observan fragmentos de DNA de alto peso molecular (> 12,000 pb) y en algunas muestras de bajo peso molecular, se incluyen las concentraciones de DNA obtenidas, mediante el protocolo de sales modificado, en unidades de ng/µl. Esc: marcador de referencia de 1 Kb.
En comparación con el protocolo de sales, la concentración de DNA obtenido con el protocolo de
extracción comercial fue de menor; en promedio se obtuvieron 526 ng/µl (Desviación estándar = 405
ng/µl), los valores mínimo y máximo fueron 49 y 1,782 ng/µl, respectivamente (Fig. 9, puntos rojos). Para
la relación 260/280, todas las extracciones obtenidas con este protocolo presentaron un valor mayor a
1.8, y el 74% de los datos tuvieron valores mayores a 2 (Fig. 9A). En cuanto a la relación 260/230, el 96%
de las muestras presentaron un valor superior a 2 (Fig. 9B).
Figura 9. Concentración e índices de calidad del DNA obtenido (A. relación 260/280 y B. relación 260/230) con el protocolo de sales (puntos azules) y el protocolo comercial (puntos rojos).
Esc Esc

24
3.3 Amplificación de los microsatélites
Para todas las muestras seleccionadas, independientemente del protocolo de extracción de DNA
empleado, se lograron amplificar los microsatélites, (Fig. 10).
Figura 10. Ejemplo de la visualización de productos de PCR en gel de agarosa al 2% durante 80 min a 60 volts en
buffer TBE 1X del grupo uno de microsatélites (Sel008, Sel011, Sel017, Sel019) de la población de cultivo de Bahía
Magdalena (BM, n=14). Esc: marcador de referencia de 100 pb, Pos: Control positivo, Neg: Control negativo.
3.4 Genotipado
Los tamaños mínimos y máximos de los productos de PCR obtenidos para cada locus microsatélite
amplificado se resumen en la tabla 5. Con base en el tamaño de los fragmentos fue posible determinar el
genotipo heterocigótico u homocigótico de cada loci para cada individuo (Fig. 11). Para el análisis de
caracterización genética se obtuvieron un total de 645 genotipos, mientras que para el análisis parental
un total de 352 genotipos (Ver tabla 4).
Tabla 5. Tamaño, en número de bases, de los fragmentos de PCR obtenidos para cada locus microsatélite amplificado.
Loci Mínimo Máximo
Sel001 206 236
Sel002 118 140
Sel008 117 177
Sel011 116 161
Sel017 132 265
Sel019 216 246
Sdu21 272 388
Sdu32 87 167
Sdu46 216 259
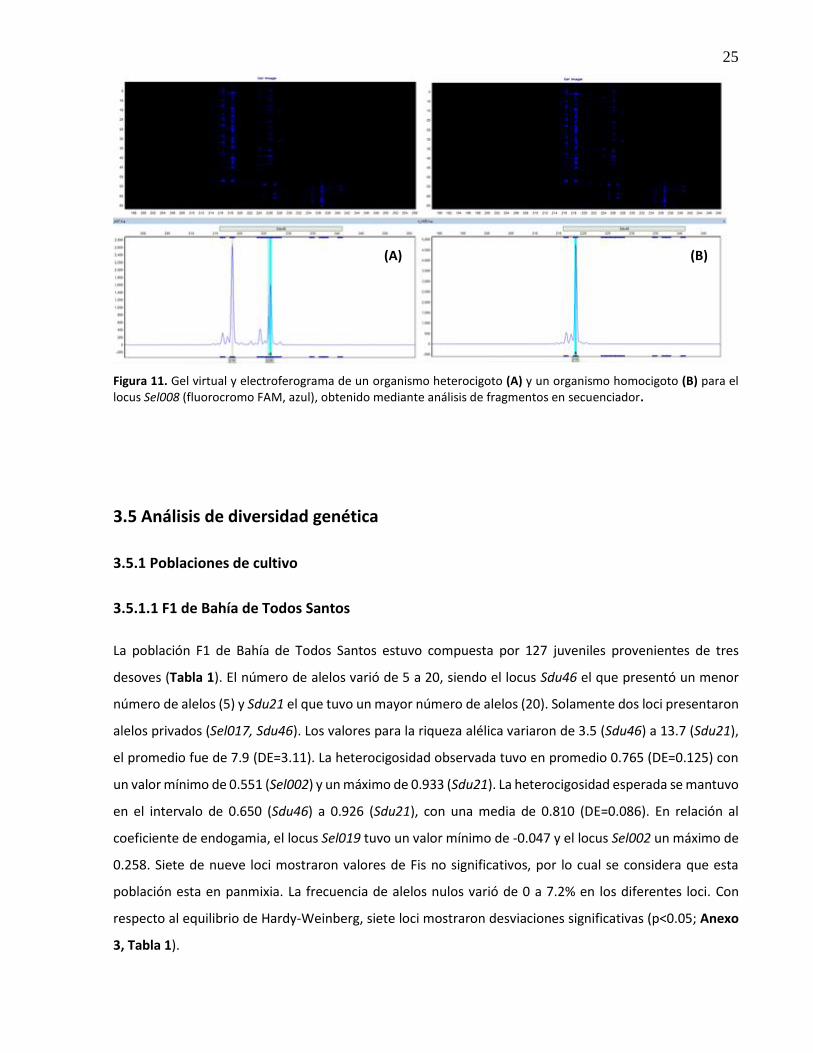
25
Figura 11. Gel virtual y electroferograma de un organismo heterocigoto (A) y un organismo homocigoto (B) para el locus Sel008 (fluorocromo FAM, azul), obtenido mediante análisis de fragmentos en secuenciador.
3.5 Análisis de diversidad genética
3.5.1 Poblaciones de cultivo
3.5.1.1 F1 de Bahía de Todos Santos
La población F1 de Bahía de Todos Santos estuvo compuesta por 127 juveniles provenientes de tres
desoves (Tabla 1). El número de alelos varió de 5 a 20, siendo el locus Sdu46 el que presentó un menor
número de alelos (5) y Sdu21 el que tuvo un mayor número de alelos (20). Solamente dos loci presentaron
alelos privados (Sel017, Sdu46). Los valores para la riqueza alélica variaron de 3.5 (Sdu46) a 13.7 (Sdu21),
el promedio fue de 7.9 (DE=3.11). La heterocigosidad observada tuvo en promedio 0.765 (DE=0.125) con
un valor mínimo de 0.551 (Sel002) y un máximo de 0.933 (Sdu21). La heterocigosidad esperada se mantuvo
en el intervalo de 0.650 (Sdu46) a 0.926 (Sdu21), con una media de 0.810 (DE=0.086). En relación al
coeficiente de endogamia, el locus Sel019 tuvo un valor mínimo de -0.047 y el locus Sel002 un máximo de
0.258. Siete de nueve loci mostraron valores de Fis no significativos, por lo cual se considera que esta
población esta en panmixia. La frecuencia de alelos nulos varió de 0 a 7.2% en los diferentes loci. Con
respecto al equilibrio de Hardy-Weinberg, siete loci mostraron desviaciones significativas (p<0.05; Anexo
3, Tabla 1).
(A) (B)

26
3.5.1.2 F1 de Bahía Magdalena
La F1 de Bahía Magdalena consideró 111 crías correspondientes a tres desoves (Tabla 1). Esta población,
presentó un rango alélico de 7 (Sel002; Sdu46) a 18 (Sel017; Sdu21). Mientras que la riqueza alélica en
promedio fue de 7.36 (DE=2.33) con valores de 3.512 a 10.826. No se observaron alelos privados en ningún
loci. La heterocigosidad observada en promedio fue de 0.771 (DE=0.110) con un intervalo que varió de
0.551 en locus Sdu21 a 0.946 en el locus Sel008. Por otra parte, la heterocigosidad esperada tuvo valores
similares, presentando una media de 0.779 (DE=0.112) con un valor mínimo de 0.514 (Sdu46) y un máximo
de 0.879 (Sdu32). Sólo los loci Sel001 y Sdu21 estuvieron en equilibrio Hardy-Weinberg, el resto presentó
desviaciones significativas (p<0.05). La frecuencia de alelos nulos varió de 0 a 7.9%. El valor del coeficiente
de endogamia osciló de -0.103 (p<0.01) en el locus Sel008 a 0.117 (p<0.05) en el locus Sel019. Siete de
nueve loci mostraron valores de Fis no significativos (p>0.05), por lo cual esta población estuvo en
panmixia (Anexo 3, Tabla 2).
3.5.1.3 F1 de Estados Unidos de América
El número de alelos en la población F1 de Estados Unidos de América (n=40) varió de 5 (Sel002) a 18
(Sel008) alelos por locus. Cuatro loci mostraron alelos privados, Sel001 y Sdu46 presentaron uno, mientras
que Sel017 y Sdu21 tuvieron 3 alelos. La riqueza alélica en promedio fue de 7.296 (DE=2.473) con valores
de 4.56 (Sel002) a 11.87 (Sel017). La heterocigosidad observada promedio fue de 0.849 (DE=0.101), el
locus Sdu46 presentó el valor mínimo (0.675) mientras que el Sel017 tuvo el valor máximo (0.949). La
heterocigosidad esperada para la población fue de 0.760 (DE=0.118) cuyo mínimo y máximo fueron 0.577
(Sel001) y 0.884 (Sel017), respectivamente. Esta población presentó un coeficiente de endogamia que
varió de 0.009 a -0.289 (p<0.001), con un valor medio de -0.114 (DE=0.084). Aunque solo el locus Sel001
presentó un valor de Fis significativamente diferente de cero (-0.289), debido a que la mayoría de los loci
no presentó valores significativos de Fis, se considera esta población en panmixia; además, en esta
población no hubo evidencia de la presencia de alelos nulos, y todos los loci evaluados se ajustaron al
equilibrio de Hardy-Weinberg (Anexo 3, Tabla 3).

27
3.5.1.4 F1 de Chile
La población de cultivo de Chile (n=50) comprendió un número alélico de 4 en el locus a Sel002 a 12 en los
loci Sdu21 y Sdu32. Con respecto a los alelos privados, solo se observaron en dos loci (Sel008 y Sel019). La
riqueza alélica poblacional fue de 5.358 (DE=2.413) oscilando de 3.2 en el locus Sel002 a 10.3 en el locus
Sdu21. El promedio de la heterocigosidad observada fue de 0.706 (DE=0.162) mientras que para la
heterocigosidad esperada fue de 0.652 (DE=0.174). De acuerdo con el equilibrio de Hardy-Weinberg, sólo
el locus Sdu32 mostró una desviación significativa (p<0.05). En cuanto al coeficiente de endogamia, este
varió de 0.027 (Sel001) a -0.231 (Sel017) y sólo el valor de Fis obtenido para el locus Sel017 fue significativo
(-0.231) con tendencia exogámica. El resto de los loci no presentaron valores de Fis significativos por lo
cual se considera a esta población en panmixia. La frecuencia de alelos nulos fue menor al 1.5% (Anexo 3,
Tabla 4).
3.5.1.5 F1 de Australia
La F1 proviente de Australia (n=50), mostró un valor mínimo de 2 alelos en el locus Sel002 y un máximo de
11 alelos en el locus Sdu21. En esta población no se observaron alelos privados en ningún loci. La riqueza
alélica tuvo un promedio de 4.5 (DE=2.241), variando de 2 a 8.9. La heterocigosidad observada registró un
promedio de 0.758 (DE=0.113) con un valor mínimo de 0.531 (Sel002) y máximo de 0.955 (Sdu32), mientras
que, la heterocigosidad esperada fue de 0.667 (DE=0.128), presentando un rango de 0.425 (Sel002) a 0.850
(Sdu21). Relativo al modelo de Hardy-Weinberg tres loci (Sel008, Sdu21 y Sdu32) estuvieron en
desequilibrio (p<0.05). En general la población tuvo una tendencia a la exogamia, cuyo valor promedio fue
de -0.140 (DE=0.135; P<0.001). Tres loci (Sel008, Sdu32 y Sdu46) presentaron valores negativos
significativos (p<0.05), que reflejan cierto grado de exogamia. Para el locus Sdu21 se estimó un 6% en la
frecuencia de alelos nulos (Anexo 3, Tabla 5).

28
3.5.2 Poblaciones silvestres
3.5.2.1 Bahía de Todos Santos, BC, México
En la tabla 6 del anexo 3, se resumen los parámetros genéticos obtenidos del análisis de la población de
Bahía de Todos Santos (n=17). En este grupo, el número de alelos varió de 4 a 20 por locus, el locus con
mayor número de alelos fue el Sel017 (20) y el de menor número fue el locus Sdu46 (4). En cuanto al
número de alelos privados por locus, solo el locus Sel002 presentó uno. La riqueza alélica fue de un valor
mínimo de 3.8 a un máximo de 18 alelos, en los loci Sdu46 y Sdu21, respectivamente. La heterocigosidad
observada fue de 0.118 a 0.941, con un promedio de 0.737 (DE=0.289); la heterocigosidad esperada varió
de 0.509 a 0.934, con un valor medio de 0.799 (DE=0.136). Con respecto al Equilibrio de Hardy-Weinberg
se observó sólo el locus Sdu46 en desequilibrio altamente significativo (p<0.001), asociado con un evidente
déficit de heterocigotos asociado posiblemente a la alta frecuencia (32.8%) de alelos nulos en este locus.
Los valores del coeficiente de endogamia variaron de -0.128 (Sel001) a 0.852 (Sdu46) con un valor
promedio de 0.130 (DE=0.283). Sólo el locus Sdu46 presentó un valor de Fis significativamente diferente
de cero, mostrando alta endogamia.
3.5.2.2 Ejido Eréndira, BC, México
La población silvestre del Ejido Eréndira estuvo compuesta por 24 organismos. El número de alelos se
mantuvo en un rango de 4 (Sel002) a 24 (Sel017). Los loci Sel017, Sdu21 y Sdu46 presentaron alelos
privados. Paralelamente, la riqueza alélica varió de 3.5 (Sel002) a 17.6 (Sel017), promediando 9.87 (DE=4.3)
alelos por locus. La heterocigosidad observada varió de 0.348 para el locus Sdu21 hasta 0.833 en los loci
Sel011 y Sel017, con un promedio de 0.664 (DE=0.179). En cuanto a la heterocigosidad esperada, sus
valores fueron de 0.568 en el locus Sel002 a 0.934 en el locus Sel017, el valor medio fue de 0.802
(DE=0.136). Sólo el locus Sdu21 tuvo desviación altamente significativa del equilibrio de Hardy-Weinberg
(p<0.001). Con relación al coeficiente de endogamia, la población presentó un nivel de endogamia
moderado (en promedio 0.189 con DE=0.175), con un rango de 0.046 a 0.629. Sin embargo, sólo el locus
Sdu21 reflejó un coeficiente de endogamia alto y significativo (0.629; p<0.001), mientras que el resto de
los loci no presentaron valores de Fis significativamente diferentes de cero. La frecuencia de alelos nulos
fue menor al 6.5%, excepto para el locus Sdu21 que fue del 32.7%.; Anexo 3 tabla 7).

29
3.5.2.3 Bahía Magdalena, BCS, México
El número de alelos en la población de Bahía Magdalena (n=30) varió de 5 en el locus Sel002 a 24 en el
locus Sdu21. En cuanto al número de alelos privados los loci: Sel008, Sel011, Sdu21, y Sdu46 presentaron
un alelo privado. Por otra parte, la riqueza alélica observada fue de 4.9 en el locus Sel002 a 24 en el locus
Sdu21. En cuanto a la heterocigosidad observada, la media obtenida fue de 0.765 (DE=0.197) cuyos valores
fueron de 0.267 en Sdu46 a 0.900 en Sel011. En contraste, el promedio de la heterocigosidad esperada fue
de 0.791 (DE=0.192) que varió de 0.324 en el locus Sdu46 a 0.939 en el locus Sdu21. Solamente el locus
Sdu46 presentó una desviación altamente significativa al equilibrio de Hardy-Weinberg (p<0.001). Este
grupo, presentó un coeficiente de endogamia promedio bajo (0.054 y DE=0.094), y puede considerarse en
panmixia dado que la mayoría de los valores de Fis no fueron significativos. Sin embargo, para el locus
Sdu21 el valor de Fis fue significativo y ligeramente endogámico (0.190). Además, cinco loci presentaron
alelos nulos con una frecuencia de 0.6 a 6.4% (Anexo 3 tabla 8).
3.5.2.4 Bahía de los Ángeles, BC, México
Para evaluar la diversidad genética de la población de Bahía de los Ángeles se usó un total de 45 individuos
silvestres. El número de alelos varió de 5 en el locus Sdu002 a 34 en el locus Sel017. Cinco loci presentaron
alelos privados (Anexo 3, Tabla 9), el locus Sel017 presentó nueve alelos privados, Sel019 presentó cinco,
Sel011 mostró dos y los loci Sdu21 y Sdu32 tuvieron solo uno. La riqueza alélica estuvo en un rango de 4.2
(Sdu002) a 17.7 (Sel017). La heterocigosidad observada en promedio fue de 0.733 (DE=0.107) variando de
0.578 en Sdu46 a 0.889 en Sel017. De manera similar, la heterocigosidad esperada varió de 0.544 en Sdu46
a 0.941 en Sel017, en promedio 0.816 (DE=0.134). En cuando al modelo de equilibrio de Hardy-Weinberg,
dos loci (Sdu21 y Sdu32) presentaron desviaciones significativas (p<0.001). En cuanto al coeficiente de
endogamia se encontró que los loci Sdu21 y Sdu32 presentaron valores de Fis significativos y con un grado
moderado de endogamia (0.361 y 0.152, respectivamente). El resto no presentó valores significativos
diferentes de cero; sin embargo, de manera global se observó una ligera tendencia endogámica con un Fis
promedio de 0.102 (DE=0.115). Siete loci tuvieron presencia de alelos nulos, variando de 1.9 (Sel008) a
21.1% (Sdu21).

30
3.5.2.5 Antofagasta, Chile
La población proveniente de Antofagasta, Chile estuvo integrada por 50 individuos. El número mínimo de
alelos fue de 4 en el locus Sel002 mientras que el número máximo fue de 18 en el locus Sdu21. Tres de los
nueve loci presentaron alelos privados, el locus Sel001 presentó dos, mientras que el Sel017 tuvo el valor
máximo de alelos, que fue de cinco (Anexo 3, Tabla 10). La riqueza alélica varió de 3.4 (Sel002) a 12.75
(Sdu21). La heterocigosidad observada fue de 0.689 (DE=0.157) con un valor mínimo de 0.480 en el locus
Sel008 y un valor máximo de 0.891 en el locus Sdu21. En cuanto a la heterocigosidad esperada se obtuvo
un promedio de 0.718 (DE=0.150) con un valor mínimo de 0.470 en el locus Sel008 y un máximo 0.915 en
Sdu21. Se presentaron desviaciones significativas (p<0.05) al equilibrio de Hardy-Weinberg en los loci
Sel001 y Sel002. El coeficiente de endogamia varió de -0.96 a 0.171, en promedio fue de 0.051, y el locus
Sel001 fue el único que presentó un nivel de endogamia moderado de 0.171 (p<0.05). La frecuencia de
alelos nulos varió de 1.4 a 6.9%.
3.5.2.6 Nueva Gales del Sur, Australia
El número de alelos en la población silvestre de Australia (n=39) varió de 4 alelos en el locus Sel002 a 21
en el locus Sdu21. Cinco loci presentaron entre uno y dos alelos privados (Anexo 3, Tabla 11). La riqueza
alélica varió de 3.27 (Sel002) a 14.64 (Sdu21). La heterocigosidad observada fue de 0.716 (DE=0.204) con
un valor mínimo de 0.395 en el locus Sel002 y un máximo de 0.923 en el locus Sdu21, mientras que la
heterocigosidad esperada fue de 0.696 (DE=0.190) con un valor mínimo de 0.351 en el locus Sel008 y un
valor máximo de 0.933 en el locus Sdu21. Todos los loci estuvieron en equilibrio de Hardy-Weinberg. En
general, esta población tuvo una tendencia a la panmixia, sin embargo, el locus Sel002 mostró un grado
de endogamia alto (0.243, p<0.05). Por otra parte, sólo en dos loci se evidenció la presencia de alelos
nulos, variando la frecuencia de 1.9 a 9.9% en Sel017 y Sel002, respectivamente.
3.5.2.7 Análisis estadístico
La prueba de Kruskall-Wallis para el número de alelos mostró diferencias significativas (p<0.01).
Posteriormente, mediante la prueba de Dunn, se identificaron diferencias significativas (p<0.05) entre la
población de cultivo australiana con las poblaciones silvestres provenientes de las localidades de Bahía
Magdalena y de Bahía de los Ángeles (Fig. 12A). Asimismo, la riqueza alélica mostró diferencias
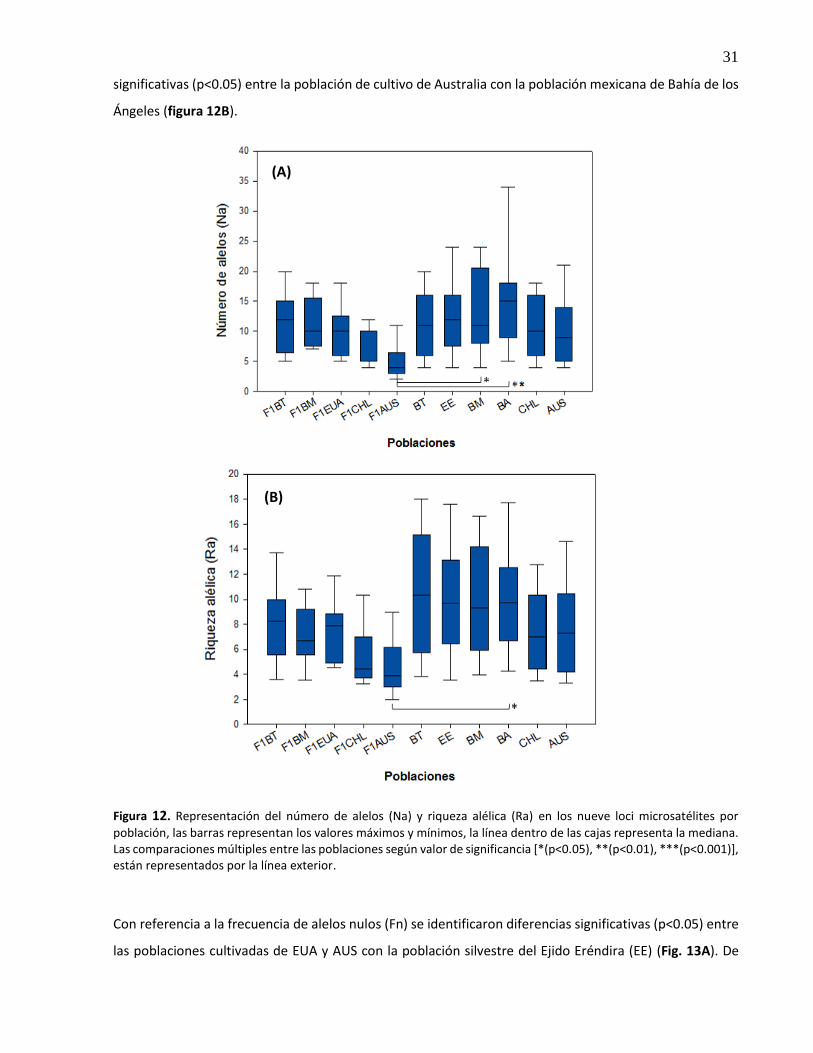
31
significativas (p<0.05) entre la población de cultivo de Australia con la población mexicana de Bahía de los
Ángeles (figura 12B).
Figura 12. Representación del número de alelos (Na) y riqueza alélica (Ra) en los nueve loci microsatélites por
población, las barras representan los valores máximos y mínimos, la línea dentro de las cajas representa la mediana. Las comparaciones múltiples entre las poblaciones según valor de significancia [*(p<0.05), **(p<0.01), ***(p<0.001)], están representados por la línea exterior.
Con referencia a la frecuencia de alelos nulos (Fn) se identificaron diferencias significativas (p<0.05) entre
las poblaciones cultivadas de EUA y AUS con la población silvestre del Ejido Eréndira (EE) (Fig. 13A). De
(A)
(B)
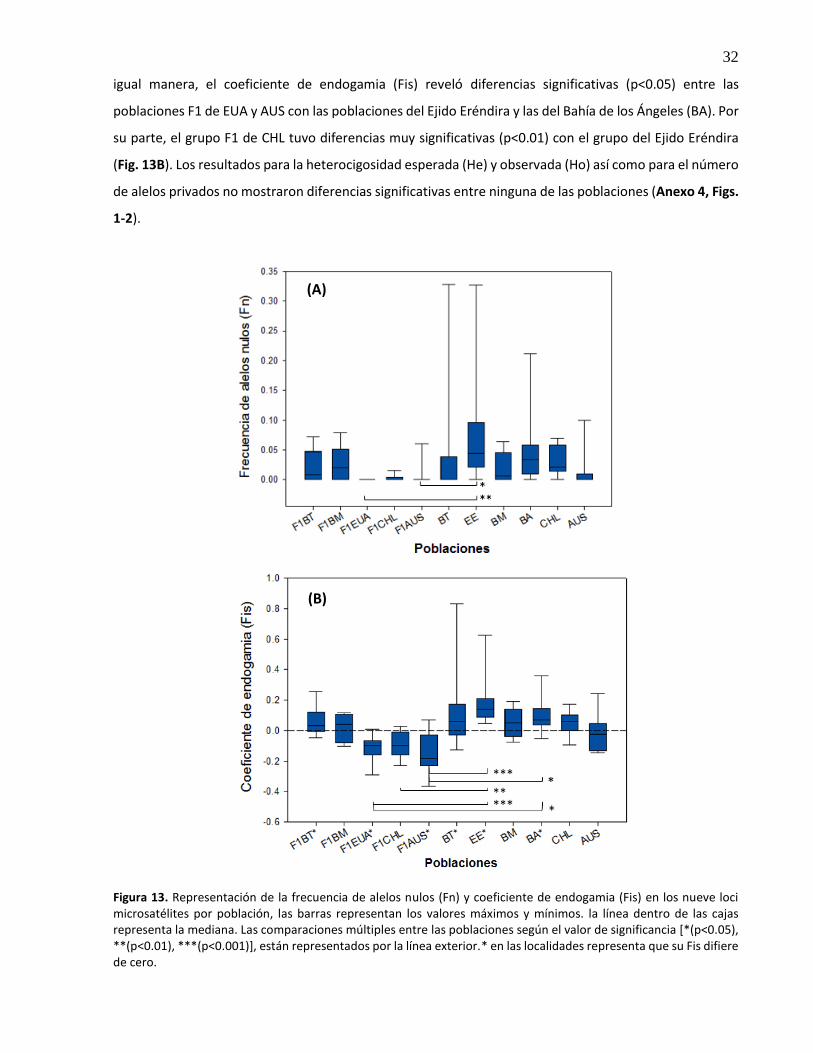
32
igual manera, el coeficiente de endogamia (Fis) reveló diferencias significativas (p<0.05) entre las
poblaciones F1 de EUA y AUS con las poblaciones del Ejido Eréndira y las del Bahía de los Ángeles (BA). Por
su parte, el grupo F1 de CHL tuvo diferencias muy significativas (p<0.01) con el grupo del Ejido Eréndira
(Fig. 13B). Los resultados para la heterocigosidad esperada (He) y observada (Ho) así como para el número
de alelos privados no mostraron diferencias significativas entre ninguna de las poblaciones (Anexo 4, Figs.
1-2).
Figura 13. Representación de la frecuencia de alelos nulos (Fn) y coeficiente de endogamia (Fis) en los nueve loci microsatélites por población, las barras representan los valores máximos y mínimos. la línea dentro de las cajas representa la mediana. Las comparaciones múltiples entre las poblaciones según el valor de significancia [*(p<0.05), **(p<0.01), ***(p<0.001)], están representados por la línea exterior.* en las localidades representa que su Fis difiere de cero.
(A)
(B)

33
3.6 Análisis de estructura genética
3.6.1 Análisis de grupos y asignación de individuos
Se realizó un análisis exploratorio de grupos y asignación de individuos a “n” componentes genéticos (K),
posteriormente se calculó el número más probable de componentes genéticos (DK) en las poblaciones
estudiadas. El primer análisis consideró solamente las muestras de origen mexicano (silvestres y de cultivo,
Fig. 14) mientras que en el segundo se incluyeron muestras de diferente origen geográfico (Fig. 15). Las
gráficas representan individualmente el porcentaje de asignación del genoma a cada uno de los
componentes genéticos K inferidos. Por ejemplo, en el análisis de K=3 (Fig. 14B) el primer individuo de la
población F1BT muestra que el 79% de su genoma corresponde a un componente genético (designado con
color azul), mientras que el 21% restante corresponde a un componente genético distinto (color verde).
Con base en lo anterior, es posible visualizar gráficamente la estructura genética de las poblaciones
considerando la prevalencia de un componente genético determinado y/o hacer inferencias sobre la
migración entre poblaciones silvestres.
En la figura 14 se resumen los resultados obtenidos en las poblaciones mexicanas considerando la
presencia de dos a cuatro componentes genéticos (K). Cuando el número de K fue igual a dos las
poblaciones silvestres del Ejido Eréndira y Bahía de los Ángeles resultaron ser muy parecidas
genéticamente, mientras que, las poblaciones de Bahía de Todos Santos y Bahía Magdalena mostraron un
patrón similar de componentes genéticos (Fig. 14A). Sin embargo, cuando se analizaron las poblaciones
considerando tres componentes genéticos (K=3, Fig. 14B) la población de Bahía de Todos Santos se
diferenció del resto. De manera similar al análisis anterior las poblaciones del Ejido Eréndira y de Bahía de
los Ángeles fueron muy parecidas. En este análisis, las poblaciones en cautiverio correspondieron a su
contraparte silvestre, es decir, los individuos de la F1BT mostraron un componente genético característico
de la población silvestre de Bahía de Todos Santos (verde), de igual manera la F1BM mostró predominancia
del color rojo que es característico de la población silvestre de Bahía Magdalena (Fig. 14B).
Al considerarse cuatro componentes genéticos (Fig. 14C), las poblaciones del Ejido Eréndira y Bahía de los
Ángeles siguieron siendo parecidas. Sin embargo, la población de Bahía de Todos Santos se subdividió,
surgió un nuevo componente (azul) en mayor proporción en esa población, pero presente también en la
población de Bahía Magdalena en donde también hubieron individuos con características genéticas
distintas (verdes), pero que tuvieron un éxito reproductivo mayor en cautiverio (Ver población F1BM, Fig.
14C). En la F1BT surgió una subdivisión, dirigida principalmente por el aporte parental de los reproductores
originarios de BT, quienes al parecer se reprodujeron en proporciones similares. La prueba de Evanno
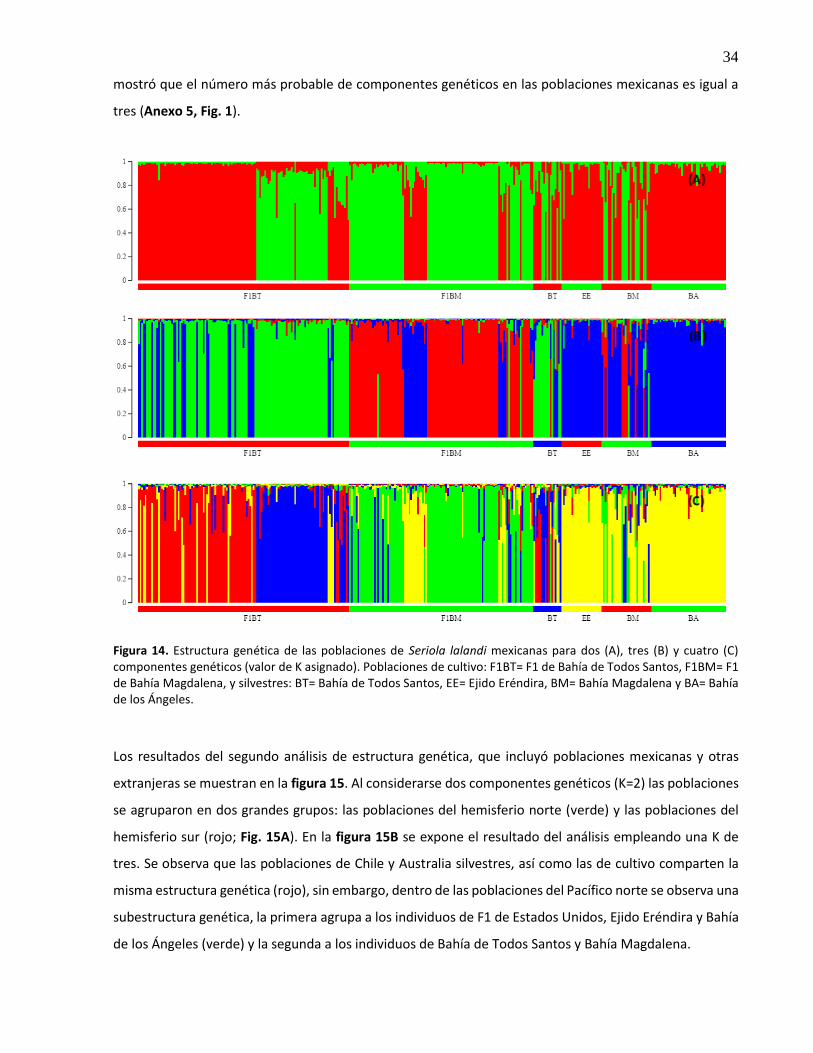
34
mostró que el número más probable de componentes genéticos en las poblaciones mexicanas es igual a
tres (Anexo 5, Fig. 1).
Figura 14. Estructura genética de las poblaciones de Seriola lalandi mexicanas para dos (A), tres (B) y cuatro (C) componentes genéticos (valor de K asignado). Poblaciones de cultivo: F1BT= F1 de Bahía de Todos Santos, F1BM= F1 de Bahía Magdalena, y silvestres: BT= Bahía de Todos Santos, EE= Ejido Eréndira, BM= Bahía Magdalena y BA= Bahía de los Ángeles.
Los resultados del segundo análisis de estructura genética, que incluyó poblaciones mexicanas y otras
extranjeras se muestran en la figura 15. Al considerarse dos componentes genéticos (K=2) las poblaciones
se agruparon en dos grandes grupos: las poblaciones del hemisferio norte (verde) y las poblaciones del
hemisferio sur (rojo; Fig. 15A). En la figura 15B se expone el resultado del análisis empleando una K de
tres. Se observa que las poblaciones de Chile y Australia silvestres, así como las de cultivo comparten la
misma estructura genética (rojo), sin embargo, dentro de las poblaciones del Pacífico norte se observa una
subestructura genética, la primera agrupa a los individuos de F1 de Estados Unidos, Ejido Eréndira y Bahía
de los Ángeles (verde) y la segunda a los individuos de Bahía de Todos Santos y Bahía Magdalena.

35
Finalmente, al emplear una K de cuatro, la población de BT se diferenció del resto de las poblaciones
mexicanas, como en el análisis anterior. Se observó además, que la población de BM estaba mezclada con
individuos de otros grupos y presentaba un componente genético característico (amarillo; Fig. 15C). Por
otro lado, las poblaciones del hemisferio sur permanecieron agrupadas (Figura 15C). La prueba de Evanno
demostró que el número más probable de componentes genéticos en este análisis fue de tres (Anexo 5,
Fig. 2).
Figura 15. Estructura genética de las poblaciones de Seriola lalandi mexicanas y extranjeras para dos (A), tres (B) y cuatro (C) componentes genéticos (valor de K asignado). Poblaciones de cultivo: F1BT= F1 de Bahía de Todos Santos, F1BM= F1 de Bahía Magdalena, F1EUA= F1 de Estados Unidos, F1CHL= F1 de Antofagasta, Chile, F1AUS= F1 de Nueva Gales del Sur, Australia, y silvestres: BT= Bahía de Todos Santos, EE= Ejido Eréndira, BM= Bahía Magdalena, BA= Bahía de los Ángeles, CHL= Chile y AUS= Australia.
(A)

36
3.6.2 Análisis discriminante de componentes principales
En la figura 16 se presenta en forma gráfica el resultado del análisis discriminante de componentes
principales, el eje de las abscisas (x) explica la varianza entre las poblaciones mientras el eje de las
ordenadas (y) explica la varianza dentro de las poblaciones. Los puntos representan a los individuos,
mientras que el ovalo agrupa al 95% de los individuos en la población. Se puede observar, que existe poca
divergencia genética entre las poblaciones del Ejido Eréndira (EE), Bahía Magdalena (BM) y Bahía de los
Ángeles (BA). Sin embargo, el grupo de Bahía de Todos Santos (BT) es el que tiene una divergencia mayor
al resto de las poblaciones silvestres. Por otro lado, las poblaciones de cultivo son las que presentan una
divergencia genética mayor.
Figura 16. Plano cartesiano del análisis discriminante de componentes principales para las poblaciones del noreste del pacífico. Poblaciones de cultivo: F1BT= F1 de Bahía de Todos Santos, F1BM= F1 de Bahía Magdalena, F1EUA= F1 de Estados Unidos; y silvestres: BT= Bahía de Todos Santos, EE= Ejido Eréndira, BM= Bahía Magdalena y BA= Bahía de los Ángeles.
El análisis discriminante de componentes principales para las poblaciones de diversas partes del mundo
(Fig. 17) evidenció una clara estructura genética entre las poblaciones muestreadas en el hemisferio norte
y las del hemisferio sur que coincidió con los resultados obtenidos mediante el programa STRUCTURE
presentados en la sección anterior (Fig. 15). Por otra parte, se puede observar una mayor diversidad

37
genética entre las poblaciones de CHL y AUS, en comparación con las poblaciones mexicanas y de EUA que
muestran ser más parecidas entre ellas.
Figura 17. Plano cartesiano del análisis discriminante de componentes principales para las poblaciones del noroeste del Pacífico (poblaciones mexicanas y de EUA) y del hemisferio sur (Chile y Australia). Poblaciones de cultivo: F1BT= F1 de Bahía de Todos Santos, F1BM= F1 de Bahía Magdalena, F1EUA= F1 de Estados Unidos, F1CHL= F1 de Antofagasta, Chile, F1AUS= F1 de Nueva Gales del Sur, Australia, y silvestres: BT= Bahía de Todos Santos, EE= Ejido Eréndira, BM= Bahía Magdalena, BA= Bahía de los Ángeles, CHL= Chile y AUS= Australia
3.6.3 Similitud genética entre poblaciones
Para la construcción de los dendrogramas se emplearon las distancias genéticas de Nei considerando los
alelos compartidos (Da) y el algoritmo del método del vecino más cercano (en inglés Neighbor-Joining o
NJ). Posteriormente, se realizaron pruebas de proporciones de bootstrapping (10000 veces) para
determinar la confiabilidad de la topología obtenida. En el caso de las poblaciones locales, en el
dendrograma se utilizó una población genéticamente distinta (enraizamiento), con la finalidad de inferir
la topología adecuada.
A nivel regional se observó un clado principal (Fig. 18), conformado por dos clados secundarios el primero
soportado con un 99% de confiabilidad agrupó a los individuos de Bahía de Todos Santos y de Bahía

38
Magdalena, el segundo, reunió a los del Ejido Eréndira y Bahía de los Ángeles con una confiabilidad del
95%. En la figura 18 se puede observar las similitudes genéticas entre las poblaciones silvestres regionales.
Figura 18. Dendrograma inferido por el método del vecino más cercano a través de la Distancia genética de Nei (Da) en las poblaciones mexicanas, anclado a un grupo externo genéticamente diferente (AUS).
En el dendrograma inferido con las poblaciones extranjeras (Fig. 19) se observaron dos clados principales
soportados con el 100% de confiabilidad que concuerdan con la ubicación geográfica de los organismos,
agrupando a los del hemisferio norte en un grupo bien definido y en otro, a los del hemisferio sur. En el
caso de las poblaciones del hemisferio sur, se encontró un ligero grado de diferenciación entre las
poblaciones silvestres y las mantenidas en cautiverio, así como entre las poblaciones de Chile y Australia.
Por otra parte, se puede observar que la composición genética entre EUA, Eréndira y Bahía de los Ángeles
fue similar, a pesar de que el clado que soportó a este grupo tuvo una confiabilidad del 58%.
Bahía de Todos Santos
Bahía Magdalena
Ejido Eréndira
Bahía de los Ángeles
Grupo externo
99
95
0.05
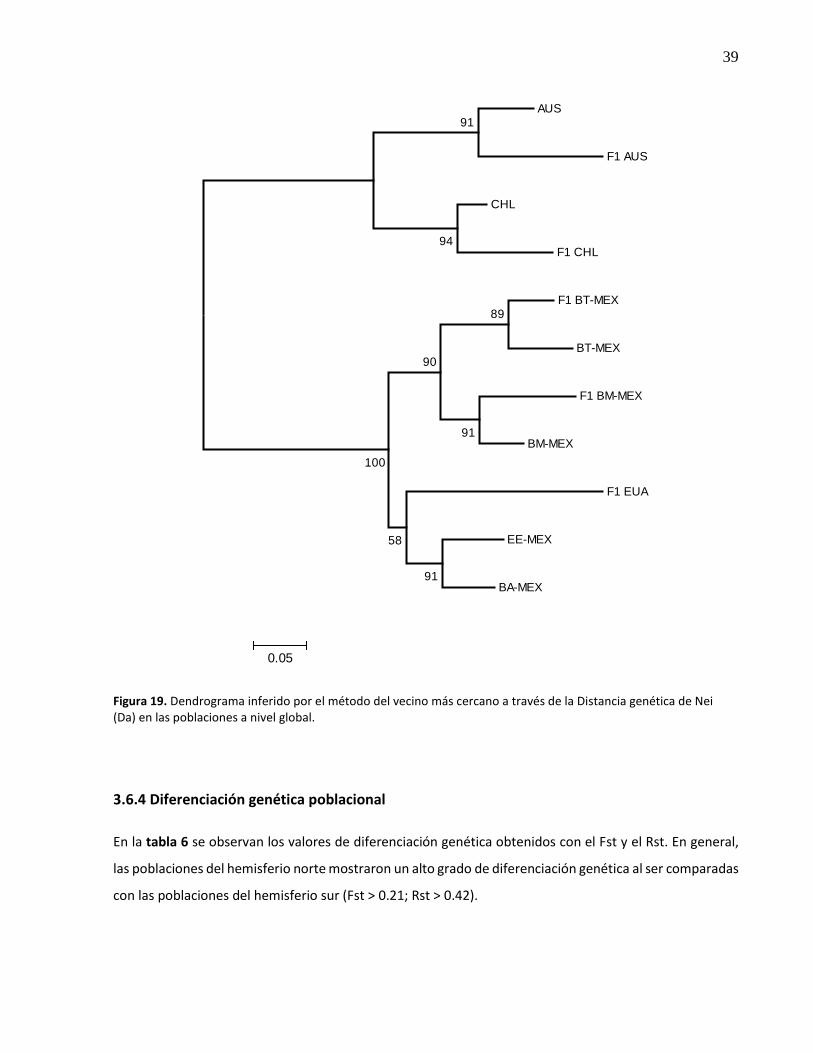
39
Figura 19. Dendrograma inferido por el método del vecino más cercano a través de la Distancia genética de Nei (Da) en las poblaciones a nivel global.
3.6.4 Diferenciación genética poblacional
En la tabla 6 se observan los valores de diferenciación genética obtenidos con el Fst y el Rst. En general,
las poblaciones del hemisferio norte mostraron un alto grado de diferenciación genética al ser comparadas
con las poblaciones del hemisferio sur (Fst > 0.21; Rst > 0.42).
AUS
F1 AUS
CHL
F1 CHL
F1 BT-MEX
BT-MEX
F1 BM-MEX
BM-MEX
F1 EUA
EE-MEX
BA-MEX
91
94
89
91
90
91
58
100
0.05
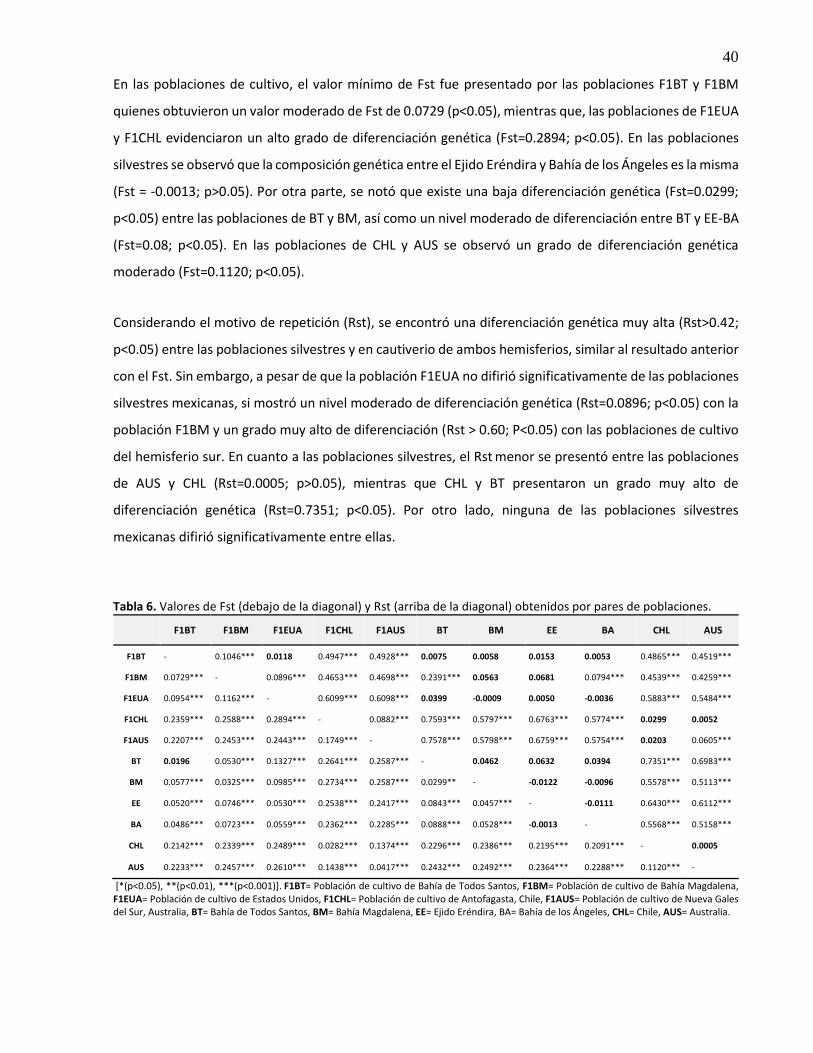
40
En las poblaciones de cultivo, el valor mínimo de Fst fue presentado por las poblaciones F1BT y F1BM
quienes obtuvieron un valor moderado de Fst de 0.0729 (p<0.05), mientras que, las poblaciones de F1EUA
y F1CHL evidenciaron un alto grado de diferenciación genética (Fst=0.2894; p<0.05). En las poblaciones
silvestres se observó que la composición genética entre el Ejido Eréndira y Bahía de los Ángeles es la misma
(Fst = -0.0013; p>0.05). Por otra parte, se notó que existe una baja diferenciación genética (Fst=0.0299;
p<0.05) entre las poblaciones de BT y BM, así como un nivel moderado de diferenciación entre BT y EE-BA
(Fst=0.08; p<0.05). En las poblaciones de CHL y AUS se observó un grado de diferenciación genética
moderado (Fst=0.1120; p<0.05).
Considerando el motivo de repetición (Rst), se encontró una diferenciación genética muy alta (Rst>0.42;
p<0.05) entre las poblaciones silvestres y en cautiverio de ambos hemisferios, similar al resultado anterior
con el Fst. Sin embargo, a pesar de que la población F1EUA no difirió significativamente de las poblaciones
silvestres mexicanas, si mostró un nivel moderado de diferenciación genética (Rst=0.0896; p<0.05) con la
población F1BM y un grado muy alto de diferenciación (Rst > 0.60; P<0.05) con las poblaciones de cultivo
del hemisferio sur. En cuanto a las poblaciones silvestres, el Rst menor se presentó entre las poblaciones
de AUS y CHL (Rst=0.0005; p>0.05), mientras que CHL y BT presentaron un grado muy alto de
diferenciación genética (Rst=0.7351; p<0.05). Por otro lado, ninguna de las poblaciones silvestres
mexicanas difirió significativamente entre ellas.
Tabla 6. Valores de Fst (debajo de la diagonal) y Rst (arriba de la diagonal) obtenidos por pares de poblaciones.
F1BT F1BM F1EUA F1CHL F1AUS BT BM EE BA CHL AUS
F1BT - 0.1046*** 0.0118 0.4947*** 0.4928*** 0.0075 0.0058 0.0153 0.0053 0.4865*** 0.4519***
F1BM 0.0729*** - 0.0896*** 0.4653*** 0.4698*** 0.2391*** 0.0563 0.0681 0.0794*** 0.4539*** 0.4259***
F1EUA 0.0954*** 0.1162*** - 0.6099*** 0.6098*** 0.0399 -0.0009 0.0050 -0.0036 0.5883*** 0.5484***
F1CHL 0.2359*** 0.2588*** 0.2894*** - 0.0882*** 0.7593*** 0.5797*** 0.6763*** 0.5774*** 0.0299 0.0052
F1AUS 0.2207*** 0.2453*** 0.2443*** 0.1749*** - 0.7578*** 0.5798*** 0.6759*** 0.5754*** 0.0203 0.0605***
BT 0.0196 0.0530*** 0.1327*** 0.2641*** 0.2587*** - 0.0462 0.0632 0.0394 0.7351*** 0.6983***
BM 0.0577*** 0.0325*** 0.0985*** 0.2734*** 0.2587*** 0.0299** - -0.0122 -0.0096 0.5578*** 0.5113***
EE 0.0520*** 0.0746*** 0.0530*** 0.2538*** 0.2417*** 0.0843*** 0.0457*** - -0.0111 0.6430*** 0.6112***
BA 0.0486*** 0.0723*** 0.0559*** 0.2362*** 0.2285*** 0.0888*** 0.0528*** -0.0013 - 0.5568*** 0.5158***
CHL 0.2142*** 0.2339*** 0.2489*** 0.0282*** 0.1374*** 0.2296*** 0.2386*** 0.2195*** 0.2091*** - 0.0005
AUS 0.2233*** 0.2457*** 0.2610*** 0.1438*** 0.0417*** 0.2432*** 0.2492*** 0.2364*** 0.2288*** 0.1120*** -
[*(p<0.05), **(p<0.01), ***(p<0.001)]. F1BT= Población de cultivo de Bahía de Todos Santos, F1BM= Población de cultivo de Bahía Magdalena, F1EUA= Población de cultivo de Estados Unidos, F1CHL= Población de cultivo de Antofagasta, Chile, F1AUS= Población de cultivo de Nueva Gales del Sur, Australia, BT= Bahía de Todos Santos, BM= Bahía Magdalena, EE= Ejido Eréndira, BA= Bahía de los Ángeles, CHL= Chile, AUS= Australia.
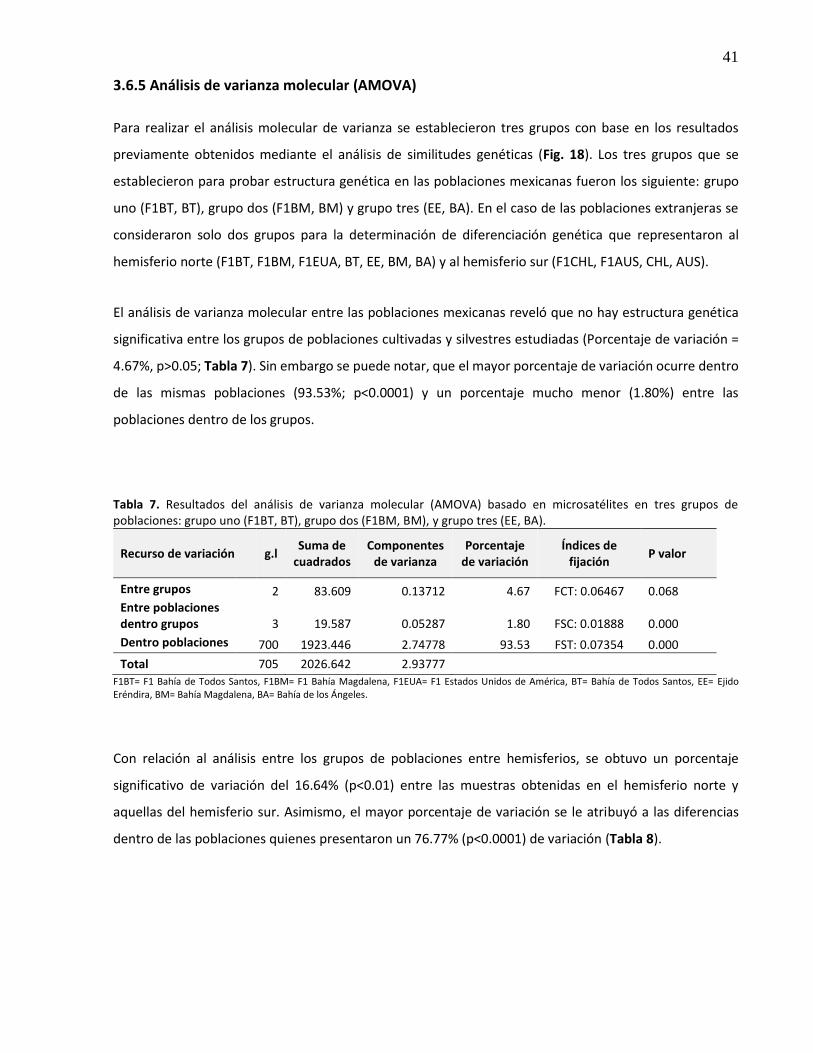
41
3.6.5 Análisis de varianza molecular (AMOVA)
Para realizar el análisis molecular de varianza se establecieron tres grupos con base en los resultados
previamente obtenidos mediante el análisis de similitudes genéticas (Fig. 18). Los tres grupos que se
establecieron para probar estructura genética en las poblaciones mexicanas fueron los siguiente: grupo
uno (F1BT, BT), grupo dos (F1BM, BM) y grupo tres (EE, BA). En el caso de las poblaciones extranjeras se
consideraron solo dos grupos para la determinación de diferenciación genética que representaron al
hemisferio norte (F1BT, F1BM, F1EUA, BT, EE, BM, BA) y al hemisferio sur (F1CHL, F1AUS, CHL, AUS).
El análisis de varianza molecular entre las poblaciones mexicanas reveló que no hay estructura genética
significativa entre los grupos de poblaciones cultivadas y silvestres estudiadas (Porcentaje de variación =
4.67%, p>0.05; Tabla 7). Sin embargo se puede notar, que el mayor porcentaje de variación ocurre dentro
de las mismas poblaciones (93.53%; p<0.0001) y un porcentaje mucho menor (1.80%) entre las
poblaciones dentro de los grupos.
Tabla 7. Resultados del análisis de varianza molecular (AMOVA) basado en microsatélites en tres grupos de poblaciones: grupo uno (F1BT, BT), grupo dos (F1BM, BM), y grupo tres (EE, BA).
Recurso de variación g.l Suma de
cuadrados Componentes
de varianza Porcentaje
de variación Índices de
fijación P valor
Entre grupos 2 83.609 0.13712 4.67 FCT: 0.06467 0.068 Entre poblaciones dentro grupos 3 19.587 0.05287 1.80 FSC: 0.01888 0.000
Dentro poblaciones 700 1923.446 2.74778 93.53 FST: 0.07354 0.000
Total 705 2026.642 2.93777
F1BT= F1 Bahía de Todos Santos, F1BM= F1 Bahía Magdalena, F1EUA= F1 Estados Unidos de América, BT= Bahía de Todos Santos, EE= Ejido Eréndira, BM= Bahía Magdalena, BA= Bahía de los Ángeles.
Con relación al análisis entre los grupos de poblaciones entre hemisferios, se obtuvo un porcentaje
significativo de variación del 16.64% (p<0.01) entre las muestras obtenidas en el hemisferio norte y
aquellas del hemisferio sur. Asimismo, el mayor porcentaje de variación se le atribuyó a las diferencias
dentro de las poblaciones quienes presentaron un 76.77% (p<0.0001) de variación (Tabla 8).
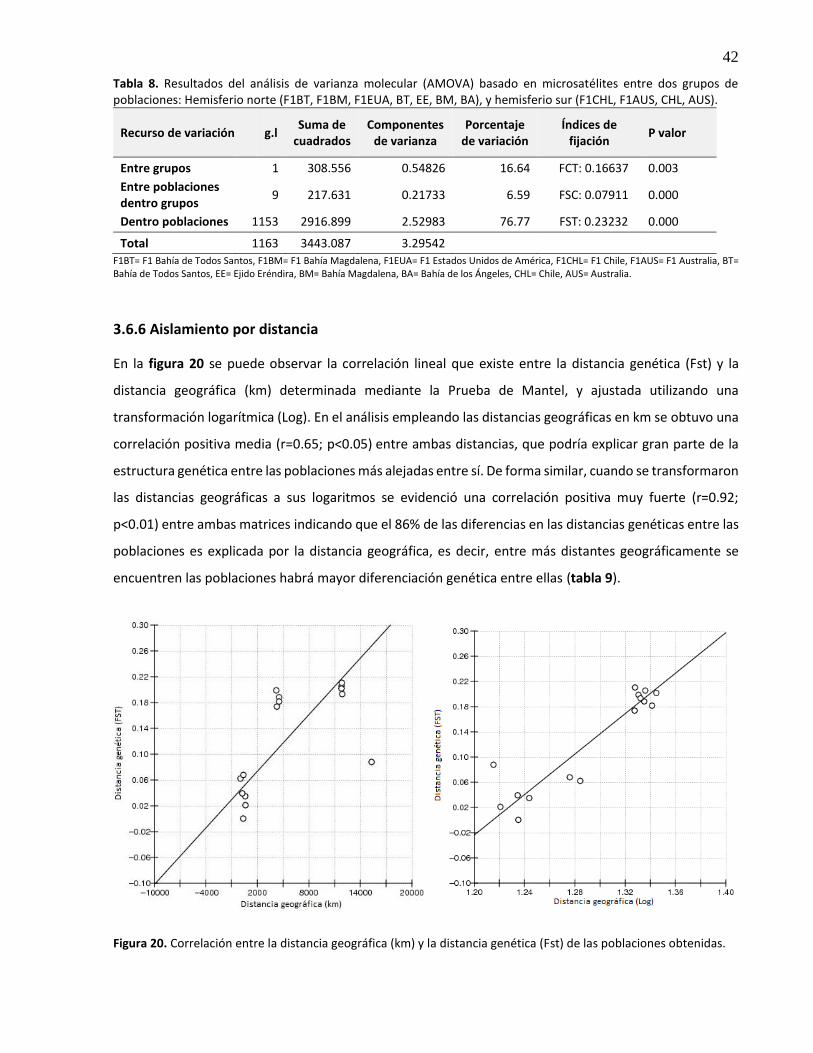
42
Tabla 8. Resultados del análisis de varianza molecular (AMOVA) basado en microsatélites entre dos grupos de poblaciones: Hemisferio norte (F1BT, F1BM, F1EUA, BT, EE, BM, BA), y hemisferio sur (F1CHL, F1AUS, CHL, AUS).
Recurso de variación g.l Suma de
cuadrados Componentes
de varianza Porcentaje
de variación Índices de
fijación P valor
Entre grupos 1 308.556 0.54826 16.64 FCT: 0.16637 0.003
Entre poblaciones dentro grupos
9 217.631 0.21733 6.59 FSC: 0.07911 0.000
Dentro poblaciones 1153 2916.899 2.52983 76.77 FST: 0.23232 0.000
Total 1163 3443.087 3.29542
F1BT= F1 Bahía de Todos Santos, F1BM= F1 Bahía Magdalena, F1EUA= F1 Estados Unidos de América, F1CHL= F1 Chile, F1AUS= F1 Australia, BT= Bahía de Todos Santos, EE= Ejido Eréndira, BM= Bahía Magdalena, BA= Bahía de los Ángeles, CHL= Chile, AUS= Australia.
3.6.6 Aislamiento por distancia
En la figura 20 se puede observar la correlación lineal que existe entre la distancia genética (Fst) y la
distancia geográfica (km) determinada mediante la Prueba de Mantel, y ajustada utilizando una
transformación logarítmica (Log). En el análisis empleando las distancias geográficas en km se obtuvo una
correlación positiva media (r=0.65; p<0.05) entre ambas distancias, que podría explicar gran parte de la
estructura genética entre las poblaciones más alejadas entre sí. De forma similar, cuando se transformaron
las distancias geográficas a sus logaritmos se evidenció una correlación positiva muy fuerte (r=0.92;
p<0.01) entre ambas matrices indicando que el 86% de las diferencias en las distancias genéticas entre las
poblaciones es explicada por la distancia geográfica, es decir, entre más distantes geográficamente se
encuentren las poblaciones habrá mayor diferenciación genética entre ellas (tabla 9).
Figura 20. Correlación entre la distancia geográfica (km) y la distancia genética (Fst) de las poblaciones obtenidas.
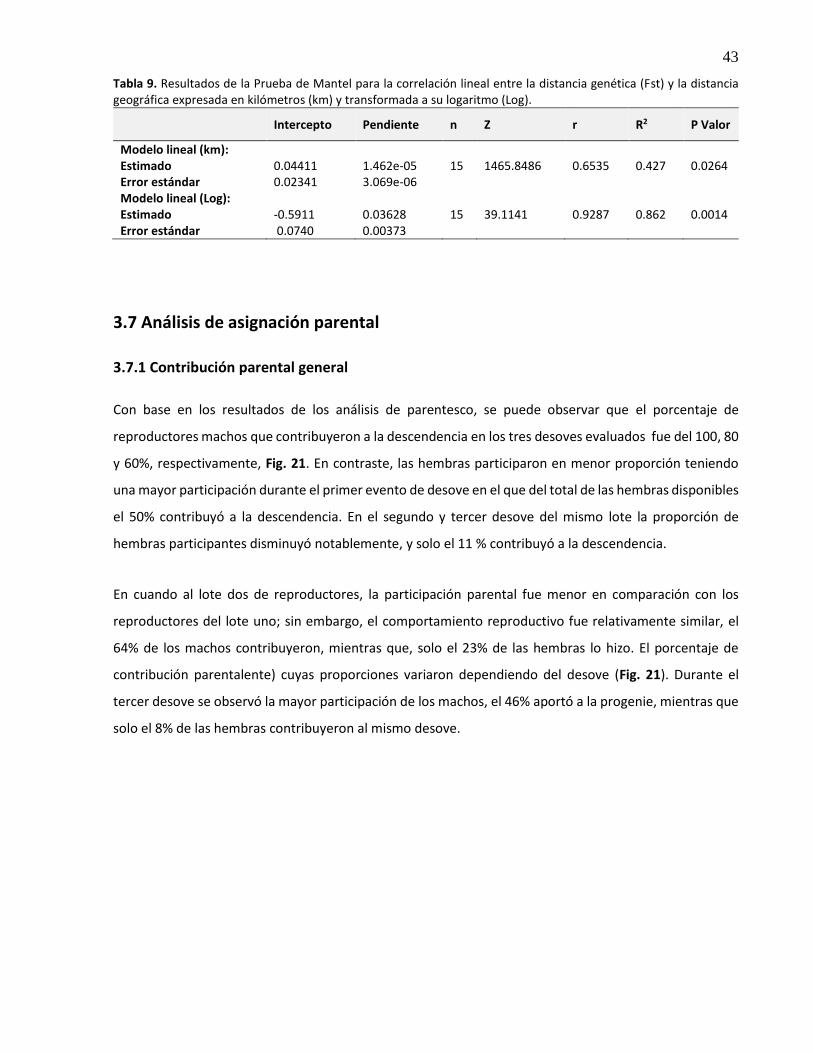
43
Tabla 9. Resultados de la Prueba de Mantel para la correlación lineal entre la distancia genética (Fst) y la distancia geográfica expresada en kilómetros (km) y transformada a su logaritmo (Log).
Intercepto Pendiente n Z r R2 P Valor
Modelo lineal (km):
Estimado 0.04411 1.462e-05 15 1465.8486 0.6535 0.427 0.0264 Error estándar 0.02341 3.069e-06
Modelo lineal (Log): Estimado -0.5911 0.03628 15 39.1141 0.9287 0.862 0.0014 Error estándar 0.0740 0.00373
3.7 Análisis de asignación parental
3.7.1 Contribución parental general
Con base en los resultados de los análisis de parentesco, se puede observar que el porcentaje de
reproductores machos que contribuyeron a la descendencia en los tres desoves evaluados fue del 100, 80
y 60%, respectivamente, Fig. 21. En contraste, las hembras participaron en menor proporción teniendo
una mayor participación durante el primer evento de desove en el que del total de las hembras disponibles
el 50% contribuyó a la descendencia. En el segundo y tercer desove del mismo lote la proporción de
hembras participantes disminuyó notablemente, y solo el 11 % contribuyó a la descendencia.
En cuando al lote dos de reproductores, la participación parental fue menor en comparación con los
reproductores del lote uno; sin embargo, el comportamiento reproductivo fue relativamente similar, el
64% de los machos contribuyeron, mientras que, solo el 23% de las hembras lo hizo. El porcentaje de
contribución parentalente) cuyas proporciones variaron dependiendo del desove (Fig. 21). Durante el
tercer desove se observó la mayor participación de los machos, el 46% aportó a la progenie, mientras que
solo el 8% de las hembras contribuyeron al mismo desove.

44
Figura 21. Porcentaje de reproductores que contribuyen a la descendencia por desove y total.
3.7.2 Aporte parental individual
3.7.2.1 Lote uno
En el primer desove natural del lote uno de reproductores de Bahía de Todos Santos, de abril de 2014
donde se evaluaron 71 crías, los resultados muestran que los nueve machos contribuyeron a la F1, y los
machos M2 y M5 en conjunto contribuyeron con el 46% de la progenie evaluada. Sin embargo, en el caso
de las hembras dos de ellas (H1, H4) contribuyeron con el 71% de las crías (Fig. 22). Para el primer desove
la mayoría de los juveniles pudieron ser asignados exitosamente.
Posterior al primer evento de desove, fueron introducidos dos reproductores provenientes de Bahía
Magdalena que se anexaron al análisis para calcular las proporciones de los desoves restantes (Fig. 22).
Durante el segundo evento de desove el macho M6 y la hembra H8 resultaron ser los que participaron en
mayor proporción con el 29 y 19%, respectivamente. Sin embargo, en particular en el segundo desove, el
porcentaje de la progenie que no puedo ser asignadas fue alto (36% padre y 81% madre). De forma similar,
en el desove tres solo una hembra (H3) contribuyó al desove en un 70%, mientras que, cuatro machos
(M6, M7, M8 y M10) participaron con una proporción homogénea individual del 20%.
100
80
60
100
50
11 11
55
36
29
46
64
15 15
8
23
0
10
20
30
40
50
60
70
80
90
100
Desove 1 Desove 2 Desove 3 Total
Po
rcen
taje
de
rep
rod
uct
ore
s q
ue
con
trib
uye
n a
la d
esce
nd
enci
a (%
)
Machos lote 1 Hembras lote 1 Machos lote 2 Hembras lote 2
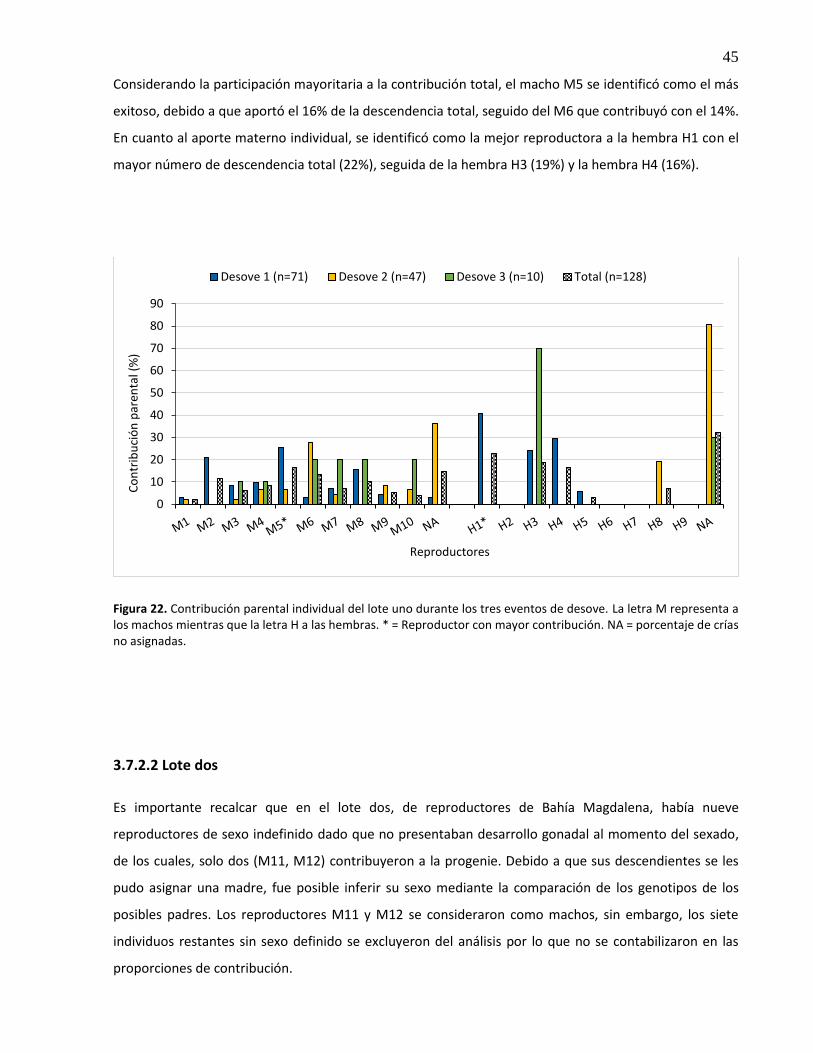
45
Considerando la participación mayoritaria a la contribución total, el macho M5 se identificó como el más
exitoso, debido a que aportó el 16% de la descendencia total, seguido del M6 que contribuyó con el 14%.
En cuanto al aporte materno individual, se identificó como la mejor reproductora a la hembra H1 con el
mayor número de descendencia total (22%), seguida de la hembra H3 (19%) y la hembra H4 (16%).
Figura 22. Contribución parental individual del lote uno durante los tres eventos de desove. La letra M representa a los machos mientras que la letra H a las hembras. * = Reproductor con mayor contribución. NA = porcentaje de crías no asignadas.
3.7.2.2 Lote dos
Es importante recalcar que en el lote dos, de reproductores de Bahía Magdalena, había nueve
reproductores de sexo indefinido dado que no presentaban desarrollo gonadal al momento del sexado,
de los cuales, solo dos (M11, M12) contribuyeron a la progenie. Debido a que sus descendientes se les
pudo asignar una madre, fue posible inferir su sexo mediante la comparación de los genotipos de los
posibles padres. Los reproductores M11 y M12 se consideraron como machos, sin embargo, los siete
individuos restantes sin sexo definido se excluyeron del análisis por lo que no se contabilizaron en las
proporciones de contribución.
0
10
20
30
40
50
60
70
80
90
Co
ntr
ibu
ció
n p
aren
tal (
%)
Reproductores
Desove 1 (n=71) Desove 2 (n=47) Desove 3 (n=10) Total (n=128)
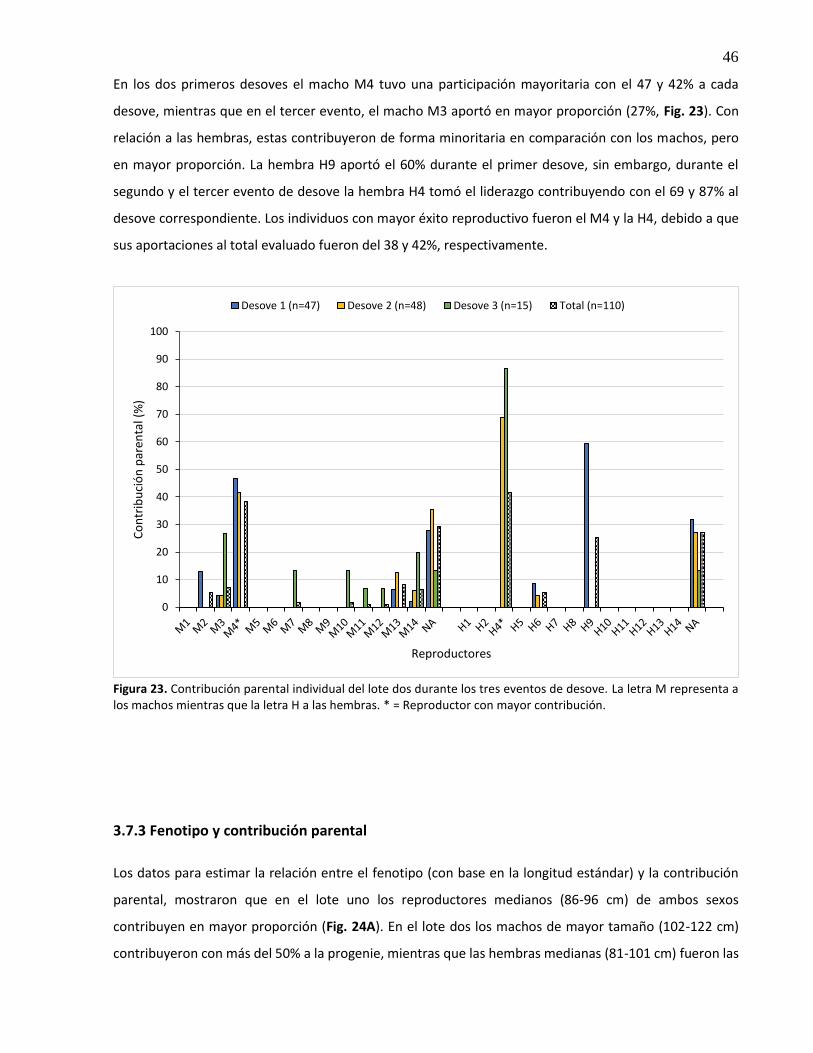
46
En los dos primeros desoves el macho M4 tuvo una participación mayoritaria con el 47 y 42% a cada
desove, mientras que en el tercer evento, el macho M3 aportó en mayor proporción (27%, Fig. 23). Con
relación a las hembras, estas contribuyeron de forma minoritaria en comparación con los machos, pero
en mayor proporción. La hembra H9 aportó el 60% durante el primer desove, sin embargo, durante el
segundo y el tercer evento de desove la hembra H4 tomó el liderazgo contribuyendo con el 69 y 87% al
desove correspondiente. Los individuos con mayor éxito reproductivo fueron el M4 y la H4, debido a que
sus aportaciones al total evaluado fueron del 38 y 42%, respectivamente.
Figura 23. Contribución parental individual del lote dos durante los tres eventos de desove. La letra M representa a los machos mientras que la letra H a las hembras. * = Reproductor con mayor contribución.
3.7.3 Fenotipo y contribución parental
Los datos para estimar la relación entre el fenotipo (con base en la longitud estándar) y la contribución
parental, mostraron que en el lote uno los reproductores medianos (86-96 cm) de ambos sexos
contribuyen en mayor proporción (Fig. 24A). En el lote dos los machos de mayor tamaño (102-122 cm)
contribuyeron con más del 50% a la progenie, mientras que las hembras medianas (81-101 cm) fueron las
0
10
20
30
40
50
60
70
80
90
100
Co
ntr
ibu
ció
n p
aren
tal (
%)
Reproductores
Desove 1 (n=47) Desove 2 (n=48) Desove 3 (n=15) Total (n=110)

47
que presentaron un mayor aporte (Fig. 24B). Sin embargo, debe considerarse que la variabilidad en
tamaños en el lote uno fue menor en relación al lote dos, por lo que el número de reproductores por
categoría fue desigual.
Figura 24. Relación entre la longitud estándar (cm) y la contribución parental en reproductores del lote uno (A) y dos (B).
Para considerar una clasificación más homogénea se realizó un nuevo agrupamiento, considerando a los
reproductores de ambos lotes. En la Figura 25 se puede observar que los reproductores medianos (79-99
cm) contribuyeron con el 58% del total de la descendencia muestreada, seguida de los reproductores
grandes (100-120 cm) que aportaron el 41%. Los reproductores de una longitud entre 58 a 78 cm tuvieron
un aporte insignificante a la descendencia (1%).
Longitud estándar (cm)
75-85 86-96 97-107
Co
ntr
ibució
n p
are
nta
l to
tal (%
)
0
20
40
60
80
100
Machos
Hembras
Longitud estándar (cm)
60-80 81-101 102-122
Co
ntr
ibució
n p
are
nta
l (%
)
0
20
40
60
80
100
Machos
Hembras(A) (B)
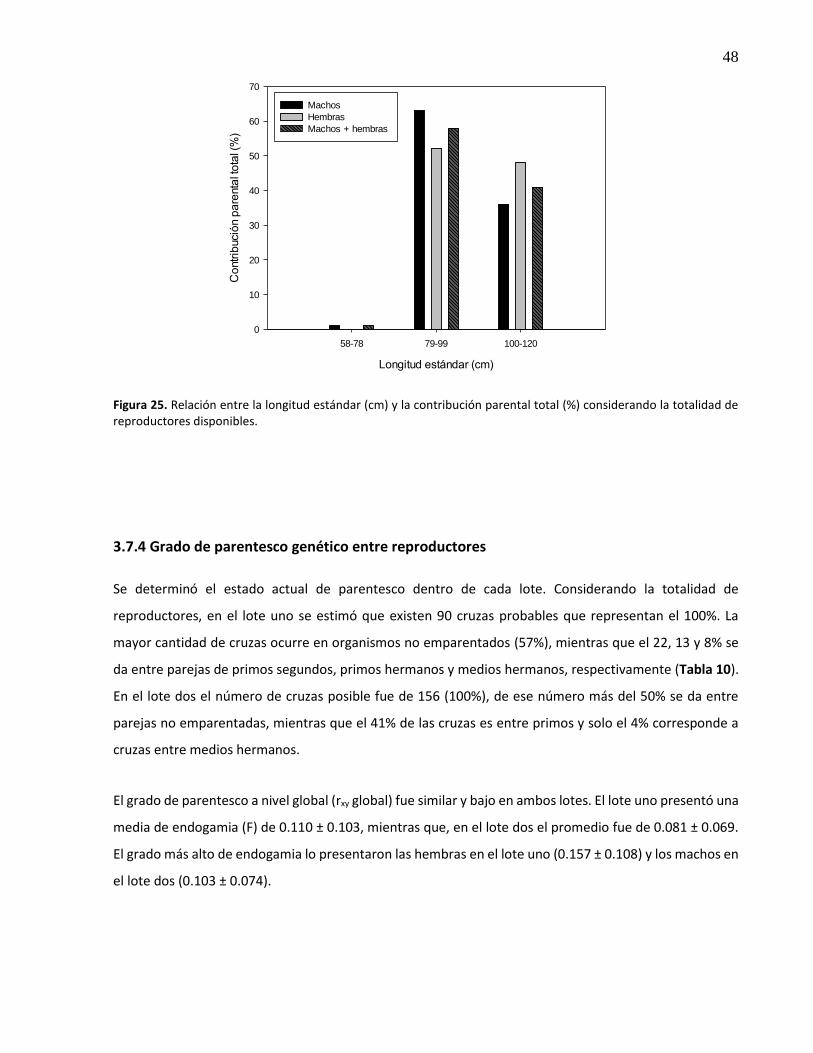
48
Figura 25. Relación entre la longitud estándar (cm) y la contribución parental total (%) considerando la totalidad de reproductores disponibles.
3.7.4 Grado de parentesco genético entre reproductores
Se determinó el estado actual de parentesco dentro de cada lote. Considerando la totalidad de
reproductores, en el lote uno se estimó que existen 90 cruzas probables que representan el 100%. La
mayor cantidad de cruzas ocurre en organismos no emparentados (57%), mientras que el 22, 13 y 8% se
da entre parejas de primos segundos, primos hermanos y medios hermanos, respectivamente (Tabla 10).
En el lote dos el número de cruzas posible fue de 156 (100%), de ese número más del 50% se da entre
parejas no emparentadas, mientras que el 41% de las cruzas es entre primos y solo el 4% corresponde a
cruzas entre medios hermanos.
El grado de parentesco a nivel global (rxy global) fue similar y bajo en ambos lotes. El lote uno presentó una
media de endogamia (F) de 0.110 ± 0.103, mientras que, en el lote dos el promedio fue de 0.081 ± 0.069.
El grado más alto de endogamia lo presentaron las hembras en el lote uno (0.157 ± 0.108) y los machos en
el lote dos (0.103 ± 0.074).
2D Graph 20
Longitud estándar (cm)
58-78 79-99 100-120
Co
ntr
ibució
n p
are
nta
l to
tal (
%)
0
10
20
30
40
50
60
70
Machos
Hembras
Machos + hembras
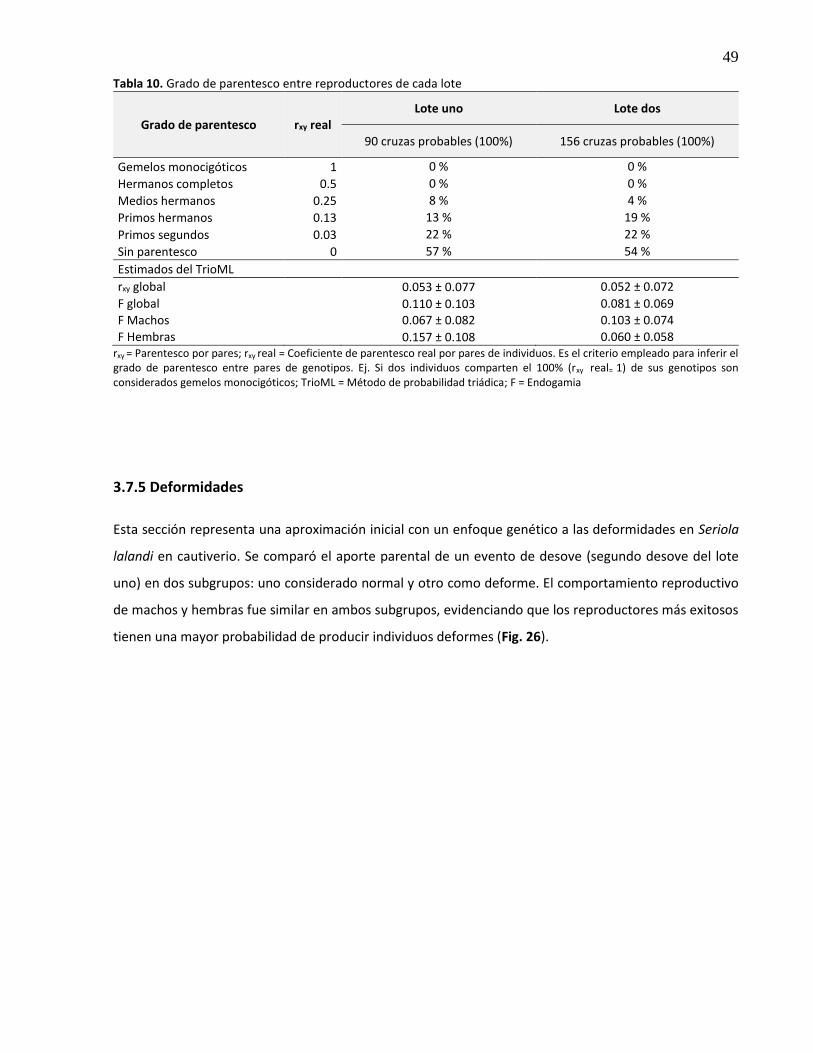
49
Tabla 10. Grado de parentesco entre reproductores de cada lote
Grado de parentesco rxy real Lote uno Lote dos
90 cruzas probables (100%) 156 cruzas probables (100%)
Gemelos monocigóticos 1 0 % 0 %
Hermanos completos 0.5 0 % 0 %
Medios hermanos 0.25 8 % 4 %
Primos hermanos 0.13 13 % 19 %
Primos segundos 0.03 22 % 22 %
Sin parentesco 0 57 % 54 %
Estimados del TrioML
rxy global 0.053 ± 0.077 0.052 ± 0.072
F global 0.110 ± 0.103 0.081 ± 0.069
F Machos 0.067 ± 0.082 0.103 ± 0.074
F Hembras 0.157 ± 0.108 0.060 ± 0.058 rxy = Parentesco por pares; rxy real = Coeficiente de parentesco real por pares de individuos. Es el criterio empleado para inferir el grado de parentesco entre pares de genotipos. Ej. Si dos individuos comparten el 100% (rxy real= 1) de sus genotipos son considerados gemelos monocigóticos; TrioML = Método de probabilidad triádica; F = Endogamia
3.7.5 Deformidades
Esta sección representa una aproximación inicial con un enfoque genético a las deformidades en Seriola
lalandi en cautiverio. Se comparó el aporte parental de un evento de desove (segundo desove del lote
uno) en dos subgrupos: uno considerado normal y otro como deforme. El comportamiento reproductivo
de machos y hembras fue similar en ambos subgrupos, evidenciando que los reproductores más exitosos
tienen una mayor probabilidad de producir individuos deformes (Fig. 26).
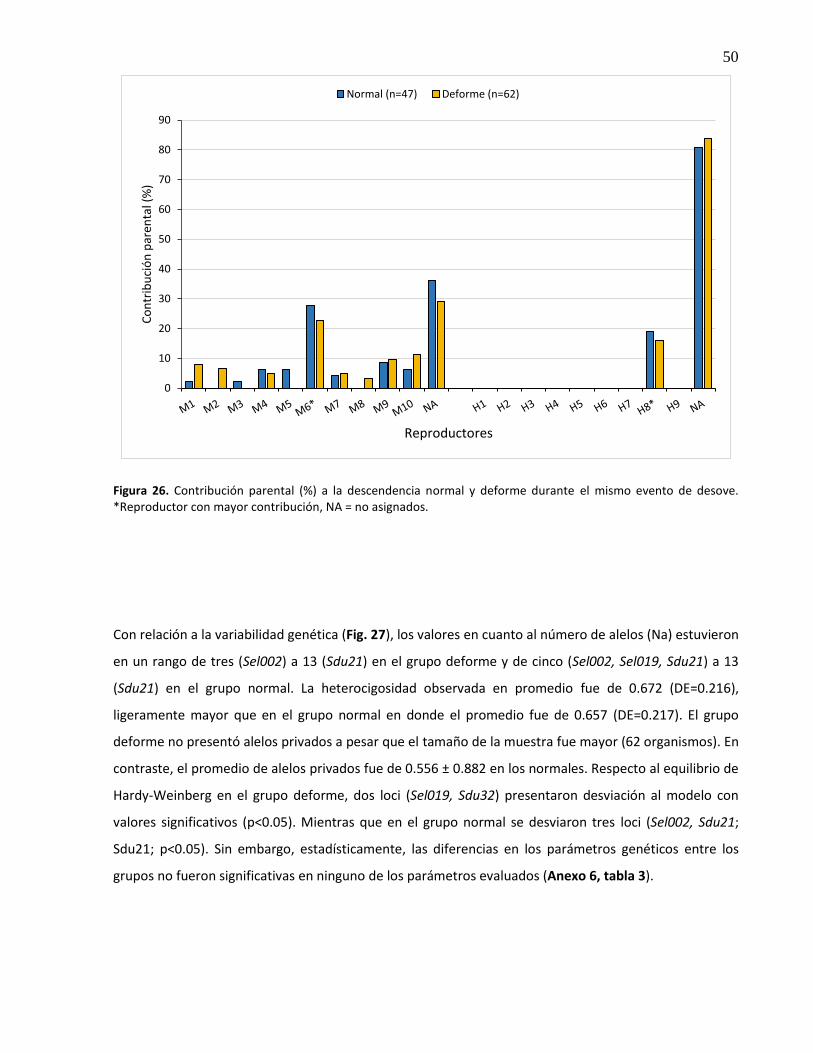
50
Figura 26. Contribución parental (%) a la descendencia normal y deforme durante el mismo evento de desove. *Reproductor con mayor contribución, NA = no asignados.
Con relación a la variabilidad genética (Fig. 27), los valores en cuanto al número de alelos (Na) estuvieron
en un rango de tres (Sel002) a 13 (Sdu21) en el grupo deforme y de cinco (Sel002, Sel019, Sdu21) a 13
(Sdu21) en el grupo normal. La heterocigosidad observada en promedio fue de 0.672 (DE=0.216),
ligeramente mayor que en el grupo normal en donde el promedio fue de 0.657 (DE=0.217). El grupo
deforme no presentó alelos privados a pesar que el tamaño de la muestra fue mayor (62 organismos). En
contraste, el promedio de alelos privados fue de 0.556 ± 0.882 en los normales. Respecto al equilibrio de
Hardy-Weinberg en el grupo deforme, dos loci (Sel019, Sdu32) presentaron desviación al modelo con
valores significativos (p<0.05). Mientras que en el grupo normal se desviaron tres loci (Sel002, Sdu21;
Sdu21; p<0.05). Sin embargo, estadísticamente, las diferencias en los parámetros genéticos entre los
grupos no fueron significativas en ninguno de los parámetros evaluados (Anexo 6, tabla 3).
0
10
20
30
40
50
60
70
80
90
Co
ntr
ibu
ció
n p
aren
tal (
%)
Reproductores
Normal (n=47) Deforme (n=62)
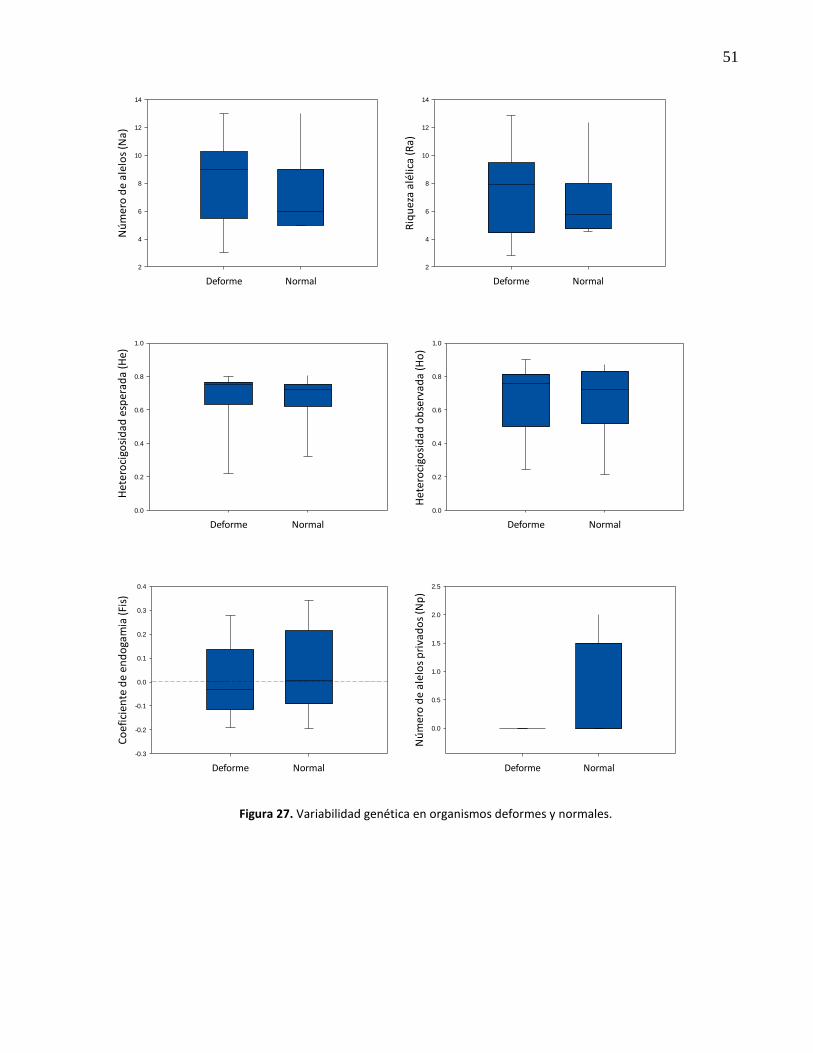
51
Figura 27. Variabilidad genética en organismos deformes y normales.
Deforme Normal
Nú
me
ro d
e a
lelo
s (N
a)
2
4
6
8
10
12
14
Deforme Normal
Riq
ue
za a
lélic
a (
Ra
)
2
4
6
8
10
12
14
Deforme Normal
Het
ero
cigo
sid
ad e
sper
ada
(He)
0.0
0.2
0.4
0.6
0.8
1.0
Deforme Normal
Het
ero
cigo
sid
ad o
bse
rvad
a (H
o)
0.0
0.2
0.4
0.6
0.8
1.0
Deforme Normal
Co
efic
ien
te d
e en
do
gam
ia (F
is)
-0.3
-0.2
-0.1
0.0
0.1
0.2
0.3
0.4
Deforme Normal
Nú
me
ro d
e a
lelo
s p
riva
do
s (N
p)
0.0
0.5
1.0
1.5
2.0
2.5

52
Capítulo 4. Discusión
En la presente investigación se describieron tres principales aspectos: 1) la diversidad genética (Capítulo
3 sección 3.5) de las poblaciones silvestres y en cautiverio de S. lalandi a escala regional y global; 2) la
estructura genética (grado de diferenciación; Capítulo 3 sección 3.6) dentro de las poblaciones mexicanas,
así como en aquellas de diferentes partes del mundo; para concluir con 3) el éxito reproductivo (Capítulo
3 sección 3.7) de los reproductores cautivos, a través de la contribución parental a la descendencia. En el
punto anterior, se realizó también una aproximación inicial al estudio de las deformidades craneales desde
una perspectiva genética.
Con base en los resultados obtenidos se puede deducir lo siguiente: 1) No existen diferencias significativas
entre las poblaciones silvestres estudiadas y la primera generación filial cultivada, 2) Los individuos de las
localidades silvestres de S. lalandi en México corresponden a una sola población genéticamente
homogénea, mientras que, las poblaciones extranjeras muestreadas (EUA, CHL, AUS) en ambos
hemisferios, presentan un grado de diferenciación genética muy alto; y finalmente 3) la contribución
parental de los reproductores en cautiverio difiere en términos de proporciones y frecuencias.
A partir de lo anterior, se discutirán los principales hallazgos de la investigación comenzando con las
generalidades respecto a los microsatélites empleados en el estudio, para posteriormente profundizar en
cada una de las secciones en relación a las preguntas de la investigación.
4.1 Generalidades de los microsatélites
La caracterización genética en poblaciones de Seriola lalandi se ha basado principalmente en el uso de
microsatélites desarrollados y aislados a partir de una especie diferente (heterólogos). Los estudios en los
que se utilizaron microsatélites de Seriola dumerili, se hicieron en poblaciones de S. lalandi de disintas
regiones geográficas como: Japón (tres microsatélites; Nugroho et al., 2001), Australia y Nueva Zelanda
(siete microsatélites; Miller et al., 2011) y Sudáfrica (seis microsatélites; Swart et al., 2016). Los estudios
que engloban poblaciones de Estados Unidos (Purcell et al., 2015) y Chile (Fernández et al., 2015)
emplearon una combinación de microsatélites de S. dumerili y S. quinqueradiata. Otros trabajos con S.
lalandi, combinan los microsatélites con otro tipo de marcadores moleculares, a partir de secuencias de

53
genes del DNA mitocondrial y DNA nuclear, para obtener una mayor cantidad de información molecular
(Swart, 2014; Purcell et al., 2015 ).
La desventaja en el uso de microsatélites heterólogos se ve reflejada en la ausencia de amplificación (alelos
nulos). En el 2013, Whatmore et al., diseñaron un set de microsatélites especialmente para S. lalandi a
partir de secuencias transcriptómicas del hígado y del sistema digestivo, lo que podría facilitar en el futuro
la identificación de loci de rasgos cuantitativos (QTL’s por sus siglas en Inglés). Sin embargo, las secuencias
de los cebadores no están depositadas en el GenBank (https://www.ncbi.nlm.nih.gov/genbank/) debido a
restricciones de la empresa que financió el desarrollo de los mismos (Dr. Knibb, University of the Sunshine
Coast, 2016. Com. Pers.).
En este estudio se emplearon seis loci microsatélites específicos para S. lalandi y tres heterólogos (aislados
de S. dumerili) para la caracterización genética de las poblaciones silvestres y en cautiverio de Seriola
lalandi, así como para el análisis de asignación parental. Aproximadamente el 80% de los loci resultaron
ser altamente informativos (Índice de información polimórfica 0.86). En general, la frecuencia de alelos
nulos fue baja (2.7% ± 2.0%), sin embargo, las poblaciones silvestres mexicanas de Bahía de Todos Santos
(BT), Ejido Eréndira (EE), y Bahía de los Ángeles (BA) presentaron una frecuencia de alelos nulos mayor al
20% en los loci heterólogos Sdu46, Sdu21 y Sdu21, respectivamente. La mayor frecuencia de alelos nulos
en los loci Sdu46 y Sdu21 (4.9 ± 1.9%), aislados y caracterizados en S. dumeriili (Renshaw et al., 2006, 2007),
especie que habita en las regiones tropicales y subtropicales, se debe a su lejanía filogenética con S. lalandi.
Recientemente, Swart y colaboradores (2015) describen las relaciones filogenéticas basadas en análisis
moleculares de las nueve especies que conforman el género Seriola, y calculan que el grupo de las especies
tropicales/subtropicales entre las que se encuentra S. dumeriili, y el grupo de las especies templadas,
donde se ubica S. lalandi se separaron hace aproximadamente 55 millones de años.
4.2 Diversidad genética
4.2.1 Caracterización genética regional de las poblaciones silvestres de Seriola lalandi
El número de alelos, así como, la heterocigosidad son considerados los parámetros genéticos principales
para cuantificar la diversidad genética (Nugroho et al., 2001). En las poblaciones de Baja California el
número de alelos varió de 4 a 34 (Anexo 3, tablas 7 a 9). Además, se observó un nivel moderado de

54
endogamia en tres de las poblaciones locales (BT Fis= 0.13; EE Fis= 0.19; BA Fis= 0.10; Anexo 3, tablas 6, 7
y 9). El grado de endogamia observado en las poblaciones silvestres puede estar asociado a dos factores
principales: la presencia de alelos nulos y al comportamiento social. La alta frecuencia de alelos nulos en
las localidades de BT (4.8 ± 10.6% en 4 loci), EE (7.9 ± 9.9% en 8 loci) y BA (5 ± 14%, en 7 loci) podría explicar
el nivel moderado de endogamia observado en dichas poblaciones, atribuible a un déficit de heterocigotos.
Este déficit pudo deberse a mutaciones en los sitios de reconocimiento de los cebadores de uno o más de
los alelos presentes analizados (Aranguren-Méndez et al., 2005).
Respecto al comportamiento social, Sakakura y Tsukamoto (1998) encontraron que la organización dentro
de los cardúmenes en Seriola quinqueradiata está determinada por el grado de agresividad de los
organismos a los cuales dividieron en tres categorías: los individuos dominantes, presentes en una
proporción de 10 a 20% , los intermedios (10 a 20%) que tienen la capacidad de asumir el liderazgo si el
grupo dominante desaparece, y finalmente, los subordinados que son el grupo mayoritario (60 a 80%). El
comportamiento gregario, además de incrementar la probabilidad de sobrevivencia individual y del
cardumen en el medio natural (comportamiento antipredatorio), posiblemente también beneficia la eficaz
transferencia de estrategias de caza a las nuevas generaciones como ocurre en otros verebrados. Sin
embargo, la gran desventaja de este comportamiento social es la mayor probabilidad de cruzamiento
entre organismos con algún grado de parentesco genético. Dado que se ha sugerido que algunas especies
de peces (ej. Phoxinus phoxinus; Barber y Wright, 2001) muestran preferencia por agrupamientos con los
que han pasado un tiempo determinado de su vida.
4.2.2 Diversidad genética en poblaciones silvestres extranjeras
La variabilidad genética en términos de heterocigosidad observada (Ho) en las poblaciones estudiadas fue
alta y varió de 0.689 a 0.716 para los nueve loci microsatélites evaluados (Anexo 3, tablas 10 y 11). En
general, este parámetro registro valores similares a los reportados en diversos estudios: Nugroho et al.
(2001) en poblaciones de S. lalandi de Australia (0.782), Nueva Zelanda (0.650) y Japón (0.750); Swart
(2014) en cinco regiones Sudafricanas (0.700); Purcell et al. (2015) en una población silvestre de las costas
de California (0.700) y Fernández et al. (2015) en dos poblaciones chilenas (0.710)
Sin embargo, la heterocigosidad observada para la población de Nueva Gales del Sur, Australia en este
estudio (0.716; Anexo 3, tablas 10 y 11) fue diferente de la reportada por Miller et al. (2011) en las
poblaciones Nueva Zelanda (0.420) y de Australia (0.420). Una diferencia importante entre el estudio de

55
Miller et al. (2001) y el presente, es el tipo y número de microsatélites analizados, en el primer caso fueron
siete loci microsatélites diseñados para S. dumerili, mientras que en este trabajo fueron nueve, seis
especficos y tres heterólogos, por lo cual los resultados obtenidos son mas informativos.
Con relación a las estimaciones de los coeficientes de endogamia (Fis) solamente la población chilena
proveniente de la región de Antofagasta, presentó una tendencia al cruzamiento aleatorio (panmixia; Fis
= 0.051). Lo anterior difiere de lo reportado por Fernández et al. (2015) en dos poblaciones silvestres
chilenas de S. lalandi capturadas en Punta Frodden y Bahía Inglesa, cuyos coeficientes de endogamia
fueron moderados (0.11 y 0.23, respectivamente).
4.2.3 Variabilidad genética en el cultivo de S. lalandi
La diversidad genética, considerando el número de alelos corregido por el tamaño de la muestra (Riqueza
alélica; Ra) en los organismos producidos en cautiverio (F1), varió de 2 a 13.747 (Anexo 3, tablas 1 a 5). En
comparación con las poblaciones silvestres, las F1 de Bahía de Todos Santos (BT), Bahía Magdalena (BM),
Antofagasta, Chile, y Nueva Gales del Sur, Australia, presentaron una reducción aproximada de la riqueza
alélica (Ra) del 23, 46, 28 y 38%, respectivamente.
Está reducción en la variabilidad genética se debe principalmente a un efecto fundador, el cual erosiona
la variabilidad genética en cautiverio mediante la selección de pocos reproductores en comparación con
el medio silvestre (Norris et al., 1999). Asimismo, la pérdida de los alelos de forma aleatoria (deriva
genética) se vio reflejada principalmente en las poblaciones F1AUS y F1BM, mediante la desaparición de
los alelos privados (presentes en menor frecuencia) en comparación con sus respectivas poblaciones
silvestres (Anexo 3, tablas 8 y 11).
Solamente F1BT evidenció un ligero grado de endogamia (Fis= 0.068) posiblemente asociado al reducido
tamaño del grupo reproductor (n=19), que presentó un coeficiente moderado de endogamia (BT Fis = 0.13)
(Tabla 10; Anexo 3, tabla 6). En contraste, la progenie F1EUA (Fis= -0.11) y F1AUS (Fis= -0.14) mostraron
un nivel moderado de exogamia que puede estar relacionado con la introducción de reproductores de
diferentes áreas geográficas y/o las estrategias de crianza selectiva, como el cruzamiento entre familias
diferentes, que siguen las tres empresas comerciales muestreadas para la producción de jurel en
cautiverio, disminuyendo así la probabilidad de cruzamiento entre parientes. Los lotes de crías F1BM y
F1CHL estuvieron en panmixia (Figura 13, Anexo 3 tabla 2 y 4), indicativo de un apareamiento aleatorio.

56
Con respecto al Equilibrio de Hardy-Weinberg (EHW), que postula que la heterocigosidad observada es
estadísticamente igual a la heterocigosidad esperada, se determinó que en los lotes producidos en
cautiverio en México (F1BT y F1BM) siete de nueve loci se encuentran en desequilibrio con un nivel de
significancia de alto a muy alto (p<0.01 a 0.001; Anexo 3, tablas 1 y 2), mientras, que la F1 de CHL y AUS,
presentaron uno y tres loci en desequilibrio, respectivamente. El lote F1EUA estuvo en Equilibrio de Hardy-
Weinberg en todos los loci (Anexo 3, tablas 3, 4 y 5).
El desequilibrio de Hardy-Weinberg en la mayoría de los loci de las poblaciones mexicanas, así como, la
reducción en la riqueza alélica (Ra) del 23% en la F1 de Bahía de Todos Santos y del 46% en la F1 de Bahía
Magdalena, son evidencia de la falta de manejo genético del, hasta hoy dia, único criadero de Jurel de
Castilla en México. Cabe resaltar, que aunque existió un cultivo piloto experimental de engorda de
organismos silvestres de esta especie en BCS en el año 2001 (Avilés y Castelló, 2004), no fue sino hasta el
año 2014, que se estableció el primer laboratorio comercial (Luening, 2014; Scarratt, 2014). Por otra parte,
en Australia se inició el cultivo comercial en el año 2000, y las empresas están fuertemente asesoradas por
investigadores de universidades reconocidas (Dr. Knibb, University of the Sunshine Coast, 2016. Com.
Pers.), siendo el mismo caso de las empresas privadas en Chile, en donde además, el gobierno tiene como
prioridad la diversificación de la acuicultura (Fernández et al., 2015; Orellana et al., 2014). Mientras, que
en el caso de EUA, el Instituto de Investigación de Hubbs-Sea World (HSWRI, por sus siglás en inglés), que
lleva a cabo la reproducción de la especie, cuenta con un programa de crianza basado en marcadores
moleculares (Stuart y Drawbridge, 2013).
La introducción de alevines de diversas partes del mundo, con la finalidad de emplearlos posteriormente
como reproductores en los criaderos de jurel, podría desencadenar dos tipos de respuestas: la primera
posibilidad es el vigor híbrido, es decir, que en la primera generación filial producto de la cruza de las
poblaciones locales y extranjeras se observe una mejora respecto a los progenitores en algunas
características deseables, por ejemplo en la tasa de crecimiento (Refstie y Gjedrem, 1975). Sin embargo,
es posible que tenga un efecto contrario debido a la depresión exogámica que puede provocar un
desequilibrio en complejos de genes coadaptados para fortalecer el sistema inmunológico (Goldberg et
al., 2005), o la pérdida de adaptaciones locales en las poblaciones (Houde et al., 2011). Un ejemplo de
depresión exogámica se observó en la lobina Micropterus salmoides donde se mezclaron reproductores
de orígenes geográficos diferentes y se demostró que la segunda generación filial (F2), expuesta a agentes
patógenos, tuvo una supervivencia significativamente inferior debido al incremento en la susceptibilidad
al patógeno (Goldberg et al., 2005).

57
En acuicultura, el vigor híbrido así como la depresión exogámica tienen consecuencias económicas, por lo
tanto, la introducción de reproductores de S. lalandi de regiones diferentes es una decisión muy seria que
debería estar respaldada por estudios específicos de cruzas controladas y seguimiento de varias líneas
genéticas y generaciones, para poder dilucidar su efecto a largo plazo. Dado que S. lalandi es un complejo
de tres especies (Martinez-Takeshita et al., 2015) el peligro mayor es que se introduzca al criadero una
especie distinta a las locales.
4.3 Estructura genética
4.3.1 Estructura genética de las poblaciones mexicanas
En el dendrograma de similitudes genéticas de las poblaciones mexicanas se observó un clado principal
con dos agrupamientos (clados secundarios): el primero entre las poblaciones de Bahía de Todos Santos
(BT) y de Bahía Magdalena (BM) y el segundo entre las poblaciones del Ejido Eréndira (EE) y el de Bahía de
los Ángeles (BA) (Fig. 18). Este último resultado fue inesperado dado que el Ejido Eréndira, ubicado en la
costa del Pacífico, está geográficamente más cerca de BT y BM, que de Bahía de los Ángeles, ubicada en el
Golfo de California (Fig. 5B). Sin embargo, cabe destacar que el muestreo de Bahía de los Ángeles se realizó
en el 2015, mientras que el de Eréndira se hizo en abril de 2016, por lo que la similitud podría deberse a
un evento migratorio, dado que que la migración del jurel de poblaciones mexicanas, incluyendo las del
Golfo de California, hacia el norte (EUA) ocurre en las épocas de primavera y verano (Baxter, 1960). El
análisis bayesiano apoya esta posibilidad ya que reveló similitudes genéticas entre las poblaciones del Ejido
Eréndira (EE) y Bahía de los Ángeles (BA) (Fig. 14B).
El análisis discriminante de componentes principales evidenció un alto flujo genético entre las poblaciones
silvestres mexicanas. Lo anterior coincidió con los valores no significativos en el índice de diferenciación
(Rst, Tabla 6), en el análisis de varianza molecular (AMOVA, Tabla 7), así como con el clado principal
observado en el dendrograma (Fig. 18). Por lo que se puede aceptar la hipótesis planteada en este trabajo
y considerar que las muestras de las diferentes localidades silvestres son, en realidad una población
genéticamente homogénea. Lo anterior difiere de lo observado por Baxter (1960) quien sugirió la
presencia de dos poblaciones en el Pacífico mexicano con base al comportamiento migratorio del jurel,
señalando a Isla Cedros, México como el punto de divergencia. La ausencia de estructura genética en las

58
poblaciones mexicanas de Seriola lalandi es atribuible a su alto potencial de migración (Baxter, 1960;
Gillanders et al., 2001)
4.3.2 Diferenciación genética en poblaciones mundiales de Seriola
En relación con el nivel de diferenciación genética entre las poblaciones mundiales de ambos hemisferios,
los resultados obtenidos a partir de los nueve loci microsatélites en los diversos análisis realizados (Análisis
bayesiano, Fig. 15; Análisis discriminante de componentes principales, Fig. 17; Dendrograma de distancias
genéticas, Fig. 19 ; Índice de diferenciación (Rst) Tabla 6; Análisis de varianza molecular, Tabla 8;
Aislamiento por distancia, Tabla 9), demostraron la presencia de dos acervos genéticos completamente
distintos (Fig. 15A), el del hemisferio norte que agrupa a las poblaciones de MEX-EUA y el del hemisferio
sur que comprende las poblaciones de CHL y AUS. Los resultados del presente estudio coinciden con
publicaciones recientes que sugieren que las diferencias genéticas a nivel geográfico son suficientes para
dividir el complejo de Seriola lalandi en varias especies (Swart, 2014; Swart et al., 2015; Martinez-Takeshita
et al., 2015; Purcell et al., 2015).
Miller et al. (2011) analizaron varios loci microsatelitales en las poblaciones de S. lalandi de Australia y
Nueva Zelanda y encontraron diferencias significativas entre las poblaciones del oeste con el resto de las
regiones en Australia y Nueva Zelanda. Esas diferencias fueron atribuidas a barreras geográficas o
ambientales existentes, o del pasado, en virtud de que varias especies de peces de la región presentan un
patrón de diferenciación genética similar. Por otra parte Nugroho et al. (2001) detectaron diferencias
genéticas significativas entre las poblaciones de S. lalandi de Austraila-Nueva Zelanda (hemisferio sur) y
de Japón (Pacífico Noroeste), lo cual justificaria el que al jurel de Australia-Nueva-Zelanda se le conociera
comúnmente como Seriola lalandi lalandi mientras que al de Japón como Seriola lalandi aureovittata.
La posibilidad de que las poblaciones de jurel del hemisferio norte y sur fueran diferentes fue también
propuesto por Ochoa-Saloma (2015) quien determinó variaciones en el tamaño genómico de dos
poblaciones de Seriola lalandi, una originaria de México, y la otra proveniente de Chile, los cuales,
presentaron un tamaño genómico de 0.82 y 0.79 pg, respectivamente; por lo que el autor sugirió se trataba
de especies distintas.
En un trabajo reciente, Martinez-Takeshita et al. (2015) analizaron la estructura genética de Seriola lalandi
de distintas regiones geográficas (México, EUA, Australia, Chile, Japón, Sudáfrica) con dos genes

59
mitocondriales (CR y COI) y cuatro genes nucleares (RAG2, EHHADH, UBE3A, MLL) y observaron la
existencia de diferencias genéticas altamente significativas, suficientes para proponer la separación del
complejo S. lalandi en tres especies distintas: Seriola aureovittata (Pacífico noroeste,Japón), Seriola
dorsalis (Pacífico noreste, México) y Seriola lalandi (Hemisferio sur). Con base en esta información los
autores concluyen que deben reclasificarse taxonómicamente las poblaciones de Japón de Seriola lalandi
a Seriola aureovittata y las del Noroeste del Pacífico de Seriola lalandi a Seriola dorsalis. Otro estudio
reciente (Purcell et al., 2015) confirmo los resultados de Martinez-Takeshita et al (2015) y detectó otro
grupo divergente en el Océano atlántico (Sudáfrica) que podría representar una cuarta especie dentro del
complejo Seriola lalandi.
La incertidumbre taxonómica en especies crípticas podría resolverse mediante estudios citogenéticos. Esto
es posible debido a que el número, tamaño, forma y estructura de los cromosomas difieren entre especies
(Cipriano et al., 2014; Nirchio y Oliveira, 2014; Perazzo et al., 2011). A la fecha, solo se ha realizado un
estudio citogenético en Seriola lalandi, en organismos de Australia, en el que se comparó el cariotipo de
Seriola quinqueradiata de Japón (Chai et al,. 2009). Ambas especies tienen 48 cromosomas pero difieren
en un par de cromosomas metacéntricos. Un estudio de este tipo en las poblaciones del Pacífico,
incrementaría las evidencias de la existencia de diferentes especies en el complejo de S. lalandi.
Entre las causas que influyen en la diferenciación genética de Seriola lalandi, se ha sugerido que las
variaciones entre las temporadas reproductivas entre los hemisferios norte y sur dificultan el flujo genético
entre ellas (Purcell et al., 2015). La reproducción de la especie en el hemisferio norte ocurre en primavera
verano (marzo – agosto; Baxter, 1960; Stuart y Drawbridge, 2013) mientras que en el hemisferio sur se
presenta de octubre a diciembre (Gillanders et al., 1999). Otros factores asociados a la divergencia entre
poblaciones de ambos hemisferios, son las condiciones térmicas y corrientes oceánicas en la línea
Ecuatorial (Figura 29) que actúan como una barrera geográfica restringiendo el intercambio genético entre
regiones oceánicas (Swart, 2014; Swart et al., 2015; Purcell et al., 2015). Swart et al. (2016) proponen que
S. aureovittata, S. dorsalis y S. lalandi, posiblemente se originaron en el oceáno Pacífico hace
aproximadamente 115 000 – 500 000 años, en el Pleistoceno tardío; debido al establecimiento del
gradiente térmico en el océano, después del periodo interglacial (Domínguez-López et al., 2010).

60
Figura 28. Barrera térmica ecuatorial. Fuente: NASA's Goddard Space Flight Center, http://svs.gsfc.nasa.gov/10953
4.4 Éxito reproductivo en cautiverio (BC)
Los cultivos de peces marinos que son, hoy en día, rentables a nivel mundial, son aquellos que han logrado
obtener de forma controlada juveniles, y no dependen del medio silvestre para la recolección de
organismos para su engorda (e.g., industria del salmón). En general, para obtener huevos en cantidad y
calidad se necesita un manejo adecuado de los reproductores, y estos reproductores, cuando se inicia el
cultivo de una nueva especie, son obtenidos mediante captura en el mar. Tal es el caso de la empresa
Ocean Baja Labs, que inicio operaciones en México en el año 2012, y adquirió su primer lote de
reproductores provenientes de Bahía Todos Santos, BC, en mayo y junio de 2013. Posteriormente, en el
año 2014 adquirió un segundo lote de reproductores en las proximidades de Bahía Magdalena, BCS.
Para el lote de reproductores originarios de Bahía de Todos Santos (lote uno) se observó una contribución
a la descendencia del 100% de los machos y el 55% de las hembras, considerando los tres desoves
evaluados (2014-2015;). A diferencia del grupo de reproductores de Bahía Magdalena (lote dos), en donde
se observó una participación de los machos del 64% y del 23% de las hembras (2015; Fig. 21). Estas
diferencias podrían estar relacionadas propiamente con las características de los reproductores
capturados y al manejo de los lotes, desde su captura hasta el primer evento de desove en cautividad.
En el caso del lote uno, cuando los reproductores ingresaron al criadero en 2013 (n=19), el peso promedio
fue de 4.21.7 kg y una biomasa total de 79.5 kg. Este lote presentó sus primeros eventos de desove

61
natural a finales de marzo de 2014, y el primer evento de desove muestreado fue el de abril de 2014,
nueve meses después de su llegada al plantel y después de haber duplicado su peso, alcanzando un
promedio de 9.51.7 kg (biomasa total de 179.7 kg). A los cuatro-cinco meses después de su ingreso al
criadero, se identificó su sexo mediante una biopsia de las gónadas, además, cabe considerar, que la
distancia de transporte desde la zona de colecta a la granja es de aproximadamente 100 kms por carretera.
En el caso del lote dos de reproductores, su ingreso fue en junio de 2014 (n=35), con un peso promedio de
103.8 kg y una biomasa total de 350 kg, y los primeros eventos de desove fueron registrados a finales de
enero de 2015, seis meses después de su arribo al plantel, cuando alcanzaron un peso de 13.65.4 kg con
una biomasa total de 420.7 kg en el estanque de mantenimiento. Cuando los reproductores ingresaron al
plantel, también fueron sexados mediante biopsia y algunos fueron inducidos al desove mediante
implantes con hormonas. La distancia de transporte de la zona de colecta a la granja es de
aproximadamente 1000 kms por carretera.
Tomando estos datos del cultivo en consideración, se observó que la biomasa y densidad de organismos
del lote dos respecto al uno fue del doble, y aunque ambos estanques mantenian una proporción de sexos
de aproximadamente 1:1, el desempeño en el estanque de menor densidad fue mejor asociado quizás al
mayor tiempo de acondicionamiento de éstos organismos a las condiciones de cultivo (9 vs 6 meses), su
menor edad al ingresar al plantel (4.2 vs 10 kgs), un menor estrés de manejo en la captura (diferencias en
zona de capturas y tiempos de confinamiento en transporte hasta plantel en Ejido Eréndira, 100 vs 1000
km), menor estrés en periodo de acondicionamiento (no hubo sexado inmediato ni inducción hormonal)
y mejores condiciones generales de cultivo, dado que la cantidad de alimento, la competencia, la carga
parasitaria/bacteriana y los parámetros de calidad de agua se ven influenciados directamente por la
biomasa y densidad de organismos en un estanque de tamaño y volumen limitado.
El impacto del manejo de los reproductores en su maduración y desove ha sido documentado en otros
peces marinos, por ejemplo en el pez Centropomus undecimalis se observa una ausencia de actividad
reproductiva en peces que son sometidos mensualmente a la toma de biopsias por canulación y muestras
de sangre, con el objeto de monitorear el ciclo reproductivo a través del perfil de hormonas esteroideas;
mientras que en tanques donde no se realizó este proceso, se registraron 16 eventos de desove durante
la temporada reproductiva (Rhody et al., 2014). Asimismo, el manejo para la inducción hormonal,
mediante inyección o implantes en la musculatura del pez, puede tener efectos negativos en la

62
maduración y desove de algunas especies dado que puede suprimir la circulación de hormonas
reproductivas en el organismo (Barton, 2002).
Cabe señalar también que para el lote uno, se registraron un total de 29 desoves en la temporada 2014
(marzo-julio) y 35 desoves en 2015 (marzo-junio); mientras que en el lote dos, se registraron sólo 22
eventos de desove durante 2015 (enero-junio). Las puestas de ambos lotes variaron entre 50 - 370 mil
huevos a lo largo de la temporada de desove, y se registraron desoves cada 2-5 días, como ha sido
reportado con anterioridad para esta especie (Moran et al. , 2007, Smith et al., 2016). Resultados similares
se han documentado para reproductores de S. dorsalis (lalandi) capturados al sur de California, donde los
periodos de desove natural se dán de marzo a septiembre en organismos mantenidos en condiciones
ambientales y fotoperiodos naturales (Smith et al., 2016). Durante las temporadas 2013 y 2014, Smith et
al. (2016) monitorearon 130 eventos de desove con 19 (2013) y 37 (2014) reproductores presentes, en
proporciones 1:1. Sus resultados muestran que una sóla hembra, contribuyó con el 40% de toda la
progenie, desovando cada 5-6 días durante ambas temporadas. Mientras que la contribución de los padres
fue relativamente similar durante las temporadas y eventos de desove. Resultados que también
concuerdan con las observaciones sobre la estrategia reproductiva de S. lalandi en cultivo en Australia,
donde el número de hembras que participan en los desoves es menor (14 al 67% en cuatro estanques
independientes), respecto al número de machos (80-100%, Knibb, et al., 2016).
El porcentaje de asignación parental total, considerando los dos lotes de reproductores y los seis desoves
evaluados, fue del 70% con una confiabilidad superior al 95% considerando los nuevos loci microsatélites.
El 30% de las muestras no pudieron ser asignadas a una pareja de reproductores. Whatmore et al., (2013),
utilizando estos mismos marcadores, más otro loci heterólogo (un total de 10 loci), fue capaz de asignar el
84% con una confiabilidad mayor al 95% y 8% con una confiabilidad entre 80 y 95%. Las diferencias entre
estos resultados pueden deberse al uso de un marcador más en este estudio, así como al control de calidad
de la asignación de genotipos realizada en una progenie específica de 90 crías provenientes de un número
conocido de reproductores, donde el 98% de los animales fue correctamente asignado a su grupo parental
conocido. Sin embargo, este resultado de asignación parental fue mayor al reportado en el salmón de río
Brycon orbignyanus, en donde se obtuvo un porcentaje de asignación de solo el 50% derivado quizás del
bajo número de alelos por el uso de cuatro loci microsatélites heterólogos (Lopera-Barrero et al., 2010).
La dificultad de asignar el 100% de la progenie en los estudios, se puede atribuir principalmente a errores
en el genotipado, ya sea por falta de producto amplificado o errores en la asignación de alelos, por lo cual
se recomienda que el genotipificado se realice dos veces de forma independiente en las muestras

63
tomadas, o bien que se cuente con un método de control de calidad de genotipificado en cruzas dirigidas;
sin embargo, el costo de los estudios aumenta significativamente y para muchas especies aún no se cuenta
con un sistema de cruzamiento para la obtención de alevines. En el caso de especies de peces marinos de
gran talla, como con las especies de Seriola spp, que además son promiscuos, es complicado tener
estanques con parejas seleccionadas. Sin embargo, una alternativa podría ser usar un número reducido de
reproductores con genotipos diferentes conocidos.
Por otra parte, en el nuevo contexto de que la especie de Seriola que se tiene en aguas mexicanas es
dorsalis y no lalandi, y las diferencias genéticas ya reportadas entre ellas, los marcadores desarrollados
por Whatmore et al., (2013) utilizados aquí deben considerarse heterólogos y no especie-específicos. En
el caso de los lotes de reproductores, para cinco de ellos no fue posible obtener su genotipo con el
marcador Sdu21, que significa un 2.5% de datos perdidos. Mientras que para los lotes F1BT se tuvieron 30
genotipos faltantes en marcadores Sel017 (1), Sel019 (7) y Sdu21 (22) y para el lote de crías F1BM se tuvo
un total de 82 genotipos faltantes en marcadores Sel001 (1), Sel017 (3), Sel019 (21) y Sdu21 (53); lo que
representa un 2.6 y un 8.2% de datos pérdidos, respectivamente. Aunado a esta pérdida de información,
también se reporta la posible presencia de alelos nulos (máximo de 7.2 y 7.9% en F1BT y F1BM). Además,
en el caso del lote dos de reproductores no fue posible obtener muestras de todos los organismos
presentes en el estanque. Tomando todo esto en consideración, el 30% de la progenie que no pudo ser
asignada responde a esta pérdida de información, y por lo tanto, debe considerarse desarrollar nuevos
marcadores microsatélites para el análisis parental en esta especie.
Sin embargo, considerando el 70% de asignación parental es factible rechazar la hipótesis planteada al
inicio del proyecto, sobre la contribución homogénea del jurel en cautiverio.
4.4.1 Relación entre el fenotipo y la contribución parental
Los reproductores con una talla, en téminos de longitud estándar, mayor a 79 cm contribuyeron con el
99% de la descendencia, mientras que el 1% restante se les atribuyó a individuos de menor tamaño. Estos
resultados sugieren, que los individuos menores a 79 cm no estaban maduros sexualmente. Comparando
con Poortenaar et al. (2001) en un estudio sobre la fisiología reproductiva en S. lalandi, las longitudes
furcales mínimas en las que se observó madurez sexual fueron 77.5 cm y 75 cm en hembras y machos,
respectivamente, por lo que las medidas obtenidas en el presente estudio fueron mayores. En el mismo

64
trabajo se observó que el 100% de los individuos alcanzaron la madurez reproductiva a los 127.5 cm en
hembras y 92.5 cm en machos.
Smith et al., (2016) concluyen que los reproductores más chicos, desovan menos veces que los grandes
durante la temporada reproductiva, y que una hembra más grande (21 kg) tiene una mayor fecundidad (
490 mil ovocitos por desove y alrededor de 23 millones de ovocitos/año) con respecto a la hembra más
chica (8.4 kg y 35 mil ovocitos por desove), y es por consecuencia la hembra más exitosa y que mas
aporta a la progenie total.
En este estudio, el lote uno de reproductores, evaluados una vez terminada su primer temporada de
desove en cautiverio (julio de 2014), presentó un peso de entre 8-17 kg en machos y 7-16 kg en hembras;
mientras que, en el lote dos, evaluado antes del inicio de la temporada de desoves (enero/2015), los
machos presentaron un peso promedio de 8-20 kg y las hembras 4-24 kg. No se observó una relación entre
el peso y la contribución parental total de los padres. El aporte parental de los reproductores varió de
acuerdo al desove, por lo que para la determinación del reproductor más exitoso se consideró el aporte
total en relación al número de crías. Los padres que contribuyeron con un mayor número de descendientes
en el lote 1 (M5/H1; Fig. 22) y en el lote 2 (M4/H4; Fig. 23) tuvieron un peso de entre 10 y 12 kgs. lo que
indica que no todos los reproductores presentaban un desarrollo gonadal adecuado para desovar y la
mayoría de las hembras se encontraban inmaduras.
Knibb et al. (2016), señala en su estudio, que de 54 reproductores disponibles, un máximo de 37 (69%)
contribuyen a la progenie, debido a que el resto se encuentran inmaduros. Asimismo señala que de 30
machos, 27 desovan y de 24 hembras, solo 10 lo hacen, por lo que menciona que existen relativamente
más machos maduros que hembras. Esto concuerda con lo puntualizado por el trabajo de Poortenaar y
colaboradores (2001) donde se evidencia que los machos alcanzan la madurez reproductiva en una menor
talla que las hembras.
4.4.2 Deformidades en el cultivo de S. dorsalis
Las deformidades están determinadas por la interacción entre factores ambientales y genéticos (Nguyen
et al., 2016). Algunos estudios sugieren que en los peces, la presencia de deformidades puede estar
asociada a una reducción en la variabilidad genética, al grado de endogamia entre progenitores y podrían
estar ligadas a una característica de interés económico (Tiira et al., 2006; Whatmore et al., 2013; Nguyen

65
et al., 2016). En el cultivo de S. lalandi las deformidades craneales (mandíbulares, operculares) son las de
mayor prevalencia (Cobcroft y Battaglene, 2013; Cobcroft et al., 2004).
Por lo anterior, en la presente sección, se exploró la posibilidadad de que las deformidades craneales se
relacionaran con el cruzamiento entre reproductores emparentados en el cultivo de S. dorsalis. En este
trabajo no se observó un patrón determinante en relación a los reproductores que produjeron larvas
deformes. El progenitor con mayor contribución de larvas normales fue el mismo que presentó una mayor
proporción de larvas deformes, por lo que los resultados no son concluyentes (Fig. 26). Por otra parte, no
se encontraron diferencias significativas en el coeficiente de endogamia entre ambos grupos así como
tampoco, en el resto de los parámetros de la diversidad genética, de los organismos deformes y sin
deformidades craneales (Fig. 27).
Nguyen et al. (2016) determinaron que las características como el crecimiento y el rendimiento de carne
en S. lalandi están correlacionados genéticamente con las deformidades mandibulares, esto significa que
a mayor crecimiento mayor porcentaje de deformidades. Por lo anterior, sugirieron considerarlas en los
planes de selección genética, aunque el porcentaje heredable en deformidades mandibulares sea del 16%.
En relación a los hallazgos observados en esta tesis, la endogamia no fue un factor determinante en la
aparición de las deformidades craneales. Lo anterior difiere a lo propuesto por Nguyen et al., (2016)
quienes reportaron variaciones en las 22 familias de hermanos completos y medios hermanos de S. lalandi
empleados en su estudio, que alcanzan hasta el 63% en el porcentaje de deformidades craneales
(mandíbula, opérculo, erosión nasal). Respecto a la diversidad genética Tiira et al., (2006) observaron una
menor variabilidad genética en larvas deformes (espiral en espina dorsal, presencia de dos cabezas) en
Salmón del atlántico (Salmo salar), así como una disminución significativa en la heterocigosidad. A partir
de esos resultados concluyeron que debido a su baja variabilidad genética Salmo salar presenta una mayor
susceptibilidad a la endogamia, y por ende, exhibe mayor probabilidad de presentar deformidades óseas.
A pesar de que los factores ambientales tienen un mayor efecto en la incidencia de deformidades, el
desarrollo diferencial de estas entre individuos, aún cuando las condiciones ambientales son las mismas,
sugiere la existencia de un factor genético asociado a ellas. Un análisis transcriptómico entre larvas
deformes y normales sometidas a las mismas condiciones experimentales podría sugerir genes asociados
a su desarrollo o regeneración. Por lo que se deben realizar nuevos análisis en el área.

66
Capítulo 5. Conclusiones
La diversidad genética de las poblaciones silvestres mexicanas es alta. Sin embargo, la perdida de
variabilidad en la primera generación en cautiverio de Bahía de Todos Santos y de Bahía Magdalena fue
del 23 y 46%, por lo que se debe implementar un manejo reproductivo adecuado.
No se encontró estructura genética significativa entre las poblaciones mexicanas, sin embargo, se debe
considerar la captura de nuevos reproductores de las localidades del Ejido Eréndira o Bahía de los ángeles
con la finalidad de preservar en cautiverio la mayor parte de la diversidad genética silvestre.
Con base en los análisis con nueve microsatélites, las poblaciones de Bahía de Todos Santos, Éjido Eréndira,
Bahía Magdalena, Bahía de los Ángeles, y California, Estados Unidos en el hemisferio norte presentaron
un alto grado de diferenciación genética en comparación con las poblaciones de Nueva Gales del Sur, AUS
y Antofagasta, CHL en el hemisferio sur, sugiriendo la presencia de dos especies distintas, de acuerdo al
concepto filogenético de especie. Dada la importancia en la conservación de las especies locales, la
población del hemisferio sur debe mantener el nombre de Seriola lalandi, mientras que la del Noreste del
Pacífico debe ser renombrada a: Seriola dorsalis como anteriormente fue descrita. Debiéndose restringir
la translocación de la población no solo del Hemisferio sur, sino también de la especie japonesa Seriola
aureovitata, por el riesgo ecológico y en la erosión genética que representan para la especie local.
La contribución parental del jurel en cautiverio es desigual. El éxito reproductivo de las hembras es inferior
al de los machos, sin embargo, las hembras contribuyen en mayor proporción a la descendencia, por lo
que el número de machos que se utilizan en los laboratorios se puede reducir. Bajo estas condiciones, las
hembras juegan un papel determinante en la conservación de la diversidad genética en cautiverio, que
debería considerarse en el manejo reproductivo futuro.
No se encontraron evidencias que relacionen la diversidad genética con deformidades, por lo que se deben
realizar nuevas aproximaciones en relación con esta problemática.

67
Literatura citada
Abbink, W., Blanco Garcia, A., Roques, J. A. C., Partridge, G. J., Kloet, K., & Schneider, O. (2012). The effect of temperature and pH on the growth and physiological response of juvenile yellowtail kingfish Seriola lalandi in recirculating aquaculture systems. Aquaculture, 330-333, 130–135. http://doi.org/10.1016/j.aquaculture.2011.11.043
Aljanabi, S. M., y Martinez, I. (1997). Universal and rapid salt-extraction of high quality genomic DNA for PCR-based techniques. Nucleic Acids Research, 25(22), 4692–4693. http://doi.org/10.1093/nar/25.22.4692
Allendorf, F. W. (1991). Ecological and Genetic Effects of Fish Introductions: Synthesis and Recommendations. Canadian Journal of Fisheries and Aquatic Sciences, 48(1), 178–181.
Aranguren-Méndez, J. A., Román-Bravo, R., Isea, W., Villasmil, Y., & Jordana, J. (2005). Los microsatélites (STR´ s), marcadores moleculares de ADN por excelencia para programas de conservación: una revisión (Microsatellites (STR’s), ADN Molecular Markers for Excellency for conservation programs: A review).
Avilés, A., & Castelló, F. (2004). Manual para el Cultivo de Seriola lalandi (Pisces: Carangidae) en Baja California Sur, México. (J. P. Cuellar & C. O. Cadena, Eds.) (1a ed.). Mexico: Instituto Nacional de la Pesca (INAPESCA).
Bailly, N. (2015). Seriola lalandi Valenciennes, 1833. Retrieved October 23, 2016, from
http://www.marinespecies.org/aphia.php?p=taxdetails&id=218436
Barber, I., & Wright, H. A. (2001).How strong are familiarity preferences in shoaling fish?. Animal Behaviour. 61(5). Doi: 10.1006/anbe.2000.1665
Barton, B. A. (2002). Stress in fishes: a diversity of responses with particular reference to changes in circulating corticosteroids. Integrative and comparative biology, 42(3), 517-525.
Baxter, J. L. (1960). A Study of The Yellowtail Seriola Dorsalis (Gill). Calif. Dept. Fish Game, Fish. Bull. l10:1-91.
Brody, T., Wohlfarth, G., Hulata, G., & Moav, R. (1981). Application of electrophoretic genetic markers to fish breeding. Aquaculture, 24, 175–186. http://doi.org/10.1016/0044-8486(81)90054-5
Chai, X., Li, X., Lu, R., & Clarke, S. (2009). Karyotype analysis of the yellowtail kingfish Seriola lalandi lalandi (Perciformes: Carangidae) from South Australia. Aquaculture Research, 40(15), 1735–1741. http://doi.org/10.1111/j.1365-2109.2009.02278.x
Charles, A. T., & Leith, J. (1999). Social and Economic Approaches to Biodiversity Conservation: An Annoted Bibliography with Emphasis on Aquaculture.
Chauhan, T., & Rajiv, K. (2010). Molecular markers and their applications in fisheries and aquaculture. Advances in Bioscience and Biotechnology, 01(04), 281–291. http://doi.org/10.4236/abb.2010.14037

68
Cipriano, R. R., Noleto, R. B., Kantek, D. L. Z., Da Silva Cortinhas, M. C., & Cestari, M. M. (2014). Comparative cytogenetic analysis of marine needlefishes (Beloniformes) from southern Brazil. Cytotechnology. http://doi.org/10.1007/s10616-014-9809-x
Cobcroft, J. M., & Battaglene, S. C. (2013). Skeletal malformations in Australian marine finfish hatcheries. Aquaculture, 396-399(2013), 51–58. http://doi.org/10.1016/j.aquaculture.2013.02.027
Cobcroft, J. M., Pankhurst, P. M., Poortenaar, C., Hickman, B., & Tait, M. (2004). Jaw malformation in cultured yellowtail kingfish (Seriola lalandi) larvae. New Zealand Journal of Marine and Freshwater Research, 38(1), 67–71. http://doi.org/10.1080/00288330.2004.9517218
Collins, J. E., White, S., Searle, S. M. J., & Stemple, D. L. (2012). Incorporating RNA-seq data into the Zebrafish Ensembl genebuild. Genome Research, 2067–2078. http://doi.org/10.1101/gr.137901.112
CONAPESCA, C. N. de A. y P. (2013). Anuario estadístico de Acuacultura y Pesca 2013. Mazatlán, Sinaloa, México.
Coombs, J. A., Letcher, B. H., & Nislow, K. H. (2008). CREATE: a software to create input files from diploid genotypic data for 52 genetic software programs. Molecular Ecology Resources, 8(3), 578–580. http://doi.org/10.1007/SpringerReference_38033
CROOKE, S. J. (1983). YELLOWTAIL, SERlOLA LALANDEI VALENCIENNES. CalCOFI Rep., Vol. XXIV.
Dekkers, J. C.M., Gibson, J. Bijma, P., Van Arendonk, J. A.M. (2004). Design and optimisation of animal breeding programmes. Iowa State University.
Domínguez-López, M., Uribe-Alcocer, M., Díaz-Jaimes, Píndaro. (2010). Phylogeography and historical demography of the Pacific Sierra mackerel (Scomberomorus sierra) in the Eastern Pacific. BMC genetics.
Earl, D. A., & vonHoldt, B. M. (2012). STRUCTURE HARVESTER: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conservation Genetics Resources, 4(2), 359–361. http://doi.org/10.1007/s12686-011-9548-7
Evanno, G., Regnaut, S., & Goudet, J. (2005). Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Molecular Ecology, 14(8), 2611–2620. http://doi.org/10.1111/j.1365-294X.2005.02553.x
Excoffier, L., Laval, G., & Schneider, S. (2005). Arlequin (version 3.0): an integrated software package for population genetics data analysis. Evolutionary Bioinformatics Online, 1(47).
Falconer, D. S., & Mackay., T. F. C. (1996). Introducción a la genética cuantitativa (4th ed.). Zaragoza, España: ACRIBIA S.A.
FAO. (1995). Code of Conduct for Responsible Fisheries. Food and Agriculture Organization of the United Nations.
FAO. (1998). FAO species identification guide for fishery purposes.The livingmarine resources of theWestern Central Pacific. K. E. Carpenter & V. H. Niem, Eds.Vol. 2. Rome.
FAO. (2016). The State of World Fisheries and Aquaculture. Rome. Retrieved from http://www.fao.org/3/a-i5555e.pdf

69
FAO/UNEP. (1981). Conservation of the genetic resources of fish: problems and recommendations. Report of the Expert Consultation on the genetic resources of fish. Rome.
Fernández, G., Cichero, D., Patel, A., & Martínez, V. (2015). Genetic structure of Chilean population of Seriola lalandi Genetic structure of Chilean populations of Seriola lalandi for the diversification of the national aquaculture in the north of Chile. J. Aquat. Res, 43(2), 374–379. http://doi.org/10.3856/vol43-issue2-fulltext-14
Frankham, R., Ballou, J. D., & Briscoe, D. A. (2002). Introduction to Conservation Genetics. Forest Ecology and Management, 617. http://doi.org/10.1016/j.foreco.2003.12.001
Gill, T. N. (1863). Catalogue of the fishes of Lower California, in the Smithsonian Institution, collected by Mr. J. Xantus. Part IV. In Proceedings of the Academy of Natural Sciences of Philadelphia (pp. 15:80–88). Retrieved from http://www.biodiversitylibrary.org/page/1958344#page/90/mode/1up
Gillanders, B. M., Ferrell, D. J., & Andrew, N. L. (1999). Aging methods for yellowtail kingfish, Seriola lalandi, and results from age- and size-based growth models. Fishery Bulletin, 97(4), 812–827.
Gillanders, B. M., Ferrell, D. J., & Andrew, N. L. (1999). Size at maturity and seasonal changes in gonad activity of yellowtail kingfish (Seriola lalandi; Carangidae) in New South Wales, Australia. New Zealand Journal of Marine and Freshwater Research, 33(3), 457–468. http://doi.org/10.1080/00288330.1999.9516891
Gillanders, B. M., Ferrell, D. J., & Andrew, N. L. (2001). Estimates of movement and life-history parameters of yellowtail kingfish (Seriola lalandi): How useful are data from a cooperative tagging programme? Marine and Freshwater Research, 52(2), 179–192. http://doi.org/10.1071/MF99153
Gjedrem, T. (2005). Selection and Breeding Programs in Aquaculture. In New York (pp. 1–364). http://doi.org/10.1111/j.1365-2109.2005.01440.x
Gjedrem, T. (2012). Genetic improvement for the development of efficient global aquaculture: A personal opinion review. Aquaculture, 344-349(0044), 12–22. http://doi.org/10.1016/j.aquaculture.2012.03.003
Gjedrem, T., & Baranski, M. (2009). Selective Breeding in Aquaculture: An Introduction, Reviews: Methods and Technologies in Fish Biology and Fisheries 10 (Vol. 10). Norway: Springer Science+Business Media B.V. http://doi.org/10.1007/978-90-481-2773-3_1
Gjedrem, T., & Kolstad, K. (2012). Development of Breeding Programs for Aquatic Species Should be Given High Priority. World Aquaculture Magazine, (September), 10–13.
Goldberg, T. L., Grant, E. C., Inendino, K. R., Kassler, T. W., Claussen, J. E., & Philipp, D. P. (2005). from Outbreeding Depression. Conservation Biology, 19(2), 455–462. http://doi.org/10.1111/j.1523-1739.2005.00091.x
Goudet, J. (1995). FSTAT (version 1.2): a computer program to calculate F-statistics. Journal of Heredity, 86(6), 485–486.
Graham, C. M. (2009). Metodologías de mejoramiento genético en la acuicultura. (FAO) (Ed.), Desarrollo de la acuicultura. Gestion de los recursos geneticos (pp. 34–59).

70
Herbinger, C. ., Doyle, R. ., Pitman, E. ., Paquet, D., Mesa, K. ., & Morris, D. B. (1995). DNA fingerprint based analysis of paternal and maternal effects on offspring growth and survival in communally reared rainbow trout. Aquaculture, 137, 245–256. http://doi.org/10.1016/0044-8486(95)01109-9
Herlin, M., Delghandi, M., Wesmajervi, M., Taggart, J. B., McAndrew, B. J., & Penman, D. J. (2008). Analysis of the parental contribution to a group of fry from a single day of spawning from a commercial Atlantic cod (Gadus morhua) breeding tank. Aquaculture, 274(2-4), 218–224. http://doi.org/10.1016/j.aquaculture.2007.11.034
Houde, A. L. S., Fraser, D. J., O’Reilly, P., & Hutchings, J. A. (2011). Relative risks of inbreeding and outbreeding depression in the wild in endangered salmon. Evolutionary Applications, 4(5), 634–647. http://doi.org/10.1111/j.1752-4571.2011.00186.x
ICES, International Council for the Exploration of the Sea. (2005). ICES Code of Practice on the Introductions and Transfers of Marine Organisms 2005.
IUCN, I. U. for C. of N., UNEP, U. N. E. P., & WWF, W. W. F. (1980). World Conservation Strategy. World Conservation Strategy: Living Resource Conservation for Sustainable Development. http://doi.org/10.2305/IUCN.CH.1980.9
Jombart, T. (2008). Adegenet: A R package for the multivariate analysis of genetic markers. Bioinformatics, 24(11), 1403–1405. http://doi.org/10.1093/bioinformatics/btn129
Jombart, T., Devillard, S., Balloux, F., Falush, D., Stephens, M., Pritchard, J., … Nei, M. (2010). Discriminant analysis of principal components: a new method for the analysis of genetically structured populations. BMC Genetics, 11(1), 94. http://doi.org/10.1186/1471-2156-11-94
Jones, O. R., & Wang, J. (2010). COLONY: a program for parentage and sibship inference from multilocus genotype data. Molecular Ecology Resources, 10(3), 551–555.
Kalinowski, S. T., & Taper, M. L. (2006). Maximum likelihood estimation of the frequency of null alleles at microsatellite loci. Conservation Genetics, 7(6), 991–995. http://doi.org/10.1007/s10592-006-9134-9
Kalinowski, S. T., Taper, M. L., & Marshall, T. C. (2007). Revising how the computer program CERVUS accommodates genotyping error increases success in paternity assignment. Molecular Ecology, 16(5), 1099–1106.
Kapuscinski., A. R. (2008). Risk assessment and monitoring in genetic improvement programmes. In F. and A. O. FAO (Ed.), Aquaculture development. 3. Genetic resource management (p. 125). Rome. http://doi.org/10.1017/CBO9781107415324.004
Knibb, W. (2000). Genetic improvement of marine fish - which method for industry ? Aquaculture Research, 31, 11–23. http://doi.org/10.1046/j.1365-2109.2000.00393.x
Knibb, W., Miller, A., Quinn, J., D'Antignana, T., Nguyen, N. H. (2016). Comparison of lines shows selection response in kingfish (Seriola lalandi). Aquaculture. Doi: 10.1016/j.aquaculture.2015.11.015
Kolkovski, S., & Sakakura, Y. (2004). Yellowtail kingfish , from larvae to mature fish – problems and opportunities. Avances en Nutrición Acuícola VII. Hermosillo, Sonora, México.

71
Lind, C. E., Ponzoni, R. W., Nguyen, N. H., & Khaw, H. L. (2012). Selective breeding in fish and conservation of genetic resources for aquaculture. Reproduction in Domestic Animals, 47(SUPPL.4), 255–263. http://doi.org/10.1111/j.1439-0531.2012.02084.x
Liu, Z. J., & Cordes, J. F. (2004). DNA marker technologies and their applications in aquaculture genetics. Aquaculture, 238(1-4), 1–37. http://doi.org/10.1016/j.aquaculture.2004.05.027
Lopera-Barrero, N. M., Vargas, L., Nardez-Sirol, R., Pereira-Ribeiro, R., Aparecido-Povh, J., Streit Jr, D. P., & Cristina-Gomes, P. (2010). Genetic Diversity and Reproductive Contribution of Brycon orbignyanus Offspring in the semi-natural reproductive system, using microsatellites markers. Agrociencia, 44, 171–181.
Luening, E. (2014). Mexican yellowtail farm debuts first harvest at Boston seafood show an Interview with Blue Goose turns its focus to fish Distributing Yellowtail. Aquaculture North America, 5(3), 11–12.
Martinez-Takeshita, N., Purcell, C. M., Chabot, C. L., Craig, M. T., Paterson, C. N., Hyde, J. R., & Allen, L. G. (2015). A Tale of Three Tails: Cryptic Speciation in a Globally Distributed Marine Fish of the Genus Seriola. Copeia, 103(2), 357–368. http://doi.org/10.1643/CI-124-224
McDonald, G. J., Danzmann, R. G., & Ferguson, M. M. (2004). Relatedness determination in the absence of pedigree information in three cultured strains of rainbow trout (Oncorhynchus mykiss). Aquaculture, 233(1-4), 65–78. http://doi.org/10.1016/j.aquaculture.2003.08.003
Miller, P. A., Fitch, A. J., Gardner, M., Hutson, K. S., & Mair, G. (2011). Genetic population structure of Yellowtail Kingfish (Seriola lalandi) in temperate Australasian waters inferred from microsatellite markers and mitochondrial DNA. Aquaculture, 319(3-4), 328–336. http://doi.org/10.1016/j.aquaculture.2011.05.036
Moran, D., Smith, C. K., Gara, B., & Poortenaar, C. W. (2007). Reproductive behaviour and early development in yellowtail kingfish (Seriola lalandi Valenciennes 1833). Aquaculture, 262(1), 95-104.
Nakada, M. (2002). Yellowtail culture development and solutions for the future. Reviews in Fisheries Science, 10(3-4), 559–575. http://doi.org/10.1080/20026491051794
Nakada, M. (2008). Capture-based aquaculture of yellowtail. In A. Lovatelli & P. . Holthus (Eds.), Capture-Based Aquaculture. Global Overview. (Vol. No. 508, pp. 199–215). Rome: FAO Fisheries Technical Paper.
Nguyen, N. H., Whatmore, P., Miller, A., & Knibb, W. (2016). Quantitative genetic properties of four measures of deformity in yellowtail kingfish Seriola lalandi Valenciennes, 1833. Journal of Fish Diseases, 39(2), 217–228. http://doi.org/10.1111/jfd.12348
Nirchio, M., & Oliveira, C. (2014). Artículo de revisión agrobiología citogenética como herramienta taxonómica en peces. Universidad de Oriente, 26(4), 361–372.
Norris, A. ., Bradley, D. ., & Cunningham, E. . (1999). Microsatellite genetic variation between and within farmed and wild Atlantic salmon (Salmo salar) populations. Aquaculture, 180(3-4), 247–264. http://doi.org/10.1016/S0044-8486(99)00212-4
Norris, A. T., Bradley, D. G., & Cunningham, E. P. (2000). Parentage and relatedness determination in farmed Atlantic salmon (Salmo salar) using microsatellite markers. Aquaculture, 182, 73–83.

72
Nugroho, E., Ferrell, D. J., Smith, P., & Taniguchi, N. (2001). Genetic divergence of kingfish from Japan , Australia and New Zealand inferred by microsatellite DNA and mitochondrial DNA control region markers. Fisheries Science, 67, 843–850.
Oosterhout, C., Hutchinson, W. F., Wills, D. P., & Shipley, P. (2004). MICROCHECKER: software for identifying and correcting genotyping errors in microsatellite data. Molecular Ecology Notes, 4(3), 535–538.
Orellana, J., Waller, U., & Wecker, B. (2014). Culture of yellowtail kingfish (Seriola lalandi) in a marine recirculating aquaculture system (RAS) with artificial seawater. Aquacultural Engineering, 58, 20–28. http://doi.org/10.1016/j.aquaeng.2013.09.004
Ottolenghi, F., Silvestri, C., Giordano, P., Lovatelli, A., & B.New, M. (2004b). Capture-based aquaculture. The fattening of eels, groupers, tunas and yellowtails. Retrieved from http://www.fishbase.org/summary/Seriola-lalandi.html
Peakall, R., & Smouse, P. E. (2006). GENALEX 6: Genetic analysis in Excel. Population genetic software for teaching and research. Molecular Ecology Notes, 6(1), 288–295. http://doi.org/10.1111/j.1471-8286.2005.01155.x
Peakall, R., & Smouse, P. E. (2012). GenALEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research-an update. Bioinformatics, 28(19), 2537–2539. http://doi.org/10.1093/bioinformatics/bts460
Perazzo, G., Noleto, R. B., Vicari, M. R., Machado, P. C., Gava, A., & Cestari, M. M. (2011). Chromosomal studies in Crenicichla lepidota and Australoheros facetus (Cichlidae, Perciformes) from extreme Southern Brazil. Reviews in Fish Biology and Fisheries, 21(3), 509–515. http://doi.org/10.1007/s11160-010-9170-x
Poortenaar, C. W., Hooker, S. H., & Sharp, N. (2001). Assessment of yellowtail kingfish (Seriola lalandi lalandi) reproductive physiology, as a basis for aquaculture development. Aquaculture, (201), 271–286.
Porta, J., Porta, J. M., Martínez-Rodríguez, G., & Alvarez, M. C. (2006). Genetic structure and genetic relatedness of a hatchery stock of Senegal sole (Solea senegalensis) inferred by microsatellites. Aquaculture, 251(1), 46–55. http://doi.org/10.1016/j.aquaculture.2005.05.019
Price, E. O. (1984). Behavioral Aspects of Animal Domestication. The Quarterly Review of Biology. Accessed: http://www.jstor.org/stable/2827868 .
Pritchard, J. K., Stephens, M., & Donnelly, P. (2000). Inference of population structure using multilocus genotype data. Genetics, 155(2), 945–959. http://doi.org/10.1111/j.1471-8286.2007.01758.x
Purcell, C. M., Chabot, C. L., Craig, M. T., Larry, N. M., & John, G. A. (2015). Developing a genetic baseline for the yellowtail amberjack species complex , Seriola lalandi sensu lato , to assess and preserve variation in wild populations of these globally important aquaculture species. Conservation Genetics, 16(6), 1475–1488. http://doi.org/10.1007/s10592-015-0755-8
Refstie, T., & Gjedrem, T. (1975). Hybrids between Salmonidae species. Hatchability and growth rate in the freshwater period. Aquaculture, 6(4), 333–342. http://doi.org/10.1016/0044-8486(75)90112-X

73
Renshaw, M. a, Patton, J. C., Rexroad, C. E., & Gold, J. R. (2006). PCR primers for trinucleotide and tetranucleotide microsatellites in greater amberjack, Seriola dumerili. Molecular Ecology Notes, 6(4), 1162–1164. http://doi.org/10.1111/j.1471-8286.2006.01474.x
Renshaw, M. a., Patton, J. C., Rexroad, C. E., & Gold, J. R. (2007). Isolation and characterization of dinucleotide microsatellites in greater amberjack, Seriola dumerili. Conservation Genetics, 8(4), 1009–1011. http://doi.org/10.1007/s10592-006-9221-y
Rhody, N. R., Puchulutegui, C., Taggart, J. B., Main, K. L., & Migaud, H. (2014). Parental contribution and spawning performance in captive common snook Centropomus undecimalis broodstock. Aquaculture, 432, 144-153
Rice, W. E. R. (1989). Analyzing tables of statistical tests. Evolution. http://doi.org/10.2307/2409177
Rimmer, M.A. and Ponia, B. (2007). A review of cage aquaculture: Oceania. Cage aquaculture – Regional reviews and global overview, pp. 208–231. FAO Fisheries Technical Paper. No. 498. Rome, FAO. 2007. 241 pp.
SAGARPA. (2012). Carta nacional acuícola. México.
Sakakura, Y., & Tsukamoto, K. (1998). Social rank in schools of juvenile yellowtail, Seriola quinqueradiata. Journal of Applied Ichthyology, 14(1-2), 69–73. Retrieved from http://hdl.handle.net/10069/35745
Sala, E., Aburto-Oropeza, O., Paredes, G., & Thompson, G. (2003). Spawning aggregations and reproductive behavior of reef fishes in the Gulf of California. Bulletin of Marine Science, 72(1), 103–121. http://doi.org/10.1126/science.1075284
Savolainen, V., Powell, M. P., Davis, K., Reeves, G., & Corthals, A. (2006). DNA and Tissue Banking for Biodiversity and Conservation Theory, Practice and Uses. Kew Publishing.
Scarratt, D. (2014). First spawnings for new hatchery in Mexico. Hatchery International. The Magazine for Fish Managers, 15(4), 11–13. Retrieved from http://hatcheryinternational.com/Profiles/first-spawnings-for-new-yellowtail-hatchery-in-mexico/
Sekino, M., Sugaya, T., Hara, M., & Taniguchi, N. (2004). Relatedness inferred from microsatellite genotypes as a tool for broodstock management of Japanese flounder Paralichthys olivaceus. Aquaculture, 233(1-4), 163–172. http://doi.org/10.1016/j.aquaculture.2003.11.008
SEPESCABC, Secretaría de Pesca y Acuicultura de Baja California (2014a). Carta Estatal Acuicola de Baja California.
SEPESCABC, Secretaría de Pesca y Acuicultura de Baja California. (2014b). Carta Estatal Pesquera de Baja California. http://doi.org/10.1017/CBO9781107415324.004
Shiraishi, T. , Ohshimo, S. and Yukami, R.(2010). Age, growth and reproductive characteristics of gold striped amberjack in the waters off western Kyushu, Japan. New Zealand Journal of Marine and Freshwater Research. 44: 2, 117 — 127. DOI: 10.1080/00288330.2010.488787
Sicuro, B., & Luzzana, U. (2016). The State of Seriola spp. Other Than Yellowtail ( S. quinqueradiata ) Farming in the World. Reviews in Fisheries Science & Aquaculture, 24(4), 314–325. http://doi.org/10.1080/23308249.2016.1187583

74
Smith, Elizabeth K.; Drawbridge, Mark MS; Hyde, John PhD; and Morse, Geoffrey PhD. (2015).Evaluation of reproductive strategies in captive California yellowtail (Seriola dorsalis) using genetic parentage
analyses.Theses. Paper 6.http://digital.sandiego.edu/theses/6
Stuart, K. R., & Drawbridge, M. A. (2013). Captive spawning and larval rearing of California yellowtail (Seriola lalandi). Aquaculture Research, 44(5), 728–737. http://doi.org/10.1111/j.1365-2109.2011.03077.x
Sumida, B. Y., Moser, H. G., Ahlstrom, E. H., Cali-, S., & La, C. (1985). Descriptions of larvae of california yellowtail , seriola lalandi , and three other carangids from the eastern tropical pacific : chloroscombrus orquina , caranx caballus , and caranx sexfasciatus Larvae are described for four species of jacks , fami- four, XXVI, 139–159.
Swart, B. L. (2014). The evolutionary history of the genus Seriola and the phylogeography and genetic diversity of S . lalandi ( yellowtail ) across its distribution range. Stellenbosch University. Retrieved from http://scholar.sun.ac.za
Swart, B. L., von der Heyden, S., Bester-van der Merwe, A., & Roodt-Wilding, R. (2015). Molecular systematics and biogeography of the circumglobally distributed genus Seriola (Pisces: Carangidae). Molecular Phylogenetics and Evolution, 93, 274–280. http://doi.org/10.1016/j.ympev.2015.08.002
Swart, B., Bester-van der Merwe, A., Kerwath, S., & Roodt-Wilding, R. (2016). Phylogeography of the pelagic fish Seriola lalandi at different scales: confirmation of inter-ocean population structure and evaluation of southern African genetic diversity. African Journal of Marine Science, 2338(November), 1–12. http://doi.org/10.2989/1814232X.2016.1238410
Symonds, J., Walker, S., Pether, S., Gublin, Y., McQueen, D., King, a, … Bruce, M. (2014). Developing yellowtail kingfish ( Seriola lalandi ) and hāpuku ( Polyprion oxygeneios ) for New Zealand aquaculture. New Zealand Journal of Marine and Freshwater Research, 48(3), 371–384. http://doi.org/10.1080/00288330.2014.930050
Takezaki, N., Nei, M., & Tamura, K. (2010). POPTREE2: Software for constructing population trees from allele frequency data and computing other population statistics with windows interface. Molecular Biology and Evolution, 27(4), 747–752. http://doi.org/10.1093/molbev/msp312
Tautz, D. (1989). Hypervariability of simple sequences as a general source for polymorphic DNA markers. Nucleic Acids Research, 17(16), 6463–6471.
Temminck, C. J., & Schlegel, H. (1845). In: Temminck and Schlegel 1843. Fauna Japonica, sive description animalium quae in itinere per Japoniam. Retrieved from http://gallica.bnf.fr/ark:/12148/bpt6k970324/f117.item.zoom
Thorpe, J. E., and j. F. Koonce, D. Borgeson, B. Henderson, A. Lamsa, P. S. Maitland, M. A. Ross, R. C. Simon, and C. Walters. (1981). Assessing and managing man's impact on fish genetic resources. Can. .I. Fish. Aquat. Sci. 38: 1899- 1907.
Tiira, K., Piironen, J., & Primmer, C. R. (2006). Evidence for reduced genetic variation in severely deformed juvenile salmonids. Canadian Journal of Fisheries and Aquatic Sciences, 63(12), 2700–2707. http://doi.org/10.1139/f06-154

75
Toro, M. ., & Caballero, a. (2005). Characterization and conservation of genetic diversity in subdivided populations. Philosophical Transactions of the Royal Society B: Biological Sciences, 360(1459), 1367–1378. http://doi.org/10.1098/rstb.2005.1680
Valenciennes, A. (1833). Histoire naturelle des poissons. Tome neuvième. Suite du livre neuvième. Des Scombéroïdes. Histoire naturelle des poissons. (G. Cuvier & A. Valenciennes, Eds.). Retrieved from http://www.biodiversitylibrary.org/item/25879#page/245/mode/1up
Wang, J. (2011). Coancestry: A program for simulating, estimating and analysing relatedness and inbreeding coefficients. Molecular Ecology Resources, 11(1), 141–145. http://doi.org/10.1111/j.1755-0998.2010.02885.x
Wasko, A. P., Martins, C., Oliveira, C., & Foresti, F. (2003). Non-destructive genetic sampling in fish. An improved method for DNA extraction from fish fins and scales. Hereditas, 138(3), 161–165. http://doi.org/10.1034/j.1601-5223.2003.01503.x
Whatmore, P., Nguyen, N. H., Miller, A., Lamont, R., Powell, D., D’Antignana, T. Knibb, W. (2013). Genetic parameters for economically important traits in yellowtail kingfish Seriola lalandi. Aquaculture, 400-401, 77–84. http://doi.org/10.1016/j.aquaculture.2013.03.002
Worm, B., Barbier, E. B., Beaumont, N., Duffy, J. E., Folke, C., Halpern, B. S., Watson, R. (2006). Impacts of biodiversity loss on ocean ecosystem services. Science (New York, N.Y.), 314(5800), 787–90. http://doi.org/10.1126/science.1132294
Yang, S. G., Ji, S. C., Lim, S. G., Hur, S. W., & Jeong, M. (2016). Management of Sexual Maturation and Natural Spawning of Captive- Reared Yellowtail Kingfish , Seriola lalandi , in an Indoor Rearing Tank, 20(2), 141–147.
Yousefian, M., & Nejati, A. (2008). Inbreeding Depression by Family Matching in Rainbow Trout (Oncorhynchus mykiss). Journal of Fisheries and Aquatic Science, 3(6), 284–391.

76
Anexos
Anexo 1
Tabla 1. Relación de reproductores de Bahía de Todos Santos (lote 1).
Identificación Sexo Peso (kg) Longitud estándar (mm)
M1 Macho 11.7 92.5
M2 Macho 11.8 93
M3 Macho 10 91
M4 Macho 9 87
M5 Macho 10.3 91.5
M6 Macho 9.3 89
M7 Macho 9.8 90
M8 Macho 8.3 82
M9 Macho 9.6 89
**M10 Macho 8.6 99
H1 Hembra 11.2 91
H2 Hembra 9.6 90
H3 Hembra 9.6 88
H4 Hembra 8 84
H5 Hembra 8.4 85
H6 Hembra 4.2 75
H7 Hembra 8.7 89
H8 Hembra 10.1 91
**H9 Hembra 11.7 105 ** Individuos originarios de Bahía Magdalena transferidos a este lote el 27 de junio de 2014.
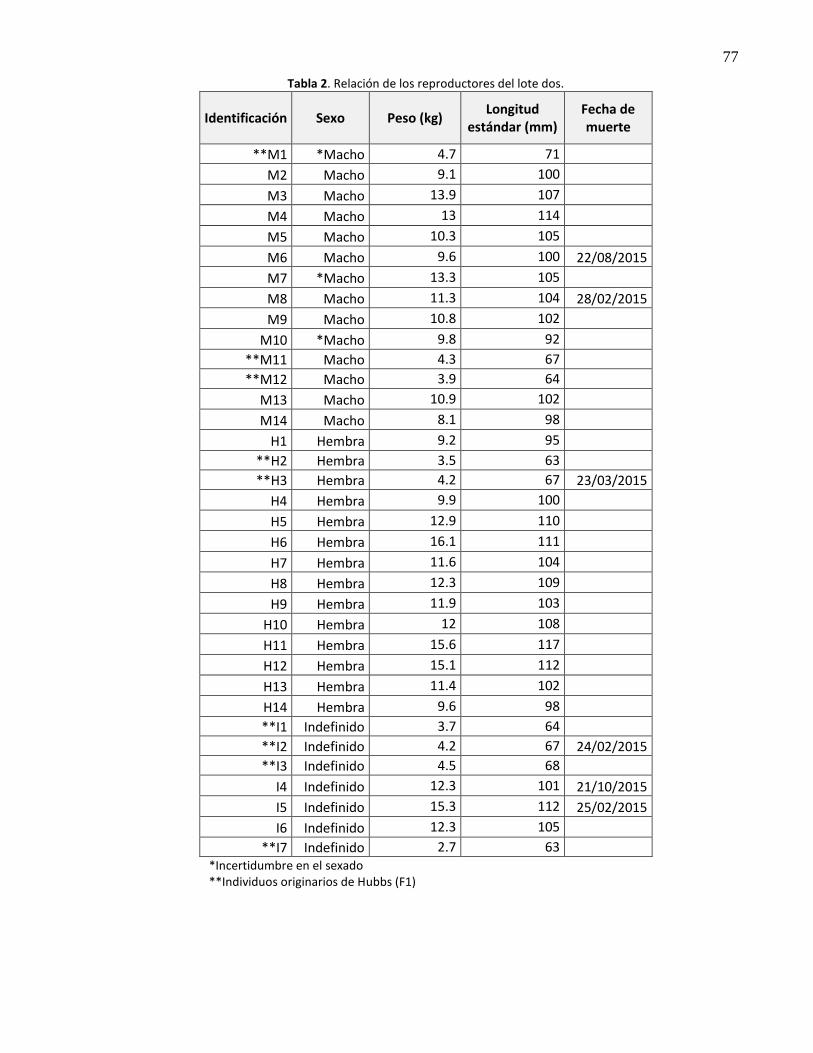
77
Tabla 2. Relación de los reproductores del lote dos.
Identificación Sexo Peso (kg) Longitud
estándar (mm) Fecha de muerte
**M1 *Macho 4.7 71
M2 Macho 9.1 100
M3 Macho 13.9 107
M4 Macho 13 114
M5 Macho 10.3 105
M6 Macho 9.6 100 22/08/2015
M7 *Macho 13.3 105
M8 Macho 11.3 104 28/02/2015
M9 Macho 10.8 102
M10 *Macho 9.8 92
**M11 Macho 4.3 67
**M12 Macho 3.9 64
M13 Macho 10.9 102
M14 Macho 8.1 98
H1 Hembra 9.2 95
**H2 Hembra 3.5 63
**H3 Hembra 4.2 67 23/03/2015
H4 Hembra 9.9 100
H5 Hembra 12.9 110
H6 Hembra 16.1 111
H7 Hembra 11.6 104
H8 Hembra 12.3 109
H9 Hembra 11.9 103
H10 Hembra 12 108
H11 Hembra 15.6 117
H12 Hembra 15.1 112
H13 Hembra 11.4 102
H14 Hembra 9.6 98
**I1 Indefinido 3.7 64
**I2 Indefinido 4.2 67 24/02/2015
**I3 Indefinido 4.5 68
I4 Indefinido 12.3 101 21/10/2015
I5 Indefinido 15.3 112 25/02/2015
I6 Indefinido 12.3 105
**I7 Indefinido 2.7 63 *Incertidumbre en el sexado **Individuos originarios de Hubbs (F1)
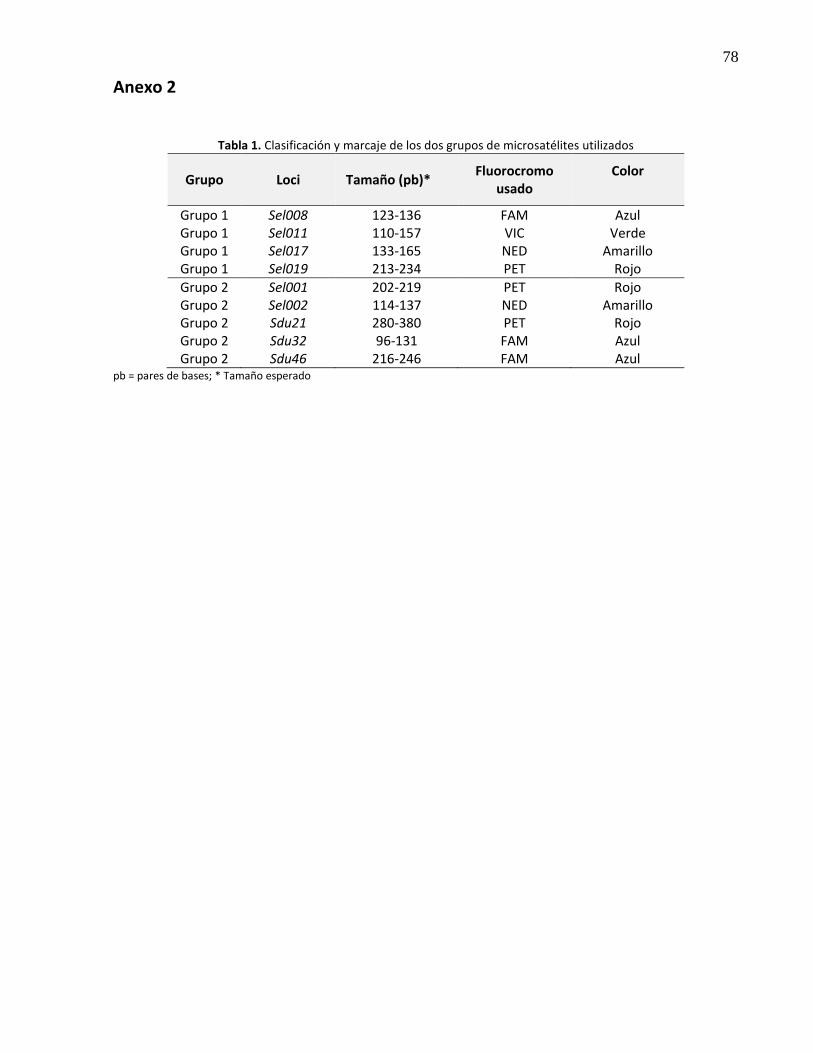
78
Anexo 2
Tabla 1. Clasificación y marcaje de los dos grupos de microsatélites utilizados
Grupo Loci Tamaño (pb)* Fluorocromo
usado Color
Grupo 1 Sel008 123-136 FAM Azul Grupo 1 Sel011 110-157 VIC Verde Grupo 1 Sel017 133-165 NED Amarillo Grupo 1 Sel019 213-234 PET Rojo
Grupo 2 Sel001 202-219 PET Rojo Grupo 2 Sel002 114-137 NED Amarillo Grupo 2 Sdu21 280-380 PET Rojo Grupo 2 Sdu32 96-131 FAM Azul Grupo 2 Sdu46 216-246 FAM Azul
pb = pares de bases; * Tamaño esperado
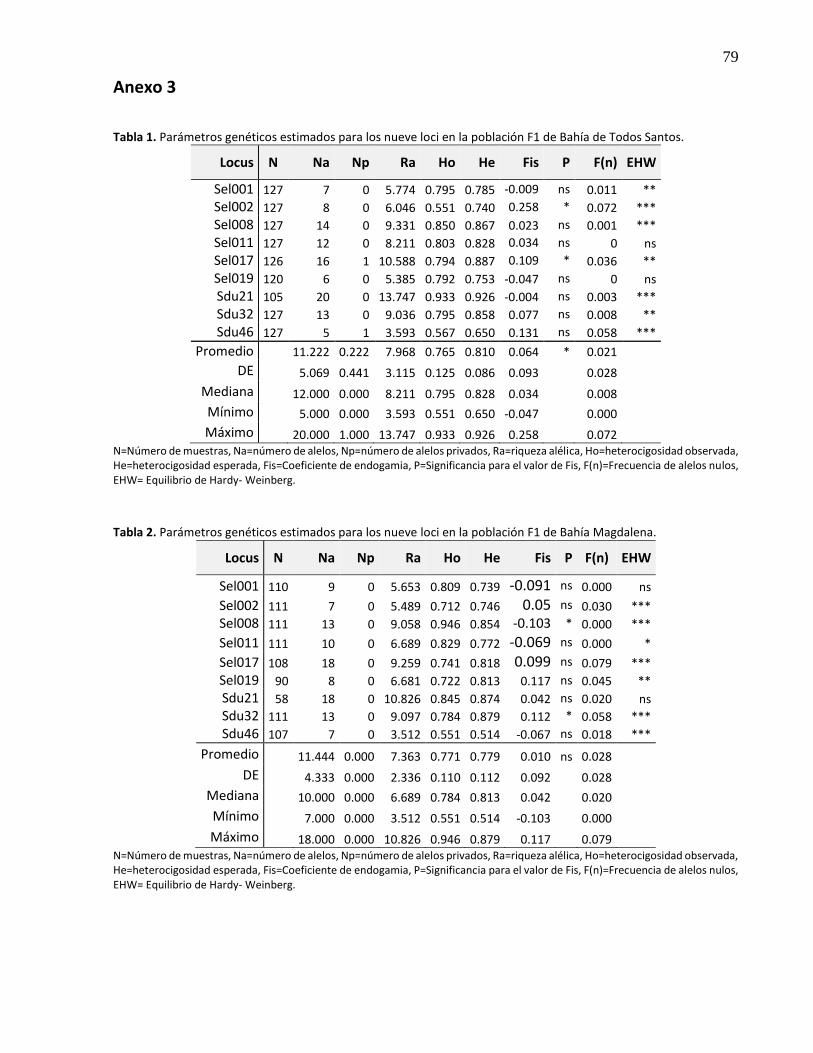
79
Anexo 3
Tabla 1. Parámetros genéticos estimados para los nueve loci en la población F1 de Bahía de Todos Santos.
Locus N Na Np Ra Ho He Fis P F(n) EHW
Sel001 127 7 0 5.774 0.795 0.785 -0.009 ns 0.011 ** Sel002 127 8 0 6.046 0.551 0.740 0.258 * 0.072 *** Sel008 127 14 0 9.331 0.850 0.867 0.023 ns 0.001 *** Sel011 127 12 0 8.211 0.803 0.828 0.034 ns 0 ns Sel017 126 16 1 10.588 0.794 0.887 0.109 * 0.036 ** Sel019 120 6 0 5.385 0.792 0.753 -0.047 ns 0 ns Sdu21 105 20 0 13.747 0.933 0.926 -0.004 ns 0.003 *** Sdu32 127 13 0 9.036 0.795 0.858 0.077 ns 0.008 ** Sdu46 127 5 1 3.593 0.567 0.650 0.131 ns 0.058 ***
Promedio 11.222 0.222 7.968 0.765 0.810 0.064 * 0.021 DE 5.069 0.441 3.115 0.125 0.086 0.093 0.028
Mediana 12.000 0.000 8.211 0.795 0.828 0.034 0.008 Mínimo 5.000 0.000 3.593 0.551 0.650 -0.047 0.000 Máximo 20.000 1.000 13.747 0.933 0.926 0.258 0.072
N=Número de muestras, Na=número de alelos, Np=número de alelos privados, Ra=riqueza alélica, Ho=heterocigosidad observada, He=heterocigosidad esperada, Fis=Coeficiente de endogamia, P=Significancia para el valor de Fis, F(n)=Frecuencia de alelos nulos, EHW= Equilibrio de Hardy- Weinberg.
Tabla 2. Parámetros genéticos estimados para los nueve loci en la población F1 de Bahía Magdalena.
Locus N Na Np Ra Ho He Fis P F(n) EHW
Sel001 110 9 0 5.653 0.809 0.739 -0.091 ns 0.000 ns
Sel002 111 7 0 5.489 0.712 0.746 0.05 ns 0.030 *** Sel008 111 13 0 9.058 0.946 0.854 -0.103 * 0.000 ***
Sel011 111 10 0 6.689 0.829 0.772 -0.069 ns 0.000 *
Sel017 108 18 0 9.259 0.741 0.818 0.099 ns 0.079 *** Sel019 90 8 0 6.681 0.722 0.813 0.117 ns 0.045 ** Sdu21 58 18 0 10.826 0.845 0.874 0.042 ns 0.020 ns Sdu32 111 13 0 9.097 0.784 0.879 0.112 * 0.058 *** Sdu46 107 7 0 3.512 0.551 0.514 -0.067 ns 0.018 ***
Promedio 11.444 0.000 7.363 0.771 0.779 0.010 ns 0.028 DE 4.333 0.000 2.336 0.110 0.112 0.092 0.028
Mediana 10.000 0.000 6.689 0.784 0.813 0.042 0.020 Mínimo 7.000 0.000 3.512 0.551 0.514 -0.103 0.000 Máximo 18.000 0.000 10.826 0.946 0.879 0.117 0.079
N=Número de muestras, Na=número de alelos, Np=número de alelos privados, Ra=riqueza alélica, Ho=heterocigosidad observada, He=heterocigosidad esperada, Fis=Coeficiente de endogamia, P=Significancia para el valor de Fis, F(n)=Frecuencia de alelos nulos, EHW= Equilibrio de Hardy- Weinberg.
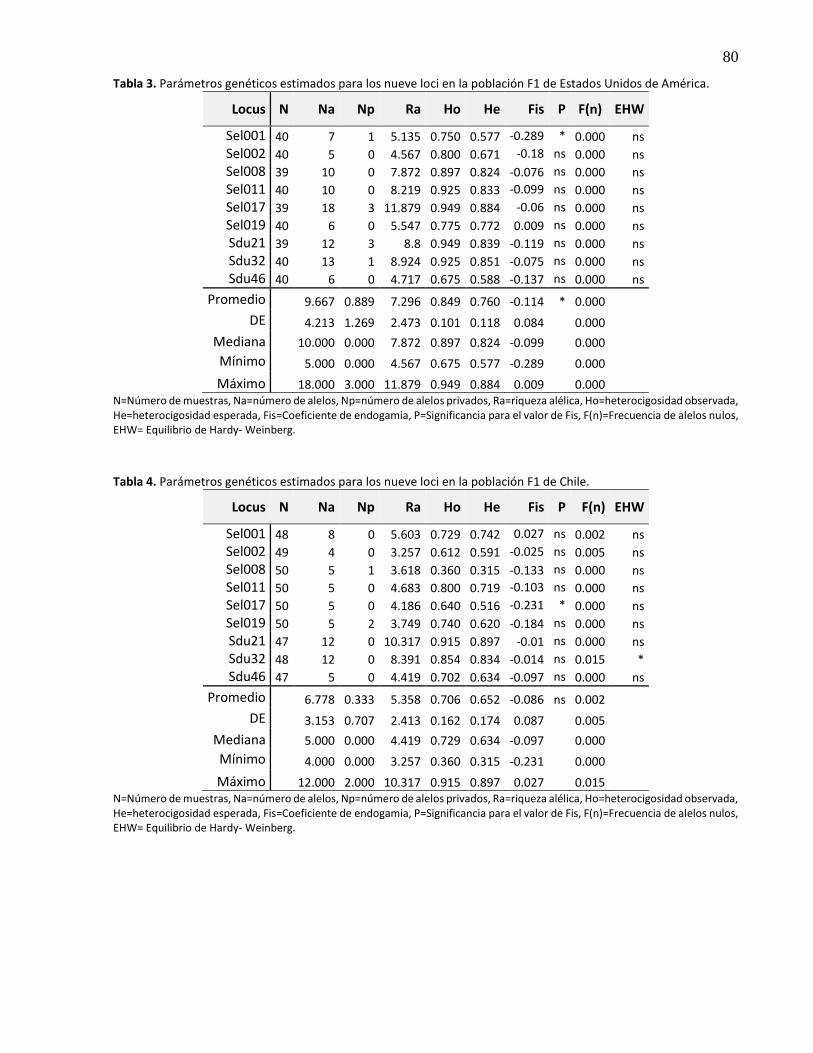
80
Tabla 3. Parámetros genéticos estimados para los nueve loci en la población F1 de Estados Unidos de América.
Locus N Na Np Ra Ho He Fis P F(n) EHW
Sel001 40 7 1 5.135 0.750 0.577 -0.289 * 0.000 ns Sel002 40 5 0 4.567 0.800 0.671 -0.18 ns 0.000 ns Sel008 39 10 0 7.872 0.897 0.824 -0.076 ns 0.000 ns Sel011 40 10 0 8.219 0.925 0.833 -0.099 ns 0.000 ns Sel017 39 18 3 11.879 0.949 0.884 -0.06 ns 0.000 ns Sel019 40 6 0 5.547 0.775 0.772 0.009 ns 0.000 ns Sdu21 39 12 3 8.8 0.949 0.839 -0.119 ns 0.000 ns Sdu32 40 13 1 8.924 0.925 0.851 -0.075 ns 0.000 ns Sdu46 40 6 0 4.717 0.675 0.588 -0.137 ns 0.000 ns
Promedio 9.667 0.889 7.296 0.849 0.760 -0.114 * 0.000
DE 4.213 1.269 2.473 0.101 0.118 0.084 0.000
Mediana 10.000 0.000 7.872 0.897 0.824 -0.099 0.000
Mínimo 5.000 0.000 4.567 0.675 0.577 -0.289 0.000
Máximo 18.000 3.000 11.879 0.949 0.884 0.009 0.000
N=Número de muestras, Na=número de alelos, Np=número de alelos privados, Ra=riqueza alélica, Ho=heterocigosidad observada, He=heterocigosidad esperada, Fis=Coeficiente de endogamia, P=Significancia para el valor de Fis, F(n)=Frecuencia de alelos nulos, EHW= Equilibrio de Hardy- Weinberg.
Tabla 4. Parámetros genéticos estimados para los nueve loci en la población F1 de Chile.
Locus N Na Np Ra Ho He Fis P F(n) EHW
Sel001 48 8 0 5.603 0.729 0.742 0.027 ns 0.002 ns Sel002 49 4 0 3.257 0.612 0.591 -0.025 ns 0.005 ns Sel008 50 5 1 3.618 0.360 0.315 -0.133 ns 0.000 ns Sel011 50 5 0 4.683 0.800 0.719 -0.103 ns 0.000 ns Sel017 50 5 0 4.186 0.640 0.516 -0.231 * 0.000 ns Sel019 50 5 2 3.749 0.740 0.620 -0.184 ns 0.000 ns Sdu21 47 12 0 10.317 0.915 0.897 -0.01 ns 0.000 ns Sdu32 48 12 0 8.391 0.854 0.834 -0.014 ns 0.015 * Sdu46 47 5 0 4.419 0.702 0.634 -0.097 ns 0.000 ns
Promedio 6.778 0.333 5.358 0.706 0.652 -0.086 ns 0.002 DE 3.153 0.707 2.413 0.162 0.174 0.087 0.005
Mediana 5.000 0.000 4.419 0.729 0.634 -0.097 0.000 Mínimo 4.000 0.000 3.257 0.360 0.315 -0.231 0.000 Máximo 12.000 2.000 10.317 0.915 0.897 0.027 0.015
N=Número de muestras, Na=número de alelos, Np=número de alelos privados, Ra=riqueza alélica, Ho=heterocigosidad observada, He=heterocigosidad esperada, Fis=Coeficiente de endogamia, P=Significancia para el valor de Fis, F(n)=Frecuencia de alelos nulos, EHW= Equilibrio de Hardy- Weinberg.

81
Tabla 5. Parámetros genéticos estimados para los nueve loci en la población F1 de Australia.
Locus N Na Np Ra Ho He Fis P F(n) EHW
Sel001 40 4 0 3.996 0.750 0.725 -0.021 ns 0.000 ns
Sel002 49 2 0 2 0.531 0.425 -0.239 ns 0.000 ns
Sel008 48 4 0 3.91 0.792 0.576 -0.365 * 0.000 *
Sel011 49 5 0 4.944 0.776 0.740 -0.038 ns 0.000 ns
Sel017 48 3 0 3 0.667 0.614 -0.076 ns 0.000 ns
Sel019 42 4 0 3.558 0.786 0.658 -0.182 ns 0.000 ns
Sdu21 40 11 0 8.951 0.800 0.850 0.071 ns 0.060 ***
Sdu32 44 8 0 7.314 0.955 0.796 -0.188 ns 0.000 ***
Sdu46 43 3 0 2.999 0.767 0.621 -0.225 ns 0.000 ns
Promedio 4.889 0.000 4.519 0.758 0.667 -0.140 * 0.007 DE 2.848 0.000 2.241 0.113 0.128 0.135 0.020
Mediana 4.000 0.000 3.910 0.776 0.658 -0.182 0.000 Mínimo 2.000 0.000 2.000 0.531 0.425 -0.365 0.000 Máximo 11.000 0.000 8.951 0.955 0.850 0.071 0.060
N=Número de muestras, Na=número de alelos, Np=número de alelos privados, Ra=riqueza alélica, Ho=heterocigosidad observada, He=heterocigosidad esperada, Fis=Coeficiente de endogamia, P=Significancia para el valor de Fis, F(n)=Frecuencia de alelos nulos, EHW= Equilibrio de Hardy- Weinberg.
Tabla 6. Parámetros genéticos estimados para los nueve loci en la población silvestre de Bahía de Todos Santos, BC, México.
Locus N Na Np Ra Ho He Fis P F(n) EHW
Sel001 17 8 0 7.47 0.941 0.813 -0.128 ns 0.000 ns
Sel002 17 5 1 4.79 0.412 0.509 0.220 ns 0.040 ns
Sel008 17 13 0 11.70 0.824 0.848 0.059 ns 0.030 ns
Sel011 17 11 0 10.33 0.941 0.848 -0.080 ns 0.000 ns
Sel017 17 20 0 17.76 0.941 0.931 0.019 ns 0.000 ns
Sel019 17 7 0 6.64 0.706 0.779 0.123 ns 0.037 ns
Sdu21 14 18 0 18.00 0.929 0.934 0.042 ns 0.000 ns
Sdu32 17 14 0 12.50 0.824 0.867 0.080 ns 0.000 ns
Sdu46 17 4 0 3.82 0.118 0.664 0.832 * 0.328 ***
Promedio 11.111 0.111 10.339 0.737 0.799 0.130 * 0.048 DE 5.622 0.333 5.190 0.289 0.136 0.283 0.106
Mediana 11.000 0.000 10.337 0.824 0.848 0.059 0.000 Mínimo 4.000 0.000 3.824 0.118 0.509 -0.128 0.000
Máximo
20.000 1.000 18.000 0.941 0.934 0.832 0.328 N=Número de muestras, Na=número de alelos, Np=número de alelos privados, Ra=riqueza alélica, Ho=heterocigosidad observada, He=heterocigosidad esperada, Fis=Coeficiente de endogamia, P=Significancia para el valor de Fis, F(n)=Frecuencia de alelos nulos, EHW= Equilibrio de Hardy- Weinberg.

82
Tabla 7. Parámetros genéticos estimados para los nueve loci en la población del Ejido Eréndira, BC, México.
Locus N Na Np Ra Ho He Fis P F(n) EHW
Sel001 24 10 0 8.482 0.750 0.836 0.124 ns 0.040 ns Sel002 24 4 0 3.517 0.500 0.568 0.14 ns 0.026 ns Sel008 24 12 0 9.671 0.750 0.860 0.149 ns 0.065 ns Sel011 24 13 0 10.423 0.833 0.854 0.046 ns 0.016 ns Sel017 24 24 2 17.619 0.833 0.934 0.129 ns 0.044 ns Sel019 24 7 0 6.154 0.750 0.777 0.056 ns 0.000 ns Sdu21 23 17 2 14.087 0.348 0.905 0.629 * 0.327 *** Sdu32 24 15 0 12.187 0.750 0.903 0.19 ns 0.085 ns Sdu46 24 8 1 6.73 0.458 0.583 0.234 ns 0.106 ns
Promedio 12.222 0.556 9.874 0.664 0.802 0.189 * 0.079 DE 5.995 0.882 4.323 0.179 0.136 0.175 0.099
Mediana 12.000 0.000 9.671 0.750 0.854 0.140 0.044 Mínimo 4.000 0.000 3.517 0.348 0.568 0.046 0.000 Máximo 24.000 2.000 17.619 0.833 0.934 0.629 0.327
N=Número de muestras, Na=número de alelos, Np=número de alelos privados, Ra=riqueza alélica, Ho=heterocigosidad observada, He=heterocigosidad esperada, Fis=Coeficiente de endogamia, P=Significancia para el valor de Fis, F(n)=Frecuencia de alelos nulos, EHW= Equilibrio de Hardy- Weinberg.
Tabla 8. Parámetros genéticos estimados para los nueve loci en la población de Bahía Magdalena, BCS, México.
Locus N Na Np Ra Ho He Fis P F(n) EHW
Sel001 30 9.000 0 8.856 0.867 0.788 -0.074 ns 0.000 ns Sel002 30 5.000 0 4.933 0.700 0.682 -0.006 ns 0.006 ns Sel008 30 18.000 1 17.516 0.867 0.896 0.049 ns 0.000 ns Sel011 30 11.000 1 10.863 0.900 0.853 -0.038 ns 0.000 ns Sel017 30 24.000 0 23.249 0.867 0.929 0.087 ns 0.015 ns Sel019 30 9.000 0 8.797 0.833 0.818 -0.039 ns 0.000 ns Sdu21 28 24.000 1 24 0.821 0.939 0.149 * 0.064 ns Sdu32 30 18.000 1 17.33 0.767 0.887 0.128 ns 0.045 ns Sdu46 30 7.000 0 6.73 0.267 0.324 0.19 ns 0.046 ***
Promedio 13.889 0.444 13.586 0.765 0.791 0.050 ns 0.020 DE 7.253 0.527 7.123 0.197 0.192 0.094 0.025
Mediana 11.000 0.000 10.863 0.833 0.853 0.049 0.006 Mínimo 5.000 0.000 4.933 0.267 0.324 -0.074 0.000 Máximo 24.000 1.000 24.000 0.900 0.939 0.190 0.064
N=Número de muestras, Na=número de alelos, Np=número de alelos privados, Ra=riqueza alélica, Ho=heterocigosidad observada, He=heterocigosidad esperada, Fis=Coeficiente de endogamia, P=Significancia para el valor de Fis, F(n)=Frecuencia de alelos nulos, EHW= Equilibrio de Hardy- Weinberg.
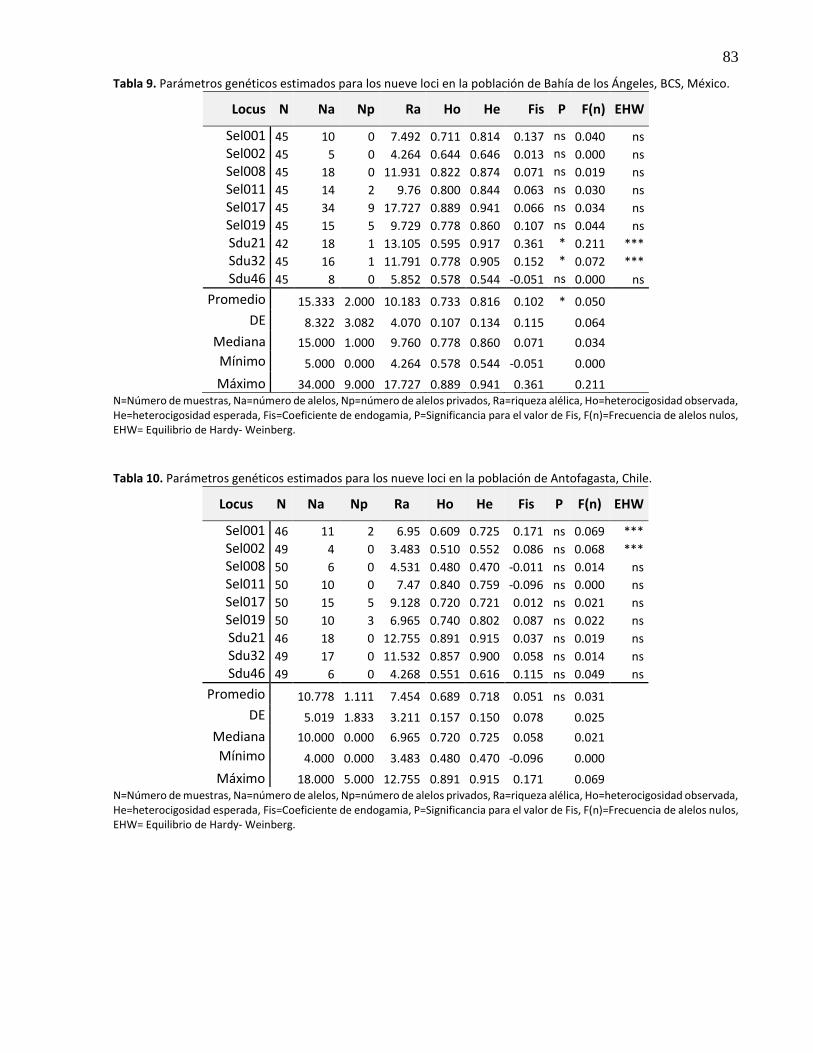
83
Tabla 9. Parámetros genéticos estimados para los nueve loci en la población de Bahía de los Ángeles, BCS, México.
Locus N Na Np Ra Ho He Fis P F(n) EHW
Sel001 45 10 0 7.492 0.711 0.814 0.137 ns 0.040 ns Sel002 45 5 0 4.264 0.644 0.646 0.013 ns 0.000 ns Sel008 45 18 0 11.931 0.822 0.874 0.071 ns 0.019 ns Sel011 45 14 2 9.76 0.800 0.844 0.063 ns 0.030 ns Sel017 45 34 9 17.727 0.889 0.941 0.066 ns 0.034 ns Sel019 45 15 5 9.729 0.778 0.860 0.107 ns 0.044 ns Sdu21 42 18 1 13.105 0.595 0.917 0.361 * 0.211 *** Sdu32 45 16 1 11.791 0.778 0.905 0.152 * 0.072 *** Sdu46 45 8 0 5.852 0.578 0.544 -0.051 ns 0.000 ns
Promedio 15.333 2.000 10.183 0.733 0.816 0.102 * 0.050 DE 8.322 3.082 4.070 0.107 0.134 0.115 0.064
Mediana 15.000 1.000 9.760 0.778 0.860 0.071 0.034 Mínimo 5.000 0.000 4.264 0.578 0.544 -0.051 0.000 Máximo 34.000 9.000 17.727 0.889 0.941 0.361 0.211
N=Número de muestras, Na=número de alelos, Np=número de alelos privados, Ra=riqueza alélica, Ho=heterocigosidad observada, He=heterocigosidad esperada, Fis=Coeficiente de endogamia, P=Significancia para el valor de Fis, F(n)=Frecuencia de alelos nulos, EHW= Equilibrio de Hardy- Weinberg.
Tabla 10. Parámetros genéticos estimados para los nueve loci en la población de Antofagasta, Chile.
N=Número de muestras, Na=número de alelos, Np=número de alelos privados, Ra=riqueza alélica, Ho=heterocigosidad observada, He=heterocigosidad esperada, Fis=Coeficiente de endogamia, P=Significancia para el valor de Fis, F(n)=Frecuencia de alelos nulos, EHW= Equilibrio de Hardy- Weinberg.
Locus N Na Np Ra Ho He Fis P F(n) EHW
Sel001 46 11 2 6.95 0.609 0.725 0.171 ns 0.069 *** Sel002 49 4 0 3.483 0.510 0.552 0.086 ns 0.068 *** Sel008 50 6 0 4.531 0.480 0.470 -0.011 ns 0.014 ns Sel011 50 10 0 7.47 0.840 0.759 -0.096 ns 0.000 ns Sel017 50 15 5 9.128 0.720 0.721 0.012 ns 0.021 ns Sel019 50 10 3 6.965 0.740 0.802 0.087 ns 0.022 ns Sdu21 46 18 0 12.755 0.891 0.915 0.037 ns 0.019 ns Sdu32 49 17 0 11.532 0.857 0.900 0.058 ns 0.014 ns Sdu46 49 6 0 4.268 0.551 0.616 0.115 ns 0.049 ns
Promedio 10.778 1.111 7.454 0.689 0.718 0.051 ns 0.031 DE 5.019 1.833 3.211 0.157 0.150 0.078 0.025
Mediana 10.000 0.000 6.965 0.720 0.725 0.058 0.021 Mínimo 4.000 0.000 3.483 0.480 0.470 -0.096 0.000 Máximo 18.000 5.000 12.755 0.891 0.915 0.171 0.069
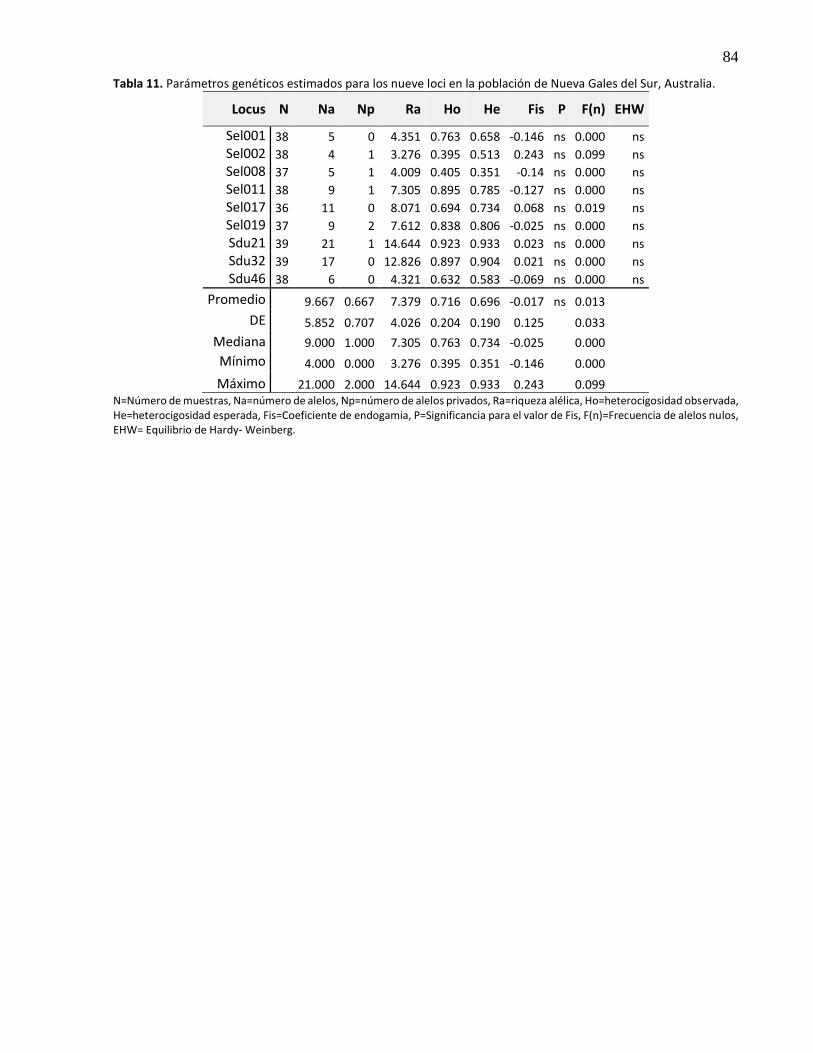
84
Tabla 11. Parámetros genéticos estimados para los nueve loci en la población de Nueva Gales del Sur, Australia.
Locus N Na Np Ra Ho He Fis P F(n) EHW
Sel001 38 5 0 4.351 0.763 0.658 -0.146 ns 0.000 ns Sel002 38 4 1 3.276 0.395 0.513 0.243 ns 0.099 ns Sel008 37 5 1 4.009 0.405 0.351 -0.14 ns 0.000 ns Sel011 38 9 1 7.305 0.895 0.785 -0.127 ns 0.000 ns Sel017 36 11 0 8.071 0.694 0.734 0.068 ns 0.019 ns Sel019 37 9 2 7.612 0.838 0.806 -0.025 ns 0.000 ns Sdu21 39 21 1 14.644 0.923 0.933 0.023 ns 0.000 ns Sdu32 39 17 0 12.826 0.897 0.904 0.021 ns 0.000 ns Sdu46 38 6 0 4.321 0.632 0.583 -0.069 ns 0.000 ns
Promedio 9.667 0.667 7.379 0.716 0.696 -0.017 ns 0.013 DE 5.852 0.707 4.026 0.204 0.190 0.125 0.033
Mediana 9.000 1.000 7.305 0.763 0.734 -0.025 0.000 Mínimo 4.000 0.000 3.276 0.395 0.351 -0.146 0.000 Máximo 21.000 2.000 14.644 0.923 0.933 0.243 0.099
N=Número de muestras, Na=número de alelos, Np=número de alelos privados, Ra=riqueza alélica, Ho=heterocigosidad observada, He=heterocigosidad esperada, Fis=Coeficiente de endogamia, P=Significancia para el valor de Fis, F(n)=Frecuencia de alelos nulos, EHW= Equilibrio de Hardy- Weinberg.
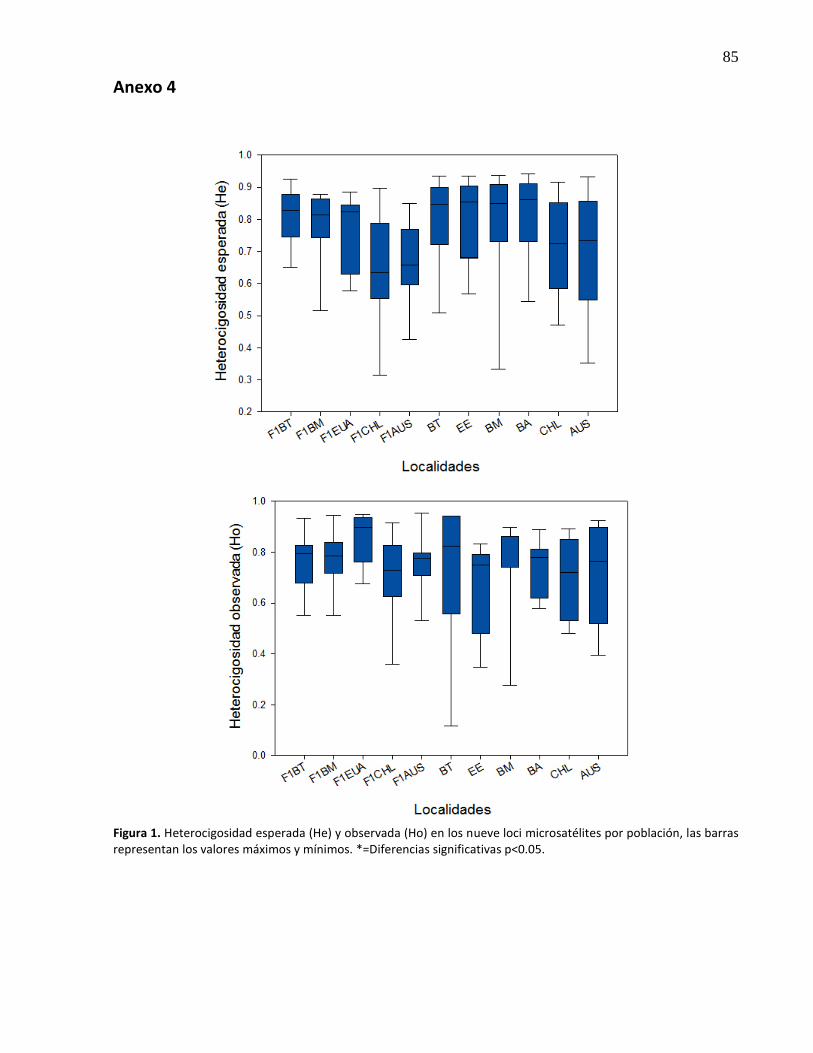
85
Anexo 4
Figura 1. Heterocigosidad esperada (He) y observada (Ho) en los nueve loci microsatélites por población, las barras representan los valores máximos y mínimos. *=Diferencias significativas p<0.05.
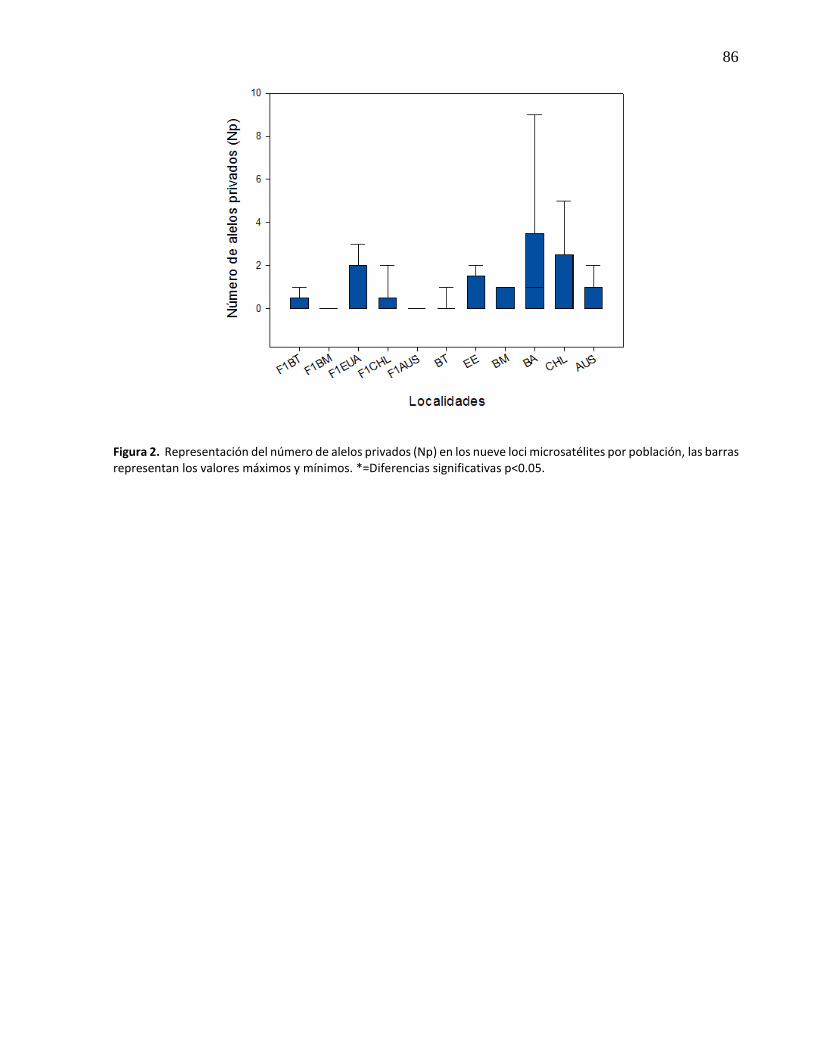
86
Figura 2. Representación del número de alelos privados (Np) en los nueve loci microsatélites por población, las barras representan los valores máximos y mínimos. *=Diferencias significativas p<0.05.
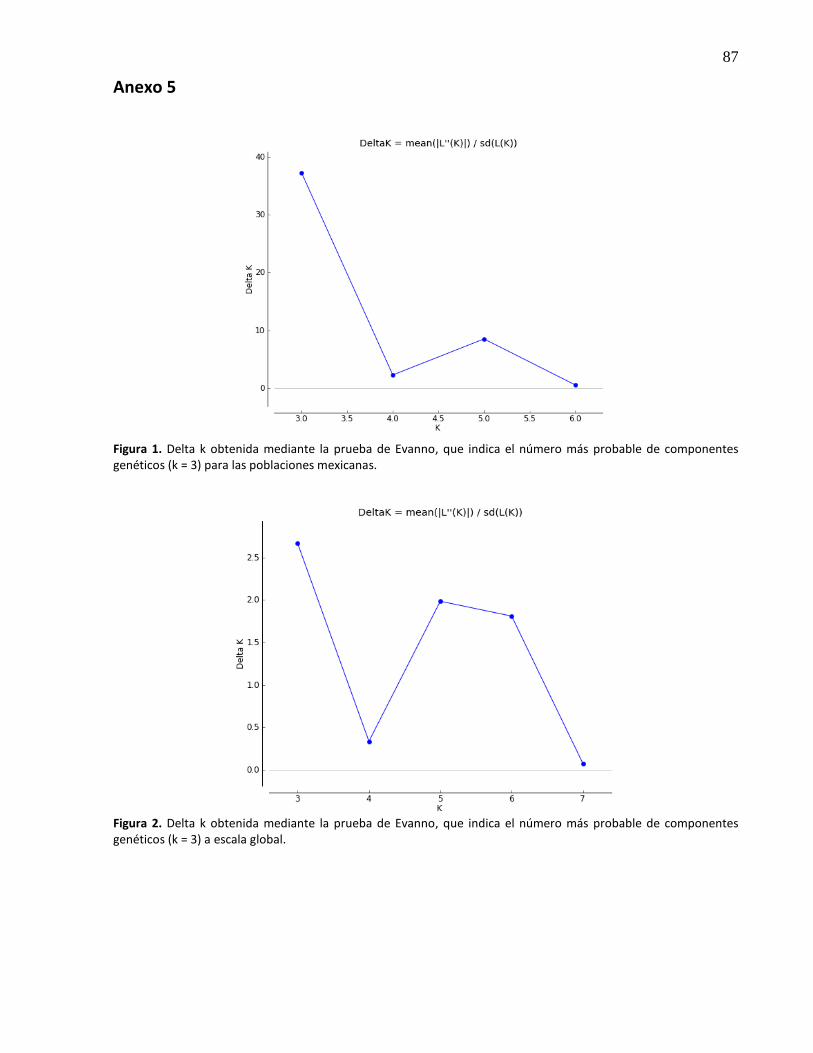
87
Anexo 5
Figura 1. Delta k obtenida mediante la prueba de Evanno, que indica el número más probable de componentes genéticos (k = 3) para las poblaciones mexicanas.
Figura 2. Delta k obtenida mediante la prueba de Evanno, que indica el número más probable de componentes genéticos (k = 3) a escala global.
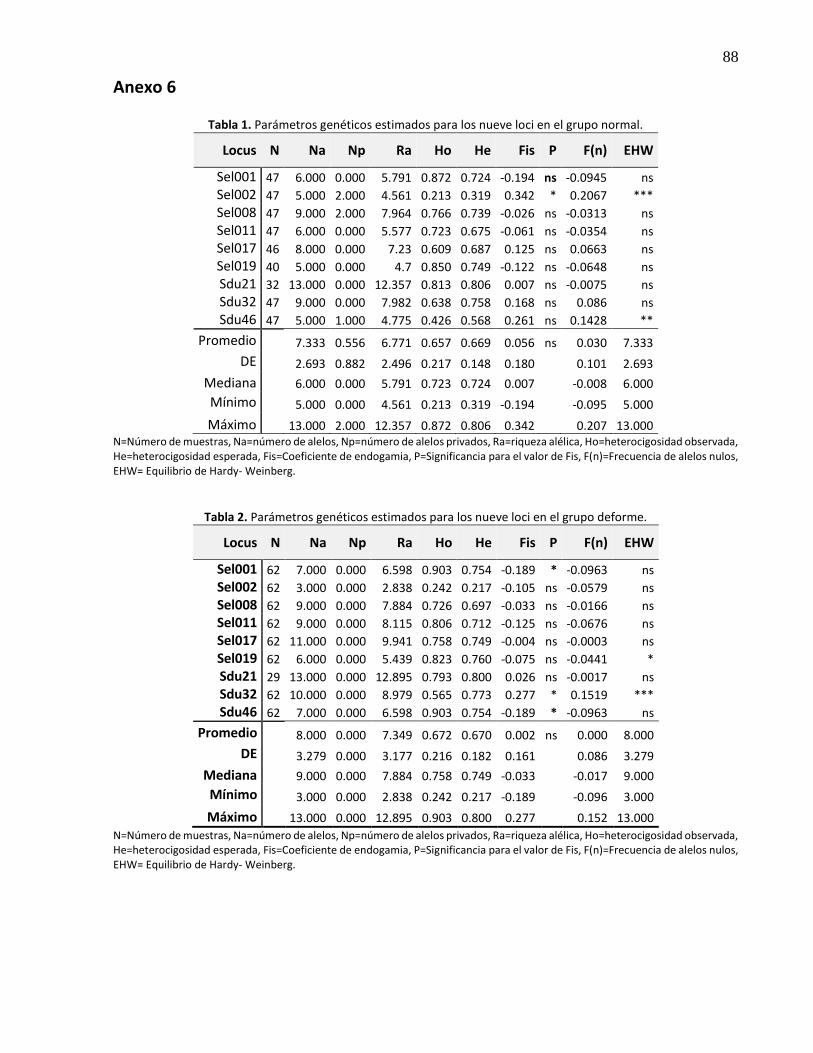
88
Anexo 6
Tabla 1. Parámetros genéticos estimados para los nueve loci en el grupo normal.
Locus N Na Np Ra Ho He Fis P F(n) EHW
Sel001 47 6.000 0.000 5.791 0.872 0.724 -0.194 ns -0.0945 ns Sel002 47 5.000 2.000 4.561 0.213 0.319 0.342 * 0.2067 *** Sel008 47 9.000 2.000 7.964 0.766 0.739 -0.026 ns -0.0313 ns Sel011 47 6.000 0.000 5.577 0.723 0.675 -0.061 ns -0.0354 ns Sel017 46 8.000 0.000 7.23 0.609 0.687 0.125 ns 0.0663 ns Sel019 40 5.000 0.000 4.7 0.850 0.749 -0.122 ns -0.0648 ns Sdu21 32 13.000 0.000 12.357 0.813 0.806 0.007 ns -0.0075 ns Sdu32 47 9.000 0.000 7.982 0.638 0.758 0.168 ns 0.086 ns Sdu46 47 5.000 1.000 4.775 0.426 0.568 0.261 ns 0.1428 **
Promedio 7.333 0.556 6.771 0.657 0.669 0.056 ns 0.030 7.333
DE 2.693 0.882 2.496 0.217 0.148 0.180 0.101 2.693
Mediana 6.000 0.000 5.791 0.723 0.724 0.007 -0.008 6.000
Mínimo 5.000 0.000 4.561 0.213 0.319 -0.194 -0.095 5.000
Máximo 13.000 2.000 12.357 0.872 0.806 0.342 0.207 13.000 N=Número de muestras, Na=número de alelos, Np=número de alelos privados, Ra=riqueza alélica, Ho=heterocigosidad observada, He=heterocigosidad esperada, Fis=Coeficiente de endogamia, P=Significancia para el valor de Fis, F(n)=Frecuencia de alelos nulos, EHW= Equilibrio de Hardy- Weinberg.
Tabla 2. Parámetros genéticos estimados para los nueve loci en el grupo deforme.
Locus N Na Np Ra Ho He Fis P F(n) EHW
Sel001 62 7.000 0.000 6.598 0.903 0.754 -0.189 * -0.0963 ns Sel002 62 3.000 0.000 2.838 0.242 0.217 -0.105 ns -0.0579 ns Sel008 62 9.000 0.000 7.884 0.726 0.697 -0.033 ns -0.0166 ns Sel011 62 9.000 0.000 8.115 0.806 0.712 -0.125 ns -0.0676 ns Sel017 62 11.000 0.000 9.941 0.758 0.749 -0.004 ns -0.0003 ns Sel019 62 6.000 0.000 5.439 0.823 0.760 -0.075 ns -0.0441 * Sdu21 29 13.000 0.000 12.895 0.793 0.800 0.026 ns -0.0017 ns Sdu32 62 10.000 0.000 8.979 0.565 0.773 0.277 * 0.1519 *** Sdu46 62 7.000 0.000 6.598 0.903 0.754 -0.189 * -0.0963 ns
Promedio 8.000 0.000 7.349 0.672 0.670 0.002 ns 0.000 8.000
DE 3.279 0.000 3.177 0.216 0.182 0.161 0.086 3.279
Mediana 9.000 0.000 7.884 0.758 0.749 -0.033 -0.017 9.000
Mínimo 3.000 0.000 2.838 0.242 0.217 -0.189 -0.096 3.000
Máximo 13.000 0.000 12.895 0.903 0.800 0.277 0.152 13.000 N=Número de muestras, Na=número de alelos, Np=número de alelos privados, Ra=riqueza alélica, Ho=heterocigosidad observada, He=heterocigosidad esperada, Fis=Coeficiente de endogamia, P=Significancia para el valor de Fis, F(n)=Frecuencia de alelos nulos, EHW= Equilibrio de Hardy- Weinberg.