BLOQUE III. ¿DÓNDE ESTÁ LA INFORMACIÓN DE … · Griffith dedujo que en las ... El ARN y sus...
32
BLOQUE III. ¿DÓNDE ESTÁ LA INFORMACIÓN DE LOS SERES VIVOS? ¿CÓMO SE EXPRESA Y SE TRASMITE? LABASE QUÍMICA DE LA HERENCIA. PRINCIPALES TEMAS DEL CURRICULUM 1. Genética molecular. 1.1. El ADN como portador de la información genética. 1.1.1. ADN y cromosomas. 1.1.2. Concepto de gen. 1.1.3. Conservación de la información: la replicación del ADN. 1.1.4. Expresión de la información genética (flujo de la información genética): transcripción y traducción en procariotas y eucariotas. 1.1.5. El código genético. 1.2. Alteraciones de la información genética. 1.2.1. Concepto de mutación. 1.2.2. Causas de las mutaciones. 1.2.3. Consecuencias de las mutaciones. 1.2.3.1. Consecuencias evolutivas. 1.2.3.2. Efectos perjudiciales. 2. Genética mendeliana 2.1. Conceptos básicos de herencia biológica. 2.1.1. Genotipo y fenotipo. 2.2. Aportaciones de Mendel al estudio de la herencia. 2.2.1. Leyes de Mendel. 2.2.2. Cruzamiento prueba y retrocruzamiento. 2.2.3. Ejemplos de herencia mendeliana en animales y plantas. 2.3. Teoría cromosómica de la herencia. 2.3.1. Los genes y los cromosomas. 2.3.2. Relación del proceso meiótico con las leyes de Mendel. 2.3.3. Determinismo del sexo y herencia ligada al sexo. II. ORIENTACIONES 1. Reconocer al ADN como molécula portadora de la información genética. Recordar que el ADN es el componente esencial de los cromosomas. 2. Entender el gen como el fragmento de ADN que constituye la más pequeña unidad funcional. 3. Relacionar e identificar el proceso de replicación del ADN como el mecanismo de conservación de la información genética. 4. Reconocer la necesidad de que la información genética se exprese y explicar brevemente los procesos de transcripción y traducción por los que se realiza dicha expresión. 5. Comprender la forma en que está codificada la información genética y valorar su universalidad. 6. Definir las mutaciones como alteraciones genéticas.
Transcript of BLOQUE III. ¿DÓNDE ESTÁ LA INFORMACIÓN DE … · Griffith dedujo que en las ... El ARN y sus...

BLOQUE III. ¿DÓNDE ESTÁ LA INFORMACIÓN DE LOS SERES VIVOS? ¿CÓMO SE EXPRESA Y SE TRASMITE? LABASE QUÍMICA DE LA HERENCIA.
PRINCIPALES TEMAS DEL CURRICULUM 1.
Genética molecular.
1.1. El ADN como portador de la información genética.
1.1.1. ADN y cromosomas.
1.1.2. Concepto de gen.
1.1.3. Conservación de la información: la replicación del ADN.
1.1.4. Expresión de la información genética (flujo de la información genética): transcripción y
traducción en procariotas y eucariotas.
1.1.5. El código genético.
1.2. Alteraciones de la información genética.
1.2.1. Concepto de mutación.
1.2.2. Causas de las mutaciones.
1.2.3. Consecuencias de las mutaciones.
1.2.3.1. Consecuencias evolutivas.
1.2.3.2. Efectos perjudiciales.
2. Genética mendeliana
2.1. Conceptos básicos de herencia biológica.
2.1.1. Genotipo y fenotipo.
2.2. Aportaciones de Mendel al estudio de la herencia.
2.2.1. Leyes de Mendel.
2.2.2. Cruzamiento prueba y retrocruzamiento.
2.2.3. Ejemplos de herencia mendeliana en animales y plantas.
2.3. Teoría cromosómica de la herencia.
2.3.1. Los genes y los cromosomas.
2.3.2. Relación del proceso meiótico con las leyes de Mendel.
2.3.3. Determinismo del sexo y herencia ligada al sexo.
II. ORIENTACIONES 1. Reconocer al ADN como molécula portadora de la información genética. Recordar que el ADN es
el componente esencial de los cromosomas.
2. Entender el gen como el fragmento de ADN que constituye la más pequeña unidad funcional.
3. Relacionar e identificar el proceso de replicación del ADN como el mecanismo de conservación de
la información genética.
4. Reconocer la necesidad de que la información genética se exprese y explicar brevemente los
procesos de transcripción y traducción por los que se realiza dicha expresión.
5. Comprender la forma en que está codificada la información genética y valorar su universalidad.
6. Definir las mutaciones como alteraciones genéticas.

7. Distinguir entre mutación espontánea e inducida y citar algunos agentes mutagénicos: rayos UV,
radiaciones ionizantes, agentes químicos y agentes biológicos.
8. Destacar que las mutaciones son necesarias pero no suficientes para explicar el proceso evolutivo.
9. Reconocer el efecto perjudicial de gran número de mutaciones y relacionar el concepto de
mutación con el de enfermedad hereditaria.
10. Definir y explicar el significado de los siguientes términos: genoma, cariotipo, gen, alelo,
locus, homocigótico, heterocigótico, herencia dominante, recesiva, intermedia (dominancia
parcial o incompleta) y codominancia.
11. Aplicar los mecanismos de la herencia mediante el estudio de las leyes de Mendel a
supuestos sencillos de cruzamientos monohíbridos y dihíbridos con genes autosómicos y genes
ligados al sexo. 12. Reconocer el proceso que siguen los cromosomas en la meiosis como
fundamento citológico de la distribución de los factores hereditarios en los postulados de Mendel.
III. OBSERVACIONES
1. Se recomienda que los procesos de replicación del ADN, transcripción y traducción se
expliquen tomando como referencia lo que acontece en una célula procariótica sin dejar de resaltar
la compartimentación asociada a estos procesos en las células eucarióticas.
2. En el proceso de replicación del ADN, se sugiere, al menos, la mención de: origen de
replicación, sentido 5´ ---> 3´, cadenas adelantada (conductora) y retrasada (retardada), cebador,
fragmento de Okazaki, ADN y ARN polimerasas y ADN ligasa.
3. En la explicación del proceso de transcripción se sugiere, al menos, la mención de: diferencia
entre cadena codificante y cadena molde del ADN, sentido 5´ ---> 3´, copia de una sola cadena del
ADN, señal de inicio (promotor), acción de la ARN polimerasa y señal de terminación.
4. En la síntesis de proteínas se sugiere la mención de, al menos: etapa de iniciación (ARN
mensajero, ARN transferente, codón de inicio, anticodón y subunidades ribosómicas); etapa de
elongación (formación del enlace peptídico y desplazamiento del ribosoma (translocación); etapa de
terminación (codón de terminación).
5. En relación con el código genético, los alumnos deben conocer, al menos, que se trata de un
código universal (aunque con excepciones) y degenerado.
6. Se sugiere el uso de diferentes tablas o imágenes del código genético donde se muestre la
asignación de aminoácidos a los 64 tripletes; tanto el modelo conocido en una tabla de doble entrada
como el modelo de círculos concéntricos, u otros similares.
7. No será necesario explicar los tipos de mutaciones, pero el alumno deberá ser capaz de
reconocer como mutaciones los cambios en una secuencia de nucleótidos y los cambios en la
dotación cromosómica, e interpretar las consecuencias de las mismas.
8. Los problemas de genética mendeliana serán incluidos en el examen como preguntas de
razonamiento o de interpretación de imágenes. En cualquier caso, los problemas versarán sobre
aspectos básicos elementales y de aplicación directa de la herencia mendeliana, no siendo materia
de examen los problemas de pedigrí. Se sugiere la realización de ejercicios relacionados con la
herencia autosómica, incluyendo los sistemas ABO y Rh (sólo alelo D) de los grupos sanguíneos y con la herencia ligada al sexo, incluyendo los relacionados con el daltonismo y la hemofilia

1. Genética molecular.
1.1. El ADN como portador de la información genética
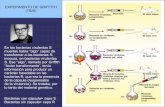
En 1928 F, Griffith buscaba una vacuna contra la neumonía provocada por la bacteria Streptococcus
pneumoniae que producía la neumonía (pulmonía), así descubrió que existían dos cepas distintas:
Smooth (liso) y Rough (rugoso). Las cepas S poseen una capa gelatinosa de polisacáridos y son
capaces de provocar la enfermedad cuando se inoculan en un animal sano. Las colonias tienen un
aspecto liso. Las cepas R no provocan la enfermedad. No tienen cápsula gelatinosa y las colonias
tienen un aspecto rugoso.
Intentando conseguir la vacuna, pensó que se podrían inmunizar ratones inyectándole bacterias
virulentas (S) muertas por el calor o bien hacerlo con bacterias vivas no virulentas (R). En sus
ensayos obtuvo algún resultado inesperado. Griffith dedujo que en las bacterias muertas había
“algo” que le denominó principio transformante, que era captado por las bacterias vivas no
virulentas y transformaba sus caracteres hereditarios convirtiéndolas en virulentas.
En 1944, Avery et col. Demostraron que el principio transformante de Griffith era el ADN, ya que
para que un extracto de células S modificaba el comportamiento de las cepas R (no virulentas), lo
único que podían añadir era su propio ADN (el de la cepa S). En el resto de organismos no
bacterianos se demostró gracias a las experiencias de Hershey y Chase que trabajaron con el
bacteriófago T2, un virus que ataca a la E. coli y que está formado exclusivamente por ADN y
proteínas, que son las dos sustancias que se sospechaban que podrían ser material hereditario.
Las proteínas del virus tiene azufre pero no fósforo y su ADN tiene fósforo pero no azufre. Marcaron
radiactivamente los fagos con P 32 y otros fagos con S35 . Cada grupo sirvió para infectar un cultivo

de bacterias diferente; posteriormente se trituraron y centrifugaron las muestras comprobándose
que el S35 estaba en el medio externo mientras que en el interno abundaba el P32. Esto es lógico
si pensamos que las bacterias habían sido infectadas por virus cuyo ADN se había recombinado con
el de las bacterias. (recuerda que el ADN tiene fósforo). Obviamente el material hereditario no podía
ser otro que el ADN

1.1.1. ADN y cromosomas.
1.1.2. Concepto de gen.
El ADN es el material del que están formados los genes y contiene la información necesaria que
permite la síntesis de todas las proteínas de un organismo. Pero esta información se tiene que
descodificar para poder ser utilizada por la célula, este proceso se realiza en dos fases:
TRANSCRIPCIÓN Y TRADUCCIÓN,
ESTO ES LO QUE CONSTITUYE EL DOGMA CENTRAL DE LA BIOLOGÍA
ADN ARNm Proteína
Autoduplicación. Mediante mecanismos de síntesis se dotan a las células hijas de la información
genética adecuada para que los caracteres se pasen de generación en generación.
Funciones biológicas del ADN y el ARN:
El ADN es el portador de la información genética. Está protegido en el núcleo en las células
eucarióticas y en la c. procariotas se encuentra en el protoplasma.
El ARN y sus diferentes tipos intervienen en la transcripción y la traducción de la información
genética.
Podemos imaginar que nuestro genoma es como una gran biblioteca compuesta por 46
estanterías, organizadas en 23 pares, cada una de las cuales contiene muchos libros con
información para sintetizar enzimas o proteínas.
Si imaginamos nuestro genoma como una gran biblioteca, decimos entonces que los pares de
estantes son los cromosomas homólogos. Cada miembro de un par de cromosomas es similar,
pero no idéntico, a su compañero. Los libros son los genes, en los cuales se guarda la información
para fabricar una proteína o una enzima. El conjunto de genes de una especie determinada se
llama genoma.
Cromosomas, genes y ADN
En los organismos eucariontes, el ADN está organizado en cromosomas. Cada especie tiene un
número característico: la cebolla tiene 16 (organizados en 8 pares), la mosca de la fruta Drosophila
melanogaster, 8, y los seres humanos, 46. De esto no se desprende que una mayor cantidad de
cromosomas equivale a ser “más inteligente” ya que las células que componen las patatas tienen 48
cromosomas.

Los seres humanos tenemos 23 pares de cromosomas: 22 de ellos se llaman
cromosomas autosómicos y se heredan uno del padre y otro de la madre. Los
cromosomas del par 23 se llaman cromosomas sexuales y son diferentes
entre sí.
En muchos organismos los cromosomas sexuales son distintos entre sí. Los
seres humanos (así como otros mamíferos) tenemos cromosomas X iguales
para el hombre y la mujer, mientras que el cromosoma Y, en los hombres, es
un poco más corto y tiene menos genes. En las aves, los machos tienen dos
cromosomas WW y las hembras uno W y otro Z.
Tipos de cromosomas dependiendo de la posición del
centrómero
En cada cromosoma, que contiene una única molécula de ADN asociada a proteínas, se pueden
distinguir las siguientes estructuras:
Centrómero. Es el punto de unión de las cromátidas hermanas.
Telómeros. Son las regiones del cromosoma ubicadas en los extremos, con estructuras de ADN
repetidas que aseguran que no se pierda información importante en cada ciclo de duplicación.
Orígenes de replicación: son los lugares donde comienza la replicación del ADN.
Si tomamos una fotomicrografía de todos los cromosomas de un ser humano, luego cortamos las
imágenes de cada cromosoma individual y las ordenamos, creamos una imagen llamada cariotipo.
Este permite detectar anomalías cromosómicas en células somáticas debidas a enfermedades
genéticas o determinados tipos de cáncer.


1.1.2. CONCEPTO DE GEN
Un gen es la unidad básica de herencia de los seres vivos. Desde el punto de vista molecular,
un gen es una secuencia lineal de nucleótidos en la molécula de ADN (o ARN en el caso de
algunos virus), que contiene la información necesaria para la síntesis de una macromolécula
con función celular específica.
El gen es considerado como la unidad de almacenamiento de información y unidad de herencia
al transmitir esa información a la descendencia. Los genes se disponen, a lo largo de cada uno
de los cromosomas. Cada gen ocupa en el cromosoma una posición determinada llamada locus.
El conjunto de cromosomas de una especie se denomina genoma.
Algunas enfermedades como la anemia drepanocítica (o anemia falciforme) pueden ser
ocasionadas por un cambio en un solo gen (uno de los 30.000 genes que constituyen el plan
para todo el cuerpo humano).
Los organismos diploides (entre ellos, casi todos los animales y plantas) disponen de dos juegos
de cromosomas homólogos, cada uno de ellos proveniente de uno de los padres. Cada par de
cromosomas tiene, pues, un par de copias de cada gen, una procedente de la madre y otra del
padre.
Los genes pueden aparecer en versiones diferentes, con variaciones pequeñas en su secuencia,
denominadas alelos. Los alelos pueden ser dominantes o recesivos. Cuando una sola copia del
alelo hace que se manifieste el rasgo fenotípico, el alelo es dominante. Cuando son precisas dos
copias del alelo (una en cada cromosoma del par), el alelo es recesivo.
Tipos de genes
La mayoría de los genes codifican proteínas, responsables de la mayor parte de las propiedades
de un organismo. Para ello, la transcripción genera una molécula de ARN que posteriormente
se traducirá en proteínas en los ribosomas. Muchos genes se encuentran constituidos por
regiones codificantes (exones) interrumpidas por regiones no codificantes (intrones) que son
eliminadas en el procesamiento del ARN. En células procariontes esto no ocurre.

La secuencia de bases presente en el ARN determina la secuencia de aminoácidos de la
proteína por medio del código genético. Otros genes no son traducidos a proteína, sino que
cumplen su función en forma de ARN. Entre éstos, encontramos genes de ARN transferente,
ARN ribosómico, ribozimas y otros ARN pequeños de funciones diversas.
Algunos genes han sufrido procesos de mutación u otros fenómenos de reorganización y han
dejado de ser funcionales, pero persisten en los genomas de los seres vivos. Al dejar de tener
función, se denominan pseudogenes, y pueden ser muy parecidos a otros genes del mismo
organismo que sean funcionales.
1.1.3. Conservación de la información: la replicación del
ADN
Sin duda alguna, el comienzo del estudio de la replicación del DNA empieza con el
descubrimiento de un enzima: La DNA polimerasa III, que en bacterias es la responsable de hacer
una copia del cromosoma de E. coli, justo antes de que comience la división celular.
Este enzima solo puede añadir en el extremo 3’ nuevos desoxirribonucleótidos; por lo tanto, la
síntesis de DNA siempre se hace en la dirección 5’ 3’. Los nuevos desoxirribonucleótidos
incorporados están trifosfatados, por esta razón se denominan dNTP, donde la N representa
cualquiera de las cuatro bases. De esta manera las reacciones de incorporación de nuevos
nucleótidos a la cadena es exergónica (ceden energía) y provoca la aparición de enlaces
fosfodiéster en una hebra creciente.
1- Comienzo de la REPLICACIÓN.
Cuando se fotografiaron bacterias se observó una especie de burbuja que comenzaba en un solo
lugar, a este punto se le denominó origen de la replicación. La burbuja crecía con el tiempo y lo
hacía bidireccionalmente pero siempre en sentido 5’ 3’ (ten en cuenta que las hebras son
antiparalelas). En los cromosomas eucariotas es prácticamente igual, solo que se hace en varios
puntos a la vez, observándose múltiples burbujas de replicación.

Un conjunto de proteínas (activadas por las proteínas que desencadenan la fase S en el ciclo
celular) reconocen los lugares donde va a comenzar la síntesis y abren una doble hélice. Una vez
que se ha abierto la cadena, un grupo de enzimas toma el control y comienza la replicación. Una
HORQUILLA DE REPLICACIÓN es una región en forma de Y donde la doble hélice parental se
divide en dos hebras simples.
2- APERTURA Y ESTABILIZACIÓN DE LA DOBLE CADENA DE DNA
a. LA HELICASA cataliza la ruptura de los puentes de hidrógeno entre nucleótidos
complementarios
b. Las proteínas de unión (SSBP) proteínas de unión al DNA monocatenario, impiden que
se vuelvan a unir para hacer una doble hélice.
c. Las tensiones que se generan al desenrollar la cadena son compensadas por la
TOPOGIRASAISOMERASA, que va cortando y pegando para que desaparezcan los giros
En este momento, la horquilla está abierta y estabilizada.
3- SINTESIS DE LA HEBRA CONDUCTORA (la que se copia de forma continua)
Recuerda que la DNA polimerasa III solo trabaja en sentido 5’ 3’, incorporando nuevos
nucleótidos en 3’. Con este enzima y la plantilla de la cadena simple de DNA, solo tiene que
ir incorporando nucleótidos complementarios. Un grupo de nucleótidos denominados
CEBADORES proporcionan los grupos –OH que se une a un dNTP complementario formando
un enlace fosfodiéster.
Cuando el cebador se añade a una plantilla de hebra simple, la DNA polimerasa III comienza
a trabajar en sentido 5’ 3’; pero antes de que comience la síntesis un enzima llamado
PRIMASA (una RNA polimerasa) tiene que sintetizar un corto fragmento de RNA que actúa
como cebador para la DNA polimerasa.
La DNA polimerasa III añade dNTP al extremo 3’ de la nueva hebra, en una secuencia
complementaria a la hebra patrón o molde, literalmente agarra al DNA, como si tu mano
agarrase una cuerda. Al final el producto obtenido es una hebra continua o hebra
conductora
4- SÍNTESIS DE LA HEBRA RETRASADA O RETARDADA o discontinua
Ya que las dos cadenas de una DNA son antiparalelas y que además la DNA polimerasa III solo
lee en sentido 5’ 3’, la hebra que se tiene que sintetizar a partir del molde se hace con un
retraso respecto a la otra hebra.
La DNA polimerasa que trabaja en la horquilla, añade dNTP de forma continua; sin embargo,
otra DNA polimerasa tiene que hacerlo alejada de la horquilla para sintetizar en sentido 5’3´,
esta hebra así sintetizada se denomina hebra retardada

Esta síntesis comienza cuando la primasa sintetiza un corto cebador de RNA. A continuación, la
DNA polimerasa III va añadiendo nucleótidos en 3’ de la hebra retrasada, mientras tanto la
helicasa sigue abriendo la doble cadena generando un hueco no copiado directamente.
¿Cómo se replicaba ese hueco?...
Reiji Okazaki y sus colaboradores imaginaron que un RNA
primasa sintetiza un cebador de RNA en la hebra
retardada provocando que la DNA polimerasa III vaya
sintetizando FRAGMENTOS CORTOS de DNA (fragmentos
de Okazaki) y que posteriormente estos fragmentos se
unirían.
La DNA polimerasa I elimina el cebador del RNA al
principio de cada fragmento y rellena con los dNTP
apropiados; después, una LIGASA cataliza la formación de
enlaces fosfodiéster entre fragmentos adyacentes.


CORRECCIÓN DE ERRORES
La actividad autocorrectora exonucleásica del ADN polimerasa es un buen mecanismo de
prevención de errores, pero aun así se comete un
error de apareamiento por cada 10 x 106 bases, así
pues, este mecanismo puede ser eficaz en bacterias,
pero es insuficiente en genoma humano con 3 x 109
pares de bases.
De cualquier manera, se ha descrito un mecanismo de
corrección POSTREPLICATIVA que da lugar a un error
por cada 10.000 106 de bases apareadas, esta
corrección se realiza gracias a un complejo de enzimas
que detecta el nucleótido mal emparejado, lo elimina
y regenera la secuencia del siguiente modo:
1) Se corta un trozo comprendido entre dos GATC
contiguas que contengan el error. Detecta la cadena
errónea porque las secuencias GATC no tienen las
bases metiladas hasta que pasa un cierto tiempo.
2) Actúa una ENDONUCLEASA que corta la cadena
en el lugar exacto del nucleótido mal emparejado
3) La ADN polimerasa regenera la secuencia
correcta y rellenan el hueco tomando como molde el
ADN parental. Se elimina el trozo erróneo y la ADN
polimerasa fábrica la secuencia correcta; después
actúa una ligasa empalmando la cadena.

1.1.4. Expresión de la información genética (flujo de la información genética):
transcripción y traducción en procariotas y eucariotas.
Uno de los principales avances en Biología, durante el siglo XX, fue encontrar la relación existente
DNA y proteínas. Para comprenderlo seguiremos el clásico esquema que es el DOGMA CENTRAL
DE LA BIOLOGÍA MOLECULAR:
TRANSCRIPCIÓN: DNA RNAm
TRADUCCIÓN: RNA proteínas
Las enzimas responsables de la síntesis de RNAm son las RNA polimerasas. Estas enzimas hacen
una síntesis copiando la “plantilla” de una hebra de DNA en dirección 5’ 3’ (acuérdate que
estos números representan los carbonos de las ribosas de los nucleótidos enganchados por un
enlace fosfodiéster)
Cuando la RNA polimerasa acopla la base de un ribonucleótido fosfato con su base
complementaria en un gen cualquiera. Cuando este ribonucleotido está en su lugar la RNA
polimerasa cataliza la formación de un enlace fosfodiéster entre el extremo 3’ de la cadena en
crecimiento de RNAm y el nuevo incorporado. Cuando este proceso se repite un número
indeterminado de veces se sintetiza un RNA complementario a la hebra de DNA.
LA cadena leída por el RNA polimerasa es la hebra no codificante, la otra hebra es la denominada
codificante porque su secuencia se empareja con la secuencia del RNA que se transcribe a partir
de la hebra no codificante. Esta cadena de RNA contiene URACILO en vez de TIMINA.
LA RNA polimerasa es un enzima globular con diversos canales internos, el centro activo del
enzima está en un cruce de estos canales.
INICIACIÓN La RNA polimerasa no puede iniciar por su cuenta la síntesis del RNAm, es
necesaria la presencia de una proteína separable del enzima denominada
SIGMA. LA RNA polimerasa y SIGMA es lo que llamábamos en bioquímica
un HOLOENZIMA.
Parece ser que SIGMA ayuda a la RNA polimerasa a encontrar lugares
específicos (promotores) donde comenzar la transcripción. Pero. ¿Cómo y
qué hacen los promotores?.......
David Pribnow, estudiando bacterias, comprobó en 1970 que los
promotores estaban localizados en la hebra codificante, tenían una
longitud de 40-50 pares de bases y en todos sus experimentos observó que
aparecía un factor común en ellas: UN SECUENCIA TATAAT, hoy se
denomina caja de Pribnow o caja -10. David Pribnow

Esta caja está situada a 10 pares de bases (cuesta ARRIBA) de donde la RNA polimerasa comienza
la transcripción.

Años más tarde se encontró una
secuencia TTGACA (caja -35), en los
promotores a -35 bases del punto +1
que es donde comienza la síntesis.
SIGMA es capaz de reconocer ambas
cajas y orientar a la RNA polimerasa
dónde y cuándo debe comenzar la
transcripción. Cuando esta proteína
contacta con el DNA, lo abre y crea dos
cadenas de ADN, la hebra no
codificante y la codificante. La no
codificante se inserta a través del
canal que conduce la hebra al centro
activo del holoenzima.
Los monómeros NTP (similares a los
dNTP descritos anteriormente) entran
por otro canal y difunden hacia el
centro activo; cuando se empareja
con su base complementaria sobre la
hebra no codificante comienza la
polimerización del RNA, esta reacción
está catalizada por la RNA polimerasa
y es exergónica y espontánea. En este
momento se libera SIGMA.
Estudios recientes han demostrado
que cada bacteria tiene distintos de
proteínas SIGMA y por lo tanto elige
distintos tipos de genes para su
transcripción. En Streptomyces se han
descrito 60 tipos de proteínas SIGMA
con ligeras variaciones, aunque todas
tienen caja -10 y -35.

ELONGACIÓN y TERMINACIÓN
En esta fase la RNA polimerasa se mueve a lo largo de la plantilla de DNA en dirección 3’- 5’
sintetizando RNAm en dirección 5’-3’. Un grupo de aminoácidos cerca del centro activo de la
Holoenzima actúan como timón y cremallera ayudan a abrir la doble hélice de DNA y a guiar
hacia los canales adecuados las hebras de DNA. Los TNP entran por otro canal y la hebra
recién sintetizada de RNA sale por otro canal.
La transcripción acaba cuando la RNA polimerasa alcanza un tramo de la secuencia de DNA
que funciona como señal de terminación de la transcripción. En el momento que se
transcribe esta secuencia el RNA se pliega hacia atrás sobre sí misma y forma una doble
hélice. Esta estructura así plegada de denomina HORQUILLA, gracias a este pliegue se separa
la RNA polimerasa del transcrito de RNA separándose físicamente.
EXONES, INTRONES Y UNIÓN DE LOS RNA TRANSCRITOS
En eucariotas el trasncrito primario de RNA contiene exones e intrones, conforme tiene lugar
la transcripción, los intrones son eliminados de la cadena.
Esta eliminación es muy compleja:
Para eliminar un intrón entre dos exones interviene una snRNP (riboproteina pequeña
nucleolar) que al encontrar la secuencia que separa los intrones se asocia a la cadena,

posteriormente intervienen otras snRNP que localizan el otro punto de separación entre
intrón y exón y forman un complejo llamado ESPLICEOSOMA. Cuando se forma este complejo
el intrón se pliega formando un bucle que corta una adenina y los exones forman un enlace
fosfodiéster provocando que la información codificante sea continua. En algunos textos este
fenómeno se denomina “SPLICING”
AÑADIR CAPERUZAS Y COLAS A LOS RNA
Tan pronto como el extremo 5’ emerge del RNA, otros enzimas añaden la denominada
caperuza 5’, esta caperuza es una molécula de metilguanosina y tres grupos fosfato.
Otros enzimas cortan el extremo 3’ y añaden un complejo de 100-250 nucleótidos de
adenina, formando la cola poli-A. Ahora el ARNm está completo para salir del núcleo. Esta
caperuza y cola sirve de señal de reconocimiento para la traducción. La cola de poli-A parece
que protege el RNA transcrito de degradaciones enzimáticas.

TRADUCCIÓN, BIOSÍNTESIS DE PROTEINAS
El código genético especifica la relación exstente entre un grupo de tres bases del RNAm
(CODON) y el aminoácido que codifica en la cadena proteica. En este proceso intervienen el
RNAm, el RNAt, los ribosomas y enzimas.
1- RNAm 2-
RNAt
Los RNA transferentes son intermediarios químicos que permiten a los aminoácidos
interactuar con una plantilla de RNAm. Los RNAt son moléculas cortas de unos 75-85
nucleótidos de longitud. Se comprobó que en algunos sitios de esta molécula se formaban
estructuras secundarias con zonas complementarias de la misma molécula, dándole aspecto
de trébol. Presentan una secuencia CCA en el extremos 3’ que es el sitio de unión al
aminoácido. El bucle del extremo opuesto sirve como anticodón, que es la zona de
acoplamiento por complementariedad de bases con el RNAm. Las bases que forman los
bucles no están apareadas. Cuando se estudió la estructura terciaria de este RNAt se
comprobó que la molécula se plegaba, obteniendo una forma de “L”.
3- Ribosomas
Son un conjunto de RNAr (ribosomal) unido a proteínas. Consta de dos subunidades,
denominadas grande y pequeña. Un diagrama tridimensional de un conjunto ribosomal;
durante la síntesis protéica, tres tipos de RNAt se alinean dentro del ribosoma. Los tres se
encuentran unidos al codón correspondiente del RNAm en la base de la estructura.
El RNAt que está situado a la derecha lleva un aminoácido, esta posición se denomina SITIO A,
(aceptor de aminoacil). El RNAt que está en el medio sujeta la cadena polipeptídica creciente
y ocupa el SITIO P, (por peptidil – piensa en la formación del enlace peptídico-). El tercer RNAt
situado a la izquierda SITIO E (de exit –salida-) +

SINTESIS DE PROTEÍNAS
Esta maquinaria molecular sintetiza proteínas en tres pasos:
1. Una aminoacil RNAt se difunde hacia el interior del sitio A, su anticodón se une al codón
del RNAm
2. Un enlace peptídico se forma entre el aminoácido sujeto por el aminoacil-RNAt en el sitio
A y el polipéptido creciente que es mantenido por un RNAt en el sitio P.
3. El ribosoma se mueve hacia adelante y los tres RNAt se mueven por detrás de la línea. El
RNAt en el sitio E se sale del complejo; el RNAt del sitio P se mueve al sitio E; el RNAt del
sitio A cambia al sitio P. El sitio A queda vacío para recibir otro RNAt cargado con su
aminoácido.
INICIACIÓN
Comienza cuando una región del RNAr de la subunidad pequeña del ribosoma se une a la
secuencia complementaria en el RNAm (secuencia de Shine-Dalgarno). El lugar está a seis
nucleótidos cuesta arriba del codón de iniciación AUG 5´ - AGGAGGU - 3’, la secuencia
complementaria en el RNAr de la subunidad pequeña se lee 3’ - UCCUCCA - 5’. La interacción
entre la subunidad pequeña y el mensajero está mediada por factores de iniciación. En los
eucariotas los factores de iniciación se unen a la caperuza 5’ de los mensajeros y guían al
ribosoma.
Cuando la secuencia de Shine-Dalgarnno se une a la subunidad pequeña, un R-aminoacil
RNAt se carga con una forma modificada de la metionina (formilmetionia) se une al codón
de iniciación AUG, Posteriormente la subunidad mayor completa el complejo.
ELONGACIÓN
Al comienzo el sitio E y A, están vacíos de RNA; por lo tanto, un codón de RNAm se encuentra
expuesto en la base del sitio A. Cuando llegan dos RNAt ocupan el sitio A y P, los aminoácidos

que llevan se colocan en el centro activo del complejo y tiene lugar la formación de un enlace
peptídico entre los aminoácidos.
Posteriormente a la formación del enlace, la cadena polipeptídica se transfiere del sitio P al
RNAt que ocupa en ese momento el sitio A. Este fenómeno se denomina translocación.
La translocación hace varias cosas a la vez: Mueve el RNAt vacio al sito E: mueve el RNAt que
contiene el polipéptido creciente en el sitio P y abre el sitio A para exponer un nuevo codón
del mensajero. Si el sitio E está ocupado por un RNAt es expulsado del complejo ribosomal
hacia el citosol.
TERMINACIÓN
Recuerda que el código genético incluye tres codones de terminación (UAA, UAG, UGA). En
la mayoría de las células ningún aminoacil-RNAt tiene un anticodón complementario. En
estas condiciones cuando una de esas tripletas queda expuesta en el sitio A, una proteína
denominada factor de liberación rellena el sitio A sin aportar ningún aminoácido. Al llegar al
centro activo del complejo ribosomal, se cataliza la hidrólisis del enlace que mantiene el
polipétido unido al RNAt y lo libera al citosol. Inmediatamente actuarán la proteína
denominadas “chaperonas” para plegar al polipéptido, adquiriendo así la funcionalidad
propia de la secuencia proteica.


1.1.5. EL CÓDIGO GENÉTICO
Este código es la clave que relaciona una secuencia nucleotídica del RNAm con una secuencia de
aminoácidos en las proteínas.
Si cada nucleótido determinara un aminoácido, solamente podríamos codificar cuatro
aminoácidos diferentes ya que en el ADN solamente hay cuatro nucleótidos distintos. Cifra muy
inferior a los 20 aminoácidos distintos que existen.
Si cada dos nucleótidos codificarán un aminoácido, el número total de dinucleótidos distintos
que podríamos conseguir con los cuatro nucleótidos diferentes (A, G, T y C) serían variaciones
con repetición de cuatro elementos tomados de dos en dos VR4,2 = 16. Por tanto, tendríamos
solamente 16 dinucleótidos diferentes, cifra inferior al número de aminoácidos distintos que
existen (20).
Si cada grupo de tres nucleótidos determina un aminoácido. Teniendo en cuenta que existen
cuatro nucleótidos diferentes (A, G, T y C), el número de grupos de tres nucleótidos distintos
que se pueden obtener son variaciones con repetición de cuatro elementos (los cuatro
nucleótidos) tomados de tres en tres: VR4,3 = 64. Por consiguiente, existe un total de 64 tripletes
diferentes, cifra más que suficiente para codificar los 20 aminoácidos distintos.
CARACTERÍSTICAS DEL CÓDIGO GENÉTICO:
• Un triplete de RNA (codón) determinan un aminoácido.
• Este código está degenerado ya que existen más codones (64) que aminoácidos (20)
• Es universal, ya que el mismo triplete codifica para el mismo aminoácido en diferentes
especies.
• Hay un triplete de iniciación, AUG, que codifica para la formil-metionina
• Existen tres tripletes o codones de terminación (UAA,UAG,UGA )
En el descubrimiento del código genético intervino Severo Ochoa, nacido en Luarca, Asturias,
aislando un enzima: polirribonucleótido fosforilasa capaz de sintetizar RNA sin molde. Sus
estudios los realizo en Alemania, Inglaterra y posteriormente en EEUU, donde se exilió en 1936
al comienzo de la guerra civil española.

REGULACIÓN DE LA EXPRESIÓN GÉNICA
En los años 60 del siglo pasado Jacob y Monod propusieron un modelo denominado operón para la regulación de la expresión génica
en las bacterias.
Un operón es un conjunto de genes que codifican proteínas que a su vez intervienen en una ruta metabólica. En cada operón se
pueden encontrar: genes reguladores, genes estructurales, la zona del promotor y la zona del operador. lacZ, lacY, lacA codifican
para la síntesis de proteínas (enzimáticas o reguladoras)
R codifican para la síntesis de una proteína represora que se puede encontrar de forma activa o inactiva y es el agente que controla materialmente la expresión. P Promotor, es la zona que se une al ARN polimerasa y decide el inicio de la transcripción. O Operador. Región entre el promotor y los genes estructurales que posee una secuencia característica reconocida por la proteína represora activa. Cuando se bloquea el operador con la proteína represora, impide el avance del ARN polimerasa y la transcripción se interrumpe,
(represión génica) Si la bacteria necesita sintetizar proteínas debe separar el operador del represor y utiliza dos caminos: la inducción
enzimática y la represión enzimática.
En el esquema anterior se representa la inducción enzimática (operón LAC): La lactosa inhibe a la proteína represora y entonces la
zona del operador queda libre y por lo tanto se sintetizan los enzimas que degradan la lactosa. Si no hay lactosa en el medio ¿Para qué
necesita la bacteria los enzimas que degradan la lactosa?
En la represión enzimática (operón HIS de histidina) se sintetiza un represor que es inactivo, esto implica que los genes se expresan
y se fabrica la histidina. Cuando hay en exceso, sus moléculas se unen a la proteína represora y la activa con lo cual se bloquea el
operador y se reprimen los genes. Si disminuye la histidina en el medio se desactiva el represor y los genes se expresan de nuevo.


1.2. Alteraciones de la información genética.
1.2.1. Concepto de mutación.
1.2.2. Causas de las mutaciones.
1.2.3. Consecuencias de las mutaciones.
1.2.3.1. Consecuencias evolutivas.
1.2.3.2. Efectos perjudiciales
1.2.1. Una de las características propias del material genético es la extraordinaria fidelidad con
la que se efectúan las copias durante los fenómenos de mitosis o meiosis; sin embargo, en
ocasiones pueden sufrir cambios que además se pueden transmitir a la descendencia. Estos
cambios son denominados MUTACIONES.
Algunas de estas mutaciones traducidas a proteínas pueden perjudicar al organismo anómalo,
incluso puede causar su muerte; sin embargo en ocasiones puede mejorar sus expectativas
vitales, manteniéndose (a partir de ese momento) en el “gen-pool” de la población. A veces las
mutaciones ni son beneficiosas ni son perjudiciales, al menos a simple vista.
1.2.2. CAUSAS DE LAS MUTACIONES
1) Por errores de lectura durante la replicación del ADN.
Los llamados errores de lectura pueden aparecer espontáneamente durante la replicación
del ADN y deberse a dos causas: a) A cambios tautoméricos:
Cada base nitrogenada puede presentarse en dos formas diferentes llamadas tautómeros
(formas tautoméricas), una normal y otra distinta. Ambas formas están equilibrio, y
espontáneamente se pasa de una a la otra, lo que se denomina cambio tautomérico. Si esto
sucede durante la replicación implica mutaciones, ya que cambia la base complementaria en
la nueva hebra de ADN. Por ejemplo, la forma normal de la G se complementa con la C,
mientras que la forma tautomérica (rara) de la G lo hace con la T. b) A cambios de fase:
Los cambios de fase son deslizamientos de la hebra que se está formando sobre la hebra
molde, de forma que quedan bucles al volverse a emparejar. El crecimiento sigue y la
diferencia queda fijada, originándose así la mutación.
2) Por lesiones fortuitas.
Son alteraciones espontáneas de la estructura de uno o varios nucleótidos, que aparecen de
forma natural. Las más frecuentes son:
a) Despurinización: Consiste en la pérdida de purinas (A, G) por rotura del enlace entre
éstas y las desoxirribosas.
b) Desaminación: Consiste en la pérdida de grupos amino en las bases nitrogenadas, que
entonces se emparejan con una distinta a la normal.
c) Dímeros de timina: Consiste en un enlace entre dos timinas contiguas, generalmente
provocados por la radiación UV solar.

3) Por transposiciones de ciertos segmentos del gen.
Son cambios de lugar espontáneos de determinados segmentos de ADN, los llamados
elementos genéticos transponibles. Éstos pueden ser menores que un gen, un gen, o un
grupo de genes (los denominados transposones).
Las transposiciones pueden producir mutaciones génicas si el elemento genético transpuesto
se sitúa dentro de un gen, o mutaciones cromosómicas si pasa a un lugar donde no hay un
gen, ya sea del mismo cromosoma o incluso a otro cromosoma.
Desde otro punto de vista podría considerarse que las mutaciones pueden ser causadas de forma
espontánea (mutaciones naturales) o ser inducidas de manera artificial (mutaciones inducidas)
mediante radiaciones y determinadas sustancias químicas a las que llamamos agentes
mutágenos. Así pues, y desde este punto de vista, las causas de las mutaciones pueden ser:
1) Las radiaciones, que, según sus efectos, pueden ser:
a) No ionizantes, como los rayos ultravioletas (UV) que son muy absorbidos por el ADN y
favorecen laa formación de enlaces covalentes entre pririmidinas contiguas (dímeros de
timin, por ejemplo) y la aparición de formas tautómeras que originan mutaciones génicas.
b) Ionizanates, como los rayos X y los rayos gamma, que son mucho más energéticos que
los UV; pueden originar formas tautómeras, romper los anillos de las bases nitrogenadas o
los enlaces fosfodiéster con la correspondiente rotura del ADN y, por tanto, de los
cromosomas.
2) Determinadas sustancias químicas que reaccionan con el ADN y que pueden provocar las
alteraciones siguientes:
a) Modificación de bases nitrogenadas. Así, el HNO2 las desamina, la hidroxilamina les
adiciona grupos hidroxilo, el gas mostaza añade grupos metilo, etilo, ...
b) Sustitución de una base por otra análoga. Esto provoca emparejamientos entre bases
distintas de las complementarias.
c) Intercalación de moléculas. Se trata de moléculas parecidas a un par de bases enlazadas,
capaces de alojarse entre los pares de bases del ADN. Cuando se produce la duplicación
pueden surgir inserciones o deleciones de un par de bases con el correspondiente
desplazamiento en la pauta de lectura.

TIPOS DE MUTACIONES:
Según las células afectadas pueden ser: Germinales y Somáticas. Resulta evidente que las mutaciones en células
somáticas solo producen alteraciones puntuales que no se transmiten a la descendencia. Si la mutación afectan a los
gametos se pueden transmitir a la descendencia y sobre ellas actuará la selección natural.
Según la extensión del material genético afectado pueden ser:
GÉNICAS: Provocan cambios en la secuencia de nucleótidos de un gen.
CROMOSÓMICAS: Afectan a la disposición de los genes de un cromosoma, pero no a la secuencia de nucleótidos.
GENÓMICAS: Son aquellas que alteran, aumentando o disminuyendo el número cromosómico de la especie.
Las causas de estas mutaciones ya las comenté en los apuntes de bioquímica (repásalo).
MUTACIONES GÉNICAS:
Se denominan mutaciones puntuales ya que afectan a un solo gen; pueden ocurrir por sustitución de bases y también
por cambios en las pautas de lectura:
TRANSICIONES: Se sustituye una púrica por otra púrica o una pirimidínica por otra pirimidínica.
TRANSVERSIONES: Se cambia una púrica por otra pirimidínica o al revés; el caso es que solo afecta a uno de
los nucleótidos y solo un triplete de bases. Puesto que el código genético esta degenerado (varios tripletes
codifican para el mismo aminoácido), puede ocurrir que esa mutación sea silenciosa y no afecte al fenotipo
del indivíduo. Si la mutación afecta a la tripleta final, puede ocurrir que la proteína sea más larga o más corta
de lo normal.
INSERCIONES Y DELECCIONES. Consiste en la pérdida o adición de algún nucleótido en la molécula de ADN. A
partir del punto de inserción o delección cambian todas las tripletas. Al traducirse en proteína puede que
sea totalmente distinta a la original.
MUTACIONES GENÓMICAS:
Son variaciones en el número normal de cromosomas de una especie. Ocurren por una incorrecta segregación de
cromosomas durante la meiosis. Dos grades grupos EUPLOIDIAS Y ANEUPLOIDIAS.
EUPLOIDIAS: Alteraciones del número normal de dotaciones cromosómicas. Existen dos tipos:
Monoploidias: Solo existe un cromosomas de cada par (solo se ha observado en algunas especies vegetales; muy raro)
Poliploidías: Tienen más de un juego completo de cromosomas, pudiendo ser triploides (3n), tetraploides (4n) o en
general poliploides. Se observan en vegetales que tienen generalmente mayor tamaño que las naturales. El trigo que
se cultiva hoy día es hexaploide.

ANEUPLOIDIAS:
Trisomías. Los portadores poseen un cromosoma de más (2n+1) si se da en autosomas. La más estudiada es la
trisomía del par 21 (mongolismo). El síndrome de Edwards (trisomía del 18); el síndrome de Patau (trisomía del 13).
Son poco frecuentes.
Si se da en cromosomas sexuales las trisomías son más frecuentes:
- Síndrome de Klinefelter (trisomía XXY). Hombres estériles, testículos poco desarrollados y con tendencia a
rasgos femeninos.
- Síndrome de triple X (trisomía XXX). Mujeres de aspecto normal, aunque en algún caso se podido describir
algún retraso mental.
- Cariotipo XYY (trisomía XYY). Hombres normales, más altos que la media con tendencia a padecer acné
intenso. En algún caso se ha descrito una vinculación con actitudes violentas.
- Monosomías: Los individuos carecen de un cromosoma de una pareja de homólogos. Su dotación cromosómica es
(2n-1). La carencia de un autosoma es letal. Sí puede faltar un cromosoma sexual (Síndrome de Turner XO). Mujeres
con escaso desarrollo de los caracteres sexuales primarios y secundarios; son estériles.
MUTACIONES CROMOSÓMICAS:
Son mutaciones que afectan a la estructura de los cromosomas; al orden de los genes, a su número, o un gen o a
grupos de genes.
Las traslocaciones e inversiones afectan poco al portador ya que no varía el número de genes. Pueden provocar
alteraciones cuando un gen se separa de las regiones que controlan su expresión (zona del operador, promotor o
zonas reguladores en la síntesis proteica). Las delecciones y duplicaciones, en cambio, aunque afecten solo a un
cromosoma de los homólogos pueden tener consecuencias graves. No basta con poseer todos los genes propios de
la especie, sino que han de estar en el número adecuado, para que existan desequilibrios en su expresión.

1.2.3. Consecuencias de las mutaciones.
1.2.3.1. Consecuencias evolutivas.
1.2.3.2. Efectos perjudiciales.
1.2.3.1. Consecuencias evolutivas
El concepto de mutación provocó una puntualización de la teoría de la Selección Natural
Darwinista. El redescubrimiento de las leyes de Mendel y la constatación de que las mutaciones
son la causa de la variabilidad genética, llevó a elaborar la teoría del neodarwinismo o teoría
sintética de la evolución biológica, Se denominó así gracias a que integra en una sola ley la
genética, la paleontología y la sistemática. Sus aspectos esenciales se deben a las aportaciones
de los siguientes autores:
Theodosius Dobzhansky que detalla en términos genéticos el proceso evolutivo.
Georges G. Simpson, paleontólogo que interpreta que las series filogenéticas fósiles es el
resultado de pequeños cambios a lo largo del tiempo; propone al igual que Darwin que la
especiación se consigue desde un antecesor común mediante pequeños cambios acumulados.
Ermst Mayr, introdujo el concepto de que la unidad evolutiva no es la especie sino la población.
Por lo tanto, en el neodarwinismo: Las mutaciones aportan la variabilidad genética sobre la que
luego actúa la selección natural en el proceso evolutivo. Algunas de estas mutaciones producen
variaciones individuales que permiten a su poseedor adaptarse a las condiciones ambientales y
tener mayores posibilidades de supervivencia, transmitiendo sus genes a la descendencia. Es
posible que las mutaciones impliquen una peor adaptación al medio ambiente y sus portadores
irán desapareciendo de la población. La evolución se produce de una manera gradual como
consecuencia de pequeños cambios que van surgiendo en una población, con lo que el proceso
para que aparezca una nueva especie es muy largo.
Alternativas al neodarwinismo:
Teoría neutralista de MOTOO KIMURA: Asegura que casi todas las mutaciones son neutrales y
su permanencia o no en la población es una cuestión de azar; así pues, existen periodos en los
que se observa una quietud en la frecuencia de mutaciones y otros periodos donde son más
evidentes. Se han construido árboles filogenéticos observando la secuencia de genes en
citocromos, comprobándose la existencia de antepasados comunes.
Teoría del equilibrio puntuado: Como no se han encontrado fósiles intermedios entre especies
conocidas, se desarrolló el modelo del equilibrio intermitente. Esta teoría asegura que existen
grandes periodos de estasis en los que no habido cambios apreciables, seguidos de periodos
cortos donde aparecen especiaciones. Esto explicaría la observación de fósiles vivientes muy
abundantes en una época y desaparecidos en otra. Por lo tanto, postula que la evolución no
puede ser gradual.
Gradualismo: Asegura que el proceso es gradual (como el punto de vista Darwinista), explicando
que el hecho de no encontrar fósiles intermedios es debido a que los organismos se

descomponen cuando mueren y sus restos no han fosilizado. A veces se encuentran fósiles
completos que avalan esta teoría.
Árbol filogenético construido con los cambios en la secuencia
de aminoácidos del citocromo C en diferentes especies.
Teoría del gen “egoísta”: Imagina a los genes como las únicas unidades de supervivencia que
usa a los organismos como máquinas automáticas para conseguir su perpetuación. El
determinismo y mecanicismo de esta teoría repugna a los neutralistas, puntualistas y
darwinistas. La competencia real se da en los genes que son la unidad básica de la evolución;
esta teoría se apoya en el darwinismo conservador ya que propone la supervivencia de los más
aptos y la evolución de los mejores adaptados. Comportamientos sociales en mamíferos
complejos como el altruismo, el instinto maternal, comportamientos heroicos etc. no son otra
cosa que intentar perpetuar los genes en la población.
Límites del neodarwinismo: los principales inconvenientes a esta teoría neodarwinista
provienen de la lentitud en la que se dan estos fenómenos y sobre todo a la hora de explicar
procesos como la macroevolución y especiación:
La microevolución como cambios pequeños y graduales se puede explicar desde una perspectiva
neodarwinista; sin embargo, la macroevolución (cambios que conciernen a grandes grupos
taxonómicos como las Clases u Órdenes) no está suficientemente esclarecidos. Respecto a la
especiación, los neodarwinistas proponen el mecanismo alopátrida; es decir, la separación

geográfica de una población. Otros autores proponen una especiación simpatrida; es decir, la
especiación gradual que ocurre cuando una especie pese a ocupar un mismo territorio
geográfico se diversifica en dos subpoblaciones debido a unos mecanismos que impiden el cruce
como son: Aislamiento ecológico, aislamiento estacional, aislamiento etológico, aislamiento
mecánico, aislamiento genético etc.
1.2.3.2 Efectos perjudiciales: MUTACIÓN Y CÁNCER
Para que una célula se transforme en tumoral y posteriormente en cancerígena es necesaria una
acumulación de mutaciones en los genes implicados en los procesos de división y proliferación
celular. Mientras las células se dividan y se mantengan unidas la situación se puede controlar,
solo aparecerá una tumoración por proliferación celular; sin embargo, cuando algunas células
se liberan de ese tumor (y a través del sistema circulatorio sanguíneo o linfático llegan a otros
tejidos y los “convence” para que hagan otro tumor), hacen una metástasis y entonces se
denominan cancerígenas.
Existen genes que están implicados en estos procesos:
Protoncogenes. Generalmente se encuentran reprimidos y su expresión podría provocar la
aparición de una proliferación celular.
Genes supresores de tumores. Son de dos tipos, los que controlan negativamente la
proliferación celular y los que están implicados en los procesos de diferenciación celular. Son los
denominados antioncogenes; generalmente se encuentran reprimidos.
Genes reparadores del ADN. Tienen como misión impedir la acumulación de mutaciones en el
ADN, generalmente están activados.
Cuando algunos de los protooncogenes muta puede transformarse en un oncogén capaz de
originar un proceso cancerígeno; si los supresores o reparadores del ADN mutan no se podría
controlar la proliferación celular.
Los primeros oncogenes fueron descubiertos en virus capaces de infectar células de mamíferos
y transformarlas en células cancerosas. Ahora se conocen más de cien de estos virus. Un
protooncogen puede transformarse en un oncogen mediante una mutación puntual (se
descubrió por primera vez en los tumores de vejiga urinaria). Se pudo comprobar que la
mutación solo cambiaba el aminoácido valina por glicina y así comenzaba el proceso tumoral.
También un protooncogen puede convertirse en oncogen cuando se asocian a nuevas regiones
reguladoras por rotura de cromosomas o transposiciones, o cuando se duplican, dando como
resultado la sobreexpresión del gen y la aparición del tumor.
Algunos retrovirus, papovirus (con ADN) y virus del herpes pueden provocar tumores. Este
aspecto ha sido comprobado en animales de granja, así que es más que probable su incidencia
en la especie humana.
Visita esta página
http://www2.uah.es/biomodel/citogene/horwitz/mshcance.htm