
Biología - yoquieroaprobar.es · Dentro de estas últimas, algunas atraviesan toda la membrana...
11
1 Biología
Transcript of Biología - yoquieroaprobar.es · Dentro de estas últimas, algunas atraviesan toda la membrana...

1B i o l o g í a

2B i o l o g í a

3B i o l o g í a

4B i o l o g í a

5© Oxford University Press España, S. A. B i o l o g í a
COMUNITAT VALENCIANA CONVOCATORIA SEPTIEMBRE 2009
S O L U C I Ó N D E L A P R U E B A D E A C C E S OAUTORA: María Purificación Hernández Nieves
Opción A� Las diferencias entre la pared celular y la membrana
celular son las siguientes:
Localización. La pared celular es una envuelta gruesaque rodea a la célula vegetal, mientras que la membra-na celular es una capa fina que se encuentra en todotipo de células (procariotas y eucariotas —animales yvegetales—).
Composición. La pared celular está compuesta porfibras de celulosa, como componente más abundante,embebidas en un entramado de hemicelulosas y pecti-nas. Presenta también glucoproteínas con consistenciade gel. Entre los constituyentes de la membrana plas-mática se encuentran las proteínas (integrales y periféri-cas), los fosfolípidos y algunos oligosacáridos.
Estructura. En las células diferenciadas, la pared celularaparece como una estructura gruesa compuesta porvarias capas, que se van depositando a medida que seproduce el crecimiento celular. De fuera adentro, estascapas son la lámina media (que puede ser compartidapor las células adyacentes de un tejido y está formadapor pectinas y proteínas, que se unen posteriormente aiones Ca2�), la pared primaria (constituida por largasfibras de celulosa cohesionadas por hemicelulosa, pecti-nas y glucoproteínas) y la pared secundaria (constituidapor una o varias capas fibrilares, semejantes en compo-sición a la pared celular primaria pero con una mayorproporción de celulosa y carentes de pectinas).
Las microfibrillas de celulosa se ordenan paralelamente,con diferente orientación, en las distintas capas. En oca-siones, entran a formar parte de su composición polí-meros como la lignina, ceras y la cutina o la suberina. Enalgunas paredes se observan, también, inclusionesminerales, principalmente carbonatos y sílice.
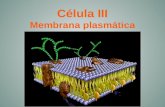
La estructura de la membrana celular es diferente.Según el modelo actualmente aceptado de Singer yNicholson o modelo de mosaico fluido, los lípidos y lasproteínas constituyen un mosaico molecular con unadisposición asimétrica en cuanto a la distribución de suscomponentes. Los lípidos mayoritarios son los fosfolípi-dos, que forman una bicapa hacia el interior de la mem-brana.
Las proteínas se distribuyen en las caras externa e inter-na de la membrana (proteínas extrínsecas) y alguna par-te se incluye en la bicapa lipídica (proteínas intrínsecas).Dentro de estas últimas, algunas atraviesan toda lamembrana (proteínas de canal). Uno de los componen-tes más llamativos de esta membrana son los glúcidos,
situados solo en la parte externa de la membrana enforma de glucolípidos y glucoproteínas y que constitu-yen el glucocálix.
Funciones
� La pared celular confiere rigidez y contribuye al man-tenimiento de la forma celular. La membrana celular,en cambio, es menos rígida. En las células animales,la rigidez la aporta la molécula de colesterol, peroesta rigidez no llega a parecerse nunca a la de lapared celular.
� La pared sirve de conexión entre las células de lostejidos vegetales, a los que dota de soporte y estruc-tura, lo que permite a la planta crecer erguida yalcanzar cierta altura. La conexión de los tejidos ani-males es la membrana celular.
� La pared celular permite a las células vegetales viviren un medio hipotónico impidiendo que se hincheny estallen. La membrana celular, en cambio, estallaríaen un medio hipotónico, por lo que su medio idóneoes el isotónico.
� La pared celular impermeabiliza la superficie vegetalen algunos tejidos; la membrana celular, no.
� La pared celular no tiene reconocimiento celular; lamembrana celular, sí. Este reconocimiento dependedel glucocálix, en concreto de las cadenas de oligosa-cáridos, que actúan como señales que deben serreconocidas para poder interrelacionarse con lascélulas que las poseen. Son los denominados recep-tores de membrana.
� La pared celular no posee fluidez; la membrana celu-lar, sí. La fluidez se debe a la presencia de los lípidos ylas proteínas, que se pueden desplazar en el plano dela bicapa lipídica. La fluidez también depende delmovimiento de difusión lateral que experimentan losfosfolípidos.
� Los lípidos de membrana son los fosfolípidos, los gluco-lípidos y el colesterol.
Los fosfolípidos pertenecen a los lípidos complejos o demembrana. Son lípidos saponificables que están forma-dos por el alcohol glicerina esterificada con dos ácidosgrasos, de los cuales el segundo es insaturado. El tercergrupo alcohol (�OH) de la glicerina se esterifica con unácido ortofosfórico (H3PO4) que, a su vez, puede estar este-rificado con un derivado aminado o de un polialcohol.
Los fosfolípidos tienen una propiedad que les permiteformar la estructura básica de las membranas celulares:su carácter anfótero o polar.
Bloque 1

fimbrias
flagelo
pared celular
cápsula
ribosomas
inclusiones
nucleoide
membranaplasmática
citoplasma
En la molécula de un fosfolípido hay dos zonas: unapolar o hidrófila y otra apolar o hidrófoba. La zona polarestá constituida por el resto de la glicerina y el ácido fos-fórico; la zona apolar la forman el resto de los dos ácidosgrasos. Debido a esto, cuando los fosfolípidos seencuentran en contacto con el agua, interaccionan conesta molécula a través de la zona hidrófila, mientras quelas cadenas de los ácidos grasos, repelidas por el agua,interaccionan con las cadenas alifáticas de las moléculasvecinas formando bicapas que se cierran sobre sí mis-mas y dan lugar a las membranas biológicas.
Bicapa de fosfolípidos.
Como se observa en la figura, las regiones polares ohidrófilas se orientan hacia el agua y las regiones hidró-fobas (lipófilas), hacia dentro.
Los glucolípidos son muy semejantes a los fosfolípidos.En las células animales suelen ser derivados de esfingo-lípidos. En las células vegetales y en las bacterias, sinembargo, derivan de los fosfoglicéridos. Se encuentranen la cara externa de la membrana plasmática, forman-do parte del glucocálix.
El colesterol es un lípido insaponificable, derivado delciclopentanoperhidrofenantreno, cuya estructura lacomponen tres anillos de ciclohexano unidos a un ciclo-pentano.
El colesterol forma parte de las membranas de las célu-las animales, a las que proporciona rigidez y, por tanto,consistencia. Se encuentra también unido a lipoproteí-nas en el plasma sanguíneo.
Además, el colesterol es una molécula precursora deotros esteroides, entre los que cabe citar las hormonassexuales, los corticoides, los ácidos biliares y el 7-deshi-drocolesterol, molécula que se transforma en vitamina D.
�
Las envueltas bacterianas son la membrana celular y lapared celular. Algunas bacterias presentan, además,cápsula. La membrana celular tiene una serie de invagi-naciones denominadas mesosomas. En alguno de ellosse encuentran enzimas respiratorias. Las bacterias foto-sintéticas tienen, en determinados mesosomas, pig-mentos fotosintéticos o cromatóforos. Otros mesoso-mas albergan enzimas respiratorias.
El citoplasma presenta una molécula de ADN, que es elúnico cromosoma que posee la bacteria. Algunas tienenpequeñas moléculas de ADN circular conocidas con elnombre de plásmidos. Los ribosomas son más peque-ños que los de la célula eucariota y se encuentran espar-cidos por todo el citoplasma. Como medio de locomo-ción presentan flagelos.
El componente mayoritario de la pared bacteriana es elpeptidoglicano o mureína. Está formado por cadenaspolisacarídicas compuestas por azúcares: N-acetilgluco-samina (NAG) y ácido N-acetilmurámico (NAM) unidospor enlaces glucosídicos. Al NAM se une, a su vez, unacorta cadena de cuatro aminoácidos. En las bacteriasGram positivas hay, además, ácidos teicoicos, y en lasGram negativas existe una membrana externa cuyocomponente mayoritario son los lipopolisacáridos.
6© Oxford University Press España, S. A. B i o l o g í a
COMUNITAT VALENCIANA CONVOCATORIA SEPTIEMBRE 2009
agua
agua
colasapolares
cabezaspolares
ácidosteicoicos
lipopolisacárido(LPS)
8 nm
peptidoglicanoo mureína
membranaplasmática
fosfolípido
lípido A proteína
porinaexterior
membranaexterna
espacioperiplásmico
interior
A B
Estructura de la pared bacteriana de tipo Gram positivo (A) y Gram negativo (B).

Opción B� Una proteína es una molécula tridimensional, con
estructura terciaria (algunas, cuaternaria), que surge porplegamiento en el espacio de una o varias cadenas poli-peptídicas.
Los principales elementos constituyentes de las proteí-nas son el C, el O, el N y el H. Estos elementos forman elmonómero de estas macromoléculas, que es el aminoá-cido. La unión de los aminoácidos mediante enlacespeptídicos da lugar a la cadena peptídica.
� Una proteína conjugada o heteroproteína es una molé-cula que presenta una parte proteica y otra no proteica,llamada grupo prostético. Todas son globulares y se cla-sifican, en función del grupo prostético, en:
� Fosfoproteínas. Su grupo prostético es el ácido fos-fórico. Son de carácter ácido. Ejemplos de estas pro-teínas son la vitelina y la caseína.
� Glucoproteínas. Su grupo prostético es un glúcido.Desempeñan funciones enzimáticas, hormonales, decoagulación, etc. Destacan las inmunoglobulinas.
� Lipoproteínas. Su grupo prostético es un lípido.Abundan en las paredes bacterianas, en las membra-nas mitocondriales y en el suero sanguíneo. Un ejem-plo de lipoproteínas son los quilomicrones.
� Nucleoproteínas. Su grupo prostético está formandopor ácidos nucleicos. Hay dos tipos: las que presen-tan ARN (que forman los ribosomas) y las que tienenADN (que forman los cromosomas).
� Cromoproteínas. Su grupo prostético es una molé-cula compleja que presenta coloración debido a la
presencia de metales. Destacan los pigmentos respi-ratorios (hemoglobina y mioglobina) y las proteínasque intervienen en la transferencia de electrones(citocromos y flavoproteínas).
� Las principales funciones biológicas de las proteínasson:
� Estructural. Las membranas biológicas tienen comocomponente estructural las proteínas. El colágeno esuna proteína fibrosa que se encuentra en la sustanciaintercelular de los tejidos conjuntivo, cartilaginoso yóseo. La queratina es un elemento constituyente delas uñas, pelos, pezuñas, escamas de los reptiles,etcétera.
� Enzimática. Las enzimas son proteínas que catalizancasi todas las reacciones químicas en la célula: sonbiocatalizadores. Un ejemplo es la lactasa, que trans-forma la lactosa en galactosa y glucosa.
� Transportadora. Algunas proteínas se unen a molé-culas o iones específicos y los transportan a otrolugar. Un ejemplo es la hemoglobina, que transportael O2 en la sangre de los vertebrados.
� Contráctil. Existen proteínas que intervienen en lossistemas motiles y contráctiles: son la actina y la mio-sina, que intervienen en la contracción muscular.
� De reserva. Algunas son reserva de aminoácidos,como la albúmina de la sangre y de la clara del hue-vo, la caseína de la leche, o las proteínas de las semi-llas de las plantas.
� De defensa. Hay proteínas que actúan como mediode defensa contra virus, bacterias y sustancias extra-ñas; son las inmunoglobulinas y los interferones.
7© Oxford University Press España, S. A. B i o l o g í a
COMUNITAT VALENCIANA CONVOCATORIA SEPTIEMBRE 2009
Opción A� Cuando hay una bacteria en el exterior de un macrófa-
go, esta célula, por expansión de su membrana, emitepseudópodos con los cuales rodea dicha bacteria y laintroduce en su interior, en una vesícula fagocítica ofagosoma. Entonces actúan los lisosomas, vesículas for-madas en el complejo de Golgi por estrangulación delos sáculos del dictiosoma.
Los lisosomas son los orgánulos implicados en la diges-tión celular, pues contienen las enzimas digestivas queintervendrán en la degradación de las partículas fagocita-das. Como los macrófagos son células fagocitarias, con-tienen un abundante número de lisosomas. El alto con-tenido de enzimas hidrolíticas se sintetiza en el retículoendoplásmico rugoso, desde el cual pasan a las vesícu-las del complejo de Golgi para formar los lisosomas.
Algunos de los lisosomas (lisosomas secundarios) llevana cabo la digestión intracelular. Para ello se unen a lavesícula digestiva o fagosoma, que contiene la bacteria
capturada por fagocitosis, y forman con ella una vacuoladigestiva, en la cual tiene lugar la degradación de labacteria por la acción de las enzimas digestivas aporta-das desde el lisosoma, ahora convertido en heteroliso-soma.
Fagocitosis de una bacteria.
� La relación que existe entre el retículo endoplásmico, elaparato de Golgi y los lisosomas es la siguiente:
A través de las vesículas de transición originadas en suscisternas, el retículo endoplásmico rugoso conecta con
Bloque 2
productos de la digestión
fagolisosoma
vacuola fagocítica
complejo de Golgi
bacteria
lisosomasprimarios

la cara cis o de formación del aparato de Golgi y le trans-fiere glicoproteínas para completar el proceso de glico-silación (adición de nuevos restos glucídicos). Es en elaparato de Golgi donde estas proteínas adquieren suforma activa. Por ejemplo, en las células pancreáticas, elretículo endoplásmico forma proinsulina, pero el proce-so se completa en el aparato de Golgi, donde se formala insulina.
El sistema retículo endoplásmico-aparato de Golgi estáimplicado en importantes procesos celulares. Por ejem-plo, participa en la formación de la pared celular vegetale interviene en el tránsito de lípidos en las glándulassebáceas, sudoríparas o de la bilis de los hepatocitos yen la formación de los lisosomas.
Opción B� Se entiende por ciclo celular el tiempo que transcurre
desde que una célula se forma, por división de una pre-existente, hasta que se divide y da origen a dos célulashijas. Comprende dos fases (interfase y división celular)y su duración depende del tipo de célula y de los facto-res ambientales. La división celular (mitosis y citocine-sis) es una de las fases de ese ciclo.
La diferencia entre la mitosis y la citocinesis es que laprimera es el proceso que conduce a la división delnúcleo celular, mientras que la segunda es el procesopor el cual se divide el citoplasma. La citocinesis tienelugar después de la mitosis y es diferente en las célulasanimales y las vegetales. En las primeras tiene lugar porestrangulación del citoplasma y en las segundas se reali-za por tabicación intracelular.
La diferencia entre centrómero y cinetocoro es que elcentrómero, denominado también constricción prima-ria, es un estrechamiento que divide el cromosoma endos fragmentos llamados brazos cromosómicos. Las cro-mátidas hermanas están unidas por el centrómero. Elcinetocoro, en cambio, es un disco proteico que se sitúaa ambos lados del centrómero, en cada cromátida. Es ellugar por donde las cromátidas quedan unidas a lasfibras cinetocóricas del huso acromático.
� Paquiteno es la etapa de mayor duración de la profase Ide la meiosis. Comienza con el apareamiento más ínti-mo de los cromosomas homólogos. En cada tétrada haydos pares de cromátidas hermanas (cromátidas de unmismo cromosoma). A continuación, se produce unintercambio entre cromátidas homólogas, llamadorecombinación genética, como consecuencia de lasroturas transversales en las cromátidas, el intercambiode segmentos y la posterior unión cruzada de los extre-mos rotos. De este modo, fragmentos que antes perte-necían a cromátidas homólogas acaban formando partedel mismo ADN. Esto se conoce como sobrecruzamien-to, entrecruzamiento o crossing-over, y los puntos dondese producen se denominan quiasmas.
� La meiosis es necesaria para asegurar el número de cro-mosomas de la especie. Esta división reduce a la mitadel número de cromosomas en los gametos o célulassexuales, y en ella tiene lugar la recombinación genéti-ca, cuya consecuencia es la aparición de nuevas combi-naciones de genes que, junto con las mutaciones, son lacausa de la variabilidad genética y, por tanto, de la evo-lución de las poblaciones.
8© Oxford University Press España, S. A. B i o l o g í a
COMUNITAT VALENCIANA CONVOCATORIA SEPTIEMBRE 2009
membrana plastidial externa
espaciointermembranoso
membrana tilacoidal
tilacoide de los grana
estromaADN tilacoideintergrana
membrana plastidial interna
espaciotilacoidal
granum
ribosomas 70 S
� �
��
�
Bloque 3
Opción A�
Esquema general de la estructura de un cloroplasto.
La fase luminosa tiene lugar en la membrana interna(tilacoide) y la fase oscura, en el estroma.
� Los principales pigmentos fotosintéticos son la clorofila ylos carotenoides. En las bacterias, las bacterioclorofilas.
Estos pigmentos captan la luz solar en la fase luminosade la fotosíntesis. Los carotenoides absorben luz a unalongitud de onda diferente a la clorofila, por lo queensanchan el abanico de luz que puede captarse. Losprincipales carotenoides son el β-caroteno y la xantofila.Las clorofilas más importantes son la clorofila a y la clo-rofila b.
Estos pigmentos se encuentran asociados a proteínasen la membrana tilacoidea, formando los denominadoscomplejos antena. En ellos, las diferentes clorofilas ycarotenoides transfieren la energía luminosa a una clo-rofila especial llamada clorofila del centro de reacción,para convertirla, más tarde, en energía química.
� La fotosíntesis es el proceso anabólico por el cual lasplantas, algas y algunas bacterias captan la energía de laluz y la convierten en energía química (ATP) y en poderreductor (NADPH2) para poder transformar los com-puestos inorgánicos (H2O, sales minerales y CO2) encompuestos orgánicos.

9© Oxford University Press España, S. A. B i o l o g í a
COMUNITAT VALENCIANA CONVOCATORIA SEPTIEMBRE 2009
La quimiosíntesis es también un tipo un tipo de nutri-ción autótrofa y permite, igualmente, obtener materiaorgánica a partir de materia inorgánica; en este caso, sinembargo, se utiliza como fuente de energía la liberadaen las reacciones químicas redox exergónicas.
Opción B� La naturaleza química de una enzima es proteica. Algu-
nas son proteínas puras y otras presentan, junto a la par-te proteica (apoenzima), una parte no proteica (el cofac-tor), que puede ser un ión metálico (Cu2+, Zn2+, etc.) ouna molécula de naturaleza orgánica (NAD, FAD, etc.).En este último caso, el cofactor recibe el nombre decoenzima.
Las enzimas son biocatalizadores que intervienen en lasreacciones a concentraciones muy bajas, acelerando lavelocidad de las reacciones en las que participan. Noexperimentan modificación alguna (se obtienen al finalde la reacción sin gastarse) y actúan en condicionesrelativamente constantes de temperatura, pH, presión,etcétera.
Las enzimas, como proteínas que son, presentan un altogrado de especificidad, que puede ser de sustrato (algu-nas muestran una especificidad absoluta sobre un sus-trato, mientras que otras muestran su especificidadsobre un grupo funcional) y de reacción (para cada tipode reacción química existe una enzima diferente, esdecir, una enzima cataliza una sola reacción química oun grupo de reacciones estrechamente relacionadasentre sí).
� La inhibición es la disminución y posterior parálisis de laactividad enzimática. Se produce cuando en la reacciónenzimática aparece un inhibidor. La inhibición puedeser de dos tipos: irreversible y reversible.
� La inhibición irreversible se caracteriza porque elinhibidor altera permanentemente la estructura de laenzima y la deja inactiva para volver a actuar. El inhi-bidor se une a la enzima mediante enlaces fuertes.
� La inhibición reversible tiene lugar cuando la enzimavuelve a tener actividad una vez que se elimina lasustancia inhibidora. La unión del inhibidor a la enzi-ma tiene lugar mediante enlaces débiles, fáciles deromper. Según el lugar de unión del inhibidor a laenzima, se distinguen dos tipos de inhibición reversi-ble: competitiva y no competitiva.
� Inhibición competitiva. En ella, el inhibidor y elsustrato son moléculas de naturaleza muy pareci-da, que entran en competencia por llegar al centroactivo de la enzima. Si este es ocupado por el sus-trato, habrá catálisis enzimática, pero si es ocupa-do por el inhibidor, habrá inhibición enzimática. Lainhibición competitiva se caracteriza porque, apa-rentemente, la enzima tiene menor afinidad por elsustrato y porque, si en el medio se pone suficien-te concentración de sustrato, este es capaz de des-plazar al inhibidor del centro activo de la enzima.
� Inhibición no competitiva. En este caso, el inhibi-dor no compite con el sustrato, sino que se une ala enzima por otra zona distinta del centro activo.Otras veces el inhibidor se une al complejo enzi-ma-sustrato una vez creado este e impide la for-mación de productos.
Para invertir una inhibición competitiva habría quesuministrar, en presencia del inhibidor, muchas molécu-las de sustrato para que este compitiera con el inhibidory lograra alcanzar la velocidad máxima que la reaccióncatalítica tendría sin su presencia. En este caso, serequerirían grandes concentraciones de sustrato. Sinembargo, en presencia de un inhibidor irreversible estono sería posible, ya que este alteraría la estructura mole-cular de la enzima, lo que imposibilitaría la unión delsustrato, con lo cual la catálisis no podría llevarse a cabo.
� Los factores que afectan a la actividad enzimática sonaquellos factores que hacen que esta pierda sus propie-dades características; por ejemplo, cambios en el pH delmedio, en la temperatura, la presión, etc., y los llamadosinhibidores enzimáticos (moléculas que, al unirse a laenzima, paralizan la catálisis enzimática).
� Variaciones de temperatura. La enzima, como todaproteína, presenta una estructura terciaria o cuater-naria. Si se produjese un cambio notable de tempera-tura, la proteína enzimática se desnaturalizaría (per-dería su estructura característica) y, por consiguiente,perdería su funcionalidad.
� Cambios de pH. Si se producen cambios bruscos depH en el medio, la enzima se desnaturaliza y pierdesu función. Sin embargo, el pH óptimo es diferentepara las enzimas de un mismo organismo. Cada unatiene un pH óptimo en el cual realiza su función coneficacia.
� El efecto de la dilución. La mayoría de las enzimas ysustratos se encuentran en la célula en concentracio-nes muy bajas, lo que supone un gran inconvenientepara la velocidad de la reacción, que se ve limitadapor la posibilidad de que el sustrato se encuentre consu enzima. Para ello, en el ambiente celular se hancreado una serie de estrategias, como la comparti-mentación celular y los complejos multienzimáticos.
� Inhibición de la actividad enzimática. Los inhibido-res enzimáticos son sustancias que disminuyen o,incluso, anulan la velocidad de la reacción.
� En la inhibición reversible, el inhibidor presentauna unión temporal con la enzima y, una vez quecesa la unión, la enzima se recupera y se reanuda lacatálisis enzimática. Se distinguen dos tipos deinhibición reversible: competitiva y no competiti-va. En la primera, el sustrato y el inhibidor presen-tan una estructura molecular muy parecida y com-piten por el centro activo de la enzima. Si elsustrato llega antes no habrá inhibición, sino catá-lisis enzimática; en cambio, si es el inhibidor el queocupa el centro activo de la enzima, habrá inhibi-

10© Oxford University Press España, S. A. B i o l o g í a
COMUNITAT VALENCIANA CONVOCATORIA SEPTIEMBRE 2009
ción enzimática. En la inhibición no competitiva, elinhibidor se une a un centro diferente al centroactivo, denominado centro regulador o alostérico.Si el inhibidor está alojado en este centro, esté ono el sustrato situado en el centro activo, se produ-ce inhibición enzimática.
� En la inhibición irreversible, la unión del inhibidorcon la enzima provoca en la estructura de esta unaserie de cambios irreversibles que anulan la capa-cidad catalítica para siempre.
Bloque 4
Opción A� Virulencia de un microorganismo es su capacidad para
provocar la aparición y el desarrollo de una enfermedadinfecciosa.
Las toxinas son sustancias producidas por ciertos pató-genos y resultan perjudiciales para la célula. Cuandoestas toxinas se liberan al exterior, durante el crecimien-to del microorganismo, se denominan exotoxinas, y, sison componentes del propio microorganismo y única-mente se liberan cuando la célula se lisa, se llamanendotoxinas.
� a) La tolerancia del sistema inmune es su capacidadpara reconocer los antígenos propios y no rechazarlos.
b) La inmunodeficiencia es la incapacidad del sistemainmune para desarrollar una respuesta inmunitariaadecuada ante antígenos extraños. Estos antígenosno pueden ser eliminados correctamente.
c) La autoinmunidad es una respuesta inmunitariaprovocada por antígenos no asociados a microorga-nismos infecciosos. Es una reacción de los mecanis-mos de defensa contra el mismo organismo que losalberga.
� Las diferencias fundamentales entre la vacunación(introducción de vacunas) y la sueroterapia (tratamientode enfermedades con suero) son las siguientes:
Como se observa, los sueros producen inmunidad pasi-va y las vacunas, inmunidad activa.
Opción B� Existen tres hipótesis que explican la replicación o auto-
duplicación del ADN:
Conservativa. En ella, la doble cadena original se man-tiene y se sintetiza otra completamente nueva.
Semiconservativa. Consiste en la formación de dosnuevas moléculas de ADN de doble hélice, en la queuna de las hebras es antigua (actúa como molde) y laotra, de nueva formación (constituida por la polimeriza-ción de nucleótidos libres dispuestos enfrente del mol-de, por complementariedad de bases nitrogenadas). Esel modelo de Watson y Crick.
Dispersiva. En cada doble hélice existen fragmentos dela cadena original y fragmentos nuevos.
� Las enzimas helicasas, topoisomerasas y girasas desen-rollan y abren la doble hélice de la molécula de ADN,para lo cual rompen los puentes de hidrógeno quemantienen unidas las cadenas y forman una horquillade replicación.
La ARN-polimerasa sintetiza una molécula pequeña deARN cebador o primer, sin el cual la ADN-polimerasa nopuede ejercer su función.
Vacunación Sueroterapia
Produce inmunidad activa. Produce inmunidad pasiva.
Se inyectan antígenosatenuados o inactivos.
Se inyectan anticuerposespecíficos.
El organismo elaboraanticuerpos que provocanuna respuesta primaria.
El organismo no elaboraanticuerpos.
Genera memoriainmunológica frente alantígeno.
No genera memoriainmunológica.
La inmunidad frente a laenfermedad espermanente.
La duración es de unosmeses.
Requiere varios días paraproducir su efecto.
El efecto es inmediato(pocas horas).
�
conservativa
�
semiconservativa
�
dispersiva

11© Oxford University Press España, S. A. B i o l o g í a
COMUNITAT VALENCIANA CONVOCATORIA SEPTIEMBRE 2009
La ADN-polimerasa sintetiza las nuevas cadenas deADN. La ADN-polimerasa III va alargando la cadena,leyendo en dirección 3’ → 5’ y uniendo nucleótidos endirección 5’ → 3’. Como las dos cadenas de ADN moldeson antiparalelas, la cadena de ADN que tiene dirección3’ → 5’ se replica de forma continua (hebra conductora),mientras que la que tiene dirección 5’ → 3’ lo hace deforma discontinua (hebra retardada). La ADN-polimera-sa I elimina los fragmentos de ARN que han actuadocomo iniciadores de la síntesis y rellena los huecos deja-dos por estos.
Por último, las ADN-ligasas unen los fragmentos de Oka-zaki y, de este modo, se forma la molécula completa.
� La síntesis de las nuevas cadenas es continua en una deellas y discontinua en la otra, debido a que las cadenasmolde de ADN son antiparalelas y la ADN-polimerasalee siempre en dirección 3’→ 5’. Esta síntesis se realizade la siguiente manera:
Primero actúa una ARN-polimerasa, que sintetiza unamolécula pequeña de ARN cebador o primer, sin el cualla ADN-polimerasa no puede iniciar su función. A conti-nuación, la ADN-polimerasa III va alargando la cadena,leyendo en dirección 3’ → 5’ y uniendo nucleótidos endirección 5’ → 3’. Como las dos cadenas de ADN molde
son antiparalelas, la cadena de ADN que tiene dirección3’ → 5’ se replica de forma continua (hebra conductora),mientras que la que tiene dirección 5’ → 3’ lo hace deforma discontinua (hebra retardada). Esta última sereplica mediante la síntesis de pequeños fragmentos,llamados fragmentos de Okazaki. Posteriormente, actúala ADN-polimerasa I, que elimina los fragmentos de ARNque han actuado como iniciadores de la síntesis y relle-na los huecos dejados por estos. Finalmente, las ADN-ligasas unen los fragmentos de Okazaki y, de este modo,se forma la molécula completa.
Replicación del ADN.
5'
5'
5'
3'
3'
3'
topoisomerasashelicasa
ADN polimerasa III
ARNcebador
fragmentosde Okazaki
hebraconductora
hebra
proteínas SSB
dirección generalde la replicación

















![[PPT]LA MEMBRANA PLASMÁTICA - Biologia Blog | … · Web viewTEMA 8. LA MEMBRANA PLASMÁTICA Y LAS ENVOLTURAS EXTERNAS 1. LA MEMBRANA PLASMÁTICA 1.1. MEMBRANA UNITARIA Membrana](https://static.fdocuments.ec/doc/165x107/5bcd58cf09d3f206798d90a2/pptla-membrana-plasmatica-biologia-blog-web-viewtema-8-la-membrana-plasmatica.jpg)